
Preliminary study for detection and classification of swallowing ...
Upload
independentCategory
view
0download
0
J Comp Physiol A (2010) 196:111–122
DOI 10.1007/s00359-009-0498-4ORIGINAL PAPER
Central regulation of the pharyngeal and upper esophageal reXexes during swallowing in the Japanese eel
Takao Mukuda · Masaaki Ando
Received: 24 March 2009 / Revised: 24 November 2009 / Accepted: 8 December 2009 / Published online: 25 December 2009© Springer-Verlag 2009
Abstract We investigated the regulation of the pharyn-geal and upper esophageal reXexes during swallowing ineel. By retrograde tracing from the muscles, the motoneu-rons of the upper esophageal sphincter (UES) were locatedcaudally within the mid-region of the glossopharyngeal-vagal motor complex (mGVC). In contrast, the motoneuronsinnervating the pharyngeal wall were localized mediallywithin mGVC. Sensory pharyngeal Wbers in the vagal nerveterminated in the caudal region of the viscerosensorycolumn (cVSC). Using the isolated brain, we recorded 51spontaneously active neurons within mGVC. Theseneurons could be divided into rhythmically (n = 8) andcontinuously (n = 43) Wring units. The rhythmically Wringneurons seemed to be restricted medially, whereas the con-tinuously Wring neurons were found caudally withinmGVC. The rhythmically Wring neurons were activated bythe stimulation of the cVSC. In contrast, the stimulation ofthe cVSC inhibited Wring of most, but not all the continu-ously Wring neurons. The inhibitory eVect was blocked byprazosin in 17 out of 38 neurons. Yohimbine also blocked
the cVSC-induced inhibition in Wve of prazosin-sensitiveneurons. We suggest that the neurons in cVSC inhibit thecontinuously Wring motoneurons to relax the UES andstimulate the rhythmically Wring neurons to constrict thepharynx simultaneously.
Keywords Swallowing · Esophagus · Medulla oblongata · Vagal nerve · Catecholamines · Japanese eel
AbbreviationsaCSF ArtiWcial cerebrospinal XuidAP Area postremaBBB Blood–brain barrierBDA-10KF Biotinylated dextran amine conjugated
with Xuorescein isothiocyanateChAT Choline acetyltransferaseCNC Commissural nucleus of the CajalcVSC Caudal region of VSCEB Evans blueGVC Glossopharyngeal-vagal motor complexmGVC Mid-region of GVCPBS Phosphate-buVered salineTH Tyrosine hydroxylaseUES Upper esophageal sphincterVSC Viscerosensory column
Introduction
Swallowing is an essential behavior associated with drink-ing or ingestion in all vertebrates, including Wsh. Thisbehavior requires the precisely coordinated control of mus-cles in the oropharyngeal region. Muscular action is coordi-nated by both the central and peripheral nervous systems.The central components involved in swallowing are well
Electronic supplementary material The online version of this article (doi:10.1007/s00359-009-0498-4) contains supplementary material, which is available to authorized users.
T. Mukuda (&) · M. AndoLaboratory of Integrative Physiology, Graduate School of Integrated Arts and Sciences, Hiroshima University, 1-7-1 Kagamiyama, Higashi-hiroshima 739-8521, Japane-mail: [email protected]
Present Address:M. AndoLaboratory of Physiology, Ocean Research Institute, The University of Tokyo, 1-15-1 Minamidai, Tokyo 194-8639, Japan
123

112 J Comp Physiol A (2010) 196:111–122
known in vertebrates, including Wsh and their morphologi-cal organization has been reported in a number of teleostspecies, such as goldWsh (Morita and Finger 1987; Goehlerand Finger 1992) and catWsh (Kanwal and Caprio 1987).The central swallowing reXex circuits are localized in thecaudal medulla. Central sensation from the oropharyngealregion is detected by the dorsal viscerosensory neurons.Conversely, motor innervation of the muscles involved inswallowing originates in the ventromedial visceromotorneurons of the glossopharyngeal and the vagal nerves.Although the direct aVerent connections to the visceromotornuclei are still unclear, anterograde tracing has revealedsome projections to the visceromotor nuclei from the vis-cerosensory neurons (Morita and Finger 1985; Goehler andFinger 1992).
In the Japanese eel, morphological and functional fea-tures of the vagal motor components are relatively wellunderstood, but not much is known about the sensoryreception. Motoneurons of swallowing-associated muscles,such as the pharyngeal wall and upper esophageal sphincter(UES), are viscerotopically arranged in the glossopharyn-geal-vagal motor complex (GVC). These motoneurons arecholinergic (Mukuda and Ando 2003a) and constrict thepharyngeal muscle and UES through acetylcholine (Kozakaand Ando 2003). Ito et al. (2006) in the eel reported thatmotor neurons in GVC Wre spontaneously at high frequencyusing the isolated brain specimens and the Wring activitywas inhibited by catecholamines.
The viscerosensory neurons form a long viscerosensorycolumn (VSC) in the dorsal medulla in the eel and the VSClacks specialized, hypertrophied glossopharyngeal andvagal lobes (Mukuda and Ando 2003b) as in the gray mul-let (Díaz-Regueira and Anadón 1992) and the rainbow trout(Folgueira et al. 2003) although it is well known to possessthe highly specialized, protruded vagal lobe in the goldWsh(Morita and Finger 1987; Goehler and Finger 1992) and thecatWsh (Kanwal and Caprio 1987). Thus, it has been diY-cult to diVerentiate individual nuclei strictly in the eel.However, in general, there has been not much eVort tosystematically study the neurophysiologic features ofthese swallowing-related nuclei, and their connections, inteleosts.
The entrance of esophagus is normally held closed bytonic contraction of the UES. This facilitates breathing inboth Wsh and mammals. The process of swallowing requirescoordination of the upper esophageal relaxation and pha-ryngeal contractile reXexes to transport pharyngeal contentsinto the esophagus. The reXexes are triggered by pharyn-geal stimulation, that is, attachment of pharyngeal contents,such as food and water, to the pharyngeal wall (Meddaet al. 1994). Thus, the central system controlling the UESrelaxation following the sensation of inputs from the phar-ynx is an important component to coordinate swallowing
reXex. However, despite their importance, not much isknown about the central regulation of these reXexes.
To reveal the central systems controlling the pharyngealcontraction and upper esophageal relaxation reXexes in theJapanese eel, we identiWed the motoneurons innervating theUES and pharyngeal wall in the mid-region of GVC(mGVC), and searched candidates of neuronal pathwaysand neurotransmitters regulating the mGVC neurons. Tobetter localize the motoneurons of the UES and pharyngealwall within mGVC than previously reported (Mukuda andAndo 2003a), we injected a retrograde Xuorescent dye,Evans blue (EB), into the concerned muscles, using animproved method. In addition, we also injected biotinylateddextran amine conjugated with Xuorescein isothiocyanate(BDA-10KF) into the vagal nerve bundle distributed in thepharyngeal wall and branchial arches to determine thereception sites for sensory inputs in the caudal medulla. Wethen recorded neuronal activity in mGVC in response toelectric Weld stimulation of the caudal region of viscerosen-sory column (cVSC). Last, we investigated the involvementof catecholamines during the control of motoneuron activityusing the receptor antagonists, prazosin and yohimbine.
Materials and methods
Cultured Japanese eels, Anguilla japonica (n = 49,»200 g), were obtained from a commercial source. Theeels were held in artiWcial seawater aquaria (20°C), withoutfood, for at least 1 week before use.
To study the distribution of the motoneurons innervatingthe pharyngeal wall and UES, we injected a retrograde Xuo-rescent tracer, EB (green excitation, red emission, 1422-52,Kanto Chemical, Tokyo, Japan), into the pharyngeal wall(n = 5) or UES (n = 44). Out of 44 eels injected with EBinto the UES, 7 eels were employed for immunohistochem-istry, 6 were for BDA-10KF staining and the rest 31 werefor electrophysiology. The eel was anesthetized by immer-sion in 0.1% tricaine methanesulfonate (A5040, Sigma, StLouis, MO) dissolved in artiWcial seawater buVered with0.3% NaHCO3. To open a mouth wide and expose the UES,a mouthpiece made by a conical sampling tube (ST-500,BIO-BIK) which the tip was cut (diameter of the cuttingend: 5 mm) was inserted into the oral cavity, 20 �l EB solu-tion (5.0 mg/ml, dissolved in phosphate buVered saline,PBS, pH 7.4) was injected into the left UES by a syringewith a thin needle (33 gages). To inject EB into the pharyn-geal wall, the mouthpiece was not necessary. After injec-tion, the animal was returned to the aquarium and kept forat least 3 days. Some of the eels injected with EB into theUES were also injected with a neuronal tracer, BDA-10KF(SP-1130, Vector, Burlingame, CA), into the pharyngealWbers of vagal nerve (n = 6), which projects to the pharyngeal
123

J Comp Physiol A (2010) 196:111–122 113
wall and branchial arches caudal to the third branchial arch,but not to the UES. Following the injection of EB, the epi-dermal layer of the branchial chamber covered by the oper-culum was removed to expose a vagal nerve trunk under theanesthesia. Under a stereoscopic microscope (MS 5, Leica,Wetzlar, Germany), 20 �l BDA-10KF solution (5.0 mg/mlin PBS) was injected into the vagal nerve via a glass capil-lary by air pressure. After injections, the eels were sutured,and returned to the aquaria to recover for 1 week.
Following the recovery period, the eel was anaesthetizedand perfused transcardially with PBS and followed by 4%paraformaldehyde in 0.1 M phosphate buVer (pH 7.4). Theskull was then fenestrated and immersed in the Wxativeovernight at 4°C. After the Wxation, the brain was isolatedfrom the skull and cryoprotected with 30% sucrose in0.1 M phosphate buVer and embedded. The transverse orhorizontal sections (14-�m thick) were made using a frozenmicrotome (CM1850, Leica) and mounted on MAS-coatedglass slides (S9441, Matsunami Glass, Osaka, Japan). Toexamine the staining by EB, the mounted sections wereimmediately photographed under a Xuorescent microscope(Eclipse E600, Nikon, Tokyo) equipped with a digital cam-era (DXM1200F, Nikon). Following this, the sections werefrozen at ¡80°C until further use.
The catecholaminergic neurons as well as the choliner-gic neurons were visualized by immunohistochemistry. Allthe operations without notation of the condition were car-ried out at room temperature. After rinsing three times inPBS, the sections were treated with a blocking solutioncontaining 5% normal donkey serum, 0.1% Triton X-100and 0.05% Tween 20 in PBS for 1 h. A rabbit polyclonalantibody raised against tyrosine hydroxylase (TH) from ratphenochromocytoma (1:500, AB152, Chemicon, Teme-cula, CA) and a goat polyclonal antibody raised againstcholine acetyltransferase (ChAT) from human placental(1:500, AB144, Chemicon) were applied for 24–48 h at4°C. After rinsing three times in PBS, the sections weretreated with the secondary antibodies, Cy2-conjugated don-key anti-rabbit IgG (1:500, 711-225-152, Jackson, WestGrove, PA) and Cy3-conjugated donkey anti-goat IgG(1:500, 705-165-147, Jackson), together with 4�,6-diami-dino-2-phenylindoledihydrochloride (1:1,000, D9564,Sigma) to counterstain, were applied for 2 h. After rinsingthree times in PBS, the sections were coverslipped. Thesections were examined and photographed under a Xuores-cent microscope (BZ-9000, Keyence, Osaka, Japan). Pri-mary antibodies employed in the present study are widelyused for immunohistochemistry in Wsh: anti-TH antibody(e.g. Sueiro et al. 2003 in dogWsh; Barreiro-Iglesias et al.2008 in lamprey; Mukuda et al. 2005 in Japanese eel;Castro et al. 2008 in trout; Ma 1994 in zebraWsh); anti-ChAT antibody (e.g. Anadón et al. 2000 in dogWsh; Pombalet al. 2001, 2003 in lamprey; Clemente et al. 2004; Arenzana
et al. 2005 in zebraWsh), and we did not observe any immu-noreactivity in the absence of the primary antibodies.
To intensify the staining of BDA-10KF, we developedevery other section of the caudal medulla with ABC kit(PK-6100, Vector). The sections were pretreated with PBScontaining 0.3% Triton X-100 and 0.05% Tween 20 for10 min. The sections were immersed in 5% H2O2 in metha-nol for 30 min, rinsed three times with PBS, and then incu-bated with the avidin–biotin complex (1:100) overnight at4°C. After rinsing, the sections were incubated with 0.05%3,3�-diaminobenzidine tetrahydrochloride (Nakarai Tesque,Tokyo, Japan) in phosphate buVer for 30 min to intensifycolorization. The sections were then treated with 0.05%3,3�-diaminobenzidine tetrahydrochloride in phosphatebuVer containing 0.01% H2O2 and 0.04% NiCl2 for 5 minin dark. Finally, the sections were rinsed, dehydrated,cleared and coverslipped. The sections were examined witha light microscope (Eclipse E600, Nikon). The remainingsections were stained with anti-TH antibody and examinedunder the Xuorescent microscope (BZ-9000, Keyence)according to the immunohistochemical procedures asdescribed before.
To prepare the specimen for electrophysiology, wedecapitated the eel injected with EB into the UES (n = 31).The muscular tissue around the skull was removed and thecranium was placed in artiWcial cerebrospinal Xuid (aCSF:124 mM NaCl, 5 mM KCl, 1.3 mM MgSO4, 1.2 mMCaCl2, 1.2 mM NaHPO4, 26 mM NaHCO3, 10 mM glu-cose) during the operations. All the bones, with the excep-tion of the palatal bone, were removed to expose the brain.The whole brain (between the olfactory bulb and therostralmost spinal cord), associated with the palatal bone,was tied to an acryl plate and set in the experimental cham-ber (»2-ml volume). The chamber was continuouslyperfused with aCSF (1.0 ml/min) and maintained at 20°C.A gas mixture of 95% O2 and 5% CO2 (pH 7.4) was bubbledinto the aCSF in the reservoir.
To record neuronal activity, we used a tungsten electrode(5-�m tip diameter; KU207-010, Unique Medical, Tokyo,Japan). The electrode was slowly inserted into mGVC torecord extracellular spiking activity. An Ag–AgCl referenceelectrode was placed in aCSF in the chamber. The neuronalactivity of the mGVC neurons was Wltered at 15–3 k Hz andthen ampliWed. The signals were digitized by an AD con-verter (Power Lab 2/25, ADInstruments, Bella Vista, Austra-lia) at a sampling rate of 40 k Hz. We recorded the electricalactivity of mGVC in an area ranging from 0 to 280 �m ros-tral to the obex. This area is dominated by motoneurons thatinnervate the UES (Mukuda and Ando 2003a). These neu-rons typically Wre spontaneously and show large amplitudespikes (>75 �V) (Ito et al. 2006).
To apply electric Weld stimulation, we used a bipolarelectrode manufactured by combining the two tungsten
123

114 J Comp Physiol A (2010) 196:111–122
electrodes mentioned above. The distance between the tipsof two electrodes was 10–30 �m. The stimulating electrodewas placed on the dorsal surface of the medulla at the levelof the obex and immediately above the cVSC, which isipsilateral to the recording site. As a stimulus, three electri-cal pulses (0.5 mA, 0.1 ms) were applied at 10 Hz.
We examined the eVects of two antagonists for catechol-amine receptors (prazosin and yohimbine) on the action ofcVSC stimulation. Drugs (1 �M Wnal concentration) wereadded to the aCSF bath via a series of manually operatedvalves. Prazosin was used as a nonspeciWc catecholaminereceptor antagonist in the eels, and yohimbine as a speciWcadrenoceptor antagonist (Ito et al. 2006).
After the electrophysiological recordings, the recordingsite was checked histologically (Supplement Fig. 2). Eachbrain was Wxed in 4% paraformaldehyde in 0.1 M phos-phate buVer (pH 7.4) overnight at 4°C, cryosectioned andimmunostained as described before.
OZine analyses of the digitized signals were made byChart Pro (ADInstruments). The single units were sepa-rated based on the distribution pattern of the amplitude andwidth of spikes during a 300-s period. Spikes with anamplitude <75 �V were omitted. To evaluate the eVect ofWeld stimulation of cVSC on mGVC, the mean spike countsduring a 5-s period at 30 s before the stimulation, immedi-ately after the stimulation, and 30 s after the stimulationwere compared. The trial was replicated Wve times in eachexperiment. Comparison of the mean spike count duringeach time epoch was performed by Tukey–Kramer post hoctest. The diVerences were considered signiWcant atP < 0.01.
Results
Motoneurons innervating the pharyngeal wall and UES
Following the injection of EB into the pharyngeal wall orUES, the neurons in mGVC which was ipsilateral to theinjection site were stained (Fig. 1a–d). As shown in a sum-mary diagram of the rostrocaudal distribution of motoneu-rons stained with EB, the neurons innervating thepharyngeal wall were distributed in the mid-region ofmGVC (Fig. 1f). In contrast, the neurons innervating theUES were distributed in more caudal region within mGVC(Fig. 1f). In the region between 238 and 280 �m rostral tothe obex, both motoneurons were co-existed (Fig. 1f). Inthe case of eels injected with BDA-10KF into the pharyn-geal branch of vagal nerve, similarly, the BDA-10KF-posi-tive motoneurons were located more rostral to EB-positiveneurons within mGVC (data not shown). All the EB-posi-tive neurons were ChAT immunoreactive (Fig. 1a, b), indi-cating that they are cholinergic.
TH-immunoreactive Wbers were found abundantlyaround the neurons innervating the UES (Fig. 1b–d) and toa lesser extent around the neurons innervating the pharyn-geal wall (Fig. 1a). A summary of the rostrocaudal distribu-tion of TH-immunoreactive neurons is shown in Fig. 1f.Although TH-immunoreactive Wbers were relatively wide-spread throughout the medulla, the strongest signal wasobserved around the mGVC neurons innervating the UESand the dorsal region along the wall of the fourth ventricle(Fig. 1c, d). The cell bodies of these TH-immunoreactiveWbers were localized in the cVSC (Fig. 1e). The TH-immu-noreactive cell bodies in cVSC were observed in the dorsalmedulla from the paired region (Supplement Fig. 1c–f) tothe obecular region (Supplement Fig. 1g–i) in cVSC. Neu-rons of the area postrema (AP) were also TH immunoreac-tive in the postobecular region, but their cell bodies werelocated outside the blood–brain barrier (BBB) (SupplementFig. 1j–p) and their Wbers were entered into the caudalregion of GVC, not into mGVC.
Sensory inputs into the dorsal medulla from the pharyngeal region
BDA-10KF-positive nerve Wbers were traced to the ipsilat-eral, but not contralateral, caudal region of the dorsalmedulla. The BDA-10KF-positive Wbers were divided intotwo types: (1) those showing thin Wbers alone (Fig. 2a, b)and (2) those forming tubercles at the end of the thin Wbers incVSC (Fig. 2a, d). The tubercles were tadpole shaped(Fig. 2d) and 4–8 �m in width. There were BDA-10KF-pos-itive cell bodies in the nodose ganglion (Fig. 2c), and theirWbers entered the medulla via the vagal nerve, coursed alongthe dorsolateral margin of the DV, and terminated in thecVSC (Fig. 2b, d). Figure 2e illustrates the TH immunostain-ing of the section immediately rostral to the one shown inFig. 2d. The cVSC contained a large number of TH-immu-noreactive neurons (Fig. 2e). The section shown in Fig. 2b isalmost same sectioning level of that in Fig. 2e. In the pairedregion of VSC, TH-immunoreactive neurons were predomi-nant more caudally (Supplement Fig. 1d–f), compared ros-trally (Supplement Fig. 1a–c). These suggest that the sensoryinputs from the pharyngeal region are transmitted into thecatecholaminergic neurons in cVSC exclusively.
EVect of viscerosensory activation on spontaneous activities of the mGVC
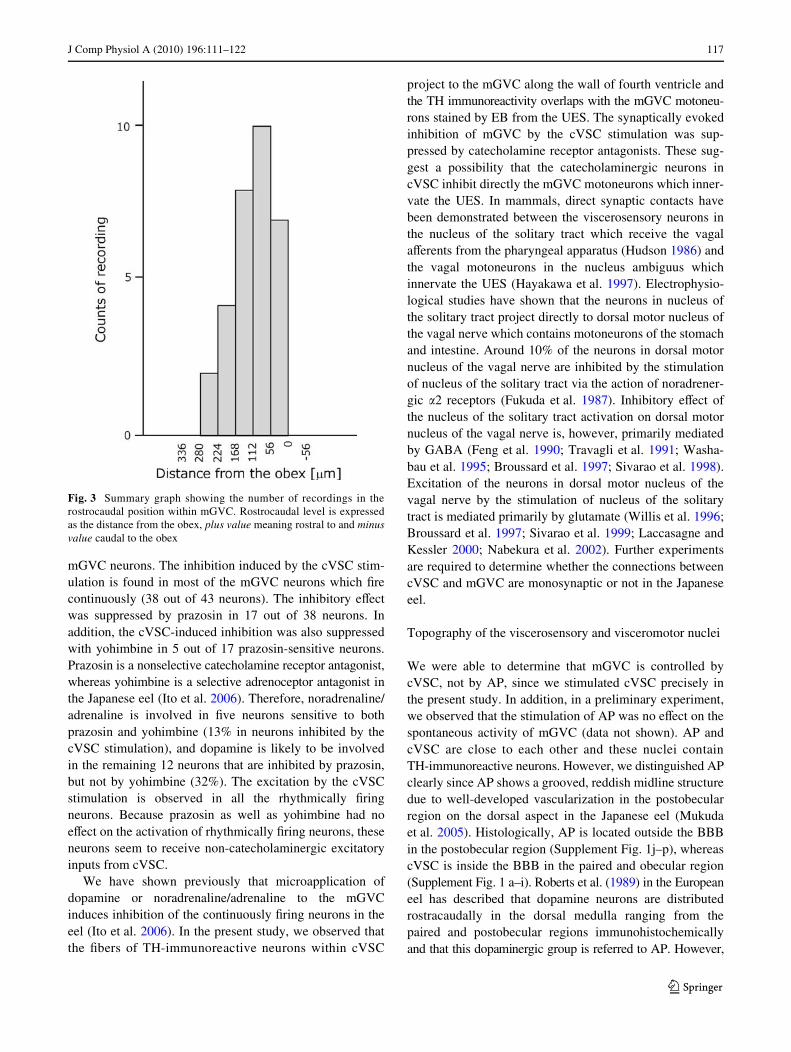
We recorded spontaneous neuronal activities in the mGVCneurons and examined the eVects of the stimulation ofcVSC in the eels which were injected EB into the UES atleast 3 days before the electrophysiological recording. Therecording positions are summarized in Fig. 3. The majorityof the recordings were made within the caudal region of
123

J Comp Physiol A (2010) 196:111–122 115
mGVC, which contains the neurons innervating the UES.We also made two recordings in the mid-region of mGVC.In this region, the motoneurons innervating the UES or thepharynx were co-located (Figs. 1f, 3).
We obtained 31 multiunit extracellular recordings frommGVC. The neurons could be divided into two categoriesbased on the spontaneous Wring patterns. The one categoryincludes neurons that show rhythmical activities of an inter-val of several seconds (Fig. 4a). The other includes the neu-rons that Wre continuously (Fig. 4b). To obtain single-unitactivity, we tried to classify the units in 31 extracellularrecordings as described in “Materials and methods” anddiscriminated 51 single units (Fig. 4c).
Eight single units displayed a “rhythmic Wring” activityand were found in the mid-region of mGVC (+224 to +280
from the obex). When the electric Weld stimulation of cVSCwas applied during the interval of the spontaneous burstinginterval, these neurons were activated immediately(Fig. 5a). As a result, the next burst was signiWcantlydelayed. However, spontaneous Wring pattern was resumedrather quickly and the burst interval was restored to pre-stimulation levels. To investigate the involvement of thecatecholamines in the excitatory eVect by the cVSC stimu-lation, similar experiments were repeated in the presence ofprazosin, a nonselective catecholamine receptor antagonistin eels or yohimbine, a selective adrenoceptor antagonist(Ito et al. 2006). The excitatory action of the cVSC stimula-tion on the rhythmically bursting neurons was not aVectedby these catecholamine receptor antagonists (data notshown).
Fig. 1 Motoneurons innervat-ing the pharyngeal wall and up-per esophageal sphincter (UES) within glossopharyngeal-vagal motor complex (GVC) and cate-cholaminergic neurons within the caudal region of viscerosen-sory column (cVSC) in the eel medulla. Representative trans-verse sections stained retro-gradely with Evans blue (EB red) injected into the pharyngeal wall (a) and UES (b). The sec-tions were also immunostained with anti-choline acetyltransfer-ase (ChAT blue) and anti-tyro-sine hydroxylase (TH green) antibodies. Fluorescent image of EB alone (left) and a merged im-age of EB, ChAT, and TH (right). To intensify contrast, Xuorescence of ChAT immuno-reactivity is expressed with blue color. c Low magniWcation view of the transverse section stained with anti-TH antibody (green) and EB (red) injected into the UES. MagniWed view of the mid-region of GVC (mGVC) (d) and cVSC (e) areas, enclosed by squares in c. f Summary of dis-tribution of the motoneurons stained with EB and TH-immu-noreactive neurons rostrocaudal-ly in the eel caudal medulla. The rostrocaudal level is expressed as the distance from the obex, plus value meaning rostral to and minus value caudal to the obex. Number of the eel examined in the analysis is indicated in parentheses. AP area postrema, cGVC caudal region of GVC, rGVC rostral region of GVC, 4V fourth ventricle
123

116 J Comp Physiol A (2010) 196:111–122
Forty-three single units displayed a “continuous Wring”activity. The neurons showing this type of activity wererecorded throughout the caudal region of the mGVC.Figure 5b illustrates the eVect of the cVSC stimulation onmGVC activity. Spontaneous Wring activity was transientlyinhibited immediately after the stimulation of cVSC (Fig. 5b).The single-unit analysis of the recorded activities is presentedin Fig. 5c. The number of spikes was decreased in all thedetected units (a–d) immediately after the stimulation(Fig. 5c). This inhibitory eVect was suppressed in the unit a, b,or d, but not in c by prazosin (Fig. 5c). Yohimbine suppressedthe inhibitory eVect in the unit a and b, but not in the unit cand d (Fig. 5c). The eVects of prazosin and yohimbine werecompletely reversed by washing out of the drugs (Fig. 5c).
The inhibitory eVect of cVSC stimulation on mGVC issummarized in Fig. 6. Forty-three units were from the con-
tinuously Wring neurons. Of these, 38 were inhibited by thecVSC stimulation and none of them were activated by thecVSC stimulation (Fig. 6). We observed no eVect in Wveunits (Fig. 6). Seventeen of the cVSC stimulation-respon-sive units were sensitive to prazosin (Fig. 6). Of these, Wveunits were also sensitive to yohimbine while the remainderswere insensitive (Fig. 6).
Discussion
Control in neuronal activities of the GVC
This is the Wrst report to show that the teleost mGVC iscontrolled by inputs from cVSC. Our results suggest thatcVSC exerts both excitatory and inhibitory control of the
Fig. 2 Sensory neurons projecting to the pharyngeal wall and bran-chial arches in the eel. a Schematic drawing of a transverse aspect inthe eel caudal medulla (Mukuda and Ando 2003b). Neurons traced bya biotinylated dextran amine conjugated with Xuorescein isothiocya-nate (BDA-10KF) injected into the vagal nerve projecting to the pha-ryngeal area. BDA-10KF-positive Wbers were observed along thedorsal margin of descending trigeminal root (DV) (n = 3) (at 56 �mrostral to the obex) (b), whereas their cell bodies were located in thenodose ganglion within the glossopharyngeal-vagal nerve (IX–X) (c).BDA-10KF-positive cell bodies in the nodose ganglion are denoted by1, 2, and 3. Asterisk indicates a region containing a predominance ofBDA-10KF-positive Wbers. Each region is shown by hatching in a. dBDA-10KF-positive Wbers with tubercles in cVSC (at 56 �m rostral to
the obex) (n = 3). The inset shows a highly magniWed view of thecVSC region enclosed by a square in the main diagram. The eel wasinjected concurrently with EB into the UES, and BDA-10KF into thevagal nerve projecting to the pharyngeal wall and branchial archesexcept for third branchial arch. e Immunohistochemistry using anti-THantibody in the section immediately rostral to d (at 70 �m rostral to theobex). Number in d and e is given for EB-positive neurons withinmGVC. f MagniWed view of the EB-positive neurons (red) numbered4, 5, and 6 within the mGVC. Cven commissura ventralis rhomben-cephali, MLF medial longitudinal fascicle, RF reticular formation, RInfraphe nucleus inferior, SGT secondary gustatory tract, TSV vestibulo-spinal tract
123
J Comp Physiol A (2010) 196:111–122 117
mGVC neurons. The inhibition induced by the cVSC stim-ulation is found in most of the mGVC neurons which Wrecontinuously (38 out of 43 neurons). The inhibitory eVectwas suppressed by prazosin in 17 out of 38 neurons. Inaddition, the cVSC-induced inhibition was also suppressedwith yohimbine in 5 out of 17 prazosin-sensitive neurons.Prazosin is a nonselective catecholamine receptor antagonist,whereas yohimbine is a selective adrenoceptor antagonist inthe Japanese eel (Ito et al. 2006). Therefore, noradrenaline/adrenaline is involved in Wve neurons sensitive to bothprazosin and yohimbine (13% in neurons inhibited by thecVSC stimulation), and dopamine is likely to be involvedin the remaining 12 neurons that are inhibited by prazosin,but not by yohimbine (32%). The excitation by the cVSCstimulation is observed in all the rhythmically Wringneurons. Because prazosin as well as yohimbine had noeVect on the activation of rhythmically Wring neurons, theseneurons seem to receive non-catecholaminergic excitatoryinputs from cVSC.
We have shown previously that microapplication ofdopamine or noradrenaline/adrenaline to the mGVCinduces inhibition of the continuously Wring neurons in theeel (Ito et al. 2006). In the present study, we observed thatthe Wbers of TH-immunoreactive neurons within cVSC
project to the mGVC along the wall of fourth ventricle andthe TH immunoreactivity overlaps with the mGVC motoneu-rons stained by EB from the UES. The synaptically evokedinhibition of mGVC by the cVSC stimulation was sup-pressed by catecholamine receptor antagonists. These sug-gest a possibility that the catecholaminergic neurons incVSC inhibit directly the mGVC motoneurons which inner-vate the UES. In mammals, direct synaptic contacts havebeen demonstrated between the viscerosensory neurons inthe nucleus of the solitary tract which receive the vagalaVerents from the pharyngeal apparatus (Hudson 1986) andthe vagal motoneurons in the nucleus ambiguus whichinnervate the UES (Hayakawa et al. 1997). Electrophysio-logical studies have shown that the neurons in nucleus ofthe solitary tract project directly to dorsal motor nucleus ofthe vagal nerve which contains motoneurons of the stomachand intestine. Around 10% of the neurons in dorsal motornucleus of the vagal nerve are inhibited by the stimulationof nucleus of the solitary tract via the action of noradrener-gic �2 receptors (Fukuda et al. 1987). Inhibitory eVect ofthe nucleus of the solitary tract activation on dorsal motornucleus of the vagal nerve is, however, primarily mediatedby GABA (Feng et al. 1990; Travagli et al. 1991; Washa-bau et al. 1995; Broussard et al. 1997; Sivarao et al. 1998).Excitation of the neurons in dorsal motor nucleus of thevagal nerve by the stimulation of nucleus of the solitarytract is mediated primarily by glutamate (Willis et al. 1996;Broussard et al. 1997; Sivarao et al. 1999; Laccasagne andKessler 2000; Nabekura et al. 2002). Further experimentsare required to determine whether the connections betweencVSC and mGVC are monosynaptic or not in the Japaneseeel.
Topography of the viscerosensory and visceromotor nuclei
We were able to determine that mGVC is controlled bycVSC, not by AP, since we stimulated cVSC precisely inthe present study. In addition, in a preliminary experiment,we observed that the stimulation of AP was no eVect on thespontaneous activity of mGVC (data not shown). AP andcVSC are close to each other and these nuclei containTH-immunoreactive neurons. However, we distinguished APclearly since AP shows a grooved, reddish midline structuredue to well-developed vascularization in the postobecularregion on the dorsal aspect in the Japanese eel (Mukudaet al. 2005). Histologically, AP is located outside the BBBin the postobecular region (Supplement Fig. 1j–p), whereascVSC is inside the BBB in the paired and obecular region(Supplement Fig. 1 a–i). Roberts et al. (1989) in the Europeaneel has described that dopamine neurons are distributedrostracaudally in the dorsal medulla ranging from thepaired and postobecular regions immunohistochemicallyand that this dopaminergic group is referred to AP. However,
Fig. 3 Summary graph showing the number of recordings in therostrocaudal position within mGVC. Rostrocaudal level is expressedas the distance from the obex, plus value meaning rostral to and minusvalue caudal to the obex
123

118 J Comp Physiol A (2010) 196:111–122
Fig. 4 Extracellular recordings from the mGVC neurons. Spon-taneous activity of mGVC neu-rons that Wred: a at a multisecond burst interval (“rhythmically Wring”) or b con-tinuously (“continuously Wr-ing”). Solid circle in the rhythmically Wring recording indicates the timing of spike appearance. c Single-unit dis-crimination of the neuronal activity shown in b. Four diVer-ent single units, denoted by a–d, were discriminated based on the width-amplitude plot (left) and frequency (right) during a 300-s period of spontaneous activity
Fig. 5 EVect of cVSC stimulation on the mGVC neurons. Activationof the rhythmically Wring (a) and inhibition of the continuously Wring(b) neurons in response to the cVSC stimulation. Arrow indicates theinitiation of Weld stimulation in cVSC (0.5 mA, 3 trains of 0.1-ms pulseduration at 10 Hz). Timing of spike appearance in the trace of therhythmically Wring neurons is indicated by solid circle (before stimu-lation) and solid triangle (after stimulation). Open circle indicatesthe putative timing of spike appearance if stimulation is not applied.
c Histogram showing the mean spike number in normal aCSF (control,wash out), in aCSF per 1 �M prazosin (Prazosin), and in aCSF per1 �M yohimbine (Yohimbine) in each unit shown in b. Mean spikenumber was calculated over Wve trials and was compared among a 5 speriod 30 s before (pre), immediately after (stim), and 30 s after (post)cVSC stimulation. Asterisk indicates signiWcant diVerences (Tukey–Kramer test, P < 0.01). Row data in the presence of prazosin or yohim-bine are shown on the right
123

J Comp Physiol A (2010) 196:111–122 119
this assignment seems to contain both AP and cVSC com-ponents since the postobecular region lacks the BBB.
In the eel VSC, the vagal lobe and commissural nucleusof the Cajal (CNC) are not discriminated strictly yet and thehistological assignment is confused since the eel VSCforms a rostrocaudal long column containing the compo-nents of vagal lobe and CNC as in the gray mullet(Díaz-Regueira and Anadón 1992) and the rainbow trout(Folgueira et al. 2003). Roberts et al. (1989) has found vagallobe in the rostral region of VSC at which the cerebellarcrest emerges in the European eel, while Ito et al. (2006) hasregarded the paired region of cVSC as vagal lobe, whichshows TH immunoreactivity in the Japanese eel. Mukudaand Ando (2003b) has referred to the postobecular regionas CNC, where nerve Wbers are distributed, but have notdescribed origin of the Wbers in the Japanese eel. IngoldWsh, on the other hand, these nuclei are clearlydistinguished and their neurohistological features are welldocumented. The vagal lobe is hypertrophied dorsolaterallyin the paired region and CNC is found in the commissuralregion exclusively (Morita and Finger 1987; Goehler andFinger 1992). CNC is known to receive viscerosensoryinputs from the pharynx (Morita and Finger 1987; Goehlerand Finger 1992) and is TH immunopositive (Morita andFinger 1987). In addition, CNC has been reported to con-nect with vagal motoneurons in goldWsh (Morita and Finger1985; Goehler and Finger 1992). In the present study, wedemonstrated that the vagal aVerent from pharynx termi-nates in the paired region of cVSC which shows TH immuno-reactivity (Fig. 2b, d, e) and that cVSC controls the vagalmotoneurons. In addition, TH-immunoreactive somata arelocalized within a rostrocaudally short range (ca. 84 �m)between the paired and commissural regions (Fig. 1f;
Supplement Fig. 1c–i). Therefore, in the eel, it may be rea-sonable that CNC is comparable to the region including thecommissural and paired regions of cVSC, but not postobe-cular region, and that vagal lobe corresponds to the rostralregion of VSC as described in European eel (Roberts et al.1989).
Some of BDA-10KF-positive Wbers end emerge astubercles in cVSC in the tracing study. Size of the tuberclesis smaller (4–8 �m) than the cell body of TH-immunoreac-tive neuron (15–20 �m). In lamprey, electrosensory nerveforms extraordinarily large terminal (10–30 �m), as well asnormal-sized terminal (1–3 �m) in the dorsal octavolater-alis nucleus (Kishida et al. 1988; Koyama et al. 1993).Shape of the large terminal is circular or ellipsoidal(Koyama et al. 1993). The size and localization are diVerentbetween the eel and lamprey, but the shape of tubercleobserved in the eel is similar to that of large terminal of thelamprey. Although Wne analyses are needed, the tubercularstructures seen in the eel VSC may be the terminals ofvagal nerve from the pharynx.
Viscerotopic arrangement of the mGVC, noted in ourprevious study (Mukuda and Ando 2003a), was conWrmedin the present study. We were also able to determine thedistribution of the motoneurons that innervate the pharyn-geal wall or UES much Wner scale than previously using animproved method of the injection into UES in the presentstudy. Longitudinally narrower distribution of the motoneu-rons of UES was found in the present study, compared pre-viously. This may be caused by the diVusion of EB injectedinto the UES as well as unexpected variation of injectionsite within the UES. By inserting a mouthpiece into oralcavity to hold mouth opening widely and expose the UES,it became possible to inject the dye into a target point in theUES correctly in the present study. The method for injec-tion into the UES is advantageous to discriminate the moto-neurons which innervate the UES and those whichinnervate the other muscles adjacent to the UES, since weobserved in an individual animal that the mGVC motoneu-rons stained with BDA-10KF injected into the vagal nerveprojecting to the pharyngeal wall and branchial arches arenot labeled with EB injected into the UES (Fig. 2d–f).
A plausible function of the neuronal circuits on swallowing
In the Japanese eel, Xow of ventilatory water across thegills is archived by combined actions of an increased intra-pharyngeal pressure and UES closure. To generate ventila-tion cycle, rhythmic pharyngeal and continuous UEScontractions are required. The rhythmic muscular contrac-tion is generated by the rhythmic Wring of the muscle moto-neurons (Burleson and Smith 2001). Although thecontraction of UES, the motoneurons of UES would Wreand release acetylcholine at the nerve endings continuously.
Fig. 6 Summary of the eVect of the cVSC stimulation on the continu-ously Wring mGVC neurons. Mean spike number during a 5 s periodeither 30 s before (pre circle), immediately after (stim triangle), or 30 safter (post rectangle) cVSC stimulation. Data are the mean of Wve trialsfor each unit. We compared the mean number of spikes between units,as in Fig. 5c
123

120 J Comp Physiol A (2010) 196:111–122
The pharyngeal wall muscle and UES in the eel are striated,and are constricted by action of acetylcholine (Kozaka andAndo 2003). We demonstrated that the motoneurons ofpharyngeal wall muscle and UES are located in the mGVCand are ChAT immunoreactive. When compared with theresults of retrograde tracing and electrophysiological exper-iments, distribution of the motoneurons of UES seem to becomparable to that of the continuously Wring neuronswithin mGVC. On the other hand, restricted distribution ofthe rhythmically Wring neurons is located in the mid-regionof mGVC where the motoneurons of pharyngeal wall andUES are co-existed.
Assuming that the neuronal activity recorded in the iso-lated brain represents that of an intact brain, we can con-clude that the continuously Wring motoneurons innervatethe UES and the rhythmically Wring motoneurons innervatethe pharyngeal wall. The diVering responsiveness of therhythmically and continuously Wring motoneurons to thecVSC stimulation forms the basis for explaining the regula-tion of the pharyngeal and UES reXexes during swallowingin the eel (Fig. 7). During swallowing, the cVSC is acti-vated by vagal inputs from the pharynx, which are initiatedby physical stimuli, such as attachment of food or water tothe pharyngeal wall. The stimulation of cVSC likely mim-ics neuronal inputs from the pharynx. To transport pharyn-geal contents into the esophagus (i.e., facilitation ofswallowing), cVSC relays signals to the motoneurons toconstrict pharynx and to relax UES, simultaneously. There-fore, cVSC may coordinate the pharyngeal and upperesophageal muscular actions during swallowing.
The entrance to the esophagus is generally held closed bythe contraction of UES in mammals. Hence, the relaxationof UES and contraction of pharynx are necessary to archiveswallowing as in the eel. Mammalian UES is composed ofseveral muscles including the cricopharyngeal muscle andaction of the muscle is the most critical for proper function-ing of UES. During the relaxation of UES, continuous Wringof the cricopharyngeal muscle ceases transiently (Shippet al. 1970; Asoh and Goyal 1978) and the break in Wringmay be induced by the inhibition of active motoneurons thatinnervate the cricopharyngeal muscle (Doty 1968; Zoun-grana et al. 1997). The cricopharyngeal muscle is innervatedby the nucleus ambiguus ipsilaterally (Bieger and Hopkins1987; Kitamura et al. 1991; Kobler et al. 1994; Bao et al.1995) and constricted by acetylcholine (Malmberg et al.1991). Nucleus ambiguus receive the projections fromnucleus of the solitary tract (Cunningham and Sawchenko2000) which contains catecholaminergic neurons, i.e. A2noradrenergic/C2 adrenergic neurons (Hollis et al. 2004).The cricopharyngeal muscle is commonly derived from thebranchial arches and is composed of striated muscles inmammals and Wsh. The similarities in ontogeny, motoneu-ron innervation, and catecholamine sensitivity between theeel UES and mammalian cricopharyngeal muscle suggestthat the mechanisms controlling UES action are similaramong both Wsh and mammals. Thus, our results may beuseful in understanding the origin of central regulation ofthe pharyngeal and UES reXexes in mammals.
In conclusion, the cVSC activates rhythmically Wringneurons and inhibits continuously Wring neurons in the eel
Fig. 7 Summary diagram of the functional arrangement of mGVC neurons
123

J Comp Physiol A (2010) 196:111–122 121
mGVC. Inhibition is mediated, in part, by catecholamines.Our results demonstrate the functional signiWcance of theconnections between the viscerosensory and visceromotorneurons previously reported in teleosts. Based on this, wepropose a model for the neuronal regulation on swallowingin Wsh, which may also be useful in understanding the ori-gin of central regulation of the pharyngoesophageal stageof swallowing in mammals.
Acknowledgments The present study was supported, in part, bySatake Foundation and by Grants-in-Aid for ScientiWc Research (C)no. 19570069 from the Ministry of Education, Culture, Sports, Scienceand Technology, Japan. Animal usage and all experimental proceduresin the present study were approved by the Committee for AnimalExperimentation of Hiroshima University and meet the guidelines ofthe Japanese Association for Laboratory Animal Science.
References
Anadón R, Molist P, Rodríguez-Moldes I, López JM, Quintela I,Cerviño MC, Barja P, González A (2000) Distribution of cholineacetyltransferase immunoreactivity in the brain of an elasmo-branch, the lesser spotted dogWsh (Scyliorhinus canicula). J CompNeurol 420:139–170
Arenzana FJ, Clemente D, Sánchez-González R, Porteros A, Aijón J,Arévalo R (2005) Development of the cholinergic system in thebrain and retina of the zebraWsh. Brain Res Bull 66:421–425
Asoh R, Goyal RK (1978) Manometry and electromyography of theupper esophageal sphincter in the opossum. Gastroenterology74:514–520
Bao X, Wiedner EB, Altschuler SM (1995) Transsynaptic localizationof pharyngeal premotor neurons in rat. Brain Res 696:246–249
Barreiro-Iglesias A, Villar-Cerviño V, Villar-Cheda B, Anadón R,Rodicio MC (2008) Neurochemical characterization of sea lampreytaste buds and aVerent gustatory Wbers: presence of serotonin, cal-retinin, and CGRP immunoreactivity in taste bud bi-ciliated cellsof the earliest vertebrates. J Comp Neurol 511:438–453
Bieger D, Hopkins DA (1987) Viscerotopic representation of the upperalimentary tract in the medulla oblongata in the rat: the nucleusambiguus. J Comp Neurol 262:546–562
Broussard DL, Li H, Altschuler SM (1997) Colocalization of GABA(A) and NMDA receptors within the dorsal motor nucleus of thevagal nerve (DMV) of the rat. Brain Res 763:123–126
Burleson ML, Smith RL (2001) Central nervous control of gill Wlamentmuscles in channel catWsh. Respir Physiol 126:103–112
Castro A, Becerra M, Anadón R, Manso MJ (2008) Distribution of cal-retinin during development of the olfactory system in the browntrout, Salmo trutta fario: comparison with other immunohisto-chemical markers. J Chem Neuroanat 35:306–316
Clemente D, Porteros A, Weruaga E, Alonso JR, Arenzana FJ, AijónJ, Arévalo R (2004) Cholinergic elements in the zebraWsh centralnervous system: histochemical and immunohistochemical analy-sis. J Comp Neurol 474:75–107
Cunningham ET Jr, Sawchenko PE (2000) Dorsal medullary pathwayssubserving oromotor reXexes in the rat: implications for the cen-tral neural control of swallowing. J Comp Neurol 417:448–466
Díaz-Regueira S, Anadón R (1992) Central projections of the vagusnerve in Chelon labrosus Risso (Teleostei, O. Perciformes). BrainBehav Evol 40:297–310
Doty RW (1968) Neural organization of deglutition. In: Code CF (ed)Handbook of physiology, Sect. 6, vol IV. American PhysiologicalSociety, Washington DC, pp 1861–1902
Feng HS, Lynn RB, Han J, Brooks FP (1990) Gastric eVects of TRHanalogue and bicuculline injected into dorsal motor vagal nucleusin cats. Am J Physiol 259:G321–G326
Folgueira M, Anadón R, Yáñez J (2003) Experimental study of theconnections of the gustatory system in the rainbow trout,Oncorhynchus mykiss. J Comp Neurol 465:604–619
Fukuda A, Minami T, Nabekura J, Oomura Y (1987) The eVects ofnoradrenaline on neurones in the rat dorsal motor nucleus of thevagus, in vitro. J Physiol 393:213–231
Goehler LE, Finger TE (1992) Functional organization of vagal reXexsystems in the brain stem of the goldWsh, Carassius auratus.J Comp Neurol 319:463–478
Hayakawa T, Zheng JQ, Yajima Y (1997) Direct synaptic projectionsto esophageal motoneurons in the nucleus ambiguus from the nu-cleus of the solitary tract of the rat. J Comp Neurol 381:18–30
Hollis JH, Lightman SL, Lowry CA (2004) Integration of systemic andvisceral sensory information by medullary catecholaminergic sys-tems during peripheral inXammation. Ann NY Acad Sci 1018:71–75
Hudson LC (1986) The origins of innervation of the canine caudal pha-ryngeal muscles: an HRP study. Brain Res 374:413–418
Ito S, Mukuda T, Ando M (2006) Catecholamines inhibit neuronalactivity in the glossopharyngeal-vagal motor complex of the Jap-anese eel: signiWcance for controlling swallowing water. J ExpZool 305:499–506
Kanwal JS, Caprio J (1987) Central projections of the glossopharyn-geal and vagal nerves in the channel catWsh, Ictalurus punctatus:clues to diVerential processing of visceral input. J Comp Neurol264:216–230
Kishida R, Koyama H, Goris RC (1988) Giant lateral-line aVerent ter-minals in the electroreceptive dorsal nucleus of lampreys. Neuro-sci Res 6:83–87
Kitamura S, Ogata K, Nishiguchi T, Nagase Y, Shigenaga Y (1991)Location of the motoneurons supplying the rabbit pharyngealconstrictor muscles and the peripheral course of their axons: astudy using the retrograde HRP or Xuorescent labeling technique.Anat Rec 229:399–406
Kobler JB, Datta S, Goyal RK, Benecchi EJ (1994) Innervation ofthe larynx, pharynx, and upper esophageal sphincter of the rat.J Comp Neurol 349:129–147
Koyama H, Kishida R, Goris R, Kusunoki T (1993) Giant terminals inthe dorsal octavolateralis nucleus of lampreys. J Comp Neurol335:245–251
Kozaka T, Ando M (2003) Cholinergic innervation to the upper esoph-ageal sphincter muscle in the eel, with special reference to drink-ing behavior. J Comp Physiol B 173:135–140
Laccasagne O, Kessler JP (2000) Cellular and subcellular distributionof the amino-3-hydroxy-5-methyl-4-isoxazole propionate recep-tor subunit GluR2 in the rat dorsal vagal complex. Neurosci99:557–563
Ma PM (1994) Catecholaminergic systems in the zebraWsh. I. Number,morphology, and histochemical characteristics of neurons in thelocus coeruleus. J Comp Neurol 344:242–255
Malmberg L, Ekberg O, Ekström J (1991) EVects of drugs and electri-cal Weld stimulation on isolated muscle strips from rabbit pharyn-goesophageal segment. Disphagia 6:203–208
Medda BK, Lang IM, Layman R, Hogan WJ, Dodds WJ, Shaker R(1994) Characterization and quantiWcation of a pharyngo-UEScontractile reXex in cats. Am J Physiol 267:G972–G983
Morita Y, Finger TE (1985) Topographic and laminar organization ofthe vagal gustatory system in the goldWsh, Carassius auratus.J Comp Neurol 238:187–201
Morita Y, Finger TE (1987) Topographic representation of the sensoryand motor roots of the vagus nerve in the medulla of goldWsh,Carassius auratus. J Comp Neurol 264:231–249
Mukuda T, Ando M (2003a) Medullary motor neurons associated withdrinking behavior of Japanese eels. J Fish Biol 62:1–12
123

122 J Comp Physiol A (2010) 196:111–122
Mukuda T, Ando M (2003b) Brain atlas of the Japanese eel: compari-son to other Wshes. Memory of Faculty of Integrated Arts and Sci-ences, Hiroshima University Series IV 29:1–25
Mukuda T, Matsunaga Y, Kawamoto K, Yamaguchi K, Ando M(2005) “Blood-contacting neurons” in the brain of the Japaneseeel Anguilla japonica. J Exp Zool A 303:366–376
Nabekura J, Ueno T, Katsurabayashi S, Furuta A, Akaike N, Okada M(2002) Reduced NR2A expression and prolonged decay ofNMDA receptor-mediated synaptic current in rat vagal motoneu-rons following axotomy. J Physiol 539:735–741
Pombal MA, Marín O, González A (2001) Distribution of cholineacetyltransferase-immunoreactive structures in the lamprey brain.J Comp Neurol 431:105–126
Pombal MA, Abalo XM, Rodicio MC, Anadón R, González A (2003)Choline acetyltransferase-immunoreactive neurons in the retinaof adult and developing lampreys. Brain Res 993:154–163
Roberts BL, Meredith GE, Maslam S (1989) Immunocytochemicalanalysis of the dopamine system in the brain and spinal cord ofthe European eel, Anguilla anguilla. Anat Embryol (Burl)180:401–412
Shipp T, Deatsch WW, Robertson K (1970) Pharyngoesophageal mus-cle activity during in man. Laryngoscope 80:1–16
Sivarao DV, Krowicki ZK, Hornby PJ (1998) Role of GABAA recep-tors in rat hindbrain nuclei controlling gastric motor function.Neurogastroenterol Motil 10:305–313
Sivarao DV, Krowicki ZK, Abrahams TP, Hornby PJ (1999) Vagally-regulated gastric motor activity: evidence for kainate and NMDAreceptor mediation. Eur J Pharmacol 368:173–182
Sueiro C, Carrera I, Rodríguez-Moldes I, Molist P, Anadón R (2003)Development of catecholaminergic systems in the spinal cord ofthe dogWsh Scyliorhinus canicula (Elasmobranchs). Brain ResDev Brain Res 142:141–150
Travagli RA, Gillis RA, Rossiter CD, Vicini S (1991) Glutamate andGABA-mediated synaptic currents in neurons of the rat dorsalmotor nucleus of the vagus. Am J Physiol 260:G531–G536
Washabau RJ, Fudge M, Price WJ, Barone FC (1995) GABA receptors inthe dorsal motor nucleus of the vagus inXuence feline lower esopha-geal sphincter and gastric function. Brain Res Bull 38:587–594
Willis A, Mihalevich M, NeV RA, Mendelowitz D (1996) Three types ofpostsynaptic glutamatergic receptors are activated in DMNX neu-rons upon stimulation of NTS. Am J Physiol 271:R1614–R1619
Zoungrana OR, Amri M, Car A, Roman C (1997) Intracellular activityof motoneurons of the rostral nucleus ambiguus during swallow-ing in sheep. J Neurophysiol 77:909–922
123
Copyright © 2022 FDOKUMEN


















![4* • Pharingeal grooves/cleft : 4 • [Pharyngeal membrane]](https://static.fdokumen.com/doc/165x107/6334ea00b9085e0bf5093ec7/4-pharingeal-groovescleft-4-pharyngeal-membrane.jpg)

