
Molecular pathogenesis of focal nodular hyperplasia and hepatocellular adenoma

Brain (1999),122,219–238
Characterization of nodular neuronal heterotopia inchildrenAnthony J. Hannan,1 Sandrine Servotte,1 Alla Katsnelson,1 Sanjay Sisodiya,2,* Colin Blakemore,1
Marian Squier3 and Zoltan Molnar1,†
1University Laboratory of Physiology, University of Oxford, Correspondence to: Dr Zolta´n Molnar, Institut de BiologieDepartments of2Neurology and3Neuropathology, Radcliffe Cellulaire et de Morphologie, Rue du Bugnon 9,Infirmary, Oxford, UK 1005 Lausanne, Switzerland
E-mail: [email protected] Dr Marian Squier, Department of Neuropathology,Radcliffe Infirmary, Woodstock Road, Oxford OX2 6HE,UKE-mail: [email protected]
*Present address: Epilepsy Research Group, Institute ofNeurology, Queen Square, London WC1N 3BG, UK†Present address: Institut de Biologie Cellulaire et deMorphologie, Lausanne, Switzerland
SummaryNeuronal heterotopia are seen in various pathologies andare associated with intractable epilepsy. We examinedbrain tissue from four children with subcortical orperiventricular nodular heterotopia of differentaetiologies: one with severe epilepsy following focal braintrauma at 17 weeks gestation, one with hemimega-lencephaly and intractable epilepsy, one with focal corticaldysplasia and intractable epilepsy, and one dysmorphicterm infant with associated hydrocephalus andpolymicrogyria. The connectivity of nodules wasinvestigated using histological and carbocyanine dye (DiI)tracing techniques. DiI crystal placement adjacent toheterotopic nodules revealed numerous DiI-labelled fibreswithin a 2–3 mm radius of the crystals. Although weobserved labelled fibres closely surrounding nodules, themajority did not penetrate them. Placement of DiI crystalswithin nodules also identified a limited number ofprojections out of the nodules and in one case there wasevidence for connectivity between adjacent nodules. Thecellular and neurochemical composition of nodules was
Keywords: epilepsy; heterotopia; calretinin; neuropeptide Y; cortex
Abbreviations: DiI 5 1,19-dioctadecyl-3,3,39,39-tetramethylindocarbocyanine perchlorate; GABA5 γ-aminobutyric acid;LBCV 5 luxol blue/cresyl violet; NPY5 neuropeptide Y
IntroductionRecent MRI studies have shown that developmental brainabnormalities of structure, or malformations of corticaldevelopment, are an important cause of refractory epilepsyin adults (Li et al., 1995); their prevalence in children with
© Oxford University Press 1999
also examined using immunohistochemistry for calretininand neuropeptide Y (NPY), which are normally expressedin GABAergic cortical interneurons. Within heterotopicnodules from all cases, numerous calretinin-positiveneurons were identified, along with a few cell bodiesand many processes positive for NPY. Calretinin-positiveneurons within nodules were less morphologically complexthan those in the cortex, which may reflect incompletedifferentiation into an inhibitory neuronal phenotype.There were also abnormal clusters of calretinin-positivecells in the overlying cortical plate, indicating that themigratory defect which produces heterotopic nodules alsoaffects development of the cortex itself. Thus, heterotopicnodules consisting of multiple neuronal cell types areassociated with malformation in the overlying corticalplate, and have limited connectivity with other brainregions. This abnormal development of connectivity mayaffect neuronal maturation and consequently the balanceof excitation and inhibition in neuronal circuits, leadingto their epileptogenic potential.
refractory epilepsy is less well studied but is increasinglyrecognized. Normal development of the cerebral cortex occursthrough a co-ordinated programme of cellular events,including cell division, migration, selective cell death and
by guest on June 17, 2016http://brain.oxfordjournals.org/
Dow
nloaded from
220 A. J. Hannanet al.
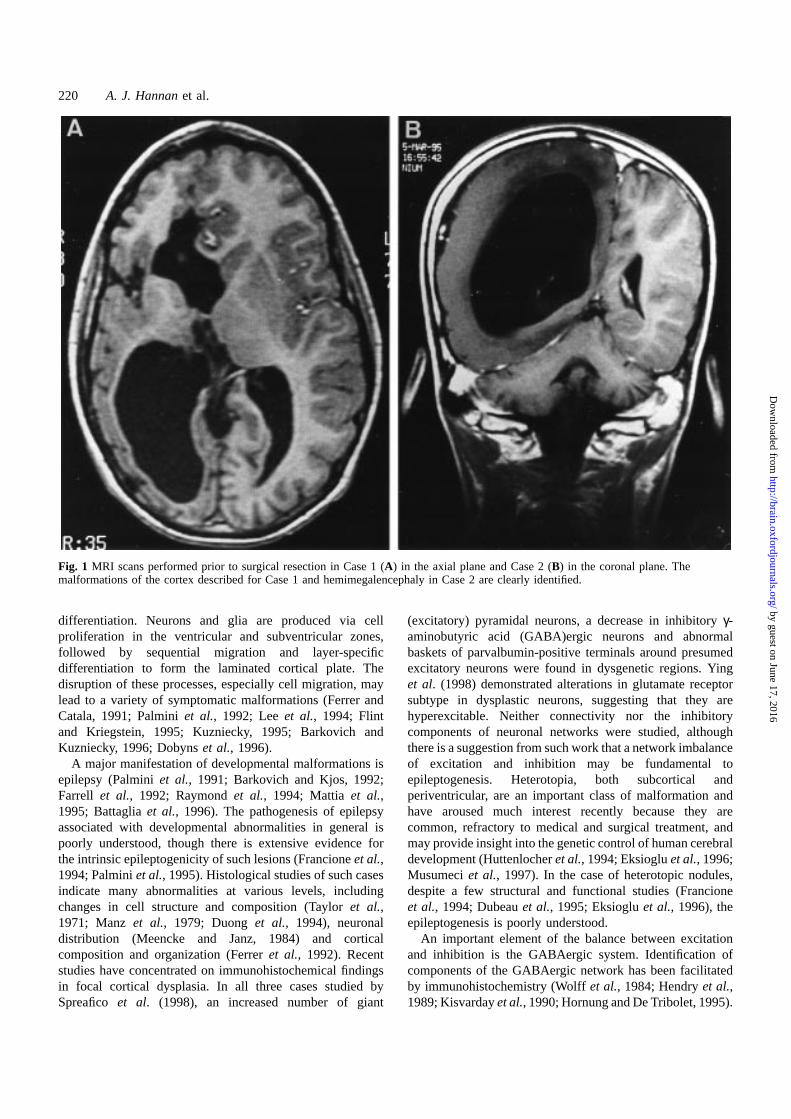
Fig. 1 MRI scans performed prior to surgical resection in Case 1 (A) in the axial plane and Case 2 (B) in the coronal plane. Themalformations of the cortex described for Case 1 and hemimegalencephaly in Case 2 are clearly identified.
differentiation. Neurons and glia are produced via cellproliferation in the ventricular and subventricular zones,followed by sequential migration and layer-specificdifferentiation to form the laminated cortical plate. Thedisruption of these processes, especially cell migration, maylead to a variety of symptomatic malformations (Ferrer andCatala, 1991; Palminiet al., 1992; Leeet al., 1994; Flintand Kriegstein, 1995; Kuzniecky, 1995; Barkovich andKuzniecky, 1996; Dobynset al., 1996).
A major manifestation of developmental malformations isepilepsy (Palminiet al., 1991; Barkovich and Kjos, 1992;Farrell et al., 1992; Raymondet al., 1994; Mattiaet al.,1995; Battagliaet al., 1996). The pathogenesis of epilepsyassociated with developmental abnormalities in general ispoorly understood, though there is extensive evidence forthe intrinsic epileptogenicity of such lesions (Francioneet al.,1994; Palminiet al., 1995). Histological studies of such casesindicate many abnormalities at various levels, includingchanges in cell structure and composition (Tayloret al.,1971; Manz et al., 1979; Duonget al., 1994), neuronaldistribution (Meencke and Janz, 1984) and corticalcomposition and organization (Ferreret al., 1992). Recentstudies have concentrated on immunohistochemical findingsin focal cortical dysplasia. In all three cases studied bySpreafico et al. (1998), an increased number of giant
(excitatory) pyramidal neurons, a decrease in inhibitoryγ-aminobutyric acid (GABA)ergic neurons and abnormalbaskets of parvalbumin-positive terminals around presumedexcitatory neurons were found in dysgenetic regions. Yinget al. (1998) demonstrated alterations in glutamate receptorsubtype in dysplastic neurons, suggesting that they arehyperexcitable. Neither connectivity nor the inhibitorycomponents of neuronal networks were studied, althoughthere is a suggestion from such work that a network imbalanceof excitation and inhibition may be fundamental toepileptogenesis. Heterotopia, both subcortical andperiventricular, are an important class of malformation andhave aroused much interest recently because they arecommon, refractory to medical and surgical treatment, andmay provide insight into the genetic control of human cerebraldevelopment (Huttenlocheret al., 1994; Eksiogluet al., 1996;Musumeciet al., 1997). In the case of heterotopic nodules,despite a few structural and functional studies (Francioneet al., 1994; Dubeauet al., 1995; Eksiogluet al., 1996), theepileptogenesis is poorly understood.
An important element of the balance between excitationand inhibition is the GABAergic system. Identification ofcomponents of the GABAergic network has been facilitatedby immunohistochemistry (Wolffet al., 1984; Hendryet al.,1989; Kisvardayet al., 1990; Hornung and De Tribolet, 1995).
by guest on June 17, 2016http://brain.oxfordjournals.org/
Dow
nloaded from
Neuronal heterotopia and epilepsy 221
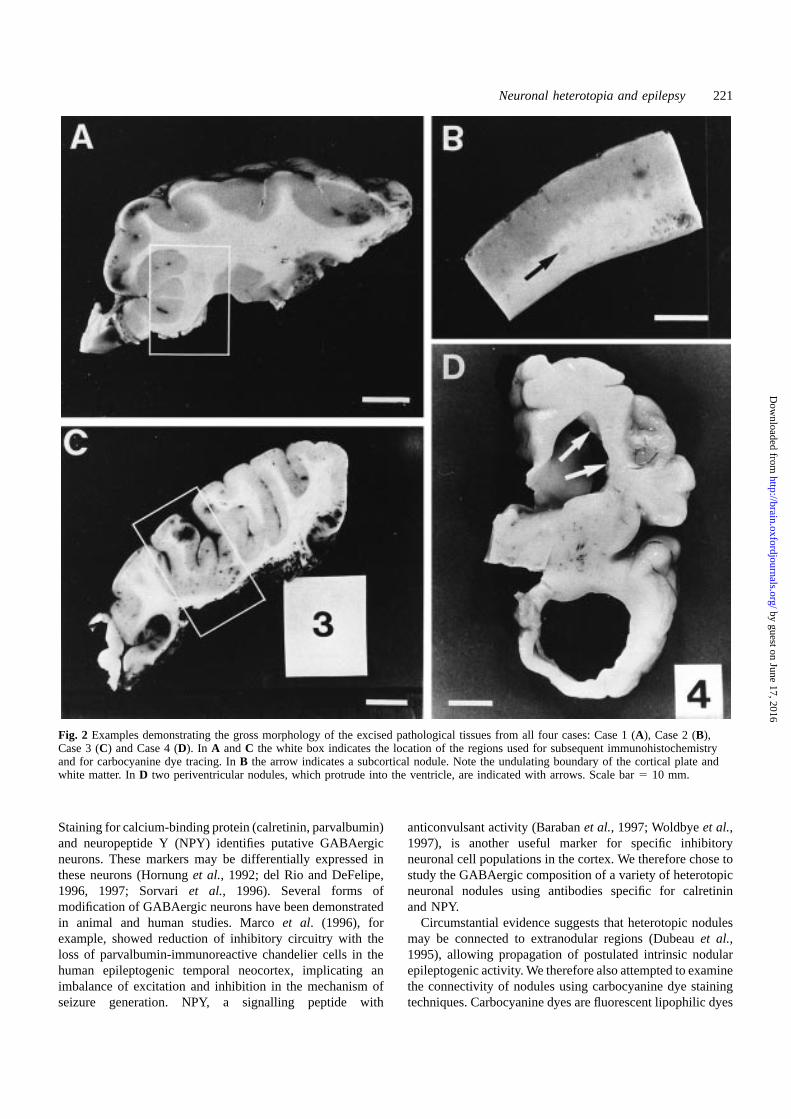
Fig. 2 Examples demonstrating the gross morphology of the excised pathological tissues from all four cases: Case 1 (A), Case 2 (B),Case 3 (C) and Case 4 (D). In A andC the white box indicates the location of the regions used for subsequent immunohistochemistryand for carbocyanine dye tracing. InB the arrow indicates a subcortical nodule. Note the undulating boundary of the cortical plate andwhite matter. InD two periventricular nodules, which protrude into the ventricle, are indicated with arrows. Scale bar5 10 mm.
Staining for calcium-binding protein (calretinin, parvalbumin)and neuropeptide Y (NPY) identifies putative GABAergicneurons. These markers may be differentially expressed inthese neurons (Hornunget al., 1992; del Rio and DeFelipe,1996, 1997; Sorvariet al., 1996). Several forms ofmodification of GABAergic neurons have been demonstratedin animal and human studies. Marcoet al. (1996), forexample, showed reduction of inhibitory circuitry with theloss of parvalbumin-immunoreactive chandelier cells in thehuman epileptogenic temporal neocortex, implicating animbalance of excitation and inhibition in the mechanism ofseizure generation. NPY, a signalling peptide with
anticonvulsant activity (Barabanet al., 1997; Woldbyeet al.,1997), is another useful marker for specific inhibitoryneuronal cell populations in the cortex. We therefore chose tostudy the GABAergic composition of a variety of heterotopicneuronal nodules using antibodies specific for calretininand NPY.
Circumstantial evidence suggests that heterotopic nodulesmay be connected to extranodular regions (Dubeauet al.,1995), allowing propagation of postulated intrinsic nodularepileptogenic activity. We therefore also attempted to examinethe connectivity of nodules using carbocyanine dye stainingtechniques. Carbocyanine dyes are fluorescent lipophilic dyes
by guest on June 17, 2016http://brain.oxfordjournals.org/
Dow
nloaded from
222 A. J. Hannanet al.
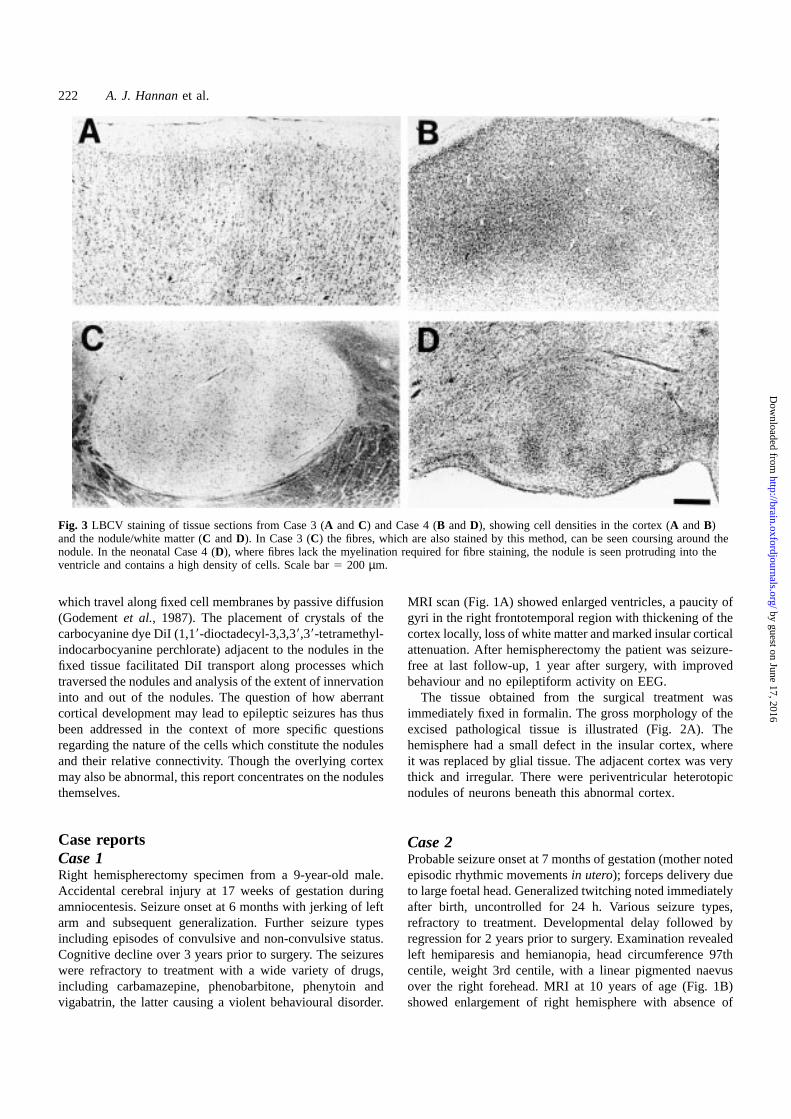
Fig. 3 LBCV staining of tissue sections from Case 3 (A andC) and Case 4 (B andD), showing cell densities in the cortex (A andB)and the nodule/white matter (C andD). In Case 3 (C) the fibres, which are also stained by this method, can be seen coursing around thenodule. In the neonatal Case 4 (D), where fibres lack the myelination required for fibre staining, the nodule is seen protruding into theventricle and contains a high density of cells. Scale bar5 200 µm.
which travel along fixed cell membranes by passive diffusion(Godementet al., 1987). The placement of crystals of thecarbocyanine dye DiI (1,19-dioctadecyl-3,3,39,39-tetramethyl-indocarbocyanine perchlorate) adjacent to the nodules in thefixed tissue facilitated DiI transport along processes whichtraversed the nodules and analysis of the extent of innervationinto and out of the nodules. The question of how aberrantcortical development may lead to epileptic seizures has thusbeen addressed in the context of more specific questionsregarding the nature of the cells which constitute the nodulesand their relative connectivity. Though the overlying cortexmay also be abnormal, this report concentrates on the nodulesthemselves.
Case reportsCase 1Right hemispherectomy specimen from a 9-year-old male.Accidental cerebral injury at 17 weeks of gestation duringamniocentesis. Seizure onset at 6 months with jerking of leftarm and subsequent generalization. Further seizure typesincluding episodes of convulsive and non-convulsive status.Cognitive decline over 3 years prior to surgery. The seizureswere refractory to treatment with a wide variety of drugs,including carbamazepine, phenobarbitone, phenytoin andvigabatrin, the latter causing a violent behavioural disorder.
MRI scan (Fig. 1A) showed enlarged ventricles, a paucity ofgyri in the right frontotemporal region with thickening of thecortex locally, loss of white matter and marked insular corticalattenuation. After hemispherectomy the patient was seizure-free at last follow-up, 1 year after surgery, with improvedbehaviour and no epileptiform activity on EEG.
The tissue obtained from the surgical treatment wasimmediately fixed in formalin. The gross morphology of theexcised pathological tissue is illustrated (Fig. 2A). Thehemisphere had a small defect in the insular cortex, whereit was replaced by glial tissue. The adjacent cortex was verythick and irregular. There were periventricular heterotopicnodules of neurons beneath this abnormal cortex.
Case 2Probable seizure onset at 7 months of gestation (mother notedepisodic rhythmic movementsin utero); forceps delivery dueto large foetal head. Generalized twitching noted immediatelyafter birth, uncontrolled for 24 h. Various seizure types,refractory to treatment. Developmental delay followed byregression for 2 years prior to surgery. Examination revealedleft hemiparesis and hemianopia, head circumference 97thcentile, weight 3rd centile, with a linear pigmented naevusover the right forehead. MRI at 10 years of age (Fig. 1B)showed enlargement of right hemisphere with absence of
by guest on June 17, 2016http://brain.oxfordjournals.org/
Dow
nloaded from

Neuronal heterotopia and epilepsy 223
detectable gyri. After right hemispherectomy there wasdevelopmental and behavioural improvement; at 12 monthsfollow-up the patient was seizure-free.
The specimen, obtained by surgical treatment at 10 yearsof age and immediately fixed in formalin, showed dilationof the lateral ventricle with reduction in volume of whitematter, which showed widespread spongy multicystic change.Several small heterotopic neuronal nodules were seen closeto the ventricular wall and subcortically. A representativearea of resected tissue from Case 2 is shown in Fig. 2B.
Case 3Left hemispherectomy specimen of a 6-year-old female withsevere intractable epilepsy, starting at 8 weeks of age,following a normal birth and delivery. Marked behaviouraldifficulties, mild right hemiparesis. Seizures refractory totreatment; vigabatrin led to non-convulsive status epilepticus.MRI brain scan revealed abnormality in the left frontal lobe,with abnormal agyric grey matter and absence of subcorticalwhite matter. She was seizure-free at 6-month follow-up.
The tissue obtained from the surgical treatment wasimmediately fixed in formalin. Examination of the fixedspecimen (Fig. 2C) revealed a large area of the frontal cortexto be thickened and irregular. There were numerous nodulesof heterotopic grey matter occupying almost the entire whitematter of the frontal lobe.
Case 4Dysmorphic term infant with hydrocephalus and poly-microgyria, who died at 5 days of age. This male infant wasborn by emergency caesarean section for foetal distress. Thebaby was hydrocephalic with bilateral cleft lip and palate,widely spaced nipples and micropenis. The brain, which wasremoved and formalin-fixed post-mortem, was found to beabnormal with grossly dilated lateral ventricles and anirregular cortical gyral pattern with multiple small gyri.Almost confluent nodules of heterotopic grey matter wereseen in the subependymal zone (Fig. 2D). Histology confirmedvirtually confluent subependymal neuronal heterotopia, poly-microgyria, agenesis of the corpus callosum and hydro-cephalus. There was aqueduct forking with a cluster ofnarrow irregular ependymal channels in the midbrain. Theinferior olives were malformed and appeared as thick C-shaped bands. There were thick bands of tissue containinggroups of neurons on the inferolateral surfaces of the medullain the site of the corpus pontobulbare. The cerebellum wasnormal. The central canal of the spinal cord was replaced bymultiple small channels. The appearances were of a complexseries of neuronal migration disorders involving cerebraland medullary migration pathways. Confluent subependymalheterotopia associated with polymicrogyria has beendescribed in three other males associated with an X-chromosome (Xq28) mutation (Finket al., 1997).
Control casesCortical tissues from a neonate and children aged 7 and 9years who had died from causes not related to neurologicaldiseases were used as a comparison for immunohisto-chemical staining.
MethodHistological analysisAs part of routine pathological diagnosis, representativeblocks of tissue were taken and 10µm paraffin-embeddedtissue sections cut. Sections were dewaxed and hydratedthrough descending changes of alcohol solutions, stainedwith haematoxylin and eosin as well as luxol fast blue andcresyl violet (LBCV), dehydrated and mounted. Sectionswere examined and bright-field photomicrographs preparedusing a light microscope (Leica, Wetzlar, Germany).
DiI analysisCrystals of DiI (Molecular Probes, Eugene, Oreg., USA)were placed in the formalin-fixed tissue blocks adjacent toor within the heterotopic nodules, using a thin stainless steelwire under an operating microscope. Tissues were incubatedat 37°C in 4% (w/v) paraformaldehyde in phosphate buffer(PB) for 6 months, or 3 months for Case 4 due to the youngerage with less myelination, allowing faster DiI membranetransport. Tissues were then embedded in 5% (w/v) agar and65–100µm sections cut on a vibratome (General Scientific,Redhill, UK). Sections were counterstained with bisbenzimide[10 min in 2.5 µg/ml solution in PBS (phosphate-bufferedsaline); Riedal-De Haen AG, Seelze-Hannover, Germany] toreveal chromatin in cell nuclei and thus the major boundariesand layers in the sections. Sections were coverslipped underHydromount (National Diagnostics). The extent of DiItransport along fibres was analysed by fluorescencemicroscopy (Leica). Colour photomicrographs were takenusing Kodak Ektachrome colour reversal film (400 ASA).
ImmunohistochemistryImmunohistochemistry was performed using antibodiesdirected against calretinin (rabbit polyclonal; SWant,Bellinzona, Switzerland) and NPY (rabbit polyclonal, Sigma-Aldrich, Poole, UK). Immunoreactivity in the heterotopiacases was compared with that of age-matched control cortexfrom a patient with no known neurological condition. Sectionsof fixed tissue were cut either on a vibratome (as describedfor DiI analysis) or a freezing microtome. For freezingmicrotome sectioning, tissues were immersed in 30% sucrosein PB overnight. Sectioned tissues (50µm) were rinsed intwo changes of PBS. Sections were blocked for 30 min with5% (v/v) normal goat serum, 0.3% Triton X-100 in PBS,and then incubated at 4°C overnight in primary antibodydiluted (1 : 5000 for calretinin and NPY) in 1% (v/v) normal
by guest on June 17, 2016http://brain.oxfordjournals.org/
Dow
nloaded from
224 A. J. Hannanet al.
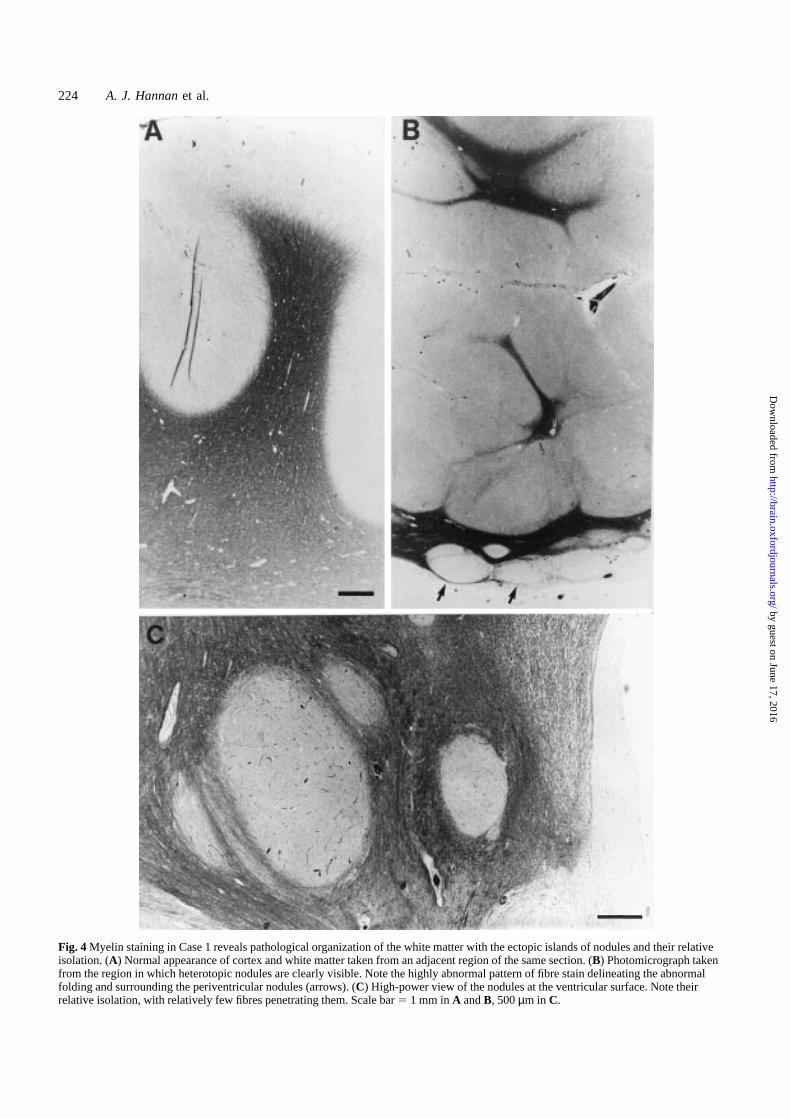
Fig. 4 Myelin staining in Case 1 reveals pathological organization of the white matter with the ectopic islands of nodules and their relativeisolation. (A) Normal appearance of cortex and white matter taken from an adjacent region of the same section. (B) Photomicrograph takenfrom the region in which heterotopic nodules are clearly visible. Note the highly abnormal pattern of fibre stain delineating the abnormalfolding and surrounding the periventricular nodules (arrows). (C) High-power view of the nodules at the ventricular surface. Note theirrelative isolation, with relatively few fibres penetrating them. Scale bar5 1 mm inA andB, 500µm in C.
by guest on June 17, 2016http://brain.oxfordjournals.org/
Dow
nloaded from
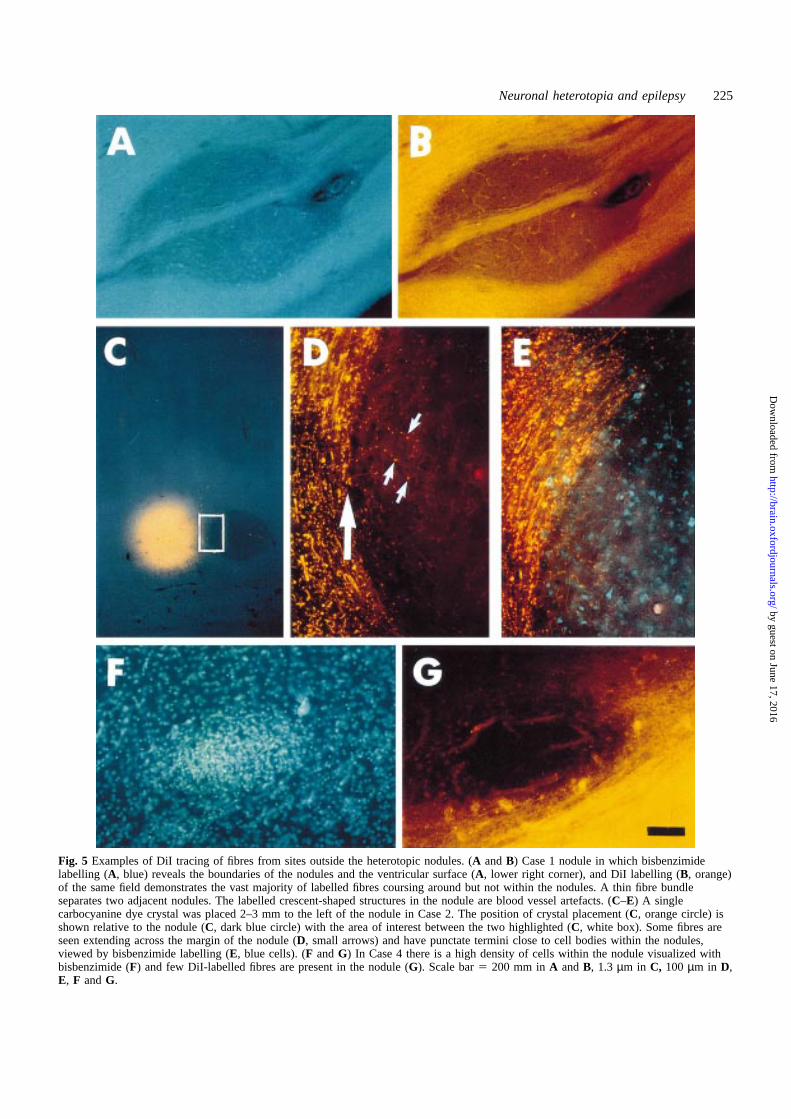
Neuronal heterotopia and epilepsy 225
Fig. 5 Examples of DiI tracing of fibres from sites outside the heterotopic nodules. (A andB) Case 1 nodule in which bisbenzimidelabelling (A, blue) reveals the boundaries of the nodules and the ventricular surface (A, lower right corner), and DiI labelling (B, orange)of the same field demonstrates the vast majority of labelled fibres coursing around but not within the nodules. A thin fibre bundleseparates two adjacent nodules. The labelled crescent-shaped structures in the nodule are blood vessel artefacts. (C–E) A singlecarbocyanine dye crystal was placed 2–3 mm to the left of the nodule in Case 2. The position of crystal placement (C, orange circle) isshown relative to the nodule (C, dark blue circle) with the area of interest between the two highlighted (C, white box). Some fibres areseen extending across the margin of the nodule (D, small arrows) and have punctate termini close to cell bodies within the nodules,viewed by bisbenzimide labelling (E, blue cells). (F andG) In Case 4 there is a high density of cells within the nodule visualized withbisbenzimide (F) and few DiI-labelled fibres are present in the nodule (G). Scale bar5 200 mm inA andB, 1.3 µm in C, 100 µm in D,E, F andG.
by guest on June 17, 2016http://brain.oxfordjournals.org/
Dow
nloaded from
226 A. J. Hannanet al.
by guest on June 17, 2016http://brain.oxfordjournals.org/
Dow
nloaded from

Neuronal heterotopia and epilepsy 227
goat serum, 0.3% Triton X-100 in PBS. Following threewashes in 0.3% Triton X-100 in PBS (10 min, roomtemperature) with shaking, sections were incubated withbiotin-conjugated secondary antibody (Vector Laboratories,Bretton, UK) at room temperature for 2 h with shaking.Sections were washed three times and then incubated withABC Elite avidin–biotin detection reagent (VectorLaboratories) for 1 h. Following a further three washes,colour detection was achieved by the addition of a solutioncontaining 0.5 mg/ml 3,39-diaminobenzidine (Sigma), 0.6%Ni(NH4) (SO4) and 0.03% H2O2. The colour reaction wasstopped with PBS and sections were dehydrated and mountedunder coverslips with DePex (Merck, Lutterworth, UK).
For control experiments, the primary antibody was replacedby 0.3% Triton X-100 in PBS and then reacted as above.These control sections showed no positive immunoreactivityfor any of the antibodies used. This protocol was also adaptedto stain paraffin wax sections (10µm) of the same tissues.Slides were dewaxed in xylene and hydrated throughdescending changes of alcohol solutions, and immunohisto-chemistry was performed as described above, with PBS aloneused as wash buffer. Bright-field photomicrographs wereprepared by light microscopy.
Morphological analysis of immunoreactiveneuronsCamera lucida drawings of immunoreactive neurons presentin the 50µm experimental sections were used to assess cellmorphology in different regions. Neurons were viewed athigh magnification (340 objective) using the light microscopeand drawn with the aid of the camera lucida attachment. Thefeatures of neuronal cell morphology that were specificallyassessed included cell size, process length and the numberand extent of dendritic branching.
ResultsHistological studiesHaematoxylin and eosin and LBCV histology was carriedout to examine the structure of the nodules and overlyingcortical plate. Heterotopic nodules in all cases consisted ofwell-defined masses of grey matter with randomly arrangedlarge and small neurons (Fig. 3). The cellular compositionof the nodules, distinct from the surrounding periventricularwhite matter, is clearly seen in Case 4 (Fig. 3D), in one regionof the almost confluent subependymal neuronal heterotopia.
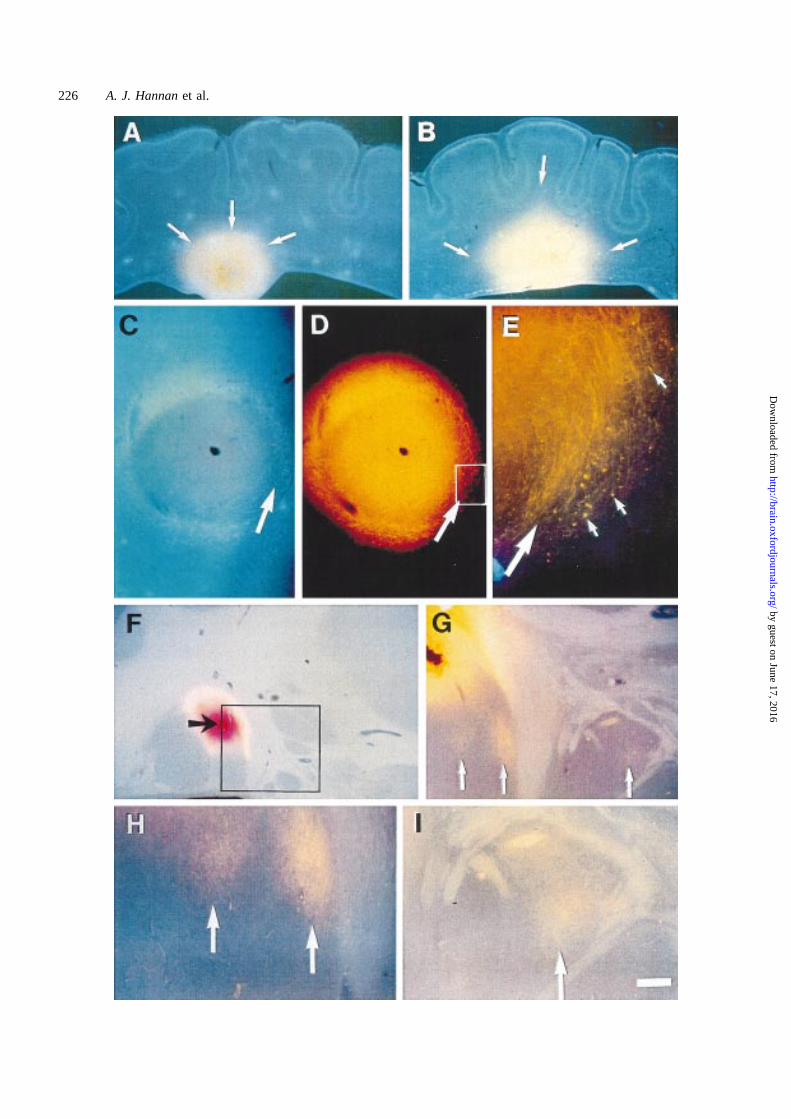
Fig. 6 DiI labelling of fibres from a single DiI crystal (orange) placed into the nodule and cells labelled for bisbenzimide (blue). (A andB) Case 4 with a large nodule close to the ventricular surface (A, arrows) very intensively labelled; few fibres are seen. There isrelatively little labelling in the surrounding tissue, in contrast with crystal placements directly into the white matter (B, arrows). (C–E) InCase 1, a nodule revealed by bisbenzimide staining (C) was labelled by DiI (D) to reveal fibres extending outside the boundary (arrow)of the nodule (E, enlarged view of boxed region inD). (F–I) In Case 3, DiI crystal placement in one of a cluster of nodules (F, arrow)labelled fibres within the nodule (G) but also connected fibres in adjacent nodules (enlarged inH and I, arrows). Scale bar5 1.3 mm inA, B andF, 400 µm in C, D andG, 100 µm in E; 200 µm in H and I.
In Case 1 (Fig. 4), staining in the white matter revealedaxons streaming around the nodules, without significantpenetration. The normal gross appearance of other parts ofthe cortex and white matter taken from the same tissuesection is illustrated (Fig. 4A). Histology of the malformedcortex showed cortical disorganization with malorientationof neurons, which were divided into irregular clusters bybands of glial cells and myelinated fibres. Neuronal and glialectopias were seen in the leptomeninges. These appearanceswere considered to be consistent with traumatic brain punctureat 17 weeks gestation and interruption of neuronal migrationand corticogenesis (Fig. 4B and C).
Similar patterns were observed in the other cases (Fig. 3and data not shown). The resected cortex from Case 2 wasthick with many small gyri. Histology showed a variety ofhistological abnormalities, including complete lack of normallamination. In some areas polymicrogyria was seen and therewere foci of meningeal glioneuronal heterotopia. Abnormalgiant dysplastic neurons were seen throughout the cortex andin the white matter. Fibre connections were seen for nodulesadjacent to the ventricular surface (Fig. 4C).
Histology from Case 3 showed the cortical malformationto have the characteristic features of polymicrogyria, withneurons forming a single band festooned beneath fusedsurface layers. The appearance was that of a focal corticaldysplasia with underlying neuronal heterotopia.
DiI tracing in and around the nodulesNeural connectivity between the nodules and theirenvironment was investigated using the technique ofcarbocyanine dye tracing, which permitted analysis of theextent of fibre projection into and out of the nodules. DiItracing of fibres from sites outside the heterotopic nodulescombined with bisbenzimide labelling (Fig. 5A) revealed theboundaries of the nodules and the ventricular surface. InCase 1, the vast majority of DiI-labelled fibres coursedaround but not within the nodules (Fig. 5B). A thin fibrebundle separated two adjacent nodules. This pattern of fibresskirting the nodule was also seen for Case 2 (Fig. 5C–E). Athigh magnification, some fibres in this region were seen topenetrate the nodule (Fig. 5D) and to have punctate terminals,possibly forming synapses, in the vicinity of cell bodieswithin the nodules (Fig. 5E). This pattern, in which a smallfraction of labelled fibres penetrated the nodules, was alsoobserved in Case 3 (data not shown). In Case 4 (Fig. 5F and
by guest on June 17, 2016http://brain.oxfordjournals.org/
Dow
nloaded from
228 A. J. Hannanet al.
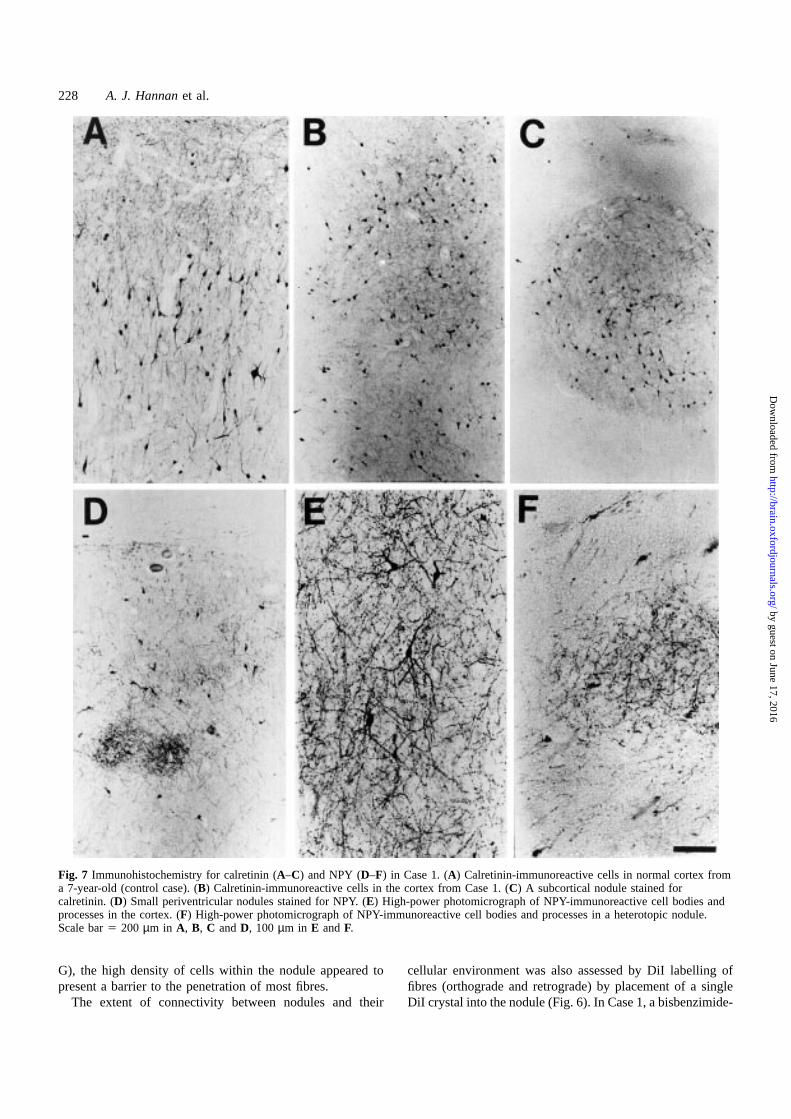
Fig. 7 Immunohistochemistry for calretinin (A–C) and NPY (D–F) in Case 1. (A) Calretinin-immunoreactive cells in normal cortex froma 7-year-old (control case). (B) Calretinin-immunoreactive cells in the cortex from Case 1. (C) A subcortical nodule stained forcalretinin. (D) Small periventricular nodules stained for NPY. (E) High-power photomicrograph of NPY-immunoreactive cell bodies andprocesses in the cortex. (F) High-power photomicrograph of NPY-immunoreactive cell bodies and processes in a heterotopic nodule.Scale bar5 200 µm in A, B, C andD, 100 µm in E andF.
G), the high density of cells within the nodule appeared topresent a barrier to the penetration of most fibres.
The extent of connectivity between nodules and their
cellular environment was also assessed by DiI labelling offibres (orthograde and retrograde) by placement of a singleDiI crystal into the nodule (Fig. 6). In Case 1, a bisbenzimide-
by guest on June 17, 2016http://brain.oxfordjournals.org/
Dow
nloaded from
Neuronal heterotopia and epilepsy 229
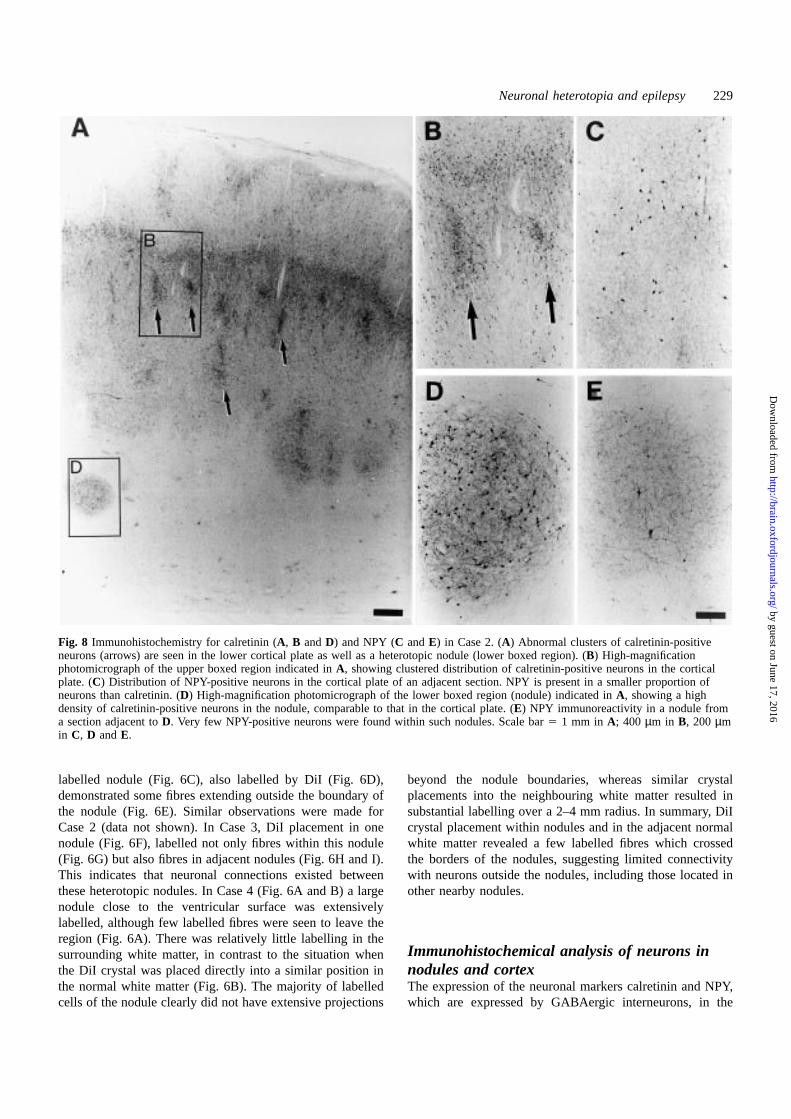
Fig. 8 Immunohistochemistry for calretinin (A, B andD) and NPY (C andE) in Case 2. (A) Abnormal clusters of calretinin-positiveneurons (arrows) are seen in the lower cortical plate as well as a heterotopic nodule (lower boxed region). (B) High-magnificationphotomicrograph of the upper boxed region indicated inA, showing clustered distribution of calretinin-positive neurons in the corticalplate. (C) Distribution of NPY-positive neurons in the cortical plate of an adjacent section. NPY is present in a smaller proportion ofneurons than calretinin. (D) High-magnification photomicrograph of the lower boxed region (nodule) indicated inA, showing a highdensity of calretinin-positive neurons in the nodule, comparable to that in the cortical plate. (E) NPY immunoreactivity in a nodule froma section adjacent toD. Very few NPY-positive neurons were found within such nodules. Scale bar5 1 mm in A; 400 µm in B, 200 µmin C, D andE.
labelled nodule (Fig. 6C), also labelled by DiI (Fig. 6D),demonstrated some fibres extending outside the boundary ofthe nodule (Fig. 6E). Similar observations were made forCase 2 (data not shown). In Case 3, DiI placement in onenodule (Fig. 6F), labelled not only fibres within this nodule(Fig. 6G) but also fibres in adjacent nodules (Fig. 6H and I).This indicates that neuronal connections existed betweenthese heterotopic nodules. In Case 4 (Fig. 6A and B) a largenodule close to the ventricular surface was extensivelylabelled, although few labelled fibres were seen to leave theregion (Fig. 6A). There was relatively little labelling in thesurrounding white matter, in contrast to the situation whenthe DiI crystal was placed directly into a similar position inthe normal white matter (Fig. 6B). The majority of labelledcells of the nodule clearly did not have extensive projections
beyond the nodule boundaries, whereas similar crystalplacements into the neighbouring white matter resulted insubstantial labelling over a 2–4 mm radius. In summary, DiIcrystal placement within nodules and in the adjacent normalwhite matter revealed a few labelled fibres which crossedthe borders of the nodules, suggesting limited connectivitywith neurons outside the nodules, including those located inother nearby nodules.
Immunohistochemical analysis of neurons innodules and cortexThe expression of the neuronal markers calretinin and NPY,which are expressed by GABAergic interneurons, in the
by guest on June 17, 2016http://brain.oxfordjournals.org/
Dow
nloaded from
230 A. J. Hannanet al.
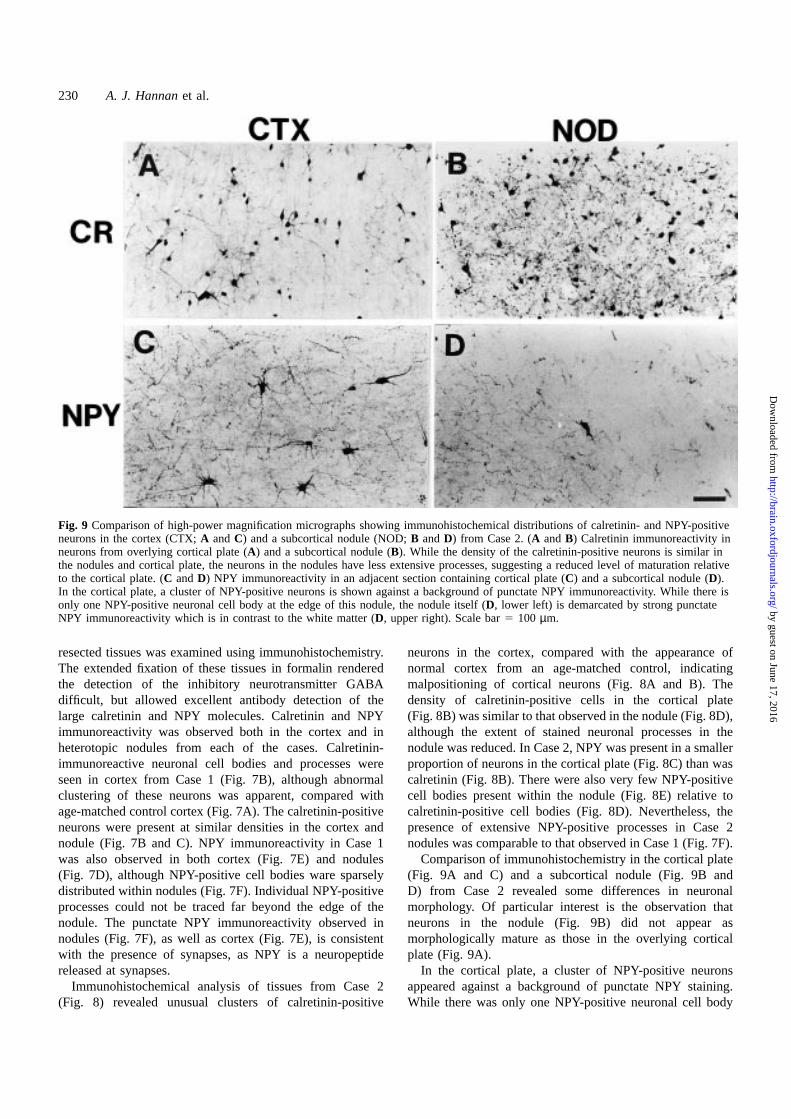
Fig. 9 Comparison of high-power magnification micrographs showing immunohistochemical distributions of calretinin- and NPY-positiveneurons in the cortex (CTX;A andC) and a subcortical nodule (NOD;B andD) from Case 2. (A andB) Calretinin immunoreactivity inneurons from overlying cortical plate (A) and a subcortical nodule (B). While the density of the calretinin-positive neurons is similar inthe nodules and cortical plate, the neurons in the nodules have less extensive processes, suggesting a reduced level of maturation relativeto the cortical plate. (C andD) NPY immunoreactivity in an adjacent section containing cortical plate (C) and a subcortical nodule (D).In the cortical plate, a cluster of NPY-positive neurons is shown against a background of punctate NPY immunoreactivity. While there isonly one NPY-positive neuronal cell body at the edge of this nodule, the nodule itself (D, lower left) is demarcated by strong punctateNPY immunoreactivity which is in contrast to the white matter (D, upper right). Scale bar5 100 µm.
resected tissues was examined using immunohistochemistry.The extended fixation of these tissues in formalin renderedthe detection of the inhibitory neurotransmitter GABAdifficult, but allowed excellent antibody detection of thelarge calretinin and NPY molecules. Calretinin and NPYimmunoreactivity was observed both in the cortex and inheterotopic nodules from each of the cases. Calretinin-immunoreactive neuronal cell bodies and processes wereseen in cortex from Case 1 (Fig. 7B), although abnormalclustering of these neurons was apparent, compared withage-matched control cortex (Fig. 7A). The calretinin-positiveneurons were present at similar densities in the cortex andnodule (Fig. 7B and C). NPY immunoreactivity in Case 1was also observed in both cortex (Fig. 7E) and nodules(Fig. 7D), although NPY-positive cell bodies ware sparselydistributed within nodules (Fig. 7F). Individual NPY-positiveprocesses could not be traced far beyond the edge of thenodule. The punctate NPY immunoreactivity observed innodules (Fig. 7F), as well as cortex (Fig. 7E), is consistentwith the presence of synapses, as NPY is a neuropeptidereleased at synapses.
Immunohistochemical analysis of tissues from Case 2(Fig. 8) revealed unusual clusters of calretinin-positive
neurons in the cortex, compared with the appearance ofnormal cortex from an age-matched control, indicatingmalpositioning of cortical neurons (Fig. 8A and B). Thedensity of calretinin-positive cells in the cortical plate(Fig. 8B) was similar to that observed in the nodule (Fig. 8D),although the extent of stained neuronal processes in thenodule was reduced. In Case 2, NPY was present in a smallerproportion of neurons in the cortical plate (Fig. 8C) than wascalretinin (Fig. 8B). There were also very few NPY-positivecell bodies present within the nodule (Fig. 8E) relative tocalretinin-positive cell bodies (Fig. 8D). Nevertheless, thepresence of extensive NPY-positive processes in Case 2nodules was comparable to that observed in Case 1 (Fig. 7F).
Comparison of immunohistochemistry in the cortical plate(Fig. 9A and C) and a subcortical nodule (Fig. 9B andD) from Case 2 revealed some differences in neuronalmorphology. Of particular interest is the observation thatneurons in the nodule (Fig. 9B) did not appear asmorphologically mature as those in the overlying corticalplate (Fig. 9A).
In the cortical plate, a cluster of NPY-positive neuronsappeared against a background of punctate NPY staining.While there was only one NPY-positive neuronal cell body
by guest on June 17, 2016http://brain.oxfordjournals.org/
Dow
nloaded from
Neuronal heterotopia and epilepsy 231
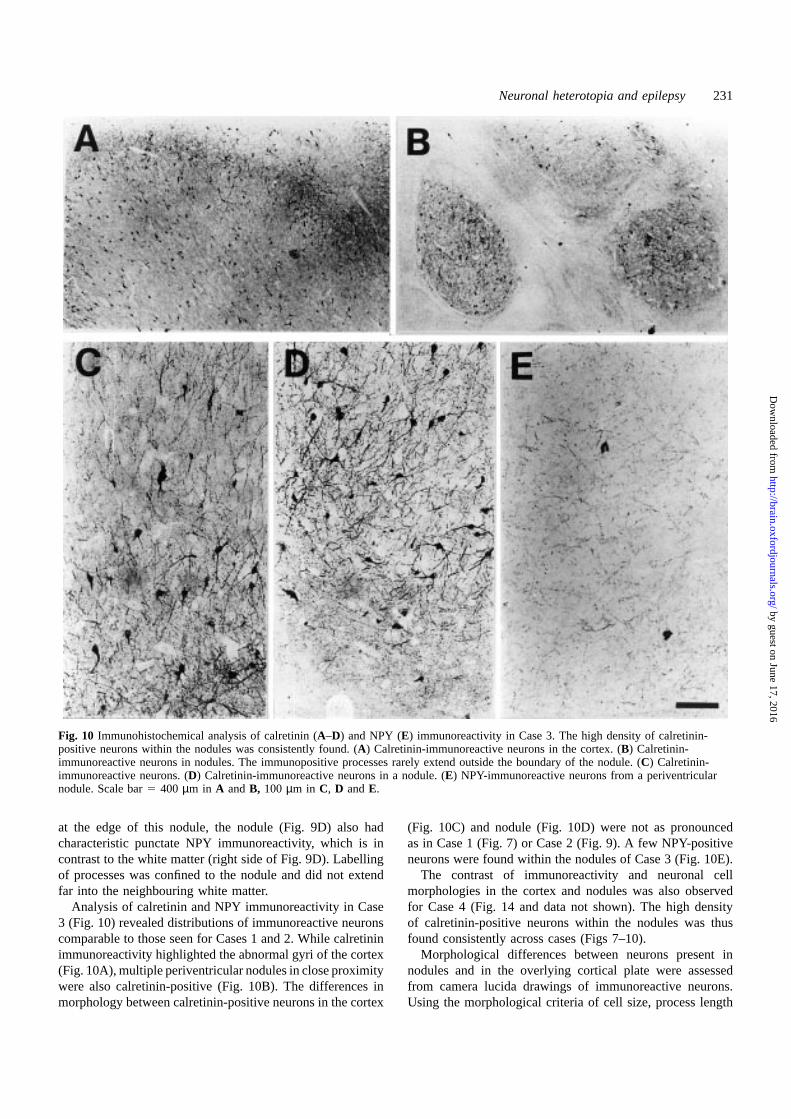
Fig. 10 Immunohistochemical analysis of calretinin (A–D) and NPY (E) immunoreactivity in Case 3. The high density of calretinin-positive neurons within the nodules was consistently found. (A) Calretinin-immunoreactive neurons in the cortex. (B) Calretinin-immunoreactive neurons in nodules. The immunopositive processes rarely extend outside the boundary of the nodule. (C) Calretinin-immunoreactive neurons. (D) Calretinin-immunoreactive neurons in a nodule. (E) NPY-immunoreactive neurons from a periventricularnodule. Scale bar5 400 µm in A and B, 100 µm in C, D andE.
at the edge of this nodule, the nodule (Fig. 9D) also hadcharacteristic punctate NPY immunoreactivity, which is incontrast to the white matter (right side of Fig. 9D). Labellingof processes was confined to the nodule and did not extendfar into the neighbouring white matter.
Analysis of calretinin and NPY immunoreactivity in Case3 (Fig. 10) revealed distributions of immunoreactive neuronscomparable to those seen for Cases 1 and 2. While calretininimmunoreactivity highlighted the abnormal gyri of the cortex(Fig. 10A), multiple periventricular nodules in close proximitywere also calretinin-positive (Fig. 10B). The differences inmorphology between calretinin-positive neurons in the cortex
(Fig. 10C) and nodule (Fig. 10D) were not as pronouncedas in Case 1 (Fig. 7) or Case 2 (Fig. 9). A few NPY-positiveneurons were found within the nodules of Case 3 (Fig. 10E).
The contrast of immunoreactivity and neuronal cellmorphologies in the cortex and nodules was also observedfor Case 4 (Fig. 14 and data not shown). The high densityof calretinin-positive neurons within the nodules was thusfound consistently across cases (Figs 7–10).
Morphological differences between neurons present innodules and in the overlying cortical plate were assessedfrom camera lucida drawings of immunoreactive neurons.Using the morphological criteria of cell size, process length
by guest on June 17, 2016http://brain.oxfordjournals.org/
Dow
nloaded from
232 A. J. Hannanet al.
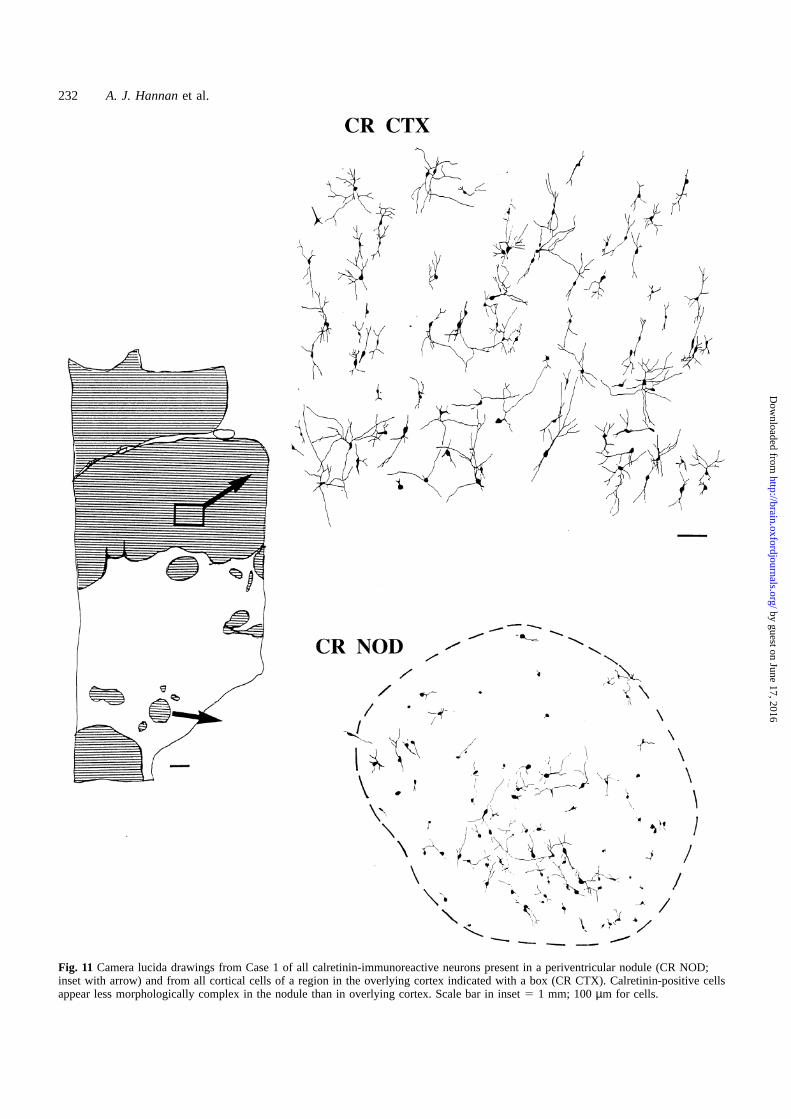
Fig. 11 Camera lucida drawings from Case 1 of all calretinin-immunoreactive neurons present in a periventricular nodule (CR NOD;inset with arrow) and from all cortical cells of a region in the overlying cortex indicated with a box (CR CTX). Calretinin-positive cellsappear less morphologically complex in the nodule than in overlying cortex. Scale bar in inset5 1 mm; 100µm for cells.
by guest on June 17, 2016http://brain.oxfordjournals.org/
Dow
nloaded from

Neuronal heterotopia and epilepsy 233
Fig. 12 Camera lucida drawings of NPY-immunoreactive neurons of Case 1 from nodules (NPY NOD), the overlying cortical plate (NPYCTX) and from the neighbouring white matter (NPY WM). Boxes in the inset indicate the site of origin of the cells drawn. Theexamples illustrate the differences in morphology. Scale bar in inset5 1 mm; 100µm for cells.
and the number of process branches, the calretinin-positiveneurons in the cortex were clearly more mature than thosewithin the nodule, as shown in Case 1 (Fig. 11). This relativeimmaturity was also seen when comparing NPY-positiveneurons in the cortex and nodule (Fig. 12). Interestingly,even the occasional white matter NPY neurons appearedmore mature than the nodular NPY cells (Fig. 12).
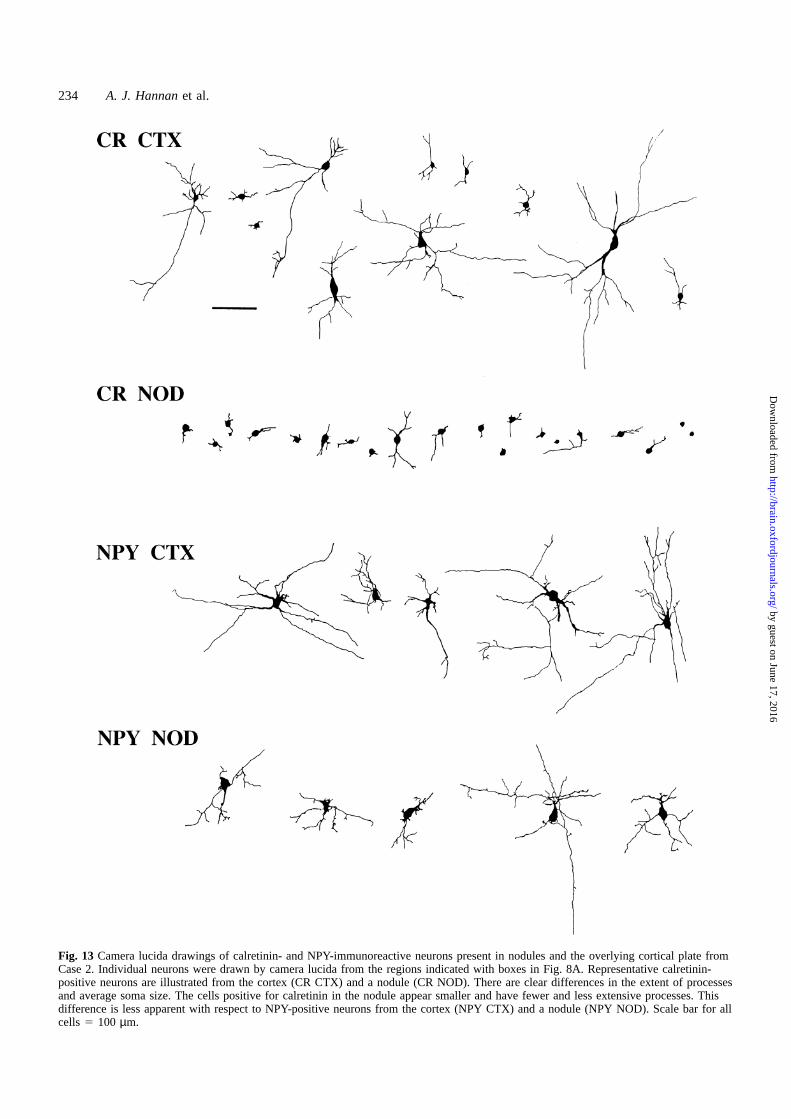
This difference in length and complexity of processes wasalso seen in Cases 2 and 3 (Figs 9A and C and 13), whencomparing calretinin-positive neurons of the cortex (Fig. 13,CR CTX) and the subcortical nodule (Fig. 13, CR NOD).The difference in neuronal morphology was still present,although less pronounced, when comparing NPY-positiveneurons from the cortex (Fig. 13, NPY CTX) and nodule(Fig. 13, NPY NOD).
The observations made in Cases 1–3 were similar to thosemade in Case 4, although, since this case was neonatal, therewas a somewhat different pattern of expression of thedevelopmentally regulated proteins calretinin and NPY.Immunoreactive neurons in the cortex of Case 4 did not have
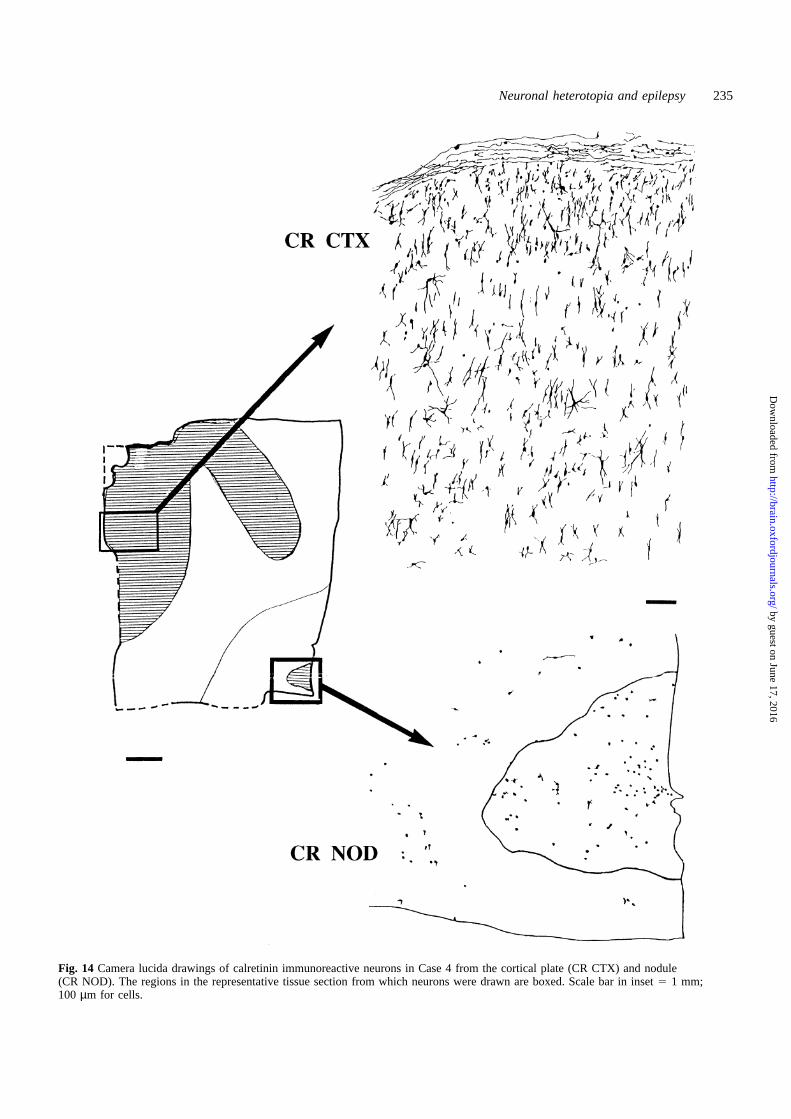
the same level of maturity as those in the cortex of the oldercases, which was not unexpected. Camera lucida drawingsof immunoreactive cells in Case 4 are shown (Fig. 14). Theseresults also demonstrate a disruption of the developmentalmaturation of neurons within the nodules, which appearedto have remained extremely immature with minimaldifferentiation.
In summary, immunohistochemical analysis of tissues fromthese cases of heterotopia reveals differences in neuronal cellmorphologies between the cortex and the underlying nodules.Of great interest is the observed reduction in the morpho-logical maturation of interneurons in the nodules relative tothat in the cortex.
DiscussionIt is widely accepted that cortical malformations can beintrinsically epileptogenic. There is clinical evidence for this,ranging from their association with epilepsy in imagingstudies (Li et al., 1995; Raymondet al., 1995), results of
by guest on June 17, 2016http://brain.oxfordjournals.org/
Dow
nloaded from
234 A. J. Hannanet al.
Fig. 13 Camera lucida drawings of calretinin- and NPY-immunoreactive neurons present in nodules and the overlying cortical plate fromCase 2. Individual neurons were drawn by camera lucida from the regions indicated with boxes in Fig. 8A. Representative calretinin-positive neurons are illustrated from the cortex (CR CTX) and a nodule (CR NOD). There are clear differences in the extent of processesand average soma size. The cells positive for calretinin in the nodule appear smaller and have fewer and less extensive processes. Thisdifference is less apparent with respect to NPY-positive neurons from the cortex (NPY CTX) and a nodule (NPY NOD). Scale bar for allcells 5 100 µm.
by guest on June 17, 2016http://brain.oxfordjournals.org/
Dow
nloaded from
Neuronal heterotopia and epilepsy 235
Fig. 14 Camera lucida drawings of calretinin immunoreactive neurons in Case 4 from the cortical plate (CR CTX) and nodule(CR NOD). The regions in the representative tissue section from which neurons were drawn are boxed. Scale bar in inset5 1 mm;100 µm for cells.
by guest on June 17, 2016http://brain.oxfordjournals.org/
Dow
nloaded from

236 A. J. Hannanet al.
direct electrical recording from abnormal tissue (Dubeauet al., 1995) and outcome after surgical resection (Palminiet al., 1995).In vitro studies of resected tissue and modelledlesions support these findings. For heterotopia in particular,there is much indirect evidence for their epileptogenicity.Heterotopic nodules contain neurons which are metabolicallyactive (Hatazawaet al., 1996; Marshet al., 1996) and maybe connected to extranodular targets that are appropriate,given the neurons’ likely birth dates (Jensen and Killackey,1984). Periventricular nodules are the most commondysgenetic findings in refractory epilepsy in adults (Raymondet al., 1995); both subcortical and periventricular noduleshave been reported very rarely in adult patients in the absenceof epilepsy or a family history of epilepsy. This circumstantialevidence is supported by the finding that surgical resectionleaving periventricular nodular tissue behind does not renderpatients seizure-free (Liet al., 1997), and by direct electricalrecordings from heterotopic nodules demonstratingindependent ictal activity in two cases (Francioneet al.,1994; Dubeauet al., 1995). Such reports are rare, since mostpatients do not undergo presurgical evaluation in view of therecognized poor seizure prognosis after surgery. The cellularbasis of this epileptogenicity, however, is not well understood.At the cellular level, we have shown a number ofabnormalities in heterotopic nodules and the overlying cortexthat may provide a preliminary structural basis forunderstanding their intrinsic epileptogenicity.
GABAergic neurons and an imbalance ofexcitatory and inhibitory mechanisms in epilepsyThere are clinical (Marcoet al., 1996; Spreaficoet al., 1998)and animal model (Buhlet al., 1996) studies which implicatedisorders of GABAergic interneurons in epilepsy. Despite thevarying macroscopic appearances from the four cases, wefound broadly similar cellular composition within the nodules.Nodules all contained GABAergic neuronal populations, asjudged by staining for calretinin and NPY. The GABAergicneurons appeared immature, consistent with the results of aproton magnetic resonance spectroscopy study whichsuggested increasedcellularactivityorpersistenceof immatureneurons in a case of grey matter heterotopia (Marshet al.,1996). During development, immature/early GABAergic cellsare known to be excitatory and later become inhibitory, viamodulation of GABAA receptor function (reviewed by Ben-Ari et al., 1997). Slowed or arrested development within thenodules, as shown by the morphological immaturity of thecells, might prevent this functional maturation of GABAergicsynapses. Excitatory pyramidal neurons, immunopositive forthe microtubule associated protein MAP2 have been identifiedwithin nodules in these cases (Mirkovicet al., 1998). Hence,the nodules themselves might have an overall excitatory bias.The refractory nature of epilepsy in Cases 1–3 could be due toimmaturity in the nodular neurotransmitter phenotypes(reviewed by Ben-Ariet al., 1997) affecting the moleculartargets of pharmaceutical intervention.
Aberrant activity of inhibitory GABAergic neurons in adultsmay be associated with epilepsy. It is thought that this is partof the epileptogenic mechanism in abnormal corticothalamicactivity in primary generalized epilepsy (Duncan andPanayiotopoulos, 1995). Vigabatrin is an antiepileptic drug thatinhibits GABA-transaminase and thus increases brain GABAlevels (Petroffet al., 1996). The immaturity of GABAergicneurons we have observed in nodules in these cases indicatesthat vigabatrin would actually increase excitation, through theaction of GABA as an excitatory transmitter in immatureneuronal networks. In this context the deterioration in clinicalcondition brought about by treatment with vigabatrin in Case1 is easily understood and mimics the aggravation of seizureactivity in primary generalized epilepsy that vigabatrin cancause.
Nodular connectivity and pathogenic implicationsOur examination of the connectivity of nodules showed thatthe nodules did have connections, although sparse, withextranodular regions. Some of this connectivity may have beenwith other adjacent nodules, as shown explicitly in one case,but the possibility of connection with the cortex cannot beexcluded as DiI labelling can only mark axons for a shortdistance. The relatively few NPY-positive neuronal cell bodieswithin the nodules and the presence of intranodular punctatestaining for NPY (which is known to be localized within axons,dendrites and synapses as well as cell bodies) suggests thatsome of the nodular NPY arose from the processes ofextranodular neuronal cell bodies, which would be consistentwith the fibre staining data. The small number of intranodularNPY-positive neuronal cell bodies is also of interest in the lightof the anticonvulsant activity of NPY (Barabanet al., 1997;Woldbyeet al., 1997). Thus, it is likely that the nodules havereciprocal connections, however sparse, with other parts of thehemisphere, including the cortex. This would allow postulatedfocal discharges arising in the nodules to spread to the cortex,which is known to be an excellent amplifier.
Cortical abnormalities associated with neuronalheterotopiaAlthough not the main focus of this study, our observationsthat there were abnormal clusters of neurons within the corticalplate itself raises the possibility that epileptic activity may alsobe due to abnormal neuronal distribution and connectivity inthe cortical plate. During brain development the defect whichleads to the formationofnodulesmayalso induceabnormalitiesof connection or cell type or number in the more normal-appearing, adjacent neocortex. It is possible that theseabnormalities could be responsible for, or contribute to, theepilepsy. In addition to these findings, quantitative MRIanalysis suggests that patients with apparently focal subcorticalheterotopia (Sisodiyaet al., 1995) and male patients withperiventricular heterotopia (unpublished results) also have
by guest on June 17, 2016http://brain.oxfordjournals.org/
Dow
nloaded from

Neuronal heterotopia and epilepsy 237
other, visually occult, structural abnormalities of the neocortex,raising the possibility of dual pathology.
In summary, we have shown that the abnormal immaturityof GABA networks within nodules could produce an excess ofexcitatory over inhibitory neuronal circuitry, thus inducingpathological activity. We have also identified connectionsbetween neurons in different nodules and shown thatintranodularneuronsareconnected toother regionsof thebrain,possibly including the cortex itself. Thus, intrinsic nodularepileptogenicity might be communicated to the cortex. Thecortex is a highly effective amplifier of synchronized activityand therefore few connections are required to set off cascadesof activity, including epileptiform discharge. Therefore wehave shown that the nodular tissue structure and compositionmight be structurally capable of acting as epileptogenicsubstrates, and that abnormal activity may be transmitted byconnections from the nodules. In addition, abnormalities ofcellular composition, as well as cytoarchitecture andinterneuronal connectivity, of the overlying cortex have alsobeen demonstrated. These changes may allow the cortex eitherto act independently as an epileptogenic substrate, providingfurtherhistological confirmationof thedual pathologyconcept,or to amplify and propagate the epileptiform activity generatedwithin the nodules themselves.
AcknowledgementsWe wish to thank Beat Ga¨hwiler, Carolyn Hannan, Anton vanDellen and Courtney Voelker for critical comments on themanuscript, and Laurence Waters and William Hinkes forphotographic assistance. This work is supported by the MRCand a Nuffield Medical Fellowship awarded to A.J.H.
ReferencesBaraban SC, Hollopeter G, Erickson JC, Schwartzkroin PA, PalmiterRD. Knock-out mice reveal a critical antiepileptic role forneuropeptide Y. J Neurosci 1997; 17: 8927–6.
Barkovich AJ, Kjos BO. Gray matter heterotopias: MR characteristicsand correlation with developmental and neurologic manifestations.Radiology 1992; 182: 493–9.
Barkovich AJ, Kuzniecky RI. Neuroimaging of focal malformationsof cortical development. [Review]. J. Clin Neurophysiol 1996; 13:481–94.
Battaglia G, Arcelli P, Granata T, Selvaggio M, Andermann F, DubeauF, et al. Neuronal migration disorders and epilepsy: a morphologicalanalysis of three surgically treated patients. Epilepsy Res 1996; 26:49–58.
Ben-Ari Y, Khazipov R, Leinekugel X, Caillard O, Gaiarsa J-L.GABAA, NMDA and AMPA receptors: a developmentally regulated‘menage a trois’. [Review]. Trends Neurosci 1997; 20: 523–9.
Buhl EH, Otis TS, Mody I. Zinc-induced collapse of augmentedinhibition by GABA in a temporal lobe epilepsy model. Science 1996;271: 369–73.
del Rio MR, DeFelipe J. Colocalization of calbindin D-28k, calretinin,
and GABA immunoreactivities in neurons of the human temporalcortex. J Comp Neurol 1996; 369: 472–82.
del Rio MR, DeFelipe J. Synaptic connections of calretinin-immunoreactive neurons in the human neocortex. J Neurosci 1997;17: 5143–54.
Dobyns WB, Andermann E, Andermann F, Czapansky-Beilman D,Dubeau F, Dulac O, et al. X-linked malformations of neuronalmigration. [Review]. Neurology 1996; 47: 331–9.
Dubeau F, Tampieri D, Lee N, Andermann E, Carpenter S, LeblancR, et al. Periventricular and subcortical nodular heterotopia: a studyof 33 patients. Brain 1995; 118: 1273–87.
Duncan JS, Panayiotopoulos CP, editors. Typical absences and relatedepileptic syndromes. London: Churchill Communications Europe;1995.
Duong T, De Rosa MJ, Poukens V, Vinters HV, Fisher RS. Neuronalcytoskeletal abnormalities in human cerebral cortical dysplasia. ActaNeuropathol (Berl) 1994; 87: 493–503.
Eksioglu YZ, Scheffer IE, Cardenas P, Knoll J, DiMario F, RamsbyG, et al. Periventricular heterotopia: an X-linked dominant epilepsylocus causing aberrant cerebral cortical development. Neuron 1996;16: 77–87.
Farrell MA, De Rosa MJ, Curran JG, Lenard Secor D, Cornford ME,Comair YG, et al. Neuropathologic findings in cortical resections(including hemispherectomies) performed for the treatment ofintractable childhood epilepsy. Acta Neuropathol (Berl) 1992; 83:246–59.
Ferrer I, Catala I. Unlayered polymicrogyria: structural anddevelopmental aspects. Anat Embryol (Berl) 1991; 184: 517–28.
Ferrer I, Pineda M, Tallada M, Oliver B, Russi A, Oller L, et al.Abnormal local-circuit neurons in epilepsia partialis continuaassociated with focal cortical dysplasia. Acta Neuropathol (Berl)1992; 83: 647–52.
Fink JM, Dobyns WB, Guerrini R, Hirsch BA. Identification of aduplication of Xq28 associated with bilateral periventricular nodularheterotopia. Am J Hum Genet 1997; 61: 379–87.
Flint AC, Kriegstein AR. Mechanisms underlying neuronal migrationdisorders and epilepsy. [Review]. Curr Opin Neurol 1997; 10: 92–7.
Francione S, Kahane P, Tassi L, Hoffmann D, Durisotti C, PasquierB, et al. Stereo-EEG of interictal and ictal electrical activity of ahistologically proved heterotopic gray matter associated with partialepilepsy. Electroencephalogr Clin Neurophysiol 1994; 90: 284–90.
Godement P, Vanselow J, Thanos S, Bonhoeffer F. A study indeveloping visual systems with a new method of staining neuronesand their processes in fixed tissue. Development 1987; 101: 697–713.
Hatazawa J, Sasajima T, Shimosegawa E, Fujita H, Okudera T, KannoI, et al. Regional cerebral blood flow response in gray matterheterotopia during finger tapping: an activation study with positronemission tomography. AJNR Am J Neuroradiol 1996; 17: 479–82.
Hendry SH, Jones EG, Emson PC, Lawson DE, Heizmann CW, StreitP. Two classes of cortical GABA neurons defined by differentialcalcium binding protein immunoreactivities. Exp Brain Res 1989; 76:467–72.
Hornung JP, De Tribolet N Chemical organization of the human
by guest on June 17, 2016http://brain.oxfordjournals.org/
Dow
nloaded from

238 A. J. Hannanet al.
cerebral cortex. In: Tracey DJ, Paxinos G, Stone J, editors.Neurotransmitters in the human brain. New York: Plenum Press; 1995.p. 41–5.
Hornung JP, De Tribolet N, Tork I. Morphology and distribution ofneuropeptide-containing neurons in human cerebral cortex.Neuroscience 1992; 51: 363–75.
Huttenlocher PR, Taravath S, Mojtahedi S. Periventricular heterotopiaand epilepsy. Neurology 1994; 44: 51–5.
Jensen KF, Killackey HP. Subcortical projections from ectopicneocortical neurons. Proc Natl Acad Sci USA 1984; 81: 964–8.
Kisvarday ZF, Gulyas A, Beroukas D, North JB, Chubb IW, SomogyiP. Synapses, axonal and dendritic patterns of GABA-immunoreactiveneurons in human cerebral cortex. [Review]. Brain 1990; 113: 793–812.
Kuzniecky RI. MRI in cerebral developmental malformations andepilepsy. [Review]. Magn Reson Imaging 1995; 13: 1137–45.
Lee N, Radtke RA, Gray L, Burger PC, Montine TJ, DeLong GR,et al. Neuronal migration disorders: positron emission tomographycorrelations. Ann Neurol 1994; 35: 290–7.
Li LM, Fish DR, Sisodiya SM, Shorvon SD, Alsanjari N, Stevens JM.High resolution magnetic resonance imaging in adults with partialor secondary generalised epilepsy attending a tertiary referral unit.J Neurol Neurosurg Psychiatry 1995; 59: 384–7.
Li LM, Dubeau F, Andermann F, Fish DR, Watson C, Cascino GD,et al. Periventricular nodular heterotopia and intractable temporal lobeepilepsy: poor outcome after temporal lobe resection. Ann Neurol1997; 41: 662–8.
Manz HJ, Phillips TM, Rowden G, McCullough DC. Unilateralmegalencephaly, cerebral cortical dysplasia, neuronal hypertrophy,and heterotopia: cytomorphometric, fluorometric cytochemical, andbiochemical analyses. Acta Neuropathol (Berl) 1979; 45: 97–103.
Marco P, Sola RG, Pulido P, Alijarde MT, Sanchez A, Ramon y CajalS, et al. Inhibitory neurons in the human epileptogenic temporalneocortex: an immunocytochemical study. Brain 1996; 119: 1327–47.
Marsh L, Lim KO, Sullivan EV, Lane B, Spielman D. Proton magneticresonance spectroscopy of a gray matter heterotopia. Neurology 1996;47: 1571–4.
Mattia D, Olivier A, Avoli M. Seizure-like discharges recorded inhuman dysplastic neocortex maintained in vitro. Neurology 1995; 45:1391–5.
Meencke HJ, Janz D. Neuropathological findings in primarygeneralized epilepsy: a study of eight cases. Epilepsia 1984; 25: 8–21.
Mirkovic N, Hannan AJ, Blakemore C, Streit P, Sisodiya S, SquierM, et al. Further characterization of neuronal heterotopia in childrenwith epilepsy. Suppl Euro J Neurosci 1998; 10: 50.
Musumeci SA, Ferri R, Elia M, Scuderi C, Del Gracco S, Azan G,et al. A new family with periventricular nodular heterotopia andpeculiar dysmorphic features: a probable X-linked dominant trait.Arch Neurol 1997; 54: 61–4.
Palmini A, Andermann F, Olivier A, Tampieri D, Robitaille Y,
Andermann E, et al. Focal neuronal migration disorders andintractable partial epilepsy: a study of 30 patients. [Review]. AnnNeurol 1991; 30: 741–9.
Palmini A, Andermann F, Tampieri D, Andermann E, Robitaille Y,Olivier A. Epilepsy and cortical cytoarchitectonic abnormalities: anattempt at correlating basic mechanisms with anatomoclinicalsyndromes. Epilepsy Res Suppl 1992; 9: 19–29.
Palmini A, Gambardella A, Andermann F, Dubeau F, da Costa JC,Olivier A, et al. Intrinsic epileptogenicity of human dysplastic cortexas suggested by corticography and surgical results. Ann Neurol 1995;37: 476–87.
Petroff OA, Rothman DL, Behar KL, Collins TL, Mattson RH. Humanbrain GABA levels rise rapidly after initiation of vigabatrin therapy.Neurology 1996; 47: 1567–71.
Raymond AA, Fish DR, Stevens JM, Sisodiya SM, Alsanjari N,ShorvonSD.Subependymalheterotopia:adistinctneuronalmigrationdisorder associated with epilepsy. [Review]. J Neurol NeurosurgPsychiatry 1994; 57: 1195–202.
Raymond AA, Fish DR, Sisodiya SM, Alsanjari N, Stevens JM,Shorvon SD. Abnormalities of gyration, heterotopias, tuberoussclerosis, focal cortical dysplasia, microdysgenesis, dysembryo-plastic neuroepithelial tumour and dysgenesis of the archicortex inepilepsy—clinical, EEG and neuroimaging features in 100 adultpatients. [Review]. Brain 1995; 118: 629–60.
SisodiyaSM,Free SL,StevensJM,FishDR, ShorvonSD.Widespreadcerebral structural changes in patients with cortical dysgenesis andepilepsy. Brain 1995; 118: 1039–50.
Sorvari H, Soininen H, Pitkanen A. Calretinin-immunoreactive cellsand fibers in the human amygdaloid complex. J Comp Neurol 1996;369: 188–208.
Spreafico R, Battaglia G, Arcelli P, Andermann F, Dubeau F, PalminiA, et al. Cortical dysplasia: an immunocytochemical study of threepatients [see comments]. Neurology 1998; 50: 27–36. Comment in:Neurology 1998; 50: 8–10.
Taylor DC, Falconer MA, Bruton CJ, Corsellis J. A. Focal dysplasiaof the cerebral cortex in epilepsy. J Neurol Neurosurg Psychiatry1971; 34: 369–87.
Woldbye DP, Larsen PJ, Mikkelsen JD, Klemp K, Madsen TM,Bolwig TG. Powerful inhibition of kainic acid seizures byneuropeptide Y via Y5-like receptors [see comments]. Nat Med 1997;3: 761–4. Comment in: Nat Med 1997; 3: 728–9.
Wolff JR, Bottcher H, Zetzsche T, Oertel WH, Chronwall BM.Development of GABAergic neurons in rat visual cortex as identifiedby glutamate decarboxylase-like immunoreactivity. Neurosci Lett1984; 47: 207–12.
Ying Z, Babb TL, Comair YG, Bingaman W, Bushey M, TouhaliskyK. Induced expression of NMDAR2 proteins and differentialexpression of NMDAR1 splice variants in dysplastic neurons ofhuman epileptic neocortex. J Neuropathol Exp Neurol 1998; 57:47–62.
Received August 3, 1998. Accepted September 28, 1998
by guest on June 17, 2016http://brain.oxfordjournals.org/
Dow
nloaded from
Copyright © 2022 FDOKUMEN




















