
Cadmium contaminated soil affects retinogenesis in lizard embryos

1 23
Archives of EnvironmentalContamination and Toxicology ISSN 0090-4341Volume 67Number 4 Arch Environ Contam Toxicol (2014)67:639-650DOI 10.1007/s00244-014-0056-0
Cadmium Neurotoxicity to a FreshwaterPlanarian
Jui-Pin Wu, Hui-Ling Lee & Mei-Hui Li

1 23
Your article is protected by copyright and all
rights are held exclusively by Springer Science
+Business Media New York. This e-offprint is
for personal use only and shall not be self-
archived in electronic repositories. If you wish
to self-archive your article, please use the
accepted manuscript version for posting on
your own website. You may further deposit
the accepted manuscript version in any
repository, provided it is only made publicly
available 12 months after official publication
or later and provided acknowledgement is
given to the original source of publication
and a link is inserted to the published article
on Springer's website. The link must be
accompanied by the following text: "The final
publication is available at link.springer.com”.

Cadmium Neurotoxicity to a Freshwater Planarian
Jui-Pin Wu • Hui-Ling Lee • Mei-Hui Li
Received: 27 January 2014 / Accepted: 26 May 2014 / Published online: 5 July 2014
� Springer Science+Business Media New York 2014
Abstract Although freshwater planarians are evolution-
arily primitive, they are some of the simplest bilateral ani-
mals possessing integrated neural networks similar to those
in vertebrates. We attempted to develop planarian Dugesia
japonica as a model for investigating the neurotoxicity of
environmental pollutants such as cadmium (Cd). This study
was therefore designed to study the effects of Cd on the
locomotor activity, neurobehavior, and neurological
enzymes of D. japonica. After planarians were exposed to Cd
at high concentrations, altered neurobehavior was observed
that exhibited concentration-dependent patterns. Morpho-
logical alterations in Cd-treated planarians included irregu-
lar shape, body elongation, screw-like hyperkinesia, and
bridge-like position. To study the direct effects of Cd on
neurological enzymes, tissue homogenates of planarians
were incubated in vitro with Cd before their activity was
measured. Results showed that acetylcholinesterase
(AChE), adenosine triphosphatase (ATPase), and mono-
amine oxidase A (MAO-A) activities were inhibited in a
concentration-dependent manner. MAO-B activity was sig-
nificantly induced by Cd at low concentrations and inhibited
at high concentrations. Changes in the in vivo activity of
AChE and ATPase were also found after planarians were
treated with Cd at a sublethal concentration (5.56 lM).
These observations indicate that neurotransmission systems
in planarians are disturbed after Cd exposure.
Freshwater free-living flatworms, or planarians, of the
phylum Platyhelminthes, are evolutionarily primitive
invertebrates that have one obvious neuronal cluster loca-
ted at the head end and two symmetric longitudinal cords
with ladder-like interconnections (Nishimura et al. 2008).
Although some investigators prefer to designate the ante-
rior neuronal cluster in planarians as a ‘‘cephalic gan-
glion,’’ it actually fulfills all of the criteria for the definition
of a brain in animals (Sarnat & Netsky 2002). It had been
even considered that the evolution of the vertebrate central
nervous system (CNS) may have begun with planarians
(Sarnat & Netsky 2002), although this is controversial in
light of the fact that planarians do not seem to be in the
direct evolutionary lineage to vertebrates (Northcutt 2012).
Biochemical substances identical to those used by mam-
mals as neurotransmitters and second messengers, as well
as their receptors, also occur in freshwater planarians
(Buttarelli et al. 2008). In addition, they have a relatively
sophisticated repertoire of behaviors (e.g., learning and
memory) and the capacity for tissue regeneration (Raffa &
Rawls 2008). Specific behavioral patterns can be clearly
observed after treating planarians with drugs that act on
neural transmission (Buttarelli et al. 2000; Carolei et al.
1975; Palladini et al. 1996; Venturini et al. 1989). These
biological characteristics and advantages make the fresh-
water planarian a simple but powerful and practical model
for research into drug action and abuse (Raffa & Rawls
2008) as well as neurological diseases (Kitamura et al.
1998).
When exposed to neurotoxic chemicals, the normal
neural activities of humans and other animals will be
altered and/or their structures damaged. The most well-
known and widely distributed neurotoxic chemicals
include, but are not limited to, certain heavy metals such as
cadmium (Cd) (Wright & Baccarelli 2007). Cd
J.-P. Wu � M.-H. Li (&)
Environmental Toxicology Laboratory, Department of
Geography, National Taiwan University, 1, Section 4, Roosevelt
Road, Taipei 106, Taiwan
e-mail: [email protected]
H.-L. Lee
Department of Chemistry, Fu Jen Catholic University, 510,
Zhongzheng Road, New Taipei City 24205, Taiwan
123
Arch Environ Contam Toxicol (2014) 67:639–650
DOI 10.1007/s00244-014-0056-0
Author's personal copy

neurotoxicity has been reported in other experimentally
treated mammals (Antonio et al. 1998; Gabbiani et al.
1967; Lafuente et al. 2001, 2005; Pari & Murugavel 2007),
fish (Beauvais et al. 2001; Pretto et al. 2010; Richetti et al.
2011; Silva & Pathiratne 2008), and taxonomically higher
invertebrates (Cunha et al. 2007). However, related infor-
mation for acoelomates and pseudocoelomates, which
constitute a large proportion of the world’s invertebrate
biota, is still limited (Du & Wang 2009). Previously, we
found Cd distributed in the head of treated D. japonica
Ichikawa and Kawakatsu 1964 at a significantly greater
concentration than the tail, and suggested that the head
might be the target of Cd toxicity after exposure (Wu et al.
2011). Although head dissolution (Calevro et al. 1998) and
some neurobehavioral changes (Grebe & Schaeffer 1991a;
Plusquin et al. 2012) after Cd treatments have been
reported in some planarian species other than D. japonica,
further investigation of the neurotoxic effects of Cd to them
at the biochemical level has never been reported.
The present study was designed to investigate the neu-
rotoxic effects of Cd to D. japonica on the neurobehavioral
and biochemical levels. First, locomotor activity and spe-
cific behavioral patterns were tested in D. japonica treated
with Cd. Second, the effects of Cd on neurological enzyme
activities, including acetylcholinesterase (AChE), adeno-
sine triphosphatase (ATPase), and monoamine oxidase
(MAO), in D. japonica were determined after in vitro and
in vivo exposures. Finally, specific behavioral patterns
observed in Cd-treated planarians were linked to alterations
of their neurological enzyme activities. The results of this
study not only provide a mechanistic understanding of Cd
neurotoxicity, but also provide helpful information on
establishing D. japonica as a research model for neuro-
toxicity of other environmental contaminants.
Materials and Methods
Chemicals
Cadmium sulfate (3CdSO4�8H2O) was purchased from
Mallinckrodt (Hazelwood, Missouri, USA). Amplex Red
reagent was obtained from Life Technologies Corporation
(Carlsbad, California, USA). Other chemicals used in this
study were purchased from Sigma-Aldrich (St Louis,
Missouri, USA).
Animals
The stock of D. japonica used in this study was obtained
from Taikong Corporation (Taipei, Taiwan) and acclimated
in our laboratory for [2 weeks. In laboratory, planarians
were maintained in dechlorinated tap water at room tem-
perature (23–28 �C) and fed fresh chicken liver once per
week followed by immediate renewal of culture waters
(Wu et al. 2012). Size-selected intact planarians (approxi-
mately 1 cm in length) were used for this study. Planarians
were starved for 3–5 days to create a uniform metabolic
state before being used in the experiments.
Locomotor Activity Test
To study the effects of Cd on locomotor activity of treated
D. japonica, planarians were divided into five groups, each
containing one individual, and exposed to 3 mL formulated
artificial freshwater (ISO water) (Wu et al. 2012) with
various concentrations of Cd at room temperature. The Cd
concentrations used in this study were 0 (control), 0.5, 5,
50, and 500 lM. After exposure for 1 h, locomotor
Table 1 Behavioral responses of planarians categorized by response
group that was adapted according to Grebe and Schaeffer (1991b)
Response
group
Behavioral
response
Description
Morphology Head/nose twist Tip of nose curls up or down
Body elongated Body length is extremely
elongated
Dumbbell-
shape/irregular
shape
Body is contracted irregularly or
becomes dumbbell-shaped
Screw-like Planarian twists around anterior–
posterior axis, and corkscrew-
shaped movement appears
Bridge-like Head is brought closer to tail
ventrally leading to the body
being shaped like a bridge
C-like Head is brought closer to tail
laterally leading to a body
shapes like the letter ‘‘C’’
Walnut-shape Planarian is usually stationary,
moving only slowly with its
body contracted and with an
obvious decrease in its length,
leading to the body being
walnut-shaped
Ornamentation Edges of planarian become
crenelated
Neurological Convulsions Violent twitching or snake-like
movement
Morbidity Labored
movement
Planarian is stationary or moving
slowly
Depression Planarian barely moves when
poked
Unconsciousness Planarian does not move even
when poked
Death Planarian does not move when
poked, and rapid tissue necrosis
is apparent.
640 Arch Environ Contam Toxicol (2014) 67:639–650
123
Author's personal copy

activities of control and treated planarians were quantified
as previously described (Raffa et al. 2001; Pagan et al.
2006). Briefly, planarians were placed individually into a
plastic petri dish (10-cm diameter) containing 10 mL ISO
water and located over graph paper with grid lines spaced
0.5 cm apart. Locomotor activity was quantified as the
number of grid lines each planarian crossed over a
10-minute observation period. Each planarian was used
only once. The experiment was repeated four times.
Neurobehavioral Test
Neurobehavioral testing of planarians after Cd treatment
was performed in 6-well cell culture plates at room tem-
perature with 1 planarian/well. Well culture waters were
replaced by 5 mL ISO water. No obvious abnormalities in
morphology or behavior were apparent in planarians after
culture waters were replaced. The test was started by
replacing the plain ISO water with 5 mL ISO water con-
taining Cd at concentrations of 0, 1.25, 2.5, 5, 10, and
20 mM. The concentrations of Cd used in neurobehavioral
test were based on the results of a preliminary test. Neu-
robehavioral responses in treated planarians was continu-
ously observed and recorded for 60 min. The experiment
was independently repeated ten times. Distinct neurobe-
havioral responses of planarians after Cd-treatment were
categorized based on Grebe and Schaeffer (1991b) with
modifications (Table 1).
Effects of Cd on In Vitro Neurological Enzyme
Activity
Planarians were homogenized with homogenizing buffer
(25 mM Tris-HCl [pH 7.0]) for AChE or 50 mM Tris-HCl
containing 250 mM sucrose and 1 mM ethylene diamine
tetraacetic acid (EDTA) (pH 7.4) for ATPase analysis.
After centrifuging at 12,0009g for 20 min at 4 �C, the
pellets were discarded and the supernatants used for
in vitro assays. For MAO, planarians were homogenized
with 250 mM sucrose (pH 7.6, adjusted by 1 M K2HPO4)
and centrifuged at 1,5009g for 20 min at 4 �C followed by
mitochondria extraction (Tipton & Dawson 1968). Mito-
chondria were resuspended in 50 mM potassium phosphate
buffer (pH 7.4) and kept on ice until in vitro assays were
performed. Supernatants from planarian tissue homoge-
nates for AChE and ATPase assays and mitochondria
extracted for MAO assay were incubated with Cd at con-
centrations of 0, 0.625, 1.25, 2.5, and 5 mM for 30 min at
25 �C before determinations of enzyme activities. These
Cd concentrations were chosen for the in vitro study based
on the neurobehavioral study, but they were lower. For
each enzyme, the experiment was repeated at least three
times.
Effects of Cd on In Vivo Neurological Enzyme Activity
To study the effects of Cd on the in vivo AChE, ATPase,
and MAO activity of treated D. japonica, 20 planarians
were divided into 2 groups, each containing 10 animals,
and exposed to 10 mL ISO water with Cd as the treatment
group or without Cd as controls. The Cd concentration used
in this study was 5.56 lM (=0.63 mg/L), a concentration
equivalent to half of the 48-hour median lethal concen-
tration (LC50) of Cd to D. japonica. This caused significant
alterations in several toxicological end points without the
death of animals during the experimental period (at least
7 days) (Wu et al. 2012). Each sample containing two
animals was killed after exposure for 1, 2, 4, and 7 days to
determine the enzyme activity of AChE, ATPase, and
MAO. This experiment was repeated six times.
Measurement of Neurological Enzyme Activity
To determine AChE activity, planarians were homogenized
with 25 mM Tris-HCl (pH 7.0) and centrifuged at
12,0009g for 20 min at 4 �C. The supernatant was used for
AChE activity measurement according to the method pre-
viously described for determining planarian ChE activity
(Wu et al. 2011) after applying a selective irreversible
butyrylcholinesterase inhibitor, tetraisopropyl pyrophos-
phoramide (iso-OMPA) before the assay was performed.
Briefly, enzymes were incubated with 10 mM iso-OMPA
for 30 min at room temperature before adding dithiobis-
nitrobenzoic acid and acetylthiocholine iodide followed by
recording the absorbance at 414 nm.
To determine ATPase activity, planarians were
homogenized with 50 mM Tris-HCl containing 250 mM
sucrose and 1 mM EDTA (pH 7.4). After centrifugation at
12,0009g for 20 min at 4 �C, the ATPase activity of the
supernatant was measured as previously described (Li
2012).
MAO activity in mitochondria extracted from planarians
was measured according to the method described by Zhou
and Panchuk-Voloshina (1997) with some modifications.
Briefly, 20 lL of resuspended mitochondria were added to
a 96-well microplate and incubated with 160 lL of work-
ing solution containing 200 lM Amplex Red reagent and
1 U/ml horseradish peroxidase in 50 mM potassium
phosphate buffer (pH 7.4) for three minutes at 25 �C.
After 20 lL of amine substrate was added into the samples,
fluorescence was measured with a fluorescence microplate
reader with its filter set for excitation and emission at 560
and 580 nm, respectively, for 30 min and corrected by
Arch Environ Contam Toxicol (2014) 67:639–650 641
123
Author's personal copy

subtracting the values derived from the blank without the
amine substrate. Amine substrates used for the determi-
nation of MAO-A and -B activity were 0.1 mM serotonin
and 20 mM phenethylamine, respectively. Serial concen-
trations of resorufin standards were prepared to establish
the standard curve. MAO activity was expressed as the
amount of resorufin (fmol) produced per mg of mito-
chondria protein per minute. Protein concentrations in the
homogenate and mitochondria extracts were measured
using Bradford’s method (Bradford 1976).
Determination of Actual Cd Concentrations in Test
Solutions
To confirm the actual Cd concentrations in test solutions,
1 mL of each solution was sampled, combined with 9 mL
0.5 % HNO3 in distilled water, and stored at 4 �C until
analysis. Cd levels were measured using an atomic
absorption spectrophotometer (932 AA, GBC Scientific
Equipment, Braeside, Australia). Before measuring the test
solution, standard curves with high correlation coefficients
(R2 = 0.9884 to 0.9975) were established after the certified
Cd standard solution (Merck KGaA, Darmstadt, Germany)
was serially diluted and determined. The detection limit of
the AAS for Cd was 0.01 mg/L. If necessary, test solutions
were diluted with 0.5 % HNO3 in double-distilled water.
Statistical Analysis
Statistical evaluations were performed with the Minitab
Statistical Program (ver. 15) (Minitab; State College, PA,
USA). Data normality was tested and confirmed with the
Anderson-Darling test. Data that met the assumptions of
normality were analyzed with one-way analysis of variance
to determine the effects of Cd treatment on locomotor
activity, neural behavior patterns, and neurological enzyme
activities. Post hoc Duncan’s multiple-range test was used
to evaluate mean differences between control and treatment
groups. Unless noted, all comparisons were considered
significantly different at p \ 0.05. Results are presented as
the mean ± SD.
Results
Cd Concentrations Measured in Test Solutions
The actual concentrations of Cd in the test solutions were
analytically determined, and the average percent nominal
concentration calculated to be 121.38 ± 13.89 %. This
indicates that measured Cd concentrations were close to
those of nominal values.
Locomotor Activity in Cd-treated D. japonica
Locomotor activity of untreated D. japonica is shown in
Fig. 1a. Untreated planarians showed a characteristic and
stable locomotor velocity during the course of the
10-minute observation period (28.58 ± 1.07 crosses/min).
In planarians exposed to Cd, inhibition of locomotor
velocity was observed at C5 lM Cd (Fig. 1b).
Neural Behavioral Patterns in Cd-treated D. japonica
We modified the categories, and only three groups of
behavioral responses were recorded: morphological
responses, neurological responses, and morbidity
(Table 1). The morphology group contained eight different
responses: head/nose twist, body elongated, dumbbell-
shape/irregular shape, screw-like, bridge-like, C-like, wal-
nut-shape, and ornamentation. The neurological group
contained only convulsions response. Morbidity group
contained four different responses: labored movement,
depression, unconsciousness, and death.
Fig. 1 a Locomotor activity of untreated D. japonica expressed as
the mean ± SD of the cumulative number of grid lines crossed per
minute. b Locomotor velocity of planarians after exposure to various
concentrations of Cd for 1 h. After exposure, locomotor velocity was
quantified over a 10-minute observation period and expressed as
mean ± SD of the number grid lines crossed per minute. *p \ 0.05
and #p \ 0.1 compared with controls
642 Arch Environ Contam Toxicol (2014) 67:639–650
123
Author's personal copy
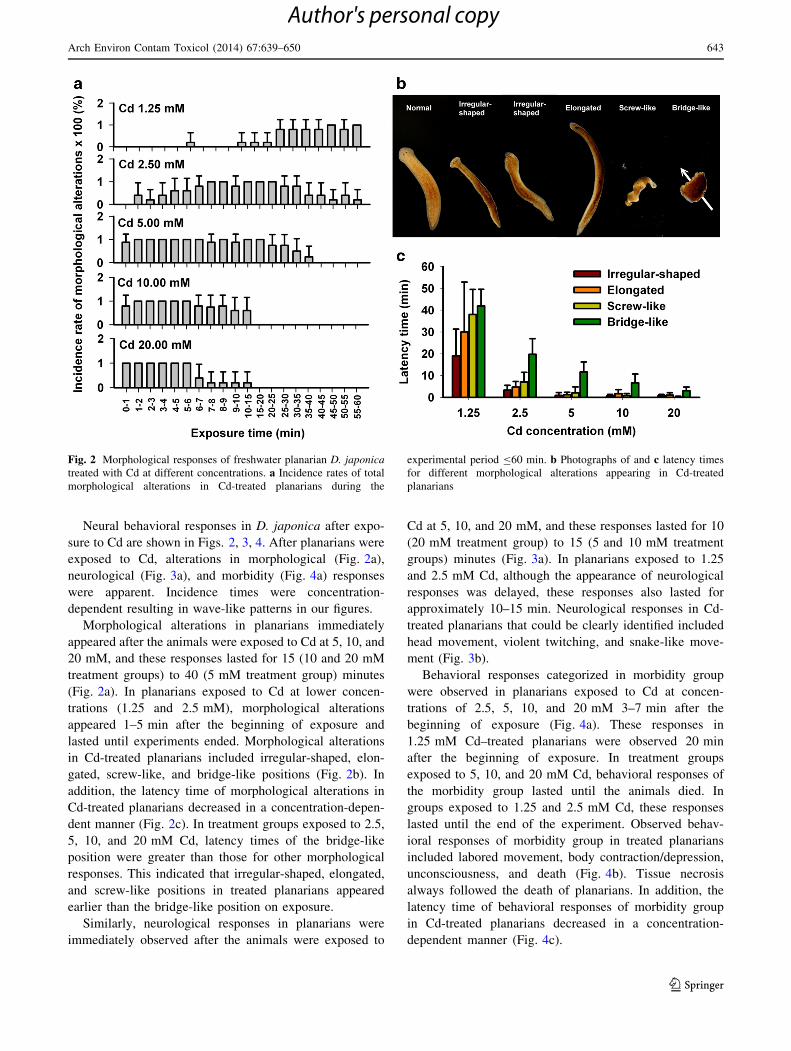
Neural behavioral responses in D. japonica after expo-
sure to Cd are shown in Figs. 2, 3, 4. After planarians were
exposed to Cd, alterations in morphological (Fig. 2a),
neurological (Fig. 3a), and morbidity (Fig. 4a) responses
were apparent. Incidence times were concentration-
dependent resulting in wave-like patterns in our figures.
Morphological alterations in planarians immediately
appeared after the animals were exposed to Cd at 5, 10, and
20 mM, and these responses lasted for 15 (10 and 20 mM
treatment groups) to 40 (5 mM treatment group) minutes
(Fig. 2a). In planarians exposed to Cd at lower concen-
trations (1.25 and 2.5 mM), morphological alterations
appeared 1–5 min after the beginning of exposure and
lasted until experiments ended. Morphological alterations
in Cd-treated planarians included irregular-shaped, elon-
gated, screw-like, and bridge-like positions (Fig. 2b). In
addition, the latency time of morphological alterations in
Cd-treated planarians decreased in a concentration-depen-
dent manner (Fig. 2c). In treatment groups exposed to 2.5,
5, 10, and 20 mM Cd, latency times of the bridge-like
position were greater than those for other morphological
responses. This indicated that irregular-shaped, elongated,
and screw-like positions in treated planarians appeared
earlier than the bridge-like position on exposure.
Similarly, neurological responses in planarians were
immediately observed after the animals were exposed to
Cd at 5, 10, and 20 mM, and these responses lasted for 10
(20 mM treatment group) to 15 (5 and 10 mM treatment
groups) minutes (Fig. 3a). In planarians exposed to 1.25
and 2.5 mM Cd, although the appearance of neurological
responses was delayed, these responses also lasted for
approximately 10–15 min. Neurological responses in Cd-
treated planarians that could be clearly identified included
head movement, violent twitching, and snake-like move-
ment (Fig. 3b).
Behavioral responses categorized in morbidity group
were observed in planarians exposed to Cd at concen-
trations of 2.5, 5, 10, and 20 mM 3–7 min after the
beginning of exposure (Fig. 4a). These responses in
1.25 mM Cd–treated planarians were observed 20 min
after the beginning of exposure. In treatment groups
exposed to 5, 10, and 20 mM Cd, behavioral responses of
the morbidity group lasted until the animals died. In
groups exposed to 1.25 and 2.5 mM Cd, these responses
lasted until the end of the experiment. Observed behav-
ioral responses of morbidity group in treated planarians
included labored movement, body contraction/depression,
unconsciousness, and death (Fig. 4b). Tissue necrosis
always followed the death of planarians. In addition, the
latency time of behavioral responses of morbidity group
in Cd-treated planarians decreased in a concentration-
dependent manner (Fig. 4c).
Fig. 2 Morphological responses of freshwater planarian D. japonica
treated with Cd at different concentrations. a Incidence rates of total
morphological alterations in Cd-treated planarians during the
experimental period B60 min. b Photographs of and c latency times
for different morphological alterations appearing in Cd-treated
planarians
Arch Environ Contam Toxicol (2014) 67:639–650 643
123
Author's personal copy

Effects of Cd Treatments on the In Vitro Activity
of Neurological Enzymes
Planarian neurological enzymes were prepared and
exposed in vitro to Cd at different concentrations followed
by measuring their activity (Fig. 5). After incubation with
Cd, AChE and ATPase activities decreased significantly
and in a concentration-dependent manner (p \ 0.1 or 0.05)
(Fig. 5a, b). In addition, a significant decrease in MAO-A
activity was apparent after the enzyme was incubated with
Cd at concentrations of 2.5 and 5 mM Cd (p \ 0.1 and
0.05, respectively) (Fig. 5c). It was interesting that pla-
narian MAO-B activity was significantly increased after
the enzyme was incubated with lower concentrations of Cd
(0.63 and 1.25 mM) (p \ 0.05) but was inhibited at a
greater concentration (5 mM) (p \ 0.1) (Fig. 5d).
In Vivo Activity of Neurological Enzymes in D.
japonica After Exposure to Sublethal Concentrations
of Cd
To investigate the effects of Cd on the in vivo activity of
neurological enzymes in planarians, animals were exposed
to sublethal concentrations of Cd for 1, 2, 4, and 7 days
followed by measurements of neurological enzyme activ-
ity. Significant decreases of AChE activity in treated pla-
narians were observed on days 4 and 7 after exposure
(Fig. 6a). Although Cd exposure did not affect planarian
ATPase activity when animals were exposed for 1 and
4 days, a significant increase in activity was observed when
the exposure was prolonged to 7 days (Fig. 6b). An obvi-
ous decrease in MAO-A activity was apparent in Cd-trea-
ted planarians after exposure for 4 days, but no significant
difference was found between the enzyme activity of
controls and treatment groups 7 days after exposure
(Fig. 6c). Cd exposure did not influence the activity of
planarian MAO-B during the entire experimental period
(Fig. 6d).
Discussion
Effects of Cd on Neural Behavior of D. japonica
We found that the differences between some of the
responses in D. japonica after exposure to Cd were not as
distinct as described by Grebe and Schaeffer (1991b). For
example, the ‘‘inch-worming’’ (hyperkinesia) response
categorized in their locomotive group is actually difficult to
clearly distinguish from the ‘‘shape change’’ response of
the morphological group. Under these circumstances,
experimental results could be influenced by observer bias.
To make the response categories more practical and the
results more consistent, we modified the categories by
discarding some that we considered equivocal and recorded
only those responses that were distinctive and clearly
identifiable.
Using the behavioral responses in our modified catego-
ries as end points, the effects of Cd on the neural behavior
of D. japonica were exhibited in a dose-dependent manner.
This was expected because, in the early 1990s, some Du-
gesia dorotocephala behavior was found to be altered by
metal exposure. This included prolonged righting (or thi-
gmotactic) times, decreased prey capture ability, decreased
motility, locomotive disorders, and changes in body shape
(Best & Morita 1991; Kapu & Shaeffer 1991). In this
study, inhibited locomotor activity in D. japonica after Cd
Fig. 3 Neurological responses of freshwater planarian D. japonica
treated with Cd at different concentrations. a Incidence rates of total
neurological alterations in Cd-treated planarians during the experi-
mental period B60 min. b Photographs of morphological alterations
appearing in Cd-treated planarians
644 Arch Environ Contam Toxicol (2014) 67:639–650
123
Author's personal copy

treatment was also observed. Altered neural behaviors and
morphological changes in Cd-treated D. dorotocephala
were reported by Grebe and Schaeffer (1991a). However,
they did not further discuss the relationships between
altered neural behaviors and potential disorder/dysfunction
of the neural system.
Observations from previous pharmacological studies
revealed that planarians show specific behavioral patterns
when normal neural transmission is altered by chemicals.
Stimulations of dopamine receptors D1 (D1R) and D2
(D2R) induce hyperkinesia in planarians leading to the
appearances of screw-like and C-like morphologies,
respectively (Palladini et al. 1996; Venturini et al. 1989). In
contrast, stimulation of ACh transmission in planarians is
followed by hypokinesia of animals in bridge-like poses
(Carolei et al. 1975). More recently, the interaction of ACh
and dopamine transmission in planarians was investigated,
and it was found that the effects of its dopaminergic
transmission appear to be quite the opposite of those of
ACh (Buttarelli et al. 2000, 2008). Neural behaviors—
including the C-like position, screw-like hyperkinesia,
writhing, and head-swinging—were observed in planarians
treated with serotonin-receptor agonists and serotonin
(Farrell et al. 2008). The results of these pharmacological
studies indicate that ACh, dopamine, and serotonin are
crucial for mediating the regulation of neuromuscular
functions in planarians.
In this study, we found that both screw-like hyperkinesia
and bridge-like position occurred in Cd-treated D. japonica
suggesting that Cd possibly exerts its effect on cholinergic,
dopaminergic, and/or serotonergic transmission in planar-
ians. Irregular-shaped and elongated body morphologies
were also observed in Cd-treated D. japonica. Although the
correlations between these two neural behavioral responses
in planarians and neural transmission were not reported and
discussed previously, they were also apparently the results
of abnormal contractions in muscular systems. The C-like
position was not observed in Cd-treated D. japonica, sug-
gesting that Cd possibly did not stimulate its D2R. Previ-
ously, the loss of the D2R was found in rat striatal
homogenate pretreated with Cd (Scheuhammer & Cherian
1985).
The occurrences of neurological responses in Cd-treated
planarians were within the time periods when their mor-
phological responses occurred. This confirmed that treated
planarians were still alive, conscious, and responsive and
that their neurotransmission and neuromuscular system
were severely disturbed during these periods. When
exposure time was prolonged, however, Cd-treated pla-
narians died, especially those exposed to Cd at
Fig. 4 Morbidity responses of freshwater planarian D. japonica
treated with Cd at different concentrations. a Incidence rates of total
morbidity responses in Cd-treated planarians during the experimental
period B60 min. b Photographs of and c latency times for different
morbidity responses appearing in Cd-treated planarians. Arrow
indicates the pharynx extended outside the body of the planarian
Arch Environ Contam Toxicol (2014) 67:639–650 645
123
Author's personal copy

concentrations [5 mM. Before death, morbidity responses
occurred after the neurological and morphological
responses, occurring in the order of labored movement,
depression, and unconsciousness.
Effects of Cd on Neurological Enzymes of D. japonica
In mammals, although the entrance of Cd into the brain is
limited by the blood–brain barrier (BBB), it disturbs CNS
functioning by altering brain tissue levels of trace elements
and provoking disturbances to crucial neural enzymes
involved in neurotransmitter release and neurotransmission
(Carageorgiou & Katramadou 2012). Planarians lack the
equivalent structure of a BBB (Raffa et al. 2007), so it is
possible that Cd enters the brain directly and causes some
influences on its function and structure. Nevertheless, we
showed that AChE, ATPase, and MAO-A and -B activities
of D. japonica were altered after these enzymes were
directly exposed to Cd in vitro. In addition, altered in vivo
activities of these neurological enzymes were observed in
planarians treated with Cd.
AChE is a crucial enzyme for the regulation of cholin-
ergic transmission by catalyzing the hydrolysis of ACh to
acetate and choline to terminate the transmission. Thus,
any fluctuation of AChE activity could cause a change in
cholinergic transmission (Carageorgiou & Katramadou
2012). Our in vitro and in vivo studies both showed that
planarian AChE activity was inhibited by Cd exposure,
which was presumably followed by uninterrupted cholin-
ergic signal transmission.
Na?, K?-ATPase is a membrane-bound enzyme
involved in several neural functions of animals such as
neural excitability (Sastry & Phillis 1977) and penetration
of neurotransmitters across membranes (Hernandez 1989;
Swann 1984). We found that the activity of planarian
ATPase was inhibited in a concentration-dependent man-
ner after in vitro incubation with Cd. Inhibited Na?, K?-
ATPase activities after exposure to high concentrations of
Cd (0.01–1 mM) were also reported in in vitro systems of
other animals (Carageorgiou et al. 2004; Chandra et al.
1984; Kinne-Saffran et al. 1993). The inhibitory mecha-
nisms of Cd on Na?, K?-ATPase show species specificity
(Kinne-Saffran et al. 1993), so further investigation is still
needed to understand the molecular mechanism in planar-
ians. Regardless of the mechanism, Na?, K?-ATPase
activity inhibited by high concentrations of Cd (0.1 mM) is
Fig. 5 Activity of a AChE, b ATPase, c MAO-A, and d MAO-B in
tissue homogenates of planarian D. japonica after in vitro pre
incubations with different concentrations of Cd for 30 min. The
activity of the enzymes in treatment groups is presented as
percentages of corresponding controls. *p \ 0.05 and #p \ 0.1
compared with controls
646 Arch Environ Contam Toxicol (2014) 67:639–650
123
Author's personal copy

accompanied by decreases in dopamine and noradrenalin
uptakes in rat brain synaptosomes (Chandra et al. 1984).
This suggests that dopamine reuptake was possibly
decreased in D. japonica after treatment with Cd at high
concentrations. In addition, increased Na?, K?-ATPase
activity was observed in brains of rats treated with 1–5 mg
Cd/kg and in in vitro brain homogenate incubated with Cd
at low concentrations (Carageorgiou et al. 2004). These
findings suggest that compared with the effect caused by
Cd at high levels, low levels of Cd increase Na?, K?-
ATPase activity. A similar phenomenon was also observed
in planarians exposed to Cd at sublethal concentrations for
7 days with their in vivo ATPase activity significantly
increasing (p \ 0.05).
MAO is a key enzyme catalyzing the metabolism of
biogenic amines, including catecholamine neurotransmit-
ters. Two isoforms of MAO, including MAO-A and -B,
were characterized according to substrate specificity and
inhibitor sensitivity (Youdim et al. 1969). The differential
effects of Cd on the activity of these two MAO isoforms
were found in rat (Leung et al. 1992). Similarly, we found
that in vitro activities of planarian MAO isoforms were
affected by Cd with different patterns. Presumably, fluc-
tuations of MAO-A and -B activity caused by Cd might
alter the levels of serotonin and dopamine, which in turn
influence serotonergic and dopaminergic neurotransmis-
sions. In contrast, the in vivo MAO activity of D.
japonica was insensitive to Cd at sublethal concentra-
tions. Because the direct effect of Cd to planarian MAO
was shown through in vitro study, the insusceptibility of
in vivo MAO activity to Cd might be a result of pro-
tection by intrinsic physiological mechanisms. Indeed, we
previously showed that metallothionein, a well-known
inducible protein possessing the ability to sequester
divalent metals in animals, was largely induced in Cd-
treated D. japonica under an identical experimental
design (Wu et al. 2012). This suggested that Cd was
sequestered and that the amount of free Cd ions within
the planarian body might be insufficient to have direct
effects on MAO enzymes.
Fig. 6 Activity of a AChE, b ATPase, c MAO-A, and d MAO-B in the planarian D. japonica treated with Cd at a concentration of 5.56 lM for
1, 2, 4, and 7 days. *p \ 0.05 compared with controls
Arch Environ Contam Toxicol (2014) 67:639–650 647
123
Author's personal copy

Correlations Between the Altered Activity
of Neurological Enzymes and Neural Behavior
Responses in Cd-treated D. japonica
Apparently all of the enzymes investigated in this study
were susceptible to direct Cd exposure and potentially
contributed to altered neurotransmission in Cd-treated D.
japonica The susceptibility to Cd of these planarian neu-
rological enzymes was on the order of ATPase = MAO-B
[ AChE [ MAO-A according to Cd concentrations that
caused significant differences in their activity. It is also
interesting that in D. japonica treated with Cd at concen-
tration [2.5 mM, screw-like hyperkinesia in planarians
appeared earlier than the bridge-like position, which was
soon followed by depression and other morbidity respon-
ses. With these observations, we propose a hypothetic
model containing three stages to link the changes on altered
activities of neurological enzymes in treated D. japonica to
those of its neurobehavioral responses (Fig. 7).
According to the model, although initial body levels of
Cd are relatively low after exposure, the first stage ATPase
activity is inhibited due to its greater susceptibility to Cd
accompanying decreased dopamine reuptake (Chandra
et al. 1984). It might be followed immediately by the
accumulation of dopamine in the synaptic cleft and con-
tinuous stimulation on dopaminergic neurotransmission,
thus leading to the induction of hyperkinesia (screw-like
morphology) in this stage. At the second stage, however, as
exposure continues, the level of Cd in the treated planarian
body is gradually increased and causes an increase in ACh
release (Meyer & Cooper 1981), inhibition of AChE
activity, and enhancement of MAO-B activity. These
alterations might be followed by stimulation of cholinergic
neurotransmission and decrease of dopaminergic
neurotransmission. That would account for the bridge-like
position in treated planarians appearing at this stage.
Finally, at the third stage, accumulated body levels of Cd
are so high that disorders in other essential physiological
functions also occur in parallel. At this stage, the responses
of planarians related to disturbed neurotransmissions (e.g.,
morphological and neurological responses) are almost
completely replaced by morbidity responses, tissue necro-
sis, and death. Therefore, although decreases in MAO-A
and -B activity observed in planarian homogenate after
in vitro exposure to Cd at high concentrations (2.5 and
5 mM) are shown, no corresponding neurobehavioral
effects are presented at this stage.
Conclusion
In this study, Cd at high concentrations was used to explore
its direct effect on neurotransmission systems and the
neural behavior of the freshwater planarian D. japonica.
Clearly, concentration-dependent patterns in behavioral
responses of Cd-treated planarians were shown. In addi-
tion, through in vitro and in vivo studies, effects of Cd on
planarian AChE, ATPase, and MAO-A, and MAO-B
activity were revealed and shown to be comparable with
those of mammalian systems. Thus, disturbed neurotrans-
mission in Cd-treated planarians is suggested and used to
propose a hypothetic model connecting the results from
behavioral and enzyme studies. However, some key
mechanistic details about the effects of Cd on planarian
neurotransmission systems—such as the molecular mech-
anisms of alterations in neurological enzymes, the patterns
of changes in neurotransmitter levels, and the ranges of
body Cd levels that separate the three response stages that
Fig. 7 Hypothetical model connecting the changes in neurological enzyme activity and neurobehavioral responses in the Cd-treated planarian D.
japonica
648 Arch Environ Contam Toxicol (2014) 67:639–650
123
Author's personal copy

we propose—are still unknown. In addition, future study of
the long-term exposure of planarians to Cd at sublethal
and/or environmentally relevant concentrations is desired
for understanding the neurotoxic effects of this metal under
real situations.
Acknowledgments The authors are grateful for funding support
under Grants No. NSC101-2811-B-002-055 and NSC101-2321-B-
002-069 from the National Science Council, Taiwan.
References
Antonio MT, Benito MJ, Leret ML, Corpas I (1998) Gestational
administration of cadmium alters the neurotransmitter levels in
newborn rat brains. J Appl Toxicol 18:83–88
Beauvais SL, Jones SB, Parris JT, Brewer SK, Little EE (2001)
Cholinergic and behavioral neurotoxicity of carbaryl and cad-
mium to larval rainbow trout (Oncorhynchus mykiss). Ecotoxicol
Environ Saf 49:84–90
Best JB, Morita M (1991) Toxicology of planarians. Hydrobiologia
227:375–383
Bradford MM (1976) A rapid and sensitive method for the
quantitation of microgram quantities of protein utilizing the
principle of protein-dye binding. Anal Biochem 72:248–254
Buttarelli FR, Pontieri FE, Margotta V, Palladini G (2000) Acetyl-
choline/dopamine interaction in planaria. Comp Biochem Phys-
iol C 125:225–231
Buttarelli FR, Pellicano C, Pontieri FE (2008) Neuropharmacology
and behavior in planarians: translations to mammals. Comp
Biochem Physiol C 147:399–408
Calevro F, Filippi C, Deri P, Albertosi C, Batistoni R (1998) Toxic
effects of aluminum, chromium, and cadmium in intact and
regenerating freshwater planarians. Chemosphere 37:651–659
Carageorgiou H, Katramadou M (2012) Aspects of cadmium
neurotoxicity. In: Li YV, Zhang JH (eds) Metal ion in stroke.
Springer Science?Business Media, New York, pp 703–750
Carageorgiou H, Tzotzes V, Pantos C, Mourouzis C, Zarros A,
Tsakiris S (2004) In vivo and in vitro effects of cadmium on
adult rat brain total antioxidant status, acetylcholinesterase,
(Na?, K?)-ATPase and Mg2?-ATPase activities: protection by
L-cysteine. Basic Clin Pharmacol Toxicol 94:112–118
Carolei A, Margotta V, Palladini G (1975) Proposal of a new model
with dopaminergic-cholinergic interactions for neuropharmaco-
logical investigations. Neuropsychobiology 1:355–364
Chandra SV, Murthy RC, Husain T, Bansal SK (1984) Effect of
interaction of heavy metals on (Na?-K?) ATPase and the uptake
of 3H-DA and 3H-NA in rat brain synaptosomes. Acta Pharmacol
Toxicol 54:210–213
Cunha I, Mangas-Ramirez E, Guilhermino L (2007) Effects of copper
and cadmium on cholinesterase and glutathione S-transferase
activities of two marine gastropods. Comp Biochem Physiol C
145:648–657
Du M, Wang D (2009) The neurotoxic effects of heavy metal
exposure on GABAergic nervous system in nematode Caeno-
rhabditis elegans. Environ Toxicol Pharmacol 27:314–320
Farrell MS, Gilmore K, Raffa RB, Walker EA (2008) Behavioral
characterization of serotonergic activation in the flatworm
Planaria. Behav Pharmacol 19:177–182
Gabbiani G, Baid D, Deziel C (1967) Toxicity of cadmium for the
central nervous system. Exp Neurol 18:154–160
Grebe E, Schaeffer DJ (1991a) Neurobehavioral toxicity of cadmium
sulfate to the planarian Dugesia dorotocephala. Bull Environ
Contam Toxicol 46:727–730
Grebe E, Schaeffer DJ (1991b) Planarians in toxicology, stan-
dardization of a rapid neurobehavioral toxicity test using
phenol in a crossover study. Bull Environ Contam Toxicol
46:866–870
Hernandez R (1989) Na?-K?-ATPase activity possibly regulated by a
specific serotonin receptor. Brain Res 408:399–402
Kapu MM, Shaeffer DJ (1991) Planarians in toxicology. Responses of
asexual Dugesia dorotocephala to selected metals. Bull Environ
Contam Toxicol 47:302–307
Kinne-Saffran E, Hulseweh M, Pfaff C, Kinne RKH (1993) Inhibition
of Na, K-ATPase by cadmium: different mechanisms in different
species. Toxicol Appl Pharmacol 121:22–29
Kitamura Y, Kakimura J, Taniguchi T (1998) Protective effect of
Talipexole on MPTP-treated planarian, a unique Parkinsonian
worm model. Jpn J Pharmacol 78:23–29
Lafuente A, Marquez N, Pazo D, Esquifino AI (2001) Cadmium
effects on dopamine turnover and plasma levels of prolactin, GH
and ACTH. J Physiol Biochem 57:231–236
Lafuente A, Gonzalez-Carracedo A, Romero A, Cabaleiro T,
Esquifino AI (2005) Toxic effects of cadmium on the regulatory
mechanism of dopamine and serotonin on prolactin secretion in
adult male rats. Toxicol Lett 155:87–96
Leung TKC, Lim L, Lai JCK (1992) Differential effects of metal ions
on type A and type B monoamine oxidase activities in rat brain
and liver mitochondria. Metab Brain Dis 7:139–146
Li M-H (2012) Survival, mobility, and membrane-bound enzyme
activities of freshwater planarian, Dugesia japonica, exposed to
synthetic and natural surfactants. Environ Toxicol Chem
31:843–850
Meyer EM, Cooper JR (1981) Correlations between Na?, K?-ATPase
activity and acetylcholine release in rat cortical synaptosomes.
J Neurochem 36:467–475
Nishimura K, Yamamoto H, Kitamura Y, Agata K (2008) Brain and
neural networks. In: Raffa RB, Rawls SM (eds) Planaria: a
model for drug action and abuse. Landes Bioscience, Austin,
pp 4–12
Northcutt RG (2012) Evolution of centralized nervous systems: two
schools of evolutionary thought. Proc Natl Acad Sci USA
109(Suppl 1):10626–10633
Pagan OR, Rowlands AL, Urban KR (2006) Toxicity and behavioral
effects of dimethylsulfoxide in planaria. Neurosci Lett
407:274–278
Palladini G, Ruggieri S, Stocchi F, De Pandis MF, Venturini G,
Margotta V (1996) A pharmacological study of cocaine activity
in planaria. Comp Biochem Physiol C 115:41–45
Pari L, Murugavel P (2007) Diallyl tetrasulfide improves cadmium
induced alterations of acetylcholinesterase, ATPase and oxida-
tive stress in brain of rats. Toxicology 234:44–50
Plusquin M, Stevens A-S, Van Belleghem F, Degheselle O, Van
Roten A, Vroonen J et al (2012) Physiological and molecular
characterization of cadmium stress in Schmidtea mediterranea.
Int J Dev Biol 56:183–191
Pretto A, Loro VL, Morsch VM, Moraes BS, Menezes C, Clasen B
et al (2010) Acetylcholinesterase activity, lipid peroxidation, and
bioaccumulation in silver catfish (Rhamdia quelen) exposed to
cadmium. Arch Environ Contam Toxicol 58:1008–1014
Raffa RB, Rawls SM (2008) Planaria: a model for drug action and
abuse. Landes Bioscience, Austin
Raffa RB, Holland LJ, Schulingkamp RJ (2001) Quantitative
assessment of dopamine D2 antagonist activity using inverte-
brate (Planaria) locomotion as a functional endpoint. J Pharma-
col Toxicol Methods 45:223–226
Raffa RB, Cavallo F, Capasso A (2007) Flumazenil-sensitive dose-
related physical dependence in planarians produced by two
benzodiazepine and one non-benzodiazepine benzodiazepine
receptor agonists. Eur J Pharmacol 564:88–93
Arch Environ Contam Toxicol (2014) 67:639–650 649
123
Author's personal copy

Richetti SK, Rosemberg DB, Ventura-Lima J, Monserrat JM, Bogo
MR, Bonan CD (2011) Acetylcholinesterase activity and anti-
oxidant capacity of zebrafish brain is altered by heavy metal
exposure. Neurotoxicology 32:116–122
Sarnat HB, Netsky MG (2002) When does a ganglion become a
brain? Evolutionary origin of the central nervous system. Semin
Pediatr Neurol 9:240–253
Sastry BSR, Phillis JW (1977) Antagonism of biogenic amine
induced depression of cerebral cortical neurons by Na?-K?-
ATPase inhibitors. Can J Physiol Pharmacol 55:170–180
Scheuhammer AM, Cherian MG (1985) Effects of heavy metal
cations, sulfhydryl reagents and other chemical agents on striatal
D2 dopamine receptors. Biochem Pharmacol 34:3405–3413
Silva KTU, Pathiratne A (2008) In vitro and in vivo effects of
cadmium on cholinesterases in Nile tilapia fingerlings: implica-
tions for biomonitoring aquatic pollution. Ecotoxicology
17:725–731
Swann AC (1984) Na?, K?-adenosine triphosphatase regulation by
the sympathetic nervous system: effects of noradrenergic
stimulation and lesion in vivo. J Pharmacol Exp Ther
228:304–311
Tipton KF, Dawson AP (1968) The distribution of monoamine
oxidase and a-glycerophosphate dehydrogenase in pig brain.
Biochem J 108:95–99
Venturini G, Stocchi F, Margotta V, Ruggieri S, Bravi D, Bellantuono
P et al (1989) A pharmacological study of planaria’s dopami-
nergic receptors. Neuropharmacology 28:1377–1382
Wright RO, Baccarelli A (2007) Metals and neurotoxicology. J Nutr
137:2809–2813
Wu J-P, Chen H-C, Li M-H (2011) The preferential accumulation of
cadmium in the head portion of the freshwater planarian,
Dugesia japonica (Platyhelminthes: turbellaria). Metallomics
3:1368–1375
Wu J-P, Chen H-C, Li M-H (2012) Bioaccumulation and toxicody-
namics of cadmium to freshwater planarian and the protective
effect of N-acetylcysteine. Arch Environ Contam Toxicol
63:220–229
Youdim MBH, Collins GGS, Sandler M (1969) Multiple forms of rat
brain monoamine oxidase. Nature 223:626–628
Zhou M, Panchuk-Voloshina N (1997) A one-step fluorometric
method for the continuous measurement of monoamine oxidase
activity. Anal Biochem 253:169–174
650 Arch Environ Contam Toxicol (2014) 67:639–650
123
Author's personal copy
Copyright © 2022 FDOKUMEN




















