
The Use of Photolabels to Probe the Ouabain Binding Site of the (Na, K)-Atpase
14
THE USE OF PHOTOLABELS TO PROBE THE OUABAIN BINDING SITE OF THE (Na, K)-ATPase* Arnold E. Ruoho and Clifford C. Hall Department of Pharmacology University of Wisconsin Madison, Wisconsin 53706 INTRODUCTION Digitalis glycosides are specific inhibitors of the ion transport (Na, K)-ATPase (ATP phosphohydratase, EC 3.6.1.3).',2It has been suggested that this enzyme is the receptor through which the cardiac glycosides produce their inotropic effect on the heart The (Na, K)-ATPase, purified from several source^,^-^ consists of at least two polypeptides: a catalytic subunit ((u). M, = 100,000, and a glycoprotein (@), M, = 50,000. Structural studies indicate that the enzyme probably contains an (u2@2'0 or (u2P4" structure. During the hydrolysis of ATP, the terminal phosphate is transferred to the a subunit of the enzyme as an aspartyl phosphate.I2 This phosphorylation is dependent upon the simultaneous presence of Na' and Mg+ +.I3 Cardiac glycosides bind to this phosphorylated form of the which has been described as a type I ~omplex."'~'~ The enzyme can be phosphorylated with inorganic phosphate in vitro with the simultaneous presence of Mg' + and cardiac glycosides." This latter cardiac glycoside-enzyme complex has been referred to as the type II c~mplex.'~,'~ Cardiac glycoside derivatives that are capable of covalent bond formation in the binding site are useful for several reasons: 1. To allow analysis of peptide fragments of the binding site, which will be exposed to denaturing conditions 2. To identify structural relationships between the various binding sites of the enzyme (e.g., ATP site and glycoside binding sites) 3. To probe the cardiac glycoside binding site with compounds that can also be used in intact atria to assess the similarity of the (Na, K)-ATPase binding site with the in sifu cardiac glycoside receptor. We prepared 3-haloacetyl derivatives of strophanthidin and hellebrigenin for affinity labeling of the binding site," but they lacked sufficient specificity for structural work (FIGURE I). One disadvantage of these reagents is the requirement for a suitably positioned nucleophile in the active site. Since the 3-haloacetyl compounds (which were inhibitors and therefore did bind to the enzyme at the specific binding site) did not result in measurable yields of covalent bond formation, it appeared that suitable nucleophilic residues were beyond bond forming distance at the 3-hydroxyl position of the steroid binding site. It was clear from these experiments that affinity probes would require either derivation at positions other than the 3-hydroxyl in order to seek out appropriate functional groups in the protein that are suitably positioned in *This work was supported by a National Institutes of Health grant, no. NS12392. 90 0077-8923/80/0346&O090 $l.75/1 0 1980. NYAS
-
Upload
independent -
Category
Documents
-
view
0 -
download
0
Transcript of The Use of Photolabels to Probe the Ouabain Binding Site of the (Na, K)-Atpase

THE USE OF PHOTOLABELS TO PROBE THE OUABAIN BINDING SITE OF THE (Na, K)-ATPase*
Arnold E. Ruoho and Clifford C. Hall
Department of Pharmacology University of Wisconsin
Madison, Wisconsin 53706
INTRODUCTION
Digitalis glycosides are specific inhibitors of the ion transport (Na, K)-ATPase (ATP phosphohydratase, EC 3.6.1.3).',2 It has been suggested that this enzyme is the receptor through which the cardiac glycosides produce their inotropic effect on the heart
The (Na, K)-ATPase, purified from several source^,^-^ consists of at least two polypeptides: a catalytic subunit ((u). M, = 100,000, and a glycoprotein (@), M , =
50,000. Structural studies indicate that the enzyme probably contains an (u2@2'0 or (u2P4" structure.
During the hydrolysis of ATP, the terminal phosphate is transferred to the a subunit of the enzyme as an aspartyl phosphate.I2 This phosphorylation is dependent upon the simultaneous presence of Na' and Mg+ + . I 3 Cardiac glycosides bind to this phosphorylated form of the which has been described as a type I ~omplex." '~ '~ The enzyme can be phosphorylated with inorganic phosphate in vitro with the simultaneous presence of Mg' + and cardiac glycosides." This latter cardiac glycoside-enzyme complex has been referred to as the type I I c ~ m p l e x . ' ~ , ' ~
Cardiac glycoside derivatives that are capable of covalent bond formation in the binding site are useful for several reasons:
1 . To allow analysis of peptide fragments of the binding site, which will be exposed to denaturing conditions
2. To identify structural relationships between the various binding sites of the enzyme (e.g., ATP site and glycoside binding sites)
3. To probe the cardiac glycoside binding site with compounds that can also be used in intact atria to assess the similarity of the (Na, K)-ATPase binding site with the in sifu cardiac glycoside receptor.
We prepared 3-haloacetyl derivatives of strophanthidin and hellebrigenin for affinity labeling of the binding site," but they lacked sufficient specificity for structural work (FIGURE I ) . One disadvantage of these reagents is the requirement for a suitably positioned nucleophile in the active site. Since the 3-haloacetyl compounds (which were inhibitors and therefore did bind to the enzyme a t the specific binding site) did not result in measurable yields of covalent bond formation, it appeared that suitable nucleophilic residues were beyond bond forming distance at the 3-hydroxyl position of the steroid binding site. It was clear from these experiments that affinity probes would require either derivation at positions other than the 3-hydroxyl in order to seek out appropriate functional groups in the protein that are suitably positioned in
*This work was supported by a National Institutes of Health grant, no. NS12392.
90 0077-8923/80/0346&O090 $l.75/1 0 1980. NYAS

Ruoho & Hall: Ouabain Binding Site 91
OH
STR: R = H HEL: R'= ~ 2 = H S I A : R =ICH,CO SBA : R = BrCH2C0
H A : R'=CH,CQ R 2 = H HDA : R' = R2= CH,CO
HBA: R '= BrCH2 CO; R 2 = H H I A : R'= ICH2 CO; R2 = H
FIGURE I . The structure of haloacetyl derivatives. STR: strophanihidin; SIA: strophanthidin- 3-iodoacetate; SBA: strophanthidin-3-bromoacetate; HEL: hellebrigenin; HA: hellebrigenin- )-acetate: HDA: hellebrigenin-3.5-diacetate; H I A : hellebrigenin-3-iodoacetate; HBA: hellebrigenin-3-bromoacetate.
the binding site to allow reaction to occur (in erect, a molecular "fishing expedition") or incorporation of groups that formed covalent bonds in a much more nonselective fashion and that were more reactive. This latter requirement is very effectively met by the use of ligands, which can be converted by photolysis into exceedingly reactive intermediates and, under appropriate conditions, insert into C-H bonds." Compounds that generate carbenes of nitrenes upon photolysis have been used for the photoaffinity labeling of a variety of ligand binding sites.2' The use of these compounds, therefore, allows a greater opportunity for covalent bond formation in a binding site, since the presence of specific reactive centers (e.g., N-H, 0 - H , S-H) is probably not necessary.
In addition to the photolabels described in this paper, other photoactivatable compounds have been reported for photolabeling of the ouabain binding site of this enzyme.22 *'
LABELING OF THE a SUBUNIT OF THE ENZYME
J'C-Diazomalonyl Cymarin
Cymarin (FIGURE 2) is a cardiac glycoside containing the 2-deoxy, 3-methoxy sugar, cymarose, in glycosidic linkage to the 3-hydroxyl of the aglycone, strophanthi- din. Cymarose contains, therefore, only one secondary hydroxyl that can be acylated with ease. ''C-diazomalonyl (I4C-DAM) chloride was prepared at a specific activity of 5 mCi/mmol.26 When this carbene generator was photolyzed with a medium pressure

92 Annals New York Academy of Sciences

cpm
15
0
100
50
a 1 b
I C
I /
0 P / :P-’
-&
Jl
LL
L
10
20
A
7
10
20
0 R d
Sli
ce
N
um
be
r
FIG
UR
E
3. T
he d
istr
ibut
ion
of r
adio
activ
ity c
oval
ently
atta
ched
to
(Na.
K)-
AT
Pase
that
had
bee
n ph
otol
yzed
in
the
pres
ence
of
“C-D
AM
cym
arin
. Sa
mpl
es of
dog
kid
ney
enzy
me
that
had
bee
n ph
otol
yzed
eith
er in
the
pre
senc
e of
the
com
plet
e M
gAT
P ph
osph
oryl
atin
g sy
stem
(0) o
r in
the
com
plet
e sy
stem
plu
s a 2
5-fo
ld e
xces
s of
cym
arin
as
prot
ecto
r (w
) w
ere
run
on S
DS
gels
, whi
ch w
ere
scan
ned,
slic
ed, a
nd c
ount
ed. T
he A
,, tr
ace
from
the
gel w
ith
the
unpr
otec
ted
sam
ple
was
div
ided
into
segm
ents
exa
ctly
as t
he g
el h
ad b
een
slic
ed. T
he m
ean
A&,,
of e
ach
of th
ese
segm
ents
was
cal
cula
ted
and
the
valu
es
plot
ted
(0).
The
uni
ts o
f ar
e ar
bitr
ary,
sin
ce th
e sc
anne
r w
as u
ncal
ibra
ted.
The
thre
e pr
otei
n co
mpo
nent
s ar
e: (
a) c
ross
-link
ed a
@ di
mer
. (b
) la
rge
chai
n, a
nd (
c) sm
all c
hain
. (Fr
om A
. E. R
uoho
and
J. K
yteZ
6; by
per
mis
sion
of
the
Proc
eedi
ngs
ofth
e N
urio
nol
Aca
dem
y of
Scie
nces
.)

94 Annals New York Academy of Sciences
mercury lamp in the type I complex with the deoxycholate (DOC) purified dog kidney (Na, K)-ATPase, specific photolabel insertion was observed into the a subunit of the enzyme only (FIGURE 3). This data leads to two conclusions:
I . The a subunit contained at least a portion of the cardiac glycoside binding site.
2. The a subunit must span the bilayer since it has been shown that cardiac glycosides inhibit only from the outside surface of the membrane and ATP phosphorylates only from the in~ide.~ '
Primarily because of the low specific radioactivity of this compound, however, it was not possible to unequivocally eliminate the possibility that the /3 subunit contributes to the binding site as well. If the ratio of labeling a:@ was 1O: l . radioactivity would not have been detectable in the p subunit region.
N A P Glycyl Digitoxigenin and N A P GlycyI Strophanthidin
The synthesis of these photolabels was accomplished in the following fashion. Nitrofluorophenylazide was reacted with glycine to produce N-nitrophenylazidogly- cine.28 The acyl halide of this compound was produced with oxalyl chloride. The 3-hydroxyl of digitoxigenin or strophanthidin was then acylated in pyridine and dioxane. The tritiated compound was synthesized by reacting 'H-digitoxigenin (7-1 7 Ci/mmol; derived by purification of the acid hydrolysate of 'H-digitoxin on thin layer chromatography) with N-nitrophenylazidoglycyl chloride (NAP glycyl chloride), as described above. The product, 'H-NAP glycyl digitoxigenin, was 99% radiopure in three solvent systems on thin layer silica gel G.
Derivation of the 3-hydroxyl of the cardiotonic steroid aglycones does not compromise their inhibition of the action of (Na. K)-ATPase. NAP glycyl digitoxi- genin (FIGURE 2) and NAP glycyl strophanthidin (FIGURE 4) retain the same potency for inhibition of (Na, K)-ATPase activity as digitoxigenin. The Is , (concentration for half maximal inhibition) for these two photolabels was found to be 1-3 x lo-' M ( F I G U R E 4). which was similar to that determined for digitoxigenin under the same experimental conditions. The nitrene generated on photolysis (as estimated from molecular models) is positioned slightly beyond the second glycosidic bond of digitoxin, the parent glycoside, and will, therefore, probe that area of the glycoside binding site. Since 'H-digitoxin could be readily purchased at high specific radioac- tivity (7.4-1 2.2 Ci/mmol), the digitoxigenin derivative was synthesized in a tritiated form as a photoaffinity label for the binding site.
Photolysis of the enzyme-photolabel complex resulted in ouabain protectable covalent labeling of the a subunit (FIGURE 5); no labeling was observed if the enzyme-photolabel complex was not photolyzed. This specific photoincorporation into the large polypeptide agrees with experiments previously r e p ~ r t e d . ~ ~ . ~ ~ . ~ ~ The labeling pattern was found to be virtually identical for both type I and type I1 complexes. No specific labeling could be detected with either type of complex in the glycoprotein (p) subunit (slices 15-17, F I G U R E 5). Because of the very high specific radioactivity of the photolabel, even 10% of that incorporated into the a subunit would have been readily detected. No more than 5% of the site-bound label was photoincorporated into the (Y
subunit. Some cross-linking of the a subunit was observed in these experiments but none of
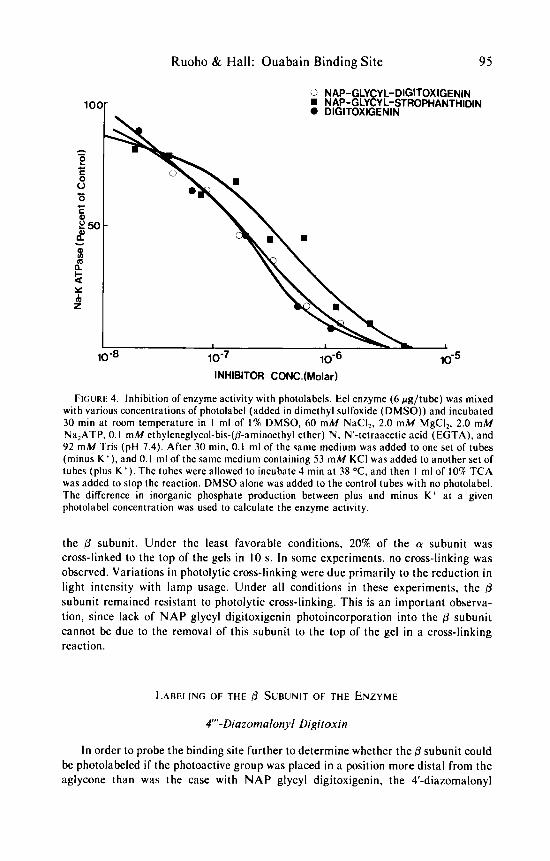
Ruoho & Hail: Ouabain Binding Site 95
o NAP-GLYCYL-DIGITOXIGENIN w NAP-GLYCY L-STROPHANTHIDIN 0 DlGlTOXlGENlN
lNHlBlTOR CONC.(Molar)
F I G U R ~ 4. Inhibition of enzyme activity with photolabels. Eel enzyme (6 pg/tube) was mixed with various concentrations of photolabel (added in dimethyl sulfoxide (DMSO)) and incubated 30 min at room temperature in I ml of 1% DMSO, 60 mM NaCI,, 2.0 m M MgCI,, 2.0 mM Na,ATP. 0. I mM ethyleneglycol-bis-(6-aminoethyl ether) N, "-tetraacetic acid (EGTA), and 92 m M Tris (pH 7.4). After 30 min, 0.1 ml of the same medium was added to one set of tubes (minus K + ) , and 0.1 ml of the same medium containing 53 mM KCI was added to another set of tubes (plus K ' ) . The tubes were allowed to incubate 4 min at 38 "C, and then 1 ml of 10% TCA was added to stop the reaction. DMSO alone was added to the control tubes with no photolabel. The difference in inorganic phosphate production between plus and minus K' at a given photolabel concentration was used to calculate the enzyme activity.
the @ subunit. Under the least favorable conditions, 20% of the a subunit was cross-linked to the top of the gels in 10 s. In some experiments, no cross-linking was observed. Variations in photolytic cross-linking were due primarily to the reduction in light intensity with lamp usage. Under all conditions in these experiments, the p subunit remained resistant to photolytic cross-linking. This is an important observa- tion, since lack of NAP glycyl digitoxigenin photoincorporation into the /3 subunit cannot be due to the removal of this subunit to the top of the gel in a cross-linking reaction.
LABELING OF THE p SUBUNIT OF THE ENZYME
4'"-Diazomalonyl Digitoxin
In order to probe the binding site further to determine whether the /3 subunit could be photolabeled i f the photoactive group was placed in a position more distal from the aglycone than was the case with NAP glycyl digitoxigenin, the 4'-diazomalonyl

96 Annals New York Academy of Sciences
Type I(Na.Mg,ATP)
Slice Number
FIGURE 5. SDS polyacrylamide gel patterns of 'H-NAP glycyl digitoxigenin covalent label- ing.
M ) was incubated with the enzyme for type 1 complex binding. After 90 s of photolysis a t C&2 OC, the eel enzyme was dissolved in 0.1 ml of 2% SDS and 5% 0-mercaptoethanol and electrophoresed on 10% acrylamide ethylene diacrylate cross-linked gels. 0: 1 O-6M 'H-NAP glycyl digitoxigenin; 0 :
M 'H-NAP glycyl digitoxigenin and lo-' M ouabain; A: M 'H-NAP glycyl digitoxigenin bound to the enzyme but not photolyzed. Coomassie staining indicated the presence of the a subunit at slices 7-10 and of the f l subunit at slices 15-18.
M ) was incubated with the enzyme for type I I complex binding. Photolysis and electrophoresis methods are as described above. 0: M 'H-NAP glycyl digitoxigenin and 10.' M ouabain. Coomassie staining indicated the presence of the a subunit a t slices 7-1 1 and of the 0 subunit a t slices 14-17.
Upper panel (type I complex): 'H-NAP glycyl digitoxigenin
Lower panel (type I I complex): 'H-NAP glycyl digitoxigenin
M 'H-NAP glycyl digitoxigenin; 0 :

Ruoho & Hall: Ouabain Binding Site 97
derivative of digitoxin was synthesized (FIGURE 2). Yoda and Y ~ d a * ~ have demon- strated that acetylation of the various hydroxyls of digitoxin by the method of Satoh and Morita” affects the dissociation rate of the enzyme-glycoside complex. Increased rates of dissociation were seen with several acetylated derivatives but most notably with the 3’monoacetyl digitoxin in both type I and type I 1 complexes and the 3”-monoacetyl digitoxin in the type I complex. The least affected, in terms of dissociation rate, was the 4-digitoxin. This implied that the 4-position on the third digitoxose is sufficiently distant from the aglycone that one might expect “exo” labeling”; that is, probing the immediate environment of the binding site by placing the reactive group (in this case, a carbene) at a suitable distance from that portion of the molecule involved in the primary binding site (the aglycone and probably the first sugar of the glycoside).
It was found that the 4”’-diazomalonyl digitoxin was as potent as digitoxin in inhibiting the (Na, K)-ATPase activity (FIGURE 6). The Iso for 4”‘-diazomalonyl digitoxin was found to be 4 x lo-’ M and, for digitoxin, 8 x lo-’ M. The binding isotherms determined for ’H-4”‘-diazomalonyl digitoxin and ’H-digitoxin were very
FIGURE 6. Inhibition of (Na, K)-ATPase with 4-diazomalonyl digitoxin and digitoxin. Eel enzyme (6 fig/tube) was mixed with various concentrations of photolabel and digitoxin and incubated 30 min at room temperature in 1 ml of 1% DMSO, 60 mM NaCI, 2.0 m M MgCI2, 210 m M NalATP. 0.1 m M EGTA, and 92 m M Tris (pH 7.4). After 30 min, 0.1 ml of the same medium was added to the minus K’ tubes, and 0.1 ml of the same medium containing 53 m M KCI was added to the plus K’. The tubes were allowed to incubate 4 min at 38 OC. after which 1 ml of 10% TCA was added to stop the reaction. DMSO alone was added to the control tube with no photolabel. The difference in inorganic phosphate production between plus and minus K’ at a given photolabel concentration was used to calculate the enzyme activity. 0: 4-diazomalonyl digitoxin; 0: digitoxin.

98 Annals New York Academy of Sciences
similar. The value estimated for the parent digitoxin and 'H-photolabel in the type I complex was 8-10 x lo-* M.
The stability of the type 1 enzyme-photolabel complex was compared to the stability of the type 1 enzyme-ouabain complex at 4 O C . As shown by Yoda,'* rhamnosides such as ouabain possess the slowest dissociation rate of the common glycosides used as inhibitors of the (Na, K)-ATPase. 'H-ouabain binding was stable for a t least 2 h at 4 O C when complexed to the purified eel enzyme. Similarly, within 2 h a t 4 "C an insignificant amount of 'H-4"'-diazomalonyl digitoxin had dissociated from the binding site. In all binding experiments, nonspecific binding of 'H- 4"'-diazomalonyl digitoxin never exceeded 5% of the total binding.
FIGURE 7 (upper panel) shows the photolytic labeling profile obtained for the type I photolabel-enzyme complex after sodium dodecylsulfate (SDS) disc gel electropho- resis. The 4-diazomalonyl derivative demonstrated ouabain protectable incorporation
Slke number
FIGURE 7. The 'H-4-diazomalonyl digitoxin labeling pattern of eel enzyme on SDS polyacrylamide gels.
Upper panel (type I complex): 0: 1.5 x M photolabel and ouabain lo-' M; .: 3 x M photolabel prephotolyzed for 10 s and then bound to the enzyme under type I conditions.
Lower panel (type I1 complex): 0: 8.3 x lo-' M photolabel; 0: 8.3 x lo-' M photolabel and ouabain lo- ' M. The slash at slice 25 indicates a change to the right hand side ordinate. Binding of the 4"'-derivative was more than 95% specific.
M photolabel; 0: 1.5 x M photolabel not photolyzed; 0 : 3 x

Ruoho & Hall: Ouabain Binding S i t e 99
into the (r subunit of the enzyme, as previously noted for N A P glycyl digitoxigenin and other photo label^.^^ However, specific incorporation into the 13 subunit was also noted with this compound. This incorporation was not detected if the enzyme-ligand complex was not photolyzed or i f the ligand was prephotolyzed before being added to the enzyme. These data support the conclusion that 4”’-diazomalonyl digitoxin is a photoaffinity label for the cardiac glycoside binding site.
The absolute amount of photoincorporation varied, in the a subunit, from 1 to 5% and, in the /3 subunit, from 0.5 to 2%. Photolytic cross-linking of the enzyme was confined entirely to the a subunit, as estimated by Coomassie staining.26 No major cross-linking of the @ subunit could be detected. I t is likely that a portion of the radioactivity that did not penetrate the gel was photoincorporated into the a subunit in a cross-linked state. Since the tritium in the peak region of the dye marker (slice 40) was also present in the prephotolyzed and unphotolyzed dark controls, we conclude that it was mainly unreacted photolabel.
A similar photoincorporation pattern was detected with the type I I photolabel complex of the enzyme (FIGURE 7, bottom panel). Again, covalent labeling could readily be detected in the (3 subunit as well as in the a subunit, suggesting that these two complexes (type I and type 11) of cardiac glycoside with the enzyme probably do not involve gross conformational dissimilarities, although subtle conformational changes probably do occur, as evidenced by different dissociation rates of cardiac glycosides from the type I and type I I complexes.” Cross-linking of the a subunit also routinely reduced the absolute amount of radioactivity detected at this position on the gels. No cross-linking of the p subunit could be detected.
It was noted that, for both complexes, a radioactive peak could be reproducibly detected on the gels a t slice 32 (FIGURE 7). Although poorly resolved from the broad unreacted photolabel peak at the bottom of the gel, this peak of radioactivity was consistently on the gels. Utilizing soluble standard protein markers (data not shown), it was determined that this peak corresponded to a molecular weight of approximately 12.000, which is the size estimated for a proteolipid component reported to be photolabeled in the enzyme purified from pig kidney2’ and from electric eeLz4 Further experiments, however, are needed to clarify the labeling with the diazomalonyl digitoxins in this region of the gel.
3”’-Diazornalonyl Digitoxin
This compound was prepared by very facile acyl migration of the diazomalonyl group, as was described by Satoh and Morita for acetyl derivatives.” Since the 4-hydroxyl of digitoxin is equatorial, the 3“’-hydroxyl is axial,” and the digitoxose rings are held in the preferred C-l (D) configuration,” substituents on the 4“’- hydroxyl and 3”’-hydroxyl will be projected in the binding site in significantly different directions. I f the residues on the p subunit that interact with the carbene generated at the 4”’-position are accessible to the carbene generated at the 3”’-position, then the labeling patterns between the 4“’ - and 3”’ -isomers should be at least qualitatively similar. I f . on the other hand, a very specific “cut off” exists in this region of the binding site, then it is possible that labeling of the 0 subunit would be either undetected or greatly reduced. I n fact, when the enzyme-3H‘3”‘-diazomalonyl digi-

100 Annals N e w York Academy of Sciences
Slice number
FIGURE 8. The 3H-3”’-diazomalonyl digitoxin labeling pattern of eel enzyme on SDS
Upper panel (type I complex): 0: 5.5 x lo-’ M photolabel; 0: 5.2 x lo-’ M photolabel and
Lower panel (type 11 complex): 0: 5.27 x lo-’ M photolabel; 0 : 2.2 x lo-’ M photolabel and
polyacrylamide gels.
lo-’ M ouabain.
lo-’ M ouabain. Binding of 3”’-derivative was greater than 98% specific.
toxin complex was photolyzed, label could again be detected in the subunit as well as in the a subunit ( F I G U R E 8). Covalent labeling efficiencies were similar to those found for the ’H-4”’-derivative. Since light flux and binding site occupancy varied, it was not possible to make accurate quantitative comparisons concerning covalent incorporation between these isomers in these experiments.
These data indicate that, within a very short distance in the “sugar ~pecific”~’ binding site region, type I and type I I complex &chain residues and a-chain residues are within at least 15 A of each other.
DISCUSSION
Four light-activatable photolabels for (Na, K)-ATPase have been prepared. Two of these compounds have been prepared as tritiated derivatives a t specific radioactiv- ities ranging from 7.4 to 12.2 Ci/mmol. The nitrophenylazido derivatives of

Ruoho 8c Hall: Ouabain Binding Site 101
digitoxigenin and strophanthidin retain the same potency for inhibition of the enzyme as the aglycone, digitoxigenin. Specific binding to both type I and type I I complexes was demonstrated utilizing radiopure tritiated N A P glycyl digitoxigenin ( 12.2 Ci/mmol). Both of these complexes were prepared with approximately 1 mole of cardiac glycoside/mole of enzyme. Photolysis of the enzyme-photolabel complex resulted in ouabain protectable covalent incorporation only into the 100,000 dalton polypeptide (FIGURE 5). Specific labeling was not detected in the glycoprotein (8) subunit regardless of the time of photolysis. This is consistent with the results of the use of other photolabels2’-26 on the ATPase, in which either the photoactivatable group extends to the general region of the second sugar residue (as estimated from molecular models) or the photoactivatable group is attached to the A/B ring junction of the agly~one.~’ No detectable photolytic cross-linking of the 8 subunit occurred in these photolysis experiments. This is an important observation, since one could argue that the lack of tritium in the region of the B subunit is due to the removal of this subunit to the top of the gel. Some photolytic cross-linking of the a subunit was found to occur, however, and this accounts for the radioactivity found at the top of the gel. The experiments performed with ’H-NAP glycyl digitoxigenin were designed to focus exclusively on the a and (3 subunits. No attempt was made to scrutinize the gels in the region of the dye marker for the labeling of the proteolipid reported by Forbush et af.*’ and Rogers and L a ~ d u n s k i . ~ ~ At this point, it can be concluded from our data and the data of that the a subunit contains the primary aglycone portion of the ouabain binding site. The j3 subunit is not directly involved in binding either the aglycone or the first and second sugars of the glycosides.
Based on the experiments with 4-diazomalonyl digitoxin and 3”‘-diazomalonyl digitoxin, it can be concluded further that the p subunit does reside close to the “sugar-specific” site. Yoda has shown that specific interactions, presumably in the form of hydrogen bonds, occur with the hydroxyls of the digitoxose portion of digit~xin.’~ This interaction has been shown to stabilize the steroid-enzyme complex by, primarily, decreasing the dissociation rate. From acetylation studies, Yoda and Yoda concluded that the 4“‘ hydroxyl was not involved in the formation of a stable cardiac glycoside-enzyme ~omplex .~’ The 3’ and 3”’ hydroxyls were involved to varying degrees in the type I and type I I complexes. Acetylation of the 3’ hydroxyl greatly increased the dissociation rate of digitoxin in the type I complex, suggesting that this hydroxyl may be involved in a binding reaction crucial to the maintenance of the stability of this complex. If the hydroxyl a t the 4 position is not integrally involved in hydrogen bonding to the protein surface, it might be expected that the reactive carbene will be “exo” labeling and thus probing the immediate environment of the sugar-specific site.3’ It is possible that the area accessible to the carbene at the 4”’ position contains large amounts of water. This may account partially for the low levels of incorporation, since the insertion of carbenes into water would serve as a competing reaction.20 Another possible side reaction of the carbene is insertion into the oxygen of the 3”’ hydroxyl. resulting in a stable six-membered ring. This possibility has not been experimentally investigated by, for instance, analyzing the photolysis products of 4”‘-diazomalonyl digitoxin or 3”’-diazomalonyl digitoxin.
subunit (FIGURE 7) with the 3H-4-diazomalonyl digitoxin was specific and did not occur if the enzyme-photolabel complex was not photolyzed. This subunit was labeled in both type I and type 11 complexes. It is likely
Photoincorporation into the

102 Annals New York Academy of Sciences
that the labeling of the a subunit occurs in an area different from that observed with NAP glycyl digitoxigenin, since the photoactive groups are a t quite different positions relative to the aglycone portion of the molecule. The tritium at the bottom of the gel (FIGURE 7) is probably unreacted photolabel, since a peak in this region of the gel is seen in the dark control. It is possible that some portion of the photolabel has inserted into the lipid that borders the sugar-specific site.
Our results are consistent with the hypothesis that the a subunit is the locus of the primary cardiac glycoside binding site. However, the secondary area of the binding site, which interacts with the sugars, borders closely with some portion of the B subunit. This implies that, in the region of the sugar-specific site, the a and @ subunits of the detergent-purified (Na, K)-ATPase of Electrophorus electricus are very intimately associated. If one compares structures utilizing molecular models of N A P glycyl digitoxigenin. nitroazidobenzoyl-ouabain (NAB-o~abain) ,~’ NAP-~uabain , ’~ and 4”‘-diazomalonyl digitoxin, the diazo group of 4-diazomalonyl digitoxin extends approximately 10 A further from the aglycone than does the azido group of N A P glycyl digitoxigenin and 5 A further than the azido groups of NAB-ouabain and NAP-ouabain. Since specific labeling of the B subunit could not be detected with NAP glycyl digitoxigenin but did occur with 4-diazomalonyl digitoxin, /3 subunit residues must border the region of the third digitoxose. This p subunit labeling cannot be detected with reactive groups placed in the region of the second sugar. Forbush et al. reported some incorporation (1%) into the @ subunit with NAB-ouabain in which the photoactive azide extends to the third glycosidic bond.” This labeling probably reflects exo labeling of the @ subunit, also, but it is difficult to make a comparison with the 4”‘-diazomalonyl digitoxin labeling because the microenvironment of the hydro- phobic azidophenyl group may be much different from that of the more polar diazomalonyl group.
Our results show that there is a possibility of systematically studying the sugar-specific region of the binding site by using 4 - , 4”-, and 4‘-diazomalonyl (or other photoactivable) derivatives of digitoxin. These compounds are synthesized by deriving the terminal equatorial hydroxyl of digitoxin, bisdigitoxoside, and monodigi- toxoside.” Acyl migration of the diazomalonyl group to the 3”’ and 3” positions will allow the facile preparation of additional compounds to probe the site. This series of compounds will be useful because the same photoactive group can be compared at different positions on the molecule. The smaller diazomalonyl functional group is also less likely to produce those local perturbations in structure that may occur with the bulkier hydrophobic nitrophenylazido group.
REFERENCES
I . SCHATZMANN. H. J. 1953. Helv. Physiol. Pharmacol. Acta 11: 346. 2. GLYNN. 1. M. 1964. Pharmacol. Rev. 1 6 3 8 1 . 3. AKERA. T. 1977. Science 198 569. 4. PETERS, T., R. H. RABEN & 0. WASSERMAN. 1974. Eur. J. Pharmacol. 26: 166. 5. SCHWARTZ. A., G. E. LINDENMAYER & J. c. ALLEN. 1975. Pharmacol. Rev. 27: 3. 6. KYTE. J. 1972. J. Biol. Chem. 24’7: 7642. 7. HOKIN. L. E., J. L. DAHL. J. D. DEUPREE. J. F. DIXON. J. F. HACKNEY & F. PERDUE. 1973.
J. Biol. Chem. 248: 2593.

Ruoho & Hall: Ouabain Binding Site 103
8.
9. 10.
1 1 . 12.
13. 14. 15. 16.
17. 18.
19.
20. 21.
22. 23. 24. 25. 26. 27. 28. 29. 30. 31.
32.
LANE. L. K., J. H. COPENHAVER. G. E. LINDENMAYER & A. SCHWARTZ. 1973. J. Biol.
JORGENSEN. P. L. 1974. Biochim. Biophys. Acta 3 5 6 53. PETERSON. G. L., R. D. EWING. S. R. HOOTMAN & F. P. CONTE. 1978. J. Biol. Chem.
HASTINGS. D. F. & J. A. REYNOLDS. 1979. Biochemistry 18 817. ALBERS. J. W. 1976. In The Enzymes of Biological Membranes, Vol. 3. A. N. Martonosi,
DAHL. J. & L. E. HOKIN. 1974. Annu. Rev. Biochem. 4 3 327. MARTSUI. H. & A. SCHWARTZ. 1968. Biochim. Biophys. Acta 151: 655. SCHWARTZ. A,. H. MATSUI & A. H. LAUGHTER. 1968. Science 160: 323. VAN WINKLE, W. B., J. C. ALLEN & A. SCHWARTZ. 1972. Arch. Biochem. Biophys.
YODA. A. 1973. Mol. Pharmacol. 9 51. SCHWARTZ. A.. G. E. LINDENMAYER. J. C. ALLEN & J. L. MCCANS. 1974. Ann. N.Y.
RUOHO. A. E.. L. E. HOKIN, R. J. HEMINGWAY & S. M. KUPCHAN. 1968. Science
KNOWLES. J. R. 1972. Acc. Chem. Res. 5 155. JAKOBY. W. B. & M. WILCHEK. In Methods in Enzymology, Vol. 46. Academic Press.
TOBIN. T., T. AKERA. T. M. BROSDY & H. R. TAREJA. 1976. Eur. J . Pharmacol. 3 5 6 9 . FORBUSH, B., J. KAPLAN & J . F. HOFFMAN. 1978. Biochemistry 117: 3667. ROGERS. T. B. & M. LAZDUNSKI. 1979. Biochemistry 18 135. ROGERS. T. B. & M. LAZDUNSKI. 1979. FEBS Lett. 98 373. RUOHO. A. & J . KYTE. 1974. Proc. Nat. Acad. Sci. USA 71: 2352. GARRAHAN. P. J . & 1. M. GLYNN. 1967. J . Physiol. 192 159. FLET-T, G. W. J., J. R. KNOWLE~ & R. R. PORTER. 1972. Biochem. J . 128499. YODA. A. & S. YODA. 1975. Mol. Pharmacol. 11: 653. SATOH. D. & J. MORITA. 1969. Chern. Pharm. Bull. 17: 1456. BAKER. B. R. 1967. In Design of Active Site-Directed Irreversible Enzyme Inhibitors:
YODA. A. 1974. Ann. N.Y. Acad. Sci. 242: 598.
Chem. 248: 7197.
2 5 3 4762.
Ed.: 283-301. Plenum Press. New York.
151: 85.
Acad. Sci. 2 4 2 577.
1 5 9 1354.
London.
17-22. John Wiley and Sons. New York.
33. SESSA. G. & G. WEISSMANN. 1970. J. Biol. Chem. 2453295.




















