
Modelling, Distributed Identification and Control of Spatially ...
Upload
independentCategory
view
1download
0
PHYSICAL REVIEW E 69, 011909 ~2004!
Impact of spatially correlated noise on neuronal firing
Sentao Wang, Feng Liu,* and Wei WangNational Laboratory of Solid State Microstructure and Department of Physics, Nanjing University, Nanjing 210093, China
Yuguo YuCenter for the Neural Basis of Cognition, Carnegie Mellon University, Pittsburgh Pennsylvania 15213, USA
~Received 6 May 2003; revised manuscript received 30 September 2003; published 29 January 2004!
We explore the impact of spatially correlated noise on neuronal firing when uncoupled Hodgkin-Huxleymodel neurons are subjected to a common subthreshold signal. Noise can play a positive role in optimizingneuronal behavior. Although the output signal-to-noise ratio decreases with enhanced noise correlation, boththe degree of synchronization among neurons and the spike timing precision are improved. This suggests thatthere can exist precisely synchronized firings in the presence of correlated noise and that the nervous systemcan exploit temporal patterns of neural activity to convey more information than just using rate codes. Themechanisms underlying these noise-induced effects are also discussed in detail.
DOI: 10.1103/PhysRevE.69.011909 PACS number~s!: 87.16.Ac, 05.40.Ca
us
u
ceilnre
imt
pe-coke
ti
o
paoo
te
o-ainio
a
tooesrtiu
. Ittive
put--
ni-al
on-are
ethec-e-n-ingralto
eseaperII,III.
in-llel
fol-
I. INTRODUCTION
Cortical spike trains with high interspike interval~ISI!variability have been observed in a wide range of stimulevoked activity of pyramidal neurons@1#. Moreover, the neu-ral responses to repeated presentations of the same stimoften vary largely from trial to trial@2#. An issue thus arisesconcerning how the nervous system can precisely proinformation. One often assumes that the response variabrepresents ‘‘neural noise’’ and that the averaged respoover some populations of neurons may suppress the inhenoise and enhance the extracted information about the stlus. But such a hypothesis is based on the condition thatresponse variation in each neuron is more or less indedent of that in its neighbors@2,3#. However, it has been demonstrated that neural populations can exhibit significantvariance both in their spontaneous and stimulus-evoactivity @4,5#.
It has been suggested that, in the presence of correlain response variation, one possible function for neuronscarry redundant message may be to improve the tempresolution in coding a rapidly changing variable@6#. That is,temporal correlations are crucial for signal processing asof the information is encoded in the temporal structureactivity patterns. Furthermore, the synchronized activitiesneurons with high temporal precision can be transmitmore efficiently than the asynchronous ones@7#. It has alsobeen argued@8# that cortical neurons might act more as cincidence detectors preferentially relaying synchronizedtivity, than as temporal integrators effectively summatingcoming synaptic inputs. However, how the synchronizatand coincidence-detection mechanisms work in as noisyenvironment as a cortical circuit remains elusive.
It is well known that cortical neurons are subjectedlarge numbers of random synaptic inputs and other endenous noise. As a first approximation, we can model thtogether as Gaussian noise. But as mentioned above, coneurons display coherence in their firing activity, and th
*Corresponding author. Email address: [email protected]
1063-651X/2004/69~1!/011909~7!/$22.50 69 0119
-
lus
ssitysentu-
hen-
-d
ontoral
rtffd
c--nn
g-e
cals
the correlation in input noise must be taken into accounthas recently been shown that noise can play a construcrole in weak signal detection, such as improving the outsignal-to-noise ratio~SNR!, in the context of stochastic resonance~SR! @9#. While we have previously discussed the impact of spatially correlated noise on the output SNR@10#,here we mainly explore its influence on neuronal synchrozation and spike timing precision, as well as their biologicrelevance.
Motivated by the aforementioned considerations, we cstruct a network composed of uncoupled neurons whichsubjected to a common subthreshold~local field potential!signal s(t) plus spatially correlated noiseh(t). The neuralbehavior depends remarkably on both noise intensityD andthe measureR of noise correlation. On the one hand, thoutput SNR, the population coherence measure, andspike timing precision all go through a maximum as a funtion of D. On the other hand, the SNR monotonically dcreases with increasingR, whereas both the degree of sychronization among the neurons and the spike timprecision are improved. This makes it possible for neunetworks to exploit precise spatiotemporal firing patternsencode the stimulus. The mechanisms underlying thnoise-induced effects are also discussed in detail. This pis organized as follows. The model is described in Sec.while the results and discussion are presented in Sec.Finally, a conclusion is given in Sec. IV.
II. MODEL
We consider a summing network composed of HodgkHuxley ~HH! model neurons which are connected in paraand converge to a summing centerS, as shown in Fig. 1.The dynamic equations for the network are presented aslows @11#:
Cm
dVi
dt52gNa
mi3hi~Vi2ENa
!2gKni4~Vi2EK!
2gl~Vi2El !1I 01s~ t !1h i~ t !, ~1!
dmi
dt5am~Vi !~12mi !2bm~Vi !mi , ~2!
©2004 The American Physical Society09-1

f
tyna
ter-
r-tion,,
nsween
sn-
aons.
thefora
is
edeme
nd(al
ult,as
if-
inin-
ul-. Inutated
ityl
h a
h
ec-gnal, in
Ho
WANG et al. PHYSICAL REVIEW E 69, 011909 ~2004!
dhi
dt5ah~Vi !~12hi !2bh~Vi !hi , ~3!
dni
dt5an~Vi !~12ni !2bn~Vi !ni , i 51, . . . ,N. ~4!
Here Cm51 mF/cm2, ENa550 mV, EK5277 mV, El
5254.4 mV, gNa5120 mS/cm2, gK536 mS/cm2, gl
50.3 mS/cm2, and am(V)50.1(V140)/(12e2(V140)/10),bm(V)54e2(V165)/18, ah(V)50.07e2(V165)/20, bh(V)51/(11e2(V135)/10), an(V)50.01(V155)/(12e2(V155)/10),andbn(V)50.125e2(V165)/80. All the currents are in units omA/cm2.
I 0 is a constant bias and taken as 1mA/cm2. s(t) is asubthreshold signal,A cos(2pfst), corresponding to the inpugenerated by the local field potential. The signal frequencset to f s550 Hz unless specified otherwise, and the sigamplitude isA51 mA/cm2. Similar to that in Refs.@10,12#,the noise term is assumed to beh i(t)5A12R« i(t)1ARx(t) with
^« i~ t !&50, ^« i~ t1!« j~ t2!&52Dd i j d~ t12t2!, ~5!
and
^x~ t !&50, ^x~ t1!x~ t2!&5Dle2lut12t2u. ~6!
Here^ & represents the ensemble average andD is referred toas noise intensity.« i(t) is the independent Gaussian whinoise, whilex(t) is the Gaussian colored noise with the corelation timel21 being 2 ms. Since
^h i~ t1!h j~ t2!&52D~12R!d i j d~ t12t2!1RDle2lut12t2u,~7!
FIG. 1. A schematic diagram of the network composed of Hneurons. The total noise is divided into two items: the commnoiseARx(t) and the independent noiseA12R« i(t).
01190
isl
the control parameterR (0<R<1) measures the noise corelation between a pair of neurons. In the above assumpwe can consider that« i(t) represents the internal noisewhile x(t) reflects the random synaptic input from neurobeyond the system under study. This is plausible whenmodel cortical neurons in the same column. It has bestressed in Ref.@5# that common input, common stimuluselectivity, and common noise are tightly linked in functioing cortical circuits.
The output of the network is defined as
I out~ t !51
N (i 51
N
u„Vi~ t !2V* …. ~8!
V* is the firing threshold taken as220 mV, andu(x)51 ifx>0 andu(x)50 if x,0. Thus the summer (S) operatesby averaging the output spike train of each unit to obtainresultant output for the entire system. The number of neurin the network is set toN5500 unless specified otherwiseThe output SNR is defined as 10 log10(S/B) with S and Brepresenting the signal peak and the mean amplitude ofnoise at the input signal frequency in the power spectrumI out(t), respectively. Numerical integration is performed bysecond-order stochastic algorithm@13#, and the time step is500/32 768 ms. An average over 50 different realizationstaken to obtain reported results.
III. RESULTS AND DISCUSSION
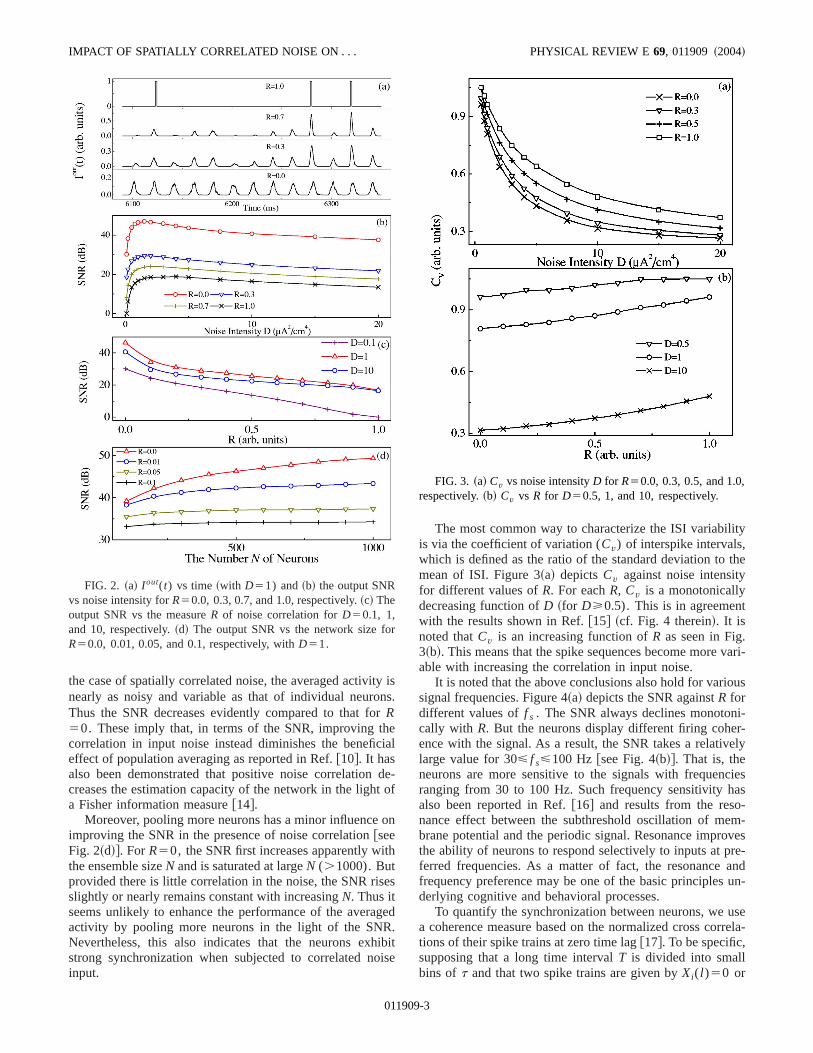
First, we investigate the influence of spatially correlatnoise on neuronal firing. ForR51, since all the neurons arsubjected to an identical input, they discharge at the satime but the firings exhibit skipping as seen in Fig. 2~a!. Thatis, I out(t) is composed of irregular sequences of ones azeros. In contrast, for the case of independent noiseR50), while one neuron is responding poorly to the signwithout spiking, others may be responding well. As a resI out(t) varies nearly periodically at the same frequencythe signal though its peak values are small. For 0,R,1,however,I out(t) exhibits apparent fluctuations between dferent driving cycles; that is,I out(t) is nearly zero in somedriving cycles, whereas it takes a relatively high valueothers. Such fluctuations become more remarkable withcreasingR. This means that more neurons tend to fire simtaneously as the correlation in input noise is enhancedother words, the neurons exhibit synchronous firing bmeanwhile the ISIs are more variable. It is noted thARx(t) has a dominant impact on neuronal firing comparto A12R« i(t) whenR.0.5.
Figure 2~b! depicts the output SNR against noise intensD for different values ofR. Each curve presents a typicacharacteristic of the SR, namely, the SNR goes througmaximum with increasingD. The optimal noise intensityslightly shifts rightward asR increases. Note that at eacnoise level the SNR is rigorously a decreasing function ofR.This is clearly shown in Fig. 2~c!. In fact, in the case ofindependent input noise, population averaging can efftively suppress the inherent noise, and thus the output sicontains more information about the stimulus. In contrast
n
9-2
tynr
hecia
dt o
o
th
e
g.ib
ois
ity
the
t
vari-
us
i-r-ely
ciesas-m-vesre-andun-
userrela-
r
IMPACT OF SPATIALLY CORRELATED NOISE ON . . . PHYSICAL REVIEW E69, 011909 ~2004!
the case of spatially correlated noise, the averaged activinearly as noisy and variable as that of individual neuroThus the SNR decreases evidently compared to that foR50. These imply that, in terms of the SNR, improving tcorrelation in input noise instead diminishes the benefieffect of population averaging as reported in Ref.@10#. It hasalso been demonstrated that positive noise correlationcreases the estimation capacity of the network in the ligha Fisher information measure@14#.
Moreover, pooling more neurons has a minor influenceimproving the SNR in the presence of noise correlation@seeFig. 2~d!#. ForR50, the SNR first increases apparently withe ensemble sizeN and is saturated at largeN (.1000). Butprovided there is little correlation in the noise, the SNR risslightly or nearly remains constant with increasingN. Thus itseems unlikely to enhance the performance of the averaactivity by pooling more neurons in the light of the SNRNevertheless, this also indicates that the neurons exhstrong synchronization when subjected to correlated ninput.
FIG. 2. ~a! I out(t) vs time~with D51) and~b! the output SNRvs noise intensity forR50.0, 0.3, 0.7, and 1.0, respectively.~c! Theoutput SNR vs the measureR of noise correlation forD50.1, 1,and 10, respectively.~d! The output SNR vs the network size foR50.0, 0.01, 0.05, and 0.1, respectively, withD51.
01190
iss.
l
e-f
n
s
ed
ite
The most common way to characterize the ISI variabilis via the coefficient of variation (Cv) of interspike intervals,which is defined as the ratio of the standard deviation tomean of ISI. Figure 3~a! depictsCv against noise intensityfor different values ofR. For eachR, Cv is a monotonicallydecreasing function ofD ~for D>0.5). This is in agreemenwith the results shown in Ref.@15# ~cf. Fig. 4 therein!. It isnoted thatCv is an increasing function ofR as seen in Fig.3~b!. This means that the spike sequences become moreable with increasing the correlation in input noise.
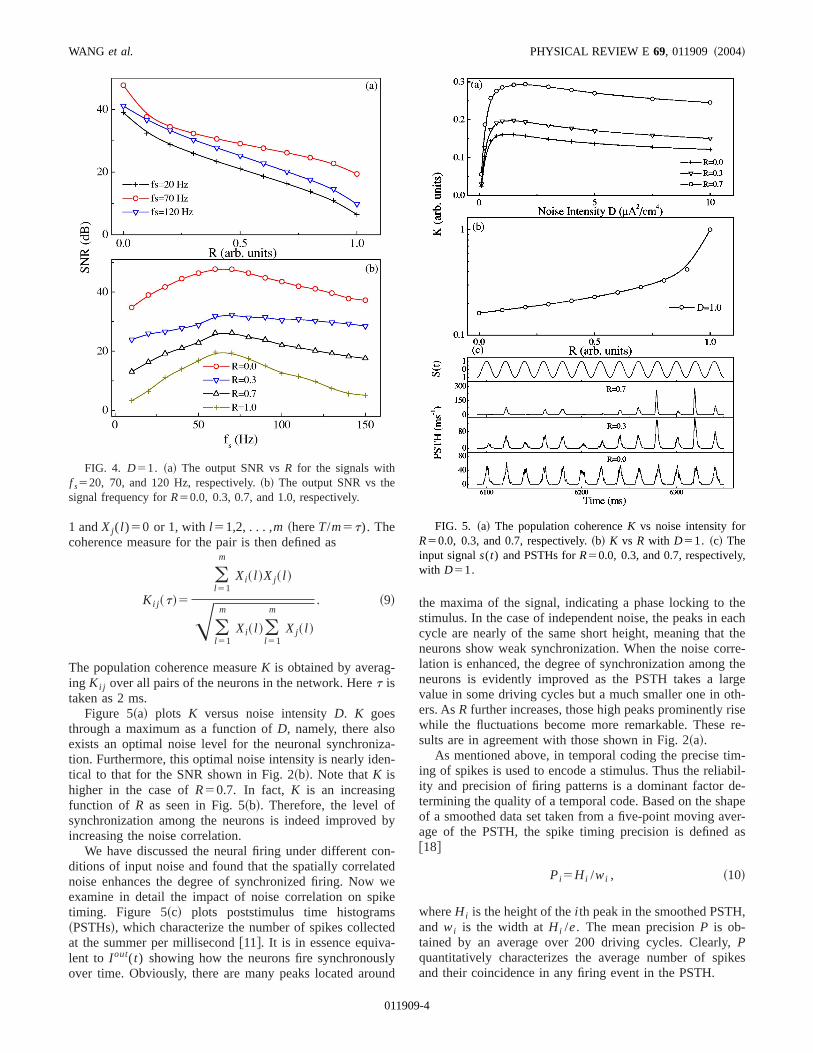
It is noted that the above conclusions also hold for variosignal frequencies. Figure 4~a! depicts the SNR againstR fordifferent values off s . The SNR always declines monotoncally with R. But the neurons display different firing coheence with the signal. As a result, the SNR takes a relativlarge value for 30< f s<100 Hz @see Fig. 4~b!#. That is, theneurons are more sensitive to the signals with frequenranging from 30 to 100 Hz. Such frequency sensitivity halso been reported in Ref.@16# and results from the resonance effect between the subthreshold oscillation of mebrane potential and the periodic signal. Resonance improthe ability of neurons to respond selectively to inputs at pferred frequencies. As a matter of fact, the resonancefrequency preference may be one of the basic principlesderlying cognitive and behavioral processes.
To quantify the synchronization between neurons, wea coherence measure based on the normalized cross cotions of their spike trains at zero time lag@17#. To be specific,supposing that a long time intervalT is divided into smallbins of t and that two spike trains are given byXi( l )50 or
FIG. 3. ~a! Cv vs noise intensityD for R50.0, 0.3, 0.5, and 1.0,respectively.~b! Cv vs R for D50.5, 1, and 10, respectively.
9-3
-
zan
b
ontew
ikste-lyun
theachtherre-thergeth-isere-
im-bil-e-apeer-as
,
ikes
,
WANG et al. PHYSICAL REVIEW E 69, 011909 ~2004!
1 andXj ( l )50 or 1, with l 51,2, . . . ,m ~hereT/m5t). Thecoherence measure for the pair is then defined as
Ki j ~t!5
(l 51
m
Xi~ l !Xj~ l !
A(l 51
m
Xi~ l !(l 51
m
Xj~ l !
. ~9!
The population coherence measureK is obtained by averaging Ki j over all pairs of the neurons in the network. Heret istaken as 2 ms.
Figure 5~a! plots K versus noise intensityD. K goesthrough a maximum as a function ofD, namely, there alsoexists an optimal noise level for the neuronal synchronition. Furthermore, this optimal noise intensity is nearly idetical to that for the SNR shown in Fig. 2~b!. Note thatK ishigher in the case ofR50.7. In fact, K is an increasingfunction of R as seen in Fig. 5~b!. Therefore, the level ofsynchronization among the neurons is indeed improvedincreasing the noise correlation.
We have discussed the neural firing under different cditions of input noise and found that the spatially correlanoise enhances the degree of synchronized firing. Nowexamine in detail the impact of noise correlation on sptiming. Figure 5~c! plots poststimulus time histogram~PSTHs!, which characterize the number of spikes collecat the summer per millisecond@11#. It is in essence equivalent to I out(t) showing how the neurons fire synchronousover time. Obviously, there are many peaks located aro
FIG. 4. D51. ~a! The output SNR vsR for the signals withf s520, 70, and 120 Hz, respectively.~b! The output SNR vs thesignal frequency forR50.0, 0.3, 0.7, and 1.0, respectively.
01190
--
y
-de
e
d
d
the maxima of the signal, indicating a phase locking tostimulus. In the case of independent noise, the peaks in ecycle are nearly of the same short height, meaning thatneurons show weak synchronization. When the noise colation is enhanced, the degree of synchronization amongneurons is evidently improved as the PSTH takes a lavalue in some driving cycles but a much smaller one in oers. AsR further increases, those high peaks prominently rwhile the fluctuations become more remarkable. Thesesults are in agreement with those shown in Fig. 2~a!.
As mentioned above, in temporal coding the precise ting of spikes is used to encode a stimulus. Thus the reliaity and precision of firing patterns is a dominant factor dtermining the quality of a temporal code. Based on the shof a smoothed data set taken from a five-point moving avage of the PSTH, the spike timing precision is defined@18#
Pi5Hi /wi , ~10!
whereHi is the height of thei th peak in the smoothed PSTHand wi is the width atHi /e. The mean precisionP is ob-tained by an average over 200 driving cycles. Clearly,Pquantitatively characterizes the average number of spand their coincidence in any firing event in the PSTH.
FIG. 5. ~a! The population coherenceK vs noise intensity forR50.0, 0.3, and 0.7, respectively.~b! K vs R with D51. ~c! Theinput signals(t) and PSTHs forR50.0, 0.3, and 0.7, respectivelywith D51.
9-4

isimthde
t-
seroseset
fi
sre
-rises
onnsic
of
ngtion
e
mitry
xed
n
tableclesimit
for
IMPACT OF SPATIALLY CORRELATED NOISE ON . . . PHYSICAL REVIEW E69, 011909 ~2004!
Figure 6~a! showsP versus noise intensityD for variousvalues ofR. Each curve displays a SR-like behavior. Thatthere exists an optimal noise level which maximizes the ting precision via the SR mechanism. This indicates thatspike timing precision in response to subthreshold periostimuli can be enhanced by input noise, as reported in R@18#. The optimal noise intensity also slightly shifts righward with increasingR. Moreover, for eachR the maximumSNR, K, and P occur at the same noise intensity. Theverify that noise can play a positive role in weak signal pcessing. The timing precision also monotonically increawith R @see Fig. 6~b!#, which means that the correlated noidoes make the spike timing more precise. It is noted thaPincreases more steeply whenR.0.5 since ARx(t), thedominant part of the noise, makes the neurons prone tosynchronously. The inset of Fig. 6~b! depictsP against thesignal frequency forD51. Clearly,P takes a relatively largevalue for 50< f s<90 Hz. This indicates that the neurontransmit these signals with a high precision and respond perentially to them.
FIG. 6. ~a! The spike timing precisionP vs noise intensity forR50.0, 0.3, and 0.7, respectively.~b! P vs R for D50.5 and 1.0,respectively. The inset isP vs the signal frequencyf s for R50.0and 0.3, respectively, withD51. ~c! P vs the network size forR50.0, 0.3, and 0.7, respectively, withD51.
01190
,-eicf.
-s
re
f-
It is worth noting that the timing precision linearly increases with the number of neurons and that the slopewith increasingR, which is clearly seen in Fig. 6~c!. This islargely different from the dependence of the output SNRN shown in Fig. 2~d!. Here we see that pooling more neurois of functional significance in effectively firing postsynaptneurons, and this effect is more prominent in the casecorrelated noise.
It is of interest to investigate the mechanism underlyithese noise-induced effects. We first discuss the bifurcain the HH neuron to a constant biasI 0 in the absence of noise(D50) and input signal (A50). As seen in Fig. 7~a!, forI 0,I c56.2 there is only a globally stable fixed point. Thbirth of stable and unstable limit cycles occurs atI c due tothe saddle-node bifurcation. ForI c,I 0,I h59.8, there exista stable fixed point, a stable limit cycle, and an unstable licycle. The unstable limit cycle constitutes the boundaseparating the attractive basins corresponding to the fi
FIG. 7. ~a! Deterministic bifurcation diagram of a HH neurounder dc current input. HereI 0 is the bifurcation parameter andV isthe membrane potential. The thick and dashed lines represent sand unstable fixed points, respectively. The filled and open cirrepresent both maxima and minima of stable and unstable lcycles, respectively. Noise-induced bifurcation diagram forR50~b! and R51 ~c!. The curves represent the upper (d) and lower(3) bounds of the stationary distribution of membrane potentialeach noise intensityD.
9-5

dy
thaiz
ly
dit
th
ry
teon
ire
.
h
aynneuik
edetenafi
seine
ndvfirr
tefirrageandb
ro-is
ofthis
onherm
on-in aons
an-ity,
. Inise-ig-bedion
r afireise
s thero-ntingan
s ay-on
ot
-ngmal
forme
hro-ur
ticsthatin-omerge-y of
rnvityym.ike
th
WANG et al. PHYSICAL REVIEW E 69, 011909 ~2004!
point and limit cycle. When the initial conditionV0 fallsinside this boundary, i.e., in the contraction region, thenamics of system will be attracted to the point attractor. IfV0
is outside the boundary, i.e., in the expansion region,system will be attracted to the stable limit cycle. As we shsee, this feature has a large impact on neuronal synchrontion. At I 05I h the Hopf bifurcation occurs and there is ona stable limit cycle thereafter.
In the presence of noise, the neurons can be evokedischarge spikes, and there exists a noise-induced transin excitability with increasingD @19#. To quantify such atransition, we compute stochastic bifurcation diagram insame way as in Refs.@19,20#. Figures 7~b! and 7~c! show thenoise-evoked transition diagrams forR50 and R51, re-spectively, providing a global view of how the stationadistributionV99 of membrane potential changes withD. Foreach fixedD, output membrane potential has been collecfor 100 s. Then the top and bottom limits of the distributiare computed in the way so that 99% of the distributionbelow the upper line and 99% is above the lower one. Thregions can be distinguished. ForR50, the distribution in-creases linearly for low noise (D<0.5). As noise intensityincreases (0.5,D<2), the distribution evidently widensThat is, a transition occurs aroundD50.5. WhenD is fur-ther increased, the distribution changes slightly. ForR51,however, the bifurcation point is shifted rightwards and tsecond region also broadens. Tanabeet al. @19# demonstratedthat noise with intensities prior to the bifurcation point mplay an important role in enhancing spike timing precisioHere we further show that the correlation of noise widethis beneficial region, which can largely enhance the nronal synchronization and lead to high variability of the sping dynamics.
Comparing Figs. 2~b!, 5~a! and 6~a! with Figs. 7~b! and7~c!, we see that the SNR,K, and P are in step with thedistribution of membrane potential varying withD. Thus wecan interpret their dependence onD and R in terms of theexcitability of neurons. In the first regime, noise-inducfluctuations improve the excitability of some units in thensemble, while their membrane potentials are locaaround the resting potential. In the presence of input sigthose neurons which are more excitable than at rest maysynchronously in some driving cycles. As noise is increawithin this range, more neurons evolve closer to the firthreshold, and enhanced response is observed such as thof the SNR andP. As the noise is further increased beyothe first regime, the membrane potential fluctuations edently increase. A fraction of neurons that are evoked toby noise alone may not respond to the input due to thefractory period, while other neurons whose membrane potials are around the resting potential may be triggered tosimultaneously by the signal. This may reduce the overesponse of the ensemble. In the third regime with larnoise intensity, the noise fluctuations become dominant,the neural coherence further decreases. Here the bounbetween attractive basins, which is related to the unstalimit cycle, plays a crucial role in the noise-induced synchnization, as reported in Ref.@21# wherein a saddle point embedded in system dynamics is responsible for the no
01190
-
ella-
toion
e
d
se
e
.s-
-
dl,red
grise
i-ee-n-ellrd
aryle-
e-
evoked synchronization. It is worth noting that in the casecorrelated noise, the first region evidently expands, andgives rise to a prominent increment inP andK with increas-ing D.
Therefore, the correlated noise has two primary effectsneuronal firing. The first is related to noise intensity. In tabsence of input signal, small noise disturbs the long-temotion of the system and results in the dominance of ctraction dynamics. The membrane potential is distributednarrow region. In this case, the noise can trigger the neurto respond synchronously to a weak signal, leading tohigher spike timing precision when noise intensity is icreased up to the transition point. For large noise intensthe membrane potential distribution is largely broadenedthis case, when the weak signal is input to the system, noinduced firings become dominant while the effect of the snal on driving the ensemble in phase has been disturheavily. This leads to a decrease of spike timing preciswith increasingD.
The second effect of noise is due to its correlation. Fofixed noise intensity, neurons with independent noise canspikes more independently, whereas correlation in nomakes the neurons prone to act together, which increaseinertia of ensemble neurons to be rest or to fire synchnously. That is, the correlation drives neurons with differeinitial conditions to converge to an identical response, beinside the contraction or expansion region. This results inenhanced neural synchronization. Periodic signal playsimilar synergic role to correlation in driving the neural dnamics in phase, improving the level of synchronizatiamong neurons.
Finally, it is worth noting that the correlation of noise nonly enhances spike timing precision@see Fig. 6~a!#, but alsoenlarges the spiking variability@see Fig. 3~b!#. The incrementin ISI variability will improve the encoding capacity of neurons. This is of functional significance when considerihow the nervous system tunes noise intensity to its optivalues. Our results may give a reasonable mechanismwhy neural responses in the cerebral cortex often becohighly variable but precise in encoding input signals@1#.
IV. CONCLUSION
We have demonstrated that there exists precisely syncnized activity in the presence of strong noise correlation. Oresults reproduce some of the typical firing characterisobserved in cortical neurons. First, it is generally agreedthe response variability originates in unreliable synapticputs @22#. We also showed that the spike sequences becmore variable in response to correlated noise. Second, lascale synchronized firings have been observed in a varietbrain areas, especially theg oscillations~at frequencies of30–70 Hz! @23#, which play functional roles such as pattesegmentation and feature binding. The frequency sensitishown in Figs. 4 and 6~b! may give us an enlightenment whthe g oscillations are so ubiquitous in the nervous systeThird, it has been found that under some conditions the sptiming can show a high precision and reproducibility withe temporal resolution being 2–3 ms@24#. Thus precise
9-6

m
-
thucinacyteonro
aibto
eto
ticcinvira
pereig
es
odicutsen-
ingeu-
lli-ork
and
actanto
ncetionan-ngsedu-ultsim-yn-
eaxver
ci-
IMPACT OF SPATIALLY CORRELATED NOISE ON . . . PHYSICAL REVIEW E69, 011909 ~2004!
temporal firing patterns can be exploited to encode the stilus.
As shown in Fig. 2~a!, although the neurons exhibit synchronized firings whenR50.7, they miss firing in manydriving cycles. But this does not necessarily mean thatneurons are poorly processing information. In contrast, ssynchronized activity may subserve information processFor example,s(t) may just represent a modulation of neurbehavior providing the system with an effect of frequenselection@16#, and the neurons may preferentially respondsynchronized synaptic inputs. Alternatively, the neural nworks can exploit precise temporal relations among neurto select responses for joint processing and to bind neutemporally into functionally coherent assemblies@7#. Fur-thermore, the combination of synchronized firing of corticneurons and high temporal precision also makes it possfor their downstream neurons to act as coincidence detecpreferentially transferring synchronized activity@8#. For in-stance, a coincidence-detection neuron can precisely dmine whether two coupled neurons receive similar levelsensory input@25#.
Although we did not directly model the random synapinput, our results suggest that the noise correlation is crufor cortical neurons to temporally process information athat the cerebral cortex may convey more informationtemporal codes than exclusively using rate codes. In contit seems plausible to assume that sensory neurons in theripheral nervous system are subject to more or less indedent noise. In that case, it is the averaging of neuralsponses that encodes a stimulus feature such as the sfrequency.
In addition, it is noted that Rudolph and Destexhe inv
u-
u
v.
01190
u-
eh
g.l
ot-s
ns
llers
er-f
aldast,pe-n--nal
-
tigated the case wherein a neuron is subjected to a periweak signal plus large numbers of random synaptic inp@26#. They reported that neuronal response can also behanced by the correlation among synaptic inputs, exhibita SR-like behavior. Their results suggested that cortical nrons are efficient in detecting such correlations within misecond time scales. This is consistent with the present wthat neurons may exploit correlated noise to encodetransmit information.
In conclusion, in this paper we have explored the impof spatially correlated noise on neuronal firing. Noise cplay a constructive role in optimizing neuronal responsesubthreshold stimuli. The results illustrate how the preseof correlated noise improves the degree of synchronizaamong the neurons and the spike timing precision but mewhile makes output spikes more variable. With a high timiresolution, temporal structures of neural activity can be uto convey more information. Thus correlated firings of nerons play functional roles in signal processing. These resare consistent with observations in cortical neurons andply that we should consider the correlation in noise or saptic input when we model the cortical dynamics. Finally, wwould like to point out that the present form of noise issimplification, and it is of interest to exploit more compleconfigurations, such as the spatially decaying functions othe population.
ACKNOWLEDGMENTS
This work was supported by the National Natural Sences Foundation of China~under Grant No. 30070208! andthe Nonlinear Science Project of NSM.
v.
,
@1# W. Softky and K. Koch, J. Neurosci.13, 334 ~1993!; M.N.Shadlen and W.T. Newsome,ibid. 18, 3870~1998!.
@2# D. Ferster, Science273, 1812~1996!.@3# E. Zohary, M.N. Shadlen, and W.T. Newsome, Nature~Lon-
don! 370, 140~1994!; M.N. Shadlen, K.H. Britten, W.T. New-some, and J.A. Movshon, J. Neurosci.16, 1486~1996!.
@4# A. Arieli, D. Shoham, R. Hildesheim, and A. Grinvald, J. Nerophysiol.73, 2072~1995!; A. Arieli, A. Sterkin, A. Grinvald,and A. Aertsen, Science273, 1868~1996!.
@5# W. Bair, E. Zohary, and W.T. Newsome, J. Neurosci.21, 1676~2001!.
@6# D. Lee, N.L. Port, W. Kruse, and A.P. Georgopoulos, J. Nerosci.18, 1161~1998!.
@7# W. Singer, Curr. Opin. Neurobiol.9, 189 ~1999!.@8# P. Konig, A.K. Engel, and W. Singer, Trends Neurosci.19, 130
~1996!.@9# L. Gammaitoni, P. Ha¨nggi, P. Jung, and F. Marchesoni, Re
Mod. Phys.70, 223 ~1998!.@10# F. Liu, B. Hu, and W. Wang, Phys. Rev. E63, 031907~2001!.@11# Y. Yu, F. Liu, J. Wang, and W. Wang, Phys. Lett. A282, 23
~2001!.@12# C. Zhou, J. Kurths, and B. Hu, Phys. Rev. Lett.87, 098101
~2001!.
-
@13# R.F. Fox, Phys. Rev. A43, 2649~1991!.@14# H. Sompolinsky, H. Yoon, K. Kang, and M. Shamir, Phys. Re
E 64, 051904~2001!.@15# J. Feng and P. Zhang, Phys. Rev. E63, 051902~2001!.@16# Y. Yu, W. Wang, J. Wang, and F. Liu, Phys. Rev. E63, 021907
~2001!; F. Liu, J.F. Wang, and W. Wang,ibid. 59, 3453~1999!.@17# X. Wang and G. Buzsaki, J. Neurosci.16, 6402~1996!.@18# X. Pei, L. Wilkens, and F. Moss, Phys. Rev. Lett.77, 4679
~1996!.@19# S. Tanabe, S. Sato, and K. Pakdaman, Phys. Rev. E60, 7235
~1999!.@20# S. Tanabe and K. Pakdaman, Biol. Cybern.85, 269 ~2000!.@21# C. Zhou and J. Kurths, Chaos13, 401 ~2003!.@22# J.A. Movshon, Neuron27, 412 ~2000!.@23# Neuron 24, 7 ~1999!, special issue on the binding problem
edited by A. Roskies.@24# W. Bair and K. Koch, Neural Comput.15, 1185 ~1996!; A.
Riehle, S. Gru¨n, M. Diesmann, and A. Aertsen, Science278,1950 ~1997!.
@25# L.F. Abbott, Nat. Neurosci.4, 115 ~2001!; J.J. Hopfield andC.D. Brody, Proc. Natl. Acad. Sci. U.S.A.97, 13 919~2000!.
@26# M. Rudolph and A. Destexhe, Phys. Rev. Lett.86, 3662~2001!; J. Comput. Neurosci.11, 19 ~2001!.
9-7
Copyright © 2022 FDOKUMEN



















