
Convergence and Segregation of Ventral Striatal Inputs and Outputs
Upload
independentCategory
view
6download
0
Photic and nonphotic inputs to the diurnal circadian clock
Colleen M. Novaka*, J. Christopher Ehlenb and H. Elliott Albersb
aMayo Clinic, Endocrine Research Unit, St Marys Hospital, Joseph 5-194, 200 1st St. SW, Rochester,MN 55905, USA; bDepartment of Biology, Georgia State University, Atlanta, GA 30302, USA
Diurnal animals occupy a different temporal niche from nocturnal animals and areconsequently exposed to different amounts of light as well as different dangers.Accordingly, some variation exists in the way that diurnal animals synchronize theirinternal circadian clock to match the external 24-hour daily cycle. First, though thebrain mechanisms underlying photic entrainment are very similar among species withdifferent daily activity patterns, there is evidence that diurnal animals are less sensitiveto photic stimuli compared to nocturnal animals. Second, stimuli other than light thatsynchronize rhythms (i.e. nonphotic stimuli) can also entrain and phase shift dailyrhythms. Some of the rules that govern nonphotic entrainment in nocturnal animals aswell as the brain mechanisms that control nonphotic influences on rhythms do notappear to apply to diurnal animals, however. Some evidence supports the idea thatarousal or activity plays an important role in entraining rhythms in diurnal animals,either during the light (active) or dark (inactive) phases, though no consistent pattern isseen. GABAergic stimulation induces phase shifts during the subjective day in bothdiurnal and nocturnal animals. In diurnal Arvicanthis niloticus (Nile grass rats), SCNGABAA receptor activation at this time results in phase delays while in nocturnalanimals phase advances are induced. It appears that the effect of GABA at this circadianphase results from the inhibition of period gene expression in both diurnal and nocturnalanimals. Nonetheless, the resulting phase shifts are in opposite directions. It is notknown what stimuli or behaviours ultimately induce changes in GABA activity in theSCN that result in alterations of circadian phase in diurnal grass rats. Taken together,studies such as these suggest that it may be problematic to apply the principlesgoverning nocturnal nonphotic entrainment and its underlying mechanisms to diurnalspecies including humans.
Keywords: entrainment; phase shift; GABA; suprachiasmatic nucleus; brain
Introduction
The majority of evidence regarding synchronization of the circadian clock to environmentalstimuli has come from nocturnal rodent models. There are many similarities in themechanisms of circadian clocks in diurnal and nocturnal species (see Smale et al., this issue).It is impossible to ignore the vastly different experiences of animals that explore theirenvironment during the daytime versus night-time, however. The differences in the naturalhistory of exposure to light and other environmental stimuli affect synchronization between
*Corresponding author. Email: [email protected]
Biological Rhythm Research
Vol. 39, No. 3, June 2008, 291–304
ISSN 0929-1016 print/ISSN 1744-4179 online
� 2008 Taylor & Francis
DOI: 10.1080/09291010701683482
http://www.informaworld.com

the circadian clock and the 24-hour cycle. In this review, we will focus on evidence regardingdiurnal/nocturnal differences in the photic and nonphotic inputs to the circadian system.
Photic synchronization
As described by Smale et al. (this issue), the suprachiasmatic nuclei (SCN) of diurnal andnocturnal animals have several striking similarities. One common feature of most speciesstudied so far is the pattern of response of the circadian system to light, the primarysynchronizer, or zeitgeber. The phase-response curve (PRC) for light of diurnal rodentssuch as Arvicanthis niloticus (unstriped Nile grass rats) is very similar to those of severalnocturnal animals, with light in the early night causing phase delays in the activity rhythm,and light in the late night causing phase advances (Figure 1) (Mahoney et al. 2001).Moreover, rhythms in the expression of clock genes such as Period 1 and 2 in the SCN ofdiurnal animals (Figure 2) approximate patterns of expression previously described innocturnal animals (Sun et al. 1997; Tei et al. 1997; Kume et al. 1999; Nuesslein-Hildesheimet al. 2000; Mrosovsky et al. 2001; Dardente et al. 2002; Lincoln et al. 2002; Oster et al.2002; Caldelas et al. 2003; Novak et al. 2006; Ramanathan et al. 2006). These data suggestthat, as a general rule, diurnal and nocturnal animals react similarly to acute and chroniclight exposure with regard to the synchronization of their daily cycle and phaseadjustment. In summary, SCN neuronal activity and clock gene expression patterns arefixed in their relationship to the light – dark cycle, and it is likely that it is some aspect ofSCN output that results in diurnal or nocturnal behavioural and endocrine rhythms.
One fundamental question has been: what is the impact of a history of long-duration,high-intensity light exposure on the brains and circadian systems of diurnal animals? First,it is readily observed that the optic structures and brain regions that receive visual andphotic input differ between diurnal and nocturnal rodents (Goel et al. 1999; Novak et al.1999; Smale & Boverhof 1999). The optic nerve, chiasm, and tract are larger in comparisonto brain size in diurnal rodents compared to nocturnal rodents (Pickard & Silverman 1981;Kita & Oomura 1982; Pickard 1982; Bronchti et al. 1991; Goel et al. 1999; Novak et al.1999; Smale & Boverhof 1999; Abrahamson & Moore 2001). In addition, retinal input to
Figure 1. The phase-response curve (PRC) to light in diurnal Nile grass rats (Arvicanthis niloticus).The PRCs of both ‘‘diurnal’’ and ‘‘nocturnal’’ chronotypes of grass rats were examined (Figure 5used with permission from (Mahoney et al. 2001)).
292 C.M. Novak et al.

visual and circadian structures is more intense compared to some (rat) but not all (Syrianhamster) nocturnal rodents (Pickard & Silverman 1981; Kita & Oomura 1982; Pickard1982; Youngstrom & Nunez 1986; Abrahamson & Moore 2001). This includes retinalprojections to the SCN and other regions such as the superior colliculus. In A. niloticus,retinohypothalamic projections innervate the SCN region and continue into thesubparaventricular zone (Smale & Boverhof 1999) This region is likely to be critical fordetermining the active phase of diurnal and nocturnal rodents (Smale et al. 2001; Moore &Danchenko 2002; Schwartz et al. 2004; Schwartz & Smale 2005; Ramanathan et al. 2006).The retinal projections to the dorsal raphe, an indirect pathway for light to affect thecircadian system (Morin & Allen 2006), are considerably denser in some diurnal andcrepuscular rodents compared to rats (Fite et al. 1999; Fite & Janusonis 2001). This featureis not apparent in other diurnal rodents, however (Major et al. 2003), and may reflectinter-species variability. It is interesting that retinal input is not greater in all brain regionsin diurnal rodents. The supraoptic nucleus receives a roughly equivalent projection patternin diurnal and nocturnal animals, while the sleep-active ventrolateral preoptic area(VLPO) contains only the occasional retinal fiber in A. niloticus. The VLPO of nocturnal
Figure 2. (Top) Daily rhythms of Per mRNA levels in the SCN of Arvicanthis niloticus after threedays in constant darkness. (A) Per1mRNA peaked during the mid-subjective day. (B) The rhythm inmPer2 mRNA levels in the SCN of A. niloticus peaked approximately four hours later.ZT ¼ zeitgeber time, with ZT 0 ¼ time of lights on before release into DD for 3 cycles. N ¼ 6/group (N ¼ 7 for ZT 0). mPer1 and hPer1: *ZTs 4 and 8 greater than all other groups. mPer2: **ZT8 greater than all groups but ZT12, p 5 0.05; *ZT 12 greater than all groups but ZTs 8 and 4,p 5 0.05. (Bottom) Light pulses significantly increased SCN Per mRNA levels during the earlysubjective night in the SCN of Arvicanthis niloticus. (A) The Per1 hybridization signal wassignificantly increased by the light pulse after 30 min and one hour. (B) mRNA levels of Per2 weresignificantly increased at one and two hours after the pulse, compared to sham-pulsed animals.N ¼ 5 for each treatment at each time point (total N ¼ 30) (*p 5 0.05) All animals were housed inconstant darkness for three cycles. (Figures 1 and 3 used with permission from (Novak et al. 2006)).
Biological Rhythm Research 293

rats (Lu et al. 1999; Novak et al. 1999) receives significant retinal inputs, which appears toexcite the neurons of this area to hasten the onset of slow-wave sleep (Lu et al. 1999). It isreasonable to suppose that in diurnal rodents, the lack of significant retinal input to theVLPO may reflect a decreased responsiveness of slow-wave sleep to light compared tonocturnal rats (Novak et al. 1999). Together, these data suggest that visual and photicinput to the brain differs in diurnal and nocturnal species and that this variation seemsdependent on region and function.
Second, because of the high intensity of light that diurnal animals are exposed to, theymay be less sensitive to the effects of light on the circadian system. For comparison, averageindoor light exposure is 100–1000 lux, whereas the average outdoor sunlight diurnal animalsare exposed to is 50 000 lux. Evidence suggests that the circadian systems of diurnal rodentsmay be less sensitive to the effects of light compared to circadian rhythms of nocturnalanimals. The intensity of light needed to phase shift the rhythms of diurnal and nocturnalanimals has not been directly compared, however. We have compared the relative sensitivityof the neurochemical mechanisms through which light acts on the SCN in diurnal andnocturnal rodents. The neuronal pathway through which light affects the SCN, theretinohypothalamic tract, is glutamatergic; activation of NMDA glutamate receptorsinduces a light-like PRC (Mintz & Albers 1997; Mintz et al. 1999). In diurnal A. niloticus,50 mM of NMDA directly microinjected into the SCN induces the maximum phase shift atnight (Novak & Albers 2002); in hamsters, only 10 mM NMDA produces maximal phaseshifts using the identical protocol (Mintz & Albers 1997; Mintz et al. 1999). This suggeststhat diurnal grass rats are less sensitive than Syrian hamsters to the neurochemical signalsthat mediate the effects of photic stimulation on the clock, which may reflect a dampenedsensitivity to light in grass rats and possibly other diurnal animals. Finally, decreased photicsensitivity in diurnal species has also been suggested by in vitro studies. SCN cells receivingretinal input respond with neurophysiological suppression more often in diurnal deguscompared to rats (Jiao et al. 1999). These studies suggest mechanisms through which lightsensitivity might vary between diurnal and nocturnal animals. Thus, potential mechanismsexist to explain diurnal/nocturnal differences in photic sensitivity.
The examination of melatonin suppression provides an alternate means to comparephotic sensitivity. The ability of light to suppress melatonin secretion also requiresretinohypothalamic projection to the SCN (Paul et al. 2003, 2004) and the activation ofNMDA receptors. Melatonin levels in nocturnal hamsters are suppressed by 50 lux of lightfor 15 min (Nelson & Takahashi 1991; Paul et al. 2003). A range of light intensitiesvarying from room light (more intense than the 50 lux cited above) to 2000 lux did notsuppress night-time melatonin levels in diurnal Eastern Chipmunks and/or Mexicanground squirrels or Richardson’s ground squirrels (Reiter et al. 1982, 1983; Reiter & Peters1984). In humans, two-hour exposure to light of 500 lux was insufficient to affect nocturnalmelatonin (Lewy et al. 1980) and it was recently suggested that the circadian system ofhumans is roughly 10 000 times less sensitive to light than the mouse circadian system(Bullough et al. 2006). Taken together, these studies suggest the possibility that diurnalanimals, including humans, may have a decreased photic sensitivity compared tonocturnal animals. Systematic studies of neural, genetic, hormonal, and behaviouralresponses of the circadian systems to light must be conducted before such a conclusion canbe made regarding general differences in photic sensitivity, however. This is a particularlyinteresting question given the importance of light in maintaining alertness and clocksynchrony in people involved in shift work and in the treatment of rhythm disruptions(Boivin & James 2002; Burgess et al. 2002; Horowitz & Tanigawa 2002; Kuller 2002; Yoonet al. 2002; Stevens 2005).
294 C.M. Novak et al.

Nonphotic synchronization
Zeitgebers other than light are also able to phase shift the circadian clock. Very few data havebeen gathered on the subject of nonphotic phase adjustment in diurnal animals incomparison to the body of evidence existing for nocturnal rodents. Moreover, many of thestudies on nonphotic stimulation in diurnal animals have used timed daily nonphoticstimulation to entrain rhythms. Most studies of nocturnal species employ delivery of a singleacute phase-shifting stimulus to generate PRCs. This difference inmethodologymakes directcomparisons to many nocturnal animals difficult. Using the literature on nocturnal animals(Hastings et al. 1998) as a starting point, we will examine the neural systems mediatingnonphotic phase alteration and compare these findings with those of diurnal rodents.
First, although melatonin is a relatively weak zeitgeber, it has a distinct PRC innocturnal rodents, inducing phase advances before the onset of subjective night (VanReeth et al. 1997). Accordingly, melatonin given during this same time period (the latesubjective day) effectively entrained the rhythms of free-running rats (Redman 1997). Anadditional period of sensitivity is sometimes found during the late subjective night(Redman 1997). This pattern of entrainment is comparable in diurnal rodents. InFunambulus pennati and Arvicanthis ansorgei, melatonin entrains circadian rhythms whenadministered near subjective dawn or dusk, with a trend for a greater sensitivity tomelatonin at subjective dusk in the F. pennati (Rajaratnam & Redman 1997; Slotten et al.2005). A similar effect was found in humans, with sensitivity to melatonin during thelight – dark transitions and with entrainment of rhythms in free-running blind subjects(Lewy et al. 1992, 1996, 2005; Lewy & Sack 1996). In general, the temporal profile ofmelatonin sensitivity seems to be similar in diurnal and nocturnal animals, althoughdiurnal animals may be more sensitive to melatonin at dawn. This apparent trend may bedue to methodological differences or variability among species, however.
The most common nonphotic stimuli investigated are those that induce phase advancesduring the subjective day in nocturnal animals (Yannielli & Harrington 2004). Serotonergicstimulation, forced arousal with a novel wheel or sleep deprivation, and application of NPYinto the SCN region all induce marked phase advances in the mid-subjective day (Ellis et al.1982; Albers et al. 1984; Hastings et al. 1998; Ehlen et al. 2001; Glass et al. 2003; Mistlbergeret al. 2003; Yannielli & Harrington 2004). The question of how this class of nonphoticstimuli act in diurnal animals is especially intriguing given the relationship between thestimulus and the activity phase. If forced arousal induces phase advances in nocturnalanimals during their subjective daytime (inactive phase), howmight diurnal animals respondto the same stimulus? Is the effect dependent on the phase of the lighting cycle or the activitycycle? As discussed above, very few PRCs to nonphotic stimuli have been generated indiurnal species. The ability of arousal to alter phase in diurnal animals has been studiedmostly using a scheduled daily activity protocol, with the goal of synchronizing the circadianactivity rhythm to these stimuli (Mistlberger 1991a, 1991b; Mistlberger et al. 1991; Hut et al.1999; Glass et al. 2001; Kas & Edgar 2001). In a diurnal primate, the marmoset, daily one-hour bouts of forced activity caused phase delays at the end of the subjective night andtemporarily entrained their daily rhythm when the forced activity coincided with thebeginning of the activity phase (Glass et al. 2001). Similarly, voluntary wheel running for ascheduled two-hour duration using Octodon degus resulted in entrainment when the wheelwas given at the end of the animals’ subjective night. These data suggest that, in diurnalspecies, (1) activity is a relatively weak zeitgeber, and (2) nonphotic stimuli induce smallphase delays at the end of the subjective night (Kas & Edgar 2001). In diurnal Europeanground squirrels, on the other hand, over half of the animals were entrained by three hours
Biological Rhythm Research 295
of confinement to a running wheel when the stimulus coincided with the end of the activityphase (Hut et al. 1999). Taken together, these studies do not yield a cohesive story explainingactivity-induced nonphotic phase synchronization in diurnal and nocturnal rodents,possibly due to inter-species variation in the effect of arousal on rhythm synchronizationunrelated to diurnality or nocturnality.
With regard to activity and rhythmicity in diurnal animals, another trend is observed.Access to a wheel can acutely alter chronotype in a subset of degus and grass rats so thatthey appear to be crepuscular or nocturnal rather than diurnal (Katona & Smale 1997;Blanchong et al. 1999; Kas & Edgar 1999). Two lines of evidence suggest that the wheeldoes not actually alter circadian phase but rather temporarily changes phase preference.First, no transient activity onsets are evidenced upon introduction of the wheel, whichwould be expected if a phase shift occurred (Blanchong et al. 1999). Second, Redlin andMrosovsky (2004) demonstrated that grass rats with access to a running wheel during aseries of 30-min light – dark transitions were active during the dark times, whereas grassrats that could not run in wheels preferred to be active during the light periods (Redlin &Mrosovsky 2004). This supports the idea that masking (the direct effect of light ordarkness on activity), rather than circadian phase or diurnality, interacts with the‘‘nonphotic’’ wheel running stimulus to alter phase preferences (Blanchong et al. 1999; Kas& Edgar 1999; Redlin & Mrosovsky 2004). It has been suggested that NPY projectionsfrom the IGL might mediate this effect in grass rats, as the Fos profile of NPY cells differsbetween day-active versus night-active grass rats; the SCN shows no apparent differencesbetween animals with different chronotypes (Labyak et al. 1997; Smale et al. 2001).
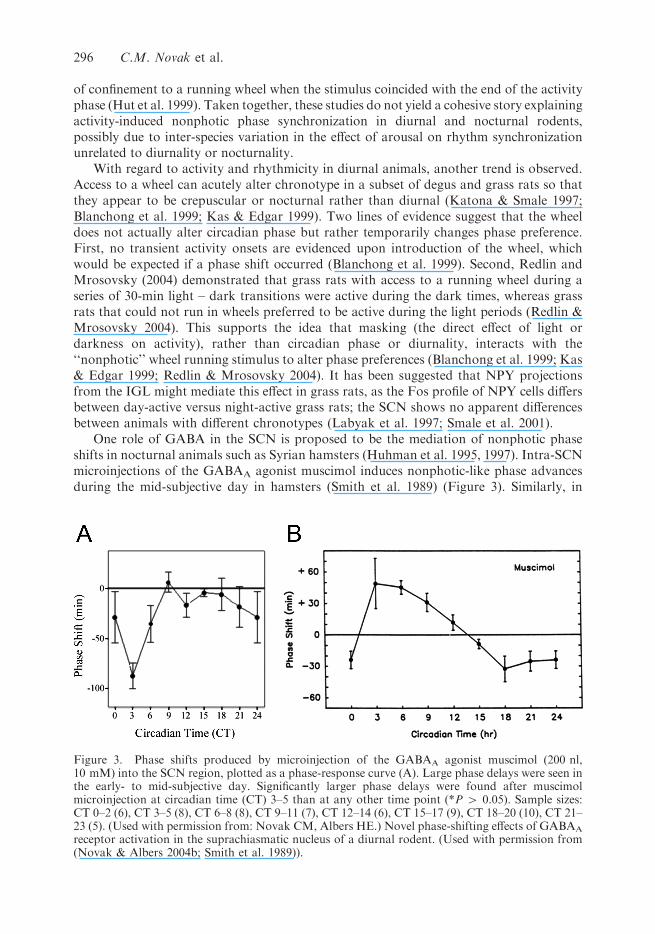
One role of GABA in the SCN is proposed to be the mediation of nonphotic phaseshifts in nocturnal animals such as Syrian hamsters (Huhman et al. 1995, 1997). Intra-SCNmicroinjections of the GABAA agonist muscimol induces nonphotic-like phase advancesduring the mid-subjective day in hamsters (Smith et al. 1989) (Figure 3). Similarly, in
Figure 3. Phase shifts produced by microinjection of the GABAA agonist muscimol (200 nl,10 mM) into the SCN region, plotted as a phase-response curve (A). Large phase delays were seen inthe early- to mid-subjective day. Significantly larger phase delays were found after muscimolmicroinjection at circadian time (CT) 3–5 than at any other time point (*P 4 0.05). Sample sizes:CT 0–2 (6), CT 3–5 (8), CT 6–8 (8), CT 9–11 (7), CT 12–14 (6), CT 15–17 (9), CT 18–20 (10), CT 21–23 (5). (Used with permission from: Novak CM, Albers HE.) Novel phase-shifting effects of GABAA
receptor activation in the suprachiasmatic nucleus of a diurnal rodent. (Used with permission from(Novak & Albers 2004b; Smith et al. 1989)).
296 C.M. Novak et al.

diurnal A. niloticus, microinjections of muscimol into the SCN produced phase shifts atroughly the same circadian time as nocturnal rodents (Novak & Albers 2004b; Smith et al.1989) (Figure 3). A major difference was also seen, however: the grass rats had large phasedelays, rather than the phase advances seen in nocturnal rodents. In each species, the phasealteration presumably functions to reset the clock in the direction of the onset of activity,phase advancing the hamsters toward subjective night and phase delaying grass ratstoward subjective dawn. Also, it appears that muscimol acts directly on rhythm-generatingcells to alter phase because blocking sodium-dependent action potentials is ineffective atinhibiting the phase-altering effects of muscimol (Gamble et al. 2003; Novak & Albers2004b). Clearly, as suggested earlier (Glass et al. 2001; Mrosovsky et al. 2001), this class ofnonphotic stimuli is similar to photic stimulation in that it is keyed to the phase of theSCN rather than to the activity phase of the animal.
Since muscimol-induced phase shifts occur during the subjective day in diurnal grassrats, it is not likely that the effect of this stimulus would be activity- or arousal-related, asin some nocturnal animals. Grass rats are already routinely active during the day and are,in general, very easily agitated (Blanchong et al. 1999). This raises the question of whatenvironmental stimulus may be responsible for inducing the GABAergic signal within theSCN resulting in these phase delays, if not arousal. Unpublished data from our laboratoryand that of Dr. Smale suggest that the following manipulations fail to alter circadian phasein grass rats during the subjective day: serotonergics, NPY at several doses (with orwithout concurrent light exposure), a NPY-receptor antagonist, novel wheels, restraintstress, carbachol, melatonin, dark pulses, wheel deprivation, or social/sexual experiencewith a novel conspecific. Indeed, with the exception of light and muscimol, these animalsare surprisingly resistant to acute phase alteration. It is interesting that the neuroanato-mical pathways that mediate nonphotic phase shifting in nocturnal animals are highlyconserved in diurnal animals including A. niloticus (Goel et al. 1999; Smale et al. 1991;Smale & Boverhof 1999; Castillo-Ruiz et al. 2006; Vidal & Lugo 2006). The question thatis then raised is, if serotonergic and NPY innervation of the SCN is not likely to mediatenonphotic synchronization in diurnal animals, what is the function of these inputs (Vidal& Lugo 2006)? As described above, NPY inputs into the diurnal clock may serve otherfunctions (Vidal & Lugo 2006). For example, it is possible that one role of NPY in grassrats is to modulate the masking effect of darkness and activity (i.e. wheel access) (Vidal &Lugo 2006). It is conceivable that serotonergic and cholinergic projections to the SCN ofdiurnal animals serve other purposes as well.
The previously described mechanism underlying photic phase shifting—the inductionof expression of clock genes such as Per1 and Per2—seems to be similar in diurnal andnocturnal animals (Dardente et al. 2002; Caldelas et al. 2003; Novak et al. 2006;Ramanathan et al. 2006). For nonphotic phase shifting in nocturnal animals, thepreponderance of information supports the idea that these phase shifts are induced bydecreasing the expression of Per1 and Per2 in the SCN (Maywood et al. 1999; Fukuharaet al. 2001; Maywood et al. 2002; Fukuhara 2004; Ehlen et al. 2006). Following theobservation that the temporal pattern of clock gene expression in the SCN of diurnalground squirrels was the same as in nocturnal rodents, it was suggested by Mrosovskyet al. (2001) that the diurnal SCN is also sensitive to nonphotic phase alteration during thesubjective day (Mrosovsky et al. 2001). This idea, based on the supposition that theuniversal mechanism driving nonphotic phase shifting is decreased Per expression, hasbeen supported by subsequent studies (Hastings et al. 1997; Mrosovsky et al. 2001; Novak& Albers 2004b; Novak et al. 2006). Further, we have found that muscimol administeredduring the day not only induced phase delays in grass rats, but also decreased SCN Per2
Biological Rhythm Research 297

mRNA levels (Novak et al. 2006). Lastly, photic and nonphotic stimuli often interact tocancel out each others’ effects (Yannielli & Harrington 2004): light-induced phase shiftsduring the subjective night are inhibited by NPY, muscimol, and nonphotic behaviouralstimulation (Gillespie et al. 1996, 1997; Weber et al. 1998; Challet et al. 2001; Yannielli &Harrington 2001; Gamble et al. 2004; Novak &Albers 2004a), and daytime nonphotic phasedelays are decreased by light exposure or light-like stimulation (Ralph & Mrosovsky 1992;Ehlen et al. 2001; Yannielli & Harrington 2001; Gamble et al. 2004). The mechanismunderlying this interaction may be the opposing effects of photic and nonphotic stimuli onthe expression of SCN clock genes such as Per1 and Per2 (Christian & Harrington 2002;Maywood et al. 2002; Edelstein et al. 2003; Mendoza et al. 2004; Sanggaard et al. 2003;Yannielli & Harrington 2004; Ehlen et al. 2006). In diurnal A. niloticus, we found that light-induced phase delays are inhibited by microinjections of muscimol into the SCN (Novak &Albers 2004a). On the other hand, unlike nocturnal animals, muscimol-induced daytimephase delays in grass rats appear to be completely unaffected by 15 min of exposure torelatively bright light (Novak & Albers 2004a). This dichotomy makes sense when onecompares the natural environment of the grass rats to that of nocturnal rodents. Grass ratsare regularly exposed to extremely bright light at least intermittently during the subjectiveday (Blanchong et al. 1999), while nocturnal rodents are typically holed up safely in aburrow during the day. If light inhibits nonphotic phase shifts in grass rats, the ability of anynonphotic stimulus to alter phase would effectively be nullified. This differential effect oflight on nonphotic or GABA-induced phase shifts also raises the possibility of mechanisticdifferences. If light increases clock gene expression in the SCN of diurnal rodents, at leastduring the night (Novak et al. 2006), why is light virtually ineffective at inhibiting GABA-induced phase delays? The cellular and neural mechanisms underlying the interactions ofphotic and nonphotic stimuli may also differ between diurnal and nocturnal animals.
Summary and questions
Whether an animal is diurnal or nocturnal, the phase of the SCN rhythm of sensitivity tozeitgebers is linked to the light – dark cycle rather than to the animal’s activity cycle.Although the functioning of the SCN is surprisingly similar among animals active duringthe day or night, one functional difference has been identified. Activation of receptors forGABA, the primary neurotransmitter of the SCN, results in phase shifts in oppositedirections in diurnal and nocturnal animals. The cellular mechanism underlying this effect,namely muscimol inhibition of Per mRNA, appears to be similar across species, however(Ehlen et al. 2006; Novak et al. 2006). Together, these studies evoke several questions:
. Is the sensitivity of the diurnal circadian system to light decreased compared tonocturnal animals? If so, what is the mechanism of this change?
. Is the ability of GABAA receptor stimulation to induce phase delays a generalproperty of diurnal circadian systems?
. How does muscimol induce phase delays in diurnal animals but advances innocturnal animals?
. If NPY and 5-HT are not integral to nonphotic phase resetting in diurnal animals,then what is the role of these systems in diurnal animals? Are they coupled to SCNGABA in nocturnal animals but decoupled in diurnal animals?
. Are there environmental or behavioural stimuli that induce daytime phase delays ingrass rats? Clearly, stimuli other than arousal or activity should be considered. Forexample, a large body of evidence suggests that social and olfactory stimuli affect the
298 C.M. Novak et al.

ability of diurnal degus to reset their circadian clock (Lee, this issue). With respect tograss rats and daytime phase delays, focusing on a behaviour associated with dawnmay provide likely candidates.
. Since most studies on nocturnal nonphotic entrainment focus on forced arousal, andlittle evidence supports a role for arousal in diurnal phase resetting, how much of theinformation regarding nonphotic phase resetting and its underlying mechanisms canbe generalized to the human population?
. What do these findings regarding nonphotic stimulation mean for human circadianrhythmicity, and are there therapeutic implications for circadian disorders?
With the emerging recognition of the importance of circadian rhythmicity in cognitiveabilities, disease, and quality of life (Gorbacheva et al. 2005; Winocur & Hasher 1999), it isespecially critical to understand how to effectively synchronize our circadian rhythms. Thisunderscores the importance of continuing to further understand the principles andmechanisms underlying phase synchronization in diurnal animals.
References
Abrahamson EE, Moore RY. 2001. Suprachiasmatic nucleus in the mouse: retinal innervation,
intrinsic organization and efferent projections. Brain Res. 916(1–2):172–191.Albers HE, Ferris CF, Leeman SE, Goldman BD. 1984. Avian pancreatic polypeptide phase shifts
hamster circadian rhythms when microinjected into the suprachiasmatic region. Science.223(4638):833–835.
Blanchong JA, McElhinny TL, Mahoney MM, Smale L. 1999. Nocturnal and diurnal rhythms in theunstriped Nile rat, Arvicanthis niloticus. J Biol Rhythms. 14(5):364–377.
Boivin DB, James FO. 2002. Circadian adaptation to night-shift work by judicious light and
darkness exposure. J Biol Rhythms. 17(6):556–567.Bronchti G, Rado R, Terkel J, Wollberg Z. 1991. Retinal projections in the blind mole rat: a WGA-
HRP tracing study of a natural degeneration. Brain Res Dev Brain Res. 58(2):159–170.
Bullough JD, Rea MS, Figueiro MG. 2006. Of mice and women: light as a circadian stimulus inbreast cancer research. Cancer Causes Control. 17(4):375–383.
Burgess HJ, Sharkey KM, Eastman CI. 2002. Bright light, dark and melatonin can promotecircadian adaptation in night shift workers. Sleep Med Rev. 6(5):407–420.
Caldelas I, Poirel VJ, Sicard B, Pevet P, Challet E. 2003. Circadian profile and photic regulation ofclock genes in the suprachiasmatic nucleus of a diurnal mammal Arvicanthis ansorgei.Neuroscience. 116(2):583–591.
Castillo-Ruiz A, Smale L, Nunez AA. 2006. Cholinergic projections to the suprachiasmatic nucleus(SCN) of the diurnal grass rat, #75. Sandestin (FL): Society for Research on BiologicalRhythms. p. 71.
Challet E, Turek FW, Laute M, Van Reeth O. 2001. Sleep deprivation decreases phase-shiftresponses of circadian rhythms to light in the mouse: role of serotonergic and metabolic signals.Brain Res. 909(1–2):81–91.
Christian CA, Harrington ME. 2002. Three days of novel wheel access diminishes light-induced phase delays in vivo with no effect on per1 induction by light. Chronobiol Int. 19(4):671–682.
Dardente H, Klosen P, Caldelas I, Pevet P, Masson-Pevet M. 2002. Phenotype of Per1- and Per2-
expressing neurons in the suprachiasmatic nucleus of a diurnal rodent (Arvicanthis ansorgei):comparison with a nocturnal species, the rat. Cell Tissue Res. 310(1):85–92.
Edelstein K, de la Iglesia HO, Schwartz WJ, Mrosovsky N. 2003. Behavioral arousal blocks light-
induced phase advances in locomotor rhythmicity but not light-induced Per1 and Fos expressionin the hamster suprachiasmatic nucleus. Neuroscience. 118(1):253–261.
Biological Rhythm Research 299

Ehlen JC, Grossman GH, Glass JD. 2001. In vivo resetting of the hamster circadian clock by 5-HT7
receptors in the suprachiasmatic nucleus. J Neurosci. 21(14):5351–5357.Ehlen JC, Novak CM, Karom MC, Gamble KL, Paul KN, Albers HE. 2006. GABAA receptor
activation suppresses Period 1 mRNA and Period 2 mRNA in the suprachiasmatic nucleus
during the mid-subjective day. Eur J Neurosci. 23(12):3328–3336.Ellis GB, McKlveen RE, Turek FW. 1982. Dark pulses affect the circadian rhythm of activity in
hamsters kept in constant light. Am J Physiol. 242(1):R44–50.Fite KV, Janusonis S. 2001. Retinal projection to the dorsal raphe nucleus in the Chilean degus
(Octodon degus). Brain Res. 895(1–2):139–145.Fite KV, Janusonis S, Foote W, Bengston L. 1999. Retinal afferents to the dorsal raphe nucleus in
rats and Mongolian gerbils. J Comp Neurol. 414(4):469–484.
Fukuhara C. 2004. Effect of dark exposure in the middle of the day on Period1, Period2, andarylalkylamine N-acetyltransferase mRNA levels in the rat suprachiasmatic nucleus and pinealgland. Brain Res Mol Brain Res. 130(1–2):109–114.
Fukuhara C, Brewer JM, Dirden JC, Bittman EL, Tosini G, Harrington ME. 2001. Neuropeptide Yrapidly reduces Period 1 and Period 2 mRNA levels in the hamster suprachiasmatic nucleus.Neurosci Lett. 314(3):119–122.
Gamble KL, Novak CM, Albers HE. 2004. Neuropeptide Y and N-methyl-D-aspartic acid interact
within the suprachiasmatic nuclei to alter circadian phase. Neuroscience. 126(3):559–565.Gamble KL, Novak CM, Paul KN, Albers HE. 2003. Tetrodotoxin blocks the circadian effects of
NMDA during the day but not at night. Neuroreport. 14(4):641–644.
Gillespie CF, Huhman KL, Babagbemi TO, Albers HE. 1996. Bicuculline increases and muscimolreduces the phase-delaying effects of light and VIP/PHI/GRP in the suprachiasmatic region. JBiol Rhythms. 11(2):137–144.
Gillespie CF, Mintz EM, Marvel CL, Huhman KL, Albers HE. 1997. GABA(A) and GABA(B)agonists and antagonists alter the phase-shifting effects of light when microinjected into thesuprachiasmatic region. Brain Res. 759(2):181–189.
Glass JD, Grossman GH, Farnbauch L, DiNardo L. 2003. Midbrain raphe modulation ofnonphotic circadian clock resetting and 5-HT release in the mammalian suprachiasmaticnucleus. J Neurosci. 23(20):7451–7460.
Glass JD, Tardif SD, Clements R, Mrosovsky N. 2001. Photic and nonphotic circadian phase
resetting in a diurnal primate, the common marmoset. Am J Physiol Regul Integr Comp Physiol.280(1):R191–197.
Goel N, Lee TM, Smale L. 1999. Suprachiasmatic nucleus and intergeniculate leaflet in the diurnal
rodent Octodon degus: retinal projections and immunocytochemical characterization. Neu-roscience. 92(4):1491–1509.
Gorbacheva VY, Kondratov RV, Zhang R, Cherukuri S, Gudkov AV, Takahashi JS, Antoch MP.
2005. Circadian sensitivity to the chemotherapeutic agent cyclophosphamide depends on thefunctional status of the CLOCK/BMAL1 transactivation complex. Proc Natl Acad Sci USA.102(9):3407–3412.
Hastings MH, Duffield GE, Ebling FJ, Kidd A, Maywood ES, Schurov I. 1997. Non-photicsignalling in the suprachiasmatic nucleus. Biol Cell. 89(8):495–503.
Hastings MH, Duffield GE, Smith EJ, Maywood ES, Ebling FJ. 1998. Entrainment of the circadiansystem of mammals by nonphotic cues. Chronobiol Int. 15(5):425–445.
Horowitz TS, Tanigawa T. 2002. Circadian-based new technologies for night workers. Ind Health.40(3):223–236.
Huhman KL, Babagbemi TO, Albers HE. 1995. Bicuculline blocks neuropeptide Y-induced phase
advances when microinjected in the suprachiasmatic nucleus of Syrian hamsters. Brain Res.675(1–2):333–336.
Huhman KL, Marvel CL, Gillespie CF, Mintz EM, Albers HE. 1997. Tetrodotoxin blocks NPY-
induced but not muscimol-induced phase advances of wheel-running activity in Syrian hamsters.Brain Res. 772(1–2):176–180.
300 C.M. Novak et al.

Hut RA, Mrosovsky N, Daan S. 1999. Nonphotic entrainment in a diurnal mammal, the European
ground squirrel (Spermophilus citellus). J Biol Rhythms. 14(5):409–419.Jiao YY, Lee TM, Rusak B. 1999. Photic responses of suprachiasmatic area neurons in diurnal degus
(Octodon degus) and nocturnal rats (Rattus norvegicus). Brain Res. 817(1–2):93–103.
Kas MJ, Edgar DM. 1999. A nonphotic stimulus inverts the diurnal-nocturnal phase preference inOctodon degus. J Neurosci. 19(1):328–333.
Kas MJ, Edgar DM. 2001. Scheduled voluntary wheel running activity modulates free-runningcircadian body temperature rhythms in Octodon degus. J Biol Rhythms. 16(1):66–75.
Katona C, Smale L. 1997. Wheel-running rhythms in Arvicanthis niloticus. Physiol Behav. 61(3):365–372.
Kita H, Oomura Y. 1982. An anterograde HRP study of retinal projections to the hypothalamus in
the rat. Brain Res Bull. 8(3):249–253.Kuller R. 2002. The influence of light on circarhythms in humans. J Physiol Anthropol Appl Human
Sci. 21(2):87–91.
Kume K, Zylka MJ, Sriram S, Shearman LP, Weaver DR, Jin X, Maywood ES, Hastings MH,Reppert SM. 1999. mCRY1 and mCRY2 are essential components of the negative limb of thecircadian clock feedback loop. Cell. 98(2):193–205.
Labyak SE, Lee TM, Goel N. 1997. Rhythm chronotypes in a diurnal rodent, Octodon degus. Am J
Physiol. 273(3 Pt 2):R1058–R1066.Lewy AJ, Ahmed S, Jackson JM, Sack RL. 1992. Melatonin shifts human circadian rhythms
according to a phase-response curve. Chronobiol Int. 9(5):380–392.
Lewy AJ, Ahmed S, Sack RL. 1996. Phase shifting the human circadian clock using melatonin.Behav Brain Res. 73(1–2):131–134.
Lewy AJ, Emens JS, Lefler BJ, Yuhas K, Jackman AR. 2005. Melatonin entrains free-running
blind people according to a physiological dose-response curve. Chronobiol Int. 22(6):1093–1106.
Lewy AJ, Sack RL. 1996. The role of melatonin and light in the human circadian system. Prog Brain
Res. 111:205–216.Lewy AJ, Wehr TA, Goodwin FK, Newsome DA, Markey SP. 1980. Light suppresses melatonin
secretion in humans. Science. 210(4475):1267–1269.Lincoln G, Messager S, Andersson H, Hazlerigg D. 2002. Temporal expression of seven clock genes
in the suprachiasmatic nucleus and the pars tuberalis of the sheep: evidence for an internalcoincidence timer. Proc Natl Acad Sci USA. 99(21):13890–13895.
Lu J, Shiromani P, Saper CB. 1999. Retinal input to the sleep-active ventrolateral preoptic nucleus in
the rat. Neuroscience. 93(1):209–214.Mahoney M, Bult A, Smale L. 2001. Phase response curve and light-induced fos expression in the
suprachiasmatic nucleus and adjacent hypothalamus of Arvicanthis niloticus. J Biol Rhythms.
16(2):149–162.Major DE, Rodman HR, Libedinsky C, Karten HJ. 2003. Pattern of retinal projections in the
California ground squirrel (Spermophilus beecheyi): anterograde tracing study using cholera
toxin. J Comp Neurol. 463(3):317–340.Maywood ES, Mrosovsky N, Field MD, Hastings MH. 1999. Rapid down-regulation of mammalian
period genes during behavioral resetting of the circadian clock. Proc Natl Acad Sci USA.96(26):15211–15216.
Maywood ES, Okamura H, Hastings MH. 2002. Opposing actions of neuropeptide Y and light onthe expression of circadian clock genes in the mouse suprachiasmatic nuclei. Eur J Neurosci.15(1):216–220.
Mendoza JY, Dardente H, Escobar C, Pevet P, Challet E. 2004. Dark pulse resetting of thesuprachiasmatic clock in Syrian hamsters: behavioral phase-shifts and clock gene expression.Neuroscience. 127(2):529–537.
Mintz EM, Albers HE. 1997. Microinjection of NMDA into the SCN region mimics the phaseshifting effect of light in hamsters. Brain Res. 758(1–2):245–249.
Biological Rhythm Research 301

Mintz EM, Marvel CL, Gillespie CF, Price KM, Albers HE. 1999. Activation of NMDA receptors
in the suprachiasmatic nucleus produces light-like phase shifts of the circadian clock in vivo. JNeurosci. 19(12):5124–5130.
Mistlberger RE. 1991a. Effects of daily schedules of forced activity on free-running rhythms in the
rat. J Biol Rhythms. 6(1):71–80.Mistlberger RE. 1991b. Scheduled daily exercise or feeding alters the phase of photic entrainment in
Syrian hamsters. Physiol Behav. 50(6):1257–1260.Mistlberger RE, Antle MC, Webb IC, Jones M, Weinberg J, Pollock MS. 2003. Circadian clock
resetting by arousal in Syrian hamsters: the role of stress and activity. Am J Physiol Regul IntegrComp Physiol. 285(4):R917–R925.
Mistlberger RE, Houpt TA, Moore-Ede MC. 1991. The benzodiazepine triazolam phase-shifts
circadian activity rhythms in a diurnal primate, the squirrel monkey (Saimiri sciureus). NeurosciLett. 124(1):27–30.
Moore RY, Danchenko RL. 2002. Paraventricular-subparaventricular hypothalamic lesions
selectively affect circadian function. Chronobiol Int. 19(2):345–360.Morin LP, Allen CN. 2006. The circadian visual system, 2005. Brain Res Brain Res Rev. 51(1):1–60.Mrosovsky N, Edelstein K, Hastings MH, Maywood ES. 2001. Cycle of period gene expression in a
diurnal mammal (Spermophilus tridecemlineatus): implications for nonphotic phase shifting. J
Biol Rhythms. 16(5):471–478.Nelson DE, Takahashi JS. 1991. Comparison of visual sensitivity for suppression of pineal
melatonin and circadian phase-shifting in the golden hamster. Brain Res. 554(1–2):272–277.
Novak CM, Albers HE. 2002. N-Methyl-D-aspartate microinjected into the suprachiasmatic nucleusmimics the phase-shifting effects of light in the diurnal Nile grass rat (Arvicanthis niloticus). BrainRes. 951(2):255–263.
Novak CM, Albers HE. 2004a. Circadian phase alteration by GABA and light differs in diurnal andnocturnal rodents during the day. Behav Neurosci. 118(3):498–504.
Novak CM, Albers HE. 2004b. Novel phase-shifting effects of GABAA receptor activation in the
suprachiasmatic nucleus of a diurnal rodent. Am J Physiol Regul Integr Comp Physiol.286(5):R820–R825.
Novak CM, Ehlen JC, Paul KN, Fukuhara C, Albers HE. 2006. Light and GABAA receptoractivation alter Period mRNA levels in the SCN of diurnal Nile grass rats. Eur J Neurosci.
24:2843–2852.Novak CM, Smale L, Nunez AA. 1999. Fos expression in the sleep-active cell group of the
ventrolateral preoptic area in the diurnal murid rodent, Arvicanthis niloticus. Brain Res.
818(2):375–382.Nuesslein-Hildesheim B, O’Brien JA, Ebling FJ, Maywood ES, Hastings MH. 2000. The circadian
cycle of mPER clock gene products in the suprachiasmatic nucleus of the siberian hamster
encodes both daily and seasonal time. Eur J Neurosci. 12(8):2856–2864.Oster H, Avivi A, Joel A, Albrecht U, Nevo E. 2002. A switch from diurnal to nocturnal activity in
S. ehrenbergi is accompanied by an uncoupling of light input and the circadian clock. Curr Biol.
12(22):1919–1922.Paul KN, Fukuhara C, Tosini G, Albers HE. 2003. Transduction of light in the suprachiasmatic
nucleus: evidence for two different neurochemical cascades regulating the levels of Per1 mRNAand pineal melatonin. Neuroscience. 119(1):137–144.
Paul KN, Gamble KL, Fukuhara C, Novak CM, Tosini G, Albers HE. 2004. Tetrodotoxinadministration in the suprachiasmatic nucleus prevents NMDA-induced reductions in pinealmelatonin without influencing Per1 and Per2 mRNA levels. Eur J Neurosci. 19(10):2808–2814.
Pickard GE. 1982. The afferent connections of the suprachiasmatic nucleus of the golden hamsterwith emphasis on the retinohypothalamic projection. J Comp Neurol. 211(1):65–83.
Pickard GE, Silverman AJ. 1981. Direct retinal projections to the hypothalamus, piriform cortex,
and accessory optic nuclei in the golden hamster as demonstrated by a sensitive anterogradehorseradish peroxidase technique. J Comp Neurol. 196(1):155–172.
302 C.M. Novak et al.

Rajaratnam SM, Redman JR. 1997. Effects of daily melatonin administration on circadian activity
rhythms in the diurnal Indian palm squirrel (Funambulus pennanti). J Biol Rhythms. 12(4):339–347.
Ralph MR, Mrosovsky N. 1992. Behavioral inhibition of circadian responses to light. J Biol
Rhythms. 7(4):353–359.Ramanathan C, Nunez AA, Martinez GS, Schwartz MD, Smale L. 2006. Temporal and spatial
distribution of immunoreactive PER1 and PER2 proteins in the suprachiasmatic nucleus andperi-suprachiasmatic region of the diurnal grass rat (Arvicanthis niloticus). Brain Res. 1073–
1074:348–358.Redlin U, Mrosovsky N. 2004. Nocturnal activity in a diurnal rodent (Arvicanthis niloticus): the
importance of masking. J Biol Rhythms. 19(1):58–67.
Redman JR. 1997. Circadian entrainment and phase shifting in mammals with melatonin. J BiolRhythms. 12(6):581–587.
Reiter RJ, Hurlbut EC, Brainard GC, Steinlechner S, Richardson BA. 1983. Influence of light
irradiance on hydroxyindole-O-methyltransferase activity, serotonin-N-acetyltransferase activ-ity, and radioimmunoassayable melatonin levels in the pineal gland of the diurnally activeRichardson’s ground squirrel. Brain Res. 288(1–2):151–157.
Reiter RJ, King TS, Richardson BA, Hurlbut EC. 1982. Studies on pineal melatonin levels in a
diurnal species, the eastern chipmunk (Tamias striatus): effects of light at night, propranololadministration or superior cervical ganglionectomy. J Neural Transm. 54(3–4):275–284.
Reiter RJ, Peters JF. 1984. Non-suppressibility by room light of pineal N-acetyltransferase activity
and melatonin levels in two diurnally active rodents, the Mexican ground squirrel (Spermophilusmexicanus) and the eastern chipmunk (Tamias striatus). Endocr Res. 10(2):113–121.
Sanggaard KM, Hannibal J, Fahrenkrug J. 2003. Serotonin inhibits glutamate- but not PACAP-
induced per gene expression in the rat suprachiasmatic nucleus at night. Eur J Neurosci.17(6):1245–1252.
Schwartz MD, Nunez AA, Smale L. 2004. Differences in the suprachiasmatic nucleus and lower
subparaventricular zone of diurnal and nocturnal rodents. Neuroscience. 127(1):13–23.Schwartz MD, Smale L. 2005. Individual differences in rhythms of behavioral sleep and its neural
substrates in Nile grass rats. J Biol Rhythms. 20(6):526–537.Slotten HA, Krekling S, Pevet P. 2005. Photic and nonphotic effects on the circadian activity rhythm
in the diurnal rodent Arvicanthis ansorgei. Behav Brain Res. 165(1):91–97.Smale L, Blanchard J, Moore RY, Morin LP. 1991. Immunocytochemical characterization of the
suprachiasmatic nucleus and the intergeniculate leaflet in the diurnal ground squirrel,
Spermophilus lateralis. Brain Res. 563(1–2):77–86.Smale L, Boverhof J. 1999. The suprachiasmatic nucleus and intergeniculate leaflet of Arvicanthis
niloticus, a diurnal murid rodent from East Africa. J Comp Neurol. 403(2):190–208.
Smale L, Castleberry C, Nunez AA. 2001. Fos rhythms in the hypothalamus of Rattus andArvicanthis that exhibit nocturnal and diurnal patterns of rhythmicity. Brain Res. 899(1–2):101–105.
Smith RD, Inouye S, Turek FW. 1989. Central administration of muscimol phase-shifts themammalian circadian clock. J Comp Physiol (A). 164(6):805–814.
Stevens RG. 2005. Circadian disruption and breast cancer: from melatonin to clock genes.Epidemiology. 16(2):254–258.
Sun ZS, Albrecht U, Zhuchenko O, Bailey J, Eichele G, Lee CC. 1997. RIGUI, a putativemammalian ortholog of the Drosophila period gene. Cell. 90(6):1003–1011.
Tei H, Okamura H, Shigeyoshi Y, Fukuhara C, Ozawa R, Hirose M, Sakaki Y. 1997. Circadian
oscillation of a mammalian homologue of the Drosophila period gene. Nature. 389(6650):512–516.
Van Reeth O, Olivares E, Zhang Y, Zee PC, Mocaer E, Defrance R, Turek FW. 1997. Comparative
effects of a melatonin agonist on the circadian system in mice and Syrian hamsters. Brain Res.762(1–2):185–194.
Biological Rhythm Research 303

Vidal L, Lugo N. 2006. Changes in neuropeptide Y immunoreactivity and transcript levels in
circadian system structures of the diurnal rodent, the thirteen-lined ground squirrel. Brain Res.1125(1):77–84.
Weber ET, Gannon RL, Rea MA. 1998. Local administration of serotonin agonists blocks light-
induced phase advances of the circadian activity rhythm in the hamster. J Biol Rhythms.13(3):209–218.
Winocur G, Hasher L. 1999. Aging and time-of-day effects on cognition in rats. Behav Neurosci.113(5):991–997.
Yannielli P, Harrington ME. 2004. Let there be ‘‘more’’ light: enhancement of light actions on thecircadian system through non-photic pathways. Prog Neurobiol. 74(1):59–76.
Yannielli PC, Harrington ME. 2001. The neuropeptide Y Y5 receptor mediates the blockade of
‘‘photic-like’’ NMDA-induced phase shifts in the golden hamster. J Neurosci. 21(14):5367–5373.Yoon IY, Jeong DU, Kwon KB, Kang SB, Song BG. 2002. Bright light exposure at night and light
attenuation in the morning improve adaptation of night shift workers. Sleep. 25(3):351–356.
Youngstrom TG, Nunez AA. 1986. Comparative anatomy of the retino-hypothalamic tract inphotoperiodic and non-photoperiodic rodents. Brain Res Bull. 17(4):485–492.
.
304 C.M. Novak et al.
Copyright © 2022 FDOKUMEN




















