
Self-assembled monolayer mechanism for corrosion inhibition of iron by imidazolines

Biosensors and Bioelectronics 20 (2005) 2053–2060
Langmuir monolayer approaches to protein recognitionthrough molecular imprinting
Xuezhong Dua,1, Vladimir Hladya, David Brittb,∗a Department of Bioengineering, University of Utah, Salt Lake City, UT 84112, USA
b Department of Biological Engineering, Utah State University, Logan, UT 84322, USA
Received 1 June 2004; received in revised form 21 August 2004; accepted 24 August 2004Available online 12 October 2004
Abstract
Surface plasmon resonance (SPR) spectroscopy and atomic force microscopy (AFM) have been employed to investigate ferritin adsorption tobinary surfactant monolayers of cationic dioctadecyldimethylammonium bromide (DOMA) and non-ionic methyl stearate (SME). Surfactantm , whichu Successivea ent, bouu c supportd SME ast adsorbingp the presenceo©
K
1
kgtnLpt1n
E
ap-l andow-
s rec-getheas-nal
hereno-
fluidbicrac-ingrfaceopy
0d
olar ratios, miscibility, and lateral mobility were controlled to define the number, size, and distribution of “binding sites” for ferritinnder the low ionic strength conditions investigated, adsorbed to the monolayers predominantly through electrostatic interactions.dsorption/desorption cycles revealed that fluid monolayers, capable of laterally restructuring during the initial protein adsorption evndp to 60% more ferritin (dependent on SME:DOMA ratios) as compared to monolayers that were immobilized on a hydrophobiuring this first adsorption step. The enhanced binding of ferritin to fluid monolayers was accentuated in films having non-ionic
he principal component. These findings support the premise that the surfactants reorganize to form favorable interactions with anrotein, leading to protein specific charge patterns, or templates, in the films. Template assessment, however, was complicated byf an irreversibly bound protein fraction, which AFM revealed to be locally ordered protein clusters.2004 Elsevier B.V. All rights reserved.
eywords:Molecular template; Mixed monolayer; Miscibility; Ferritin; SPR; AFM
. Introduction
In nature, molecular recognition occurs through “lock andey” interactions where binding sites on receptor moleculesenerally consist of a pocket that is complementary to the
arget in terms of size and chemistry. The molecular mecha-isms of specific binding and recognition were postulated byinus Pauling to occur through an induced fit, where highlylastic “generic” antibodies assumed a specific (complemen-
ary) shape in response to the presence of an antigen (Pauling,940). Although incorrect, this mechanism has inspired aovel approach of designing molecular function in synthetic
∗ Corresponding author. Tel.: +1 435 797 2158; fax: +1 435 797 1248.E-mail address:[email protected] (D. Britt).
1 Present address: Department of Chemistry, Nanjing University, Minist.duc., Key Lab Mesoscop Chem., Nanjing 210093, PR China.
materials through nanoscale imprinting, with promisingplications in sensing, separation, processing of chemicabiological molecules, and catalyzing reactions. To date, hever, imprinted materials have generally had poor succesognizing with high specificity complex and relatively larbiological materials such as proteins that first inspiredmolecular imprinting methods. To overcome limitationssociated with bulk imprint materials, a quasi two-dimensioapproach to molecular imprinting is presented here wprotein adsorption to an insoluble binary surfactant molayer imparts a complementary charge pattern in themonolayer, which is “frozen” upon transfer to a hydrophosupport. The planar imprint configuration facilitates chaterization of protein desorption and rebinding kinetics ussurface plasmon resonance (SPR), while the protein sudistribution is readily assessed with atomic force microsc(AFM).
956-5663/$ – see front matter © 2004 Elsevier B.V. All rights reserved.oi:10.1016/j.bios.2004.08.044

2054 X. Du et al. / Biosensors and Bioelectronics 20 (2005) 2053–2060
Surface imprinting of a fluid membrane is a biomimeticapproach that does not involve a pre-binding of recognitionelements (functionalized monomers) to the target as is tradi-tionally done in molecularly imprinted polymers; rather theprotein must “select” the recognition elements presented in afluid monolayer floating at the air/water interface of a Lang-muir trough. This approach has been previously applied intemplate-mediated crystal nucleation and biomineralization.(Gidalevitz et al., 1994; Litvin et al., 1995) as well as in two-dimensional crystallization of proteins and macromolecules(Crowther and Klug, 1975; Ugiriz and Kornberg, 1983). Thereverse action, i.e. the templating of a lipid monolayer by themolecules bound in an adlayer, has been less explored andgenerally only for small molecules or chemicals (Martın etal., 1996; Last et al., 2001).
Although an abundance of literature exists on the inter-actions of proteins with Langmuir monolayers, liposomes,and membranes, the nanoscale monolayer restructuring thatmay occur in the presence of a protein adlayer has notbeen successfully applied for protein imprinting. Early workby Mayer and Nelsestuen demonstrated calcium and pro-thrombin induced phase separation in mixed phosphoser-ine:phosphocholine vesicles (Mayer and Nelsestuen, 1981).While the Arnold group, drawing from the principles of metalaffinity chromatography, has shown that histidine residueso pro-t eadg al.,1 rso tion-a tedt om-p dings intot ntt tionr iteso e. Tod fluidm pro-t
tingf nicm ion( idueso men-t oughi ichi nitort ervea thati e ins m-p rfacep lat-i s in-
creases as the charge density in the monolayer films is dilutedby increasing the ratio of non-ionic to cationic surfactants.Thus, a monolayer composed entirely of cationic surfactantsis less favorable to adsorption of a negatively charged pro-tein than a mixed surfactant monolayer containing neutralsurfactants. However, protein–protein interactions interplaywith protein–surfactant interactions in defining the adsorp-tion, desorption, and readsorption trends, thus complicatingassessment of templating.
2. Materials and methods
2.1. Langmuir monolayers and protein
Dioctadecyldimethylammonium bromide (DOMA) (99%Tech Grade, TCI) and methyl stearate (SME) (99% GC,Fluka) were prepared as 1 mM solutions in chloroform (spec-trograde, Merck) and stored at 4◦C prior to use. SME andDOMA mixtures were prepared volumetrically from thestock solutions, leading to high miscibility when spread asmonolayers (Goodman et al., 2004; Britt et al., 2000). Ascontrols, poorly miscible SME:DOMA were prepared by se-quentially spreading the stock solutions, thus restricting allmixing to the plane of the air/water interface and creatinga y ex-c hesefi inter-a leso n att di-m sev unto ce.T -t theS Sur-f l thed sf truc-t pho-bS -t vea -j lowedt ub-p iluteH hichh n ex-c int entsw antm (pH52
n the protein coat can be used to target and orienteins to lipid monolayers containing metal-chelating hroups (Pack et al., 1997; Ng et al., 1995; Maloney et996; Mallik et al., 1994). Similarly, Sasaki and coworkebserved a redistribution (dispersion) of mannose funclized lipids upon binding of a tetravalent lectin, attribu
o possible steric interactions between the protein–lipid clexes and the fixed spatial distribution of mannose binites on the protein, as well as possible protein insertionhe membrane (Bondurant et al., 2003). These studies poio the potential of inducing a fundamental structure/funcelationship between the distribution of fixed binding sr residues on a protein surface and a fluid membranate, however, protein induced in-plane templating ofonolayers leading to significantly enhanced (specific)
ein readsorption has not been achieved.Here, we demonstrate a proof of concept by investiga
erritin readsorption on two-component cationic/non-ioonolayers that were initially fluid during the first adsorpt
templating) step. The excess of negatively charged resn the protein surface are anticipated to induce a comple
ary charge pattern in the monolayer that is preserved thrmmobilization of the film on a hydrophobic support, whs an alkane–thiol modified gold SPR sensor used to mohe protein binding kinetics. With this approach, we obsmodest (∼15%) enhanced rate of protein readsorption
s accompanied by a rather large (up to 60%) increasurface capacity of the imprinted film for the protein coared to control monolayers immobilized on the SPR surior to the first adsorption step. In support of the temp
ng premise, the enhanced rebinding to templated film
surface with heterogeneity of a size-scale that greatleeded the size of the protein. Protein insertion into tlms was used as a simple method to assess proteinctions with films having dramatically differing size-scaf heterogeneity. For SPR analysis of protein adsorptio
he air/water interface, miniature PTFE troughs havingensions of 4 cm× 2 cm × 0.5 cm and a total subpha
olume of 5 mL were custom built to minimize the amof protein needed and facilitate diffusion to the interfahe trough walls were undercut by 30◦ to eliminate forma
ion of meniscus, presenting a planar interface on whichPR sensor was lowered to measure protein kinetics.
actants were slowly spread using a microsyringe untiesired surface pressure of 30 mN m−1 was achieved, thu
orming a condensed monolayer capable of laterally resuring, yet resistant to protein penetration into the hydroic tail region. Horse spleen ferritin (102 mg mL−1, Type I,igma) was diluted to 1 mg mL−1 in double-distilled wa
er prior to injection of 5�L into the subphase to achiefinal concentration of 1.0�g mL−1. The protein was in
ected through ports connected to the subphase and alo adsorb for up to 2 h, followed by exchange of the shase with water having a pH adjusted to 2.7–2.8 with dCl. This procedure reversed the charge on ferritin, was an isoelectric point of 4.5. The subphase was thehanged for pH 5.5 water prior to reintroducing proteinhe subphase for readsorption experiments. All experimere carried out at 20◦C. The subphase on which surfactonolayers were prepared was double-distilled water.5) having an initial surface tension near 72 dyne cm−1 at0◦C.

X. Du et al. / Biosensors and Bioelectronics 20 (2005) 2053–2060 2055
2.2. Monolayer transfer and AFM
A KSV trough with dipping well (KSV Instruments Ltd.)was used to prepare monolayers for AFM characterization ofthe native films as well as to assess protein distribution on thefluid and immobile monolayers. For these experiments, thesurfactants were spread to low surface concentrations fol-lowed by symmetric compression at a rate of 2 mm min−1
until the surface pressure of 30 mN m−1 was attained. Thesurface pressure versus molecular area isotherms for SME,DOMA, and the mixtures have been previously published(Goodman et al., 2004; Britt et al., 2000), and are not pre-sented here. For protein adsorption to fluid monolayers, 1 mLof ferritin solution (1.0 mg mL−1) was injected into the wa-ter subphase using a syringe inserted underneath the barriersto give a final concentration of 1.0�g mL−1. The ferritin ad-sorption to the monolayer was allowed to proceed for 2 h, fol-lowed by film transfer onto an octadecyltrichlorosilane (OTS)modified silicon wafer by a modified Langmuir–Schaeffermethod: the hydrophobic wafer was horizontally pressed intothe subphase through the interface covered with the mono-layer and placed into a small beaker present in the subphase(Buijs et al., 1998). The remaining surfactants were sweptfrom the interface, and the beaker containing the wafer andwater was removed from the trough. The wafer was main-t ing.F vedb asesfr ofm inia-t nsferm
2
ouss c.).P on-s tinfi ndera tor sketw ayert
2
-s sorp-t ,2 firstm thiol( ca-t orw lled
Fig. 1. Schematic of protein induced template formation in fluid films vs.an otherwise identical immobilized control film. The gold surface of theSpreetaTM SPR sensor is hydrophobically modified with octadecanethiol soas to favorably interact with the hydrophobic tails of the surfactants.
water (refractive index,n = 1.33) and a SPR baseline wasobtained in water. The SPR sensor was then dried and po-sitioned above the monolayer free air/water interface. Oncethe monolayer was spread to the desired surface pressure andequilibrated, the sensor was slowly lowered into contact withthe monolayer and brought to a depth of ca. 2 mm using a mi-cromanipulator. This procedure was equivalent to a horizon-tal monolayer transfer to a solid substrate, but without everremoving the substrate (i.e. the sensor) carrying the trans-ferred monolayer from the trough. This method is indicatedin Fig. 1 as applied to templating (monolayer fluid duringinitial adsorption step) and control (monolayer immobilizedduring initial adsorption) experiments. Upon contact of theSPR sensor with the monolayer, a step increase of the SPRsignal was recorded and a new SPR baseline was establishedfor a period of 10 min to ensure the integrity of the trans-ferred monolayer prior to injecting the protein (control) orexchanging the subphase to induce protein desorption (tem-plate). After each experiment, the SPR sensor was cleaned
ained hydrated during further processing and AFM imagerritin adsorption to laterally immobile films was achiey transferring the monolayers on protein free subph
ollowed by exchange of the water with a 1.0�g mL−1 fer-itin solution through the AFM flow cell. AFM analysisonolayers and proteins prepared on the custom built m
ure troughs was also performed using the horizontal traethod for comparison.
.3. AFM
AFM images were performed in contact mode in aqueolution with a Nanoscope II SPM (Digital Instruments Inark Scientific Instruments cantilevers with a low force ctant of 0.01 N m−1 were used to avoid disrupting the ferrilms as much as possible. Cantilevers were irradiated uhigh-intensity UV lamp for 15 min just prior to imaging
emove organic contaminants from the tip. A silicon gaas used to seal the AFM flow cell to the hydrated monol
ransferred on the OTS-silicon.
.4. In situ SPR at the air/water interface
An integrated optics SPR sensor (SpreetaTM, Texas Intruments) was used to quantitatively measure protein adion to each monolayer as previously described (Jogikalmath003). The sensing Au surface of the SPR probe wasade hydrophobic by immersion in a 1 mM octadecane
ODT) solution in ethanol for 10 min. This surface modifiion resulted in a water contact angle of 107◦. The SPR sensas then pre-initialized in air, calibrated in double-disti

2056 X. Du et al. / Biosensors and Bioelectronics 20 (2005) 2053–2060
using oxygen plasma (Plasmod, Tegal Corp.). The sensorresponse was unaffected by repeated plasma cleaning andODT surface modification procedures. The relative changesin refractive index near the sensor surface due to changes insubphase refractive index were used to calculate the effec-tive thickness of the adsorbed protein layer from which theprotein surface concentration was determined as previouslydescribed byJung et al. (1998).
3. Results and discussion
3.1. Surfactant miscibility and protein adsorption
Tensiometer measurements of ferritin insertion into pureand binary films of SME and DOMA prepared at initial sur-face pressures of 11 mN m−1 (amenable to protein insertion)are presented inFig. 2(a). Minimal ferritin insertion into theneutral SME monolayers is contrasted with significant bind-ing to the cationic DOMA monolayer, as anticipated for anadsorption mechanism driven primarily by electrostatic inter-actions. This behavior is also observed at high surface pres-sures (30 mN m−1; Fig. 2(b)) where ferritin cannot insert intothe monolayers, and adsorption must be measured using SPR.Under these conditions, ferritin adsorption to SME is com-p MEa fp -v areda le ofm n thed vingt thefi ead-i ts inm ur-r .,2 im-p twop cribedb
Γ
wa purefi anti di-t of these Am lm,t ptiont f thea ms.
Fig. 2. Influence of monolayer composition and packing density on proteinadsorption. (a) Ferritin insertion into single and two-component monolayerfilms of cationic DOMA and non-ionic SME, prepared at initial surfacepressures of∼11 mN m−1; final ferritin concentration, 10�g mL−1. Theelectrostatic nature of the adsorption process is evident by the high affinityfor the pure cationic DOMA monolayer, with minimal adsorption to the purenon-ionic SME. Two component monolayers of SME and DOMA (2:1 mo-lar ratios) display protein adsorption behavior dependent on the size-scaleof the surfactant miscibility. The film prepared by sequentially spreadingthe two surfactants at the air/water interface is phase separated and the ad-sorption behavior to this film is a simple average of the adsorption to thepure components as predicted by Eq.(1). An adsorption that exceeds thatpredicted by Eq.(1) is observed for the film prepared from the pre-mixedSME:DOMA solution. See text for details. (b) SPR sensorgrams (angle shiftvs. time) of ferritin binding to pure SME and DOMA monolayers. Surfacepressure, 30 mN m−1; final ferritin concentration, 1�g mL−1 in water sub-phase, pH 5.5. Protein adsorption is restricted to the surfactant head groupsas insertion into the films is inhibited under these conditions.
The deviation from ideal adsorption is greatest for the two-component film prepared from a pre-mixed solution, and is at-tributed to a matching of size-scales of heterogeneity betweensorbent surface and adsorbing particle. This behavior may befurther enhanced if the surface is somewhat plastic, as in thecase of a floating surfactant monolayer. Template inductionis then anticipated, and has been demonstrated to be energet-ically favorable through thermodynamic models (May et al.,2000; Harries et al., 2002). For the present case of ferritin ina salt free medium at pH 5.5, the ferritin coat has more neg-atively charged residues than positively charged ones, andadsorption to cationic monolayers is favored over adsorp-tion to anionic or neutral monolayers at both low (Fig. 2(a))and high (Fig. 2(b)) packing densities. Nonetheless, the ba-sic protein residues cannot be ignored as the protein comes
letely diminished, suggesting ferritin interaction with St lower surface pressures (Fig. 2(a)) may be the result orotein affinity for the hydrophobic tails.Fig. 2(a) also reeals that ferritin adsorption to binary monolayers prept high and low miscibility is dependent on the size-scaonolayer heterogeneity. This phenomenon is evident iiminished adsorption to the sequentially spread film ha
he same SME:DOMA content (but not distribution) aslm prepared from a pre-mixed solution. Sequential sprng of the surfactants at the air/water interface resul
icron-sized domains comprised primarily of DOMA sounded by a predominantly SME phase (Goodman et al004). Thus, ferritin binding to this film approaches a sle average of the individual binding processes to theure components. Such an adsorption process is desy:
12 = χ1(Γ1) + χ2(Γ2) (1)
hereΓ 12 is the adsorption to the two-component film,Γ 1ndΓ 2 are the respective adsorption processes to the twolms, andχ1 andχ2 are the mole fractions of each surfactn the two-component film. Departure from this strictly adive adsorption processes is anticipated as the size-scaleurface heterogeneity approaches that of the protein (Hladyt al., 2000), which is the case here for the SME:DOMonolayer prepared from a pre-mixed solution. For this fi
he surfactants are intimately mixed, and protein adsoro the film exceeds that which is predicted by the sum odsorption events on the respective pure component fil

X. Du et al. / Biosensors and Bioelectronics 20 (2005) 2053–2060 2057
Table 1Ferritin equilibrium surface concentrations (from SPR) and surface coverages (from AFM) on indicated SME:DOMA mixtures
SME:DOMA Surface concentrationa (�g cm−2), surface coverageb (%)
Initial binding Rebinding I Rebinding II
Fluid film Control Fluid on 1st Control Fluid on 1st Control
2:1 0.478 (57.4) 0.534 (64.1) 0.445 (53.4) 0.492 (59.0) 0.408 (49.0) 0.485 (58.2)3:1 0.526 (63.1) 0.479 (57.5) 0.379 (45.5) 0.403 (48.4) 0.337 (40.4) 0.347 (41.6)6:1 0.535 (64.2) 0.439 (52.7) 0.417 (50.0) 0.336 (40.3) 0.357 (42.8) 0.282 (33.8)9:1 0.543 (65.2) 0.372 (44.6) 0.426 (51.1) 0.266 (31.9) 0.381 (45.7) 0.229 (27.5)
a Determined using Jung’s method (Jung et al., 1998).b Estimated from a 0.5�m × 0.5�m scan area and on the assumption of a close-packed arrangement of ferritin particles having diameter = 12.5 nm. A
broadening effect of the AFM tip may lead to overestimation of the actual surface concentrations.
into close proximity with the monolayer, where these posi-tively charged residues will repel the cationic DOMA headgroups just as the acidic residues will attract DOMA. Thus,an in-plane restructuring, or templating, of the monolayer isenvisioned to occur through protein adsorption to the binarymonolayer. For this to occur, the entropic penalty associatedwith protein induced de-mixing of the surfactants must beless than the favorable reduction in interfacial free energythrough physical bond formation between protein and headgroups. This hypothesis was indirectly tested as describednext through protein adsorption, desorption, and readsorp-tion to fluid versus immobilized monolayers.
Fmfec
3.2. Protein adsorption: fluid versus immobilizedmonolayers
For template induction in monolayer films, protein pen-etration into the film is undesirable as this may resultin denaturation. Thus, the monolayers must be preparedat surface pressures of 30 mN m−1, where the surfactantsstill retain lateral mobility, yet the protein is unable to in-sert into the film. The disadvantage is that simple ten-siometer measurements do not report on protein adsorp-tion, thus an optical technique such as SPR is bettersuited.
ig. 3. SPR adsorption/desorption sensorgrams (refractive index vs. time) oonolayers prepared at several molar ratios (SME:DOMA): (a) 2:1, (b) 3:1,
or weak HCl (pH 2.7–2.8) to induce a limited desorption (complete desorxchanging the subphase for pH 5.5 distilled water followed by injection of feroncentration in the subphase was 1�g mL−1.
f ferritin interaction with fluid (dashed line) and immobilized control(solid line)(c) 6:1, (d) 9:1. Following a plateau in adsorption, the subphase was exchangedption could not be achieved). Readsorption to the monolayer was initiated byritin. In all experiments, the surface pressure was 30 mN m−1 and the final ferritin

2058 X. Du et al. / Biosensors and Bioelectronics 20 (2005) 2053–2060
Fig. 4. Typical AFM topography images of ferritin equilibrium distribution on a SME:DOMA mixed monolayer, molar ratio 2:1; surface pressure, 30 mN m−1;final ferritin concentration, 1�g mL−1. Monolayer fluid during initial adsorption.
SPR sensorgrams for ferritin adsorption to mixedSME:DOMA monolayers (prepared from pre-mixed solu-tion) are given inFig. 3. Several molar ratios were inves-tigated (increasing SME content from a to d inFig. 3) inan effort to determine the optimal monolayer compositionfor templating. For the initial protein adsorption on the fluidmonolayers, the binding kinetics are not attained as the SPRprobe cannot be in contact with the monolayer if it is to re-main fluid; however, the equilibrium adsorption value is at-tained by placing the SPR probe in contact with the film after1.5 h adsorption time. The surface concentration of boundferritin on the monolayers at adsorption equilibrium are nearthe jamming limit for a random sequential adsorption process(Johnson et al., 2000) indicating a dense surface coverage ofprotein that is confirmed using AFM, as shown inFig. 4. Theferritin surface concentrations calculated from SPR and AFMimages are listed inTable 1, where several trends are noted:first, a significant fraction of the protein is irreversibly boundto the films; second, the film surface capacity diminishesand the irreversibility of binding increases with each sub-sequent rebinding/desorption process; third, the adsorptionto the control (immobilized) monolayers decreases with de-creasing monolayer charge density (decreasing DOMA con-tent), whereas this trend is reversed on the fluid monolayersas illustrated inFig. 5.
ongp thep tiono nita elyt ablet n theA ayeri tion.L siblyb iteso ctiono
Fig. 5. Surface concentration of bound ferritin on SME:DOMA mixedmonolayers as a function of molar fraction of SME. Adsorption to the fluidmonolayers (not in contact with the SPR probe during the initial adsorptionstep) increases with mole fraction SME, whereas an opposite trend is ob-served for adsorption to the monolayers that were immobilized on the SPRprobe during the initial adsorption step.
coworkers, where the final surface concentration of ferritinon hydrophobic surfaces was shown to be strongly pathwaydependent (Hook et al., 1998). This was displayed through amultistep adsorption process where incremental increases inprotein surface concentration were attained by increasing thesolution ionic strength in each step (while keeping proteinbulk concentration constant); however, in this multistepadsorption process, the final surface concentration was lessthan that attained by a single-step adsorption process wherethe most concentrated ionic strength solution was usedfrom the beginning. This was termed a surface “memory”effect, which arises from multiple pathways to proteinadsorption, protein–protein interactions, and protein lateral(re)distribution. Such a surface memory effect, however,where protein–protein interactions dominate, is undesirableas it complicates assessment of an artificial recognitionelement through template induction. Nonetheless, theopposing adsorption trends for the fluid and immobilizedfilms (seeFig. 5) support the templating hypothesis.
The first two observations are attributable to strrotein–protein interactions that interplay withrotein–monolayer interactions. Although associaf the proteins in a salt-free solution that is a full pH ubove the protein isoelectric point is not favorable, it is lik
hat once adsorbed to the interface, the proteins areo assemble into the well organized arrays observed iFM images. Thus, protein–protein and protein–monol
nteractions combine to prevent complete desorpikewise, upon readsorption, the presence of the irreveround fraction of ferritin reduces the number of binding sn the surface beyond that of the area this protein fraccupies. This is similar to behavior reported by Hook and
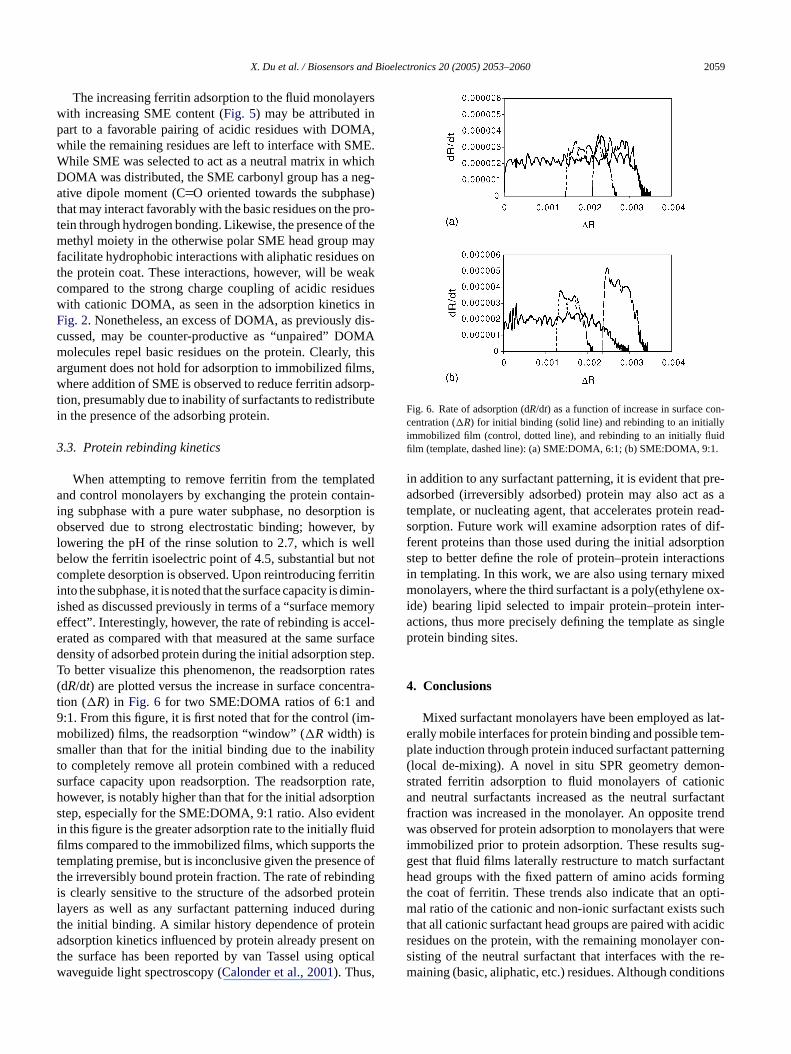
X. Du et al. / Biosensors and Bioelectronics 20 (2005) 2053–2060 2059
The increasing ferritin adsorption to the fluid monolayerswith increasing SME content (Fig. 5) may be attributed inpart to a favorable pairing of acidic residues with DOMA,while the remaining residues are left to interface with SME.While SME was selected to act as a neutral matrix in whichDOMA was distributed, the SME carbonyl group has a neg-ative dipole moment (CO oriented towards the subphase)that may interact favorably with the basic residues on the pro-tein through hydrogen bonding. Likewise, the presence of themethyl moiety in the otherwise polar SME head group mayfacilitate hydrophobic interactions with aliphatic residues onthe protein coat. These interactions, however, will be weakcompared to the strong charge coupling of acidic residueswith cationic DOMA, as seen in the adsorption kinetics inFig. 2. Nonetheless, an excess of DOMA, as previously dis-cussed, may be counter-productive as “unpaired” DOMAmolecules repel basic residues on the protein. Clearly, thisargument does not hold for adsorption to immobilized films,where addition of SME is observed to reduce ferritin adsorp-tion, presumably due to inability of surfactants to redistributein the presence of the adsorbing protein.
3.3. Protein rebinding kinetics
When attempting to remove ferritin from the templateda tain-i ion iso r, byl ellb notc rritini imin-i morye cel-e urfaced tep.T rates( ntra-t d9 im-ms lityt ceds rate,h tions enti uidfi thet ce oft ingi teinl ringt eina nt ont pticalw
Fig. 6. Rate of adsorption (dR/dt) as a function of increase in surface con-centration (�R) for initial binding (solid line) and rebinding to an initiallyimmobilized film (control, dotted line), and rebinding to an initially fluidfilm (template, dashed line): (a) SME:DOMA, 6:1; (b) SME:DOMA, 9:1.
in addition to any surfactant patterning, it is evident that pre-adsorbed (irreversibly adsorbed) protein may also act as atemplate, or nucleating agent, that accelerates protein read-sorption. Future work will examine adsorption rates of dif-ferent proteins than those used during the initial adsorptionstep to better define the role of protein–protein interactionsin templating. In this work, we are also using ternary mixedmonolayers, where the third surfactant is a poly(ethylene ox-ide) bearing lipid selected to impair protein–protein inter-actions, thus more precisely defining the template as singleprotein binding sites.
4. Conclusions
Mixed surfactant monolayers have been employed as lat-erally mobile interfaces for protein binding and possible tem-plate induction through protein induced surfactant patterning(local de-mixing). A novel in situ SPR geometry demon-strated ferritin adsorption to fluid monolayers of cationicand neutral surfactants increased as the neutral surfactantfraction was increased in the monolayer. An opposite trendwas observed for protein adsorption to monolayers that wereimmobilized prior to protein adsorption. These results sug-gest that fluid films laterally restructure to match surfactanth ingt opti-m ucht cidicr con-s re-m ions
nd control monolayers by exchanging the protein conng subphase with a pure water subphase, no desorptbserved due to strong electrostatic binding; howeve
owering the pH of the rinse solution to 2.7, which is welow the ferritin isoelectric point of 4.5, substantial butomplete desorption is observed. Upon reintroducing fento the subphase, it is noted that the surface capacity is dshed as discussed previously in terms of a “surface meffect”. Interestingly, however, the rate of rebinding is acrated as compared with that measured at the same sensity of adsorbed protein during the initial adsorption so better visualize this phenomenon, the readsorptiondR/dt) are plotted versus the increase in surface conceion (�R) in Fig. 6 for two SME:DOMA ratios of 6:1 an:1. From this figure, it is first noted that for the control (obilized) films, the readsorption “window” (�Rwidth) is
maller than that for the initial binding due to the inabio completely remove all protein combined with a reduurface capacity upon readsorption. The readsorptionowever, is notably higher than that for the initial adsorptep, especially for the SME:DOMA, 9:1 ratio. Also evidn this figure is the greater adsorption rate to the initially fllms compared to the immobilized films, which supportsemplating premise, but is inconclusive given the presenhe irreversibly bound protein fraction. The rate of rebinds clearly sensitive to the structure of the adsorbed proayers as well as any surfactant patterning induced duhe initial binding. A similar history dependence of protdsorption kinetics influenced by protein already prese
he surface has been reported by van Tassel using oaveguide light spectroscopy (Calonder et al., 2001). Thus,
ead groups with the fixed pattern of amino acids formhe coat of ferritin. These trends also indicate that anal ratio of the cationic and non-ionic surfactant exists s
hat all cationic surfactant head groups are paired with aesidues on the protein, with the remaining monolayeristing of the neutral surfactant that interfaces with theaining (basic, aliphatic, etc.) residues. Although condit

2060 X. Du et al. / Biosensors and Bioelectronics 20 (2005) 2053–2060
were selected to favor electrostatic interactions between pro-tein and monolayer and disfavor protein–protein interactions,AFM analysis revealed protein clusters that may have formedthrough surfactant mediated aggregation. Such aggregationwould be favored on the fluid monolayers, which may havecontributed to the enhanced adsorption observed on thesefilms. These results demonstrate the feasibility of protein in-duced patterning of monolayer films; however, future effortswill need to focus on tuning the monolayer properties to hin-der protein–protein aggregation leading to irreversible ad-sorption.
Acknowledgments
This study was supported by NSF (9986289) and NIH(EB000278) grants. Travel support to DWB from the UtahState University Center for Integrated Biosystems is grate-fully acknowledged. The authors thank Paul van Tassel forinsight into protein readsorption behavior and Hemang Patelfor the tensiometer measurements.
References
B 003.uponmuir
B lti-ility599.
B tionmuir
C f pro-669.
C ularAnn.
G L.,bing116,
G ritt,y-
ers using electrostatic force microscopy. Langmuir 20, 3684–3689.
Harries, D., May, S., Ben-Shaul, A., 2002. Adsorption of charged macro-molecules on mixed fluid membranes. Colloids Surf. A 208, 41–50.
Hlady, V., Ho, C.-H., Britt, D.W., 2000. Quantitative analysis of pro-tein adsorption kinetics. In: Kallay, N. (Ed.), Interfacial Dynamics.Surfactant Science Series. M. Dekker Inc., NY, pp. 405–418.
Hook, F., Rodahl, M., Brzezinski, P., Kasemo, B., 1998. Measurementsusing the quartz crystal microbalance technique of ferritin monolayerson methyl-thiolated gold: dependence of energy dissipation and sat-uration coverage on salt concentration. J. Colloid Interface Sci. 208,63.
Jogikalmath, G., 2003. Study of polystyrene-poly (ethylene oxide) diblockcopolymer monolayers as barriers to protein adsorption. Ph.D. Thesis.Department of Materials Science and Engineering, University of Utah.
Johnson, C.A., Yuan, Y., Lenhoff, A.M., 2000. Adsorbed layers of ferritinat solid and fluid interfaces studied by atomic force microscopy. J.Colloid Interface Sci. 223, 261.
Jung, L.S., Campbell, C.T., Chinowsky, T.M., Mar, M.N., Yee, S.S., 1998.Quantitative interpretation of the response of surface plasmon reso-nance sensors to adsorbed films. Langmuir 14, 5636.
Last, J.A., Waggoner, T.A., Sasaki, D.Y., 2001. Lipid membrane re-organization induced by chemical recognition. Biophys. J. 81 (5),2737–2742.
Litvin, A., Samuleson, L.A., Charych, D.H., Spevak, W., Kaplan, D.L.,1995. Influence of supramolecular template organization on mineral-ization. J. Phys. Chem. 99, 12065–12068.
Mallik, S., Plunkett, S.D., Dhal, P.K., Johnson, R.D., Pack, D., Shnek,D., Arnold, F.H., 1994. Towards materials for the specific recognitionand separation of proteins. New J. Chem. 18, 299–304.
M res-lipid
M gble
M nds on
M ateral
N rings to
P cedamol.
P n of
U ech-
ondurant, B., Last, J.A., Waggoner, T.A., Slade, A., Sasaki, D.Y., 2Optical and scanning probe analysis of glycolipid reorganizationconcanavalin A binding to mannose-coated lipid bilayers. Lang19, 1829–1837.
ritt, D.W., Hlady, V., Mobius, D., 2000. Ferritin adsorption to mucomponent monolayers: influence of lipid charge density, misciband fluidity on adsorption. Phys. Chem. Chem. Phys. 2, 4594–4
uijs, J., Britt, D.W., Hlady, V., 1998. Human growth hormone adsorpkinetics and conformation on self-assembled monolayers. Lang14, 335–341.
alonder, C., Tie, Y., Van Tassel, P.R., 2001. History dependence otein adsorption kinetics. Proc. Natl. Acad. Sci. USA 98, 10664–10
rowther, R.A., Klug, A., 1975. Structural analysis of macromolecassemblies by image reconstruction from electron micrographs.Rev. Biochem. 44, 161–182.
idalevitz, D., Weissbuch, I., Kjaer, K., Als-Nielsen, J., Leiserowitz,1994. Design of two dimensional crystals as models for prothe structure of the solid–liquid interface. J. Am. Chem. Soc.3271–3278.
oodman, T., Bussmann, E., Williams, C., Taveras, M., BD., 2004. Miscibility analysis of two-component lipid monola
aloney, K.M., Shnek, D.R., Sasaki, D.Y., Arnold, F.H., 1996. Fluocent signaling of ligand binding and assembly in metal-chelatingmembranes. Chem. Biol. 3, 185–192.
artın, M.T., Prieto, I., Camacho, L., Mobius, D., 1996. Partial stackinof a water-soluble porphyrin in complex monolayers with insolulipid. Langmuir 12, 6554–6560.
ay, S., Harries, D., Ben-Shaul, A., 2000. A Lipid demixing aprotein–protein interactions in the adsorption of charged proteinmixed membranes. Biophys. J. 79, 1747–1760.
ayer, L., Nelsestuen, G., 1981. Calcium and prothrombin induced lphase separation in membranes. Biochemistry 20, 2457–2463.
g, K., Pack, D.W., Sasaki, D.Y., Arnold, F.H., 1995. Engineeprotein–lipid interactions: targeting of histidine-tagged proteinmetal-chelating lipid monolayers. Langmuir 11, 4048–4055.
ack, D.W., Ng, K., Maloney, K.M., Arnold, F.H., 1997. Ligand-indureorganization and assembly in synthetic lipid membranes. SuprSci. 4, 3–10.
auling, L.A., 1940. Theory of the structure and process of formatioantibodies. J. Am. Chem. Soc. 62, 2643–2657.
giriz, E.E., Kornberg, R.D., 1983. Two-dimensional crystallization tnique for imaging macromolecules. Nature 301, 125–129.
Copyright © 2022 FDOKUMEN




















