
Coarse neural tuning for print peaks when children learn to read
10
Coarse neural tuning for print peaks when children learn to read Urs Maurer, 1,2 Silvia Brem, Felicitas Kranz, Kerstin Bucher, Rosmarie Benz, Pascal Halder, Hans-Christoph Steinhausen, and Daniel Brandeis ⁎ ,1 Department of Child and Adolescent Psychiatry, University of Zurich, Brainmapping Research, Neumunsterallee 9, CH-8032 Zurich, Switzerland Received 7 February 2006; revised 9 June 2006; accepted 23 June 2006 Available online 21 August 2006 Adult readers exhibit increased fast N1 activity to wordlike strings in their event-related brain potential. This increase has been linked to visual expertise for print, implying a protracted monotonic develop- ment. We investigated the development of coarse neural tuning for print by studying children longitudinally before and after learning to read, and comparing them to skilled adults. The coarse N1 tuning, which had been absent in nonreading kindergarten children, emerged in less than 2 years after the same children had mastered basic reading skills in 2nd grade. The N1 became larger for words than symbol strings in every child, and this coarse tuning was stronger for faster readers. Fast brain processes thus specialize rapidly for print when children learn to read, and play an important functional role in the fluency of early reading. Comparing 2nd graders with adults revealed a further decrease of the coarse N1 tuning in adults, presumably reflecting further reading practice. This constitutes a prominent nonlinear development of coarse neurophysiological specialization for print. The maximum tuning in novice readers possibly reflects the high sensitivity of their neural network for visual aspects of print, and a more selective tuning in expert adult readers. © 2006 Elsevier Inc. All rights reserved. Introduction Skilled adult readers activate fast visual brain processes which are tuned for print (Cohen and Dehaene, 2004; McCandliss et al., 2003). Such fast visual specialization is a critical first step in categorizing, identifying, and recognizing visual stimuli for which an individual has developed expertise (Palmeri et al., 2004). Since learning to read represents a developmental milestone in literate societies, we examined the development of this fast visual specialization thought to emerge many years after reading is first learned (McCandliss and Noble, 2003), along with effortless and fluent reading (Cohen and Dehaene, 2004). We did this by studying the same children immediately before and shortly after basic reading acquisition and by comparing them to adult readers (Maurer et al., 2005a), in order to determine whether this fast specialization would emerge after 1.5 years of reading training in school, and how it would develop from novice reading in 2nd grade to expert reading in adulthood. These children had only minimal fast visual specialization for words when tested as nonreading kindergartners 3 (Maurer et al., 2005a), indicating that most of the specialization indeed develops after reading training starts. In adult readers, visual specialization for print over visually matched strings of symbols is first reflected by increased neurophysiological activity between 140 and 200 ms in the occipitotemporal N1 or N170 component of the event-related potential (ERP) and its magnetic field analogue (Bentin et al., 1999; Brem et al., 2005; Maurer et al., 2005a,b; Schendan et al., 1998; Tarkiainen et al., 1999). This N1 specialization reflects coarse automatic visual categorization, as it extends to implicit reading tasks, and to consonant strings and even pseudofonts (Bentin et al., 1999; Maurer et al., 2005a; Schendan et al., 1998; Tarkiainen et al., 1999). It has also been linked to fluent reading because it is reduced in dyslexic adults (Helenius et al., 1999). The N1 specialization presumably originates from inferior occipito- temporal regions predominantely of the left hemisphere (Brem et al., 2006; Nobre et al., 1994; Tarkiainen et al., 1999). Visual N1 specialization also extends to categories of personal expertise, with bird-experts having larger N1 amplitudes to bird than dog stimuli and dog-experts showing the reverse pattern (Tanaka and Curran, 2001), and plays an important role for face processing (Gauthier et al., 2003). It can develop rapidly with practice for novel forms (Curran et al., 2002; Rossion et al., 2002) or symbols (Brem et al., 2005) in adults. Developmental ERP studies in children who had already mastered basic reading skills demonstrate that the word N1 does not reflect the maturation of specific phonological or semantic processes (Grossi et al., 2001; Holcomb et al., 1992). The word N1 www.elsevier.com/locate/ynimg NeuroImage 33 (2006) 749 – 758 ⁎ Corresponding author. Fax: +41 43 499 2604. E-mail address: [email protected] (D. Brandeis). 1 Contributed equally to this work. 2 Now at the Sackler Institute for Developmental Psychobiology, Weill Medical College of Cornell University, New York, USA. Available online on ScienceDirect (www.sciencedirect.com). 3 In Switzerland, children do not receive early literacy instruction in kindergarten, and reading instruction starts with school at age 7. 1053-8119/$ - see front matter © 2006 Elsevier Inc. All rights reserved. doi:10.1016/j.neuroimage.2006.06.025
Transcript of Coarse neural tuning for print peaks when children learn to read

www.elsevier.com/locate/ynimg
NeuroImage 33 (2006) 749–758Coarse neural tuning for print peaks when children learn to read
Urs Maurer,1,2 Silvia Brem, Felicitas Kranz, Kerstin Bucher, Rosmarie Benz, Pascal Halder,Hans-Christoph Steinhausen, and Daniel Brandeis⁎,1
Department of Child and Adolescent Psychiatry, University of Zurich, Brainmapping Research, Neumunsterallee 9, CH-8032 Zurich, Switzerland
Received 7 February 2006; revised 9 June 2006; accepted 23 June 2006Available online 21 August 2006
Adult readers exhibit increased fast N1 activity to wordlike strings intheir event-related brain potential. This increase has been linked tovisual expertise for print, implying a protracted monotonic develop-ment. We investigated the development of coarse neural tuning forprint by studying children longitudinally before and after learning toread, and comparing them to skilled adults. The coarse N1 tuning,which had been absent in nonreading kindergarten children, emergedin less than 2 years after the same children had mastered basic readingskills in 2nd grade. The N1 became larger for words than symbolstrings in every child, and this coarse tuning was stronger for fasterreaders. Fast brain processes thus specialize rapidly for print whenchildren learn to read, and play an important functional role in thefluency of early reading. Comparing 2nd graders with adults revealeda further decrease of the coarse N1 tuning in adults, presumablyreflecting further reading practice. This constitutes a prominentnonlinear development of coarse neurophysiological specialization forprint. The maximum tuning in novice readers possibly reflects the highsensitivity of their neural network for visual aspects of print, and amore selective tuning in expert adult readers.© 2006 Elsevier Inc. All rights reserved.
Introduction
Skilled adult readers activate fast visual brain processes whichare tuned for print (Cohen and Dehaene, 2004; McCandliss et al.,2003). Such fast visual specialization is a critical first step incategorizing, identifying, and recognizing visual stimuli for whichan individual has developed expertise (Palmeri et al., 2004). Sincelearning to read represents a developmental milestone in literatesocieties, we examined the development of this fast visualspecialization thought to emerge many years after reading is firstlearned (McCandliss and Noble, 2003), along with effortless and
⁎ Corresponding author. Fax: +41 43 499 2604.E-mail address: [email protected] (D. Brandeis).
1 Contributed equally to this work.2 Now at the Sackler Institute for Developmental Psychobiology, Weill
Medical College of Cornell University, New York, USA.Available online on ScienceDirect (www.sciencedirect.com).
1053-8119/$ - see front matter © 2006 Elsevier Inc. All rights reserved.doi:10.1016/j.neuroimage.2006.06.025
fluent reading (Cohen and Dehaene, 2004). We did this bystudying the same children immediately before and shortly afterbasic reading acquisition and by comparing them to adult readers(Maurer et al., 2005a), in order to determine whether this fastspecialization would emerge after 1.5 years of reading training inschool, and how it would develop from novice reading in 2ndgrade to expert reading in adulthood. These children had onlyminimal fast visual specialization for words when tested asnonreading kindergartners3 (Maurer et al., 2005a), indicating thatmost of the specialization indeed develops after reading trainingstarts.
In adult readers, visual specialization for print over visuallymatched strings of symbols is first reflected by increasedneurophysiological activity between 140 and 200 ms in theoccipitotemporal N1 or N170 component of the event-relatedpotential (ERP) and its magnetic field analogue (Bentin et al.,1999; Brem et al., 2005; Maurer et al., 2005a,b; Schendan et al.,1998; Tarkiainen et al., 1999). This N1 specialization reflectscoarse automatic visual categorization, as it extends to implicitreading tasks, and to consonant strings and even pseudofonts(Bentin et al., 1999; Maurer et al., 2005a; Schendan et al., 1998;Tarkiainen et al., 1999). It has also been linked to fluent readingbecause it is reduced in dyslexic adults (Helenius et al., 1999). TheN1 specialization presumably originates from inferior occipito-temporal regions predominantely of the left hemisphere (Bremet al., 2006; Nobre et al., 1994; Tarkiainen et al., 1999).
Visual N1 specialization also extends to categories of personalexpertise, with bird-experts having larger N1 amplitudes to birdthan dog stimuli and dog-experts showing the reverse pattern(Tanaka and Curran, 2001), and plays an important role for faceprocessing (Gauthier et al., 2003). It can develop rapidly withpractice for novel forms (Curran et al., 2002; Rossion et al., 2002)or symbols (Brem et al., 2005) in adults.
Developmental ERP studies in children who had alreadymastered basic reading skills demonstrate that the word N1 doesnot reflect the maturation of specific phonological or semanticprocesses (Grossi et al., 2001; Holcomb et al., 1992). The word N1
3 In Switzerland, children do not receive early literacy instruction inkindergarten, and reading instruction starts with school at age 7.

Fig. 1. Experimental task. Children and adults detected immediaterepetitions (17%) of blocked word, pseudoword, symbol (Brem et al.,2005; Maurer et al., 2005a, similar to Helenius et al., 1999; Tarkiainen et al.,1999) and picture presentations on a screen 120 cm away from the subject.We focused on the word–symbol comparison expected to reveal the largesteffects due to learning to read.
750 U. Maurer et al. / NeuroImage 33 (2006) 749–758
displays a lateral occipitotemporal topography consistent withvisual specialization by age 11 (Brandeis et al., 1994), butcontinues to increase in speed and to change in distribution untiladulthood, like subsequent ERP components reflecting lexical(Taylor, 1988), phonologic (Grossi et al., 2001), and semantic(Holcomb et al., 1992) processing.
Although the coarse N1 specialization for print that is found inadults (Bentin et al., 1999; Brem et al., 2005; Maurer et al., 2005a,b) presumably develops with learning to read and is indeed absentin kindergarten (Maurer et al., 2005a), no study so far hasinvestigated how it develops in novice readers.
The only developmental ERP study which examined the N1specialization focused on fine-tuning within orthographic patterns.This study found no clear N1 specialization for words overconsonant strings with the initial learning to read (between age 4and 7) or in 10 year olds, suggesting a slow development of N1fine-tuning (Posner and McCandliss, 2000). However, the coarserN1 tuning for words over symbols may well index a fasterdevelopment, as it is larger and more robust across differentlanguages, mainly reflecting visual expertise for print (Maureret al., 2005b).
In the present follow-up of our longitudinal study, weobtained the ERP responses of 20 children shortly after learningto read in school3 (age 8.26±0.40 years, 2nd grade), while theywere engaged in detecting immediate repetitions of words,pseudowords, symbols, and pictures (see Fig. 1). These childrenhad already been examined just before learning to read (age6.47±0.39 years, kindergarten) with this implicit reading task(Brem et al., 2005; Maurer et al., 2005a) which could also beperformed by our nonreading kindergartners and which providesa low-level visual control condition controlling for generalmaturation. To assess explicit reading skills in the 2nd grade,we used a standardized reading test (Landerl et al., 1997). Thechildren (and the adult controls) were the same as in Maurer et al.(2005a) except for 3 children who did not participate anymore inthis follow-up. As before, we mainly relied on spatial ERPanalysis using Global Field Power (GFP) to measure the electricfield strength and 3-dimensional centroids to summarize thetopography of the field. These two complementary summarymeasures do not require pre-selection of individual channels andhave already proven sensitive to implicit reading.
As we have shown previously, for most kindergartners, the N1response in the ERP task did not yet differentiate words fromsymbol strings, and even children with high letter knowledgeshowed only minimal N1 specialization with an immaturetopography (Maurer et al., 2005a). These results imply that themature print-specific N1 tuning develops with learning to read, butthey leave open when and how this would occur.
While the subtle N1 specialization for word forms (overconsonant strings) emerges over more than five years (McCandlissand Noble, 2003) after the start of reading training, i.e. only oncereading becomes increasingly proficient (Cohen and Dehaene,2004; Price and Devlin, 2004), we expected a more rapiddevelopment of the coarse N1 specialization for print over symbolstrings based on training studies with adults suggesting rapideffects of visual expertise (Brem et al., 2005; Curran et al., 2002;Rossion et al., 2002). Instead of a monotonic increase of tuningduring childhood and adolescence peaking in adulthood, wehypothesized that the largest sensitivity of the N1 specializationmay characterize the increased plasticity and emerging visualexpertise during the initial phase of reading acquisition. A special
role of early learning in developing expertise can be expected fromadult training studies and from computational modeling (Palmeriet al., 2004).
Thus, after the same children had mastered initial basic readingskills, we tested (A) whether the coarse specialization of thevisual N1 had already emerged, (B) whether it would befunctionally related to reading performance, and (C) how itwould relate to the adults’ specialization regarding strength,distribution, and specificity (generalization to novel word forms,i.e. pseudowords).
Materials and methods
Twenty-five children without familial history of dyslexia(Maurer et al., 2003b) were recorded at the end of kindergartenbefore starting their official reading training in school. All parentssigned an information and consent form explaining the study andstating that the child was free to leave the study at any time and forany reasons. In the middle of the 2nd grade (1.8 years later), 20 ofthe children (80%) again participated in a follow-up recording. Onboth occasions, visual and auditory acuity was assessed, andparents filled out questionnaires checking for deviant behavior andneurological disorders. In kindergarten intelligence (Weiss andOsterland, 1997), phonological abilities (Jansen et al., 1999), andword and letter knowledge were tested (Maurer et al., 2005a). In2nd grade, the children were tested for reading and spelling(Landerl et al., 1997), and their parents filled out questionnairesabout the childTs handedness (Oldfield, 1971).
Four children had already been excluded as early readers,because they could read more than 1 of the 9 test words inkindergarten (Maurer et al., 2005a). The 20 children (9 males and11 females) were 6.47 (±0.39) years old in kindergarten and 8.26(±0.40) years in 2nd grade. In kindergarten, the children couldname 11.7 (±6.6) letters and could read 0.1 (±0.3; maximum 1)words out of 9 tested.
The data of 13 adults (7 men, 6 women, mean±SD years=26.5±3.3) with the same task that were published earlier (Maurer et al.,2005a) were used to test how the novice readers in the 2nd gradewould differ from adult expert readers.
Participants were seated in a video-controlled, electricallyshielded, soundproof, and air-conditioned recording room 1.2 maway from the computer screen. The visual word processingexperiment was one of several short experiments, which were

751U. Maurer et al. / NeuroImage 33 (2006) 749–758
pseudorandomized in order of presentation. Electrode positionswere measured with a 3D digitizer. As compensation, each childreceived a present after the study. The entire session lasted about3.5 h in kindergarten and about 3 h in 2nd grade.
The stimuli of the word, pseudoword, symbol (Fig. 1), andpicture conditions were shown in black on a white background(Maurer et al., 2005a) in the center of the screen. The 72 stimuliper condition were shown in two blocks of 36 stimuli andcontained 17% immediate repetitions serving as targets. The blocksequence was counterbalanced (2×4 blocks). The participants wereasked to press a mouse button with their preferred hand after animmediate stimulus repetition. The stimulus duration was 700 msfollowed by a 1350 ms interstimulus interval (ISI).
Words, pseudowords, and symbol strings were matched forcharacter size (including ascenders and descenders), font size, andstring length, and ranged from 3.3 to 7.5 cm (1.6° to 3.6°). Words(concrete nouns) and pseudowords were in lower case but startedwith an uppercase letter, like all German nouns. Pictures weretaken from the Snodgrass pictures (Snodgrass and Vanderwart,1980).
The 43-channel ERPs were recorded at 500 Hz/channel withfilter settings 0.1–70 Hz and with calibrated technical zerobaselines. Caps (FMS, Munich) were used for the montage whichincluded all 10–20 system electrodes plus additional electrodes:Fpz (recording reference), Oz, FT9/10, FC5/6, TP9/10, CP5/6,PO9/10, AF1/2, FC1/2, C1/2, CP1/2, PO1/2, and two EOGelectrodes below the outer canthus of each eye. O1'/2' and Fp1'/2'were placed 2 cm more laterally for more even coverage. In the2nd grade recording, four additional occipital channels (POz, Iz,OI1/2) were used, but excluded from statistical analysis. Theywere used, however, for map illustrations. Impedance was keptbelow 20 kΩ (Ferree et al., 2001). The continuous EEG wastransformed to the average reference4 (Lehmann and Skrandies,1980) and corrected for horizontal and vertical eye movementsand in some cases for slow wave artifacts. An advanced methodwhich minimizes topographic EEG distortions was used (multiplesource eye correction method; Berg and Scherg, 1994). Correctedfiles were digitally lowpass filtered (30 Hz, 48 dB/oct), down-sampled to 256 Hz, and segmented (−125 ms prior and 1125 msfollowing the stimulus). Trials with artifacts exceeding±100 μVin any channel (3 children±125 μV, 1 child±150 μV inkindergarten; 3 children±125 μV, 2 children±150 μV in 2nd
grade) were automatically rejected before averaging. Averagingwas done separately for each age and condition including onlynontarget stimuli. Difference ERPs between conditions at eachage was computed individually, before computing grand averagesof ERPs and ERP-differences.
Adaptive segmentation according to GFP minima (Brandeis etal., 1998; Brandeis et al., 1994; Maurer et al., 2005a) was done
4 As a reviewer correctly pointed out, computing an average referencewith a different number of channels (43-channels in children inkindergarten and in the adults vs. 47-channels in the children in 2ndgrade) could introduce systematic differences. However, recomputing allanalyses with an average reference based on only 43-channels in all groupsdid not significantly change the results. The two only effects (that are notcentral to the paper) changing in significance level (one from p<0.01 top<0.001 and one from p<0.05 to p<0.1) are indicated in Tables 2 and3. The average reference for 2nd grade ERPs remains based on 47-channelsfor spatially more accurate illustrations.
for the averaged word and symbol grand means from kindergar-ten and 2nd grade (time range 0–850 ms), and separately foradults as described in Maurer et al. (2005a). The resulting ERPsegments are typically equivalent to the classical ERP compo-nents, but require no pre-selection of representative channels. Foreach segment mean, GFP and 3-dimensional (3D) location forpositive and negative centroids (center of gravity) were computedat the individual level for each age and condition separately(Brandeis et al., 1994; Lehmann, 1990; Maurer et al., 2003a,b).GFP is a time varying measure of map amplitude computed asthe spatial RMS (Lehmann and Skrandies, 1980), whereas thepositive and negative 3D centroids represent the map topography,computed separately for positive and negative field regions fromthe corresponding electrode positions (in x-, y-, and z-Talairachspace; Talairach and Tournoux, 1988) that are weighted by theirvoltage values.
For the word–symbol comparison, we focused on the P1 andN1 segment, but the analyses for the remaining segments areincluded in the supplementary material.
Two analyses per segment were computed for strength (GFP)and topography (3D centroids). GFP was analyzed in a Multi-variate Analysis of Variance (MANOVA) for repeated measureswith within subject factors “age” (kindergarten, 2nd grade) and“wordlike” (words and symbols). An analysis in the supplementarymaterial over all segments included the additional within subjectfactor “segment” (5 segments). The 3D centroids were analyzed inan analogous MANOVA, but with the additional factor “polarity”(positive vs. negative centroid) which is reported only wheninteracting with other factors. The three centroid coordinates (x-, yQ,and z-axes) were treated as multivariate dependent measures. Thesignificance level for MANOVA was set to 0.05.
In addition, t-maps of condition differences are used forillustration. Such t-maps are useful in interpreting GFP andcentroid differences and allow comparison with conventionalanalyses of selected electrodes. For comparison with conventionalwaveform analyses, we also illustrate the waveforms at the typicalfrontal and occipitotemporal channels.
Planned comparisons with t tests (t maps at N1 peak) were alsoused to investigate N1 responses from the additional pseudowordcondition. The significance level in t maps was set to p<0.01 tocorrect for multiple comparisons.
Behavioral accuracy (percentage of detected repetitions) andreaction time (to detected targets) were analyzed by computing 2MANOVAs for repeated measures with within subject factors“age” (kindergarten, 2nd grade) and “wordlike” (words vs.pseudowords vs. symbols vs. pictures). Two children missed alltargets in one condition in kindergarten, and were excluded fromreaction time analyses including this condition.
Results
Behavioral data
The children had learned to read normally after the 1.5 years ofbasic reading training, with mean reading speed at the 57th, andmean accuracy at the 52nd percentile (Landerl et al., 1997).
Effects of learning to read were also found for the behavioralresponses in the ERP task. The children had become much moreaccurate at detecting repetitions of print (words, pseudowords), whiletheir accuracy increased only moderately for the other stimuli(symbols, pictures; age×wordlike,F(3,17)=15.0, p<0.001; Table 1).
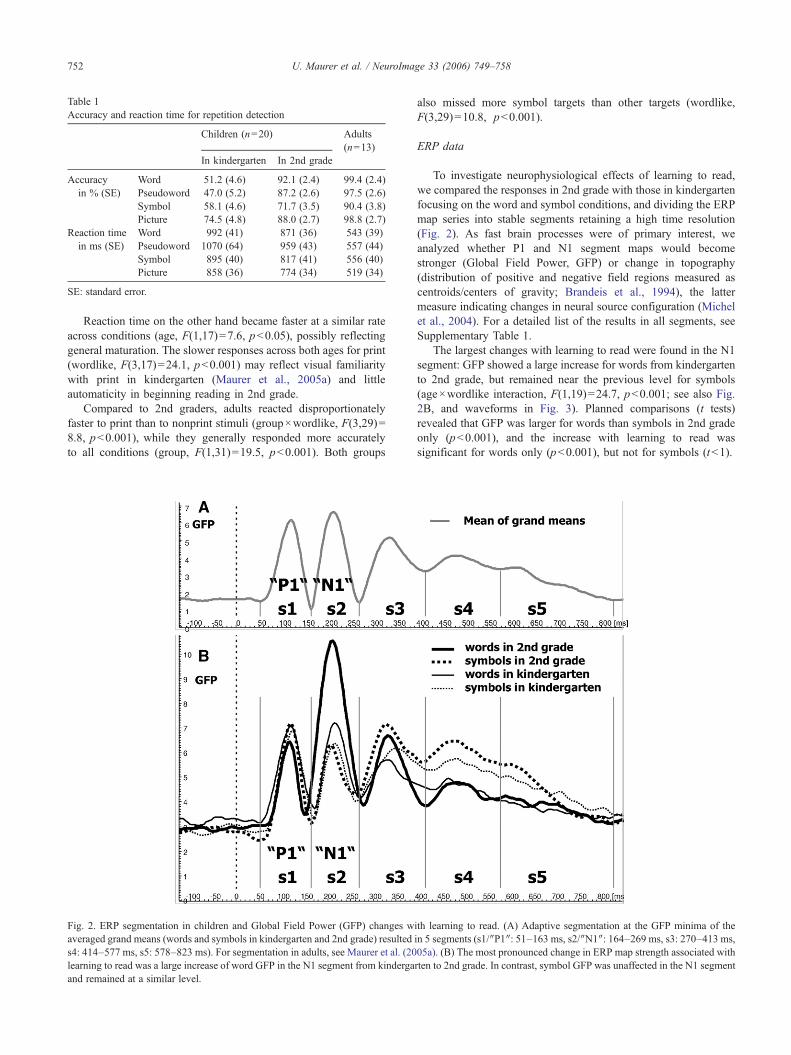
Table 1Accuracy and reaction time for repetition detection
Children (n=20) Adults(n=13)
In kindergarten In 2nd grade
Accuracyin % (SE)
Word 51.2 (4.6) 92.1 (2.4) 99.4 (2.4)Pseudoword 47.0 (5.2) 87.2 (2.6) 97.5 (2.6)Symbol 58.1 (4.6) 71.7 (3.5) 90.4 (3.8)Picture 74.5 (4.8) 88.0 (2.7) 98.8 (2.7)
Reaction timein ms (SE)
Word 992 (41) 871 (36) 543 (39)Pseudoword 1070 (64) 959 (43) 557 (44)Symbol 895 (40) 817 (41) 556 (40)Picture 858 (36) 774 (34) 519 (34)
SE: standard error.
752 U. Maurer et al. / NeuroImage 33 (2006) 749–758
Reaction time on the other hand became faster at a similar rateacross conditions (age, F(1,17)=7.6, p<0.05), possibly reflectinggeneral maturation. The slower responses across both ages for print(wordlike, F(3,17)=24.1, p<0.001) may reflect visual familiaritywith print in kindergarten (Maurer et al., 2005a) and littleautomaticity in beginning reading in 2nd grade.
Compared to 2nd graders, adults reacted disproportionatelyfaster to print than to nonprint stimuli (group×wordlike, F(3,29)=8.8, p<0.001), while they generally responded more accuratelyto all conditions (group, F(1,31)=19.5, p<0.001). Both groups
Fig. 2. ERP segmentation in children and Global Field Power (GFP) changes waveraged grand means (words and symbols in kindergarten and 2nd grade) resulteds4: 414–577 ms, s5: 578–823 ms). For segmentation in adults, see Maurer et al. (20learning to read was a large increase of word GFP in the N1 segment from kindergaand remained at a similar level.
also missed more symbol targets than other targets (wordlike,F(3,29)=10.8, p<0.001).
ERP data
To investigate neurophysiological effects of learning to read,we compared the responses in 2nd grade with those in kindergartenfocusing on the word and symbol conditions, and dividing the ERPmap series into stable segments retaining a high time resolution(Fig. 2). As fast brain processes were of primary interest, weanalyzed whether P1 and N1 segment maps would becomestronger (Global Field Power, GFP) or change in topography(distribution of positive and negative field regions measured ascentroids/centers of gravity; Brandeis et al., 1994), the lattermeasure indicating changes in neural source configuration (Michelet al., 2004). For a detailed list of the results in all segments, seeSupplementary Table 1.
The largest changes with learning to read were found in the N1segment: GFP showed a large increase for words from kindergartento 2nd grade, but remained near the previous level for symbols(age×wordlike interaction, F(1,19)=24.7, p<0.001; see also Fig.2B, and waveforms in Fig. 3). Planned comparisons (t tests)revealed that GFP was larger for words than symbols in 2nd gradeonly (p<0.001), and the increase with learning to read wassignificant for words only (p<0.001), but not for symbols (t<1).
ith learning to read. (A) Adaptive segmentation at the GFP minima of thein 5 segments (s1/′′P1′′: 51–163 ms, s2/′′N1′′: 164–269 ms, s3: 270–413 ms,05a). (B) The most pronounced change in ERP map strength associated withrten to 2nd grade. In contrast, symbol GFP was unaffected in the N1 segment
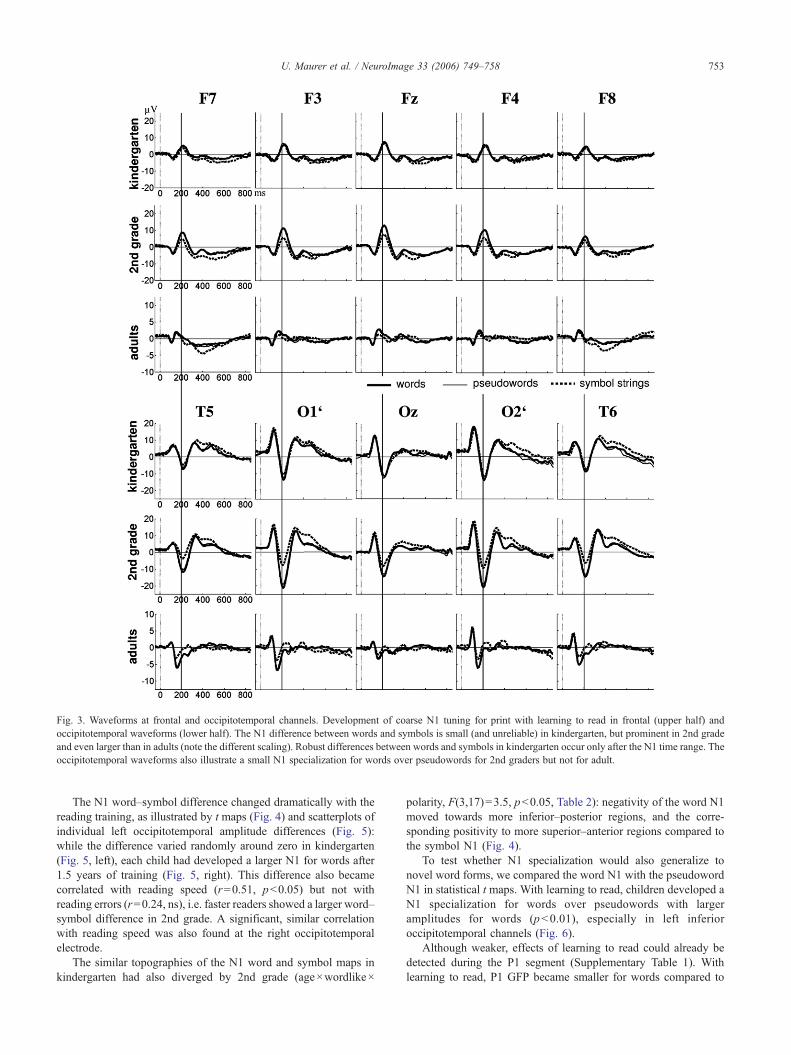
Fig. 3. Waveforms at frontal and occipitotemporal channels. Development of coarse N1 tuning for print with learning to read in frontal (upper half) andoccipitotemporal waveforms (lower half). The N1 difference between words and symbols is small (and unreliable) in kindergarten, but prominent in 2nd gradeand even larger than in adults (note the different scaling). Robust differences between words and symbols in kindergarten occur only after the N1 time range. Theoccipitotemporal waveforms also illustrate a small N1 specialization for words over pseudowords for 2nd graders but not for adult.
753U. Maurer et al. / NeuroImage 33 (2006) 749–758
The N1 word–symbol difference changed dramatically with thereading training, as illustrated by t maps (Fig. 4) and scatterplots ofindividual left occipitotemporal amplitude differences (Fig. 5):while the difference varied randomly around zero in kindergarten(Fig. 5, left), each child had developed a larger N1 for words after1.5 years of training (Fig. 5, right). This difference also becamecorrelated with reading speed (r=0.51, p<0.05) but not withreading errors (r=0.24, ns), i.e. faster readers showed a larger word–symbol difference in 2nd grade. A significant, similar correlationwith reading speed was also found at the right occipitotemporalelectrode.
The similar topographies of the N1 word and symbol maps inkindergarten had also diverged by 2nd grade (age×wordlike×
polarity, F(3,17)=3.5, p<0.05, Table 2): negativity of the word N1moved towards more inferior–posterior regions, and the corre-sponding positivity to more superior–anterior regions compared tothe symbol N1 (Fig. 4).
To test whether N1 specialization would also generalize tonovel word forms, we compared the word N1 with the pseudowordN1 in statistical t maps. With learning to read, children developed aN1 specialization for words over pseudowords with largeramplitudes for words (p<0.01), especially in left inferioroccipitotemporal channels (Fig. 6).
Although weaker, effects of learning to read could already bedetected during the P1 segment (Supplementary Table 1). Withlearning to read, P1 GFP became smaller for words compared to
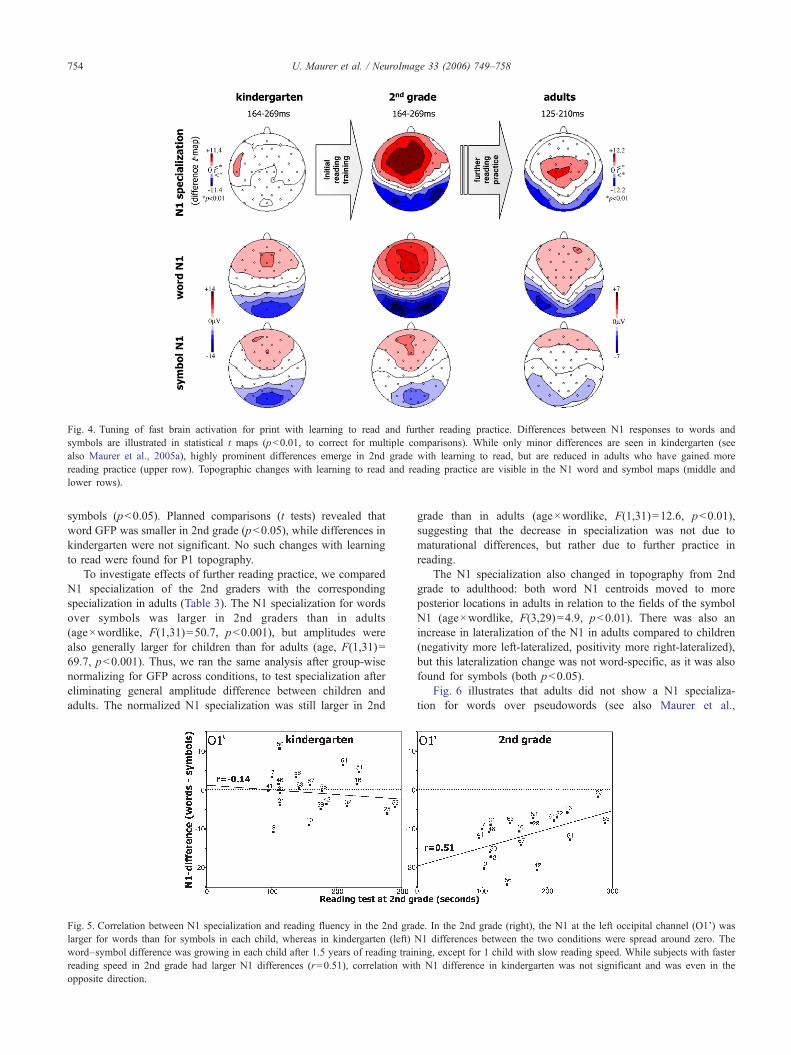
Fig. 4. Tuning of fast brain activation for print with learning to read and further reading practice. Differences between N1 responses to words andsymbols are illustrated in statistical t maps (p<0.01, to correct for multiple comparisons). While only minor differences are seen in kindergarten (seealso Maurer et al., 2005a), highly prominent differences emerge in 2nd grade with learning to read, but are reduced in adults who have gained morereading practice (upper row). Topographic changes with learning to read and reading practice are visible in the N1 word and symbol maps (middle andlower rows).
754 U. Maurer et al. / NeuroImage 33 (2006) 749–758
symbols (p<0.05). Planned comparisons (t tests) revealed thatword GFP was smaller in 2nd grade (p<0.05), while differences inkindergarten were not significant. No such changes with learningto read were found for P1 topography.
To investigate effects of further reading practice, we comparedN1 specialization of the 2nd graders with the correspondingspecialization in adults (Table 3). The N1 specialization for wordsover symbols was larger in 2nd graders than in adults(age×wordlike, F(1,31)=50.7, p<0.001), but amplitudes werealso generally larger for children than for adults (age, F(1,31)=69.7, p<0.001). Thus, we ran the same analysis after group-wisenormalizing for GFP across conditions, to test specialization aftereliminating general amplitude difference between children andadults. The normalized N1 specialization was still larger in 2nd
Fig. 5. Correlation between N1 specialization and reading fluency in the 2nd gralarger for words than for symbols in each child, whereas in kindergarten (left)word–symbol difference was growing in each child after 1.5 years of reading trainreading speed in 2nd grade had larger N1 differences (r=0.51), correlation witopposite direction.
grade than in adults (age×wordlike, F(1,31)=12.6, p<0.01),suggesting that the decrease in specialization was not due tomaturational differences, but rather due to further practice inreading.
The N1 specialization also changed in topography from 2ndgrade to adulthood: both word N1 centroids moved to moreposterior locations in adults in relation to the fields of the symbolN1 (age×wordlike, F(3,29)=4.9, p<0.01). There was also anincrease in lateralization of the N1 in adults compared to children(negativity more left-lateralized, positivity more right-lateralized),but this lateralization change was not word-specific, as it was alsofound for symbols (both p<0.05).
Fig. 6 illustrates that adults did not show a N1 specializa-tion for words over pseudowords (see also Maurer et al.,
de. In the 2nd grade (right), the N1 at the left occipital channel (O1') wasN1 differences between the two conditions were spread around zero. Theing, except for 1 child with slow reading speed. While subjects with fasterh N1 difference in kindergarten was not significant and was even in the
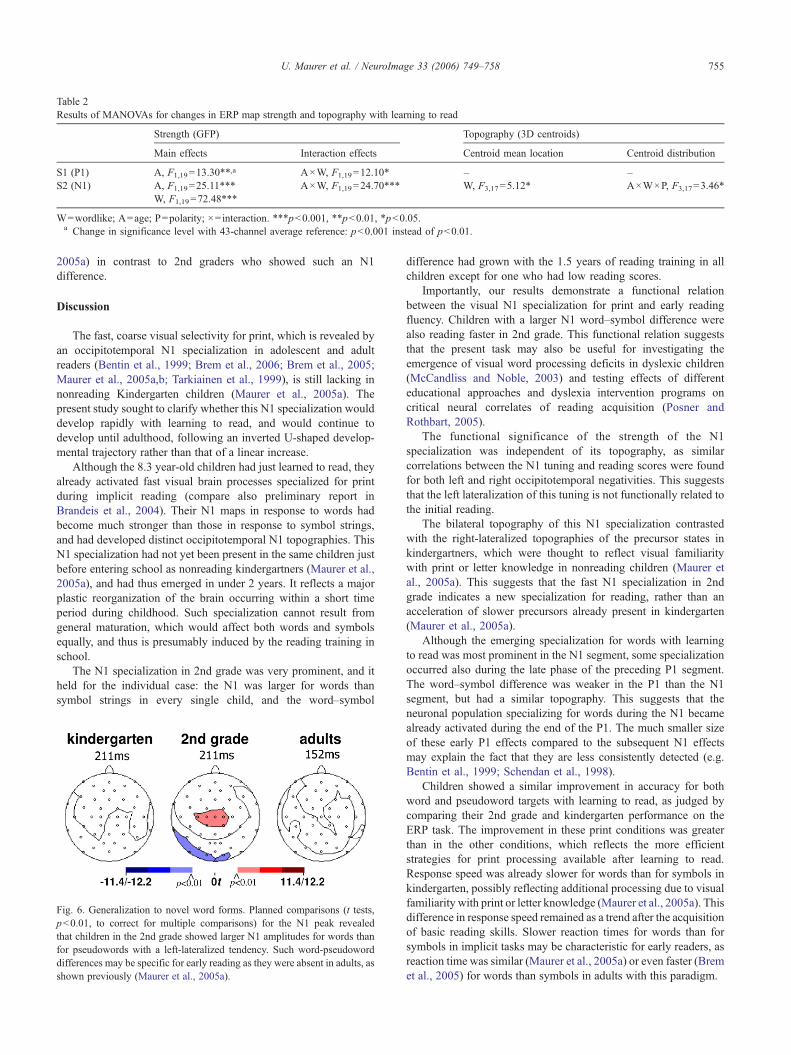
Table 2Results of MANOVAs for changes in ERP map strength and topography with learning to read
Strength (GFP) Topography (3D centroids)
Main effects Interaction effects Centroid mean location Centroid distribution
S1 (P1) A, F1,19=13.30**,a A×W, F1,19=12.10* – –S2 (N1) A, F1,19=25.11*** A×W, F1,19=24.70*** W, F3,17=5.12* A×W×P, F3,17=3.46*
W, F1,19=72.48***
W=wordlike; A=age; P=polarity; ×=interaction. ***p<0.001, **p<0.01, *p<0.05.a Change in significance level with 43-channel average reference: p<0.001 instead of p<0.01.
755U. Maurer et al. / NeuroImage 33 (2006) 749–758
2005a) in contrast to 2nd graders who showed such an N1difference.
Discussion
The fast, coarse visual selectivity for print, which is revealed byan occipitotemporal N1 specialization in adolescent and adultreaders (Bentin et al., 1999; Brem et al., 2006; Brem et al., 2005;Maurer et al., 2005a,b; Tarkiainen et al., 1999), is still lacking innonreading Kindergarten children (Maurer et al., 2005a). Thepresent study sought to clarify whether this N1 specialization woulddevelop rapidly with learning to read, and would continue todevelop until adulthood, following an inverted U-shaped develop-mental trajectory rather than that of a linear increase.
Although the 8.3 year-old children had just learned to read, theyalready activated fast visual brain processes specialized for printduring implicit reading (compare also preliminary report inBrandeis et al., 2004). Their N1 maps in response to words hadbecome much stronger than those in response to symbol strings,and had developed distinct occipitotemporal N1 topographies. ThisN1 specialization had not yet been present in the same children justbefore entering school as nonreading kindergartners (Maurer et al.,2005a), and had thus emerged in under 2 years. It reflects a majorplastic reorganization of the brain occurring within a short timeperiod during childhood. Such specialization cannot result fromgeneral maturation, which would affect both words and symbolsequally, and thus is presumably induced by the reading training inschool.
The N1 specialization in 2nd grade was very prominent, and itheld for the individual case: the N1 was larger for words thansymbol strings in every single child, and the word–symbol
Fig. 6. Generalization to novel word forms. Planned comparisons (t tests,p<0.01, to correct for multiple comparisons) for the N1 peak revealedthat children in the 2nd grade showed larger N1 amplitudes for words thanfor pseudowords with a left-lateralized tendency. Such word-pseudoworddifferences may be specific for early reading as they were absent in adults, asshown previously (Maurer et al., 2005a).
difference had grown with the 1.5 years of reading training in allchildren except for one who had low reading scores.
Importantly, our results demonstrate a functional relationbetween the visual N1 specialization for print and early readingfluency. Children with a larger N1 word–symbol difference werealso reading faster in 2nd grade. This functional relation suggeststhat the present task may also be useful for investigating theemergence of visual word processing deficits in dyslexic children(McCandliss and Noble, 2003) and testing effects of differenteducational approaches and dyslexia intervention programs oncritical neural correlates of reading acquisition (Posner andRothbart, 2005).
The functional significance of the strength of the N1specialization was independent of its topography, as similarcorrelations between the N1 tuning and reading scores were foundfor both left and right occipitotemporal negativities. This suggeststhat the left lateralization of this tuning is not functionally related tothe initial reading.
The bilateral topography of this N1 specialization contrastedwith the right-lateralized topographies of the precursor states inkindergartners, which were thought to reflect visual familiaritywith print or letter knowledge in nonreading children (Maurer etal., 2005a). This suggests that the fast N1 specialization in 2ndgrade indicates a new specialization for reading, rather than anacceleration of slower precursors already present in kindergarten(Maurer et al., 2005a).
Although the emerging specialization for words with learningto read was most prominent in the N1 segment, some specializationoccurred also during the late phase of the preceding P1 segment.The word–symbol difference was weaker in the P1 than the N1segment, but had a similar topography. This suggests that theneuronal population specializing for words during the N1 becamealready activated during the end of the P1. The much smaller sizeof these early P1 effects compared to the subsequent N1 effectsmay explain the fact that they are less consistently detected (e.g.Bentin et al., 1999; Schendan et al., 1998).
Children showed a similar improvement in accuracy for bothword and pseudoword targets with learning to read, as judged bycomparing their 2nd grade and kindergarten performance on theERP task. The improvement in these print conditions was greaterthan in the other conditions, which reflects the more efficientstrategies for print processing available after learning to read.Response speed was already slower for words than for symbols inkindergarten, possibly reflecting additional processing due to visualfamiliarity with print or letter knowledge (Maurer et al., 2005a). Thisdifference in response speed remained as a trend after the acquisitionof basic reading skills. Slower reaction times for words than forsymbols in implicit tasks may be characteristic for early readers, asreaction time was similar (Maurer et al., 2005a) or even faster (Bremet al., 2005) for words than symbols in adults with this paradigm.

Table 3Results of MANOVAs for changes in N1 map strength and topography with further reading practice
Strength (GFP) Topography (3D centroids)
Not normalized Normalized for age effects Centroid mean location Centroid distribution
S2 (N1) A, F1,31=69.74*** A, F3,29=5.28** A×P, F3,29=2.94*,a
W, F1,31=97.97*** W, F1,31=82.34 *** W, F3,29=11.53*** W×P, F3,29=18.00***A×W, F1,31=50.74*** A×W, F1,31=12.62** A×W, F3,29=4.94**
W=wordlike; A=age; P=polarity; ×=interaction. ***p<0.001, **p<0.01, *p<0.05.a Change in significance level with 43-channel average reference: p<0.1 instead of p<0.05.
756 U. Maurer et al. / NeuroImage 33 (2006) 749–758
Our results indicate that learning to read leads to a remarkableneurophysiological specialization for print after only 1.5 years ofinitial reading training. This specialization, however, differed fromthe adults’ specialization in important ways, namely its neurophy-siological characteristics and its sensitivity to novel word forms.
In addition to decreasing latencies in adults suggesting fasterword processing with increasing age, further development ofreading skill was reflected by changes in amplitudes andtopography of the N1 specialization. While reading skills areknown to increase during the subsequent years of reading practice,the basic N1 specialization instead had decreased in adults in termsof both amplitude and significance, and thus represents a stronglynonlinear development. This decrease of the N1 specialization isnot due to the general attenuation of ERP amplitudes duringdevelopment, as children still had a larger N1 specialization thanadults when standardized for equal group mean GFP or measuredwith t statistics. While nonspecific maturation can affect both meanN1 amplitude and N1 specialization if measured as an absoluteamplitude difference, it cannot account for a change in the relativeN1 amplitude of words vs. symbols. Thus, after the initial increasewith learning to read, further reading practice during childhood oradolescence is associated with a reduction of the N1 specialization.This reduction was accompanied by topographic changes reflectingchanges in the distribution of the neural networks involved. Thismay reflect changes in the network structure due to activation ofadditional or fewer regions, but it may also be due to changes in therelative activation among different parts of the same network. Thespatial resolution of ERPs is not well suited to differentiate thesealternatives, and this question might be answered in future studiesusing techniques with higher spatial resolution.
In addition to these neurophysiological differences, the new N1specialization in children differed also functionally from theadults’ specialization. The small “lexical” N1 increase for wordsover pseudowords in 2nd grade contrasts with the lack of sucheffects in adults (Bentin et al., 1999; Maurer et al., 2005a). Thiseffect may relate to two alternative explanations: fine-tuning fororthographic patterns and automaticity in grapheme–phonemeconversion. In contrast to earlier studies that showed a larger N1for the less familiar consonant strings than for words (Compton etal., 1991; McCandliss et al., 1997), the present study found theopposite effect for the less familiar peudowords. The fine-tuningexplanation may thus not apply to the present results. On the otherhand, a similarly left-lateralized increase for words over pseudo-words was found in adults with English but not with Germanusing the same paradigm (Maurer et al., 2005b), a result that hasbeen explained with the “grapheme–phoneme automaticity”account: In implicit reading tasks, grapheme-to-phoneme conver-sion strategies might be engaged to a lesser degree – especially inthe fast N1 activity – if such processing is less automatized, asmay be the case in languages with inconsistent grapheme-to-
phoneme mapping (Maurer et al., 2005b). The same mechanismmay also occur in languages with more consistent mapping duringearly reading.
At any rate, the size of this “lexical” effect in children wasmuch smaller than the N1 specialization for words over symbols.This suggests that already the N1 of beginning readers mainlyreflects coarse-tuning for print, while other effects like fine-tuningfor orthographic patterns and automaticity of higher-orderlinguistic processes only modulate this basic N1 effect.
The development of N1 specialization for print in the presentstudy contradicts assumptions about how specialized visual wordprocessing would develop (Cohen and Dehaene, 2004; McCandlissand Noble, 2003). Instead of a linear increase with more proficientreading, the development is strongly nonlinear: the N1 specializa-tion peaks after learning to read in beginning readers and thendecreases with further reading practice in adults following aninverted U-shaped developmental time-course.
Of course, the present results based on three age groups cannotdelineate the exact time-course of this development and the N1specialization in 2nd grade may already occur after the actual peakor, conversely, still precede the maximal specialization. However,support for an early peak of the N1 specialization comes fromanother developmental study showing that the size of the N1specialization in adolescents and adults no longer differs (Brem etal., 2006). This suggests that the size of N1 the specialization doespeak before adolescence.
A dissociation between early “novice” learning and late“expert” learning has been postulated for the development of per-ceptual expertise in the brain based on adults’ data and com-putational modeling (Palmeri et al., 2004). Several possiblemechanisms have been suggested that could account for suchchanges, such as changes from broadly to finely tuned receptivefields, or qualitative changes of representations (Palmeri et al.,2004). While the precise mechanisms need to be studied at thecellular level, the present study provides neurophysiological datathat strongly support a dissociation between novel and expertlearning which might also be understood from a systems levelperspective: we propose that under the influence of readingtraining, an extensive neural network becomes sensitive to print innovice readers, reflected by the strong increase of N1 specializationfrom kindergarten to 2nd grade. With further reading practice,sensitivity for some visual aspects of print may decrease in parts ofthis network, resulting in more selective activation in adults.
Increasing fine-tuning of cognitive functions has beenassociated with maturation during development (Brem et al.,2006; Casey et al., 2005). The present results suggest that duringdevelopment similar processes as found for maturation do alsoapply for skill learning.
In summary, visual expertise for print develops rapidly duringthe initial phase of learning to read, and does so by tuning fast

757U. Maurer et al. / NeuroImage 33 (2006) 749–758
visual processes. The N1 responses to words and symbol strings,which had not differed in most kindergartners, diverged clearly foreach individual case after the same children had learned to read. Itbecame stronger in faster readers, demonstrating its functional rolein the early fluency of reading. Comparison with adults' dataindicates that the novel specialization for print in beginningreaders undergoes further changes as readers become moreproficient with further practice. The degree of the N1 tuning andits sensitivity for novel word forms were maximal in beginningreaders. This nonlinear relation between the development of visualspecialization for print and reading skills may reflect an initialrecruitment of more general brain processes related to learning andplasticity which precede the subsequent full specializationinvolving more selective brain processes.
Acknowledgments
This research was supported by Swiss National ScienceFoundation grant 32-59276, the “Stiftung für wissenschaftlicheForschung, Universität Zürich”, and the NCCR on NeuralPlasticity and Repair. We thank Bruce D. McCandliss and JasonD. Zevin for their helpful comments on the manuscript.
Appendix A. Supplementary data
Supplementary data associated with this article can be found, inthe online version, at doi:10.1016/j.neuroimage.2006.06.025.
References
Bentin, S., Mouchetant-Rostaing, Y., Giard, M.H., Echallier, J.F., Pernier, J.,1999. ERP manifestations of processing printed words at differentpsycholinguistic levels: time course and scalp distribution. J. Cogn.Neurosci. 11 (3), 235–260.
Berg, P., Scherg, M., 1994. A multiple source approach to the correction ofeye artifacts. Electroencephalogr. Clin. Neurophysiol. 90 (3), 229–241.
Brandeis, D., Vitacco, D., Steinhausen, H.C., 1994. Mapping brain electricmicro-states in dyslexic children during reading. Acta Paedopsychiatr.56 (3), 239–247.
Brandeis, D., van Leeuwen, T.H., Rubia, K., Vitacco, D., Steger, J., Pascual-Marqui, R.D., Steinhausen, H.C., 1998. Neuroelectric mapping revealsprecursor of stop failures in children with attention deficits. Behav. BrainRes. 94 (1), 111–125.
Brandeis, D., Brem, S., Bucher, K.R.B, Kranz, F., Maurer, U., 2004. Fastvisual brain specialization for print with learning to read. NeuroImage 22(Suppl. 1), S29.
Brem, S., Lang-Dullenkopf, A., Maurer, U., Halder, P., Bucher, K.,Brandeis, D., 2005. Neurophysiological signs of rapidly emergingvisual expertise for symbol strings. NeuroReport 16 (1), 45–48.
Brem, S., Bucher, K., Halder, P., Summers, P., Dietrich, T.,Martin, E., Brandeis,D., 2006. Evidence for developmental changes in the visual wordprocessing network beyond adolescence. NeuroImage 29 (3), 822–837.
Casey, B.J., Tottenham, N., Liston, C., Durston, S., 2005. Imaging thedeveloping brain: what have we learned about cognitive development?Trends Cogn. Sci. 9 (3), 104–110.
Cohen, L., Dehaene, S., 2004. Specialization within the ventral stream: thecase for the visual word form area. NeuroImage 22 (1), 466–476.
Compton, P.E., Grossenbacher, P., Posner, M.I., Tucker, D.M., 1991. Acognitive-anatomical approach to attention in lexical access. J. Cogn.Neurosci. 3 (4), 304–312.
Curran, T., Tanaka, J.W., Weiskopf, D.M., 2002. An electrophysiologicalcomparison of visual categorization and recognition memory. Cogn.Affect. Behav. Neurosci. 2 (1), 1–18.
Ferree, T.C., Luu, P., Russell, G.S., Tucker, D.M., 2001. Scalp electrodeimpedance, infection risk, and EEG data quality. Clin. Neurophysiol.112 (3), 536–544.
Gauthier, I., Curran, T., Curby, K.M., Collins, D., 2003. Perceptualinterference supports a non-modular account of face processing. Nat.Neurosci. 6 (4), 428–432.
Grossi, G., Coch, D., Coffey-Corina, S., Holcomb, P.J., Neville, H.J., 2001.Phonological processing in visual rhyming: a developmental ERP study.J. Cogn. Neurosci. 13 (5), 610–625.
Helenius, P., Tarkiainen, A., Cornelissen, P., Hansen, P.C., Salmelin, R.,1999. Dissociation of normal feature analysis and deficient processing ofletter-strings in dyslexic adults. Cereb. Cortex 9 (5), 476–483.
Holcomb, P.J., Coffey, S.A., Neville, H.J., 1992. Visual and auditorysentence processing: a developmental analysis using event-related brainpotentials. Dev. Neuropsychol. 8 (2–3), 203–241.
Jansen, H., Mannhaupt, G., Marx, H., Skowronek, H., 1999. BISCBielefelder Screening zur Frueherkennung von Lese-Rechtschreibsch-wierigkeiten (Bielefeld Screening Battery for Early Diagnosis ofProblems in Reading and Writing). Hogrefe, Göttingen.
Landerl, K., Wimmer, H., Moser, E., 1997. SLRT Salzburger Lese-undRechtschreibtest (Salzburg Reading and Orthography Test). Huber,Bern.
Lehmann, D., 1990. Past, present and future of topographic mapping. BrainTopogr. 3 (1), 191–202.
Lehmann, D., Skrandies, W., 1980. Reference-free identification ofcomponents of checkerboard-evoked multichannel potential fields.Electroencephalogr. Clin. Neurophysiol. 48 (6), 609–621.
Maurer, U., Bucher, K., Brem, S., Brandeis, D., 2003a. Development of theautomatic mismatch response: from frontal positivity in kindergartenchildren to the mismatch negativity. Clin. Neurophysiol. 114 (5), 808–817.
Maurer, U., Bucher, K., Brem, S., Brandeis, D., 2003b. Altered responses totone and phoneme mismatch in kindergartners at familial dyslexia risk.NeuroReport 14 (17), 2245–2250.
Maurer, U., Brem, S., Bucher, K., Brandeis, D., 2005a. Emergingneurophysiological specialization for letter strings. J. Cogn. Neurosci.17 (10), 1532–1552.
Maurer, U., Brandeis, D., McCandliss, B., 2005b. Fast, visual specializationfor reading in English revealed by the topography of the N170 ERPresponse. Behav. Brain Func. 1 (1), 13.
McCandliss, B.D., Noble, K.G., 2003. The development of readingimpairment: a cognitive neuroscience model. Ment. Retard. Dev.Disabil. Res. Rev. 9 (3), 196–204.
McCandliss, B.D., Posner, M.I., Givon, T., 1997. Brain plasticity in learningvisual words. Cogn. Psychol. 33 (1), 88–110.
McCandliss, B.D., Cohen, L., Dehaene, S., 2003. The visual word formarea: expertise for reading in the fusiform gyrus. Trends Cogn. Sci. 7 (7),293–299.
Michel, C.M., Murray, M.M., Lantz, G., Gonzalez, S., Spinelli, L., Grave dePeralta, R., 2004. EEG source imaging. Clin. Neurophysiol. 115 (10),2195–2222.
Nobre, A.C., Allison, T., McCarthy, G., 1994. Word recognition in thehuman inferior temporal lobe. Nature 372 (6503), 260–263.
Oldfield, R.C., 1971. The assessment and analysis of handedness: theEdinburgh inventory. Neuropsychologia 9 (1), 97–113.
Palmeri, T.J., Wong, A.C., Gauthier, I., 2004. Computational approaches tothe development of perceptual expertise. Trends Cogn. Sci. 8 (8),378–386.
Posner, M., McCandliss, B.D., 2000. Brain circuitry during reading. In:Klein, R., McMullen, P. (Eds.), Converging Methods for UnderstandingReading and Dyslexia. MIT Press, Cambridge, pp. 305–337.
Posner, M.I., Rothbart, M.K., 2005. Influencing brain networks: implica-tions for education. Trends Cogn. Sci. 9 (3), 99–103.
Price, C.J., Devlin, J.T., 2004. The pro and cons of labelling a leftoccipitotemporal region: “the visual word form area”. NeuroImage 22(1), 477–479.
Rossion, B., Gauthier, I., Goffaux, V., Tarr, M.J., Crommelinck, M.,2002. Expertise training with novel objects leads to left-lateralized

758 U. Maurer et al. / NeuroImage 33 (2006) 749–758
facelike electrophysiological responses. Psychol. Sci. 13 (3),250–257.
Schendan, H.E., Ganis, G., Kutas, M., 1998. Neurophysiological evidencefor visual perceptual categorization of words and faces within 150 ms.Psychophysiology 35 (3), 240–251.
Snodgrass, J.G., Vanderwart, M., 1980. A standardized set of 260 pictures:norms for name agreement, image agreement, familiarity, and visualcomplexity. J. Exp. Psychol. Hum. Learn. 6 (2), 174–215.
Talairach, J., Tournoux, P., 1988. Co-planar Stereotaxic Atlas of the HumanBrain. Thieme, Stuttgart.
Tanaka, J.W., Curran, T., 2001. A neural basis for expert object recognition.Psychol. Sci. 12 (1), 43–47.
Tarkiainen, A., Helenius, P., Hansen, P.C., Cornelissen, P.L., Salmelin, R.,1999. Dynamics of letter string perception in the human occipitotem-poral cortex. Brain 122 (Pt 11), 2119–2132.
Taylor, M.J., 1988. Developmental changes in ERPs to visual languagestimuli. Biol. Psychol. 26 (1–3), 321–338.
Weiss, R., Osterland, J., 1997. CFT 1, Grundintelligenztest Skala 1—Culture Fair Intelligence Test, Scale 1 (Cattell, R.B., 1966), GermanVersion. Hogrefe, Göttingen.




















