
Characterization and role of carbonic anhydrase in the calcification process of the azooxanthellate...
13
Abstract In zooxanthellate corals, the photosynthetic fixation of carbon dioxide and the precipitation of CaCO 3 are intimately linked both spatially and tem- porally making it difficult to study carbon transport mechanisms involved in each pathway. When studying Tubastrea aurea, a coral devoid of zooxanthellae, we can focus on carbon transport mechanisms involved only in the calcification process. We performed this study to characterize T. aurea carbonic anhydrase and to determine its role in the calcification process. We have shown that inhibition of tissular carbonic anhy- drase activity affects the calcification rate. We have measured the activity of this enzyme both in the tissues and in the organix matrix extracted from the skeleton. Our results indicate that organic matrix proteins, which are synthesized by the calcifying tissues, are not only structural proteins, but they also play a crucial catalytic role by eliminating the kinetic barrier to interconver- sion of inorganic carbon at the calcification site. By immunochemistry we have demonstrated the presence of a protein both in the tissues and in the organic matrix, which shares common features with prokary- otic carbonic anhydrases. Keywords Carbonic anhydrase Á Carbon Á Calcification Á Coral Á Biomineralization Á Organic matrix Abbreviations CA Carbonic anhydrase BSA Bovine serum albumin DIC Dissolved inorganic carbon DTT Dithiothreitol EDTA Ethylenediaminetetraacetate FSW Filtered seawater PBS Phosphate buffered saline PAF Paraformaldehyde PIC Protease inhibitor cocktail RT Room temperature SOM Soluble organic matrix SDS Sodium dodecyl sulphate TBS Tris buffered saline DIC Dissolved inorganic carbon Introduction Scleractinians (stony corals) are coelenterates that form aragonitic calcium carbonate (CaCO 3 ) skeletons. They are classically functionally divided into two groups: the hermatypic (reef-building) and the ahermatypic (non-reef-building) corals. The vast majority of the hermatypic corals are found in shallow, tropical oceans and characteristically contain within their tissues large populations of symbiotic dinoflagellates called Communicated by S.A. Poulet, Roscoff. S. Tambutte ´(&) Á E. Tambutte ´ Á D. Zoccola Á N. Caminiti Á S. Lotto Á A. Moya Á D. Allemand Centre Scientifique de Monaco, av. Saint Martin, MC 98 000, Monaco e-mail: stambutte@centrescientifique.mc J. Adkins Department of Geology and Planetary Sciences, MS100-23, Caltech, 1200 E. California Blvd., Pasadena, CA 91125, USA D. Allemand UMR 1112 UNSA-INRA Faculte ´ des Sciences, Parc Valrose, B.P.71, 06108 Nice Cedex 2, France Mar Biol DOI 10.1007/s00227-006-0452-8 123 RESEARCH ARTICLE Characterization and role of carbonic anhydrase in the calcification process of the azooxanthellate coral Tubastrea aurea Sylvie Tambutte ´ Eric Tambutte ´ Didier Zoccola Natacha Caminiti Severine Lotto Aure ´lie Moya Denis Allemand Jess Adkins Received: 8 May 2006 / Accepted: 9 August 2006 Ó Springer-Verlag 2006
-
Upload
independent -
Category
Documents
-
view
3 -
download
0
Transcript of Characterization and role of carbonic anhydrase in the calcification process of the azooxanthellate...

Abstract In zooxanthellate corals, the photosynthetic
fixation of carbon dioxide and the precipitation of
CaCO3 are intimately linked both spatially and tem-
porally making it difficult to study carbon transport
mechanisms involved in each pathway. When studying
Tubastrea aurea, a coral devoid of zooxanthellae, we
can focus on carbon transport mechanisms involved
only in the calcification process. We performed this
study to characterize T. aurea carbonic anhydrase and
to determine its role in the calcification process. We
have shown that inhibition of tissular carbonic anhy-
drase activity affects the calcification rate. We have
measured the activity of this enzyme both in the tissues
and in the organix matrix extracted from the skeleton.
Our results indicate that organic matrix proteins, which
are synthesized by the calcifying tissues, are not only
structural proteins, but they also play a crucial catalytic
role by eliminating the kinetic barrier to interconver-
sion of inorganic carbon at the calcification site. By
immunochemistry we have demonstrated the presence
of a protein both in the tissues and in the organic
matrix, which shares common features with prokary-
otic carbonic anhydrases.
Keywords Carbonic anhydrase � Carbon �Calcification � Coral � Biomineralization �Organic matrix
Abbreviations
CA Carbonic anhydrase
BSA Bovine serum albumin
DIC Dissolved inorganic carbon
DTT Dithiothreitol
EDTA Ethylenediaminetetraacetate
FSW Filtered seawater
PBS Phosphate buffered saline
PAF Paraformaldehyde
PIC Protease inhibitor cocktail
RT Room temperature
SOM Soluble organic matrix
SDS Sodium dodecyl sulphate
TBS Tris buffered saline
DIC Dissolved inorganic carbon
Introduction
Scleractinians (stony corals) are coelenterates that form
aragonitic calcium carbonate (CaCO3) skeletons. They
are classically functionally divided into two groups:
the hermatypic (reef-building) and the ahermatypic
(non-reef-building) corals. The vast majority of the
hermatypic corals are found in shallow, tropical oceans
and characteristically contain within their tissues
large populations of symbiotic dinoflagellates called
Communicated by S.A. Poulet, Roscoff.
S. Tambutte (&) � E. Tambutte � D. Zoccola �N. Caminiti � S. Lotto � A. Moya � D. AllemandCentre Scientifique de Monaco, av. Saint Martin,MC 98 000, Monacoe-mail: [email protected]
J. AdkinsDepartment of Geology and Planetary Sciences, MS100-23,Caltech, 1200 E. California Blvd., Pasadena, CA 91125,USA
D. AllemandUMR 1112 UNSA-INRA Faculte des Sciences,Parc Valrose, B.P.71, 06108 Nice Cedex 2, France
Mar Biol
DOI 10.1007/s00227-006-0452-8
123
RESEARCH ARTICLE
Characterization and role of carbonic anhydrasein the calcification process of the azooxanthellate coralTubastrea aurea
Sylvie Tambutte Æ Eric Tambutte Æ Didier Zoccola Æ Natacha Caminiti ÆSeverine Lotto Æ Aurelie Moya Æ Denis Allemand Æ Jess Adkins
Received: 8 May 2006 / Accepted: 9 August 2006� Springer-Verlag 2006

zooxanthellae. In these zooxanthellate scleractinians,
the photosynthetic fixation of carbon dioxide (CO2)
and precipitation of CaCO3 are intimately linked both
at spatial (cell to ecosystem) and temporal (day–night)
scales rendering it difficult to study the carbon transport
mechanisms involved in each pathway. On the other
hand, the vast majority of ahermatypic corals are de-
void of zooxanthellae. Thus when studying these corals,
it is possible to focus on carbon transport mechanisms
involved only in calcification processes.
In corals, skeleton formation is a process of
‘‘extracellular biologically-controlled biomineraliza-
tion’’ and as such involves a mineral fraction and an
organic matrix. The means by which corals may
influence CaCO3 precipitation include (1) control of
the levels of inhibitors, promoters, and regulators of
calcification by the means of a set of macromolecules
(called the organic matrix) surrounding the crystal or
included within the mineral and (2) availability of
substrates. Since coral skeleton formation results from
the delivery of calcium and inorganic carbon to the
site of calcification, these two substrates are crucial to
study. Recently, most of the research involving coral
calcification has focused either upon the structure and
composition of organic matrices of skeletons (Gautret
et al. 1997, 2000; Cuif et al. 1999, 2003; Dauphin
2001) or on the uptake and mechanisms of deposition
of calcium ions (Wright and Marshall 1991; Tambutte
et al. 1995, 1996). However, for invertebrate miner-
alization, carbonate ions are as important as calcium
ions. Pearse (1970) established that skeletal carbonate
can originate from two different carbon sources: sol-
uble carbonates from sea water or CO2 produced by
animal metabolism. Furla et al. (2000) demonstrated
that in the zooxanthellate coral Stylophora pistillata,
the major source of DIC for coral calcification is
metabolic CO2 and not inorganic carbon originating
from seawater. Similar results were obtained in the
non-zooxanthellate octocoral Leptogorgia virgulata
and Corallium rubrum respectively by Lucas and
Knapp (1997) and Allemand and Grillo (1992). In
addition, these last authors have shown that DIC
supply is rate-limiting for calcification. However,
these conclusions can not be generalized to all corals
since Adkins et al. (2003) demonstrated that there is
little or no metabolic CO2 in the skeleton of the
deep-sea non-zooxanthellate coral, Desmophyllum
cristagalli.
Carbonic anhydrases are ubiquitous enzymes
known to act as catalysts for the interconversion
between CO2 and HCO3–. Since the limiting step in
the conversion from CO2 to carbonate ion is the
hydration step, CA can play an important role when
calcification is carbon limited. In avians, CA facili-
tates eggshell formation (Nys and de Laage 1984) and
in fishes, CA is supposed to play an important role in
otolith formation (Payan et al. 1997; Tohse and
Mugiya 2001; Tohse et al. 2004). In the case of
invertebrates, this enzyme has been found to play a
role in the calcification of calcareous sponges (Jones
and Ledger 1986), scleractinian corals (Goreau 1959;
Isa and Yamazato 1984; Marshall 1996, Furla et al.
2000; Al-Horani et al. 2003), octocorallians (Kingsley
and Watabe 1987; Allemand and Grillo 1992; Lucas
and Knapp 1996, 1997; Rahman et al. 2005, 2006),
molluscs and echinoderms (Miyamoto et al. 1996;
Mitsunaga et al. 1986). Carbonic anhydrase has been
described in many tissues but its presence in extra-
cellular calcified structures suggests that this enzyme
could also play an important role during the precipi-
tation step of the mineral. The aim of this study was
to characterize T. aurea carbonic anhydrase and
determine its role in the calcification process.
Materials and methods
Biological material
Parent coral colonies of Tubastrea aurea (Cœlentera-
ta:Anthozoa:Scleractinia), indigenous to the Indo-Pa-
cific, and sea anemones Aiptasia pulchella were
maintained at the Centre Scientifique de Monaco in
the following conditions: semi-open circuit, Mediter-
ranean sea water heated to 26 ± 0.2�C, and fed with
Artemia nauplii twice a week. T. aurea was maintained
in low light conditions at a constant irradiance
of 15 lmol photons m–2 s–1 on a 12 h/12 h light/dark
cycle, and A. pulchella was maintained in the same
conditions but with a constant irradiance of 250 lmol
photons m–2 s–1. For calcification rate experiments,
coral colonies were cut with a bone cutter in order to
obtain fragments of three to four polyps called nub-
bins. The sectioned skeleton was coated with epoxy
resin so that only the tissues were in contact with
seawater. Nubbins were used for experiments after a
period of 4–5 weeks, when new tissues entirely covered
the junction between the resin and the skeleton.
Cleaning was performed daily.
Preparation of tissues for carbonic anhydrase
activity assay
Six to seven polyps were cut from parent colonies, put
on ice and homogenized with a mortar in about 3 ml
cold veronal buffer prepared according to Weis et al.
Mar Biol
123

(1989): 25 mM veronal containing 5 mM ethylene-
diaminetetraacetate (EDTA), 5 mM dithiothreitol
(DTT), 10 mM MgSO4 with pH adjusted to 8.2. Pro-
tease inhibitor cocktail (PIC, SIGMA) 0.1% was then
added. The mixture was sonicated for 1 min and then
centrifuged at 765g for 20 min at 4�C. The supernatant
was centrifuged again in the same conditions and then
aliquoted in Eppendorf tubes for storage at –80�C.
Preparation of organic matrix for carbonic
anhydrase activity assay
Polyps were cleaned by removing soft tissues with 2N
NaOH for 2 h at 70�C. The skeletons were rinsed with
ultrapure water, dried at 60�C overnight and ground to
a fine powder with a mortar. The powder was demin-
eralized for one night at 4�C in 0.5 M EDTA con-
taining 0.1% PIC and 5 mM phenanthrolin. After
complete dissolution of aragonite, centrifugation
(10 min, 10,000g, 4�C) allowed soluble and insoluble
matrices to be separated. To desalt soluble compo-
nents, the supernatant containing the soluble organic
matrix (SOM) was filtered and concentrated using
Centricon� (Amicon, cut-off 5 kDa) according to the
manufacturer’s instructions. The retentate was ali-
quoted and stored at –80�C.
In vitro assay for carbonic anhydrase activity
The in vitro assay for CA activity is described in detail
by Weis et al. (1989). Briefly CA activity in crude ho-
mogenates was measured by the decrease of pH
resulting from the hydration of CO2 to HCO3– and H+
after the addition of substrate. All experiments were
performed at 4�C. CO2–saturated distilled H2O served
as a substrate and was prepared by passing gaseous
CO2 through an airstone for 30 min (pH 3.5). To run
the assay, 5 ml veronal buffer (pH 8.2) were trans-
ferred to a small beaker and 1 ml of homogenate
diluted in veronal buffer was added to obtain quanti-
ties of proteins ranging from 0 to 12 mg for tissues and
0–16 lg for organic matrix. The mixture was constantly
stirred with a magnetically driven stirring bar. Four ml
of substrate was then added rapidly and the decrease in
pH was recorded by a Ag/AgCl pH probe immersed
in the mixture and connected to a Metler DL70 pH
meter fitted with a chart recorder. As a control for
the non-catalyzed reaction, the same experiment was
performed without homogenate.
Carbonic anhydrase activity was calculated as
(t0 – t)/t, where t0 is the time needed for the non-
catalyzed reaction and t is the time for the catalyzed
reaction to obtain a pH decrease from 8 to 7.5. Units of
enzyme activity (EU) were normalized to the weight of
soluble proteins.
Inhibition of carbonic anhydrase activity
To test the effect of the inhibitor ethoxyzolamide on
CA activity, the assay was performed as described
above, but 0–10 lM ethoxyzolamide was added to the
veronal buffer before addition of tissue. Results are
expressed as percent inhibition calculated from 100 –
[(CA activity in presence of inhibitor/CA activity
in absence of inhibitor) · 100)]. IC50 represents the
concentration of inhibitor, which inhibits half of the
enzyme activity measured in the absence of inhibitor.
Effect of inhibition of CA activity on calcification
rate
Measurement of calcification rate was made according
to the method of Tambutte et al. (1995) adapted for
higher volumes. Measurements were made at equiva-
lent times of day in order to avoid possible variation
caused by endogenous circadian rhythms (Buddemeier
and Kinzie 1976). Nubbins grown on epoxy resin were
incubated for 2 h 15 min in 60 ml beakers containing
approximatively 800 kBq of 45Ca (as 45CaCl2, NEZ013,
Perkin Elmer) dissolved in filtered seawater. For
inhibition experiments, 10 lM ethoxyzolamide was
added in the incubation medium. Water motion was
provided during each incubation by small stirring bars
in order to reduce as much as possible diffusion limi-
tation by boundary layers. Exposure to air was limited
to less than 5 s during transfer to the incubation
beakers and incubations were made under low light
conditions.
At the end of the labelling period, each nubbin was
immersed for 20 s in a beaker containing 1 l FSW.
Labelled nubbins were then incubated in a beaker
containing 150 ml FSW for 180 min to monitor 45Ca
efflux into the rinse medium. Water motion was pro-
vided in the efflux medium by stirring bars. Upon
completion of the efflux, nubbins were dissolved
completely over a period of 20 min in approximately
5 ml of 1 N NaOH at 90�C. Each skeleton was then
rinsed six times in 5 ml of distilled water, dried and
dissolved in 10 ml of 6 N HCl overnight (‘‘HCl-soluble
fraction’’).
Radioactive samples were added to 4 ml Ultima
Gold (Packard) and emissions were measured using a
liquid scintillation analyzer (Tricarb, 2100TR, Pack-
ard).
Mar Biol
123

For the efflux, results are expressed as dpm mg–1
protein. For the calcification rate, results are expressed
as nmol Ca2+ h–1 mg–1 protein in tissues or nmol
Ca2+ h 1 g–1 skeleton and represent means ± S.D. for
at least three measurements. Calculation of the half-
time for calcium washout (T1/2) and its corresponding
rate constant as well as calculation of the size of the
coelenteric pool were made according to Tambutte
et al. (1995). In the presence of ethoxyzolamide, results
are expressed as percentage inhibition of calcification
rate.
A control experiment was performed with killed
nubbins (paraformaldehyde) in order to determine non
specific binding of 45Ca.
Extraction of proteins for immunoblotting
• Proteins from tissues: Six polyps of T. aurea were
cut from a colony with a bone cutter and rinsed in
FSW. They were then homogenized with a mortar
maintained on ice, in about 8 ml of extraction
buffer (50 mM Tris, 100 mM NaCl, 5 mM EDTA,
1% Triton X100, 0.1% PIC, and 5 mM phenanthr-
olin). The mixture was twice centrifuged at 765g for
10 min at 4�C to eliminate skeletal debris. The
supernatant was maintained on ice for 20 min while
vortexing every 5 min, the time necessary for the
buffer to extract proteins. The supernatant was
aliquoted in Eppendorf tubes for storage at –80�C
before experiments. Proteins were also extracted
from the sea anemone A. pulchella with the same
protocol except that they were directly homoge-
nized in the extraction buffer and sonicated.
• Proteins from organic matrix: The protocol was the
same as the preparation of organic matrix for the
carbonic anhydrase activity assay (see paragraph
above).
Dot blot
The activity of the antibody was examined by a dot-
blot assay on proteins extracted either from whole
tissues (40 lg of proteins) or organic matrix (12 lg of
proteins). Experiments were performed at room tem-
perature.
Briefly, the samples were deposited on nitrocellulose
membranes which were saturated with 1% BSA for 1 h
in TBS (140 mM NaCl, 5 mM Tris, pH 7.4) and la-
belled for 1 h with primary antibodies. The primary
antibodies were either (1) anti phycoerythrin (Ab-
Cam), 1:20,000 dilution or (2) rabbit anti-human
erythrocyte carbonic anhydrase II antibody (Rockland
immunochemicals), 1:10,000 dilution or (3) rabbit anti
N-terminal b-carbonic anhydrase from Synecchococcus
sp (generous gift from Mak Saito and Francois Morel),
1:25,000 dilution, in TBS-BSA 1%). Membranes were
then rinsed and incubated for 1 h with secondary
antibodies (horseradish peroxidase-linked anti-rabbit
IgG, Sigma, 1:2,000 dilution in TBS-BSA 1%).
Immunoreactive dots were then revealed with ECL kit
(GE Healthcare). Controls were made with the pre-
immune serum as the primary antibody.
Electrophoresis, protein transfer and Western blot
Proteins extracted either from whole tissues (100 lg
of proteins) or just the organic matrix (20 lg of pro-
teins) were homogenized in Laemmli sample buffer
(Laemmli 1970). Samples were resolved in SDS–
PAGE (12% acrylamide for resolving gel, 4% acryl-
amide for stacking gel) using a Mini Protean II
apparatus (BIORAD). Proteins were then electro-
phoretically transfered from unstained gels onto
PVDF membranes using a transfer apparatus (Mini
Transblot Cell, BIORAD). After transfer, membranes
were saturated with 5% skimmed milk in TBS con-
taining 0.1% Tween and labelled for 1 h with primary
antibodies either (1) anti phycoerythrin 1:20,000
dilution, or (2) anti-human erythrocyte carbonic an-
hydrase II antibody, 1:10,000 dilution, or (3) rabbit
anti-b carbonic anhydrase from Synecchococcus sp.,
1:10,000 dilution, in TBS containing 1% skimmed
milk and 0.1% Tween� 20). Membranes were then
rinsed and incubated for 1 h with secondary anti-
bodies (horseradish peroxidase-linked anti-rabbit IgG,
Sigma, 1:2,500 dilution in TBS containing 1% skim-
med milk and 0.1% Tween� 20). Immunoreactive
dots were then revealed with an ECL kit (GE
Healthcare). Controls were made with the preimmune
serum as the primary antibody.
Preparation of samples for immunolocalization
• Demineralized samples: One polyp was fixed in 3%
paraformaldehyde in S22 buffer (NaCl 450 mM,
KCl 10 mM, MgCl2 58 mM, CaCl2 10 mM, Hepes
100 mM, pH 7.8) at 4�C overnight and then
decalcified using 0.5 M EDTA in Ca-free S22 with
3% PAF at 4�C. It was then dehydrated in an
ethanol series, cleared with xylene and embedded
in Paraplast. Cross sections (7 lm thick) were cut
and mounted on silane-coated glass slides.
• Mineralized samples: One polyp including skele-
ton was fixed in 3% paraformaldehyde in S22
Mar Biol
123

buffer (NaCl 450 mM, KCl 10 mM, MgCl258 mM, CaCl2 10 mM, Hepes 100 mM, pH 7.8)
at 4�C overnight. It was then dehydrated in
ethanol and embedded in LR White resin. Sec-
tions were cut with a low speed saw (Buehler,
Isomet) in thick slices (about 1 mm), etched with
EDTA 1% for 1 h to expose antigenic epitopes
and rinsed in ultrapure water.
Immunolocalization of carbonic anhydrase
Deparaffinized sections of tissues or samples of skele-
ton prepared as described above were saturated with
5% BSA in 0.05 M PBS, pH 7.4, containing 0.2%,
teleostean gelatin, 0.2% Triton X100. The samples
were then incubated with primary antibodies from
rabbit anti-b carbonic anhydrase 1:1,000 dilution in
BSA-saturated PBS solution (PBS 0.05 M, pH 7.4,
containing 0.2%, teleostean gelatin, 0.2% Triton X100,
5% BSA), 1 h at RT and overnight at 4�C in moist
chamber. After rinsing in BSA-saturated PBS solution,
they were incubated with biotinylated anti-rabbit
antibodies (GE Healthcare 1:250 dilution, 1 h at RT)
as secondary antibodies. They were finally incubated
for 20 min with streptavidin-Alexa Fluor 568 (Molec-
ular probes, 1:50 dilution) and DAPI (2 lg ml–1, SIG-
MA). Controls were routinely performed with the
rabbit preimmune serum as the primary antibody.
Samples were embedded in Pro-Long antifade medium
(Molecular probes) and observed with a confocal laser
scanning microscope (Leica, TCS4D) at 568 nm exci-
tation, 600 nm emission.
Histology
Cross sections of demineralized samples or thick slices
of mineralized samples (see paragraphs above for
preparation) were stained with hemalun, eosin, and
acetified anilin blue solutions.
Media and chemicals
Unless otherwise specified, all chemicals were obtained
from Sigma or Biorad and were of analytical grade.
FSW was obtained by filtering seawater on 0.22 lm
Millipore membranes.
The carbonic anhydrase inhibitor ethoxyzolamide
was dissolved in DMSO to a concentration of 60 mM
and buffered with 1 M Tris to pH 8.2.
Protein concentration was measured in a microplate
using the BCA Protein Assay Kit (Uptima�UP40840A). BSA was used as a standard.
Statistical analysis of the data
The effect of the carbonic anhydrase inhibitor, eth-
oxyzolamide, on calcification rate was tested using a t
test (software Jump 5.1, SAS Institute, Cary, USA).
Results are considered statistically significant when
P < 0.05.
Results
The approach we have taken towards the long-range
goal of understanding the mechanisms of biomineral-
ization in corals has been to characterize Tubastrea
aurea carbonic anhydrase and then to determine its
role in the calcification process. We report results of
the measurement of enzyme activity in tissues and or-
ganic matrix and the effect of inhibition of carbonic
anhydrase on the calcification rate. We have revealed
by Western blotting and immunohistochemistry the
presence of a protein both in the tissues and in the
organic matrix, which reacts with an antiserum against
prokaryotic carbonic anhydrases.
Carbonic anhydrase activity
Measurements performed with varying concentrations
of tissues or organic matrix showed that, in both cases,
the activity increases as a linear function of protein
quantity (Fig. 1a, b). When normalized to mg of pro-
teins, a mean value of 0.084 units of enzyme activity
was obtained for tissues and a mean value of 61.6 units
of enzyme activity was obtained for organic matrix.
This difference in the activity value may be due to a
standardization artefact (see Discussion). Dose–re-
sponse experiments were performed with the homo-
genates of tissues (Fig. 2a) or the organic matrix
(Fig. 2b) in the presence of the carbonic anhydrase
inhibitor, ethoxyzolamide. The IC50 was 200 times
higher for the organic matrix (600 nM) than for the
tissues (3 nM). Organic matrix boiled at 100�C for
10 min did not show any carbonic anhydrase activity.
Effect of inhibition of carbonic anhydrase activity
on the calcification rate
In order to determine if carbonic anhydrase was
involved in the calcification process, we measured
the uptake and deposition of calcium in the absence
and in the presence of the carbonic anhydrase inhibitor
ethoxyzolamide. We first determined the efflux time
corresponding to the emptying of 45Ca from the coel-
enteric compartment (Tambutte et al. 1995). Figure 3
Mar Biol
123

shows the kinetics of 45Ca efflux from nubbins loaded
during 135 min in labelled seawater. 45Ca released
from nubbins displays a saturation curve with a plateau
reached within 100 min of efflux. Semi-logarithmic
treatment of the results (inset of Fig. 3) indicates a
T1/2 (half-time of exchange) for calcium washout of
25 min corresponding to a rate constant of 0.028 min–1.
The volume of the coelenteric compartment calculated
at equilibrium gives a value of 12.71 ± 3.6 ll mg–1
protein.
Following this experiment, an efflux time of 180 min
was chosen to completely rinse the coelenteric cavity.
We determined that incorporation of 45Ca in killed
samples was less than 1% and thus could be considered
as negligible. We measured the calcification rate
(deposition of calcium in the skeleton) in the absence
and in the presence of ethoxyzolamide (Fig. 4). The
results show that the incorporation of Ca2+ in the HCl-
soluble pool corresponding to the skeleton has a value of
5.88 ± 0.81 nmol Ca2+ h–1 mg–1 protein or if expressed
per g of skeleton, 0.43 ± 0.09 lmol Ca2+ h–1 g–1skele-
ton. Ethoxyzolamide significantly affected this uptake
(t test, P = 0.004).
1.2
1.0
0.8
0.6
0.4
0.2
0.0
CA
act
ivit
y (E
U)
1614121086420
Quantity of proteins (mg 10-3)
1.2
1.0
0.8
0.6
0.4
0.2
0.0
CA
act
ivit
y (E
U)
1614121086420
Quantity of proteins (mg)
A
B
Fig. 1 Carbonic anhydrase activity in tissues (a) and organicmatrix (b) of T. aurea expressed as a function of the quantity ofproteins in the samples. Each point represents the mean of threevalues
100
80
60
40
20
0Per
cen
tag
e o
f in
hib
itio
n
100x10-3806040200
Concentration of EZ (µM)
A
100
80
60
40
20
0Per
cen
tag
e o
f in
hib
itio
n
1086420
Concentration of EZ (µM)
B
Fig. 2 Inhibition of carbonic anhydrase activity of T. aurea inthe presence of ethoxyzolamide. a Tissues. b Organic matrix.Each point represents the mean of three values
1.6x106
1.4
1.2
1.0
0.8
0.6
0.4
0.2
0.0
dp
m.m
g p
rote
in-1
200150100500Time (min)
15.0
14.0
13.0
12.0
Ln
(Q
e-Q
)
100806040200Time (min)
Fig. 3 Kinetics of 45Ca efflux from T. aurea nubbins loaded for2 h in labelled seawater. Data are expressed in dpm (disintegra-tions per minute) and normalized per g of dried skeleton. Eachpoint represents the mean of three values. Inset: Ln(Qe – Q) as afunction of time. Qe represents the equilibrium value and Q thevalue for each time
Mar Biol
123
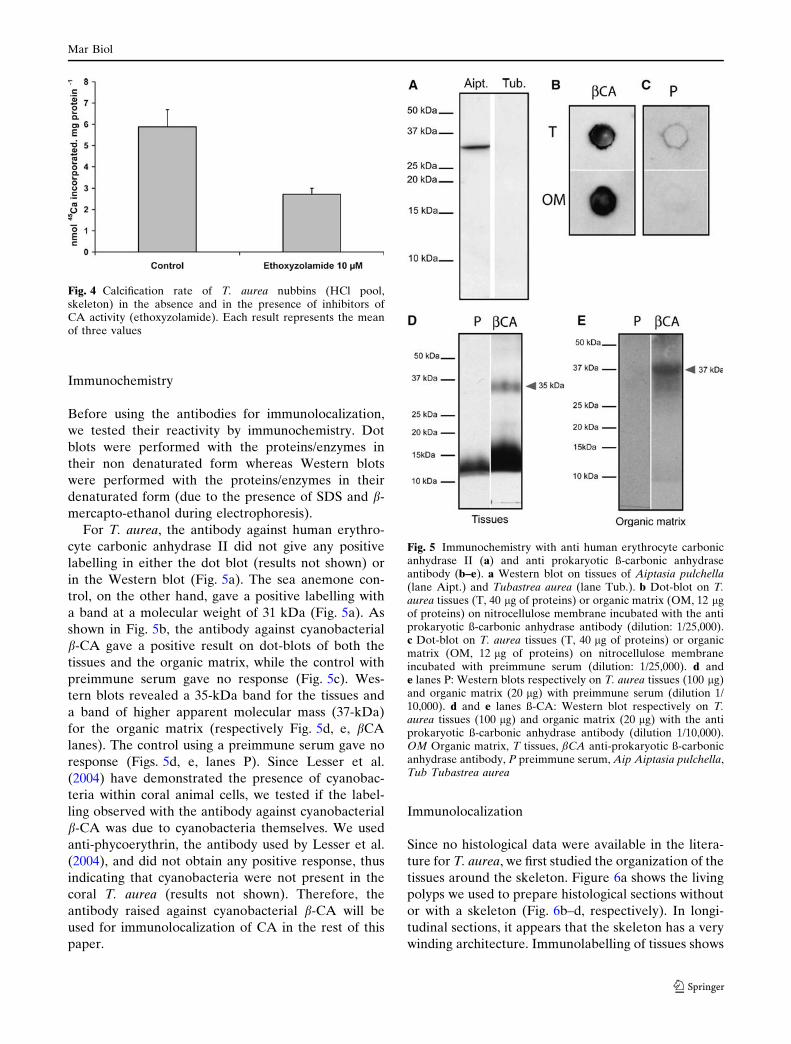
Immunochemistry
Before using the antibodies for immunolocalization,
we tested their reactivity by immunochemistry. Dot
blots were performed with the proteins/enzymes in
their non denaturated form whereas Western blots
were performed with the proteins/enzymes in their
denaturated form (due to the presence of SDS and b-
mercapto-ethanol during electrophoresis).
For T. aurea, the antibody against human erythro-
cyte carbonic anhydrase II did not give any positive
labelling in either the dot blot (results not shown) or
in the Western blot (Fig. 5a). The sea anemone con-
trol, on the other hand, gave a positive labelling with
a band at a molecular weight of 31 kDa (Fig. 5a). As
shown in Fig. 5b, the antibody against cyanobacterial
b-CA gave a positive result on dot-blots of both the
tissues and the organic matrix, while the control with
preimmune serum gave no response (Fig. 5c). Wes-
tern blots revealed a 35-kDa band for the tissues and
a band of higher apparent molecular mass (37-kDa)
for the organic matrix (respectively Fig. 5d, e, bCA
lanes). The control using a preimmune serum gave no
response (Figs. 5d, e, lanes P). Since Lesser et al.
(2004) have demonstrated the presence of cyanobac-
teria within coral animal cells, we tested if the label-
ling observed with the antibody against cyanobacterial
b-CA was due to cyanobacteria themselves. We used
anti-phycoerythrin, the antibody used by Lesser et al.
(2004), and did not obtain any positive response, thus
indicating that cyanobacteria were not present in the
coral T. aurea (results not shown). Therefore, the
antibody raised against cyanobacterial b-CA will be
used for immunolocalization of CA in the rest of this
paper.
Immunolocalization
Since no histological data were available in the litera-
ture for T. aurea, we first studied the organization of the
tissues around the skeleton. Figure 6a shows the living
polyps we used to prepare histological sections without
or with a skeleton (Fig. 6b–d, respectively). In longi-
tudinal sections, it appears that the skeleton has a very
winding architecture. Immunolabelling of tissues shows
Fig. 4 Calcification rate of T. aurea nubbins (HCl pool,skeleton) in the absence and in the presence of inhibitors ofCA activity (ethoxyzolamide). Each result represents the meanof three values
Fig. 5 Immunochemistry with anti human erythrocyte carbonicanhydrase II (a) and anti prokaryotic ß-carbonic anhydraseantibody (b–e). a Western blot on tissues of Aiptasia pulchella(lane Aipt.) and Tubastrea aurea (lane Tub.). b Dot-blot on T.aurea tissues (T, 40 lg of proteins) or organic matrix (OM, 12 lgof proteins) on nitrocellulose membrane incubated with the antiprokaryotic ß-carbonic anhydrase antibody (dilution: 1/25,000).c Dot-blot on T. aurea tissues (T, 40 lg of proteins) or organicmatrix (OM, 12 lg of proteins) on nitrocellulose membraneincubated with preimmune serum (dilution: 1/25,000). d ande lanes P: Western blots respectively on T. aurea tissues (100 lg)and organic matrix (20 lg) with preimmune serum (dilution 1/10,000). d and e lanes ß-CA: Western blot respectively on T.aurea tissues (100 lg) and organic matrix (20 lg) with the antiprokaryotic ß-carbonic anhydrase antibody (dilution 1/10,000).OM Organic matrix, T tissues, ßCA anti-prokaryotic ß-carbonicanhydrase antibody, P preimmune serum, Aip Aiptasia pulchella,Tub Tubastrea aurea
Mar Biol
123

that the antibody binds to the tissues facing the skeleton
(Fig. 7a). Preimmune serum was used as a control to
evaluate non-specific immunoreactivity (Fig. 7b), the
faint fluorescence observed was due to the autofluo-
rescence of the tissues (the same faint fluorescence is
observed in control experiments with no preimmune
and no antibody, results not shown). At higher magni-
fication (Fig. 7c, d) it appears that labelling of the tis-
sues is localized in only one layer of cells. When
performed on the skeleton, the labelling appears as a
reticulate network on the envelopes surrounding the
fibres (Fig. 7e). Figure 7f shows that the preimmune
serum gave no background signal on the skeleton.
Discussion
In the present work, we investigated carbonic anhydrase
from the tissues and the skeletal fraction of
T. aurea by (1) measuring the enzyme activity, (2)
determining its involvement in biomineralization, (3)
determining some of its biochemical properties.
Measurement of carbonic anhydrase activity
While it appears that cellular carbonic anhydrase plays a
key role in the availability of carbon involved in differ-
ent physiological processes, there are few studies deal-
ing with the presence and the role of extracellular CA in
the calcium carbonate deposition process. In calcified
skeletal structures of invertebrates, where calcium car-
bonate is the major component, a set of molecules
grouped under the term of ‘‘organic matrix’’ are always
present (Weiner 1984; Wheeler and Sikes 1984;
Constantz and Weiner 1988; Falini et al. 1996). The
roles of these molecules in the calcification process
are numerous: initiation/inhibition of crystal growth,
crystal morphology, and calcium binding (for review
see Wheeler et Sikes 1984; Watanabe et al. 2003).
Miyamoto et al. (1996) were the first to discover a
carbonic anhydrase domain within nacrein, a soluble
organic matrix protein of the nacreous layer in the
mollusc Pinctada fucata. Since then, a cDNA that
encodes a shell matrix protein composed of carbonic
anhydrase-like domains has been cloned in the oyster
Pinctada maxima by Kono et al. (2000). Watanabe et al.
(2003) have also found an internal sequence in T. aurea
that exhibits similarity to a part of the carbonic anhy-
drase sequences. CA activity was also recently found in
various biominerals (Borelli et al. 2003; Rahman 2005,
2006) suggesting a widespread distribution of CA in
calcium carbonate biominerals. In the present work we
measured carbonic anhydrase activity in the organic
matrix of T. aurea. The value of this activity is higher for
organic matrix than for tissues, which can be due either
to (1) a truly higher activity in the organic matrix, (2) a
different CA between tissues and the organic matrix, (3)
an artefact with standardization methods related to
protein assays. Indeed, invertebrate organic matrix
proteins have special biochemical features which render
their characterization difficult, for example when per-
forming staining after electrophoresis (Gotliv et al.
2003). A similar reason could explain the problems
encountered when using protein assays on the organic
matrix that probably underestimate the protein content
(unpublished results). Nevertheless the important point
to consider is that a carbonic anhydrase is present in the
organic matrix and that this enzyme possesses an
activity. Carbonic anhydrase activity in the organic
matrix is inhibited by ethoxylamide and the inhibition
constant IC50 in this matrix is 200 times higher than for
tissues. It is important to note the high resistance of the
organic matrix-linked CA, because its activity is pre-
served after the long demineralization and purification
steps necessary to obtain organic matrix itself.
Determination of CA involvement
in biomineralization
We looked at the role of CA in biomineralization by
using a pharmacological approach. We first measured
the calcification rate in control conditions and then tes-
ted the effect of CA inhibitors. We determined that the
value of calcium deposition in the skeleton is similar (i.e.
0.43 ± 0.09 lmol Ca2+ h–1 g–1 skeleton) to the value
obtained by Marshall (1996) on the same species (i.e.
0.48 ± 0.03 lmol Ca2+ h–1 g–1 skeleton). The volume of
the coelenteric cavity (i.e. 12.71 ± 3.6 ll mg–1 protein) is
comparable to the value obtained for the scleractinian
coral S. pistillata (i.e. 7.3 ± 1.2 ll mg–1 protein, Tambutte
et al. 1995). Our results on the inhibition of calcium
deposition into the skeleton in the presence of ethoxy-
zolamide indicate that CA is involved in the calcification
process. This type of inhibition of the calcification rate
was also observed in hermatypic corals (Goreau 1959;
Tambutte et al. 1996; Furla et al. 2000), sea urchin
spines (Heatfield 1970), barnacle shells (Yule et al. 1982),
molluscans (Wilbur and Jodrey 1955), crustaceans
(Roer 1980) and the red coral Corallium rubrum
(Allemand et al. 1992). Using the same kind of approach
with 45Ca, Kingsley and Watabe (1987) obtained the
opposite results in the gorgonian Leptogorgia virgulata
with an increase of Ca uptake in the spicules and the
axes in the presence of carbonic anhydrase inhibitors.
Nevertheless, while the mechanism of CA action seems
to differ depending on the species, in all cases, the results
show that CA is involved in the calcification process
Mar Biol
123

Depending on the source of carbon used at the site
of calcification, two hypothesis can be proposed:
1. If seawater HCO3– is the source of DIC, CA may
catalyze its conversion to CO2 to buffer the acidity
produced by the conversion of HCO3– into CO3
2–, as
already suggested by Sikes et al. (1980) for cocco-
lithophorids, following the equations:
HCO3- CO3
2- + H+
H+ + HCO3- CA CO2 + H2O
_________________________2 HCO3
- CO32- + CO2 + H2O
2. If intracellular CO2 is the form of DIC used at the
site of calcification, then the extracellular organic
matrix-linked CA may help in converting this CO2,
which diffuses from the tissue to the skeletogenic
fluid due to the probable high pH of this calcifying
region (Furla et al. 2000; Al-Horani et al. 2003),
into HCO3–, following the equations:
CO2 + H2OCA H+ + HCO3
-
HCO3- CO3
2- + H+
_________________________CO2 + H2O 2H+ + CO3
2-
The two H+ produced by these sets of reactions may
then be removed from the site of calcification by the
Ca2+-ATPase present within the calicoblastic epithe-
lium (Zoccola et al. 2004) which catalyzes the exchange
2H+/Ca2+.
Determination of some CA properties
Since we have shown that CA is present in both the
tissues and the organic matrix and we demonstrated
that this enzyme plays a role in the calcification process,
we tried to constrain some of its biochemical properties
and its localization both in the tissues and the organic
matrix. Five groups of cellular CA are described in the
literature (Hewett-Emmett and Tashian 1996; Cox
et al. 2000; Lane et al. 2000, 2005) (1) a-CA, mostly
found in eukaryotes, (2) b-CA, characteristic of eu-
bacteria and plants (3) c-CA, characteristic of archae-
bacteria (4) d-CA and e-CA described in the marine
diatom Thalassiosira weisflogii. The enzymes of the
type alpha, beta, and gamma use zinc as cofactor
whereas delta-CA can switch between zinc, cadmium
and cobalt, and epsilon-CA uses cadmium. Weis and
Reynolds (1999) have shown, by Western blotting with
an antibody raised against human CA, the presence of
an a-carbonic anhydrase in the tissues of a sea anem-
Fig. 6 Structure of T. aureapolyps. a Fresh polyps.b Longitudinal section of ademineralized polyp showingthe tissues. c Longitudinalsection of a nondemineralized polyp.d Higher magnification ofsection C showing tissuesabove the skeleton. Coecoenosarc (tissue betweenpolyps), Mo mouth, Po polyp,Sk skeleton, SW seawater
Mar Biol
123

one. We found the same result in the present study
using the sea anemone A. pulchella. Watanabe et al
(2003) found in T. aurea an internal sequence of a
matrix protein that exhibits sequence similarity with a-
CA sequences. However, in the present study, in T.
aurea, we could not detect CA using an immunological
approach with an anti a-CA antibody. Thus, if an a-CA
is present in T. aurea, it possesses epitopes in the cat-
alytic site that are not recognized by the antibody
against human a-CA. This result could be due to a
difference in structure between mammal a-CA and
coral a-CA. On the other hand, Western blotting with
an antibody raised against a carbonic anhydrase from
Synecchococcus sp. shows that there is a protein that
immunoreacts with this antibody raised against a
prokaryotic CA in both the tissues and in the organic
matrix. Since no labelling was obtained with the anti-
body against phycoerythrin, a cyanobacterial marker,
we can suggest that the labelling observed with the
antibody against the prokaryotic CA is specific and not
due to the presence of cyanobacteria in coral tissues.
The presence of a protein, which shares properties
with enzymes found in prokaryotes appears surpris-
ing. However, prokaryotic-like proteins have already
Fig. 7 Immunolocalization ofprokaryotic ß-carbonicanhydrase respectively in thetissues (a–d) and in theskeleton (e and f) of T. aurea.a, e In orange: carbonicanhydrase antibody coupledto Alexafluor 568. b Controlwith preimmune serum. c, dIn orange: carbonic anhydraseantibody coupled toAlexafluor 568 merged withDAPI staining showing thenuclei in blue. f Control withpreimmune serum showing nosignal in the skeleton. SKSkeleton
Mar Biol
123

been described in Cnidarians. For example, Richier
et al. (2003) found an extra-mitochondrial, monomeric
Mn-superoxide dismutase and a Fe-superoxide dismu-
tase, both enzymes characteristic of prokaryotes, within
the tissues of a sea anemone. More recently, by study-
ing 26,845 ESTs from a coral and a sea anemone,
Technau et al. (2005) found that about 1.3–2.7% of
cnidarian proteins only matched with non-metazoan
sequences (i.e. fungi, prokaryotes, plants, and protists),
many matching only with bacterial sequences. Among
these bacterial sequences, they identified the bacterial
universal stress protein (UspA). To explain this result,
these authors suggested either a conservation of ancient
genes within the genomes of basal metazoans or lateral
gene transfer. It is noteworthy that such observations
are not only limited to cnidarians since in tunicates it
has also recently been suggested that enzymes involved
in cellulose biosynthesis are likely acquired by hori-
zontal transfer from bacteria (Sasakura et al. 2005).
The protein characterized in our Western-blots by
its immunoreactivity with the prokaryotic CA antibody
has an apparent molecular mass in the tissues of
35 kDa, which is smaller than the one found in the
organic matrix (37 kDa). This difference can be ac-
counted for by oligosaccharide chains (Waheed et al.
1992; Wilson et al. 2000) since, in corals, matrices are
highly glycosylated (Dauphin 2001). When chemical
fluorochrome staining or immunolocalization was per-
formed with this antibody on the skeleton, labelling
displayed a pattern typical of the organic matrix
(Gautret et al. 2000; Puverel et al. 2005). The existence
of a protein positively labelled by the same antibody
both in the tissues and in the organic matrix suggests
that high homologies exist between these two proteins,
the latter could be the secreted form of the former. In
this case, however, since we have determined that
carbonic anhydrases react differently to inhibitors, it is
probable that these two enzymes are different iso-
forms. Another possibility is that the pharmacologic
differences between these two isoforms only result
from a modification, before secretion of the protein, by
glycosylation. Further characterization is needed to
solve this point. Furthermore, the differences in
molecular weight observed in Western-blot confirms
that, as in zooxanthellate corals (Puverel et al. 2005),
organic matrix proteins of azooxanthellate corals do
not result from the trapping of the whole soft tissues as
suggested by Constantz (1986). By immunohisto-
chemistry, we showed that only the tissues facing the
skeleton (i.e. the calcifying tissues) are labeled with the
antibody raised against prokaryotic CA, suggesting
that this protein is involved in the biomineralization
process.
Conclusions
Our work was performed on the azooxanthellate coral
T. aurea, allowing us to study the transport of carbon
used for calcification while eliminating the complica-
tions due to photosynthesis. We demonstrated the
presence of an active carbonic anhydrase both in the
tissues and in the organic matrix with a direct role of
this enzyme in the calcification process. We suggest
that this activity may be due a protein, which shares
common features with prokaryotic CA. This protein
shows similar features in the tissues and in the organic
matrix suggesting that the calcifying tissues could be
responsible for the secretion of this protein. This result
does not exclude the possibility that other types of CA
are responsible for the activity observed in the tissue
and the organic matrix. Our results also demonstrate
that in corals, organic matrix proteins are not only
structural proteins but also catalytic proteins and pro-
vide a crucial enzyme to eliminate the kinetic barrier in
the conversion of inorganic carbon. This new under-
standing of the chemistry in the calcifying region is
essential to account for the mechanisms underlying the
carbon and oxygen isotope fractionations seen in
skeletal carbonates (Adkins et al. 2003).
Acknowledgments We thank Prof. Francois Morel fromPrinceton University and Mak Saıto from the Woods HoleOceanographic Institution for providing the antibody, anti-b-carbonic anhydrase from Synecchococcus sp. This study wasconducted as part of the Centre Scientifique de Monaco 2000–2004 research program. It was supported by the Government ofthe Principality of Monaco and by the California Institute ofTechnology, USA.
References
Adkins JF, Boyle EA, Curry WB, Lutringer A (2003) Stableisotopes in deep-sea corals and a new mechanism for ‘‘vitaleffects’’. Geochim Cosmochim Acta 67:1129–1143
Al-Horani FA, Al-Moghrabi SM, de Beer D (2003) Themechanism of calcification and its relation to photosynthesisand respiration in the scleractinian coral Galaxea fascicu-laris. Mar Biol 142:419–426
Allemand D, Grillo M-C (1992) Biocalcification mechanisms ingorgonians. 45Ca uptake and deposition by the mediter-ranean red coral Corallium rubrum. J Exp Zool 292:237–246
Borelli G, Mayer-Gostan N, Merle P-L, de Pontual H, Boeuf G,Allemand D, Payan P (2003) Composition of biomineralorganic matrices with special emphasis on turbot (Psettamaxima) otolith and endolymph. Calcified Tissue Int72:717–725
Buddemeier RW, Kinzie RA (1976) Coral growth. OceanogrMar Biol Annu Rev 14:183–225
Constantz BR. (1986) Coral skeleton construction: a physio-chemically dominated process. Palaios 1:152–157
Mar Biol
123

Constantz B, Weiner S (1988) Acidic macromolecules associatedwith the mineral phase of scleractinian coral skeletons.J Exp Zool 248:253–258
Cox EH, McLendon GL, Morel F, Lane T, Prince RC, PickeringIJ, George GN (2000) The active site of Thalassiosiraweissflogii carbonic anhydrases 1. Biochem 39:12128–12130
Cuif JP, Dauphin Y, Doucet J, Salome M, Susini J (2003)XANES mapping of organic sulfate in three scleractiniancoral skeletons. Geochim Cosmochim Acta 67:75–83
Cuif JP, Dauphin Y, Gautret P (1999) Compositional diversity ofsoluble mineralizing matrices in some recent coral skeletonscompared to fine-scale growth structures of fibres: discussionof consequences for biomineralization and diagenesis. Int JEarth Sci 88:582–592
Dauphin Y (2001) Comparative studies of skeletal solublematrices from some Scleractinian corals and Molluscs. IntJ Biol Macromol 28:293–304
Falini G, Albeck S, Weiner S, Addadi L (1996) Control ofaragonite or calcite polymorphism by mollusk shell macro-molecules. Science 271:67–69
Furla P, Galgani I, Durand I, Allemand D (2000) Sources andmechanisms of inorganic carbon transport for coral calcifi-cation and photosynthesis. J Exp Biol 203:3445–3457
Gautret P, Cuif JP, Freiwald A (1997) Composition of solublemineralizing matrices in zooxanthellate and non-zooxan-thellate scleractinian corals: biochemical assessment ofphotosynthetic metabolism through the study of a skeletalfeature. Facies 36:189–194
Gautret P, Cuif JP, Stolarski J (2000) Organic components of theskeleton of scleractinian corals—evidence from in situacridine orange staining. Acta Palaeontol Pol 45:107–118
Goreau TF (1959) The physiology of skeleton formation incorals. I. A method for measuring the rate of calciumdeposition by corals under different conditions. Biol BullMar Biol Lab Woods Hole 116:59–75
Gotliv BA, Addadi L, Weiner S (2003) Mollusk shell acidicproteins: in search of individual functions. Chembiochem4:522–529
Heatfield BM (1970) Calcification in echinoderms: effects oftemperature and acetazolamide on incorporation of cal-cium-45 in vitro by regenerating spines of Strongylocentrotuspurpuratus. Biol Bull 139:151–163
Hewett-Emmet D, Tashian RE (1996) Functional diversity,conservation and convergence in the evolution of thea-,b-,c,-carbonic anhydrase gene families. Mol PhylogenEvol 5:50–77
Isa Y, Yamazato K (1984) The distribution of carbonic anhydrasein a staghorn coral Acropora hebes (Dana). Galaxea 3:25–36
Jones WC, Ledger PW (1986) The effect of acetazolamide andvarious concentrations of calcium on spicule secretion in thecalcareous sponge Sycon ciliatum. Comp Biochem Physiol84A:149–158
Kingsley RJ, Watabe N (1987) Role of carbonic anhydrase incalcification in the gorgonian Leptogorgia virgulata. J ExpZool 241:171–180
Kono M, Hayashi N, Samata T (2000) Molecular mechanism ofthe nacreous layer formation in Pinctada maxima. BiochemBiophys Res Comm 269:213–218
Laemmli UK (1970) Cleavage of structural proteins during theassembly of the head bacteriophage T4. Nature 227:680–685
Lane TW, Morel F (2000) A biological function for cadmium inmarine diatoms. PNAS 97(9):4627–4631
Lane TW, Saito MA, George GN, Pickering IJ, Prince RC,Morel MM (2005) A cadmium enzyme from a marinediatom. Nature 435(7038):42
Lesser MP, Mazel CH, Gorbunov MY, Falkowski PG (2004).Nitrogen-fixing cyanobacteria in corals. Science 305:997–1000
Lucas JM, Knapp LW (1996) Biochemical characterization ofpurified carbonic anhydrase from the octocoral Leptogorgiavirgulata. Mar Biol 126:471–477
Lucas JM, Knapp LW (1997) A physiological evaluation ofcarbon sources for calcification in the octocoral Leptogorgiavirgulata (Lamarck). J Exp Biol 200:2653–2662
Marshall AT (1996) Calcification in hermatypic and ahermatypiccorals. Science 271:637–639
Mitsunaga K, Akasaka K, Shimada H, Fujino Y, Yasumasu I,Numandi H (1986) Carbonic anhydrase activity in develop-ing sea urchin embryos with special reference to calcificationof spicules. Cell Differ 18:257–262
Miyamoto H, Miyashita T, Okushima M, Nakano S, Morita T,Matsushiro A (1996) A carbonic anhydrase from thenacreous layer in oyster pearls. Proc Natl Acad Sci USA93:9657–9660
Nys Y, de Laage X (1984) Effects of suppression of egg shellcalcification and of 1,25 (OH)2D3 on Mg2+, Ca2+ and Mg2+
HCO3– ATPase, alkaline phosphatase, carbonic anhydrase
and CaBP levels. II. The laying intestine. Comp BiochemPhysiol 78A:839–844
Payan P, Kossmann H, Watrin A, Mayer-Gostan N, Boeuf G(1997) Ionic composition of endolymph in teleosts: originand importance of endolymph alkalinity. J Exp Biol200:1905–1912
Pearse VB (1970) Incorporation of metabolic CO2 into coralskeleton. Nature 228:383
Puverel S, Tambutte E, Zoccola D, Domart-Coulon I, BouchotA, Lotto S, Allemand D, Tambutte S (2005) Antibodiesagainst the organic matrix in scleractinians: a new tool tostudy coral biomineralization. Coral Reefs 24:149–156
Rahman A, Isa Y, Uehara T (2005). Proteins of calcifiedendoskeleton. II. partial amino acid sequences of endoskel-etal proteins and the characterization of proteinaceousorganic matrix of spicules from the alcyonarian, Synulariapolydactyla. Proteomics 5:1–9
Rahman A, Isa Y, Uehara T (2006). Studies of two closelyrelated species of Octocorallians: biochemical and molecu-lar characteristics of the organic matrices of endoskeletalsclerites. Mar Biotech 8:415–424
Richier S, Merle PL, Furla P, Pigozzi D, Sola F, Allemand D(2003) Characterization of superoxide dismutases in anoxia-and hyperoxia-tolerant symbiotic cnidarians. Biochim Bio-phys Acta 1621(1):84–91
Roer RD (1980) Mechanisms of resorption and deposition ofcalcium in the carapace of the crab Carcinus maenas. J ExpBiol 88:205–218
Sasakura Y, Nakashima K, Awazu S, Matsuoka T, Nakayama A,Azuma J, Satoh N (2005) Transposon-mediated insertionalmutagenesis revealed the functions of animal cellulosesynthetase in the ascidian Ciona intestinalis. PNAS102(42):15134–15139
Sikes CS, Roer RD, Wilbur KM (1980) Photosynthesis andcocolith formation: Inorganic carbon sources and net inor-ganic reaction of deposition. Limnol Oceanogr 25:248–261
Tambutte E, Allemand D, Bourge I, Gattuso J-P, Jaubert J(1995) An improved 45Ca protocol for investigating physi-ological mechanisms in coral calcification. Mar Biol122:453–459
Tambutte E, Allemand D, Mueller E, Jaubert J (1996) Acompartmental approach to the mechanism of calcificationin hermatypic corals. J Exp Biol 199:1029–1041
Mar Biol
123

Technau U, Rudd S, Maxwell P, Gordon PMK, Saina M, GrassoLC, Hayward DC, Sensen CW, Saint R, Holstein TW, BallEE, Miller D (2005) Maintenance and ancestral complexityand non-metazoan genes in two basal cnidarians. TrendsGenet 21(12):633–639
Tohse H, Ando H, Mugiya Y (2004) Biochemical properties andimmunohistochemical localization of carbonic anhydrase inthe sacculus of the inner ear in the salmon Oncorhyncusmasou. Comp Biochem Physiol 137A:87–94
Tohse H, Mugiya Y (2001) Effects of enzyme and anion transportinhibitors on in vitro incorporation of inorganic carbon andcalcium into endolymph and otoliths in salmon Oncorhyn-chus masou. Comp Biochem Physiol 128A:177–184
Waheed A, Zhu XL, Sly WS (1992) Membrane-associatedcarbonic anhydrase from rat lung. J Biol Chem 267:3308–3311
Watanabe T, Fukuda I, China K, Isa Y (2003) Molecularanalyses of protein components of the organic matrix in theexoskeleton of two scleractinian coral species. Comp Bio-chem Physiol 136B:767–774
Weiner S (1984) Organization of organic matrix components inmineralized tissues. Amer Zool 24:945–951
Weis VM, Reynolds WS (1999) Carbonic anhydrase expressionand synthesis in the sea anemone Anthopleura elegantissima
are enhanced by the presence of dinoflagellate symbionts.Physiol Biochem Zool 72:307–316
Weis VM, Smith GJ, Muscatine L (1989) A ‘‘CO2 supply’’mechanism in zooxanthellate cnidarians: role of carbonicanhydrase. Mar Biol 100:195–202
Wheeler AP, Sikes CS (1984) Regulation of carbonate calcifica-tion by organic matrix. Am Zool 24:933–944
Wilbur KM, Jodrey LH (1955) Studies on shell formation. V.The inhibition of shell formation by carbonic anhydraseinhibitors. Biol Bull 108:359–365
Wilson JM, Randall DJ, Vogl AW, Harris J, Sly WS, Iwama GK(2000) Branchial carbonic anhydrase is present in thedogfish, Squalus acanthias. Fish Physiol Biochem 22:329–336
Wright OP, Marshall AT (1991) Calcium transport across theisolated oral epithelium of scleractinian corals. Coral Reefs10:37–40
Yule AB, Crisp DJ, Cotton IH (1982) The action of acetazol-amide on calcification in juvenile Balanus balanoides. MarBiol Lett 3:273–288
Zoccola D, Tambutte E, Kulhanek E, Puverel S, Scimeca J-C,Allemand D Tambutte S (2004). Molecular cloning andlocalization of a PMCA P-type calcium ATPase from thecoral Stylophora pistillata. Biochim Biophys Acta 1663:117–126
Mar Biol
123




















