
Changes in granule cells of the ferret olfactory bulb associated with imprinting on prey odours
12
114 Brain Research, 373 (198~) 114 123 Elsevier BRE 11674 Changes in Granule Cells of the Ferret Olfactory Bulb Associated with Imprinting on Prey Odours B. REHN 1.4,W. BREIPOHL 1.2,A.S. MENDOZA L3and R. APFELBACH 4 qnstitut f(ir Anatomie, Universittitsklinikum Essen, 4300 Essen (F. R.G.), 2Department of Anatomy, University of Queensland, St. Lucia~Brisbane, Qld. (Australia), 31nstitut fiir Anatomie, Medizinische Hochschule Li~eck, 2400 Lf~beck and 41nstitutfiir Biologie, Universitiit Tiibingen, 7400 Tabingen (F. R. G.) (Accepted September 18th, 1985) Key words: olfactory bulb development - - olfactory granule cell - - odour imprinting - - ferret The maturation of the granule cells of the ferret olfactory bulb around the time of odour imprinting has been examined. Rapid Gotgi impregnation studies revealed a temporal overshoot in the development of the spines on the external and internal dendrites of the granule cells. In contrast, the number of somatic spines decreased continuously. Electron microscopical examinations of the synaptic contacts in the external plexiform layer revealed that the time course of synapse and reciprocal synapse formation was similar to that of the formation of the spines on the external dendrites. The results were taken as evidence that both the Golgi and the electron micro- scopical investigations described the same developmental process of postnatal synaptic rearrangement. INTRODUCTION Various investigations of the morphological and functional maturation of the mammalian central ner- vous system show that development is not complete at birth6,7,9.10.14,28,32, 51. Information concerning the. olfactory system is incomplete. Since it could be shown 3.5 that the period between day 60 and day 90 is crucial for imprinting ferrets to prey odours, we wanted to know whether there are concomitant mor- phological changes in the olfactory system of this spe- cies at this stage of development. In the light of studies on the plasticity of the mamma- lian olfactory systemS,33, 45 we were especially inter- ested in the postnatal maturation of granule cells dur- ing periods of odour imprinting. In the ferret, as in other mammals, granule cells are intrinsic neurons which modulate the output of the relay neurons (mi- tral and tufted cells) and granule cells continue to be formed in postnatal lifel, s,2s. According to Altman's principle of hierarchy I it must be presumed that the maturation of granule cells (microneurons which lack an axon) is especially susceptible to environmental influences. Therefore the present investigation ad- dresses the question: do the granule cells of ferret ol- factory bulb go through a specific process of morpho- logical maturation. MATERIALS AND METHODS A total of 18 female ferrets (Mustela putorius furo L.) aged 30, 60, 90, 120 and 150 days, bred andmain- tained in captivity, were used. The animals were an- aesthetized with Nembutal (0.06 mg/g body weight i.p.) and tissues fixed by peffusion intracardialty with 3,2% glutaraldehyde and 2.6% paraformaldehyde in 0.09 M sodium cacodylate buffer (pH 7.35) at room temperature. The olfactory epithelium proprium (OEP) of the nasal septum and the main olfactory bulb (MOB) were dissected free and stored in the same fixative at 4 °C for several hours (electron mi- croscopical procedure) or even longer (light micro- scopical procedure). Light microscopy To quantify age-related changes in the number of olfactory knobs per unit area the following procedure was used so as to exclude regional differences in the Correspondence: B. Rehn, Institut for Anatomie, Universit~itsklinikum Essen, Hufelandstr. 55; 4300 Essen, F.R.G. 0006-8993/86/$03.50 © 1986 Elsevier Science Publishers B.V. (Biomedical Division)
Transcript of Changes in granule cells of the ferret olfactory bulb associated with imprinting on prey odours

114 Brain Research, 373 (198~) 114 123 Elsevier
BRE 11674
Changes in Granule Cells of the Ferret Olfactory Bulb Associated with Imprinting on Prey Odours
B. REHN 1.4, W. BREIPOHL 1.2, A.S. MENDOZA L3 and R. APFELBACH 4
qnstitut f(ir Anatomie, Universittitsklinikum Essen, 4300 Essen (F. R.G.), 2Department of Anatomy, University of Queensland, St. Lucia~Brisbane, Qld. (Australia), 31nstitut fiir Anatomie, Medizinische Hochschule Li~eck, 2400 Lf~beck and
41nstitut fiir Biologie, Universitiit Tiibingen, 7400 Tabingen (F. R. G.)
(Accepted September 18th, 1985)
Key words: olfactory bulb development - - olfactory granule cell - - odour imprinting - - ferret
The maturation of the granule cells of the ferret olfactory bulb around the time of odour imprinting has been examined. Rapid Gotgi impregnation studies revealed a temporal overshoot in the development of the spines on the external and internal dendrites of the granule cells. In contrast, the number of somatic spines decreased continuously. Electron microscopical examinations of the synaptic contacts in the external plexiform layer revealed that the time course of synapse and reciprocal synapse formation was similar to that of the formation of the spines on the external dendrites. The results were taken as evidence that both the Golgi and the electron micro- scopical investigations described the same developmental process of postnatal synaptic rearrangement.
INTRODUCTION
Various investigations of the morphological and
functional maturation of the mammalian central ner-
vous system show that development is not complete at birth6,7,9.10.14,28,32, 51. Information concerning the.
olfactory system is incomplete. Since it could be
shown 3.5 that the period between day 60 and day 90 is
crucial for imprinting ferrets to prey odours, we wanted to know whether there are concomitant mor-
phological changes in the olfactory system of this spe- cies at this stage of development.
In the light of studies on the plasticity of the mamma- lian olfactory systemS,33, 45 we were especially inter-
ested in the postnatal maturation of granule cells dur-
ing periods of odour imprinting. In the ferret, as in
other mammals, granule cells are intrinsic neurons which modulate the output of the relay neurons (mi- tral and tufted cells) and granule cells continue to be formed in postnatal lifel, s,2s. According to Altman's
principle of hierarchy I it must be presumed that the maturation of granule cells (microneurons which lack
an axon) is especially susceptible to environmental influences. Therefore the present investigation ad-
dresses the question: do the granule cells of ferret ol-
factory bulb go through a specific process of morpho- logical maturation.
MATERIALS AND METHODS
A total of 18 female ferrets (Mustela putorius furo
L.) aged 30, 60, 90, 120 and 150 days, bred andmain- tained in captivity, were used. The animals were an-
aesthetized with Nembutal (0.06 mg/g body weight i.p.) and tissues fixed by peffusion intracardialty with
3,2% glutaraldehyde and 2.6% paraformaldehyde in 0.09 M sodium cacodylate buffer (pH 7.35) at room temperature. The olfactory epithelium proprium (OEP) of the nasal septum and the main olfactory
bulb (MOB) were dissected free and stored in the
same fixative at 4 °C for several hours (electron mi- croscopical procedure) or even longer (light micro- scopical procedure).
Light microscopy
To quantify age-related changes in the number of olfactory knobs per unit area the following procedure was used so as to exclude regional differences in the
Correspondence: B. Rehn, Institut for Anatomie, Universit~itsklinikum Essen, Hufelandstr. 55; 4300 Essen, F.R.G.
0006-8993/86/$03.50 © 1986 Elsevier Science Publishers B.V. (Biomedical Division)

density of sensory cells per unit area of the epitheli- um. Per animal 5 small pieces of tissue (ca. 1000 x 1000gm) were dissected from different regions of the OEP and embedded in Epon using standard proce- dures. From each tissue block a series of 1 gm semi- thin sections were taken over a length of 500/zm and every 10th section was stained with Azur-II and methylene blue. From each of these series 10 sections were randomly chosen and the number of the olfacto- ry knobs per 250/zm length (i.e. a total of 2500/~m) of the epithelial surface was counted under the light mi- croscope. For counting the randomly selected slides of all series were mixed with their original code, not known to the examiner.
The following light microscopical Golgi procedure was used, as described by Fairen et a1.19. After fixa- tion, one olfactory bulb from each animal was im- mersed in a solution of 2.4% potassium dichromate in distilled water containing 0.2% osmium tetroxide for 3 days. The bulbs were subsequentely washed in a 0.75% solution of fresh silver nitrate and stored in the same solution for about 3 days in the dark at room temperature. The bulbs were then embedded in Agar, cut by hand (perpendicular to the granule cell layer) with a razor blade into slices about 100-300 pm thick which were dehydrated and mounted on slides with DePeX. A minimum of 100 granule cells completely impregnated were evaluated per olfacto- ry bulb, regardless of their location within the gran- ule cell layer. The material was examined with the light microscope (magnification x 800). For count- ing, the slides of all series were mixed with their origi- nal code, not known to the examiner.
Three criteria were taken to characterize the stage of maturation.
(1) All clearly countable granule cell bodies were classified in two groups (cells with spines/cells with- out spines) according to the number of their visible somatic spines.
(2) Spines were counted on different, randomly se- lected granule cell dendrites over a length of 50/zm. To obtain comparable information on different stages of development, counts of spines were made in 3 distinct regions of the dendrites (for orientation see Fig. 4).
(3) Photomicrographs (final magnification x 1250) were used to examine the shape of spines of the ex- ternal dendrites of granule cells. Spines lying in the
115
optical plane were measured with a magnifying lens fitted with a scale bar.
Electron microscopy After fixation samples from the rostral, middle and
caudal third of the bulbs were cut into small pieces, washed in Hanks solution and postfixed in 1% osmi- um tetroxide (ref. 15) for 3 h, washed and stained with 1% uranyl acetate in 0.2 M sodium acetate buf- fer (pH 5), dehydrated in a graded series of methanol and embedded in Epon. Semithin sections (sagittal through the MOB), 1/zm thick, were cut from each block and stained with Azur-II-methylene blue to serve as an aid to orientation when examining thin sections. Thin sections were stained with lead citrate and examined with a Philips 400 electron microscope at 60-80 kV. From each animal 3 tissue blocks (ros- tral, middle and caudal of the MOB) were examined. Photographs of the outer half of the external plexi- form layer (EPL) beneath the olfactory glomeruli and the periglomular cells were obtained (magnifica- tion x 15,000) from 5 to 10 randomly selected areas (10 x 10 pm in size) per block. Synapses were counted and classified using a magnifying lens. Dur- ing counting, the photographs of all series were mixed and their protocol numbers were covered.
Statistics All counts were carried out using standard mor-
phometric procedures. Samples were randomly se- lected and identified by a code unknown to the exam- iner. The significance of developmental differences (criteria: changes in the number of spines per 50/zm of dendrite and number of synapses per 10 x 10/tm test area in the EPL) was evaluated according to a homogeneity test of Kolmogoroff and Smirnoff as de- scribed by Sachs 46. This test applies if two indepen- dent samples are to be compared from populations with continuous or discrete distribution, but both of the same type, whether they were drawn from the same or different populations. P-values equal or less than 0.05 were considered as statistically significant.
RESULTS
Olfactory epithelium of the nasal septum The morphological composition of the OEP is simi-
lar to that of other mammals. It is composed of a thin

116
layer of small, dark and light staining basal cells in which mitotic figures can occasionally be seen, a broader medial layer of sensory cell perikarya and a small apical layer of supporting cell perikarya. Be- tween day 30 and day 60 the nasal septum continues to increase remarkably in size and the area covered by the OEP enlarges by almost 50%. In older animals there are no obvious further increases in the size of the area covered by the OEP. The growing number of olfactory knobs per micron of the epithelial sur- face between day 30 and day 60 indicates that the OEP in this phase not only enlarges rapidly in area but also has a greater density of sensory cells per unit area (Fig. 1). Counting of the olfactory knobs can be taken as a measure of the number of sensory cells. Comparing the number of olfactory knobs for the dif- ferent developmental stages investigated, it can be concluded that close to the final number of sensory cells is reached between day 60 and day 90 (Fig. 1).
General morphology of the main olfactory bulb The MOB of the ferret is composed of relay neu-
rones (mitral and tufted cells) and interneurones (periglomular and granule cells). The cell bodies of these neuronal elements and their processes are ar- ranged in discrete layers (Figs. 2 and 3a, b), as is typical
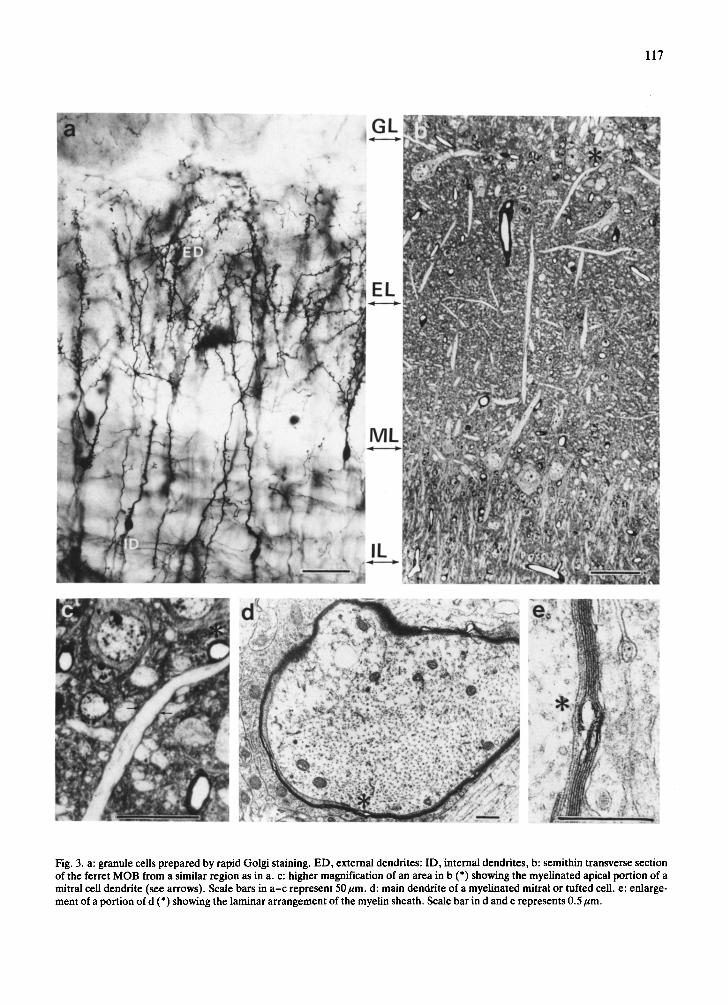
for other mammals. Exceptionally, it seems that the primary dendrites of mitral and tufted cells in ferrets are not only surrounded by loose folds of gliai mem- brane but that these folds of membrane are often packed down like a typical myelin sheath, especially
• mean number of olfacfory knobs
' ~ ' n-z2~ III / ' ° ° ~ - ,,,
-,o ~.~/
~ n= 1722
(~ ' per each age I "30 | 5 different areas
I of fhe OEPfSeptum'Nosi) age [days posfnafal]
30 60 90 120 150 I I I I J
Fig. 1. Age-related nummerical increase of olfactory knobs on the epithelial surface of the OEP. Data sampling procedure is given in Materials and Methods.
in the upper third of the dendrites near the glomeru- lar boundaries (Figs. 3d, e). However, this 'myelini- zation' of the dendrites, as first described by Pinch- ing 37 for the rhesus monkey, appears to be restricted to older animals (120 days and older).
The primary dendrites of the mitral cells and of the tufted cells extend to the olfactory glomeruli. The secondary dendrites of these cell types overlap only to a limited extent. Those of the mitral cells branch mainly within the internal half of the EPL and those of the tufted cells branch predominantly in the outer half of the EPL. The external dendrites of the gran- ule cells project to the EPL and ramify there to vari- ous degrees (Figs. 2 and 3a). The shorter internal dendrites of the granule cells branch within the gran- ule cell layer (Figs. 2 and 3a). The external dendrite (ED) of cells from deeper in the granule cell layer
. ~ Bin, sensory
FL P layer oJ
6L m. g ~ o m e r u ~
layer
EL " exferr~ plexifor M L P mifra! ( layer
/L mmm
interna~
plexifor granule
Fig. 2. Schematic drawing of the layers of neuronal elements in the MOB of the ferret as revealed by rapid Goigi impregnation.
117
Fig. 3. a: granule cells prepared by rapid Golgi staining. ED, external dendrites: ID, internal dendrites, b: semithin transverse section of the ferret MOB from a similar region as in a. c: higher magnification of an area in b (*) showing the myelinated apical portion of a mitral cell dendrite (see arrows). Scale bars in a -c represent 50/zm. d: main dendrite of a myelinated mitral or tufted cell. e: enlarge- ment of a portion of d (*) showing the laminar arrangement of the myelin sheath. Scale bar in d and e represents 0.5/zm.

118
branch predominantly in the inner half of the EPL.
Cells located closer to the band of mitral cell bodies bear ED which branch mainly in the outer half of the EPL. In contrast to the smooth surface of mitral and
tufted cell soma and dendrites, granule cells possess a rich variety of spines on their dendrites and to a cer- tain degree, also on their cell soma.
The term spine is used here for all spine-like proc- esses because the discrimination and classification
into spines, gemmules or other protrusions is not pos- sible at the light microscopical level. The spines of ED make contact with the dendrites and soma of mi- tral and tufted cells as can easily be seen in Golgi preparations. Electron microscopical investigations
Fig. 4, Schematic drawing of a typical granule cell. ED, exter- nal dendrite; GS, granule cell soma; ID, internal dendrite. The regions where the number of spines were quantified are num- bered.
demonstrate that these mitral to granule and granule to mitral cell contacts are dendrodendritic synapses, These synaptic contacts form a special type of syn-
apse, a paired reciprocal synapse (synonymous with reciprocal synapses).
In brief, the general morphological composition of the ferret MOB resembles that seen in other mam- mals.
Light microscopy of granule cells As mentioned, the proliferation in the number of
granule cells occurs mainly postnatally and therefore it is to be expected that granule cells may exhibit clear changes in morphology during postnatal devel-
opment. Using the Golgi staining method, the num- ber of spines on the dendrites and soma were there- fore counted and taken as a criterion for the stage of granule cell maturation. To check for local differ- ences in the number of the spines on the cell surface it proved useful to calculate their number in 4 different regions of the granule cell: soma, endings of ED, middle portion of ED, and basal portion of internal dendrite (ID) (Fig. 4).
Granule cell soma Granule cell bodies with no visible spines on them
and others with different numbers of spines occur ir- respective of the stage of postnatal development of the animal. By classifying the population of cells into those with and those without somatic spines and esti-
[%1 percenPage of granule cells with s o m a t i c s p i n e s
7O I
q ' I
30 celt bodies ] ~ ' ~ ' t t - . ~
age [days postnatal7 BO 60 90 120 1510 B~
, J A J II
Fig. 5. Age-related reduction in the number of granule cells possessing somatic spines expressed as a precentage 0fthe total number of granule cell bodies counted in each age group (n = 550).
f 19 [mean frequency of spines per 50pro of denaPffel
18 • • I 3,6<srt,S,7 17 • 16 / /,,5<S~>&,7
15 : 5,2<s~.5.9
e 0 S 4,S,s~,S,9
~3csli~7 age [days postnatalJ
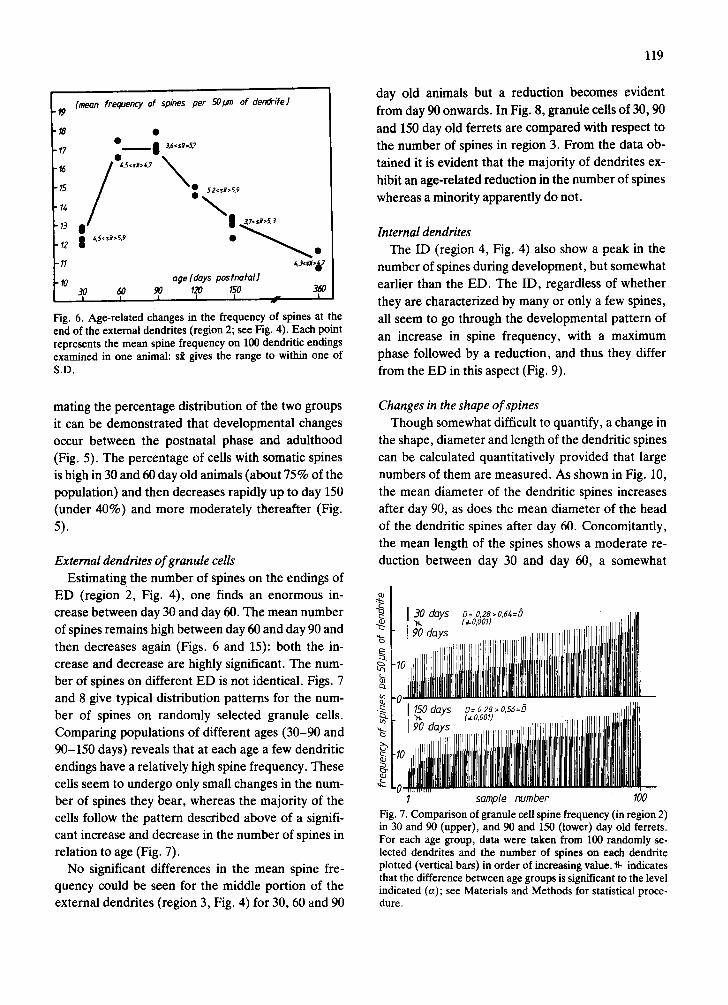
Fig. 6. Age-related changes in the frequency of spines at the end of the external dendrites (region 2; see Fig. 4). Each point represents the mean spine frequency on 100 dendritic endings examined in one animal: s~ gives the range to within one of S.D.
mating the percentage distribution of the two groups it can be demonstrated that developmental changes occur between the postnatal phase and adulthood (Fig. 5). The percentage of cells with somatic spines is high in 30 and 60 day old animals (about 75% of the population) and then decreases rapidly up to day 150 (under 40%) and more moderately thereafter (Fig. 5).
External dendrites of granule cells Estimating the number of spines on the endings of
ED (region 2, Fig. 4), one finds an enormous in- crease between day 30 and day 60. The mean number of spines remains high between day 60 and day 90 and then decreases again (Figs. 6 and 15): both the in- crease and decrease are highly significant. The num- ber of spines on different ED is not identical. Figs. 7 and 8 give typical distribution patterns for the num- ber of spines on randomly selected granule cells. Comparing populations of different ages (30-90 and 90-150 days) reveals that at each age a few dendritic endings have a relatively high spine frequency. These cells seem to undergo only small changes in the num- ber of spines they bear, whereas the majority of the cells follow the pattern described above of a signifi- cant increase and decrease in the number of spines in relation to age (Fig. 7).
No significant differences in the mean spine fre- quency could be seen for the middle portion of the external dendrites (region 3, Fig. 4) for 30, 60 and 90
119
day old animals but a reduction becomes evident from day 90 onwards. In Fig. 8, granule cells of 30, 90 and 150 day old ferrets are compared with respect to the number of spines in region 3. From the data ob- tained it is evident that the majority of dendrites ex- hibit an age-related reduction in the number of spines whereas a minority apparently do not.
Internal dendrites The ID (region 4, Fig. 4) also show a peak in the
number of spines during development, but somewhat earlier than the ED. The ID, regardless of whether they are characterized by many or only a few spines, all seem to go through the developmental pattern of an increase in spine frequency, with a maximum phase followed by a reduction, and thus they differ from the ED in this aspect (Fig. 9).
Changes in the shape of spines Though somewhat difficult to quantify, a change in
the shape, diameter and length of the dendritic spines can be calculated quantitatively provided that large numbers of them are measured. As shown in Fig. 10, the mean diameter of the dendritic spines increases after day 90, as does the mean diameter of the head of the dendritic spines after day 60. Concomitantly, the mean length of the spines shows a moderate re- duction between day 30 and day 60, a somewhat
1 sattlale number 100 Fig. 7. Comparison of granule cell spine frequency (in region 2) in 30 and 90 (upper), and 90 and 150 (lower) day old ferrets. For each age group, data were taken from 100 randomly se- lected dendrites and the number of spines on each dendrite plotted (vertical bars) in order of increasing value. ~- indicates that the difference between age groups is significant to the level indicated (a); see Materials and Methods for statistical proce- dure.
'0
0
ca.
days
120
0= 0,28< 0,07= 0 (a o ~ o l i
1 samole number 100
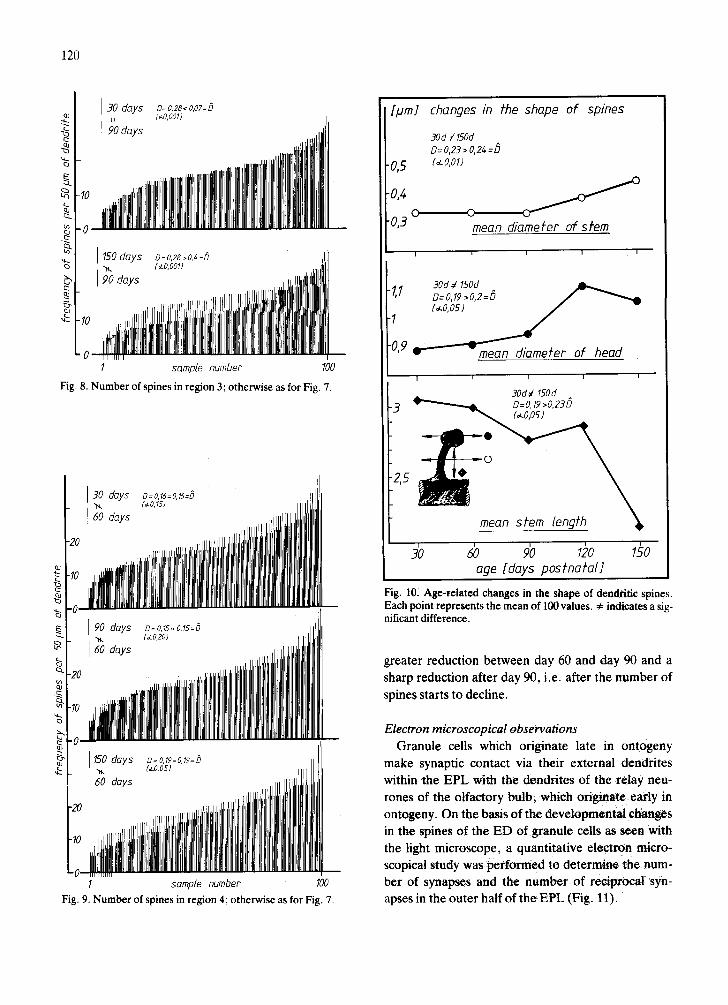
Fig. 8. Number of spines in region 3; otherwise as for Fig. 7.
1 sample number 100 Fig. 9. Number of spines in region 4; otherwise as for Fig. 7.
[pm] changes in fhe shape of spines 30d ¢ 150d D=O,23 >O,2Z,=D
• 0,5 (~.o, ol)
O' 0 "u,J mean diamefer of sfem
-1,1 30d ¢ 1SOd D= 0.19,0,2=D
"1 (,LO05) J
.0,9 ~ r "mean die~nefer of head
I i i I !
30d~l 150d o=o, ~ >o,2a5 -3 . ~
Dan sfem lengfh
age [days postnatal] I I I I i l t i i
Fig. 10. Age,related changes in the shape of dendritic spines. Each point represents the mean of 100 values. 4= indicates a sig- nificant difference.
greater reduction between day 60 and day 90 and a sharp reduction after day 90, i.e. after the number of
spines starts to decline.
Electron microscopical observations
Granule cells which originate late in ontogeny
make synaptic contact via their external dendrites within the EPL with the dendrites of the relay neu- tones of the olfactory bulb, which originate early in ontogeny. On the basis of the developmental changes in the spines of the E D of granule cells as seen with
the light microscope, a quantitative electron micro- scopical study was performed to determine the num- ber of synapses and the number of reciprocal syn- apses in the outer half of t h e E P L (Fig. 11).

121
Fig. 11. Electron micrograph taken from the outer half of the external plexiform layer of the MOB, as used for quantifying the number of synapses. Synapses are indicated by arrows, reciprocal synapses by asterisks (see insert). Scale bar, 1 gm in both cases.
The number of synapses in the outer half of the EPL increases between day 30 and day 90 by about 40%. Between day 90 and day 150 the mean number of synapses per 100/~m 2 decreases by about 25% (Fig. 12).
The estimation of the number of reciprocal syn- apses (RS) gives a relative value since only those whose synaptic thickenings lay perpendicular to the
plane of the section can be identified. Despite this de- ficiency in morphometric method, the data indicate that a moderate increase in the number of RS occurs between day 30 and day 90 followed by a slight de- crease (Fig. 13). There are fewer areas without any RS in 90 day old animals than in 30 or 150 day old ani- mals, whereas areas with 2 or more RS follow the contrary pattern.

122
synoptic
e
densify
I
[ n u m b e r o f synapses per 10x1Opm fesf areas]
N
age [days posfnafal] 30 6O 9O 120 150
, I I I I , I ,
Fig. 12. Changes in the density of synapses with age. Shaded areas show the range of mean values obtained from a total of 130 sample areas.
Excluding the temporary numerical overshoot de-
velopment between day 6Oand day 90, the number of
synapses increases between day 30 and day 150 by a
factor of 1.3, while the number of RS increases by a
factor up to about 3 (Fig. 15). The relationship be-
tween the frequency of spines on the external den-
dritic endings and the increase in the number of syn-
apses, particularly the RS, indicates that the percent-
E
i
age re/afed disfribufion ofareas wifh differenf densifies of reciprocal synapses:
areas ~,#h ~ 01 . . . .
Fig. 13. Changes in the density of paired reciprocal synapses (RS) with age (n = 20 areas for each age group). Note the de- crease and increase in the number of areas containing no RS (black bars).
i i
30 dais ]
-Io ] , lllll
6o I
- 2 0
i
Fig. ]4, Comparison of changes in spine frequency (thin verti- cal bars) and density of reciprocal synapses (thick vertical bars) with age. Spine frequencies are arranged in order of increasing value (20 randomly selected dendritic terminations were exam- ined for each age group). The density of reciprocal synapses is given as a percentage of the total number of synapses in each of the sample areas (total of 20 randomly selected areas per age group): also arranged in order of increasing value (areas con- taining no reciprocal synapses are not represented by a bar).
% [percenfage ~]spines and synapsesJ
3 20 I
/" ~ i f l v e for ~ O- reciprocal . . . . 10 / imprinting by synopses
/ preZ ~ r ,~
0 [ . . . . . . . . '1 age [days posfnafal] 30 ~ 120 150
I I I
Fig. 15. Changes in the relative frequency of spines (in region 2) and the relative density of the synapses and reciprocal syn- apses in the outer half of the external plexiform layer. The shaded area indicates the time during which imprinting on prey odours occurs (the sensitive phase).
-30

age of the latter is higher in phases of high spine fre- quency and becomes reduced when spine frequency decreases again (Figs. 14, 15).
DISCUSSION
In general, there is good evidence for assuming that different subsystems of the maturing CNS have an inherent tendency to develop a transitory (postna- tal) excess of spines and synaptic contacts 10,11,17,43,45.
During the course of further maturation, the multiple contacts between various effector nerve cells which serve a given target loose their equivalence and begin to compete against one another and may then be- come reduced in number 40. The elimination and sta- bilization of spines and synaptic contacts can be greatly influenced by interaction with the environ- ment, as for example has been detailed for the visual system9, 50-52. For the olfactory system the pattern of synaptic connectivity of the MOB of vertebrates is fairly well-known and has been described in detail in several species25,38,39,42, 49. However descriptions of
the postnatal maturation of synapses in the MOB are scarce22,23,4L There are even fewer morphological in- vestigations which help to explain the electrophysio- logicallyl8, T M and behaviourally2,4,13,16,27, 48 char-
acterised maturation of the MOB. Nevertheless, it is known that the histology2,26,31, 32,45, and thus the syn-
aptic organisation of the MOB of rodents, can be modified by sensory influences such as long-term ex- posure to odours or odour deprivation.
Imprinting of ferrets on prey odours occurs between the 60th and 90th day of postnatal life 3. During this period the rate of receptor cell proliferation slows down asymptotically (see Fig. 1) and a transient in- crease occurs in the total brain weight 4. In addition, the light microscopical data presented here show a transient increase in the number of spines during this critical period, both for the external and internal granule cell dendrites. Moreover, the electron micro- scopical data demonstrate that the formation of spines in the EPL is accompanied by the formation of synapses. Thus it would appear that the light and electron microscopical investigations describe as- pects of the same process.
It is also significant that the spine maturation ob- served here parallels, to some extent, the maturation of the related behaviour. The spines of the granule
123
cells make synaptic contacts via so-called reciprocal synapses24. 25. The mature granule to mitral cell con-
tacts and granule to tufted cell contacts are inhibito- ry 41, whereas the reverse contacts are excitatory. Other functional aspects of the reciprocal synapses in the EPL with respect to the processing of olfactory input are discussed by Shepherd 49. During devel-
opment the excitatory and inhibitory components of the reciprocal synapses mature consecutively in the mouse 22. Other developmental studies on reciprocal synapses in the EPL of the MOB show that their abundance (as a percentage of the total number of EPL synapses) changes considerably during postna- tal life. In ducks, the ratio of excitatory versus inhibi- tory synapses is about 1:1, whereas in adult ducks it has changed to 0.8 (ref. 44). This decline in the ratio of excitatory to inhibitory synapses is explained by these authors as being due to the later maturation of inhibitory synapses. Our results with ferrets are com- parable to those with mice in so far as.there is a tran- sient peak or maximum in the total number of syn- apses. The inhibitory synapses (represented here only by the reciprocal synapses) arise later than the excitatory ones (see Fig. 15). It can be concluded from our own data and from that in the literature that different regions of a given cell type may apparently have a different time course of synaptic maturation. Thus morphological and functional maturation of a neuronal network apparently does not simply involve a selective stabilisation of developing synapse 12 and the (selective) elimination of synapse 40. One may add to this the specification of the population of neu- ronal contacts, i.e. the number and location of excita- tory and inhibitory synapses with a given cell type, as a determinant of functional and morphological matu- rity. Other factors may involve the modulation of es- tablished synapses, i.e. changes in the shape of spines (see results) and concomitant with that, in the total area of synaptic contact per spine, further the matu- ration of different neurotransmitters and the trans- formation of one synaptic type into another20.
Mature granule cells have specific neuronal inputs to their external, somatic and internal regions. Dif- ferent types of granule cells have been distinguished according to their superficial, medial or deep loca- tion within the wall of the MOB, their branching pat- tern in the EPL34, 36, their content of glutamate decar- boxylase 35 and ultrastructure. As yet unpublished

124
data of ours, obta ined from mice and the ferret , also
suggest the existence of different types of granule
cells. Regardless of the type of granule cell, contra-
lateral afferents from the anterior commissure make
synaptic contact with the internal dendri te as well as
the somatic region of granule cell 21. The results pre-
sented above suggest that the control of the granule
cells per formed by the contralateral anter ior olfacto-
ry nucleus via the extrinsic fibres of the anter ior com-
missure seems to be established earl ier compared to
the inhibitory control of the relay neurones by the
granule cells. The gradual matura t ion and the differ-
ent t ime course of matura t ion of the different syn-
apses on the granule cells suggests moreover that the
influence of the granule cells on the total electrical
activity of the olfactory bulb must change during
postnatal maturat ion. In mice it has been shown in-
deed 1s,47 that quali tative changes in the matura t ion of
the electrophysiological responses in the olfactory
bulb occur duri.ng the postnatal per iod, al though this
cannot as yet be l inked to a specific cell type.
In conclusion the data presented here indicate that
the maturat ion of spines and synapses on granule
cells of the ferret MOB differ with respect to their lo-
cation (ED, ID and cell body) and t ime course (tran-
sient overshoo t and continuous decline respectively).
Thus the functional specificity of the granule cells in
the MOB of ferrets in terms of its ' input ' and 'ou tpu t '
and in terms of its 'exci tat ion ' and ' inhibi ta t ion ' need
not be reached at the same time in all its parts. It is
obvious that imprinting to prey odours occurs when
the number of spines on the ED of granule cells and
synapses in the EPL reach a maximum. At the same
time most dendrodendr i t ic connections of the gran-
ule cells mediat ing the inhibition of the mitral cells
are made. This is considered to be crucial for olfacto-
ry processing in the adult MOB.
ACKNOWLEDGEMENTS
The authors thank Dr. Militzer, Mrs. K/istner and
Mr. Waldmann for taking care of the animals and for
their assistance with the perfusion of them. We are
further indebted to Mrs. Kremer and Mrs. Busch for
careful technical assistance, to Mrs. Mlynek for pho-
tographic assistance, Mr. Rauwolf for his help in the
use of computer programs and Drs. Whit t le and
Gemmel for fruitful discussions and help with the
translat ion of the manuscript . Suppor ted by
Deutsche Forschungsgemeinschaft (Ap 14/8-6; Br
358/5-2).
REFERENCES
1 Altman, J., Autoradiographic and histological studies of postnatal neurogenisis. IV. Cell proliferation and migration in the anterior forebrain, with special reference to persist- ing neurogenesis in the olfactory bulb, J. Comp. Neurol., 137 (1969) 433-458.
2 Alberts, J.R., Ontogeny of olfaction: issues and perspec- tives from contemporary research. In W. Breipohl (Ed.), OIfaction and Endocrine Regulation, IRL Press, London, 1982, pp. 161-172.
3 Apfelbach, R., A sensitive phase for the development of ol- factory preference in ferrets (Mustela putorius f. furo L.), Z. Siiugetierkd, 43 (1978) 289-295.
4 Apfelbach, R. and Fotiadis, S., Zur postnatalen Hirnge- wichtsentwicklung des Frettchens (Mustela putorius f. furo L.), Verh. Dtsch. Zool, Ges. (1980) 325.
5 Apfelbach, R. and Rehn, B., Nahrungspr~igung und En- twicklung des Riechsystems beim Frettchen, Verb. Dtsch. Zool. Ges., (1983).
6 Balazs, R., Influence of metabolic factors on brain devel- opment, Br. Med. Bull., 30 (1974) 126-134.
7 Barlow, H.B. and Pettigrew, J.D., Lack in specificity of neurons in the visual cortex of young kittens, J. Physiol. (London), 218 (1971) 98-100.
8 Bayer, S.A., Quantitative [3H]thymidine-radiographic studies in the rat olfactory bulb, Exp. Brain Res.. 50 (1983) 329-340.
9 Blakemore. C. and Van Sluyters. R.C., Innate and envir- onmental factors in the development of the kittens visual cortex. J. Physiol. CLondon), 248 {1975) 663-716.
10 Boothe. R.G., Greenough, W.T.. Lund, J.S. and Wrege, K., A quantitative investigation of spine and dendrite de- velopment of neurons in visual cortex (area 17) of Macaca nemestrina monkeys, J. Comp. Neurol.. 186 (1979) 473-490.
11 Brunjes, P.C., Schwark, H.D. and Greenough, W.T., Ol- factory granule cell development in normal and hyper- thyroid rats, Dev. Brain Res., 5 (1982) 149-159.
12 Changeux. J-P. and Danchin. A.. Selective stabilisation of developing synapses as a mechanism for the specification of neuronal networks. Nature (London), 264 (1976) 23-30.
13 Cornwell-Jones, C.A.. Olfactory sensitive periods in albino rats and golden hamsters. J. Comp. Physiol. Psychol.. 93 (1979) 668-676.
14 Cragg, B.G.. The effects of vision and dark-rearing on the size and density of synapses in the lateral geniculate nucleus measured by electron microscopy, Brain Research, 13 (1969) 53-67.
15 Dalton, A.T., A chrome-osmium fixative for electron mi-

croscopy, Anat. Rec., 121 (1955) 281. 16 Devor, M. and Schneider, G.E., Attraction to home-cage
odors in hamster pups: specifity and changes with age, Be- hay. Biol., 10 (1974) 211-221.
17 Duffy, C.J. and Rakic, P., Differentation of granule cell dendrites in the dentate gyrus of the rhesus monkey: a quantitative Golgi study, J. Comp. Neurol., 214 (1983) 224-237.
18 Eckert, M. and Schmidt, U., The influence of permanent odor stimuli on the postnatal development of neural activity in the olfactory bulb of laboratory mice, Develop, Brain Res., submitted.
19 Fair6n, A., Peters, A. and Saldanha, J., A new procedure for examining Golgi impregnated neurons by light and elec- tron microscopy, J. Neurocytol., 6 (1977) 311-337.
20 Giildner, F.H., Ingham, C.A., Plasticity in synaptic apposi- tions of optic nerve afferents under different lighting condi- tions, Neurosci. Lett., 14 (1979) 235-240.
21 Heimer, L., Synaptic distibution of centripetal and centri- fugal nerve fibres in the olfactory system of the rat. An ex- perimental anatomical study, J. Anat., 103 (1968) 413-432.
22 Hinds, J.W. and Hinds, P.L., Synapse formation in the mouse olfactory bulb. 1. Quantitative studies, J. Comp. Neurol., 169 (1976) 15-40.
23 Hinds, J.W. and McNelly, N.A., Aging in the rat olfactory system: correlation of changes in the olfactory epithelium and olfactory bulb, J. Comp. Neurol., 203 (1981) 441-453.
24 Hirata, Y., Some observations on the fine structure of the synapses in the olfactory bulb of the mouse, with particular reference to atypical synaptic configurations, Arch. Histol. Jpn, 24 (1964) 293-302.
25 Jackowski, A., Parnavelas, J.G. and Lieberman, A.R., The reciprocal synapse in the external plexiform layer of the mammalian olfactory bulb, Brain Research, 159 (1978) 17-28.
26 Laing, G.D. and Panhuber, H., Neural and behavioral changes in rats following continuous exposure to an odor, J. Comp. Physiol., 124 (1978)259-265.
27 Leon, M., Coopersmith, R., Ulibarri, C., Porter, R.H. and Powers, B., Development of olfactory bulb organization in precocial and altricial rodents, Dev. Brain Res., 12 (1984) 45-53.
28 Mair, R.C., Gellman, R.L. and Gesteland, R.C., Postnatal proliferation and maturation of olfactory bulb neurons in the rat, Neuroscience, 7 (1982) 3105-3116.
29 Mair, R.C. and Gesteland, R.C., Response properties of mitral cells in the olfactory bulb of neonatal rat, Neurosci- ence, 7 (1982) 3117-3125.
30 Math, F. and Davrainville, J.L., Electrophysiological study on the postnatal development of mitral cell activity in the rat olfactory bulb. Influence of undernutrition, Brain Re- search, 194 (1980) 223-227.
31 Meisami, E., Effects of olfactory deprivation on postnatal growth of the rat olfactory bulb utilizing a new method for production of neonatal unilateral anosmia, Brain Research, 107 (1976) 437-444.
32 Meisami, E. and Mousavi, R., Lasting effects of early olfac- tory deprivation on the growth, DNA, RNA and protein content and Na-K-ATPase and AChE activity of the rat ol- factory bulb, Develop. Brain Res., 2 (1982) 217-229.
33 Mendoza, A.S., Rehn, B. and Apfelbach, R., Postnatale Synapsenausbildung im Bulbus olfactorius von Frettchen.
125
Befunde an der externen plexiformen Schickt, Verh. Anat. Ges., in press.
34 Mori, K., Kishi, K. and Ojima, H., Distribution of den- drites of mitral, displaced mitral, tufted and granule cells in the rabbit olfactory bulb, J. Comp. Neurol., 219 (1983) 339-355.
35 Mugnaini, E., Wouterlood, F.G., Dahl, A.L. and Oertel, W.H., Immunocytochemical identification of GABAergic neurons in the main olfactory bulb of the rat, Arch. Ital. Biol. 122 (1984) 83-113.
36 Orona, E., Scott, J.W. and Rainer, E.C., Different granule cell populations innervate superficial and deep regions of the external plexiform layer in the rat olfactory bulb, J. Comp. Neurol., 217 (1983) 227-237.
37 Pinching, A.J., Myelinated dendritic segments in the mon- key olfactory bulb, Brain Research, 29 (1971) 133-138.
38 Price, J.L. and Powell, T.P.S., The synaptology of the granule cells of the olfactory bulb, J. Cell Sci., 7 (1970) 125-155.
39 Price, J.L. and Powell, T.P.S., An electron microscopic study of the termination of the afferent fibres to the olfacto- ry bulb from the cerebral hemisphere, J. Cell Sci., 7 (1970) 157-187.
40 Purves, D. and Lichtman, J.W., Elimination of synapses in the developing nervous system, Science, 210 (1980) 153-157.
41 Rail, W., Shepherd, G.M., Reese, T.S. and Brightman, M.W., Dendrodendritic synaptic pathway for inhibition in the olfactory bulb, Exp. Neurol., 14 (1966) 44-56.
42 Ramon y Cajal, S., Histologie du Systeme Nerveux de l'Homme et des Vertebres, Maloine, Paris, 1911.
43 Rausch, G. and Scheich, H., Dendritic spine loss and en- largement during maturation of the speech control system in the mynah bird (Gracula religiosa), Neurosci. Lett., 29 (1982) 129-133.
44 Rebiere, A. and Dainat, J., Quantitative study of synapse formation in the duck olfactory bulb, J. Comp. Neurol., 203 (1981) 103-120.
45 Rehn, B., Breipohl, W., Schmidt, U., Mendoza, A.S. and Apfelbach, R.A., Effect of olfactory blockade on the post- natal development of the olfactory bulb granule cells in mouse: a Golgi study, Chem. Senses Flavor, 8 (1984) 262.
46 Sachs, L., Applied Statistics, Handbook of Techniques, Springer, New York, 1982, pp. 291-293.
47 Schiller, H.J., Elektrophysiologische Untersuchungen zur Ontogenetischen Entwicklung der Olfaktorischen Sekundiir- neurone bei der Labormaus, Ph. D. Thesis, University of Bonn, 1984.
48 Schleidt, M. and Hold, B., Human odor and identity. In W. Breipohl (Ed.), Olfaction and Endocrine Regulation, IRL Press, London, 1982, pp. 181-194.
49 Shepherd, G.M., Synaptic organization of the mammalian olfactory bulb, Physiol. Rev., 52 (1972) 864-917.
50 Valverde, F., Apical dendritic spines of the visual cortex and light deprivation in the mouse, Exp. Brain Res., 3 (1967) 337-352.
51 Wiesel, T.N. and Hubel, D.H., Effects of visual depriva- tion on morphology and physiology of cells in the cat's later- al geniculate body, J. NeurophysioL, 26 (1963) 978-993.
52 Winfield, D.A., The postnatal development of synapses in the different laminae of the visual cortex in the normal kit- ten and in kittens with eyelid suture, Dev. Brain Res., 9 (1983) 155-169.




















