
Flagellin of Pseudomonas aeruginosa inhibits Na+ transport in airway epithelia

Review
Molecular handling of cadmium in transporting epithelia
Rudolfs K. Zalups* and Sarfaraz Ahmad
Division of Basic Medical Sciences, Mercer University School of Medicine, Macon, GA 31207, USA
Received 15 August 2002; accepted 21 October 2002
Abstract
Cadmium (Cd) is an industrial and environmental pollutant that affects adversely a number of organs in humans and other mammals,including the kidneys, liver, lungs, pancreas, testis, and placenta. The liver and kidneys, which are the primary organs involved in theelimination of systemic Cd, are especially sensitive to the toxic effects of Cd. Because Cd ions possess a high affinity for sulfhydryl groupsand thiolate anions, the cellular and molecular mechanisms involved in the handling and toxicity of Cd in target organs can be defined largelyby the molecular interactions that occur between Cd ions and various sulfhydryl-containing molecules that are present in both theintracellular and extracellular compartments. A great deal of scientific data have been collected over the years to better define the toxiceffects of Cd in the primary target organs. Notwithstanding all of the new developments made and information gathered, it is surprising thatvery little is known about the cellular and molecular mechanisms involved in the uptake, retention, and elimination of Cd in target epithelialcells. Therefore, the primary purpose of this review is to summarize and put into perspective some of the more salient current findings,assertions, and hypotheses pertaining to the transport and handling of Cd in the epithelial cells of target organs. Particular attention has beenplaced on the molecular mechanisms involved in the absorption, retention, and secretion of Cd in small intestinal enterocytes, hepatocytes,and tubular epithelial cells lining both proximal and distal portions of the nephron. The purpose of this review is not only to provide asummary of published findings but also to provide speculations and testable hypotheses based on contemporary findings made in other areasof research, with the hope that they may promote and serve as the impetus for future investigations designed to define more precisely thecellular mechanisms involved in the transport and handling of Cd within the body.© 2003 Elsevier Science (USA). All rights reserved.
Keywords: Cadmium transport; Epithelial cells; Membrane metal-transporters; Ca channels; Endocytosis; Intestines; Liver; Kidneys; Albumin; Metallothio-nein; Glutathione; Cysteine; Nonprotein Thiols
Introduction
Cadmium (Cd) is a group IIB metal that has an atomicweight of 112.41 g/mol and it exists in the 0 or 2� oxidationstate. It is found naturally in the earth’s crust and is usuallypresent in the environment as an inorganic salt (such ascadmium oxide (CdO), cadmium chloride (CdCl2), or cad-mium sulfate (CdSO4); ATSDR, 1999). Although Cd maychange chemical forms, the metal ion itself is not removedfrom the environment (Morselt, 1991). Accordingly, as the
environment continues to be contaminated with this metal,there is an increasing risk of humans and other mammalsbeing exposed to Cd. There are estimates that about 25,000to 30,000 tons of Cd are released into the environment eachyear, with the major contributions coming from humanactivities (4,000 to 13,000 tons per year), such as miningand the burning of fossil fuels (ATSDR, 1999).
Cigarette smoke is by far the largest source of Cd expo-sure in the general human population. Each cigarette maycontain from 1 to 2 �g of Cd, and 40–60% of the Cd ininhaled smoke generally passes through the pulmonary ep-ithelium into systemic circulation (ATSDR, 1999; Elinderet al., 1976; Lewis et al., 1972). Among nonsmokers, in-gestion of food contaminated with Cd is a major source ofCd exposure. Fish, organ meat (especially from liver and
* Corresponding author. Division of Basic Medical Sciences, MercerUniversity School of Medicine, 1550 College Street, Macon, GA 31207,USA. Fax: �1-478-301-5489.
E-mail address: [email protected]. (R.K. Zalups).
R
Available online at www.sciencedirect.com
Toxicology and Applied Pharmacology 186 (2003) 163–188 www.elsevier.com/locate/tap
0041-008X/03/$ – see front matter © 2003 Elsevier Science (USA). All rights reserved.doi:10.1016/S0041-008X(02)00021-2

kidneys), and grain and cereal products usually contributethe greatest amount of Cd to the diet. Potatoes, leafy veg-etables, and root vegetables can also contain relatively highlevels of Cd (ATSDR, 1999).
In humans and other mammals, Cd affects adversely anumber of organs and tissues, including the kidneys, liver,lung, pancreas, testis, placenta, and bone (ATSDR, 1999;Bhattacharyya et al., 2000; Diamond and Zalups, 1998;Friberg et al., 1986; Goyer et al., 1984; Habeebu et al.,1998; Jarup et al., 1998; Kamiyama et al., 1995; Kazantzis,1978; Lui et al., 1998a, 1998b, 2000; Min et al., 1986, 1996;Nordberg and Nordberg, 2000; Nordberg et al., 1985;Oteiza et al., 1999; Sarkar et al., 1998; Zalups et al., 1992),with the liver and kidneys being two of the primary organsin which the toxic effects of Cd are expressed. In addition tothe direct cytotoxic effects that can lead to apoptotic and/ornecrotic events, Cd can have potent carcinogenic effects intarget organs (Waalkes et al., 1992). In fact, based onepidemiological and toxicological studies in humans andexperimental animals, the International Agency for Re-search on Cancer (IARC) has classified Cd as a Category Icarcinogen (IARC, 1993). Recent findings indicate that thecarcinogenic effects of Cd are related to the activation ofprotooncogenes (Joseph et al., 2001, 2002). Overall, Cd hasbeen ranked as high as seventh on the Top 20 Hazardous
Substances Priority List by the Agency for Toxic Sub-stances and Disease Registry and the U.S. EnvironmentalProtection Agency (Fay and Mumtaz, 1996).
Cellular injury induced by Cd is dependent on a numberof factors, including dose, route of exposure, and duration ofexposure. In order to understand how Cd intoxicates targetcells, it is necessary to gain a detailed understanding of themechanisms involved in the transport of Cd into those cellsand the subsequent bonding interactions that occur with Cdbefore and after it has entered into the intracellular milieu.In particular, it is important to gain a thorough understand-ing of the bonding interactions that occur between Cd andthe abundant low-molecular-weight, intracellular thiols(such as GSH and metallothionein (MT)) and critical nu-cleophilic binding sites on proteins that play vital roles inmaintaining normal homeostasis in cells. Fig. 1 illustratesthe complex nature of the handling of Cd in the body afteroral/gastrointestinal and pulmonary exposure to Cd. Thisfigure illustrates some of the potential species of Cd thatmay be involved in the uptake, accumulation, and elimina-tion of Cd in the primary target organs.
Although a considerable body of scientific data on themolecular interactions and handling of Cd in target epithe-lial cells has been accrued over the years, many importantquestions about the mechanisms involved in the transport
Fig. 1. A summary of the potential forms of Cd present in the small and large intestines, liver, biliary tree, and kidneys after oral or pulmonary exposure tocadmium salts or Cd-protein complexes. This figure provides a schematic presentation of the potential pathways involved in the handling and excretion ofthe different forms of Cd that may be present in the relevant compartments of the body. MT, metallothionein; Cys, cysteine; GSH, glutathione; N-Acetyl-Cys,N-acetylcysteine; homoCys, homocysteine.
164 R.K. Zalups, S. Ahmad / Toxicology and Applied Pharmacology 186 (2003) 163–188

and handling of Cd in these cells remain undefined. There-fore, the primary purpose of this review is to summarize andto put into perspective some of the more salient currentfindings, assertions, and hypotheses pertaining to the han-dling of Cd in the epithelial cells of target organs, includingmolecular mechanisms involved in the absorption, trans-port, and elimination of Cd.
Exposure and absorption of cadmium
Humans are generally exposed to Cd by two main routes,inhalation and ingestion. Absorption of Cd by skin is rela-tively insignificant, although small amounts of Cd can beabsorbed percutaneously during long periods of exposure(Wester et al., 1992). The body burden of Cd is derivedprimarily from ingestion of food and drinking water con-taminated with Cd. CdCl2 is the principal form of Cd asso-ciated with oral exposure, as it is highly soluble in water. Bycontrast, CdO is the main form of Cd associated with inha-lation exposure (ATSDR, 1999; Oberdorster, 1992). Inter-estingly though, a significant fraction of inhaled Cd ends upin the gastrointestinal tract as a result of mucociliary clear-ance and subsequent ingestion. In fact, Moore et al. (1973)have demonstrated that as much as 60% of the inhaled doseof Cd ended up being translocated to the gastrointestinaltract in rats exposed acutely to aerosols containing Cdcarbonate. Pulmonary absorption of Cd is relatively moreefficient than the absorption of Cd along the gastrointestinaltract. It has been shown that the efficiency of gastrointesti-nal absorption of Cd is only about 1–2% in mice and rats(Decker et al., 1958; Ragan 1977), 0.5–3% in monkeys(Nordberg et al., 1971), 2% in goats (Miller et al., 1969),and 5% in pigs and lambs (Cousins et al., 1973; Doyle et al.,1974). Among most species of mammals studied, the effi-ciency of gastrointestinal absorption of Cd appears to begreatest (nearly 16%) in cattle (Miller et al., 1969).
Cellular uptake and transport of cadmium in targetorgans
In response to the deficiency in knowledge of the han-dling of Cd by target epithelial cells, a number of hypoth-eses have been proposed recently. One hypothesis, whichhas recently gained particular favor with a number of in-vestigators, states that selective uptake of Cd involves, atleast in part, mechanisms whereby Cd interacts with, andcompetes for, binding site(s) on membrane proteins in-volved in the transport of essential elements (such as cal-cium (Ca), iron (Fe), and zinc (Zn)) into target epithelialcells, possibly through some form of ionic mimicry. An-other contemporary hypothesis states that, when Cd formsS-conjugates with low-molecular-weight thiols, such aswith GSH and cysteine (Cys), these conjugates serve asmolecular homologs or mimics at the sites of specific trans-
port proteins involved normally in the absorptive transportof amino acids, oligopeptides, organic anions, organic cat-ions, or other important homeostatic molecules. Anotherhypothesis that has gained considerable attention over re-cent years states that endocytosis of proteins containing Cdis a mechanism by which Cd ions gain entry into certaintransporting epithelia. For example, it is believed by somethat, when the Cd ions bind to MT, albumin, or otherproteins, the conjugates formed have the propensity to serveas substrates for absorptive and/or receptor-mediated endo-cytotic transport. It is noteworthy to point out that two ofthese hypotheses incorporate, at least by implication, theconcept of ionic and/or molecular homology or mimicry.
In the context of the present review, molecular mimicryrefers to a process by which an essential homeostatic mol-ecule, which is normally found within the body (such as anamino acid), bonds to Cd and forms a molecular species thatcan act as a functional homolog or mimic of either theendogenous molecule bound to the metal or a completelydifferent endogenous homeostatic molecule. To illustratethe concept of molecular homology or mimicry, Zalups andcolleagues have proposed that dicysteinylmercury (Cys-Hg-Cys) may serve as a molecular homolog or mimic of theamino acid cystine at the site(s) of transporter(s) that arenormally involved in the absorption of this amino acid inrenal proximal tubular epithelia (Cannon et al., 2000, 2001).It has also been suggested that the Cys S-conjugate ofmethylmercury can act as a molecular mimic of the aminoacid methionine at the site of amino acid transporters in theluminal plasma membrane of endothelial cells lining theblood–brain barrier (Aschner and Clarkson, 1989; Kerper etal., 1992).
For the purpose of this review, the concept of ionicmimicry or homology refers to situations in which the ionic,divalent form of Cd can serve as a functional homolog ofthe ionic, divalent form of other elements that are trans-ported by specific membrane proteins. For example, sinceCd2� has a smaller ionic radius than Ca2�, and since Cdions have been shown to effectively inhibit the movement ofCa ions through Ca channels in excitable cells (Lansman etal., 1986; Taylor, 1988), Cd2� can be thought of as apotential ionic “mimic” or homolog of Ca2� at certain typesof Ca channels. As it turns out, both Ca and Cd ions canenter and traverse through Ca channels, although Cd ionsmove through the channels at much slower rates than Caions. Experimental findings indicate that Ca channels canalso serve as molecular conduits permitting the extracellularto intracellular flux of Cd ions in some types of epithelialcells (Blazka and Shaikh, 1991; Friedman and Gesek, 1994;Hinkle et al., 1987, 1992). Many of these findings, however,need to be put into perspective, inasmuch as they wereextrapolated from cultured epithelial cells bathed in protein-free physiological salt solutions, which clearly does notrepresent the conditions in which Cd is presented to thevarious target epithelial cells in vivo. Despite this shortcom-ing, the importance of these findings dictates that any as-
165R.K. Zalups, S. Ahmad / Toxicology and Applied Pharmacology 186 (2003) 163–188

sessment of the mechanisms involved in the transport of Cdin epithelial cells warrants a consideration of the potentialroles of the different Ca channels.
Gastrointestinal handling of Cd
In humans, the efficiency of gastrointestinal absorptionof Cd has been reported to be approximately 3–8% of theingested load (ATSDR). Based on data from rodents, mostof the absorption of Cd appears to occur primarily in theduodenum and early jejunum (Andersen et al., 1994). Sev-eral dietary elements and constituents, such as Fe, Zn, Ca,and protein content, can influence greatly the absorption ofCd along the gastrointestinal tract. For instance, it has beendemonstrated that the intestinal absorption of inorganic Cddecreases in rats by as much as 80% following dietarysupplementation with Ca and phosphorous (Pi), Zn, or Fe(Groten et al., 1991b). Several investigators have assertedthat dietary Fe has an especially prominent effect on theintestinal absorption of Cd (Schumann et al., 1996), andhave determined that there is a correlation between Fe statusand the intestinal absorption of Cd in humans and othermammals (Berglund et al., 1994; Choudhury et al., 2001;Flanagan et al., 1978; Fox et al., 1980). This relationship isparticularly evident in anemic adult females (Choudhury etal., 2001). As a matter of fact, the bioavailability of Cd froma single dietary load has been shown to be almost threetimes greater in women than in men (Shaikh and Smith,1980).
Intestinal absorption of Cd is characterized by a rapidrate of accumulation of Cd within the intestinal mucosa anda low rate of diffusive transfer into systemic circulation(Elsenhans et al., 1997). The intestinal uptake of Cd and itssubsequent distribution to target tissues is greatly dependenton the chemical form of Cd presented to the intestinalepithelium. For example, it has been shown that intestinalaccumulation and retention of Cd in rats was greater afteroral exposure to CdCl2 than oral exposure to Cd-MT (Gro-ten et al., 1992). However, it appears that the ratio of theconcentration of Cd in the kidney to the concentration of Cdin the liver was higher after oral administration of Cd-MTthan after oral administration of CdCl2 (Groten et al.,1991a). This effect was likely due to the fact that Cd boundto MT (Cd-MT) or cysteine (Cd-(Cys)2) is in a form thatpermits Cd ions to be delivered to, and taken up morereadily by, the target epithelial cells in the kidneys (Mu-rakami et al., 1987; Cherian and Shaikh, 1975; Tanaka et al.,1975). In support of this hypothesis are data showing thatrenal injury in rats is more severe following exposure toCd-MT or Cd-(Cys)2 than after exposure to CdCl2 (Cherian,1980a; Maitani et al., 1985; Min et al., 1986).
It is particularly interesting to note that the role of MT inthe distribution and accumulation of Cd in the kidneys andliver is mixed after oral/intestinal exposure to Cd. Kimura etal. (1998) have shown that, following a low oral dose of Cd,
that hepatic levels and the sum of hepatic and renal levelswere significantly greater in MT-I/II knockout (null) micethan in corresponding wild-type mice. When the dose wasincreased twofold, no differences in the disposition of Cdwere detected between the two groups. Oral pretreatmentwith Zn appeared to limit the amount of orally administeredCd that was available to systemic organs in the MT-nullmice. These findings indicate that the MT induced by cy-tosolic Zn in enterocytes bound to and trapped the Cd thatwas taken up from the intestinal lumen.
In contrast to the findings of Kimura et al. (1998), Liu etal. (2001) have demonstrated, in both MT-null and wild-type mice, that approximately 0.25–0.75% of the dose waspresent in the liver and 0.05– 0.15% of the dose was presentin the kidneys 4 h subsequent to single oral doses of Cd.These findings indicate that, at the doses studied, MT doesnot appear to play a significant role in the acute intestinalabsorption of Cd and the subsequent disposition of Cd in theliver and kidneys. These investigators also found that, in-dependent of dose, approximately 60% of the dose localizedin the liver and about 5% of the dose localized in thekidneys in both MT-null and wild-type mice 4 h after anintravenous dose of Cd, which is consistent with acute renaland hepatic disposition of Cd in rats administered Cd intra-venously (Zalups, 2000a).
Transport of Cd into enterocytes from the intestinal lumen
Foulkes and colleagues (Foulkes, 1985, 1988, 2000;Foulkes and McMullen, 1986a) have proposed a two-stepprocess for the absorptive movement of Cd ions from theintestinal lumen into enterocytes. Using everted sacs formedfrom portions of the jejunum, they showed that absorptionof Cd into enterocytes was preceded by the movement of Cdions into a compartment that was accessible to chelators, butwas insensitive to temperature. The Cd associated with thiscompartment likely represented nonspecific binding of Cdto the luminal plasma membrane. The second step in theabsorptive process appeared to involve the slower move-ment of Cd into a temperature-sensitive compartment thatwas not accessible to chelators. This second step likelyrepresented the actual movement or transport of Cd acrossthe luminal plasma membrane into the enterocytes.
Although the specific mechanism(s) involved in the ab-sorptive transport of Cd is/are not known presently, there isa growing body of evidence indicating that the divalentmetal transporter 1 (DMT1) likely plays a key role intransporting Cd into enterocytes (Fig. 2). DMT1 is a proton-coupled, membrane potential-sensitive, transport proteinthat is capable of transporting a number of divalent cations.In enterocytes, DMT1 (also known as DCT1 or Nramp2)has been shown to be present in the luminal plasma mem-brane, where it is believed to serve as a major pathway forthe absorption of dietary non-heme Fe (Ferguson et al.,2001; Tandy et al., 2000). The level of expression of thistransporter decreases along the length of the gastrointestinal
166 R.K. Zalups, S. Ahmad / Toxicology and Applied Pharmacology 186 (2003) 163–188

Fig. 2. A diagrammatic representation of the potential mechanisms involved in the luminal and basolateral transport of cadmium (Cd) in enterocytes of thesmall intestine. Depending on the dietary source of Cd, there are a number of different forms of Cd that can be presented to the luminal membrane ofenterocytes. One of the potential mechanisms involved in the uptake of Cd by enterocytes is endocytosis of proteins, including metallothionein (MT), to whichCd is bound. However, there is very little information about the role of endocytosis in the luminal uptake of Cd by enterocytes. Following ingestion of aprotein-containing meal, oligopeptides and amino acids are formed in the lumen of the small intestine by pancreatic enzymes and enzymes localized on theluminal plasma membrane on enterocytes. The oligopeptides and amino acids formed in the lumen of the early portions of the small intestines are absorbedquickly and efficiently by both sodium (Na�)-dependent and Na�-independent transporters. Since amino acid transporters have been implicated in the luminalabsorptive transport of mercuric conjugates of cysteine (Cys) in the renal proximal tubule, and since some of the same transport systems are also present inenterocytes, it is reasonable to postulate that certain amino acid and oligopeptide transport systems may be involved in the absorptive transport ofCys-containing oligopeptide S-conjugates and/or Cys-S-conjugates of Cd along the small intestine. There is a significant body of evidence indicating that therecently characterized divalent metal transporter 1, DMT1, can transport Cd into enterocytes. This transporter, which is expressed in the luminal plasmamembrane, is a proton-coupled transporter that appears to be involved in the intestinal absorption of non-heme ferrous (2�) iron (Fe). When Cd binds toDMT1, through some form of ionic homology or “mimicry,” it is transported into the cytosolic compartment of enterocytes. Recent findings indicate thatthe luminal uptake of Cd can also occur through one of the luminal transporters of zinc (Zn). One possible Zn transporter that may be involved in the luminaluptake of Cd is ZTL1, which has been localized recently in the luminal plasma membrane of enterocytes. It is also possible that some forms of Cd may “leak”from the luminal compartment into the basolateral compartment through the relatively leaky junctional complexes between adjacent enterocytes in the earlyportions of the small intestine. Intercellular leak of Cd is also likely to occur when enterocytes begin to become intoxicated by Cd. Calcium (Ca) channelsmay serve as an additional mechanism by which Cd ions gain access into the intracellular milieu from the luminal compartment. Enterocytes are known torespond to the biologically active form of vitamin D, i.e., 1,25-dihydoxycholecalciferol, by promoting the absorption of Ca. One of the factors associatedwith this absorption is the synthesis of calbindin-D, which is a calcium-binding protein that assists in the luminal to basal trafficking of Ca in enterocytes.Some of the Ca absorbed at the luminal membrane of enterocytes enters through Ca channels, is delivered to the basolateral membrane by calbindin-D, andis exported out of the enterocyte by a Ca-stimulated ATPase. Assuming that Cd can serve as an ionic homolog or mimic of Ca, it seems possible that Cdions may enter into enterocytes through Ca channels and are transferred to the basolateral membrane by calbindin-D, where some of Cd ions may be exportedout of the enterocyte by one or more mechanisms present in the basolateral membrane, including the Ca2�-ATPase. Recent molecular evidence indicates thatan Fe transporter, homologous to DMT1, is present in the basolateral plasma membrane of enterocytes and is believed to be involved in the cytosolic toextracellular transport of Fe. This transporter has been referred to as metal transporter protein 1 (MTP1). Since both Fe and Cd appear to be transported byDMT1, MTP1 may also transport Cd ions from within the cytosolic compartment into the basolateral compartment. Basolateral amino acid transporters andorganic anion transporters are also possible transporters that may transport Cd both into and out of enterocytes at the basolateral membrane. As theintracellular pool of Cd accumulates within the enterocytes, this pool can interact with various intracellular components and compartments within the cells.Some of the Cd ions that enter into enterocytes induce the transcription of the genes for MT-1 and MT-2, by mechanisms that have not yet been defined fully.Increases in the cellular content of MT protein result from the translation of the increased amounts of mRNA for MT-1 and MT-2 that are induced followingexposure to Cd. The induced MT protein serves as a sink to bind some of the intracellular Cd, which results in increased retention of Cd within the enterocytes.If the intracellular pool of exchangeable Cd increases beyond what the protective elements inside the enterocytes can handle, oxidative stress is induced,which in turn can alter mitochondrial respiratory activity and lead to lipid peroxidation in the plasma membrane and other perturbations in cellularmetabolism. All of these effects can, and do, lead to the induction of cell death by either necrosis or apoptosis, which results in the release of Cd (much ofwhich is in the form of Cd-MT) from within the enterocyte. Pathways depicted by solid black arrows are supported either directly or indirectly byexperimental evidence. The thicker solid black lines denote pathways for which there is substantive evidentiary support. By contrast, the dashed linesrepresent pathways of secondary importance or pathways that are based primarily on indirect findings or speculation. Question marks associated with apathway denote uncertainty for the occurrence of that pathway in vivo.

tract, with the highest levels expressed in the proximalduodenum and the lowest levels expressed toward the distalcolon. In addition to being expressed in enterocytes, DMT1is expressed by hepatocytes and by various tubular epithe-lial cells in the kidneys. It also appears to be expressed bycells in the lung, heart, brain, and testis (Gunshin et al.,1997). It is of particular interest that DMT1 has an unusualability to transport a broad range of metal ions, includingFe2�, Zn2�, Cu2�, Cd2�, Mn2�, Co2�, Ni2�, and Pb2�.
Using an immortalized line of human intestinal (Caco-2)cells that express DMT1, Tallkvist et al. (2001) and Elismaand Jumarie (2001) recently have provided substantive ev-idence supporting the hypothesis that DMT1 transports bothFe and Cd in a competitive manner, possibly through amechanism of ionic “mimicry.” They showed that treatmentof the immortalized intestinal cells with Fe inhibited theuptake of Cd and decreased the expression of the gene forDMT1, which correlated with decreased uptake of Cd. Ad-ditional data implicating DMT1 as a transporter of Cdcomes from experiments in which Xenopus laevis oocyteswere manipulated genetically to express DMT1 (Zoller etal., 1999).
Elisma and Jumarie (2001) have also demonstrated thatZn and Cd compete for a luminal membrane transporter inenterocytes that is not DMT1. Their data indicate that Cdmay also enter into enterocytes through one of the transportsystems involved in the luminal uptake of Zn, perhaps athZTL1 (Fig. 2), which is a human, zinc-regulated, trans-porter of zinc expressed in the luminal membrane of en-terocytes (Cragg et al., 2001). Up until the recent charac-terization of hZTL1, a luminal membrane zinc transporterhad not been identified in enterocytes. On the other hand,the zinc transporter ZNT1 had been identified in the baso-lateral membrane of enterocytes (Palmiter and Findley,1995). Data obtained with apical membrane vesicles iso-lated from the small intestine of the pig also provide supportfor a common site of transport or binding of Zn and Cd(Tacnet et al., 1990, 1991). These data show that Zn bindskinetically to a distinct site on the plasma membrane, whichcan be inhibited by Cd, but not by Ca. The binding of Zn orCd to the intravesicular portions of the plasma membraneoccurred with similar affinities.
Experimental evidence indicates that, when the intestinalepithelium is exposed to Cd-MT, some of the Cd-MTcrosses the epithelium intact and enters into the capillariesof the lamina propria to be delivered into the portal circu-lation (Cherian, 1979; Cherian et al., 1978; Sugawara andSugawara, 1991). In order to explain these findings, Cd-MTmust cross the intestinal epithelium either by passingthrough the relatively leaky junctional complexes betweenadjacent enterocytes and/or by a mechanism in whichCd-MT is taken up by an endocytotic mechanism and theneither is transported across the basolateral membrane by anexocytotic mechanism or is released into the basolateralcompartment subsequent to the death of the enterocytesinduced by the Cd-MT (Fig. 2). Although endocytosis of
Cd-MT by enterocytes is a distinct possibility, basolateralexocytosis, however, seems highly improbable, inasmuch asMT does not contain a leader sequence that would promotethe exocytosis of this protein from cells. Moreover, endo-cytosed proteins are generally not exocytosed out of trans-porting epithelial cells, but rather are degraded intracellu-larly in lysosomes via hydrolytic enzymes. It is possible,however, that Cd-MT that is transported into and/or isformed within enterocytes is released when the cells be-come senescent or intoxicated. Release of Cd-MT underthese circumstances could provide a direct pathway forCd-MT to enter into the capillaries situated in the laminapropria (Fig. 2). In fact, data provided by Kimura et al.(1998) indicate that Cd-MT originating from senescent orintoxicated enterocytes likely serves as a molecular shuttlethat delivers Cd directly to the kidneys. Yet another possi-bility incorporates the recent hypothesis of Prozialeck(2000), who purports that Cd can disrupt junctional com-plexes between epithelial cells by altering the Ca-depen-dent, E-cadherin/�-catenin system, which is part of thezonula adherins of the junctional complex. Disruption ofintercellular junctions would clearly provide a path forCd-MT or other forms of Cd to pass between adjacententerocytes and enter the lamina propria of the intestinalmucosa (Fig. 2).
Inasmuch as endocytosis of Cd-MT has been implicatedas a potential means by which Cd can enter into enterocytes,there remains the distinct possibility that endocytosis of Cdbound to smaller polypeptides of larger proteins may alsoserve as a mechanism to permit the entry Cd into entero-cytes. Despite the fact that proteins present in the lumen ofthe small intestine are broken down to polypeptides, oli-gopeptides, and amino acids by the actions of pancreaticenzymes and luminal membrane peptidases, some of the Cdingested with proteins may be absorbed by the endocytosisof a Cd-polypeptide complex formed during the digestiveprocess (Figs. 1 and 2). Due to the great abundance ofamino acid and small peptide transporters in the enterocytesof the small intestine (Ganapathy et al., 2001), it is alsopossible that these transporters may participate in the lumi-nal uptake of Cd as S-conjugates of Cys or Cys-containingoligopeptides, which are likely present in the intestinallumen after a meal (Fig. 2). An additional source of CysS-conjugates of Cd likely comes from the enzymatic deg-radation of hepatically secreted GSH S-conjugates of Cd.The corresponding cysteinylglycine and Cys S-conjugatesof Cd formed along the biliary tree by the activities of�-glutamyltransferase and cysteinylglycinase are deliveredto the lumen of the duodenum at the sphincter of Odi (Figs.1 and 2). Since there is a strong body of evidence implicat-ing specific amino acid transporters (some of which arepresent in enterocytes) in the uptake of mercuric conjugatesof Cys in proximal tubular epithelial cells (Cannon et al.,2000, 2001; Zalups, 2000b), it seems possible that CysS-conjugates of Cd may be taken up by enterocytes by thesame or similar amino acid transport systems.
168 R.K. Zalups, S. Ahmad / Toxicology and Applied Pharmacology 186 (2003) 163–188

As a final comment on the inward transport of Cd intoenterocytes, there is currently a lack of substantive evidenceimplicating ion channels, especially Ca channels, in theabsorption of Cd ions along any portion of the small or largeintestines. However, there are in vivo data from rats pro-vided by Felley-Bosco and Diezi (1992) indicating thatsome factors associated with intestinal Ca metabolism canaffect greatly the absorption of Cd by enterocytes. Morespecifically, these investigators have demonstrated thatchronic restriction of dietary intake of Ca enhanced greatlythe intestinal absorption of orally administered Cd andcaused a significant increase in the accumulation of Cd andMT in target organs, such the kidneys and liver. These datalead one to postulate that, with a deficient supply of Ca inthe intestinal lumen, Cd ions can gain entry into enterocytesvia one or more of the luminal transporters of Ca, such as Cachannels or Ca carriers (Fig. 2). It also seems possible thatthe Ca-binding protein, calbindin-D, expressed in responseto 1,25-dihydroxy-vitamin D3 within enterocytes, may fa-cilitate the absorption of Cd by delivering Cd to the baso-lateral membrane of the enterocytes (Fig. 2).
Retention of Cd in enterocytes
It has been postulated that MT that is induced in entero-cytes (and perhaps other cells) plays an important role in theretention of Cd within the mucosa of the small intestineafter the ingestion of Cd (Cherian et al., 1978; Foulkes andMcMullen, 1986b) (Fig. 2). Retention of Cd by enterocyteswould reduce the amount of Cd entering into systemiccirculation, which in turn would decrease the load of Cddelivered to target organs, such as the liver and kidneys(Min et al., 1991). In support of this hypothesis are dataindicating that the synthesis of MT is induced in the mucosaof the small intestine after enteric exposure to Cd (Cherianet al., 1978; Ouellette et al., 1982). Moreover, Lehman andKlaassen (1986) have shown that approximately 60% of theCd present in the mucosa of the small intestine is bound toMT shortly after enteric exposure to this metal. Further-more, Phillpotts (1984) has demonstrated that a largeamount of Cd taken up by enterocytes after oral adminis-tration of Cd is retained in the mucosa of the small intestine.Despite all these findings, the precise role that MT (as wellas other intracellular ligands) play(s) in the retention andintracellular distribution and trafficking of Cd within en-terocytes remains poorly defined.
Basolateral transport of Cd out of enterocytes
Very little is known about the mechanisms by which Cdis transported across the basolateral membrane of entero-cytes into the capillaries of the lamina propria. Based onrecent findings, however, there is one transporter that hasthe potential for being involved in the basolateral export ofCd from within enterocytes. This transporter is the newlyidentified metal transport protein 1 (MTP1) (Fig. 2), which
is an Fe transporter homologous to DMT1, which has beenidentified in the mouse (Abboud and Haile, 2000). Immu-nohistochemical localization experiments using polyclonalantibodies indicate the presence of MTP1 in the basolateralmembrane and cytoplasm of enterocytes. Moreover, it ap-pears that the expression of MTP1 is regulated by Fe status.Thus, based on these findings and the evidence demonstrat-ing that DMT1 is capable of transporting Cd, it would seemlogical to postulate that MTP1 is a potential transporter ofCd. It should be stressed, however, that a number of factorspertaining to the intracellular ligands that bind Cd insideenterocytes play important roles in how Cd ions can beexported from the cell. Clearly, this is an area that requiresextensive study in the future.
Hepatic handling of Cd
After Cd enters into systemic circulation, from the lungsor intestines, it is delivered to target organs. Much of the Cdabsorbed in the intestines is delivered first to the liver viaportal circulation, where Cd is taken up from the sinusoidalcapillaries by hepatocytes through mechanisms that havenot yet been defined well. Regardless of oral, pulmonary, orparenteral exposure, the liver is by far the primary organthat takes up the greatest quantity of Cd during the initialhours after exposure, especially after parenteral exposure(ATSDR, 1999; Liu et al., 2001; Zalups, 2000a). In fact,Zalups (2000a) has demonstrated recently in rats exposedintravenously to a low dose of Cd (5 �mol/kg), in the formof CdCl2, GSH, or Cys S-conjugates of Cd, that the liverwas so effective in clearing the blood of Cd that less than1% of the dose was remaining in the total blood volume 1 hafter exposure. The amounts of Cd taken up by the liveraccounted for as much as 50–60% of the administered dose.Of the Cd remaining in blood shortly after exposure, ap-proximately 50% is distributed among the cellular constit-uents of blood, mainly in the erythrocytes. The uptake of Cdby erythrocytes appears to be mediated by an anion ex-changer in the plasma membrane (Dawson and Ballatori,1995).
The form of Cd that is delivered into the portal bloodfrom the intestines is likely a mixture of species. Due to thehigh concentration of albumin in the plasma (3–5 g/dL) andthe fact that each molecule of albumin possesses one re-duced sulfhydryl group (Brown and Shockley, 1982), albu-min probably serves as the primary molecule that deliversCd from the sites of entry into systemic circulation to thetarget epithelial cells in organs affected adversely by thismetal (Nordberg and Nordberg, 1988). Cd appears to bindto albumin with an apparent dissociation constant of 10�4
(Rao and Lal, 1958; Verma et al., 1982). Other moleculesthat may serve as carriers of Cd in the blood include low-molecular-weight thiols such as Cys, homoCys (hCys), andGSH, which are in the plasma of blood at low micromolarconcentrations (Lash and Jones, 1985) and are known to
169R.K. Zalups, S. Ahmad / Toxicology and Applied Pharmacology 186 (2003) 163–188

form linear II coordinate covalent complexes with Cd ions(Rabenstein, 1989; Rabenstein et al., 1983). MT and trans-ferrin are other molecules that likely transport Cd in theblood after oral exposure(s) to Cd. On the other hand, it isnot clear presently whether any fraction of Cd in blood isbound to inorganic anions, such as SO4
�2 or PO4�2.
Jonah and Bhattacharyya (1989) have demonstrated thatthe species of Cd entering into systemic circulation shortlyafter low oral doses of Cd changes over time from one thatfavors being taken up in the liver to one that favors beingtaken up in the kidneys. With continuous oral exposure toCd, the ratio of the quantity of Cd in the liver to that in thekidneys drops significantly over time, from about 8–10:1 toas low as 1:1. Some investigators suggest that, over time,Cd-MT ends up being an important chemical form of Cdthat is delivered into the blood from the intestines.
Gompertz et al. (1983) have shown that the concentrationof Cd rises faster in the liver than in the kidneys afterexposure. Cd that is taken up by the liver and kidneys canremain in these organs for as long as several years (ATSDR,1999). As Cd localizes preferentially in the liver, it inducesthe synthesis of MT (mainly MT-I and MT-II) throughmechanisms that are not yet defined fully. This induction ofMT appears to provide hepatocytes with a source of pro-tection from the cytotoxic effects of Cd (Goering and Klaas-sen, 1984). However, upon repeated exposures, hepatocytesmay become overloaded with Cd ions, which can over-whelm the cytoprotective mechanisms within the cells andlead to oxidative stress, lipid peroxidation (Gill et al., 1989;Goering et al., 1995; Hussain et al., 1987; Rikans andYamano, 2000; Sarkar et al., 1995; Shaikh et al., 1999;Stohs and Bagchi, 1995; Stohs et al., 2001), and finallyhepatocellular death (Dudley et al., 1982, 1984). It shouldbe pointed out, however, that, unless individuals have beenexposed to large doses of Cd, hepatic injury in humans isnot commonly associated with either pulmonary or oralexposure to Cd (ATSDR, 1999).
Studies by Kotsonis and Klaassen (1977, 1978) indicatethat the liver accumulates substantial amounts of Cd aftereither acute or chronic exposure to hepatotoxic doses of Cd.Numerous investigators believe that, when hepatocellularnecrosis and/or apoptosis is/are induced by Cd, a significantamount of the metal in the necrotic and/or apoptotic hepa-tocytes is released into hepatic circulation in the form ofCd-MT. Some of this Cd-MT is delivered via systemiccirculation to the kidneys, where it is filtered freely at theglomerular filtration barrier. It is also believed that somefraction of the filtered Cd-MT is taken up via endocytosis bythe epithelial cells lining the proximal convoluted tubule(Foulkes, 1978), which are the primary renal tubular cellsaffected adversely by Cd-MT (Zalups et al., 1992).
While studying the uptake and accumulation of Cd inWRL-68 cells (which are derived from the human liver),Souza et al. (1997) discovered that Cd transport occurred bytemperature-insensitive processes, temperature-sensitiveprocesses (probably ion channels, such as Ca channel), and
carriers that involved interaction with sulfhydryl groups.Previously, Failla et al. (1979) had also suggested that theuptake of Cd in isolated hepatocytes might occur via SH-dependent carrier mechanisms. This conclusion was basedprimarily on their observation that the uptake of Cd inisolated hepatocytes was decreased significantly followingtreatment with N-ethylmaleimide, which is an alkylator ofsulfhydryl groups.
Hepatocellular uptake of Cd from sinusoidal blood
Assuming that the majority of the Cd in sinusoidal bloodis bound mainly to proteins (such as albumin, ferritin, trans-ferrin, �-globulins, and MT) and nonprotein thiols, there areseveral mechanisms that can potentially explain the uptakeof Cd across the sinusoidal membrane of hepatocytes.
First of all, fluid-phase, absorptive, and receptor-medi-ated endocytotic processes need to be considered carefullyas being potential mechanisms involved in the uptake of Cdfrom sinusoidal blood (Fig. 3). Fluid-phase and receptor-mediated endocytotic processes account for a significantamount of fluid uptake and membrane turnover in hepato-cytes, although fluid phase endocytosis may account for agreater level of fluid uptake and membrane turnover thanreceptor-mediated endocytosis (Oka et al., 1989). One of thebetter characterized receptor-mediated, endocytotic pro-cesses in hepatocytes pertains to the uptake of Fe. Most ofthe Fe taken up by hepatocytes involves the endocytosis oftransferrin and ferritin by receptor-mediated processes(Mack et al., 1983; Morgan and Baker, 1986; Osterloh andAisen, 1989). Due to the binding affinity between Fe-bind-ing proteins and other cations, it is possible that Cd maygain entry into hepatocytes via endocytosis mediated by oneof the Fe-binding proteins. As a matter of fact, ferritin hasbeen shown to bind Cd, Zn, Be, and Al. Thus, endocytosisof Cd-ferritin complexes may indeed serve as a route ofentry of Cd into hepatocytes, Joshi et al. (1989) have actu-ally suggested that ferritin may serve as a detoxifying pro-tein due its ability to bind a number of cationic forms ofseveral elements. An additional receptor involved in medi-ating endocytosis that needs to be considered in the hepa-tocellular uptake of Cd is the asialoglycoprotein receptor(Stockert et al., 1980). This receptor mediates the endocy-tosis of substrates such as asialotransferrin. Inasmuch asalbumin is such an abundant and important carrier moleculein blood, there is a distinct possibility that complexes of Cdbound to albumin serve as substrates that are taken up intohepatocytes by endocytosis.
Specific membrane transporters present in the sinusoidalplasma membrane are also potential candidates for playinga role in the uptake of Cd into hepatocytes. One transportprotein that is potentially involved in the hepatic uptake ofCd, at least under certain circumstances, is DMT1 (Fig. 3).Although there has been a paucity of data on the localizationof DMT1 in the liver, Trinder et al. (2000) have recentlyprovided evidence that DMT1 protein is expressed in the
170 R.K. Zalups, S. Ahmad / Toxicology and Applied Pharmacology 186 (2003) 163–188

Fig. 3. A diagrammatic representation of the mechanisms involved in the transport of cadmium at the sinusoidal and canalicular plasma membranes ofhepatocytes. The liver takes up and accumulates more Cd than any other organ, especially following exposure to inorganic salts of Cd. Shortly after exposureto Cd, especially after parental exposure, most of the Cd in blood is bound to plasma proteins, such as albumin and ferritin. Another smaller pool of Cd inblood is likely in the form of conjugates of cysteine (Cys), GSH, and potentially other low-molecular-weight thiols (R-Cd-R). A tremendous level offluid-phase, absorptive, and receptor-mediated endocytosis occurs in hepatocytes. Therefore, endocytosis of one or more protein conjugates of Cd may serveas an important mechanism by which Cd is taken up into hepatocytes. The proton-coupled divalent metal transporter DMT1 is not only expressed inenterocytes, but it also appears to be expressed in the sinusoidal plasma membrane of hepatocytes. Therefore, based on findings showing that DMT1 cantransport Cd ions into enterocytes, this transporter may provide a route of entry for Cd into hepatocytes. Metal transporter protein 1 (MTP1) has also beenlocalized on the sinusoidal plasma membrane. Based on the current knowledge regarding the function of this transporter, it is possible that Cd ions may beexported from within the cytosol of hepatocytes into the sinusoidal blood by MTP1. DMT1 and MTP1 may serve as counterbalance transporters to helpmaintain the hepatocellular homeostasis of Fe. A likely mechanism by which Cd ions gain access into the intracellular milieu of hepatocytes is through Cachannels. Data from cultured hepatocytes indicate that agonists of Ca channels increase the uptake of Cd and antagonists of Ca channels decrease the uptakeof Cd into hepatocytes. Thus, Cd likely serves as an ionic homolog or “mimic” of Ca at the Ca channels in the sinusoidal membrane. Inasmuch as Cd doesnot exist as an unbound ion in the plasma, there must be some form of ligand-exchange reaction that occurs at the site of Ca channels or one of the metalion transporters that allows Cd ions to enter into hepatocytes via one of these transport systems. Another set of transporters that may be involved in uptakeof Cd from the sinusoidal blood are the organic anion and amino acid transporters. Very little information is available on whether these carrier proteins cantransport Cd into hepatocytes. Once Cd enters into hepatocytes, it becomes part of an exchangeable and nonexchangeable pool of intracellular Cd. Much ofthe Cd entering into hepatocytes can serve as a potential source of Cd ions that can induce the transcription of the genes for MT-1 and MT-2. Induction ofMT-1 and MT-2 in hepatocytes is one of the prominent effects detected after exposure to salts of Cd. The large amount of induction of MT in hepatocytesand the incorporation of Cd into MT is believed to serve as a protective mechanism to prevent the direct interactions between Cd ions and critical nucleophilicsites within the cells. However, if the intracellular pool of exchangeable Cd increases beyond what the protective elements inside the hepatocytes can handle,oxidative stress is induced. This in turn can lead to lipid peroxidation in the plasma membrane and the induction of cell death by either necrosis or apoptosis.As hepatocytes undergo cell death, they release Cd, much of which is in the form of Cd-MT, into the sinusoidal and canalicular compartments. The Cd-MTreleased into the sinusoidal blood then can be carried to the kidneys, where it is filtered freely at the glomerular filtration barrier and appears to be taken upby proximal tubular epithelial cells. There are some investigators that believe that the actual nephropathy induced by Cd is due largely to the absorption andsubsequent degradation of Cd-MT by the epithelial cells lining the proximal convoluted tubules. Since hepatocytes are the greatest producers of GSH in thebody, it is likely that a significant amount of Cd ions that enter into hepatocytes interact with the large intracellular pool of GSH. Experimental evidenceindicates that Cd is secreted into the bile by hepatocytes. It appears that Cd is likely transported into the biliary canaliculi as an S-conjugate of GSH. Althoughthe mechanisms responsible for this transport are not known, it is possible that the ATP-binding cassette, multiple drug- resistance protein 2 (MRP2), and/ora GSH-transporter is/are involved. MRP2 is a likely candidate for transporting GSH S-conjugates of Cd since this transporter has been shown to be atransporter of various organic GSH S-conjugates into the biliary compartment. Once GSH S-conjugates of Cd are secreted into the biliary canaliculi, theyare likely broken down rapidly to the corresponding Cys S-conjugates by the actions of �-glutamyltransferase and cysteinylglycinase located on thecanalicular plasma membrane. Therefore, Cys S-conjugates of Cd are likely the primary form of Cd delivered into lumen of the duodenum via the commonbile duct. Pathways depicted by solid black arrows are supported either directly or indirectly by experimental evidence. The thicker solid black lines denotepathways for which there is substantive evidentiary support. By contrast, the dashed lines represent pathways of secondary importance or pathways that arebased primarily on indirect findings or speculation. Question marks associated with a pathway denote uncertainty for the occurrence of that pathway in vivo.

sinusoidal membrane of hepatocytes in rats, during bothcontrol and iron-loading states. However, they found thatvery little DMT1 was expressed in hepatocytes during gen-eralized iron deficiency. Overall, their findings stronglysupport the growing body of literature implicating DMT1 inthe absorptive transport of Fe in certain epithelia. Inasmuchas DMT1 has been localized in the sinusoidal membrane,and since this transporter has been implicated in the uptakeof both Fe and Cd in cultured intestinal (Caco-2 cells;Elisma and Jumarie, 2001; Tallkvist et al., 2001) and distaltubular (Madin–Darby canine kidney (MDCK); Olivi et al.,2001) cells, there appears to be strong support for thehypothesis that DMT1 may play an important role in thehepatocellular uptake of Cd from portal and systemic bloodunder certain homeostatic conditions.
Another means by which Cd may enter into hepatocytesin vivo is through Ca channels (Fig. 3). Blazka and Shaikh(1991) were one of the first groups to hypothesize that Cdions can gain entry into hepatocytes by passing through Cachannels present in the sinusoidal membrane. These inves-tigators demonstrated in vitro that the uptake of Cd ions intohepatocytes, in primary culture, was diminished signifi-cantly in the presence of the Ca channel antagonist vera-pimil or diltiazem. They also showed that the uptake of Cdwas increased significantly in the presence vasopressin,which can act as a Ca channel agonist. Based on thesefindings, the investigators concluded that Cd can enter intohepatocytes through voltage-gated L-type Ca channels.More recently, Souza et al. (1997) have provided additionalin vitro evidence supporting the role of Ca channels in theuptake of Cd in an established human line of hepatic epi-thelial (WRL-68) cells. By using Ca channel antagonists,these investigators demonstrated that approximately one-third of the Cd entry that occurred in the cultured hepato-cytes (exposed to 10 �M Cd) was due to Cd ions enteringthrough Ca channels.
Total body Zn status is clearly an important factor influ-encing the disposition of Cd in target organs. Numerousdata show that increases in cellular contents of Zn mayreduce the absorption and accumulation of Cd and mayprevent or reduce the adverse effects of Cd. By contrast, Zndeficiency has been shown to lead to increased accumula-tion and toxicity of Cd in target organs, such as the liver(Brzoska and Moniuszko-Jakoniuk, 2001; Coyle et al.,2000). Mishima et al. (1997) have documented that theaccumulation of Cd in Chang liver cells decreased signifi-cantly when the cells were pretreated with Zn. In addition,they demonstrated that Zn pretreatment afforded these cul-tured hepatocytes protection from the toxic effects of Cd,perhaps through the actions of Zn-induced MT.
Retention of Cd by hepatocytes
After acute exposure to inorganic forms of Cd, the pre-ponderance of the dose accumulates in the liver (Kotsonisand Klaassen, 1977; Zalups, 1997, 2000a). Accumulation
and retention of Cd in the liver after acute exposure appearsto be related primarily to the hepatocellular content of MTand the ability of MT to bind and sequester Cd in the cytosol(Cherian, 1977, 1980a, 1980b; Goering and Klaassen, 1983;Liu et al., 1995). It should be stressed that Cd is a potentinducer of MT in hepatocytes (Fig. 3). The large amounts ofMT protein produced in hepatocytes after exposure to Cdare likely directly related to the large amounts of Cd takenup the cells. As much as a 15-fold increase in the hepaticconcentration of MT has been documented during the initial24 h following an intraperitoneal injection of Cd (Goering etal., 1985). In hepatocytes, it appears that MT plays animportant protective role against the injurious effects of Cd.This was made evident recently in MT-I/II-knockout mice.These mice were shown to be more sensitive to the hepa-totoxic effects of Cd than corresponding wild-type mice(Habeebu et al., 2000).
Outward transport of Cd into hepatic sinusoids
As is the case in most epithelia, very little is known aboutthe transport of toxic metals out of hepatocytes into sinu-soidal blood. Transporters such as the organic anion trans-porting polypeptides OATP1 and OATP2 (Jacquemin et al.,1994; Kullak-Ublick et al., 1994), the organic cation trans-porter OCT1 (Grundemann et al., 1994), and the amino acidand oligopeptide transporters, which are located in the si-nusoidal plasma membrane, all have the potential for trans-porting metals, such as Cd, into or out of hepatocytes (Fig.3). Another transporter that has been localized recently inthe sinusoidal plasma membrane of hepatocytes is the pu-tative Fe-transporter MTP1 (Abboud and Haile, 2000) (Fig.3). Based on the putative actions of this transporter inenterocytes, and based on its structural and functional sim-ilarities to DMT1, it seems possible that this transporter maybe involved in the efflux of Fe and Cd from within hepa-tocytes. Obviously, significant research is warranted to de-termine the potential roles of these transporters in the he-patic handling of Cd.
Transport of Cd into biliary canaliculi
One of the primary routes of excretion of Cd after acuteparenteral exposure is via the feces (Zalups, 1997, 2000a).In order for Cd to gain access to the luminal compartment ofthe small intestines, the metal must be secreted into thebiliary canaliculi by hepatocytes and then delivered into theduodenum via the common bile duct and/or must be se-creted into the intestinal lumen by currently undefinedmechanism(s). Several lines of in vivo evidence obtainedfrom rats treated with CdCl2 support the hypothesis that atleast some of the Cd that enters into hepatocytes is secretedinto the bile (Cherian, 1977; 1980a, 1980b; Cherian andVostal, 1977; Cikrt and Tichy, 1974; Graf and Sies, 1984;Gregus and Klaassen, 1986). Cherian and Vostal (1977)have in fact demonstrated, in rats treated with CdCl2, that
172 R.K. Zalups, S. Ahmad / Toxicology and Applied Pharmacology 186 (2003) 163–188

Cd was secreted into the biliary compartment 5 h aftertreatment and that the secreted Cd was associated primarilywith a low-molecular-weight compound (less than 4000Da), possibly GSH (Fig. 3). In fact, preliminary evidenceobtained by thin-layer chromatography indicated that the Cdsecreted into the bile was in the form of a GSH conjugate.Hepatocytes contain millimolar concentrations of GSH (Za-lups et al., 1999a, 1999b, 1999c) and are the primary cellsin the body where conjugation reactions with GSH occur(Lash et al., 1995). As a result of the large pool of GSH inhepatocytes, there is a significant probability for GSH tointeract with and bind to Cd that is taken up from thesinusoidal blood. Hepatocytes possess the ability to trans-port effectively a number of conjugates into the canalicularcompartment, including GSH S-conjugates of Cd (Gregusand Klaassen, 1986; Gregus and Varga, 1985) and conju-gates of glucuronides and sulfate. One of the key transport-ers present in canalicular membrane of hepatocytes that isinvolved in the process is the multidrug resistance-associ-ated protein MRP2, also known as canalicular multispecificorganic anion transporter (cMOAT) or canalicular MRP(cMRP) (Fig. 3). There is also a specific transporter of GSHpresent in the canalicular membrane (Ballatori and Dutczak,1994; Fernandez-Checa et al., 1992, 1993) that may trans-port metals bound to GSH (Fig. 3).
In addition to the findings of Cherian (Cherian, 1977,1980a, 1980b; Cherian and Vostal, 1977), there is morerecent evidence implicating the hepatobiliary secretion ofGSH S-conjugates of Cd. This evidence comes mainly fromthe studies with mutant EHBR and GY rats, in which theactivity of MRP2 and ability to secrete GSH S-conjugatesinto the bile are lacking. For example, Sugawara et al.(1996) have demonstrated that biliary secretion of Cd inmutant EHBR rats was only about 2% of that in normal rats.Moreover, Dijkstra et al. (1996) have demonstrated that thesecretion of both GSH and Cd into the biliary compartmentwere negligible in Cd-treated mutant GY rats. Additionalevidence implicating the hepatocellular formation of GSHconjugates of Cd and their subsequent secretion into the bilecomes from the study of Gregus and Varga (1985), whichshowed that the biliary excretion of Cd was reduced signif-icantly by prior depletion of GSH.
Cherian (1980a) has documented that the status of MT inhepatocytes plays a significant role in the secretion of Cdinto the biliary tree. More specifically, it was shown thatbiliary secretion of Cd was reduced greatly by prior expo-sure to Cd and that this reduction in the hepatobiliarysecretion of Cd was related to an increase in the level of MTinduced in the liver by the prior exposure to Cd. Thus, theMT that was induced by Cd appears to have served as amolecular sink trapping the Cd in the cytosolic compart-ment and preventing the Cd ions from interacting with themolecular species and transporters involved in the secretionof Cd into the biliary canaliculi (Fig. 3).
Renal handling of Cd
In humans exposed to Cd via oral and/or pulmonaryroutes, the kidney is by far the primary organ affectedadversely by Cd (ATSDR, 1999). Risk assessment dataindicate that when the renal concentration of Cd begins toexceed 50 �g/g kidney (wet weight) following exposure(s)to Cd, there is a significant risk for the induction of renaltubular injury and impaired renal function. In addition, thereare findings indicating that ingestion of approximately 30�g Cd/day may result in mild forms of renal dysfunction inabout 1% of the adult population, depending on individualvariations in absorption and sensitivity to the toxic effects ofCd (Satarug et al., 2000). Despite all of the studies impli-cating the kidney as a target organ where the toxic effects ofCd are expressed, very little is known about the mechanismsparticipating in the renal handling of Cd.
Factors that have contributed to the difficulty in under-standing the renal handling of Cd include the tremendousstructural and functional heterogeneity that exists along thelength of the nephron and the fact that there are at least twodifferent types of nephrons in most mammals. Based oncurrent deficiencies in the scientific literature, the followingobjectives are deemed to be of paramount importance forgaining a more complete understanding of the renal han-dling of Cd: (1) to determine the precise chemical forms ofCd that exist in the plasma and their ability to be filtered atthe glomerulus following the different types of exposure;(2) to determine which forms of Cd are actually presented tothe luminal and basolateral surfaces of target epithelial cellsin vivo; (3) to determine more precisely which specificsegments of the nephron are involved in the in vivo renaltubular transport and handling of the different forms of Cd;and (4) to determine the molecular and cellular mechanismsinvolved in the transport of Cd at the luminal and basolat-eral membranes of the target epithelial cells involved inhandling this metal.
Some investigators have suggested that, under conditionsof chronic exposure to Cd, complexes of Cd-MT (formed inhepatocytes in response to the uptake of Cd) are releasedfrom necrotic hepatocytes and are delivered (via systemiccirculation) to the kidneys, where it appears they are takenup and induce proximal tubular injury and death (Dudley etal., 1985; Nomiyama et al., 1998; Webb, 1986). Althoughthere is evidence indicating that Cd-MT is delivered to thekidneys and that it induces proximal tubular injury anddeath (Cherian and Nordberg, 1983; Dorian et al., 1992;Felley-Bosco and Diezi, 1987, 1989; Nordberg et al., 1975;Zalups et al., 1992), it is clearly not the only species of Cdthat is delivered to or taken up within the kidneys (Fig. 4).This is particularly evident after an acute pulmonary orparenteral exposure to a nonhepatotoxic dose of a Cd salt,such as CdCl2. There is little doubt that thiol-containingmolecules form conjugates with Cd, and, as suggestedabove, thiol-containing molecules such as albumin, GSH,and/or Cys likely serve as molecular shuttles that deliver Cd
173R.K. Zalups, S. Ahmad / Toxicology and Applied Pharmacology 186 (2003) 163–188

Fig. 4. Diagrammatic representation of mechanisms involved in the luminal and basolateral handling of cadmium (Cd) in proximal tubular epithelial cells.In this scheme, cysteine (Cys), GSH, and albumin S-conjugates Cd, as well as complexes of Cd bound to metallothionein (MT) are shown to be present inthe lumen of the proximal tubule. Although the precise chemical species of Cd that are filtered at the glomerulus are not known currently, these conjugateseither have been shown to be filtered into the proximal tubular lumen or have been implicated in the absorptive luminal transport of Cd. This scheme showsthat a significant fraction of filtered Cd that is bound to proteins (such albumin and/or MT) is transported into the intracellular compartment during theendocytosis of the protein(s) to which the Cd ion(s) is/are bound. Experimental findings obtained with mercuric conjugates of GSH indicate that if GSHS-conjugates of Cd (primarily of the form GSH-Cd-GSH) are filtered into the proximal tubular lumen, they are most likely degraded rapidly in the tubularlumen to cysteinylglycine S-conjugates of Cd (Cys-Gly-Cd- Cys-Gly) by the brush-border enzyme �-glutamyltransferase, and then to the Cys S-conjugateof Cd (Cys- Cd-Cys) by another brush-border enzyme, cysteinylglycinase. The resulting Cys S-conjugates of Cd and the pool of filtered Cys S-conjugatesof Cd may enter the proximal tubular epithelial cells via amino transporters in the luminal plasma membrane. Support for this notion comes from currentevidence indicating that both sodium-dependent and sodium-independent amino transport systems are involved in the luminal uptake of mercuric conjugatesof Cys (Cys-Hg-Cys). Inasmuch as the cysteinyl Cd complexes appear similar in structure to the amino acid cystine or Cys, it is possible that these conjugatesof Cd serve as molecular homologs or “mimics” of cystine or cysteine at the site of one or more of the amino acid transporters involved in the absorptivetransport of these amino acids. Cd may also be taken up at the luminal membrane of proximal tubular epithelial cells by calcium (Ca) channels. Althoughit is highly improbable that Cd exists as an unbound cation in the proximal tubular lumen, Cd ions may be delivered to the binding sites of Ca channels inthe form of thiol- or protein-conjugates, where the Cd ions can undergo a ligand-exchange reaction and bond to the entry site of the Ca channel, where theycan then gain access to the intracellular milieu. It is also possible that some forms of Cd actually “leak” through the junctional complexes into the basolateralcompartment because the intercellular junctions between adjacent proximal tubular epithelial cells are relatively leaky, especially in the early segments ofthe proximal tubule. This may be particularly true when the epithelial cells begin to become intoxicated by Cd. Support for this hypothesis comes fromexperiments showing that Cd can disrupt the intercellular junctions of an immortalized line of porcine proximal tubular epithelial (LLC-PK1) cells in cultureby binding to the extracellular Ca-binding sites of the E-cadherin/catenin network (perhaps as a result of ionic “mimicry”) of the zonula adherens, which isan integral component of the junctional complex. Two sets of transport systems are displayed as being potentially involved in the inward transport of Cd atthe basolateral membrane. One of these is the system of organic anion and dicarboxylate transporters. Experimental evidence indicates that intracellularproduction and reclamation of �- ketoglutarate (�-KG) generates a chemical gradient facilitating the movement of this dicarboxylate out of the cell byexchanging with extracellular organic anions at the site of the organic anion/dicarboxylate exchanger (OAT1). After it is transported out of proximal tubularcells, �-ketoglutarate is taken back up into the cell across the basolateral membrane via a dicarboxylate sym-port involving the cotransport of sodium (Na�).This sym-port is driven by the Na� - gradient generated by the Na� -K� -ATPase localized in the basolateral membrane. Although there are currently littleto no data implicating the role of OAT1 in the basolateral uptake of Cd by proximal tubular epithelial cells, there is a strong body of both in vivo and invitro evidence implicating this transporter in the basolateral uptake of mercuric conjugates of GSH, Cys, and N-acetylcysteine (NAC). Since Cys S-conjugatesof Cd may act as molecular homologs of amino acids, it is possible that these conjugates may gain entry into the proximal tubular epithelial cells via aminoacid transporters located on the basolateral membrane. Current evidence indicates that, when Cd is coadministered to rats with GSH or Cys, there is asignificant increase in the renal basolateral uptake and accumulation of Cd, perhaps as a result of these carrier systems. It is highly unlikely that complexesof Cd-MT are taken up at the basolateral membrane, inasmuch as proximal tubular epithelial cells lack the endocytotic machinery to accommodate such
174 R.K. Zalups, S. Ahmad / Toxicology and Applied Pharmacology 186 (2003) 163–188

to the epithelial cells that take up and transport Cd withinthe kidneys. Support for this hypothesis comes in part fromthe in vivo data of Zalups (2000a), which show that the netrenal uptake and accumulation of Cd increased by at least50% (relative to animals given CdCl2) when Cd was ad-ministered as a conjugate of Cys or GSH.
Intrarenal handling of Cd after exposure to Cd salts orCd-MT
When Cd is injected intravenously (bypassing gastroin-testinal and/or pulmonary routes of exposure) into rats asCdCl2, only a very small percentage of the dose becomeslocalized in the combined renal mass, while between 50 and60% of the dose localizes in the liver (Zalups 2000a).Approximately 1.5–1.8% of the administered dose of Cdaccumulates in the total renal mass during the first hour afterexposure to a 5 or 10 �mol/kg dose of CdCl2 (Zalups,2000a; Zalups and Barfuss, 2002a). During the remainder ofthe first 24 h following intravenous exposure, the renalburden of Cd increases slightly, to no more than about 2%of the dose (Zalups, 1997, 2000a; Zalups and Barfuss,2002a). The majority of Cd that accumulates in the kidneysis localized in segments of nephron situated in the renalcortex and outer stripe of the outer medulla, presumably inproximal tubular segments. Most of these findings are con-sistent with the micropuncture findings of Felley-Bosco andDiezi (1989), which show that when Cd-acetate was infusedinto the circulation of rats; a relatively low level of Cd wasfiltered into the proximal tubular lumen (at the glomerulus)and that a small amount of the Cd that had been filtered wasabsorbed mainly in proximal tubular segments in the cortex.
In another study by Felley-Bosco and Diezi (1987), thein vivo microperfusion technique was used to characterizethe luminal handling of various forms of Cd, which weremicroperfused into the lumen of superficial proximal and/ordistal tubules of the rat nephron. In experiments in which109Cd was microinjected into superficial proximal convo-luted tubules as 109CdCl2, the fractional absorption of Cdwas approximately 70%. By contrast, almost 90% of the Cdmicroinjected (as 109CdCl2) into superficial distal tubules
was recovered in the pelvic urine from the ipsilateral kid-ney, indicating that most of the absorption of Cd ionsoccurred in proximal tubular segments. When 109Cd-MTwas microinjected into the lumen of superficial proximalconvoluted tubules, the fractional uptake of Cd, relative tothe recovery of [3H]-inulin in the pelvic urine, was between17 ((Cd-MT) was 0.19 mM) and 8% ((Cd-MT) was 1.5mM). This finding tends to indicate that Cd-MT is notabsorbed very efficiently along the proximal tubule. Mini-mal absorption of Cd also occurred following proximal ordistal microinjection of a chelate of Cd-pentetic acid. Over-all, these data confirm that the molecular species of Cddelivered to the luminal compartment of both proximal anddistal portions of the nephron affects greatly the absorptivehandling of Cd by the epithelial cells in those segments.
In a more recent study, Zalups (2000a) had demonstratedthat there were both luminal and basolateral mechanismsinvolved in the in vivo renal tubular uptake of Cd in rats(Fig. 4). In addition, the findings from this study showedthat, when Cd was coadministered with Cys or GSH, theuptake of Cd at both luminal and basolateral membranesincreased significantly. These findings indicate that Cys andGSH S-conjugates of Cd may serve as transportable sub-strates and/or may provide an efficient means of deliveringCd to the luminal and basolateral transporters of Cd presentalong the nephron (Fig. 4). Felley-Bosco and Diezi (1987)have also studied the renal tubular transport of Cd when itwas co-perfused with Cys. They demonstrated that the frac-tional absorption of Cd increased (to 82%) along the prox-imal tubule when Cd and Cys were co-microperfused invivo into the lumen of the early proximal convoluted tu-bules of the rat kidney. However, when the ratio of Cd tocysteine was decreased to 1:5–10, the fractional uptake ofCd in the microinjected proximal tubules appeared to dropbelow 60%.
Co-administration of inorganic mercury (Hg) with Cysor GSH has also been shown to cause significant increasesin the renal uptake and net accumulation of Hg during theinitial hour after treatment in both normal rats and rats thathave undergone an acute bilateral ureteral ligation prior tothe injection of Hg (Zalups, 1998a, 1998b; Zalups and
transport. The scheme presented also shows that a pool of Cd accumulates within the proximal tubular epithelial cells from the different sites of entry andthat this pool can interact with various components within the cells. The pool of Cd that accumulates during and after periods of exposure provides a sourceof Cd ions that can induce the transcription of the genes for MT-1 and MT-2, by mechanisms that have not yet been defined fully. Numerous lines of evidenceindicate that expression of renal MT genes, and especially those in proximal tubular epithelial cells, increases markedly after exposure to Cd. If theintracellular pool of exchangeable Cd increases beyond what the protective elements inside the proximal tubular epithelial can handle, oxidative stress isinduced, which in turn can alter mitochondrial respiratory activity and lead to lipid peroxidation in the plasma membrane and other perturbations in cellularmetabolism. All of these effects can, and do, lead to the induction of cell death by either necrosis or apoptosis, which results in the release of Cd from withinthe necrotic or apoptotic proximal tubular epithelial cells into the tubular lumen. One intracellular molecule that protects cells from oxidative injury and bindsCd is GSH. This molecule is the most abundant nonprotein thiol in the body and is present in proximal tubular epithelial cells at concentrations of between3 and 5 mM. Some of the Cd that forms S- conjugates with GSH may be transported into the proximal tubular lumen by the ATP binding cassette proteinMRP2, which has been shown to be expressed in the luminal plasma membrane of proximal tubular epithelial cells and has been shown to transport GSHS-conjugates of various molecules. Pathways depicted by solid black arrows are supported either directly or indirectly by experimental evidence. The thickersolid black lines denote pathways for which there is substantive evidentiary support. By contrast, the dashed lines represent pathways of secondary importanceor pathways that are based primarily on indirect findings or speculation. Question marks associated with a pathway denote uncertainty for the occurrence ofthat pathway in vivo.
175R.K. Zalups, S. Ahmad / Toxicology and Applied Pharmacology 186 (2003) 163–188

Barfuss, 1995b, 1995c, 1998b). The fact that co-adminis-tration of Cys or GSH (in a 4:1 ratio) with Cd or Hg resultsin the increased accumulation of the respective metal in thekidneys leads one to consider the possibility that there maybe common mechanisms involved in some aspects of therenal tubular uptake of Cd and Hg, despite the fact that theoverall disposition of injected Cd in the body is greatlydifferent from that of injected Hg. Since both Cd and Hghave a strong affinity for sulfhydryl groups, especially insmall molecules such as Cys, GSH, and MT, it is notunreasonable to hypothesize that one or more of the mech-anisms participating in the renal tubular uptake of Cd in-volves either the transport of a Cd-thiol complex (such as aCys of GSH S-conjugate of Cd) or some sort of bondinginteraction between the Cd in the Cd-thiol complexes andspecific nucleophilic sites on the luminal and/or basolateralplasma membrane to release Cd to be taken up by anothermechanism, such as a Ca channel. In addition, the basicpremise of this hypothesis does not preclude the potentialfor there being separate independent mechanisms involvedin the renal uptake and accumulation of Cd that are dissim-ilar from those involved in the renal uptake and accumula-tion of Hg.
Intrarenal localization of Cd after exposure to Cd salts orCd-MT
Dorian et al. (1992) have provided light microscopic,autoradiographic findings showing the intrarenal dispositionof 109Cd in mice treated intravenously with 109CdCl2. Fol-lowing treatment, Cd distributed evenly throughout thelengths of the proximal tubules in the renal cortex and outermedulla. At a lower dose (0.1 mg Cd/kg), Cd was localizedin both proximal and distal segments of the nephron. Afteradministration of a large dose (3 mg Cd/kg), Cd becamelocalized mainly in proximal tubular segments, but the con-centration of Cd in the epithelial cells of both pars convo-luta and recta segments of proximal tubules appeared to besimilar, as well as being distributed evenly throughout ofcytoplasm.
In this same study, Dorian et al. (1992) also studied therenal disposition of 109Cd in mice treated intravenously with109Cd-MT. Their findings showed 109Cd became localizedalmost exclusively in the epithelial cells lining the S1 and S2
segments of the proximal tubule situated in the renal cortex.The relative concentration of 109Cd in the basal and apicalportions of the affected cells was similar when a nonneph-rotoxic dose of Cd-MT was injected. By contrast, 109Cd wasdistributed mainly in the apical portions of the cells when anephrotoxic dose had been administered. The findings fromthis study also showed that the renal burden of Cd increasedrapidly until approximately 85% of the administered dose ofCd was present in the kidneys. Subsequently, the renalcontent of Cd remained constant for up to 7 days aftertreatment. The data from this study support the hypothesisthat the nephropathy induced by Cd-MT may be due (at
least in part) to the preferential uptake of Cd-MT by theepithelial cells lining the S1 and S2 (convoluted) segmentsof the proximal tubule, presumably by an endocytotic mech-anism.
Filterability of Cd at the renal glomerulus
In order for Cd to be taken up from the luminal com-partment of the nephron, it has to first pass through theglomerular filter and/or be secreted by one or more of thedifferent tubular epithelial cells. Therefore, it is of greatimportance to determine and to gain a thorough understand-ing of the molecular species of Cd that are filtered at theglomerulus following the different types of exposure.
Currently, there is evidence indicating that when Cd-MToriginates from the intestines or liver (or parenteral admin-istration) it is delivered to the glomerular filtration barrier ofcortical or juxtamedullary nephrons, where it is filtered at alevel proportional to the filtration fraction, which in humansis about 20% of the delivered load. The ability of Cd-MT tofilter freely at the glomerulus is largely a factor of its smallsize (a molecular mass of approximately 6–7 kDa). Gener-ally, molecules having a molecular weight of less than 66kDa can pass through the glomerular filtration barrier. Thefraction of Cd-MT that is not filtered and is present in theefferent ends of the glomerular capillaries is delivered to theperitubular and vasa recta capillaries via the efferent arte-riole. These capillaries course subadjacently along the entirelength of the nephron (associated with the glomerulus fromwhich the vessels arose) and provide a compartment wheresolutes (including Cd) can be added to and/or taken from theblood. The amount of Cd-MT remaining in the blood at thedistal end of the peritubular capillary network is then shut-tled out of the renal parenchyma and is added to systemiccirculation via venous circulation.
Unlike the unimpeded glomerular filtration of Cd-MT,only a small percentage of the Cd (in plasma) that is boundto albumin is likely filtered into lumen of the proximaltubule, mainly because the glomerular-sieving coefficientfor albumin is very low. On the other hand, when Cd isbound to nonprotein thiols, such as Cys and/or GSH, thecomplexes formed can be assumed to be freely filterable atthe glomerulus (Fig. 4). What is presently unclear, however,is the actual amount, or percentage, of Cd in blood that isbound to low-molecular ligands, such as Cys and GSH, aftervarious types of exposures to Cd.
The only study in which data on the filterability of Cd (atthe renal glomerulus) have been obtained is the micropunc-ture study of Felley-Bosco and Diezi (1989). In this study,it was demonstrated that the ratio of the concentration of Cdin the ultrafiltrate in Bowman’s space to the concentrationof Cd in plasma was 0.2 during infusion of Cd-acetate and1.0 during infusion of Cd-dithiopentetic acid (DTPA).These findings indicate that only about 20% of the Cd thatis infused in the form of Cd-acetate is filterable at theglomerulus, while 100% of the Cd in the form of Cd-DTPA
176 R.K. Zalups, S. Ahmad / Toxicology and Applied Pharmacology 186 (2003) 163–188

is freely filterable. The findings obtained with Cd-acetateare probably more representative of the glomerular filtrationof Cd in the form of inorganic salts of Cd, while the findingsobtained with Cd-DTPA likely reflect the glomerular han-dling of Cd-MT or Cd S-conjugates of Cys or GSH. It wasalso shown that many of the Cd ions that were ultrafilteredinto the lumen of superficial nephrons were taken up mainlyby the pars convoluta of proximal tubules. No transepithe-lial movement of Cd-DTPA was detected, likely because ofthe polar nature of DTPA.
Luminal uptake and secretion of Cd along proximalsegments of the nephron
Using the isolated perfused tubule technique, Robinsonand colleagues (1993) discovered that all three segments(i.e., S1, S2, and S3 segments) of the rabbit proximal tubuleabsorbed Cd avidly when Cd ions were perfused through thelumen in the form of CdCl2. S1 segments of the proximaltubule accumulated Cd more rapidly and developed a moresevere form of tubular injury (at concentrations greater than500 �M) than the other two segments. These findings areconsistent with the histopathological documentation that thepars convoluta of the proximal tubule is the most sensitivesegment of the nephron to the toxic effects of Cd. In addi-tion, under the conditions studied, saturation of absorptivetransport could not be demonstrated in S1, but was demon-strable in S2 and S3 segments. Their findings also showedthat only about 10% of the Cd taken up from the lumen wastransported across the basolateral membrane, indicating that90% of the absorbed Cd was retained in the proximaltubular epithelial cells.
Endo and colleagues (Endo, 2002; Endo et al., 1997,1998a, 1998b, 1998c, 1998d, 1999) have suggested thatuptake of Cd in culture, by an immortalized line of highlytransformed porcine proximal tubular (LLC-PK1) cells ex-posed to CdCl2, occurs through a mechanism that is bothtemperature and pH dependent and can be inhibited byZnCl2 or CuCl2. They have also suggested that Cd is takenup at the apical plasma membrane, in part, by an inorganicanion-exchanger. Moreover, as a result of efflux experi-ments in the presence of tetraethylammonium (TEA), theseinvestigators have suggested that Cd is secreted from withinLLC-PK1 cells, across the apical plasma membrane, by aproton-driven antiport (H�/Cd2�) of the organic cationtransport system (Fig. 4). Similar findings apparently havebeen obtained using luminal membrane vesicles isolatedfrom the kidneys and small intestine of rats.
Although the aforementioned findings of Robinson et al.(1993) and Endo and colleagues (Endo, 2002; Endo et al.,1997, 1998a, 1998b, 1998c, 1998d, 1999) provide somepotential insights into the proximal tubular transport andhandling of Cd ions, it is highly improbable that Cd isfiltered and delivered into the luminal compartment of prox-imal tubules as CdCl2 in vivo. It is more likely that Cd ispresented to the luminal membrane of proximal tubular
epithelial cells as some thiol-conjugate(s), including Cdconjugates of albumin and other proteins. Therefore, theactual biological (as well as clinical) significance of theaforementioned findings is yet to be determined.
As has been alluded to above, much of the Cd present inplasma, shortly after exposure to inorganic forms of themetal, is likely bound to albumin. Cd is probably bound tothe sulfur atom of the single reduced sulfhydryl grouppresent on the molecule. Therefore, it is likely that com-plexes of albumin- Cd are delivered to and interact with theglomerular filter. Due to the fact the filtered load of albumincan be as great as 9 g/day (despite the low sieving coeffi-cient for albumin at the renal glomerular filtration barrier)(Maack, 1992), it would seem reasonable to predict thatsome level of ultrafiltration of Cd conjugates of albuminoccurs, perhaps at a level proportional to the concentrationof Cd-albumin complexes in plasma. Moreover, inasmuchas the majority of filtered albumin is normally absorbed byendocytosis along the early portions of the proximal con-voluted tubule (Maack, 1992), it seems highly probable thatfiltered albumin conjugates of Cd are also taken up by thesesegments of the nephron during the endocytosis of albumin(Fig. 4).
Assuming that some level of ultrafiltration of Cd-albu-min complexes occurs, some of the Cd may dissociate fromalbumin, at higher affinity binding or transport sites on theluminal membrane, prior to the Cd being taken up withalbumin. Even though the ultrafiltration of Cd bound toalbumin seems probable, the predicted filtered load of Cd inthis form would likely be very low due to the rapid extrac-tion of Cd from the plasma by the liver. In one respect, thisis consistent with the very low renal burden of Cd that hasbeen detected in rats after parenteral exposure to CdCl2(Zalups, 1997, 2000a; Zalups and Barfuss, 2002a). Obvi-ously, a great deal of research is needed to define the speciesof Cd filtered at the glomerulus and the intraluminal han-dling of Cd bound to various substrates.
As has been stated above, Cd-MT is another form of Cdthat is filtered at the glomerulus (Fig. 4). Due to the size ofthis complex, the only ways that Cd could enter into apicalcompartment of proximal tubular epithelial cells while in-corporated in MT is either by the endocytosis of the Cd-MTcomplex or by an absorptive mechanism involving Cd thatdissociates from MT in the proximal tubular lumen. Thelatter possibility seems unlikely because each Cd ion in ametallothionein molecule is coordinately bonded to foursulfur atoms and these coordinate bonds are stable at the pHof the luminal fluid in the pars convoluta (which is similarto that in plasma). Indeed, most of the experimental evi-dence indicates that some level of absorptive endocytosis offiltered Cd-MT occurs in vivo in proximal tubular segments(Felley-Bosco and Diezi, 1987; Nomiyama and Foulkes,1977) (Fig. 4). Some investigators have postulated that thenephropathy induced by chronic exposure to Cd is linked tothe endocytosis of Cd-MT (Cherian et al., 1976; Dudley etal., 1985), which appears to arise from necrotic or apoptotic
177R.K. Zalups, S. Ahmad / Toxicology and Applied Pharmacology 186 (2003) 163–188

hepatocytes intoxicated by Cd (Dudley et al., 1985; Tanakaet al., 1975). In support of this hypothesis, Nomiyama et al.(1998) have provided data showing a relationship betweenincreased plasma levels of Cd-MT and hepatic injury in-duced in rabbits treated chronically with a daily dose of Cd(0.3 mg Cd/kg sc). Perhaps more importantly, they notedthat this relationship correlated with levels of induced renaldysfunction. It should be kept in mind, however, that cir-culating Cd-MT arising from intestinal sources (Cherian,1979; Cherian et al., 1978; Sugawara and Sugawara, 1991)also likely contributes to the proximal tubular nephropathyassociated with oral exposure to Cd.
It is interesting, however, that findings from earlier stud-ies by Zalups et al. (1995) and Liu et al. (1994) have shownthat the rates of uptake of metals in the form of metal-MTcomplexes at the luminal membrane ranges from low tonegligible in in vitro preparations of proximal tubules ortheir epithelial cells. Moreover, Felley-Bosco and Diezi(1987) have shown that, when 109Cd-MT was microinjectedinto the lumen of superficial proximal tubules, the fractionaluptake of Cd was very low (8–17%). This finding tends toindicate that Cd-MT is not absorbed very efficiently alongthe proximal tubule in vivo. Furthermore, Ishido et al.(1999) have shown that Cd-MT induces apoptosis in thekidneys of rats, but not in cultured LLC-PK, cells, whichadds to the significant body of findings indicating that themechanisms involved in the endocytosis of Cd-MT in prox-imal tubular epithelial cells in vivo have not yet been char-acterized fully. It is also interesting that recent data of Liuet al. (1998a) tend to indicate that the acute administrationof Cd-MT may not be an ideal model system to study thenephropathy induced by chronic exposure to Cd salts. Over-all, these data indicate that the in vivo endocytosis ofCd-MT complexes likely involves factors that have not yetbeen identified.
The recent findings of Zalups (2000a) showing that therenal uptake and accumulation of Cd were increased whenCd was coadministered intravenously with Cys or GSH,tend to indicate that there is at least one mechanism, otherthan endocytosis, that is involved in the luminal absorptionof Cd along the nephron. Since Cd and Hg are both groupIIB metals, it is possible that Cd may enter into proximaltubular epithelial cells by one or more of the same mecha-nisms involved in the proximal tubular uptake of Hg. Forexample, experimental findings from the laboratories ofZalups and Barfuss (Zalups, 1995; Zalups and Barfuss,1998b; Cannon et al., 2000, 2001) demonstrate that specificamino acid transporters are the primary mechanisms respon-sible for transporting mercuric conjugates of Cys (i.e., Cys-Hg-Cys) across the luminal plasma membrane into proximaltubular epithelial cells. All of their data gathered to dateindicate that the absorption Cys-Hg-Cys occurs via a mech-anism involving molecular “mimicry” or homology, inwhich the mercuric Cys-Hg-Cys acts as a molecular ho-molog or “mimic” of the amino acid cystine and/or Cys atthe site of the amino acid carrier proteins (such as systems
ASC, b0,�, and/or L) involved in the luminal absorption ofthese amino acids. Based on the findings from these studies,it seems possible that Cys S-conjugates of Cd may also betransported by one or more of these same transporters (Fig.4). One should also consider that GSH S-conjugates of Cdthat enter into the early portions of the proximal tubule areprobably degraded rapidly to Cys conjugates of Cd via theactions of the brush-border enzymes �-gutamyltransferaseand cysteinylglycinase, which have been shown to degrademercuric conjugates of GSH to the corresponding mercuricconjugates of Cys in proximal tubules (reviewed in Zalupsand Lash, 1994; Zalups 2000b) (Fig. 4).
Absorptive transport of Cd in distal segments of thenephron
Up until the autoradiographic findings of Dorian et al.(1992), most studies on the renal disposition and transportof Cd have focused on the uptake of Cd by proximal tubularsegments of the nephron. The findings of Dorian et al.(1992) have served as one of the first lines of evidencesupporting the possibility that Cd may be taken up at sitesbeyond the proximal tubule. As a result of these findings,and even more recent ones provided by other investigators,there is a growing body of evidence implicating distal por-tions of the nephron in the renal handling of Cd (Felley-Bosco and Diezi, 1987; Ferguson et al., 2001; Friedman andGesek, 1994; Olivi et al., 2001). Therefore, one needs toconsider the mechanisms by which Cd might gain entry intothose cells, whether it is from the luminal and/or basolateralsurface(s). Potential candidates for transporters involved inthe luminal uptake of Cd in distal segments of the nephroninclude DMT1 and Ca channels (Fig. 5), which have beenlocalized in epithelial cells of distal portions of the nephronand collecting duct.
Some of the evidence supporting the potential handlingof Cd along the distal nephron comes indirectly from theassessment of the renal tubular handling of Fe and thepotential role of DMT1 in that transport. Using anesthetizedrats, Wareing et al. (2000) studied the handling of Fe thathad been microinjected into proximal or distal convolutedtubules. Approximately 19 and 46% of the microinjected55Fe was recovered in the urine following microinjectioninto the surface convolutions of proximal tubules and distaltubules, respectively. The investigators concluded that Fe istransported mainly along ascending thick limbs of the loopof Henle. Using the microperfusion technique, this samegroup of investigators later confirmed that ascending thicklimbs of the loop of Henle do indeed absorb Fe (approxi-mately 50% of the perfused load). They also found thataddition of Cu, but not Zn, to the luminal perfusate de-creased the rate of absorption of Fe, which indicated that Cumay compete with Fe, but not Zn, at a luminal transporter inthe ascending thick limb. Subsequently, these investigatorscarried out experiments to localize DMT1 in the kidneys ofrats using an anti-DMT-1 polyclonal antibody (Ferguson et
178 R.K. Zalups, S. Ahmad / Toxicology and Applied Pharmacology 186 (2003) 163–188
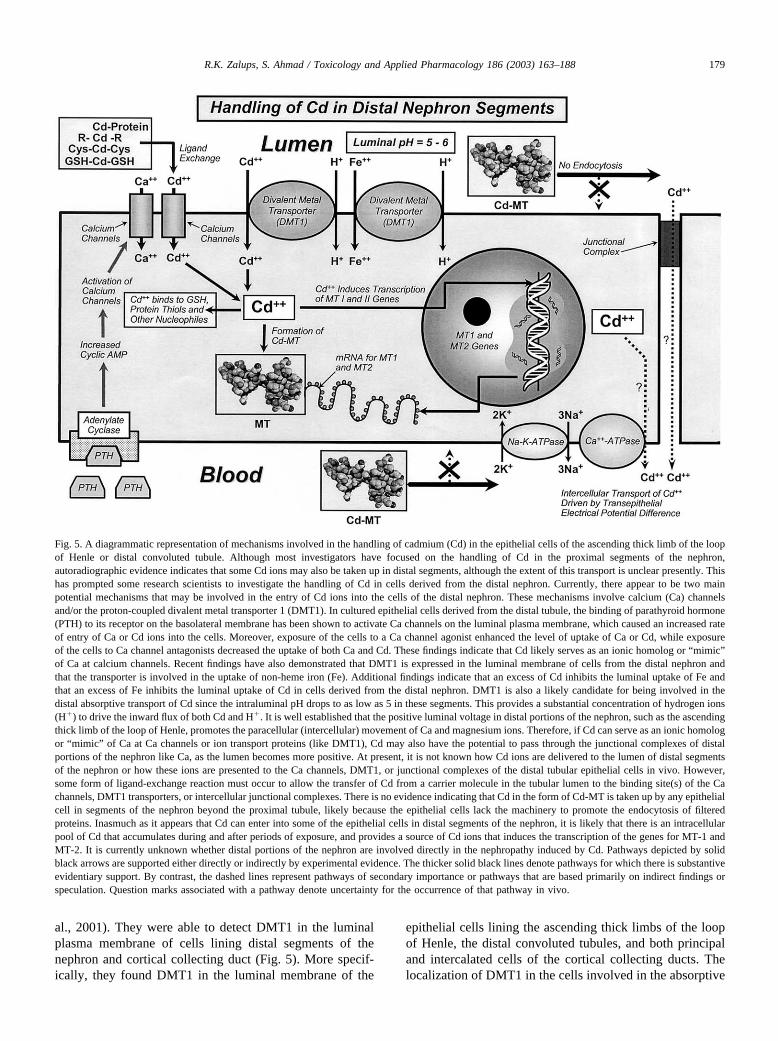
al., 2001). They were able to detect DMT1 in the luminalplasma membrane of cells lining distal segments of thenephron and cortical collecting duct (Fig. 5). More specif-ically, they found DMT1 in the luminal membrane of the
epithelial cells lining the ascending thick limbs of the loopof Henle, the distal convoluted tubules, and both principaland intercalated cells of the cortical collecting ducts. Thelocalization of DMT1 in the cells involved in the absorptive
Fig. 5. A diagrammatic representation of mechanisms involved in the handling of cadmium (Cd) in the epithelial cells of the ascending thick limb of the loopof Henle or distal convoluted tubule. Although most investigators have focused on the handling of Cd in the proximal segments of the nephron,autoradiographic evidence indicates that some Cd ions may also be taken up in distal segments, although the extent of this transport is unclear presently. Thishas prompted some research scientists to investigate the handling of Cd in cells derived from the distal nephron. Currently, there appear to be two mainpotential mechanisms that may be involved in the entry of Cd ions into the cells of the distal nephron. These mechanisms involve calcium (Ca) channelsand/or the proton-coupled divalent metal transporter 1 (DMT1). In cultured epithelial cells derived from the distal tubule, the binding of parathyroid hormone(PTH) to its receptor on the basolateral membrane has been shown to activate Ca channels on the luminal plasma membrane, which caused an increased rateof entry of Ca or Cd ions into the cells. Moreover, exposure of the cells to a Ca channel agonist enhanced the level of uptake of Ca or Cd, while exposureof the cells to Ca channel antagonists decreased the uptake of both Ca and Cd. These findings indicate that Cd likely serves as an ionic homolog or “mimic”of Ca at calcium channels. Recent findings have also demonstrated that DMT1 is expressed in the luminal membrane of cells from the distal nephron andthat the transporter is involved in the uptake of non-heme iron (Fe). Additional findings indicate that an excess of Cd inhibits the luminal uptake of Fe andthat an excess of Fe inhibits the luminal uptake of Cd in cells derived from the distal nephron. DMT1 is also a likely candidate for being involved in thedistal absorptive transport of Cd since the intraluminal pH drops to as low as 5 in these segments. This provides a substantial concentration of hydrogen ions(H�) to drive the inward flux of both Cd and H�. It is well established that the positive luminal voltage in distal portions of the nephron, such as the ascendingthick limb of the loop of Henle, promotes the paracellular (intercellular) movement of Ca and magnesium ions. Therefore, if Cd can serve as an ionic homologor “mimic” of Ca at Ca channels or ion transport proteins (like DMT1), Cd may also have the potential to pass through the junctional complexes of distalportions of the nephron like Ca, as the lumen becomes more positive. At present, it is not known how Cd ions are delivered to the lumen of distal segmentsof the nephron or how these ions are presented to the Ca channels, DMT1, or junctional complexes of the distal tubular epithelial cells in vivo. However,some form of ligand-exchange reaction must occur to allow the transfer of Cd from a carrier molecule in the tubular lumen to the binding site(s) of the Cachannels, DMT1 transporters, or intercellular junctional complexes. There is no evidence indicating that Cd in the form of Cd-MT is taken up by any epithelialcell in segments of the nephron beyond the proximal tubule, likely because the epithelial cells lack the machinery to promote the endocytosis of filteredproteins. Inasmuch as it appears that Cd can enter into some of the epithelial cells in distal segments of the nephron, it is likely that there is an intracellularpool of Cd that accumulates during and after periods of exposure, and provides a source of Cd ions that induces the transcription of the genes for MT-1 andMT-2. It is currently unknown whether distal portions of the nephron are involved directly in the nephropathy induced by Cd. Pathways depicted by solidblack arrows are supported either directly or indirectly by experimental evidence. The thicker solid black lines denote pathways for which there is substantiveevidentiary support. By contrast, the dashed lines represent pathways of secondary importance or pathways that are based primarily on indirect findings orspeculation. Question marks associated with a pathway denote uncertainty for the occurrence of that pathway in vivo.
179R.K. Zalups, S. Ahmad / Toxicology and Applied Pharmacology 186 (2003) 163–188

transport of Fe not only provides a molecular mechanismfor that transport but also provides a mechanism for theputative transport of Cd in these regions of the nephron,mainly because of the growing body of evidence implicat-ing both Cd and Fe as substrates that compete for DMT1.These investigators did not, however, find evidence forDMT1 being localized on the luminal plasma membrane ofproximal tubular epithelial cells, although they did findevidence for DMT1 in their cytoplasm. By contrast, Can-onne-Hergaux and Gros (2002) were able to provide evi-dence, with polyclonal anti-DMT1 antibodies, that DMT1was expressed in the brush-border membrane of proximaltubular epithelial cells in the kidneys of mice. Their findingsalso indicate that dietary Fe did not greatly affect the ex-pression of DMT1 in proximal tubular cells as it does inenterocytes, and they concluded that the regulation of theexpression of DMT1 in the kidneys is different from that inthe intestines. The disparity between the findings of Can-onne-Hergaux and Gros (2002) and Ferguson et al. (2001)may reflect differences in the renal expression of DMT1 andperhaps in the manner in which certain divalent cations arehandled among different species.
Other than the findings of Dorian et al. (1992), the onlysubstantive line of in vivo evidence implicating distal ab-sorption of Cd in the kidneys comes from the microinjectionstudy of Felley-Bosco and Diezi (1987). Their findingsshowed that a small amount of microinjected Cd (less than10%) is apparently absorbed in distal portions of thenephron. Most of the other evidence implicating the absorp-tive transport of Cd in epithelial cells of the distal nephroncomes from cultured epithelial cells derived from distalportions of the nephron. Using an immortalized line of cellsderived from cortical ascending thick limbs of the loop ofHenle and distal convoluted tubules from the kidneys ofmice, Friedman and Gesek (1994) were one of the firstgroups to provide a detailed characterization of the handlingof Cd in a line of cells derived from distal portions of thenephron. Their findings tend to implicate the roles of bothCa channels and a membrane transport protein that can beinhibited by Fe (presumably DMT1). More specifically, thefindings pertaining to Ca channels show that stimulation ofthe cultured cells with parathyroid hormone (PTH) resultedin a marked increase in the luminal uptake of Cd (Fig. 5),which could be inhibited by the Ca channel antagonistnifedipine and could be enhanced by using the Ca channelagonist BAY K 8644. Interestingly, use of nifedipine orBAY K8644 did not affect the uptake of Cd in cells notexposed to PTH, which indicates that a mechanism otherthan Ca channels is likely responsible for the uptake of Cdunder those conditions. The transport protein that is mostlikely responsible for the uptake of Cd in the cells notexposed to PTH is DMT1, inasmuch as Fe was found to bea potent cationic inhibitor of the uptake of Cd and sinceDMT1 has been detected in the luminal membrane of theepithelial cells lining the ascending thick limbs of the loop
of Henle and the distal convoluted tubule (Ferguson et al.,2001).
Absorptive transport of Cd has also been documented ina line of immortalized canine renal epithelial cells derivedfrom the distal nephron (i.e., MDCK cells). Olivi et al.(2001) have demonstrated recently that the uptake of bothCd and Fe by MDCK cells could be altered significantly bychanging membrane potential. They also found that Fecould inhibit the uptake of Cd and that Cd could inhibit theuptake Fe in these cells. Moreover, the uptake of bothmetals was increased at low pH, which generates a strongerproton gradient to drive the proton-coupled DMT1 symport.More direct evidence implicating DMT1 in the transport ofFe and Cd has been provided by using a line of human renalfibroblasts (HEK-293) that overexpress DMT1. HEK-293cells that overexpressed DMT1 demonstrated greater ratesof uptake of Cd. In addition, activation of protein kinase Cresulted in both an increase in the uptake of Cd and Fe inMDCK cells and an increase in the cellular content ofmRNA for DMT1. Collectively, the findings of Felley-Bosco and Diezi (1987), Dorian et al. (1992), Friedman andGesek (1994), Ferguson et al. (2001), and Olivi et al. (2001)provide sufficient evidence indicating that some level ofabsorptive uptake of Cd likely occurs in distal portions ofthe nephron under certain conditions and that DMT1 and Cachannels are involved that process (Fig. 5). Moreover, sincethe pH of the tubular fluid decreases along distal portions ofthe nephron (to about pH 5–6), as a result of normal acid-ification processes, the elevated concentration of H� in thetubular fluid of distal portions of the nephron likely pro-motes the activity of DMT1 in those segments (Fig. 5).
It is not obvious at present whether the absorption of Cdin distal segments of nephron has a significant impact on thenephropathy induced by Cd.
Retention of Cd within proximal tubular epithelial cells
Once Cd ions gain entry into the cytosolic compartmentof proximal tubular epithelial cells, one would predict thatthey would tend to bind to intracellular protein-thiols,largely because of their abundance in the intracellular mi-lieu. Recent findings obtained from LLC-PK cells exposedto Cd indicate that the Cd ions taken up across the luminalmembrane were bound primarily to cellular proteins shortlyafter exposure, as might have been predicted (Felley-Boscoand Diezi, 1991). Within 3 h, the intracellular burden of Cdwas shown to be associated with a low-molecular-weightpool of molecules. It is likely that MT was one of theprimary ligands in that pool (Fig. 4), in that significantinduction of MT was demonstrated in the LLC-PK1 cellswithin 3–6 h of exposure.
In some in vitro preparations (such as in isolated per-fused tubules (Robinson et al., 1993)), however, it has beendifficult to get a precise assessment of the intracellulardistribution of Cd in proximal tubular cells. This is espe-cially the case when acid-precipitation methods are used to
180 R.K. Zalups, S. Ahmad / Toxicology and Applied Pharmacology 186 (2003) 163–188

isolate and separate cellular proteins, since a significantamount of Cd can be liberated from cellular proteins andadded to an acid-soluble compartment during the extractionand isolation of proteins. Such events make it impossible togain an accurate representation of the intracellular localiza-tion of Cd in renal or other epithelial cells.
Secretion of Cd into the tubular lumen
As mentioned above, Endo and colleagues (Endo, 2002;Endo et al., 1997, 1998a, 1998b, 1998c, 1998d, 1999), havepostulated that, when LLC-PK1 cells are exposed to CdCl2,they have the ability to secrete Cd at the apical plasmamembrane by a proton-driven H�/Cd2� exchanger. It is notclear, though, whether such a mechanism is functional inproximal tubular epithelial cells under in vivo conditions,especially when considering that the urinary excretion of Cdis extremely low after acute exposures to Cd salts.
If there is some secretion of Cd along proximal tubularcells in vivo, the activity of MRP2 needs to be considered asa potential mechanism (Fig. 4). It would seem that proximaltubular epithelial cells possess the potential to secrete GSHS-conjugates of Cd into the lumen since these epithelialcells contain high intracellular concentrations of GSH (Za-lups and Lash, 1990; Zalups et al., 1999a, 1999b, 1999c),and MRP2, which has been is localized in the luminalmembrane proximal tubular epithelial cells, has been shownto transport various GSH S-conjugates (Schaub et al., 1997,1999). Further studies are clearly needed to address thishypothesis.
Basolateral uptake of Cd along the nephron
One of the first investigators to suggest that there may bea basolateral mechanism in the renal uptake of Cd isFoulkes (1974). His suggestion was based on indirect find-ings from single-pass experiments in the rabbit. Subse-quently, Diamond and colleagues (1986) provided in vitroevidence for a basolateral mechanism using the isolatedperfused rat kidney. Specifically, they showed that occlu-sion of the ureter did not decrease significantly the netaccumulation Cd in isolated kidneys perfused in vitro with1 �M CdCl2 in a protein-free buffer.
In a more recent study, Liu et al. (1994) have demon-strated that, when LLC-PK1 cells were exposed to CdCl2 orCd-MT at their basolateral membrane, there was a signifi-cant level of association of Cd with these cells. A number ofother studies have also examined basolateral handling of Cdin vitro using LLC-PK1 cells grown on a permeable mem-brane insert (Bruggeman et al., 1992; Kimura et al., 1996;Prozialeck, 2000; Prozialeck et al., 1993; Prozialeck andLamar, 1993). The findings from some of these studiesindicate that the level of association of Cd is greater in thesecells following basolateral exposure to Cd than luminalexposure Cd. Regrettably, some these findings are cloudedby the occurrence of cellular intoxication induced by the Cd
(Prozialeck et al., 1993; Prozialeck and Lamar, 1993). Al-though their findings tend to indicate that Cd can bind to thebasolateral surface of, and/or be taken up into, the culturedcells, it is not clear whether the findings reflect the mannerby which cadmium is taken up at the basolateral membranein vivo.
Zalups (2000a) has provided strong in vivo evidenceindicating that basolateral uptake of Cd does occur in thekidneys. The findings show that a significant level of renaluptake and accumulation of Cd occurred in rats treated withmannitol and bilateral ureteral ligation to reduce glomularfiltration rate to negligible levels. In fact, the renal burden ofCd in these animals was as much as 70–80% of that incontrol rats. These data, therefore, lead one to believe thatbetween 70 and 80% of the acute renal uptake and accu-mulation of Cd that occurs in the kidneys of rats, followingexposure to CdCl2, is likely due to one or more basolateralmechanism(s). Zalups (2000a) has also demonstrated that,when a low dose of CdCl2 was co- administered with Cys orGSH, both luminal and basolateral uptake of Cd increasedby at least 50%. These findings indicate that Cys or GSHS-conjugates of Cd are likely transportable substrates at thebasolateral membrane and/or they provide a more efficientmeans of delivering Cd to the site(s) of the transporters thattake up Cd (Fig. 4). Although it appears that basolateraluptake of Cd occurs in both the renal cortex and outer stripeof the outer medulla, it is unclear which specific segments ofthe nephron are involved in this process. However, since thethree segments of the proximal tubule are present in thesetwo zones of the kidney, they would appear to be logicalchoices for initial investigations.
In a series of recent studies, Zalups and colleagues haveprovided an extensive amount of both in vivo and in vitrodata implicating the classical p-aminohippurate-sensitiveorganic anion transport system and, more specifically, theorganic anion exchanger (OAT1) in the basolateral uptakeof mercuric conjugates of Cys, N-acetylcysteine (NAC), andGSH (Aslamkhan et al., 2002, 2003; Zalups; 1995, 1998a,1998b; Zalups and Barfuss, 1995a, 1995b, 1998a, 1998b,2002b; Zalups and Minor, 1995). Due to the fact that bothCd and Hg are group IIB metals that have a strong affinityfor SH-groups and that the basolateral uptake of both metalsapparently increases when they are bonded to Cys or GSH,it is quite reasonable to postulate that the organic aniontransport system may be involved in the basolateral uptakeof Cys or GSH S- conjugates of Cd along the three segmentsof the proximal tubule (Fig. 4). However, there is also thedistinct possibility that one or more completely differentmechanisms, not involving the organic anion transporters,are involved in the basolateral uptake of Cd.
Elimination and excretion of cadmium
Cd that is absorbed enterically is eliminated very slowlyby urinary and fecal excretory mechanisms. The estimated
181R.K. Zalups, S. Ahmad / Toxicology and Applied Pharmacology 186 (2003) 163–188

half-life of absorbed Cd in mice, rats, rabbits, and monkeyshas been calculated to be from about several months up toseveral years (Kjellstorm and Nordberg, 1985). It has beenobserved that the urinary and fecal excretion of absorbed Cdin animals is approximately equal (Kjellstrom and Nord-berg, 1978), although fecal excretion predominates afterparenteral exposure to inorganic salts of Cd and urinaryexcretion of Cd predominates after parenteral exposure toCd-MT. Findings from animal experiments indicate that theurinary excretion of Cd increases when Cd-induced renaltubular damage occurs (Friberg, 1984). The most likelyexplanation for this response is that Cd is added to theluminal compartment of nephrons from necrotic or apopto-tic proximal tubular epithelial cells (Zalups et al., 1992).
Biliary and fecal excretion of Cd
Following oral/gastrointestinal exposure to Cd, it is ob-vious that the large fractions of Cd, which are not absorbedenterically (greater than 90% of the dose in most mammals),pass through the lumen of both the small and large intestinesand are excreted in the feces (Fig. 1). It is for this reason thatNordberg and Nordberg (1988) have suggested that fecalexcretion can be used as an indicator of oral exposure to Cd.Interestingly, though, fecal excretion is also the principalroute for the elimination of Cd following parenteral expo-sures to CdCl2 (Zalups, 1997, 2000a). As much as 20% ofthe dose is excreted in the feces by rats within 3 to 4 dayssubsequent to intravenous exposure to CdCl2 (Zalups,1997).
Based on the previous findings of Cikrt and Tichy(1974), intestinal secretion of Cd cannot be ruled out as apotential mechanism involved in the fecal excretion of Cd.However, most investigators believe that the major sourceof Cd that is excreted in the feces is derived from thehepatocellular secretion of Cd into the biliary system (Fig.1 and 3). This mechanism most likely involves the hepaticsecretion of some thiol conjugate of Cd.
Due to the presence of MRP2 and specific transporters ofGSH, there seems to be a strong likelihood that GSH S-conjugates of Cd are secreted into the biliary compartment(Fig. 3). Assuming that this is the case, these conjugateswould most likely undergo rapid enzymatic degradation tothe corresponding Cys S-conjugate via the actions of �-glu-tamyltransferase and dehydropeptidases, which are presentin great abundance on the canalicular plasma membrane ofhepatocytes. This predicted degradation is based on in vitrodata showing that methylmercuric S-conjugates of GSHwere degraded to the corresponding Cys conjugates, whilethe mercuric ion remained bonded to the sulfur atom on thecysteinyl residue, in the presence of purified enzymes (Na-ganuma et al., 1988). Once in the biliary ductal system,most of the GSH and/or Cys conjugates would be deliveredinto the lumen of the duodenum at the sphincter of Odi,where they may serve as absorptive substrates at the site ofone or more of the amino acid or peptide transporters
located on the luminal membrane of enterocytes. At present,there is a lack of experimental evidence supporting this typeof enteric recycling of Cd.
Urinary excretion of Cd
Excretion of Cd in the urine is greatly dependent on thetype of exposure. Following acute parenteral exposure toinorganic forms of Cd, the urinary elimination of Cd isnegligible. Zalups (1997) has demonstrated in normal, uni-nephrectomized, and 75% nephrectomized rats that lessthan 1% of an intravenous dose (8.9 �mol Cd/kg) wasexcreted by the end of the first week after exposure. Al-though the cumulative urinary excretion of Cd was ex-tremely low in all three groups of rats, the 75% nephrecto-mized rats excreted significantly more Cd than theuninephrectomized or normal control rats during each dayof the 7-day study.
When data on the renal accumulation and urinary excre-tion of Cd are assessed together following a low parenteraldose of Cd, they indicate the following: (1) The filtered loadof Cd, over the initial hours and days subsequent to expo-sure, is very low, accounting for no more than 1–2% of thedose, (2) very little of the filtered load escapes absorptionalong the nephron, and (3) there is very little to no netsecretion of Cd along the nephron.
Following exposure to Cd-MT (or chronic exposure toCd salts), Cd in the form of Cd-MT filters readily into theluminal compartment and the urinary excretion of Cd in-creases, relative to the level of Cd excreted in the urinefollowing an acute exposure to CdCl2. By having Cd in amore readily filterable form, more Cd gets delivered to theluminal surface of proximal tubular epithelial cells, whereCd-MT complexes are presumably absorbed by some endo-cytotic process. Interestingly, Zalups et al. (1995) havedemonstrated that the luminal uptake of Hg-MT was verylow to negligible when the complexes were perfusedthrough the lumen of S1, S2, or S3 segments of the rabbitproximal tubule. By contrast, Zalups et al. (1993) have alsoshown that both the renal accumulation of Hg and theurinary excretion of Hg increased greatly during the initial72 h in rats injected intravenously with a single nonneph-rotoxic dose of Hg-MT relative to that in rats that weretreated with the same dose of Hg in the form of HgCl2. Bythe end of the initial 3 days after treatment, the urinaryexcretion of Hg was almost ninefold greater in rats treatedwith Hg-MT than in the rats treated with HgCl2. In addition,as mentioned above, Felley-Bosco and Diezi (1987) havealso demonstrated that the fractional uptake of Cd was verylow when 109Cd-MT was microinjected into the lumen ofsuperficial proximal tubules. All of these findings tend toindicate that Cd-MT and other divalent cationic forms ofMT are not absorbed very efficiently along the proximaltubule in vivo. Thus, it appears that, when divalent metalcomplexes of MT are delivered to the kidneys, the com-plexes are filtered readily into the tubular lumen, and there
182 R.K. Zalups, S. Ahmad / Toxicology and Applied Pharmacology 186 (2003) 163–188

is a significant level of tubular absorption and accumulationof the metal, although the absorptive uptake is not highlyefficient, allowing a significant amount of the filtered load tobe excreted in the urine.
Ottenwalder and Simon (1987) have demonstrated afourfold increase in the urinary excretion of Cd in rats givenN-acetylcysteine (NAC; up to 100 mg/kg daily on 6 con-secutive days ip) after treatment with CdCl2. It is not clearfrom this study how treatment with NAC promoted theurinary excretion of Cd, but it is likely that the Cd thatentered into the luminal compartment was in the form of anonabsorbable, highly polar, conjugate of NAC, whichwould promote the urinary excretion of Cd. Evidence sup-porting this hypothesis comes from two recent studies inwhich the renal tubular absorption of Hg was very low whenthe Hg was in the form of a mercuric conjugate of NAC(Zalups, 1998b; Zalups and Barfuss, 1998a).
It has been also noted that repeated daily oral adminis-tration of certain chelating agents (such as diethylenetri-aminepentaacetic, EDTA, and 2,3-dimercaptosuccinic acid(DMPS)) after exposure to Cd caused the urinary excretionof Cd to increase (Klaassen et al., 1984). Despite thesefindings, there is considerable uncertainly about the putativepalliative effects of widely used dithiol chelating agents,such as DMPS or dimercaptosuccinic acid (DMSA), on therenal burden of Cd and urinary excretion of Cd. Preliminarydata from Zalups and colleagues indicate that treatment witheither DMPS or DMSA does not reduce significantly therenal burden of Cd in rats or mice treated with CdCl2. Onlyfurther research will be able to determine whether there areindeed beneficial effects of treating humans exposed to Cdwith these dithiol chelating agents.
Urinary excretion of Cd is also dependent on the structuraland physiological integrity of the proximal tubular epithelium.When tubular pathology is induced by Cd (or other nephro-toxicants), the urinary excretion of Cd increases, largely due todecreased absorption of filtered species of Cd plus the releaseof Cd from necrotic or apoptotic tubular epithelial cells. Thispoint is exemplified in a study by Zalups et al. (1992), in whichthe cumulative urinary excretion of Cd (factored by renalmass) was greater in uninephrectomized rats than correspond-ing control rats 24 h after the two groups received one of threedoses of Cd-MT (factored by kg body wt). At each of threedoses of Cd-MT studied, uninephrectomized rats developed amore severe form of the nephropathy induced by Cd-MT (asdetermined histopathologically and by the urinary excretion ofselective plasma solutes and cellular enzymes) than corre-sponding control rats with two kidneys. These findings clearlyshowed that, as renal injury is induced or made more severe,there is a corresponding increase in the urinary excretion of Cd.
Summary
Within this review, a concerted attempt has been made toconsolidate, as much as possible, the concepts, facts, and
speculations about how Cd is handled and transported byprimary target epithelial cells in the small intestine, liver,and kidneys. It is quite evident from the current body ofliterature that, notwithstanding all of the recent scientificdevelopments and the large number of studies carried out todate, speculation and indirect evidence serve as the primarysources of information on the mechanisms involved in theuptake and transport of Cd in the target epithelial cells.Consequently, a great deal of research needs to be con-ducted before a significant impact can be made on thepaucity of knowledge pertaining to the mechanisms in-volved in the handling and transport of Cd in target organs.
Acknowledgments
Support for this review comes mainly from grantsawarded by the National Institute of Environmental HealthSciences of the National Institutes of Health (ES05157,ES11288, and ES05980).
References
Abboud, S., Haile, D.J., 2000. A novel mammalian iron-regulated proteininvolved in intracellular iron metabolism. J. Biol. Chem. 275, 19906–19912.
Andersen, O., Nielsen, J.B., Sorensen, J.A., Scherrebeck, L., 1994. Exper-imental localization of intestinal uptake sites for metals (Cd, Hg, Zn,Se) in vivo in mice. Environ. Health Perspect 102 (Suppl. 3), 199–206.
Aschner, M., Clarckson, T.W., 1989. Methyl mercury uptake across bovinebrain capillary endothelial cells in vitro: the role of amino acids.Pharmacol. Toxicol. 64, 293–297.
Aslamkhan, A.G., Han, Y.H., Pritchard, J.B., Barfuss, D.W., Zalups, R.K.,2002. Renal organic anion transporter is a mechanism for the renaluptake of mercuric conjugates of cysteinyl homologs. Toxicol. Sci. 66,199.
Aslamkhan, A.G., Han, Y.-H., Yang, X.-P., Zalups, R.K., Pritchard, J.B.,2003. Human renal organic anion transporter 1-dependent uptake andtoxicity of mercuric thiol conjugates in Madin–Darby canine kidneycells . Mol. Pharmacol. 63, 1–7.
ATSDR, 1999. Toxicological profile for cadmium. Agency for ToxicSubstances and Disease Registry, Atlanta, GA.
Avissar, N.E., Ryan, C.K., Ganapathy, V., Sax, H.C., 2001. Na�-depen-dent neutral amino acid transporter ATB0 is a rabbit epithelial cellbrush-border protein. Am. J. Physiol. 281, C963–C971.
Ballatori, N., Dutczak, W.J., 1994. Identification and characterization ofhigh and low affinity transport systems for reduced glutathione in livercell canalicular membranes. J Biol. Chem. 269, 19731–19737.
Bhattacharyya, M.H., Wilson, A.K., Tajan, S.S., Jonah, M., 2000. Bio-chemical pathways in cadmium toxicity, in: Zalups, R.K., Koropatnick,J. (Eds.), Molecular Biology and Toxicology of Metals, Taylor andFrancis, London, pp. 34–74.
Berglund, M., Akesson, A., Nermell, B., Vahter, M., 1994. Intestinalabsorption of dietary cadmium in women depends on body iron storesand fiber intake. Environ. Health Perspect. 102, 1058–1066.
Blazka, M.E., Shaikh, Z.A., 1991. Differences in cadmium and mercuryuptakes by hepatocytes: role of calcium channels. Toxicol. Appl. Phar-macol. 110, 355–363.
Brown, J.R., Shockley, P., 1982. Serum albumin: structure and character-ization of its ligand binding sites, in: Jost, P.C., Griffith, O.H. (Eds.),Lipid–Protein Interactions, Vol 1, Wiley, New York, pp. 25–68.
183R.K. Zalups, S. Ahmad / Toxicology and Applied Pharmacology 186 (2003) 163–188

Bruggeman, I.M., Temmink, J.H.M., Van Bladeren, P.J., 1992. Effect ofglutathione on apical and basolateral uptake and toxicity of CdCl2 inkidney cells (LLC-PK1). Toxicol. in Vitro 6, 195–200.
Brzoska, M.M., Moniuszko-Jakoniuk, J., 2001. Interactions between cad-mium and zinc in the organism. Food Chem. Toxicol. 39, 967–980.
Cannon, V.T., Barfuss, D.W., Zalups, R.K., 2000. Molecular homology(�mimicry�) and the mechanisms involved in the luminal uptake ofinorganic mercury in the proximal tubule of the rabbit. J. Am. Soc.Nephrol. 11, 394–402.
Cannon, V.T., Zalups, R.K., Barfuss, D.W., 2001. Amino acid transportersinvolved in luminal transport of mercuric conjugates of cysteine inrabbit proximal tubule. J. Pharmacol. Exp. Ther. 298, 780–789.
Canonne-Hergaux, F., Gros, P., 2002. Expression of the iron transporterDMT1 in kidney from normal and anemic mice. Kidney Int. 62,147–156.
Cherian, M.G., 1977. Biliary excretion of cadmium in rat. II. The role ofmetallothionein in the hepatobiliary transport of cadmium. J. Toxicol.Environ. Health 2, 955–961.
Cherian, M.G., 1979. Metabolism of orally administered cadmium-metal-lothionein in mice. Environ. Health Perspect. 28, 127–130.
Cherian, M.G., 1980a. Biliary excretion of cadmium in rat. III. Effects ofchelating agents and change in intracellular thiol content on biliarytransport and tissue distribution of cadmium. J. Toxicol. Environ.Health 6, 379–391.
Cherian, M.G., 1980b. Biliary excretion of cadmium in rat. IV. Mobiliza-tion of cadmium from metallothionein by 2,3-dimercaptopropanol. J.Toxicol. Environ. Health 6, 393–401.
Cherian, M.G., Goyer, R.A., Delaquerriere-Richardson, L., 1976. Cad-mium metallothionein induced nephropathy. Toxicol. Appl. Pharmacol.38, 399–408.
Cherian, M.G., Goyer, R.A., Valberg, L.S., 1978. Gastrointestinal absorp-tion and organ distribution of oral cadmium chloride and cadmium-metallothionein in mice. J. Toxicol. Environ. Health 4, 861–868.
Cherian, M.G., Nordberg, M., 1983. Cellular adaptation in metal toxicol-ogy and metallothionein. Toxicology 82, 70–77.
Cherian, M.G., Shaikh, Z.A., 1975. Metabolism of intravenously injectedcadmium-binding protein. Arch. Biochem. Biophys. 82, 70–77.
Cherian, M.G., Vostal, J.J., 1977. Biliary excretion of cadmium in rat. I.Dose-dependent biliary excretion and the form of cadmium in the bile.J. Toxicol. Environ. Health 2, 945–954.
Choudhury, H., Harvery, T., Thayer, W.C., Lockwood, T.F., Stiteler,W.M., Goodrum, P.E., Hassett, J.M., Diamon, G.L., 2001. Urinarycadmium elimination as a biomarker of exposure for evaluating acadmium dietary exposure-biokinetics model. J. Toxicol. Environ.Health 63, 321–350.
Cikrt, M., Tichy, M., 1974. Excretion of cadmium through bile and intes-tinal wall in rats. Br. J. Ind. Med. 31, 134–139.
Cousins, R.J., Barber, A.K., Trout, J.R., 1973. Cadmium toxicity in grow-ing swine. J. Nutr. 103, 964–972.
Coyle, P., Niezing, G., Shelton, T.L., Philcox, J.C., Rofe, A.M., 2000.Tolerance to cadmium hepatotoxicity by metallothionein and zinc: invivo and in vitro studies with MT-null mice. Toxicology 150, 53–67.
Cragg, R.A., Christie, G.R., Phillips, S.R., Russi, R.M., Kury, S., Mathers,J.C., Taylor, P.M., Ford, D., 2001. A novel zinc-regulated human zinctransporter, hZTL1, is localized to the enterocytes apical membrane.J. Biol. Chem. 277, 22789–22797.
Dawson, D.C., Ballatori, N., 1995. Membrane transporters as sites ofaction and routes of entry for toxic metals, in: Goyer, R.A., Cherian,M.G. (Eds.), Toxicology of Metals, Springer-Verlag, Berlin, pp. 53–76.
Decker, L.E., Byerrum, R.U., Decker, C.F., et al., 1958. Chronic toxicitystudies. I. Cadmium administered in drinking water to rats. Am. Arch.Ind. Health 18, 228–231.
Diamond, G.L., Cohen, J.J., Weinstein, S.L., 1986. Renal handling ofcadmium in perfused rat kidney and effects on renal function and tissuecomposition. Am. J. Physiol. 251, F784–F794.
Diamond, G.L., Zalups, R.K., 1998. Understanding renal toxicity of heavymetals. Toxicol. Pathol. 26, 92–103.
Dijkstra, M., Havinga, R., Vonk, R.J., Kuipers, F., 1996. Bile secretion ofcadmium, silver, zinc and copper in the rat: involvement of varioustransport systems. Life Sci. 59, 1237–1246.
Dorian, C., Gattone II, V.H., Klaassen, C.D., 1992. Renal cadmium dep-osition and injury as a result of accumulation of cadmium-metallothio-nein (CdMT) by the proximal convoluted tubules: a light microscopyautoradiography study with 109CdMT. Toxicol. Appl. Pharmacol. 114,173–181.
Doyle, J.J., Pfander, W.H., Grebing, S.E., Pierce II, J.O., 1974. Effect ofdietary cadmium on growth, cadmium absorption and cadmium tissuelevels in growing lambs. J. Nutr. 104, 160–166.
Dudley, R.E., Gammal, L.M., Klaassen, C.D., 1985. Cadmium-inducedhepatic and renal injury in chronically exposed rats: likely role ofhepatic cadmium-metallothionein in nephrotoxicity. Toxicol. Appl.Pharmacol. 77, 414–426.
Dudley, R.E., Svoboda, D.J., Klaassen, C.D., 1982. Acute exposure tocadmium causes severe liver injury in rats. Toxicol. Appl. Pharmacol.65, 302–313.
Dudley, R.E., Svoboda, D.J., Klaassen, C.D., 1984. Time course of cad-mium-induced ultrastructural changes in rat liver. Toxicol. Appl. Phar-macol. 76, 150–160.
Elinder, C-G., Kjellstrom, T., Lind, B., Linnman, L., Friberg, L., 1976.Cadmium in kidney cortex, liver and pancreas from Swedish autopsies.Arch. Environ. Health 31, 292–302.
Elisma, F., Jumarie, C., 2001. Evidence for cadmium uptake throughNramp2: metal speciation studies with Caco-2 cells. Biochem Biophys.Res. Commun. 20, 662–668.
Elsenhans, B., Strugala, G.J., Schafer, S.G., 1997. Small-intestinal absorp-tion of cadmium and the significance of mucosal metallothionein. Hum.Exp. Toxicol. 16, 429–434.
Endo, T., 2002. Transport of cadmium across the apical membrane ofepithelial cells lines. Comp. Biochem. Physiol. C. Toxicol. Pharmacol.131, 223–239.
Endo, T., Kimura, O., Hatakeyama, M., Takada, M., Sakata, M., 1998d.Effects of zinc and copper on cadmium uptake by brush border mem-brane vesicles. Toxicol. Lett. 91, 111–120.
Endo, T., Kimura, O., Sakata, M., 1998a. Bidirectional transport of cad-mum across apical membrane of renal epithelial cell lines via H�-antiporter and inorganic anion exchanger. Toxicology 131, 183–192.
Endo, T., Kimura, O., Sakata, M., 1998b. pH-dependent transport ofcadmium in rat renal brush border membrane vesicles: cadmium effluxvia H�-antiport. Toxicol. Lett. 99, 99–107.
Endo, T., Kimura, O., Sakata, M., 1998c. Cadmium uptake from apicalmembrane of LLC-PK1 cells via inorganic anion exchanger. Pharma-col. Toxicol. 82, 230–235.
Endo, T., Kimura, O., Sakata, M., 1999. Further analysis of cadmiumuptake from apical membrane of LLC-PK1 cells via inorganic anionexchanger. Pharmacol. Toxicol. 84, 187–192.
Failla, M.L., Cousins, R.J., Mascenik, M.J., 1979. Cadmium accumulationand metabolism by rat liver parenchymal cells in primary monolayerculture. Biochim. Biophys. Acta 583, 63–72.
Fay, R.M., Mumtaz, M.M., 1996. Development of a priority list of chem-ical mixtures occurring at 1188 hazardous waste sites, using the HazDatdatabase. Food Chem. Toxicol. 34, 1163–1165.
Felley-Bosco, E., Diezi, J., 1987. Fate of cadmium in rat renal tubules: amicroinjection study. Toxicol. Appl. Pharmacol. 91, 204–211.
Felley-Bosco, E., Diezi, J., 1989. Fate of cadmium in rat renal tubules: amicropuncture study. Toxicol. Appl. Pharmacol. 98, 243–251.
Felley-Bosco, E., Diezi, J., 1991. Cadmium uptake and induction of me-tallothionein synthesis in a renal epithelial cell line (LLC-PK1). Arch.Toxicol. 65, 160–163.
Felley-Bosco, E., Diezi, J., 1992. Dietary calcium restriction enhancescadmium-induced metallothionein synthesis in rats. Toxicol. Lett. 60,139–144.
Ferguson, C.J., Wareing, M., Ward, D.T., Green, R., Smith, C.P., Riccardi,D., 2001. Cellular localization of divalent metal transporter DMT-1 inrat kidney. Am. J. Physiol. 280, F803–F814.
184 R.K. Zalups, S. Ahmad / Toxicology and Applied Pharmacology 186 (2003) 163–188

Fernandez-Checa, J.C., Ookhtens, M., Kaplowitz, N., 1993. Selective in-duction by phenobarbital of the electrogenic transport of glutathioneand organic anions in rat liver canalicular membrane vesicles. J Biol.Chem. 268, 10836–10841.
Fernandez-Checa, J.C., Takikawa, H., Horie, T., Ookhtens, M., Kaplowitz,N., 1992. Canalicular transport of reduced glutathione in normal andmutant Eisai hyperbilirubinemic rats. J Biol. Chem. 267, 1667–1673.
Flanagan, P.R., McLellan, J.S., Haist, J., Cherian, G., Chamberlain, M.J.,Valberg, L.S., 1978. Increased dietary cadmium absorption in mice andhuman subjects with iron deficiency. Gastroenterology 74, 841–846.
Foulkes, E.C., 1974. Excretion and retention of cadmium, zinc, and mer-cury by rabbit kidney. Am. J. Physiol 227, 1356–1360.
Foulkes, E.C., 1978. Renal tubular transport of cadmium-metallothionein.Toxicol. Appl. Pharmacol. 45, 505–512.
Foulkes, E.C., 1985. Interactions between metals in rat jejunum: implica-tions on the nature of cadmium uptake. Toxicology 37, 117–125.
Foulkes, E.C., 1988. On the mechanism of transfer of heavy metals acrosscell membranes. Toxicology 52, 263–272.
Foulkes, E.C., 2000. Transport of toxic heavy metals across cell mem-branes. Proc. Soc. Exp. Biol. Med. 223, 234–240.
Foulkes, E., Blanck, S., 1990. Acute cadmium uptake by rabbit kidneys:mechanism and effects. Toxicol. Appl. Pharmacol. 102, 464–473.
Foulkes, E.C., McMullen, D.M., 1986a. On the mechanism of nickelabsorption in the rat jejunum. Toxicology 38, 35–42.
Foulkes, E.C., McMullen, D.M., 1986b. Endogenous metallothionein asdeterminant of intestinal cadmium absorption: a reevaluation. Toxicol-ogy 38, 285–291.
Fox, M.R., Jacobs, R.M., Jones, A.O., Fry, B.E., Jr., Stone, C.L., 1980.Effects of vitamin C and iron and cadmium metabolism. Ann. NYAcad. Sci. 355, 249–261.
Friberg, L., 1984. Cadmium and the kidney. Environ. Health Perspect. 54,1–11.
Friberg, L., Elinder, C.G., Kjellstrom, T., et al., 1986. General summaryand conclusions and some aspects of diagnosis and treatment of chroniccadmium poisoning, in: Friberg, L., Elinder, C.G., Kjellstrom, T., et al.(Eds.), Cadmium and health: A toxicological and epidemiological ap-praisal, Vol. 2, CRC Press, Boca Raton, FL, pp. 247–255.
Friedman, P.A., Gesek, F.A., 1994. Cadmium uptake by kidney distalconvoluted tubule cells. Toxicol. Appl. Pharmacol. 128, 257–263.
Ganapathy, V., Ganapathy, M.E., Leibach, F.H., 2001. Intestinal transportof peptides and amino acids, in: Barrett, K.E., Donowitz, M. (Eds.),Current Topics in Membranes, Academic Press, New York, pp. 379–412.
Gill, K.D., Pal, R., Nath, R., 1989. Effect of chronic cadmium exposure onlipid peroxidation and peroxidation in liver and kidneys in rats. Med.Sci. Res. 17, 921–924.
Goering, P.L., Klaassen, C.D., 1983. Altered subcellular distribution ofcadmium following cadmium pretreatment: possible mechanism oftolerance to cadmium-induced lethality. Toxicol. Appl. Pharmacol. 70,195–203.
Goering, P.L., Klaassen, C.D., 1984. Resistance to cadmium-induced hep-atotoxicity in immature rats. Toxicol. Appl. Pharmacol. 74, 321–329.
Goering, P.L., Tandon, S.K., Klaassen, C.D., 1985. Induction of hepaticmetallothionein in mouse liver following administration of chelatingagents. Toxicol. Appl. Pharmacol. 80, 467–472.
Goering, P.L., Waalkes, M.P., Klaassen, C.D., 1995. Toxicology of cad-mium, in: Goyer, R.A., Cherian, M.G. (Eds.), Toxicology of Metals:Biochemical Aspects, Handbook of Experimental Pharmacology, Vol.115, Springer-Verlag, New York, pp. 189–213.
Gompertz, D., Chettle, D.R., Fletcher, J.G., Mason, H., Perkins, J., Scott,M.C., Smith, N.J., Topping, M.D., Blindt, M., 1983. Renal dysfunctionin cadmium smellers: relation to in-vivo liver and kidney cadmiumconcentrations. Lancet 1, 1185–1187.
Goyer, R.A., Cherian, M.G., Delaquerriere-Richardson, L., 1984. Correla-tion of parameters of cadmium exposure with onset of cadmium-induced nephropathy in rats. J. Environ. Pathol. Toxicol. Oncol. 5,89–100.
Graf, P., Sies, H., 1984. Hepatic uptake of cadmium and its biliary releaseas affected by dithioerythritol and glutathione. Biochem. Pharmacol.33, 639–643.
Gregus, Z, Klaassen, C.D., 1986. Disposition of metals in rats: a compar-ative study of fecal, urinary, and biliary excretion and tissue distribu-tion of eighteen metals. Toxicol. Appl. Pharmacol. 85, 24–38.
Gregus, Z., Varga, F., 1985. Role of glutathione and hepatic glutathioneS-transferase in the biliary excretion of methyl mercury, cadmium andzinc: a study with enzyme inducers and glutathione depletors. ActaPharmacol. Toxicol. 56, 398–403.
Groten, J.P., Luten, J.B., van Bladeren, P.J., 1992. Dietary iron lowers theintestinal uptake of cadmium-metallothionein in rats. Eur. J. Pharma-col. 228, 23–28.
Groten, J.P., Sinkeldam, E.J., Luten, J.B., van Bladeren, P.J., 1991a.Cadmium accumulation and metallothionein concentrations after4-week dietary exposure to cadmium chloride or cadmium-metallothio-nein in rats. Toxicol. Appl. Pharmacol. 111, 504–513.
Groten, J.P., Sinkeldam, E.J., Muys, T., Luten, J.B., van Bladeren, P.J.,1991b. Interaction of dietary Ca, P, Mg, Mn, Cu, Fe, Zn and Se with theaccumulation and oral toxicity of cadmium in rats. Food Chem. Toxi-col. 29, 249–258.
Grundemann, D., Gorboulev, V., Gambaryan, S., Veyhl, M., Koepsell, H.,1994. Drug excretion mediated by a new prototype of polyspecifictransporter. Nature 372, 549–552.
Gunshin, H., Mackenzie, B., Berger, U.V., Gunshin, Y., Romero, M.F.,Boron, W.F., Nussberger, S., Gollan, J.L., Hediger, M.A., 1997. Clon-ing and characterization of a mammalian proton-coupled metal-iontransporter. Nature 388, 482–488.
Habeebu, S.S., Liu, J., Klaassen, C.D., 1998. Cadmium-induced apoptosisin mouse liver. Toxicol. Appl. Pharmacol. 149, 203–209.
Habeebu, S.S., Liu, J., Liu, Y., Klaassen, C.D., 2000. Metallothionein-nullmice are more sensitive than wild-type mice to liver injury induced byrepeated exposure to cadmium. Toxicol. Sci. 55, 223–232.
Hinkle, P.M., Kinsella, P.A., Osterhoudt, K.C., 1987. Cadmium uptake andtoxicity via voltage-sensitive calcium channels. J. Biol. Chem. 262,16333–16337.
Hinkle, P.M., Shanshalam II, E.D., Nelson, E.J., 1992. Measurement ofintracellular cadmium with fluorescent dyes: further evidence for therole of calcium channels in cadmium uptake. J. Biol. Chem. 267,25553–25559.
Hussain, T., Shukla, G.S., Chandra, S.V., 1987. Effects of cadmium onsuperoxide dismutase and lipid peroxidation in liver and kidney ofgrowing rats: in vivo and in vitro studies. Pharmacol. Toxicol. 60,355–358.
IARC. 1993. Cadmium and certain cadmium compounds, in: IARC mono-graphs on the evaluation of the carcinogenic risk of chemicals tohumans: Beryllium, cadmium, mercury and exposures in the glassmanufacturing industry. IARC monographs, Vol. 58, World HealthOrganization. International Agency for Research on Cancer, Lyon,France, pp. 119–146, 210–236.
Ishido, M., Tohyama, C., Suzuki, T., 1999. Cadmium-bound metallothio-nein induces apoptosis in rat kidneys, but not in cultured kidneyLLC-PK1 cells. Life Sci. 64, 797–804.
Jaquemin, E., Hagenbuch, B., Steiger, B., Wolkoff, A.W., Meier, P.J.,1994. Expression cloning of a rat liver (Na�)-independent organicanion transporter. Proc. Natl. Acad. Sci. USA 91, 133–137.
Jarup, L., Berglund, M., Elinder, C.G., Nordberg, G., Vahter, M., 1998.Health effects of cadmium exposure: a review of the literature and arisk estimate. Scand J. Work Environ. Health 1, 1–51.
Jonah, M.M., Bhattacharyya, M.H., 1989. Early changes in the tissuedistribution of cadmium after oral but not intravenous cadmium expo-sure. Toxicology 58, 325–338.
Joseph, P., Lei, Y.X., Whong, W.Z., Ong, T.M., 2002. Oncogenic potentialof mouse translation elongation factor-1delta: a novel cadmium-re-sponsive proto-oncogene. J. Biol. Chem. 277, 6131–6136.
Joseph, P., Muchnok, T.K., Klishis, M.L., Roberts, J.R., Antonini, J.M.,Whong, W.Z., Ong, T., 2001. Cadmium-induced cell transformation
185R.K. Zalups, S. Ahmad / Toxicology and Applied Pharmacology 186 (2003) 163–188

and tumorigenesis are associated with transcriptional activation ofc-fos, c-jun, and c-myc proto-oncogenes: role of cellular calcium andreactive oxygen species. Toxicol. Sci. 61, 295–303.
Joshi, J.G., Sezekan, S.R., Fleming, J.T., 1989. Ferritin: a general metaldetoxicant. Biol. Trace Elem. Res. 21, 105–110.
Kamiyama, T., Miyakawa, H., Li, J.P., Akiba, T., Liu, J.H., Liu, J.,Marumo, F., Sato, C., 1995. Effects of one-year cadmium exposure onlivers and kidneys and their relation to glutathione levels. Res. Com-mun. Mol. Pathol. Pharmacol. 88, 177–186.
Kazantzis, G., 1978. Some long-term effect of cadmium on the humankidney, in: Cadmium77. Proceedings of the 1st International CadmiumConference in San Francisco, 1977, Metal Bulletin Ltd., London, pp.194–198.
Kerper, L.E., Ballatori, N., Clarkson, T.W., 1992. Methylmercury transportacross the blood brain barrier by an amino acid carrier. Am. J. Physiol.262, R761–R765.
Kimura, O., Endo, T., Sakata, M., 1996. Comparison of cadmium uptakesfrom apical and basolateral membranes of LLC-PK1 cells. Toxicol.Appl. Pharmacol. 137, 301–306.
Kimura, T., Itoh, N., Min, K.S., Fujita, I., Muto, N., Tanaka, K., 1998.Tissue accumulation of cadmium following oral administration to met-allothionein-null mice. Toxicol. Lett. 99, 85–90.
Kjellstrom, T., Nordberg, G.F., 1978. A kinetic model of cadmium me-tabolism in the human being. Environ. Res. 16, 248–269.
Kjellstrom, T., Nordberg, G.F. (1985). Kinetic model of cadmium metab-olism, in: Friberg, L., Elinder, C.G., Kjellstorm, T. (Eds.), Cadmiumand health: A toxicological and epidemiological appraisal. Vol. I.Exposure, dose and metabolism. CRC Press, Boca Raton, FL, pp.179–197.
Klaassen, C.D., Waalkes, M.P., Cantilena Jr., L.R., 1984. Alteration oftissue disposition of cadmium by chelating agents. Environ. HealthPerspect. 54, 233–242.
Kotsonis, F.N., Klaassen, C.D., 1977. Toxicity and distribution of cad-mium administered to rats at sublethal doses. Toxicol. Appl. Pharma-col. 41, 667–680.
Kotsonis, F.N., Klaassen, C.D., 1978. The relationship of metallothioneinto the toxicity of cadmium after prolonged oral administration to rats.Toxicol. Appl. Pharmacol. 46, 39–54.
Kullack-Ublick, G.A., Hagenbuch, B., Steiger, B., Wolkoff, A.W., Meier,P.J., 1994. Functional characterization of the basolateral rat liver or-ganic anion transporting polypeptide. Hepatology 20, 411–416.
Lansman, J.B., Hess, P., Tsien, R.W., 1986. Blockade of current throughsingle calcium channels by Cd2�, Mg2�, and Ca2�. J. Gen. Physiol. 88,321–347.
Lash, L.H., Jones, D.P., 1985. Distribution of oxidized and reduced formsof glutathione and cyst(e)ine in rat plasma. Arch. Biochem. Biophys.240, 583–592.
Lash, L.H., Xu, Y., Elfarra, A.A., Duescher, R.J., Parker, J.C., 1995.Glutathione-dependent metabolism of trichloroethylene in isolatedliver and kidney cells of rats and its role in mitochondrial and cellulartoxicity. Drug Metab. Dispos. 23, 846–853.
Lehman, L.D., Klaassen, C.D., 1986. Dosage-dependent disposition ofcadmium administered orally to rats. Toxicol. Appl. Pharmacol. 84,159–167.
Lewis, G.F., Coughlin, L., Jusko, W., Hartz, S., 1972. Contribution ofcigarette smoking of cadmium accumulation in man. Lancet 1, 291–292.
Liu, J., Liu, Y., Klaassen, C.D., 1994. Nephrotoxicity of CdCl2 and Cd-metallothionein in cultured rat kidney proximal tubules and LLC-PK1cells. Toxicol. Appl. Pharmacol. 128, 264–270.
Liu, Y., Liu, J., Iszard, M.B., Andrews, G.K., Palmiter, R.D., Klaassen,C.D., 1995. Transgenic mice that overexpress metallothionein-I areprotected from cadmium lethality and hepatotoxicity. Toxicol. Appl.Pharmacol. 135, 222–228.
Liu, J., Habeebu, S.S., Liu, Y., Klaassen, C.D., 1998a. Acute CdMTinjection is not a good model to study chronic Cd nephropathy: com-
parison of chronic CdCl2 and CdMT exposure with acute CdMTinjection in rats. Toxicol. Appl. Pharmacol. 153, 48–58.
Liu, J., Liu, Y., Habeebu, S.S., Klaassen, C.D., 1998b. Susceptibility ofMT-null mice to chronic CdCl2-induced nephrotoxicity indicates thatrenal injury is not mediated by the CdMT complex. Toxicol. Sci. 46,197–203.
Liu, J., Liu, Y., Habeebu, S.M., Waalkes, M.P., Klaassen, C.D., 2000.Chronic combined exposure to cadmium and arsenic exacerbates neph-rotoxicity, particularly in metallothionein-I/II null mice. Toxicology147, 157–166.
Liu, J., Liu, Y., Klaassen, C.D., 2001. Metallothionein-null and wild-typemice show similar cadmium absorption and tissue distribution follow-ing oral cadmium administration. Toxicol. Appl. Pharmacol. 175, 253–259.
Maack, T., 1992. Renal handling of proteins and polypeptides, in:Windhager, E.E. (Ed.), Handbook of Physiology, Section 8, RenalPhysiology, Vol. II, Oxford University Press, New York, pp. 2039–2082.
Maitani, T., Saito, Y., Suzuki, K.T., 1985. Cadmium found in non-solublefraction of kidney homogenates and its relation to renal dysfunctionafter cadmium-cysteine administration. Toxicology 37, 27–38.
Mack, U., Powell, L.W., Halliday, J.W., 1983. Detection and isolation ofa hepatic membrane receptor for ferritin. J. Biol. Chem. 258, 4672–2675.
Miller, W.J., Blackmon, D.M., Gentry, R.P., Pate, F.M., 1969. Effect ofdietary cadmium on tissue distribution of 109cadmium following asingle oral dose in young goats. J. Dairy Sci. 52, 2029–2035.
Min, K.S., Fujita, Y., Onosaka, S., Tanaka, K., 1991. Role of intestinalmetallothionein in absorption and distribution of orally administeredcadmium. Toxicol. Appl. Pharmacol. 109, 7–16.
Min, K.S., Kobayashi, K., Onosaka, S., Ohta, N., Okada, Y., Tanaka, K.,1986. Tissue distribution of cadmium and nephropathy after adminis-tration of cadmium in several chemical forms. Toxicol. Appl. Pharma-col. 86, 262–270.
Min, K.S., Onosaka, S., Tanaka, K., 1996. Renal accumulation of cadmiumand ephropathy following long-term administration of cadmium-met-allothionein. Toxicol. Appl. Pharmacol. 141, 102–109.
Mishima, A., Yamamoto, C., Fujiwara, Y., Kaji, T., 1997. Tolerance tocadmium cytotoxicity is induced by zinc through non-metallothioneinmechanisms as well as metallothionein induction in cultured cells.Toxicology 118, 85–92.
Moore Jr., W., Stara, J.F., Crocker, W.C., Malanchuk, M., Iltis, R., 1973.Comparison of 115m cadmium retention in rats following differentroutes of administration. Environ. Res. 6, 473–478.
Morgan, E.H., Baker, E., 1986. Iron uptake and metabolism by hepato-cytes. Fed. Proc. 45, 2810–2816.
Morselt, A.F., 1991. Environmental pollutants and diseases: a cell biolog-ical approach using chronic cadmium exposure in the animal model asa paradigm case. Toxicology 70, 1–132.
Murakami, M., Sano, K., Webb, M., 1987. The effect of L-cysteine on theportion-selective uptake of cadmium in the renal proximal tubule.Arch. Toxicol. 60, 365–369.
Naganuma, A., Oda-Urano, N., Tanaka, T., Imura, N., 1988. Possible roleof hepatic glutathione in transport of methylmercury into mouse kid-ney. Biochem. Pharmacol. 37, 291–296.
Nomiyama, K., Foulkes, E., 1977. Reabsorption of filtered cadmium me-tallothionein in the rabbit kidney. Proc. Soc. Exp. Biol. Med. 156,97–99.
Nomiyama, K., Nomiyama, H., Kameda, N., 1998. Plasma cadmium-metallothionein, a biological exposure index for cadmium-inducedrenal dysfunction, based on the mechanism of its action. Toxicology129, 157–168.
Nordberg, G.F., Goyer, R., Nordberg, M., 1975. Comparative toxicity ofcadmium-metallothionein and cadmium chloride on mouse kidney.Arch. Pathol. 99, 192–197.
Nordberg, G.F., Kjellstrom, T., Nordberg, M. 1985. Kinetics and metab-olism, in: Friberg, L., Elinder, C.G., Kjellstrom, T. (Eds.), Cadmium
186 R.K. Zalups, S. Ahmad / Toxicology and Applied Pharmacology 186 (2003) 163–188

and Health: A Toxicological and Epidemiological Appraisal. Vol. I.Exposure, Dose, and Metabolism. CRC Press, Boca Raton, FL, pp.103–178.
Nordberg, G.F., Nordberg, M. 1988. Biological monitoring of cadmium,in: Clarkson, T.W., Friberg, L., Nordberg, G.F., Sager, P.R. (Eds.),Biological Monitoring of Toxic Metals. Plenum Press, New York, pp.151–168.
Nordberg, M., Nordberg, G.F., 2000. Toxicological aspects of metallothio-nein. Cell Mol. Biol. 46, 451–463.
Nordberg, G.F., Piscator, M., Nordberg, M., 1971. On the distribution ofcadmium in blood. Acta Pharmacol. Toxicol. 30, 289–295.
Oberdorster, G., 1992. Pulmonary deposition, clearance and effects ofinhaled soluble and insoluble cadmium compounds. IARC Sci. Publ.118, 189–204.
Oka, J.A., Christensen, M.D., Weigel, P.H., 1989. Hyperosmolarity inhib-its galactosyl receptor-mediated but not fluid phase endocytosis inisolated rat hepatocytes. J. Biol. Chem. 264, 12016–12024.
Olivi, L., Sisk, J., Bressler, J., 2001. Involvement of DMT1 in uptake of Cdin MDCK cells: role of protein kinase C. Am. J. Physiol. 281, C793–C800.
Osterloh, K., Aisen, P., 1989. Pathways in the binding and uptake offerritin by hepatocystes. Biochim. Biophys. Acta 1011, 40–45.
Oteiza, P.I., Adonaylo, V.N., Keen, C.L., 1999. Cadmium-induced testesoxidative damage in rats can be influenced by dietary zinc intake.Toxicology 137, 13–22.
Ottenwalder, H., Simon, P., 1987. Differential effect of N-acetylcysteine onexcretion of the metals Hg, Cd, Pb and Au. Arch. Toxicol. 60, 401–402.
Ouellette, A.J., Aviles, L., Burnweit, C.A., Frederick, D., Malt, R.A., 1982.Metallothionein mRNA induction in mouse small bowel by oral cad-mium and zinc. Am. J. Physiol. 243, G396–G403.
Palmiter, R.D., Findley, S.D., 1995. Cloning and functional characteriza-tion of a mammalian zinc transporter that confers resistance to zinc.EMBO J. 14, 639–649.
Phillpotts, C.J., 1984. The autoradiographic localisation of retained orallyadministered cadmium tracer within Paneth cells of rat duodenum.Toxicology 33, 59–66.
Prozialeck, W.C., 2000. Evidence that E-cadherin may be a target forcadmium toxicity in epithelial cells. Toxicol. Appl. Pharmacol. 164,231–249.
Prozialeck, W.C., Lamar, P.C., 1993. Surface binding and uptake of cad-mium (Cd2�) by LLC-PK1 cells on permeable membrane supports.Arch. Toxicol. 67, 113–119.
Prozialeck, W.C., Wellington, D.R., Lamar, P.C., 1993. Comparison of thecytotoxic effects of cadmium chloride and cadmium-metallothionein inLLC-PK1 cells. Life Sci. 53, 337–342.
Rabenstein, D.L., 1989. Metal complexes of glutathione and their biolog-ical significance, in: Dolphin, D., Auramovibc, O., Poulson, R. (Eds.),Glutathione: Chemical, Biochemical and Medical Aspects Coenzymesand Cofactors, Vol. 3, Wiley, New York, pp. 147–186.
Rabenstein, D.L., Isab, A.A., Kadima, W., Mohanakrishnan, P., 1983. Aproton nuclear magnetic resonance study of the interaction of cadmiumwith human erythrocytes. Biochim. Biophys. Acta 762, 531–541.
Ragan, H.A., 1977. Effects of iron deficiency on the absorption anddistribution of lead and cadmium in rats. J. Lab. Clin. Med. 90,700–696.
Rao, M.S.N., Lal, H., 1958. Metal protein interactions in buffer solutions.III. Interaction of Cu2�, Zn2�, Cd2�, Co2�, and Ni2� with native andmodified serum albumins. J. Am. Chem. Soc. 80, 3226–3235.
Rikans, L.E., Yamano, T., 2000. Mechanism of cadmium-mediated acutehepatotoxicity. J. Biochem. Mol. Toxicol. 14, 110–117.
Robinson, M.K., Barfuss, D.W., Zalups, R.K., 1993. Cadmium transportand toxicity in isolated perfused segments of the renal proximal tubule.Toxicol. Appl. Pharmacol. 121, 103–111.
Sarkar, S., Yadav, P., Bhatnagar, D., 1998. Lipid peroxidative damage oncadmium exposure and alterations in antioxidant system in rat eryth-rocytes: a study with relation to time. Biometals 11, 153–157.
Sarkar, S., Yadav, P., Trivedi, R., Bansal, A.K., Bhatnagar, D., 1995.Cadmium-induced lipid peroxidation and the status of the antioxidantsystem in rat tissues. J. Trace Elem. Med. Biol. 9, 144–149.
Satarug, S., Haswell-Elkins, M.R., Moore, M.R., 2000. Safe levels ofcadmium intake to prevent renal toxicity in human subjects. Br. J. Nutr.84, 791–802.
Schaub, T.P., Kartenbeck, J., Konig, J., Spring, H., Dorsam, J., Staehler,G., Storkel, S., Thon, W.F., Keppler, D., 1999. Expression of the MRP2gene-encoded conjugate export pump in human kidney proximal tu-bules and in renal cell carcinoma. J. Am. Soc. Nephrol. 10, 1159–1169.
Schaub, T.P., Kartenbeck, J., Konig, J., Vogel, O., Witzgall, R., Kriz, W.,Keppler, D., 1997. Expression of the conjugate export pump encodedby the mrp2gene in the apical membrane of kidney proximal tubules.J. Am. Soc. Nephrol. 8, 1213–1221.
Schumann, K., Friebel, P., Schmolke, G., Elsenhans, B., Schumann, K.,Friebel, P., Schmolke, G., Elsenhans, B., 1996. State of iron repletionand cadmium tissue accumulation as a function of growth in young ratsafter oral cadmium exposure. Arch. Environ. Contam. Toxicol. 31,483–487.
Shaikh, Z.A., Smith, J.C., 1980. Metabolism of orally ingested cadmium inhumans, in: Mechanisms of Toxicity and Hazard Evaluation. Cadmiumand Health: A Toxicological and Epidemiological Appraisal. Elsevier/North-Holland Biomedical Press, New York, pp. 247–255.
Shaikh, Z.A., Vu, T.T., Zaman, K., 1999. Oxidative stress as a mechanismof chronic cadmium-induced hepatotoxicity and renal toxicity andprotection by antioxidants. Toxicol. Appl. Pharmacol. 154, 256–263.
Souza, V., Bucio, L., Gutierrez-Ruiz, M.C., 1997. Cadmium uptake by ahuman hepatic cell line (WRL-68 cells). Toxicology 120, 215–220.
Stockert, R.J., Haimes, H.B., Morell, A.G., Novikoff, P.M., Quintana, N.,Sternlieb, I., 1980. Endocytosis of asialoglycoprotein-enzyme conju-gates by hepatocytes. Lab. Invest. 43, 556–563.
Stohs, S.J., Bagchi, D., 1995. Oxidative mechanisms in the toxicity ofmetal ions. Free Radical Biol. Med. 18, 321–336.
Stohs, S.J., Bagchi, D., Hassoun, E., Bagchi, M., 2001. Oxidative mecha-nisms in the toxicity of chromium and cadmium ions. J. Environ.Pathol. Toxicol. Oncol. 20, 77–88.
Sugawara, N., Lai, Y.R., Arizono, K., Ariyoshi, T., 1996. Biliary excretionof exogenous cadmium, and endogenous copper and zinc in the Eisaihyperbilirubinuric (EHB) rat with a near absence of biliary glutathione.Toxicology 112, 87–94.
Sugawara, N., Sugawara, C., 1991. Gastrointestinal absorption of Cd-metallothionein and cadmium chloride in mice. Arch. Toxicol. 65,689–692.
Tacnet, F., Watkins, D.W., Ripoche, P., 1990. Studies of zinc transport intobrush-border membrane vesicles isolated from pig small intestine.Biochim. Biophys. Acta 1024, 323–330.
Tacnet, F., Watkins, D.W., Ripoche, P., 1991. Zinc binding in intestinalbrush-border membrane isolated from pig. Biochim. Biophys. Acta1063, 51–59.
Tallkvist, J., Bowlus, C.L., Lonnerdal, B., 2001. DMT1 gene expressionand cadmium absorption in human absorptive enterocytes. Toxicol.Lett. 122, 171–177.
Tanaka, K., Sueda, K., Onosaka, S., Okahara, K., 1975. Fate of 109Cd-labeled metallothionein in rats. Toxicol. Appl. Pharmacol. 33, 258–266.
Tandy, S., Williams, M., Leggett, A., Lopez-Jimenez, M., Dedes, M.,Ramesh, B., Srai, S.K., Sharp, P., 2000. Nramp2 expression is associ-ated with pH-dependent iron uptake across the apical membrane ofhuman intestinal Caco-2 cells. J. Biol. Chem. 1475, 1023–1029.
Taylor, W.R., 1988. Permeation of barium and cadmium through slowlyactivating calcium channels in cat sensory neurons. J. Physiol. (Lond.)407, 433–452.
Trinder, D., Oates, P.S., Thomas, C., Sadleir, J., Morgan, E.H., 2000.Localization of divalent metal transporter 1 (DMT1) to the microvillusmembrane of rat duodenal enterocytes in iron deficiency, but to hepa-tocytes in iron overload. Gut 46, 270–276.
187R.K. Zalups, S. Ahmad / Toxicology and Applied Pharmacology 186 (2003) 163–188

Verma, M.O., Sharma, R.P., Bourcier, D.R., 1982. Macromolecular inter-actions with cadmium and the effects of zinc, copper, lead and mercuryions. Biol. Trace Elem. Res. 3, 35–42.
Waalkes, M.P., Coogan, T.P., Barter, R.A., 1992. Toxicological principlesof metal carcinogenesis with special emphasis on cadmium. Crit. Rev.Toxicol. 22, 175–201.
Wareing, M., Ferguson, C.J., Green, R., Riccardi, D., Smith, C.P., 2000. Invivo characterization of renal iron transport in anesthetized rat.J. Physiol. (Lond.) 524, 581–586.
Webb, M., 1986. Role of metallothionein in cadmium metabolism, in:Foulkes, E.C. (Ed.), Cadmium, Springer-Verlag, Berlin/New York,pp. 281–337.
Wester, R.C., Maibach, H.I., Sedik, L., Melendres, J., DiZio, S., Wade, M.,1992. In vitro percutaneous absorption of cadmium from water and soilinto human skin. Fundam. Appl. Toxicol. 19, 1–5.
Zalups, R.K., 1995. Organic anion transport and action of �-glutamyl-transpeptidase in the kidney linked mechanistically to renal tubularuptake of inorganic mercury. Toxicol. Appl. Pharmacol. 132, 289–298.
Zalups, R.K., 1997. Influence of different degrees of reduced renal mass onthe renal and hepatic disposition of administered cadmium. J. Toxicol.Environ. Health 51, 245–264.
Zalups, R.K., 1998a. Basolateral uptake of inorganic mercury in the kid-ney. Toxicol. Appl. Pharmacol. 151, 192–199.
Zalups, R.K., 1998b. Basolateral uptake of mercuric conjugates of N-acteylcysteine and cysteine in the kidney involves the organic aniontransport system. J. Toxicol. Environ. Health 55, 13–29.
Zalups, R.K., 2000a. Evidence for basolateral uptake of cadmium in thekidneys of rats. Toxicol. Appl. Pharmacol. 164, 15–23.
Zalups, R.K., 2000b. Molecular interactions with mercury in the kidney.Pharmacol. Rev. 52, 113–143.
Zalups, R.K., Barfuss, D.W., 1995a. Accumulation and handling of inor-ganic mercury in the kidney after coadministration with glutathione. J.Toxicol. Environ. Health 44, 385–399.
Zalups, R.K., Barfuss, D.W., 1995b. Renal disposition of mercury in ratsafter intravenous injection of inorganic mercury and cysteine. J. Toxi-col. Environ. Health 44, 401–413.
Zalups, R.K., Barfuss, D.W., 1995c. Pretreatment with p-aminohippurateinhibits renal uptake and accumulation of injected inorganic mercury.Toxicology 103, 23–35.
Zalups, R.K., Barfuss, D.W., 1998a. Participation of mercuric conjugatesof cysteine, homocysteine and N-acetylcysteine in mechanisms in-volved in the renal tubular uptake of inorganic mercury. J. Am. Soc.Nephrol. 9, 551–561.
Zalups, R.K., Barfuss, D.W., 1998b. Small aliphatic dicarboxylic acidsinhibit renal uptake of administered mercury. Toxicol. Appl. Pharma-col. 148, 183–193.
Zalups, R.K., Barfuss, D.W., 2002a. Simultaneous coexposure to inorganicmercury and cadmium: a study of the renal and hepatic disposition ofmercury and cadmium. J. Toxicol. Environ. Health, Part A 65, 101–120.
Zalups, R.K., Barfuss, D.W., 2002b. Renal organic anion transport system:A mechanism for the basolateral uptake of mercury-thiol conjugatesalong the pars recta of the proximal tubule. Toxicol. Appl. Pharmacol.(in press).
Zalups, R.K., Minor, K.H., 1995. Luminal and basolateral mechanismsinvolved in the renal tubular uptake of inorganic mercury. J. Toxicol.Environ. Health 46, 73–100.
Zalups, R.K., Barfuss, D.W., Lash, L.H., 1999a. Disposition of inorganicmercury following biliary obstruction and chemically-induced gluta-thione depletion: dispositional changes one hour after the intravenousadministration of mercuric chloride. Toxicol. Appl. Pharmacol. 154,135–144.
Zalups, R.K., Barfuss, D.W., Lash, L.H., 1999b. Effects of biliary ligationand modulation of GSH-status on the renal and hepatic disposition ofinorganic mercury in rats. Toxicologist 48, 330–331.
Zalups, R.K., Barfuss, D.W., Lash, L.H., 1999c. Relationships betweenalterations in glutathione metabolism and the disposition of inorganicmercury in rats: effects of biliary ligation and chemically inducedmodulation of glutathione status. Chem.–Biol. Interact. 123, 171–195.
Zalups, R.K., Cherian, M.G., Barfuss, D.W., 1993. Mercury-metallothio-nein and the renal accumulation and handling of mercury. Toxicology83, 61–78.
Zalups, R.K., Cherian, M.G., Barfuss, D.W., 1995. Lack of luminal orbasolateral uptake and transepithelial transport of mercury in isolatedperfused proximal tubules exposed to mercury-metallothionein. J.Toxicol. Environ. Health 44, 101–113.
Zalups, R.K., Gelein, R.M., Cherian, M.G., 1992. Shifts in the dose–effectrelationship for the nephropathy induced by cadmium-metallothioneinin rats after a reduction of renal mass. J. Pharmacol Exp. Ther. 262,1256–1266.
Zalups, R.K., Lash, L.H., 1990. Effects of uninephrectomy and mercuricchloride on renal glutathione homeostasis. J. Pharmacol. Exp. Ther.254, 962–970.
Zalups, R.K., Lash, L.H., 1994. Recent advances in the study of transportand toxicity of mercury in the kidney. J. Toxicol. Environ. Health 42,1–44.
Zoller, H., Pietrangelo, A., Vogel, W., Weiss, G., 1999. Duodenal metal-transporter (DMT-1, NRAMP-2) expression in patients with hereditaryhaemochromatosis. Lancet 353, 2120–2123.
188 R.K. Zalups, S. Ahmad / Toxicology and Applied Pharmacology 186 (2003) 163–188
Copyright © 2022 FDOKUMEN




















