
Caloric restriction and melatonin substitution: Effects on murine circadian parameters
7
Research report Caloric restriction and melatonin substitution: Effects on murine circadian parameters David Resuehr 1 , James Olcese * Institute for Hormone and Fertility Research, Centre for Innovative Medicine, University of Hamburg, Falkenried 88, 20251 Hamburg, Germany Accepted 22 April 2005 Available online 23 May 2005 Abstract Aging effects have been reported in endocrine, metabolic and behavioral circadian rhythms. The effects of age on the circadian system have been investigated primarily in rats and hamsters and only seldom in mice. Our aim was to assess the effects of two common ‘‘anti- aging’’ treatments, namely caloric restriction (CR) and melatonin substitution, on the circadian system of mice. Animals were subjected to phase delays of the light – dark cycle and constant darkness (DD). The most pronounced change in the murine circadian system was the length of the endogenous period, s , which increased with age regardless of treatment. CR had diverse effects e.g., enabling a more rapid phase shift response while concomitantly leading to a fragmented circadian phenotype with considerable activity during the rest (light) phase. Melatonin enforced the adaptation to the light/dark cycle, thus facilitating a rapid reentrainment to phase delayed lighting conditions. Interestingly, the melatonin-substituted animals displayed an increase in locomotor activity under constant darkness and in 50% of all cases a biphasic (split) activity pattern. These results contribute to the phenotypic evaluation of two very different approaches to intervene in the age-related degeneration of the mammalian circadian system. As both CR and melatonin have negative and positive effects on the behavioral expression of clock function (i.e., fragmentation of rhythms vs. faster reentrainment), their usefulness in managing age-related circadian disorders may be limited. D 2005 Elsevier B.V. All rights reserved. Theme: Neuronal basis of behavior Topic: Biological rhythms and sleep Keywords: Melatonin; Caloric restriction; Circadian period s ; SCN; Locomotor activity; DD; LD 1. Introduction The synchronization of cellular, physiological and/or behavioral rhythms to a cyclically changing environment is adaptive, ubiquitous and probably evolved early in the history of life. One of the most prominent rhythms among animals is the circadian day/night activity rhythm [34]. In the aging animal, the circadian system undergoes many changes at multiple levels. Adaptation to altered daylengths takes longer, activity rhythms are more frag- mented and the endogenous circadian period (s ) changes as compared to young animals [21,29]. Disruptions in the circadian system lead to a reduced life expectancy in many animal models [11,15,16]. Melatonin has diverse biological functions, one of which is to serve as an output signal from the circadian clock [26]. In many animal species, it also controls more slowly changing processes, such as fertility and fur growth (e.g., in hamsters, sheep, [12,37]). The synthesis and release of pineal melatonin are greatly reduced in the aging individual when compared to youthful secretory levels [25]. Melatonin substitution in aging animals increases the maximum life 0006-8993/$ - see front matter D 2005 Elsevier B.V. All rights reserved. doi:10.1016/j.brainres.2005.04.063 * Corresponding author. Current address: Department of Biomedical Sciences, Florida State University College of Medicine, Tallahassee, FL 32306, USA. Fax: +1 850 644 5781. E-mail address: [email protected] (J. Olcese). URL: http://www.med.fsu.edu (J. Olcese). 1 The data represent a portion of the doctoral dissertation of D.R. submitted to the University of Hamburg, Department of Biology. Brain Research 1048 (2005) 146 – 152 www.elsevier.com/locate/brainres
Transcript of Caloric restriction and melatonin substitution: Effects on murine circadian parameters

www.elsevier.com/locate/brainres
Brain Research 1048
Research report
Caloric restriction and melatonin substitution: Effects on murine
circadian parameters
David Resuehr1, James Olcese*
Institute for Hormone and Fertility Research, Centre for Innovative Medicine, University of Hamburg, Falkenried 88, 20251 Hamburg, Germany
Accepted 22 April 2005
Available online 23 May 2005
Abstract
Aging effects have been reported in endocrine, metabolic and behavioral circadian rhythms. The effects of age on the circadian system
have been investigated primarily in rats and hamsters and only seldom in mice. Our aim was to assess the effects of two common ‘‘anti-
aging’’ treatments, namely caloric restriction (CR) and melatonin substitution, on the circadian system of mice. Animals were subjected to
phase delays of the light–dark cycle and constant darkness (DD). The most pronounced change in the murine circadian system was the length
of the endogenous period, s, which increased with age regardless of treatment. CR had diverse effects e.g., enabling a more rapid phase shift
response while concomitantly leading to a fragmented circadian phenotype with considerable activity during the rest (light) phase. Melatonin
enforced the adaptation to the light/dark cycle, thus facilitating a rapid reentrainment to phase delayed lighting conditions. Interestingly, the
melatonin-substituted animals displayed an increase in locomotor activity under constant darkness and in 50% of all cases a biphasic (split)
activity pattern. These results contribute to the phenotypic evaluation of two very different approaches to intervene in the age-related
degeneration of the mammalian circadian system. As both CR and melatonin have negative and positive effects on the behavioral expression
of clock function (i.e., fragmentation of rhythms vs. faster reentrainment), their usefulness in managing age-related circadian disorders may
be limited.
D 2005 Elsevier B.V. All rights reserved.
Theme: Neuronal basis of behavior
Topic: Biological rhythms and sleep
Keywords: Melatonin; Caloric restriction; Circadian period s; SCN; Locomotor activity; DD; LD
1. Introduction
The synchronization of cellular, physiological and/or
behavioral rhythms to a cyclically changing environment is
adaptive, ubiquitous and probably evolved early in the
history of life. One of the most prominent rhythms among
animals is the circadian day/night activity rhythm [34].
0006-8993/$ - see front matter D 2005 Elsevier B.V. All rights reserved.
doi:10.1016/j.brainres.2005.04.063
* Corresponding author. Current address: Department of Biomedical
Sciences, Florida State University College of Medicine, Tallahassee, FL
32306, USA. Fax: +1 850 644 5781.
E-mail address: [email protected] (J. Olcese).
URL: http://www.med.fsu.edu (J. Olcese).1 The data represent a portion of the doctoral dissertation of D.R.
submitted to the University of Hamburg, Department of Biology.
In the aging animal, the circadian system undergoes
many changes at multiple levels. Adaptation to altered
daylengths takes longer, activity rhythms are more frag-
mented and the endogenous circadian period (s) changes ascompared to young animals [21,29]. Disruptions in the
circadian system lead to a reduced life expectancy in many
animal models [11,15,16].
Melatonin has diverse biological functions, one of which
is to serve as an output signal from the circadian clock [26].
In many animal species, it also controls more slowly
changing processes, such as fertility and fur growth (e.g.,
in hamsters, sheep, [12,37]). The synthesis and release of
pineal melatonin are greatly reduced in the aging individual
when compared to youthful secretory levels [25]. Melatonin
substitution in aging animals increases the maximum life
(2005) 146 – 152

D. Resuehr, J. Olcese / Brain Research 1048 (2005) 146–152 147
span [31] and can reduce the severity of other senescence-
accompanied changes [17,33].
Another approach toward reducing age-related physical
degeneration is caloric restriction (CR) which has been
shown in many model systems to delay the onset and
severity of age-related diseases [41,42]. However, the effect
of CR on the circadian system has not been investigated at
the behavioral level in terms of free-running periods or in
terms of reentrainment efficiencies after phase shifting nor
in terms of a comparison to melatonin effects on these
parameters [8,9]. In the present study, mice were maintained
for 10 months (beginning at 5 months of age) under one of
three treatment groups: CR, melatonin substitution or a
controlled normocaloric diet. During this time entrainment
to light:dark (LD) cycles, as well as free-running periods
under constant darkness (DD) and phase shifting efficiency
were assessed in all groups.
2. Materials and methods
2.1. Animals
Mice of the hybrid strain B6C3F1 were obtained at the
age of 6 weeks (N = 80) (Charles River, Wilmington, MA).
This strain has often been used in studies of aging [7,18]. In
contrast to many other laboratory mouse strains [13], the
B6C3F1 strain (derived from a C57BL/6 � C3H cross) is
melatonin-proficient (Olcese et al., unpublished). Young
control mice (N = 20) were kept for longitudinal assessment
of locomotor activity during the course of the study.
Additionally, beginning at 5 months of age, another 60
mice were weighed and sorted into three groups of 20
animals each: controls (CT), melatonin-substituted and CR.
All groups of mice had the same average initial body
weight. Body weight was then measured weekly for the first
14 weeks and every 2 weeks thereafter to assess the impact
of CR.
Animals were housed individually in Plexiglas cages
with overhead infrared sensors to record locomotor activity
(see below for details). Ambient humidity was 68%, and
room temperature was 20 T 2 -C. At the beginning of the
experiment, the light cycle was set to 06:00 h lights on and
18:00 h lights off. For subsequent experimental purposes,
the light cycle was occasionally modified as described
below. Periods of constant darkness were imposed for a
minimum of 3 weeks to determine endogenous period s.
2.2. Feeding
Specially formulated mouse chow was purchased from
Teklad (Madison, Wisconsin). Control and melatonin-
substituted animals were fed standard chow (AIN-93M
adult maintenance precision pellets). CR animals were fed
an especially prepared, vitamin- and mineral-enhanced
restriction diet (based on AIN-93M) to assure proper
nutrition in the face of caloric restriction. This diet has
been employed frequently in studies on the effects of caloric
restriction [5,18,42]. At the beginning of the experiment,
feeding was ad libitum. The amount of food consumed on a
daily basis was assessed by offering each individual a pre-
weighed amount and measuring the uneaten amount
remaining on the following day. The average amount
consumed was 4.3 g/day which was defined as 100%.
Thereafter, food was provided three times a week (a
commonly used feeding strategy [6]). On Monday and
Wednesday, a 2-day portion was given, while on Friday the
mice were fed a 3-day amount of chow. In the case of the
CR mice, initial food amounts were reduced biweekly by
20% until a level of 60% was achieved which was then
maintained over the course of the study (65.8 kcal/week).
The melatonin mice received the same amount of food as
the control mice, whereas the former received melatonin in
their drinking water at a concentration of 5 Ag/ml.
Melatonin (Sigma, St. Louis, MO) stock solutions were
dissolved in 96% ethanol which were then diluted 1:1000 in
water. The CR and CT animals also received alcohol in their
drinking water (0.1%) as a vehicle control. The drinking
bottles were painted black to protect melatonin from
photodegradation.
2.3. Locomotor activity recordings
To record individual locomotor activity, passive infrared
detectors (Conrad-Elektronik, Art.-Nr.192236-3F) were
mounted on top of all cages (N = 20 per group). The
exact timing of the light–dark cycle in the facilities was
recorded with a photodiode (Conrad-Elektronik, Art.-Nr.
606863-3F). Locomotor activity and ambient lighting
signals were measured as TTL-Signals (electrical currents
between 0 V and 5 V) and relayed to a personal computer
(IBM PC 286, 500 MB HDD, 32 MB RAM) via a digital
I/O-Card (Conrad-Elektronik, Model Nr. PIO 24/48 II).
The absence of activity ‘‘0’’ was defined as 0–0.8 V,
whereas 2.4–5 V was defined as ‘‘1’’—or activity. A
refractory period of 7 s after each signal was established in
the computer program, during which time no further
signals could be recorded. Signals were recorded in 6
min bins per recording interval, i.e., 240 intervals per day.
The recording and analysis of locomotor activity and the
determination of endogenous period s (using a chi square
periodogram) were performed with a Quickbasic computer
program written by Dr. Haiko Dernbach (Tiermedizinische
Hochschule Hannover, Germany). Reentrainment analyses
after phase shifts were conducted by calculating the
amount of activity in the 40 6 min bins immediately
following the onset of darkness before and after shifting. A
threshold level was determined by forming the average
activity under normal LD conditions i.e., before the shift
(N = 20 animals per tested group). Attainment of this
amount of nocturnal activity after shifting was considered
as the threshold for reentrainment.

Fig. 1. Total average activity of animals from different treatment groups at
the age of 14 months (N = 10 per group) under LD (12 h light: 12 h dark) or
DD (constant dark) conditions. Night LD refers to nocturnal activity under
LD, whereas day LD refers to daytime activity. Note the higher total
activity of melatonin mice under DD (P < 0.05) and the elevated daytime
activity of CR mice under LD (P < 0.05). The data represented here are
derived from the electronic files displayed in actogram form in Fig. 2.
Fig. 2. Representative actograms of mouse locomotor activity at the ages of 7 (A–
DD (*) and subsequent reentrainment (triangle) (A–C). (A) Seven-month-old
melatonin-treated animal displaying biphasic activity. In panels (D–F), the light p
(arrow Y). The DD in all actograms began at the * and lasted until the reentrainm
(E) CR animal aged 11 months. (F) Melatonin animal aged 11 months.
D. Resuehr, J. Olcese / Brain Research 1048 (2005) 146–152148
Statistical data analysis was performed on a Dell
Dimension 2400\ PC using GraphPad Prism\ and MS
Excel\.
3. Results
The untreated CT group had an average body weight of
29.1 T 0.4 g, the melatonin-substituted animals had average
body weight of 29.4 T 0.4 g and the CR animals had an
average body weight of 28.6 T 0.3 g at the beginning of the
experiment. At the end of the experiment (40th week), the
control animals had an average body weight of 41.0 T 0.9 g
and the melatonin-substituted animals had an average body
weight of 35.0 T 0.9 g. The weight difference at the end of
the experiment between the CT and melatonin groups was
statistically significant after a two sided t test (P < 0.001).
The body weight of the CR animals decreased to 20.5 T 0.3
g in the 10th week and increased thereafter to 24.1 T 0.4 g inthe 40th week (P < 0.001).
The activity in all groups of mice in the dark phase was
higher than during the light phase (Fig. 1, P < 0.05).
However, the nocturnal activity of the CR animals was a
little lower (�11%, P < 0.05), while the daytime activity
was higher (+43%, P < 0.05) when compared to CT or
C) and 11 months (D–F). The light cycle was 06:00–18:00 h followed by
control animal. (B) Seven-month-old CR animal. (C) Seven-month-old
eriod was 10:00–22:00 h followed by a 4 h phase delay to 14:00–02:00 h
ent to the pre-DD conditions (triangle). (D) Control animal aged 11 months.
D. Resuehr, J. Olcese / Brain Research 1048 (2005) 146–152 149
melatonin animals. The total activity increased (+26.3%,
P < 0.05) during DD conditions in the melatonin-treated
animals (Fig. 1). There was no significant inter-group
difference in total activity between the examined groups
of the same age (Fig. 1, LD 24 h). Older control mice
(21 months) displayed significantly less activity under all
lighting conditions (data not shown).
CT animals displayed a robust circadian rhythmicity
under LD conditions and a free-running period <24 h in DD
(Figs. 2A and D). During LD conditions, but even more so
during DD conditions, there was a tendency for the main
bout of activity to split slightly into two separate bouts. The
major trait of the CR group was a strongly fragmented
activity rhythm with significant activity during the dark/
light transition (Figs. 2B and E upper portion of picture,
above * and Y). As was the case with control animals, CR
mice individuals often displayed two activity peaks, which
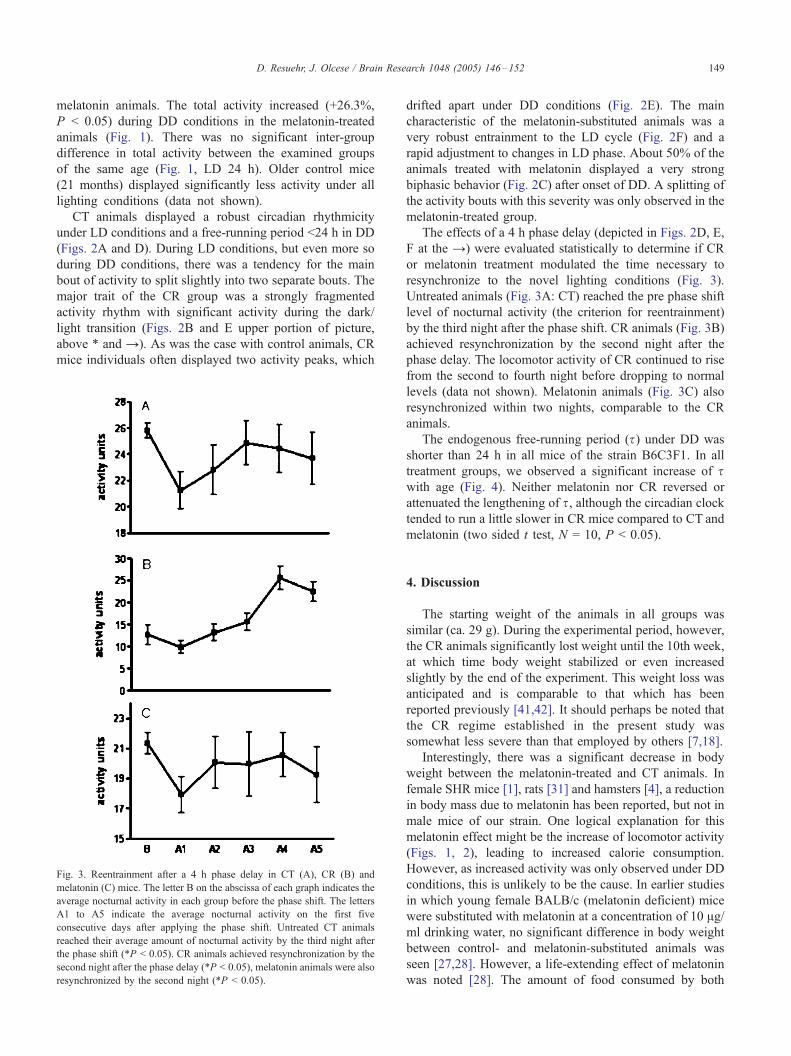
Fig. 3. Reentrainment after a 4 h phase delay in CT (A), CR (B) and
melatonin (C) mice. The letter B on the abscissa of each graph indicates the
average nocturnal activity in each group before the phase shift. The letters
A1 to A5 indicate the average nocturnal activity on the first five
consecutive days after applying the phase shift. Untreated CT animals
reached their average amount of nocturnal activity by the third night after
the phase shift (*P < 0.05). CR animals achieved resynchronization by the
second night after the phase delay (*P < 0.05), melatonin animals were also
resynchronized by the second night (*P < 0.05).
drifted apart under DD conditions (Fig. 2E). The main
characteristic of the melatonin-substituted animals was a
very robust entrainment to the LD cycle (Fig. 2F) and a
rapid adjustment to changes in LD phase. About 50% of the
animals treated with melatonin displayed a very strong
biphasic behavior (Fig. 2C) after onset of DD. A splitting of
the activity bouts with this severity was only observed in the
melatonin-treated group.
The effects of a 4 h phase delay (depicted in Figs. 2D, E,
F at the Y) were evaluated statistically to determine if CR
or melatonin treatment modulated the time necessary to
resynchronize to the novel lighting conditions (Fig. 3).
Untreated animals (Fig. 3A: CT) reached the pre phase shift
level of nocturnal activity (the criterion for reentrainment)
by the third night after the phase shift. CR animals (Fig. 3B)
achieved resynchronization by the second night after the
phase delay. The locomotor activity of CR continued to rise
from the second to fourth night before dropping to normal
levels (data not shown). Melatonin animals (Fig. 3C) also
resynchronized within two nights, comparable to the CR
animals.
The endogenous free-running period (s) under DD was
shorter than 24 h in all mice of the strain B6C3F1. In all
treatment groups, we observed a significant increase of swith age (Fig. 4). Neither melatonin nor CR reversed or
attenuated the lengthening of s, although the circadian clock
tended to run a little slower in CR mice compared to CT and
melatonin (two sided t test, N = 10, P < 0.05).
4. Discussion
The starting weight of the animals in all groups was
similar (ca. 29 g). During the experimental period, however,
the CR animals significantly lost weight until the 10th week,
at which time body weight stabilized or even increased
slightly by the end of the experiment. This weight loss was
anticipated and is comparable to that which has been
reported previously [41,42]. It should perhaps be noted that
the CR regime established in the present study was
somewhat less severe than that employed by others [7,18].
Interestingly, there was a significant decrease in body
weight between the melatonin-treated and CT animals. In
female SHR mice [1], rats [31] and hamsters [4], a reduction
in body mass due to melatonin has been reported, but not in
male mice of our strain. One logical explanation for this
melatonin effect might be the increase of locomotor activity
(Figs. 1, 2), leading to increased calorie consumption.
However, as increased activity was only observed under DD
conditions, this is unlikely to be the cause. In earlier studies
in which young female BALB/c (melatonin deficient) mice
were substituted with melatonin at a concentration of 10 Ag/ml drinking water, no significant difference in body weight
between control- and melatonin-substituted animals was
seen [27,28]. However, a life-extending effect of melatonin
was noted [28]. The amount of food consumed by both
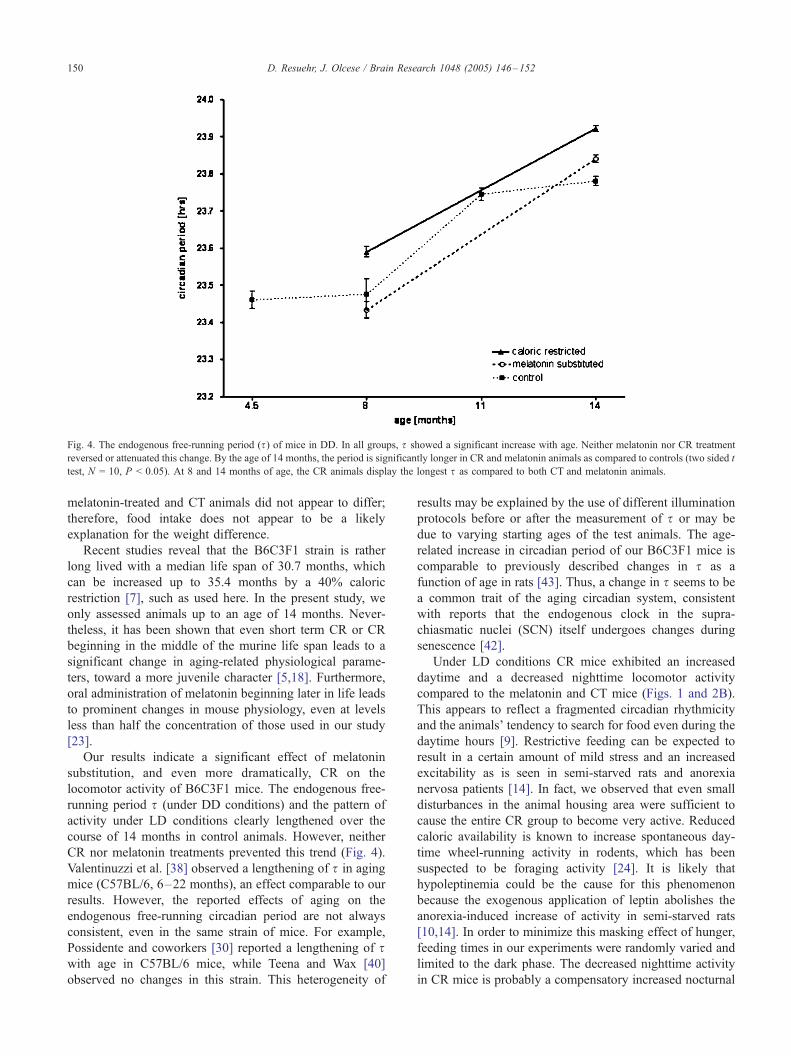
Fig. 4. The endogenous free-running period (s) of mice in DD. In all groups, s showed a significant increase with age. Neither melatonin nor CR treatment
reversed or attenuated this change. By the age of 14 months, the period is significantly longer in CR and melatonin animals as compared to controls (two sided t
test, N = 10, P < 0.05). At 8 and 14 months of age, the CR animals display the longest s as compared to both CT and melatonin animals.
D. Resuehr, J. Olcese / Brain Research 1048 (2005) 146–152150
melatonin-treated and CT animals did not appear to differ;
therefore, food intake does not appear to be a likely
explanation for the weight difference.
Recent studies reveal that the B6C3F1 strain is rather
long lived with a median life span of 30.7 months, which
can be increased up to 35.4 months by a 40% caloric
restriction [7], such as used here. In the present study, we
only assessed animals up to an age of 14 months. Never-
theless, it has been shown that even short term CR or CR
beginning in the middle of the murine life span leads to a
significant change in aging-related physiological parame-
ters, toward a more juvenile character [5,18]. Furthermore,
oral administration of melatonin beginning later in life leads
to prominent changes in mouse physiology, even at levels
less than half the concentration of those used in our study
[23].
Our results indicate a significant effect of melatonin
substitution, and even more dramatically, CR on the
locomotor activity of B6C3F1 mice. The endogenous free-
running period s (under DD conditions) and the pattern of
activity under LD conditions clearly lengthened over the
course of 14 months in control animals. However, neither
CR nor melatonin treatments prevented this trend (Fig. 4).
Valentinuzzi et al. [38] observed a lengthening of s in aging
mice (C57BL/6, 6–22 months), an effect comparable to our
results. However, the reported effects of aging on the
endogenous free-running circadian period are not always
consistent, even in the same strain of mice. For example,
Possidente and coworkers [30] reported a lengthening of swith age in C57BL/6 mice, while Teena and Wax [40]
observed no changes in this strain. This heterogeneity of
results may be explained by the use of different illumination
protocols before or after the measurement of s or may be
due to varying starting ages of the test animals. The age-
related increase in circadian period of our B6C3F1 mice is
comparable to previously described changes in s as a
function of age in rats [43]. Thus, a change in s seems to be
a common trait of the aging circadian system, consistent
with reports that the endogenous clock in the supra-
chiasmatic nuclei (SCN) itself undergoes changes during
senescence [42].
Under LD conditions CR mice exhibited an increased
daytime and a decreased nighttime locomotor activity
compared to the melatonin and CT mice (Figs. 1 and 2B).
This appears to reflect a fragmented circadian rhythmicity
and the animals’ tendency to search for food even during the
daytime hours [9]. Restrictive feeding can be expected to
result in a certain amount of mild stress and an increased
excitability as is seen in semi-starved rats and anorexia
nervosa patients [14]. In fact, we observed that even small
disturbances in the animal housing area were sufficient to
cause the entire CR group to become very active. Reduced
caloric availability is known to increase spontaneous day-
time wheel-running activity in rodents, which has been
suspected to be foraging activity [24]. It is likely that
hypoleptinemia could be the cause for this phenomenon
because the exogenous application of leptin abolishes the
anorexia-induced increase of activity in semi-starved rats
[10,14]. In order to minimize this masking effect of hunger,
feeding times in our experiments were randomly varied and
limited to the dark phase. The decreased nighttime activity
in CR mice is probably a compensatory increased nocturnal

D. Resuehr, J. Olcese / Brain Research 1048 (2005) 146–152 151
rest phase due to excessive daytime activity. The total
locomotor activity of all three treatment groups under LD
conditions is however not significantly different. In contrast,
under DD conditions, many of the melatonin-treated
animals displayed an increase in total activity (Figs. 1,
2C), in many cases due to rhythm splitting (see discussion
below).
The time required to reentrain to a shifted LD after DD
has been reported to increase with age in mice [38]. In the
present study, we observed that the melatonin-treated and
CR animals adjusted to a 4 h phase delay in the lighting
conditions faster than control mice (Figs. 2A–C and 3A–
C). However, the CR animals were unusual in that 3–5 days
after the phase delay they showed a remarkable excess of
activity (Fig. 3B). This overcompensation is likely to be
artificial and may be related to the increased excitability of
the CR mice subsequent to refeeding after a 3-day (week-
end) interval.
Melatonin’s effects on the circadian system are well
known. In several species, melatonin induces phase shifts
and entrainment of the circadian clock [32]. Melatonin
treatment can also entrain the circadian system of humans
[36]. In the present study, melatonin facilitated adjustment
to a 4 h phase delay, such that mice were resynchronized
within 48 h as opposed to 72 h for the control animals.
Similar positive effects of melatonin on synchronization to
new lighting conditions have been reported previously in
hamsters [39].
Quite unexpectedly, this mouse strain frequently devel-
oped a ‘‘split’’ free-running rhythm under DD. Splitting is
commonly seen in nocturnal rodents when they are
maintained under LL conditions [21]. It is defined as the
spontaneous separation of the main activity bout into two
smaller bouts with different free-running periods. Pro-
nounced splitting of the free-running locomotor activity
rhythm was seldom seen in either CT or CR animals (Figs.
2A, B, D, E); however, it was common in the melatonin-
treated animals, especially under DD conditions. Only
unsplit animals were used for the determination and
graphical illustration of s (Fig. 4). As depicted in Fig. 2C,
the rhythm splitting in melatonin-treated animals was often
substantial, resulting in two free-running rhythms with
different periods (23 vs. 23.6 h, calculated by v2 periodo-
gram analysis). Only 20% of the CT animals displayed
biphasic free-running activity under DD, while the inci-
dence of splitting in the melatonin animals was 50%. In
view of its known effects on the SCN clock [19, 20], it
seems conceivable that pharmacological levels of melatonin,
self-administered in a regular nocturnal manner to B6C3F1
mice, may paradoxically facilitate the uncoupling of dual
SCN oscillators under Zeitgeber-free conditions (i.e., DD),
which would manifest itself as rhythm splitting. Further
studies at the molecular level are needed to better under-
stand this unusual behavior. It would be informative to
determine whether the SCN expression of the MT1 and/or
MT2 melatonin receptor is vital for the splitting phenom-
enon that we observed in the B6C3F1 mice. This could, for
example, be assessed by backcrossing this strain with
melatonin receptor knockout mice [35].
In conclusion, the present study has compared two
anti-aging treatments, which are likely to influence the
endogenous circadian system. Melatonin’s use as a
chronobiotic has been well explored in rodents and
humans [2,3]. Timed restricted feeding under the con-
ditions of caloric restriction [22] is a potent synchronizer
of peripheral and central oscillators. In the present report,
we deliberately alternated the feeding times and fed mice
in the dark phase to avoid offering a light coupled
‘‘Zeitgeber’’ i.e., feeding in the light phase which could
lead to animals associating light with food. Nevertheless,
on a behavioral level, we could detect a direct effect of
caloric restriction on the circadian clock (in terms of
locomotor activity), rendering it more sensitive to
changed light cycles. A comparable effect was seen in
animals administered melatonin. The mechanisms through
which melatonin and caloric restriction exert their action
on the circadian clock however are probably completely
different, that is, melatonin is likely to work through G-
protein-coupled receptors [35], while caloric restriction is
thought to operate via other pathways, such as the
orexigenic system and leptin [10,14].
Acknowledgments
We would like to thank Dr. Haiko Dernbach for
assistance in setting up the locomotor activity electronics
and for his generosity in donating the software for the
activity recordings. We are also grateful to Jenny Behrens
and Helena Fischer for assistance in maintaining the
animals. This project was financed by the Leidenberger-
Muller Foundation and the Graduiertenkolleg 336 (DFG).
References
[1] V.N. Anisimov, I.N. Alimova, D.A. Baturin, I.G. Popovich, M.A.
Zabezhinski, S.V. Rosenfeld, K.G. Manton, A.V. Semenchenko, A.I.
Yashin, Dose-dependent effect of melatonin on life span and
spontaneous tumor incidence in female SHR mice, Exp. Gerontol.
38 (2003) 449–461.
[2] J. Arendt, D.J. Skene, Melatonin as a chronobiotic, Sleep Med. Rev. 9
(2005) 25–39.
[3] J. Arendt, S. Deacon, J. English, S. Hampton, L. Morgan, Melatonin
and adjustment to phase shift, J. Sleep Res. 4 (1995) 74–79.
[4] S.D. Bilbo, R.J. Nelson, Melatonin regulates energy balance and
attenuates fever in Siberian hamsters, Endocrinology 143 (2002)
2527–2533.
[5] S.X. Cao, J.M. Dhahbi, P.L. Mote, S.R. Spindler, Genomic profiling of
short- and long-term caloric restriction effects in the liver of aging
mice, Proc. Natl. Acad. Sci. U. S. A. 98 (2001) 10630–10635.
[6] J.M. Dhahbi, P.L. Mote, J. Wingo, J.B. Tillman, R.L. Walford, S.R.
Spindler, Calories and aging alter gene expression for gluconeogenic,
glycolytic, and nitrogen-metabolizing enzymes, Am. J. Physiol. 277
(1999) E352–E360.

D. Resuehr, J. Olcese / Brain Research 1048 (2005) 146–152152
[7] J.M. Dhahbi, H.J. Kim, P.L. Mote, R.J. Beaver, S.R. Spindler,
Temporal linkage between the phenotypic and genomic responses
to caloric restriction, Proc. Natl. Acad. Sci. U. S. A. 101 (2004)
5524–5529.
[8] P.H. Duffy, R. Feuers, K.D. Nakamura, J. Leakey, R.W. Hart, Effect of
chronic caloric restriction on the synchronization of various physio-
logical measures in old female Fischer 344 rats, Chronobiol. Int. 7
(1990) 113–124.
[9] P.H. Duffy, R.J. Feuers, R.W. Hart, Effect of chronic caloric restriction
on the circadian regulation of physiological and behavioral variables
in old male B6C3F1 mice, Chronobiol. Int. 7 (1990) 291–303.
[10] C. Exner, J. Hebebrand, H. Remschmidt, C. Wewetzer, A. Ziegler, S.
Herpertz, U. Schweiger, W.F. Blum, G. Preibisch, G. Heldmaier, M.
Klingenspor, Leptin suppresses semi-starvation induced hyperactivity
in rats: implications for anorexia nervosa, Mol. Psychiatry 5 (2000)
476–481.
[11] E. Filipski, V.M. King, X. Li, T.G. Granda, M.C. Mormont, B.
Claustrat, M.H. Hastings, F. Levi, Disruption of circadian coordination
accelerates malignant growth in mice, Pathol. Biol. (Paris) 51 (2003)
216–219.
[12] T. Gerlach, J.E. Aurich, Regulation of seasonal reproductive activity in
the stallion, ram and hamster, Anim. Reprod. Sci. 58 (2000) 197–213.
[13] M. Goto, I. Oshima, T. Tomita, S. Ebihara, Melatonin content of
the pineal gland in different mouse strains, J. Pineal Res. 7 (1989)
195–204.
[14] J. Hebebrand, C. Exner, K. Hebebrand, C. Holtkamp, R.C. Casper, H.
Remschmidt, B. Herpertz-Dahlmann, M. Klingenspor, Hyperactivity
in patients with anorexia nervosa and in semistarved rats: evidence for
a pivotal role of hypoleptinemia, Physiol. Behav. 79 (2003) 25–37.
[15] M.W. Hurd, M.R. Ralph, The significance of circadian organization for
longevity in the golden hamster, J. Biol. Rhythms 13 (1998) 430–436.
[16] A. Klarsfeld, F. Rouyer, Effects of circadian mutations and LD
periodicity on the life span of Drosophila melanogaster, J. Biol.
Rhythms 13 (1998) 471–478.
[17] G.C. Koster-van Hoffen, M. Mirmiran, N.P. Bos, W. Witting, P.
Delagrange, B. Guardiola-Lemaitre, Effects of a novel melatonin
analog on circadian rhythms of body temperature and activity in young,
middle-aged, and old rats, Neurobiol. Aging 14 (1993) 565–569.
[18] C.K. Lee, D.B. Allison, J. Brand, R. Weindruch, T.A. Prolla,
Transcriptional profiles associated with aging and middle age-onset
caloric restriction in mouse hearts, Proc. Natl. Acad. Sci. U. S. A. 99
(2002) 14988–14993.
[19] C. Liu, D.R. Weaver, X. Jin, L.P. Shearman, R.L. Pieschl, V.K.
Gribkoff, S.M. Reppert, Molecular dissection of two distinct actions of
melatonin on the suprachiasmatic circadian clock, Neuron 19 (1997)
91–102.
[20] A.J. McArthur, A.E. Hunt, M.U. Gillette, Melatonin action and signal
transduction in the rat suprachiasmatic circadian clock: activation of
protein kinase C at dusk and dawn, Endocrinology 138 (1997)
627–634.
[21] J.D. McAuley, J.P. Miller, E. Beck, Z.M. Nagy, K.C. Pang, Age-
related disruptions in circadian timing: evidence for ‘‘split’’ activity
rhythms in the SAMP8, Neurobiol. Aging 23 (2002) 625–632.
[22] J. Mendoza, C. Graff, H. Dardente, P. Pevet, E. Challet, Feeding cues
alter clock gene oscillations and photic responses in the supra-
chiasmatic nuclei of mice exposed to a light/dark cycle, J. Neurosci.
25 (2005) 1514–1522.
[23] Y. Okatani, A. Wakatsuki, R.J. Reiter, Y. Miyahara, Hepatic
mitochondrial dysfunction in senescence-accelerated mice: correction
by long-term, orally administered physiological levels of melatonin,
J. Pineal Res. 33 (2002) 127–133.
[24] J.M. Overton, T.D. Williams, Behavioral and physiologic responses to
caloric restriction in mice, Physiol. Behav. 81 (2004) 749–754.
[25] S.F. Pang, F. Tang, P.L. Tang, Negative correlation of age and the
levels of pineal melatonin, pineal N-acetylserotonin, and serum
melatonin in male rats, J. Exp. Zool. 229 (1984) 41–47.
[26] S. Perreau-Lenz, A. Kalsbeek, J. Van Der Vliet, P. Pevet, R.M. Buijs,
In vivo evidence for a controlled offset of melatonin synthesis at dawn
by the suprachiasmatic nucleus in the rat, Neuroscience 130 (2005)
797–803.
[27] W. Pierpaoli, W. Regelson, Pineal control of aging: effect of melatonin
and pineal grafting on aging mice, Proc. Natl. Acad. Sci. U. S. A. 91
(1994) 787–791.
[28] W. Pierpaoli, A. Dall’Ara, E. Pedrinis, W. Regelson, The pineal
control of aging. The effects of melatonin and pineal grafting on the
survival of older mice, Ann. N. Y. Acad. Sci. 621 (1991) 291–313.
[29] C.S. Pittendrigh, S. Daan, Circadian oscillations in rodents: a
systematic increase of their frequency with age, Science 186
(1974) 548–550.
[30] B. Possidente, S. McEldowney, A. Pabon, Aging lengthens circadian
period for wheel-running activity in C57BL mice, Physiol. Behav. 57
(1995) 575–579.
[31] B. Prunet-Marcassus, M. Desbazeille, A. Bros, K. Louche, P.
Delagrange, P. Renard, L. Casteilla, L. Penicaud, Melatonin reduces
body weight gain in Sprague Dawley rats with diet-induced obesity,
Endocrinology 144 (2003) 5347–5352.
[32] J.R. Redman, Circadian entrainment and phase shifting in mammals
with melatonin, J. Biol. Rhythms 12 (1997) 581–587.
[33] R.J. Reiter, Oxidative damage in the central nervous system:
protection by melatonin, Prog. Neurobiol. 56 (1998) 359–384.
[34] S.M. Reppert, D.R. Weaver, Coordination of circadian timing in
mammals, Nature 418 (2002) 935–941.
[35] S.M. Reppert, D.R. Weaver, T. Ebisawa, Cloning and characterization
of a mammalian melatonin receptor that mediates reproductive and
circadian responses, Neuron 13 (1994) 1177–1185.
[36] R.L. Sack, R.W. Brandes, A.R. Kendall, A.J. Lewy, Entrainment of
free-running circadian rhythms by melatonin in blind people, N. Engl.
J. Med. 343 (2000) 1070–1077.
[37] S.J. Schoeman, M.A. Botha, The effect of melatonin treatment on
reproductive efficiency in an accelerated lambing system, J. S. Afr.
Vet. Assoc. 66 (1995) 230–234.
[38] V.S. Valentinuzzi, K. Scarbrough, J.S. Takahashi, F.W. Turek, Effects
of aging on the circadian rhythm of wheel-running activity in
C57BL/6 mice, Am. J. Physiol. 273 (1997) R1957–R1964.
[39] O. Van Reeth, L. Weibel, E. Olivares, S. Maccari, E. Mocaer, F.W.
Turek, Melatonin or a melatonin agonist corrects age-related changes
in circadian response to environmental stimulus, Am. J. Physiol.,
Regul. Integr. Comp. Physiol. 280 (2001) R1582–R1591.
[40] T.M. Wax, Runwheel activity patterns of mature–young and senescent
mice: the effect of constant lighting conditions, J. Gerontol. 30 (1975)
22–27.
[41] R. Weindruch, The retardation of aging by caloric restriction: studies
in rodents and primates, Toxicol. Pathol. 24 (1996) 742–745.
[42] R. Weindruch, Caloric restriction and aging, Sci. Am. 274 (1996)
46–52.
[43] W. Witting, M. Mirmiran, N.P. Bos, D.F. Swaab, The effect of old age
on the free-running period of circadian rhythms in rat, Chronobiol. Int.
11 (1994) 103–112.




















