
Calbindin-containing interneurons are a target for VIP-immunoreactive synapses in rat primary...
11
Calbindin-Containing Interneurons Are a Target for VIP-Immunoreactive Synapses in Rat Primary Somatosensory Cortex JOCHEN F. STAIGER, 1 * CARMEN MASANNECK, 1 AXEL SCHLEICHER, 1 AND WERNER ZUSCHRATTER 2 1 Heinrich-Heine-University Du ¨ sseldorf, C. & O. Vogt-Institute for Brain Research, D-40001 Du ¨ sseldorf 2 Leibniz-Institute for Neurobiology, D-39008 Magdeburg, Germany ABSTRACT Inhibitory interneurons in cerebral cortex are morphologically and physiologically ex- tremely heterogeneous. This greatly interferes with an understanding of their functions. Progress has been made by classifying these neurons with the aid of molecular markers, e.g., neuropeptides or calcium-binding proteins, which are reliably expressed by certain subpopu- lations. We have used this approach to demonstrate an output of a subpopulation of cortical interneurons which express vasoactive intestinal polypeptide (VIP). By double immunostain- ing and correlated light and electron microscopy, we show that calbindin (CB)-containing interneurons located in layers II–VI of rat barrel cortex are targets of symmetric VIP- immunoreactive synapses. All CB-immunoreactive interneurons showed numerous contacts of VIP boutons on proximal and distal dendritic segments. A great majority of CB- immunoreactive interneurons (214/222) displayed such close appositions with VIP boutons on their soma as well. Quantification revealed that the number of VIP-immunoreactive boutons on CB-immunoreactive somata and dendrites of specified order is comparable for the different cortical layers. In conclusion, all calbindin-containing cortical interneurons seem to be under direct influence of other GABAergic interneurons expressing the peptide VIP. An indirect functional consequence of this may be disinhibition of pyramidal cells, which are considered the major target of calbindin interneurons. However, since the examined types of interneu- rons are intricately embedded in networks of yet different interneurons, the outcome of these multiple inhibitory interactions is likely to be less simplistic. It may be related to the timing of pyramidal cell discharge within and across layers of cortical columns. J. Comp. Neurol. 468:179 –189, 2004. © 2003 Wiley-Liss, Inc. Indexing terms: barrel cortex; cortical circuitry; inhibitory interneurons; symmetric synapses; correlated light and electron microscopy A major step forward in the understanding of cortical circuits which process sensory information is the elucida- tion of specific inputs, intrinsic physiology, and specific outputs of identified cell types. Recent studies have shown an enormity of specificity for excitatory as well as inhibi- tory interneurons with respect to their morphology, elec- trophysiology, and neurochemistry (DeFelipe, 1993; Kawaguchi, 1995; Cauli et al., 1997; Gupta et al., 2000; Thomson et al., 2002). Circuitry organization revealed a cell type-specific pattern for a special class of interneurons containing the neuropeptide VIP (Rozov et al., 2001; Staiger et al., 2002). These mostly bipolar GABAergic neurons have long been recognized to reflect the basic cortical organization into columnar modules due to the vertical orientation of their dendritic and axonal trees (Morrison et al., 1984; Bayraktar et al., 1997, 2000). This latter feature makes them a prime candidate for intra- Grant sponsor: Deutsche Forschungsgemeinschaft; Grant number: Sta 431/2-4. Present address for Carmen Masanneck: Neuroaxo Biotec Ltd., Merow- inger Platz 1a, 40225 Du ¨ sseldorf, Germany. JFS and CM contributed equally to this work. *Correspondence to: Jochen F. Staiger, C. & O. Vogt-Institute for Brain Research, Heinrich-Heine-University Du ¨ sseldorf, P.O.B 101007, D-40001 Du ¨ sseldorf; Germany. E-mail: [email protected] Received 1 April 2003; Revised 14 July 2003; Accepted 21 August 2003 DOI 10.1002/cne.10953 Published online the week of November 17, 2003 in Wiley InterScience (www.interscience.wiley.com). THE JOURNAL OF COMPARATIVE NEUROLOGY 468:179 –189 (2004) © 2003 WILEY-LISS, INC.
-
Upload
independent -
Category
Documents
-
view
2 -
download
0
Transcript of Calbindin-containing interneurons are a target for VIP-immunoreactive synapses in rat primary...

Calbindin-Containing Interneurons Are aTarget for VIP-Immunoreactive Synapses
in Rat Primary Somatosensory Cortex
JOCHEN F. STAIGER,1* CARMEN MASANNECK,1 AXEL SCHLEICHER,1
AND WERNER ZUSCHRATTER2
1Heinrich-Heine-University Dusseldorf, C. & O. Vogt-Institute for Brain Research,D-40001 Dusseldorf
2Leibniz-Institute for Neurobiology, D-39008 Magdeburg, Germany
ABSTRACTInhibitory interneurons in cerebral cortex are morphologically and physiologically ex-
tremely heterogeneous. This greatly interferes with an understanding of their functions.Progress has been made by classifying these neurons with the aid of molecular markers, e.g.,neuropeptides or calcium-binding proteins, which are reliably expressed by certain subpopu-lations. We have used this approach to demonstrate an output of a subpopulation of corticalinterneurons which express vasoactive intestinal polypeptide (VIP). By double immunostain-ing and correlated light and electron microscopy, we show that calbindin (CB)-containinginterneurons located in layers II–VI of rat barrel cortex are targets of symmetric VIP-immunoreactive synapses. All CB-immunoreactive interneurons showed numerous contactsof VIP boutons on proximal and distal dendritic segments. A great majority of CB-immunoreactive interneurons (214/222) displayed such close appositions with VIP boutons ontheir soma as well. Quantification revealed that the number of VIP-immunoreactive boutonson CB-immunoreactive somata and dendrites of specified order is comparable for the differentcortical layers. In conclusion, all calbindin-containing cortical interneurons seem to be underdirect influence of other GABAergic interneurons expressing the peptide VIP. An indirectfunctional consequence of this may be disinhibition of pyramidal cells, which are consideredthe major target of calbindin interneurons. However, since the examined types of interneu-rons are intricately embedded in networks of yet different interneurons, the outcome of thesemultiple inhibitory interactions is likely to be less simplistic. It may be related to the timingof pyramidal cell discharge within and across layers of cortical columns. J. Comp. Neurol.468:179–189, 2004. © 2003 Wiley-Liss, Inc.
Indexing terms: barrel cortex; cortical circuitry; inhibitory interneurons; symmetric synapses;
correlated light and electron microscopy
A major step forward in the understanding of corticalcircuits which process sensory information is the elucida-tion of specific inputs, intrinsic physiology, and specificoutputs of identified cell types. Recent studies have shownan enormity of specificity for excitatory as well as inhibi-tory interneurons with respect to their morphology, elec-trophysiology, and neurochemistry (DeFelipe, 1993;Kawaguchi, 1995; Cauli et al., 1997; Gupta et al., 2000;Thomson et al., 2002). Circuitry organization revealed acell type-specific pattern for a special class of interneuronscontaining the neuropeptide VIP (Rozov et al., 2001;Staiger et al., 2002). These mostly bipolar GABAergicneurons have long been recognized to reflect the basiccortical organization into columnar modules due to thevertical orientation of their dendritic and axonal trees
(Morrison et al., 1984; Bayraktar et al., 1997, 2000). Thislatter feature makes them a prime candidate for intra-
Grant sponsor: Deutsche Forschungsgemeinschaft; Grant number: Sta431/2-4.
Present address for Carmen Masanneck: Neuroaxo Biotec Ltd., Merow-inger Platz 1a, 40225 Dusseldorf, Germany.
JFS and CM contributed equally to this work.*Correspondence to: Jochen F. Staiger, C. & O. Vogt-Institute for Brain
Research, Heinrich-Heine-University Dusseldorf, P.O.B 101007, D-40001Dusseldorf; Germany. E-mail: [email protected]
Received 1 April 2003; Revised 14 July 2003; Accepted 21 August 2003DOI 10.1002/cne.10953Published online the week of November 17, 2003 in Wiley InterScience
(www.interscience.wiley.com).
THE JOURNAL OF COMPARATIVE NEUROLOGY 468:179–189 (2004)
© 2003 WILEY-LISS, INC.

and interlaminar inhibitory mechanisms (Porter et al.,1998; Rozov et al., 2001) within a functional column, afeature, however, which they share with other types ofvertically organized interneurons (Kawaguchi and Kub-ota, 1997; Thomson and Bannister, 2003).
A growing interest in these VIP-containing interneu-rons has led to detailed descriptions of their morphology(Bayraktar et al., 2000), action potential firing patterns(Porter et al., 1998), chemical signature (Cauli et al.,2000), and modulation by transmitter-specified pathways(Kawaguchi, 1997; Porter et al., 1999; Ferezou et al.,2002), as well as specific thalamic and local cortical con-nectivity (Staiger et al., 1996, 2002). However, a preciseevaluation of their target structures which would allow abetter characterization of their roles in cortical informa-tion processing is still largely missing. At the electronmicroscopic level, profiles of presumably excitatory as wellas inhibitory target neurons were found (Hajos et al.,1988; Peters, 1990).
Other studies examined the neuronal populations in-nervated by calretinin-containing neurons. This class ofinhibitory interneurons is of special interest here, becausethey colocalize VIP to a very large extent (Rogers, 1992;Kubota et al., 1994). As for VIP neurons, mainly otherGABAergic cells, many of them containing calretininthemselves, were found as targets. This was shown forneocortex (Gabbott and Bacon, 1996; Meskenaite, 1997;Gonchar and Burkhalter, 1999) as well as archicortex(Gulyas et al., 1996). In the hippocampus, models do existthat VIP/calretinin coexpressing neurons may have a rolein synchronizing other ensembles of inhibitory interneu-rons, thus orchestrating the temporal structure of neuro-nal interactions which is thought to bring about thetarhythm and other synchronized population patterns(Freund and Buzsaki, 1996). It is interesting to note inthis context that behaviorally relevant rhythmic signalsare indeed occurring during the processing of tactile in-formation in primary somatosensory cortex (Jones andBarth, 1997, 1999). Since cortical and hippocampal VIPneurons show numerous common properties, a similarfunction might also pertain to the neocortical cell type.This was hypothesized previously by us (Staiger et al.,1997).
Here we characterize, at the population level, a majortarget of VIP-immunoreactive synapses, namely, inhibi-tory interneurons immunoreactive for the calcium-bindingprotein calbindin. By quantitatively mapping close spatialappositions of VIP-immunoreactive boutons withcalbindin-immunoreactive somata and dendrites, we showan abundance of such contacts at the light microscopiclevel. We further demonstrate the synaptic nature of thesecontacts by correlated light and electron microscopy,which implies a functional role of these connections.
MATERIALS AND METHODS
Perfusion of the animals and histologicalprocessing
All procedures were performed according to the Germanlaw on animal welfare, which includes all adequate mea-sures to minimize pain or discomfort. Male Wistar rats(250–350 g) from the local breeding facility of theHeinrich-Heine-University were used in the present study(n � 12). Animals were deeply anesthetized with pento-
barbital and perfused through the aorta, first with �30 mlof physiological saline, second with 100 ml of 5% acrolein,and third with 4% freshly depolymerized paraformalde-hyde containing 0.2% picric acid (both in 0.1 M phosphatebuffer (PB), pH 7.4) by hydrostatic pressure. Subse-quently, the brain was dissected out of the skull. After apostfixation of 2 hours in the last fixative, a block ofprimary somatosensory cortex was trimmed. The blockwas glued to the stage of a vibratome (Series 1000, TPI;Polyscience Europe, Eppelheim, Germany) with cyanoac-rylate and coronal sections of 50 �m thickness were cut.Sections were extensively rinsed in PB, cryoprotected (in asolution containing 25% saccharose and 10% glycerol in0.01 M PB), and freeze-thawed over liquid nitrogen threetimes. To remove unbound aldehydes from the tissue, allsections were immersed in 1% NaBH4 in 0.01 M PB for 10minutes.
Immunohistochemistry
Several rinses with 0.1 M PB (4 � 30 minutes) and 0.05M TRIS-buffered saline (TBS; 2 � 15 minutes), both at pH7.4, followed. Between all subsequent steps tissue wasrinsed with TBS (4 � 15 minutes) unless otherwise noted.As a first step, a preincubation of 45 minutes in 10%normal goat serum (Vector, Burlingame, CA) diluted inTBS was performed. As a second step, the tissue wasexposed to a mixture of primary antibodies against VIP(raised in rabbit; 1:80,000), which was generously pro-vided by Dr. T. Gorcs (Gulyas et al., 1990) and calbindin-D28k (CB; raised in mouse, 1:100,000; Sigma, Deissen-hofen, Germany), diluted with TBS. The immunoreactionlasted for 60 hours (at 6°C) under gentle agitation. As athird step, incubation of biotinylated goat anti-rabbit se-rum (1:200, in TBS; Vector) for 2 hours at room tempera-ture followed. As a fourth step, an incubation of peroxi-dase coupled to an avidin-biotin complex (ABC, 1:400, inTBS; Vector) was performed for 2 hours at room temper-ature. As a fifth step, sections were then rinsed (2 � 15minutes each) in TBS and 0.05 M TRIS-buffer (TB, pH8.2). Subsequently, the buffer was exchanged with achromogen solution consisting of 0.015% 3,3�-diaminobenzidine (DAB; Sigma) and 0.4% ammoniumnickel sulfate (Fluka, Buchs, Switzerland). After a pre-incubation of 10 minutes the reaction was started byadding hydrogen peroxide to a final concentration of0.006%. The reaction was stopped by rinsing with TBwhen visual inspection confirmed that no further in-crease in the intensity of blue staining of neuronalprocesses occurred. As a sixth step, the ABC-blockingkit was used according the manufacturer’s instructions(Vector), with the components dissolved in TBS, pH 8.2.As a seventh step, biotinylated horse antimouse serum(1:200, in TBS; Vector) was added to the sections for 2hours at room temperature. As an eighth step, anotherincubation of ABC (1:400, in TBS; Vector) was performed for2 hours at room temperature. As a final step, sections werethen rinsed (2 � 15 minutes each) in TBS and TB, pH 7.6.The visualization of the bound peroxidase was accomplishedby a preincubation in 0.5% DAB for 10 minutes, to which0.01% hydrogen peroxide was then added. The reactionlasted until a satisfactory labeling of the respective anti-gen was achieved (�3–15 minutes) and resulted in abrown staining of the target cells.
For the immunofluorescent approach, tissue was notfreeze-thawed but treated with 0.4% Triton X-100
180 J.F. STAIGER ET AL.

(Sigma). Sections were incubated in a mixture of the sameantisera against VIP (1:20,000) and CB (1:40,000) asabove, for 60 hours at 6° C. Incubation of biotinylated goatantirabbit serum (1:200, in TBS; Vector) for 2 hours atroom temperature followed. Then a mixture ofstreptavidin-coupled Alexa 488 (1:200; Molecular Probes,Leiden, Netherlands) and goat antimouse tagged with Cy3(1:400; Jackson, Dianova, Hamburg, Germany) in TBS for16 hours at 6° C was incubated. Slices were extensivelywashed with TBS between the steps and finally with TB.After termination of the incubations, the slices weremounted on glass slides and coverslipped with Vectashield(Vector).
Immunocytochemical controls, in the present study,only included a replacement of the primary antiserum byappropriately diluted normal serum, since the specificityof the antisera was extensively tested and approved by themanufacturers. This procedure always resulted in the ab-sence of any staining.
Further tissue processing and evaluation
After permanent staining, the sections were dehydratedin an ascending series of ethanol and embedded in Dur-cupan ACM (Fluka) for correlated light and electron mi-croscopy. Finally, selected neurons were reembedded,trimmed, serially cut ultrathin (Leica UCT, Vienna, Aus-tria), mounted on Formvar-coated single-slot copper grids,and examined with an electron microscope (AB-912; LEO,Oberkochen, Germany) to determine the synaptic natureof light microscopically classified contacts. Reconstruc-tions of calbindin-immunoreactive neurons and mappingof the number of VIP-immunoreactive contacts were madewith Neurolucida (Microbrightfield Europe, Magdeburg,Germany) attached to a Nikon Eclipse 800 (Nikon, Dus-seldorf, Germany) using a 100� oil-immersion objective(NA � 1.4). In the present study, “contact” or “close spatialapposition” are used as terms for light microscopic obser-vations, whereas “synaptic contact” or “synapse” are as-signed for electron microscopic observations.
The fluorescently labeled sections were examined with aconfocal microscope (Leica TCS 4D; Bensheim, Germany)equipped with a 40� oil-immersion objective (NA � 1.0)and appropriate filter sets to separate the two fluoro-chromes. A maximum intensity projection was calculatedfor each image stack after scanning and stored togetherwith the original data. For evaluation, individual focalplanes of the two-channel scans were analyzed with re-spect to visible contacts between the VIP-immunoreactiveaxons and the immunohistochemically labeled CB targetcells.
All light microscopic photographs were acquired asz-stacks with a digital camera (CoolSNAP; Visitron,Puchheim, Germany), attached to an Axioplan 2 micro-scope (Zeiss, Gottingen, Germany). Such stacks were pro-cessed with MetaView 4.5 (Universal Imaging, West Ches-ter, PA) using the minimum-projection in order to achievean “extended-focus view.” All electron microscopic photo-graphs were captured with a CCD-camera as well (Gatan,Munchen, Germany). The resulting files were importedinto PhotoShop 6.0 (Adobe, San Jose, CA) and optimizedfor contrast, brightness, and sharpness. No other manip-ulations were performed. The micrographs were arrangedand lettered with Illustrator 9.0 (Adobe).
Quantification and statistics
A single section of 50 �m thickness through the postero-medial barrel subfield, representing the large mystacialvibrissae, was selected from each of six animals thatshowed in the peroxidase stainings the best color contrastbetween VIP (blue-black) and CB (brown). Since the cal-bindin immunoreactivity faintly reveals barrel-like struc-tures, it was possible to center a columnar field of interest(of 500 �m width) onto a barrel. In this barrel-relatedcolumn, all calbindin-immunoreactive neurons which pos-sessed at least two primary dendrites and 200 �m of totaldendritic length were reconstructed with Neurolucida.The column was further divided into layer compartments(supragranular, granular, and infragranular) according tothe relative thickness of the cortical layers as calculatedfrom standard rat stereotaxical atlases (Paxinos andWatson, 1998; Swanson, 1998). For a third of these neu-rons, randomly selected to avoid bias in sampling, a cer-tain prominent subtype or a certain staining intensity, thecontacts formed by VIP boutons on soma and dendrites ofCB neurons were mapped with Neurolucida. The recon-structions were quantitatively evaluated with Neuroex-plorer (Microbrightfield Europe) and data were trans-ferred to a spreadsheet.
The obtained values were tested for statistically signif-icant differences in the defined layer compartments byperforming multivariate ANOVA (randomized block de-sign, blocking variable “animal”; Bonferroni corrected) us-ing the software package SysStat 10 (SPSS, Chicago, IL).All data are given as mean � SD.
RESULTS
Immunohistochemistry of VIP neurons
VIP-immunoreactive (VIP�) neurons with their so-matic, dendritic and axonal patterns in rat somatosensorycortex have been described in detail recently (Bayraktar etal., 2000). Briefly, most interneurons showed a bipolarmorphology (Figs. 1, 2A). Staining of somato-dendritic andaxonal profiles was most pronounced in layers II/III andIV. In the present study, however, axonal density onlyslightly decreased in layers V and VI (Fig. 2). This wasachieved by the addition of acrolein to the fixation proce-dure, as compared to previous studies using only parafor-maldehyde. However, this also resulted in a more unpre-dictable preservation of the ultrastructure.
An observation in the present specimens, confirmingour previous studies, was that VIP� boutons reliably con-tacted VIP� somata and dendrites in all cortical layers inhigh numbers. This is exemplified for bipolar VIP� neu-rons located in layers II/III (Fig. 1). The minimum-intensity projections of stacks display several of the de-tectable close spatial appositions formed by VIP�
varicosities with different subcellular compartments of atarget neuron (Fig. 1B). That these light microscopicallydetectable contacts form synapses at the ultrastructurallevel is demonstrated for another bipolar VIP� neuron,shown by a conventional photomontage (Fig. 1C). Thecorrelated electron micrograph (Fig. 1D) presents thesame perisomatic distribution of the VIP� boutons, whichwere nearly always found to form synapses when comingin contact with a target structure (Fig. 1E).
181TARGETS OF CORTICAL VIP SYNAPSES

Fig. 1. Interconnectivity of VIP� interneurons. A: Minimum-in-tensity projection of an �25 �m-thick stack through supragranularlayers II/III. Arrows point to two VIP� neurons, the upper left beinga multipolar and the lower right being a typical bipolar-bituftedinterneuron. Note some pericellular basket-like arrangements ofVIP� boutons (asterisks). B: Higher magnification of the stackedbipolar-bitufted interneuron allows the observation of some of thecontacts (arrowheads) formed by VIP� boutons on the soma anddendrites of this interneuron, as detected in the single optical slice
micrographs. C: Another bipolar VIP� interneuron in layers II/IIIwhich is stained more faintly. This allows appreciating the VIP�
boutons (arrowheads) very clearly, in this light and the correlatedelectron micrograph (D). D: The same cell as in (C) presents a typicalultrastructural feature of GABAergic interneurons, i.e., the deep in-dentation of the nuclear membrane. E: One example of the identifiedsymmetrical synapses for this neuron formed by bouton b1 which islabeled by an arrow from C–E. Scale bars � 25 �m in A,B; 10 �m inC; 5 �m in D; 0.5 �m in E.

Immunohistochemistry of calbindin neurons
The general distribution and morphology of CB-immunoreactive (CB�) somata and dendrites was essen-tially in agreement with previous studies (Celio, 1990; van
Brederode et al., 1991). Briefly, all cortical layers (exceptlayer I) contained CB� cell bodies (Figs. 2, 3). Most ofthem were found in layers V and VI (infragranular layers),followed by layers II and III (supragranular layers), and
Fig. 2. Confocal imaging of VIP� (green) and CB� (red) containingneurons in rat barrel cortex. A: Low-power micrograph shows thepattern and typical morphology of VIP� and CB� cells in the supra-granular layers. Note the smaller ovoid somata of the mainly bipolarVIP� neurons and the larger often multipolar CB� interneurons (ar-rows) which are easily distinguished from the numerous lightly la-beled CB� pyramidal cells. B: High-power view of a bitufted CB�
neuron which is contacted by numerous VIP� boutons, some of which
are marked by arrowheads. C: Multipolar CB� neuron with numerousprimary dendrites is embedded in a dense plexus of VIP� axons andboutons. D: A horizontally oriented (multipolar) CB� neuron in layerVI is frequently contacted by VIP� axons in a climbing-fiber-likemanner (arrowheads). All images are maximum intensity projectionsof z-stacks. Roman numerals indicate cortical layers. Scale bars � 100�m in A; 25 �m in B–D.
183TARGETS OF CORTICAL VIP SYNAPSES
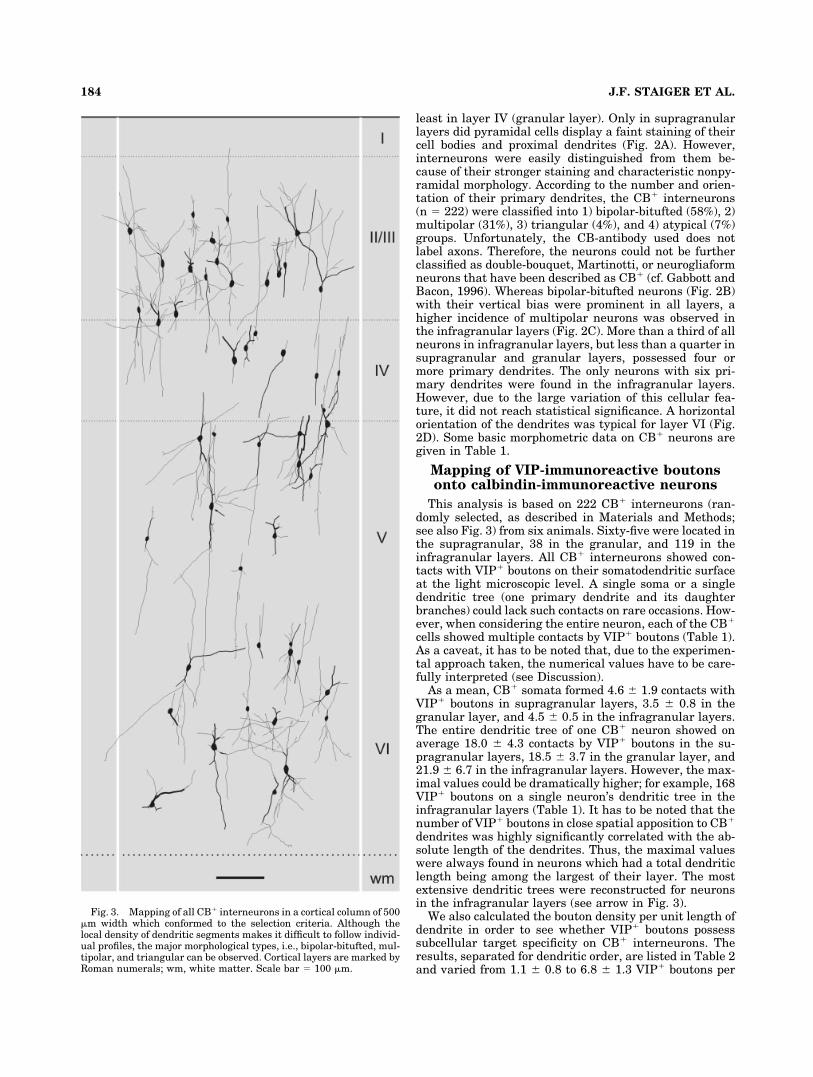
least in layer IV (granular layer). Only in supragranularlayers did pyramidal cells display a faint staining of theircell bodies and proximal dendrites (Fig. 2A). However,interneurons were easily distinguished from them be-cause of their stronger staining and characteristic nonpy-ramidal morphology. According to the number and orien-tation of their primary dendrites, the CB� interneurons(n � 222) were classified into 1) bipolar-bitufted (58%), 2)multipolar (31%), 3) triangular (4%), and 4) atypical (7%)groups. Unfortunately, the CB-antibody used does notlabel axons. Therefore, the neurons could not be furtherclassified as double-bouquet, Martinotti, or neurogliaformneurons that have been described as CB� (cf. Gabbott andBacon, 1996). Whereas bipolar-bitufted neurons (Fig. 2B)with their vertical bias were prominent in all layers, ahigher incidence of multipolar neurons was observed inthe infragranular layers (Fig. 2C). More than a third of allneurons in infragranular layers, but less than a quarter insupragranular and granular layers, possessed four ormore primary dendrites. The only neurons with six pri-mary dendrites were found in the infragranular layers.However, due to the large variation of this cellular fea-ture, it did not reach statistical significance. A horizontalorientation of the dendrites was typical for layer VI (Fig.2D). Some basic morphometric data on CB� neurons aregiven in Table 1.
Mapping of VIP-immunoreactive boutonsonto calbindin-immunoreactive neurons
This analysis is based on 222 CB� interneurons (ran-domly selected, as described in Materials and Methods;see also Fig. 3) from six animals. Sixty-five were located inthe supragranular, 38 in the granular, and 119 in theinfragranular layers. All CB� interneurons showed con-tacts with VIP� boutons on their somatodendritic surfaceat the light microscopic level. A single soma or a singledendritic tree (one primary dendrite and its daughterbranches) could lack such contacts on rare occasions. How-ever, when considering the entire neuron, each of the CB�
cells showed multiple contacts by VIP� boutons (Table 1).As a caveat, it has to be noted that, due to the experimen-tal approach taken, the numerical values have to be care-fully interpreted (see Discussion).
As a mean, CB� somata formed 4.6 � 1.9 contacts withVIP� boutons in supragranular layers, 3.5 � 0.8 in thegranular layer, and 4.5 � 0.5 in the infragranular layers.The entire dendritic tree of one CB� neuron showed onaverage 18.0 � 4.3 contacts by VIP� boutons in the su-pragranular layers, 18.5 � 3.7 in the granular layer, and21.9 � 6.7 in the infragranular layers. However, the max-imal values could be dramatically higher; for example, 168VIP� boutons on a single neuron’s dendritic tree in theinfragranular layers (Table 1). It has to be noted that thenumber of VIP� boutons in close spatial apposition to CB�
dendrites was highly significantly correlated with the ab-solute length of the dendrites. Thus, the maximal valueswere always found in neurons which had a total dendriticlength being among the largest of their layer. The mostextensive dendritic trees were reconstructed for neuronsin the infragranular layers (see arrow in Fig. 3).
We also calculated the bouton density per unit length ofdendrite in order to see whether VIP� boutons possesssubcellular target specificity on CB� interneurons. Theresults, separated for dendritic order, are listed in Table 2and varied from 1.1 � 0.8 to 6.8 � 1.3 VIP� boutons per
Fig. 3. Mapping of all CB� interneurons in a cortical column of 500�m width which conformed to the selection criteria. Although thelocal density of dendritic segments makes it difficult to follow individ-ual profiles, the major morphological types, i.e., bipolar-bitufted, mul-tipolar, and triangular can be observed. Cortical layers are marked byRoman numerals; wm, white matter. Scale bar � 100 �m.
184 J.F. STAIGER ET AL.

100 �m length of CB� dendrites. The respective neuronalpopulations within supragranular, granular, and infra-granular layers possessed rather similar numbers of VIP�
boutons per 100 �m dendritic length for each order testedseparately. This suggests that the innervation densitydoes not obviously differ in relation to the location of CB�
neurons in certain layers. Nevertheless, a tendency fordendritic segments of the second and third order to receivemore boutons than those of first or fourth order can beseen. Furthermore, a drop in innervation density wasobserved for dendritic segments of higher orders (fifth andsixth).
Correlated light and electron microscopicanalysis
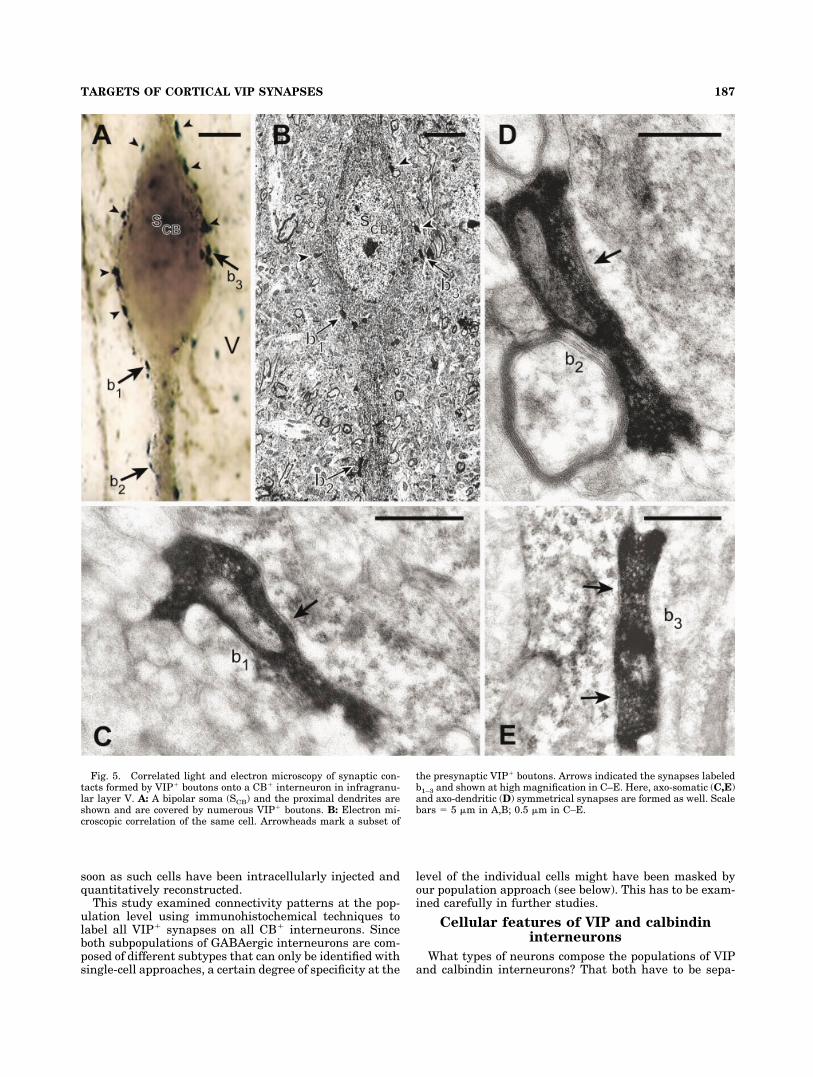
Five CB� neurons contacted by numerous VIP� boutonswere selected for correlated light and electron microscopicanalysis. No matter whether these neurons were locatedin supragranular (Fig. 4) or infragranular layers (Fig. 5),virtually all somatic (45/47; �96%) and most dendriticcontacts (54/71; �76%) contained in the reembedded tis-sue were verified as symmetric synapses at the ultrastruc-tural level. It was observed that somatic synapses seemedto be underestimated (due to the masking effect of thedense somatic staining) and dendritic synapses overesti-mated (probably due to a spatial proximity-coincidenceeffect) with light microscopic analysis alone.
For a bitufted CB� neuron in layers II/III (Fig. 4), axo-somatic (Fig. 4C,D) and axo-dendritic (Fig. 4E) synapseswere found. For a bipolar CB� neuron in layer V (Fig. 5),these types of synapses were also observed (Figs. 5C–E).Moreover, due to the superior ultrastructural preserva-tion in this specimen, the symmetric nature (i.e., an ap-proximately equal thickness of pre- and postsynapticmembranes) of these VIP� synapses is more clearly ap-preciable.
DISCUSSION
We have shown, at the population level, a consistentand strong innervation of calbindin-immunoreactive in-terneurons by VIP-immunoreactive symmetric synapses
in rat primary somatosensory cortex by using correlatedlight and electron microscopy. The occurrence of multiplesynapses on the soma as well as on the dendritic tree ofeach calbindin interneuron was not dependent on the lam-inar location of these cells. This suggests that this type ofinhibitory–inhibitory connectivity performs a similarfunction in all layers of a cortical column.
Technical considerations
We used an unbiased selection of the cells included inthis study. This resulted in a rather heterogeneous popu-lation of studied neurons ranging from those that justfulfilled the minimum criteria defined to those that mightcome closer to a complete representation of the CB� neu-rons. Since we further used a single section approach,probably all neurons examined were only partially recon-structed (cf. Gulyas et al., 1999). Thus, all absolute valueshave to be interpreted with caution. However, the evalu-ation of the connectivity pattern between VIP� boutonsand CB� interneurons should not be compromised seri-ously by these methodological limitations. Moreover, ourapproach to determine bouton density on dendriticbranches of a specified order allows the calculation of theabsolute number of VIP� synapses on CB� neurons as
Fig. 4. Correlated light and electron microscopy of synaptic con-tacts formed by VIP� boutons onto CB� interneurons in supragranu-lar layers. A: The large brownish soma (SCB) and the proximal den-drites of a bipolar-bitufted neuron are contacted by numerous darklystained VIP� boutons (some marked by arrowheads). The VIP� bou-tons indicated by arrows and labeled b1–3 are visible also in B–E.B: Low-power electron micrograph shows the ultrastructural appear-ance of the identical cell as in A. Note the deep indentation of thenucleus, which is a typical feature of inhibitory interneurons. Notethat A is a minimum intensity projection of �20 �m depth, whereas Bis an ultrathin section of 60 nm. Thus, only a subset of light micro-scopic boutons is directly correlated with presynaptic electron micro-scopic profiles. High-power images of symmetric synapses formed byVIP� presynaptic boutons with (C,D) somatic and (E) dendriticpostsynaptic membrane of the CB� target cell. Scale bars � 10 �m inA; 5 �m in B; 0.5 �m in C–E.
TABLE 1. Quantification of Soma Area, Number of Primary Dendrites and Dendritic Length for CB� Neurons and of VIP� Boutons on Soma as Well asDendritic Tree, in Supragranular (SG), Granular (GR), and Infragranular (IG) Layers
SG GR IG
Mean � SD Min Max Mean � SD Min Max Mean � SD Min Max
Soma area (�m2) 124.1 � 24.1 70.5 203.1 112.3 � 8.5 70.9 205.9 140.6 � 18.8 70.1 251.6Primary dendrites 2.9 � 0.4 2 5 2.9 � 0.3 2 5 3.2 � 0.3 2 6Dendritic length (�m) 442.1 � 93.8 203 1334 578.7 � 73.8 211 1333 552.0 � 111.6 201 1582VIP boutons per soma 4.6 � 1.9 0 26 3.5 � 0.8 1 11 4.5 � 0.5 0 22VIP boutons per dendritic tree 18.0 � 4.3 2 64 18.5 � 3.7 3 85 21.9 � 6.7 3 168
No statistically significant differences between layers were found. Means and SD describe our sample of six animals (n � 6). Minimum (Min) and Maximum (Max) values arecalculated from the total sample of all individual cells (n � 222; SG � 65, GR � 38, IG � 119).
TABLE 2. Density of VIP� Boutons in Apparent Contact With CB� Dendrites, Specified for Dendritic Order (Number per 100 �m of Dendritic Length)
Layers
Dendritic order
1st 2nd 3rd 4th 5th 6th
SG 5.3 � 0.9 (n � 189) 6.1 � 1.9 (n � 237) 6.4 � 2.7 (n � 160) 4.4 � 1.5 (n � 80) 2.0 � 2.1 (n � 24) 2.0 � 1.8 (n � 12)GR 5.0 � 0.3 (n � 105) 6.0 � 1.9 (n � 148) 4.9 � 1.7 (n � 112) 4.3 � 1.7 (n � 70) 3.2 � 2.2 (n � 28) 1.1 � 0.8 (n � 18)IG 5.7 � 1.5 (n � 383) 6.8 � 1.3 (n � 498) 6.6 � 2.9 (n � 372) 4.5 � 1.6 (n � 184) 2.8 � 1.9 (n � 80) 3.0 � 3.5 (n � 36)
Total number of segments in sample is given in parentheses.
185TARGETS OF CORTICAL VIP SYNAPSES

Figure 4
186 J.F. STAIGER ET AL.
soon as such cells have been intracellularly injected andquantitatively reconstructed.
This study examined connectivity patterns at the pop-ulation level using immunohistochemical techniques tolabel all VIP� synapses on all CB� interneurons. Sinceboth subpopulations of GABAergic interneurons are com-posed of different subtypes that can only be identified withsingle-cell approaches, a certain degree of specificity at the
level of the individual cells might have been masked byour population approach (see below). This has to be exam-ined carefully in further studies.
Cellular features of VIP and calbindininterneurons
What types of neurons compose the populations of VIPand calbindin interneurons? That both have to be sepa-
Fig. 5. Correlated light and electron microscopy of synaptic con-tacts formed by VIP� boutons onto a CB� interneuron in infragranu-lar layer V. A: A bipolar soma (SCB) and the proximal dendrites areshown and are covered by numerous VIP� boutons. B: Electron mi-croscopic correlation of the same cell. Arrowheads mark a subset of
the presynaptic VIP� boutons. Arrows indicated the synapses labeledb1–3 and shown at high magnification in C–E. Here, axo-somatic (C,E)and axo-dendritic (D) symmetrical synapses are formed as well. Scalebars � 5 �m in A,B; 0.5 �m in C–E.
187TARGETS OF CORTICAL VIP SYNAPSES

rate, nonoverlapping populations is suggested by the factthat no colocalizations of VIP and calbindin have beendescribed so far by immunocytochemical methods (Rogers,1992; Kubota et al., 1994), and rarely by single-cell RT-PCR (Cauli et al., 1997). The morphological diversity ofVIP neurons has so far precluded a complete description ofall cell types. However, the most consistent types foundwere bipolar cells with a preferential translaminar orga-nization of the axonal tree and small multipolar (basket)cells with a preferential local distribution of axonal collat-erals (Kawaguchi and Kubota, 1996; Bayraktar et al.,2000). Such a distinction also exists in the hippocampus(Acsady et al., 1996), thus suggesting that this is a basicfeature of diversity of these cells. Since bipolar cells targetmore distal dendrites and basket cells have perisomati-cally localized synapses, a convergence of projections ofboth cell types onto calbindin neurons might best explainthe ubiquitous occurrence of multiple somatic as well asproximal and distal dendritic VIP synapses on CB cells.Certainly, it will be an important next step to analyzewhat types and numbers of presynaptic VIP neurons con-verge with how many synapses on a single specified cal-bindin neuron.
Calbindin neurons are also heterogeneous with respectto their morphology and their physiology (van Brederodeet al., 1991; Kawaguchi and Kubota, 1993; Gabbott andBacon, 1996; Cauli et al., 1997). They show double bou-quet, Martinotti, and neurogliaform cells as the most con-sistent subtypes. However, the lack of axonal staining inour preparations precluded an identification of thesetypes. Therefore, it is not possible to relate the innervationpattern by VIP boutons specifically to these varieties ofcalbindin interneurons.
Inhibitory cortical circuits involving VIPneurons
Are VIP-immunoreactive neurons specialized to controlother inhibitory interneurons? In the cerebral cortex theanswer certainly is less clear than in the archicortex. There,because of its simpler lamination it was established thatcalretinin-containing interneurons—which colocalize VIP toa very large extent—extensively innervate two other typesof GABAergic cells: 1) VIP-immunoreactive basket cells and2) calbindin-immunoreactive, putative dendrite-targetingcells (Gulyas et al., 1999). The latter connectivity might bedirectly comparable to the one described in the presentstudy, because VIP and calretinin are also frequently colo-calized in neocortex (Rogers, 1992; Kubota et al., 1994). Inmonkey prefrontal cortex the following type of connectionhas been described (Gabbott and Bacon, 1996): calretininneurons heavily contact calbindin cells more distally andparvalbumin neurons more proximally. Our previous re-sults in rat barrel cortex (Staiger et al., 1997) show thatparvalbumin neurons abundantly innervate VIP cells. Inthe present investigation, as a side result, we have furtherdemonstrated a frequent interconnectivity among VIPneurons themselves.
There is still much to be learned about the fine-grainedcell type-specific connectivity of cortical GABAergic inter-neurons. An interesting recent study adds a further levelof complexity by showing the possibility of unique regula-tory mechanisms for presynaptic transmitter release inthis type of connection (Dalezios et al., 2002). Specifically,those authors found that mGluR7a-enriched VIP-immunoreactive terminals show numerous synaptic con-
tacts with mGluR1�/somatostatin-immunopositive targetneurons in rat somatosensory cortex. Since somatostatinfrequently colocalizes with calbindin in regular-spikingnonpyramidal cells (Cauli et al., 1997), this may be thesame type of connectivity as described in the presentstudy.
It is difficult to functionally define this sort of multipleinhibitory connections in simple terms of linear inhibition-disinhibition mechanisms. It seems much more likely thatthe net effect of such multiple inhibitory interactionswould be a precise timing of pyramidal cell ensembles,causing them to oscillate at different frequencies (Traub etal., 1998; Deans et al., 2001). Since all these interneurontypes also directly target pyramidal cells (Hajos et al.,1988; DeFelipe et al., 1989; Kisvarday et al., 1993; Gab-bott and Bacon, 1997), this could impose an oscillatorybehavior with differential temporal and spatial signaturesonto selected networks of excitatory cells, enabling themto perform their many functions in cortical sensory infor-mation processing (Jones and Barth, 1997; Engel et al.,2001; Ahrens et al., 2002).
ACKNOWLEDGMENTS
We thank Mr. U. Opfermann-Emmerich for invaluabletechnical assistance and for sharing PhotoShop expertise.We also thank Dr. Dirk Schubert for critically reading thearticle.
LITERATURE CITED
Acsady L, Gorcs TJ, Freund TF. 1996. Different populations of vasoactiveintestinal polypeptide-immunoreactive interneurons are specialized tocontrol pyramidal cells or interneurons in the hippocampus. Neuro-science 73:317–334.
Ahrens KF, Levine H, Suhl H, Kleinfeld D. 2002. Spectral mixing ofrhythmic neuronal signals in sensory cortex. Proc Natl Acad Sci U S A99:15176–15181.
Bayraktar T, Staiger JF, Acsady L, Cozzari C, Freund TF, Zilles K. 1997.Co-localization of vasoactive intestinal polypeptide, gamma-aminobutyric acid and choline acetyltransferase in neocortical inter-neurons of the adult rat. Brain Res 757:209–217.
Bayraktar T, Welker E, Freund TF, Zilles K, Staiger JF. 2000. Neuronsimmunoreactive for vasoactive intestinal polypeptide in the rat pri-mary somatosensory cortex: morphology and spatial relationship tobarrel-related columns. J Comp Neurol 420:291–304.
Cauli B, Audinat E, Lambolez B, Angulo MC, Ropert N, Tsuzuki K, HestrinS, Rossier J. 1997. Molecular and physiological diversity of corticalnonpyramidal cells. J Neurosci 17:3894–3906.
Cauli B, Porter JT, Tsuzuki K, Lambolez B, Rossier J, Quenet B, AudinatE. 2000. Classification of fusiform neocortical interneurons based onunsupervised clustering. Proc Natl Acad Sci U S A 97:6144–6149.
Celio MR. 1990. Calbindin D-28k and parvalbumin the rat nervous system.Neuroscience 35:375–475.
Dalezios Y, Lujan R, Shigemoto R, Roberts JDB, Somogyi P. 2002. Enrich-ment of mGluR7a in the presynaptic active zones of GABAergic andnon-GABAergic terminals on interneurons in the rat somatosensorycortex. Cereb Cortex 12:961–974.
Deans MR, Gibson JR, Sellitto C, Connors BW, Paul DL. 2001. Synchro-nous activity of inhibitory networks in neocortex requires electricalsynapses containing Connexin36. Neuron 31:477–485.
DeFelipe J. 1993. Neocortical neuronal diversity: chemical heterogeneityrevealed by colocalization studies of classic neurotransmitters, neu-ropeptides, calcium-binding proteins, and cell surface molecules. CerebCortex 3:271–289.
DeFelipe J, Hendry SHC, Jones EG. 1989. Synapses of double bouquet cellsin monkey cerebral cortex visualized by calbindin immunoreactivity.Brain Res 503:49–54.
Engel AK, Fries P, Singer W. 2001. Dynamic predictions: oscillations andsynchrony in top-down processing. Nat Rev Neurosci 2:704–716.
188 J.F. STAIGER ET AL.

Ferezou I, Cauli B, Hill EL, Rossier J, Hamel E, Lambolez B. 2002. 5-HT3receptors mediate serotonergic fast synaptic excitation of neocorticalvasoactive intestinal peptide/cholecystokinin interneurons. J Neurosci22:7389–7397.
Freund TF, Buzsaki G. 1996. Interneurons of the hippocampus. Hippocam-pus 6:347–470.
Gabbott PLA, Bacon SJ. 1996. Local circuit neurons in the medial prefron-tal cortex (areas 24a, b,c, 25 and 32) in the monkey. 1. Cell morphologyand morphometrics. J Comp Neurol 364:567–608.
Gabbott PLA, Bacon SJ. 1997. Vasoactive intestinal polypeptide contain-ing neurones in monkey medial prefrontal cortex mPFC — colocalisa-tion with calretinin. Brain Res 744:179–184.
Gonchar Y, Burkhalter A. 1999. Connectivity of GABAergic calretinin-immunoreactive neurons in rat primary visual cortex. Cereb Cortex9:683–696.
Gulyas AI, Gorcs TJ, Freund TF. 1990. Innervation of different peptide-containing neurons in the hippocampus by GABAergic septal afferents.Neuroscience 37:31–44.
Gulyas AI, Hajos N, Freund TF. 1996. Interneurons containing calretininare specialized to control other interneurons in the rat hippocampus.J Neurosci 16:3397–3411.
Gulyas AI, Megias M, Emri Z, Freund TF. 1999. Total number and ratio ofexcitatory and inhibitory synapses converging onto single interneuronsof different types in the CA1 area of the rat hippocampus. J Neurosci19:10082–10097.
Gupta A, Wang Y, Markram H. 2000. Organizing principles for a diversityof GABAergic interneurons and synapses in the neocortex. Science287:273–278.
Hajos F, Zilles K, Schleicher A, Kalman M. 1988. Types and spatial dis-tribution of vasoactive intestinal polypeptide (VIP)-containing syn-apses in the rat visual cortex. Anat Embryol 178:207–217.
Jones MS, Barth DS. 1997. Sensory evoked high frequency (gamma band)oscillating potentials in somatosensory cortex of the unanesthetizedrat. Brain Res 768:167–176.
Jones MS, Barth DS. 1999. Spatiotemporal organization of fast ( 200 Hz)electrical oscillations in rat vibrissa/barrel cortex. J Neurophysiol 82:1599–1609.
Kawaguchi Y. 1995. Physiological subgroups of nonpyramidal cells withspecific morphological characteristics in layer II/III of rat frontal cor-tex. J Neurosci 15:2638–2655.
Kawaguchi Y. 1997. Selective cholinergic modulation of cortical GABAergiccell subtypes. J Neurophysiol 78:1743–1747.
Kawaguchi Y, Kubota Y. 1993. Correlation of physiological subgroupings ofnonpyramidal cells with parvalbumin- and calbindin D28k-immunoreactive neurons in layer V of rat frontal cortex. J Neuro-physiol 70:387–396.
Kawaguchi Y, Kubota Y. 1996. Physiological and morphological identifica-tion of somatostatin- or vasoactive intestinal polypeptide-containingcells among GABAergic cell subtypes in rat frontal cortex. J Neurosci16:2701–2715.
Kawaguchi Y, Kubota Y. 1997. GABAergic cell subtypes and their synapticconnections in rat frontal cortex. Cereb Cortex 7:476–486.
Kisvarday ZF, Beaulieu C, Eysel UT. 1993. Network of GABAergic largebasket cells in cat visual cortex (area 18) — implication for lateraldisinhibition. J Comp Neurol 327:398–415.
Kubota Y, Hattori R, Yui Y. 1994. Three distinct subpopulations ofGABAergic neurons in rat frontal agranular cortex. Brain Res 649:159–173.
Meskenaite V. 1997. Calretinin immunoreactive local circuit neurons inarea 17 of the cynomolgus monkey, Macaca fascicularis. J Comp Neu-rol 379:113–132.
Morrison JH, Magistretti PJ, Benoit R, Bloom FE. 1984. The distributionand morphological characteristics of the intracortical VIP-positive cell:an immunohistochemical analysis. Brain Res 292:269–282.
Paxinos G, Watson C. 1998. The rat brain in stereotaxic coordinates. SanDiego: Academic Press.
Peters A. 1990. The axons terminals of vasoactive intestinal polypeptide(VIP)-containing bipolar cells in rat visual cortex. J Neurocytol 19:672–685.
Porter JT, Cauli B, Staiger JF, Lambolez B, Rossier J, Audinat E. 1998.Properties of bipolar VIPergic interneurons and their excitation bypyramidal neurons in the rat neocortex. Eur J Neurosci 10:3617–3628.
Porter JT, Cauli B, Tsuzuki K, Lambolez B, Rossier J, Audinat E. 1999.Selective excitation of subtypes of neocortical interneurons by nicotinicreceptors. J Neurosci 19:5228–5235.
Rogers JH. 1992. Immunohistochemical markers in rat cortex: co-localization of calretinin and calbindin-D28k with neuropeptides andGABA. Brain Res 587:147–157.
Rozov A, Jerecic J, Sakmann B, Burnashev N. 2001. AMPA receptorchannels with long-lasting desensitization in bipolar interneurons con-tribute to synaptic depression in a novel feedback circuit in layer 2/3 ofrat neocortex. J Neurosci 21:8062–8071.
Staiger JF, Zilles K, Freund TF. 1996. Innervation of VIP-immunoreactiveneurons by the ventroposteromedial thalamic nucleus in the barrelcortex of the rat. J Comp Neurol 367:194–204.
Staiger JF, Freund TF, Zilles K. 1997. Interneurons immunoreactive forvasoactive intestinal polypeptide (VIP) are extensively innervated byparvalbumin-containing boutons in rat primary somatosensory cortex.Eur J Neurosci 9:2259–2268.
Staiger JF, Schubert D, Zuschratter W, Kotter R, Luhmann HJ, Zilles K.2002. Innervation of interneurons immunoreactive for VIP by intrinsi-cally bursting pyramidal cells and fast-spiking interneurons in infra-granular layers of juvenile rat neocortex. Eur J Neurosci 16:11–20.
Swanson LW. 1998. Brain maps: structure of the rat brain. Amsterdam:Elsevier Science.
Thomson AM, Bannister AP. 2003. Interlaminar connections in the neo-cortex. Cereb Cortex 13:5–14.
Thomson AM, West DC, Wang Y, Bannister AP. 2002. Synaptic connec-tions and small circuits involving excitatory and inhibitory neurons inlayers 2-5 of adult rat and cat neocortex: triple intracellular recordingsand biocytin labelling in vitro. Cereb Cortex 12:936–953.
Traub RD, Spruston N, Soltesz I, Konnerth A, Whittington MA, JefferysJGR. 1998. Gamma-frequency oscillations: a neuronal population phe-nomenon, regulated by synaptic and intrinsic cellular processes, andinducing synaptic plasticity. Prog Neurobiol 55:563–575.
van Brederode JFM, Helliesen MK, Hendrickson AE. 1991. Distribution ofthe calcium-binding proteins parvalbumin and calbindin-D28k in thesensorimotor cortex of the rat. Neuroscience 44:157–171.
189TARGETS OF CORTICAL VIP SYNAPSES




















