
New urinary EPO drug testing method using two-dimensional gel electrophoresis

Characterization of bacterial and fungalcommunities in composted biosolids overa 2 year period using denaturing gradient gelelectrophoresis
Amy Novinscak, Nadine J. DeCoste, Celine Surette, and Martin Filion
Abstract: Composting is a microbial process that converts organic waste into a nutrient-rich end product used in horticul-tural and agricultural applications. The diversity and long-term succession of microorganisms found in composted biosolidshas been less characterized than other composts. In this study, bacterial and fungal communities found in composted bio-solids aging from 1 to 24 months were studied using denaturing gradient gel electrophoresis (DGGE) and sequencing. Theresults revealed high levels of diversity, where 53 bacterial species belonging to 10 phyla and 21 fungal species belongingto 4 phyla were identified. Significant differences were observed when comparing the bacterial DGGE patterns of youngcompost samples, whereas no differences were observed in samples over 8 months. For fungal patterns, no significant dif-ferences were observed during the first 4 months of composting, but the diversity then significantly shifted until 24 months.The results indicate that patterns of bacterial species vary during the first few months of composting, whereas fungal pat-terns generally vary throughout the whole process, except during early stages. The description of the main microbialgroups found in composted biosolids could find various applications, including the discovery of biotechnologically relevantmicroorganisms and the development of novel markers allowing quantitative monitoring of key microorganisms.
Key words: bacteria, biosolids, compost, DGGE, fungi.
Resume : Le compostage est un processus microbien qui convertit des dechets organiques en produits riches en nutrimentsutilises en horticulture et en agriculture. La diversite et la succession a long terme de microorganismes retrouves dans lesbiosolides de compostage ont ete moins bien caracterisees que chez d’autres types de composts. Dans cette etude, les com-munautes bacteriennes et fongiques de biosolides de compostage ages de 1 a 24 mois ont ete etudiees par electrophoresesur gel en gradient denaturant (DGGE) et par sequencage. Les resultats ont revele un haut degre de diversite alors que53 especes bacteriennes appartenant a 10 phyla, et 21 especes fongiques appartenant a 4 phyla ont ete identifiees. Des dif-ferences significatives ont ete observees lorsque les patrons bacteriens obtenus par DGGE a partir de jeunes echantillonsde compost etaient compares les uns aux autres, tandis qu’il n’y avait plus de difference parmi les echantillons ages deplus de 8 mois. Il n’y avait pas de differences significatives entre les patrons obtenus de champignons au cours de 4 pre-miers mois de compostage, mais la diversite augmentait ensuite significativement jusqu’au 24ieme mois. Les resultats indi-quent que les patrons des especes bacteriennes varient durant les premiers mois de compostage alors que les patronsfongiques varient tout au long du processus, a l’exception des etapes precoces. La description des principaux groupes mi-crobiens trouves dans les biosolides de compostage pourrait trouver differentes applications notamment pour la decouvertede microorganismes pertinents d’un point de vue biotechnologique et pour le developpement de nouveaux marqueurs per-mettant de suivre quantitativement des microorganismes cles.
Mots-cles : bacteries, biosolides, compost, DGGE, champignons.
[Traduit par la Redaction]
Introduction
Solid organic waste obtained from wastewater treatment,also known as biosolids, are a source of concern for publichealth and the environment because they often contain hu-man pathogenic microorganisms and undesirable chemicals,
including trace metals and pharmaceutical products (Du-montet et al. 1999). Biosolids can be eliminated by inciner-ation, direct land application, burying in landfills, andcomposting (Hoitink and Fahy 1986). Of these methods,composting seems the most viable alternative because ofnegative environmental issues associated with other disposal
Received 20 August 2008. Revision received 26 November 2008. Accepted 11 December 2008. Published on the NRC Research PressWeb site at cjm.nrc.ca on 20 April 2009.
A. Novinscak, N.J. DeCoste, and M. Filion.1 Department of Biology, Universite de Moncton, 165 Massey Ave, Moncton, NB E1A 3E9,Canada.C. Surette. Department of Chemistry and Biochemistry, Universite de Moncton, 165 Massey Ave, Moncton, NB E1A 3E9, Canada.
1Corresponding author (e-mail: [email protected]).
375
Can. J. Microbiol. 55: 375–387 (2009) doi:10.1139/W08-152 Published by NRC Research Press
methods (US EPA 1999). Composting is a microbial-mediated transformation that converts organic waste intostabilized humified products (Finstein and Morris 1975).Composted biosolids can be beneficial in agricultural, hor-ticultural, or land reclamation applications as the end prod-uct is rich in nutrients and comparable to commerciallyproduced fertilizers. The application of compost to soil hasshown additional beneficial effects, such as the suppressionof certain plant pathogens (Boulter et al. 2002). Compost-ing also releases less CO2 than incineration, which is bene-ficial for the environment (Ishii et al. 2000).
Identification of key microorganisms involved in the com-posting of biosolids can lead to the development of more ef-ficient composting practices (Herrmann and Shann 1997),which can, for example, alleviate odors associated withcomposted biosolids production or optimize the ratio of theinitial substrates used, reduce processing time and achievehigher quality end products (Finstein and Morris 1975; Ivorset al. 2000). Because of the particular nature of the sub-strate, a detailed survey of the microorganisms present andtheir role in the process could also lead to specific applica-tions, such as bioprospecting for microbes or enzymes usefulin biotechnological, industrial, and environmental applica-tions (Tyagi et al. 2002).
The microbial species present during composting areknown to vary according to different temperature-dependentphases (Finstein and Morris 1975). During the first fewhours to days, bacteria and mesophilic/thermotolerant fungiare the principal active degraders of the organic waste andtheir metabolic activity leads to a rapid increase in temper-ature (Ryckeboer et al. 2003b). The total diversity decreasesas the temperature increases, but the diversity of thermoto-lerant and thermophilic bacteria and fungi increases duringthis period (Finstein and Morris 1975). During the maturingphase, various mesophilic microorganisms recolonize or col-onize de novo the compost (Boulter et al. 2000). To date,most of the studies assessing the microbial diversity ob-served during composting, and especially for composted bio-solids, have only been performed during the first few weeksof the process (Ishii et al. 2000; Peters et al. 2000) and have
mainly concentrated on prokaryotic organisms (Nakasaki etal. 1985; Peters et al. 2000; Ishii and Takii 2003). The di-versity of microorganisms present during later phases ofcomposting is not well characterized (Ishii et al. 2000).
Traditionally, studies investigating the microbial diversitypresent in composts have mainly relied on the use of cul-ture-dependent methods (Kane and Mullins 1973; Nakasakiet al. 1985; Strom 1985). Although very useful, these meth-ods are known to detect a small portion of the total micro-bial community present in environmental samples; as littleas 5% has been reported for sewage sludge (Wagner et al.1993). Because of this drawback, recent studies are nowmainly relying on culture-independent methods (Hellmann etal. 1997; Herrmann and Shann 1997; Ishii et al. 2000;Peters et al. 2000; Dees and Ghiorse 2001). Phospholipidfatty acid analysis (PLFA) has been successful at describ-ing microbial community changes occurring over short pe-riods of time in composts, but the profiles obtained tend toonly represent broad microbial or well-characterized groups(Herrmann and Shann 1997). Denaturing gradient gel elec-trophoresis (DGGE) targeting taxonomical relevant molecu-lar markers, such as the 16S rRNA gene in bacteria andthe 18S rRNA gene or the intergenic transcribed spacers(ITS region) in fungi, has also been used to describe themicrobial diversity in composts (Gardes and Bruns 1993;Heuer et al. 1997; Anderson et al. 2003). The microbialdiversity found using this approach was greater than thatfound using other approaches (Muyzer et al. 1993; Wagneret al. 1993; Beffa et al. 1996). DGGE is also well adaptedto study the genetic diversity of complex microbial com-munities found in different environments (Muyzer et al.1993; Muyzer 1999; de Lipthay et al. 2004). Very fewstudies have used DGGE or other culture-independentmethods for studying the microbial diversity found in com-posted biosolids (Ueno et al. 2001; Ishii and Takii 2003).These studies have only concentrated on the bacterial spe-cies present or on the first few weeks of composting,which only gives a partial representation of the total mi-crobial diversity present in such an environment. Addi-tional research addressing the presence of both prokaryotic
Table 1. Variations in the physical and chemical parameters of the composted biosolids sampled in this study.
Compost age (months)
Physical or chemical parameter 1 4 8 13 18 24Temperature range (8C) 63–79 45–64 34–42 19–28 19–27 19–28pH 7.7 7.4 7.8 7.4 7.4 7.1Dry matter (%) 41.8 28.4 35.5 36.3 38.0 40.9C/N ratio 23.7 20.2 17.6 19.5 18.2 21.6Carbon (%) 14.3 9.9 10.6 10.9 12.4 10.7Nitrogen (%) 0.6 0.5 0.6 0.6 0.7 0.5Phosphorus (%) 0.2 0.1 0.3 0.3 0.2 0.3Potassium (%) 0.1 0.1 0.1 0.1 0.1 0.1Calcium (%) 1.2 0.9 1.4 2.2 1.3 1.4Magnesium (%) 0.1 0.1 0.2 0.2 0.2 0.2Copper (ppm) 36.3 43.7 64.8 70.6 52.4 64.9Zinc (ppm) 81.3 69.7 118.0 110.7 102.6 124.3Boron (ppm) 2.5 2.0 2.0 2.2 3.0 2.8Iron (ppm) 5558.9 3933.1 6763.9 6450.2 5954.1 8802.5Manganese (ppm) 311.7 270.7 345.4 389.9 400.6 377.2
376 Can. J. Microbiol. Vol. 55, 2009
Published by NRC Research Press

and eukaryotic microorganisms found in composted biosol-ids over longer periods of time is clearly needed.
In this study, a culture-independent PCR–DGGE and a se-quencing approach were combined to compare the bacterialand fungal communities in composted biosolids samplesvarying in age, for a period of up to 24 months. The 16SrRNA gene and the ITS region were targeted, respectively,as bacterial and fungal taxonomical relevant markers.
Materials and methods
Composted biosolids sampling and DNA extractionSamples of composted biosolids were obtained from a pri-
vate composting facility located in South Branch, NewBrunswick. The biosolids originated from a wastewatertreatment plant handling residential and industrial waste forthe Greater Moncton region, with a population of~125 000 habitants. The biosolids were dewatered and lime-treated before being used for composting. All windrow pilessampled were composed of 1:1 biosolids from this waste-water treatment plant and wood chips and bark obtainedfrom a wood processing plant. Six different windrow piles(approximately 15 m � 3 m � 3 m) aged, respectively,1 month, 4 months, 8 months, 13 months, 18 months, and24 months were sampled in July 2005 and 5 samples weretaken from each pile, varying in depth from 50 to 100 cm.Each sample consisted of a mixture of 5 subsamples takenfrom different locations on each pile to take into accountthe possible spatial distribution of the microorganisms. Thetemperature of each subsample from every pile was meas-ured using a thermometer (Table 1). Chemical analysis (pH,% dry matter, C/N ratio, % carbon, % nitrogen, % phospho-rus, % potassium, % calcium, % magnesium, copper (ppm),zinc (ppm), boron (ppm), iron (ppm), and manganese (ppm))were performed for each sampling date (Soil and Feed Test-ing Laboratory, Charlottetown, Prince Edward Island)(Table 1). The 30 samples were placed on ice until arrivalat the laboratory. Each sample was frozen then lyophilizedfor 24 h in a ModulyoD freeze dryer (Thermo Electron Cor-poration, Nepean, Ontario) and stored at –80 8C. GenomicDNA extractions were performed using the Mobio Power-Soil DNA extraction kit (Mobio Inc., Carlsbad, California).All DNA samples were diluted 1:2 in sterile ddH2O beforeproceeding to PCR amplifications.
Fig. 1. Denaturing gradient gel electrophoresis (DGGE) patterns ofthe 16S rDNA gene fragments amplified by PCR from differentcomposted biosolids samples. The first and last lane of each gel(lanes 1, 12, 13, 24, 25, and 36) represents a bacterial DNA laddercomprised of 16S rDNA fragments PCR amplified from Proteusvulgaris, Lactobacillus acidophilus, Pseudomonas fluorescens, andKlebsiella pneumonia. Lanes 2–6 represent the 1 month compostsamples, lanes 7–11 represent the 4 month samples, lanes 14–18represent the 8 month samples, lanes 19–23 represent the 13 monthsamples, lanes 26–30 represent the 18 month samples, and lanes31–35 represent the 24 month samples. Bands that were success-fully sequenced are numbered. Relatedness of the DGGE patternsof the 6 compost ages and their replicates are displayed using theunweighted pair-group method using arithmetic averages (UPGMA)dendrogram and Pearson’s correlation coefficient (R).
Novinscak et al. 377
Published by NRC Research Press
PCR amplificationPCR amplifications of the bacterial 16S rRNA gene were
performed using primers F984-GC and R1378 as previouslydescribed (Heuer et al. 1997). The fungal ITS region wasamplified using a nested approach and 2 primers sets con-sisting of EF4 (Smit et al. 1999) and ITS4 (White et al.1990) followed by ITS1F-GC (Gardes and Bruns 1993) andITS2 (White et al. 1990) as previously described (Andersonet al. 2003). Primers F984-GC and ITS1F-GC had a 40 bpGC-clamp at their 5’ end. All amplifications contained 1�ThermoPol reaction buffer (New England Biolabs, Picker-ing, Ontario), 1.5 mmol/L of MgCl2, 0.5 mmol/L of each pri-mer (Alpha DNA, Montreal, Quebec), 200 mmol/L of dNTP(New England Biolabs), 1.25 units of Taq DNA polymerase(New England Biolabs), and sterile ddH2O in a final volumeof 50 mL. For the first PCR amplification round of fungalDNA, 1 mL of 10 mg/mL BSA (New England Biolabs) wasadded per reaction. The cycling protocol for the 16S rRNAgene consisted of an initial denaturation at 94 8C for 5 minfollowed by 35 cycles of denaturation at 94 8C for 1 min,annealing at 55 8C for 1 min, and extension at 72 8C for2 min followed by a final 5 min extension at 72 8C. For thefungal ITS region, the initial cycling protocol consisted ofan initial denaturation at 94 8C for 5 min followed by40 cycles of denaturation at 94 8C for 30 s, annealing at55 8C for 30 s, and extension at 72 8C for 1 min followedby a final 5 min extension at 72 8C. The amplification reac-tion was then diluted 1:10 before performing the secondPCR amplification. The nested PCR protocol consisted ofan initial denaturation at 94 8C for 5 min followed by35 cycles of denaturation at 94 8C for 30 s, annealing at55 8C for 30 s, extension at 72 8C for 30 s, and a final ex-tension of 5 min at 72 8C. The molecular sizes of the ampli-
fied fragments were checked by electrophoresis on a 1.5%agarose gel with 1� TAE buffer. The gels were stained for10 min in ethidium bromide, destained for 10 min in dis-tilled water, and then visualized and the images captured us-ing a Kodak ID Gel Logic 200 (Kodak Inc., Rochester, NewYork). Samples were purified using the MinElute PCR puri-fication kit (Qiagen, Missisauga, Ontario) before being usedfor DGGE analysis.
Construction of DNA ladders for DGGE analysisDNA ladders were constructed for the DGGE analysis of
bacterial and fungal DNA. Genomic DNA from the bacteriaProteus vulgaris, Lactobacillus acidophilus, Pseudomonasfluorescens, and Klebsiella pneumonia was extracted frompure cultures and amplified by PCR using the previouslymentioned protocol. PCR reactions were purified using theMinElute PCR purification kit (Qiagen) before being com-bined and used in DGGE gels as DNA ladders. FungalDNA ladders were constructed similarly to the bacterial lad-ders using DNA extracted from Epicoccum nigrum, Scleroti-nia sclerotiorum, and Phoma pomorum.
DGGE analysesDGGE analyses were performed using a DCode Universal
mutation detection system (Bio-Rad, Hercules, California).Six percent (m/v) polyacrylamide (37.5:1 acrylamide–bisacrylamide) gels were prepared with a 40%–70% ureaand formamide linear gradient for bacterial amplicons anda 25%–55% linear gradient for fungal amplicons (100%denaturant consisting of 7 mol/L urea and 40% (v/v) for-mamide). A total of 10 mL of PCR product was loadedinto each lane of the gel and electrophoresis was per-formed in 1� TAE buffer for 16 h at 60 8C and 80 V for
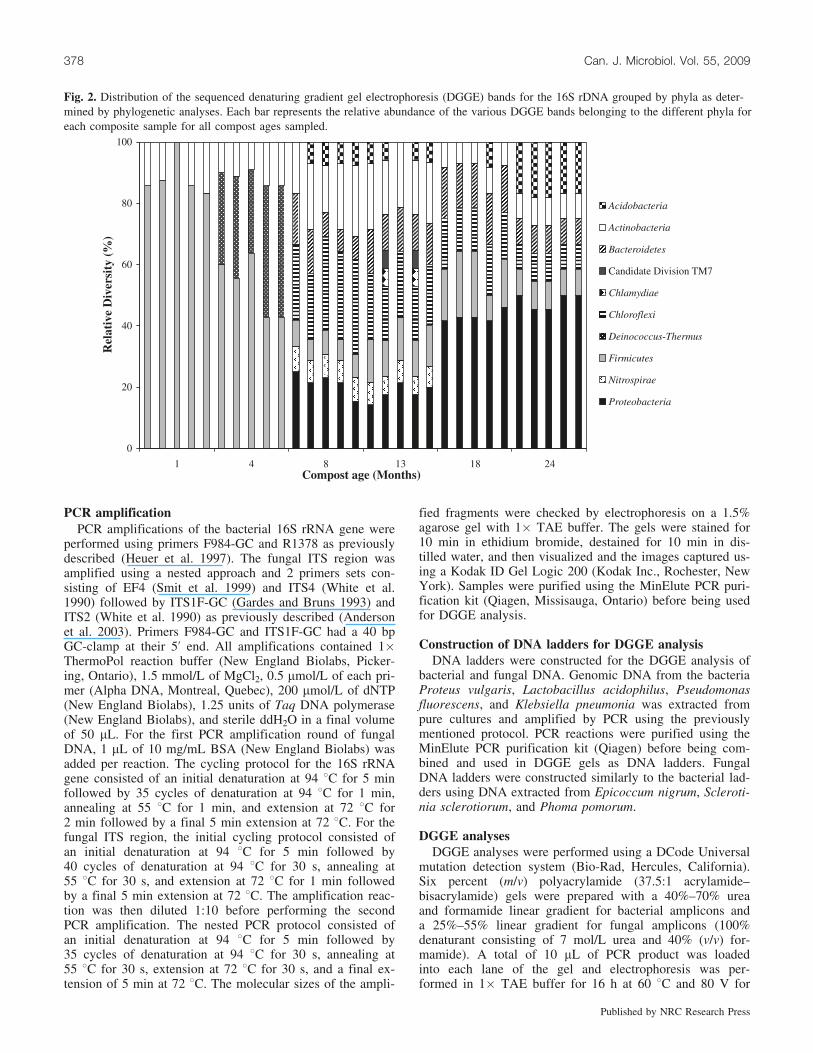
Fig. 2. Distribution of the sequenced denaturing gradient gel electrophoresis (DGGE) bands for the 16S rDNA grouped by phyla as deter-mined by phylogenetic analyses. Each bar represents the relative abundance of the various DGGE bands belonging to the different phyla foreach composite sample for all compost ages sampled.
378 Can. J. Microbiol. Vol. 55, 2009
Published by NRC Research Press

bacterial amplicons or 60 V for fungal amplicons. Follow-ing electrophoresis, gels were stained for 10 min in ethi-dium bromide and destained for 10 min in distilled water.The gels were visualized and images captured using aKodak ID Gel Logic 200 (Kodak Inc.).
GelCompar II (version 5.10) (Applied Maths, Austin,Texas) was used to analyze the fingerprints in each gel aspreviously described (Rademaker and de Bruijn 2008).Pearson’s correlation coefficient (R) was used to create asimilarity matrix and the unweighted pair-group method us-
Fig. 3. Unrooted neighbour-joining tree representing the phylogenetic relationship of the 53 cloned 16S rRNA denaturing gradient gel elec-trophoresis (DGGE) bands to the most closely related identified sequences obtained from BLASTn searches. Letters following cloned 16SrRNA DGGE bands represent the compost age groups in which the bands were present. The letter A represents 1 month; B, 4 months; C,8 months; D, 13 months; E, 18 months; and F represents 24 months of composting. The numbers at the nodes of the tree are bootstrapvalues for each node with 100 bootstrap resampling (values <50 are omitted). The bar represents 0.1 substitutions per site.
Novinscak et al. 379
Published by NRC Research Press
ing average linkages (UPGMA) was used to calculate theclustering of the band patterns.
For comparison between the ages of compost samples, thenatural variability (similarity of DGGE patterns within atreatment) and the variability between samples were com-pared as described by Smalla et al. (2001). If the medianvalues of 2 treatments differed more than expected from nat-ural variability (if there is no overlap of the interquartileranges), then a significant effect was assumed.
Cloning and sequencing of DGGE bandsFor each sampling date, 1 sample of genomic DNA corre-
sponding to the best representation of the diversity found inthe DGGE band profiles for that sample was used to amplifythe 16S rRNA gene or the ITS region, using primers 968Fand R1378 or primers ITS1F and ITS2, respectively. Primers968F and ITS1F are identical to F984-GC and ITS1F-GCbut do not have GC clamps at their 5’ end. The PCR reac-tions were purified using the MinElute PCR purification kit(Qiagen) and shotgun cloned into Top10 Escherichia colicells using the TOPO-TA Invitrogen cloning kit (InvitrogenInc., Burlington, Ontario). Ninety-six clones per samplewere selected and grown overnight in LB broth supple-mented with ampicillin (10 mg/mL) before performing plas-mid DNA extractions using the Direct Prep 96 Miniprepplasmid extraction kit (Qiagen). The cloned fragments werethen reamplified using the initial primer sets (F984GC–R1378 and ITS1FGC–ITS2), and the fragments were run onDGGE gels with the genomic DNA samples used to gener-ate the clone library. Plasmid DNA of each clone that corre-sponded to a band in the original genomic DNA sample wassubmitted to direct sequencing using the Big Dye terminatorsequencing kit version 3.1 (Applied Biosystems, Foster City,
California) and primer R1378 (for bacteria) or primer ITS2(for fungi) on a 3130xl ABI Analyzer (Applied Biosystems).
Sequence analysis and nucleotide sequence accessionnumbers
DNA sequences were edited using BioEdit version 7.0.4.1(Hall 1999). Similarity searches comparing nucleotide se-quences with the GenBank database were performed usingthe basic local alignment search tool (BLASTn), and themost homologous sequences of identified species were used(when available) to construct multiple-sequence alignmentsusing ClustalX (version 1.83) (Altschul et al. 1990). Neigh-bour-joining analyses were performed using PAUP (version4.0b10) with 100 bootstrap replicates. Phylogenetic treeswere edited using Treeview (version 1.6.6). The nucleotidesequences identified in this study have been deposited inthe National Center for Biotechnology Information (NCBI)database under accession Nos. EU096239–EU096312.
ResultsAll data (except temperature) for chemical parameters of
the composted biosolids samples under study were fitted toregression models using the Curve Fit setting of SPSS(SPSS Inc., Chicago, Illinois). None of the models observedfor all chemical parameters were significant, therefore, nosignificant changes were observed in these parameters(Table 1). However, trends were observed for certain chemi-cal parameters when plotting graphs of each parameter. ThepH seemed to decrease over time, but showed a peak at13 months. The percentage of dry matter decreased in thefirst 2 samples, then reached a plateau after 13 months. Aslight decrease of the C/N ratio was observed until18 months, where the ratio then increased from 18.2 to 21.6.A decrease in the temperature of the compost piles was ob-
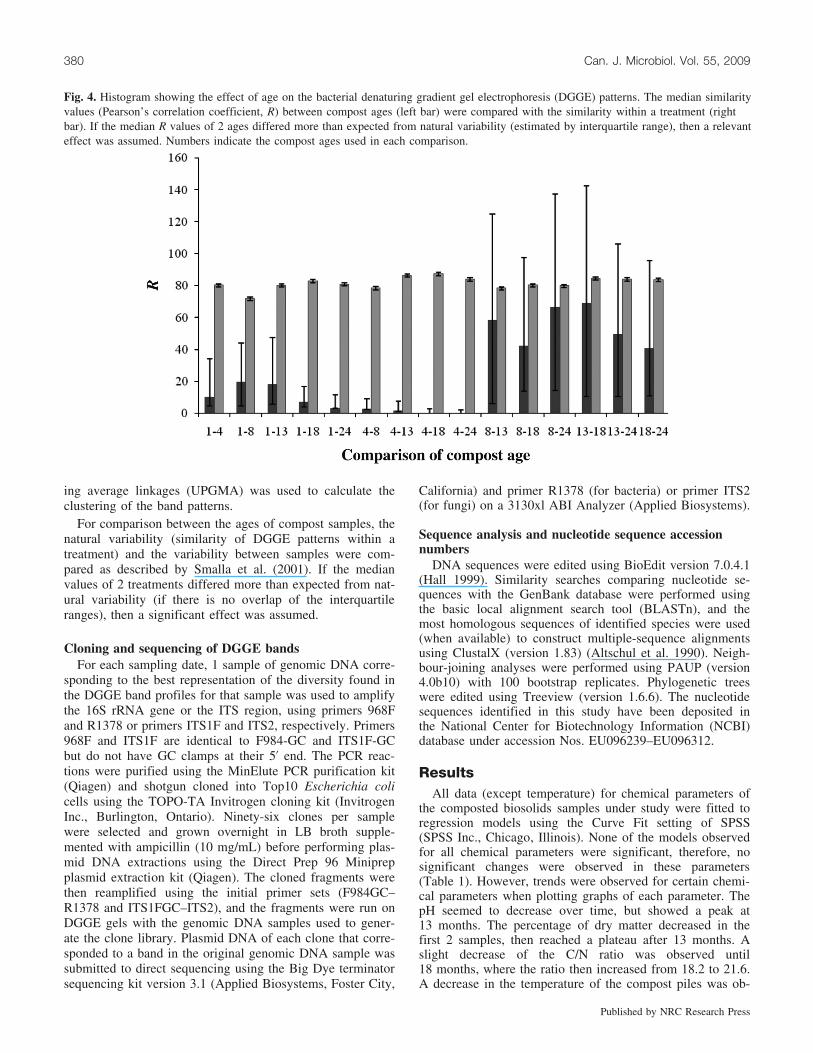
Fig. 4. Histogram showing the effect of age on the bacterial denaturing gradient gel electrophoresis (DGGE) patterns. The median similarityvalues (Pearson’s correlation coefficient, R) between compost ages (left bar) were compared with the similarity within a treatment (rightbar). If the median R values of 2 ages differed more than expected from natural variability (estimated by interquartile range), then a relevanteffect was assumed. Numbers indicate the compost ages used in each comparison.
380 Can. J. Microbiol. Vol. 55, 2009
Published by NRC Research Press

served between the first and 13th month of composting,whereas the temperature reached a plateau from the 13thmonth to the end of the composting period under study.
DNA isolated from all composted biosolids samples wassuccessfully amplified using the different primer sets usedin this study. Preliminary experiments using a direct PCRapproach targeting the ITS region of fungi were unsuccess-ful in generating amplicons of the correct size, thus, a nestedPCR approach was used. In general, DGGE banding patternsfor bacterial and fungal amplicons were similar between thedifferent replicates for each sampling date, demonstratingthe low variability of the DNA extraction, PCR amplifica-tion, and DGGE analysis.
DGGE analysis of the bacterial communities showed com-plex banding patterns throughout the process (Fig. 1). Fifty-three dominant DGGE bands were isolated, sequenced, andtheir closest matches identified using the BLASTn search al-gorithm. Phylogenetic analyses indicated that the dominantbacteria detected were taxonomically diverse and could begrouped into 10 phyla (Figs. 2 and 3). All banding patternsfor each replicate clustered together. The banding patternsfor the young compost (1 and 4 months), the 13 and18 month samples, and the 8 and 24 month samples wereclosely related and clustered together (Fig. 1). The youngcompost formed a separate group clustering away from theother DGGE profiles.
Based on the DGGE profiles and the identification ofphyla of the selected bands, a shift in the bacterial commun-ity was observed after 4 months of composting. All bacteriadetected during the first 4 months belonged to the same 3phyla, Deinococcus-Thermus, Firmicutes, and Actinobacte-ria. Whereas Deinococcus-Thermus were not detected after4 months of composting, Firmicutes and Actinobacteriawere detected throughout the whole composting period understudy. The banding patterns were more complex after4 months and visually showed a higher number of bands.Bacteria belonging to the phyla Bacteroidetes, Chloroflexi,Acidobacteria, and Proteobacteria were detected from 8 to24 months. Finally, Nitrospirae were detected at 8 and13 months of composting, whereas Chlamydiae and Candi-date Division TM7 were only detected in 13-month-old com-post. The identification of the DGGE bands also showed thatspecies within the various phyla varied with composting time(Fig. 3).
To quantify the possible effect of compost age on the bac-terial population present, R was calculated for each pair oflanes to observe the similarity between 2 lanes. For statisti-cal comparison of the variation between compost ages, thesimilarity of DGGE profiles within a compost age and be-tween compost ages was compared. One and 4 month com-post were significantly different from each other and also
Fig. 5. Denaturing gradient gel electrophoresis (DGGE) patterns ofthe PCR-amplified intergenic transcribed spacers (ITS) region frag-ments from composted biosolids samples of different ages. The firstand last lane of each gel (lanes 1, 12, 13, 24, 25, and 36) representa fungal DNA ladder comprised of PCR-amplified ITS region frag-ments from Epicoccum nigrum, Sclerotina sclerotiorum, andPhoma pomorum. Lanes 2–6 represent the 1 month compost sam-ples, lanes 7–11 represent the 4 month samples, lanes 14–18 repre-sent the 8 month samples, lanes 19–23 represent the 13 monthsamples, lanes 26–30 represent the 18 month samples, and lanes31–35 represent the 24 month samples. Bands that were success-fully sequenced are numbered. The relatedness of the DGGE pat-terns of the 6 compost ages and their replicates is displayed usingthe unweighted pair-group method using arithmetic averages(UPGMA) dendrogram and Pearson’s correlation coefficient (R).
Novinscak et al. 381
Published by NRC Research Press
when compared with all other compost ages. No differenceswere observed between the older compost samples (8–24 months), indicating that the bacterial communities ofthese 4 ages were similar (Fig. 4). Therefore, after 8 monthsof composting, the banding pattern of the samples did notdiffer significantly.
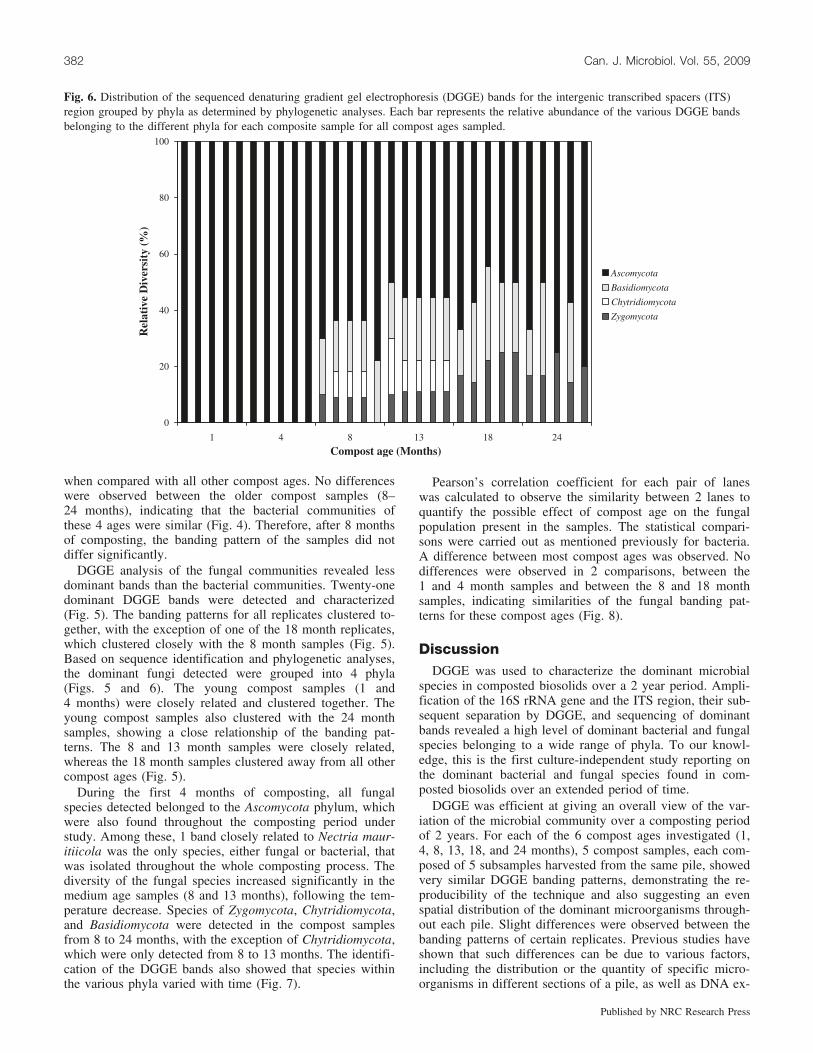
DGGE analysis of the fungal communities revealed lessdominant bands than the bacterial communities. Twenty-onedominant DGGE bands were detected and characterized(Fig. 5). The banding patterns for all replicates clustered to-gether, with the exception of one of the 18 month replicates,which clustered closely with the 8 month samples (Fig. 5).Based on sequence identification and phylogenetic analyses,the dominant fungi detected were grouped into 4 phyla(Figs. 5 and 6). The young compost samples (1 and4 months) were closely related and clustered together. Theyoung compost samples also clustered with the 24 monthsamples, showing a close relationship of the banding pat-terns. The 8 and 13 month samples were closely related,whereas the 18 month samples clustered away from all othercompost ages (Fig. 5).
During the first 4 months of composting, all fungalspecies detected belonged to the Ascomycota phylum, whichwere also found throughout the composting period understudy. Among these, 1 band closely related to Nectria maur-itiicola was the only species, either fungal or bacterial, thatwas isolated throughout the whole composting process. Thediversity of the fungal species increased significantly in themedium age samples (8 and 13 months), following the tem-perature decrease. Species of Zygomycota, Chytridiomycota,and Basidiomycota were detected in the compost samplesfrom 8 to 24 months, with the exception of Chytridiomycota,which were only detected from 8 to 13 months. The identifi-cation of the DGGE bands also showed that species withinthe various phyla varied with time (Fig. 7).
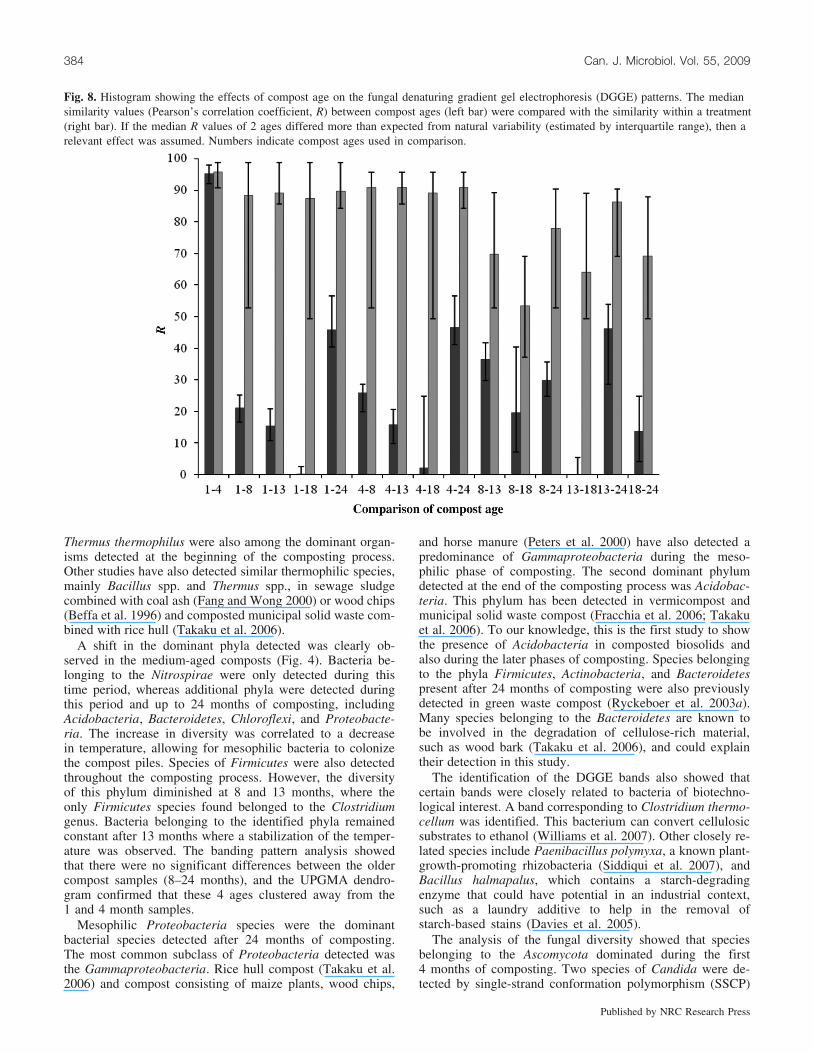
Pearson’s correlation coefficient for each pair of laneswas calculated to observe the similarity between 2 lanes toquantify the possible effect of compost age on the fungalpopulation present in the samples. The statistical compari-sons were carried out as mentioned previously for bacteria.A difference between most compost ages was observed. Nodifferences were observed in 2 comparisons, between the1 and 4 month samples and between the 8 and 18 monthsamples, indicating similarities of the fungal banding pat-terns for these compost ages (Fig. 8).
DiscussionDGGE was used to characterize the dominant microbial
species in composted biosolids over a 2 year period. Ampli-fication of the 16S rRNA gene and the ITS region, their sub-sequent separation by DGGE, and sequencing of dominantbands revealed a high level of dominant bacterial and fungalspecies belonging to a wide range of phyla. To our knowl-edge, this is the first culture-independent study reporting onthe dominant bacterial and fungal species found in com-posted biosolids over an extended period of time.
DGGE was efficient at giving an overall view of the var-iation of the microbial community over a composting periodof 2 years. For each of the 6 compost ages investigated (1,4, 8, 13, 18, and 24 months), 5 compost samples, each com-posed of 5 subsamples harvested from the same pile, showedvery similar DGGE banding patterns, demonstrating the re-producibility of the technique and also suggesting an evenspatial distribution of the dominant microorganisms through-out each pile. Slight differences were observed between thebanding patterns of certain replicates. Previous studies haveshown that such differences can be due to various factors,including the distribution or the quantity of specific micro-organisms in different sections of a pile, as well as DNA ex-
Fig. 6. Distribution of the sequenced denaturing gradient gel electrophoresis (DGGE) bands for the intergenic transcribed spacers (ITS)region grouped by phyla as determined by phylogenetic analyses. Each bar represents the relative abundance of the various DGGE bandsbelonging to the different phyla for each composite sample for all compost ages sampled.
382 Can. J. Microbiol. Vol. 55, 2009
Published by NRC Research Press

traction and PCR amplification variations (Muyzer 1999; deLipthay et al. 2004).
On the fungal DGGE gels, sequencing of multiple bandslocated very closely for a given sample revealed DNA se-quences with identical homology, as obtained for bandsclosely related to Nectria spp., Thermomyces spp., Coprinusspp., and Arthrobotrys spp. Previous studies have shown thata single species can generate more than 1 fragment duringDGGE owing to multiple copies of the same DNA sectionwith slight genetic variations (Nubel et al. 1996). Multipleband patterns corresponding to the same bacterial specieswere not observed on the bacterial DGGE gels.
Clear changes in the microbial populations were observedduring the 24 month period under study. Most of thesechanges seemed closely related to the temperature variations
observed over composting time. The diversity of bacterialand fungal species significantly increased after the decreasein temperature observed in the medium-aged compost (8 and13 months), suggesting the end of the thermophilic phase. Asimilar trend was also reported in studies conducted on com-posted pig slurry and shredded straw (Klamer and Baath1998), green waste and shredded wood (Blanc et al. 1999),and green waste (Ryckeboer et al. 2003a).
The bacteria observed at the beginning of the compostingprocess between 1 and 4 months, when temperatures of>45 8C were recorded, showed that Firmicutes, Deinococcus-Thermus, and Actinobacteria were the dominant bacterialphyla present in the thermophilic phase. Most species identi-fied belonged to the Firmicutes phylum and consisted ofBacillus or Bacillus-like species. Different isolates of
Fig. 7. Unrooted neighbour-joining tree representing the phylogenetic relationship of the 21 cloned intergenic transcribed spacers (ITS) re-gion denaturing gradient gel electrophoresis (DGGE) bands to the most closely related identified sequences obtained from BLASTnsearches. Letters following the cloned ITS region DGGE bands represent the compost age groups in which the bands were present. Theletter A represents 1 month; B, 4 months; C, 8 months; D, 13 months; E, 18 months; and F represents 24 months of composting. Thenumbers at the nodes of the tree are bootstrap values for each node with 100 bootstrap resampling (values <50 are omitted). The bar repre-sents 0.1 substitutions per site.
Novinscak et al. 383
Published by NRC Research Press
Thermus thermophilus were also among the dominant organ-isms detected at the beginning of the composting process.Other studies have also detected similar thermophilic species,mainly Bacillus spp. and Thermus spp., in sewage sludgecombined with coal ash (Fang and Wong 2000) or wood chips(Beffa et al. 1996) and composted municipal solid waste com-bined with rice hull (Takaku et al. 2006).
A shift in the dominant phyla detected was clearly ob-served in the medium-aged composts (Fig. 4). Bacteria be-longing to the Nitrospirae were only detected during thistime period, whereas additional phyla were detected duringthis period and up to 24 months of composting, includingAcidobacteria, Bacteroidetes, Chloroflexi, and Proteobacte-ria. The increase in diversity was correlated to a decreasein temperature, allowing for mesophilic bacteria to colonizethe compost piles. Species of Firmicutes were also detectedthroughout the composting process. However, the diversityof this phylum diminished at 8 and 13 months, where theonly Firmicutes species found belonged to the Clostridiumgenus. Bacteria belonging to the identified phyla remainedconstant after 13 months where a stabilization of the temper-ature was observed. The banding pattern analysis showedthat there were no significant differences between the oldercompost samples (8–24 months), and the UPGMA dendro-gram confirmed that these 4 ages clustered away from the1 and 4 month samples.
Mesophilic Proteobacteria species were the dominantbacterial species detected after 24 months of composting.The most common subclass of Proteobacteria detected wasthe Gammaproteobacteria. Rice hull compost (Takaku et al.2006) and compost consisting of maize plants, wood chips,
and horse manure (Peters et al. 2000) have also detected apredominance of Gammaproteobacteria during the meso-philic phase of composting. The second dominant phylumdetected at the end of the composting process was Acidobac-teria. This phylum has been detected in vermicompost andmunicipal solid waste compost (Fracchia et al. 2006; Takakuet al. 2006). To our knowledge, this is the first study to showthe presence of Acidobacteria in composted biosolids andalso during the later phases of composting. Species belongingto the phyla Firmicutes, Actinobacteria, and Bacteroidetespresent after 24 months of composting were also previouslydetected in green waste compost (Ryckeboer et al. 2003a).Many species belonging to the Bacteroidetes are known tobe involved in the degradation of cellulose-rich material,such as wood bark (Takaku et al. 2006), and could explaintheir detection in this study.
The identification of the DGGE bands also showed thatcertain bands were closely related to bacteria of biotechno-logical interest. A band corresponding to Clostridium thermo-cellum was identified. This bacterium can convert cellulosicsubstrates to ethanol (Williams et al. 2007). Other closely re-lated species include Paenibacillus polymyxa, a known plant-growth-promoting rhizobacteria (Siddiqui et al. 2007), andBacillus halmapalus, which contains a starch-degradingenzyme that could have potential in an industrial context,such as a laundry additive to help in the removal ofstarch-based stains (Davies et al. 2005).
The analysis of the fungal diversity showed that speciesbelonging to the Ascomycota dominated during the first4 months of composting. Two species of Candida were de-tected by single-strand conformation polymorphism (SSCP)
Fig. 8. Histogram showing the effects of compost age on the fungal denaturing gradient gel electrophoresis (DGGE) patterns. The mediansimilarity values (Pearson’s correlation coefficient, R) between compost ages (left bar) were compared with the similarity within a treatment(right bar). If the median R values of 2 ages differed more than expected from natural variability (estimated by interquartile range), then arelevant effect was assumed. Numbers indicate compost ages used in comparison.
384 Can. J. Microbiol. Vol. 55, 2009
Published by NRC Research Press

early in the composting of maize plants, wood chips, andhorse manure (Peters et al. 2000). Another study analyzingthe fungal diversity found during the initial temperature in-crease of composted dog food, used to simulate food waste,showed that 11 of the 13 isolates detected belonged to thephylum Ascomycota (Hansgate et al. 2005). The dominanceof Ascomycota species in various types of compost suggeststhat these species are initial fungal colonizers of composts.However, of the various fungal isolates detected in thesestudies, none correspond to the same genera identified in thisstudy. This could in part be due to the initial material usedfor composting, suggesting that fungal populations eitheroriginally present in various types of organic material or rap-idly colonizing them vary more than bacterial populations.
The medium-aged compost samples, similarly to bacteria,showed a significant shift in diversity, where species belong-ing to the Zygomycota, Chytridiomycota, and Basidiomycotawere also detected. PLFA used on shredded straw and horsemanure over a 100 day period showed that the fungal popu-lation increased when the temperature of the compost de-creased below 50 8C. They also observed the fruiting bodiesof Coprinus cinereus on the compost (Klamer and Baath1998). Coprinus spp. were also detected in this study after8 months of composting. Our results also corroborate an-other PLFA study conducted on composted municipal wastethat showed a general increase in fungal diversity after theend of the thermophilic phase (Hellmann et al. 1997). Com-posting rice straw also showed an increase of diversity duringthe maturing phase where species belonging to the Asco-mycota, Zygomycota, Chytridiomycota, and Basidiomycotawere detected (Cahyani et al. 2004). Overall, the numberof DGGE bands detected and the inherent fungal diversitywas far less than for bacteria, a finding also reported inprevious studies performed on composts obtained fromdog food pellets and pine chips (Hansgate et al. 2005) andgreen waste (Ryckeboer et al. 2003a).
Bands closely related to the bacterial pathogen Legionellamicdadei and the fungal pathogen Scedosporium apiosper-mum were detected at 18 and 8 months, respectively. Thesepathogens were not detected during any other compostingtime under study. Interestingly, no Escherichia or Salmo-nella species were detected in the compost samples. Thesepathogenic species are known to be shed in feces, and couldbe present in composted biosolids (Dudley et al. 1980).Their absence does not indicate that the composted biosolidsunder study were Escherichia- and Salmonella-free, but thatthey were not dominant species. Another explanation couldbe due to the lime treatment that the biosolids underwent.The increase in pH could have been elevated enough to dis-courage the growth of these bacterial species as well as re-duce the microbial diversity in the initial composting phases.Other studies on lime-added composted biosolids haveshown that the presence of lime created a decrease in thenumber of thermophilic bacteria and Actinomycetes foundin the samples (Wong and Fang 2000).
Studies have shown that microorganisms found in differ-ent composting systems can vary based on the initial materi-als composted (Ishii and Takii 2003), as well as thecomposting process used (Fracchia et al. 2006). Therefore,differences observed between the microorganisms detectedin this study and others could be explained by this factor.
As well, caution should be used when comparing small-scaleand larger scale composting processes. Larger scale com-posting systems can take longer to achieve higher tempera-tures (Klamer and Baath 1998), therefore, the presence ofvarious microorganisms could differ based on the length oftime needed to achieve the various phases.
The description of the main microbial groups found incomposted biosolids could find different applications, in-cluding the discovery of biotechnologically relevant micro-organisms, the development or adaptation of culture mediato study less-characterized microorganisms present in com-posts, and the design of novel molecular primers and probesallowing quantitative monitoring of key microorganismsover time. The detection of specific microbial species, usingculture-independent as well as culture-dependent methods,could also lead to a better identification of the differentcomposting phases and a better determination of the matur-ing process. However, the main challenge that still remainswill consist of determining and better understanding the dif-ferent roles of the various microorganisms that are responsi-ble for this complex process.
AcknowledgementsA MITACS scholarship was awarded to A. Novinscak.
The authors thank The Greater Moncton SewerageCommission for financial support and technical assistance,J. Boudreau for technical assistance, and Dr. G. Moreau forhis assistance with statistical analyses.
ReferencesAltschul, S.F., Gish, W., Miller, W., Myers, E.W., and Lipman,
D.J. 1990. Basic local alignment search tool. J. Mol. Biol. 215:403–410. PMID:2231712.
Anderson, I.C., Campbell, C.D., and Prosser, J.I. 2003. Diversity offungi in organic soils under a moorland – Scots pine (Pinussylvestris L.) gradient. Environ. Microbiol. 5: 1121–1132.doi:10.1046/j.1462-2920.2003.00522.x. PMID:14641592.
Beffa, T., Blanc, M., Lyon, P.F., Vogt, G., Marchiani, M., Fischer,J.L., and Aragno, M. 1996. Isolation of Thermus strains fromhot composts (60 to 80 8C). Appl. Environ. Microbiol. 62:1723–1727. PMID:8633870.
Blanc, M., Marilley, L., Beffa, T., and Aragno, M. 1999. Thermo-philic bacterial communities in hot composts as revealed bymost probable number counts and molecular (16S rDNA) meth-ods. FEMS Microbiol. Ecol. 28: 141–149. doi:10.1111/j.1574-6941.1999.tb00569.x.
Boulter, J.I., Boland, G.J., and Trevors, J.T. 2000. Compost: astudy of the development process and end-product potential forsuppression of turfgrass disease. World J. Microbiol. Biotechnol.16: 115–134. doi:10.1023/A:1008901420646.
Boulter, J.I., Boland, G.J., and Trevors, J.T. 2002. Assessment ofcompost for suppression of Fusarium Patch (Microdochiumnivale) and Typhula Blight (Typhula ishikariensis) snow moldsof turfgrass. Biol. Control, 25: 162–172. doi:10.1016/S1049-9644(02)00060-9.
Cahyani, V.R., Matsuya, K., Asakawa, S., and Kimura, M. 2004.Succession and phylogenetic profile of eukaryotic communitiesin the composting process of rice straw estimated by PCR–DGGE analysis. Biol. Fertil. Soils, 40: 334–344. doi:10.1007/s00374-004-0783-x.
Davies, G.J., Brzozowski, A.M., Dauter, Z., Rasmussen, M.D.,Borchert, T.V., and Wilson, K.S. 2005. Structure of a Bacillus
Novinscak et al. 385
Published by NRC Research Press

halmapalus family 13 a-amylase, BHA, in complex with anacarbose-derived nonasaccharide at 2.1 A resolution. ActaCrystallogr. Sect. D Biol. Crystallogr. 61: 190–193. doi:10.1107/S0907444904027118. PMID:15681870.
de Lipthay, J.R., Enzinger, C., Johnsen, K., Aamand, J., andSørensen, S.J. 2004. Impact of DNA extraction method on bac-terial community composition measured by denaturing gradientgel electrophoresis. Soil Biol. Biochem. 36: 1607–1614. doi:10.1016/j.soilbio.2004.03.011.
Dees, P.M., and Ghiorse, W.C. 2001. Microbial diversity in hotsynthetic compost as revealed by PCR-amplified rRNAsequences from cultivated isolates and extracted DNA. FEMSMicrobiol. Ecol. 35: 207–216. doi:10.1111/j.1574-6941.2001.tb00805.x. PMID:11295460.
Dudley, D.J., Guentzel, M.N., Ibarra, M.J., Moore, B.E., and Sagik,B.P. 1980. Enumeration of potentially pathogenic bacteria fromsewage sludges. Appl. Environ. Microbiol. 39: 118–126.PMID:6243900.
Dumontet, S., Dinel, H., and Baloda, S.B. 1999. Pathogen reduc-tion in sewage sludge by composting and other biological treat-ments: a review. Biol. Agric. Hortic. 16: 409–430.
Fang, M., and Wong, J.W.C. 2000. Changes in thermophilic bac-teria populations and diversity during composting of coal flyash and sewage sludge. Water Air Soil Pollut. 124: 333–343.doi:10.1023/A:1005284629867.
Finstein, M.S., and Morris, M.L. 1975. Microbiology of municipalsolid waste composting. Adv. Appl. Microbiol. 19: 113–151.doi:10.1016/S0065-2164(08)70427-1. PMID:242194.
Fracchia, L., Dohrmann, A.B., Giovanna Martinotti, M., andTebbe, C.C. 2006. Bacterial diversity in a finished compost andvermicompost: differences revealed by cultivation-independentanalyses of PCR-amplified 16S rRNA genes. Appl. Microbiol.Biotechnol. 71: 942–952. doi:10.1007/s00253-005-0228-y.PMID:16395545.
Gardes, M., and Bruns, T.D. 1993. ITS primers with enhanced spe-cificity for basidiomycetes — application to the identification ofmycorrhizae and rusts. Mol. Ecol. 2: 113–118. doi:10.1111/j.1365-294X.1993.tb00005.x. PMID:8180733.
Hall, T.A. 1999. BioEdit: a user-friendly biological sequence align-ment editor and analysis program for Windows 95/98/NT.Nucleic Acids Symp. Ser. 41: 95–98.
Hansgate, A.M., Schloss, P.D., Hay, A.G., and Walker, L.P. 2005.Molecular characterization of fungal community dynamics in theinitial stages of composting. FEMS Microbiol. Ecol. 51: 209–214. doi:10.1016/j.femsec.2004.08.009. PMID:16329869.
Hellmann, B., Zelles, L., Palojarvi, A., and Bai, Q.Y. 1997. Emis-sion of climate-relevant trace gases and succession of microbialcommunities during open-window composting. Appl. Environ.Microbiol. 63: 1011–1018. PMID:16535535.
Herrmann, R.F., and Shann, J.F. 1997. Microbial communitychanges during the composting of municipal solid waste. Microb.Ecol. 33: 78–85. doi:10.1007/s002489900010. PMID:9039768.
Heuer, H., Krsek, M., Baker, P., Smalla, K., and Wellington, E.M.H.1997. Analysis of actinomycete communities by specific amplifi-cation of genes encoding 16s rRNA and gel-electrophoreticseparation in denaturing gradients. Appl. Environ. Microbiol. 63:3233–3241. PMID:9251210.
Hoitink, H.A.J., and Fahy, P.C. 1986. Basis for the control of soil-borne plant pathogens with composts. Annu. Rev. Phytopathol.24: 93–114. doi:10.1146/annurev.py.24.090186.000521.
Ishii, K., and Takii, S. 2003. Comparison of microbial communitiesin four different composting processes as evaluated by denatur-ing gradient gel electrophoresis. J. Appl. Microbiol. 95: 109–119. doi:10.1046/j.1365-2672.2003.01949.x. PMID:12807460.
Ishii, K., Fukui, M., and Takii, S. 2000. Microbial succession dur-ing a composting process as evaluated by denaturing gradientgel electrophoresis analysis. J. Appl. Bacteriol. 89: 768–777.
Ivors, K.L., Collopy, P.D., Beyer, D.M., and Kang, S. 2000. Identi-fication of bacteria in mushroom compost using ribosomal RNAsequence. Compost Sci. Util. 8: 247–253.
Kane, B.E., and Mullins, J.T. 1973. Thermophilic fungi in a muni-cipal waste compost system. Mycologia, 65: 1087–1100. doi:10.2307/3758290. PMID:4585097.
Klamer, M., and Baath, E. 1998. Microbial community dynamicsduring composting of straw material studied using phospholipidfatty acid analysis. FEMS Microbiol. Ecol. 27: 9–20. doi:10.1111/j.1574-6941.1998.tb00521.x.
Muyzer, G. 1999. DGGE/TGGE: a method for identifying genesfrom natural ecosystems. Curr. Opin. Microbiol. 2: 317–322.doi:10.1016/S1369-5274(99)80055-1. PMID:10383868.
Muyzer, G., de Waal, E.C., and Uitterlinden, A.G. 1993. Profilingof complex microbial populations by denaturing gradient gelelectrophoresis analysis of polymerase chain reaction-amplifiedgenes coding for 16S rRNA. Appl. Environ. Microbiol. 59:695–700. PMID:7683183.
Nakasaki, K., Sasaki, M., Shoda, H., and Kubota, H. 1985. Changein microbial numbers during thermophilic composting of sewagesludge with reference to CO2 evolution rate. Appl. Environ. Mi-crobiol. 49: 37–41. PMID:16346706.
Nubel, U., Engelen, B., Felske, A., Snaidr, J., Wieshuber, A.,Amann, R.I., et al. 1996. Sequence heterogeneities of genes en-coding 16S rRNAs in Paenibacillus polymyxa detected by tem-perature gradient gel electrophoresis. J. Bacteriol. 178: 5636–5643. PMID:8824607.
Peters, S., Koschinsky, S., Schwieger, F., and Tebbe, C.C. 2000.Succession of microbial communities during hot composting asdetected by PCR-single-strand-conformation polymorphism-based genetic profiles of small-subunit rRNA genes. Appl. En-viron. Microbiol. 66: 930–936. doi:10.1128/AEM.66.3.930-936.2000. PMID:10698754.
Rademaker, J.L.W., and de Bruijn, F.J. 2008. Computer-assistedanalysis of molecular fingerprints and database construction. InMolecular microbial ecology manual. Edited by G.A. Kowalchuk,F.J. de Bruijn, I.M. Head, A.D.L. Akkermans, and J.D. van Elsas.Springer, Dordrecht, The Netherlands. pp. 1397–1406.
Ryckeboer, J., Mergaert, J., Coosemans, J., Deprins, K., and Swings,J. 2003a. Microbiological aspects of biowaste during compostingin a monitored compost bin. J. Appl. Microbiol. 94: 127–137.doi:10.1046/j.1365-2672.2003.01800.x. PMID:12492933.
Ryckeboer, J., Mergaert, J., Vaes, K., Klammer, S., De Clercq, D.,Coosemans, J., et al. 2003b. A survey of bacteria and fungi oc-curring during composting and self-heating processes. Ann. Mi-crobiol. 53: 349–410.
Siddiqui, Z.A., Baghel, G., and Akhtar, M.S. 2007. Biocontrol ofMeloidogyne javanica by Rhizobium and plant growth-promotingrhizobacteria on lentil. World J. Microbiol. Biotechnol. 23:435–441. doi:10.1007/s11274-006-9244-z.
Smalla, K., Weiland, G., Buchner, A., Zock, A., Parzy, J., Kaiser,S., et al. 2001. Bulk and rhizosphere soil bacterial communitiesstudied by denaturing gradient gel electrophoresis: plant-dependent enrichment and seasolan shifts revealed. Appl. En-viron. Microbiol. 67: 4742–4751. doi:10.1128/AEM.67.10.4742-4751.2001. PMID:11571180.
Smit, E., Leeflang, P., Glandorf, B., Van Elsas, J.D., and Wernars,K. 1999. Analysis of fungal diversity in the wheat rhizosphereby sequencing of cloned PCR-amplified genes encoding 18SrRNA and temperature gradient gel electrophoresis. Appl. En-viron. Microbiol. 65: 2614–2621. PMID:10347051.
386 Can. J. Microbiol. Vol. 55, 2009
Published by NRC Research Press

Strom, P.F. 1985. Identification of thermophilic bacteria in solid-waste composting. Appl. Environ. Microbiol. 50: 906–913.PMID:4083886.
Takaku, H., Kodaira, S., Kimoto, A., Nashimoto, M., and Takagi,M. 2006. Microbial communities in the garbage compostingwith rice hull as an amendment revealed by culture-dependentand -independent approaches. J. Biosci. Bioeng. 101: 42–50.doi:10.1263/jbb.101.42. PMID:16503290.
Tyagi, R.D., Sikati Foko, V., Barnabe, S., Vidyarthi, A.S., Valero,J.R., and Surampalli, R.Y. 2002. Simultaneous production ofbiopesticide and alkaline proteases by Bacillus thuringiensisusing sewage sludge as a raw material. Water Sci. Technol. 46:247–254. PMID:12479478.
Ueno, Y., Haruta, S., Ishii, M., and Igarashi, Y. 2001. Microbialcommunity in anaerobic hydrogen-producing microflora en-riched from sludge compost. Appl. Microbiol. Biotechnol. 57:555–562. doi:10.1007/s002530100806. PMID:11762604.
US EPA. 1999. Biosolids generation, use and disposal in the Uni-ted States. US EPA Municipal and Industrial Solid Waste Divi-sion, Office of Solid Waste, Washington, D.C.
Wagner, M., Amann, R., Lemmer, H., and Schleifer, K.H. 1993.Probing activated sludge with oligonucleotides specific forProteobacteria: inadequacy of culture-dependent methods fordescribing microbial community structure. Appl. Environ.Microbiol. 59: 1520–1525. PMID:8517747.
White, T.J., Bruns, S.L., and Taylor, J. 1990. Amplification and di-rect sequencing of fungal ribosomal RNA genes for phyloge-netics. In PCR protocols: a guide to methods and applications.Edited by M.A. Innis, D.H. Gelfand, J.J. Sninsky, and T.J.White. Academic Press, San Diego, California.
Williams, T.I., Combs, J.C., Lynn, B.C., and Strobel, H.J. 2007.Proteomic profile changes in membranes of ethanol-tolerantClostridium thermocellum. Appl. Microbiol. Biotechnol. 74:422–432. doi:10.1007/s00253-006-0689-7. PMID:17124583.
Wong, J.W.C., and Fang, M. 2000. Effects of lime addition onsewage sludge composting process. Water Res. 34: 3691–3698.doi:10.1016/S0043-1354(00)00116-0.
Novinscak et al. 387
Published by NRC Research Press
Copyright © 2022 FDOKUMEN



















