
Chromosome-induced microtubule assembly mediated by TPX2 is required for spindle formation in HeLa...
9
articles NATURE CELL BIOLOGY ADVANCE ONLINE PUBLICATION www.nature.com/naturecellbiology 41 Chromosome-induced microtubule assembly mediated by TPX2 is required for spindle formation in HeLa cells Oliver J. Gruss*†, Malte Wittmann*†‡, Hideki Yokoyama*, Rainer Pepperkok*, Thomas Kufer§, Herman Silljé§, Eric Karsenti*, Iain W. Mattaj* and Isabelle Vernos*¶ *EMBL, Meyerhofstrasse 1, 69 117 Heidelberg, Germany §Max-Planck-Institute for Biochemistry, Department of Cell Biology, Am Klopferspitz 18a, 82152 Martinsried, Germany ‡Current address: Zentrum für Molekulare Biologie, Im Neuenheimer Feld 282, 69120 Heidelberg, Germany †These authors contributed equally to this work ¶e-mail: [email protected] Published online: 21 October 2002; DOI: 10.1038/ncb870 In Xenopus laevis egg extracts, TPX2 is required for the Ran-GTP-dependent assembly of microtubules around chro- mosomes. Here we show that interfering with the function of the human homologue of TPX2 in HeLa cells causes defects in microtubule organization during mitosis. Suppressing the expression of human TPX2 by RNA interference leads to the formation of two microtubule asters that do not interact and do not form a spindle. Our results suggest that in vivo, even in the presence of duplicated centrosomes, spindle formation requires the function of TPX2 to generate a stable bipolar spindle with overlapping antiparallel microtubule arrays. This indicates that chromosome- induced microtubule production is a general requirement for the formation of functional spindles in animal cells. O nce in every cell cycle microtubules become organized into a bipolar spindle that segregates the replicated chromosomes to the two daughter cells 1,2 . Two models, which are not mutu- ally exclusive, have been proposed to explain how spindles assem- ble in animal cells. In the first, dynamic microtubules emanating from the separated centrosomes are selectively stabilized at the kinetochores 3 . In the second, chromosomes are the source of microtubule assembly and organization 4 . A centrosome-independ- ent pathway exists in some organisms and types of cell, including in female germ-cell meiosis in some metazoa 4 . This assembly pathway can be recapitulated in Xenopus egg extracts. In these systems, microtubules grow around chromosomes and become organized into a bipolar spindle through a self-organization process driven by microtubule motors 4–6 . Insight into the mechanism by which chromosomes can pro- mote microtubule assembly in mitosis has been obtained from dif- ferent studies investigating the role of the small GTPase Ran in spin- dle formation 7,8 . Ran in its GTP-bound form (Ran-GTP) induces microtubule assembly in M-phase extracts from Xenopus eggs 9–13 . The relative proportion of the active (Ran-GTP) and inactive (Ran- GDP) forms of Ran depends on the activities of the guanine nucleotide-exchange factor RCC1 and the GTPase-activating pro- tein RanGAP1 together with its cofactors RanBP1 and RanBP2 (ref. 7). Because a fraction of RCC1 is bound to chromatin and RanGAP1 and RanBP1 are soluble, Ran-GTP is enriched locally around chromosomes and is converted rapidly to Ran-GDP away from them 14 . As a result, chromosomes create a local environment that is favourable for microtubule nucleation and stabilization 4,9,10,15 . Two microtubule-associated proteins (MAPs), NuMA and TPX2, have been identified as potential downstream effectors of Ran-GTP in microtubule assembly in these extracts 16–18 . Depletion of TPX2 from egg extract prevents microtubule nucleation and spindle formation around chromatin beads 16 . Conversely, TPX2 gives rise to Ran-GTP-independent microtubule assembly when it is added in excess over its Ran-GTP-regulated inhibitors importin- α and importin-β (ref. 16). These results indicate that in egg extracts TPX2 may be required for microtubule assembly around chromosomes. In addition, other experiments have shown that the production of Ran-GTP is required for spindle assembly in these extracts even when centrosomes are present 15 . Taken together, these data indicate that in Xenopus egg extracts chromosome-induced microtubule assembly through Ran-GTP and TPX2 is essential for spindle assembly whether centrosomes are present or not. It is important to determine whether this is a general principle for spindle assembly or a particular pathway that is restricted to specific types of cell, such as female meiotic cells. Some experimen- tal data are consistent with generality, as they suggest that a centro- some-independent pathway for spindle assembly also exists in somatic cells. Indeed, eliminating one or both centrosomes from metazoan cells either through laser ablation or by genetic methods does not prevent spindle assembly 19–24 . To examine further the general relevance of chromatin-mediat- ed microtubule assembly, we have characterized the function of human TPX2 (hTPX2) in HeLa cells. We show that hTPX2 is a true functional homologue of TPX2. HeLa cells that do not express hTPX2 are blocked in mitosis with two microtubule asters that can- not connect and cannot form a spindle. Our data indicate that spindle formation in somatic cells requires a population of micro- tubules whose assembly is dependent on chromosomes and medi- ated by Ran-GTP and TPX2. Results GFP–hTPX2 rescues TPX2 depletion from Xenopus egg extracts. Human TPX2 has been previously purified from HeLa cell nuclear fractions and found to mediate Ran-GTP-dependent microtubule assembly in Xenopus egg extracts 16 . To examine the functional rela- tionship between TPX2 and hTPX2, we tested the ability of a fusion of green fluorescent protein and hTPX2 (GFP–hTPX2) to rescue spindle assembly on the addition of sperm chromatin to a Xenopus egg extracts depleted of TPX2. Depletion of TPX2 greatly reduced the ability of Xenopus egg extracts to mediate spindle formation © 2002 Nature Publishing Group
-
Upload
independent -
Category
Documents
-
view
3 -
download
0
Transcript of Chromosome-induced microtubule assembly mediated by TPX2 is required for spindle formation in HeLa...

articles
NATURE CELL BIOLOGY ADVANCE ONLINE PUBLICATION www.nature.com/naturecellbiology 41
Chromosome-induced microtubuleassembly mediated by TPX2 is requiredfor spindle formation in HeLa cells
Oliver J. Gruss*†, Malte Wittmann*†‡, Hideki Yokoyama*, Rainer Pepperkok*, Thomas Kufer§, Herman Silljé§, Eric Karsenti*, Iain W. Mattaj* and Isabelle Vernos*¶
*EMBL, Meyerhofstrasse 1, 69 117 Heidelberg, Germany
§Max-Planck-Institute for Biochemistry, Department of Cell Biology, Am Klopferspitz 18a, 82152 Martinsried, Germany
‡Current address: Zentrum für Molekulare Biologie, Im Neuenheimer Feld 282, 69120 Heidelberg, Germany
†These authors contributed equally to this work
¶e-mail: [email protected]
Published online: 21 October 2002; DOI: 10.1038/ncb870
In Xenopus laevis egg extracts, TPX2 is required for the Ran-GTP-dependent assembly of microtubules around chro-mosomes. Here we show that interfering with the function of the human homologue of TPX2 in HeLa cells causesdefects in microtubule organization during mitosis. Suppressing the expression of human TPX2 by RNA interferenceleads to the formation of two microtubule asters that do not interact and do not form a spindle. Our results suggestthat in vivo, even in the presence of duplicated centrosomes, spindle formation requires the function of TPX2 togenerate a stable bipolar spindle with overlapping antiparallel microtubule arrays. This indicates that chromosome-induced microtubule production is a general requirement for the formation of functional spindles in animal cells.
Once in every cell cycle microtubules become organized into abipolar spindle that segregates the replicated chromosomesto the two daughter cells1,2. Two models, which are not mutu-
ally exclusive, have been proposed to explain how spindles assem-ble in animal cells. In the first, dynamic microtubules emanatingfrom the separated centrosomes are selectively stabilized at thekinetochores3. In the second, chromosomes are the source ofmicrotubule assembly and organization4. A centrosome-independ-ent pathway exists in some organisms and types of cell, including infemale germ-cell meiosis in some metazoa4. This assembly pathwaycan be recapitulated in Xenopus egg extracts. In these systems,microtubules grow around chromosomes and become organizedinto a bipolar spindle through a self-organization process driven bymicrotubule motors4–6.
Insight into the mechanism by which chromosomes can pro-mote microtubule assembly in mitosis has been obtained from dif-ferent studies investigating the role of the small GTPase Ran in spin-dle formation7,8. Ran in its GTP-bound form (Ran-GTP) inducesmicrotubule assembly in M-phase extracts from Xenopus eggs9–13.The relative proportion of the active (Ran-GTP) and inactive (Ran-GDP) forms of Ran depends on the activities of the guaninenucleotide-exchange factor RCC1 and the GTPase-activating pro-tein RanGAP1 together with its cofactors RanBP1 and RanBP2(ref. 7). Because a fraction of RCC1 is bound to chromatin andRanGAP1 and RanBP1 are soluble, Ran-GTP is enriched locallyaround chromosomes and is converted rapidly to Ran-GDP awayfrom them14. As a result, chromosomes create a local environmentthat is favourable for microtubule nucleation and stabilization4,9,10,15.
Two microtubule-associated proteins (MAPs), NuMA andTPX2, have been identified as potential downstream effectors ofRan-GTP in microtubule assembly in these extracts16–18. Depletionof TPX2 from egg extract prevents microtubule nucleation andspindle formation around chromatin beads16. Conversely, TPX2gives rise to Ran-GTP-independent microtubule assembly when itis added in excess over its Ran-GTP-regulated inhibitors importin-α and importin-β (ref. 16). These results indicate that in egg
extracts TPX2 may be required for microtubule assembly aroundchromosomes. In addition, other experiments have shown that theproduction of Ran-GTP is required for spindle assembly in theseextracts even when centrosomes are present15. Taken together, thesedata indicate that in Xenopus egg extracts chromosome-inducedmicrotubule assembly through Ran-GTP and TPX2 is essential forspindle assembly whether centrosomes are present or not.
It is important to determine whether this is a general principlefor spindle assembly or a particular pathway that is restricted tospecific types of cell, such as female meiotic cells. Some experimen-tal data are consistent with generality, as they suggest that a centro-some-independent pathway for spindle assembly also exists insomatic cells. Indeed, eliminating one or both centrosomes frommetazoan cells either through laser ablation or by genetic methodsdoes not prevent spindle assembly19–24.
To examine further the general relevance of chromatin-mediat-ed microtubule assembly, we have characterized the function ofhuman TPX2 (hTPX2) in HeLa cells. We show that hTPX2 is a truefunctional homologue of TPX2. HeLa cells that do not expresshTPX2 are blocked in mitosis with two microtubule asters that can-not connect and cannot form a spindle. Our data indicate thatspindle formation in somatic cells requires a population of micro-tubules whose assembly is dependent on chromosomes and medi-ated by Ran-GTP and TPX2.
ResultsGFP–hTPX2 rescues TPX2 depletion from Xenopus egg extracts.Human TPX2 has been previously purified from HeLa cell nuclearfractions and found to mediate Ran-GTP-dependent microtubuleassembly in Xenopus egg extracts16. To examine the functional rela-tionship between TPX2 and hTPX2, we tested the ability of a fusionof green fluorescent protein and hTPX2 (GFP–hTPX2) to rescuespindle assembly on the addition of sperm chromatin to a Xenopusegg extracts depleted of TPX2. Depletion of TPX2 greatly reducedthe ability of Xenopus egg extracts to mediate spindle formation
© 2002 Nature Publishing Group

articles
NATURE CELL BIOLOGY ADVANCE ONLINE PUBLICATION www.nature.com/naturecellbiology42
(Fig. 1), as reported previously16,25.Adding recombinant GFP–hTPX2 to the depleted extracts at a
final concentration of 60–100 nM restored spindle assembly con-siderably (Fig. 1, bottom). In addition, GFP–hTPX2 accumulated
at the spindle poles (Fig. 1, bottom) similar to the location ofendogenous Xenopus TPX2 (ref. 25). These results indicate that thetwo proteins can be considered as true functional homologues.Subcellular localization of hTPX2 in HeLa cells. To characterizethe function of hTPX2, we first determined its subcellular localiza-tion in mitotic HeLa cells by immunofluorescence microscopy withan affinity-purified polyclonal antibody against hTPX2 (Fig. 2a).During prophase, hTPX2 concentrated at the centre of the micro-tubule asters and faintly decorated microtubules growing betweenthe two asters. As the cells progressed into prometaphase hTPX2increased in intensity at the spindle poles, and in metaphase it alsoextended along some spindle microtubules. At the onset of
∆TPX2
Mock
∆TPX2 + GFP–hTPX2
Mock ∆TPX2 ∆TPX2 + GFP–hTPX2
Per
cent
age
of in
tact
spi
ndle
s
0
10
20
30
40
50
60
70
Figure 1 Recombinant human TPX2 rescues spindle assembly in eggextracts. TPX2 was depleted from Xenopus egg extracts using specific antibodies,and spindle assembly was monitored on the addition of de-membranated spermnuclei and rhodamine-tubulin (red). In extracts treated with control antibodies (mock),spindles assembled normally around single chromosome clusters with an efficiencyof 43.1 ± 4.6% (n = 88). In TPX2-depleted extracts (∆TPX2), this efficiency wasreduced to 12.8 ± 8.6% (n = 85) and many abnormal structures were observed.Recombinant hTPX2 fused to EGFP (green, ∆TPX2 + GFP–hTPX2) restored the abilityof the TPX2-depleted extracts to form bipolar spindles with an efficiency of42.8 ±18.9% (n = 61), and GFP–hTPX2 localized to the spindle poles. DNA is shown in blue. Scale bar, 10 µm. Error bars indicate the s.d. from three samples.
Pro
phas
eM
etap
hase
Ana
phas
eTe
loph
ase
a
b
Tubulin hTPX2 DNA Overlay
Figure 2 Localization of hTPX2 in HeLa cells. a, Exponentially growing HeLacells were fixed with glutaraldehyde and processed for immunofluorescence with anaffinity-purified antibody against hTPX2 and a monoclonal antibody against α-tubulin.DNA was visualized with Hoechst. Confocal slices were 1.2 µm thick. In the overlay,microtubules are green, hTPX2 is red and DNA is blue. b, HeLa cells expressingGFP–hTPX2 were fixed with glutaraldehyde and processed for immunofluorescencewith an antibody against α-tubulin. DNA was visualized with Hoechst. Overlays ofcells in interphase, metaphase, anaphase and telophase are shown. Microtubulesare green, GFP–hTPX2 is red and DNA is blue. In the left image, only one of the cellsis transfected and GFP–hTPX2 accumulates in the nucleus. Scale bars, 10 µm.
© 2002 Nature Publishing Group

articles
NATURE CELL BIOLOGY ADVANCE ONLINE PUBLICATION www.nature.com/naturecellbiology 43
anaphase hTPX2 remained concentrated at the spindle poles, and itstarted to relocalize to the spindle midzone in late anaphase. Intelophase, hTPX2 was no longer found near centrosomes but hadrelocalized completely to the spindle midzone. Notably, hTPX2 wasnot detected on astral microtubules in any stage of mitosis.
To confirm this localization pattern, we expressed GFP–hTPX2in HeLa cells. The recombinant protein was transported efficientlyinto the nucleus in interphase and showed a very similar localizationpattern to the endogenous protein in mitosis (Fig. 2b). Time-lapsevideo microscopy on transfected cells indicated that the relocaliza-tion of GFP–hTPX2 from the spindle poles to the midbody in lateanaphase was very rapid (data not shown). The intracellular local-ization of hTPX2 is thus very similar to that of TPX2 in cultures ofXenopus cells25.Expression of hTPX2 is regulated by the cell cycle. Previous data26,as well as our localization studies, suggested that the expression ofhTPX2 might be regulated during the cell cycle. We examined thispossibility by monitoring the protein in a synchronously growingpopulation of HeLa cells. Cells were arrested at the G1/S boundaryby a double thymidine block. After release (time 0), samples weretaken every 2 h for fluorescence-activated cell sorting (FACS),immunofluorescence (Fig. 3a) and western blot analysis of whole-cell lysates (Fig. 3b). FACS analysis indicated that before release72% of the cells were arrested in G1/S. Cells progressed into the cellcycle synchronously reaching a peak of mitosis after 10–12 h (datanot shown).
Immunofluorescence analysis showed that hTPX2 was localizedin the nuclei during S phase and accumulated there throughout G2(Fig. 3a, 0 and 8 h). Ten hours after release, a high percentage ofcells had entered mitosis and hTPX2 was localized on the mitoticspindles (Fig. 3a, 10 h). After mitosis hTPX2 disappeared (Fig. 3a,16 h). Western blot analysis confirmed these results and showedthat the cellular concentration of hTPX2 increased between S phaseand mitosis (Fig. 3b, 0–12 h) and dropped at the end of mitosis(Fig. 3b, 14 and 16 h). As a loading control, α-tubulin was seen toincrease constantly over time, as expected (Fig. 3a, bottom). After
24 h and the completion of one whole cycle, hTPX2 re-accumulat-ed (Fig. 3b, 24 h). These results confirm that the expression ofhTPX2 is regulated tightly during the cell cycle.Overexpression of GFP–hTPX2 blocks cells in a prometaphase-like state. Although some cells expressing GFP–hTPX2 proceedednormally through the cell cycle (36.5%), a high proportion oftransfected cells (63.5%) that overexpressed the recombinant pro-tein showed markedly altered morphologies. Among those, manycorresponded to interphase cells (35.9% of overexpressing cells),with GFP–hTPX2 localized in both the nucleus and the cytoplasm.In these cells, microtubules were decorated by GFP–hTPX2 andwere heavily bundled (data not shown). In addition, microtubuleslost their typical interphase centrosome-dependent organization(data not shown). This marked reorganization is often seen in cellsoverexpressing MAPs27. Of the rest of the overexpressing cells,57.9% were classified as apoptotic, and 6.1% were arrested in mito-sis and had condensed chromosomes scattered on a radial micro-tubule aster. These microtubules often emanated from a ring-likestructure in the centre of the cell and had no detectable micro-tubule-organizing centres. Much GFP–hTPX2 was concentrated inthis ring and extended along some microtubule arrays (Fig. 4). Weconclude that the overexpression of GFP–hTPX2 blocks bipolarspindle formation in somatic cells.Antibodies specific for hTPX2 inhibit spindle formation. To testwhether the presence of TPX2-specific antibodies would interferewith spindle assembly, we first added affinity-purified antibodiesagainst hTPX2 to a Xenopus egg extract and analysed the spindlestructures formed there and in control reactions (Fig. 5a). ThehTPX2-specific antibodies caused severe defects in spindle forma-tion (6% of spindle formation versus 72% in the presence of con-trol antibodies; Fig. 5a) that were very similar to those observed inTPX2-depleted extracts16,25.
To study the role of hTPX2 during mitosis in living cells, weinjected the same hTPX2-specific antibodies into HeLa cells. First,HeLa cells were synchronized in G1/S phase by a double thymidineblock. Between 3 and 4 h after release, antibodies against hTPX2 or
0 2 4 6 8 10 12 14 16 24 Time (h)
hTPX2
Tubulin
Mr(K)
115
82
62
50
a
b
Time (h) 0 8 10 16 24
Figure 3 Localization of hTPX2 during the cell cycle in HeLa cells. HeLa cellswere arrested in S phase and released at time 0. TPX2 expression was monitoredby western blot (a) and immunofluorescence (b) at the indicated times using affinity-purified antibodies against hTPX2 protein and monoclonal antibodies against α-tubu-lin. a, A coverslip of synchronously growing cells was lysed with SDS sample buffer,
total lysates were run on an SDS gel, and TPX2 (top) and tubulin (bottom) detectedby western blotting. b, Samples were fixed using 3% paraformaldehyde and stainedwith the same antibodies against TPX2 (red) and tubulin (green). Staining of DNA by4′,6-diamidino-2-phenylindole (DAPI) is blue. Scale bar, 10 µm.
© 2002 Nature Publishing Group
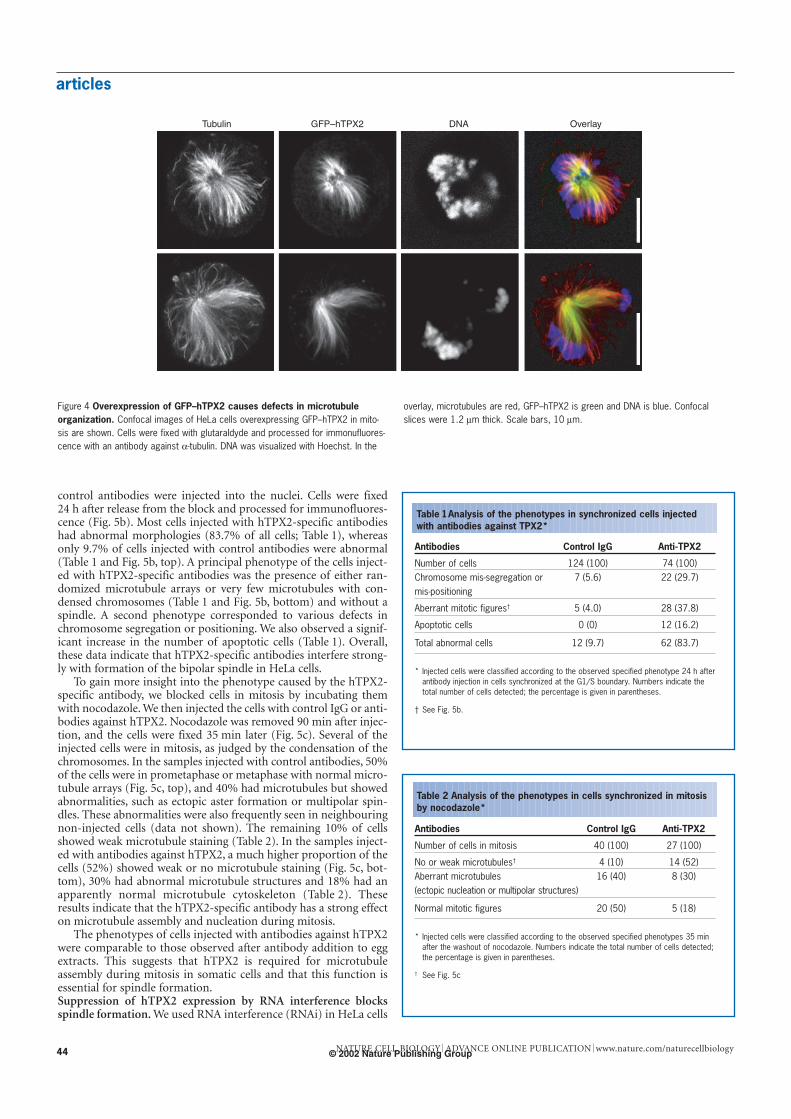
articles
NATURE CELL BIOLOGY ADVANCE ONLINE PUBLICATION www.nature.com/naturecellbiology44
control antibodies were injected into the nuclei. Cells were fixed24 h after release from the block and processed for immunofluores-cence (Fig. 5b). Most cells injected with hTPX2-specific antibodieshad abnormal morphologies (83.7% of all cells; Table 1), whereasonly 9.7% of cells injected with control antibodies were abnormal(Table 1 and Fig. 5b, top). A principal phenotype of the cells inject-ed with hTPX2-specific antibodies was the presence of either ran-domized microtubule arrays or very few microtubules with con-densed chromosomes (Table 1 and Fig. 5b, bottom) and without aspindle. A second phenotype corresponded to various defects inchromosome segregation or positioning. We also observed a signif-icant increase in the number of apoptotic cells (Table 1). Overall,these data indicate that hTPX2-specific antibodies interfere strong-ly with formation of the bipolar spindle in HeLa cells.
To gain more insight into the phenotype caused by the hTPX2-specific antibody, we blocked cells in mitosis by incubating themwith nocodazole. We then injected the cells with control IgG or anti-bodies against hTPX2. Nocodazole was removed 90 min after injec-tion, and the cells were fixed 35 min later (Fig. 5c). Several of theinjected cells were in mitosis, as judged by the condensation of thechromosomes. In the samples injected with control antibodies, 50%of the cells were in prometaphase or metaphase with normal micro-tubule arrays (Fig. 5c, top), and 40% had microtubules but showedabnormalities, such as ectopic aster formation or multipolar spin-dles. These abnormalities were also frequently seen in neighbouringnon-injected cells (data not shown). The remaining 10% of cellsshowed weak microtubule staining (Table 2). In the samples inject-ed with antibodies against hTPX2, a much higher proportion of thecells (52%) showed weak or no microtubule staining (Fig. 5c, bot-tom), 30% had abnormal microtubule structures and 18% had anapparently normal microtubule cytoskeleton (Table 2). Theseresults indicate that the hTPX2-specific antibody has a strong effecton microtubule assembly and nucleation during mitosis.
The phenotypes of cells injected with antibodies against hTPX2were comparable to those observed after antibody addition to eggextracts. This suggests that hTPX2 is required for microtubuleassembly during mitosis in somatic cells and that this function isessential for spindle formation.Suppression of hTPX2 expression by RNA interference blocksspindle formation. We used RNA interference (RNAi) in HeLa cells
Tubulin GFP–hTPX2 DNA Overlay
Figure 4 Overexpression of GFP–hTPX2 causes defects in microtubuleorganization. Confocal images of HeLa cells overexpressing GFP–hTPX2 in mito-sis are shown. Cells were fixed with glutaraldyde and processed for immonufluores-cence with an antibody against α-tubulin. DNA was visualized with Hoechst. In the
overlay, microtubules are red, GFP–hTPX2 is green and DNA is blue. Confocalslices were 1.2 µm thick. Scale bars, 10 µm.
Table 1 Analysis of the phenotypes in synchronized cells injectedwith antibodies against TPX2*
Antibodies Control IgG Anti-TPX2
Number of cells 124 (100) 74 (100)Chromosome mis-segregation or 7 (5.6) 22 (29.7)mis-positioning
Aberrant mitotic figures† 5 (4.0) 28 (37.8)
Apoptotic cells 0 (0) 12 (16.2)
Total abnormal cells 12 (9.7) 62 (83.7)
* Injected cells were classified according to the observed specified phenotype 24 h afterantibody injection in cells synchronized at the G1/S boundary. Numbers indicate thetotal number of cells detected; the percentage is given in parentheses.
† See Fig. 5b.
Table 2 Analysis of the phenotypes in cells synchronized in mitosisby nocodazole*
Antibodies Control IgG Anti-TPX2
Number of cells in mitosis 40 (100) 27 (100)
No or weak microtubules† 4 (10) 14 (52)Aberrant microtubules 16 (40) 8 (30)(ectopic nucleation or multipolar structures)
Normal mitotic figures 20 (50) 5 (18)
* Injected cells were classified according to the observed specified phenotypes 35 minafter the washout of nocodazole. Numbers indicate the total number of cells detected;the percentage is given in parentheses.
† See Fig. 5c
© 2002 Nature Publishing Group

articles
NATURE CELL BIOLOGY ADVANCE ONLINE PUBLICATION www.nature.com/naturecellbiology 45
to determine whether an absence of hTPX2 during mitosis wouldhave any effect on spindle formation. A small interfering RNA(siRNA) duplex targeted to a region located 160 base-pairs (bp)downstream of the start codon in hTPX2 mRNA was transfected intoHeLa cells. We verified the reduction in hTPX2 expression by westernblot analysis of lysates of siRNA-transfected cells 24 and 48 h aftertransfection (Fig. 6a). At 24 h, the expression of hTPX2 was reducedto 37.1% of that of a control asynchronous population. After 48 h,this expression was reduced further to 21.5%. This indicated that thesiRNA efficiently suppressed the expression of hTPX2 in HeLa cells.
Next, we scored the number of cells in different cell-cycle stages.After 36 h, 84 ± 8.6% (mean ± s.d.) of the cells transfected with
hTPX2 siRNA and 94 ± 2.8% of the control cells were in interphase(Fig. 6b and data not shown). Most of the non-interphase cells inthe sample transfected with hTPX2 siRNA showed a prometaphase-like appearance (12.2 ± 6.6%), whereas only 1.2 ± 0.8% of controlcells were in a similar configuration (Fig. 6b and data not shown).Although the number of cells arrested at that particular stage of thecell cycle was variable, this phenotype was observed consistently inall experiments (data not shown). Notably, later stages of mitosiswere hardly detected in cells transfected with hTPX2 siRNA(0.2 ± 0.3%), whereas a normal number of mitotic figures wasobserved in control cells (1.8 ± 0.6%).
We examined further the phenotype of cells arrested in aprometaphase-like state by confocal microscopy. All of these cells
Con
trol
ant
ibod
yA
nti-T
PX
2
TubulinDNAInjected Abs
Con
trol
ant
ibod
yA
nti-T
PX
2
b
c
Anti-TPX2Control antibodyNo additiona
Efficiencyof spindle formation (%) 81.1 ± 3.7 71.1 ± 16.7 5.5 ± 5.1
DNA + tubulin
Figure 5 Antibodies against hTPX2 inhibit spindle formation. a, Spindleassembly was monitored in Xenopus egg extracts as described in Fig. 1, in thepresence of 0.5 µg µl−1 antibodies against TPX2 or 0.5 µg µl−1 control antibodies,or in extracts alone. Tubulin is red, DNA is blue. Scale bar, 10 µm. The percentageefficiency of correct spindle formation was calculated from at least 140 micro-tubule assemblies around single chromatin structures in four samples. The means± s.d. are shown. b, c, Antibodies against TPX2 or control antibodies (control IgG)were injected into synchronized HeLa cells. After antibody injection, cells wereanalysed by immunofluorescence using antibodies against α-tubulin (green in over-lays), DAPI to stain DNA (blue in overlays) or secondary antibodies against theinjected antibodies, as indicated. Scale bar, 10 µm. b, HeLa cells were arrested inS phase using a double thymidine block and released synchronously by thymidinewashout. The indicated antibodies were injected 3–4 h after release, and cells werefixed 24 h after release. c, HeLa cells were arrested in early mitosis with nocoda-zole for 12 h. Mitotic cells were injected with the indicated antibodies, incubatedfor 90 min with nocodazole, released from the block by nocodazole washout, andfixed 35 min later.
Tubulin hTPX2 DNA Overlayc
hTPX2
Tubulin
24 h 48 h
C CRN
Ai
RN
Ai
a
0
Prom
etap
hase
-like
Inte
rpha
se
Prom
etap
hase
-like
Inte
rpha
se
40
60
Per
cent
age
of c
ells
0
80
100
20
Control cells TPX RNAi
Con
trol
cel
l(p
rom
etap
hase
)
TPX2RNAi
b
Figure 6 Suppression of hTPX2 expression by siRNA blocks spindle forma-tion. a, Western blot of HeLa cell lysates, prepared 24 and 48 h after transfectionwith either siRNA (RNAi) or control RNA (C), probed with antibodies against hTPX2and α-tubulin. The reduction in the amount of tubulin in cells transfected with TPX2siRNA is probably due to the observed growth arrest and to apoptosis in this popu-lation of cells. b, Quantification of the prometaphase-like state phenotype observedin HeLa cells transfected with siRNA. The cell-cycle stage was determined in con-trol cells and in cells transfected with TPX2 siRNA, and the percentages of inter-phase cells and cells in a prometaphase-like state were plotted. Shown are themeans from three independent experiments in which 200 cells were counted. Errorbars indicate the s.d. c, Triple immunofluorescence analysis of HeLa cells transfect-ed with siRNA. Cells were fixed with glutaraldehyde and processed for immunofluo-rescence with antibodies against hTPX2 and α-tubulin. DNA was visualised withHoechst. Confocal slices were 1.2 µm thick. In the overlay, microtubules are green,hTPX2 is red and DNA is blue. Scale bars, 10 µm.
© 2002 Nature Publishing Group
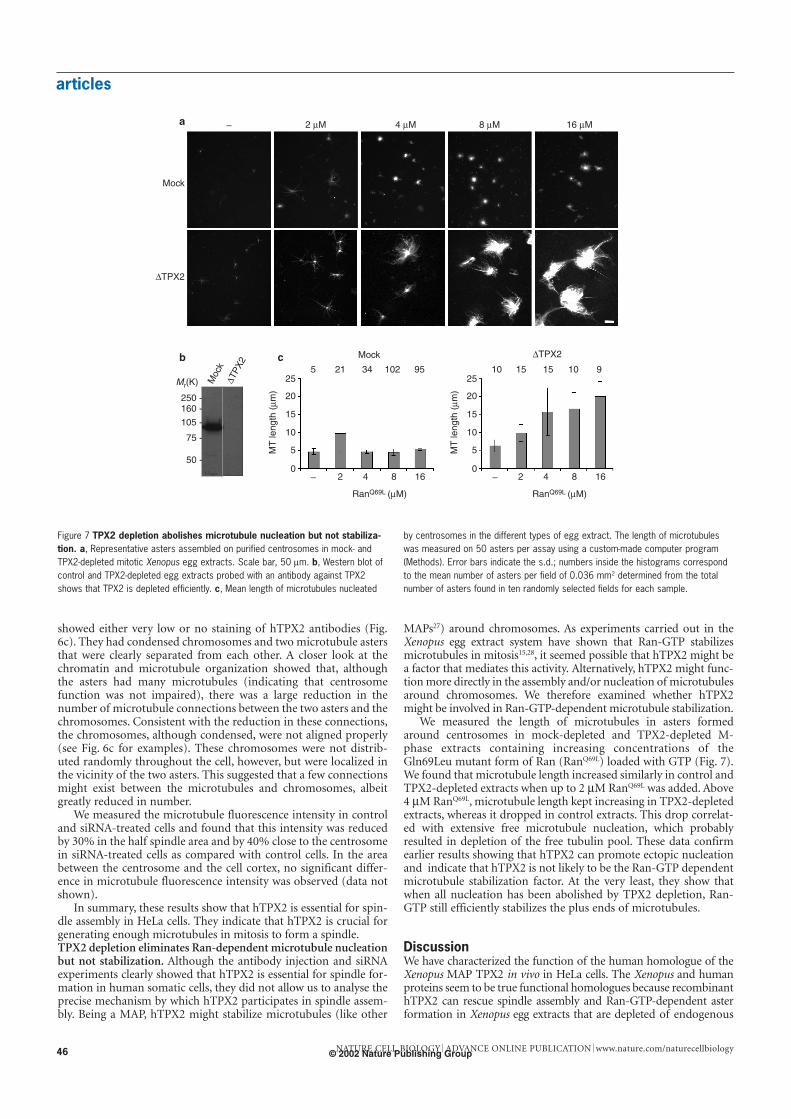
articles
NATURE CELL BIOLOGY ADVANCE ONLINE PUBLICATION www.nature.com/naturecellbiology46
showed either very low or no staining of hTPX2 antibodies (Fig.6c). They had condensed chromosomes and two microtubule astersthat were clearly separated from each other. A closer look at thechromatin and microtubule organization showed that, althoughthe asters had many microtubules (indicating that centrosomefunction was not impaired), there was a large reduction in thenumber of microtubule connections between the two asters and thechromosomes. Consistent with the reduction in these connections,the chromosomes, although condensed, were not aligned properly(see Fig. 6c for examples). These chromosomes were not distrib-uted randomly throughout the cell, however, but were localized inthe vicinity of the two asters. This suggested that a few connectionsmight exist between the microtubules and chromosomes, albeitgreatly reduced in number.
We measured the microtubule fluorescence intensity in controland siRNA-treated cells and found that this intensity was reducedby 30% in the half spindle area and by 40% close to the centrosomein siRNA-treated cells as compared with control cells. In the areabetween the centrosome and the cell cortex, no significant differ-ence in microtubule fluorescence intensity was observed (data notshown).
In summary, these results show that hTPX2 is essential for spin-dle assembly in HeLa cells. They indicate that hTPX2 is crucial forgenerating enough microtubules in mitosis to form a spindle.TPX2 depletion eliminates Ran-dependent microtubule nucleationbut not stabilization. Although the antibody injection and siRNAexperiments clearly showed that hTPX2 is essential for spindle for-mation in human somatic cells, they did not allow us to analyse theprecise mechanism by which hTPX2 participates in spindle assem-bly. Being a MAP, hTPX2 might stabilize microtubules (like other
MAPs27) around chromosomes. As experiments carried out in theXenopus egg extract system have shown that Ran-GTP stabilizesmicrotubules in mitosis15,28, it seemed possible that hTPX2 might bea factor that mediates this activity. Alternatively, hTPX2 might func-tion more directly in the assembly and/or nucleation of microtubulesaround chromosomes. We therefore examined whether hTPX2might be involved in Ran-GTP-dependent microtubule stabilization.
We measured the length of microtubules in asters formedaround centrosomes in mock-depleted and TPX2-depleted M-phase extracts containing increasing concentrations of theGln69Leu mutant form of Ran (RanQ69L) loaded with GTP (Fig. 7).We found that microtubule length increased similarly in control andTPX2-depleted extracts when up to 2 µM RanQ69L was added. Above4 µM RanQ69L, microtubule length kept increasing in TPX2-depletedextracts, whereas it dropped in control extracts. This drop correlat-ed with extensive free microtubule nucleation, which probablyresulted in depletion of the free tubulin pool. These data confirmearlier results showing that hTPX2 can promote ectopic nucleationand indicate that hTPX2 is not likely to be the Ran-GTP dependentmicrotubule stabilization factor. At the very least, they show thatwhen all nucleation has been abolished by TPX2 depletion, Ran-GTP still efficiently stabilizes the plus ends of microtubules.
DiscussionWe have characterized the function of the human homologue of theXenopus MAP TPX2 in vivo in HeLa cells. The Xenopus and humanproteins seem to be true functional homologues because recombinanthTPX2 can rescue spindle assembly and Ran-GTP-dependent asterformation in Xenopus egg extracts that are depleted of endogenous
– 2 µM 4 µM 8 µM 16 µM
Mock
∆TPX2
Mr(K)
250 -160 -
105 -
75 -
50 -
Moc
k∆T
PX2
25
20
15
10
5
0
25
20
15
10
5
0
5 21 34 102 95 10 15 15 10 9
Mock
– 2 4 8 16 – 2 4 8 16
∆TPX2
RanQ69L (µM) RanQ69L (µM)
MT
leng
th (
µm)
MT
leng
th (
µm)
a
b c
Figure 7 TPX2 depletion abolishes microtubule nucleation but not stabiliza-tion. a, Representative asters assembled on purified centrosomes in mock- andTPX2-depleted mitotic Xenopus egg extracts. Scale bar, 50 µm. b, Western blot ofcontrol and TPX2-depleted egg extracts probed with an antibody against TPX2shows that TPX2 is depleted efficiently. c, Mean length of microtubules nucleated
by centrosomes in the different types of egg extract. The length of microtubuleswas measured on 50 asters per assay using a custom-made computer program(Methods). Error bars indicate the s.d.; numbers inside the histograms correspondto the mean number of asters per field of 0.036 mm2 determined from the totalnumber of asters found in ten randomly selected fields for each sample.
© 2002 Nature Publishing Group
articles
NATURE CELL BIOLOGY ADVANCE ONLINE PUBLICATION www.nature.com/naturecellbiology 47
TPX2 (ref. 16). In the egg extract system, TPX2 is required for spin-dle assembly because it is needed for the formation of microtubulesaround chromosomes and, in turn, this population of microtubulesis essential for spindle assembly16. But it has been unclear whethera similar centrosome-independent pathway of spindle assemblyexists in vertebrate somatic cells. Indeed, the predominant view hasbeen that a mechanism based on the selective stabilization of cen-trosome-nucleated microtubules around the chromosomes is themain organizing principle in spindle assembly.
To clarify this issue we investigated the effect of interfering withhTPX2 function in vivo in somatic cells in culture. Our resultsshowed that hTPX2 is essential for spindle assembly in HeLa cells.Altering the amounts of hTPX2 by either overexpression or deple-tion, as well as by antibody microinjection led to a failure in spin-dle assembly. Antibody injection experiments indicated that hTPX2is involved in microtubule assembly. This was confirmed by RNAiexperiments, which generated a very marked phenotype: twomicrotubule asters formed around the duplicated centrosomes butfailed to interact stably, either with chromosomes or between them-selves. A significant reduction in the microtubule density towardsthe chromosomes was observed. As a consequence, a functionalspindle did not form, leading to a strong mitotic block.
Ran-GTP is known to regulate the activity of TPX2 in Xenopus eggextracts. The phenotype that we observed in HeLa cells depleted ofhTPX2 is reminiscent of that obtained in Caenorhabditis elegansembryos depleted of Ran by RNAi (ref. 29 and Askjaer et al. (see noteadded in proof)) or in Xenopus egg extracts containing the dominant-negative Thr24Asn form of Ran (RanT24N)15. This latter experiment15
showed that Ran has various effects on microtubules during mitosis,including the stabilization of mitotic microtubules15,28. Thus, it is pos-sible that TPX2, when released from its interaction with importin-αand importin-β by Ran-GTP, might stabilize astral microtubules.Such an activity alone could be responsible for promoting micro-tubule nucleation around chromosomes and for the stabilization ofastral microtubules on the chromosomes. This would mean that thelack of normal spindle assembly in the siRNA-treated or antibody-injected HeLa cells would have resulted from a lack of selective stabi-lization of astral microtubules around the chromosomes.
Alternatively, it is possible that TPX2 might promote micro-tubule assembly by stimulating microtubule nucleation itself. We
found that when TPX2 was depleted from egg extracts in such a waythat all Ran-GTP-dependent nucleation was eliminated, the astralmicrotubules were still stabilized in a Ran-GTP-dependent manner.This supports the idea that in mitotic HeLa cells the nucleation ofmicrotubules around chromosomes is mediated by hTPX2 and thata bipolar spindle cannot assemble when this population of nucleat-ed microtubules is absent in cells that do not express hTPX2.
In TPX2-depleted cells, numerous microtubules are nucleatedby the centrosomes and centrosome duplication and separationseems to proceed normally. The centrosomal microtubules shouldbe stabilized through the effect of Ran-GTP as shown here and pre-viously15,28. But these centrosomal microtubules do not seem toestablish stable interactions with the chromosomes and do notorganize a spindle. This indicates that in the absence of hTPX2 nei-ther kinetochores nor chromokinesins can capture and stabilize asufficient number of centrosome-nucleated microtubules to makea spindle. Chromokinesins are likely to be the mediators of micro-tubule attachment to chromosome arms6, but in fact the mecha-nism by which chromokinesins such as Xklp1 participate in spindleassembly may be through binding and moving microtubules nucle-ated by chromosomes rather than centrosomes6.
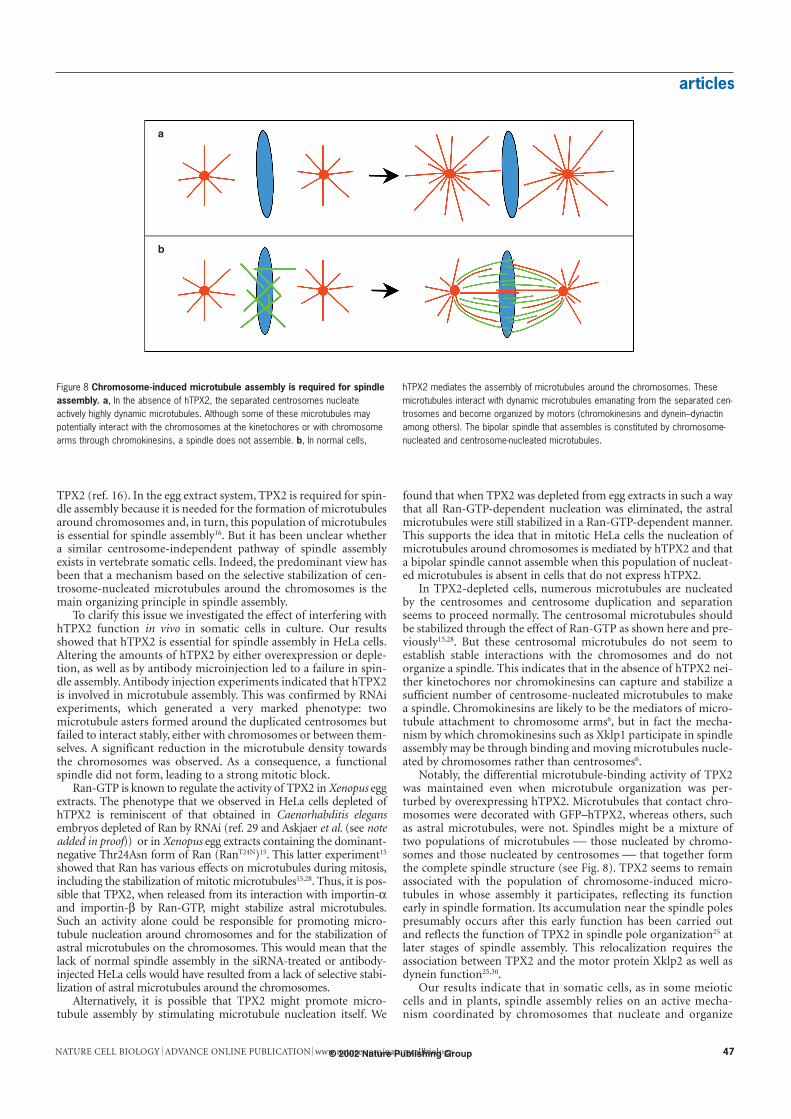
Notably, the differential microtubule-binding activity of TPX2was maintained even when microtubule organization was per-turbed by overexpressing hTPX2. Microtubules that contact chro-mosomes were decorated with GFP–hTPX2, whereas others, suchas astral microtubules, were not. Spindles might be a mixture oftwo populations of microtubules those nucleated by chromo-somes and those nucleated by centrosomes that together formthe complete spindle structure (see Fig. 8). TPX2 seems to remainassociated with the population of chromosome-induced micro-tubules in whose assembly it participates, reflecting its functionearly in spindle formation. Its accumulation near the spindle polespresumably occurs after this early function has been carried outand reflects the function of TPX2 in spindle pole organization25 atlater stages of spindle assembly. This relocalization requires theassociation between TPX2 and the motor protein Xklp2 as well asdynein function25,30.
Our results indicate that in somatic cells, as in some meioticcells and in plants, spindle assembly relies on an active mecha-nism coordinated by chromosomes that nucleate and organize
a
b
Figure 8 Chromosome-induced microtubule assembly is required for spindleassembly. a, In the absence of hTPX2, the separated centrosomes nucleateactively highly dynamic microtubules. Although some of these microtubules maypotentially interact with the chromosomes at the kinetochores or with chromosomearms through chromokinesins, a spindle does not assemble. b, In normal cells,
hTPX2 mediates the assembly of microtubules around the chromosomes. Thesemicrotubules interact with dynamic microtubules emanating from the separated cen-trosomes and become organized by motors (chromokinesins and dynein–dynactinamong others). The bipolar spindle that assembles is constituted by chromosome-nucleated and centrosome-nucleated microtubules.
© 2002 Nature Publishing Group

articles
NATURE CELL BIOLOGY ADVANCE ONLINE PUBLICATION www.nature.com/naturecellbiology48
microtubules into the bipolar spindle apparatus. They stronglysuggest that this is a general pathway and that the function of TPX2is conserved in all vertebrate cells. Whether TPX2 is regulateddirectly by Ran-GTP in all of these cells remains to be determined.Note added in proof: the reference by Askjaer et al. is now in the press(Askjaer, P., Galy, V., Hannak, E. & Mattaj, I. W. The Ran GTPasecycle and importins α and β are essential for spindle formation andnuclear envelope assembly in living Caenorhabditis elegans embryos.MBC Online (cited 3 September 2002); DOI: 10.1091/mbc.E02-06-0346 (2002).
MethodsTPX2 cDNA constructsFor recombinant expression in Escherichia coli, the human TPX2 cDNA was cloned into a vector based
on pQE 70 (Qiagen, Hilden, Germany). An amino-terminal protein-A-binding domain (zz-domain)
followed by enhanced GFP (EGFP) was fused to the human TPX2 cDNA followed by a histidine tag
from the pQE vector. For transfection experiments, we amplified the human TPX2 cDNA by poly-
merase chain reaction with primers that introduced HindIII and BamH1 restriction sites and cloned it
into pEGFP-C3 (Clontech, Palo Alto, CA).
Recombinant protein expressionWe expressed TPX2 and zz-tagged TPX2 in E. coli and purified them on TALON beads (Clontech) as
described16. RanQ69L was expressed, purified and loaded with GTP as described31.
Western blotting and SDS–PAGETo analyse the expression of TPX2 throughout the cell cycle by western blotting, a cover slip with syn-
chronized HeLa cells (see below) was taken at each indicated time point after release from S phase. We
lysed cells by adding 20 µl of sample buffer32, ran samples on 12% SDS32 gels and then transferred
them onto Nitrocellulose (Schleicher und Schuell, Dassel, Germany). TPX2 and tubulin were detected
with the antibodies described below.
Antibodies, microinjectionsAntibodies against TPX2 were generated in rabbits using recombinantly expressed full-length hTPX2
as an antigen in accordance with standard procedures. Polyclonal sera were affinity purified using the
same recombinant TPX2 protein covalently linked to AffiGel 10 (Biometra, Göttingen, Germany).
Monoclonal antibodies against β-tubulin were purchased from Sigma (#T9026; Munich, Germany).
HeLa cells were arrested for 12 h using 40 ng ml−1 nocodazole33, injected with rabbit IgG (Sigma) or
TPX2 antibodies (both at 3 mg ml−1 in PBS), and incubated for 90 min in Dulbecco’s modified Eagle’s
medium in the presence of nocodazole. Cells were released by washing out the nocodazole twice care-
fully with pre-warmed medium. After 35 min, cells were fixed in 3% paraformaldehyde and analysed
by immunofluorescence. Only cells with condensed chromatin were considered to be mitotic and
counted. Alternatively, cells were synchronized by a double thymidine block as described below. We
injected cells 3–4 h after release from S phase with affinity-purified antibodies against TPX2 or rabbit
IgG as described34. We injected 150 cells in three different experiments and grew the cells for another
20 h (24 h time point) before fixation as above. Injected cells were detected by immunofluorescence
using specific secondary antibodies.
Egg extract preparation and depletionWe prepared CSF-arrested egg extracts as described35. TPX2 depletion was essentially done as
described16 using affinity-purified polyclonal antibodies against TPX2 (antibodies against Xenopus or
human TPX2 worked equally well), or rabbit IgG (Sigma) as a control, pre-coupled to magnetizable
protein A beads (Dynal, Oslo, Norway). We incubated 100 µl of CSF-arrested extracts with 50 µl of
beads that had been pre-incubated with antibodies at a concentration of 1 µg of antibody per
microlitre of bead slurry. Cycled spindle assembly experiments were done as described in Xenopus egg
extracts containing 0.2 mg ml−1 rhodamine-conjugated tubulin36.
Centrosomal asters were assembled as described25. In brief, recombinant RanQ69L preloaded with
GTP was added dose dependently to 10 µl of control or TPX2-depleted CSF-arrested extracts contain-
ing 0.2 mg ml−1 rhodamine-conjugated tubulin. Asters were assembled by addition of human centro-
somes purified from human KE37 lymphoid cells as described37. The reactions were incubated for
30 min at 20 °C, fixed and centrifuged onto coverslips30. We acquired images on a Zeiss Axioskop 2
microscope (Oberkochen, Germany). The average length of the microtubules in each aster was deter-
mined using a custom macro written using Matlab (available from I.V. on request) on the basis of pro-
files obtained by adding the pixel values on each of several circles positioned at increasing distances
from the aster centre. We measured 50 asters for each RanQ69L concentration. We quantified the num-
ber of asters by counting the total number of asters in ten random fields of 0.036mm2 per condition.
Cell culture and transfectionHeLa cells (CCL2; ATCC, Manassas, VA) were grown in 10% fetal calf serum (FCS) and 2 mM L-glut-
amine in modified Eagle’s medium at 37 °C and 5% CO2 and were kept below passage 25. Cells were
arrested in S phase using a double thymidine (2 mM) block as described38 and released from S phase
by washing away the thymidine (referred to as time 0). We monitored cell-cycle progression by FACS
analysis in accordance with standard protocols. Transfections of HeLa with TPX2 cDNA constructs
were done with Efectene (Qiagen) in accordance with the manufacturer’s recommendations.
Immunofluorescence and microscopyHeLa cells were fixed in methanol at –20 °C, in 0.25% glutaraldehyde or in 4% formaldehyde and
processed for imunofluorescence as described25. We examined fixed cells on a Zeiss Axioskop fluores-
cence microscope or on a Zeiss LSM 510 confocal microscope. Images were captured with the F-View
black and white camera (Soft Imaging System, Münster, Germany). For time-lapse video microscopy,
cells were grown on glass bottom Microwell petri dishes (MatTek, Ashland, MA), and placed in car-
bonate-free culture medium with 5% FCS and 20 mM HEPES buffer at pH 7.4. Imaging was done
using the Ultraview Software on a Perkin Elmer spinning disc microscope (Nikon, Düsseldorf,
Germany). Maximum brightness four-dimensional projections were made using Interactive Data lan-
guage (Research Systems Inc., Victoria, Australia).
RNA interferenceWe carried out RNAi experiments with siRNA as described39. The region of hTPX2 cDNA targeted for
siRNA was 5′-GAAUGGAACUGGAGGGCUU-3′, corresponding to a region located 160-bp down-
stream from the start codon. Synthetic sense and antisense oligonucleotides (Dharmacon, Lafayette,
CO) were annealed in 100 mM potassium acetate, 30 mM HEPES-KOH (pH 7.4), 2 mM magnesium
acetate for 1 min at 90 °C and 1 h at 37 °C, and frozen. Oligos were transfected into cells with
Oligofectamine (Invitrogen, Karlsruhe, Germany) in accordance with the supplier’s recommendations.
In brief, 4.5 µl of 20 µM siRNA duplex was mixed with 150 µl of Opti-MEM (GibcoBRL, Karlsruhe,
Germany), and 9 µl of Oligofectamine with 36 µl of Opti-MEM. These solutions were incubated for
7–10 min, mixed together and incubated for 20–25 min before being added to the cells. Cells were col-
lected for immunoblotting or fixed for immunostaining 24 and 48 h after transfection.
RECEIVED 29 APRIL 2002, REVISED 7 AUGUST 2002, ACCEPTED26 AUGUST 2002,
PUBLISHED 21 OCTOBER 2002.
1. Wittmann, T., Hyman, A. & Desai, A. The spindle: a dynamic assembly of microtubules and
motors. Nature Cell Biol. 3, E28–E34 (2001).
2. Compton, D. A. Spindle assembly in animal cells. Annu. Rev. Biochem. 69, 95–114 (2000).
3. Kirschner, M. & Mitchison, T. Beyond self-assembly: from microtubules to morphogenesis. Cell 45,
329–342 (1986).
4. Karsenti, E. & Vernos, I. The mitotic spindle: a self-made machine. Science 294, 543–547 (2001).
5. Heald, R. et al. Self-organization of microtubules into bipolar spindles around artificial chromo-
somes in Xenopus egg extracts. Nature 382, 420–425 (1996).
6. Walczak, C. E., Vernos, I., Mitchison, T. J., Karsenti, E. & Heald, R. A model for the proposed roles
of different microtubule-based motor proteins in establishing spindle bipolarity. Curr. Biol. 8,
903–913 (1998).
7. Hetzer, M., Gruss, O. J. & Mattaj, I. W. The Ran GTPase as a marker for chromosome position in
spindle formation and nuclear envelope assembly. Nature Cell Biol. 4, E177–E184 (2002).
8. Dasso, M. Running on Ran: nuclear transport and the mitotic spindle. Cell 104, 321–324 (2001).
9. Carazo-Salas, R. E. et al. Generation of GTP-bound Ran by RCC1 is required for chromatin-
induced mitotic spindle formation. Nature 400, 178–181 (1999).
10. Kalab, P., Pu, R. T. & Dasso, M. The Ran GTPase regulates mitotic spindle assembly. Curr. Biol. 9,
481–484 (1999).
11. Ohba, T., Nakamura, M., Nishitani, H. & Nishimoto, T. Self-organization of microtubule asters
induced in Xenopus egg extracts by GTP-bound Ran. Science 284, 1356–1358 (1999).
12. Wilde, A. & Zheng, Y. Stimulation of microtubule aster formation and spindle assembly by the
small GTPase Ran. Science 284, 1359–1362 (1999).
13. Zhang, C., Hughes, M. & Clarke, P. R. Ran-GTP stabilises microtubule asters and inhibits nuclear
assembly in Xenopus egg extracts. J. Cell Sci. 112, 2453–2461 (1999).
14. Kalab, P., Weis, K. & Heald, R. Visualization of a Ran-GTP gradient in interphase and mitotic
Xenopus egg extracts. Science 295, 2452–2456 (2002).
15. Carazo-Salas, R. E., Gruss, O. J., Mattaj, I. W. & Karsenti, E. Ran-GTP coordinates regulation of
microtubule nucleation and dynamics during mitotic-spindle assembly. Nature Cell Biol. 3,
228–234 (2001).
16. Gruss, O. J. et al. Ran induces spindle assembly by reversing the inhibitory effect of importin α on
TPX2 activity. Cell 104, 83–93 (2001).
17. Nachury, M. V. et al. Importin β is a mitotic target of the small GTPase Ran in spindle assembly.
Cell 104, 95–106 (2001).
18. Wiese, C. et al. Role of importin-β in coupling Ran to downstream targets in microtubule assem-
bly. Science 291, 653–656 (2001).
19. Khodjakov, A., Cole, R. W., Oakley, B. R. & Rieder, C. L. Centrosome-independent mitotic spindle
formation in vertebrates. Curr. Biol. 10, 59–67 (2000).
20. Vaizel-Ohayon, D. & Schejter, E. D. Mutations in centrosomin reveal requirements for centrosomal
function during early Drosophila embryogenesis. Curr. Biol. 9, 889–898 (1999).
21. Megraw, T. L., Kao, L. R. & Kaufman, T. C. Zygotic development without functional mitotic centro-
somes. Curr. Biol. 11, 116–120 (2001).
22. Bonaccorsi, S., Giansanti, M. G. & Gatti, M. Spindle assembly in Drosophila neuroblasts and gan-
glion mother cells. Nature Cell Biol. 2, 54–56 (2000).
23. Bonaccorsi, S., Giansanti, M. G. & Gatti, M. Spindle self-organization and cytokinesis during male
meiosis in asterless mutants of Drosophila melanogaster. J. Cell Biol. 142, 751–761 (1998).
24. Llamazares, S., Tavosanis, G. & Gonzalez, C. Cytological characterisation of the mutant phenotypes
produced during early embryogenesis by null and loss-of-function alleles of the gammaTub37C
gene in Drosophila. J. Cell Sci. 112, 659–667 (1999).
25. Wittmann, T., Wilm, M., Karsenti, E. & Vernos, I. TPX2, A novel Xenopus MAP involved in spindle
pole organization. J. Cell Biol. 149, 1405–1418 (2000).
26. Heidebrecht, H. J. et al. p100: a novel proliferation-associated nuclear protein specifically restricted
to cell cycle phases S, G2, and M. Blood 90, 226–233 (1997).
27. Cassimeris, L. & Spittle, C. Regulation of microtubule-associated proteins. Int. Rev. Cytol. 210,
163–226 (2001).
28. Wilde, A. et al. Ran stimulates spindle assembly by altering microtubule dynamics and the balance
of motor activities. Nature Cell Biol. 3, 221–227 (2001).
29. Bamba, C., Bobinnec, Y., Fukuda, M. & Nishida, E. The GTPase Ran regulates chromosome posi-
tioning and nuclear envelope assembly in vivo. Curr. Biol. 12, 503–507 (2002).
30. Wittmann, T., Boleti, H., Antony, C., Karsenti, E. & Vernos, I. Localization of the kinesin-like pro-
tein Xklp2 to spindle poles requires a leucine zipper, a microtubule-associated protein, and dynein.
J. Cell Biol. 143, 673–685 (1998).
31. Weis, K., Dingwall, C. & Lamond, A. I. Characterization of the nuclear protein import mechanism
© 2002 Nature Publishing Group

using Ran mutants with altered nucleotide binding specificities. EMBO J. 15, 7120–7128 (1996).
32. Laemmli, U. K. Cleavage of structural proteins during the assembly of the head of bacteriophage
T4. Nature 277, 680–685 (1970).
33. Zieve, G. W., Turnbull, D., Mullins, J. M. & McIntosh, J. R. Production of large numbers of mitotic
mammalian cells by use of the reversible microtubule inhibitor nocodazole. Nocodazole accumu-
lated mitotic cells. Exp. Cell. Res. 126, 397–405 (1980).
34. Pepperkok, R., Saffrich, R. & Ansorge, W. in Cell Biology: A Laboratory Handbook (ed. Celis, J. E.)
23–36 (Academic, San Diego, CA, 1998).
35. Murray, A. in Xenopus laevis: Practical Uses in Cell and Molecular Biology (eds Kay, B. K. & Peng, H.
B.) 581–605 (Academic, San Diego, 1991).
36. Sawin, K. E. & Mitchison, T. J. Mitotic spindle assembly by two different pathways in vitro. J. Cell
Biol. 112, 925–940 (1991).
37. Bornens, M., Paintrand, M., Berges, J., Marty, M. C. & Karsenti, E. Structural and chemical charac-
terization of isolated centrosomes. Cell Motil. Cytoskel. 8, 238–249 (1987).
38. Stein, G. S. et al. in Cell Biology: A Laboratory Handbook (ed. Celis, J. E.) 282–287 (Academic, San Diego, 1994).
39. Elbashir, S. M. et al. Duplexes of 21-nucleotide RNAs mediate RNA interference in cultured mam-
malian cells. Nature 411, 494–498 (2001).
ACKNOWLEDGEMENTS
We thank T. Zimmermann for help with confocal microscopes and image acquisition, and E. Nigg for
sharing unpublished data. This work was funded by the European Molecular Biology Laboratory
(EMBL). O.J.G. was supported in part by a Deutsche Forschungsgemeinschaft fellowship and in part
by an EMBL postdoctoral fellowship.
Correspondence and requests for materials should be addressed to I.V.
COMPETING FINANCIAL INTERESTS
The authors declare that they have no competing financial interests.
articles
NATURE CELL BIOLOGY ADVANCE ONLINE PUBLICATION www.nature.com/naturecellbiology 49© 2002 Nature Publishing Group




















