
Changes in plant chemical defenses and nutritional quality as a function of ontogeny in Plantago...
11
Oecologia (2012) 168:471–481 DOI 10.1007/s00442-011-2114-x 123 PLANT-ANIMAL INTERACTIONS - ORIGINAL PAPER Changes in plant chemical defenses and nutritional quality as a function of ontogeny in Plantago lanceolata (Plantaginaceae) Carolina Quintero · M. Deane Bowers Received: 10 July 2010 / Accepted: 26 July 2011 / Published online: 13 September 2011 © Springer-Verlag 2011 Abstract Numerous empirical studies have examined ontogenetic trajectories in plant defenses but only a few have explored the potential mechanisms underlying those patterns. Furthermore, most documented ontogenetic tra- jectories in plant defenses have generally concentrated on aboveground tissues; thus, our knowledge regarding whole plant trends in plant defenses throughout development or potential allocation constraints between growth and defenses is limited. Here, we document changes in plant biomass, nutritional quality and chemical defenses for below- and aboveground tissues across seven age classes of Plantago lanceolata (Plantaginaceae) to evaluate: (1) par- tial and whole plant ontogenetic trajectories in constitutive chemical defenses and nutritional quality, and (2) the role of resource allocation constraints, namely root:shoot (R:S) ratios, in explaining whole plant investment in chemical defenses over time. Overall investment in iridoid glyco- sides (IGs) signiWcantly increased, while water and nitro- gen concentrations in shoot tissues decreased with plant age. SigniWcant variation in IG content between shoot and root tissues across development was observed: allocation of IGs into root tissues linearly increased from younger to older plants, while non-linear shifts in allocation of IGs during ontogeny were observed for shoot tissues. Finally, R:S ratios only weakly explained overall allocation of resources into defenses, with young stages showing a positive relationship, while older stages showed a negative relationship between R:S ratios and IG concentrations. Ontogenetic trajectories in plant quality and defenses within and among plant tissues can strongly inXuence insect herbivores’ performance and/or predation risk; thus, they are likely to play a signiWcant role in mediating species interactions. Keywords Iridoid glycosides · Constitutive defenses · Root:shoot ratios · Resource allocation · Root defenses Introduction Plant susceptibility to herbivory, as well as the impact that herbivore damage has on plant Wtness, vary markedly over a plant’s lifetime. For example, variation in herbivore dam- age from the seedling stage to senescence can change by several orders of magnitude for the same plant–herbivore interaction (e.g., Pires and Price 2000; Fonseca et al. 2006; Loney et al. 2006), and may even involve considerable changes in the composition of the herbivore community inXicting damage throughout plant ontogeny (e.g., Waltz and Whitham 1997; Boege 2005; Thomas et al. 2010). Sim- ilarly, the impact of herbivore damage on plant Wtness also depends on the ontogenetic stage being attacked, ranging from complete compensation by the host plant to long-last- ing reductions in Xower production and fruit set (Warner and Cushman 2002; Boege et al. 2007; Hanley and Fegan 2007). As a result, the selective pressure that herbivores exert on plant traits that confer resistance should vary with plant ontogeny, leading to qualitative and/or quantitative changes in plant resistance traits during plant development. Despite the relevance that assessing developmental changes in plant defenses may have for the improvement of plant Communicated by Jacqui ShykoV. C. Quintero (&) · M. D. Bowers Department of Ecology and Evolutionary Biology and University of Colorado Museum, University of Colorado, Campus Box 334, Boulder, CO 80309-0260, USA e-mail: [email protected]
Transcript of Changes in plant chemical defenses and nutritional quality as a function of ontogeny in Plantago...

Oecologia (2012) 168:471–481
DOI 10.1007/s00442-011-2114-xPLANT-ANIMAL INTERACTIONS - ORIGINAL PAPER
Changes in plant chemical defenses and nutritional quality as a function of ontogeny in Plantago lanceolata (Plantaginaceae)
Carolina Quintero · M. Deane Bowers
Received: 10 July 2010 / Accepted: 26 July 2011 / Published online: 13 September 2011© Springer-Verlag 2011
Abstract Numerous empirical studies have examinedontogenetic trajectories in plant defenses but only a fewhave explored the potential mechanisms underlying thosepatterns. Furthermore, most documented ontogenetic tra-jectories in plant defenses have generally concentrated onaboveground tissues; thus, our knowledge regarding wholeplant trends in plant defenses throughout development orpotential allocation constraints between growth anddefenses is limited. Here, we document changes in plantbiomass, nutritional quality and chemical defenses forbelow- and aboveground tissues across seven age classes ofPlantago lanceolata (Plantaginaceae) to evaluate: (1) par-tial and whole plant ontogenetic trajectories in constitutivechemical defenses and nutritional quality, and (2) the roleof resource allocation constraints, namely root:shoot (R:S)ratios, in explaining whole plant investment in chemicaldefenses over time. Overall investment in iridoid glyco-sides (IGs) signiWcantly increased, while water and nitro-gen concentrations in shoot tissues decreased with plantage. SigniWcant variation in IG content between shoot androot tissues across development was observed: allocation ofIGs into root tissues linearly increased from younger toolder plants, while non-linear shifts in allocation of IGsduring ontogeny were observed for shoot tissues. Finally,R:S ratios only weakly explained overall allocation ofresources into defenses, with young stages showing a
positive relationship, while older stages showed a negativerelationship between R:S ratios and IG concentrations.Ontogenetic trajectories in plant quality and defenseswithin and among plant tissues can strongly inXuenceinsect herbivores’ performance and/or predation risk; thus,they are likely to play a signiWcant role in mediating speciesinteractions.
Keywords Iridoid glycosides · Constitutive defenses · Root:shoot ratios · Resource allocation · Root defenses
Introduction
Plant susceptibility to herbivory, as well as the impact thatherbivore damage has on plant Wtness, vary markedly overa plant’s lifetime. For example, variation in herbivore dam-age from the seedling stage to senescence can change byseveral orders of magnitude for the same plant–herbivoreinteraction (e.g., Pires and Price 2000; Fonseca et al. 2006;Loney et al. 2006), and may even involve considerablechanges in the composition of the herbivore communityinXicting damage throughout plant ontogeny (e.g., Waltzand Whitham 1997; Boege 2005; Thomas et al. 2010). Sim-ilarly, the impact of herbivore damage on plant Wtness alsodepends on the ontogenetic stage being attacked, rangingfrom complete compensation by the host plant to long-last-ing reductions in Xower production and fruit set (Warnerand Cushman 2002; Boege et al. 2007; Hanley and Fegan2007). As a result, the selective pressure that herbivoresexert on plant traits that confer resistance should vary withplant ontogeny, leading to qualitative and/or quantitativechanges in plant resistance traits during plant development.Despite the relevance that assessing developmental changesin plant defenses may have for the improvement of plant
Communicated by Jacqui ShykoV.
C. Quintero (&) · M. D. BowersDepartment of Ecology and Evolutionary Biology and University of Colorado Museum, University of Colorado, Campus Box 334, Boulder, CO 80309-0260, USAe-mail: [email protected]
123

472 Oecologia (2012) 168:471–481
defense theories, as well as our understanding of ecologicalinteractions, relatively little is known about how allocationof resources to plant defenses varies throughout plantdevelopment (Boege and Marquis 2005; Hanley et al.2007).
Numerous models have been proposed to explain pat-terns of variation in plant constitutive chemical defensesas a function of plant development. With the exception ofboreal woody plants, where the ontogenetic trajectories inplant defenses have been proposed to strongly reXect theselective pressure exerted by mammalian herbivores(Bryant et al. 1992), most arguments propose that intrin-sic forces, namely resource acquisition and allocationconstraints, are responsible for ontogenetic variability inplant defenses (Bryant et al. 1991; Herms and Mattson1992; Boege and Marquis 2005). Among these, the theo-retical model proposed by Boege and Marquis (2005) isthe more comprehensive and detailed model, encompass-ing all stages from cotyledon to senescent plants at thewhole plant level. This model predicts that multipleresource allocation and architectural constraints through-out plant development will lead to a non-linear relation-ship between plant age and defenses. SpeciWcally, themodel predicts relatively higher levels of defenses inyounger over older seedlings until resources stored incotyledons and seeds are depleted, followed by a decreasein defenses in response to large root:shoot (R:S) ratios anda disproportional allocation of resources to the productionof photosynthetic area. Once R:S ratios start to decrease,growth rate might be more limited than photosyntheticrate allowing for a signiWcant increase in defensesthroughout juvenile stages. Finally, when plants reachreproductive maturity, constraints due to reproductionmay again result in decreased levels of defenses or theymay remain constant depending on the plant’s reproduc-tive strategy (Boege and Marquis 2005). A recent meta-analysis conducted by Barton and Koricheva (2010),including 116 studies and 153 plant species, concludedthat ontogenetic trajectories in plant defenses tend toshow non-linear patterns. However, while non-linearityappears to be the norm, the shape of the relationshipseems to depend on plant life form (i.e. woody plants,herbs or grasses) and defensive traits (i.e. physical traits,chemical traits, tolerance) (Barton and Koricheva 2010).
The absence of a generalized pattern describing theontogeny of plant defenses suggests that allocation con-straints and costs of defenses through plant developmentare not consistent across all plant life forms and defensivetraits. Nevertheless, despite the large number of studiesdocumenting ontogenetic trajectories in plant defenses,only a few have tested the potential mechanisms drivingthese patterns, such as those looking at age-speciWc alloca-tion costs and trade-oVs between growth and defenses
(e.g., Goodger et al. 2006; Barton 2007; Boege et al. 2007;Orians et al. 2010). Hence, the potential of diVerences inallocation constraints among grasses, herbs and woodyplants, as well as among diVerent kinds of defensive traits,as the source of discrepancies in ontogenetic patterns ofdefenses remains unclear. Furthermore, inconsistencyamong results from investigations of ontogenetic patternsof plant defense might reXect the lack of studies assessingontogenetic trajectories at the whole plant level. Most stud-ies to date concentrate on aboveground tissues, especiallyleaves, while ontogenetic trajectories in root defenses havebeen largely ignored. However, the few studies that haveexplored ontogenetic trajectories in below- and above-ground tissues have consistently documented dissimilarontogenetic patterns of defenses among tissues (Hartmannand Zimmer 1986; Williams and Ellis 1989; Bellostas et al.2007; Beninger et al. 2009). Therefore, without a completeassessment of defensive traits at the whole plant level,meaningful variability within and among tissues duringplant development may obscure overall patterns of plantinvestment in defenses over time, as well as potential trade-oVs between growth and defenses.
In sum, in the last two decades, numerous attempts topredict theoretical patterns and drivers of ontogeneticvariation in constitutive plant defenses have been made;yet few studies to date have empirically tested the rela-tionship between plant development and defenses acrossmultiple developmental stages and plant tissues, and/ortested the mechanisms driving these patterns. In thisstudy, we used seven pre-reproductive age classes ofPlantago lanceolata L. (Plantaginaceae) to investigateontogenetic variation in below- and aboveground consti-tutive plant chemical defenses (iridoid glycosides), andthe role of resource allocation constraints as the poten-tial mechanism driving these ontogenetic trajectories. Inaddition, we assessed ontogenetic variation in otherplant traits relevant to herbivores, including leaf waterand nitrogen concentration, as well as below- and above-ground biomass. Because our interest was to assess over-all variation in constitutive trajectories at the wholeplant level, we did not assess here within-tissue varia-tion (e.g., young, intermediate and old leaf ages) whichmay add considerable variation in levels of defenses andnutritional quality for herbivores (e.g., Bowers andStamp 1992, 1993). In particular, we addressed the fol-lowing three questions: (1) how do plant traits relevantto herbivores, including constitutive chemical defensesand nutritional quality, change throughout plant devel-opment?; (2) do plant chemical defenses in below- andaboveground tissues follow similar ontogenetic trajecto-ries?; and (3) can R:S ratios help to predict the alloca-tion of resources to constitutive chemical defenses at thewhole plant level?
123

Oecologia (2012) 168:471–481 473
Methods
Study system
Plantago lanceolata is a cosmopolitan, short-lived peren-nial herb with a rosette growth form introduced to NorthAmerica from Eurasia more than 200 years ago (Caverset al. 1980). This plant species produces, in addition to irid-oid glycosides (Ronsted et al. 2000), a number of bioactivephenolic compounds such as Xavonoids and phenyletha-noid glycosides (Tamura 2001; Galvez et al. 2005). How-ever, iridoid glycosides are the primary allelochemicalsfound in large concentrations in both below- and above-ground tissues and inXuencing feeding preferences of gen-eralist and specialist rhizophagous and folivorousherbivores (e.g., Bowers and Puttick 1989; Biere et al.2004; De Deyn et al. 2009); thus, here we speciWcallyfocused on the ontogenetic trajectories of these compounds.
Iridoid glycosides (IGs) are a group of cyclopentanoidmonoterpene-derived compounds that occur in over 50families of plants (Jensen 1991). Plantago lanceolata pro-duces primarily two iridoid glycosides: aucubin and catal-pol, and amounts of these compounds may be as high as10–12% dry weight (Bowers and Stamp 1992, 1993). Highconcentrations of IGs, especially of catalpol, indicate notonly a greater biosynthetic investment by the plant but mayalso result in a higher unpalatability and/or toxicity to gen-eralist herbivores (Bowers and Stamp 1993). Although welack speciWc knowledge regarding IG production sites (i.e.shoots vs. roots), these carbon-based defenses are synthe-sized through the isoprenoid biosynthetic pathway and areusually stored in cell vacuoles (Croteau 1987). They arealso likely phloem mobile (Gowan et al. 1995; Beningeret al. 2007), and concentrations in leaves and roots inP. lanceolata were found to be positively correlated (DeDeyn et al. 2009). Previous studies have identiWed plant ageas an important intrinsic factor aVecting P. lanceolata chem-istry (e.g., Bowers and Stamp 1993; Fuchs and Bowers 2004;Barton 2007); and these ontogenetic trajectories in IG pro-duction signiWcantly vary among both maternal families andpopulations (Bowers and Stamp 1993; Barton 2007).
In addition, P. lanceolata has been incorporated into thediet of many North American insect herbivores, and thusmultiple specialist and generalist herbivores can aVect andbe aVected by IG content in this species. For example, highIG concentrations usually deter generalist herbivores feed-ing on shoot or root tissues (e.g., Bowers and Puttick 1989;Biere et al. 2004; De Deyn et al. 2004; Wurst and van derPutten 2007). Yet several specialist herbivores use IGs asfeeding and/or oviposition stimulants (e.g., Prudic et al.2005; Talsma et al. 2008), and some have the ability tosequester these compounds from leaves and roots and usethem for their own protection against natural enemies (e.g.,
Strohmeyer et al. 1998; Baden and Dobler 2009; Opitzet al. 2010). Furthermore, although most herbivores associ-ated with this species are either rhizophagous or folivorous,there is at least one example of a species of Longitarsus(Chrysomelidae) Xea beetle that feeds on and sequestersIGs from both tissues, as larvae and adults (Willinger andDobler 2001; Baden and Dobler 2009). As a result, ontoge-netic trajectories in constitutive variation of shoot and rootIGs in this species can have profound consequences on theidentity, abundance and performance of the herbivore com-munity associated with it throughout its lifetime.
Ontogenetic variation in plant traits
In order to evaluate the eVect of plant ontogeny on plantquality and chemical defenses, seven distinct age classes ofPlantago lanceolata (3, 5, 8, 10, 12, 15, and 18 weeks oldsince germination) were grown at the University of Colo-rado greenhouse during the summer of 2008. The Wrst twostages (3 and 5 weeks old) correspond to two distinctiveseedling stages, the next three stages (8, 10 and 12 weeksold) correspond to juvenile stages, and the last two ageclasses (15 and 18 weeks old) to pre-reproductive stages.All age classes were harvested simultaneously in order tocontrol for confounding environmental factors that couldinXuence plant performance such as photoperiod and tem-perature. Synchronization among ontogenetic stagesoccurred by germinating seeds at intervals of 15–30 daysfrom March to July. In this way, although average environ-mental conditions from sowing to harvest varied among ageclasses, we ensured that all individuals were exposed to thesame conditions at harvest time. Since IG concentrationsin this as well as in other IG-containing plant species canvary substantially over days and even hours (Hogedal andMolgaard 2000; Fuchs and Bowers 2004), or be inXuencedby temperature (Tamura 2001), simultaneous harvest acrossall age classes should minimize variables other than plantage that could inXuence IG content.
Seeds were collected from >20 maternal plants from apopulation in Boulder County, Colorado, mixed, and ran-domly assigned to treatments. Seeds were germinated inFafard mix and transplanted to a growth medium of equalparts Metro Mix 350, sterilized sand, and turface after15 days. Plants were grown in extra-deep 13 l pots in orderto minimize root-binding eVects. We grew 20 plants in eachage class, for a total of 140 plants. Replicates were ran-domly placed on a single 1.2 £ 8 m greenhouse bench,exposed to natural daylight, and watered daily. Fertilizerwas supplied to all plants every 3–4 days throughout thelength of the experiment. The fertilizer solution was thestandard solution used in the University of Colorado green-houses, which was Scotts Peter’s Excel (Scotts-SierraHorticultural Products, Marysville, OH, USA) mixed at a
123

474 Oecologia (2012) 168:471–481
ratio of 15–5–15 N–P–K which also provides trace amountsof micronutrients. Temperatures Xuctuated over the courseof the season but greenhouse temperatures did not exceed32°C or fall below 15°C at any time. Extra-deep pots wereused in order to ensure natural growth rate of root as well asshoot tissues and to prevent pot-binding, which can aVectplant secondary metabolite production (Baldwin 1988).However, we believe that the combination of abundantresources and nutrient availability together with moderategreenhouse temperatures delayed the onset of Xowering(usually recorded at 15 weeks after sowing under green-house conditions; personal observation) during this Wrstgrowing season.
At harvest, plants were separated into roots and shoots.Shoot tissues represented mostly leaves but in a few of theoldest plants (n = 14) these also contained immature inXo-rescences accounting for less than 1% of the total shoot bio-mass. Statistical analyses with and without these 14individual plants yielded similar results; thus, we presentthe results for the complete dataset. All plant tissues wereweighed fresh, oven-dried at 50°C for 48 h to a constantmass, and weighed again to the nearest 0.01 g. The ratiobetween root and shoot dry biomass was calculated for eachindividual plant.
Analyses of plant water, nitrogen and iridoid glycoside content
To assess variation in plant quality throughout develop-ment, we measured plant water content and nitrogen con-tent. Percent water was calculated as [(wet weight¡dryweight)/wet weight] £ 100. Nutritional quality was mea-sured as total leaf nitrogen, and quantiWed by Micro-Dumascombustion on a NA1500 C/H/N analyzer, using a samplesize of ten plants per age class. Due to the low biomass ofplants at the 3-week stage, we were unable to measure leafnitrogen content for those plants.
To assess concentrations of iridoid glycosides, rootsand shoots were separately ground into a Wne powder, and2–50 mg subsamples (entire above- and/or belowgroundtissue for some seedlings) were processed for IG extractionand analyzed by gas chromatography following previouslydescribed methods (Bowers and Stamp 1993; Barton 2007).BrieXy, samples were extracted overnight in 95% metha-nol, the methanol extract was Wltered to remove the plantmaterial and the residue evaporated to dryness. Phenyl-�-D-glucose (PBG) was added as an internal standard and thesamples were then partitioned between water and ether toremove hydrophobic compounds. An aliquot of the remain-ing solution was removed, evaporated, and derivatized withTri-Sil-Z™ (Pierce Chemical) and injected into a HP7890A gas chromatograph (Agilent Technology) using anAgilent DB-1 column (30 m, 0.320 mm, 0.25 �m particle
size). Amounts of aucubin and catalpol were quantiWedusing ChemStation B-03-01 software and they are pre-sented as percent dry weight for comparative purposes(e.g., Fuchs and Bowers 2004; Barton 2007; Wurst and vander Putten 2007).
Statistical analyses
Developmental variation in root and shoot biomass, R:Sratios and percentage of water and nitrogen in abovegroundtissues were analyzed using one-way ANOVAs, followedby Bonferroni post hoc tests to distinguish mean diVerencesamong age classes if overall signiWcant diVerences werefound. Variation in production of IGs across plant ontogenywas Wrst assessed simultaneously for both plant tissues,shoots and roots, and both iridoids, aucubin and catalpol.Because aucubin and catalpol are correlated and their con-centrations are also potentially correlated between shootand root tissues, we used multivariate analysis of variance(MANOVA) to examine variation in these four variablesdue to plant age. When signiWcant eVects were detected byMANOVA, we followed up with univariate ANOVAs foreach of the four variables. Bonferroni post hoc tests wereused to assess signiWcant diVerences among the seven ageclasses. Lastly, we did a repeated measures ANOVA inorder to test overall variation in total IGs (aucubin + catal-pol) among age classes, between plant tissues (shoots vs.roots; the repeated measure), and their interaction. Water,nitrogen, and IG concentrations as well as R:S ratios werearcsine square-root transformed to meet assumptions ofnormality. Biomass measures did not require transforma-tion.
To assess whether investment in chemical defenses atthe whole plant level as well as for below- and above-ground tissues showed a linear or non-linear increase withplant age, we regressed the concentration of total IGs(aucubin + catalpol) at the whole plant level (shoots +roots), as well as at the shoot and root tissue level sepa-rately, on plant age and tested whether the linear and qua-dratic slopes associated with age diVer signiWcantly fromzero. Because age is treated as a continuous variable, forthese analyses we Wrst needed to center this predictor vari-able (Judd et al. 2009). Then, to evaluate whether the shapeof the curve of total IG content as a function of plant agediVers between shoot and root tissues, we Wrst computed anew dependent variable that describes the diVerence inchemical defenses between tissues (i.e. within-subjectdiVerence variable IGdiV = IGs shoots ¡ IGs roots) andregressed it on plant age. When a within-subject diVerenceis regressed on a between-subject predictor, the resultingslopes will be testing for an interaction between the within-subject independent variable (tissue) and the between-sub-ject one (age). Thus, the linear and quadratic term in this
123

Oecologia (2012) 168:471–481 475
case will assess whether the linear or quadratic eVect of ageon plant defenses interacts with plant tissue (i.e. whetherthe linear and quadratic slopes for shoot and root tissuesdiVer from each other) (Judd et al. 2009).
Finally, in order to assess whether R:S ratios can help toexplain total IG production across developmental stages,we performed a linear regression to test the prediction of anegative relationship between R:S ratios and plant invest-ment in defenses as proposed by Boege and Marquis(2005). In addition, in order to test whether the relationshipbetween R:S ratios and IG concentrations shifts with plantage, we performed an analysis of covariance with totalplant IG levels as a function of age using R:S ratios as acovariate and testing for an age by R:S ratio interaction. Forthe Wrst regression we used all seven plant stages (n = 140),then after we conWrmed a signiWcant interaction betweenage and R:S ratios (see “Results”), we performed two sepa-rated linear regressions for younger life stages (age classes3, 5, and 8 week-old, n = 60) and older life stages (age clas-ses 10, 12, 15, and 18 week-old, n = 80).
Results
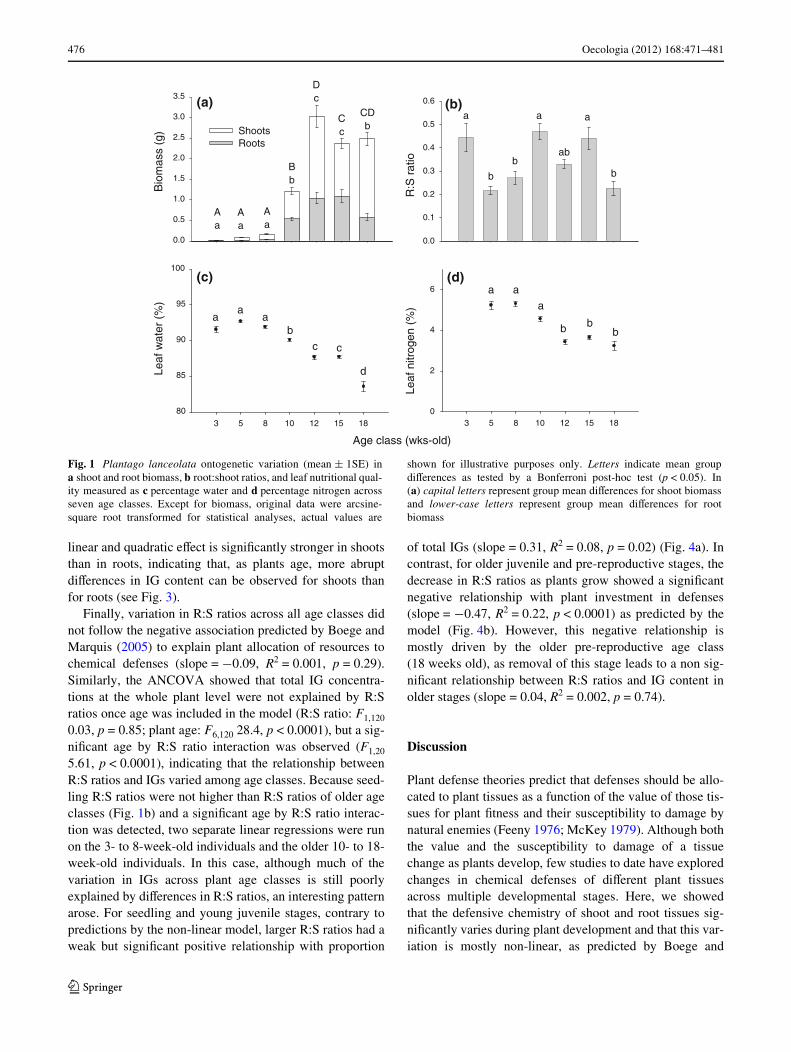
There were pronounced diVerences in plant biomass, nutri-tional quality and chemical defenses among the seven Plan-tago lanceolata age classes. Plant age signiWcantlyinXuenced total plant biomass (F6,132 = 103.4, p < 0.0001)and R:S ratios (F6,132 = 8.9, p < 0.0001) (Fig. 1a, b). Asexpected, overall biomass of both shoot and root tissuesincreased with plant age; however, some exceptions can beobserved such as lower biomass in shoots of the 15-week-old class and in roots of the 18-week-old class than previ-ous age classes (Fig. 1a). Given our experimental design,this unexpected variation is unlikely to be the result ofpotential genetic biases or limited resources altering growthrates. Furthermore, because we did not follow individualplants during a growing season but rather harvested all ageclasses simultaneously from cohorts germinated at diVerenttimes from March to July, these changes in biomass cannotbe interpreted as loss of biomass. Instead, they may haveemerged as a result of diVerential average environmentalconditions (i.e. day length and temperature) shaping devel-opmental trajectories in biomass partitioning, as seen inother systems (McConnaughay and Coleman 1999; Moriu-chi and Winn 2005). Nevertheless, after performing all sta-tistical analyses with and without these age classes, becauseoverall patterns remained the same (except where indi-cated), we present here the results of the complete data set.
Plant age signiWcantly inXuenced shoot nutritional qual-ity measured as percentage of water (F6,133 = 85.5,p < 0.0001) (Fig. 1c), and nitrogen (F5,54 = 34.2,p < 0.0001) (Fig. 1d), both of which declined as plants
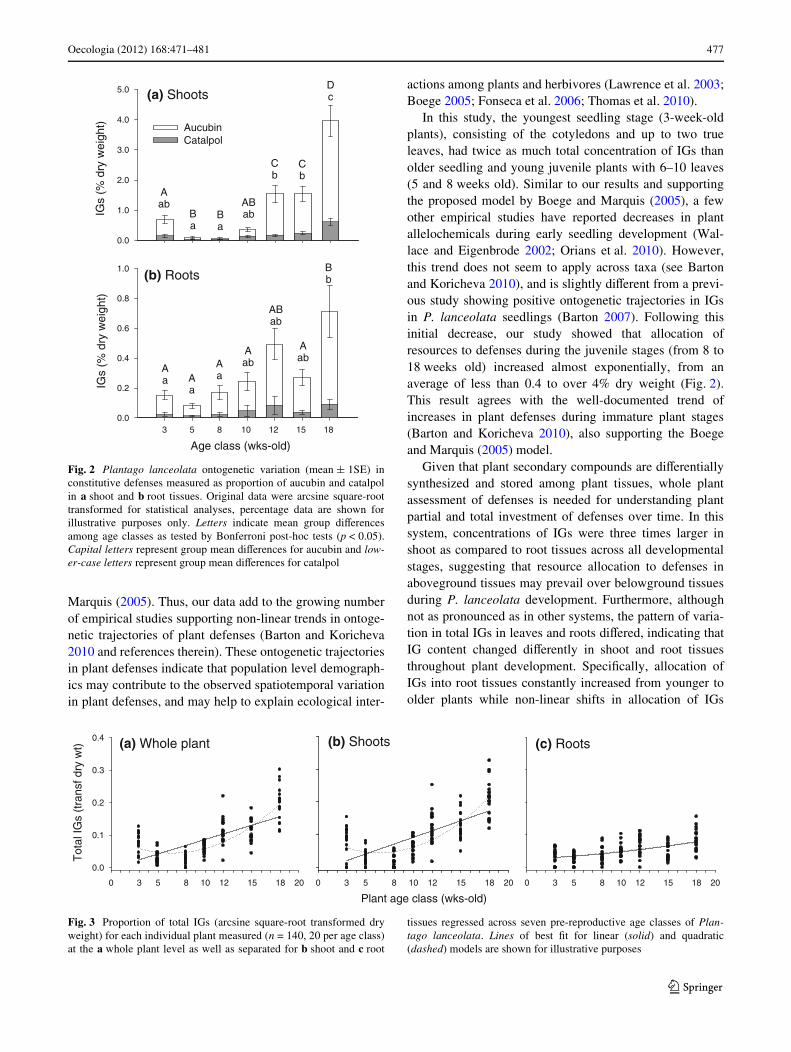
aged. Mean water (Fig. 1c) and nitrogen (Fig. 1d) concen-trations were signiWcantly higher for seedling and youngjuvenile stages (i.e. 3–8 weeks old) than older juvenileand pre-reproductive plants (i.e. 10–18 weeks old). In con-trast, constitutive production of total iridoid glycosides(aucubin + catalpol) increased over time, ranging from aslow as 0.003 to as high as 8.86% dry weight from seedlingto pre-reproductive plants, respectively. Aucubin and catal-pol in shoot and root tissues varied considerably throughoutplant development as shown by the MANOVA (Wilks’� = 0.229, F4,126 = 9.67, p < 0.0001) (Fig. 2). UnivariateANOVAs also demonstrated a signiWcant eVect of plant ageon shoot aucubin and catalpol concentrations (F6,129 =43.31, p < 0.0001 and F6,129 = 11.63, p < 0.0001, respectively)as well as on root aucubin and catalpol concentrations(F6,129 = 16.44, p < 0.0001 and F6,129 = 5.26, p < 0.0001,respectively).
In turn, repeated measures ANOVA demonstrated thattotal IG concentration was signiWcantly inXuenced by plantage (F6,129 = 38.6, p < 0.0001) and plant tissue (F1,129 =104.3, p < 0.0001), with higher IG concentrations in shootcompared to root tissues and overall increase in IGs asplants aged, although in a non-linear fashion (Fig. 2). Fur-thermore, there was a signiWcant interaction between plantage and tissue (F6,129 = 26.02, p < 0.0001), indicating thatdiVerences in IG content between shoot and root tissueschanged throughout plant development. Thus, three maintrends emerged from our data. First, concentrations of IGsin shoot tissues were, on average, three times higher thanwere concentrations in root tissues. Second, mean groupdiVerences in aucubin and catalpol among all seven ageclasses followed similar trends for both shoot (Fig. 2a) androot tissues (Fig. 2b). Third, aucubin concentrations inshoot tissues drive most of the changes in total IGs through-out plant ontogeny.
Regression models demonstrated that constitutive varia-tion in total IGs increased non-linearly with plant age forshoots and linearly for roots, indicating that ontogeneticpatterns in defenses between tissues were dissimilar(Fig. 3). At the whole plant level, ontogenetic trajectories inconstitutive IG production were also non-linear (Table 1;Fig. 3a). This non-linear trend was mostly driven by shootssince partial investment in defenses for shoot tissues alsoshowed a signiWcant linear and quadratic term (Table 1;Fig. 3b), but roots, with signiWcantly less variation acrossage classes than shoots, showed a signiWcant linear increasein IGs with plant age but not a signiWcant quadratic trend(Table 1; Fig. 3c). Furthermore, the shape of the curvesdescribing IG content as plants age diVered between tis-sues, demonstrated by a signiWcant interaction betweenplant tissue and the linear or quadratic eVect of age on plantdefenses (for IGdiV linear t = 7.72, p < 0.0001; IGdiV qua-dratic t = 6.37, p < 0.0001). SpeciWcally, we found that the
123
476 Oecologia (2012) 168:471–481
linear and quadratic eVect is signiWcantly stronger in shootsthan in roots, indicating that, as plants age, more abruptdiVerences in IG content can be observed for shoots thanfor roots (see Fig. 3).
Finally, variation in R:S ratios across all age classes didnot follow the negative association predicted by Boege andMarquis (2005) to explain plant allocation of resources tochemical defenses (slope = ¡0.09, R2 = 0.001, p = 0.29).Similarly, the ANCOVA showed that total IG concentra-tions at the whole plant level were not explained by R:Sratios once age was included in the model (R:S ratio: F1,120
0.03, p = 0.85; plant age: F6,120 28.4, p < 0.0001), but a sig-niWcant age by R:S ratio interaction was observed (F1,20
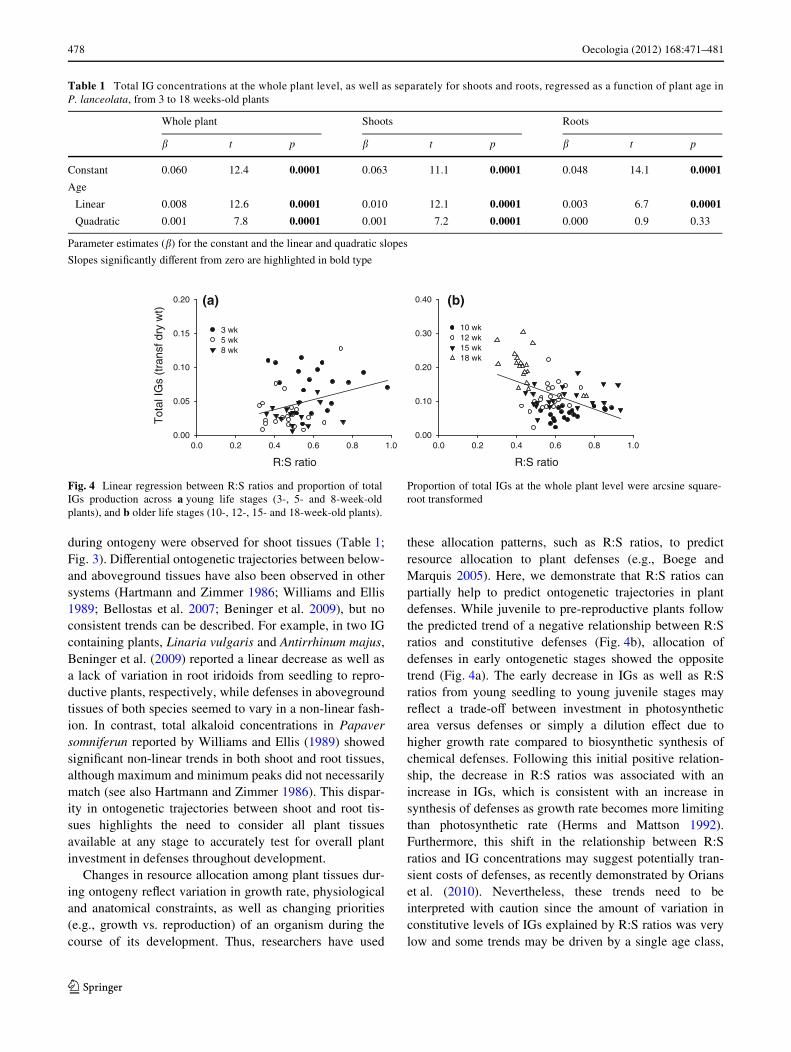
5.61, p < 0.0001), indicating that the relationship betweenR:S ratios and IGs varied among age classes. Because seed-ling R:S ratios were not higher than R:S ratios of older ageclasses (Fig. 1b) and a signiWcant age by R:S ratio interac-tion was detected, two separate linear regressions were runon the 3- to 8-week-old individuals and the older 10- to 18-week-old individuals. In this case, although much of thevariation in IGs across plant age classes is still poorlyexplained by diVerences in R:S ratios, an interesting patternarose. For seedling and young juvenile stages, contrary topredictions by the non-linear model, larger R:S ratios had aweak but signiWcant positive relationship with proportion
of total IGs (slope = 0.31, R2 = 0.08, p = 0.02) (Fig. 4a). Incontrast, for older juvenile and pre-reproductive stages, thedecrease in R:S ratios as plants grow showed a signiWcantnegative relationship with plant investment in defenses(slope = ¡0.47, R2 = 0.22, p < 0.0001) as predicted by themodel (Fig. 4b). However, this negative relationship ismostly driven by the older pre-reproductive age class(18 weeks old), as removal of this stage leads to a non sig-niWcant relationship between R:S ratios and IG content inolder stages (slope = 0.04, R2 = 0.002, p = 0.74).
Discussion
Plant defense theories predict that defenses should be allo-cated to plant tissues as a function of the value of those tis-sues for plant Wtness and their susceptibility to damage bynatural enemies (Feeny 1976; McKey 1979). Although boththe value and the susceptibility to damage of a tissuechange as plants develop, few studies to date have exploredchanges in chemical defenses of diVerent plant tissuesacross multiple developmental stages. Here, we showedthat the defensive chemistry of shoot and root tissues sig-niWcantly varies during plant development and that this var-iation is mostly non-linear, as predicted by Boege and
Fig. 1 Plantago lanceolata ontogenetic variation (mean § 1SE) ina shoot and root biomass, b root:shoot ratios, and leaf nutritional qual-ity measured as c percentage water and d percentage nitrogen acrossseven age classes. Except for biomass, original data were arcsine-square root transformed for statistical analyses, actual values are
shown for illustrative purposes only. Letters indicate mean groupdiVerences as tested by a Bonferroni post-hoc test (p < 0.05). In(a) capital letters represent group mean diVerences for shoot biomassand lower-case letters represent group mean diVerences for rootbiomass
Leaf
wat
er (
%)
80
85
90
95
100
aa
ab
c c
d
(c)
Age class (wks-old)
3 5 8 10 12 15 18 3 5 8 10 12 15 18Le
af n
itrog
en (
%)
0
2
4
6(d)
a a
a
b bb
Bio
mas
s (g
)
0.0
0.5
1.0
1.5
2.0
2.5
3.0
3.5
ShootsRoots
R:S
rat
io
0.0
0.1
0.2
0.3
0.4
0.5
0.6
a
b
b
a
ab
a
b
(a) (b)
Aa
Aa
Aa
Bb
Dc
Cc
CDb
123
Oecologia (2012) 168:471–481 477
Marquis (2005). Thus, our data add to the growing numberof empirical studies supporting non-linear trends in ontoge-netic trajectories of plant defenses (Barton and Koricheva2010 and references therein). These ontogenetic trajectoriesin plant defenses indicate that population level demograph-ics may contribute to the observed spatiotemporal variationin plant defenses, and may help to explain ecological inter-
actions among plants and herbivores (Lawrence et al. 2003;Boege 2005; Fonseca et al. 2006; Thomas et al. 2010).
In this study, the youngest seedling stage (3-week-oldplants), consisting of the cotyledons and up to two trueleaves, had twice as much total concentration of IGs thanolder seedling and young juvenile plants with 6–10 leaves(5 and 8 weeks old). Similar to our results and supportingthe proposed model by Boege and Marquis (2005), a fewother empirical studies have reported decreases in plantallelochemicals during early seedling development (Wal-lace and Eigenbrode 2002; Orians et al. 2010). However,this trend does not seem to apply across taxa (see Bartonand Koricheva 2010), and is slightly diVerent from a previ-ous study showing positive ontogenetic trajectories in IGsin P. lanceolata seedlings (Barton 2007). Following thisinitial decrease, our study showed that allocation ofresources to defenses during the juvenile stages (from 8 to18 weeks old) increased almost exponentially, from anaverage of less than 0.4 to over 4% dry weight (Fig. 2).This result agrees with the well-documented trend ofincreases in plant defenses during immature plant stages(Barton and Koricheva 2010), also supporting the Boegeand Marquis (2005) model.
Given that plant secondary compounds are diVerentiallysynthesized and stored among plant tissues, whole plantassessment of defenses is needed for understanding plantpartial and total investment of defenses over time. In thissystem, concentrations of IGs were three times larger inshoot as compared to root tissues across all developmentalstages, suggesting that resource allocation to defenses inaboveground tissues may prevail over belowground tissuesduring P. lanceolata development. Furthermore, althoughnot as pronounced as in other systems, the pattern of varia-tion in total IGs in leaves and roots diVered, indicating thatIG content changed diVerently in shoot and root tissuesthroughout plant development. SpeciWcally, allocation ofIGs into root tissues constantly increased from younger toolder plants while non-linear shifts in allocation of IGs
Fig. 2 Plantago lanceolata ontogenetic variation (mean § 1SE) inconstitutive defenses measured as proportion of aucubin and catalpolin a shoot and b root tissues. Original data were arcsine square-roottransformed for statistical analyses, percentage data are shown forillustrative purposes only. Letters indicate mean group diVerencesamong age classes as tested by Bonferroni post-hoc tests (p < 0.05).Capital letters represent group mean diVerences for aucubin and low-er-case letters represent group mean diVerences for catalpol
3 5 8 10 12 15 180.0
0.2
0.4
0.6
0.8
1.0
IGs
(% d
ry w
eigh
t)
0.0
1.0
2.0
3.0
4.0
5.0
AucubinCatalpol
(a) Shoots
(b) Roots
Aab
Ba
Ba
ABab
Cb
Cb
Dc
Bb
Aa A
a
Aa
Aab
ABab
Aab
Age class (wks-old)
IGs
(% d
ry w
eigh
t)
Fig. 3 Proportion of total IGs (arcsine square-root transformed dryweight) for each individual plant measured (n = 140, 20 per age class)at the a whole plant level as well as separated for b shoot and c root
tissues regressed across seven pre-reproductive age classes of Plan-tago lanceolata. Lines of best Wt for linear (solid) and quadratic(dashed) models are shown for illustrative purposes
0 3 5 8 10 12 15 18 20
0.0
0.1
0.2
0.3
0.4 (a) Whole plant
Plant age class (wks-old)
0 3 5 8 10 12 15 18 20
(c) Roots
0 3 5 8 10 12 15 18 20
Tot
al IG
s (t
rans
f dry
wt) (b) Shoots
123
478 Oecologia (2012) 168:471–481
during ontogeny were observed for shoot tissues (Table 1;Fig. 3). DiVerential ontogenetic trajectories between below-and aboveground tissues have also been observed in othersystems (Hartmann and Zimmer 1986; Williams and Ellis1989; Bellostas et al. 2007; Beninger et al. 2009), but noconsistent trends can be described. For example, in two IGcontaining plants, Linaria vulgaris and Antirrhinum majus,Beninger et al. (2009) reported a linear decrease as well asa lack of variation in root iridoids from seedling to repro-ductive plants, respectively, while defenses in abovegroundtissues of both species seemed to vary in a non-linear fash-ion. In contrast, total alkaloid concentrations in Papaversomniferun reported by Williams and Ellis (1989) showedsigniWcant non-linear trends in both shoot and root tissues,although maximum and minimum peaks did not necessarilymatch (see also Hartmann and Zimmer 1986). This dispar-ity in ontogenetic trajectories between shoot and root tis-sues highlights the need to consider all plant tissuesavailable at any stage to accurately test for overall plantinvestment in defenses throughout development.
Changes in resource allocation among plant tissues dur-ing ontogeny reXect variation in growth rate, physiologicaland anatomical constraints, as well as changing priorities(e.g., growth vs. reproduction) of an organism during thecourse of its development. Thus, researchers have used
these allocation patterns, such as R:S ratios, to predictresource allocation to plant defenses (e.g., Boege andMarquis 2005). Here, we demonstrate that R:S ratios canpartially help to predict ontogenetic trajectories in plantdefenses. While juvenile to pre-reproductive plants followthe predicted trend of a negative relationship between R:Sratios and constitutive defenses (Fig. 4b), allocation ofdefenses in early ontogenetic stages showed the oppositetrend (Fig. 4a). The early decrease in IGs as well as R:Sratios from young seedling to young juvenile stages mayreXect a trade-oV between investment in photosyntheticarea versus defenses or simply a dilution eVect due tohigher growth rate compared to biosynthetic synthesis ofchemical defenses. Following this initial positive relation-ship, the decrease in R:S ratios was associated with anincrease in IGs, which is consistent with an increase insynthesis of defenses as growth rate becomes more limitingthan photosynthetic rate (Herms and Mattson 1992).Furthermore, this shift in the relationship between R:Sratios and IG concentrations may suggest potentially tran-sient costs of defenses, as recently demonstrated by Orianset al. (2010). Nevertheless, these trends need to beinterpreted with caution since the amount of variation inconstitutive levels of IGs explained by R:S ratios was verylow and some trends may be driven by a single age class,
Table 1 Total IG concentrations at the whole plant level, as well as separately for shoots and roots, regressed as a function of plant age inP. lanceolata, from 3 to 18 weeks-old plants
Parameter estimates (�) for the constant and the linear and quadratic slopes
Slopes signiWcantly diVerent from zero are highlighted in bold type
Whole plant Shoots Roots
� t p � t p � t p
Constant 0.060 12.4 0.0001 0.063 11.1 0.0001 0.048 14.1 0.0001
Age
Linear 0.008 12.6 0.0001 0.010 12.1 0.0001 0.003 6.7 0.0001
Quadratic 0.001 7.8 0.0001 0.001 7.2 0.0001 0.000 0.9 0.33
Fig. 4 Linear regression between R:S ratios and proportion of totalIGs production across a young life stages (3-, 5- and 8-week-oldplants), and b older life stages (10-, 12-, 15- and 18-week-old plants).
Proportion of total IGs at the whole plant level were arcsine square-root transformed
(a)
R:S ratio
0.0 0.2 0.4 0.6 0.8 1.0
Tot
al IG
s (t
rans
f dry
wt)
0.00
0.05
0.10
0.15
0.20
3 wk5 wk8 wk
(b)
R:S ratio
0.0 0.2 0.4 0.6 0.8 1.00.00
0.10
0.20
0.30
0.40
10 wk 12 wk15 wk18 wk
123

Oecologia (2012) 168:471–481 479
indicating that production of IGs depends on several otherfactors besides the allocation of biomass to diVerent tissues.Alternatively, lack of strong correlations between R:Sratios and IG concentrations may be due to partial assess-ment of P. lanceolata overall investment in defenses.Because P. lanceolata also invest in phenolic compounds(Galvez et al. 2005), as well as in physical defenses such asleaf toughness and trichomes (Schipper and OlV 2000; de laFuente 2002), we cannot rule out the potential predictedvalue of R:S ratios in overall defenses.
Whatever the mechanism, the ontogenetic variation inIGs described here, encompassing four- to Wvefoldincreases in shoot and root tissues, can have important con-sequences for plant–herbivore interactions. Laboratory andWeld experiments have shown that specialist butterXies pre-fer to oviposit on P. lanceolata tissues with higher IG con-tent (Prudic et al. 2005; Talsma et al. 2008), and that larvaeof these herbivores grew better on diets with higheramounts of IGs (e.g., Bowers and Puttick 1989; Harveyet al. 2005; Saastamoinen et al. 2007). In contrast, general-ist herbivores and pathogenic fungi damaging below- oraboveground tissues are usually deterred by high IG con-tent (Strohmeyer et al. 1998; Biere et al. 2004; De Deynet al. 2004; Harvey et al. 2005; Wurst and van der Putten2007). As a consequence, the relative palatability ofP. lanceolata to below- and aboveground herbivores andpathogens will signiWcantly vary through the season, andmay determine the impact of these antagonists on plantWtness. Lastly, for sequestering specialist herbivores, onto-genetic trajectories in plant defenses can, in turn, modifyherbivore unpalatability and thus can indirectly mediate tri-trophic interactions (e.g., Strohmeyer et al. 1998; Harveyet al. 2005; Baden and Dobler 2009; Smilanich et al. 2009).Thus, mainly for specialist herbivores that consume andsequester IGs from single or multiple tissues, these ontoge-netic trajectories in plant constitutive defenses may impactherbivore population dynamics and community structurethrough seasonal changes in host selection, herbivore per-formance, and levels of resistance to natural enemies.
Ontogenetic trajectories in leaf nutritional quality werealso evident, but the trend was opposite to the onedescribed for chemical defenses, showing a decrease in leafwater and nitrogen concentration as plants aged. Similardecreases in nutritional quality have been observed innumerous plant taxa (e.g., Gaudet et al. 2001; Bowers andStamp 1993; Goodger et al. 2006; Merilo et al. 2009),directly inXuenced by plant age or indirectly mediatedthrough their correlation with other ontogenetic shifts inleaf anatomy and physiology (Apple et al. 2002; Ishidaet al. 2005). In our study, ontogenetic variation in nutri-tional quality encompassed an average decrease of 10% inpercentage water and 30% in nitrogen concentration from3- to 18-week-old plants (Fig. 1c, d). This variation can
also have important implications for plant-insect interac-tions since diets with low water and nitrogen concentrationstypically have negative impacts on herbivore host selection,growth rates, and Wtness (Mattson 1980; Awmack andLeather 2002). SpeciWcally for this system, similar varia-tion in leaf nitrogen content (»30%) among individuals ofP. lanceolata resulted in a signiWcant eVect on ovipositionpreferences of the specialist Buckeye butterXy (Junoniacoenia, Nymphalidae), with females showing preferencefor high-nitrogen plants (Prudic et al. 2005). Thus, theontogenetic trajectories in nutritional quality showed heremay also help to explain temporal variation in species inter-actions as seen in other systems (e.g., Gaudet et al. 2001;Barrett and Agrawal 2004).
In summary, developmental variation in chemicaldefenses across Plantago lanceolata ontogeny appears tobe similar, and sometimes greater, in magnitude thangenetic or environmental variation in a single age class(Bowers et al. 1992; Bowers and Stamp 1993; Barton2007). Recent empirical studies have shown similar pat-terns in other systems (e.g., Donaldson et al. 2006; Holeskiet al. 2009; Lawrence et al. 2003). Thus, these studies high-light the importance of separating the proportion of varia-tion in plant defenses that has a developmental basis fromstrict genetic or environmentally dependent variation, inorder to improve our understanding of the evolution ofplant defenses. Furthermore, these ontogenetic trajectoriesin plant defenses and nutritional quality are likely to inXu-ence below- and aboveground herbivores alike, providing apotential mechanism to explain temporal variation in herbi-vore community composition and species interactions overtime.
Acknowledgments We thank K. Barton, K. Boege, Y. Linhart, theBowers lab, and three anonymous reviewers for valuable commentsand suggestions on this manuscript. In addition, we gratefullyacknowledge A. Hill, L. Mulder, and T. J. Lemieux for greenhouse andlaboratory assistance; and N. Robinson and C. M. Judd for their assis-tance with data analyses. Funding for this project was provided byBeverly Sears Graduate Student Grant, the Undergraduate ResearchOpportunity Program and the Department of Ecology and Evolution-ary Biology at the University of Colorado, and a National ScienceFoundation grant DEB 0909717.
References
Apple M, Tiekotter K, Snow M, Young J, Soeldner A, Phillips D,Tingey D, Bond BJ (2002) Needle anatomy changes with increas-ing tree age in Douglas-Wr. Tree Physiol 22:129–136
Awmack CS, Leather SR (2002) Host plant quality and fecundity inherbivorous insects. Annu Rev Entomol 47:817–844
Baden CU, Dobler S (2009) Potential beneWts of iridoid glycosidesequestration in Longitarsus melanocephalus (Coleoptera,Chrysomelidae). Basic Appl Ecol 10:27–33
Baldwin IT (1988) Damage-induced alkaloids in tobacco: pot-boundplants are not inducible. J Chem Ecol 14:1113–1120
123

480 Oecologia (2012) 168:471–481
Barrett RDH, Agrawal AA (2004) Interactive eVects of genotype, envi-ronment, and ontogeny on resistance of cucumber (Cucumis sati-vus) to the generalist herbivore, Spodoptera exigua. J Chem Ecol30:37–51
Barton KE (2007) Early ontogenetic patterns in chemical defense inPlantago (Plantaginaceae): genetic variation and trade-oVs. Am JBot 94:56–66
Barton KE, Koricheva J (2010) The ontogeny of plant defense and her-bivory: characterizing general patterns using meta-analysis. AmNat 175:481–493
Bellostas N, Sorensen JC, Sorensen H (2007) ProWling glucosinolatesin vegetative and reproductive tissues of four Brassica species ofthe U-triangle for their biofumigation potential. J Sci Food Agric87:1587–1594
Beninger CW, Cloutier RR, Monteiro MA, Grodzinski B (2007) Thedistribution of two major iridoids in diVerent organs of Antirrhi-num majus L. at selected stages of development. J Chem Ecol33:731–747
Beninger CW, Cloutier RR, Grodzinski B (2009) A comparison of an-tirrhinoside distribution in the organs of two related Plantagina-ceae species with diVerent reproductive strategies. J Chem Ecol35:1363–1372
Biere A, Marak HB, van Damme JMM (2004) Plant chemical defenseagainst herbivores and pathogens: generalized defense or trade-oVs? Oecologia 140:430–441
Boege K (2005) Herbivore attack in Casearia nitida inXuenced byplant ontogenetic variation in foliage quality and plant architec-ture. Oecologia 143:117–125
Boege K, Marquis RJ (2005) Facing herbivory as you grow up: theontogeny of resistance in plants. Trends Ecol Evol 20:441–448
Boege K, Dirzo R, Siemens D, Brown P (2007) Ontogenetic switchesfrom plant resistance to tolerance: minimizing costs with age?Ecol Lett 10:177–187
Bowers MD, Puttick GM (1989) Iridoid glycosides and insect feedingpreferences—Gypsy moths (Lymantria dispar, Lymantriidae)and Buckeyes (Junonia coenia, Nymphalidae). Ecol Entomol14:247–256
Bowers MD, Stamp NE (1992) Chemical variation within and betweenindividuals of Plantago lanceolata (Plantaginaceae). J ChemEcol 18:985–995
Bowers MD, Stamp NE (1993) EVects of plant age, genotype, and her-bivory on Plantago performance and chemistry. Ecology74:1778–1791
Bowers MD, Collinge SK, Gamble SE, Schmitt J (1992) EVects ofgenotype, habitat, and seasonal variation on iridoid glycosidecontent of Plantago lanceolata (Plantaginaceae) and the implica-tions for insect herbivores. Oecologia 91:201–207
Bryant JP, Kuropat PJ, Reichardt PB, Clausen TP (1991) Controls overthe allocation of resources by woody plants to chemical anti-her-bivore defense. In: Palo RT, Robbins CT (eds) Plant defensesagainst mammalian herbivory. CRC Press, Boca Raton, pp 83–103
Bryant JP, Reichardt PB, Clausen TP, Provenze FD, Kuropat PJ (1992)Woody plant-mammal interactions. In: Rosenthal GA, Beren-baum MR (eds) Herbivores: their interactions with plant second-ary metabolites. Academic, San Diego, pp 344–371
Cavers PB, Bassett IJ, Crompton CW (1980) The biology of Canadianweeds. 47. Plantago lanceolata L. Can J Plant Sci 60:1269–1282
Croteau R (1987) Biosynthesis and catabolism of monoterpenoids.Chem Rev 87:929–954
De Deyn GB, Raaijmakers CE, van Ruijven J, Berendse F, van der Put-ten WH (2004) Plant species identity and diversity eVects ondiVerent trophic levels of nematodes in the soil food web. Oikos106:576–586
De Deyn GB, Biere A, van der Putten WH, Wagenaar R, KlironomosJN (2009) Chemical defense, mycorrhizal colonization and
growth responses in Plantago lanceolata L. Oecologia 160:433–442
de la Fuente MAS (2002) Variation in plant antiherbivore defenses:causes and consequences. PhD dissertation, University of Colo-rado, Boulder
Donaldson JR, Stevens MT, Barnhill HR, Lindroth RL (2006) Age-re-lated shifts in leaf chemistry of clonal aspen (Populus tremulo-ides). J Chem Ecol 32:1415–1429
Feeny PP (1976) Plant apparency and chemical defence. Recent AdvPhytochem 10:1–40
Fonseca CR, Fleck T, Fernandes GW (2006) Processes driving ontoge-netic succession of galls in a canopy. Biotropica 38:514–521
Fuchs A, Bowers MD (2004) Patterns of iridoid glycoside productionand induction in Plantago lanceolata and the importance of plantage. J Chem Ecol 30:1723–1741
Galvez M, Martin-Cordero C, Houghton PJ, Ayuso MJ (2005) Antiox-idant activity of methanol extracts obtained from Plantago spe-cies. J Agric Food Chem 53:1927–1933
Gaudet DA, Laroche A, Puchalski B (2001) EVect of plant age on wa-ter content in crowns of fall rye and winter wheat cultivars diVer-ing in snow mold resistance. Can J Plant Sci 81:541–550
Goodger JQD, Gleadow RM, Woodrow IE (2006) Growth cost andontogenetic expression patterns of defence in cyanogenic Euca-lyptus spp. Trees 20:757–765
Gowan E, Lewis BA, Turgeon R (1995) Phloem transport of antirrhi-noside, an iridoid glycoside, in Asarina scandens (Scrophularea-ceae). J Chem Ecol 21:1781–1788
Hanley ME, Fegan EL (2007) Timing of cotyledon damage aVectsgrowth and Xowering in mature plants. Plant Cell Environ30:812–819
Hanley ME, Lamont BB, Fairbanks MM, RaVerty CM (2007) Plantstructural traits and their role in anti-herbivore defence. PerspectPlant Ecol Evol Syst 8:157–178
Hartmann T, Zimmer M (1986) Organ-speciWc distribution and accu-mulation of pyrrolizidine alkaloids during the life history of twoannual Senecio species. J Plant Physiol 122:67–80
Harvey JA, Van Nouhuys S, Biere A (2005) EVects of quantitative var-iation in allelochemicals in Plantago lanceolata on developmentof a generalist and a specialist herbivore and their endoparasi-toids. J Chem Ecol 31:287–302
Herms DA, Mattson WJ (1992) The dilemma of plants—to grow or de-fend. Q Rev Biol 67:283–335
Hogedal BD, Molgaard P (2000) HPLC analysis of the seasonal anddiurnal variation of iridoids in cultivars of Antirrhinum majus.Biochem Syst Ecol 28:949–962
Holeski LM, Kearsley MJC, Whitham TG (2009) Separating ontoge-netic and environmental determination of resistance to herbivoryin cottonwood. Ecology 90:2969–2973
Ishida A, Yakaki K, Lai Hoe A (2005) Ontogenetic transition of leafphysiology and anatomy from seedlings to mature trees of a rainforest pioneer tree, Macaranga gigantean. Tree Physiol 25:513–522
Jensen SR (1991) Plant iridoids, their biosynthesis and distribution inangiosperms. In: Harborne JB, Tomas-Barberan FA (eds) Annualproceedings of the phytochemical society of Europe: ecologicalchemistry and biochemistry of plant terpenoids. Oxford Univer-sity Press, Oxford, pp 133–158
Judd CM, McClelland GH, Ryan CS (2009) Data analysis: a modelcomparison approach, 2nd edn. Routledge Taylor and Francis,New York
Lawrence R, Potts BM, Whitham TG (2003) Relative importance ofplant ontogeny, host genetic variation, and leaf age for a commonherbivore. Ecology 84:1171–1178
Loney PE, McArthurt C, Potts BM, Jordan GJ (2006) How does ontog-eny in a Eucalyptus species aVect patterns of herbivory by Brush-tail Possums? Funct Ecol 20:982–988
123

Oecologia (2012) 168:471–481 481
Mattson WJ (1980) Herbivory in relation to plant nitrogen-content.Annu Rev Ecol Syst 11:119–161
McConnaughay KDM, Coleman JS (1999) Biomass allocation inplants: ontogeny or optimality? A test along three resource gradi-ents. Ecology 80:2581–2593
McKey D (1979) The distribution of secondary compounds withinplants. In: Rosenthal GA, Janzen DH (eds) Herbivores: theirinteraction with secondary plant metabolites. Academic, London,pp 55–133
Merilo E, Tulva I, Raim O, Kukit A, Sellin A, Kull O (2009) Changesin needle nitrogen partitioning and photosynthesis during80 years of tree ontogeny in Picea abies. Trees 23:951–958
Moriuchi KS, Winn AA (2005) Relationships among growth, develop-ment and plastic response to environment quality in a perennialplant. New Phytol 166:149–158
Opitz SEW, Jensen SR, Muller C (2010) Sequestration of glucosin-olates and iridoid glucosides in sawXy species of the genusAthalia and their role in defense against ants. J Chem Ecol36:148–157
Orians CM, Hochwender CG, Fritz RS, Snall T (2010) Growth andchemical defense in willow seedlings: trade-oVs are transient.Oecologia 163:283–290
Pires CSS, Price PW (2000) Patterns of host plant growth and attackand establishment of gall-inducing wasp (Hymenoptera: Cynipi-dae). Environ Entomol 29:49–54
Prudic KL, Oliver JC, Bowers MD (2005) Soil nutrient eVects on ovi-position preference, larval performance, and chemical defense ofa specialist insect herbivore. Oecologia 143:578–587
Ronsted N, Gobel E, Franzyk H, Jensen SR, Olsen CE (2000) Chemo-taxonomy of Plantago. Iridoid glucosides and caVeoyl phenyleth-anoid glycosides. Phytochemistry 55:337–348
Saastamoinen M, van Nouhuys S, Nieminen M, O’Hara B, Suomi J(2007) Development and survival of a specialist herbivore, Meli-taea cinxia, on host plants producing high and low concentrationsof iridoid glycosides. Ann Zool Fenn 44:70–80
Schipper P, OlV H (2000) Biomass partitioning, architecture and turn-over of six herbaceous species from habitats with diVerent nutri-ent supply. Plant Ecol 149:219–231
Smilanich MA, Dyer LA, Chambers JQ, Bowers MD (2009) Immuno-logical cost of chemical defence and the evolution of herbivorediet breadth. Ecol Lett 12:612–621
Strohmeyer HH, Stamp NE, Jarzomski CM, Bowers MD (1998) Preyspecies and prey diet aVect growth of invertebrate predators. EcolEntomol 23:68–79
Talsma JHR, Torri K, van Nouhuys S (2008) Host plant use by theHeath fritillary butterXy, Melitaea athalia: plant habitat, speciesand chemistry. Arthropod-Plant Interact 2:63–75
Tamura Y (2001) EVects of temperature, shade, and nitrogen applica-tion on the growth and accumulation of bioactive compounds incultivars of Plantago lanceolata L. Jpn J Crop Sci 70:548–553
Thomas SC, Sztaba AJ, Smith SM (2010) Herbivory patterns in maturesugar maple: variation with vertical canopy strata and tree ontog-eny. Ecol Entomol 35:1–8
Wallace SK, Eigenbrode SD (2002) Changes in the glucosinolate–myrosinase defense system in Brassica juncea cotyledons duringseedling development. J Chem Ecol 28:243–256
Waltz AM, Whitham TG (1997) Plant development aVects arthropodcommunities: opposing impacts of species removal. Ecology78:2133–2144
Warner PJ, Cushman JH (2002) InXuence of herbivores on a perennialplant: variation with life history stage and herbivore species. Oec-ologia 132:77–85
Williams RD, Ellis BE (1989) Age and tissue distribution of alkaloidsin Papaver somniferum. Phytochemistry 28:2085–2088
Willinger G, Dobler S (2001) Selective sequestration of iridoid glyco-sides from their host plants in Longitarsus Xea beetles. BiochemSyst Ecol 29:335–346
Wurst S, van der Putten WH (2007) Root herbivore identity matters inplant-mediated interactions between root and shoot herbivores.Basic Appl Ecol 8:491–499
123




















