
Interaction between oil-collecting bees and seven species of Plantaginaceae
11
Flora 208 (2013) 401–411 Contents lists available at ScienceDirect Flora jou rn al hom epage: www.elsevier.com/locate/flora Interaction between oil-collecting bees and seven species of Plantaginaceae Aline C. Martins a,∗ , Antonio J.C. Aguiar b , Isabel Alves-dos-Santos c a Pós-graduac ¸ ão em Entomologia, Faculdade de Filosofia, Ciências e Letras de Ribeirão Preto, Ribeirão Preto, SP, Brazil b Departamento de Zoologia, Instituto de Ciências Biológicas, Universidade de Brasilia, Brasília, DF, Brazil c Departamento de Ecologia, Instituto de Biociências, Universidade de São Paulo, São Paulo, SP, Brazil a r t i c l e i n f o Article history: Received 28 January 2013 Accepted 24 June 2013 Available online 1 August 2013 Keywords: Angelonia Mutualism Neotropical Pollination Apidae Oil-collecting bees a b s t r a c t Oil-bee/oil-flower mutualism evolved through multiple gains and losses of the ability to produce floral oil in plants and to collect it in bees. Around 2000 plant species are known to produce floral oils that are collected by roughly 450 bee species, which use them for the construction of nests and for the larval food. The Plantaginaceae contain several Neotropical species that produce floral oils, the main reward offered by these plants. In the genera Angelonia, Basistemon, Monopera and Monttea, mainly associated with Cen- tris bees, the floral oil is produced in trichomes that are located in the inner corolla. The pollinators of a few species in this neotropical clade of Plantaginaceae are known, and the role of flower morphology as well as the requirements from pollinators and the role of other groups of bees in the pollination of these flowers remains unclear. In this paper we provide a list of the flower visitors of seven Plantaginaceae species (six Angelonia species and Basistemon silvaticus) analyzing their behavior to highlight the legiti- mate pollinators and illustrating little known aspects of flower morphology and oil-collecting apparatuses of the bees. Two general morphological patterns were observed in the Angelonia flowers: deep corolla tube with short lobes, and short corolla tube with long lobes. Corolla tubes of different length result in pollen adherence to different parts of the insect body. The six Angelonia species and B. silvaticus flowers were visited by 25 oil-collecting bee species (10 Centris, 11 Tapinotaspidini and 4 Tetrapedia species), the majority acting as legitimate visitors. The flowers were also visited by illegitimate bee pollinators, which collected pollen but do not transfer it to the female organ. Specialized collectors of Plantaginaceae floral oils present modifications on the first pair of legs, mainly in the basitarsi but also extended to the tarsomeres. The new records of Tapinotaspidini and Centridini species acting as specialized pollinators of Plantaginaceae suggest that there is a geographic variation in the pollinators of the same plant species, and that the evolutionary scenario of the historical relationships between oil-collecting bees and floral oil producing plants is more complex than previously considered. © 2013 Elsevier GmbH. All rights reserved. Introduction The mutualism of oil-collecting bees and floral-oil producing plants, discovered 40 years ago by Stefan Vogel, evolved through a set of morphological, behavioral and chemical adaptations (Vogel, 1969, 1974). Floral oil production and oil-collecting behavior has been gained and lost many times in the evolutionary history of bees and plants (Renner and Schaefer, 2010). This resource is produced in glands called ‘elaiophores’ in approximately 2000 species, spread over 11 families of angiosperms, and are collected by 447 species ∗ Corresponding author. Current address: Univ. Fed. Paraná, Dept. Zool., Lab. Biol. Comparada Hymenoptera, Caixa Postal 19020, BR-81531980, Curitiba, Paraná, Brazil. Tel.: +55 4133611589. E-mail address: aline [email protected] (A.C. Martins). of apid bees (Rasmussen and Olesen, 2000; Simpson et al., 1977; Simpson and Neff, 1981; Alves-dos-Santos et al., 2007; Buchmann, 1987; Cocucci et al., 2000; Machado, 2004; Neff and Simpson, 1981; Renner and Schaefer, 2010). Oil-collecting behavior in bees is a characteristic of species in the genera Centris (230 species), Epicharis (31), Tetrapedia (28), Ctenoplectra (19), Macropis (16), Redi- viva (24) and 12 genera (95 species) within Tapinotaspidini (Moure et al., 2007; Renner and Schaefer, 2010, and references therein). Solitary females oil-bees use floral oils to surface the nest and cell walls, and also to feed larvae, mixed with pollen (Alves-dos- Santos et al., 2007; Buchmann, 1987; Cane et al., 1983; Schäffler and Dötterl, 2011), while the males (Tetrapediini and Tapintoaspi- dini tribes) collect them for still unknown purposes (Aguiar and Melo, 2009; Cappellari et al., 2011; Vogel and Machado, 1991). Females usually present more conspicuous organs for oil collection compared to males (Cocucci et al., 2000; Neff and Simpson, 1981). 0367-2530/$ – see front matter © 2013 Elsevier GmbH. All rights reserved. http://dx.doi.org/10.1016/j.flora.2013.07.001
Transcript of Interaction between oil-collecting bees and seven species of Plantaginaceae

IP
Aa
b
c
a
ARAA
KAMNPAO
I
ps1baio
BB
0h
Flora 208 (2013) 401– 411
Contents lists available at ScienceDirect
Flora
jou rn al hom epage: www.elsev ier .com/ locate / f lora
nteraction between oil-collecting bees and seven species oflantaginaceae
line C. Martinsa,∗, Antonio J.C. Aguiarb, Isabel Alves-dos-Santosc
Pós-graduac ão em Entomologia, Faculdade de Filosofia, Ciências e Letras de Ribeirão Preto, Ribeirão Preto, SP, BrazilDepartamento de Zoologia, Instituto de Ciências Biológicas, Universidade de Brasilia, Brasília, DF, BrazilDepartamento de Ecologia, Instituto de Biociências, Universidade de São Paulo, São Paulo, SP, Brazil
r t i c l e i n f o
rticle history:eceived 28 January 2013ccepted 24 June 2013vailable online 1 August 2013
eywords:ngeloniautualismeotropicalollinationpidaeil-collecting bees
a b s t r a c t
Oil-bee/oil-flower mutualism evolved through multiple gains and losses of the ability to produce floraloil in plants and to collect it in bees. Around 2000 plant species are known to produce floral oils that arecollected by roughly 450 bee species, which use them for the construction of nests and for the larval food.The Plantaginaceae contain several Neotropical species that produce floral oils, the main reward offeredby these plants. In the genera Angelonia, Basistemon, Monopera and Monttea, mainly associated with Cen-tris bees, the floral oil is produced in trichomes that are located in the inner corolla. The pollinators of afew species in this neotropical clade of Plantaginaceae are known, and the role of flower morphology aswell as the requirements from pollinators and the role of other groups of bees in the pollination of theseflowers remains unclear. In this paper we provide a list of the flower visitors of seven Plantaginaceaespecies (six Angelonia species and Basistemon silvaticus) analyzing their behavior to highlight the legiti-mate pollinators and illustrating little known aspects of flower morphology and oil-collecting apparatusesof the bees. Two general morphological patterns were observed in the Angelonia flowers: deep corollatube with short lobes, and short corolla tube with long lobes. Corolla tubes of different length result inpollen adherence to different parts of the insect body. The six Angelonia species and B. silvaticus flowerswere visited by 25 oil-collecting bee species (10 Centris, 11 Tapinotaspidini and 4 Tetrapedia species),the majority acting as legitimate visitors. The flowers were also visited by illegitimate bee pollinators,
which collected pollen but do not transfer it to the female organ. Specialized collectors of Plantaginaceaefloral oils present modifications on the first pair of legs, mainly in the basitarsi but also extended to thetarsomeres. The new records of Tapinotaspidini and Centridini species acting as specialized pollinatorsof Plantaginaceae suggest that there is a geographic variation in the pollinators of the same plant species,and that the evolutionary scenario of the historical relationships between oil-collecting bees and floraloil producing plants is more complex than previously considered.ntroduction
The mutualism of oil-collecting bees and floral-oil producinglants, discovered 40 years ago by Stefan Vogel, evolved through aet of morphological, behavioral and chemical adaptations (Vogel,969, 1974). Floral oil production and oil-collecting behavior haseen gained and lost many times in the evolutionary history of bees
nd plants (Renner and Schaefer, 2010). This resource is producedn glands called ‘elaiophores’ in approximately 2000 species, spreadver 11 families of angiosperms, and are collected by 447 species∗ Corresponding author. Current address: Univ. Fed. Paraná, Dept. Zool., Lab.iol. Comparada Hymenoptera, Caixa Postal 19020, BR-81531980, Curitiba, Paraná,razil. Tel.: +55 4133611589.
E-mail address: aline [email protected] (A.C. Martins).
367-2530/$ – see front matter © 2013 Elsevier GmbH. All rights reserved.ttp://dx.doi.org/10.1016/j.flora.2013.07.001
© 2013 Elsevier GmbH. All rights reserved.
of apid bees (Rasmussen and Olesen, 2000; Simpson et al., 1977;Simpson and Neff, 1981; Alves-dos-Santos et al., 2007; Buchmann,1987; Cocucci et al., 2000; Machado, 2004; Neff and Simpson,1981; Renner and Schaefer, 2010). Oil-collecting behavior in beesis a characteristic of species in the genera Centris (230 species),Epicharis (31), Tetrapedia (28), Ctenoplectra (19), Macropis (16), Redi-viva (24) and 12 genera (95 species) within Tapinotaspidini (Moureet al., 2007; Renner and Schaefer, 2010, and references therein).Solitary females oil-bees use floral oils to surface the nest andcell walls, and also to feed larvae, mixed with pollen (Alves-dos-Santos et al., 2007; Buchmann, 1987; Cane et al., 1983; Schäfflerand Dötterl, 2011), while the males (Tetrapediini and Tapintoaspi-
dini tribes) collect them for still unknown purposes (Aguiar andMelo, 2009; Cappellari et al., 2011; Vogel and Machado, 1991).Females usually present more conspicuous organs for oil collectioncompared to males (Cocucci et al., 2000; Neff and Simpson, 1981).
4 lora 2
i(g(wrt(t(MC
o3(sAan2i(imvauAwatsdaao
TTa
N
02 A.C. Martins et al. / F
In Plantaginaceae, floral-oil production emerged only oncen the Neotropical clade, previously placed in ScrophulariaceaeAlbach et al., 2005; Olmstead et al., 2001), which comprises theenera Angelonia, Basistemon, Monttea, Monopera and MelospermaRenner and Schaefer, 2010). In this clade the floral oil productionas also lost at least twice: in Melosperma, which offers nectar as
eward (Sérsic and Cocucci, 1996), and in some species of Basis-emon, in which the pollinators’ rewards are probably only pollenRenner and Schaefer, 2010). Floral oil production in species of Plan-aginaceae is known due to some studies of Angelonia and MontteaMachado et al., 2002; Simpson et al., 1990; Vogel, 1974; Vogel and
achado, 1991), and scarce information on Basistemon (Vogel andocucci, 1995) and Monopera (Aguiar and Melo, 2009).
The present study focused on two Neotropical genera of floral-il producing Plantaginaceae: Angelonia, which contains about0 described species, and occurs from Mexico to southern BrazilBarringer, 1981); and Basistemon, which contains seven describedpecies, and occurs in Chaco and Andean vegetation areas in Southmerica (Barringer, 1985). Angelonia flowers are highly specializednd are pollinated mainly by Centris bees, but also by Tapinotaspisordestina and Caenonomada aff. unicalcarata (Aguiar and Melo,009; Machado et al., 2002; Vogel and Machado, 1991), while there
s only one known pollinator for Basistemon flowers, Arhysoceble sp.Tapinotaspidini) (Vogel and Cocucci, 1995). The aim of this studys to compare the pollination strategies of Angelonia and Basiste-
on species, and record their flower visitors, the behavior of theisitors during visits, and the morphology of the flowers and theirssociated pollinators. Among the seven studied species, there isntil now no record of the pollinators or reproductive aspects of. eryostachis, A. goyazensis and B. silvaticus, and some visitors onlyere recorded in A. integerrima flowers (Vogel, 1974; Gonc alves
nd Melo, 2005; Pinheiro et al., 2008). But no extensive study ofhe reproductive systems or mutualistic interaction between thesepecies has been undertaken before. New cases of interaction were
iscovered and are presented here, with a focus on the efficiencynd interdependence of the partners and reciprocal morphologicaldaptations, as well as the visiting behavior of oil-collecting beesn Plantaginaceae flowers.able 1he species of Angelonia and Basistemon (Plantaginaceae) studied, listing their main traits,nd the fieldwork dates and effort (in number of hours of observation).
Species Main traits Geographic occurrence Study site
A. cornigera Solitary flowersBilobate proximal calliRounded oil sacs
Northeastern Brazil(Caatinga)
São MigueTapuio (PI)
A. eriostachys Terminal racemesDiscrete oil sacs
Between theJequitinhonha and SanFrancisco Rivers
Rio Preto S(MG)
A. goyazensisTerminal racemes,bifid proximal callus
High-altitude Cerrado(GO, MG)
Rio Preto S(MG)
Chapada dContagemBiological R(DF)
A. integerrima Terminal racemesFlower white to blue
South Brazil, Paraguayand Argentina
Morro do O(RS)
A. pubescens Laterally expanded oilsacs
Northeastern Brazil;Caatinga enclaves (GO)
Teresina d(GO)
A. salicariifolia Terminal racemes, bifidproximal callus
South America (widelydistributed)
Porto Murt(MS)
B. silvaticus White or purpleflowersDensely thricomatic
Bolivia and Brazil (MS) Porto Murt(MS)
otes: Abbreviation of Brazilian states: DF: Distrito Federal; MG: Minas Gerais; GO: Goiás
08 (2013) 401– 411
Materials and methods
Study species
Six species of Angelonia and Basistemon silvaticus were studiedin several localities in Brazil (Table 1). Except for the Parque NaturalMorro do Osso site, which is an area of open fields in the AtlanticForest biome, all the other localities belong to the domain of openvegetation biomes, characterized by seasonal drought, a significantrate of endemism and highly endangered species (Werneck, 2011).
Fieldwork
The behavior of the bees visiting the flowers was observeddirectly, roughly between 8 am and 3 pm, the times varying inaccordance with local weather conditions. The bees’ behavior wasdocumented with photographs and video. During the observations,the resource gathered, the visitation time and the floral organsthe bees came into contact with were all noted. Species that cameinto contact with fertile organs during visitation were consideredpollinators, and the other species were considered illegitimate vis-itors. Information about flower anthesis, the location of resources(oil and/or pollen), colors and the scents released was also noted.The captured bees were deposited in the Entomological collec-tions of the Universidade de São Paulo (CEPANN) and Universidadede Brasilia. Voucher specimens of the seven plant species weredeposited at the Herbarium of the Escola Superior de AgriculturaLuís de Queiroz (ESA) – Universidade de São Paulo.
Floral morphology
Flower samples were collected and fixed in FAA (37% formalde-hyde, acetic acid and 50% ethanol, 1:1:2) for morphological studiesand for making illustrations with a camera lucida adapted to a
stereomicroscope. Between 7 and 18 flowers were examined perspecies. The morphology of flowers from Angelonia and Basistemonare described using the categories given in Vogel (1974), Vogel andMachado (1991) and Vogel and Cocucci (1995). This informationgeographic occurrence, study sites with coordinates, the vegetational type present,
Coordinatesand altitude
Vegetationtype
Observationeffort (h)
Dates ofobservation
l do 5◦30′S,41◦19′W,200 m
Seasonally DryTropical Forest(Caatinga)
15 21–23.4.2008
tate Park 18◦12′S,43◦19′W,1500 m
TropicalSavanna(Cerrado)
44 28–30.4.2008,1–6.9.2008,11–12.9.2008;27.4–1.5.2009
tate Park 18◦12′S,43◦19′W,1500 m
TropicalSavanna(Cerrado)
14 28–30.4.2008;27.4–1.5.2009
a
eserve
15◦38′S,47◦51′W,1200 m
TropicalSavanna(Cerrado)
8 20.12.2011
sso Park 30◦07′S,51◦14′W,143 m
Atlantic Forest(Grassland)
55 7–11.12.2007;9–14.11.2008
e Goiás 13◦34′S,47◦11′W,405 m
Seasonally DryTropical Forest(Caatinga)
16 1–2.4.2010
inho 21◦37′S,57◦49′W, 90 m
Chaco 10 29–30.1.2008
inho 21◦37′S,57◦49′W, 90 m
Chaco 25 24–30.1.2008
; MS: Mato Grosso do Sul; PI: Piauí; RS: Rio Grande do Sul.

A.C. Martins et al. / Flora 208 (2013) 401– 411 403
Fig. 1. Main morphological traits of the Angelonia salicariifolia (a, b) flower. (a) Frontal view. (b) Lateral view. (st) stamens, (pc) proximal callus, (dc) distal callus (pt) palate,(
idslaaoa
M
pEdfltit(
R
F
aeAmfieocecec
rt
sl) superior lobes, (ml) medium lobes, (E) elaiophore and (L) labellum.
ncludes the size and shape of the corolla lobes and oil sacs, theensity and position of the elaiophores and their accessibility, mea-ured in relation to the corolla’s opening, the presence of calli in theabelum, the size and shape of calli on the lower lip, the palate size,nd the presence of glandular hairs other than elaiophores (Fig. 1and b). The terminology used to describe the floral organs is basedn the literature (Barringer, 1981; Souza, 1996; Vogel, 1974; Vogelnd Machado, 1991).
orphology of the oil-collecting apparatus
The oil-collecting apparatuses of Centris hyptidoides, Chale-ogenus sp. and Caenonomada bruneri were examined in a Scanninglectron Microscope LEO 440. These species were chosen forescription because they were observed only in Plantaginaceaeowers so far and they are probably adapted to oil gathering inhese flowers. The anterior legs were removed from pinned spec-mens, and they were cleaned, dried and metalized in gold. Theerminology utilized follows that proposed by Neff and Simpson1981) and Cocucci et al. (2000), with some modifications.
esults
loral morphology
Two general patterns in the floral morphology of Angeloniare distinguished: 1. a deep corolla tube and short lobes (e.g. A.ryostachys) (Fig. 2); 2. a short corolla tube and long lobes (e.g.. salicariifolia), giving the flower the appearance of being open-outhed (Fig. 2b). Basistemon silvaticus flowers are similar to the
rst pattern. The nototribic position of fertile organs in the flow-rs of Angelonia and Basistemon induces the depositing of pollenn the mesoscutum or front head of visitors. In flowers with deeporolla (A. integerrima and A. eriostachys) pollen transfer and adher-nce occurs on the mesoscutum, while in the flowers with shortorolla the anterior surface of head is the place where the adher-nce of pollen takes place. All studied flowers present a protandric
ondition of reproductive organs’ maturation.The shape of oil sacs varies among the species studied andelates to the corolla types. In the flowers with deeper corolla,he oil sacs are slightly evident (Fig. 2a), while in the open-mouth
flowers the oils sacs are evident, rounded and, in some cases, projectlaterally (e.g. A. cornigera) (Fig. 3b). In the latter group of flowers,the elaiophore is easily accessed close to the corolla opening,although this varies among the species. For example, in A. salicari-ifolia, the edges of the elaiophore area coincide with the floweropening. In the flowers with deeper corolla, the elaiophore area isdistant from the corolla opening and the fertile organs, and requiresthe pollinator to enter the flower with the whole body. This alsooccurs in B. silvaticus, but in this case the trichomes are more con-centrated on the discrete sacs, and in other parts of the inner corolla,including regions more accessible to the visitors.
The most conspicuous differences observed between the sixspecies of Angelonia relate to the shape and size of the proximalcallus, the distal callus and the palate (Fig. 3). Angelonia goyazensis,A. cornigera and A. salicariifolia present a long and bifid proximalcallus, which is bilobed in A. cornigera, with the corolla color dif-fering between species (Fig. 3b, d and e). In A. integerrima and A.eriostachys, the proximal callus consists of a white protuberance(Fig. 3c and f). The proximal callus is preceded by the palate, withthe concave area variable between species and especially conspicu-ous in A. salicariifolia, because of its green color (Fig. 3e). The palateis located between the proximal callus and the elaiophores. Thedistal callus is in juxtaposition with the proximal callus. In A. goy-azensis, the distal callus is excrescent (Fig. 3d). In A. integerrima it issimilar but less protuberant, and all the labellum has a wavy aspect(Fig. 3f). In A. salicariifolia and A. cornigera, the proximal callus isnot evident and it forms part of the curvature of the corolla. Whilein the other species of Angelonia the proximal callus is prominent,in A. eryostachys it is the distal callus that is conspicuous, beingwhite in color. The flowers of B. silvaticus possess only one cal-lus in the median region of the corolla, near the elaiophore area(Fig. 3a).
The flowers of the Plantaginaceae studied have non-oil-producing trichomes, possibly associated with scent production,but their function was not investigated. Sweetish odor was detectedon the flowers of A. goyazensis, A. salicariifolia and A. integerrima.Colors of the species studied varied from white to purple. Some
species have spots (A. salicariifolia, Angelonia integerrima), and oth-ers just have one color, such as lilac (A. eryostachys), purple (A.cornigera), two simultaneous tones of lilac (more frequent) or white(A. goyazensis and B. silvaticus).
404 A.C. Martins et al. / Flora 208 (2013) 401– 411
Fig. 2. Two types of flower in the Angelonia genus, in relation to the total corolla length. (a) In the A. eryostachys flower the corolla tube is long and the lobes are short, whilein the A. goyazensis flower (b) the tube is short, the lobes are long, and the flower resembles an open mouth. This second type is common in the majority of Angelonia species(scale = 5 mm). (sl) superior lobes, (ml) medium lobes, (E) elaiophore, (L) labellum, (pc) proximal callus and st (stamens).
Fig. 3. Specialized structures in Angelonia and Basistemon flowers indicated by arrows. (a) Medium callus in B. silvaticus. (b) Proximal bifid (1) and bilobed (2) callus in A.cornigera. (c) Distal callus in A. eriostachys. (d) Proximal callus (1) and distal callus (2) in A. goyazensis. (e) Proximal callus (1) and distal callus (2) in A. salicariifolia. (f) Proximalcallus (1) and distal callus (2) in A. integerrima.
A.C. Martins et al. / Flora 208 (2013) 401– 411 405
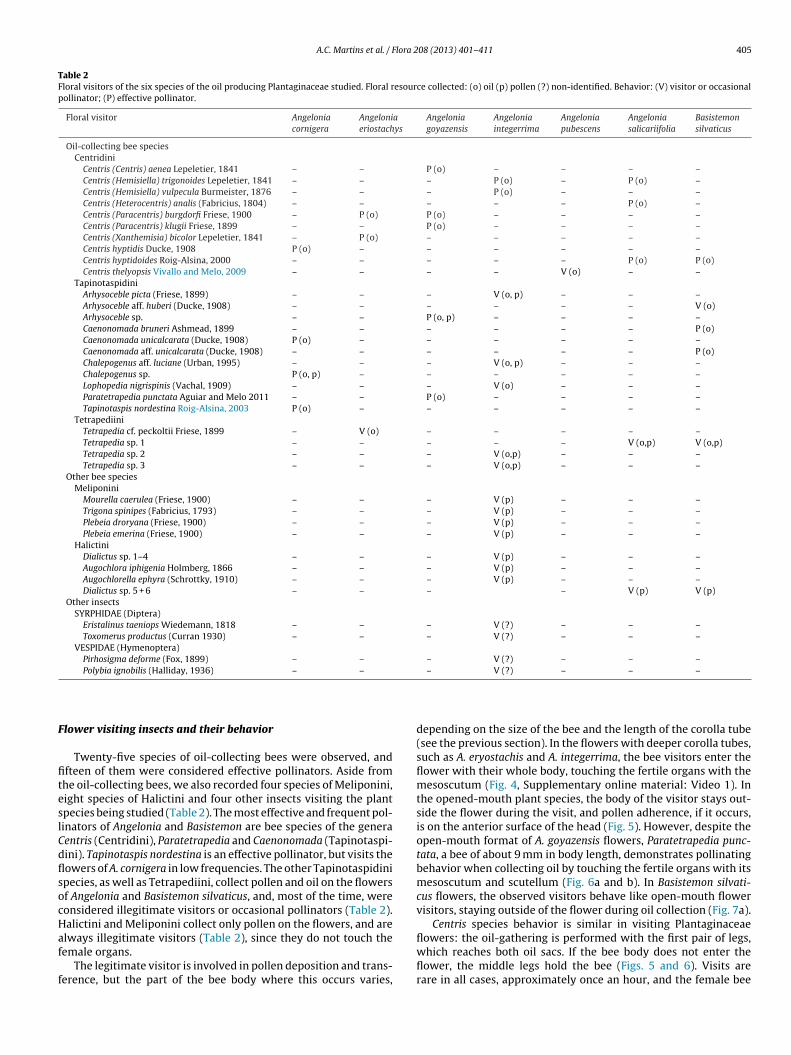
Table 2Floral visitors of the six species of the oil producing Plantaginaceae studied. Floral resource collected: (o) oil (p) pollen (?) non-identified. Behavior: (V) visitor or occasionalpollinator; (P) effective pollinator.
Floral visitor Angeloniacornigera
Angeloniaeriostachys
Angeloniagoyazensis
Angeloniaintegerrima
Angeloniapubescens
Angeloniasalicariifolia
Basistemonsilvaticus
Oil-collecting bee speciesCentridini
Centris (Centris) aenea Lepeletier, 1841 – – P (o) – – – –Centris (Hemisiella) trigonoides Lepeletier, 1841 – – – P (o) – P (o) –Centris (Hemisiella) vulpecula Burmeister, 1876 – – – P (o) – – –Centris (Heterocentris) analis (Fabricius, 1804) – – – – – P (o) –Centris (Paracentris) burgdorfi Friese, 1900 – P (o) P (o) – – – –Centris (Paracentris) klugii Friese, 1899 – – P (o) – – – –Centris (Xanthemisia) bicolor Lepeletier, 1841 – P (o) – – – – –Centris hyptidis Ducke, 1908 P (o) – – – – – –Centris hyptidoides Roig-Alsina, 2000 – – – – – P (o) P (o)Centris thelyopsis Vivallo and Melo, 2009 – – – – V (o) – –
TapinotaspidiniArhysoceble picta (Friese, 1899) – – – V (o, p) – – –Arhysoceble aff. huberi (Ducke, 1908) – – – – – – V (o)Arhysoceble sp. – – P (o, p) – – – –Caenonomada bruneri Ashmead, 1899 – – – – – – P (o)Caenonomada unicalcarata (Ducke, 1908) P (o) – – – – – –Caenonomada aff. unicalcarata (Ducke, 1908) – – – – – – P (o)Chalepogenus aff. luciane (Urban, 1995) – – – V (o, p) – – –Chalepogenus sp. P (o, p) – – – – – –Lophopedia nigrispinis (Vachal, 1909) – – – V (o) – – –Paratetrapedia punctata Aguiar and Melo 2011 – – P (o) – – – –Tapinotaspis nordestina Roig-Alsina, 2003 P (o) – – – – – –
TetrapediiniTetrapedia cf. peckoltii Friese, 1899 – V (o) – – – – –Tetrapedia sp. 1 – – – – – V (o,p) V (o,p)Tetrapedia sp. 2 – – – V (o,p) – – –Tetrapedia sp. 3 – – – V (o,p) – – –
Other bee speciesMeliponini
Mourella caerulea (Friese, 1900) – – – V (p) – – –Trigona spinipes (Fabricius, 1793) – – – V (p) – – –Plebeia droryana (Friese, 1900) – – – V (p) – – –Plebeia emerina (Friese, 1900) – – – V (p) – – –
HalictiniDialictus sp. 1–4 – – – V (p) – – –Augochlora iphigenia Holmberg, 1866 – – – V (p) – – –Augochlorella ephyra (Schrottky, 1910) – – – V (p) – – –Dialictus sp. 5 + 6 – – – – V (p) V (p)
Other insectsSYRPHIDAE (Diptera)
Eristalinus taeniops Wiedemann, 1818 – – – V (?) – – –Toxomerus productus (Curran 1930) – – – V (?) – – –
VESPIDAE (Hymenoptera)Pirhosigma deforme (Fox, 1899) – – – V (?) – – –Polybia ignobilis (Halliday, 1936) – – – V (?) – – –
F
fiteslCdflsocHaf
f
lower visiting insects and their behavior
Twenty-five species of oil-collecting bees were observed, andfteen of them were considered effective pollinators. Aside fromhe oil-collecting bees, we also recorded four species of Meliponini,ight species of Halictini and four other insects visiting the plantpecies being studied (Table 2). The most effective and frequent pol-inators of Angelonia and Basistemon are bee species of the generaentris (Centridini), Paratetrapedia and Caenonomada (Tapinotaspi-ini). Tapinotaspis nordestina is an effective pollinator, but visits theowers of A. cornigera in low frequencies. The other Tapinotaspidinipecies, as well as Tetrapediini, collect pollen and oil on the flowersf Angelonia and Basistemon silvaticus, and, most of the time, wereonsidered illegitimate visitors or occasional pollinators (Table 2).alictini and Meliponini collect only pollen on the flowers, and are
lways illegitimate visitors (Table 2), since they do not touch theemale organs.The legitimate visitor is involved in pollen deposition and trans-erence, but the part of the bee body where this occurs varies,
depending on the size of the bee and the length of the corolla tube(see the previous section). In the flowers with deeper corolla tubes,such as A. eryostachis and A. integerrima, the bee visitors enter theflower with their whole body, touching the fertile organs with themesoscutum (Fig. 4, Supplementary online material: Video 1). Inthe opened-mouth plant species, the body of the visitor stays out-side the flower during the visit, and pollen adherence, if it occurs,is on the anterior surface of the head (Fig. 5). However, despite theopen-mouth format of A. goyazensis flowers, Paratetrapedia punc-tata, a bee of about 9 mm in body length, demonstrates pollinatingbehavior when collecting oil by touching the fertile organs with itsmesoscutum and scutellum (Fig. 6a and b). In Basistemon silvati-cus flowers, the observed visitors behave like open-mouth flowervisitors, staying outside of the flower during oil collection (Fig. 7a).
Centris species behavior is similar in visiting Plantaginaceae
flowers: the oil-gathering is performed with the first pair of legs,which reaches both oil sacs. If the bee body does not enter theflower, the middle legs hold the bee (Figs. 5 and 6). Visits arerare in all cases, approximately once an hour, and the female bee
406 A.C. Martins et al. / Flora 208 (2013) 401– 411
Fig. 4. Centris (Xanthemisia) bicolor visiting Angelonia eryostachys. The whole bodyof the bee enters the flower to access the elaiophores, touching the fertile organs.Scale = 2 mm.
Fig. 5. Centris thelyopsis collecting oil in Angelonia pubescens flower and contactingthe fertile organs.
Fig. 7. Centris (Hemisiella) vulpecula female collecting oil in the flower of A. inte-gerrima, which was intentionally sectioned to show the bee’s position and behaviorduring oil collection.
Fig. 6. Paratetrapedia punctata visiting an Angelonia goyazensis flower in Chapada da Contagem. (a) The female bee is collecting oil and touching the fertile organs. (b) Thepollen adhers to the bee’s mesoscutum before leaving the flower.

A.C. Martins et al. / Flora 2
Fa
sacmmiibfbnoestVfledso(
rior tarsomeres 2–4 are also pilose and are used in oil-collecting
ig. 8. Basistemon silvaticus visitors. (a) Caenonomada bruneri collecting oil with itsnterior pair of legs. (d) Dialictus sp. collecting pollen.
earches for oil among the flowers, flying rapidly around the plant,nd only entering open flowers. After gathering the oil, all oil-ollecting species transfers it to its posterior legs in flight returningany times to the flower continually collecting more (Supple-entary online material: Videos 1–3). When the flowers of A.
ntegerrima were cut transversally and exposed to the female bee,t was observed that the bee also used the mandibles to hold itsody inside the flower, while exploiting the elaiophore with theorelegs (Fig. 8). It was observed that Centris hyptidis also held theody with the mandibles in Angelonia cornigera, however it wasot possible to identify exactly which flower structure was heldn. In Basistemon flowers, where the sacs are not well defined, alllaiophore areas are explored with both forelegs, while the midlegsupport the body on the flower and the front legs come into con-act with the fertile organs (Fig. 8; Supplementary online materialideo 2). Centris hyptidoides females act as pollinator of B. silvaticusowers in most of the visits, however a different kind of floral oilxploitation was also observed. In this specific behavior the femaleso not contact the fertile organs of B. silvaticus, since their legs are
tretched-out toward the elaiophores, maintaining the whole bodyutside the flower while the midlegs provide support on the leavesSupplementary online material: Video 3).08 (2013) 401– 411 407
The oil-collecting Tapinotaspidini bees, Arhysoceble, Chale-pogenus and Lophopedia, collect oil and pollen in Angelonia and/orB. silvaticus. All species in these genera are about 6 mm long or less,and they do not touch the fertile organs of the flowers during oilgathering. The Tapinotaspidini bees use the forelegs, alternatelyor at the same time, for foraging on the oil sacs. In the samevisit these bees can also collect pollen, landing on the anthersin an upside-down position. However, Paratetrapedia punctata,Tapinotaspis nordestina and species of Caenonomada do not col-lect pollen on the Plantaginaceae species, and during oil-collectionthese bees act as effective pollinators of the flowers. In A. cornigeraand A. goyazensis, the pollen adheres to the labrum, the frons andclypeus of the female, except with Paratetrapedia punctata (Fig. 7).In B. silvaticus the pollen adherence occurs on the mesoscutum ofCaenonomada bruneri and C. aff. unicalcarata (Fig. 8a).
Tetrapedia females collect both oil and pollen on the flowersof A. eriostachys, A. integerrima, A. salicariifolia and B. silvaticus.These bees are not big enough (ca. 10 mm) to touch the fertileorgans. Indeed they are rare visitors and their detailed behaviorwas not observed. Pollen robbers were registered in almost all stud-ied species, and all belong to the subfamily Halictinae and subtribeMeliponina (Table 2). They collect pollen with their forelegs, afterlanding on the stamen in an upside-down position (Fig. 8b). Theprotandric condition of the flowers inhibits pollination by visitorspresenting this behavior because the pollen-offering flowers haveno receptive stigma.
Males of the oil-collecting bees were commonly observed,searching for females on the flowers of Angelonia and B. silvaticus:this behavior was observed in Centris (H.) analis, C. (H.) vulpecula,Centris hyptidoides, Chalepogenus sp., and Caenonomada unical-carata. They were never observed landing on the flowers to collectany resource, so they obviously do not act as pollinators.
Aside from bees, other insects were also observed visiting, butnot pollinating A. integerrima, such as the syrphid flies Eristalinustaeniops and Toxomerus productus (Syrphidae) and the wasp speciesPirhosigma deforme and Polybia ignobilis (Vespidae). These insectsfly around the flowers, landing on them less frequently, and do notcollect oil or pollen. The chrysomelid beetles Trichaltica sp. werealso observed on A. integerrima, feeding on pollen.
Morphology of the oil-collecting apparatus
The oil-collecting apparatuses of Caenonomada bruneri and C.aff. unicalcarata are identical and restricted to the anterior pair oflegs. In both species they are located on the anterior basitarsus andtarsomeres 2–3, and are composed of two kinds of setae: thick, longand leaf-like setae with acute apexes, and thin, short and branchedsetae. In the basitarsus, the second type of setae are distributedover the ventral surface, while the first type of setae are groupedon the dorsal margin, forming a comb-like structure (Fig. 9a andb). On tarsomeres 2–3, the thin and branched setae are intermixedwith simple, leaf-like setae (Fig. 9a).
Centris hyptidoides possesses oil-collecting structures restrictedto the forelegs. On the middle legs, a structure resembling a combcan be observed, but it is composed of scattered setae and probablynot functional. On the anterior basitarsus, the oil-collecting appa-ratus is composed of a primary comb, specialized setae (Fig. 9e) anda vestigial secondary comb. The primary comb is curved and com-posed of simple, large setae, with rounded apexes (Fig. 9e). Thereare numerous simple, leaf-like setae on the dorsal surface. The ven-tral surface of the basitarsus is covered with numerous plumosesetae, intermixed with long, simple and thick setae. The ante-
(Fig. 9d); the anterior margin and dorsal surface possesses manylong, leaf-like setae, with rounded apexes (Fig. 9c), similar to thoselocated on the dorsal surface of the basitarsus. A set of modified

408 A.C. Martins et al. / Flora 208 (2013) 401– 411
Fig. 9. Scanning electron micrograph showing the oil-collecting apparatuses of pollinators specialized on Plantaginaceae: Caenonomada bruneri (a–b) and Centris hyptidoides(c–e). (a). The third tarsomere of the anterior leg, in ventral view, showing the dorsal setae (ds) which form a set of setae resembling a comb (scale = 0.05 mm). (b) Tarsomeres2–3 of the anterior leg (scale = 0.2 mm). (c) Second tarsomere, detail of the anterior leg (scale = 0.05 mm) showing the anterior surface (as) covered in long, leaf-like setae,that are curved, with an oval apex. (d). Tarsomeres 2–4 showing simple and curved setae, with concave, spoon-like apexes (scale = 0.2 mm). (e) Anterior basitarsus in frontalv and sa mose
str
tOssas(d
iew, showing a unique comb of setae (ac) – specific to the Centris group hyptidis –re numerous simple and leaf-like setae, and, in the ventral part (vp), there are plu
etae is located on the posterior margin of the tarsomeres, andhese setae are simple and spoon-like, with a concave surface andounded apexes.
In Chalepogenus sp., the oil-collecting apparatus is located onhe basitarsus and tarsomeres 2–3 of the forelegs (Fig. 10a–c).n the basitarsus, the dorsal surface is covered in long, thick
etae, while on the ventral surface there are long, thin, branchedetae (Fig. 10b). Tarsomeres 2–3 present specialized setae, which
re sparsely distributed but in higher concentration on tar-omere 2 (Fig. 10a). These setae are strongly curved and leaf-likeFig. 10c) and on their concave surface there is a longitudinaluct.etae in a position posterior to the comb (pgs). Dorsally, to the primary comb, theresetae mixed with simple and thick setae. (scale = 0.2 mm).
Discussion
From oil-collecting specialized bees to pollen robbers: theflower visitors of Angelonia and Basistemon
Angelonia and Basistemon flowers are highly specialized, andvisited by a specialized guild of pollinators, i.e. “oil-bees”, whichbehave in a similar way and exert similar selection pressures on the
flowers, characterizing a functional specialization (Fenster et al.,2004). However, the same plant species can be visited by differ-ent guilds of insects (A. integerrima was visited in the presentstudy by 14 different bees, 2 wasps and 2 Syrphidae species),
A.C. Martins et al. / Flora 208 (2013) 401– 411 409
Fig. 10. Scanning electron micrograph of the oil-collecting apparatuses of Chalepogenus sp., pollinator of A. cornigera. (a) Anterior tarsomeres 2–4 showing specialized setaefor oil-collection and the ventral surface covered with leaf-like setae (scale = 0.03 mm). (b) Anterior basitarsus (bta) showing anterior surface covered by simple, wide andt cale =
l
stvt2bpTodaiatpaes
f1ettm1E
hick setae (cs), and the ventral surface covered with thin and branching setae (cr) (songitudinal channel (scale = 0.01 mm).
uggesting a more generalist system from an ecological perspec-ive (Waser et al., 1996). A generalization in regard to the number ofisitors was observed on a geographical scale in systems which fea-ured the specialization of oil-flower-bee mutualism (Cosacov et al.,008). In our sampling, Angelonia cornigera flowers were pollinatedy the Tapinotaspidini bees Caenonomada unicalcarata and Chale-ogenus sp., while in other geographical regions Centris hyptidis andapinotaspis nordestina were pointed out as specialized pollinatorsf this plant species (Machado et al., 2002). Similarly the widely-istributed Angelonia salicariifolia was visited by generalist, andlso widely-distributed, bees in different areas: by Centris fuscatan a Seasonally Dry Tropical Forest area (Vogel and Machado, 1991)nd by Centris (Heterocentris) analis in a Chaco area, as reported inhe present study. In accordance with the geographical mosaic pro-osed by Thompson (2005), the flower visitors in a specific localityre part of a wider spectrum of possible interactions, when consid-red alongside the geographical distribution of widely distributedpecies, such as A. salicariifolia.
Pollination of Plantaginaceae by Centris bees was known alreadyor Angelonia and Monttea species from previous studies (Vogel,974; Simpson et al., 1990; Vogel and Machado, 1991; Machadot al., 2002). It is reinforced by the present observations, in whichheir role in pollination also of Basistemon is presented for the first
ime. It is assumed that Malpighiaceae and Centris bees are pri-arily associated as pollination partners (Vogel, 1974; Anderson,979), both having arisen synchronously in the Early Cenozoicra (Renner and Schaefer, 2010). Malpighiaceae taxa have been
0.02 mm). (c) Tarsomere 3 detail, showing curved setae (cc) which possess a median
a unique floral oil resource for many groups of the genus Cen-tris (Machado, 2004). However some younger oil-producing plantfamilies, such as Calceolariaceae, Krameriaceae and Plantaginaceaeare associated with distinct subgenera of Centris by new kindsof exploitation of flowers with different morphological patterns(Simpson et al., 1977; Cosacov et al., 2009).
Another group of visitors of Plantaginaceae oil-flowers, thetapinotaspidine bees are also mainly associated with Malpighi-aceae flowers (Vogel, 1974) but some genera of them present aclose relationship with trichomatic oil glands, for example Cal-ceolariaceae flowers (Vogel, 1974). A special case of adaptationof a Plantaginaceae species associated to a tapinotaspine bee wasreported in Tapinotaspis nordestina which explores oil glands inAngelonia cornigera with its long midlegs (Machado et al., 2002).Long midlegs are present in all species of Tapinotaspis occur-ring in Chaco and Pampa vegetation (Roig-Alsina, 2003) and areassociated with the morphologically different oil flowers Nierem-bergia/Solanaceae (Cocucci, 1991) and/or Sisyrinchium/Iridaceae(Cocucci et al., 2000). Cocucci et al. (2000) stated that Caenono-mada species are only known to visit the genera Angelonia, Aguiarand Melo (2009) observed them visiting Monopera, and now thisis documented also for Basistemon. Most probably they visit onlyPlantaginaceae species for oil-collecting. Cocucci et al. (2000)
suggested that Caenonomada bruneri collects oil from Cypella(Iridaceae), based on indirect observation of pollen from theseplants on the insect’s bodies, but this observation was neverconfirmed.
4 lora 2
iitM2tIpat2
Ff
sdoflsdwtaMrwAoaaoab
twatsvo
temstn2l1osbcaoei
ap(
10 A.C. Martins et al. / F
In Angelonia flowers, Tetrapedia bees collect floral oil sporad-cally, and in other floral-oil hosts these bees were reported asllegitimate visitors, exploring the elaiophores without having con-act with the fertile organs (Sazima and Sazima, 1989; Vogel and
achado, 1991; Singer and Cocucci, 1999; Sigrist and Sazima,005). Other bees that collect pollen also act as illegitimate visi-ors in Basistemon, Angelonia and Monttea (Simpson et al., 1990).n A. pubescens, stingless bees also collect oil, although they do notossess specialized structures for collecting this resource (Vogelnd Machado, 1991). They probably use it in nesting construc-ion, with a similar role as played by plant resins (Michener,007).
loral morphology of Angelonia and Basistemon: whicheatures are required from pollinators?
Angelonia eryostachys has a flower structure resembling the B.ilvaticus flowers, both lacking developed calli and possessing lesseveloped oil sacs, in contrast to the other five studied speciesf Angelonia (Vogel, 1974; Vogel and Machado, 1991). Among theoral-oil producing Plantaginaceae, Angelonia presents the mostpecialized flowers, and most Angelonia species have two well-efined oil sacs. Well-developed calli, other structure associatedith oil-collecting visitors, can also be observed in flowers of
his genus (Vogel, 1974; Vogel and Machado, 1991). Angeloniand Basistemon are both monophyletic, as well as Monttea andelosperma, according to a phylogenetic analysis including few
epresentatives of them and any Monopera (Albach et al., 2005),hich latter is known to be closely related morphologically tongelonia (Barringer, 1983). The relationships between the floral-il producing Plantaginaceae species remain poorly understood yet,nd it is difficult to establish homologies between their floral char-cters. Furthermore, the non-oil-producing Basistemon species arenly known from one taxonomic study (Barringer, 1981), and only
few observations of flower visitors to the genus Monopera haveeen published (Aguiar and Melo, 2009).
Basistemon silvaticus presents widely distributed elaiophores inhe inner corolla, differing from other species in the genus, forhich the elaiophores are restricted to the corolla base (B. argutus),
nd can be slightly concentrated (B. pulchellus) or highly concen-rated (B. spinosus) in the inner corolla (Barringer, 1985). Otherpecies of Basistemon lack glandulary hairs (B. bogotensis, B. peru-ianus, B. intermedius, B. klugii) and also nectaries, probably offeringnly pollen to visitors.
The morphology of Angelonia and Basistemon flowers resembleshat of the African floral-oil producing species of the related withach other genera Colpias and Diascia. The African genera requireorphological different features from the pollinators and it is pos-
ible to do a parallel comparison with Angelonia and Basistemon (forhe phylogenetic unrelatedness of African Scrophulariaceae and theewly circumscribed Plantaginaceae family see Olmstead et al.,001 and Oxelman et al., 2005). In Diascia a long pair of anterior
egs to reach the long spurs are required (Steiner and Whitehead,990) as well as a mode of fixing the body on the flower duringil-collection (Steiner, 1990), similarly to the situation with thehort-tube Angelonia flowers. In Colpias the right size of thorax andody length (Steiner and Whitehead, 2002) fitting the flowers isomparable with the situation of long-tube flowers of Angeloniand B. silvaticus. Long legs to exploit Angelonia oil sacs were onlybserved in Centris hyptidis and Tapinotaspis nordestina (Machadot al., 2002), a special and extreme adaptation that deserves furthernvestigations.
Grasping with the mandibules in specific parts of the flowers is common behavior in many oil-bee-flower interactions, for exam-le the insects cling in this way to the flag petals of MalpighiaceaeAnderson, 1979) and Krameriaceae flowers (Simpson et al., 1977)
08 (2013) 401– 411
and the Diascia window (Steiner, 1990). The use of the proximal cal-lus as a support for bees was hypothesized by Vogel and Machado(1991) and refuted in the same paper through the observation thatthe bee’s head remained positioned toward the anthers during oilcollection. They suggested that the colors of the proximal callus andgullet form visual cues for the bee (Vogel and Machado, 1991). Wesuggest that the presence of these structures helps pollinators inentering the flower in the correct position, as the callus decreasesthe distance between the landing area and the anthers. The pollentransfer, in this way, occurs through the mesoscutum or frons ofthe female bee (Vogel and Machado, 1991; Machado et al., 2002).Vogel and Cocucci (1995), however, noted the absence of calli inBasistemon flowers, although they also noted that Arhysoceble aff.huberi is correctly positioned when pollinating Basistemon, similarto the pollinators of Angelonia.
If the callus structures are not used by bees to grasp themwith their mandibules, in which structure do those bees fix theirbodies in short tube Angelonia? In Angelonia integerrima, it wasobserved that the crest between oil sacs is the structure whereCentris vulpecula fixes its body with its mandibles, despite the bodybeing supported by the deep corolla. Also, in Angelonia cornigera,Centris hyptidis was recorded with opened mandibles, biting justbelow the anthers, a position that would also be the crest betweenoil sacs. In conclusion we hypothesize that the anthers should bethe fixation point in short corolla tubes Angelonia and the mediumcallus in Basistemon silvaticus. The crest between oil sacs in longcorolla tubes Angelonia should have the same function.
Morphological features of oil-collecting apparatuses in beesadapted to the exploitation of Plantaginaceae oil sacs
The oil-collecting apparatuses of Centris – consisting of combsof setae on the anterior and midlegs – are primarily associ-ated to Malpighiaceae flowers (Neff and Simpson, 1981). Somemodified kinds of setae occur in species of the Centris subgenusWagenknechtia and the hyptidis group, both are associated withPlantaginaceae (Simpson et al., 1990; Machado et al., 2002). Themorphology of Centris hyptidoides, as described here, gives ussome clues about the evolutionary association between the hyptidisgroup (Vivallo and Melo, 2009) and oil-producing Plantaginaceae.Regarding the oil combs of the females of C. hyptidoides, they arerestricted to the anterior legs, a characteristic primarily associ-ated with Angelonia flowers in the hyptidis group (Machado et al.,2002). However, C. hyptidoides does not present a first pair of legsas extraordinarily long as the closely related C. hyptidis (Machadoet al., 2002), or as long as those of other species in this group, suchas C. thelyopsis (Vivallo and Melo, 2009). The oil-collecting appara-tuses of the species described are restricted to the anterior pair oflegs and extend to the tarsomeres, differing from the common pat-tern of combs of setae on the basistarsus of the anterior and midlegs(Neff and Simpson, 1981). The modifications are probably associ-ated with the gathering of oil in oil-sacs, since only the anteriorlegs are effectively used for this purpose. In Colpias mollis (Scro-phulariaceae), the floral oils are produced in similar oil sacs, andits pollinator, Rediviva albifasciata, also possesses an oil-collectingapparatus on the tarsomeres (Steiner and Whitehead, 2002).
Acknowledgements
We thank IBAMA, SMA-RS and IEF-MG for permission to workin their Conservation Units. For taxonomic identification we are
grateful to: Vinicius C. Souza (Plantaginaceae), Amanda C. Pires andMírian N. Morales (Syrphidae), Marcel G. Hermes (Vespidae), JoãoM.F. de Camargo (Meliponini), Rodrigo B. Gonc alves (Halictinae)and Felipe Vivallo (Centris). To the colleagues that helped with the
lora 2
fidPtPtd
A
t
gwtttt
fpislf
fowd
R
A
A
A
A
B
B
B
B
C
C
C
C
C
C
F
A.C. Martins et al. / F
eldwork: Jéssica Gillung, Nara Mota, Victor Giorni, Lauren Por-any, Camillo Pedrolo, Verônica Silveira. We also thank FAPESP andROAP-CAPES for their financial support and CNPq for a scholarshipo ACM and IAS. Thanks to Maria C. Gaglianone and Emerson R.ansarin for the suggestions in an early version of the manuscript,o Daniel Yencken for his revision of the text, and to Gabriel Rezendee Paula who kindly took it on to make the drawings.
ppendix A. Supplementary data
Supplementary data associated with this article can be found, inhe online version, at http://dx.doi.org/10.1016/j.flora.2013.07.001.
Video 1 Centris (Hemisiella) vulpecula visiting Angelonia inte-errima. The female bee chooses an opened flower in the raceme,ith the forelegs it scrapes the elaiophores in the oil saccs. Between
he visits to one and another flower, the bee transfers the oil tohe posterior legs in flight. It is noticeable that the bee mesoscu-um touches the fertile organ during oil-collecting, allowing pollenransfer between flowers.
Video 2 Centris hyptidoides visiting Basistemon silvaticus. Theemale bee holds its body in the flower with middle and posteriorair of legs, while the forelegs are scraping the elaiophores in the
nner corolla (all green area). Note the movements of anterior legscraping the oil glands and the transference of oil for the posterioregs during oil-collection and the contact of clipeo and labrum withertile organs.
Video 3 Centris hyptidoides visiting Basistemon silvaticus. Theemale bee behaves differently in this video: while collecting oil,nly the forelegs remain in the flower scraping the elaiophore area,hile the whole body stays out of the flower. This kind of behavioroes not enable the pollen adherence to any part of bee body.
eferences
guiar, A.J.C., Melo, G.A.R., 2009. Notes on oil sources for the bee genus Caenonomada(Hymenoptera, Apidae, Tapinotaspidini). Rev. Bras. Entomol. 53, 154–156.
lbach, D.C., Meudt, H.M., Oxelman, B., 2005. Piecing together the “new” Plantagi-naceae. Am. J. Bot. 92, 297–315.
lves-dos-Santos, I., Gaglianone, M.C., Machado, I.C., 2007. História natural das abel-has coletoras de óleo. Oecol. Bras. 11, 544–557.
nderson, W.R., 1979. Floral conservatism in Neotropical Malpighiaceae. Biotropica11, 219–223.
arringer, K., 1981. A revision of Angelonia (Scrophulariaceae). University of Con-necticut, 135 pp (Ph.D. Dissertation).
arringer, K., 1983. Monopera, a new genus of Scrophulariaceae from South America.Brittonia 35, 111–114.
arringer, K., 1985. Revision of the genus Basistemon (Scrophulariaceae). Syst. Bot.10, 125–133.
uchmann, S., 1987. The ecology of oil flowers and their bees. Annu. Rev. Ecol. Syst.18, 343–369.
ane, J.H., Eickwort, G.C., Wesley, F.R., Spielholz, J., 1983. Foraging, grooming andmate-seeking behaviors of Macropis nuda (Hymenoptera, Melittidae) and useof Lysimachia ciliata (Primulaceae) oils in larval provisions and cell linings. Am.Midl. Nat. 110, 257–264.
appellari, S.C., Melo, G.A.R., Aguiar, A.J.C., Neff, J.L., 2011. Floral oil collection by maleTetrapedia bees (Hymenoptera: Apidae: Tetrapediini). Apidologie 43, 39–50.
ocucci, A.A., 1991. Pollination biology of Nierembergia (Solanaceae). Plant Syst. Evol.174, 17–35.
ocucci, A.A., Sérsic, A., Roig-Alsina, A., 2000. Oil-collecting structures in Tapino-taspidini: their diversity, function and probable origin. Mitt. Münchn. Entomol.Ges. 90, 51–74.
osacov, A., Nattero, J., Cocucci, A., 2008. Variation in pollinator assemblages andpollen limitation in a locally specialized system: the oil-producing Nierembergialinariifolia (Solanaceae). Ann. Bot. 102, 723–734.
osacov, A., Sérsic, A.N., Sosa, V., De-Nova, J.A., Nylinder, S., Cocucci, A.A., 2009.New insights into the phylogenetic relationships, character evolution, and
phytogeographic patterns of Calceolaria (Calceolariaceae). Am. J. Bot. 96,2240–2255.enster, C.B., Armbruster, W.S., Wilson, P., Dudash, M.R., Thompson, J.D., 2004. Pol-lination syndromes and floral specialization. Annu. Rev. Ecol. Evol. Syst. 35,375–403.
08 (2013) 401– 411 411
Gonc alves, R.B., Melo, G.A.R., 2005. A comunidade de abelhas (Hymenoptera, Apidaes. l.) em uma área restrita de campo natural no Parque Estadual de Vila Velha,Paraná: diversidade, fenologia e fontes florais de alimento. Rev. Bras. Entomol.49, 557–571.
Machado, I.C., 2004. Oil-collecting bees and related plants: a review of thestudies in the last twenty years and case histories of plants occurring inNE Brazil. In: Freitas, B.M., Pereira, J.O.P. (Eds.), Solitary Bees: Conservation,Rearing and Management for Pollination. University of Ceará, Fortaleza, pp.255–280.
Machado, I.C., Vogel, S., Lopes, A.V., 2002. Pollination of Angelonia cornigera Hook.(Scrophulariaceae) by long-legged, oil collecting bees in NE Brazil. Plant Biol. 4,352–359.
Michener, C.D., 2007. The Bees of the World. John Hopkins Univ. Press, Baltimore,MD, USA.
Moure, J.S., Urban, D., Melo, G.A.R. (Eds.), 2007. Catalogue of the bees (Hymenoptera,Apoidea) in the Neotropical Region. Sociedade Brasileira de Entomologia,Curitiba.
Neff, J., Simpson, B., 1981. Oil-collecting structures in the Anthophoridae(Hymenoptera): morphology, function, and use in systematics. J. Kansas Ento-mol. Soc. 54, 95–123.
Olmstead, R.G., dePamphilis, C.W., Wolfe, A.D., Young, N.D., Elisons, W.J., Reeves,P.A., 2001. Disintegration of the Scrophulariaceae. Am. J. Bot. 88, 348–361.
Oxelman, B., Kornhall, P., Olmstead, R.G., Bremer, B., 2005. Further disintegration ofScrophulariaceae. Taxon 54, 411–425.
Pinheiro, M., Abrão, B.E., Harter-Marques, B., Miotto, S.T.S., 2008. Floral resourcesused by insects in a grassland community in Southern Brazil. Rev. Bras. Bot. 31,469–489.
Rasmussen, C., Olesen, J.M., 2000. Oil-flowers and oil-collecting bees. In: Totland, Ø.,et al. (Eds.), The Scandinavian Association for Pollination Ecology honours KnutFægri. Norweg Acad. Sci. Lett., Oslo, pp. 23–31.
Renner, S.S., Schaefer, H., 2010. The evolution and loss of oil-offering flowers: newinsights from dated phylogenies for angiosperms and bees. Phil. Trans. Roy. Soc.London B 365, 423–435.
Roig-Alsina, A., 2003. A revision of the bee genus Tapinotaspis Holmberg(Hymenoptera, Apidae, Tapinotaspidini) Mitt. Münchn. Entomol. Ges. 93, 45–53.
Sazima, M., Sazima, I., 1989. Oil-gathering bees visit flowers of eglandular morphsof the oil-producing Malpighiaceae. Bot. Acta 102, 106–111.
Schäffler, I., Dötterl, S., 2011. A day in the life of an oil bee: phenology, nesting, andforaging behavior. Apidologie 42, 409–424.
Sérsic, A.N., Cocucci, A.A., 1996. A remarkable case of ornithophily in Calceo-laria: food bodies as rewards for a non-nectarivorous bird. Bot. Acta 109,172–176.
Sigrist, M.R., Sazima, M.R., 2005. Pollination and reproductive biology of twelvespecies of Neotropical Malpighiaceae: stigma morphology and its implicationfor breeding system. Ann. Bot. 94, 22–41.
Simpson, B.B., Neff, J.L., 1981. Floral rewards: alternatives to pollen and nectar. Ann.Miss. Bot. Gard. 68, 301–322.
Simpson, B.B., Neff, J.L., Seigler, D., 1977. Krameria, free-fatty acids and oil-collectingbees. Nature 267, 150–151.
Simpson, B.B., Neff, J.L., Dieringer, G., 1990. The production of floral oil by Monttea(Scrophulariaceae) and the function of tarsal pads in Centris bees. Plant Syst.Evol. 173, 209–222.
Singer, R.B., Cocucci, A.A., 1999. Pollination mechanisms in four sympatric southernBrazilian Epidendroideae orchids. Lindleyana 14, 47–56.
Souza, V.C., 1996. Levantamento das espécies brasileiras de Scrophulariacae. Tesede doutorado. Universidade de São Paulo.
Steiner, K.E., 1990. The Diascia window: an orientation cue for oil collecting bees.Bot. J. Linnean Soc. 102, 175–195.
Steiner, K., Whitehead, V.B., 1990. Pollinator adaptation to oil-secreting flowers:Rediviva and Diascia. Evol 44, 1701–1707.
Steiner, K., Whitehead, V.B., 2002. Oil secretion and the pollination of Colpias mollis(Scrophulariaceae). Plant Syst. Evol. 235, 53–66.
Thompson, J.N., 2005. The Geographic Mosaic of Coevolution. University of ChicagoPress, Chicago.
Vivallo, F., Melo, G.A.R., 2009. Taxonomy and geographic distribution of the species ofCentris of the hyptidis group (Hymenoptera: Apidae: Centridini), with a descrip-tion of a new species from Central Brazil. Zootaxa 2075, 33–44.
Vogel, S., 1969. Flowers offering fatty oil instead of nectar. In: Abstracts XI. Int. Bot.Congress, Seattle, p. 229.
Vogel, S., 1974. Ölblumen und ölsammelnde Bienen. Trop. subtrop. Pflanzenwelt 7.Steiner, Wiesbaden.
Vogel, S., Cocucci, A., 1995. Pollination of Basistemon (Scrophulariaceae) byoil-collecting bees in Argentina. Flora 190, 353–363.
Vogel, S., Machado, I.C., 1991. Pollination of four sympatric species of Angelonia(Scrophulariaceae) by oil-collecting bees in NE Brazil. Plant Syst. Evol. 178,153–178.
Waser, N.M., Chittka, L., Price, M.V., Williams, N.M., Ollerton, J., 1996. Generalizationin pollination systems and why it matters. Ecology 77, 1043–1060.
Werneck, F.P., 2011. The diversification of eastern South American open vegeta-tion biomes: historical biogeography and perspectives. Quaternary Sci. Rev. 30,1630–1648.



















