BRAIN CHOLINESTERASE ACTIVITY AND BRAIN AND LIVER RESIDUES IN WILD BIRDS OF A FOREST SPRAYED WITH...
10
Environmenral ToxicologyandChemistry, Vol. 3, pp. 79-88, 1984 Printed in the USA. Pergamon Press Ltd. 0730.7268/84 $3.00+ .OO Copyright 0 1984 SETAC BRAIN CHOLINESTERASE ACTIVITY AND BRAIN AND LIVER RESIDUES IN WILD BIRDS OF A FOREST SPRAYED WITH ACEPHATE JOSEPH G. ZINKL,' PAUL D. MACK and MICHAEL E. MOUNT Department of Clinical Pathology School of Veterinary Medicine University of California Davis. California 95616 PATRICK J. SHEA Pacific Southwest Forest and Range Experiment Station Forest Service US. Department of Agriculture Davis. California 956 16 (Received 24 February 1983; Accepted 31 August 1983) Abstract - In May I98 I, acephate (U,S-dimethyl-acetylphosphoramidothioate) was applied at 0.85 kg [a.i. (active ingredient)]/ha in a ponderosa pine (Pinusponderosa Douglas)forest of northern Arizona for suppression of a pandora moth (Coloradiu pandora Blake) infestation. Brain cholinesterase (ChE)activity was determined in six speciesof wild birds, and brain and liver residues were determined in five species. Brain ChE was not inhibited on the day of spraying, but ChE was significantly inhibited 2 d after acephate application. The speciesof birds which presumably had the smallest territories and therefore would be more likely to be confined to the relatively small spray plots (about 250 ha) had thegreatest ChE inhibition, both in magnitude and in numbers of birds with inhibition. Acephate and methamidophos (US-dimethyl-phosphoramidothioate) residues in brain and liver were highest on the day of spraying and decreased to undetectable concentrations 14 d after application. No correlation between brain ChE activities and brain or liver residue Goncentrations was found. The results of this study suggest that brain ChE inhibition found after acephate application is a better indicator of exposure than is tissue residue analysis. Brain ChE inhibition is probably due toa metabolite of acephate that is not detected by the methodsused. Birds exposed to acephate in the wild may suffer as yet undetermined sublethal effects, but it is unlikely that acephate applied at 0.85 kg/ha or less will kill many birds. Keywords - Cholinesterase Acephate Methamidophos Birds Pandora moth [3]. Brain ChE inhibition is not readily found on the day of spraying but begins to Application of the organophosphate occur on the day after application and be- comes most severe at two or three days after application [ 1-31. Brain ChE depression can be detected for three to four weeks, al- though its magnitude and the percentage of birds with depressed activities declines. When such findings are compared with lab- INTRODUCTION insecticide acephate (0,s-dimethyl-acetyl- phosphoramidothioate) in forests has been shown to cause brain cholinesterase (ChE) depression in wild birds [ 1-31 and mammals *To whom correspondence may be addressed. 79
Transcript of BRAIN CHOLINESTERASE ACTIVITY AND BRAIN AND LIVER RESIDUES IN WILD BIRDS OF A FOREST SPRAYED WITH...

Environmenral ToxicologyandChemistry, Vol. 3, pp. 79-88, 1984 Printed in the USA. Pergamon Press Ltd.
0730.7268/84 $3.00+ .OO Copyright 0 1984 SETAC
BRAIN CHOLINESTERASE ACTIVITY AND BRAIN AND LIVER RESIDUES IN WILD BIRDS OF A FOREST SPRAYED WITH
ACEPHATE
JOSEPH G. ZINKL,' PAUL D. MACK and MICHAEL E. MOUNT
Department of Clinical Pathology School of Veterinary Medicine
University of California Davis. California 95616
PATRICK J. SHEA
Pacific Southwest Forest and Range Experiment Station Forest Service
US. Department of Agriculture Davis. California 956 16
(Received 24 February 1983; Accepted 31 August 1983)
Abstract - In May I98 I , acephate (U,S-dimethyl-acetylphosphoramidothioate) was applied at 0.85 kg [a.i. (active ingredient)]/ha in a ponderosa pine (Pinusponderosa Douglas) forest of northern Arizona for suppression of a pandora moth (Coloradiu pandora Blake) infestation. Brain cholinesterase (ChE) activity was determined in six speciesof wild birds, and brain and liver residues were determined in five species. Brain ChE was not inhibited on the day of spraying, but ChE was significantly inhibited 2 d after acephate application. The speciesof birds which presumably had the smallest territories and therefore would be more likely to be confined to the relatively small spray plots (about 250 ha) had thegreatest ChE inhibition, both in magnitude and in numbers of birds with inhibition. Acephate and methamidophos (US-dimethyl-phosphoramidothioate) residues in brain and liver were highest on the day of spraying and decreased to undetectable concentrations 14 d after application. No correlation between brain ChE activities and brain or liver residue Goncentrations was found. The results of this study suggest that brain ChE inhibition found after acephate application is a better indicator of exposure than is tissue residue analysis. Brain ChE inhibition is probably due toa metabolite of acephate that is not detected by the methodsused. Birds exposed to acephate in the wild may suffer as yet undetermined sublethal effects, but it is unlikely that acephate applied at 0.85 kg/ha or less will kill many birds.
Keywords - Cholinesterase Acephate Methamidophos Birds Pandora moth
[3]. Brain ChE inhibition is not readily found on the day of spraying but begins to
Application of the organophosphate occur on the day after application and be- comes most severe at two or three days after application [ 1-31. Brain ChE depression can be detected for three to four weeks, al- though its magnitude and the percentage of birds with depressed activities declines. When such findings are compared with lab-
INTRODUCTION
insecticide acephate (0,s-dimethyl-acetyl- phosphoramidothioate) in forests has been shown to cause brain cholinesterase (ChE) depression in wild birds [ 1-31 and mammals
*To whom correspondence may be addressed. 79

80 J. G. ZINKL ET AL.
oratory studies of the toxicity of acephate, discrepancies are found. The amount of acephate necessary to cause brain ChE inhibition in the caged birds equal to the inhibition found in the birds of the forest is many times the amount that the forest birds probably could ingest during feeding [4]. Metharnidophos (0,s-dimethyl-phosphora- midothioate), the deacylated metabolite of acephate, is more toxic to birds than is ace- phate 151, and it has been found in a few birds after an acephate application [3]. However, even if most of the acephate in an opera- tional application were converted to metharnidophos, it is unlikely that birds could ingest a sufficient quantity of metha- midophos to cause the magnitude of brain ChE inhibition found after acephate spray- ing [4]. Furthermore, limited data from field applications of acephate indicate that there is no correlation between brain ChE reduc- tion and residues of acephate or methamidophos in the brain [3].
This study was performed to determine whether brain ChE actjvity was related to brain or liver concentrations of acephate or metamidophos. An additional objective was to determine how extensive ChE depression is within a population of birds exposed to acephate. To do this, the study concentrated on the more abundant species.
MATERIALS AND METHODS
Insecticide application
In May 1981, the US. Forest Service sprayed a ponderosa pine (Pinus ponderosa Douglas) forest in the Kiabab National For- est of northern Arizona with acephate to suppress a pandora moth (Coloradiapando- ra Blake) infestation. Acephate was applied by fixed-wing aircraft at 0.85 kg (a.i.)/ha (0.75 lblacre) to five of eight plots of approxi- mately 250 ha (600 acres) each. Acephate was formulated in water and applied at 7.85 Llha (1 gallacre). All plots were sprayed be- tween 6:OO a.m. and 1O:OO a.m.; two plots were treated on May 13 and one each on
May 14, 15 and 19. The three unsprayed plots served as controls, were at least 1 km from the sprayed plots and had habitats similar to those of the sprayed plots.
Bird collection
The major species of birds collected for determining ChE activities were grey- headed juncos (Junco caniceps), western bluebirds (Sialia mexicana), Grace’s war- blers (Dendroica graciae) and yellow- rumped warblers (Dendroica coronata). In addition, pygmy nuthatches (Sittapygmaea) and American robins (Turdus migratorius) were collected in limited numbers. The birds were collected in the morning by shooting them with shotguns. Birds were collected from both the sprayed plots and control areas before the spray dates, on the day of spraying (about 2 h after the application was completed) and at intervals thereafter up to 21 dafter the spray. We attempted to collect four birds of each species at each time.
After collecting the birds, we placed them in ice chests containing crushed ice. After one day’s collection was complete, the birds were transported to Jacob Lake, Arizona, where, among other procedures, their heads and livers were frozen in dry ice. Periodi- cally, the frozen heads and livers were transported in dry ice to Davis, California, by either automobile or commercial airline, In all cases, the parts remained frozen throughout storage and transportation. At Davis they were stored in an ultra-cold freezer ( - 7OOC) until analysis.
Cholinesterase analysis
After thawing the heads, the brains were dissected from the skulls and cut in half along the midline. One-half of each brain was refrozen for later use in residue analysis; the other half was weighed and homogen- ized with 9 volumes of 0.05 M Tris buffer (pH 8.0). Cholinesterase activity was de- termined by the Ellman [6] technique as modified for brain tissue by Dieter and Ludke [7].

Brain cholinesterase in birds exposed to acephate 81
Residue analysis
The analytical procedure for acephate and methamidophos was an organic extrac- tion, cleanup on silica gel, followed by gas-liquid chromatography (Varian 3700). Methylene chloride was used to extract the residues from liver and brain tissues and cleanup was performed by previously described methods 181. Acephate and methamidophos were separated using a 46-cm (1 8-in) glass column (2 mm id.) with 0.7 % Reoplex on 100- 120 mesh Gas Chrom Q packing material. A thermionic specific detector was used for residue detection. Limits of detection were 0.05 ppm for acephate and 0.005 ppm for metha- midophos. Recovery of the insecticides in spiked samples was between 90 and 95%.
Statistical analysis Samples generally consisted of four birds.
Sampling times varied among species, but in general consisted of groups of birds taken from treatment and control plots before spraying, on the day of spraying (day 0) and at various times after the spraying. The birds used for controls were collected from the control plots throughout the study period and from the spray plots before the insecti- cide was applied. In a few cases, the birds from two consecutive days were combined to produce larger groups. Details on the group sizes and collection days can be found in Tables 1 through 1 1.
For grey-headed juncos, western blue- birds, Grace’s warblers and yellow-rumped warblers, one-way analysis of variance was used to determine if brain ChE activities differed between groups of birds. When the F test indicated significant brain ChE activi- ty differences existed, Scheffe’s [9] method for pairwise comparison of means was used to determine if brain ChE activities of ace- phate-exposed birds and control birds accounted for the differences. For robins and pygmy nuthatches, Student’s t test was used, since there were only one control group and one exposed group of birds for each species.
In addition to the statistical method used to determine if groups of birds had different brain ChE activities, a clinical technique was used to determine if individual birds had decreased ChE activities. Using this method a bird’s ChE activity was considered de- creased if it was depressed more than two standard deviations below the mean of unexposed birds or 20% below the mean, whichever was greater [lo].
Linear regression analysis was used to determine if there was a correlation between brain ChE activities and brain or liver residue concentrations.
RESULTS
Brain cholinesterase activity
Within each species there were groups that showed significant brain ChE depres- sion (Tables 1-6). Depressed ChE activity was not found on the day of spraying. Birds collected two days after the spray had the greatest depression. The greatest magnitude of brain ChE depression occurred two days after spraying and was in grey-headed juncos, Grace’s warblers, yellow-rumped warblers, and pygmy nuthatches (Tables 1 and 3-5). hterestingly, birds collected on day 3 generally did not have depressed activ- ity. Most of these birds were collected on May 16, immediately after a snow storm. A few single birds with depressed brain ChE activities were found throughout the rest of the study (Tables 1-4). However, only in yellow-rumped warblers on post- spraying days 7 and 8 was the group brain ChE activity depressed (Table 4).
Of the 124 birds collected from the sprayed plots, five had brain ChE activities depressed more than 50%. These birds were two grey-headed juncos collected on day 2, two yellow-rumped warblers collected on days 2 and 5 and a pygmy nuthatch col- lected on day 2. The birds with the greatest ChE depression, both in magnitude and in numbers of birds with depression, were the grey-headed juncos, Grace’s warblers,
82 J. G. Z I N K L ET AL.
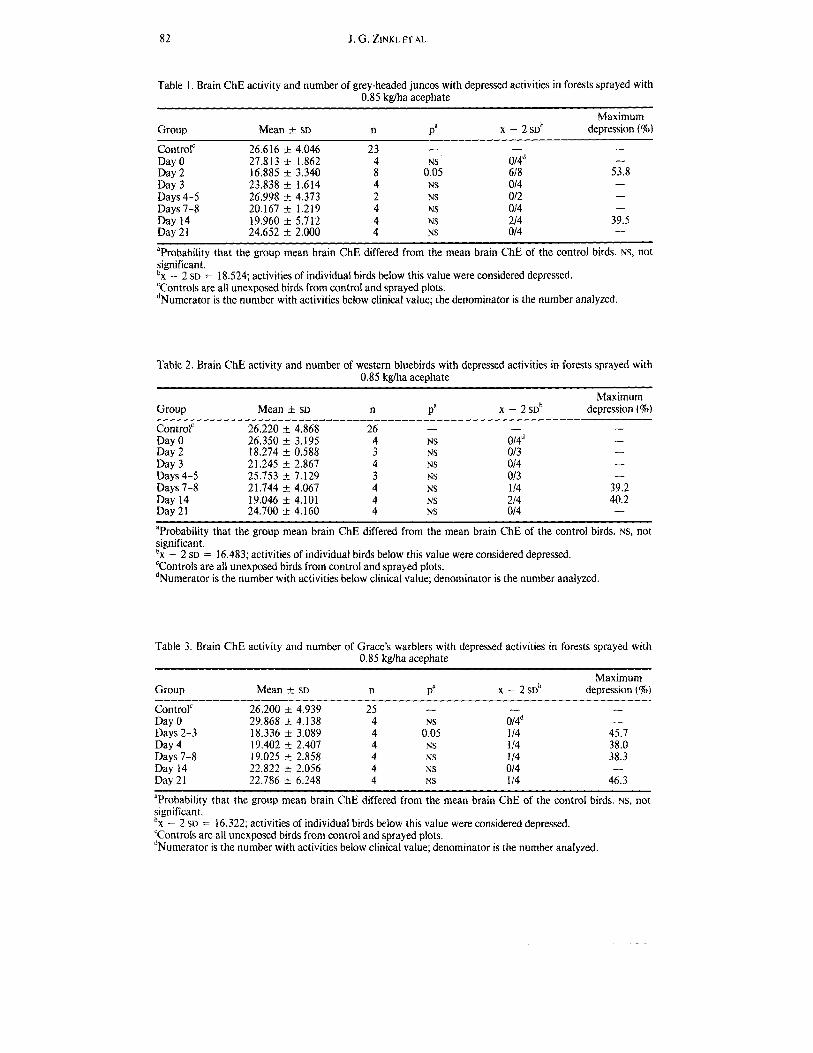
Table 1. Brain ChE activity and number of grey-headed juncos with depressed activities in forests sprayed with 0.85 kdha aceuhate
Group Mean -t SD n Pa X - 2SDb - - Control' 26.616 t 4.046 23
Day 0 27.813 t 1.862 4 NS 0/4d Day 2 16.885 t 3.340 8 0.05 618 Day 3 23.838 t 1.614 4 NS 014
Days 7-8 20.167 k 1.219 4 NS 014 Day 14 19.960 zk 5.712 4 NS 214 Day 21 24.652 k 2.000 4 NS 014
Days 4-5 26.998 k 4.373 2 NS 012
Maximum depression (%)
- -
53.8
- 39.5 -
aProbability that the group mean brain ChE differed from the mean brain ChE of the control birds. NS, not significant. b~ - 2 SD = 18.524; activities of individual birds below this value were considered depressed. 'Controls are all unexposed birds from control and sprayed plots. dNumerator is the number with activities below clinical value; the denominator is the number analyzed.
Table 2. Brain ChE activity and number of western bluebirds with depressed activities in forests sprayed with 0.85 kglha acephate
Maximum Group Mean -t SD n Pa X - 2 SDb depression (%)
- - Control' 26.220 t 4.868 26 Day 0 26.350 t 3.195 4 NS 014d Day 2 18.274 t 0.588 3 NS 013 Day 3 21.245 _+ 2.867 4 NS 014 Days 4-5 25.753 k 7.129 3 NS 013 Days 7-8 21.744 k 4.067 4 NS 1 14 Day 14 19.046 _t 4.101 4 NS 214 Day 21 24.700 k 4.160 4 NS 014
- -
39.2 40.2 -
aProbability that the group mean brain ChE differed from the mean brain ChE of the control birds. NS, not significant . b~ - 2 SD = 16.483; activities of individual birds below this value were considered depressed. 'Controls are all unexposed birds from control and sprayed plots. dNumerator is the number with activities below clinical value; denominator is the number analyzed.
Table 3. Brain ChE activity and number of Grace's warblers with depressed activities in forests sprayed with 0.85 kg/ha acephate
Group Mean t SD n P' X - 2SDb depression (YO) Maximum
- - - Control' 26.200 t 4.939 25 Day 0 29.868 _t 4.138 4 NS 014d - Days 2-3 18.336 _t 3.089 4 0.05 114 45.7 Day 4 19.402 +. 2.407 4 NS 1 I4 38.0 Days 7-8 19.025 k 2.858 4 NS I 14 38.3 Day 14 22.822 -t 2.056 4 NS 014 - Day 21 22.786 -t 6.248 4 NS 114 46.3
'Probability that the group mean brain ChE differed from the mean brain ChE of the control birds. NS, not significant. b~ - 2 SD = 16.322; activities of individual birds below this value were considered depressed. 'Controls are all unexposed birds from control and sprayed plots. dNumerator is the number with activities below clinical value; denominator is the number analyzed.
Brain cholinesterase in birds exposed to acephate 83
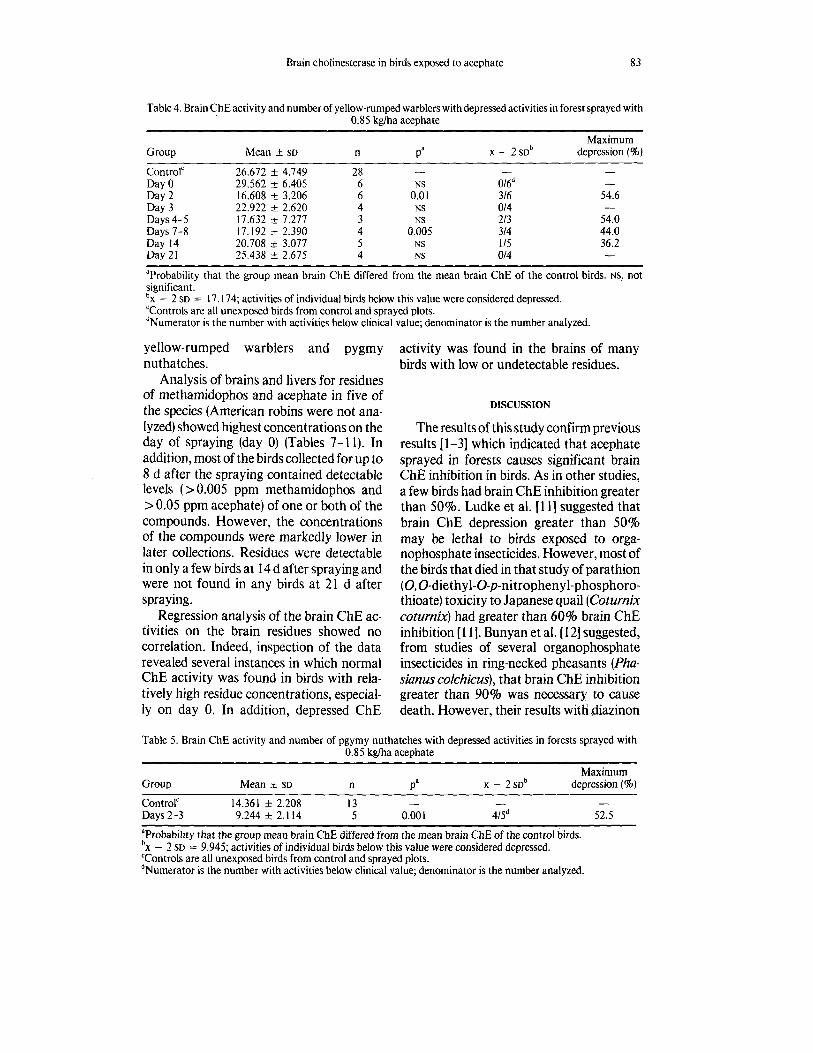
Table 4. Brain ChE activity and number of yellow-rumped warblers with depressed activities in forest sprayed with 0.85 kglha acephate
Maximum Group Mean t SD n Pa X - 2SDb depression (%)
- - - Control' 26.672 f 4.749 28
Day 2 16.608 5 3.206 6 0.0 1 316 54.6 Day 3 22.922 * 2.620 4 NS 014 Days 4-5 17.632 k 7.277 3 NS 213 54.0 Days 7-8 17.192 t 2.390 4 0.005 314 44.0 Day 14 20.708 k 3.077 5 NS 115 36.2 Day 21 25.438 t 2.675 4 NS 014 - "Probability that the group mean brain ChE differed from the mean brain ChE of the control birds. NS, not significant . b~ - 2 SD = 17.174; activities of individual birds below this value were considered depressed. 'Controls are all unexposed birds from control and sprayed plots. dNumerator is the number with activities below clinical value; denominator is the number analyzed.
Day 0 29.562 f 6.405 6 NS 0/6d - -
yellow-rumped warblers and pygmy nuthatches.
Analysis of brains and livers for residues of methamidophos and acephate in five of the species (American robins were not ana- lyzed) showed highest concentrations on the day of spraying (day 0) (Tables 7-1 1). In addition, most of the birds collected for up to 8 d after the spraying contained detectable levels (> 0.005 ppm methamidophos and > 0.05 ppm acephate) of one or both of the compounds. However, the concentrations of the compounds were markedly lower in later collections. Residues were detectable in only a few birds at 14 d after spraying and were not found in any birds at 21 d after spraying.
Regression analysis of the brain ChE ac- tivities on the brain residues showed no correlation. Indeed, inspection of the data revealed several instances in which normal ChE activity was found in birds with rela- tively high residue concentrations, especial- ly on day 0. In addition, depressed ChE
activity was found in the brains of many birds with low or undetectable residues.
DISCUSSION
The results of this study confirm previous results [ 1-31 which indicated that acephate sprayed in forests causes significant brain ChE inhibition in birds. As in other studies, a few birds had brain ChE inhibition greater than 50%. Ludke et al. [l 11 suggested that brain ChE depression greater than 50% may be lethal to birds exposed to orga- nophosphate insecticides. However, most of the birds that died in that study of parathion (0, O-diethyl-O-p-nitrophenyl-phosphoro- thioate) toxicity to Japanese quail (Coturnix coturnix) had greater than 60% brain ChE inhibition [ 1 I]. Bunyan et al. [ 121 suggested, from studies of several organophosphate insecticides in ring-necked pheasants (Phu- siunus colchicus), that brain ChE inhibition greater than 90% was necessary to cause death. However, their results with diazinon
Table 5. Brain ChE activity and number of pgymy nuthatches with depressed activities in forests sprayed with 0.85 kglha acephate
Maximum Group Mean k SD n Pa X - 2SDb depression (8) Control' 14.361 & 2.208 13 Days 2-3 9.244 1+_ 2.114 5 0.001 41Sd 52.5
aProbability that the group mean brain ChE differed from the mean brain ChE of the control birds. b~ - 2 SD = 9.945; activities of individual birds below this value were considered depressed. 'Controls are all unexposed birds from control and sprayed plots. dNumerator is the number with activities below clinical value; denominator is the number analyzed.
- - -
84 J . G. ZINKL ETAL
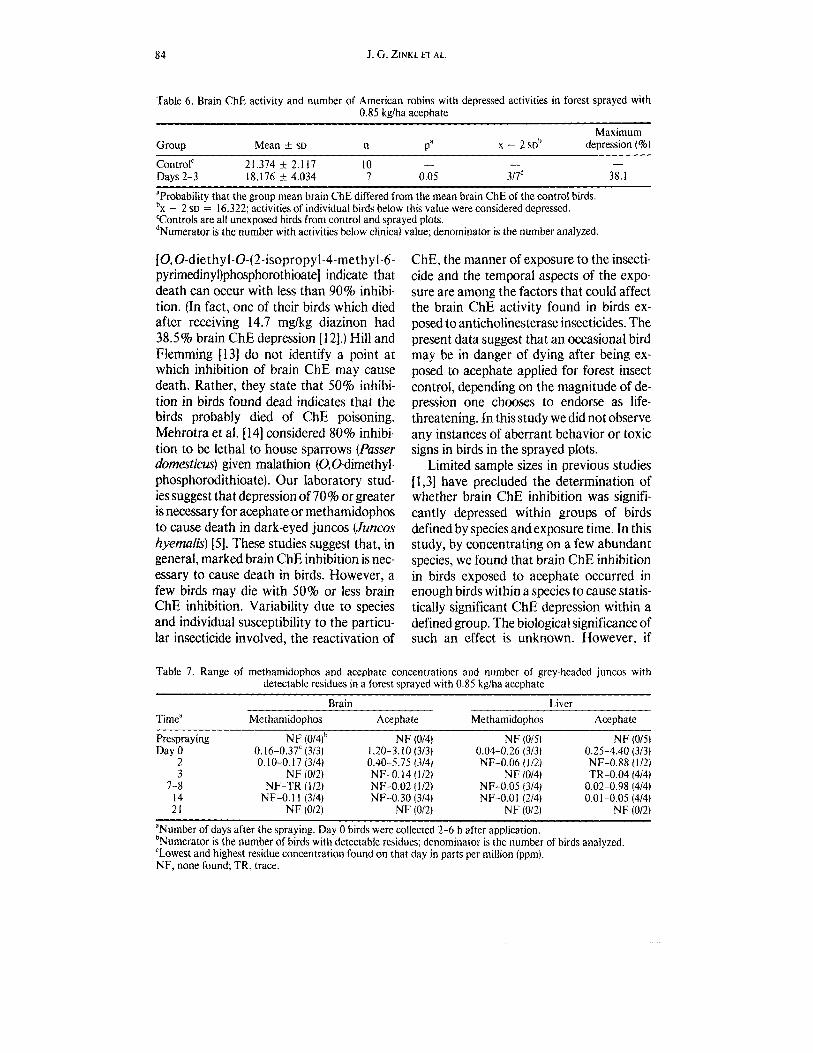
Table 6. Brain ChE activity and number of American robins with depressed activities in forest sprayed with 0.85 kglha acephate
Maximum Group Mean k SD n Pa X - 2SDb depression (%)
ControlC 21.374 t 2.1 17 10 Days 2-3 18.176 t 4.034 7 0.05 3/Id 38.1
'Probability that the group mean brain ChE differed from the mean brain ChE of the control birds. b~ - 2 SD = 16.322; activities of individual birds below this value were considered depressed. 'Controls are all unexposed birds from control and sprayed plots. dNumerator is the number with activities below clinical value; denominator is the number analyzed.
- - -
[O, O-diethyl-O-(2-isopropyl-4-methyl-6- pyrimedinyl)phosphorothioate] indicate that death can occur with less than 90% inhibi- tion. (In fact, one of their birds which died after receiving 14.7 mglkg diazinon had 38.5% brain ChE depression [ 121.) Hill and Flemming [13] do not identify a point at which inhibition of brain ChE may cause death. Rather, they state that 50% inhibi- tion in birds found dead indicates that the birds probably died of ChE poisoning. Mehrotra et al. [ 141 considered 80% inhibi- tion to be lethal to house sparrows (Passer domesticus) given malathion (0,Odimethyl- phosphorodithioate). Our laboratory stud- ies suggest that depression of 70% or greater is necessary for acephate or methamidophos to cause death in dark-eyed juncos (Juncos hyemalis) IS]. These studies suggest that, in general, marked brain ChE inhibition is nec- essary to cause death in birds. However, a few birds may die with 50% or less brain ChE inhibition. Variability due to species and individual susceptibility to the particu- lar insecticide involved, the reactivation of
ChE, the manner of exposure to the insecti- cide and the temporal aspects of the expo- sure are among the factors that could affect the brain ChE activity found in birds ex- posed to anticholinesterase insecticides. The present data suggest that an occasional bird may be in danger of dying after being ex- posed to acephate applied for forest insect control, depending on the magnitude of de- pression one chooses to endorse as life- threatening. In this study we did not observe any instances of aberrant behavior or toxic signs in birds in the sprayed plots.
Limited sample sizes in previous studies [ 1,3] have precluded the determination of whether brain ChE inhibition was signifi- cantly depressed within groups of birds defined by species and exposure time. In this study, by concentrating on a few abundant species, we found that brain ChE inhibition in birds exposed to acephate occurred in enough birds within a species to cause statis- tically significant ChE depression within a defined group. The biological significance of such an effect is unknown. However, if
Table 7. Range of methamidophos and acephate concentrations and number of grey-headed juncos with detectable residues in a forest sprayed with 0.85 kglha acephate
Brain Liver Timea Metharnidophos Acep hate
Prespraying NF (014)b NF (014) Day 0 0.16-0.37' (313) 1.20-3.10 (3/3)
2 0.10-0.17 (314) 0.40-5.75 (314) 3 NF (012) NF-0.14 (112)
7-8 NF-TR (112) NF-0.02 (112) 14 NF-0.11 (3/4) NF-0.30 (314) 21 NF (012) NF (012)
Methamidophos Acephate
NF (015) NF (015) 0.04-0.26 (313) 0.25-4.40 ( 3 / 3 ) NF-0.06 (112) NF-0.88 (112)
NF (014) TR-0.04 (4/4) NF-0.05 (314) 0.02-0.98 (4/4) NF-0.01 (214) 0.01-0.05 (414)
NF (012) NF (012)
'"umber of days after the spraying. Day 0 birds were collected 2-6 h after application. bNumerator is the number of birds with detectable residues; denominator is the number of birds analyzed. 'Lowest and highest residue concentration found on that day in parts per million (ppm). NF, none found; TR, trace.
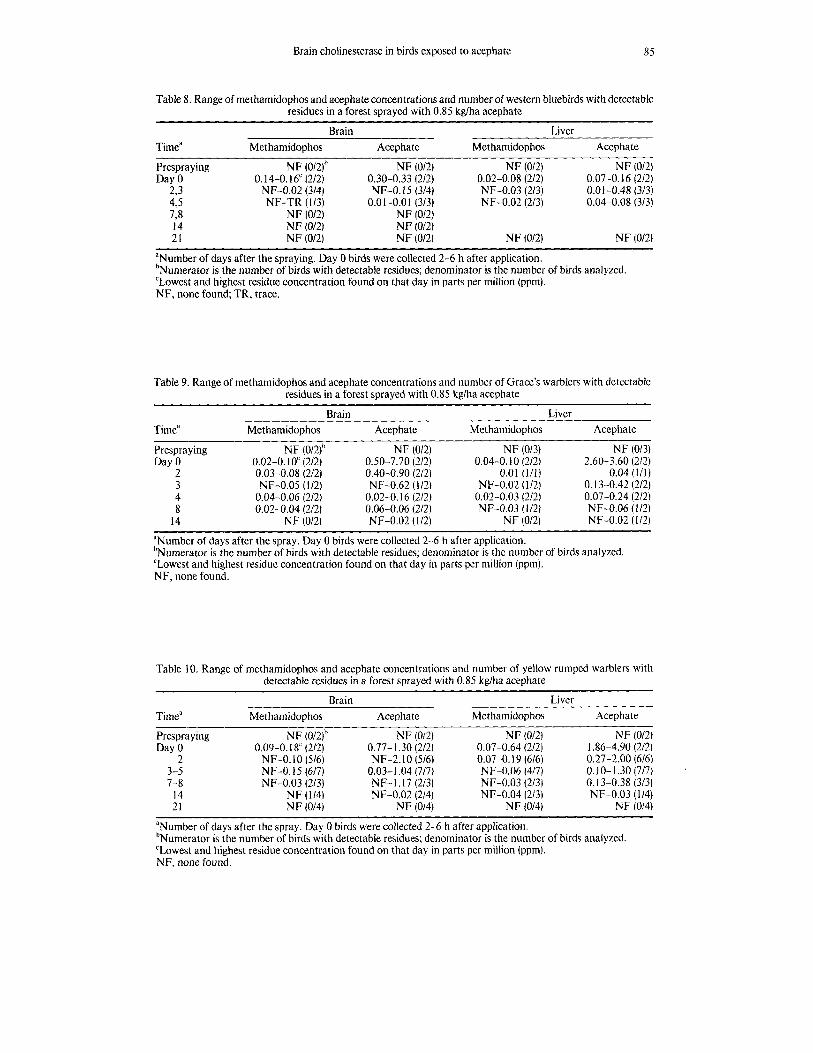
Brain cholinesterase in birds exposed to acephate 85
Table 8. Range of methamidophos and acephate concentrations and number of western bluebirds with detectable residues in a forest sprayed with 0.85 kglha acephate
Brain Liver Timea Methamidophos Acephate Methamidophos Acephate
Prespray ing NF (0/2ib NF (012) NF (012) NF (012) Day 0 0.14-0.16‘ (212) 0.30-0.33 (212) 0.02-0.08 (2/2) 0.07-0.16 (212)
NF-0.02 (314) NF-0.15 (314) NF-0.03 (213) 0.01-0.48 (313) NF-TR (113) 0.01-0.01 (3/3) NF-0.02 (2/3) 0.04-0.08 (313)
2,3 4 s 7,s N F (012) NF (012) 14 N F (012) NF (012) 21 N F (012) NF (012) NF (012) NF (0/2)
aNumber of days after the spraying. Day 0 birds were collected 2-6 h after application. bNumerator is the number of birds with detectable residues; denominator is the number of birds analyzed. ‘Lowest and highest residue concentration found on that day in parts per million (ppm). NF, none found; TR, trace.
Table 9. Range of methamidophos and acephate concentrations and number of Grace’s warblers with detectable residues in a forest sprayed with 0.85 kglha acephate
Brain Liver Time“ Methamidophos Acephate Met hamidophos
Prespraying NF (012)’ NF (012) NF (013) Day 0 0.02-0.10‘ (212) 0.50-7.70 (212) 0.04-0.10 (212)
2 0.03-0.08 (212) 0.40-0.90 (212) 0.01 (111) 3 NF-0.05 (112) NF-0.62 (112) NF-0.02 (1/2) 4 0.04-0.06 (212) 0.02-0.16 (2/2) 0.02-0.03 (212) 8 0.02-0.04 (212) 0.06-0.06 (212) NF-0.03 ( 112)
NF-0.02 (112) NF (012) 14 NF (0121
Acephate
NF (013)
0.04 (1/1) 2.60-3.60 (212)
0.13-0.42 (212) 0.07-0.24 (212) NF-0.06 (112) NF-0.02 (112)
‘Number of days after the spray. Day 0 birds were collected 2-6 h after application. ’Numerator is the number of birds with detectable residues; denominator is the number of birds analyzed ‘Lowest and highest residue concentration found on that day in parts per million (ppm). NF. none found.
Table 10. Range of methamidophos and acephate concentrations and number of yellow-rumped warblers with detectable residues in a forest sprayed with 0.85 kg/ha acephate
Brain Liver Timea Methamidophos Acephate Methamidophos Acephate
Prespra ying NF (012)h NF (012) NF (012) NF (012) Day 0 0.09-0.18‘ (212) 0.77-1.30 (212) 0.07-0.64 (212) 1.86-4.90 (212)
0.07-0.19 (616) 0.27-2.00 (616) 3-5 NF-0.15 (617) 0.03-1.04 (717) NF-0.06 (4/7) 0.10- 1.30 (717) 7-8 NF-0.03 (213) NF- 1.17 (213) NF-0.03 (213) 0.13-0.38 (313)
14 NF (114) NF-0.02 (214) NF-0.04 (213) NF-0.03 (114)
2 NF-0.10 (516) NF-2.10 (516)
21 NF (014) NF (014) NF (014) NF (014)
aNumber of days after the spray. Day 0 birds were collected 2-6 h after application. ‘Numerator is the number of birds with detectable residues; denominator is the number of birds analyzed ‘Lowest and highest residue concentration found on that day in parts per million (ppm). NF, none found.

J. G. ZINKL ET AL. 86
Table 11. Range of methamidophos and acephate concentrations and number of pygmy nuthatches with detectable residues in a forest sprayed with 0.85 kg/ha acephate
Brain Liver Timea Methamidophos Acephate Methamidophos Acephate
Prespraying NF (0/4)b NF (014) NF (014) NF (014) Day 2 NF-0.06' (112) NF-0.35 (112) 0.18-0.19 (212) 0.96-2.10 (212)
3 NF-0.04 (214) NF-0. I2 (214) NF-0.04 (214) NF-0.18 (214)
"Number of days after the spray. Day 0 birds were collected 2-6 h after application. hNumerator is the number of birds with detectable residues; denominator is the number of birds analyzed. 'Lowest and highest residue concentration found on that day in parts per million (ppm). NF, none found.
moderate ChE depression can cause behav- ior modifications that might affect individu- al survival, then the effect on a species having a small population or one confined to a distinct habitat might be severe. Several studies have suggested that one effect of ChE inhibitors might be the alteration of foraging ability [ 15-1 71.
The plots sprayed in this study were rela- tively small (about 250 ha). However, they were probably sufficiently large to encom- pass the range of individuals of some of the species. Such birds would be exposed to ace- phate and its metabolites constantly after spraying until degradation occurred. As ex- pected, those species of birds that probably have smaller territories (grey-headed juncos, yellow-rumped warblers, Grace's warblers and pygmy nuthatches) had greater brain ChE inhibition than did more mobile species (American robins and western bluebirds). Therefore, it seems likely that the effect on the more confined species is a better indica- tor of the possible effects that may occur when very large areas are sprayed during insect control programs.
Previous work suggesting that acephate itself is not the major brain ChE inhibitor in birds exposed during spraying of the insecti- cide was reaffirmed by this study. From studies in which brain ChE activity was de- termined in American robins, bobwhite quail and blackbirds orally dosed with ace- phate and methamidophos, Booth et al. [4] concluded that a bird would have to con- sume several times its body weight of contaminated food for either of these
compounds to depress brain ChE activity to a level found in birds from forest applica- tions of acephate. Comparison of the present study and our previous field studies [ 1,3] with our laboratory studies with ace- phate and methamidophos [4] suggests that after application of 0.57 to 2.26 kglha ace- phate (0.5 to 2.0 lb/acre) it would be difficult for a bird to ingest or absorb sufficient acephate to produce 20 to 50% brain ChE inhibition. Furthermore, several birds in this study had high brain acephate and or met hamidophos concentrations but little or no brain ChE inhibition, an unlikely possi- bility if either of these compounds was the brain ChE inhibitor. Therefore, it ap- pears that acephate must be converted to a more potent ChE inhibitor (but not methamidophos) in the forest. Eto et al. [ 181 indicated that biological conversion to a sulfoxide derivative was necessary for insecticidal activity of acephate and methamidophos. Bull [19] showed that cot- ton leaves metabolized acephate to methamidophos and at least four other com- pounds, two of which were not toxic. Wer- ner [20] also found that acephate was metabolized to methamidophos, and he found an unknown compound with insecti- cidal activity. To date, no other insecticidal compound has been positively identified, and we did not detect any compounds other than acephate or methamidophos with our residue analysis techniques.
Further evidence suggesting that ace- phate per se is not the brain ChE inhibitor is provided by the time course of ChE inhibi-

Brain cholinesterase in birds exposed to acephate 87
tion that occurs after a spraying. In this study and in other studies [ 1,3], ChE inhibi- tion was not found on the day of spraying but became evident one or two days after spraying, even though residue concentra- tions were relatively high. Usually, maxi- mum ChE inhibition was found at two to three days after spraying, suggesting that a potent ChE inhibitor is produced from ace- phate one to three days after application.
It has been suggested that, when birds have brain ChE inhibition, the cause of the inhibition should be confirmed by tissue or stomach contents analysis [ 131. This study suggests that after acephate application tis- sue analysis would often fail to reveal either acephate or methamidophos because birds with brain ChE inhibition often had no de- tectable residues. These results indicate that brain ChE activity is a better indicator of exposure to acephate than is tissue residue analysis. Unfortunately, we were not able to analyze stomach contents because they were used in a dietary study [ 171. Residues of acephate and methamidophos have been found in foliage for up to 14 d after applica- tion of 0.57 kglha acephate (0.5 lblacre) in forests (R. B. Roberts, U.S. Forest Service, personal communication, 1977). Perhaps residue analysis of stomach con- tents would have greater utility than tissue residue analysis for confirming acephate exposure.
These studies confirm that acephate sprayed in forests causes brain ChE inhi- bition in birds. It appears that neither acephate nor methamidophos is the brain ChE inhibitor. The inhibitor is probably a metabolite which develops after the forests are sprayed with acephate. Brain ChE is the best way to determine if exposure has occurred. The magnitude of inhibition suggests that a few individuals may be in danger of being killed, but it is unlikely that a large number of birds will be killed after proper application of acephate. The magni- tude of depression for potential lethality is unclear, but brain ChE depression greater
than 50 to 70% should be viewed as poten- tially harmful. If sublethal effects such as decreased foraging ability occur when brain ChE is inhibited by less than 50%, then substantial numbers of birds could be affect- ed after exposure to acephate application in forests.
REFERENCES
1.
2.
3.
4.
5 .
6 .
7.
8.
9.
10.
Zinkl, J.G., C.J. Henny and P.J. Shea. 1979. Brain cholinesterase activities of passerine birds in forests sprayed with cholinesterase inhibiting insecticides. In Animals as Monitors of Environ- mental Pollutants. National Academy of Sciences, Washington, DC. Julin, A.M. and F.J. Gramlich. 1978. Effects of Orthene on songbird brain cholinesterase in Maine, 1977. Forest Insect and Disease Manage- ment Evaluation Report. Northeastern Area, State and Private Forestrv. U.S. DeDartment of Agriculture, Bromall, PA. Zinkl. J.G.. R.B. Roberts. C.J. Hennv and D.J. Lenhart. 1980. Inhibition of brain chdlinesterase activity in forest birds and squirrels exposed to aerially applied acephate. Bull. Environ. Contam. Toxicol. 241676-683. Booth, G.M., J. Woodfield and D. Simmons. 1977. The effects of Orthenem and Monitor@ on small mammal and bird brain cholinesterase activi- ty. Report to the US. Forest Service. Douglas Fir Tussock Moth Program. Activity Number FS-PNW-Grant No. 49. Zinkl, J.G., R.B. Roberts, P.J. Shea and T. Lasmanis. 1981. Toxicity of acephate and methamidophos to dark-eyed juncos. Arch. Envi- ron. Contam. Toxicol. 10: 185-192. Ellman, G.L., K.D. Courtney, V. Andres, Jr., and R.M. Featherstone. 1961. A new and rapid colorometric determination of acetylcholines- terase activity. Biochem. Pharmacol. 7:88-95. Dieter, M.P. and J.L. Ludke. 1975. Studies on combined effects of organophosphates and heavy metals in birds. 1. Plasma and brain cholinesterase in coturnix quail fed methyl mercury and orally dosed with parathion. Bull. Environ. Contam. Toxicol. 13:257-262. Mount, M.E. and F.W. Oehme. 1980. Microprocedure for determination of carbaryl in blood and tissues. J. Anal. Toxicol. 4286-292. Kleinbaum, D.G. and L.L. Kupper. 1977. Applied Regression Analysis and Other Multivariable Methods. Duxbury Press, North Scituate, MA, pp. 27 1-276. Zinkl. J.G..C.J.Hennvand L.R.DeWeese. 1977. Brain'cholinesterase aitivity of birds from forests sprayed with trichlorfon (Dylox) and carbaryl (Sevin-4-oil). Bull. Environ. Contam. Toxicol. 17379-386.

88 J. G . ZINKL ET AL.
1 1 . Ludke, J.L.,E.F.HillandM.P.Dieter. 1975.Cho- linesterase (ChE) response and related mortality among birds fed ChE inhibitors. Arch. Environ. Contam. Toxicol. 3: 1-2 1,
12. Bunyan, P.J., D.M. Jennings and A. Taylor. 1968. Organophosphate poisoning: Diagnosis of poison- ing in pheasants owing to a number of common pesticides. J. Agric. Food Chem. 16:332-339.
13. Hill, E.F. and W.J. Fleming. 1982. Anticholines- terase poisoning of birds: Field monitoring and di- agnosis of acute poisoning. Environ. Toxicol. Chem. 1:27-38.
14. Mehrotra, K.N., Y.P. Beri, S.S. Misra and A. Phokela. 1967. Physiological effects of malathion on the house sparrow Passer domesticus. Indian J. Exp. Biol. 5 2 19-22 1,
15. Pope, G.G. and P. Ward. 1972. The effects of small applications of an organophosphorous poisoning, fenthion, on the weaver bird Que/ea quelea. Pestic. Sci. 3: 197-205.
16. Grue, C.E. and B.K. Shipley. 1981. Interpreting population estimates of birds following pesticide applications-Behavior of male starlings exposed to an organophosphate pesticide. Stud. Avian Bio/.
17. Mack, P.D. 1982. The effect of an orga- nophosphate pesticide, acephate, on the diets of six bird species in a ponderosa pine forest. M.S. thesis. University of California, Davis, CA.
18. Eto, M., S. Okatbe, Y. Ozoe and K. Maekawa. 1977. Oxidative activation of 0,s-dimethyl phosphoramidothiolate. Pestic. Biochem. Physiol.
19. Bull, D.L. 1979. Fate and efficacy of acephate after application to plants and insects. J. Agric. Food Chem. 27:268-272.
20. Werner, R.D. 1974. Distribution and toxicity of root-absorbed I4C-Orthene and its metabolites in loblolly pine seedlings. J. Econ. Entorno/.
61292-296.
71367-377.
67~588-591.




















