
CHARACTERIZATION OF (Na+ + K + )-ATPase LIPOSOMES II. EFFECT OF a-SUBUNIT DIGESTION ON INTRAMEMBRANE...
9
262 Biochimica et Biophysica Acta, 773 (1984) 262-270 Elsevier BBA 72128 CHARACTERIZATION OF (Na+ + K+ )-ATPase LIPOSOMES II. EFFECT OF a-SUBUNIT DIGESTION ON INTRAMEMBRANE PARTICLE FORMATION AND Na +,K +-TRANSPORT BEATRICE M. ANNER a, H. PING TING-BEALL b and J. DAVID ROBERTSON h " Department of Pharmacology, Centre Médical Universitaire, CH-I2H Geneva 4 (Switzerland) and h Department of Anatomy, Duke University Médical Cenîer, Durham, NC 27710 (U.S.A.) (Received December 6th, 1983) Keywords: (No+ + K +)-ATPase; Liposome; Subunit modification; Trypsin digestion; Membrane reconstitution; Particle density; Freeze-fracture; (Rabbit kidney) The effect of thé protein structure of (Na+ + K*)-ATPase on its incorporation into liposome membranes was investigated as follows:the catalytic a-subunit of (Na + + K + )-ATPase was split into low-molecular weight fragments by trypsin treatment and thé digested enzyme was reconstituted at thé same protein concentration as intact control enzyme. The reconstitution process was quantified by thé average number of intramembrane particles appearing on concave and convex fracture faces after freeze-fracture of thé (Na + + K+)-ATPase liposomes. The number of intramembrane particles as well as their distribution on concave and convex fracture faces is not modified by thé proteolysis. In contrast, thé ATPase activity and thé transport capacity of thé (Na ' + K + )-ATPase decrease progessively with increasing incubation times in thé présence of trypsin and are abolished when thé original 100000 molecular weight a-subunit is no longer visible by sodium dodecylsulfate gel electrophoresis. Apparently, functional (Na+ + K+)-ATPase with intact protein structure and digested, non functional enzyme consisting of fragments of thé a-subunit reconstitute in thé same manner and to thé same extent as judged by freeze-fracture analysis. We conclude that, while trypsin treatment modifies thé (Na* + K+ )-ATPase molécule in a functional sensé, il appears not to modify its interaction with thé bilayer in producing intramembrane particles. On thé basis of our results, we propose a lipid-lipid interaction mechanism for reconstitution of (Na+ + K+ )-ATPase. Introduction In thé preceeding publication [1] we found that inhibition of (Na + + K + )-ATPase with ouabain and vanadate had essentially no effect on thé enzyme's ability to reconstitute. In constrast, in- tense détergent-treatment abolished thé enzyme's capacity to reconstitute in lipid bilayers and to produce intramembrane particles [1]. We attribute this effect to a disruption of a lipid shell around thé portions of thé protein normally embedded in thé lipid bilayer. 0005-2736/84/S03.00 © 1984 Elsevier Science Publishers B.V. In view of thèse findings, we wondered whether intact protein structure is as important for recon- stitution as thé structure of thé lipid shell around thé protein. The best way to modify selectively thé protein part of thé (Na + + K+)-ATPase molécule is proteolysis [2-4]. We therefore treated thé (Na + + K+>-ATPase molécules with trypsin and reconstituted thé mod- ified molécules into liposomes. We then de- termined vesicle size, intramembrane particle frequency and distribution with respect to concave and convex fracture faces as well as thé Na + ,K + -
Transcript of CHARACTERIZATION OF (Na+ + K + )-ATPase LIPOSOMES II. EFFECT OF a-SUBUNIT DIGESTION ON INTRAMEMBRANE...

262 Biochimica et Biophysica Acta, 773 (1984) 262-270Elsevier
BBA 72128
CHARACTERIZATION OF (Na+ + K + )-ATPase LIPOSOMES
II. EFFECT OF a-SUBUNIT DIGESTION ON INTRAMEMBRANE PARTICLE FORMATIONAND Na+,K +-TRANSPORT
BEATRICE M. ANNER a, H. PING TING-BEALL b and J. DAVID ROBERTSON h
" Department of Pharmacology, Centre Médical Universitaire, CH-I2H Geneva 4 (Switzerland) and h Department of Anatomy,Duke University Médical Cenîer, Durham, NC 27710 (U.S.A.)
(Received December 6th, 1983)
Keywords: (No+ + K +)-ATPase; Liposome; Subunit modification; Trypsin digestion; Membrane reconstitution; Particle density;Freeze-fracture; (Rabbit kidney)
The effect of thé protein structure of (Na+ + K*)-ATPase on its incorporation into liposome membraneswas investigated as follows:the catalytic a-subunit of (Na++ K + )-ATPase was split into low-molecularweight fragments by trypsin treatment and thé digested enzyme was reconstituted at thé same proteinconcentration as intact control enzyme. The reconstitution process was quantified by thé average number ofintramembrane particles appearing on concave and convex fracture faces after freeze-fracture of thé(Na++ K+)-ATPase liposomes. The number of intramembrane particles as well as their distribution onconcave and convex fracture faces is not modified by thé proteolysis. In contrast, thé ATPase activity and thétransport capacity of thé (Na ' + K + )-ATPase decrease progessively with increasing incubation times in théprésence of trypsin and are abolished when thé original 100000 molecular weight a-subunit is no longervisible by sodium dodecylsulfate gel electrophoresis. Apparently, functional (Na+ + K+)-ATPase with intactprotein structure and digested, non functional enzyme consisting of fragments of thé a-subunit reconstitute inthé same manner and to thé same extent as judged by freeze-fracture analysis. We conclude that, whiletrypsin treatment modifies thé (Na* + K + )-ATPase molécule in a functional sensé, il appears not to modifyits interaction with thé bilayer in producing intramembrane particles. On thé basis of our results, we proposea lipid-lipid interaction mechanism for reconstitution of (Na+ + K + )-ATPase.
Introduction
In thé preceeding publication [1] we found thatinhibition of (Na+ + K + )-ATPase with ouabainand vanadate had essentially no effect on théenzyme's ability to reconstitute. In constrast, in-tense détergent-treatment abolished thé enzyme'scapacity to reconstitute in lipid bilayers and toproduce intramembrane particles [1]. We attributethis effect to a disruption of a lipid shell aroundthé portions of thé protein normally embedded inthé lipid bilayer.
0005-2736/84/S03.00 © 1984 Elsevier Science Publishers B.V.
In view of thèse findings, we wondered whetherintact protein structure is as important for recon-stitution as thé structure of thé lipid shell aroundthé protein. The best way to modify selectively théprotein part of thé (Na+ + K+)-ATPase moléculeis proteolysis [2-4].
We therefore treated thé (Na+ + K+>-ATPasemolécules with trypsin and reconstituted thé mod-ified molécules into liposomes. We then de-termined vesicle size, intramembrane particlefrequency and distribution with respect to concaveand convex fracture faces as well as thé Na+ ,K+-

263
transport capacity of thé reconstituted liposomes.We wished to know whether or not thé fragmenteda-subunits would produce a larger number ofsmaller intramembrane particles than thé intactmaterial. Thus we compared equal amounts oftreated enzyme incorporated into liposome for in-tramembrane particle counts. We found that théNa+ ,K +-transport capacity of liposomes recon-stituted with digested a-subunit is abolished butnot thé appearance and distribution of in-tramembrane particles.
Materials and Methods
Purified (Na+ + K + )-ATPase was obtainedfrom thé outer medulla of rabbit kidneys by thémethod of J0rgensen [5], Pure egg phosphati-dylcholine (PC) was purchased from Supelco.Trypsin from bovine pancréas (Type III) andsoybean trypsin inhibitor were from Sigma.
Reconstituted (Na+ + K + )-ATPase liposomeswere prepared as described in thé preceeding pub-lication [1] and in Réf. 6. The (Na+ + K + )-ATPasewas trypsin treated in thé présence of NaCl or KC1as described by Jorgensen [3] except that thé con-centration of thé enzyme and thé trypsin/enzymeratio were variée as indicated in thé correspondingfigure legends. Gel electrophoresis in thé présenceof SDS was performed according to thé method ofWeber and Osborne [7]. The morphology of thé(Na+ + K + )-ATPase liposomes was determined asdescribed in thé preceeding publication. Proteinconcentrations were determined by thé procédureof Lowry et al. [8].
The liposomes were prepared in a solution con-taining 50 mM NaCl, 50 mM KC1, 5 mM MgCl2,30 mM imidazole or histidine buffer, 1 mM di-thiothreitol and 1 mM EDTA adjusted to pH 7.15by HC1. The passive transmembranous ion fluxeswere measured as follows : 10 i*,\f solution (hav-ing thé same composition as thé solution in whichthé liposomes were suspended) were added to sea-lable conical plastic tubes containing 0.6 fiCi 22Naor 0.3 jiiCi K6Rb. It was previously demonstratedthat 86Rb can be used as a convenient tracer forK+ fluxes in this System (see Refs. 13, 15, 16, 17 inthé preceeding publication [1]). One volume of théradioactive stock solution was then added to twovolumes of liposomes and thé mixture was in-
cubated at room température. Active Na +-influxinto thé liposomes was performed by adding 30mM ATP to thé radioactive stock solution, yîeld-ing a final ATP concentration of 10 mM. Tofollow active K+-efflux, thé liposomes were firstequilibrated with 86Rb and a concentrated ATPsolution was added to thé 86Rb loaded liposomesto yield a final external ATP concentration of 10mM. The ion fluxes were stopped by putting thésamples on ice and adding a 50-fold excess ofice-cold stop solution containing 100 mM Tris and30 mM imidazole buffer adjusted to pH 7.15 byHC1. The internai isotope content was determinedas described in Réf. 6.
Résulta
Digestion of thé a-subunit by trypsinFig. 1 shows thé protein profiles of a control (a)
and three trypsin-treated (b, c, d) soluble (Na+ 4-K+)-ATPase préparations (supernatant fractions)that were incorporated into liposomes. In prépara-tion b about 65% of thé a-subunit was digested.After a prolonged incubation in thé présence oftrypsin and NaCl (c) a trace of thé a-subunitremained, whereas in KC1 (d) thé a-subunit wascompletely digested within 60 min. Whether théband remaining at thé place of thé /î-subunit isonly undigested jS-subunit or contains in additiona newly formed, trypsin résistant 60000 mol. wt.fragment of thé a-subunit is currently under inves-tigation.
Effect of a-subunit digestion on intramembrane par-ticle formation and distribution
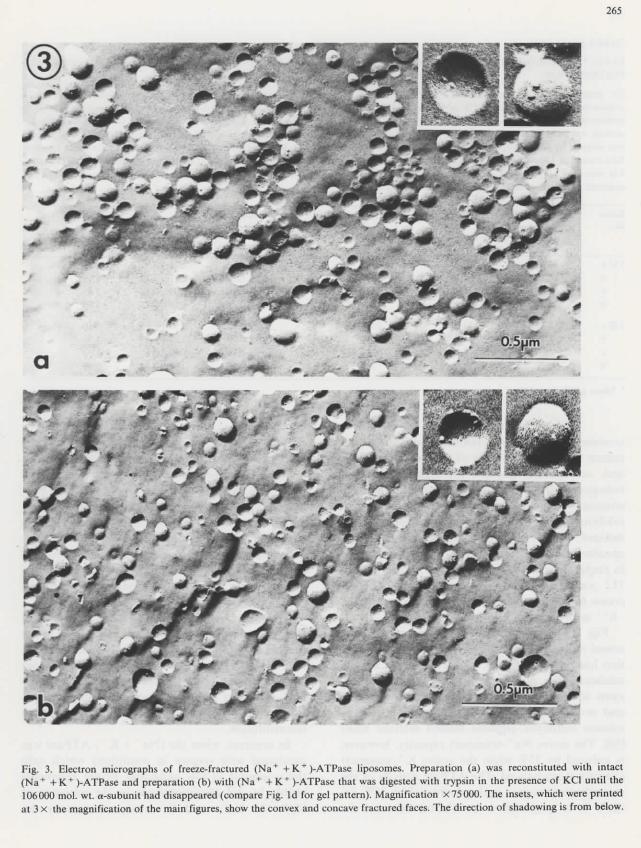
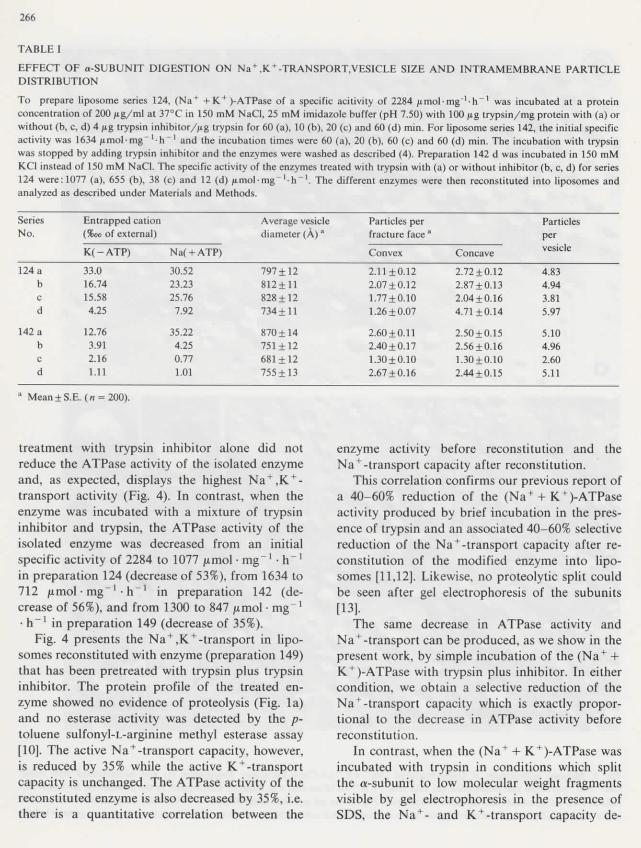
When each of thé three trypsin-treated ATPasepréparations was reconstituted into liposomesneither thé size distribution of thé vesicles, northeir intramembrane particle content, nor thé in-tramembrane particle distribution on concave andconvex membrane fracture-faces, nor thé form andsîze of thé particles were significantly différentfrom those observed with thé reconstitued controlpréparation (Table I and Fig. 2). Thus, as far asthèse parameters are concerned, thé liposomescontaining thé various digested forms of thé a-sub-unit display thé same ultrastructure as théliposomes reconstituted with native enzyme.
However, thé intramembrane particles formed

264
1-2
1-0-
0-8-
0-6
0-4
0-2
0-6-
02
06-
0-2
0 6
<K
02
RELATIVE MOBILITV
Fig. 1. Digestion of thé a-subunit. Préparation (a) was in-cubated at a protein concentration of 200 Mg protein/ml at37°C in 150 mM NaCl, 25 mM imidazole buffer (pH 7.5), 100/tg trypsin/mg protein and 4 /ig trypsin inhibilor/>g trypsinfor 60 min. Préparations (b) and (c) were incubated in thé sameconditions, except thaï trypsin inhibitor was added after a 20(b) and 60 (c) min incubation period in thé présence of trypsin.Préparation (d) was incubated for 60 min under thé sameconditions, but in thé présence of 150 mM KCI instead ofNaCl. The treated enzymes were washed as described in Réf. 4.They were then solibilized in a 1 % cholate solution as describedin Réf. 7 and aliquots of thé 100000 X g supernatants contain-ing 20 p g protein were used for SDS-polyacrylamide gelelectrophoresis. The remaining supernatants were processedfurther to liposome séries 142. The Coomassie blue stainedprotein profiles were obtained in a ZK 4 Zeiss gel scanneraccessory fitted to a Zeiss PMQ spectrophotometer. The appâ-tent molecular weights were 106000 for thé large a-subunit and68000 for thé ,8-subunit as compared to molecular weightmarkers of BDH Chemicals Ltd.
by (Na+ + K + )-ATPase with completely digesteda-subunits are not functional with regard to activeion transport. In fact they are not only inactive,
uu
a o-
60-
b o-
80-
c o-
60-
d 0-
80-(mm) '
(A) C
\
-
M
I
2 i
]
i'
.
6
] l > . - 0-
160-
I,- o-
160i
,- 0-
160-
I i i - 0-
6 10 12
Illll.x.-.-
-
Illn.... . - --
In---
niii-i..-0 2 i* 6 8 10 12
400 800 1200 1600 ,PARTICLES/
/ESICLE DIAMETER
Fig. 2. Histograms of vesicle size and particle distribution ofliposomes reconstituted with trypsinized (Na+ +K + )-ATPase.Liposomes were reconstituted as described in Réf. 7 with(Na+ + K+)-ATPase treated for 60 min with trypsin plustrypsin inhibitor (a) or with trypsin alone for 10 (b), 20 (c) or60 (d) min before reconstitution. The incubation conditions areindicated in thé legend to Fig. 1, thé protein profile in Fig. 1and thé transport properties of thé enzymes in Fig. 4. Aliquotsof thé liposomes were processed for électron microscopy asdescribed under Materials and Methods and 200 convex frac-ture faces (above zéro Une) and 200 concave fracture faces(below zéro line) were analyzed for vesicle size and particle
distribution.
but présent leak pathways. The number of pas-sively entrapped potassium ions and of activelyincorporated sodium ions decrease proportionallyto thé degree of proteolysis and thé associateddrop in ATPase activity (Table I and Réf. 9).
Effect of a-subunit digestion on Na * ,K +-transportExperiments were performed to test thé effect
of trypsin inhibitor alone, trypsin inhibitor plustrypsin or trypsin alone on thé transport proper-ties of (Na+ + K + )-ATPase. The enzyme was firstpretreated in thèse three conditions and was thenreconstituted at equal amounts of protein. Pré-
265
Fig. 3. Electron micrographs of freeze-fractured (Na+ + K+)-ATPase liposomes. Préparation (a) was reconstituted with intact(Na+ +K+)-ATPase and préparation (b) with (Na+ + K+)-ATPase that was digested with trypsin in thé présence of KC1 until thé106000 mol. wt. a-subunit had disappeared (compare Fig. Id for gel pattern). Magnification X75000. The insets, which were printedat 3 X thé magnification of thé main figures, show thé convex and concave fractured faces. The direction of shadowing is from below.
266
TABLE I
EFFECT OF o-SUBUNIT DIGESTION ON Na+,K + -TRANSPORT,VESICLE SIZE AND INTRAMEMBRANE PARTICLEDISTRIBUTION
To prépare liposome séries 124, (Na+ +K+)-ATPase of a spécifie acitivity of 2284 / imo]-mg" ' -h~ ' 1 was incubated at a proteinconcentration of 200 |iig/ml at 37°C in 150 mM NaCl, 25 mM imidazole buffer (pH 7.50) with 100 pg trypsin/mg protein with (a) orwithout (b, c, d) 4 pg trypsin inhibilor/jig trypsin for 60 (a), 10 (b), 20 (c) and 60 (d) min. For liposome séries 142, thé initial spécifieactivity was 1634 /nnol'Ulg -h"1 and thé incubation times were 60 (a), 20 (b), 60 (c) and 60 (d) min. The incubation with trypsinwas stopped by adding trypsin inhibitor and thé enzymes were washed as descnbed (4). Préparation 142 d was incubated in 150 mMKCI instead of 150 mM NaCl. The spécifie activity of thé enzymes treated with trypsin with (a) or without inhibitor (b, c, d) for séries124 were:1077 (a), 655 (b), 38 (c) and 12 (d) pmol-mg'1-!! ', The différent enzymes were ttien reconstituted into liposomes andanalyzed as described under Materials and Methods.
SériesNo.
124 abcd
142 abcd
Entrapped cation(%oo of external)
K(-ATP)
33.016.7415.584.25
12.763.912.161.11
Na( + ATP)
30.5223.2325.76
7.92
35.224.250.771.01
Average vesiclediameter (A) *
797+12812+11828±12734 + 11
870+14751 ±12681±12755 ±13
Particles perfracture face "
Convex
2.11+0.122.07 + 0.121.77 + 0.101.26 + 0.07
2.60 + 0.112.40 + 0.171.30 ±0.102.67 ±0.1 6
Concave
2.72+0.122.87 + 0.132.04 ±0.1 64.71 ±0.14
2.50±0.152.56 ±0.161.30 ±0.102.44±0.15
Particlespervesicle
4.834.943.815.97
5.104.962.605.11
Mean±S.E. (n - 200).
treatment with trypsin inhibitor alone did notreduce thé ATPase activity of thé isolated enzymeand, as expected, displays thé highest Na+ ,K + -transport activity (Fig. 4). In contrast, when théenzyme was incubated with a mixture of trypsininhibitor and trypsin, thé ATPase activity of théisolated enzyme was decreased from an initialspécifie activity of 2284 to 1077 jtimol • mg"1 • h"1
in préparation 124 (decrease of 53%), from 1634 to712 /j,mol • mg~' • h~ ' in préparation 142 (de-crease of 56%), and from 1300 to 847 )nmol • mg~ '• h~ ' in préparation 149 (decrease of 35%).
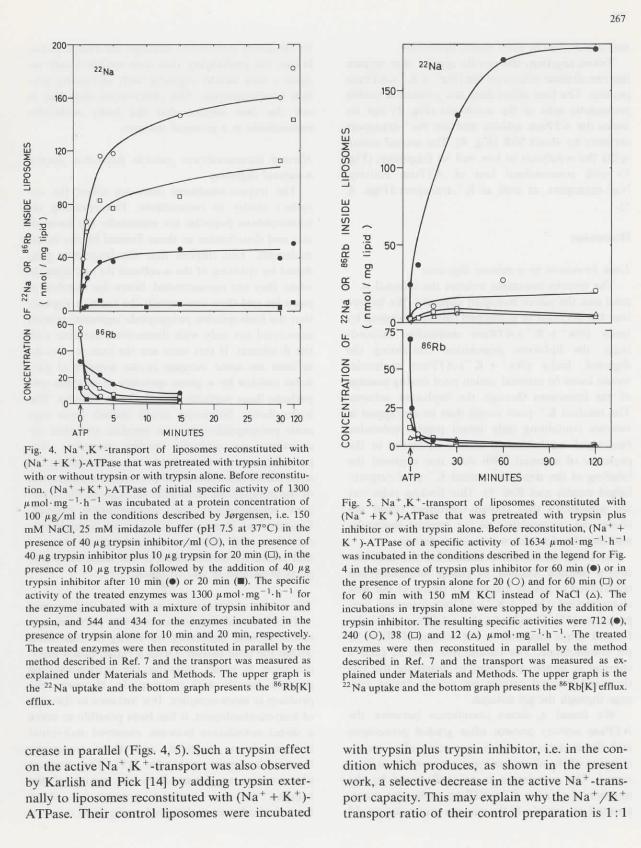
Fig. 4 présents thé Na+ ,K + -transport in lipo-somes reconstituted with enzyme (préparation 149)that has been pretreated with trypsin plus trypsininhibitor. The protein profile of thé treated en-zyme showed no évidence of proteolysis (Fig. la)and no esterase activity was detected by thé p-toluene sulfonyl-L-arginine methyl esterase assay[10]. The active Na+-transport capacity, however,is reduced by 35% while thé active K + -transportcapacity is unchanged. The ATPase activity of théreconstituted enzyme is also decreased by 35%, i.e.there is a quantitative corrélation between thé
enzyme activity before reconstitution and théNa +-transport capacity after reconstitution.
This corrélation confirms our previous report ofa 40-60% réduction of thé (Na++ K + )-ATPaseactivity produced by brief incubation in thé prés-ence of trypsin and an associated 40-60% sélectiveréduction of thé Na + -transport capacity after re-constitution of thé modified enzyme into lipo-somes [11,12], Likewise, no proteolytic split couldbe seen after gel electrophoresis of thé subunits[13].
The same decrease in ATPase activity andNa+-transport can be produced, as we show in théprésent work, by simple incubation of thé (Na+ +K + )-ATPase with trypsin plus inhibitor. In eithercondition, we obtain a sélective réduction of théNa+-transport capacity which is exactly propor-tional to thé decrease in ATPase activity beforereconstitution.
In contrast, when thé (Na+ + K+)-ATPase wasincubated with trypsin in conditions which splitthé a-subunit to low molecular weight fragmentsvisible by gel electrophoresis in thé présence ofSDS, thé Na + - and K +-transport capacity de-
267
30 120
ATP
Fig. 4. Na + ,K + -transport of liposomes reconstituted with(Na* + K+ )-ATPase that was pretreated with trypsin inhibitorwith or without trypsin or with trypsin alone. Before reconstitu-tion. (Na+ +K+)-ATPase of initial spécifie activity of 1300Hmol-mg"1-!!"1 was incubated at a protein concentration of100 ^g/ml in thé conditions described by Jorgensen, i.e. 150mM NaCl, 25 mM imidazote buffer (pH 7.5 at 37°C) in théprésence of 40 jug trypsin inhibitor/ml (O), in thé présence of40 ftg trypsin inhibitor plus 10 ftg trypsin for 20 min (D), in théprésence of 10 pg trypsin followed by thé addition of 40 jtgtrypsin inhibitor after 10 min (•) or 20 min (•). The spécifieactivity of thé treated enzymes was 1300 jumol-mg"1^"1 forthé enzyme incubated with a mixture of trypsin inhibitor andtrypsin, and 544 and 434 for thé enzymes incubated in théprésence of trypsin alone for 10 min and 20 min, respectively.The treated enzymes were then reconstituted in parallel by thémethod described in Réf. 7 and thé transport was measured asexplained under Materials and Methods. The upper graph isthé 22Na uptake and thé bottom graph présents thé 86Rb[K]efflux.
1/1LU
IOCL
LJQ
l/)
150-
•gQ.
60
MINUTES
90 120
Fig. 5. Na+ ,K + -transport of liposomes reconstituted with(Na+ +K+)-ATPase that was pretreated with trypsin plusinhibitor or with trypsin alone. Before reconstitution, (Na+ +K+)-ATPase of a spécifie activity of 1634 j u . m o l - m g ~ ' - h ~ l
was incubated in thé conditions described in thé legend for Fig.4 in thé présence of trypsin plus inhibitor for 60 min (•) or inthé présence of trypsin alone for 20 (O) and for 60 min (a) orfor 60 min with 150 mM KC1 instead of NaCl (A). Theincubations in trypsin alone were stopped by thé addition oftrypsin inhibitor. The resulting spécifie activities were 712 (•),240 (O), 38 (D) and 12 (A) ^mol-mg-'-h'1 . The treatedenzymes were then reconstitued in parallel by thé methoddescribed in Réf. 7 and thé transport was measured as ex-plained under Materials and Methods. The upper graph is thé2 2Na uptake and thé bottom graph présents thé 86Rb[K] efflux.
crease in parallel (Figs. 4, 5). Such a trypsin effecton thé active Na+ ,K + -transport was also observedby Karlish and Pick [14] by adding trypsin exter-nally to liposomes reconstituted with (Na+ + K + )-ATPase. Their control liposomes were incubated
with trypsin plus trypsin inhibitor, i.e. in thé con-dition which produces, as shown in thé présentwork, a sélective decrease in thé active Na +-trans-port capacity. This may explain why thé Na+/K +
transport ratio of their control préparation is 1:1

268
and not 3 : 2 as in their native liposomes.Taken together, thé results suggest that trypsin
has two distinct effects on thé (Na+ + K + )-ATPaseprotein. The first effect does not produce a visibleproteolytic split of thé a-subunit (Fig. 1) but re-duces thé ATPase activity and thé Na+-transportcapacity by about 50% (Fig. 4). The second actionsplits thé a-subunit to low mol wt fragments (Fig.1) with concomitant loss of ATPase activity,Na+-transport, as well as K+-transport (Figs. 4,5).
Discussion
Leak formation by a-subunit digestionThe trypsin-treatment reduces thé internai K + -
pool and thé active transport capacity. We believethat thé decrease in entrapped cations is caused byleaky (Na+ + K + )-ATPase molécules. Accord-ingly, thé liposome population containing thédigested, leaky (Na+ + K + )-ATPase moléculeswould loose its internai cation pool during passageof thé liposomes through thé Sephadex column.The residual K^-pool would then be entrapped invesicles containing only intact pump molécules.Prolonged incubation of altered liposones in théprésence of external 86Rb does not augment thélabeling of thé residual internai K + -pool (unpub-lished results and Réf. 9). This finding rules outthé possibility that trypsin-treatment could de-crease thé permeability of thé enzyme moléculesand thereby slow down isotope exchange.
To bring additional évidence for thé leak-the-ory, a defined number of pump molécules wastransferred to planar bilayers by fusion of our wellcharacterized (Na+ + K+)-ATPase-liposome pré-parations [15]. The (Na+ + K f)-ATPase moléculesformed single channels in thé bilayer of 40 to 50pS conductance. The cation flux rate in such achannel is largely sufficient to eliminate thé fewthousand internai cations per vesicle during pas-sage through thé gel column.
We found a direct corrélation between théATPase activity présent after graded proteolysisand thé transport activity appearing after recon-stitution of thé altered enzyme préparation in lipo-somes. The corrélation is not influenced by thénumber of (Na+ + K+)-ATPase molécules incor-porated per vesicle. If thé enzyme molécules would
be randomly distributed amongst thé vesicle popu-lation, thé probability that each vesicle would re-ceive a leak would augment with increasing pro-tein concentrations. The observation that this isnot thé case implies that thé leaky moléculesreconstitute in a grouped manner.
Normal intramembrane partide formation despitea-subunit digestion
The trypsin-treatment does not affect thé en-zyme's ability to reconstitute. The resulting in-tramembrane particles are essentially thé same ofsize and distribution as those formed by thé intactmolécules. This implies that thé fragments pro-duced by splitting of thé a-subunit do not separatewhen they are reconstituted. Since thé number ofparticles and their size remain thé same, it appearsthat thé hydrophobic polypeptide segments remainassociated not only with themselves, but also withthé /?-subunit. If this were not thé case we shouldat least see some increase in thé number of par-ticles yielded by a given quantity of enzyme andperhaps thèse particles should also be smaller. Thehydrophobic fragments could indeed form sep-arate polypeptide-detergent micelles and then re-constitute as multiple smaller particles. Ap-parently, thé boundary lipid layer surrounding théa2^2 dimer stays there and prevents thé séparationof thé polypeptide fragments throughout thé déter-gent solubilization and reconstitution process.Deamer and Yamanaka [16] treated sarcoplasmicreticulum with thé proteolytic Nagarase enzymeand found no modification of thé intramembraneparticle pattern in this situation.
The nature of intramembrane particles in liposomesIt is believed that intramembrane particles rep-
resent protein molécules traversing or embeddedwithin thé lipid bilayer. Often a rather Hteral inter-prétation is made equating thé intramembraneparticle directly with polypeptide. However, théproblem is more complex. For instance in thé caseof bacteriorhodopsin, it has been possible to makea direct corrélation between observed individualimtramembrane particles and three transversebacteriorhodopsin molécules [17]. The fractureplane seems to départ from thé center of thébilayer to run around groups of threebacteriorhodopsin molécules leaving them intact in

269
thé PF face. It thus appears that in général théfracture plane does not break polypeptide chains.Récent work by Pinto da Silva and his group bythé 'fracture-label' technique applied to erythro-cyte membranes supports thé idea that thé fractureplane tends to deviate around transverse proteinmolécules leaving thé whole molécule within oneor thé other fracture face [18]. In thé case of thénative (Na+ + K + )-ATPase molécule, where thereis a pronounced structural asymmetry (19-21], îtseems reasonable to suppose such an asymmetricalfracture plane. The fact that we find thé samenumber of intramembrane particles on thé convexand concave fracture faces of liposomes wouldthen indicate a random orientation of thé (Na+ +K + )-ATPase molécules.
One might expect that each intramembrane par-ticle would hâve a corresponding pit in thé oppo-site fracture face. However, we hâve seen no pitsin any of our préparations. The failure to démon-strate pits to correspond to intramembrane par-ticles has puzzled freeze-fracture électron micros-copists for many years [22]. It is commonly statedthat it is very easy to obliterate pits by contamina-tion and not so easy to obliterate intramembraneparticles. There are other explantions, such as thénotion that thé intramembrane particle does notrepresent thé protein molécule and its associatedlipîd directly but rather a disturbance in thé lipidbilayer caused by thé présence of thé protein lead-ing to sélective décoration of thé fracture face inthé région of thé disturbance [23]. This mechanismwould produce intramembrane particles but notpits and would also provide an explanation for thérelatively large size of intramembrane particleswhich often exceeds estimâtes of thé mass of théprotein that could be involved.
A lipid-lipid interaction mechanism for reconstitu-tion of (Na+ + K +)-A TPase
In visualizîng thé mechanism whereby thé (Na +
+ K + )-ATPase liposomes form we may regard théenzyme molécules as condensation centers for pré-cipitation of thé phospholipîd molécules fromsolution. Thus, it seems reasonable to suppose thatthé phospholipid molécules would attach to théparts of thé enzyme molécule that are alreadycoated with a shell of lipid from thé native mem-brane. A protein-associated lipid shell of 50 to 60
molécules per a2/22 dimer has been demonstratedby électron spin résonance [24,25]. If we assumethat this enzyme phospholipid retains its essentialbilayer configuration, wîth thé détergent moléculesproviding an interface between thé carbon chainsand thé water, thé newly added phospholipidmolécules would be expected to replace détergentmolécules and attach themselves to thé enzymelipids with thé carbon chains and head groupslining up with their counterparts so as to form anextending bilayer.
Acknowledgement
Mrs. Marlis Moosmayer provided outstandingassistance with regard to (Na++ K+)-ATPasepurification, liposome préparation and transportmeasurement. The work was supported by grantNo. 3.155-0.81 of thé Swiss National ScienceFoundation to B.M.A and thé U.S. National In-stitute of Health GM-27804 and GM/AM-28224to H.P.T.-B. and J.D.R., respectively.
Références
1 Anner, B.M., Robertson, J.D. and Ting-Beall, H.R (1984)Biochim. Biophys. Acta 773, 253-261
2 Giotta, G.J. (1975) J. Biol. Chem. 250, 5159-51643 J0rgensen, P.L. (1975) Biochim. Biophys. Acta 401, 399-4154 J0rgensen, P.L., Karlish, S.J.D. and Gitler, C. (1982) J. Biol.
Chem. 257, 7435-74425 Jergensen, P.L. (1974) Methods Enzymol. 32, 277-2906 Anner, B.M. and Moosmayer, M. (1982) Biochem. Biophys.
Methods 5, 299-3067 Weber, K. and Osborn, M. (1969) J. Biol. Chem. 244,
4460-44128 Lowry, O.H., Rosebrough, N.J., Farr, A.L. and Randall,
R.J. (1952) J. Biol. Chem. 193, 265-2759 Anner, B.M. (1983) FEBS Lett. 158, 7-11
10 Walsh, K.A. (1970) Methods Enzymol. 19, 41-6311 Anner, B.M. and Jergensen, P.L. (1979) in Na+K-
ATPase : Structure and Kinetics (Skou, J.C. and Norby,J.G., eds.), pp. 87-97, Académie Press, London/New York
12 J0rgensen, P.L. and Anner, B.M. (1979) Biochim. Biophys.Acta 555, 485-492
13 J0rgensen, P.L. (1977) Biochim. Biophys. Acta 466, 97-10814 Karlish, S.J.D. and Pick, U. (1981) J. Physiol. 312, 505-52915 Reinhardt, R., Lindenmann, B. and Anner, B.M. (1984)
Biochim. Biophys. Acta 774, in thé press (BBA 70167)16 Deamer, D.W. and Yamanaka, N. (1975) Biophys. J. 15,
llOa17 Robertson, J.D. (1981) in Demyelinating Disease and Basic
and Clinical Electrophysiology (Waxman, S.G. and Ritchie,J.M., eds.), pp. 419-477, Raven Press, New York

270
18 Pinto da Silva, P., Parkison, C. and Dwyer, N. (1981) Proc. 23 Robertson, J.D. (1978) in Physiology of Membrane Dis-Natl. Acad. Sci. U.S.S. 78, 343-347 orders (Andreoli, T.E., Hoffman, J.E. and Fanestil, D.D.,
19 Repke, K.R.H. (1982) Ann. N.Y. Acad. Sci. 402, 272-286 eds.), pp. 61-93, Plénum Médical Book Co., New York20 J0rgensen, P.L., Skriver, E., Herbert, H. and Maunsbach, B. 24 Brotherus, J.R., Jost, P.C., Griffith, O.H., Keana, J.F.W.
(1982) Ann. N.Y. Acad. Sci. 402, 207-225 and Hokin, L.E. (1980) Proc. Natl. Acad. Sci. U.S.A. 77,21 Ting-Beall, H.P., Holland, V.F., Freytag, J.W. and Hast- 272-276
ings, D.F. (1983) Biophys. J. 41,5a 25 Marsh, D., Watts, A., Pâtes, R.D. and Uhl, R. (1982)22 Roberison, J.D. (1978) in Physiology of Membrane Dis- Biophys. J. 37, 265-274
orders (Andreoli, T.E., Hoffman, J.E. and Fanestil, D.D.,eds.), pp. 1-26, Plénum Médical Book Co., New, York









![FYdgYdcYf]k Yf\ FYdgYj]f]k - Samagra](https://static.fdokumen.com/doc/165x107/631f358f13819e2fbb0fa1db/fydgydcyfk-yf-fydgyjfk-samagra.jpg)










