
A reporter model to visualize imprinting stability at the Dlk1 ...
15
STEM CELLS AND REGENERATION TECHNIQUES AND RESOURCES REPORT A reporter model to visualize imprinting stability at the Dlk1 locus during mouse development and in pluripotent cells Emily Swanzey and Matthias Stadtfeld* ABSTRACT Genomic imprinting results in the monoallelic expression of genes that encode important regulators of growth and proliferation. Dysregulation of imprinted genes, such as those within the Dlk1- Dio3 locus, is associated with developmental syndromes and specific diseases. Our ability to interrogate causes of imprinting instability has been hindered by the absence of suitable model systems. Here, we describe a Dlk1 knock-in reporter mouse that enables single-cell visualization of allele-specific expression and prospective isolation of cells, simultaneously. We show that this ‘imprinting reporter mouse’ can be used to detect tissue-specific Dlk1 expression patterns in developing embryos. We also apply this system to pluripotent cell culture and demonstrate that it faithfully indicates DNA methylation changes induced upon cellular reprogramming. Finally, the reporter system reveals the role of elevated oxygen levels in eroding imprinted Dlk1 expression during prolonged culture and in vitro differentiation. The possibility to study allele-specific expression in different contexts makes our reporter system a useful tool to dissect the regulation of genomic imprinting in normal development and disease. KEY WORDS: Genomic imprinting, Dlk1, Fluorescent reporter, Allele-specific expression, DNA methylation, Pluripotency INTRODUCTION More than 100 mammalian genes are expressed in a predominantly monoallelic fashion in a paradigmatic epigenetic event referred to as genomic imprinting (Bartolomei and Ferguson-Smith, 2011). Imprinted genes, such as those within the commonly studied Dlk1-Dio3 gene cluster, are regulated by gender-specific DNA methylation marks at imprinting control regions (ICRs) (Duffie and Bourc’his, 2013; Sanli and Feil, 2015). Failure to preserve allele- specific imprinted gene expression, such as by the acquisition of hypermethylation at the ICR (referred to as loss-of-imprinting or LOI), can have detrimental developmental consequences and is a hallmark of cancer (Peters, 2014). The factors that contribute to the epigenetic instability of imprinted genes remain largely elusive, partly because of the absence of suitable model systems. At present, imprinting is typically studied by assessing DNA methylation levels or nucleotide polymorphisms in imprinted transcripts. These tools, however, are restricted to either bulk populations (Babak et al., 2015; Hammoud et al., 2013) and/or retrospective analysis (Ginart et al., 2016). Dlk1 is a paternally expressed protein-coding gene within Dlk1- Dio3 (da Rocha et al., 2008) that regulates fetal growth. We and others have previously shown that the ICR controlling Dlk1-Dio3, called the intergenic differentially methylated region (IG-DMR), frequently becomes hypermethylated in a context-dependent manner during in vitro reprogramming of somatic cells (Carey et al., 2011; Stadtfeld et al., 2010). This yields induced pluripotent stem cells (iPSCs) with LOI and upregulation of Dlk1 that is indicative of expression from both alleles without paternal bias (Stadtfeld et al., 2010). Together, these observations suggest that the insertion of fluorescent reporter genes into the endogenous Dlk1 locus might enable novel approaches to study imprinted gene expression in living cells and tissues. RESULTS AND DISCUSSION A reporter model for allele-specific expression of Dlk1 To generate a mouse reporter system for imprinted Dlk1 expression, we inserted the coding sequence for the green/yellow fluorescent protein Venus or the red fluorescent protein tdTomato (Tomato), into the three prime untranslated region (3′UTR) of endogenous Dlk1 (Fig. 1A). In accordance with widespread Dlk1 expression at perinatal stages (da Rocha et al., 2007), embryonic day (E)16.5 mice that had inherited reporter alleles from the father exhibited strong fluorescence (Fig. 1B). Reporter gene expression from the maternal allele was greatly diminished but detectable above background levels (Fig. 1B). The brightness of the paternal reporter allele allowed direct visualization of Dlk1 expression in living pups. This revealed expression in the growing limbs and the trunk until postnatal day (P)2, when it rapidly declined and became nearly undetectable by P4 (Fig. 1C). These findings are in accordance with previous studies (Lui et al., 2008), indicating that our reporter model recapitulates expression hallmarks of Dlk1. Identification of organ-specific expression patterns at the single-cell level We next sought to test whether the dual-reporter system could provide insight into allele-specific Dlk1 expression during development. Therefore, we analyzed E16.5 lung, skin and liver, representing tissues for which a developmental function of Dlk1 has been reported (Driskell et al., 2013; Tanimizu et al., 2003; Weng et al., 2009; Wu et al., 2008). Flow cytometric analysis revealed strong paternal reporter expression and detectable levels of maternal expression in some cells (Fig. 2A). The number of such bi-allelic cells ranged from rare (∼4% of Dlk1 + cells) in lung, to predominant (>95%) in liver (Fig. 2A,B). Importantly, the reporter insertions did not alter the expression levels of Dlk1 or the reciprocally expressed Gtl2 gene in the tissues analyzed (Fig. S1). Of note, while the reporter alleles revealed relatively homogenous expression profiles Received 5 April 2016; Accepted 29 September 2016 The Helen L. and Martin S. Kimmel Center for Biology and Medicine, Skirball Institute of Biomolecular Medicine, Department of Cell Biology, NYU School of Medicine, New York, NY 10016, USA. *Author for correspondence ([email protected]) M.S., 0000-0002-5852-9906 This is an Open Access article distributed under the terms of the Creative Commons Attribution License (http://creativecommons.org/licenses/by/3.0), which permits unrestricted use, distribution and reproduction in any medium provided that the original work is properly attributed. 4161 © 2016. Published by The Company of Biologists Ltd | Development (2016) 143, 4161-4166 doi:10.1242/dev.138255 DEVELOPMENT
-
Upload
khangminh22 -
Category
Documents
-
view
2 -
download
0
Transcript of A reporter model to visualize imprinting stability at the Dlk1 ...

STEM CELLS AND REGENERATION TECHNIQUES AND RESOURCES REPORT
A reporter model to visualize imprinting stability at the Dlk1 locusduring mouse development and in pluripotent cellsEmily Swanzey and Matthias Stadtfeld*
ABSTRACTGenomic imprinting results in the monoallelic expression of genesthat encode important regulators of growth and proliferation.Dysregulation of imprinted genes, such as those within the Dlk1-Dio3 locus, is associated with developmental syndromes and specificdiseases. Our ability to interrogate causes of imprinting instability hasbeen hindered by the absence of suitable model systems. Here, wedescribe a Dlk1 knock-in reporter mouse that enables single-cellvisualization of allele-specific expression and prospective isolation ofcells, simultaneously. We show that this ‘imprinting reporter mouse’can be used to detect tissue-specific Dlk1 expression patterns indeveloping embryos. We also apply this system to pluripotent cellculture and demonstrate that it faithfully indicates DNA methylationchanges induced upon cellular reprogramming. Finally, the reportersystem reveals the role of elevated oxygen levels in eroding imprintedDlk1 expression during prolonged culture and in vitro differentiation.The possibility to study allele-specific expression in different contextsmakes our reporter system a useful tool to dissect the regulation ofgenomic imprinting in normal development and disease.
KEY WORDS: Genomic imprinting, Dlk1, Fluorescent reporter,Allele-specific expression, DNA methylation, Pluripotency
INTRODUCTIONMore than 100 mammalian genes are expressed in a predominantlymonoallelic fashion in a paradigmatic epigenetic event referred to asgenomic imprinting (Bartolomei and Ferguson-Smith, 2011).Imprinted genes, such as those within the commonly studiedDlk1-Dio3 gene cluster, are regulated by gender-specific DNAmethylation marks at imprinting control regions (ICRs) (Duffie andBourc’his, 2013; Sanli and Feil, 2015). Failure to preserve allele-specific imprinted gene expression, such as by the acquisition ofhypermethylation at the ICR (referred to as loss-of-imprinting orLOI), can have detrimental developmental consequences and is ahallmark of cancer (Peters, 2014). The factors that contribute to theepigenetic instability of imprinted genes remain largely elusive,partly because of the absence of suitable model systems. At present,imprinting is typically studied by assessing DNAmethylation levelsor nucleotide polymorphisms in imprinted transcripts. These tools,
however, are restricted to either bulk populations (Babak et al.,2015; Hammoud et al., 2013) and/or retrospective analysis (Ginartet al., 2016).
Dlk1 is a paternally expressed protein-coding gene within Dlk1-Dio3 (da Rocha et al., 2008) that regulates fetal growth. We andothers have previously shown that the ICR controlling Dlk1-Dio3,called the intergenic differentially methylated region (IG-DMR),frequently becomes hypermethylated in a context-dependentmanner during in vitro reprogramming of somatic cells (Careyet al., 2011; Stadtfeld et al., 2010). This yields induced pluripotentstem cells (iPSCs) with LOI and upregulation of Dlk1 that isindicative of expression from both alleles without paternal bias(Stadtfeld et al., 2010). Together, these observations suggest that theinsertion of fluorescent reporter genes into the endogenous Dlk1locus might enable novel approaches to study imprinted geneexpression in living cells and tissues.
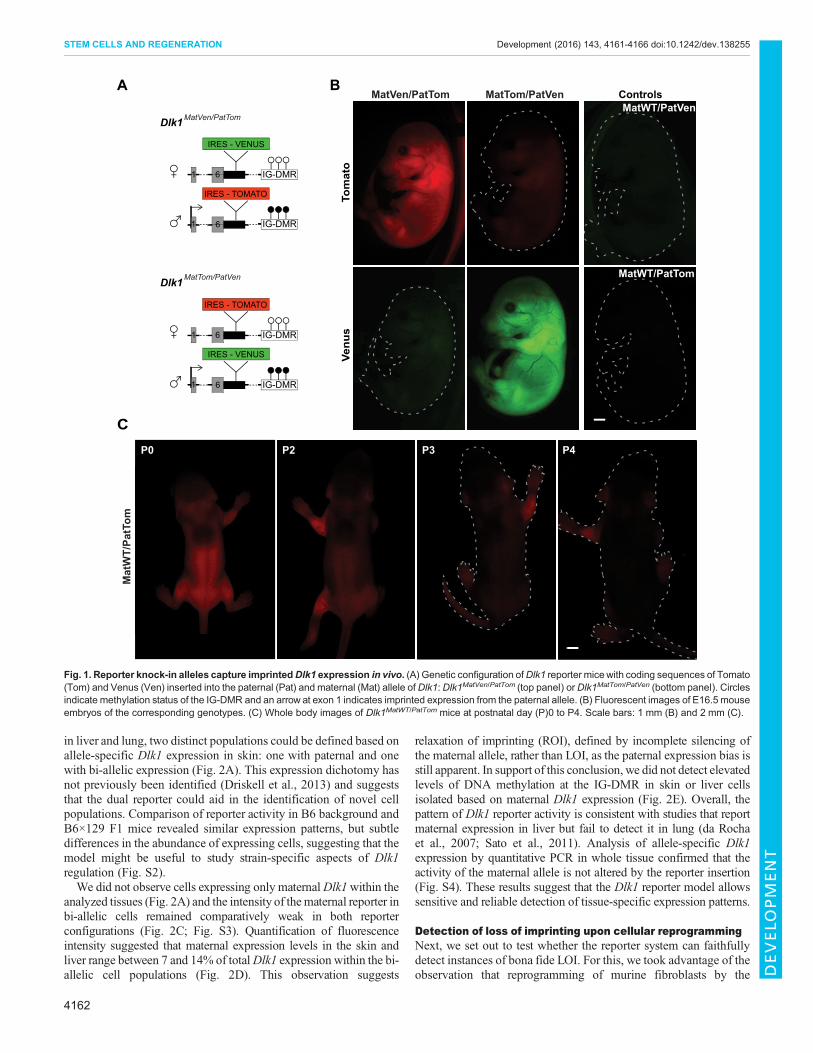
RESULTS AND DISCUSSIONA reporter model for allele-specific expression of Dlk1To generate a mouse reporter system for imprinted Dlk1 expression,we inserted the coding sequence for the green/yellow fluorescentprotein Venus or the red fluorescent protein tdTomato (Tomato),into the three prime untranslated region (3′UTR) of endogenousDlk1 (Fig. 1A). In accordance with widespread Dlk1 expression atperinatal stages (da Rocha et al., 2007), embryonic day (E)16.5 micethat had inherited reporter alleles from the father exhibited strongfluorescence (Fig. 1B). Reporter gene expression from the maternalallele was greatly diminished but detectable above backgroundlevels (Fig. 1B). The brightness of the paternal reporter alleleallowed direct visualization of Dlk1 expression in living pups. Thisrevealed expression in the growing limbs and the trunk untilpostnatal day (P)2, when it rapidly declined and became nearlyundetectable by P4 (Fig. 1C). These findings are in accordance withprevious studies (Lui et al., 2008), indicating that our reporter modelrecapitulates expression hallmarks of Dlk1.
Identification of organ-specific expression patterns at thesingle-cell levelWe next sought to test whether the dual-reporter system couldprovide insight into allele-specific Dlk1 expression duringdevelopment. Therefore, we analyzed E16.5 lung, skin and liver,representing tissues for which a developmental function ofDlk1 hasbeen reported (Driskell et al., 2013; Tanimizu et al., 2003; Wenget al., 2009; Wu et al., 2008). Flow cytometric analysis revealedstrong paternal reporter expression and detectable levels of maternalexpression in some cells (Fig. 2A). The number of such bi-alleliccells ranged from rare (∼4% of Dlk1+ cells) in lung, to predominant(>95%) in liver (Fig. 2A,B). Importantly, the reporter insertions didnot alter the expression levels of Dlk1 or the reciprocally expressedGtl2 gene in the tissues analyzed (Fig. S1). Of note, while thereporter alleles revealed relatively homogenous expression profilesReceived 5 April 2016; Accepted 29 September 2016
The Helen L. and Martin S. Kimmel Center for Biology and Medicine, SkirballInstitute of Biomolecular Medicine, Department of Cell Biology, NYU School ofMedicine, New York, NY 10016, USA.
*Author for correspondence ([email protected])
M.S., 0000-0002-5852-9906
This is an Open Access article distributed under the terms of the Creative Commons AttributionLicense (http://creativecommons.org/licenses/by/3.0), which permits unrestricted use,distribution and reproduction in any medium provided that the original work is properly attributed.
4161
© 2016. Published by The Company of Biologists Ltd | Development (2016) 143, 4161-4166 doi:10.1242/dev.138255
DEVELO
PM
ENT
in liver and lung, two distinct populations could be defined based onallele-specific Dlk1 expression in skin: one with paternal and onewith bi-allelic expression (Fig. 2A). This expression dichotomy hasnot previously been identified (Driskell et al., 2013) and suggeststhat the dual reporter could aid in the identification of novel cellpopulations. Comparison of reporter activity in B6 background andB6×129 F1 mice revealed similar expression patterns, but subtledifferences in the abundance of expressing cells, suggesting that themodel might be useful to study strain-specific aspects of Dlk1regulation (Fig. S2).We did not observe cells expressing only maternalDlk1within the
analyzed tissues (Fig. 2A) and the intensity of thematernal reporter inbi-allelic cells remained comparatively weak in both reporterconfigurations (Fig. 2C; Fig. S3). Quantification of fluorescenceintensity suggested that maternal expression levels in the skin andliver range between 7 and 14% of totalDlk1 expression within the bi-allelic cell populations (Fig. 2D). This observation suggests
relaxation of imprinting (ROI), defined by incomplete silencing ofthe maternal allele, rather than LOI, as the paternal expression bias isstill apparent. In support of this conclusion, we did not detect elevatedlevels of DNA methylation at the IG-DMR in skin or liver cellsisolated based on maternal Dlk1 expression (Fig. 2E). Overall, thepattern of Dlk1 reporter activity is consistent with studies that reportmaternal expression in liver but fail to detect it in lung (da Rochaet al., 2007; Sato et al., 2011). Analysis of allele-specific Dlk1expression by quantitative PCR in whole tissue confirmed that theactivity of the maternal allele is not altered by the reporter insertion(Fig. S4). These results suggest that the Dlk1 reporter model allowssensitive and reliable detection of tissue-specific expression patterns.
Detection of loss of imprinting upon cellular reprogrammingNext, we set out to test whether the reporter system can faithfullydetect instances of bona fide LOI. For this, we took advantage of theobservation that reprogramming of murine fibroblasts by the
A
61
IRES - VENUS
IRES - TOMATO
61
61
IRES - VENUS
IRES - TOMATO
IG-DMR 61
CP0 P2 P4P3
Tom
ato
Venu
s
BControls
Dlk1 MatVen/PatTom
Dlk1 MatTom/PatVen
MatVen/PatTom MatTom/PatVen
MatWT/PatTom
MatWT/PatVen
Mat
WT/
PatT
om
IG-DMR
IG-DMR
IG-DMR
Fig. 1. Reporter knock-in alleles capture imprintedDlk1 expression in vivo. (A) Genetic configuration ofDlk1 reporter micewith coding sequences of Tomato(Tom) and Venus (Ven) inserted into the paternal (Pat) and maternal (Mat) allele ofDlk1:Dlk1MatVen/PatTom (top panel) orDlk1MatTom/PatVen (bottom panel). Circlesindicate methylation status of the IG-DMR and an arrow at exon 1 indicates imprinted expression from the paternal allele. (B) Fluorescent images of E16.5 mouseembryos of the corresponding genotypes. (C) Whole body images of Dlk1MatWT/PatTom mice at postnatal day (P)0 to P4. Scale bars: 1 mm (B) and 2 mm (C).
4162
STEM CELLS AND REGENERATION Development (2016) 143, 4161-4166 doi:10.1242/dev.138255
DEVELO
PM
ENT

transcription factors, Oct4, Klf4, Sox2 andMyc (together referred toas OKSM), frequently yields induced pluripotent stem cells (iPSCs)with DNA hypermethylation of the IG-DMR and upregulation ofDlk1 (Liu et al., 2010; Stadtfeld et al., 2010). This abnormalityresults in reduced developmental potential but can be prevented byaddition of ascorbic acid (AA) to the reprogramming medium(Stadtfeld et al., 2012). We therefore anticipated thatreprogramming of Dlk1 reporter fibroblasts in basal conditionswould predominantly yield cells with equal paternal and maternalexpression, while reprogramming in the presence of AA wouldprevent the occurrence of such cells (Fig. 3A).In agreement with low levels of Dlk1 transcription in pluripotent
cells (Kota et al., 2014), no reporter gene expression could bedetected in naïve iPSCs. However, exposure to the differentiation-inducing agent retinoic acid (RA) yielded cells with readilydetectable reporter gene fluorescence. As shown in Fig. 3B, cellsderived in basal conditions almost exclusively expressed bothreporter alleles, whereas monoallelic fluorescence was prevalent incells obtained in the presence of AA. In contrast to our observationsin the embryo, paternal and maternal expression levels in cellsgenerated in basal reprogramming conditions were indistinguishable(compare Fig. 2C and Fig. 3C). As this is consistent with imprintingloss, we will refer to such cells as ‘LOI’ cells, while we call thosewith paternal-only or bi-allelic with low maternal expression ‘MOI’
cells (see Fig. 3B). In agreement with this categorization, iPSCs witha high degree of LOI expression showed strongly elevated levels ofDNA methylation at the IG-DMR (Fig. 3E; Fig. S5).
These observations suggest that the Dlk1 reporters accuratelyreflect changes in imprinting status during reprogramming. We alsoevaluatedDlk1 imprinting in iPSCs derived by combinedmodulationof TGFβ andWnt signaling in the presence of AA (referred to as ‘3c’conditions) (Vidal et al., 2014), which increases reprogrammingefficiencies 10- to 20-fold (Fig. 3D). We found that differentiation ofiPSCs derived in 3c mirrored the results obtained in AA conditions,with the majority of cells exhibiting normal DNA methylation levelsandMOI expression ofDlk1 (Fig. 3E; Figs S5,S6). This suggests thatthis highly efficient reprogramming condition might provide atractable method to study factors that contribute to imprintingmaintenance.
Modulation of allele-specific Dlk1 expression by oxygenlevels during iPSC culture and differentiationDespite the striking difference in maternal reporter expressionbetween cells derived in basal and 3c conditions, we noticed thatdifferentiation of AA and 3c iPSCs frequently yielded a small subsetof LOI cells (Fig. 3B). Such cells were rare when differentiatingfreshly generated iPSCs but became more frequent with iPSCs athigher passage, along with a number of cells with maternal-only
EFluorescence intensity
maternalpaternal
negative
Liver
0-103
103
104
105
0
20
40
60
80Cou
nt
Skin
0
20
40
60
D
MOI LOI 1 2%
CpG
met
hyla
tion
1 2 0
20
40
60
80
100
10
20
30
40
50
60
70
80
bi-allelic paternal only
% p
ositi
ve c
ells
of t
otal
Lung Skin LiverEmbryo: 1 2 3 1 2 3 1 2 3
0
B CLu
ng
0.25 2.16
49.648.0
0.11 8.22
41.450.3
Skin
1.15 50.2
1.3547.3
Live
rA
Skin Liver
Tom
ato
Venus
MatTom/PatVen
1.14 0.34
60.737.8
0.12 0.10
45.054.8
0.30 0.21
51.348.2
MatWT/PatVen
skin liver0
5
10
15
%m
ater
nal e
xpre
ssio
n of
tota
l
Fig. 2. Quantification of allele-specific expression in developing tissues. (A) Flow cytometric analysis of fetal tissues from Dlk1MatWT/PatVen (left column)control mice orDlk1MatTom/PatVen (right column) dual-reporter mice at E16.5. (B) Percentage ofDlk1+ cells with paternal-only and bi-allelic expression in fetal lung,skin and liver. (C) Mean fluorescent intensity of paternal and maternal Dlk1Tom in bi-allelic fetal skin and liver cells. (D) Quantification of relative strength ofmaternal to totalDlk1 in bi-allelic cells as measured by flow cytometry. Error bars indicate s.e. (n=3 embryos). (E) DNAmethylation at the IG-DMR in FACS-sortedfetal skin and liver cells with active maternal Dlk1 allele from two embryos. Methylation levels in iPSCs with established LOI or MOI were used as controls. Errorbars indicate s.e. (n=28 CpGs analyzed).
4163
STEM CELLS AND REGENERATION Development (2016) 143, 4161-4166 doi:10.1242/dev.138255
DEVELO
PM
ENT

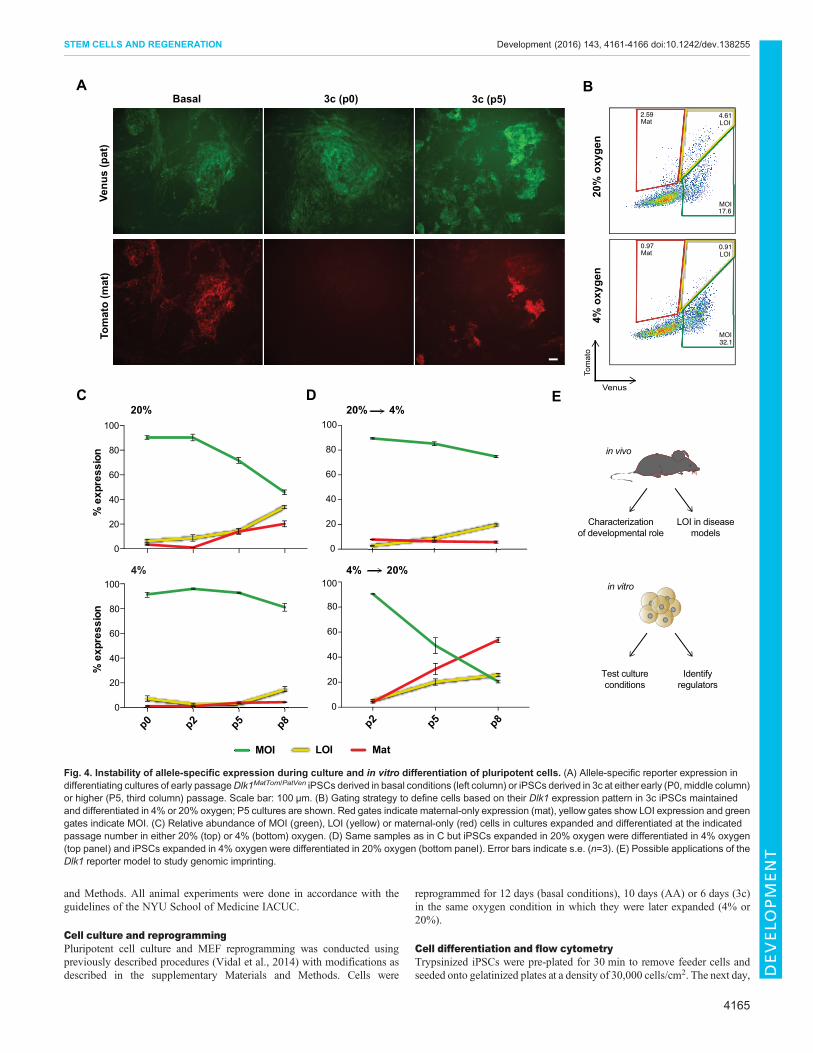
expression (Fig. 4A). All in vitro experiments described thus farwere conducted in standard culture conditions, including 20%oxygen levels. In light of the reported impact of molecular oxygenon epigenetic processes in pluripotent cells, including imprinting(Xie et al., 2014) and X-chromosome inactivation (Lengner et al.,2010), we tested the impact of oxygen concentration on allele-specific Dlk1 expression. Taking advantage of the 3c system, whichenables analysis at earlier passage, we compared iPSCs that wereexpanded and differentiated in atmospheric (20%) andphysiological (4%) oxygen levels. We observed no significantdifference with early passage iPSCs, but a gradual increase in cellswith LOI and maternal-only reporter expression in 20% oxygenconditions at higher passage (Fig. 4B,C). In contrast, predominantlyMOI expression was maintained in 4% oxygen (Fig. 4B,C). Whencells expanded in 20% oxygen were differentiated in 4% oxygen,the frequency of cells with LOI and maternal-only expressionremained low, while their numbers strongly increased in the reverseconditions (expansion in 4%, followed by differentiation in 20%oxygen) (Fig. 4C; Fig. S7). This indicates an unexpectedsusceptibility of the maternal Dlk1 allele to become reactivatedupon prolonged expansion and differentiation of iPSCs at elevatedoxygen levels. Because imprint dysregulation has been recognized
as a concern for the quality of pluripotent cell lines (Greenberg andBourc’his, 2015), we anticipate that theDlk1 reporter system will beuseful in optimizing stem cell derivation and culture conditions.
In summary, we have developed a reporter model that serves as asensitive indicator for allele-specific expression of Dlk1 and allowsprospective isolation of even small subsets of cells with paternal, bi-allelic, LOI or maternal-only expression patterns. Our observationsare consistent with tight control of Dlk1 imprinting during earlydevelopmental stages in the tissues analyzed, but suggest frequenterosion during in vitro manipulations. The system described hereshould therefore provide a novel means to systematically studyDlk1-Dio3 regulation in diseased tissues, pluripotent cell cultureand during development (Fig. 4E). This should aid in refining ourunderstanding of the molecular processes involved in theestablishment and maintenance of imprinting.
MATERIALS AND METHODSTransgenic miceAll animals were on a B6 background, unless otherwise indicated. Forreprogramming experiments, Dlk1 reporter mice were crossed with animalscarrying an inducible OKSM transgene (Stadtfeld et al., 2010). Dlk1reporter mice were generated as described in the supplementary Materials
BAA
24.2
2.43
Basal2.89
28.3
C
0
20
40
60
80
% L
OI
AA 3cBasal
0
20
40
60
80
% C
pG m
ethy
latio
n 1 2 3 4 1 2 3 1 2 3
A
D
0
10
20
30
40
50
%re
prog
ram
min
g ef
ficie
ncy
****
AA 3cBasal0-103
103
104
105
Fluorescence intensity
0
50
100
150
Cou
nt
E
PatTomWT
MatTom LOIMatTom MOI
AA
LOI
MOI
Rosa26M2-rtTa
tetOOKSM
Dlk1-PatTomato
Venus
Dlk1-Mat +Dox
MOI
LOI
MOI
LOIBasal
Tom
ato
Venus
Dlk1 locus
****
Fig. 3. Detection of LOI induced by cellular reprogramming. (A) Transgenic alleles in fibroblasts isolated from reprogrammable imprinting reporter animals. Theexpected behavior of the reporter genes in cells derived upon reprogramming in basal or AA conditions is illustrated. (B) Representative FACS plots ofRA-differentiated iPSCs derived via basal reprogramming (left) or in AA conditions (right). Yellow and green gates indicate LOI- and MOI-expressing cells,respectively. (C) Mean fluorescence intensity of Tomato in the indicated genotypes. (D) Reprogramming efficiencymeasured as percentage of stable iPSC coloniesper input MEFs in basal conditions, AA or 3c. ****P<0.00005 with a one-way ANOVA and Tukey’s multiple comparison test. Error bars indicate standard error (n=3).(E) LOI-level Dlk1 expression (top panel) and DNA methylation at the IG-DMR (bottom panel) in iPSC cultures derived in the indicated conditions.
4164
STEM CELLS AND REGENERATION Development (2016) 143, 4161-4166 doi:10.1242/dev.138255
DEVELO
PM
ENT
and Methods. All animal experiments were done in accordance with theguidelines of the NYU School of Medicine IACUC.
Cell culture and reprogrammingPluripotent cell culture and MEF reprogramming was conducted usingpreviously described procedures (Vidal et al., 2014) with modifications asdescribed in the supplementary Materials and Methods. Cells were
reprogrammed for 12 days (basal conditions), 10 days (AA) or 6 days (3c)in the same oxygen condition in which they were later expanded (4% or20%).
Cell differentiation and flow cytometryTrypsinized iPSCs were pre-plated for 30 min to remove feeder cells andseeded onto gelatinized plates at a density of 30,000 cells/cm2. The next day,
AVe
nus
(pat
)Basal
Tom
ato
(mat
)3c (p5) 3c (p0)
MatLOI
C
0
20
40
60
80
100
% e
xpre
ssio
n
20%
4%
p0 p2 p5 p80
20
40
60
80
100
% e
xpre
ssio
n
p2 p5 p80
20
40
60
80
100
0
20
40
60
80
100
D20% 4%
4% 20%
in vivo
in vitro
LOI in diseasemodels
Characterizationof developmental role
Identifyregulators
Test cultureconditions
E
B
20%
oxy
gen
4% o
xyge
n
4.612.59
17.6MOI
LOIMat
94750.910.97
32.1MOI
LOIMat
MOI
Tom
ato
Venus
Fig. 4. Instability of allele-specific expression during culture and in vitro differentiation of pluripotent cells. (A) Allele-specific reporter expression indifferentiating cultures of early passageDlk1MatTom/PatVen iPSCs derived in basal conditions (left column) or iPSCs derived in 3c at either early (P0,middle column)or higher (P5, third column) passage. Scale bar: 100 µm. (B) Gating strategy to define cells based on their Dlk1 expression pattern in 3c iPSCs maintainedand differentiated in 4% or 20% oxygen; P5 cultures are shown. Red gates indicate maternal-only expression (mat), yellow gates show LOI expression and greengates indicate MOI. (C) Relative abundance of MOI (green), LOI (yellow) or maternal-only (red) cells in cultures expanded and differentiated at the indicatedpassage number in either 20% (top) or 4% (bottom) oxygen. (D) Same samples as in C but iPSCs expanded in 20% oxygen were differentiated in 4% oxygen(top panel) and iPSCs expanded in 4% oxygen were differentiated in 20% oxygen (bottom panel). Error bars indicate s.e. (n=3). (E) Possible applications of theDlk1 reporter model to study genomic imprinting.
4165
STEM CELLS AND REGENERATION Development (2016) 143, 4161-4166 doi:10.1242/dev.138255
DEVELO
PM
ENT

fibroblast mediumwith 0.4 µg/ml retinoic acid was added, followed by dailymedia changes and imaging using a Nikon Eclipse TiE inverted microscopewith filters to detect Venus (excitation, 500/20 nm; emission, 535/30 nm)and Tomato (excitation, 545/30 nm; emission, 620/60 nm). Forquantification of Dlk1 reporter expression, dissociated cultures wereacquired on an LSRII cytometer (BD Biosciences) and analyzed withFlowJo software (Tree Star).
Tissue isolation and reporter detectionMouse embryos isolated at E16.5 were imaged using a Nikon SMZ1500Stereo Fluorescence Microscope. For flow cytometry, isolated tissues wereincubated for 30 min at 37°C in 0.25% trypsin. During this time, tissueswere disassociated using progressively smaller pipet tips (1000 μl to200 μl), followed by analysis on an LSRII or sorting on a FACSAria (BDBiosciences).
DNA methylation analysisGenomic DNA was isolated using proteinase K in lysis buffer, pH 8(100 mM Tris-HCl, 5 mM EDTA, 0.2% SDS, 200 mM NaCl) andreconstituted in TE buffer, pH 7.5 (10 mM Tris-HCl, 1 mM EDTA).DNA pyrosequencing and next-generation bisulfite sequencing wasconducted using the ADS1452 assay, which covers the IG-DMR(chr.12:109,528,253-109,528,471 in mm10) (EpigenDX).
Gene expression analysisRNA isolated using the miRNeasy kit (Qiagen) was used for cDNApreparation with the Transcriptor HIFI cDNA synthesis kit (Roche).Samples were run on a LightCycler 480 Real-Time PCR System (Roche).Relative allele-specific expression was assessed using qRT-PCR withprimers as described in the supplementary Materials and Methods.
AcknowledgementsWe would like to thank Mike Gregory and the NYUSOM Flow Cytometry Core forhelp with cell sorting, Sang Yong Kim for blastocyst injections, Bhishma Amlani forhelp with statistical analysis, and Effie Apostolou and all members of the Stadtfeldlab for discussion.
Competing interestsThe authors declare no competing or financial interests.
Author contributionsE.S. and M.S. conceived the study, designed experiments, interpreted results andwrote the manuscript. E.S. conducted all experiments.
FundingM.S. is an American Society of Hematology fellowand was supported by grants fromthe Sidney Kimmel Foundation for Cancer Research, the March of DimesFoundation, the NewYork State Department of Health (C092546 and C028130) andthe National Institutes of Health (1R21 HD079883-01 and 1R01 GM111852-01).Deposited in PMC for immediate release.
Supplementary informationSupplementary information available online athttp://dev.biologists.org/lookup/doi/10.1242/dev.138255.supplemental
ReferencesBabak, T., DeVeale, B., Tsang, E. K., Zhou, Y., Li, X., Smith, K. S., Kukurba, K. R.,Zhang, R., Li, J. B., van der Kooy, D. et al. (2015). Genetic conflict reflected intissue-specific maps of genomic imprinting in human and mouse. Nat. Genet. 47,544-549.
Bartolomei, M. S. and Ferguson-Smith, A. C. (2011). Mammalian genomicimprinting. Cold Spring Harb. Perspect. Biol. 3, a002592.
Carey, B. W., Markoulaki, S., Hanna, J. H., Faddah, D. A., Buganim, Y., Kim, J.,Ganz, K., Steine, E. J., Cassady, J. P., Creyghton, M. P. et al. (2011).
Reprogramming factor stoichiometry influences the epigenetic state andbiological properties of induced pluripotent stem cells. Cell Stem Cell 9, 588-598.
da Rocha, S. T., Tevendale, M., Knowles, E., Takada, S., Watkins, M. andFerguson-Smith, A. C. (2007). Restricted co-expression of Dlk1 and thereciprocally imprinted non-coding RNA, Gtl2: implications for cis-acting control.Dev. Biol. 306, 810-823.
da Rocha, S. T., Edwards, C. A., Ito, M., Ogata, T. and Ferguson-Smith, A. C.(2008). Genomic imprinting at the mammalian Dlk1-Dio3 domain. Trends Genet.24, 306-316.
Driskell, R. R., Lichtenberger, B. M., Hoste, E., Kretzschmar, K., Simons, B. D.,Charalambous, M., Ferron, S. R., Herault, Y., Pavlovic, G., Ferguson-Smith,A. C. et al. (2013). Distinct fibroblast lineages determine dermal architecture inskin development and repair. Nature 504, 277-281.
Duffie, R. and Bourc’his, D. (2013). Parental epigenetic asymmetry in mammals.Curr. Top. Dev. Biol. 104, 293-328.
Ginart, P., Kalish, J. M., Jiang, C. L., Yu, A. C., Bartolomei, M. S. and Raj, A.(2016). Visualizing allele-specific expression in single cells reveals epigeneticmosaicism in an H19 loss-of-imprinting mutant. Genes Dev. 30, 567-578.
Greenberg, M. V. C. and Bourc’his, D. (2015). Cultural relativism: maintenance ofgenomic imprints in pluripotent stem cell culture systems. Curr. Opin. Genet. Dev.31, 42-49.
Hammoud, S. S., Cairns, B. R. and Carrell, D. T. (2013). Analysis of gene-specificand genome-wide sperm DNA methylation. Methods Mol. Biol. 927, 451-458.
Kota, S. K., Lleres, D., Bouschet, T., Hirasawa, R., Marchand, A., Begon-Pescia,C., Sanli, I., Arnaud, P., Journot, L., Girardot, M. et al. (2014). ICR noncodingRNA expression controls imprinting and DNA replication at the Dlk1-Dio3 domain.Dev. Cell 31, 19-33.
Lengner, C. J., Gimelbrant, A. A., Erwin, J. A., Cheng, A. W., Guenther, M. G.,Welstead, G. G., Alagappan, R., Frampton, G. M., Xu, P., Muffat, J. et al.(2010). Derivation of pre-X inactivation human embryonic stem cells underphysiological oxygen concentrations. Cell 141, 872-883.
Liu, L., Luo, G.-Z., Yang, W., Zhao, X., Zheng, Q., Lv, Z., Li, W., Wu, H.-J., Wang,L., Wang, X.-J. et al. (2010). Activation of the imprinted Dlk1-Dio3 regioncorrelates with pluripotency levels of mouse stem cells. J. Biol. Chem. 285,19483-19490.
Lui, J. C., Finkielstain, G. P., Barnes, K. M. and Baron, J. (2008). An imprintedgene network that controls mammalian somatic growth is down-regulated duringpostnatal growth deceleration in multiple organs. Am. J. Physiol. Regul. Integr.Comp. Physiol. 295, R189-R196.
Peters, J. (2014). The role of genomic imprinting in biology and disease: anexpanding view. Nat. Rev. Genet. 15, 517-530.
Sanli, I. and Feil, R. (2015). Chromatin mechanisms in the developmental control ofimprinted gene expression. Int. J. Biochem. Cell Biol. 67, 139-147.
Sato, S., Yoshida, W., Soejima, H., Nakabayashi, K. and Hata, K. (2011).Methylation dynamics of IG-DMR and Gtl2-DMR during murine embryonic andplacental development. Genomics 98, 120-127.
Stadtfeld, M., Apostolou, E., Akutsu, H., Fukuda, A., Follett, P., Natesan, S.,Kono, T., Shioda, T. and Hochedlinger, K. (2010). Aberrant silencing ofimprinted genes on chromosome 12qF1 in mouse induced pluripotent stem cells.Nature 465, 175-181.
Stadtfeld, M., Apostolou, E., Ferrari, F., Choi, J., Walsh, R. M., Chen, T., Ooi,S. S., Kim, S. Y., Bestor, T. H., Shioda, T. et al. (2012). Ascorbic acid preventsloss of Dlk1-Dio3 imprinting and facilitates generation of all-iPS cell mice fromterminally differentiated B cells. Nat. Genet. 44, 398-405.
Tanimizu, N., Nishikawa, M., Saito, H., Tsujimura, T. and Miyajima, A. (2003).Isolation of hepatoblasts based on the expression of Dlk/Pref-1. J. Cell Sci. 116,1775-1786.
Vidal, S. E., Amlani, B., Chen, T., Tsirigos, A. and Stadtfeld, M. (2014).Combinatorial modulation of signaling pathways reveals cell-type-specificrequirements for highly efficient and synchronous iPSC reprogramming. StemCell Rep. 3, 574-584.
Weng, T., Gao, L., Bhaskaran, M., Guo, Y., Gou, D., Narayanaperumal, J.,Chintagari, N. R., Zhang, K. and Liu, L. (2009). Pleiotrophin regulates lungepithelial cell proliferation and differentiation during fetal lung development viabeta-catenin and Dlk1. J. Biol. Chem. 284, 28021-28032.
Wu, Q., Kawahara, M. and Kono, T. (2008). Synergistic role of Igf2 and Dlk1 in fetalliver development and hematopoiesis in bi-maternal mice. J. Reprod. Dev. 54,177-182.
Xie, P., Sun, Y., Ouyang, Q., Hu, L., Tan, Y., Zhou, X., Xiong, B., Zhang, Q., Yuan,D., Pan, Y. et al. (2014). Physiological oxygen prevents frequent silencing of theDLK1-DIO3 cluster during human embryonic stem cells culture. Stem Cells 32,391-401.
4166
STEM CELLS AND REGENERATION Development (2016) 143, 4161-4166 doi:10.1242/dev.138255
DEVELO
PM
ENT

SUPPLEMENTARY METHODS
A reporter model to visualize imprinting stability at the Dlk1 locus during mouse development and in pluripotent cellsEmily Swanzey and Matthias Stadtfeld
Generation of Dlk1 reporter mice
To generate targeting vectors for insertion of reporter genes into the 3’ UTR of Dlk1, a
6.7kb EcoRV fragment was isolated from BAC RP23-117C15 and cloned into
pBluescript together with a floxed PGK-neoR cassette. In the resulting pBS-Dlk1, a
SnaBI site was introduced at the site corresponding to position chr12:109,460,514 in
mm10 using site-directed mutagenesis (Agilent QuikChange II). This site was used to
insert the coding sequence of IRES-Venus or IRES-Tomato. V6.5 ESCs were
electroporated with linearized (by SacII digestion) targeting vectors, followed by
selection in G418-containing media. Correctly targeted clones were identified by
Southern blotting of AseI-digested genomic DNA, using a 770bp HincII/AseI fragment
probe isolated from pBS-Dlk1. Following confirmation, the PGK-neoR cassette was
removed by transient transduction with Adeno-Cre. Resultant ESCs were injected into
diploid blastocysts and transgenic chimeric mice bred with B6 animals to establish
homozygous Dlk1 reporter mouse strains.
Cell culture ESCs and iPSCs were cultured in KO-DMEM with 15% FBS, L-Glutamine, penicillin-
streptomycin, non-essential amino acids, 2-Mercaptoethanol and 1000 U/ml LIF on
irradiated feeder cells. MEFs were cultured in DMEM with 10% FBS, L-Glutamine,
penicillin, streptomycin, non-essential amino acids and 2-Mercaptoethanol. For
reprogramming, MEFs were seeded in ESC media on a layer of irradiated feeder cells
in presence of 1 µg/ml doxycycline. If applicable, L-Ascorbic acid (50 µg/ml),
CHIR99021 (3 µM) and TGFβ RI Kinase Inhibitor II (0.25 µM) were added.
Development 143: doi:10.1242/dev.138255: Supplementary information
Dev
elo
pmen
t • S
uppl
emen
tary
info
rmat
ion

Primers for quantitative PCR Dlk1: GCGTGCTGTCCTGTGTGC and TGTCAGCCTCGCAGAATCCATAC
Gtl2: TTGCACATTTCCTGTGGGAC and AAGCACCATGAGCCACTAGG
Dlk1 3’ UTR: CCCCAGGCCCTTCTCTATTA and GTAGCATGGCACACAGCAAC
Venus: GGGACGTGGTTTTCCTTTGAA and AGATCAGCTTCAGGGTCAGC
GAPDH: AGGTCGGTGTGAACGGATTTG and TGTAGACCATGTAGTTGAGGTCA
Calculation of relative allele-specific expression by qRT-PCR Allele-specific expression was calculated from Dlk1MatWT/PatVen and Dlk1MatVen/PatWT
samples. WT expression was measured from the WT primer amplification of the
unmodified alleles and Venus expression was detected from Venus primer amplification.
Ct values were normalized to GAPDH, followed by calculation of MatWT expression as
a percentage of total WT (MatWT + PatWT) or MatVen expression as a percentage of
total Venus (MatVen + PatVen).
Development 143: doi:10.1242/dev.138255: Supplementary information
Dev
elo
pmen
t • S
uppl
emen
tary
info
rmat
ion
SUPPLEMENTARY FIGURES
A reporter model to visualize imprinting stability at the Dlk1 locus during mouse development and in pluripotent cellsEmily Swanzey and Matthias Stadtfeld
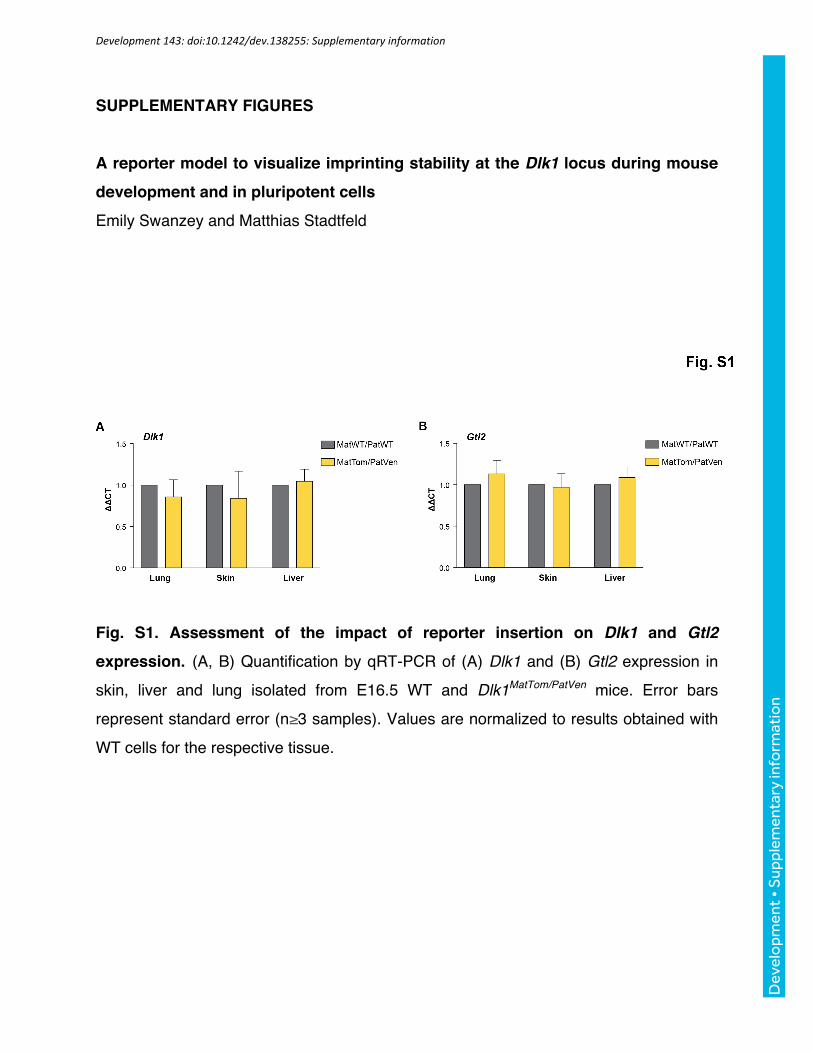
Fig. S1. Assessment of the impact of reporter insertion on Dlk1 and Gtl2
expression. (A, B) Quantification by qRT-PCR of (A) Dlk1 and (B) Gtl2 expression in
skin, liver and lung isolated from E16.5 WT and Dlk1MatTom/PatVen mice. Error bars
represent standard error (n≥3 samples). Values are normalized to results obtained with
WT cells for the respective tissue.
Development 143: doi:10.1242/dev.138255: Supplementary information
Dev
elo
pmen
t • S
uppl
emen
tary
info
rmat
ion

Fig. S2. Quantification of allele-specific Dlk1 expression in different mouse strains. (A) Flow cytometric analysis of E16.5 mouse lung (first row), skin (middle) and
liver (bottom) of Dlk1MatWT/PatTom and Dlk1MatTom/PatWT reporter mice in a C57BL/6 (B6)
background, compared to F1 crosses with 129/SV-E (129) mice. Gating to identify
Tomato+ cells is shown. (B) Quantification of Tomato+ cells identified as shown in A. (*)
indicates p<0.05 and (**) p<0.005 with a one-way ANOVA and Bonferroni multiple
comparison test (n≥3 samples).
Development 143: doi:10.1242/dev.138255: Supplementary information
Dev
elo
pmen
t • S
uppl
emen
tary
info
rmat
ion

Fig. S3. Measurement of allele-specific Dlk1 reporter gene activity by flow
cytometry. Histograms showing fluorescent intensities of paternal and maternal Dlk1-
Venus expression in E16.5 skin (left panel) and liver (right panel) cells (compare to the
results obtained with the Tomato reporter configuration shown in Fig. 2C).
Development 143: doi:10.1242/dev.138255: Supplementary information
Dev
elo
pmen
t • S
uppl
emen
tary
info
rmat
ion

Fig. S4. Assessment of the impact of reporter insertion on allele-specific Dlk1
expression. (A) Schematic showing the strategy to measure allele-specific Dlk1
expression with qRT-PCR, in the presence or absence of reporter insertion. The black
box represents the 3’ UTR; a white bar, the reporter insertion site; grey arrows, the
primer positions to measure the Dlk1WT allele; and green arrows, primers to measure
the Dlk1Ven allele. In Dlk1MatWT/PatVen mice, paternal expression can be detected with
primers that are specific to the Venus insertion and maternal expression can be
detected with WT primers that flank the insertion site. The WT primers are unable to
amplify the reporter insertion sequence under qRT-PCR conditions, due to its size. In
Dlk1MatVen/PatWT mice, the WT primers detect paternal expression and the Venus primers
detect maternal expression. (B) Representative qRT-PCR measurements showing the
effectiveness of the strategy outlined in C in liver cells. Note that WT primers are unable
to detect any transcript in Dlk1MatVen/PatVen (V/V) cells while Venus primer are unable to
detect transcript in Dlk1WT/WT (-/-) cells. (n = 3 samples). (C) Maternal Dlk1 expression
represented as a percentage of total expression in E16.5 skin, liver and lung by qRT-
PCR. Error bars represent standard error (n≥3 samples).
Development 143: doi:10.1242/dev.138255: Supplementary information
Dev
elo
pmen
t • S
uppl
emen
tary
info
rmat
ion

Fig. S5. Statistical assessment of allele-specific expression and DNA methylation
differences. Mean percentage of LOI Dlk1 expression (upper panel) and mean DNA
methylation at the IG-DMR (lower panel) in matching, independent iPSC cultures
derived in basal conditions, with only AA or with AA, iAlk5 and CHIR99021 (3c). (*)
indicates p<0.05 with a one-way ANOVA and Turkey’s multiple comparison test. Error
bars represent standard error (n≥3).
Development 143: doi:10.1242/dev.138255: Supplementary information
Dev
elo
pmen
t • S
uppl
emen
tary
info
rmat
ion

Fig. S6. Quantification of DNA methylation at the IG-DMR in iPSCs derived in
basal and 3c conditions. (A) Representation of next generation bisulfite sequencing
reads for iPSCs derived by reprogrammed MEFs in basal (left) and 3c (right) conditions.
Each row represents a single read and each column represents a CpG. Methylated
CpGs are indicated by black boxes and unmethylated are white. For clarity purposes,
only the first 60 reads obtained from each sample are shown. (B) Quantification of the
percentage of sequencing reads with high (>75% CpGs methylated), intermediate (25-
75%) or low (<25%) methylation at the IG-DMR. The even distribution of predominantly
methylated and unmethylated DNA in the 3c sample suggests that most cells in this
condition maintained their germline imprinting status. (C) Quantification of the
percentage of cells with LOI in the samples shown in (A) and (B) by flow cytometry.
Development 143: doi:10.1242/dev.138255: Supplementary information
Dev
elo
pmen
t • S
uppl
emen
tary
info
rmat
ion

Fig. S7. Effect of oxygen levels on allele-specific Dlk1 expression during
differentiation. (A) Representative FACS plots of retinoic acid-differentiated
Dlk1MatTom/PatVen iPSCs at p8 that were derived in 3c conditions and cultured in 20%
oxygen and differentiated in 4% (upper panel) or cultured in 4% oxygen and
differentiated in 20% (lower panel). Red gates indicate maternal-only expression (mat),
yellow gates show LOI expression and green indicates MOI. Note the elevated levels of
maternal Dlk1 expression in cultures differentiated in 20% oxygen. (B) Representative
images of cultures corresponding to FACS plots shown in (A), with iPSCs cultured in
20% oxygen and differentiated in 4% (upper row) and cultured in 4% and differentiated
in 20% (bottom row). A yellow arrow points to a group of LOI cells and a red arrow
indicates maternal-only expression. Scale bar = 100 μm.
Development 143: doi:10.1242/dev.138255: Supplementary information
Dev
elo
pmen
t • S
uppl
emen
tary
info
rmat
ion




















