
Salt-induced oxidative stress mediated by activated oxygen species in pea leaf mitochondria
9
PHYSIOLOGIA PLANTARUM 89: 103-110. 1993 Primed in Denmark - all rights reserved Salt-induced oxidative stress mediated by activated oxygen species in pea leaf mitochondria Jose A. Hernandez, Francisco J. Corpas, Manuel Gomez, Luis A. del Rio and Francisca Sevilla Hernandez, J.A., Corpas, EJ., Gomez, M., del Rio, L. A. and Sevilla, E 1993. Salt-induced oxidative stress mediated by activated oxvgen species in pea leaf mi- tochondria. - Physiol. Plant. 89: 103-110. The effect in vivo of salt stress on the activated oxygen metabolism of mitochondria, was studied in leaves from two NaCI-treated cultivars of Pisum sativum L. with different sensitivity to NaCl. In mitochondria from NaCI-sensitive plants, salinity brought about a significant decrease of Mn-SOD (EC 1.15.1.1) Cu,Zn-SOD I (EC 1.15.1.1) and fumarase (EC 4.2.1.2) activities. Conversely, in salt-tolerant plants NaCl treatment produced an increase in the mitochondrial Mn-SOD activity and, to a lesser extent, in fumarase activity. In mitochondria from both salt-treated cultivars, the internal H^O, concentration remained unchanged. The NADH- and succinate- dependent generation of O\ radicals by submitochondrial particles and the lipid peroxidation of mitochondrial membranes, increased as a result of salt treatment, and these changes were higher in NaCl-sensitive than in NaCt-tolerant plants. Ac- cordingly, the enhanced rates of superoxide production by mitochondria from salt- sensitive plants were concomitant with a strong decrease in the mitochondrial Mn- SOD activity, whereas NaCl-tolerant plants appear to have a protection mechanism against salt-induced increased Oi" production by means of the induction of the mitochondrial Mn-SOD activity. These results indicate that in the subcellular toxicity of NaCl in pea plants, at.i"he level of mitochondria, an oxidative stress mechanism mediated by superoxide radicals is involved, and also imply a function for mi- tochondrial Mn-SOD in the molecular mechanisms of plant tolerance to NaCl. Key words - Activated oxygen, cytochrome c oxidase, fumarase, H^O,, mitochon- dria, pea. Pisum sativum. salt stress, superoxide dismutase, superoxide radicals. J.A. Hernandez and E. Sevilla (corresponding author), Unidad de Alimentacion v Desarrollo de la Planta, CEBAS, CSfC, Apdo f95, E-30080 Murcia, Spain; E J. Corpas, M. Gomez and L.A. del Rio, Unidad de Bioquimica Vegetal, Estacion Experimental del Zaidin, CSIC, Apdo 419, E-18080 Granada, Spain. oxide radicals (O; ; Rabinowitch and Fridovich 1983, Introduction Halliwell and Guttertdge 1989). In the mitochondrial Saline environments can induce a wide number of re- respiratory chain there are two sites of superoxide pro- sponses in plants ranging from readjustments of trans- duction which are located at the flavoprotein NADH port and metabolic processes to growth inhibition dehydrogenase and at the ubiquinone-cytochrome b (Lambers 1985). Under saline conditions, plant respira- segments (Rich and Bonner 1978. Boveris 1984). Super- tion undergoes different changes depending upon leaf oxide dismutases (SOD; EC 1.15.1.1) are a group of age (Brown et al. 1987, Jolivet et al. 1990), and detailed metalloenzymes that catalyze the disproportionation of knowledge of the respiratory metabolism during salt Oj" radicals to H^O, and O,, and play an important role stress conditions is still lacking. In plant cells, mitochon- in protecting cells against superoxide-derived oxidative dria are important intracellular generators of super- damages (Rabinowitch and Fridovich 1983, Fridovich Received 10 November, 1992; revised 1 April. 1993 Physiol. Plant. 89. 1993 103
Transcript of Salt-induced oxidative stress mediated by activated oxygen species in pea leaf mitochondria

PHYSIOLOGIA PLANTARUM 89: 103-110. 1993Primed in Denmark - all rights reserved
Salt-induced oxidative stress mediated by activated oxygen speciesin pea leaf mitochondria
Jose A. Hernandez, Francisco J. Corpas, Manuel Gomez, Luis A. del Rio and Francisca Sevilla
Hernandez, J.A., Corpas, EJ. , Gomez, M., del Rio, L. A. and Sevilla, E 1993.Salt-induced oxidative stress mediated by activated oxvgen species in pea leaf mi-tochondria. - Physiol. Plant. 89: 103-110.
The effect in vivo of salt stress on the activated oxygen metabolism of mitochondria,was studied in leaves from two NaCI-treated cultivars of Pisum sativum L. withdifferent sensitivity to NaCl. In mitochondria from NaCI-sensitive plants, salinitybrought about a significant decrease of Mn-SOD (EC 1.15.1.1) Cu,Zn-SOD I (EC1.15.1.1) and fumarase (EC 4.2.1.2) activities. Conversely, in salt-tolerant plantsNaCl treatment produced an increase in the mitochondrial Mn-SOD activity and, to alesser extent, in fumarase activity. In mitochondria from both salt-treated cultivars,the internal H^O, concentration remained unchanged. The NADH- and succinate-dependent generation of O\ radicals by submitochondrial particles and the lipidperoxidation of mitochondrial membranes, increased as a result of salt treatment,and these changes were higher in NaCl-sensitive than in NaCt-tolerant plants. Ac-cordingly, the enhanced rates of superoxide production by mitochondria from salt-sensitive plants were concomitant with a strong decrease in the mitochondrial Mn-SOD activity, whereas NaCl-tolerant plants appear to have a protection mechanismagainst salt-induced increased Oi" production by means of the induction of themitochondrial Mn-SOD activity. These results indicate that in the subcellular toxicityof NaCl in pea plants, at.i"he level of mitochondria, an oxidative stress mechanismmediated by superoxide radicals is involved, and also imply a function for mi-tochondrial Mn-SOD in the molecular mechanisms of plant tolerance to NaCl.
Key words - Activated oxygen, cytochrome c oxidase, fumarase, H^O,, mitochon-dria, pea. Pisum sativum. salt stress, superoxide dismutase, superoxide radicals.
J.A. Hernandez and E. Sevilla (corresponding author), Unidad de Alimentacion vDesarrollo de la Planta, CEBAS, CSfC, Apdo f95, E-30080 Murcia, Spain; E J.Corpas, M. Gomez and L.A. del Rio, Unidad de Bioquimica Vegetal, EstacionExperimental del Zaidin, CSIC, Apdo 419, E-18080 Granada, Spain.
oxide radicals (O; ; Rabinowitch and Fridovich 1983,Introduction Halliwell and Guttertdge 1989). In the mitochondrialSaline environments can induce a wide number of re- respiratory chain there are two sites of superoxide pro-sponses in plants ranging from readjustments of trans- duction which are located at the flavoprotein NADHport and metabolic processes to growth inhibition dehydrogenase and at the ubiquinone-cytochrome b(Lambers 1985). Under saline conditions, plant respira- segments (Rich and Bonner 1978. Boveris 1984). Super-tion undergoes different changes depending upon leaf oxide dismutases (SOD; EC 1.15.1.1) are a group ofage (Brown et al. 1987, Jolivet et al. 1990), and detailed metalloenzymes that catalyze the disproportionation ofknowledge of the respiratory metabolism during salt Oj" radicals to H^O, and O,, and play an important rolestress conditions is still lacking. In plant cells, mitochon- in protecting cells against superoxide-derived oxidativedria are important intracellular generators of super- damages (Rabinowitch and Fridovich 1983, Fridovich
Received 10 November, 1992; revised 1 April. 1993
Physiol. Plant. 89. 1993 103

1986). The production and/or participation of activatedoxygen species in different plant stress situations as wellas the induction of SOD isozymes by Oi -generatingmetals and environmental contaminants have been re-ported (Rabinowitch and Fridovich 1983, Sevilla et al.1987. del Rio et al. 1991. Palma et al. 1991). Some plantspecies have evolved biotypes which are tolerant to highlevels of xenobiotics and have increased SOD activity(Rabinowitch and Fridovich 1983). Recently, in leavesof Vigna and Oryza plants it has been reported thatNaCl decreased both SOD and catalase activities and.at the same time, the H^O^-producing glycolate oxidaseactivity was increased, suggesting that H^O, could playan important role in the mechanism of salt injury(Singha and Choudhud 1990). Similarly, drought condi-tions induced in wheat chloroplasts the formation ofO r radicals which could be responsible for the primarypathologies observed in drought-stressed plants (Priceet al. 1989).
The effect of salt stress on the rate of mitochondrialoxygen uptake and oxidative phosphorylation has beenstudied in mitochondria from pea and barley plants(Brown et al. 1987, Jolivet et al. 1990). However, thereis no information on the activated oxygen metabolism ofmitochondria during saline stress of plants. Informationregarding endogenous antioxidant enzymes and produc-tion of oxyradicals in salt-stressed mitochondria isnecessary to understand important processes takingplace in these organelles, such as lipid peroxidation.redox changes in NADH. and activated oxygen-derivedprotein fragmentation (Fridovich 1986. Marcillat et al.1988). Likewise, this knowledge could allow deeperinsights into the molecular mechanisms of plant toler-ance to salt-induced oxidative stress.
Pea leaves contain three electrophoretically distinctSODs. a Mn-containing SOD and two Cu.Zn-contain-ing SODs (I and 11; del Rio et al. 1985). By studies ofsubcellular distribution of SODs in pea leaves the pres-ence of Mn-SOD and Cu.Zn-SOD I has been demon-strated in mitochondria (Sandalio et al. 1987). In thepresent work, the existence of a relationship betweenNaCl toxicity and oxidative stress mediated by activatedoxygen species was studied using leaf mitochondria pu-rified from two NaCl-treated cultivars of Pisum sativumL. having different sensitivities to NaCl.
Abbreviations - BSA. bovine serum albumin; Cu.Zn-SOD,Cu.Zn-containing superoxide dismutase; Cyt c oxidase. cyto-chrome c oxidase; Mn-SOD, Mn-containing superoxide dismu-tase; NBT. nitroblue tetrazolium; MDA. malondialdehyde;SHAM, salicylhydroxamic acid; SMP, submitochondrial par-ticle; SOD, superoxide dismutase.
Materiais and methods
Plant material
Seeds of different commercial cultivars of pea {Pisumsativum L.). obtained from Royal Sluis (Enkhuizen,The Netherlands), were surface-sterilized (ethanol, 3
min) and germinated and grown in vermiculite with 0.5mM CaSO4 for 3 days. Healthy and vigorous seedlingswere selected and grown in a growth chamber (Convi-ron PGW-36) in nutrient solutions containing differentconcentrations of NaCl (Merck). First, plants were cul-tivated in aerated optimum nutdent solutions (del Rioet al. 1985) for 7 days. The growth chamber was set to28/18°C, 80% RH, and 16-h photopedod. Light (203\imol m- s ' PAR) was supplied by cool-white fluo-rescent tubes (Sylvania F96T12VHO/CW) and incan-descent lamps (Philips 40 W Arlite). After 7 days, plantswere transplanted to similar media supplemented withNaCl concentrations of 30-300 mM. and were grown for14 days. Pots containing 4 plants in 2.5 1 nutrientsolution were arranged in blocks at random, and eachtreatment (NaCl level and cultivar) was replicated 4times. All the experiments were repeated 3 times ondifferent days. Concentrations of NaCl and micronu-trients in nutrient media were determined potentio-metdcally and by atomic absorption spectrometry. re-spectively. Plants were harvested and plant growth wasdetermined by measuring fresh weight of leaves, freshweight of shoots, foliar area (in cm-), dry and freshweight of roots. Statistical analysis of results from the 3different experiments was carried out according to Dun-can's multiple range test.
Purification of mitochondria
All operations were performed at (>-4°C. In leaves ofeach pea cultivar selected, mitochondria were purifiedby differential and density-gradient centrifugation indiscontinuous gradients of Percoll (15-53%. v/v) as de-scribed by Sandalio et al. (1987). After centrifugation,gradients were fractionated by upward displacementwith 45% sucrose (w/w) using an Isco fractionator.model 185, equipped with an optical unit and an ab-sorbance detector. The transmittance of eluted sampleswas monitored at 280 nm and enzyme activities weremeasured in the different fractions collected (1.5 ml).The integrity of the outer mitochondrial membrane wasestimated from the succinate;cytochrome c oxidoreduc-tase (EC 1.3.99.1) activity in isotonic and hypotonicmedium (Douce et al. 1972).
SMPs were prepared essentially as described by Nohland Hegner (1978), Suspensions of mitochondria (5 ml)were sonicated at 0°C for 1 min, with pulses of 3 s, in0,05 M Tds-HCl, pH 7,8, containing 0.23 M mannitoland 0.07 M sucrose, according to Boveds (1984). Thesuspension was centrifuged at 12000 g for 15 min andthe precipitate was discarded. The SMPs were sedi-mented by centrifugation at 114000 g for 30 min. andwere then washed once with the same buffer. The su-pernatants obtained were used for the determination ofSOD activity. The SMPs were resuspended in a freshlyprepared 0.25 M sucrose solution and used immediatelyfor the assay of superoxide production. By followingthis procedure the possible interferences of peroxisomal
104 Physiol. Plant. 89. 1993

• 1 LSD 0001
1mM NaCl GR CH
I 150 70
Na Cl, mM210
Eig. 1. Plant growth of a sensitive (hatched columns) and amoderately NaCl-tolerant (filled columns) cultivar of Pisumsativum L. expressed as fresh weight of leaves after a growthperiod of 14 days in tiutrient solutions containing a range ofNaCl concentrations. Columns represent the average of threeindependent experiments ± SE, as described in Materials andmethods. Bars indicate the LSD (P <O.(K)1) for each combina-tion of NaCl concentration and cultivar, aecording to Duncan'smultiple range test. GR, tolerant cultivar Granada: CH, sensi-tive cultivar Challis.
soluble activities on the subsequent assays with SMPswere negligible. On the basis of catalase activity valuesdetermined in mitochondrial suspensions and in SMPs,the peroxisomal contamination of SMPs was about 2%.
Enzyme assays
Total SOD (EC 1.15.1.1) activity of samples was deter-mined by its ability to inhibit the reduction of ferricyto-chrome c by superoxide radicals generated by xanthine/xanthine oxidase, as described by McCord and Fri-dovich (1969). SOD isozymes (Cu,Zn-SODs and Mn-SOD) were separated by polyacrylamide cylindric gelelectrophoresis on 10% gels, at pH 8.9, according toDavis (1964). Prior to electrophoresis, organelle sam-ples were diluted 2-5 fold with 50 mM potasium phos-phate, pH 7.8, containing Triton X-l()0(0.r/o final con-centration) and stirred to solubilize bound superoxidedismutases. SODs were localized on the gels by themethod of the NBT reduction by O r radicals generatedphotochemically (Weisiger and Fridovich 1973). Theisozyme activity was quantitated by recording the trans-
mittance of the gels in a Shimadzu CS-9000 densitom-eter and integrating the activity areas under the trans-mittance peaks. Cyt c oxidase (EC 1.9.3.1) was deter-mined as described by Schnarrenberger et al. (1971).Fumarase (EC 4.2.1.2) was assayed according to Walkand Hock (1977).
Electron microscopy
Samples of leaf tissue from plants treated and nottreated with NaCl (controls) were fixed and processedfor electron microscopy as described by Palma et al.(1991), and were examined with a Zeiss EM 10°C elec-tron microscope at 60 kV.
Assays
Superoxide radical generation by SMPs was determinedby the method of the superoxide dismutase-inhibitableoxidation of epinephrine to adrenochrome usingNADH and succinate as respiratory substrates, accord-ing to Boveris (1984). The assays were carried out at25°C and the reaction mixture (3 ml) contained 20 mMTris-HCI. pH 7.0, air-saturated; 1 mM epinephrine;0.23 M mannitol; 0.07 M sucrose; and SMPs (8-10 ngprotein). The reaction was started by adding eitherNADH or succinate and was followed at 480 nm versusa mixture with identical composition but containing 1(jtM Cu.Zn-SOD from bovine erythrocytes (SigmaChemical Co.). Rotenone (1 ^M) and antimycin (1 \iM)were used as specific inhibitors to assay NADH- andsiTccinate-dependent generation of O^' radicals, respec-tively (Boveris 1984). The endogenous phenol oxidaseof mitochondria was inhibited by using either 1 mMKCN or 1 mM SHAM (Rich and Bonner 1978). Theamount of O r radicals produced was calculated as de-scribed by Boveris (1984) using for epinephrine an e4x,iof 4.0 mM ' cm ' (Green et al. 1956).
Lipid peroxidation of mitochondrial membranes wasestimated by determining the concentration of malon-dialdehyde with thiobarbituric acid, according to Buegeand Aust (1978). The concentration of H^O, in purifiedintact mitochondria was determined by a peroxidase-coupled assay using 4-aminoantipyrine and phenol as
Tab. 1. Nutrient and chlorophyll contents in leaves of control and NaCl-treated pea plants. Differences from control values weresignificant at: P <0.()()l (A); 0.001 < P <0.01 (B); 0.01 < P <0.05 (C); NS, not significant. Mean values ± SEM of at least 6different leaf extracts each of control and NaCl-treated plants.
Cultivarand treatment
Content, mmol (g DW)
Na K Ca Mg Cl
Chlorophyllmg(gEwV'
NaCl-tolcrantControl70 mM NaCl
NaCl-sensitiveControl70 mM NaCl
0.15 ±0.04Ll l+0.09 A
0.12 + 0.031.43 ±0.02 A
1.22±0.t)81.09 ±0.03 NS
1.25 ±0.051.10 ±0.03 B
0.41 ±0.030.33 ± 0.01 NS
0.38 ±0.010.30 ± 0.02 C
0.26 ± O.(X)80.20 ±0.003 B
0.24 ±0.010.16 ±0.02 C
0.12 ±0.010.90±0.t)2 A
0.19±0.0I1.82 ±0.03 A
3.57 ±0.582.17 ± 0.24 C
3.06 ±0.31
Physiol. Plant. 89. 1993 105
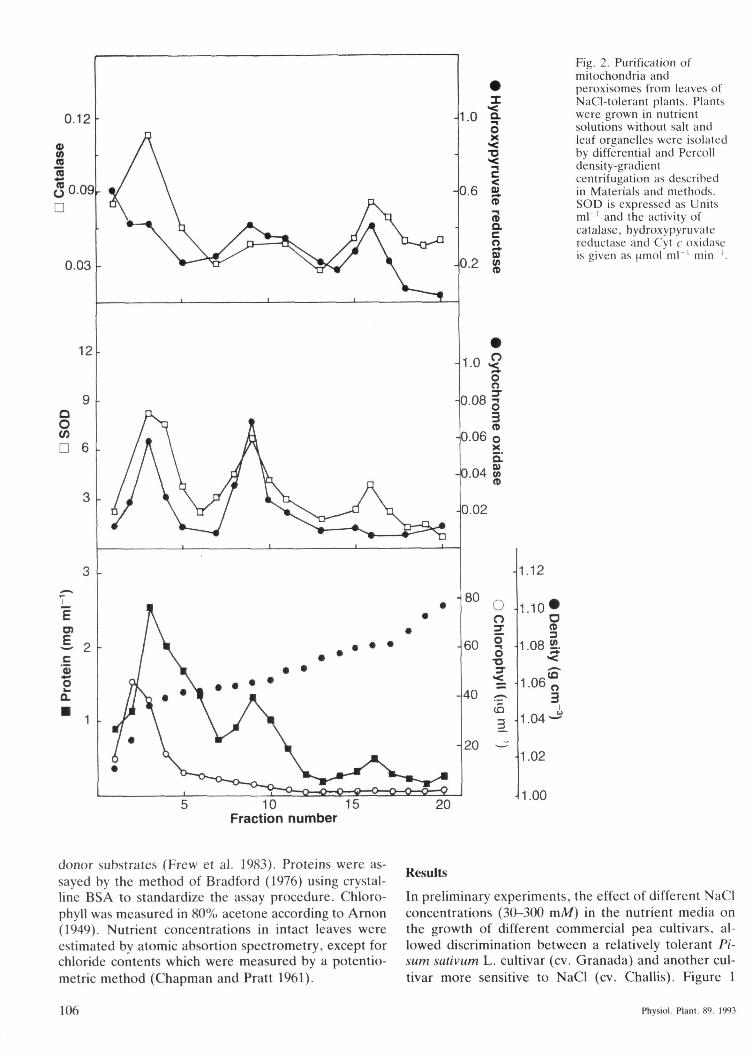
Eig, 2, Purification ofmitochondria andperoxisomes from leaves ofNaCl-tolerant plants. Plantswere grown in nutrientsolutions without salt andleaf organelles were isolatedby differential and Percolldensity-gradientcentrifugation as describedin Materials and methods,SOD is expressed as Unitsm l ' and the activity ofcatalase, hydroxypyruvatereductase and Cyt c oxidaseis given as (imol m l ' min ',
10 15Fraction number
1,00
donor substrates (Frew et a), 1983), Proteins were as-sayed by the method of Bradford (1976) using crystal-line BSA to standardize the assay procedure. Chloro-phyll was measured in 80% acetone according to Arnon(11949). Nutrient concentrations in intact leaves wereestimated by atomic absortion spectrometry, except forchloride contents which were measured by a potentio-metric method (Chapman and Pratt 1961).
Results
In preliminary experiments, the effect of different NaClconcentrations (30-300 mM) in the nutrient media onthe growth of different commercial pea cultivars, al-lowed discrimination between a relatively tolerant Pi-sum sativum L, cultivar (cv, Granada) and another cul-tivar more sensitive to NaCl (cv, Challis). Figure 1
106 Physiol, Plant, 89, 1993

Tab. 2. Enzymatie activities in leaf mitochondda of control- and NaCl-treated plants. For the determination of enzyme activities,mitochondria were broken by osmotic shock. Mean values ± SEM from at least 8 separate mitochondria preparations each ofcontrol and NaCl-treated plants are shown. Differences from control values were significant at; P <O.(X)1 (A); 0.001 < P <0 01(B); 0.01 < P <0.05 (C); NS. not significant.
Cultivar
NaCl-tolerantControl70 mM NaCl
NaCl-sensitiveControl70 mM NaCl
Activity.
Mn-SOD
132 ±8226 ± 18 B
208 ±7136 ±7 A
U (mg protein) '
Cu.Zn-SOD 1
24.9± 1.529.2 ± 2.4 NS
45.3 ±1.514.5 ± 0.8 A
Activity, nmol
Cyt c oxidase
595 ±16612 ±22 NS
455 ± 43338 ± 24 NS
(mg protein) ' min '
Fumarase
287 ±14478 ± 32 C
236 ±8130 ±7 C
shows the effect of different NaCl concentrations on thegrowth of the two Pisum sativum cultivars. At 70 mMNaCl. the growth of cv. Granada was only slightly af-fected, whereas growth of cv. Challis was significantlyinhibited. After treatment with 70 mM NaCl. the foliarconcentration of Na* and C\ increased in the two culti-vars, but the uptake of both nutrients from the solutionswas higher in cv. Challis than in cv. Granada (Tab. 1).Coinciding with these increases, there was a loss ofchlorophyll in both salt-treated cultivars, the chloro-phyll concentration being significantly lower in NaCl-sensitive than in NaCl-tolerant plants. (Tab. 1).
A purification procedure of leaf mitochondria fromNaCl-tolerant plants (cv. Granada), representative ofdifferent independent experiments carded out, is shownin Fig. 2. As indicated by the Cyt c oxidase activity,mitochondria were located in the density-gradient infractions 8 to 10. at an average equilibrium density of1.062 g cm"\ which is nearly identical to the densityreported for mitochondria isolated from spinach andpea leaves using the same method (Schwitzguebel andSiegenthaler 1984. Sandalio et al. 1987). Salt treatmentdid not modify the equilibrium density values obtained
Tab. 3. Superoxide production in SMPs from plants treatedwith NaCl. SMPs were prepared from purified intact mitochon-dria as described in Materials and methods. NADH (50 \iM)and succinate (7 mM) were used as substrates to assay Oiproduction. Reactions mixtures contained 10 [xg of mitochon-drial membranes. Mean values ± SEM of at least 6 differentmitochondria preparations each of control and NaCl-treatedplants, are presented. Differences from control values weresignificant at P <0.001 (A).
Cultivarand treatment
NaCl-tolerantControl70 mM NaCl
NaCl-sensitiveControl70 mM NaCl
Physiol. Plant. 89, 1993
O: \ nmol (mg
NADH-dependentgeneration
2.36 ±0.135.57 ±0.22 A
3.57 ±0.147.96 ±0.22 A
protein) ' min '
Succinate-dependentgeneration
3.14±0.195.]0±0.16 A
2.61 ±0.215.77 ±0.21 A
for mitochondria isolated from NaCl-sensitive andNaCl-tolerant plants. Peroxisomes were well separatedfrom mitochondria and banded in fraction 15 to 18 at anaverage density of about 1.092 g cm \ characteristic ofthese intact organelles in Percoil solutions (Schwitz-guebel and Siegenthaler 1984. Sandalio et al. 1987).Similar results were obtained when leaves of NaCl-sensitive plants were used.
Mitochondria were sometimes contaminated withperoxisomal activity. The recovery of catalase activity inmitochondrial preparations was about 15%. This con-tamination could probably derive from partially brokenperoxisomes co-eluting with the mitochondrial fraction(;Schmitt and Edwards 1982. Sandalio et al. 1987) butdid not interfere with any of the determinations cardedout with either intact mitochondria or SMPs.
Under normal growth conditions, the activity of Mn-SOD and Cu,Zn-SOD I isozymes were higher in mi-tochondria from NaCl-sensitive plants (cv. Challis) thanin those from NaCl-tolerant plants (cv. Granada; Tab.2). In contrast, the activity of Cyt c oxidase andfumarase were slightly higher in the NaCl-tolerant culti-var (Tab. 2). The intactness of leaf mitochondria puri-fied from salt-tolerant and salt-sensitive plants was notaffected by NaCl and was in most cases 80%.
Treatment of plants with NaCl affected total super-oxide dismutase (Mn-SOD plus Cu,Zn-SOD I) andfumarase activities of leaf mitochondria differently de-pending on the pea cultivar. Mn-SOD was increased byNaCl in the tolerant cultivar and Cu,Zn-SOD I did notshow statistically significant variations, whereas in thesensitive cultivar NaCl produced a significant inhibitionof Mn-SOD and Cu.Zn-SOD I activities. Fumarase ac-tivity was also increased by salinity in mitochondria ofNaCl-tolerant plants, whereas in NaCl-sensitive plantsthis activity was inhibited by salt treatment. However,in leaf mitochondda from both NaCl-tolerant and NaCl-sensitive plants, no significant changes were detected inthe activity of Cyt c oxidase.
In SMPs, the NADH-induced production of O r radi-cals was doubled in both pea cultivars as a result of salttreatment. Similar results were obtained when succinate
107

Tab. 4. Lipid peroxidation and hydrogen peroxide concentra-tion in mitochondria of control and NaCl-treated plants. Lipidperoxidation and hydrogen peroxide concentration were deter-mined in intact mitochondria. Mean values ± SEM of at least sixdifferent mitochondria preparations each of control and NaCl-treated plants are presented. Differences from control valueswere significant at: P < 0.01 (A): O.OK /> < O.t)5 (B): NS, notsignificant.
Cultivarand treatment
MDA, pmol(mg protein) '
H,O:, nmol(mg protein)"'
NaCl-tolerantControl70 mM NaCl
NaCI-.sensitiveControl70 mM NaCl
513 ±20716 ±55 B
383 ± 23920 ± 62 A
1.60 ±0.131.41 ±().t)4NS
0.99 ± 0.051.05 ± 0.08 NS
was used as respiratory inducer of O; generation (Tab.3). However, the effect of salt on the succinate-de-pendent O r production by SMPs was comparativelyhigher in NaCl-sensitive than in NaCl-tolerant plantsand this difference was statistically significant accordingto Duncan's multiple range test (0.01 < P <0.05; Tab.3).
Salinity apparently did not change the mitochondrialconcentration of H^O,, which was very similar to thecontrols in NaCl-tolerant and in NaCl-sensitive plants(Tab. 4). In contrast, lipid peroxidation of membranes,measured as the formation of malondialdehyde, wasincreased in mitochondria from plants treated withNaCl, and lipid peroxidation was greater in NaCl-sensi-tive than in NaCl-tolerant plants (Tab. 4).
The cytomorphology of leaves from plants grownwith NaCl and leaves from control plants, was studiedby electron microscopy. NaCl treatment produced al-terations in the ultrastructure of leaf cells particularly inthe membranes and thylakoidal structure of chloro-plasts, which were notably disorganized. The ultrastruc-ture of mitochondria was not greatly affected by NaCland the only change observed was that NaCl-sensitiveplants treated with salt had mitochondria with a reducednumber of cristae and a lower electron density com-pared to NaCl-tolerant plants (data not shown).
Discussion
In NaCl-sensitive plants, the addition of 70 mM NaCl tothe nutrient solutions resulted in a considerable de-crease in the fresh weight of leaves (Fig. 1), leaf area(not shown) and chlorophyll content (Tab. 1), and theoldest leaflets started to develop necrotic margins 14days after sait addition. The reduction in leaf chloro-phyll concentration is often observed as a result of saltstress in sensitive plants (Seeman and Critchley 1985).In both salt-treated cultivars, leaf Na* and C r concen-trations were increased. However, NaCl-tolerant plantswere relatively more efficient Cl -excluders than theNaCl-sensitive plants.
The results reported here show that growth of twoFisum sativum cultivars with salt, induced different re-sponses in the specific activities of intra-mitochondrialenzymes involved in the oxygen metabolism, and thatthese responses were related to the distinct susceptibil-ity of both cultivars to NaCl. In mitochondria of NaCl-sensitive plants, salinity brought about a significant de-crease of Mn-SOD, Cu,Zn-SOD I and fumarase activi-ties. However, in mitochondria of NaCl-tolerant plantssalt treatment induced an increase in the activities ofisozyme Mn-SOD and fumarase. The only mitp^hon-drial activity of both cultivars that did not show sig-nificant variations with salt was Cyt c oxidase. Thestimulation by NaCl of fumarase activity in mitochon-dria from salt-tolerant plants seems to be an indicationthat the tricarboxylic acid cycle is affected by salinity.
In plants resistant to different types of environmentalstress conditions, increased levels of enzymes involvedin superoxide detoxification have been found (Rabino-witch and Fridovich 1983, del Rio et al. 1991). Over-expression of Mn-SOD in mitochondria and chloro-plasts from transgenic tobacco plants can reduce cellulardamage mediated by oxygen radicals (Bowler et al.1991). In pea plants, clofibrate induces peroxisome pro-liferation and oxidative stress mediated by activatedoxygen species in peroxisomes, and these effects occurconcomitant with a decrease in the specific activities ofcatalase and Mn-SOD, the main peroxisomal defensesagainst W.O, and O r (Palma et al. 1991). All theseresults are in good agreement with those of the presentwork on the response of mitochondrial SOD isozymesto salt stress. The higher activity of Mn-SOD found intolerant plants under salt-stress, suggests that mitochon-dria, and Mn-SOD in particular, might have a certainrole in the molecular mechanisms responsible for planttolerance to NaCl.
Isolated plant mitochondria, unlike animal mitochon-dria, contain a rotenone-insensitive NAD(P)H dehy-drogenase on the outer surface of the inner membranewhich oxidises external NAD(P)H in a Ca-+-dependentmanner (M0l]er and Palmer 1982, M0ller and Lin1986). This rotenone-insensitive pathway is differentfrom the rotenone-resistant NADH-dehydrogenase onthe matrix side of the inner membrane that does notconserve energy (M0ller and Palmer 1982). In SMPsfrom the two pea cultivars, salt treatment stimulatedboth the NADH- and succinate-dependent generationof Oj" radicals. As a result of salt stress, O^' generationrose 2-3 times in mitochondria from NaCl-sensitiveplants, and in these conditions a significant decrease inMn-SOD and Cu,Zn-SOD I activities took place. Thisindicates that in salt-sensitive plants, contrary to salt-tolerant plants, the ability of mitochondria to removeO r radicals is diminished under conditions of enhancedproduction of these activated oxygen species due tosalinity stress. This could also explain the higher rates oflipid peroxidation found in mitochondrial membranesfrom NaCl-sensitive plants. Sjolin and M0ller (1991)
108 Physiol. Plant. 89, i993

have demonstrated that salts stimulate electron trans-port in SMPs from potato through electrostatic chargescreening and this could be the reason for the enhancedproduction of Q; radicals observed in pea leaf SMPsfrom plants grown with NaCl.
In Jerusalem artichoke tuber and in potato tuber, theisolation of SMPs of well defined polarities, right-sideout (RQ-SMP) and inside-out (IQ-SMP) has been de-scribed and these SMPs showed different characteristicsin relation to NADH oxidation (M0ller et al. 1987). Inour Vv rk, sidedness of the pea leaf SMPs has not beendetermined, and accordingly the generation rate of Q rradicals in the dehydrogenase region (complex I) is notknown exactly. We intend to carry out studies on theNADH-induced Or production by RQ-SMPs or IQ-SMPs obtained from leaves of control and salt-stressedpea plants.
The results reported in this paper for pea leaf mito-chondria contrast to those recently found in leaf peroxi-somes from pea plants grown in the presence of NaCl.In peroxisomal membranes, the NADH-dependentgeneration of Qj^ radicals and the lipid peroxidationwas not altered by NaCl, and the Mn-SQD activitypresent in the peroxisomal matrix did not show sig-nificant variations (Corpas et al. 1993). This indicatesthat the salt-induced changes in Mn-SQD activity re-ported in this paper are due exclusively to the mitochon-drial Mn-SQD isozyme and not to an eventual con-tamination by the less abundant peroxisomal Mn-SQD.
Low levels of Q r radicals can damage enzymes andmembranes per se (Fridovich 1986). However, the mainoxidative threat for mitochondria and other cell orga-nelles could stem from the generation of the highlyreactive hydroxyl radicals ('QH) by reaction of Qjradicals with H^Q ,̂ by a metal-catalyzed site-specificHaber-Weiss reaction (Halliwell and Gutteridge 1989).Hydroxyl radicals are strong oxidizing species that canseverely damage biological membranes and react withmany other compounds present in biological systems,including DNA (Halliwell and Gutteridge 1989). Theformation of QH radicals could be favoured by thepresence of different iron- and copper-containing en-zyme complexes in the inner mitochondrial membrane(Zhang et al, 1990). Therefore, NaCl-tolerant plantscould have evolved a protection mechanism against theincreased production in mitochondria of Qj -dependenttoxic species by high salt concentrations by inducing themitochondrial Mn-SQD activity. In this way, toxiclevels of Q; radicals in these cell organelles could beremoved, and the eventual formation of highly toxicQH radicals avoided.
The H2Q2 concentration in purified intact leaf mi-tochondria was not apparently altered by salinity, but itmust be noted that the concentration of this metabolitedetermined in mitochondria under our experimentalconditions does not reflect the steady-state concentra-tion of H2Q2 in vivo. Mitochondrial membranes arepermeable to H2Q2 (Boveris 1984), and this diffusion
could be increased as a result of NaCl-indueed leakinessof the mitochondrial membrane (Schmitt and Dizengre-mel 1989). Superoxide radicals are the main precursorsof mitochondrial H2Q2 (Boveris 1984, Puntarulo et al,1988) and Qj" generation in these organelles is stimul-ated by NaCl as has been reported in this paper. Theidea that mitochondrial membranes are an importantsource of intracellular H2Q2 has been proposed (Boveris1984, Puntarulo et al. 1988, 1991). Under conditions ofsevere salt stress, an increased export of H2Q2 frommitochondria to the eytosol could be potentially danger-ous for different compartments of the plant cell, due tothe possible formation of the strong oxidizing QH radi-cals by reaction of H2Q2 with Qj" through a metal-catalyzed site-specific Haber-Weiss reaction (Halliwelland Gutteridge 1989). Hydroxyl radicals ( QH) damagebiological membranes and react with most compoundspresent in biological systems (Halliwell and Gutteridge1989).
Qur present results indicate that in the toxicity ofNaCl in plant cells, at the level of mitochondria, anoxidative stress mechanism mediated by superoxideradicals is involved. Although further experiments arestill necessary, mitochondrial Mn-SQD appears to havea function in the molecular mechanism of plant toler-ance to salt stress in Pisum sativum plants, possiblyparticipating in conjuction with other detoxifying en-zymes.
Acknowledgements - The authors are grateful to Mrs D. Lapazfor technical assistance. J.A.H. acknowledges a research fel-lowship from the Instituto de Fomento, Comunidad de Murcia,Spain. This work was supported by grant PB87-0404-02 fromthe DGICYT (Spain).
References
Arnon, D. I. 1949. Copper enzymes in isolated chloroplasts.Polyphenoloxidase in Beta vulgaris. - Plant Physiol. 24:1-15.
Boveris, A. 1984. Determination of the production of super-oxide radicals and hydrogen peroxide in mitochondria. -Methods Enzymol. 105: 429-435.
Bowler, C , Slooten, L., Vandenbrauden, S., De Rycke, R.,Botterman. J.. Sybesma, G., Van Montagu, M. & Inze, D.1991. Manganese superoxide dismutase can reduce cellulardamage mediated by oxygen radicals in transgenic plants. -EMBO J. 10: 1723-1732.
Bradford, M.M. 1976. A rapid and sensitive method for thequantitation of microgram quantities of protein utilizing theprinciple of protein-dye binding. - Anal. Biochem. 72:248-254.
Brown, S., Day, D. A. & Critchley, C. 1987. Salt tolerance -does leaf respiration have a contribution to make? - InPlant Mitochondria: Structural, Functional and Physiolog-ical Aspects (A.L. Moore and R.B. Bechey, eds), pp.393-396. Plenum Press, New York, NY. ISBN 0-306-42572-6.
Buege, J.A. & Aust, S. D. 1978. Microsomal lipid peroxida-tion. - Methods Enzymol. 52: 302-310.
Chapman, H.D. & Pratt, P. R 1961. Methods of analysis forsoil, plants and water. - University of California (River-side), Division of Agriculture.
Corpas, F. J., Gomez. M., Hernandez. J. A. & del Rio, L. A.1993. Metabolism of activated oxygen in leaf peroxisomes
8 Physiol, Plant. 89. t993 109

from two Fisum sativum L, cultivars with different sensi-tivity to sodium chloride, - J, Plant Physiol, 141: 160-165,
Davis, B, J, 1964, Disc gel electrophoresis, II, - Ann, NY,Acad, Sci, 121: 404-427,
del Rio, L, A,, Sandalio, L, M,, Yanez, J. & Gomez, M, 1985,Induction of a manganese-containing superoxide dismutasein leaves of Fisum .sativum L, by high nutrient levels of zincand manganese, - J, Inorg, Biochem, 24: 25-34,
- , Sevilla, E, Sandalio, L, M, & Palma, J,M, 1991, Nutri-tional effect and expression of superoxide dismutases: in-duction and gene expression, diagnostics, prospective pro-tection against oxygen toxicity, - Eree Rad, Res, Commun,12-13: 819-828,
Douce, R,, Christensen, E L , & Bonner, W, D, Jr, 1972,Preparation of intact plant mitochondria, - Biochim, Bio-phys, Acta 275: 148-160,
Frew, J, E,, Jones, P, & Scholes, G, 1983, Spectrophotometricdeterminations of hydrogen peroxide and organic hydro-peroxides at low concentrations in aqueous solutions, -Anal, Chim, Acta 155: 139-150,
Eridovich, 1, 1986, Biological effects of the superoxide radical,- Arch, Biochem, Biophys, 247: 1-11,
Green, S, Mazur, A, & Shorr, E, 1956, Mechanism of thecatalytic oxidation of adrenaline by ferritin, - J, Biol,Chem, 220: 237-255,
Halliwcll, B, & Guttcridge, J,M,C, 1989, Eree Radicals inBiology and Medicine, pp, 86-179, - Oxford UniversityPress, London, ISBN 0-19-855291-2,
Jolivet, Y,, Pireaux, J, C, & Dizengremel, P, 1990, Changes inproperties of barley leaf mitochondria from NaCI-treatedplants, - Plant Physiol, 94: 641-646,
Lambers, H, 1985, Respiration in intact plants and tissues: itsregulation and dependence on environmental factors, me-tabolism and invaded organisms, - In Encyclopedia of PlantPhysiology, New Series, Vol, 18 (R, Douce and D, A, Day,eds), pp, 418-473, Springer-Verlag, Berlin, ISBN 3-540-139.35-4,
Marcillat, O,, Zhang, Y,, Lin, SW, & Davies, J,A, 1988,Mitochondria contain a proteolytic system which can recog-nize and degrade oxidatively-denaturated proteins, - Bio-chem, J, 2.54: 677-683,
McCord, J, M, & Eridovich, I, 1969, Superoxide dismutase: ancnzymic function crythrocyprein (hemocuprein), - J, Biol,Chem, 244: 6049-6055,
M0llcr, LM, & Palmer, J,M, 1982, Direct evidence for thepresence of a rotenone-resistant NADH dehydrogenase onthe inner surface of the inner membrane of plant mitochon-dria, - Physiol Plant, 54: 267-274,
- & Lin, W, 1986, Membrane-bound NAD(P)H dehydro-gcnases in higher plant cells, - Annu, Rev, Plant Physiol,37: 309-324,
- , Liden, A, C , Ericson, 1, & Gardestrom, P, 1987, Isolationof submitochondrial particles with different polarities, -Methods Enzymol, 148: 442-453,
Nohl, H, & Hegner, D, 1978, Do mitochondria produce oxy-gen radicals in vivo? - Eur, J, Biochem, 82: :563-567,
Palma, J, M,, Garrido, M,, Rodiguez-Garcia, M, I, & del Rio,L, A, 1991, Peroxisome proliferation and oxidative stress
mediated by activated oxygen species in plant peroxisomes,- Arch, Biochem, Biophys, 287: 68-74,
Price, A, H,, Atherton, N, M, & Hendry, G, A, E, 1989, Plantsunder drought-stress generate activated oxygen, - EreeRad, Res, Commun, 8: 61-66,
Puntarulo, S,, Sanchez, R, A, & Boveris, A, 1988, Hydrogenperoxide metabolism in soybean axes at the onset of germi-nation, - Plant Physiol, 86: 626-630,
- , Galleano, M,, Sanchez, R, A, & Boveris, A, 1991, Super-oxide anion and hydrogen peroxide metabolism in soybeanembrionic axes during germination, - Biochim, Biophys,Acta, 1074: 277-283,
Rabinowitch, H, D, & Eridovich, I, 1983, Superoxide radicals,superoxide dismutase and oxygen toxicity in plants, -Photochem, Photobiol, 37: 679-690,
Rich, P, R, & Bonner, W, D, Jr, 1978, The sites of superoxideanion generation in higher plant mitochondria, - Arch,Biochem, Biophys, 188: 206-213,
Sandalio, L, M,, Palma, J, M, & del Rfo, L, A, 1987, Localiza-tion of manganese superoxide dismutase in peroxisomesisolated from Fisum sativum L, - Plant, Sci, 51: 1-8,
Schmitt, M,R, & Edwards, G,E, 1982, Isolation and puri-fication of intact peroxisomes from green leaf tissue, - PlantPhysiol, 70: 1213-1217,
Schmitt, N, & Dizengremel, P, 1989, Effect of osmotic stresson mitochondria isolated from etiolated mung bean andsorghum seedlings, - Plant Physiol, Biochem, 27: 17-26,
Schnarrenberger, C A , , Oeser, A, & Tolbert, N, E, 1971,Development of microbodies in sunflower cotyledons andcastor bean endosperm during germination, - Plant Phy-siol, 48: 566-574,
Schwitzguebel, J, P, & Siegenthalcr, P, A, 1984, Purification ofperoxisomes and mitochondria from spinach leaf by Percollgradient centrifugation, - Plant Physiol, 75: 670-674,
Seeman, J, R, & Critchley, C, 1985, Effects of salt stress on thegrowth, ion content, stomatal behaviour and photosyn-thetic capacity of a salt-sensitive species, Fhaseolus vulgarisL, - Planta 164: 151-162,
Sevilla, E, Hellin, E, & del Rio, L, A, 1987, Induction of ironand manganese superoxide dismutases in Citrus limonumleaves by iron, - Life Sci, Adv, Ser, B6: 133-139,
Singha, S, & Choudhuri, M, A, 1990, Effect of salinity (NaCl)stress on H^Oi metabolism in Vigna and Oryza seedlings, -Biochem, Physiol, Pflanz, 186: 69^74,
Sjolin, A,M, & M0ller, LM, 1991, The effect of polyaminesand other cations on NADH oxidation on the inner surfaceof the inner mitochondrial membrane, - Plant Physiol,Biochem, 29: 607-613,
Walk, S,J, & Hock, B, 1977, Glyoxysomal malate dehydro-genase of watermelon cotyledon: de novo synthesis on cy-toplasmic ribosomes, - Planta 134: 277-285,
Weisiger, R,A, & Eridovich, I, 1973, Superoxide dismutase:Organelle specificity, - J, Biol, Chem, 248: 3582-3592,
Zhang, Y,, Marcillat, O,, Giulivi, C . Ernster, L, & Davies,K,J, A, 1990, The oxidative inactivation of mitochondrialelectron transport chain components and ATPase, - J, Biol,Chem, 265: 16330-16336,
Edited by P, Gardestrom
110 Physiol, Plant, 89, t993





















