
Cadmium causes the oxidative modification of proteins in pea plants
10
Plant, Cell and Environment (2002) 25, 677–686 © 2002 Blackwell Science Ltd 677 Blackwell Science, LtdOxford, UK PCEPlant, Cell and Environment0016-8025Blackwell Science Ltd 2002 255May 2002 850 Cadmium-induced protein oxidation M. C. Romero-Puertas et al. 10.1046/j.0016-8025.2002.00850.x Original Article677686BEES SGML Correspondence: Luisa M. Sandalio. Fax: +34 958129600; e-mail: [email protected] Cadmium causes the oxidative modification of proteins in pea plants M. C. ROMERO-PUERTAS, 1 J. M. PALMA, 1 M. GÓMEZ, 2 L. A. DEL RÍO 1 & L. M. SANDALIO 1 1 Departamento de Bioquímica, Biología Celular y Molecular de Plantas, and 2 Departamento de Agroecología y Protección Vegetal, Estación Experimental del Zaidín, Consejo Superior de Investigaciones Científicas, Apartado 419, E-18080 Granada, Spain ABSTRACT In pea (Pisum sativum L.) leaves from plants grown in the presence of 50 µ µM CdCl 2 the oxidative production of car- bonyl groups in proteins, the rate of protein degradation and the proteolytic activity were investigated. In leaf extracts the content of carbonyl groups measured by derivatization with 2,4-dinitrophenylhydrazine (DNPH), was two-fold higher in plants treated with Cd than in con- trol plants. The identification of oxidized proteins was car- ried out by sodium dodecyl sulphate-polyacrylamide gel electrophoresis of proteins derivatized with DNPH and immunochemical detection with an antibody against DNPH. The intensity of the reactive bands was higher in plants exposed to Cd than in controls. By using different antibodies some of the oxidized proteins were identified as Rubisco, glutathione reductase, manganese superoxide dis- mutase, and catalase. The incubation of leaf crude extracts with increasing H 2 O 2 concentrations showed a progressive enhancement in carbonyl content and the pattern of oxi- dized proteins was similar to that found in Cd-treated plants. Oxidized proteins were more efficiently degraded, and the proteolytic activity increased 20% due to the metal treatment. In peroxisomes purified from pea leaves a rise in the carbonyl content similar to that obtained in crude extracts from Cd-treated plants was observed, but the func- tionality of the peroxisomal membrane was not apparently affected by Cd. Results obtained demonstrate the partici- pation of both oxidative stress, probably mediated by H 2 O 2 , and proteolytic degradation in the mechanism of Cd toxic- ity in leaves of pea plants, and they appear to be involved in the Cd-induced senescence previously reported in these plants. Key-words: antioxidants; cadmium; carbonyl groups; oxidative stress; peroxisomes; proteases; reactive oxygen species; Rubisco; senescence. INTRODUCTION Cadmium is a heavy metal that is toxic for humans, animals and plants, and is one of the widespread trace pollutants with a long biological half-life (Wagner 1993). This metal enters the environment mainly from industrial processes and phosphate fertilizers and is transferred to animals and humans through the food chain (Wagner 1993). The toxic action of cadmium is still not completely understood and apparently different mechanisms are involved. Cadmium produces alterations in the functionality of membranes by inducing lipid peroxidation (Fodor, Szabó-Nagy & Erdei 1995), and disturbances in chloroplast metabolism by inhib- iting chlorophyll biosynthesis and reducing the activity of enzymes involved in CO 2 fixation (Stobart et al. 1985; De Filippis & Ziegler 1993). Some studies have suggested that an oxidative stress could be involved in Cd toxicity by either inducing oxygen free radical production or by decreasing enzymatic and non-enzymatic antioxidants (Somashekaraiah, Padmaja & Prasad 1992; Stohs & Bagchi 1995; Shaw 1995). Recently, we have demonstrated that cadmium produces a concentration-dependent imbalance in the antioxidant status of pea plants (Sandalio et al. 2001). Reactive oxygen species (ROS) are known to cause the oxidative modification of proteins and the generation of reactive aldehydes as by-products of lipid peroxidation. Proteins can be affected by ROS either directly by oxida- tion of amino acid side chains, or by secondary reactions with aldehydic products of lipid peroxidation or glycosyla- tion, giving rise to the production of carbonyl-groups in the protein molecule (Reinheckel et al. 1998). The presence of these reactive groups can be determined by derivatization with 2,4-dinitrophenylhydrazine (DNPH) and has been considered as a good index of oxidative stress (Levine et al. 1994). Modified proteins can undergo a change in their hydrophobicity producing protein aggregates, or giving rise to the formation of peptide fragments (Pacifici & Davies 1990). Several authors have demonstrated that oxidatively modified proteins are selectively used as target substrates for proteases, and have proposed the use of protein degra- dation as an index of oxidative stress (Pacifici & Davies 1990). Peroxisomes are cell organelles that produce H 2 O 2 , and O 2 – radicals, and very probably nitric oxide (NO·) (del Río et al. 2002; Corpas, Barroso & del Río 2001). However, these organelles also contain a battery of antioxidative enzymes, including superoxide dismutase (SOD), catalase (CAT), the ascorbate-glutathione cycle, and several NADP-dehydrogenases (del Río et al. 1998, 2002). Differ- ent evidence obtained in recent years have shown that leaf peroxisomes have a ROS-mediated metabolic function in
-
Upload
independent -
Category
Documents
-
view
1 -
download
0
Transcript of Cadmium causes the oxidative modification of proteins in pea plants

Plant, Cell and Environment
(2002)
25,
677–686
© 2002 Blackwell Science Ltd
677
Blackwell Science, LtdOxford, UKPCEPlant, Cell and Environment0016-8025Blackwell Science Ltd 2002255May 2002850Cadmium-induced protein oxidationM. C. Romero-Puertas
et al.
10.1046/j.0016-8025.2002.00850.xOriginal Article677686BEES SGML
Correspondence: Luisa M. Sandalio. Fax: +34
958129600; e-mail: [email protected]
Cadmium causes the oxidative modification of proteins in pea plants
M. C. ROMERO-PUERTAS,
1
J. M. PALMA,
1
M. GÓMEZ,
2
L. A. DEL RÍO
1
& L. M. SANDALIO
1
1
Departamento de Bioquímica, Biología Celular y Molecular de Plantas, and
2
Departamento de Agroecología y Protección Vegetal, Estación Experimental del Zaidín, Consejo Superior de Investigaciones Científicas, Apartado 419, E-18080 Granada, Spain
ABSTRACT
In pea (
Pisum sativum
L.) leaves from plants grown in thepresence of 50
µµµµ
M
CdCl
2
the oxidative production of car-bonyl groups in proteins, the rate of protein degradationand the proteolytic activity were investigated. In leafextracts the content of carbonyl groups measured byderivatization with 2,4-dinitrophenylhydrazine (DNPH),was two-fold higher in plants treated with Cd than in con-trol plants. The identification of oxidized proteins was car-ried out by sodium dodecyl sulphate-polyacrylamide gelelectrophoresis of proteins derivatized with DNPH andimmunochemical detection with an antibody againstDNPH. The intensity of the reactive bands was higher inplants exposed to Cd than in controls. By using differentantibodies some of the oxidized proteins were identified asRubisco, glutathione reductase, manganese superoxide dis-mutase, and catalase. The incubation of leaf crude extractswith increasing H
2
O
2
concentrations showed a progressiveenhancement in carbonyl content and the pattern of oxi-dized proteins was similar to that found in Cd-treatedplants. Oxidized proteins were more efficiently degraded,and the proteolytic activity increased 20% due to the metaltreatment. In peroxisomes purified from pea leaves a rise inthe carbonyl content similar to that obtained in crudeextracts from Cd-treated plants was observed, but the func-tionality of the peroxisomal membrane was not apparentlyaffected by Cd. Results obtained demonstrate the partici-pation of both oxidative stress, probably mediated by H
2
O
2
,and proteolytic degradation in the mechanism of Cd toxic-ity in leaves of pea plants, and they appear to be involved inthe Cd-induced senescence previously reported in theseplants.
Key-words
: antioxidants; cadmium; carbonyl groups;oxidative stress; peroxisomes; proteases; reactive oxygenspecies; Rubisco; senescence.
INTRODUCTION
Cadmium is a heavy metal that is toxic for humans, animalsand plants, and is one of the widespread trace pollutantswith a long biological half-life (Wagner 1993). This metal
enters the environment mainly from industrial processesand phosphate fertilizers and is transferred to animals andhumans through the food chain (Wagner 1993). The toxicaction of cadmium is still not completely understood andapparently different mechanisms are involved. Cadmiumproduces alterations in the functionality of membranes byinducing lipid peroxidation (Fodor, Szabó-Nagy & Erdei1995), and disturbances in chloroplast metabolism by inhib-iting chlorophyll biosynthesis and reducing the activity ofenzymes involved in CO
2
fixation (Stobart
et al
. 1985; DeFilippis & Ziegler 1993). Some studies have suggested thatan oxidative stress could be involved in Cd toxicity byeither inducing oxygen free radical production or bydecreasing enzymatic and non-enzymatic antioxidants(Somashekaraiah, Padmaja & Prasad 1992; Stohs & Bagchi1995; Shaw 1995). Recently, we have demonstrated thatcadmium produces a concentration-dependent imbalancein the antioxidant status of pea plants (Sandalio
et al
. 2001).Reactive oxygen species (ROS) are known to cause the
oxidative modification of proteins and the generation ofreactive aldehydes as by-products of lipid peroxidation.Proteins can be affected by ROS either directly by oxida-tion of amino acid side chains, or by secondary reactionswith aldehydic products of lipid peroxidation or glycosyla-tion, giving rise to the production of carbonyl-groups in theprotein molecule (Reinheckel
et al
. 1998). The presence ofthese reactive groups can be determined by derivatizationwith 2,4-dinitrophenylhydrazine (DNPH) and has beenconsidered as a good index of oxidative stress (Levine
et al
.1994). Modified proteins can undergo a change in theirhydrophobicity producing protein aggregates, or giving riseto the formation of peptide fragments (Pacifici & Davies1990). Several authors have demonstrated that oxidativelymodified proteins are selectively used as target substratesfor proteases, and have proposed the use of protein degra-dation as an index of oxidative stress (Pacifici & Davies1990).
Peroxisomes are cell organelles that produce H
2
O
2
, andO
2–
radicals, and very probably nitric oxide (NO·) (del Río
et al
. 2002; Corpas, Barroso & del Río 2001). However,these organelles also contain a battery of antioxidativeenzymes, including superoxide dismutase (SOD), catalase(CAT), the ascorbate-glutathione cycle, and severalNADP-dehydrogenases (del Río
et al
. 1998, 2002). Differ-ent evidence obtained in recent years have shown that leafperoxisomes have a ROS-mediated metabolic function in

678
M. C. Romero-Puertas
et al.
© 2002 Blackwell Science Ltd
,
Plant, Cell and Environment
,
25
, 677–686
plant abiotic stress (del Río
et al
. 2002). In recent studiescarried out in the authors’ laboratory with pea plants grownwith cadmium, results showed that this heavy metalinduced a concentration-dependent oxidative stress inleaves (Sandalio
et al
. 2001). At subcellular level, cadmiumproduced important alterations in the oxidative metabo-lism of peroxisomes and induced senescence symptoms inthese organelles (Romero-Puertas
et al
. 1999; McCarthy
etal
. 2001).In the present study, the effect of growing pea plants
with CdCl
2
was studied in leaf extracts and peroxisomes iso-lated from pea leaves in order to determine whether theoxidative protein modification and proteolytic degradationare involved in the mechanism of Cd toxicity.
MATERIALS AND METHODS
Plant material and growth conditions
Pea (
Pisum sativum
L., cv Lincoln) plants were obtainedfrom Royal Sluis (Enkhuizen, Holland). The plants weregrown in the greenhouse in aerated full nutrient media for14 d. The nutrient solution had a pH of 5·5 and the follow-ing composition in m
M
: NO
3–
, 12; H
2
PO
4–
, 4·0; SO
4
=
, 1·5; K
+
,5·0; Ca
2+
, 4·5; Mg
2+
, 1·5, and in
µ
M
: Fe, 91·3; B, 46; Cu, 1·1;Zn, 2·3; Mn, 9·3. Then media were supplemented with50
µ
M
CdCl
2
and plants were grown for another 14 d.Leaves were washed and homogenized in 10 m
M
sodiumphosphate buffer (pH 7·4), containing 1 m
M
EDTA, 2 m
M
dithiothreitol, 0·2% (v/v) Triton X-100, and 1 m
M
phenyl-methane sulphonyl fluoride (PMSF) (1/4; w/v). Homoge-nates were filtered and centrifuged at 27 000
×
g
for 20 minand the supernatants were used for the determinations.PMSF was not added to the buffers for homogenates thatwere to be used for protease activity determination.
Spectrophotometric determination of protein-bound carbonyls
For carbonyl quantification, the reaction with DNPH wasused basically as described by Levine
et al
. (1994). For eachdetermination two replicates and their respective blankswere used. Samples containing at least 0·5 mg protein wereincubated with 0·03% (v/v) Triton X-100 and 1% (w/v)streptomycin sulphate for 20 min to remove the nucleicacids. After centrifuging at 2000
×
g
, supernatants (200
µ
L)were mixed with 300
µ
L of 10 m
M
DNPH in 2
M
HCl. Theblank was incubated in 2
M
HCl. After 1 h incubation atroom temperature, proteins were precipitated with 10%(w/v) trichloroacetic acid (TCA) and the pellets werewashed three times with 500
µ
L of ethanol/ethylacetate (1/1).The pellets were finally dissolved in 6
M
guanidine hydro-chloride in 20 m
M
potassium phosphate buffer (pH 2·3)and the absorption at 370 nm was measured. Proteinrecovery was estimated for each sample by measuring theabsorption at 280 nm. Carbonyl content was calculatedusing a molar absorption coefficient for aliphatic hydra-zones of 22 000
M
−
1
cm
−
1
(Levine
et al
. 1994).
As a positive control assay, leaf extracts from untreatedpea plants (controls) were incubated for 2 h with differentH
2
O
2
concentrations (0–20 m
M
). The carbonyl content wasdetermined as described above.
Immunochemical detection of carbonyls
Proteins (0·5 mg) were incubated with 10 m
M
DNPH(100
µ
L) in 2
M
HCl for 1 h at room temperature. Sampleswere precipitated with 10% (w/v) TCA and pellets werewashed with ethanol/ethylacetate (1/1) three times. The pel-lets were re-suspended in 10 m
M
sodium phosphate buffer(pH 7·4) (Yan, Orr & Sohal 1998).
Proteins were separated by sodium dodecyl sulphate(SDS)-polyacrylamide gel electrophoresis (PAGE) in aMini-Protean II slab cell (Bio-Rad, Hercules, CA, USA) in12% (w/v) separating and 4% (w/v) stacking polyacryla-mide gels. Two gels were run simultaneously, one for pro-tein staining with Coomassie Brilliant Blue R-250 or silvernitrate and the other for immunochemical detection.Proteins were transferred onto polyvinylidene fluoride(PVDF) (Millipore Co., Bedford, MA, USA) membranesin a Bio-Rad Semi-Dry Transfer Cell according to Corpas
etal
. (1998). Oxidized proteins were detected with antibodiesagainst DNPH from Sigma-Aldrich Co. (St Louis, MO,USA) (1/40 000 dilution) and visualized either by anenhanced chemiluminiscence method (Corpas
et al
. 1998)or with alkaline phosphatase using 5-bromo-4-chloro-3-indolyl phosphate/nitrotetrazolium blue chloride (BCIP/NBT) as substrate.
Protein immunoprecipitation
Proteins (50–100
µ
g), derivatized with DNPH as mentionedabove, were precipitated by incubation overnight at 0
°
Cwith anti-DNPH-IgG (1/1000 dilution) in phosphate-buff-ered saline (PBS). Then, samples were loaded on a two-layer sucrose gradient (0·5 and 1·0
M
sucrose layers in PBS)in 0·5 mL Eppendorf tubes and were centrifuged at 5000
×
g
for 10 min. Supernatants were discarded and the pelletswere re-suspended in 0·1 N NaOH, 0·1 N HCl, 0·1% (w/v)SDS and 0·025% (w/v) bromophenol blue. The proteinswere separated by SDS-PAGE and transferred onto PVDFmembranes as described above. Oxidized proteins wereidentified by using antibodies against the following pro-teins: Mn-SOD, fructose-1,6-bisphosphatase, Rubisco, andglutathione reductase (GR) from pea leaves, and Cu, Zn-SOD and catalase from watermelon and cucumber cotyle-dons, respectively.
Purification of peroxisomes
Peroxisomes were purified from pea leaves by differentialand sucrose density-gradient centrifugation (35–60%; w/w)and were recovered from gradients by puncture with asyringe as described by López-Huertas, Sandalio & del Río(1995). To carry out assays of membrane permeability inintact peroxisomes, the organelles were purified by Percoll

Cadmium-induced protein oxidation
679
© 2002 Blackwell Science Ltd
,
Plant, Cell and Environment
,
25
, 677–686
density-gradient centrifugation (Sandalio, Palma & del Río1987) instead of sucrose gradients to avoid osmotic shock.The intactness of peroxisomes was determined by hydrox-ypyruvate reductase latency as described by Sandalio
et al
.(1987). Permeability was assayed by measuring the activityof glycolate oxidase (Kerr & Groves 1975). Commercialenzyme (0·027 units) was added to purified peroxisomesand the leakage of its substrate (glycolate) was monitoredby spectrophotometry following the increase of the absor-bance at 324 nm over 1 h. The slopes obtained were analy-sed by the Student’s
t
-test.
Proteolytic activity and protein degradation
Proteolytic activity in crude extracts was determined spec-trophotometrically by following the digestion of azocaseinat 340 nm (Distefano
et al
. 1997). Endoprotease isozymeswere detected by electrophoretic separation on SDS/poly-acrylamide gels (8%; w/v) containing 0·05% (w/v) gelatin asdescribed by Distefano
et al
. (1997). After electrophoresisthe proteases were renatured by rising gels with 2·5% (v/v)Triton X-100 and then were incubated with 250 m
M
Tris-HCl (pH 7·5) at 37
°
C for 4 h. The gels were stained with0·1% (w/v) Coomassie Brilliant Blue and the achromaticactivity bands were quantified by scanning on a Shimadzu(Kyoto, Japan) CS-9000 densitometer.
To determine protein degradation, crude extracts frompea leaves were incubated at 37
°
C for 16 h and then theprotein pattern was determined by SDS-PAGE and stain-ing with Coomassie blue. Simultaneously, protein-boundcarbonyl groups in proteolytically degraded proteins weredetected by Western blot and immunochemically stainedwith anti-DNPH as described above.
Protein determination
Protein concentration was measured according to themethod of Bradford (1976) using bovine serum albumin asstandard. Protein recovery after derivatization with DNPHwas estimated by measuring the absorbance at 280 nm.
RESULTS
Cadmium-dependent oxidative damage to proteins
In leaf extracts from pea plants grown in 50
µ
M
CdCl
2
anearly two-fold increase of carbonyl groups productiontook place (Fig. 1A). To determine whether Cd produced aselective protein modification, the oxidatively modifiedproteins were detected by immunochemical methods usingan antibody against DNPH. As shown in Fig. 1B, in bothcontrol and Cd-treated homogenates, proteins containingcarbonyl-groups showed molecular masses in the range 30–120 kDa, but the number and intensity of cross-reactingbands was considerably higher in homogenates from Cd-treated plants (Fig. 1B). The specificity of the antibodyreaction was checked by using controls without DNPH,
which did not give any labelling (data not shown).TheDNPH-reactive proteins analysed by densitometric scan-ning showed a two-fold increase in Cd-treated plants versusthe control plants, what is in accordance with the resultsobtained by the spectrophotometric method (Fig. 1A). Thepeak areas obtained by densitometric scanning revealedthat in both control and Cd-treated extracts, the 55 kDaprotein was the most abundant followed by those of molec-ular masses of 42, 60, 70 and approximately 120 kDa. How-ever, the 42 and 120 kDa proteins were more oxidized bythe Cd treatment in comparison with proteins from controlextracts.
Figure
1.
Oxidative protein modification in extracts from control and Cd-treated pea plants. (A) Oxidative proteins were determined as DNPH-reactive protein carbonyls by a spectrophotometric method. Data are mean
±
SE from at least three independent samples. Data were analysed by Duncan’s multiple-range test (
P
<
0·05). (B) Immunochemical detection of protein carbonyls in pea leaf extracts. DNPH derivatized proteins (10
µ
g) were individualized by SDS-PAGE (12% polyacrylamide, w/v) and transferred to PVDF membranes. Oxidized proteins were detected by using a commercial antibody against DNPH (1/40
000 dilution) and reactive bands were detected with luminol. Derivatized proteins were detected in the polyacrylamide gels by Coomassie blue staining. Molecular mass markers are indicated in the panel on the left.

680
M. C. Romero-Puertas
et al.
© 2002 Blackwell Science Ltd
,
Plant, Cell and Environment
,
25
, 677–686
Identification of oxidatively modified proteins
To identify the oxidized proteins, DNPH-derivatized sam-ples were immunoprecipitated with anti-DNPH-IgG, sepa-rated by SDS-PAGE and transferred onto PVDFmembranes. Proteins were identified by using specific anti-bodies against different enzymes. Controls were carried outby testing the antibodies before and after DNPH derivati-zation and immunoprecipitation (data not shown). Asshown in Fig. 2 only the antibodies against GR, CAT andMn-SOD gave positive reactions among the immunopre-cipitated proteins from Cd-treated plants. The Mn-SODand CAT protein bands showed a reduction as a result ofthe cadmium treatment. GR (57 kDa) and CAT (53 kDa)could correspond to the DNPH reactive bands close to the57 kDa band detected by Western blot (Fig. 1B). On thecontrary, CuZn-SOD (16·5 kDa) and fructose-1,6-bisphosphatase (43 kDa) were not recognized in the immu-noprecipitated fraction. Based on its molecular mass andWestern blot data of pea leaf extracts, the major DNPHreactive band of 55 kDa was attributed to the Rubisco largesubunit (RLS). As shown in Fig. 3, several bands of cross-reactivity with the antibody against Rubisco were detectedin control and Cd-treated plant extracts; the 55 kDa beingthe RLS and the 16 kDa the small subunit (RSS). The restof polypeptides detected between both subunits could cor-respond to proteolytic fragments of the RLS. The proteincontent of the RLS detected with the antibody againstRubisco was reduced in Cd-treated plants, whereas the RSSprotein content did not change as a result of the treatment
(Fig. 3). However, the peak area of the DNPH cross-reac-tive bands obtained for RLS was bigger in Cd-treatedplants, which is indicative of a higher degree of proteinoxidation (Fig. 1B).
Effect of H
2
O
2
on carbonyl formation in pea leaf extracts
To check the possible involvement of ROS in carbonylproduction, crude extracts from control pea leaves wereincubated with different H
2
O
2
concentrations (0–20 m
M
)for 2 h. The amount of carbonyl groups measured spectro-photometrically increased with the H
2
O
2
concentration(Fig. 4A), and the analysis by Western blot showed a pro-gressive enhancement in the reactivity against the DNPHantibody (Fig. 4B). Furthermore, the pattern of carbonyl-reactive proteins was basically the same as that obtained inleaf extracts from Cd-treated plants (Fig. 1B).
Effect of cadmium on the endoprotease activity and protein degradation
The endoprotease (EP) activity of pea leaf extracts wasdetermined spectrophotometrically by following the diges-tion of azocasein at 340 nm. The results obtained did notshow any significant change in the total EP activity as a con-sequence of Cd treatment (Fig. 5A). The analysis of EPactivity by SDS-PAGE in gelatin-containing gels showedthree different bands of activity (EP1–EP3) in both controland Cd-treated plants. The densitometric scanning of gelsshowed that there was a more than three-fold increase inthe activity of isozymes EP2 plus EP3, as a result of the Cdtreatment (Fig. 5B)
Figure
2.
Identification of specific oxidized proteins in pea leaf extracts. Proteins were derivatized and immunoprecipitated with anti-DNPH-IgG as indicated in Materials and Methods. Immunoprecipitated proteins were subjected to SDS-PAGE (12% polyacrylamide, w/v) and transferred onto PVDF membranes. Proteins were identified by using different antibodies. Lanes 1 and 2, non-derivatized control and Cd-treated plant extracts; lanes 3 and 4, derivatized and immunoprecipitated control and Cd-treated plant extracts. CAT and FBPase were detected with NBT/BCIP and the rest of proteins were detected with luminol.
Figure
3.
Immunochemical detection of Rubisco. Proteins from pea leaf extracts were individualized by SDS-PAGE and transferred onto PVDF membranes as described in Fig.
2. Proteins were detected by Western blotting and stained with NBT/BCIP. RLS, Rubisco large subunit; RSS, Rubisco small subunit. Molecular markers are indicated on the left.

Cadmium-induced protein oxidation
681
© 2002 Blackwell Science Ltd
,
Plant, Cell and Environment
, 25, 677–686
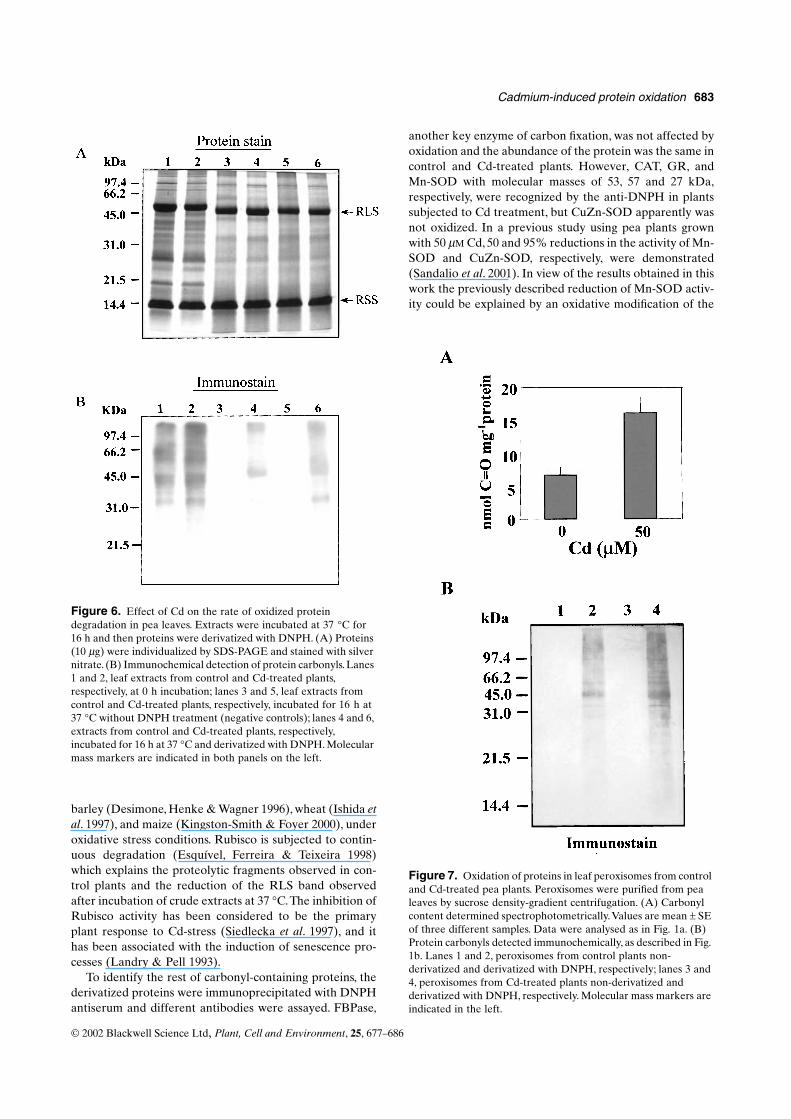
The rate of protein degradation by endogenous pro-teases was studied in leaf extracts from control and Cd-treated plants by SDS-PAGE and protein and immu-nochemical staining. Leaf extracts were incubated at 37 °Cfor 16 h and the proteins were then separated by SDS-PAGE and stained with silver nitrate (Fig. 6A). For carbo-nyl detection, samples were derivatized and detected asdescribed in Materials and Methods (Fig. 6B). After 16 hincubation, the pattern of proteins showed a slight reduc-tion in the number and intensity of bands as a consequenceof protein degradation by endoproteases, and the polypep-tide pattern did not show any specific modification due to
Cd treatment. The proteins most affected by degradationwere those in the molecular mass range 20–60 kDa. TheRLS band was reduced with the incubation time in bothcontrol and Cd-treated plants and its mobility was slightlyincreased (Fig. 6A). In Western blot analyses, the quantifi-cation of carbonyl content by densitometric scan showed areduction in the cross-reactivity with the anti-DNPH being77% in control plants (lane 4 versus lane 1) and 78% in Cd-treated plants (lane 6 versus lane 2). The majority ofdegraded proteins gave reactivity with the anti-DNPH,especially the 60 kDa protein which practically disappearedafter 16 h incubation.
Cadmium-dependent oxidative damage to proteins in isolated peroxisomes
The results obtained in purified peroxisomes were similarto those found in leaf homogenates. The total content ofcarbonyl groups in peroxisomes from Cd-treated plants,measured spectrophotometrically, was more than two-foldhigher than that in control plants (Fig. 7A). The pattern ofmodified proteins obtained by Western blot and immuno-labelling with the anti-DNPH showed cross-reactivitybands in the range 40–60 kDa in both control and Cd-treated plants, and the most prominent bands were those of45 and 55 kDa (Fig. 7b). The quantification of all theDNPH-reactive bands by densitometric scanning showed a2·4-fold increase in Cd-treated plants.
To investigate whether protein oxidation could affect theintactness of peroxisomal membranes the integrity oforganelles was studied by determining the latency ofhydroxypyruvate reductase activity and the leakage of sol-utes from the isolated peroxisomes. The integrity of leafperoxisomal membranes did not seem to be affected by Cdsince the intactness of organelles was about 70% in bothcontrol and Cd-treated plants (Table 1). The leakage of sub-strates from intact peroxisomes, measured by using exoge-nous glycolate oxidase, followed the same pattern incontrol and Cd-treated plants (Fig. 8). No differences wereobserved in the slopes of glycolate oxidase activity in con-trol and Cd-treated peroxisomes. Similar results wereobtained using exogenous hydroxypyruvate reductaseinstead of glycolate oxidase to measure the leakage of thesubstrate (data not shown). This means that apparently thepermeability of peroxisomal membranes was not affectedby the treatment with Cd.
DISCUSSION
In previous studies carried out in the authors’ laboratory itwas demonstrated that Cd can induce oxidative stress inpea leaves (Dalurzo et al. 1997; Romero-Puertas et al. 1999;Sandalio et al. 2001). The aim of the present study was toinvestigate the rate of protein oxidation by Cd in bothwhole leaf and in purified leaf peroxisomes, and to identifythe pattern of modified proteins. As oxidized proteins can
Figure 4. Oxidation of proteins in pea leaf extracts by different H2O2 concentrations. Leaf crude extracts were incubated for 2 h with H2O2 (0–20 mM) and proteins were derivatized with DNPH, as described in Materials and Methods. (A) Carbonyl content determined spectrophotometrically. Values are mean ± SE of three different samples Data were analysed as in Fig. 1A. (B) Immunochemical detection of protein carbonyls. DNPH derivatized proteins (10 µg) were individualized by SDS-PAGE and stained as described in Fig. 2. Molecular mass markers are indicated in the panel on the left.

682 M. C. Romero-Puertas et al.
© 2002 Blackwell Science Ltd, Plant, Cell and Environment, 25, 677–686
be the target for proteases, the study was also focused onthe endoprotease activity and the oxidatively modified pro-tein degradation of leaf extracts.
Oxidative stress is known to cause oxidative proteinmodification characterized by the production of carbonylgroups in the molecule which can be determined by deriva-tization with DNPH. The presence of these reactive groupshas been considered as a good index of oxidative stress(Levine et al. 1994). By using this measurement as an indexof ROS-induced cellular injury, it can be concluded thatcadmium promotes oxidative stress in pea plants. The basallevels of the carbonyl group content observed in controlplants could be the result of ROS generation as by-productsof normal physiological processes which produce small butdetectable levels of oxidatively modified proteins in the tis-sue. Increases of carbonyl contents have also been observedin maize seedlings after chilling-induced oxidative stress(Prasad 1996; Kingston-Smith & Foyer 2000), and insenescing nodules from pea and bean (Matamoros et al.1999).
The increase in the level of oxidized proteins as an effectof Cd treatment, can be explained by several factors. In aprevious study it was demonstrated that Cd produced areduction of chlorophyll content as well as disturbances inthe nutritional status of pea plants (Sandalio et al. 2001).This situation could promote metabolic disturbances inphotosystems I and II, leading to the generation of ROS inchloroplasts (Sandalio et al. 2001). The antioxidant capacity
was also affected by the Cd treatment, with the SOD andCAT activities being specifically depressed and this wascorrelated with an increase in the lipid peroxidation rate(Sandalio et al. 2001). The depletion of GSH due to the for-mation of Cd-phytochelatin complexes has been reportedto contribute to oxidative stress by diminishing the plantantioxidative capacity (Gallego, Benavides, & Tomaro1996). A two-fold increase in the H2O2 content has beenobserved in leaf peroxisomes from pea plants treated with50 µM Cd and this was associated with a rise in the activityof glycolate oxidase, one of the sources of H2O2 in peroxi-somes (Romero-Puertas et al. 1999)
The pattern of oxidized proteins determined by immun-ostaining with anti-DNPH IgG showed a Cd-inducedincrease of cross-reactivity. The main carbonyl-containingproteins were those of 55, 42, 60, 70, and approximately120 kDa. In chloroplasts isolated from wheat leaves underwater-stress, an increase in oxidative damage has also beenobserved and the polypeptides identified by anti-DNPHhad similar molecular masses to those reported in thepresent study, especially those of 68, 54 and 41 kDa (Tam-bussi et al. 2000). The major 55 kDa carbonyl-containingprotein was identified as the RLS, a key enzyme of photo-synthetic carbon fixation. Smaller fragments were also rec-ognized by the antibody with molecular masses of 48, 34, 24and 16 kDa. The smallest one corresponds to the RSSwhereas the rest are probably breakdown products of theRLS. Similar cleavage fragments have been observed in
Figure 5. Endoprotease activity (EP) in pea leaf extracts. (A) Total proteolytic activity was determined spectrophotometrically following the azocasein digestion at 340 nm. Values are mean ± SE of three different samples. Data were analysed as in Fig. 1A. (B) Endoprotease isozymes (EP1–EP3) were detected by SDS-PAGE (8% polyacrylamide, w/v) containing 0·05% gelatin and were stained with 0·1% (w/v) Coomassie blue. Molecular mass markers are indicated on the right. Proteolytic activity in the gels was quantified by densitometric scanning and expressed as arbitrary units (a u).
Cadmium-induced protein oxidation 683
© 2002 Blackwell Science Ltd, Plant, Cell and Environment, 25, 677–686
barley (Desimone, Henke & Wagner 1996), wheat (Ishida etal. 1997), and maize (Kingston-Smith & Foyer 2000), underoxidative stress conditions. Rubisco is subjected to contin-uous degradation (Esquível, Ferreira & Teixeira 1998)which explains the proteolytic fragments observed in con-trol plants and the reduction of the RLS band observedafter incubation of crude extracts at 37 °C. The inhibition ofRubisco activity has been considered to be the primaryplant response to Cd-stress (Siedlecka et al. 1997), and ithas been associated with the induction of senescence pro-cesses (Landry & Pell 1993).
To identify the rest of carbonyl-containing proteins, thederivatized proteins were immunoprecipitated with DNPHantiserum and different antibodies were assayed. FBPase,
another key enzyme of carbon fixation, was not affected byoxidation and the abundance of the protein was the same incontrol and Cd-treated plants. However, CAT, GR, andMn-SOD with molecular masses of 53, 57 and 27 kDa,respectively, were recognized by the anti-DNPH in plantssubjected to Cd treatment, but CuZn-SOD apparently wasnot oxidized. In a previous study using pea plants grownwith 50 µM Cd, 50 and 95% reductions in the activity of Mn-SOD and CuZn-SOD, respectively, were demonstrated(Sandalio et al. 2001). In view of the results obtained in thiswork the previously described reduction of Mn-SOD activ-ity could be explained by an oxidative modification of the
Figure 7. Oxidation of proteins in leaf peroxisomes from control and Cd-treated pea plants. Peroxisomes were purified from pea leaves by sucrose density-gradient centrifugation. (A) Carbonyl content determined spectrophotometrically. Values are mean ± SE of three different samples. Data were analysed as in Fig. 1a. (B) Protein carbonyls detected immunochemically, as described in Fig. 1b. Lanes 1 and 2, peroxisomes from control plants non-derivatized and derivatized with DNPH, respectively; lanes 3 and 4, peroxisomes from Cd-treated plants non-derivatized and derivatized with DNPH, respectively. Molecular mass markers are indicated in the left.
Figure 6. Effect of Cd on the rate of oxidized protein degradation in pea leaves. Extracts were incubated at 37 °C for 16 h and then proteins were derivatized with DNPH. (A) Proteins (10 µg) were individualized by SDS-PAGE and stained with silver nitrate. (B) Immunochemical detection of protein carbonyls. Lanes 1 and 2, leaf extracts from control and Cd-treated plants, respectively, at 0 h incubation; lanes 3 and 5, leaf extracts from control and Cd-treated plants, respectively, incubated for 16 h at 37 °C without DNPH treatment (negative controls); lanes 4 and 6, extracts from control and Cd-treated plants, respectively, incubated for 16 h at 37 °C and derivatized with DNPH. Molecular mass markers are indicated in both panels on the left.

684 M. C. Romero-Puertas et al.
© 2002 Blackwell Science Ltd, Plant, Cell and Environment, 25, 677–686
enzyme during cadmium treatment, whereas the reductionin the activity and protein content of CuZn-SOD could bedue to modifications of its biosynthesis at translational ortranscriptional level. The differential response of bothSODs against oxidative modifications could be due to dif-ferences in their primary and three-dimensional structure.In fact it has been observed previously that during the puri-fication of both enzymes from pea leaves, Mn-SOD wasmore sensitive to oxidation than CuZn-SOD (Palma et al.1998). Antibodies against CAT recognized a band of53 kDa in both control and cadmium-treated plants. Thepresence of oxidized CAT protein in the control plantscould be due to the high turnover of this protein innormal conditions, such as has been described by Streb,Michael-Knauf, & Feierabend (1993). With regard to GR,Cd produced the oxidative modification of this protein,although apparently it did not affect the active centre of themolecule, as no changes were previously observed in theGR activity of pea plants treated with Cd (Sandalio et al.2001). However, proteolytic degradation of the oxidizedGR can not be excluded and the absence of changes in
protein and activity of GR by Cd could be explained bytranscriptional or translational modifications to keep anadequate GR level to protect against Cd-stress.
To demonstrate that ROS are involved in the oxidativemodification of proteins, pea leaf extracts were incubatedwith increasing H2O2 concentrations and the resultsobtained mimicked those observed in plants grown with50 µM Cd. These results indicate that an oxidative stressimposed by cadmium could be responsible for protein oxi-dation in vivo.
It is well established that oxidatively modified proteinsare preferred substrates for proteolytic degradation andthis fact is correlated with an increase in the rate of proteinturnover (Pacifici & Davies 1990). For this reason, proteindegradation and proteolytic activity have been proposed asan index of oxidative stress. In this work, the proteolyticactivity was analysed to determine whether cadmium couldinduce endoprotease activity in the leaves of pea plantstreated with this metal. However, only a small increase inthe EP activity measured by native-PAGE was observed.No changes in protease activity have been found in poplarleaves under pro-oxidant conditions induced by ozoneexposition (Landry & Pell 1993). In maize seedlings sub-jected to low temperatures, which also gave rise to an accu-mulation of oxidized proteins, the protease activity was notmodified either (Prasad 1996). In spite of the absence ofchanges in total EP activity of leaf extracts due to Cd treat-ment, differences have been reported at subcellular level.In leaf peroxisomes from pea plants, aminopeptidase andendopeptidase activities were increased by cadmium treat-ment and this fact was associated with symptoms of senes-cence induced by this metal in peroxisomes (McCarthy etal. 2001).
Hydrogen peroxide is a well-known oxidant and one ofthe main products of peroxisomal metabolism. In leaf per-oxisomes purified from Cd-treated pea plants it wasrecently demonstrated that the H2O2 content increasedtwo-fold in comparison with control plants (Romero-Puer-tas et al. 1999). Accordingly, the effect of Cd-treatment onthe carbonyl-group content of the leaf peroxisomes wasstudied. The results obtained suggest that Cd produces oxi-dative damage in peroxisomes, although the functionalityof their membranes was not affected in terms of integrityand leakage of substrates, and this agrees with previousresults in pea leaves showing that Cd treatment did notproduce the lipid peroxidation of peroxisomal membranes(Romero-Puertas et al. 1999). The absence of oxidativedamage in peroxisomal membranes of Cd-treated plantscould be explained by the presence in these membranes ofthe efficient H2O2-scavenger ascorbate peroxidase, whoseactivity was increased two-fold by Cd treatment (Romero-Puertas et al. 1999), although changes in the lipidcomposition of peroxisomal membranes induced by Cdshould be also taken into account. Kinetic studies carriedout in rat liver mitocondria have demonstrated that pro-tein oxidation preceded formation of thiobarbituric acidreacting substances under oxidative stress conditions(Reinheckel et al. 1998). This could explain the absence of
Figure 8. Glycolate leakage in leaf peroxisomes from control and Cd-treated pea plants. Peroxisomes were purified from pea leaves by Percoll density-gradient centrifugation. Under isotonic conditions commercial GOX (0·027 units) was added to the suspension of peroxisomes and the absorbance corresponding to the reaction product (glyoxylate phenylhydrazone) was recorded for 1 h at room temperature. The graphic is representative of three independent experiments. �, peroxisomes from control plants; �, peroxisomes from Cd-treated plants.
Table 1. Latency of hydroxypyruvate reductase in intact leaf peroxisomes from control and Cd-treated pea leaves. Peroxisomes were purified from pea leaves by Percoll density-gradient centrifugation The hydroxypyruvate reductase activity was determined in isotonic medium (IM) in the absence and presence of 0·05% (v/v) Triton X-100
Cd (µM)
Hydroxypyruvate reductase (mmol hydroxypyruvate mL−1 min−1)
Latency (%)IM IM + Triton-100
0 1·09 3·84 71·650 1·86 6·00 69·0

Cadmium-induced protein oxidation 685
© 2002 Blackwell Science Ltd, Plant, Cell and Environment, 25, 677–686
oxidative damage in peroxisomal lipid membraneswhereas proteins are probably more affected. In mito-chondria from rabbit chondrocytes cell cultures, senes-cence produces an increase of ROS production as well ascarbonyl content and peroxides without changes in thestructure and functionality of mitochondrial membrane(Pfeuty & Gueride 2000). In spite of that, we cannotexclude the diffusion of H2O2 outside peroxisomes. Theseorganelles could also contribute to H2O2 production bydismutation of the O2
– produced in the cytosolic side ofperoxisomal membranes (López-Huertas et al. 1999)which would escape to the APX activity. A slight Cd-induced increase of O2
– production in peroxisomal mem-branes has been previously observed (Romero-Puertas etal. 1999). Oxidized proteins in peroxisomes can be effi-ciently cleaved by proteases present in these organelles(Distefano et al. 1997). The increase in proteolytic activityin peroxisomes was also associated with metabolic changesthat take place in these organelles that are induced by Cd,mainly the conversion of leaf peroxisomes into glyoxy-somes, which is characteristic of leaf senescence (McCar-thy et al. 2001).
In conclusion, in pea plants Cd induces an increase inthe level of oxidized proteins, probably mediated by H2O2,which affects important leaf antioxidative enzymes, andperhaps the photosynthetic carbon assimilation. Cadmiumalso causes a slight increase in proteolytic activity,although apparently this is not enough to degrade all theoxidized proteins whose accumulation to toxic levels couldproduce different cellular damages (Stadtman & Oliver1991). The biochemical changes observed in this work inpea plants appear to be associated with the oxidative stressand the induction of senescence produced by Cd toxicity(Sandalio et al. 2001; McCarthy et al. 2001). The oxidativedeterioration is considered an intrinsic feature of thesenescence process of leaves (del Río et al. 1998), butother symptoms of senescence such as photosynthesis ces-sation, or disintegration of organelle structure, have alsobeen described in Cd-treated pea plants (Sandalio et al.2001). The induction of senescence by Cd is a complexprocess and different factors are involved, including nutri-tional disturbances which can affect important metabolicprocesses such as photosynthesis. Cadmium can alsoinduce senescence by stimulating the biosynthesis of ethyl-ene, as has been described by Rascio et al. (1993). Theidentification of the specific oxidized proteins of leavesdescribed in this work, can improve our knowledge of themechanisms involved in Cd toxicity.
ACKNOWLEDGMENTS
M.C.R.-P. acknowledges a FPI fellowship from the Junta deAndalucía. The authors thank Dr Nishimura (NationalInstitute for Basic Biology, Okazaki, Japan), Dr R. Her-moso (Estación Experimental del Zaidín, CSIC, Granada,
Spain), and Dr P.M. Mullineaux (John Innes Centre, Nor-wich, UK) for their generous donation of the antibodiesagainst pumpkin CAT, pea Rubisco and fructose-1,6-bisphosphatase, and pea GR, respectively. This work wassupported by the European Union (Contrat HPRN-CT-2000–00094), grants 1FD97-0889-02 and PB98-0493-01from the DGESIC (Ministry of Education and Culture)and the Junta de Andalucía (Research group CVI 0192).
REFERENCES
Bradford M.M. (1976) A rapid and sensitive method for the quan-titation of microgram quantities of protein utilizing the principleof protein-dye binding. Analytical Biochemistry 72, 248–254.
Corpas F.J., Barroso J.B. & del Río L.A. (2001) Peroxisomes as asource of reactive oxygen species and nitric oxide signal mole-cules in plant cells. Trends in Plant Science 6, 145–150.
Corpas F.J., Barroso J.B., Sandalio L.M., Distefano S., Palma J.M.,Lupiáñez J.A. & del Río L.A. (1998) A dehydrogenase-medi-ated recycling system of NADPH in plant peroxisomes. Bio-chemical Journal 330, 777–784.
Dalurzo H.C., Sandalio L.M., Gómez M. & del Río L.A. (1997)Cadmium infiltration of detached pea leaves: effect on its acti-vated oxygen metabolism. Phyton (Austria) 37, 59–64.
De Filippis L.F. & Ziegler H. (1993) Effect of sublethal concentra-tions of zinc, cadmium and mercury on the photosynthetic car-bon reduction cycle of Euglena. Journal of Plant Physiology 142,167–172.
Desimone M., Hemke A. & Wagner E. (1996) Oxidative stressinduces partial degradation of the large subunit of ribulose-1,5-bisphosphate carboxylase/oxygenase in isolated chloroplasts ofbarley. Plant Physiology 111, 789–796.
Distefano S., Palma J.M., Gómez M. & del Río L.A. (1997) Char-acterization of endopeptidases from plant peroxisomes. Bio-chemical Journal 327, 399–405.
Esquível M.G., Ferreira R.B. & Teixeira A.R. (1998) Protein deg-radation in C3 and C4 plants with particular reference to ribu-lose bisphosphate carboxylase and glycolate oxidase. Journal ofExperimental Botany 49, 807–816.
Fodor A., Szabó-Nagy A. & Erdei L. (1995) The effects of cad-mium on the fluidity and H+-ATPase activity of plasma mem-brane from sunflower and wheat roots. Journal of PlantPhysiology 14, 787–792.
Gallego S.M., Benavides M.P. & Tomaro M.L. (1996) Effect ofheavy metal ion excess on sunflower leaves: evidence forinvolvement of oxidative stress. Plant Science 121, 151–159.
Ishida H., Nishimori Y., Sugisawa M., Makino A. & Mae T. (1997)The large subunit of ribulose-1,5 bisphosphate carboxylase/oxygenase is fragmented into 37 kDa and 16 kDa polypeptidesby active oxygen in the lysates of chloroplasts from primaryleaves of wheat. Plant and Cell Physiology 38, 471–479.
Kerr M.V. & Groves D. (1975) Purification and properties of gly-colate oxidase from Pisum sativum leaves. Phytochemistry 14,359–362.
Kingston-Smith A.H. & Foyer C.H. (2000) Bundle sheath proteinsare more sensitive to oxidative damage than those of the meso-phyll in maize leaves exposed to paraquat or low temperatures.Journal of Experimental Botany 51, 123–130.
Landry L.G. & Pell E.J. (1993) Modification of rubisco and alteredproteolytic activity in O3-stressed hybrid poplar (Populus max-imowizii x trichoacrpa). Plant Physiology 101, 1355–1362.
Levine R.L., Willians J.A., Stadtman E.R. & Shacter E. (1994)

686 M. C. Romero-Puertas et al.
© 2002 Blackwell Science Ltd, Plant, Cell and Environment, 25, 677–686
Carbonyl assays for determination of oxidatively modified pro-teins. Methods in Enzymology 233, 346–363.
López-Huertas E., Sandalio L.M. & del Río L.A. (1995) Integralmembranes polypeptides of pea leaf peroxisomes: characteriza-tion and response to plant stress. Plant Physiology and Biochem-istry 33, 295–302.
López-Huertas E., Corpas F.J., Sadalio L.M. & del Rio L.A. (1999)Characterization of membrane polypeptides from pea leaf per-oxisomes in superoxide radical generation. Biochemical Journal337, 531–536.
Matamoros M.A., Baird L.M., Escuredo P.R., Dalton D.A.,Minchin F.R., Iturbe-Ormaetxe I., Rubio M.C., Moran J.F.,Gordon A.J. & Becana M. (1999) Stress-induced legume rootnodule senescence. Physiological, and structural alterations.Plant Physiology 121, 97–111.
McCarthy I., Romero-Puertas M.C., Palma J.M., Sandalio L.M.,Corpas F.J., Gómez M. & del Río L.A. (2001) Cadmium inducessenescence symptoms in leaf peroxisomes of pea plants. Plant,Cell and Enviroment 24, 1065–1073.
Pacifici R.E. & Davies K.J.A. (1990) Protein degradation as anindex of oxidative stress. Methods in Enzymology 186, 485–502.
Palma J.M., López-Huertas E., Corpas F.J., Sandalio L.M., GómezM. & del Río L.A. (1998) Peroxisomal manganese superoxidedismutase: purification and properties of the isozyme from pealeaves. Physiologia Plantarum 104, 720–726.
Pfeuty A. & Gueride M. (2000) Peroxide accumulation withoutmajor mitochondrial alteration in replicative senescence. FEBSLetters 468, 43–47.
Prasad T.K. (1996) Mechanisms of chilling-induced oxidativestress injury and tolerance: changes in antioxidant system, oxi-dation of proteins and lipids and protease activities. Plant Jour-nal 10, 1017–1026.
Rascio N., Dalla Vecchia D., Ferreti M., Merlo L. & Ghisi R. (1993)Some effects of cadmium on maize plants. Archives of Environ-mental Contamination and Toxicology 25, 244–249.
Reinheckel T., Noack H., Lorenz S., Wiswedel I. & Augustin W.(1998) Comparison of protein oxidation and aldehyde formationduring oxidative stress in isolated mitochondria. Free RadicalResearch 29, 297–305.
del Río L.A., Pastori G.M., Palma J.M., Sandalio L.M., Sevilla F.,Corpas F.J., Jiménez A., López-Huertas E. & Hernández J.A.(1998) The activated oxygen role of peroxisomes in senescence.Plant Physiology 116, 1195–1200.
del Río L.A., Sandalio L.M., Palma J.M., Corpas F.J., López-Huertas E., Romero-Puertas M.C. & McCarthy I. (2002) Perox-isomes, reactive oxygen metabolism, and stress-related enzymeactivities. In Plant Peroxisomes (eds A. Baker & I. Graham).Kluwer Academic Publishers, Dordrecht, The Netherlands, inpress.
Romero-Puertas M.C., McCarthy I., Sandalio L.M., Palma J.M.,Corpas F.J., Gómez M. & del Río L.A. (1999) Cadmium toxicityand oxidative metabolism of pea leaf peroxisomes. Free RadicalResearch 31, S25–S32.
Sandalio L.M., Dalurzo H.C., Gómez M., Romero-Puertas M.C. &del Río L.A. (2001) Cadmium-induced changes in the growthand oxidative metabolism of pea plants. Journal of ExperimentalBotany 52, 2115–2126.
Sandalio L.M., Palma J.M. & del Río L.A. (1987) Localization ofmanganese superoxide dismutase in peroxisomes isolated fromPisum sativum L. Plant Science 51, 1–8.
Shaw B.P. (1995) Effect of mercury and cadmium on the activitiesof antioxidative enzymes in the seedling of Phaseolus aureus.Biologia Plantarum 37, 587–596.
Siedlecka A., Krupa Z., Samuelson G., Öquist G. & Gardeströn P.(1997) Primary carbon metabolism in Phaseolus vulgaris plantsunder Cd/Fe interaction. Plant Physiology and Biochemistry 35,951–957.
Somashekaraiah B.V., Padmaja K. & Prasad A.R.K. (1992) Phy-totoxicity of cadmium ions on germinating seedlings of mungbean (Phaseolus vulgaris): involvement of lipid peroxides inchlorophyll degradation. Physiologia Plantarum 85, 85–89.
Stadtman E.R. & Oliver C.N. (1991) Metal-catalyzed oxidation ofproteins. Physiological consequences. Journal of BiologicalChemistry 266, 2005–2008.
Stobart A.K., Griffits W., Bukhari I.A. & Sherwood R.P. (1985)The effect of Cd2+ on the biosynthesis of chlorophyll in leaves ofbarley. Physiologia Plantarum 63, 293–298.
Stohs S.J. & Bagchi D. (1995) Oxidative mechanism in the toxic-ity of metal ions. Free Radical Biology and Medicine 18, 321–336.
Streb P., Michael-Knauf A. & Feierabend J. (1993) Preferentialphotoinactivation of catalase and photoinhibition of photosys-tem II are common early symptoms under various osmoticand chemical stress conditions. Physiologia Plantarum 88,590–598.
Tambussi E.A., Bartoli C.G., Beltrano J., Guiamet J.J. & ArausJ.L. (2000) Oxidative damage to tylakoid proteins in water-stressed leaves of wheat (Triticum aestivum). Physiologia Plan-tarum 108, 398–404.
Wagner G.J. (1993) Accumulation of cadmium in crop plants andits consequences to human health. Advances in Agronomy 51,173–212.
Yan L.-J., Orr W.C. & Sohal R.S. (1998) Identification of oxidizedproteins based on sodium dodecyl sulfate-polyacrylamide gelelectrophoresis, immunochemical detection, isoelectric focusing,and microsequencing. Analytical Biochemistry 263, 67–71.
Received 29 November 2001; received in revised form 10 January2002; accepted for publication 10 January 2002




















