
Salt-induced oxidative stress in chloroplasts of pea plants
17
ELSEVIER Salt-induced oxidative stress in chloroplasts of pea plants “T A CiENCE Plant Science 105 (1995) 151-167 J.A. Hernhdeza, E. Olmosa, F.J. Corpasb, F. Sevilla*“, L.A. de1 Riob aDeparmmento de Nutricidn y Fisioiogia Vegetal, Centro de Edafoiogia y Biologia Aplicada del Segura, CSIC, Apdo. 195, E-30080 Murcia, Spain bDepartamento de Bioquimica. Biologia Celular y Molecular de Plantas. Estacibn Experimental del Zaidin. CSIC, Apdo. 419. E-18008 Granada. Spain Received 8 August 1994; revision received 13 December 1994; accepted 13 December 1994 Abstract The possible involvement of activated oxygen species in the mechanism of damage by NaCl stress was studied in chloroplasts from leaves of two cultivars of pea (Pistun safivum L.) with differential sensitivity to NaCl. Intact organelles were purified by centrifugation in density-gradients of Percoll. In chloroplasts from tolerant plants, NaCl stress produced a significant increase of CuZn-SOD II and ascorbate peroxidase activities as well as in ascorbate con- tent, while in those from sensitive plants NaCl produced increases in the HzO, content and lipid peroxidation and no changes were observed in the enzymatic activities. Chlorophyll content significantly decreased in chloroplasts from sen- sitive plants and chloroplast integrity was lower in sensitive than in tolerant plants. Electron microscopy showed that the thylakoidal structure of chloroplast was notably disorganized in the NaCl-treated leaves. In purified chloroplasts, an increase in the number and size of plastoglobuli was produced by NaCl in chloroplasts from tolerant plants and to a lesser extent, in chloroplasts from sensitive plants. The relative starch content only decreased in chloroplasts from tolerant plants by NaCI-treatment. Results obtained suggest that in the cellular toxicity of NaCl in pea plants, superoxide- and H202-mediated oxidative damage in chloroplasts may play an important role. Keywords: Pisum sativum; Salt stress; Oxidative stress; Chloroplast; Ascorbate peroxidase; Superoxide dismutase; Ac- tivated oxygen 1. Introduction In plant cells, chloroplasts, mitochondria and peroxisomes are important intracellular generators of activated oxygen species like 02: and Hz02 [l-3]. In chloroplasts, O2 : and O2 :-derived HzOz are mainly produced by the electron accep- l Corresponding author. tor of photosystem I [ 1,4]. In addition, singlet oxygen is commonly produced in illuminated chlo- roplasts by the transfer of an electron from an excited chlorophyll molecule to molecular O2 [ 1,4]. These activated oxygen species are highly reactive and, in the absence of any protective mechanism can damage different aspects of cell structure and function [5,6]. The metalloenzymes superoxide dismutases 0168-9452/95/$09.50 0 1995 Elsevier Science Ireland Ltd. All rights reserved SSDI 0168-9452(94)04047-K
Transcript of Salt-induced oxidative stress in chloroplasts of pea plants

ELSEVIER
Salt-induced oxidative stress in chloroplasts of pea plants
“T A CiENCE
Plant Science 105 (1995) 151-167
J.A. Hernhdeza, E. Olmosa, F.J. Corpasb, F. Sevilla*“, L.A. de1 Riob
aDeparmmento de Nutricidn y Fisioiogia Vegetal, Centro de Edafoiogia y Biologia Aplicada del Segura, CSIC, Apdo. 195, E-30080 Murcia, Spain
bDepartamento de Bioquimica. Biologia Celular y Molecular de Plantas. Estacibn Experimental del Zaidin. CSIC, Apdo. 419. E-18008 Granada. Spain
Received 8 August 1994; revision received 13 December 1994; accepted 13 December 1994
Abstract
The possible involvement of activated oxygen species in the mechanism of damage by NaCl stress was studied in chloroplasts from leaves of two cultivars of pea (Pistun safivum L.) with differential sensitivity to NaCl. Intact organelles were purified by centrifugation in density-gradients of Percoll. In chloroplasts from tolerant plants, NaCl stress produced a significant increase of CuZn-SOD II and ascorbate peroxidase activities as well as in ascorbate con- tent, while in those from sensitive plants NaCl produced increases in the HzO, content and lipid peroxidation and no changes were observed in the enzymatic activities. Chlorophyll content significantly decreased in chloroplasts from sen- sitive plants and chloroplast integrity was lower in sensitive than in tolerant plants. Electron microscopy showed that the thylakoidal structure of chloroplast was notably disorganized in the NaCl-treated leaves. In purified chloroplasts, an increase in the number and size of plastoglobuli was produced by NaCl in chloroplasts from tolerant plants and to a lesser extent, in chloroplasts from sensitive plants. The relative starch content only decreased in chloroplasts from tolerant plants by NaCI-treatment. Results obtained suggest that in the cellular toxicity of NaCl in pea plants, superoxide- and H202-mediated oxidative damage in chloroplasts may play an important role.
Keywords: Pisum sativum; Salt stress; Oxidative stress; Chloroplast; Ascorbate peroxidase; Superoxide dismutase; Ac- tivated oxygen
1. Introduction
In plant cells, chloroplasts, mitochondria and peroxisomes are important intracellular generators of activated oxygen species like 02: and Hz02 [l-3]. In chloroplasts, O2 : and O2 :-derived HzOz are mainly produced by the electron accep-
l Corresponding author.
tor of photosystem I [ 1,4]. In addition, singlet oxygen is commonly produced in illuminated chlo- roplasts by the transfer of an electron from an excited chlorophyll molecule to molecular O2 [ 1,4]. These activated oxygen species are highly reactive and, in the absence of any protective mechanism can damage different aspects of cell structure and function [5,6].
The metalloenzymes superoxide dismutases
0168-9452/95/$09.50 0 1995 Elsevier Science Ireland Ltd. All rights reserved SSDI 0168-9452(94)04047-K

152 J.A. Herncinde-_ et al. /Plant Science 105 11995) 151-167
(SOD; EC 1.15.1.1) converts 02: to Hz02. The hydrogen peroxide is scavenged by ascorbate per- oxidase (APX; EC 1.11.1.11) using ascorbate as an electron donor as a part of a system for the protec- tion of chloroplasts and other cellular compart- ments from oxidative damage (the ascorbate- glutathione cycle) [7,8]. The activities of SOD, ascorbate peroxidase and other enzymes associ- ated with the removal of oxyradical species have been reportedly influenced by various environ- mental factors. Among these factors, herbicidal action, xenobiotics, air pollutants, low tempera- ture, high light intensity, metal deficiency and tox- icity and water stress have been identified [3,9-L?].
In plants, the effect of salinity on important metabolic processes located in chloroplasts and mitochondria is well established [ 13-171. How- ever, little is known about the effect of salt stress on the activated oxygen metabolism of chloro- plasts, mitochondria, and peroxisomes, and the endogenous antioxidants of these cell organelles. This knowledge can supply information on the possible involvement of activated oxygen species in the mechanism of damage by NaCl stress, and also could allow deeper insights into the molecular mechanisms of plant tolerance to salt-induced oxi- dative stress.
In previous studies with leaves of pea (Pisum sativum L.) the presence of three electrophoretical- ly distinct SODS, an Mn-containing SOD and two CuZn-containing SODS (I and II), was reported [ 181. Mn-SOD has been demonstrated to be pre- sent both in mitochondria and peroxisomes [19,20], whereas CuZn-SOD II was located in chloroplasts [21], and CuZn-SOD I was distri- buted between mitochondria and the cytosol [20].
In recent experiments conducted with leaf mito- chondria and peroxisomes from pea plants grown with NaCl, we have demonstrated that NaCl stress induced an enhancement in the 02’ production by submitochondrial particles as well as a strong decrease in the mitochondrial Mn-SOD activity in sensitive plants [22]. However, in pea peroxisomes neither Mn-SOD activity nor NADH-dependent generation of 0,: were altered by salinity [23].
In this work, using chloroplasts purified from two cultivars of pea with different sensitivities to
NaCl, the effect of salt stress on various functional enzymes, enzymatic and chemical antioxidants, lipid peroxidation of membranes, and Hz02 con- centration in those organelles is reported. Results are compared with those responses obtained in leaf mitochondria and peroxisomes from the same pea cultivars treated with NaCl.
2. Material and methods
2.1. Plant material Seeds of various commercial cultivars of pea
(Pisum sativum L.) obtained from Royal Sluis (Enkhuizen, The Netherlands), were surface- sterilized (ethanol, 3 min) and germinated in ver- miculite with 0.5 mM CaS04 for 3 days. Healthy and vigorous seedlings were selected and grown in a growth chamber (Conviron PGW-36) in an aerated nutrient solution [24] for 7 days. The growth chamber was set to 28/18”C, 80% RH, and a 16-h photoperiod (203 pmol m-* s-l PAR). Plants were then transplanted to a similar medium supplemented with NaCl concentrations of 30-300 mM, and were grown for another 14 days. Pots containing four plants in 2.5 1 nutrient solu- tion were arranged in blocks at random, and each treatment (NaCl level and cultivar) was replicated four times. All the experiments were repeated three times in different periods. Plants were harvested and plant growth was determined by measuring fresh and dry weight of leaves, fresh weight of shoots, foliar area (in cm2), and dry and fresh weight of roots. Statistical analysis of results from the three different experiments was carried out ac- cording to Duncan’s multiple range test.
2.2. Isolation and purification of chloroplasts All operations were performed at 0-4°C. Intact
purified chloroplasts were obtained by differential and densitydgradient centrifugation in discon- tinuous gradients of Percoll, essentially as describ- ed by Palma et al. [21]. In brief, leaves (So-100 g) were blended at high speed for 2-3 s in a medium containing 0.35 M mannitol, 30 mM MOPS buffer (pH 7.5), 4 mM L-cysteine, 1 mM EDTA and 0.2% BSA (leaf to medium ratio 1:4, w/v), using a Polytron tissue homogenizer. Homogenates were filtered through four layers of nylon cloth and

J.A. Hermindez et al. /Plant Science IO5 (1995) 151-167 153
were centrifuged at 2200 x g for 30 s, and the pel- let was suspended in 0.3 M mannitol, 20 mM MOPS buffer (pH 7.2), 1 mM EDTA, 0.2% BSA. The suspension was centrifuged at 2200 x g for 30 s. Volumes (2.5 ml) of washed chloroplast suspen- sions in the same medium were layered on top of discontinuous density gradients of Percoll (21~60%; v/v) with a composition analogous to that described by Palma et al. [21]. Gradients were centrifuged at 13 000 x g for 30 min in a Beckman ultracentrifuge using a 60 Ti rotor. After cen- trifugation, the gradients were fractionated by up- ward displacement with 45% sucrose (w/w) using an Isco fractionatior model 185 equipped with an optical unit and absorbance detector. The transmittance of eluted samples was monitored at 280 nm and enzyme activities were measured in the different fractions collected ( 1.5 ml).
2.3. Enzyme assays
Total SOD activity of samples was determined as described by McCord and Fridovich [25]. SOD isozymes (CuZn-SODS and Mn-SOD) were separated by polyacrylamide gel electrophoresis on 10% cylindric gels, at pH 8.9, according to Davis [26]. Prior to electrophoresis, organelle sam- ples were diluted 2-5-fold with 50 mM K- phosphate, pH 7.8, containing Triton X-100 (0.1% final concentration; v/v) and stirred to solubilize membrane-bound SODS. SOD-isozymes were localized on the gels by the photochemical method of Weissiger and Fridovich [27]. The isozyme ac- tivity was quantified by recording the transmit- tance of gels in a Shimadzu CS-9000 densitometer. In fractions from Percoll density-gradient cen- trifugation the SOD activity determinations were carried out quickly and samples were appropriate- ly diluted in order to reverse the Percoll inhibitory action on SODS [21]. Catalase (EC 1.11.1.6) cytochrome c oxidase (EC 1.9.3.1) and chlorophyll were used as markers for peroxisomes, mitochon- dria and chloroplasts, respectively and were assayed as described in Hernandez et al. [22] and Corpas et al. [23]. Ascorbate peroxidase was deter- mined according to Hossain and Asada [28]. Mea- surement of the density of the Percoll was carried out at room temperature using a refractometer.
2.4. Other assays The amount of lipid peroxidation products of
chloroplastic membranes was estimated by deter- mining the concentration of thiobarbituric acid reacting substances (TBARS) according to Buege and Aust 1291. The concentration of H202 in purified intact organelles was determined im- mediately after their purification by a peroxidase- coupled assay with 4-aminoantipyrine and phenol as donor substrates [30]. In chloroplasts, ascor- bate concentration was determined according to Law et al. [31], and a-tocopherol was extracted from thylakoid membranes and determined by HPLC using a fluorescence detector [32]. The activity of photosystem II (PSII) was determined by the method of Lilley et al. [33]. Proteins were assayed by the method of Bradford [34] and chlorophyll was measured according to Arnon [35]. Sodium and potassium concentrations in leaves were estimated by flame emission spectro- metry, and chloride was measured by a poten- tiometric method i [36]. For determining the nutrient contents in chloroplast suspensions, the extraction method of Schriippel-Meier and Kaiser [37] was used. Nutrient concentrations were estimated by atomic absorption spectrophoto- metry, except for chloride which was measured by a calorimetric method [38].
The integrity of isolated and purified chloro- plasts was determined by measuring the ferri- cyanide reduction spectrophotometrically at 410 run [32].
2.5. Electron microscopy Samples of leaf tissue from NaCl-treated and
control plants were fixed and processed for elec- tron microscopy as described by Palma et al. [ 111, and were examined with a Zeiss EM 1OC electron microscope at 60 kV.
Isolated chloroplasts were washed with washing media and centrifugated as above (2000 x g, 30 s).
Pellets were resuspended in 1 ml of washing medi- um containing 0.2% (v/v) glutaraldehyde (Fluka) and centrifuged in microfuge (5000 x g, 30 s). Pellets were fixed during 2 h 30 min at 4°C in 0.1 M cacodilate buffer (Sigma) pH 7.2; containing 0.3 M mannitol (Sigma) and 2% (v/v) glutaraldehide. After fixation, chloroplast pellets were washed
154 J.A. Hermindez et al. /Plant Science I05 (1995) 151-167
three times in 0.1 M cacodilate buffer, pH 7.2. Postlixation was performed in 1% osmium tetrox- ide (Serva) (w/v) prepared in the same buffer for 2 h 30 min at 4°C.
Dehydrations were done in an alcohol series and samples were imbedded in Spurr’s resin [39]. Sam- ples were cut using an ultramicrotome Reiss, mounted on copper grids and counterstained with acetate uranile. Samples were examined with both a Zeiss ElOM and E-109 electron microscope at 60 kV.
All data were determined over 50 chloroplasts with a magnification of x 3500. Chloroplast di- ameter was measured as the major diameter and the number of plastoglobuli was counted in each using a magnification of x 7000. Starch grains were analysed as the percentage of chloroplasts in which they were present.
3. Results
In a first set of experiments, different pea cultivars were used and by studying the effect on plant growth of a range of NaCl concentrations (30-300 mM) in the nutrient media, we were able to discriminate one relatively NaCl-tolerant cultivar (cv. Granada) and another cultivar more sensitive to NaCl (cv. Challis). In these two se- lected cultivars, the effect of NaCl on the growth,
assesed by leaf fresh weight, leaf dry weight and leaf area, was studied. At 70 mM NaCl, the growth of the tolerant cultivar was only slightly affected, whereas that of the sensitive cultivar was significantly inhibited (Table 1). These growth re- sults were similar to those using a dry weight basis. After treatment with 70 mM NaCl leaf dry weight decreased to about 44% in tolerant and 62% in sen- sitive plants, relative to control leaves (Table 1). The foliar concentration of Na+ and Cl- increas- ed in both cultivars, but the uptake of these nutrients from the solution was higher in sensitive than in tolerant plants (Table 2). Leaves from both NaCl-treated plants had decreased iron concentra- tions and the same was true for the copper content of sensitive plants, whereas in tolerant plants the copper level increased as a result of NaCl treat- ment (Table 2). Coinciding with these nutrient content modifications, there was a loss of chlorophyll in both cultivars, the leaf chlorophyll concentration being significantly lower in sensitive (from 3.06 to 1.71 mg/g fresh wt.) than in tolerant plants (from 3.57 to 2.17 mg/g fresh wt.).
3.1. Chtoroplusts The distribution of leaf chloroplasts from toler-
ant plants over the Percoll gradient after cen- trifugation is shown in Fig 1. Using this method, two intact chloroplast bands were obtained (frac-
Table I Plant growth of a sensitive and a moderately NaCl-tolerant cultivar of Pisum sarivum L. after a growth period of 14 days in nutrient
solutions containing a range of NaCl concentrations
NaCl cv. Challis cv. Granada
(mM) LA LFW LDW LA LFW LDW
0 100.00 f 0.76 100.00 f 3.15 100.00 f 0.78 100.00 zrz 1.28 100.00 f 0.03 100.00 l 0.44 30 99.08 f I.61 83.76 f 0.37.. 69.92 f 0.26* 98.66 zt 0.45 85.78 f 0.33; 71.20 f 0.33* 50 77.89 l 1.74; 70.29 f 2.21* 60.54 zt 0.16* 90.79 zt 1.87** 81.48 f 4.29* 62.90 f 2.43. 70 41.29 zt 1.89’ 46.30 f 2.58* 38.28 f 1.04* 61.53 zt 1.23; 78.67 f 1.32’ 55.92 f 0.44*
105 14.23 f 0.31’ 13.28 zt l.lO* 17.70 f 0.78’ 25.95 f 0.36* 32.89 f 0.99’ 26.02 f 0.55’ 140 8.18 za 0.24* 3.87 f 0.02’ 16.92 f 0.39* 17.42 zt 0.36* 20.66 f 0.02; 18.88 l 0.55; 210 7.89 f 0.18* 3.32 zt 0.06; 14.45 zt 0.26* 10.32 zt 0.14’ 7.27 f 0.05* 14.51 f 0.33*
Values represent residual growth (%) of the control. Mean values f S.E.M. from three independent experiments are shown. Challis.
sensitive cultivar; Granada, tolerant cultivar.
Abbreviations: LA, leaf area (cm*/plant); LFW, leaf fresh weight (g/plant); LDW. leaf dry weight (giplant).
*Differences from control values were significant at P < 0.001. **Differences from control values were significant at P < 0.01.

Tab
le
2
Nut
rien
t co
nten
ts
in
leav
es
of
cont
rol
and
NaC
I-tr
eate
d pl
ants
Cul
tivar
an
d tr
eat-
N
a (m
mol
/g
K
(mm
ohg
Cl
(mm
ohg
Fe
(gm
ol/g
C
u (p
mol
lg
men
t dr
y w
t.)
dry
wt.)
dr
y w
t.)
dry
wt.)
dr
y w
t.)
NaC
I-to
lera
nt
Con
trol
0.
15
f 0.
04
1.22
f
0.08
0.
12
f 0.
01
2.83
f
0.30
0.
32
zt 0
.01
70 m
M
NaC
l I.
1 I l
0.09
* 1.
09
f 0.
03
0.90
f
0.02
; 2.
01 l
0.20
***
0.89
f
0.16
***
NaC
I-se
nsiti
ve
Con
trol
0.
12 l
0.03
1.
25
f 0.
05
0.19
f
0.01
3.
40
f 0.
35
1.48
f
0.05
70 m
M
NaC
l 1.
43
f 0.
02*
1.10
f
0.03
**
1.82
zt
0.0
3*
2.21
f
0.33
***
0.44
f
0.04
*
Val
ues
are
mea
ns
A S
.E.M
. of
at
le
ast
six
diff
eren
t le
af
extr
acts
ea
ch
of
cont
rol
and
NaC
l-tr
eate
d pl
ants
.
*Dif
fere
nces
fr
om
cont
rol
valu
es
wer
e si
gnif
ican
t at
P
< 0.
001.
**D
iffe
renc
es
from
co
ntro
l va
lues
w
ere
sign
ific
ant
at
P c
0.01
.
***D
iffe
renc
es
from
co
ntro
l va
lues
w
ere
sign
ific
ant
at: P < 0.05.
Mn
(gm
ol/g
Z
n (p
moV
g
dry
wt.)
dr
y w
t.)
1.93
l
0.43
1.
09
f 0.
12
1.93
f
0.23
0.
99
zt 0
.06
1.93
zt
0.2
3 I.
18
+ 0.
10
2.00
f
0.27
1.
04
zt 0
.06
156 J.A. Hernhdez et al. /Plant Science IOS (199s) 151-167
t 11.360
5-
ii k!
CJ 3-
a0
1.
1
1 5 10 15 20
Fmction number
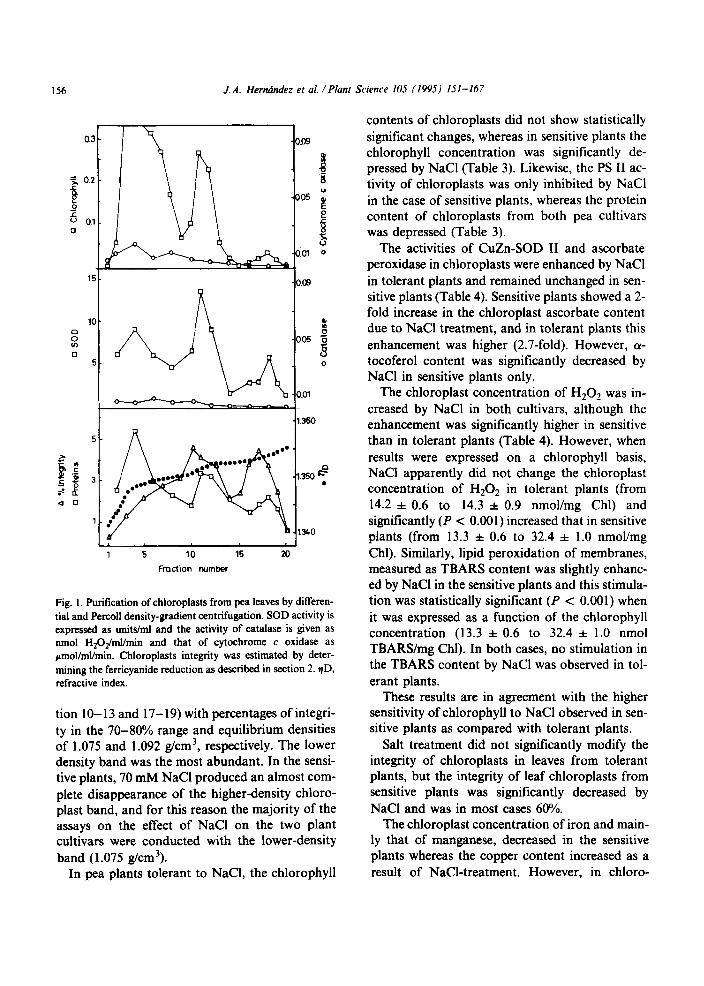
Fig. 1. Purification of chloroplasts from pea leaves by differen- tial and Percoll density-gradient centrifugation. SOD activity is expressed as units/ml and the activity of catalase is given as nmol H~O#nl/min and that of cytochrome c oxidase as ~mol/ml/min. Chloroplasts integrity was estimated by deter- mining the ferricyanide reduction as described in section 2. qD, refractive index.
tion IO- 13 and 17- 19) with percentages of integri- ty in the 70-80% range and equilibrium densities of 1.075 and 1.092 g/cm3, respectively. The lower density band was the most abundant. In the sensi- tive plants, 70 mM NaCl produced an almost com- plete disappearance of the higher-density chloro- plast band, and for this reason the majority of the assays on the effect of NaCl on the two plant cultivars were conducted with the lower-density band (1.075 g/cm3).
In pea plants tolerant to NaCl, the chlorophyll
contents of chloroplasts did not show statistically significant changes, whereas in sensitive plants the chlorophyll concentration was significantly de- pressed by NaCl (Table 3). Likewise, the PS II ac- tivity of chloroplasts was only inhibited by NaCl in the case of sensitive plants, whereas the protein content of chloroplasts from both pea cultivars was depressed (Table 3).
The activities of CuZn-SOD II and ascorbate peroxidase in chloroplasts were enhanced by NaCl in tolerant plants and remained unchanged in sen- sitive plants (Table 4). Sensitive plants showed a 2- fold increase in the chloroplast ascorbate content due to NaCl treatment, and in tolerant plants this enhancement was higher (2.7-fold). However, (Y- tocoferol content was significantly decreased by NaCl in sensitive plants only.
The chloroplast concentration of H,Oz was in- creased by NaCl in both cultivars, although the enhancement was significantly higher in sensitive than in tolerant plants (Table 4). However, when results were expressed on a chlorophyll basis, NaCl apparently did not change the chloroplast concentration of H202 in tolerant plants (from 14.2 f 0.6 to 14.3 i 0.9 nmoYmg Chl) and significantly (P < 0.001) increased that in sensitive plants (from 13.3 i 0.6 to 32.4 f 1.0 nmol/mg Chl). Similarly, lipid peroxidation of membranes, measured as TBARS content was slightly enhanc- ed by NaCl in the sensitive plants and this stimula- tion was statistically significant (P < 0.001) when it was expressed as a function of the chlorophyll concentration (13.3 f 0.6 to 32.4 f 1 .O nmol TBARS/mg Chl). In both cases, no stimulation in the TBARS content by NaCl was observed in tol- erant plants.
These results are in agreement with the higher sensitivity of chlorophyll to NaCl observed in sen- sitive plants as compared with tolerant plants.
Salt treatment did not significantly modify the integrity of chloroplasts in leaves from tolerant plants, but the integrity of leaf chloroplasts from sensitive plants was significantly decreased by NaCl and was in most cases 60%.
The chloroplast concentration of iron and main- ly that of manganese, decreased in the sensitive plants whereas the copper content increased as a result of NaCl-treatment. However, in chloro-
J.A. Hermindez et al. /Plant Science IO5 (1995) 151-167 157
Table 3 Photosystem II activity, chlorophyll and protein content in leaf chloroplasts of control and NaCl-treated plants
Cultivar and Chlorophyll Protein PS II activity treatment (m&ml) (mg/ml) (pm01 O,/mg Chl/h)
NaCl-tolerant Control 0.406 f 0.06 2.96 f 0.08 57.4 f 6.9 70 mM NaCl 0.263 & 0.03 I.44 f O.l6* 57.1 f 3.4
NaCI-sensitive Control 0.433 f 0.05 3.31 f 0.22 63.5 * 2.5 70 mM NaCl 0.161 f 0.02* 1.05 f 0.09’ 47.5 f 3.481
Mean values f S.E.M. from at least 6 separate chloroplasts preparations each of control and NaCl-treated plants are shown. *Differences from control values were significant at P c 0.001. **Differences from control values were significant at P < 0.01.
plasts from tolerant plants a decrease in manganese only was observed (Table 5). Unlike that found in leaves, the chloroplast concentration of Cl- was not modified by NaCl, but the Na+ content was increased in chloroplasts from sensi- tive plants.
3.2. Electron microscopy The cytomorphological effects of NaCl were
essentially the same in the two cultivars. NaCl treatment produced alterations in the ultrastruc- ture of leaf cells. Cellular membranes were notably disorganized in the NaCl-treated leaves, and the
chloroplasts showed disarranged thylakoids and increased numbers of plastoglobuli (Figs. 2 and 3).
Electron microscopy of the two chloroplasts bands showed the absence of other organelles like peroxisomes and mitochondria as contaminants, indicating the high level of purification obtained. In general, centrifugation brought about a slight displacement of the thylakoids and lamelae sys- tems, although the organelle integrity was not af- fected. The higher density band showed a higher content of starch granules compared with the chlo- roplast population of lower density (Table 6). In the latter, the major diameter was higher than that
Table 4 Hydrogen peroxide, antioxidant contents, lipid peroxidation, and enzymatic activities in leaf chloroplasts of control and NaCl-treated plants
Hz% (nmohmg protein)
TBARS (nmohmg protein)
Ascorbate (nmohmg protein)
a-Tocopherol
Wmg protein)
CuZn-SOD II (Uimg protein)
Ascorbate peroxidase (nmoUmg protein/min)
NaCI-tolerant Control 70 mM NaCl
NaCI-sensitive Control 70 mM NaCl
1.74 f 0.10 0.52 f 0.04 75.5 l 4.9 1.57 f 0.23 5.13 f 0.33 23.77 f 2.01 2.40 * 0.13* 0.55 * 0.07 203.4 & 9.7* 1.27 f 0.05 8.46 f 0.55* 47.94 f 0.75’
1.84 + 0.13 0.84 f 0.10 68.0 f 9.8 1.78 j: 0.16 6.20 f 0.40 27.04 f 1.32 3.95 l 0.41+ I.11 * 0.11 145.1 + 15.2’; 0.83 * O.l3* 6.50 f 0.27 29.81 f 1.56
Hydrogen peroxide, ascorbate content and lipid peroxidation were estimated in intact chloroplasts. The a-tocopherol content was determined in thylakoid membranes. For the determination of enzyme activities, chloroplasts were broken by osmotic shock. Mean values f S.E.M. from at least six separate chloroplasts preparations each of control and NaCI-treated plants are shown. *Differences from control values were significant at P < 0.001. **Differences from control values were significant at P < 0.01.

Tab
le 5
M
iner
al c
onte
nts
in c
hlor
opla
sts
susp
ensi
ons
of c
ontr
ol
and
NaC
I-tr
eate
d pe
a pl
ants
Cul
tivar
and
N
a (m
g/m
g K
fm
@m
g C
l (m
g/m
g Fe
(C
cg/m
g C
u W
mg
Mn
(pgl
mg
Zn
Wm
g tr
eatm
ent
chlo
roph
yll)
ch
loro
phyl
l)
chlo
roph
yll)
ch
loro
phyl
l)
chlo
roph
yll)
ch
loro
phyl
l)
chlo
roph
yll)
NaC
l-to
lera
nt
Con
trol
0.
23 z
t 0.
03
0,36
z+z
0.04
0.
11 f
0.
01
3.94
zt
0.32
0.
51 f
0.
11
0.88
zk
0.05
2.
29 z
t 0.
10
70 m
M N
aCl
0.22
*
0.01
0.
27 f
0.
02
0.12
f
0.02
3.
11 f
0.
34
0.51
f
0.03
0.
66 f
0.
03*
2.05
zt
0.10
N
aCI-
sens
itive
C
ontr
ol
0.21
f
0.01
0.
28 z
iz 0.
02
0.09
f
0.01
3.
01 z
t 0.
38
0.69
f
0.03
0.
68 f
0.
02
2.87
zt
0.15
70
mM
NaC
I 0.
37 l
0.02
* 0.
34 f
0.
04
0.11
f
0.01
1.
73 f
0.07
” 0.
60 f
0.
10
nd
3.68
f
0.26
**
Mea
n va
lues
f
S.E
.M.
of a
t le
ast
thre
e di
ffer
ent
chlo
ropl
asts
ex
trac
ts
each
of
cont
rol
and
NaC
l-tr
eate
d pl
ants
, ar
e pr
esen
ted.
nd
, no
t de
tect
ed.
*Dif
fere
nces
fro
m c
ontr
ol
valu
es w
ere
sign
ific
ant
at P
<
0.
001.
**
Dif
fere
nces
fr
om c
ontr
ol
valu
es w
ere
sign
ific
ant
at P
<
0.
05.

J.A. Hermindez et al. /Plant Science IO5 (1995) 151-167
Fig. 2. Electron micrographs from leaves of Pisum sarivum (cv. Granada). Plants were grown without NaCl (A, x 28 Ooo) and with
70 mM NaCl (B, x 28 000). G, grana; M, mitochondria; S, starch; arrow, plastoglobuli.

Fig. 3. Electron micrographs from leaves of Pisum surivum (cv. Challis). Plants were grown without NaCl (A, x 28 000) and with 70 mM NaCl (B, x 28 000).
J.A. Hernhdez et al. /Plant Science IO5 (1995) 151-167
Table 6 Morphometric characteristics of the two chloroplasts bands of control and NaCl-treated plants
Cultivar
NaCl-tolerant
NaCl-sensitive
Chloroplast population
Lower-density
Higher-density
Lower-density
Higherdensity
Treatment
Control 70 mM NaCl Control 70 mM NaCl Control 70 mM NaCl Control 70 mM NaCl
Diameter Plastoglobuli
(ccm) number
5.4 * 0.15 8.7 zt 0.48 4.6 f 0.08. 18.6 l 0.862 4.7 f 0.13 9.1 f 0.44 4.5 * 0.09 12.3 zt 0.61* 5.7 f 0.07 8.3 zt 0.44 5.7 l 0.1 I 10.4 zt 0.72+’ 4.7 f 0.11 3.5 l 0.10 nd nd
Starch (%)
20.3 12.5 53.7 42.8 20.2 26.0 57.5 nd
Mean values f S.E.M. from at least 25 separate chloroplasts preparations each of control and NaCl-treated plants are shown. nd, not determined. *Differences from control values were significant at P < 0.001. **Differences from control values were significant at P < 0.05.
on the higher-density chloroplast band (Table 6). In tolerant plants, salt treatment produced a reduction in the major chloroplast diameter, but that did not change in the same lower-density band of sensitive plants (Table 6). In these low-density chloroplast bands from both plants, there was an increase in the number and size of plastoglobuli by NaCl, which was more significant in the tolerant plants (Figs. 4A,C and 5A,C). In these plants, the same NaCl effect was found in the higher-density chloroplast population (Figs. 4B,D and 5B).
4. Discussion
In NaCl-sensitive plants, the addition of 70 mM NaCl to the nutrient solutions resulted in a con- siderable decrease in the fresh and dry weight of leaves, leaf area and chlorophyll content, and the oldest leaflets started to develop necrotic margins within 14 days of salt addition. The reduction in leaf chlorophyll concentration is a general phe- nomenon in sensitive plants subjected to salt stress 1401.
In glyophytes the inhibition of photosynthetic capacity under salinity and drought-stress condi- tions is due, in part, to the closure of stomata to limit transpiration, although direct salt effects on several biochemical and physiological processes are known too [40]. In recent years, the over- production of activated oxygen species in chloro-
plasts of plants under drought-stress has been described [41], and a similar mechanism may operate under salt conditions. As soon as the CO2 concentration decreases inside the chloroplasts, as a result of stomata closure, there is also a lower availability of NADP+ to accept electrons from PS I, thus initiating O2 reduction with the con- comitant generation of activated oxygen species [42]. The results reported here show that in chloro- plasts from salt-sensitive pea plants, salinity pro- duces important decreases in the chlorophyll content and PS II activity and, at the same time, stimulates the amount of TBARS and the Hz02 concentration. This suggests that salinity induces an overproduction of O2 7 and O2 :-derived H202 in salt-sensitive plants, which is only partially counterbalanced by the ascorbate and (Y- tocopherol levels, and the CuZn-SOD II and as- corbate peroxidase activities present in chloro- plasts of these plants. These results support the idea proposed by Singha and Choudhuri [43] that H202 could play an important role in the mecha- nism of salt injury and the same has also been pro- posed for drought-stress damage [44]. However, in chloroplasts from tolerant plants, NaCl produced a significant increase in CuZn-SOD II and ascor- bate peroxidase activities. This could represent an important protection mechanism against salt- induced 02: and H202 production in this organelle, as evidenced by the unchanged level of lipid peroxidation and PS II activity, as well as by
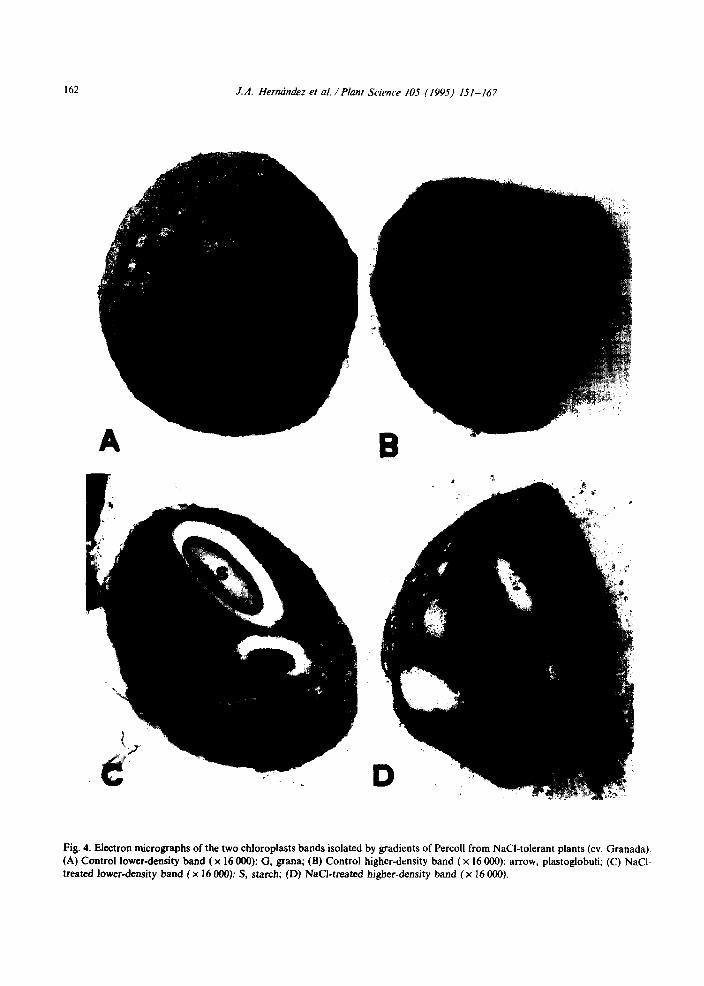
162 J.A. Hernhdez et al. / Planz Science 105 (1995) 151-167
Fig. 4. Electron micrographs of the two chloroplasts bands isolated by gradients of Percoll from NaCI-tolerant plants (cv. Granada). (A) Control lower-density band (x I6 000): G, grana; (B) Control higher-density band ( x I6 000): arrow, plastoglobuli; (C) NaCl- treated lower-density band (x I6 000): S, starch; (D) NaCI-treated higher-density band (x 16 000).

J.A. Hernhdez et al. /Plant Science 105 (1995) 151-167 163
Fig. 5. Electron micrographs of the two chloroplasts bands isolated by gradients of Percoll from NaCl-sensitive plants (cv. Challis).
(A) Control lower-density band (x 16 000): G, grana; (B) Control higher-density band ( x 16 000): arrow, plastoglobuli; (C) NaCl-
treated lower-density band ( x 16 000): S, starch.

164 J.A. Hernrindez et al. /Plant Science IO5 (1995) 151-167
the lower Hz02 production as compared with the sensitive plants.
In plants resistant to different types of en- vironmental stress conditions, increased levels of enzymes involved in O2 : detoxification have been found [3,9,12,45&J. In pea plants, clotibrate in- duces peroxisome proliferation and oxidative stress mediated by activated oxygen species in peroxisomes, and these effects are parallel to a decrease in the specific activities of catalase and Mn-SOD, the main peroxisomal defenses against H,Oz and O2 : [1 11. All these results are in agree- ment with those reported in this work on the re- sponse of chloroplast ascorbate peroxidase and CuZn-SOD, and with our recent results on mito- chondria from the same pea cultivars under salt stress [22]. Likewise, the induction of two CuZn- SODS in a selected calli from leaves of Pisum sativum cv. Challis, resistant to 85.5 mM NaCl has recently been described [47].
In mitochondria from tolerant plants, NaCl induced an increase in the activity of Mn-SOD [22]. Moreover, NaCl enhanced the rate of 02: production by submitochondrial particles in both sensitive and tolerant cultivars [22]. However, only in mitochondria from sensitive plants, a significant decrease in Mn-SOD and CuZn-SOD I activities took place. This showed that in salt-sensitive plants, unlike salt-tolerant plants, the ability of mitochondria to scavenge 02: radicals was diminished under conditions of enhanced produc- tion of these activated oxygen species due to salt stress. This could also explain the higher amount of TBARS found in mitochondrial membranes from NaCl-sensitive plants [22]. Salt-sensitivity of Mn-SOD and CuZn-SOD I has also been found in Vigna unguiculata, a moderately salt-sensitive plant [48].
Low levels of 02: per se can damage enzymes and membranes [49]. However, the main oxidative threat for chloroplast and other cell organelles stems from the generation of the highly reactive hydroxyl radicals (*OH) by a metal-catalysed site- specific Haber-Weiss reaction [7]. Hydroxyl radi- cals are strong oxidizing species. Therefore, NaCl- tolerant plants could have evolved a protection mechanism against the increased production of OzT -dependent toxic species in chloroplasts by
increasing the CuZn-SOD and ascorbate peroxi- dase activities. In this way, toxic levels of 02; radicals and H202 in these cell organelles could be removed, and the eventual formation of highly toxic -OH radicals avoided.
Conversely, in leaf peroxisomes isolated from the same pea cultivars, we have observed that the Mn-SOD activity and the NADH-dependent generation of 02: by peroxisomal membranes was not altered by NaCl [23]. The observed decrease in the internal H,02 concentration in leaf peroxisomes by NaCl suggested that an in- creased diffusion of H202 into the cytosol could take place in leaf peroxisomes from NaCl-treated plants [23]. This may indicate that in chloroplasts, as in the case of peroxisomes and mitochondria, severe NaCl stress could induce the export of H,O, into the cytosol with the subsequent toxic oxidative reactions in this compartment.
The content of NaCl in leaves of the tolerant cultivar was lower than that of the sensitive one. This is an important factor, since a lower uptake might explain the lower damage in the tolerant cultivar. Chloride toxicity has been reported in many NaCl-sensitive plants including leguminous plants [40]. The level of Cl- in chloroplasts from the sensitive cultivar was only slightly increased despite the large increase in leaf Cl- contents, which suggests that the additional Cl- in the leaf was restricted to the cytoplasm. This has generally been demonstrated in the case of glycophyte plants [40]. On the contrary, the level of Na+ in chloro- plasts from sensitive plants was markedly increas- ed by NaCl and therefore, the slight replacement of K+ by Na+ observed in whole leaves was not apparent in chloroplasts. In fact, an increase in the Na+ concentration could be sufficient to affect chloroplastic SOD activity. In recent in vitro experiments carried out with cowpea protoplasts we demonstrated that Na+ rather than Cl- was an effective inhibitor of Mn-SOD and CuZn-SODS
1481. The splitting of the intact-chloroplasts into two
bands after centrifugation in discontinuous Percoll density gradients has also been reported in Pisum sativum [50], Vigna unguiculata and Phaseolus vulgaris [51]. This fact has been suggested to be probably an artefact of the ‘turnover effect’ at den-

J.A. Hermindez et 01. /Plant Science 105 (1995) 151-167 165
sity discontinuities [50]. However, morphometric results of the two chloroplast populations have shown differences in their starch grain contents as well as in the higher diameters, these unlike the starch grains being higher in the lower-density than in the higher-density chloroplasts. Thus, the distinct densities shown by the two chloroplast populations could be related to these mor- phometric differences.
As to the effects of NaCl, it was observed that the enhancement of the antioxidant defense system in chloroplasts of tolerant plants did not prevent cytomorphological damage. The NaCl-induced cytomorphological alterations in chloroplasts were apparently similar in tolerant and sensitive plants. However, results of chloroplast integrity indicate some differences in damage, at the level of the ex- ternal chloroplast membrane. Its integrity was lower in sensitive than in tolerant plants. These degenerative changes of chloroplasts induced by NaCl are reminiscent of those produced by iron and manganese deficiencies [52], and agree with ultrastructural results described in NaCl-treated Oryza plants [53] and Euglena gracilis [54].
An increase in the number and size of chloro- plast plastoglobuli was produced by NaCl in both pea plants although it was more intense in the tol- erant cultivar. This response has also been reported by Nazarenko and Serebryakova [54]. Plastoglobuli are ubiquitous in both chloroplasts and chromoplasts, and a probable role in salt tol- erance for these has been proposed [55].
The results described in this work indicate that in the cellular toxicity of NaCl in pea plants, chlo- roplasts have a role in an oxidative stress mecha- nism mediated by 02: and/or HzOz, similar to that found for mitochondria and, to a lesser ex- tent, for peroxisomes. As to the molecular mecha- nism of plant tolerance to salt stress, CuZn-SOD and ascorbate peroxidase appear to have a role in preventing NaCl-induced oxidative stress in chlo- roplasts.
Acknowledgements
The authors thank Dr M. Gomez Ortega (E.E.Z.) for his valuable help in the selection of pea plants with different sensitivities to NaCl. This
work was supported by grant PB87-0404 from the DGICYT and the Junta de Andalucia (research group #33 15), Spain. J.A. Hemandez is grateful to the Instituto de Fomento, Comunidad Autonoma de Murcia, Spain, for a research fellowship. L.A. de1 Rio acknowledges financial support from the British Council.
References
VI
I21
I31
141
151
(61
I71
181
[91
[lOI
1111
1121
1131
M.L. Sahn, Chloroplasts and mitochondrial mechanisms for protection against oxygen toxicity. Free Radical Res. Commun., 12-13 (1991) 851-858. P.R. Rich and W.D. Jr. Bonner, The sites of superoxide anion generation in higher plant mitochondria. Arch. B&hem. Biophys., 188 (1978) 206-213. L.A. de1 Rio, F. Sevilla, L.M. Sandalio and J.M. Palma, Nutritional effect and expression of superoxide dis- mutases: induction and gene expression; diagnostics; pro- spective protection against oxygen toxicity. Free Radical Res. Commun., 12-13 (1991) 819-828. K. Asada and M. Takahashi, Production and scavenging of active oxygen in photosynthesis, in: D.J. Kyle, C.B. Osmond and C.J. Arntzen (Eds.), Photoinhibition, Elsevier Science, Amsterdam, 1987, pp. 227-287. 1. Fridovich, Superoxide dismutases. Progr. Nucleic Acid Res., 40 (1991) 220-253. E.F. Eltsner, Mechanisms of oxygen activation in dif- ferent compartment of plant cells, in: E.J. Pell and F.L. Steffen (Eds.), Active oxygen/oxidative stress and plant metabolism, American Society of Plant Physiology, Rockville, MD, ISBN: O-943088-22-4. B. Halliwell and J.M.C. Gutteridge, Free Radicals in Biology and Medicine, Oxford University Press, London, 1986. K. Asada, Ascorbate peroxidase - a hydrogen peroxide -scavenging enzyme in plants. Physiol. Plant., 85 (1992) 235-241. H.D. Rabinowitch and I. Fridovich, Superoxide radicals, superoxide dismutase and oxygen toxicity in plants. Photochem. Photobiol., 37 (1983) 679-690. F. Sevilla, E. Hellin, C.F. Alcaraz and L.A. de1 Rio, In- duction of iron and manganese superoxide dismutases in Citrus limonum leaves by iron. Life Sci. Adv., Ser., B, 6 (1987) 133-139. J.M. Palma, M. Garrido, M.I. Rodriguez-Garcia and L.A. de1 Rio, Peroxisome proliferation and oxidative stress mediated by activated oxygen species in plant peroxisomes. Arch. B&hem. Biophys., 287 (1991) 68-74. A.S. Gupta, R.P. Webb, A.S. Holaday and R.D. Allen, Overexpression of superoxide dismutase protects plants from oxidative stress induction of ascorbate peroxidase in superoxide dismutase-overexpressing plants. Plant Physiol., 103 (1993) 1067-1073. M.C. Ball and J.M. Anderson, Sensitivity of

166 J.A. Herncindez et al. I Plant Science 10s (1995) 151-167
photosystem II to NaCl in relation to salinity tolerance. Comparative studies with thylakoids of the salt-tolerant mangrove, Avicennia marina, and the salt-sensitive pea, Pisurn sativum. Aust. J. Plant Physiol., 13 (1986) 689-698.
[l4] J.M. Cheeseman, Mechanisms of salinity tolerance in plants. Plant Physiol., 87 (1988) 547-550.
[l5] S. Brown, D.A. Day and C. Critchley, Salt-tolerance - does leaf respiration have a contribution to make? in: A.L. Moore and R.B. Bechey (Eds.), Plant Mitochon- dria: Structural, Functional and Physiological Aspects, Plenum Press, New York, 1987, pp. 383-396.
[I61 Y. Jolivet. J.C. Pireaux and P. Dizengremel, Changes in properties of barley leaf mitochondria isolated from NaCl-treated plants. Plant Physiol., 94 (1990) 641-646.
1171 H. Lambers, Respiration in intact plant and tissues. Its regulation and dependence on environmental factors, metabolism and invaded organisms, in: R. Douce and D.A. Day (Eds.), Encyclopedia of Plant Physiology, Vol. 18, Springer-Verlag, Berlin, 1985, pp, 418-473.
[IS] L.A. del Rio, F. Sevilla, M. Gomez, J. Yaiez and J. Lopez-gorge, Superoxide dismutase: an enzyme system for the study of micronutrient interactions in plants. Planta, I40 (1978) 221-225.
1191 L.A. del Rio, D.S. Lyon, I. Olah, B. Glick and M.L. Salin, Immunocytochemical evidence for a peroxisomal localization of manganese superoxide dismutase in leaf protoplasts from a higher plant. Planta, I58 (1983) 216-224.
[ZO] L.M. Sandalio, J.M. Palma and L.A. del Rio, Localiza- tion of manganese superoxide dismutase in peroxisomes isolated from Pisum sativwn L. Plant Sci., 51 (1987) 1-8.
[2l] J.M. Palma, L.M. Sandalio and L.A. del Rio, Manganese superoxide dismutase and higher plant chloroplasts: a reappraisal of a controverted cellular localization. J. Plant Physiol., I25 (1986) 427-439.
[22] J.A. Hemindez, F.J. Corpas, M. Gomez, L.A. del Rio and F. Sevilla, Salt-induced oxidative stress mediated by activated oxygen species in pea leaf mitochondria. Physiol. Plant., 89 (1993) 103-l IO.
1231 F.J. Corpas, M. Gomez, J.A. Hemindez and L.A. del Rio, Metabolism of activated oxygen in leaf peroxisomes from two Pisum sativwn L. cultivars with different sen- sivity to sodium chloride. J. Plant Physiol., I41 (1993) 160-165.
[24] L.A. del Rio, L.M. Sandalio, J. YaGez. and M. Gomez, Induction of a manganese-containing superoxide dismutase in leaves of Pisum sativwn L. by high nutrient levels of zinc and manganese. J. Inorg. Biochem., 24 (1985) 25-34.
[ZS] J.M.McCord and I. Fridovich, Superoxide dismutase: an enzymic function for erythrocuprein. J. Biol. Chem., 244 (1969) 6049-6055.
[26] B.J. Davies, Disc gel electrophoresis. Ann. N. Y. Acad. Sci. I21 (1964) 404-427.
[27] R.A. Weiss&r and I. Fridovich, Superoxide dismutase: organelle specificity. J. Biol. Chem., 248 (1973) 3582-3592.
1281 M.A. Hossain and K. Asada, Inactivation of ascorbate peroxidase in spinach chloroplasts on dark addition of hydrogen peroxide: its protection by ascorbate. Plant Cell Physiol., 25 (1984) 1285-1295.
[29] J.A. Buege and S.D. Aust, Microsomal lipid peroxida- tion. Methods Enzymol., 52 (1978) 302-310.
[30] J.E. Frew, P. Jones and G. Scholes, Spectrophotometric determinations of hydrogen peroxide and organic hydro- peroxides at low concentrations in aqueous solutions. Anal. Chim. Acta, I55 (1983) 139-150.
1311 M.Y. Law, S.A. Charles and B. Halliwell, Glutathione and ascorbic acid in spinach (Spinacea oleruceu) chloro- plasts. The effect of hydrogen peroxide and paraquat. Biochem. J. 210 (1983) 899-903.
[32] K. Tanaka, R. Masuda, T. Sugimoto, K. Omasa and T. Sakaki, Water deficiency-induced changes in the contents of defensive substances against active oxygen in spinach leaves. Agric. Biol. Chem., 54 (1990) 2629-2634.
[33] R.McC. Lilley, M.P. Fitzgerald, K.G. Rienits and D.A. Walker, Criteria of intactness and the photosynthetic ac- tivity of spinach chloroplast preparations. New Phytol., 75 (1975) I-IO.
[34] M.M. Bradford, A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Anal. Biochem., 72 (1976) 248-254.
[35] D.I. Amon, Copper enzymes in isolated chloroplasts. Polyphenol oxidase in Eeru vulgaris. Plant Physiol.. 24 (1949) l-15.
(361 H.D. Chapman and P.F. Pratt, Methods of Analysis for Soil, Plants and Water. Division of Agriculture, Univer- sity of California (Riverside). USA, 1961.
[37] G. Schr6pper-Meier and M. Kaiser, Ion homeostasis in chloroplasts under salinity and mineral deficiency. I. Sol- ute concentration in leaves and chloroplasts from spinach plants under NaCl or NaN03 salinity. Plant Physiol., 87 (1988) 822-827.
138) J. Didier de St-Amand, S. Gas and H. Lefranc, Methodes de dosages effectties par autoanalyse au laboratorie de diagnostic foliare de L’Orstom. Cah. Orstom, Dot. Tech. no. 9, 1968.
[39] A.R. Spurr, A low viscosity epoxy resin embedding medi- um for electron microscopy. J. Ultrastruct. Res., 26 (1969) 31-43.
[40] J.R. Seeman and Ch. Critchley, Effects of salt stress on the growth, ion content, stomata1 behaviour and photo- synthetic capacity of a salt-sensitive species, Phaseoh
vulgaris L. Planta, 164 (1985) 151-162. 141) A.H. Price, N.M. Atherton and G.A.F. Hendry, Plants
underdrought-stress generate activated oxygen. Free Radical Res. Commun., 8 (1989) 61-66.
[42] B. Halliwell, The toxic effects of oxygen on plant tissues, in: L.W. Oberley (Ed.), Superoxide Dismutase, Vol. I, CRC Press, Boca Raton, FL, 1982, pp. 89-123.
[43] S. Singha and M.A. Choudhuri, Effect of salinity (NaCI) on H2G2 mechanism in Vigna and Oryza seedlings. Biochem. Physiol. Pflanz., I86 (1990) 69-74.
[44] G. Pastori and VS. Trippi, Antioxidative protection in a

J.A. Hernrinde: et al. /Plant Science 105 (1995) 151-167 167
drought-resistant maize during leaf senescence. Physiol. Plant., 87 (1993) 227-231.
[45] C. Bowler, L. Slooten, S. Vandenbrauden, R. de Rycke, J. Botterman, G. Sybesma, M. van Montagu and D. Inze, Manganese superoxide dismutase can reduce cellular damage mediated by oxygen radicals in transgenic plants. EMBO J., 10 (1991) 1723-1732.
[46] R. Mittler and B.A. Zilinskas, Regulation of pea cytosolic ascorbate peroxidase and other antioxidant en- zymes during the progression of drought stress and following recovery from drought. Plant J., 5 (1994) 397-405.
1471 E. Olmos, J.A. Hernandez, F. Sevilla and E. Hellin, In- duction of several antioxidant enzymes in the selection of a salt-tolerant cell line of Pisum sativum. J. Plant Physiol., 44 (1994) 594-598.
(481 J.A. Hemandez, L.A. del Rio and F. Sevilla, Salt stress- induced changes in superoxide dismutase isozymes in leaves and mesophyll protoplasts from Vigna unguiculata (L.) Walp. New Phytol., 126 (1994) 37-44.
[49] I. Fridovich, Biological effects of the superoxide radical. Arch. B&hem. Biophys., 247 (1986) I-11.
[50] D.M. Mousdale and J.R. Coggins, Subcellular localiza-
tion of the common shikimate enzymes in Pisum sativum
L. Planta, 163 (1985) 241-249. [51] F.J. Corpas, L.M. Sandalio, J.M. Palma, E.O. Leidi, J.A.
Hernindez, F. Sevilla and del Rio, Subcellular distribu- tion of superoxide dismutase in leaves of ureide- producing leguminous plants. Physiol. Plant., 8 I (I 99 I ) 285-291.
[52] Ch. Hecht-Buchholz, Light and electron microscopic in- vestigations of the reaction of various genotypes to nutri- tional disorders. Plant Soil, 72 (I 983) I5 I - 165.
[53] T.J. Flowers, E. Duque, M.A. Hajibagheri, T.P. McGlonigle and A.R. Yeo, The effect of salinity on leaf ultrastructure and net photosynthesis of two varieties of rice: further evidence for a cellular component of salt- resistance. New Phytol., 100 (1985) 37-43.
1541 L.V. Nazarenko and V.N. Serebryakova, Ultrastructural changes in Euglena cells of different nutritional types under the influence of salinity. Fiziol. Rast., 37 (1990) 142-146.
[55] J. Burguess, An Introduction to Plant Cell Development, 1st Edn., Cambridge University Press, Cambridge, UK. 1985, pp. 42-47.




















