
BIOSCOURING OF COTTON FABRICS
82
Anita K. Losonczi BIOSCOURING OF COTTON FABRICS Ph.D. Thesis Supervisor: Emília Csiszár Budapest University of Technology and Economics Department of Plastics and Rubber Technology 2004 Budapest University of Technology and Ecomomics
-
Upload
khangminh22 -
Category
Documents
-
view
0 -
download
0
Transcript of BIOSCOURING OF COTTON FABRICS

Anita K. Losonczi
BIOSCOURINGOF COTTON FABRICS
Ph.D. Thesis
Supervisor:
Emília Csiszár
Budapest University of Technology and EconomicsDepartment of Plastics and Rubber Technology
2004
Budapest University of Technology and Ecomomics

® Anita K. Losonczi
Supervisor: Emília Csiszár
All rights reserved
Published by Budapest University of Technology and EconomicsDepartment of Plastics and Rubber Technology
1111 Budapest, Mûegyetem rkp. 3.

CONTENTS
PREFACE · · · · · · · · · · · · · · · · · · · · · · · · · · · · · · · · · · · · · · · · · · · · · · · · · 5LIST OF ORIGINAL PUBLICATIONS · · · · · · · · · · · · · · · · · · · · · · · · 6ABBREVIATIONS· · · · · · · · · · · · · · · · · · · · · · · · · · · · · · · · · · · · · · · · · · 8
1. INTRODUCTION · · · · · · · · · · · · · · · · · · · · · · · · · · · · · · · · · · · · · · · 9
2. LITERATURE2.1. Structure of cotton · · · · · · · · · · · · · · · · · · · · · · · · · · · · · · · · · · · 112.2. Noncellulosic constituents of cotton · · · · · · · · · · · · · · · · · · · · · 122.3. Enzymes and their use in processing of cellulosics · · · · · · · · · 162.4. Mode of action of pectinases, cellulases and xylanases · · · · · · 182.5. Biopreparation of cotton · · · · · · · · · · · · · · · · · · · · · · · · · · · · · · 20
2.5.1. Bioscouring with pectinases . . . . . . . . . . . . . . . . . . . . . . 202.5.2. Bioscouring with cellulases . . . . . . . . . . . . . . . . . . . . . . . 222.5.3. Bioscouring with proteases . . . . . . . . . . . . . . . . . . . . . . . 222.5.4. Bioscouring with lipases . . . . . . . . . . . . . . . . . . . . . . . . . 232.5.5. Combined enzyme systems in bioscouring of cotton . . 232.5.6. Effect of reaction parameters on the efficiency of
bioscouring . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 242.6. Aims of the study · · · · · · · · · · · · · · · · · · · · · · · · · · · · · · · · · · · · 26
3. MATERIALS AND METHODS3.1. Substrates and enzymes · · · · · · · · · · · · · · · · · · · · · · · · · · · · · · · 273.2. Treatments· · · · · · · · · · · · · · · · · · · · · · · · · · · · · · · · · · · · · · · · · · 273.3. Analysis of enzymes and substrates· · · · · · · · · · · · · · · · · · · · · · 29
4. RESULTS AND DISCUSSION4.1. Bioscouring of cotton fabric· · · · · · · · · · · · · · · · · · · · · · · · · · · · 29
4.1.1. Wettability (II) . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 294.1.2. Colour differences, colour evenness
and bleachability (II) . . . . . . . . . . . . . . . . . . . . . . . . . . . . 294.1.3. Dyeability with reactive dye (II) . . . . . . . . . . . . . . . . . . 31
4.2. Bioscouring with EDTA addition · · · · · · · · · · · · · · · · · · · · · · · 334.2.1. Characteristics of cotton fabric (I) . . . . . . . . . . . . . . . . . 34
3

4.3. Role of EDTA in bioscouring of cellulosics · · · · · · · · · · · · · · · 364.3.1. Effect of EDTA on enzyme activity (III) . . . . . . . . . . . 364.3.2. Effect of EDTA on seed-coat fragment
degradation (III) and linen properties (IV) . . . . . . . . . 395. CONCLUSIONS · · · · · · · · · · · · · · · · · · · · · · · · · · · · · · · · · · · · · · · · 436. REFERENCES · · · · · · · · · · · · · · · · · · · · · · · · · · · · · · · · · · · · · · · · · 45
APPENDICESEnzymes and chelating agent in cotton pretreatment· · · · · · · · · · · 49Bleachability and dyeing properties of biopretreated
and conventionally scoured cotton fabrics · · · · · · · · · · · · · · · · 59Role of EDTA chelating agent in bioscouring of cotton · · · · · · · · 67Influence of EDTA complexing agent on biopreparation
of linen fabric · · · · · · · · · · · · · · · · · · · · · · · · · · · · · · · · · · · · · · · 77
4
Contents

PREFACE
This work was carried out at Budapest University of Technology and Eco-nomics, Department of Plastics and Rubber Technology during the years2000–2004.
I thank Professor Béla Pukánszky for providing excellent working facili-ties.
I warmly thank the personnel of the Laboratory of Textile Chemistry for afriendly and supporting atmosphere. I express my sincere thanks to my su-pervisor Dr. Emília Csiszár for introducing me to the fascinating textilebiotechnology and for her support, ideas and constructive criticismduring these years.
I express my warm thanks to Dr. György Szakács from the Department ofAgricultural Chemical Technology for his useful help and support duringmy studies.
I also express my warm thanks to Dr. László Bezúr from the Departmentof General and Analytical Chemistry for the ICP-OES measurements.
I also thank my other co-authors for all their help.
Finally, I express my warmest thanks to my husband, my parents and mybrother for their patience and support throughout this work.
5

LIST OF ORIGINAL PUBLICATIONS
This work is based on the following papers, referred to in the text by theRoman numerals given below.I. Csiszár, E., Losonczi, A., Szakács, G., Rusznák, I., Bezúr, L. and Reicher,
J. (2001) Enzymes and Chelating Agent in Cotton Pretreatment. J.Biotechnol. 89, 271–279.
II. Losonczi, A., Csiszár, E., Szakács G. and Kaarela, O. (2004) Bleachabilityand Dyeing Properties of Biopretreated and Conventionally Scoured Cot-ton Fabrics. Textile Res. J. 74(6), 501–508.
III. Losonczi, A., Csiszár, E., Szakács, G. and Bezúr, L. Role of EDTA Chelat-ing Agent in Bioscouring of Cotton. Textile Res. J. (accepted).
IV. Csiszár, E., Losonczi, A., Szakács, G., Bezúr, L. and Kustos, K. Influence ofEDTA Complexing Agent on Biopreparation of Linen Fabric. Biocatalysis& Biotransformation (accepted).
The author in the present thesis had main responsibility for the practicalwork in all publications.
Publications in Hungarian Journals1. Losonczi, A. and Csiszár, E. (2004) Komplexképzõk és alkalmazásuk a
textiliparban. Magyar Textiltechnika LVII(3), 54–57.
2. Losonczi, A. and Csiszár, E. (2003) A pektinek jellemzõje és enzimesdegradációja. Papíripar XLVII(3), 96–100.
3. Losonczi, A., Csiszár, E. and Szakács, Gy. (2003) Enzimes pamut-elõkészítés komplexképzõ jelenlétében. 2. rész. Magyar TextiltechnikaLVI(1), 22–25.
4. Losonczi, A., Csiszár, E. and Szakács, Gy. (2002) Komplexképzõk szerepeaz enzimes pamutelõkészítésben. In IN-TECH-ED ’02 Mûszaki Fejlesztés ésOktatás a Textil- és ruházati Iparban. TMTE 2002, 208-214.
5. Losonczi, A., Csiszár, E. and Szakács, Gy. (2002) Enzimes pamut-elõkészítés komplexképzõ jelenlétében. 1. rész. Magyar TextiltechnikaLV(6), 186-189.
6. Csiszár, E., Losonczi, A. and Szakács, Gy. (2000) Biotechnológia alkal-mazása a textiliparban. Magyar Textiltechnika LIII(2), 27–34.
Oral and Poster Presentations1. Losonczi, A., Csiszár, E., Szakács, G. and Bezúr, L. (2004) Role of EDTA
Chelating Agent in Bioscouring of Cotton and Linen Fabrics. 3rd AnnualWorkshop COST Action 847. Graz, Austria, June, 14–16.
6
List of Original Publications

2. Losonczi, A., Kaarela, O., Csiszár, E. and Szakács, G. (2004) Bleachability andDyeing Properties of Conventionally Scoured versus Biopretreated CottonFabrics. 3rd Annual Workshop COST Action 847. Graz, Austria, June, 14–16.
3. Losonczi, A., Kareela, O., Csiszár, E. and Szakács, G. (2004) Bleachabilityand Dyeing Properties of Biopretreated and Conventionally Scoured Cot-ton Fabrics. COST Action 847, WG 2 and WG 3 Meeting. Maribor, Slovenia,February, 26–27.
4. Losonczi, A., Csiszár, E. and Szakács Gy. (2003) A bioelõkészítés hatása apamutszövetek fehéríthetõségére és színezhetõségére. MTA TermészetesPolimerek Munkabizottság ülése. Budapest, March, 17.
5. Losonczi, A., Csiszár, E. and Szakács, G. (2002) Hydrogen Peroxide Bleach-ing of Bioscoured Cellulosic Substrates. 19th IFATCC Congress. Paris,France, October, 16–18.
6. Losonczi, A., Kaarela, O., Csiszár, E. and Szakács, G. (2002) Dyeability,Dyeing Evenness and Washfastness Properties of Bioscoured Cotton Fab-rics. COST Action 847, 2nd Annual Workshop. Como, Italy, October, 10–11.
7. Losonczi, A., Szakács, G. and Csiszár, E. (2002) Biopreparation of Cottonwith Trichoderma Enzymes. 7th International Trichoderma-GliocladiumWorkshop. Cancum, Mexico, June, 25–29.
8. Losonczi, A. and Csiszár, E. (2002) Komplexképzõk szerepe az enzimespamutelõkészítésben. IN-TECH-ED 02., 4. Nemzetközi Konferencia. Buda-pest, April, 25–26.
9. Losonczi, A., Csiszár, E. and Szakács, G. (2001) Enzymes and ChelatingAgent in Cotton Pretreatment. 1st Annual Workshop COST Action 847.Funchal, Madeira, Portugal, October, 3–5.
10. Losonczi, A., Csiszár, E., Szakács, G. and Rusznák, I. (2001) Biopreparationof cotton. 221st ACS National Meeting. San Diego, CA, April, 1–5.
11. Csiszár, E. and Losonczi, A. (2001) Környezetbarát technológiák a textil-iparban. Ipari Nyílt Nap. Budapesti Mûszaki és GazdaságtudományiEgyetem, February, 28.
12. Losonczi, A. and Csiszár, E. (2000) Enzimek alkalmazása a pamutelõkészí-tésben. MTA Természetes Polimerek Munkabizottság ülése. Budapest, Sep-tember, 25.
13. Losonczi, A. and Csiszár, E. (2000) Enzimek a pamut elõkészítésében.MTA Mûanyag Munkabizottság ülése. Budapest, May, 17.
14. Csiszár, E., Szakács, G., Losonczi, A. and Rusznák, I. (2000) Enzymes inCotton Pretreatment. Symposium on Biotechnology in the Textile Industry.Povoa do Varzim, Portugal, May, 3–7.
7
List of Original Publications

ABBREVIATIONS
CBH Cellobiohydrolase
�Cab* Chroma difference
�Eab* Colour difference
�Eab* Colour evenness
EDTA Ethylenediaminetetraacetic acid
EG Endoglucanase
EGU Endoglucanase Unit
FPA Filter Paper Activity
FPU Filter Paper Unit
IU International Unit
�L* Lightness difference
8
Abbreviations

1. INTRODUCTION
Preparation and bleaching are among the most energy- and chemical in-tensive steps of the conventional cotton finishing. About 75 % of the or-ganic pollutants arising from textile finishing are derived from prepara-tion of cotton goods. In the conventional preparatory process concen-trated sodium hydroxide solution and additional hydrogen peroxideand/or sodium hypochlorite solutions are applied for removing the impu-rities from raw cotton. Cotton fibres contain approximately 10 % byweight of noncellulosic substances, such as waxes, fats, pectins, proteins,noncellulosic polysaccharides, water-soluble inorganics, lignin-contain-ing impurities and colouring materials. Most of them are located in theouter layer of the fibre. By removing the impurities, the preparatory pro-cess yields an adequately absorbent and appropriately white material withcellulose content of 99 %, but the process generated huge amount of efflu-ent. On the fibre level, oxidative damage may occur and be reflected in alower degree of polymerisation and decreased tensile strength.
Biopreparation may be a valuable and environmentally friendly alterna-tive to harsh alkaline chemicals for preparing of cotton. In bioscouringthe removal of the noncellulosic substances can be achieved mainly byhydrolytic enzymes, such as pectinases, xylanases and cellulases. Thebioprocess has several advantages over conventional chemical scouring.Enzymes operate under mild conditions (pH, temperature) with low wa-ter consumption and act only on specific substrates.
Numerous studies have been carried out on biopreparation of cotton inthe preceding 3–4 years. The results clearly show that in bioscouring ofcotton, degradation of pectic substances is one of the most essential pro-cesses. Enzymatic degradation of pectin accelerates the removal of waxymaterials from the cotton primary wall, thus produces water wettable cot-ton. However, the degree of whiteness is often less and the process is notsuitable for removing seed-coat fragments and mote adequately. By nowit is also clear that the greatest obstacle to commercialisation ofbiopreparation is the removal of matter (stalks, leaves and seed-coatfragments) of vegetable origin.
In spite of the extensive information available and the large number of pa-pers published, numerous questions have remained open concerning theproperties of bioscoured cotton substrates generally and the degradationof pectin and seed-coat fragments particularly. It became clear soon that
9

if we intend to explain certain phenomena, help process development orcontribute to the solution of problems emerging in the practice, we haveto concentrate on the basic problem of cotton bioscouring. As a conse-quence, the main goal of this thesis is to carry out detailed studies on cer-tain aspects of the degradation of pectin and seed-coat fragments.
Desized cotton fabric was treated with different commercial enzymes un-der a wide range of conditions and the wettability, whiteness, colour even-ness and weight loss were studied. Effect of complexing agent on the effi-ciency of the process was also determined. Attempts were made for deter-mination of the role of complexing agents in the degradation of pectin. Ina further step degradation of seed-coat fragment was investigated in de-tails. Fabrics subsequent to biopreparation are generally further sub-jected to bleaching, dyeing or printing. Therefore, it was particularly es-sential to characterise the bleachability and dyeing properties of thebiopretreated substrates. The results were compared with those obtainedin a conventional alkaline scouring.
10
1. Introduction

2. LITERATURE
2.1. Structure of cotton
Cotton is still the most important of the raw materials for the textile indus-try. Worldwide about 50 % of the fibres consumed is cotton (Lewin andPearce, 1998). Cotton grows as unicellular fibre on seeds (seed hair fibre).The mature cotton fibre forms a highly convoluted flat ribbon, varying inwidth between 12 and 20 �m. Cotton fibres have a fibrillar structure.Their morphology is illustrated in Figure 1. A thin cuticle that mainly con-sists of waxes and pectins protects the outside of the fibre. The primarywall in mature fibres is only 0.5–1 �m thick and contains about 50 % of cel-lulose. Noncellulosic constituents consist of pectins, fats and waxes, pro-teins and natural colorants. The secondary wall, containing about92–95% cellulose, is built of concentric layers with alternating S- andZ-shaped twists. The layers consist of densely packed elementary fibrils,organised into microfibrils and macrofibrils. They are held together bystrong hydrogen bonds. The lumen forms the centre of the fibres(Buschle-Diller, 2003).
Cotton is composed almost entirely of the polysaccharide cellulose. Cellu-lose is the most abundant renewable polymer today. Chemical composi-tion of cellulose is simple; it is a linear (1�4)-linked polymer of�-D-glucopyranose (Lewin and Pearce, 1998). The degree of polymerisa-tion (DP) of cellulose varies with its source and the processing stage ofthe cellulosic material. The DP of native cellulose may be as high as 14000, but the different purification treatments can reduce this to1000–2000. Three free hydroxyl groups in C2, C3 and C6 per
11
Figure 1. Structure of cotton (Nevell, 1995).

anhydroglucose unit are available for formation of strong inter- andintramolecular hydrogen bonds. Each cellulose chain has a non-reducingend group at C4 and a reducing end at C1. The latter exhibits the charac-teristics of both an alcohol and an aldehyde under appropriate conditions(Nevell, 1995).
2.2. Noncellulosic constituents of cotton
Raw cotton fibre, after ginning and mechanical cleaning, is essentially90–95 % cellulose (typical cotton fibre). The mature cotton fibre has anoncellulosic covering called cuticle that contains waxes, pectins, and pro-teins left over from biosynthesis. This cuticle is deposited on the primarywall, the first part of the cell wall to be formed (Lewin and Pearce, 1998).The primary wall is about 0.1 �m thick and comprises only about 1 % ofthe total thickness of cotton fibre. It is illustrated schematically by thecut-out diagram given in Figure 2.
The major portion of the noncellulosic constituents of cotton fibre is pres-ent in or near the primary wall. The exact percentage of each componentof the primary wall is determined by the type of the cotton plant, its origin,the growth conditions, and the degree of maturity (Lewin and Pearce,1998). The composition of a mature cotton fibre is given in Table 1.
12
2. Literature
Figure 2. Cotton fibre primary wall matrix, illustrating the concentrationgradient of noncellulosic material

Table 1. Composition of the components identified in cotton fibresand in the cuticle (Lewin and Pearce, 1998; Hardin et al., 2004).
ComponentsComposition
of the whole fibre[% of the dry weight]
Cuticle[%]
Cellulose 88 – 96
Protein 1.1 – 1.9 30.4
Pectin 0.7 – 1.2 19.6
Wax 0.4 – 1.0 17.4
Ash 0.7 – 1.6 6.5
Others (e.g. seed-coat fragment) 0.5 – 1.0
Of all the noncellulosic constituents, the nitrogen-containing compoundsconstitute the largest percentage when expresses as percent protein. Thenitrogenous material occurs principally in the lumen of the fibre, mostlikely being protoplasmic residue; a small portion is also extracted fromthe primary wall. Cotton fibre and its primary wall both contain pro-teins/peptides, free amino acids, and most likely nonprotein nitrogen.The free amino acids that have been detected are glutamic acid, asparticacid, valine, serine, and threonine (Lewin and Pearce, 1998). It is sug-gested that the brown colour of cotton and dark brown colour of kier li-quors are associated with proteins (Ward, 1955).
Pectin is located mostly in the primary wall of the fibre. It is composed of ahigh proportion of D-galacturonic acid residues, joined together by�(1�4) linkages. The carboxylic acid groups of some of the galacturonicacid residues are partly esterified with methanol (Dey and Harborne,1997). Pectic molecule can be called a block-copolymer with alternatingthe esterified and the non-esterified blocks (Jarvis et al., 1984). In the pri-mary cell wall pectin is covalently linked to cellulose or in other plants tohemicellulose, or that is strongly hydrogen-bonded to other components.On the other hand, pectin undergoes self-interaction to some extent, itcan forms crossbridges by the help of calcium ions (Dey and Harborne,1997). The crosslinks are formed between the non-esterified or littlemethyl-esterified galacturonan blocks with negative charge and calciumion with positive charge by electrostatic interactions (“egg-box” model)(Demarty et al., 1984; Dey and Harborne, 1997).
13
2.2. Noncellulosic constituents of cotton

All the waxes take place on the surfaces of the cotton fibre, in the primarywall, which is responsible for the hydrophobic nature of unscoured cot-ton. The waxes consist of various hydrocarbons, fatty alcohols, fatty acidsand their esters. The presence of waxes is detrimental in the chemical pro-cessing of cotton yarns and fabrics because it interferes with wetting of thefibre and penetration of reagents (bleaching agent, dyes, etc.). Afterscouring the wax content of the fibre is about 0.15 %. Because of itsinhomogeneous distribution, the fibre has a hydrophilic character (Lewinand Pearce, 1998; Trotman, 1968).
The inorganic salts (phosphates, carbonates and oxides) and salts of or-ganic acids present in raw fibre are reported as percent ash. The ash itselfis highly alkaline. The ash contents of cottons are highly variable in com-position and quantity, arising in differences in soil and agricultural prac-tices. During the production of cotton, the plant absorbs potassium andother metals as normal nutrients. The following metals are found in rawcotton fibre: potassium, calcium, magnesium, sodium, iron, manganese,copper, zinc. Metals in cotton are of importance of processors, becausethey can contribute to problems in bleaching and dyeing. Decompositionof peroxide can be catalysed by iron and copper ions, which may induce fi-bre damage. Insoluble calcium and magnesium salts can interfere withdyeing (Lewin and Pearce, 1998).
14
2. Literature
Table 2. Chemical composition of cotton seed-coat fragments(Csiszár et al., 1987).
Components Amount [%]
Wax 5.6
Lignin 28.3
Holocellulose, including 41.3
�-cellulose 23.0
pentosans 7.2
Ash 12.8
Moisture 8.3
Others 3.7

Seed-coat fragments (Figure 3) present on the surface of cotton fabric arethe most resistant impurities of cotton. Seed-coat fragments are part ofthe outer layer of cotton seed and they are formed from mature or imma-ture seeds during mechanical processing. They are usually black or darkbrown and may or may not have fibres and linters attached (Annual Bookof ASTM Standards, 1980; Verschraege, 1989). Chemically seed-coatfragments consist of two main components: lignin and holocellulose (Ta-ble 2). The inorganic content of seed-coat fragments is significant, thereare potassium, calcium, and magnesium compounds in large scale(Csiszár et al., 1987).
Similarly to other plants, hemicelluloses are also present in seed-coat frag-ment. They are polymeric molecules that contain components other thanglucose, such as xylose or galactose residues. Shorter oligomeric mole-cules are also present. These molecules coexist readily with cellulose buthave different properties, especially solubility (Lewin and Pearce, 1998).
Seed coat fragments are more troublesome in pretreatment than theother non-cellulosic constituents of the cotton fibres. Although, the differ-ent mechanical cleaning steps remove majority of the seed-coat frag-ments, a certain amount of these impurities may adhere to the fibres andmay be present even in the final fabric. In the traditional chemical pro-cesses their removal requires more concentrated chemical solutions andlonger steaming periods.
15
2.2. Noncellulosic constituents of cotton
Figure 3. Cotton seed-coatfragment on the surfaceof cotton fabric(light microscopy).

2.3. Enzymes and their use in processing of cellulosics
Enzymes are high molecular weight proteins that are produced by livingorganisms. They are capable of catalysing chemical reactions in biologicalprocesses and hence are known as ‘biocatalysts’. Generally enzymes con-tain an ‘active site’, where the reaction is carried out, and a ‘binding site’,which links to the substrate. In order to catalyse a reaction, the enzymemolecule has to form a complex with the substrate. The enzyme has aquite specific three-dimensional shape. This shape and other factors, suchas the location of the active site on the enzyme, control the substratespecificity of the molecule.
The enzyme is connected to the substrate by ‘lock and key mechanism’(Figure 4). The bioreaction takes place in the enzyme-substrate complex re-ducing the activation energy, and as a consequence, the reaction speed ismultiplied. Finally, the complex disintegrates releasing the reaction prod-ucts, and the original enzyme is available again. Generally enzymes work inmild reaction conditions: atmospheric pressure, low temperature and slightacidic, neutral or alkaline pH. They are very sensitive molecules, thereforethe assurance of the optimal reaction conditions is needed during thewhole process (Shukla et al., 2000; Etters and Annis, 1998).
The efficiency of the enzyme-catalysed reactions depends on a number offactors. Enzymes can work with maximum activity only in a specific nar-row range of temperature. Increasing the temperature, the activity in-
16
2. Literature
Figure 4. “Lock and key” mechanism for enzyme action.Enzyme is absorbed at substrate, followed by release of products
of reaction and enzyme (Etters and Annis, 1998).

creases due to increased possibilities of the enzymes coming closer to sub-strate molecules. On the other hand, denaturation of the enzyme proteindue to heat is also accelerated. Most enzymatic activities are extremelysensitive to pH. Any increase or decrease of pH results in lowered reac-tion rates. A variation of pH during the reaction may bring about an alter-ation of the protein structure with a denaturing effect or the ionisation ofthe active site. In order to avoid deviations of the pH during the reactionit is suggested to buffer the medium. Certain enzymes require some spe-cific bivalent metallic cations as activators that stabilise the structure ofthe enzyme-substrate complex. Certain chemicals such as alkalis, acid lib-erating agents, heavy metal cations, oxidising or reducing agents, how-ever, acts as inhibitors of the enzymes blocking or destroying the usefulgroups of the molecules (Shukla et al., 2000).
Textile industry uses various chemical agents in the different wet pro-cesses. These chemicals, after their use, cause pollution in the effluents;some of them are corrosive that could damage equipment and the sub-strate itself. However, by introducing enzymatic processes an environ-ment friendly production can be ensured.
The serious wastewater pollution caused by conventional textile finishinghas oriented the research towards application of enzymes in textile wetprocesses. The enzymes are non-toxic and environmentally sound. Beingnatural products, they are completely biodegradable and accomplishtheir work quietly and efficiently without leaving any pollutant behind.One of the oldest technology being used today is based on amy-lase-catalysed hydrolysis of the starch size. And in the last two decades sev-eral other enzymatic processes have also been developed for the differentwet processing of textile goods in wide-ranging operations from cleaningpreparations to finishing processes. Cellulases, hemicellulases andpectinases (hydrolyses) acting on native cellulosic fibres (cotton, flax,hemp, jute, etc.) became the target enzymes in textile bioprocessing.
The production of “aged” denim garments with cellulases is the most suc-cessful enzyme process that has emerged in the textile industry in the lastdecades. Each year approx. 1 billion jeans are produced and a big part ofit is further finished to stonewashed fashion. Cellulase enzyme removesthe cotton fragments from the surface of the garment and facilitates theabrasion of indigo dye from the fibre surface. Traditional stonewashingprocess carried out by pumice stones was vastly replaced by enzymaticbiostoning and fading or combined procedures (Lange et al., 1993).
17
2.3. Enzymes and their use in processing of cellulosics

Biofinishing or biopolishing represents another application of cellulasesfor non-denim cellulosic fabrics and garments. To various extents, all cel-lulosic materials have a tendency to fuzz formation. Fuzz is the designa-tion for short fibres protruding from the surface of the yarn and fabrics.Traditionally, chemical agents (cationic surface-active compounds) areused to decrease the fuzz formation. In the environmentally-friendly pro-cess cellulosic fabrics are subjected to a treatment with cellulolytic en-zymes, and the action of cellulases removes the small loose fibre ends thatprotrude from the fabric surface as well as improves handle properties ofthe fabrics (Cavaco-Paulo et al., 1998).
Xylanase enzymes have not been applied in the textile procedures before,but they are well-known in the pulp and paper industry. Xylanase treat-ment of the pulp means an indirect bleaching method, rendering thefibres more accessible to bleaching chemicals and leading to a more effi-cient delignification. Lignin-carbohydrate linkages have been proposedto restrict the chemical removal of lignin from the pulp. Xylanase en-zymes attack xylan, chemically bonded to lignin or degrade reprecipitatedxylan, deposited on the fibre surface during cooking, resulting in an im-proved removal of coloured lignin substances. Xylanase enzyme-aidedbleaching is one of the most successful and widely used environmentallyfriendly biotechnological processes (Viikari et al., 1996).
Recent results indicate that certain enzymes may be used effectively inthe cleaning procedure of cotton. The scientific interest in this process isreflected in the number of papers published during recent years describ-ing biopreparation results obtained, using various enzymes from differentsources. But enzymatic biopreparation of cotton represents a fairly newapproach and is still mostly in the development stage.
In the following part the characteristic features of pectinases, cellulasesand xylanases, as well as the mode of their action on cellulosics are dis-cussed in details, since these enzymes are used in the experiments.
2.4. Mode of action of pectinases, cellulases and xylanases
Pectinases catalyse the degradation of pectin. The total degradation is re-sulted by the harmonised work of several enzymes with different activi-ties. These enzymes are in synergism with each other. There is anondepolymerase in pectin degrading enzyme system: pectin-esterase.This enzyme catalyses the cleavage of ester bond of poligalacturonan, as a
18
2. Literature

consequence the degree of esterification decreases. Free carboxyl groupand methyl alcohol are produced in the reaction (Bailey and Pessa, 1990;Henriksson et al., 1999). Polygalacturonase, which is a depolymerase en-zyme, catalyses the cleavage of �(1�4) bonds in pectic polymer chain, re-leasing water and reducing groups at the chain ends. Exopoly-galacturonase works at the end of the chain, while endopolygalacturonaseworks randomly within the chain (Bailey and Pessa, 1990; Schejter andMarcus, 1988). Transeliminase or shortly lyase (depolymerase) catalysesthe cleavage of �(1�4) bonds in polygalacturonan chain without releas-ing water and creating a double bond between the C4 and C5 atoms.Endo and exo enzymes work within or at the end of the chain, respec-tively, similar to the polygalacturonases (Sakamoto et al., 1994).
Cellulases catalyse the degradation of cellulose. All cellulases have anidentical chemical specificity towards the �(1�4) glycosidic bonds, butthey differ in terms of the site of attack on the solid substrates. This differ-ence in their mode of action has been defined as exoglucanase andendoglucanase activities. The exoglucanases, or cellobiohydrolases(CBH) cleave cellobiose units from the end of the polysaccharide chainsand typically exhibit relatively high activities on crystalline cellulose.Trichoderma reesei produces two different cellobiohydrolases. CBH I ispresumed to attack the cellulose chains from their reducing ends andCBH II from their non-reducing ends. The mechanism of action ofcellobiohydrolyses has been investigated in detailed. These enzymescatalyse the hydrolysis of the glycosidic bonds by general acid catalysis.The reaction involves two essential carboxylates, the proton donor andthe nucleophile. Depending on the spatial arrangement of these two resi-dues, the reaction proceeds either by retention or by inversion ofanomeric configuration at the C1 carbon. T. reesei CBH I has been shownto be a retaining enzyme, whereas CBH II is an inverting enzyme (Koivulaet. al., 1998).
The endoglucanases (EG) make more random cuts in the middle of thelong chains, thereby producing new chain ends for the cellobiohydrolasesto act upon. The endoglucanases seem to prefer the amorphous regionsof cellulose and in contrast to the cellobiohydrolases, can also hydrolysesubstituted celluloses, such as carboxymethylcellulose and hydroxyethyl-cellulose. Finally, �-glucosidases cleave cellobiose and other soluble oligo-saccharides to glucose, which is an important step since cellobiose is anend-product inhibitor of many cellulases (Koivula et. al., 1998).
19
2.4. Mode of action of pectinases, cellulases and xylanases

Xylanases catalyse the hydrolysis of xylan, the major constituent ofhemicellulose. Xylans are heteropolysaccharides with a homopolymericbackbone chain of (1�4)-linked �-D-xylopyranose units. The backbonemay consist of O-acetyl, �-L-arabinofuranosyl, �(1�2)-linkedglucuronic or 4-O-methylglucuronic acid substituents. Two types ofxylanases are distinguished, one is a non-branching, which does not liber-ate arabinose, while the other is a debranching, which liberates arabinosefrom the side chain substituents in addition to cleaving main chain link-ages. Endo- and exoxylanases work within or at the end of the chain, re-spectively. The hydrolysis reaction of xylan catalysed by xylanases, pro-ceeds through an acid-base mechanism with the retention of anomericconfiguration in the product. Xylans are present in a partly acetylatedform in various plants. The O-acetyl groups present at C2 and C3 posi-tions of xylosyl residues, inhibit xylanases in completely degradation,probably by steric hindrance. The synergetic action of acetylxylan esterasesand xylanases is therefore essential for the complete hydrolysis ofacetylxylans (Kulkarni et al., 1999).
2.5. Biopreparation of cotton
Raw cotton contains about 90 % of cellulose and various noncellulosicssuch as waxes, pectins, proteins, fats, lignin-containing impurities andcolouring matter. The goal of the cotton preparatory process is to removethe hydrophobic and noncellulosic components and produce highly absor-bent fibres that can be dyed and finished uniformly. In the conventionalenergy and chemical intensive process concentrated sodium hydroxide so-lution and additional hydrogen peroxide and/or sodium hypochlorite solu-tions are applied for removing the impurities from greige cotton. Themild reaction conditions offered by enzymatic treatment provide an envi-ronmentally friendly alternative. Pectinases, cellulases, proteases and li-pases have been investigated most commonly and compared to alkalinescouring.
2.5.1. Bioscouring with pectinases
Some of the researchers have reported that pectinase treatment alone re-sults in adequate wettability (Traore and Buschle-Diller, 2000; Buchertand Pere, 2000; Li and Hardin, 1998b; Hartzell and Hsieh, 1998b), how-ever, others have found only a little improvement in water absorbency(Hartzell and Hsieh, 1998a). Tzanov and his co-workers investigated twotypes of petinases: an acidic and an alkaline pectinase. Both pectinases
20
2. Literature

are found to be equally efficient in terms of improved absorbency as theconventional alkaline scouring. The alkaline pectinase has similar perfor-mance to the acidic pectinase, however the former works on lower concen-tration, which is beneficial from the economic point of view (Tzanov etal., 2001). Yachmenev and his co-workers obtained better absorbencyand whiteness after the treatment with alkaline pectinase than with acidicpectinase (Yachmenev et al., 2001).
The pectin content of cotton fibre can be decreased by about 30 % duringthe pectinase treatment. Removal of pectin results in lower amounts ofwaxes on the cotton surface and subsequently in improved water absor-bency of the fabric, which supports the hypothesis of chemical linkage be-tween pectin and waxes. The enzymatic treatment has no effect on the ten-sile strength (Buchert and Pere, 2000), moreover it enhances slightly thewhiteness (Traore and Buschle-Diller, 2000).
Pectin acts as a sort of cement or matrix that stabilises the primary cellwall of the cotton fibres. During incubation the enzymes will degrade pec-tin, thereby destabilising the structure in the outer layers. The weakenedouter layers can be removed in a subsequent wash process (Nierstrasz andWarmoeskerken, 2003).
Pectinase treatment modifies the morphology of cotton fibres. Raw cot-ton has smooth appearance with characteristic parallel ridges andgrooves. The smooth surface without irregularities is a reflection of theamorphous stage of the cuticle. After pectinase treatment the fibre sur-face becomes perforated at first, and further treatment results in cellulosefibrils protruding from the surface of the fibre. The cause of the changescan be proposed as pectinase digesting the pectins, which are part of theconnecting structure holding the cuticle on the fibre surface (Li andHardin, 1998a).
Pectinase treatment subsequent to a hot water pretreatment can furtherimprove the wettability of the fabric. Waxes have a melting point about70�C, therefore during pretreatment they melt and disperse into the treat-ment bath or they are redistributed on the fibre surface. The thickness ofthe fabrics increases, moreover lightness and weight loss are slightly in-crease by the water pretreatment. Subsequent pectinase treatment resultsin further changes neither in thickness, nor lightness. Yarn strength is notaltered by the enzyme treatment (Hartzell and Hsieh, 1998a, 1998b).
21
2.5. Biopreparation of cotton

Sorption and dyeing properties of the pectinase scoured cotton fabricshave also been investigated and compared with those of the fabricspretreated with alkaline solution (Etters et al., 1999a, 1999b). Resultsprove that there is no difference in water sorption between thebioscoured and conventionally scoured fabrics. However, the wickingrate of the bioscoured fabric is greater than that of the alkaline scouredfabric. Dying with direct dyes does not result significant difference in col-our depth (K/S) and colour yield ((K/S)/c) between alkaline scoured andbioscoured samples. Uniformity of the bioscoured and dyed samples isbetter than that of the conventionally scoured and dyed substrates.
Novozymes, Bayer and Dexter Chemical Corporation have introduced anenzymatic alternative for scouring woven and knitted cotton fabrics in thetextile industry on the basis of an alkaline pectinase (Bioprep 3000) pro-duces by a genetically modified Bacillus strain. The bioscouring process re-sults in textiles being softer than those scoured in the conventional so-dium hydroxide process, however the degree of whiteness is often less andthe process is not suitable for removing seed-coat fragments and moteadequately (Nierstrasz and Warmoeskerken, 2003).
2.5.2. Bioscouring with cellulases
Cellulase enzyme treatment improves significantly the water wettabilityand water retention capacity of cotton fabrics (Li and Hardin, 1998b;Hartzell and Hsieh, 1998a). Comparing cellulase to the noncellulosic en-zymes, it is obvious that cellulose causes greater weight loss and decreasesthe yarn tenacity. Pretreatment with hot water enhances the effectivenessof cellulase, similar to pectinase. Morphology of the fibres changes signifi-cantly after the treatment. Progressive hydrolysis of cellulose after cuticleremoval results in protruding fibrils and cavities on the surfaces of thefibres. Cracks are more frequently observed in cellulase treated fibre thanin noncellulosic enzyme treated fibre (Traore and Buschle-Diller, 2000).Cellulases peel off the cuticle by digesting primary wall cellulose, thenthey have the opportunity to act on the internal cellulose layers and digestthem gradually (Li and Hardin, 1998a).
2.5.3. Bioscouring with proteases
Proteases are enzymes that catalyse the hydrolysis of proteins. Differentkinds of proteases have been investigated in cotton bioscouring (Hartzelland Hsieh, 1998a; Buchert and Pere, 2000). Results prove that proteaseshave only a slight effect on improving wettability, however 50 % of the pro-
22
2. Literature

tein can be removed from the fibres. Protease appears to be effective in re-moving the proteinaceous materials only from the lumen but not from thesurface. Proteins of the surface layers are imbedded beneath the pectinmaterials, therefore they are inaccessible for the enzymes. However, Linand his co-worker have shown that there are some proteases, which im-prove fabric wettability to the same level as alkaline scouring. These pro-teases are capable of accessing the surface proteins without any pretreat-ment (e.g. hot water) to alter the surface waxes to dislodge thehydrophobic surface compounds (Lin and Hsieh, 2001).
Protease treatment causes no change in the tensile strength and surfacefriction (Buchert and Pere, 2000; Lin and Hsieh, 2001). Moreover, someproteases improve the lightness of the fabric supporting the role of pro-tein in the coloration of cotton (Buchert and Pere, 2000). Others produceslight yellowness (Lin and Hsieh, 2001).
2.5.4. Bioscouring with lipases
Lipases catalyse the hydrolysis of fats. Common fats are esters of glyceroland fatty acids. Lipases attack the ester bonds regenerating water-solubleglycerol and water-insoluble fatty acid. Lipase treatment does not im-prove water wetting and retention properties of cotton. It may be ex-plained by either the low accessibility of waxes in the fibres, or the less sus-ceptibility of waxes to enzymatic action (Hartzell and Hsieh, 1998a;Buchert et al., 1998). Lipase treatment results in reduced friction coeffi-cient. It does not affect the yarn strength (Buchert et al., 1998).
2.5.5. Combined enzyme systems in bioscouring of cotton
Using different kind of enzymes in the same treatment bath synergetic ef-fect can be observed in the bioscouring process. Pectinase and cellulaseenzymes are applied together most frequently. The combined treatmentimproves the wetting properties of the cotton fabrics significantly and re-quires lower amounts of enzymes (Li and Hardin, 1998b). Cellulase appar-ently assists the action of pectinase by increasing the accessibility to thepectin materials. Removing the pectins should release othernoncellulosic components residing on the fibre surfaces (Hartzell andHsieh, 1998a). In other words by cleaving the biological glue (pectates) inthe primary cell wall, pectinase makes the noncellulosics more easilyremovable (Durden et al., 2001).
23
2.5. Biopreparation of cotton

Pectinase in combination with cellulase or lipase or xylanase was also in-vestigated. After each combined treatment better absorbency is obtainedthan the pectinase treatment alone. The best wettability is measuredwhen all the enzymes are applied in the same treatment bath. Analysis ofthe pectin with Ruthenium Red does not indicate significant differencesin pectin removal. All samples show small amounts of pectins and pro-teins unevenly distributed on the fibre surface. The whiteness of the fab-rics is slightly enhanced, and the dyeing with direct dyes yields uniform re-sults and comparable colour strength. Each sample shows significantlysofter hand with basically unchanged tensile properties, except for thetreatments where cellulase is also present in the treatment bath (Traoreand Buschle-Diller, 2000).
2.5.6. Effect of reaction parameters on the efficiency of bioscouring
Many reports confirm that the surfactant has a major role during enzy-matic process (Li and Hardin, 1998b; Lenting et al., 2002; Sawada et al.,1998, 2001). Ionic surfactants are particularly effective enzymedenaturants. Anionic surfactants may form complexes with proteins anddisrupt their structure. Cationic surfactants may also form complexeswith proteins, but reduced affinity. However, nonionic surfactants arecompatible with enzymes, they do not reduce the catalytic activity, unlesstheir concentrations in the solution greatly exceed their critical micelleconcentration. The surfactant reduces the surface tension of cotton assist-ing enzymes to penetrate micropores or cracks and helping them to orientthemselves in favourable positions for the catalytic functions. After the re-action the enzymes re-enter the solution to bind to new sites again withassistance of the nonionic surfactants.
Lenting and his co-workers have reported recently that after thepectinase treatment of cotton fabrics in the presence of a surfactant, themeasured wetting time is the result of two contrary phenomena: the ab-sorption of the surfactant on the hydrophobic cuticle and the removal ofthe cuticle substances upon pectin hydrolysis. Moreover the impact of rins-ing temperature after enzymatic treatment is more than that of thesurfactant. Increasing the rinsing temperature the hydrophobic materialremoval overrules the impact of surfactant absorption and the absor-bency improves. This result is related to the melting point of the differentwax compounds present in the cuticle. A combination of a high surfactantconcentration in the enzymatic incubation and high temperatures in therinsing process afterward could result in adequate absorbency. The mostcrucial factor in the whole bioscouring process is the efficient removal of
24
2. Literature

the enzyme-weakened cuticle compounds in the rinsing phase. In combi-nation with an optimal rinsing process, a processing time of severalminutes of enzymatic degradation of pectin will be sufficient (Lenting etal., 2002).
Effect of mechanical agitation on the effectiveness of the enzyme treat-ment has also investigated in details (Hartzell and Durrant, 1998c; Li andHardin, 1998a; Traore and Buschle-Diller, 2000; Cavaco-Paulo et al.,1996). Generally it is found that agitation may increase the enzyme activ-ity and the efficiency of scouring. However, using mechanical agitationwith high shear forces, the enzyme can be deactivated, presumably by dis-rupting the characteristic three dimensional structure that makes the en-zyme substrate specific. The positive effect of agitation may be attributedto the changes in boundary layer. Sorption or desorption of diffusants onthe fibre can be influenced by the rate of flow of the processing bath pastthe fibre surface. The reason for this effect is found in the existence of aflow related hydrodynamic boundary layer. As agitation increases bound-ary layer effects become small and diffusion of the decomposition prod-ucts from the reaction sites is not impeded by boundary layers, resulting infaster and more efficient catalytic action (Li and Hardin, 1998a).
Harzell and her co-worker have generated fabric-to-fabric agitation treat-ing two samples in a single reaction container with pectinase enzyme. Thewater absorbency values were equal to those obtained from single treatedfabrics, despite a 50 % reduction in enzyme concentration. Furthermorefabric-to-fabric agitation does not cause any additional loss in strength, te-nacity or elongation (Hartzell and Durrant, 1998c).
Another possibility for improving the enzyme transport to the substratesurface beside mechanical agitation is the sonication. Introducing ultra-sonic energy to the treatment bath results in very small bubbles in the liq-uid, which generate powerful shock waves when they collapse. It causespowerful agitation in the liquid border layer and as a consequence, it im-proves the transport of the bulky enzyme molecules towards the fibre sur-face and increases the overall reaction rate. At laboratory scale, introduc-ing ultrasonic energy into the reaction chamber during enzymatic scour-ing of raw cotton fabric significantly improves the pectinase efficiency.Adequate water absorbency was observed without loss of tensile strengthof the fabrics (Yachmenev et al., 2001).
25
2.5. Biopreparation of cotton

2.6. Aims of the study
The aim of this work was to investigate the enzymatic degradation of theimpurities in greige cotton fabric in order to achieve an adequately absor-bent and appropriately white substrate with minimum impacts such asstrength and weight losses.
The specific topics of the study were:
� To investigate the effect of different commercial enzymes, i.e. cellu-lase, hemicellulase-pectinase and xylanase enzymes on biopreparationof cotton fabrics.
� To study the synergetic effect between enzyme and chelator on degrada-tion of the pectin and lignin-containing impurities (seed-coat frag-ments) of cotton.
� To investigate the bleachability and dyeing properties of the biopre-treated cotton fabric.
26
2. Literature

3. MATERIALS AND METHODS
3.1. Substrates and enzymes
Greige cotton print cloth fabric obtained from Testfabric Inc., NJ. was usedfor the tests after amylase enzymatic desizing. Spinning blowroom waste (acombined waste produced during the mechanical steps of cotton purifica-tion) was selected to model the impurities. It consists of stalks, leaves andseed-coat fragments of the cotton Before enzymatic treatment, the sub-strate was washed in distilled water to extract the water-soluble constitu-ents and air dried. 100 per cent greige linen fabric, plain-weave, supplied byPannon-Flax Linen Weaving Co. Gyõr, Hungary, was also tested as an ad-ditional substrate for studying the effect of EDTA on the pectindegradation in bioscouring.
The applied enzymes were an acidic cellulase (Celluclast 1.5 L, Tricho-derma reesei origin), an acidic pectinase (Viscozyme 120 L, Aspergillus sp.origin) and a pure xylanase (Pulpzyme HC, Bacillus sp. origin). They weregifts from Novozymes, Copenhagen, Denmark. Enzyme activities aregiven in chapter 4.3.1. in Table 3.
3.2. Treatments
Enzymatic treatments were carried out in non-agitated and agitated sys-tems, respectively, at pH 5.0 for the enzymes Celluclast 1.5L andViscozyme 120L, or at pH 7.0 for the enzyme Pulpzyme HC; enzyme con-centration of 1–4 g/l; treatment times of 1 h; incubation temperature of 50°C; and a nonionic surfactant concentration of 1 g/l. In the agitated systemthe enzymatic treatment was accomplished in a Scourotester laboratoryunit. When the effect of the chelating agent was tested, ethylene-diaminetetraacetic acid (EDTA) was added to the enzyme solution (I). In-vestigating the role of EDTA, different kinds of treatments have been ap-plied: EDTA-only, enzyme-only, EDTA pretreatment and subsequent en-zymatic treatment, without drying between the two steps and enzyme andEDTA applied together in one bath (III, IV). After enzymatic treatmentthe substrates were washed in hot distilled water to deactivate the enzymeand then air-dried. Reference treatments were carried out similarlywithout enzyme addition.
27

Caustic scouring was performed in a Mathis Labomat Typ BFA12 equip-ment with 50 g/l sodium hydroxide solution. Hydrogen peroxide bleachingwas carried out in the same equipment, the bleaching solution contained2 ml/l of 30 % hydrogen peroxide (II).
Dyeing was carried out in the Mathis Labomat by bath exhaustion proce-dure, with a liquor ratio of 1:20. A heterobifunctional reactive dye (DyeA, Sumitomo Co. Japan) was used at four dye concentrations (0.2, 0.5, 1.0and 2.0 %) based on the weight of the cotton fabric. After dyeing the sam-ples were rinsed in distilled water and air dried (II).
3.3. Analysis of enzymes and substrates
The enzyme activities were measured by internationally recognised meth-ods (Bailey and Nevalainen, 1981; Bailey et al. 1992; Ghose et al., 1987;Kubicek et al., 1982; Miller et al., 1959; Schejter and Marcus, 1988). Theinfluence of EDTA on the activity of the enzymes was tested by perform-ing the standard activity assay in the presence of EDTA at the EDTA-en-zyme ratio of 1; 3; 5; 6; 8.6 mM EDTA/ml enzyme.
Weight loss of the substrates was determined by weighing the samples(i.e. fabrics, seed-coat fragments) before and after the treatment follow-ing 24 h of conditioning at 20 °C and 65% rh. Release of reducing sugarsduring the enzymatic treatments was determined in the reaction liquor us-ing the 3,5-dinitrosalicylic acid reagent (Miller et al., 1959). Fabricwettability was determined by water dropping test, counting the elapsedseconds between the contact of the water drop with the fabric and the dis-appearance of the drop into the fabric. Colour measurement was per-formed by using a Hunterlab Color QUEST (D65/10�) colorimeter. Thecolour evaluation was done according to the CIELab colour space. Thesubstrates were characterised by L* a* b* colour coordinates. The effi-ciency of the processes (e. g.: pretreatment, bleaching), as well as the di-vergence between the reference and enzymatic treated samples were char-acterised by the colour difference (�Ea,b
*) values. Colour evenness of thetreated fabrics and fastness to washing of the dyed samples were also inves-tigated (II). The metal ion content of the substrates and the enzymes wasdetermined by inductively coupled plasma optical emission method(ICP-OES) (I, III, IV).
3. Materials and Methods
28

4. RESULTS AND DISCUSSION
4.1. Bioscouring of cotton fabric
Greige cotton contains various noncellulosics such as waxes, pectins, pro-teins, organic acids, lignin-containing impurities and colouring matter.The goal of the cotton preparation process is to remove these hydropho-bic noncellulosic components and produce highly absorbent fibres withappropriate whiteness. Wettability, absorbency, whiteness and unifor-mity of the pretreated fabrics have significant effect on the efficiency ofthe subsequent processes, therefore these parameters have beeninvestigated after bioscouring in details.
4.1.1. Wettability (II)
Greige cotton fabric is hydrophobic and it can be characterized by a wet-ting time of more than 60 seconds. Desized cotton fabric is also hydropho-bic, but it has a measurable wetting time near 10 seconds owing to thedesizing process with amylase and surfactant. All of the enzyme treatedfabrics are hydrophilic and each has a wetting time less than one second.It can be stated that the enzymatic treatments in the presence of an appro-priate surfactant improve water wettability of the cotton fabric to a level,which is adequate to dyeing, printing and finishing, as well as result in ho-mogeneously absorbent fabric. The water absorbency achieved with en-zyme is sufficient and equal to that obtained by conventional scouringprocess.
4.1.2. Colour differences, colour evenness and bleachability (II)
Removal of natural colouring matters from the cotton fibres was evalu-ated by colour measurement of the fabric. Conventional scouring resultsin extremely high colour difference (8.4) with desized fabric. An analysisof the constituent of �Eab
* reveals that alkaline treatment increases thelightness and decreases the colour content significantly (Figure 5). Col-our difference values of the enzyme treated samples with desized fabric,however, are significantly lower (0.4–1.1), indicating less effective re-moval of natural colouring matters from cotton fibres.
Comparing the colour of enzyme treated fabrics with that of the alkalinescoured fabric, we see dramatic colour differences (Figure 6). Convention-ally scoured fabric is significantly lighter and less coloured than all of thepretreated samples.
29

30
4. Results and Discussion
Figure 5. Lightness and chroma differences of the conventionally scouredand biopretreated samples with desized fabric.
Figure 6. Colour difference between biopretreated and conventionally scouredfabrics after bioscouring and subsequent hydrogen peroxide bleaching.

Treatments with enzymes do not influence the colour evenness of the fab-rics and do not cause inhomogenities in the substrates. Most of the calcu-lated �E
ab
* values are 0.2, thus, human eye can not make differences incolour of the pretreated fabric samples. It means that there are not visu-ally noticeable differences in colour of the pretreated samples.
The effect of hydrogen peroxide bleaching applied after the differentpretreatments (enzymatic, caustic scouring) is examined on the fabricproperties. Comparing the colour differences between enzyme treatedand conventionally scoured fabrics, measured after pretreatment as wellas after hydrogen peroxide bleaching (Figure 6), it becomes obvious thathydrogen peroxide bleaching significantly reduced the colour differencesbetween the alkaline and biopretreated samples. The results also showthat bleachability of the biopretreated fabrics is significantly better thanthat of the conventionally scoured sample. Colour evenness of thebleached fabrics is 0.2 or below, so all the treated fabrics arehomogeneous in colour.
It can be summarised that application of a hydrogen peroxide bleachingstep subsequent to bioscouring is beneficial, because it reduces the largecolour differences that exist between the conventionally scoured andbiopretreated cotton fabric samples.
4.1.3. Dyeability with reactive dye (II)
Insufficient preparation can cause uneven dyeing, therefore careful pre-treatment before dyeing is very important. It should ensure that the sub-strate has a high and uniform dye uptake and absorbency (Hickman,1995). Although our results prove also that absorbency and colour even-ness of the fabrics pretreated with enzyme are good enough and equal tothose of the conventionally scoured fabric, it is worthwhile to characterisethe dyeing behaviour of the biopretreated fabrics, and to compare theircolour with that of the alkaline scoured and dyed fabric in detail. Elucida-tion of the effect of the subsequent bleaching step on the dyeing behav-iour of the biopretreated fabrics is also very important.
Figures 7a and b illustrate the colour differences at 0.2 and 2 % dye con-centrations, respectively. It is obvious from the data that without bleach-ing, the colour differences that exist between the alkaline scoured andbioscoured samples after dyeing are significantly greater than those forthe dyed fabrics, which are bleached after the pretreatment. Data alsoshow that colour differences depend largely on dye concentration. The
31
4.1. Bioscouring of cotton fabric

32
4. Results and Discussion
Figure 7. Colour differences between bioscoured andconventionally scoured cotton fabrics after reactive dyeing.
Effect of dye concentration (a) 0.2 %, b) 2 %) and bleaching.

higher the dye concentration, the lower the colour difference between thealkaline scoured and bioscoured samples after dyeing.
Visual evaluation of the dyed fabrics shows that without bleaching, the col-our differences at the dye concentrations of 0.2 and 0.5 % are perceptibleand too great to be acceptable. However, at higher dye concentrations(i.e., 1 and 2 %), despite the measured higher colour difference values,the bioscoured samples conform to the conventionally scoured and dyedsample. Hydrogen peroxide bleaching between pretreatment and dyeing,at all the dye concentrations investigated, yields colour differences thatare not perceptible, and the pretreated samples match the alkalinescoured sample.
All the dyed samples (with or without bleaching) appeare homogeneousin colour and free from any defects related to dyeing process. Most of the�E
ab
* values are lower than 0.5, which is considered to be a visually unno-ticeable colour difference. We can therefore summarise that biopre-treatment results in homogeneously scoured fabrics that can be dyed uni-formly. This means also that enzyme treated samples are ready for subse-quent dyeing without further oxidative bleaching.
Even a perfect dyeing of a biopretreated sample may be rendered worth-less if the fastness properties of the fabrics are poor. The results show thatwash fastness values of the enzyme treated and conventionally scouredfabrics, with or without application of a bleaching step, do not differ andare optimal (grade 5) both for staining and colour change. Wash fastnesstests cannot determine differences between samples pretreated in differ-ent ways, or those bleached prior to dyeing.
4.2. Bioscouring with EDTA addition
Both in bioscouring of cotton and enzyme-retting of flax, degradation ofpectic substances is the most essential process. Pectin acts as a glue in theprimary wall of cotton, keeping together the noncellulosic components,and also in flax stems, where pectin fixes the bast fibres to the epidermisand to the core as well. Pectin interacts itself forming calcium crossbridges, which link the pectin components to one another. The calciumion can bind pectin and other polysaccharides too. Consequently, re-moval of calcium ions can accelerate the degradation of calcium-richpectic substances.
33
4.2. Bioscouring with EDTA addition

Application of chelating substances to desiccated flax to enhance micro-bial-retting was first reported several years ago (Sharma, 1986). Compre-hensive research works on enzyme retting of flax have shown that amongseveral chelators tested, ethylenediaminetetraacetic acid (EDTA) is themost efficient chelator in improving enzyme retting of flax (Akin et al.,1998, 2002; Adamsen et al., 2002; Henriksson et al., 1997). Our initial as-sumption was that chelators could accelerate the removal of thenoncellulosic components of cotton fibre.It has been demonstrated recently that metal ion content of cottonseed-coat fragments – the most resistant impurities of cotton fabric – is ex-tremely high comparing to that of the cotton fibre itself (I). According toICP-OES determinations the amount of calcium ions in loose cotton fibreis about 500 ppm, whereas in seed-coat fragments is more than 25000ppm. Therefore the enzymatic degradation of cotton seed-coat fragmentscan also be accelerated by chelating.
4.2.1. Characteristics of cotton fabric (I)
Biopreparation of desized cotton fabrics with different enzymes for 1hour was investigated. The weight loss values of cotton fabrics are in therange of 0–3.0 %. Celluclast 1.5L, which exhibited high filter paper activ-ity, causes the highest cotton cellulose degradation. Except for this en-zyme, agitation has only a slight effect on the enzyme action. Convention-ally scouring gives a significantly greater weight loss, 5.6 %. Addition ofEDTA to the enzyme solution results in higher weight losses of the cottonfabric. The small weight loss values caused by the enzymes show that thenoncellulosic constituents of the cuticle are removed without significantcotton cellulose degradation.
Desized cotton fabric has an averaged lightness value of 87.5. The samevalue for conventionally scoured fabric is 90.8. Biopreparation with differ-ent enzymes with or without EDTA addition has only a slight effect on fab-ric lightness.
4.2.2. Seed-coat fragment degradation (I)
Biopreparation of cotton seed-coat fragments has been carried out simi-larly to the treatment of desized cotton fabric. Degradation of the impuri-ties is characterised by weight loss. Figure 8 shows that treatment ofseed-coat fragments with different enzymes results in significant weightloss. The treating solutions has become brown, indicating that somechromophores from the impurity have been released. The seed-coat frag-ment degradation can be enhanced significantly by EDTA addition. Themost remarkable degradation effect is observed when EDTA is combined
34
4. Results and Discussion
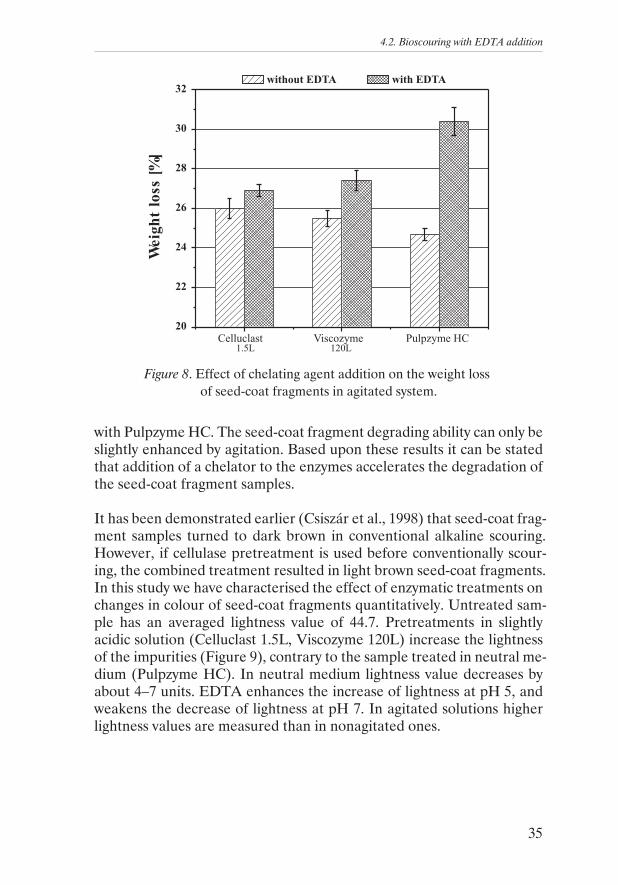
with Pulpzyme HC. The seed-coat fragment degrading ability can only beslightly enhanced by agitation. Based upon these results it can be statedthat addition of a chelator to the enzymes accelerates the degradation ofthe seed-coat fragment samples.
It has been demonstrated earlier (Csiszár et al., 1998) that seed-coat frag-ment samples turned to dark brown in conventional alkaline scouring.However, if cellulase pretreatment is used before conventionally scour-ing, the combined treatment resulted in light brown seed-coat fragments.In this study we have characterised the effect of enzymatic treatments onchanges in colour of seed-coat fragments quantitatively. Untreated sam-ple has an averaged lightness value of 44.7. Pretreatments in slightlyacidic solution (Celluclast 1.5L, Viscozyme 120L) increase the lightnessof the impurities (Figure 9), contrary to the sample treated in neutral me-dium (Pulpzyme HC). In neutral medium lightness value decreases byabout 4–7 units. EDTA enhances the increase of lightness at pH 5, andweakens the decrease of lightness at pH 7. In agitated solutions higherlightness values are measured than in nonagitated ones.
35
4.2. Bioscouring with EDTA addition
Figure 8. Effect of chelating agent addition on the weight lossof seed-coat fragments in agitated system.

4.3. Role of EDTA in bioscouring of cellulosics
Results in the previous chapter show that EDTA improves the effective-ness of commercial enzymes, as well as accelerates both the removal ofthe impurities from cotton fabric and the degradation of seed-coat frag-ments separated from the fabric. In this chapter it is explored how theEDTA can improve the efficiency of the enzyme process in bioscouring.The effects of EDTA on the apparent activities of commercial pectinaseand xylanase enzymes, and on the degradation of non-cellulosic constitu-ents in bioscouring are investigated in detail.
4.3.1. Effect of EDTA on enzyme activity (III)
Viscozyme 120L is a pectinase rich enzyme with significant 1,4-�-endo-glucanase activity (Table 3). The enzyme complex shows negligiblexylanase, cellulase and �-glucosidase activities too. Pulpzyme HC is almosta pure xylanase enzyme with practically zero cellulase (FPA), �-glucosidaseand pectinase activities under standard assay conditions; a very smallamount of 1,4-�-endoglucanase activity can be detected (Table 3). Influ-ence of EDTA on the enzyme activities of the two enzymes was investi-gated in more detail. EDTA with concentrations of 0–8.6 mM/ml enzymewas added to the enzyme solution and the standard activity assays were per-formed.
36
4. Results and Discussion
Figure 9. Effect of chelating agent addition on the lightnessof seed-coat fragments. Data based on the untreated sample.

Table 3. Activities of commercial enzymes used throughout the experiments. a
EnzymesCellulase
(FPA)[FPU/ml]
1,4-�-endo-glucanase[EGU/ml]
�-glucosi-dase
[IU/ml]
Pectinase
[IU/ml]
Xylanase
[IU/ml]
Celluclast1.5L
61(0.2)
22700(560)
14(0.2)
114(6)
417(7)
Viscozyme120L
2(0.3)
8600(80)
4(0.2)
7900(380)
110(3)
PulpzymeHC
0 50(0.8) 0 0 13800
(550)
a Standard deviations in brackets.
The influence of EDTA on the main activities of the enzymes investigatedis shown in Figure 10 and 11. For Viscozyme 120L, the pectinase activity(Figure 10) are only slightly inhibited by EDTA similarly to the1,4-�-endoglucanase and xylanase activities. On the other hand, EDTAhas no effect on cellulase (FPA) and �-glucosidase activities.
For Pulpzyme HC, EDTA does not inhibit the xylanase activity signifi-cantly (Figure 11). Although the data vary near the initial activity valuewith the EDTA concentration, but no trend can be observed. EDTAcauses measurable decrease in 1,4-�-endoglucanase activity.
It can be summarised from the results that EDTA does not inhibit signifi-cantly the main activities of the enzymes investigated, thus pectinase activ-ity of Viscozyme 120L and xylanase activity of Pulpzyme HC retain al-most unchanged in the presence of EDTA.
There are many so-called metallo-enzymes that contain metal ion in theirstructure (Kobayashi et al., 1999, 2001; Sakamoto et al., 1994; Spurway etal., 1997). By removing the metal ion by chelating, tertiary structure of theenzyme can be destroyed. As a consequence, the activity may decrease ordisappear. Analysis of the two enzymes used in this research by ICP-OESmethod show that they contain approximately equal amounts of phospho-rus, and magnesium. For Viscozyme 120L, the amount of sodium andpotassium ions, and for Pulpzyme HC, the amount of calcium ion issignificant.
37
4.3. Role of EDTA in bioscouring of cellulosics

38
4. Results and Discussion
Figure 10. Effect of EDTA on pectinase activity of Viscozyme 120L enzyme.
Figure 11. Effect of EDTA on xylanase activity of Pulpzyme HC enzyme.

Our results prove that there is no significant activity loss in the presenceof EDTA if Viscozyme 120L or Pulpzyme HC enzymes are used togetherwith EDTA. Thus, these metal ions are not part of the enzyme structure,and EDTA does not change the tertiary structure of the enzymes by che-lating. The origin of metal ions in the commercial enzymes may be attrib-uted to the medium ingredients used in fermentation and to the additivesduring the downstream processing.
4.3.2. Effect of EDTA on seed-coat fragment degradation (III) and linenproperties (IV)
Seed-coat fragment samples have been subjected to different treatments,i.e. EDTA-only, enzyme-only, enzyme subsequent to the EDTA pretreat-ment and co-application of enzyme and EDTA in one bath. The overalldegradation of the solid materials was characterised by the weight loss(Table 4). While the weight loss values for the two enzyme treatments areessentially the same, the pectinase treatment in acidic buffer decreasesthe calcium content enormously, while the xylanase treatment at pH 7causes only a milder decrease. For EDTA + Pulpzyme HC two-step treat-ment, the effect of EDTA is more remarkable than the action of enzyme.
Table 4. Effect of EDTA on cotton bioscouring. Degradation of cottonseed-coat fragments: weight loss and residual calcium ion content.
Treatments Weight loss [%] Calcium ion [ppm]
- (untreated) -— 34900
EDTA a 10.6 18500
Viscozyme 120L b 14.8 17700
EDTA + Viscozyme 120L c 22.5 13200
Viscozyme 120L with EDTA d 21.5 11900
Pulpzyme HC b 14.1 31100
EDTA + Pulpzyme HC c 25.0 16000
Pulpzyme HC with EDTA d 22.2 16800
a Treatment with EDTA. b Treatment with enzyme. c Pretreatment with EDTA and subsequent en-
zyme treatment. d Enzyme treatment with EDTA.
While weight loss data characterised the overall degradation effect of theapplied treatments, reducing sugars in the treatment filtrates reveal the ef-
39
4.3. Role of EDTA in bioscouring of cellulosics

fectiveness of the enzyme action. As Figures 12 shows, the EDTA pre-treatment decreases significantly the hydrolysis level in the subsequent en-zyme treatment, indicating changes in substrate structure by extraction ofthe divalent ions. Removal of the calcium cross bridges by EDTA pretreat-ment can cause a temporarily open structure, which can collapse by theend of the pretreatment, resulting in a less accessible structure for the en-zyme macromolecules applied subsequently. On the other hand, additionof EDTA into the enzyme solution can accelerate the hydrolysis degreeenormously (Figures 12). Removal of calcium ions from calciumcross-bridges in hemicelluloses by EDTA results in somewhat free and ac-cessible areas in the substrate where the enzyme present is prompt to actin catalysis of chain-cutting process. Thus, in this case the enzymes are ca-pable of accessing the macromolecules released in the simultaneouschelating process.
Taken altogether, EDTA has significant effect on the efficiency of the en-zymes i.e. pectinase and xylanase, since in both pectin and xylan polymersthere are divalent ion cross-bridges. Their removal modifies the polymerstructure and alters its accessibility for the enzymes.
We have also investigated the ‘EDTA-enzyme-substrate’ interaction withan additional substrate, namely all-linen fabric, for confirming the results
40
4. Results and Discussion
Figure 12. Reducing sugar liberation after enzymatic treatment of cottonseed-coat fragments. Effect of EDTA on the enzyme action.

noted above. The pectin-rich linen contains only 70 % of cellulose, and ahigh portion of noncellulosic matter. The influence of EDTA on the en-zyme performance, as well as on the degradation of the noncellulosic con-
41
4.3. Role of EDTA in bioscouring of cellulosics
Figure 13. Effect of EDTA on the residual calcium ion contentof biopretreated linen fabrics.
Figure 14. Effect of EDTA on reducing sugar liberationin biopreparation of linen fabrics.

stituents in biopreparation was investigated in detail. The main goal ofthe study was the comparison of the degradation efficiency of thenoncellulosic constituents of linen in biopreparation with or withoutEDTA.
Results prove that adding EDTA into the enzyme solution acceleratesthe degree of hydrolysis, and results in a higher reducing sugar liberationand more efficient calcium ion extraction, indicating a synergetic effect ofenzyme and EDTA in one treating bath. However, when EDTA is ap-plied as a pretreatment, a decrease in the efficiency of the subsequent en-zymatic hydrolysis is observed (Figure 13 and 14). These results confirmour earlier findings in the field of cotton biopreparation that EDTA modi-fies the substrate structure by removing the calcium ions from thecross-bridges of pectin.
42
4. Results and Discussion

5. CONCLUSIONS
Biotechnology offers a wide range of alternative environmentally-friendly processes for the textile industry to complement or improve theconventional technologies. Biopreparation of the cellulosic fibres is an en-zyme-aided process by which the noncellulosic ‘impurities’ (i.e. waxes,pectic substances, proteins, lignin-containing and colouring materials,etc.) are removed mainly by pectinase rich enzymes. Enzymes act undermild conditions (pH, temperature) with low water consumption; whilethe conventional alkaline scouring carried out with hot caustic soda is un-questionably an energy, water and chemical-intensive process. Thus,bioscouring provides an environmentally friendly alternative to theconventional alkaline process.
This study has been among the first attempts to study the effects of differ-ent enzymes with a chelator, and their synergetic action on bioprepa-ration of cotton fabric. The results obtained provide new information onthe effects of commercial enzymes on the properties of cotton and linenfabrics and degradation of cotton seed-coat fragments, as well as onbetter understanding of the mechanism of the chelator-aided bioprepa-ration.
The most important conclusions of this study can be briefly summarisedin the following main points:
1. Besides cellulases and pectinases, xylanase enzyme can also be used ef-fectively in biopreparation of cotton fabric. Biopretreatment results in hy-drophilic and homogeneously absorbent fabric with excellent colour even-ness.
2. Whiteness of the biopretreated fabrics is less than that of the conven-tionally scoured samples. However, a hydrogen peroxide bleaching ap-plied subsequent to the biopretreatment overcomes the colour differ-ences between conventionally scoured and bioscoured samples. Hydro-gen peroxide bleaching does not cause divergences and perceptiblecolour differences in the samples.
3. Biopretreated fabrics can be dyed with a reactive dye subsequent tothe enzyme treatment without further oxidative bleaching. At higher dyeconcentrations, there is no perceptible colour difference between thebiopretreated and alkaline scoured fabrics in dyed state. In pale and me-
43

dium dyeing, however, the colour difference is great and perceptible.Bleaching applied subsequent to bioscouring significantly decreases thecolour difference between the dyed samples pretreated in different ways.
4. Efficiency of the biopreparation process can be enhanced signifi-cantly by adding ethylenediaminetetraacetic acid (EDTA) chelator to theenzyme solution. EDTA accelerates the degradation of cotton seed-coatfragments, the most resistant impurities of cotton. Furthermore, EDTAimproves the lightening and weakens the darkening effects of the enzymetreatments carried out in acidic or neutral medium, respectively.
5. Results on ‘enzyme-EDTA-substrate’ interaction investigated inbiopreparation of cotton and linen prove that application of EDTA in dif-ferent concentrations does not inhibit the main activities of the hydrolyticenzymes (pectinase, xylanase), but does not increase them either. EDTAmodifies the substrate structure by removing the calcium ions from thecross-bridges, linked the macromolecules in pectin.
44
5. Conclusions

6. REFERENCES
Adamsen, A.P.S., Akin, D.E. and Rigsby, L.L. (2002) Chelating Agent and En-zyme Retting of Flax. Textile Res. J. 72(4), 296–302.
Akin, D.E., Henriksson, G., Morrison, W.H. and Eriksson, K.L. (1998) Enzy-matic Retting of Flax, Enzyme Application for Fibre Processing. In ACS Symp.Series 687 (eds. K-E.L. Eriksson and A. Cavaco-Paulo). Am. Chem. Soc., Wash-ington, D.C., pp. 269–278.
Akin, D.E., Foulk, J.A. and Dodd, R.B. (2002) Influence on Flax Fibres of Com-ponents in Enzyme Retting Formulations. Textile Res. J. 72(6), 510–514.
Annual Book of ASTM Standards (1980) Standard Test Methods for Seed-coatFragments and Funiculi in Cotton Fibre Samples. American Society for TestingMaterials, Easton, USA, pp. 2496–2580.
Bailey, M.J. and Pessa, E. (1990) Strain and Process for Production ofPolygalacturonase. Enzyme Microb. Technol. 12, 266–271.
Bailey, M.J. and Nevalainen, K.M.H. (1981) Induction, Isolation and Testing ofStable Trichoderma reesei Mutants. Enzyme Microb. Technol. 3, 153–157.
Bailey, M.J., Biely, P. and Poutanen, K.J. (1992) Interlaboratory Testing andMethods for Assay of Xylanase Activity. J. Biotechnol. 23, 257–270.
Buchert, J. and Pere, J. (2000) Scouring of Cotton with Pectinases, Proteases, andLipases. Textile Chemist and Colorist � American Dyestuff Reporter 32(5), 48–52.
Buchert, J., Pere, J., Puolakka, A. and Nousiainen, P. (1998) Enzymatic Scouringof Cotton. In Book of Papers, AATCC International Conference & Exhibition. Am.Assoc. Text. Chem. Color., Philadelphia, pp. 493–499.
Buschle-Diller, G. (2003) Substrates and their Structure. In Textile Processingwith Enzymes. (eds.: A. Cavaco-Paulo and G.M Gübitz). pp. 42–85.
Cavaco-Paulo, A. (1998) Enzyme Applications in Fiber Processing. In ACS Sym-posium Series 687 (eds. K-E.L. Eriksson and A. Cavaco-Paulo). Am. Chem. Soc.,Washington, D.C., pp. 180–189.
Cavaco-Paulo, A., Almeida, L., and Bishop, D. (1996) Effects of agitation andendoglucanase pretreatment on the hydrolysis of cotton fabrics by a totalcellulase. Textile Res. J. 66(5), 287–294.
Csiszár, E., Rusznák, I. and Reicher, J. (1987) Wirksamere Alkalishe Beuche derBaumwolle. Melliand Textilber. 6, 411– 415.
Csiszár, E., Szakács, G., and Rusznák, I. (1998) Combining Traditional CottonScouring with Cellulase Enzymatic Treatment. Textile Res. J. 68(3), 163–167.
Demarty, M., Morvan, C. and Thellier, M. (1984) Calcium and the Cell Wall.Plant, Cell and Environment 7, 441–448.
45

Dey, P.M. and Harborne, J.B. (1997) Plant Biochemistry. Academic Press, SanDiego, London, Boston, New York.
Durden, D.K., Etters, J.N., Sarkar, A.K., Henderson, L.A. and Hill, J.E. (2001)Advances in Commercial Biopreparation of Cotton with Alkaline Pectinase.AATCC Review 8, 28–31.
Etters, J.N. and Annis, P.A. (1998) Textile Enzyme Use: A Developing Technol-ogy. American Dyestuff Reporter 5, 18–23.
Etters, J.N. (1999a) Cotton Preparation with Alkaline Pectinase: An Environmen-tal Advance. Textile Chemist and Colorist � American Dyestuff Reporter 1(3), 33–36.
Etters, J.N., Condon, D., Husain, A. and Lange, N.K. (1999b) Dyeing Propertiesof Caustic Scoured Versus Alkaline Pectinase Prepared Fabric. Colourage (an-nual), 41–46.
Ghose, T.K. (1987) Measurement of Cellulase Activities. Pure and Appl. Chem.59, 257–268.
Hardin, I.R., Wilson, S.S., Lu, Y. and Lu, W. (2004) Biopreparation of Cotton:Progress and Challanges. 3rd Annual Workshop COST Action 847. Graz, Austria,14–16 June.
Hartzell, M.M. and Hsieh, Y-L. (1998a) Enzymatic Scouring to Improve CottonFabric Wettability. Textile Res. J. 68(4), 233–241.
Hartzell, M.M. and Hsieh, Y-L. (1998b) Pectin-Degrading Enzymes for Scour-ing Cotton. In ACS Symposium Series 687 (eds. K. Eriksson and A. Cavaco-Paulo). Washington, D.C., pp. 212–227.
Hartzell, M.M. and Durrant, S.K. (1998c) Improving the Efficiency of PectinaseScouring with Agitation to Improve Cotton Fabric Wettability. In Book of Papers,AATCC International Conference & Exhibition. Philadelphia, Am. Assoc. Text.Chem. Color., pp. 310–318.
Henriksson, G., Akin, D.E., Slomczinsky, D. and Eriksson, K-E.L. (1999) Produc-tion of Highly Efficient Enzymes for Flax Retting by Rhizomucor Pusillus. J.Biotech. 68, 115–123.
Henriksson, G., Akin, D.E., Rigsby, L.L., Patel, N. and Eriksson, K-E.L. (1997)Influence of Chelating Agents and Mechanical Pretreatment on Enzymatic Ret-ting of Flax. Textile Res. J. 67(11), 829–836.
Hickman, W.S. (1995) Preparation. In Cellulosics Dyeing (ed. J. Shore). Societyof Dyers and Colourists, The Alden Press, Oxford, pp. 81–151.
Jarvis, M.C. (1984) Structure and Properties of Pectin Gels in Plant Cell Walls.Plant, Cell and Environment 7, 153–164.
Kobayashi, T., Hatada, Y., Higaki, N., Lusterio, D.D., Ozawa T., Koike, K.,Kawai, S. and Ito, S. (1999) Enzymatic Properties and Deduced Amino Acid Se-
6. References
46

quence of a High-alkaline Pectate Lyase from an Alkaliphilic Bacillus Isolate.Biochim. Biophys. Acta. 1427, 145–154.
Kobayashi, T., Higaki, N., Suzumatsu, A., Sawada, K., Hagihara, H., Kawai, S. andIto, S. (2001) Purification and Properties of a High-molecular–weight, AlkalineExopolygalacturonase from a Strain of Bacillus. Enzyme Microb. Technol. 29, 70–75.
Koivula, A., Linder, M. and Teeri, T.T. (1998) Structure-function Relationshipsin Trichoderma Cellulolytic Enzymes. In Trichoderma and Gliocladium Vol. 2 En-zymes, Biological Control and Commercial Applications (eds. G.E. Harman andC.P. Kubicek). Taylor&Francis, pp. 3–23.
Kubicek, C.P. (1982) �-Glucosidase Excretion by Trichoderma pseudokoningiiCorrelation with Cell Wall Bound �-1,3-glucanase Activities. Arch. Microbiol.132, 349–354.
Kulkarni, N., Shendye, A. and Rao, M. (1999) Molecular and BiotechnologicalAspects of Xylanases. FEMS Microbiology Review 23, 411–456.
Lange, N.K. (1993) Trichoderma reesei Cellulases and Other Hydrolyses. In Pro-ceedings of the Second TRICEL Symposium (eds. P. Suominen andT. Reinikainen) Espoo, Finland Foundation for Biotechnical and Industrial Fer-mentation Research 8, pp. 263–272.
Lenting, H.B.M., Zwier, E. and Nierstrasz, V.A. (2002) Identifying Important Pa-rameters for a Continuous Bioscouring Process. Textile Res. J. 72(9), 825–831.
Lewin, M. and Pearce, E.M. (1998) Handbook of Fibre Chemistry. Marcel DekkerInc., New York.
Li, Y. and Hardin, I.R. (1998a) Treating Cotton with Cellulases and Pectinases:Effects on Cuticle and Fibre Properties. Textile Res. J. 68(9), 671–679.
Li, Y. and Hardin, I.R. (1998b) Enzymatic Scouring of Cotton - Surfactants, Agi-tation, and Selection of Enzymes. Textile Chemist and Colorist 30(9), 23–29.
Lin, C-H. and Hsieh, Y-L. (2001) Direct Scouring of Greige Cotton Fabrics withProteases. Textile Res. J. 71(5), 425–434.
Miller, G.L. (1959) Use of Dinitro-Salicylic as Reagent for the Determination ofReducing Sugars. Anal. Chem. 31, 426–428.
Nevell, T.P. (1995) Cellulose: Structure, Properties and Behaviour in the Dyeing Pro-cesses. In Cellulose Dyeing (ed. J. Shore). Society of Dyers and Colourists, pp. 1–26
Nierstrasz, V.A. and Warmoeskerken, M.M.C.G. (2003) Process Engineeringand Industrial Enzyme Applications. In Textile processing with enzymes(eds. A. Cavaco-Paulo and G.M. Gübitz). Woodhead Publishing Ltd., Cam-bridge, England, pp. 129–131.
Sakamoto, T., Hours, R.A. and Sakai, T. (1994) Purification, Characterisationand Production of Pectic Transeliminases with Protopectinase Activity from Ba-cillus subtilis. Biosci. Biotech. Biochem. 58(2), 353–358.
47
6. References

Sawada, K., Tokino, S., Ueda, M. and Wang, X.Y. (1998) Bioscouring of Cottonwith Pectinase Enzyme. JSDC 114, 333–336
Sawada, K. and Ueda, M. (2001) Enzyme Processing of Textiles in ReverseMicellar Solution. J. of Biothechnol. 89, 263–269.
Schejter, A. and Marcus, L. (1988) Isoenzymes of Pectinesterase andPoly-galacturonase from Botrytis cinerea Pers. In Biomass, Part B (Lignin, Pectinand Chitin), Methods in Enzymology, Vol. 161 (eds. W.A. Wood and S.T.Kellogg). Acadamic Press Inc., San Diego, California, pp. 366–372.
Sharma, H.S.S. (1986) Effects of the Application of Chemical Additives to Desic-cated Flax on Retting. Biotechnol. Lett. 8, 219–223.
Shukla, R., Sharma, U. and Kulkarni, S. (2000) Enzymes and Their Use in Tex-tile Processes. Colourage 2, 19–24.
Skoog, D.A., West, D.M. and Holler, J.F. (1988) Fundamentals of AnalyticalChemistry. Sauders College Publishing, International Edition, New York, USA.
Spurway, T.D., Morland, C., Coopers, A., Sumner, I., Hazlewood, G.P.,O’Donell, A.G., Pickersgill, R.W. and Gilbert, H.J. (1997) Calcium Protects aMesophilic Xylanase from Proteinase Inactivation and Thermal Unfolding.J. Biol. Chem. 272(28), 17523–17530.
Traore, M.K. and Buschle-Diller, G. (2000) Environmentally Friendly ScouringProcesses. Textile Chemist and Colorist � American Dyestuff Reporter 32(12),40–43.
Trotman, E.R. (1968) Textile Scouring and Bleaching. Charles Griffin & Com-pany Ltd., London.
Tzanov, T., Calafell, M., Guebitz, G.M. and Cavaco-Paulo, A. (2001) Bio-prepa-ration of Cotton Fabrics. Enzyme and Microbial Technology 29, 357–362.
Verschraege, L. (1989) Cotton Fibre Impurities. Neps, Motes and Seed-coatFragments. In ICAC Review Articles on Cotton Production Research No. 1. CABInternational, Wallingford, UK.
Viikari, L., Suurnäkki, A., Buchert, J. (1996) Enzyme-Aided Bleaching of KraftPulps: Fundamental Mechanisms and Practical Applications. In Enzymes forPulp and Paper Processing (eds. T. W. Jeffries and L. Viikari). American Chemi-cal Society, Washington, D.C., 15–24.
Ward, K. (1955) Chemistry and Chemical Technology of Cotton. Interscience Pub-lishers Inc., New York, London.
Yachmenev, V.G., Bertoniere, N.R. and Blanchard, E.J. (2001) Effect ofSonication on Cotton Preparation with Alkaline Pectinase. Textile Res. J. 71(6),527–533.
48
6. References

Appendix I.
49

Appendix I.
50

Enzymes and chelating agent in cotton pretreatment
51

Appendix I.
52

Enzymes and chelating agent in cotton pretreatment
53

Appendix I.
54

Enzymes and chelating agent in cotton pretreatment
55

56
Appendix I.

Enzymes and chelating agent in cotton pretreatment
57


Appendix II.
59

Appendix II.
60

Bleachability and Dyeing Properties of Biopretreated and Conventionally
61

Appendix II.
62

Bleachability and Dyeing Properties of Biopretreated and Conventionally
63

Appendix II.
64

Bleachability and Dyeing Properties of Biopretreated and Conventionally
65

Appendix II.
66

Role of EDTA Chelating Agentin Bioscouring of Cotton
A. LOSONCZI, E. CSISZÁR,1 G. SZAKÁCS AND L. BEZÚR
Budapest University of Technology and Economics,H-1521 Budapest, Hungary
ABSTRACT
Our previous work demonstrated that ethylenediamine-tetra-acetic acid(EDTA) markedly enhances the apparent activity of commercialxylanase and pectinase enzymes in cotton bioscouring, accelerating deg-radation and removal of impurities. In this study, we explore how theEDTA can improve the efficiency of the enzyme process in cottonbioscouring. Effects of EDTA on the enzyme action, as well as on the deg-radation of seed-coat fragments – the most resistant natural impuritiesof cotton to be degraded in bioscouring - are investigated. Results showthat application of EDTA in different concentrations does not inhibitthe main activities of the enzymes investigated, thus pectinase activity ofViscozyme 120L, and xylanase activity of Pulpzyme HC retain almost un-changed. On the other hand, EDTA modifies the substrate structure byremoving the calcium ions from the cross-bridges that link themacromolecules in pectin to one another or pectin to other polysaccha-rides. While EDTA pretreatment decreases significantly the efficiencyof enzymatic hydrolysis monitored by reducing sugar liberation, simulta-neous application of EDTA and enzyme in one treating bath acceleratesthe degree of hydrolysis very much indicating a synergistic effect of en-zyme and EDTA.
Recent results in textile biotechnologyproves that enzymes can be used effectivelyin the preparatory processes of natural cellu-losic fibers like cotton and flax. Inbioscouring of cotton the removal of the
noncellulosic ‘impurities” (i.e. waxes, pecticsubstances, proteins, lignin-containing andcoloring materials, etc) can be achievedmainly by hydrolytic enzymes, such aspectinases, xylanases and cellulases. Enzy-matic degradation of pectin accelerates theremoval of waxy materials from the cottonprimary wall, thus produces water wettablecotton. The bioprocess has several advan-
67
Appendix III.
1 To whom correspondence should be ad-dressed: e-mail [email protected]
Textile Res. J. (accepted)

tages over traditional chemical scouring. En-zymes operate under mild conditions (pH,temperature) with low water consumptionand act only on specific substrates. On theother hand, traditional alkaline scouringcarried out with hot caustic soda is unques-tionably an energy-, water- and chemical-in-tensive process [17].
Enzymes can also be used with good re-sults in the retting process of flax. Attemptswere made to eliminate the disadvantagesof the current commercial method of fieldretting and to improve the quality and con-sistency of the fibers. As a result of the re-search focused on the development of alter-native retting methods a specific en-zyme-retting has been worked out. In theprocess pectinase-rich enzyme mixtures areused to separate the fibers from non-fibertissues and to produce quality fibres fromflax. Enzyme-retting is more controllableand environmentally-friendly than the con-ventional dew-retting process [1, 20].
Both in bioscouring of cotton and en-zyme-retting of flax, degradation of pecticsubstances is the most essential process. Pec-tin generally builds up from D-galacturonicacid residues, which are present in a linearbackbone. Some of the carboxylic acidgroups are esterified with methanol. Thepectic polymer can be called a block copoly-mer with alternating methyl-esterified andnon-esterified blocks. The pectin is cova-lently linked to hemicellulose or to cellu-lose, or there are strong hydrogen-bonds be-tween pectin and other components in thecell wall. On the other hand, pectin inter-acts itself forming calcium cross bridges,which link the pectin components to one an-other. The crosslinks are formed betweenthe non-esterified or little methyl-esterifiedgalacturonan blocks with negative chargeand calcium ion with positive charge by elec-trostatic interactions. The calcium ion canbind pectin and other polysaccharides too.It generates coordination linkages withhydroxylic group of polysaccharides andionic linkages with the carboxylic group ofpectin [12, 10, 8]. An “egg-box” model was
proposed to show how calcium ionsstrengthen plant cell walls [11]. Conse-quently, removal of calcium ions can accel-erate the degradation of calcium-rich pecticsubstances.
Application of chelating substances todesiccated flax to enhance microbial-rettingwas first reported several years ago [19].Comprehensive research works on enzymeretting of flax have shown that among sev-eral chelators tested, ethylenediaminetetra-acetic acid (EDTA) is the most efficientchelator in improving enzyme retting offlax. Recently we have shown that additionof EDTA markedly enhances the apparentenzyme activity in cotton bioscouring [5].EDTA improves the effectiveness of com-mercial xylanase and pectinase enzymes, aswell as accelerates both the removal of theimpurities from cotton fabric and the degra-dation of seed-coat fragments separatedfrom the fabric. Application of EDTA inthe enzyme solution has also significant ef-fect on the efficiency of the subsequentchemical process.
In the present paper we explore how theEDTA can improve the efficiency of the en-zyme process in cotton bioscouring. The ef-fects of EDTA on the apparent activities ofcommercial pectinase and xylanase en-zymes, and on the degradation of non-cellu-losic constituents in bioscouring are investi-gated in detail. The main goals of this studywere the quantitative characterization of en-zyme activities in the presence of EDTAand comparison of the degradation effi-ciency of the non-cellulosic constituents ofcotton in bioscouring with or withoutEDTA. Influence of EDTA in different con-centrations on enzyme activities i.e. filter pa-per (cellulase), 1,4-�-endoglucanase,�-glucosidase, xylanase and pectinase activi-ties, is investigated in detail. Degradation ofseed-coat fragments – the most resistant im-purities of greige cotton - is characterized incotton bioscouring by weight loss and reduc-ing sugar liberation, as well as by changes incalcium ion content.
68
Appendix III TEXTILE RESEARCH JOURNAL (accepted)

Experimental
Viscozyme 120L (Aspergillus sp. origin),a pectinase-rich enzyme and Pulpzyme HC(Bacillus sp. origin), a pure xylanase werekindly supplied by Novozymes. The activi-ties were measured according to the meth-ods described in Refs. 9, 15, 3, 2, 22. Resultsare shown in Table I. The influence ofEDTA on the activity of the enzymes wastested by performing the standard activityassay in the presence of EDTA, at theEDTA-enzyme ratio of 1; 3; 5; 6; 8.6 (mMEDTA/ml enzyme). All activity values re-ported in this paper are mean values of trip-licate experiments. (Table I)
Spinning blowroom waste, a combinedwaste produced during the mechanicalsteps of cotton purification was selected tomodel the seed-coat fragment impurity. Itconsists of stalks leaves and seed-coat frag-ments of cotton [7]. Before use, the sub-strate was washed in distilled water at roomtemperature for 30 minutes to extract thewater-soluble constituents and air-dried.Prior to the treatments the sample was con-ditioned at 65% RH and 21�C for 24 hours.
Four sets of treatments have been ap-plied: (a) EDTA-only, (b) enzyme-only, (c)EDTA pretreatment and subsequent en-zyme treatment, without drying betweenthe two steps and (d) enzyme and EDTA ap-plied together in one bath. EDTA treat-ment of seed-coat fragment samples (a) wascarried out in phosphate buffer (pH 7) at50�C for 15 minutes in agitated system(Scourotester laboratory unit, 40 � 2 rpm).The liquor ratio was 1:100 and EDTA con-centration was 0.5 mM/g substrate. Enzyme
treatment (b) was performed in agitated sys-tem at pH 5 (0.05 M acetate buffer) forViscozyme 120 L, or at pH 7 (0.05 M phos-phate buffer) for Pulpzyme HC. The liquorratio was 1:100, enzyme concentrationswere 2 and 4 ml/L respectively, treatmenttime was 1 hour, incubation temperaturewas 50°C, and the concentration of thenonionic surfactant (Felosan TAK-NO,CHT R. Beitlich GmbH, Germany) was 1ml/L. After enzymatic treatment, the sub-strate was washed twice in hot distilled wa-ter for 10 seconds each to deactivate the en-zyme and then air-dried for 24 hours. In theexperiment (c) the EDTA pretreatment ofseed-coat fragment samples was carried outas described above (step a). Subsequent tofiltration the substrate was treated with en-zyme according to the method describedearlier (step b). When enzyme and EDTAwere used together (d) 0.5 mM EDTA/g sub-strate was added to the enzyme solution.The treatment was carried out similarly asdescribed above (step b).
Weight loss was determined by weigh-ing the samples before and after the treat-ment following 24 hours of conditioning at20�C and 65% RH. Release of reducing sug-ars during the enzymatic treatments was de-termined in the reaction liquor using the3,5-dinitrosalicylic acid reagent [16]. Ele-mental analysis of the enzymes and calciumion content of the substrate i.e. cottonseed-coat fragments, were determined by In-duced Coupled Plasma Optical EmissionSpectroscopy (ICP-OES) method as de-scribed earlier [5].
69
Role of EDTA Chelating Agent in Bioscouring of Cotton
Table I. Activities of commercial enzymes used throughout the experiments. a
Enzymes Cellulase(FPA),FPU/ml
1,4-�-endo-glucanase,
EGU/ml
�-glucosidase,
IU/ml
Pectinase
IU/ml
Xylanase,
IU/ml
Viscozyme 120L 2 (0.3) 8600 (80) 4 (0.2) 7900 (380) 110 (3)
Pulpzyme HC 0 50 (0.8) 0 0 13800 (550)
a Standard deviations in brackets.

Results and Discussion
INFLUENCE OF EDTA ON ENZYME ACTIVITY
Viscozyme 120L is a pectinase rich en-zyme with significant 1,4-�-endoglucanaseactivity. The enzyme complex shows negligi-ble xylanase, cellulase and �-glucosidase ac-tivities too. Pulpzyme HC is almost a purexylanase enzyme with practically zerocellulase (FPA), �-glucosidase andpectinase activities under standard assayconditions; a very small amount of1,4-�-endoglucanase activity can be de-tected (Table I). Influence of EDTA on theenzyme activities of the two enzymes was in-vestigated in more detail. EDTA with con-centrations of 0 - 8.6 mM/ml enzyme wasadded to the enzyme solution and the stan-dard activity assays were performed.
For Viscozyme 120L, the measured en-zyme activities as a function of EDTA con-centration are presented in Figures 1-5. Asshown in Figures 1 and 2, EDTA has no ef-fect on cellulase (FPA) and �-glucosidaseactivities. On the other hand,1,4-�-endoglucanase (Figure 3), xylanase(Figure 4) and pectinase (Figure 5) activi-ties are slightly inhibited by EDTA. With anEDTA concentration of 1 mM/ml enzyme,1,4-�-endoglucanase activity is reduced byapprox. 9% (from 8600 to 7800 EGU/ml,
Figure 3). By increasing the EDTA concen-tration from 1 to 8.6 mM/ml only a minorfurther negative effect on1,4-�-endoglucanase activity occurred. Forxylanase activity, by adding 1 and 3 mMEDTA to the enzyme solution the initial ac-tivity (107 IU/ml) has been reduced to 103and 93 IU/ml (Figure 4), respectively. Fur-ther increase in EDTA concentration from3 to 8.6 mM/ml does not result in any fur-ther xylanase activity loss. It can be statedfrom the data in Figure 5 that EDTA doesnot affect significantly the pectinase activ-ity, although a slight decrease can be mea-sured at the first two EDTA concentrations(i.e. 1 and 3 mM/ml). At 3 mM EDTA/ml en-zyme concentration the initial pectinase ac-tivity (7900 UI/ml) is reduced only by 8.8%.
For Pulpzyme HC, EDTA does not in-hibit the xylanase activity significantly (Fig-ure 6). Although the data vary near the ini-tial activity value with the EDTA concentra-tion, but no trend can be observed. EDTAcauses measurable decrease in4-�-endoglucanase activity (Figure 7). Itcan be summarized from the results thatEDTA does not inhibit significantly themain activities of the enzymes investigated,thus pectinase activity of Viscozyme 120Land xylanase activity of Pulpzyme HC re-tain almost unchanged in the presence ofEDTA.
70
Appendix III.
Table II. Analysis of Viscozyme 120 L and Pulpzyme HC enzymes by ICP-OES method.
Constituents Viscozyme 120L Pulpzyme HC
Concentration, ppm
Al 2.3 18.5
Ca 13.2 2290
Fe 11.6 7.6
K 2860 56
Mg 101 114
Na 39600 344
P 1160 1250
S 721 427

There are many so-called metallo-en-zymes that contain metal ion in their struc-ture. By removing the metal ion by chelat-ing, tertiary structure of the enzyme can bedestroyed. As a consequence, the activitymay decrease or disappear. It is also demon-strated that several bacterial pectate lyasesrequire calcium ion for enzyme activity andthey are inhibited by EDTA, but the inhibi-tion can be reduced by addition of an excessof calcium ion [18, 13]. EDTA can also in-hibit the activity of exopolygalacturonase by20-80% depending on the reaction condi-tions. However, the enzyme activity is not re-
71
Role of EDTA Chelating Agent in Bioscouring of Cotton
FIGURE 1. Effect of EDTA on filter paper activityof Viscozyme 120 L enzyme.
FIGURE 2. Effect of EDTA on �-glucosidase ac-tivity of Viscozyme 120 L enzyme.
FIGURE 3. Effect of EDTA on 1,4-�-endoglu-canase activity of Viscozyme 120 L enzyme.
FIGURE 4. Effect of EDTA on xylanase activity ofViscozyme 120 L enzyme.
FIGURE 5. Effect of EDTA on pectinase activityof Viscozyme 120 L enzyme.

stored at all by adding different divalentions, such as calcium [14].
Among xylanases only one was found tohave a calcium binding domain. However,the activity of the enzyme was not signifi-cantly influenced by either the disruption ofthe metal binding domain, or EDTA treat-ment. It was also suggested that endo-xylanase from Aspergillus versicolor requiresmetal ion for enzyme action, because in thepresence of EDTA the activity decreased al-most by 50%. [21, 4].
Analysis of the two enzymes used in thisresearch by ICP-OES method show that theycontain approximately equal amounts ofphosphorus, and magnesium (Table II). ForViscozyme 120L, the amount of sodium andpotassium ions, and for Pulpzyme HC, theamount of calcium ion is significant. Our re-sults prove that there is no significant activityloss in the presence of EDTA if Viscozyme120 L or Pulpzyme HC enzymes are used to-gether with EDTA. Thus, these metal ionsare not part of the enzyme structure, andEDTA does not change the tertiary struc-ture of the enzymes by chelating. The originof metal ions in the commercial enzymesmay be attributed to the medium ingredientsused in fermentation and to the additives dur-ing the downstream processing.
EFFECT OF EDTA ON COTTONBIOSCOURING (SEED-COAT FRAGMENTDEGRADATION)
Seed coat fragment is a portion of a cot-ton seed, usually black or dark brown incolor, broken from mature or immatureseed, and to which fibers and linters may beor may not be attached. We found in our ear-lier experiments [6, 7] that mechanically at-tached (vegetable origin) contaminants i.e.seed-coat fragments, pieces of stem and leaftrash, are the most resistant impurities ofcotton. They are more troublesome in pre-treatment than the other non-cellulosic con-stituents of the cotton fibers. Although, thedifferent mechanical cleaning steps removemajority of the seed-coat fragments, a cer-tain amount of these impurities may adhereto the fibers and may be present even in thefinal fabric. They differ in chemical composi-tion, color and morphology from cotton cel-lulose. In the traditional chemical processestheir removal requires more concentratedchemical solutions and longer steaming peri-ods. Their removal is the greatest obstacleto commercialization of biopreparation pro-cess. Whether either the traditional chemi-cal or the enzymatic process is used for prep-aration of cotton fabrics, special attention
72
Appendix III.
Table III. Effect of EDTA on cotton bioscouring. Degradation of cotton seed-coat fragments:weight loss and residual calcium ion content. a
Treatments Weight loss, % Calcium ion, ppm
- (untreated) - 34900
EDTA b 10.6 (0.7) 18500
Viscozyme 120L c 14.8 (0.5) 17700
EDTA + Viscozyme 120L d 22.5 (0.7) 13200
Viscozyme 120L with EDTA e 21.5 (0.6) 11900
Puplzyme HC c 14.1 (0.8) 31100
EDTA + Pulpzyme HCd 25.0 (0.6) 16000
Puplzyme HC with EDTA e 22.2 (0.6) 16800
a Standard deviations in brackets. b Treatment with EDTA. c Treatment with enzyme. d
Pretreatment with EDTA and subsequent enzyme treatment. e Enzyme treatment withEDTA.

has to be paid to the degradation ofseed-coat fragments.
Metal ion content of cotton seed-coatfragments is extremely high comparing tothat of the cotton fiber itself [5]. Accordingto ICP-OES determinations, for examplethe amount of calcium ions in loose cottonfiber is about 500 ppm, whereas the quan-tity of the same cation in the seed-coat frag-ments is as high as 35000 ppm (Table III).Pectin substances in plant usually form ag-gregates with calcium ions that crosslinkpectin chains to each other and to other
polymers of the lignin-holocellulose com-posite. Therefore, removal of calcium ionsfrom the substrate with chelators may accel-erate the enzyme action and promote thedegradation of seed-coat fragments inbiopreparation process.
Seed-coat fragment samples were sub-jected to a cold distilled water extractionfirst and then to different treatments, i.e.EDTA-only, enzyme-only, enzyme subse-quent to the EDTA pretreatment and co-ap-plication of enzyme and EDTA in one bath.The overall degradation of the solid materi-
73
Role of EDTA Chelating Agent in Bioscouring of Cotton
FIGURE 6. Effect of EDTA on xylanase activity ofPulpzyme HC enzyme.
FIGURE 7. Effect of EDTA on 1,4-�-endoglu-canase activity of Pulpzyme HC enzyme.
FIGURE 8. Treatment of cotton seed-coat fragments by pectinase (a) and xylanase (b). The enzymeshave been used directly to the substrate, or after EDTA pretreatment of the fragments, or together(in one bath) with EDTA.

als were characterized by the weight loss(Table III). EDTA treatment alone causes10.6% loss in weight, and the initial amountof calcium ions has been reduced by 47%owing to the chelating process. Bothpectinase and xylanase treatments result inhigher weight losses at 14.8 and 14.1%, re-spectively. Note that while the weight lossvalues for the two enzyme treatments are es-sentially the same, the pectinase treatmentin acidic buffer decreases the calcium con-tent enormously from 34900 to 17700 ppm,while the xylanase treatment at pH 7 causesonly a milder decrease (to 31100 ppm).
The two-step process, i.e. EDTA pre-treatment and a subsequent enzyme treat-ment proves to be the most effective in deg-radation of seed-coat fragment samples,yielding 22.5 and 25.0% weigh loss valuesfor Viscozyme 120L and Pulpzyme HC, re-spectively. Both the EDTA pretreatmentand the subsequent acidic pectinase treat-ment (EDTA + Viscozyme 120L) contrib-ute significantly to release the calcium fromthe substrate, leaving the residual calciumcontent of 13200 ppm. For EDTA +Pulpzyme HC two-step treatment, the ef-fect of EDTA is more remarkable than theaction of enzyme. For simultaneous treat-ment with enzyme and EDTA, the weightloss values are higher as compared to the sin-gle use of enzyme or EDTA, however thesefigures are less than those from the two-stepEDTA + enzyme treatment.
While weight loss data characterizedthe overall degradation effect of the appliedtreatments, reducing sugars in the treat-ment filtrates reveal the effectiveness of theenzyme action. As Figures 8a and b show,the EDTA pretreatment decreases signifi-cantly the hydrolysis level in the subsequentenzyme treatment, indicating changes insubstrate structure by extraction of the diva-lent ions. The amount of solubilized sugarsdecreases by 70 and 22% for pectinase andxylanase, respectively. Removal of the cal-cium cross bridges by EDTA pretreatmentcan cause a temporarily open structure,which can collapse by the end of the pre-
treatment, resulting in a less accessiblestructure for the enzyme macromoleculesapplied subsequently.
On the other hand, addition of EDTAinto the enzyme solution can accelerate thehydrolysis degree enormously (Figures 8aand b). Amount of reducing sugars in-creases by 72 and 38 % in the bath ofpectinase and xylanase enzymes, respec-tively, indicating synergistic effect of the en-zymes and EDTA applied together in onetreating bath. Removal of calcium ionsfrom calcium cross-bridges inhemicelluloses by EDTA results in some-what free and accessible areas in the sub-strate where the enzyme present is promptto act in catalysis of chain-cutting process.Thus, in this case the enzymes are capableof accessing the macromolecules releasedin the simultaneous chelating process.
Taken altogether, EDTA has signifi-cant effect on the efficiency of the enzymesi.e. pectinase and xylanase, since in both pec-tin and xylan polymers there are divalention cross-bridges. Their removal modifiesthe polymer structure and alters its accessi-bility for the enzymes.
Conclusions
The main goal of this study was to exam-ine how EDTA is effective in cottonbioscouring with commercial pectinase andxylanase enzymes. Influence of EDTA onthe enzyme action and enzymatic degrada-tion of the chosen substrate i.e. cottonseed-coat fragment was investigated in de-tail. Our findings show that EDTA added indifferent concentrations to the enzymes,does not have any inhibitory or stimulatingeffect on the main activities, i.e. pectinaseand xylanase for Viscozyme 120L andPulpzyme HC, respectively.
For enzymatic degradation of cottonseed-coat fragments, EDTA modifies thesubstrate structure by removing the calciumions from the cross-bridges that link themacromolecules in pectin to one another or
74
Appendix III.

pectin to other polysaccharides. WhileEDTA pretreatment decreases significantlythe efficiency of enzymatic hydrolysis moni-tored by reducing sugar liberation, simulta-neous application of EDTA and enzyme inone treating bath accelerates the degree ofhydrolysis very much. EDTA applied simul-taneously with the enzyme, seems to assistin the creation of free and accessible areasfor the enzyme present, indicating synergis-tic effect of the enzyme and EDTA. How-ever, EDTA pretreatment results in a lessaccessible substrate for the enzyme appliedsubsequently, most likely due to the col-lapse of the temporarily open structure
ACKNOWLEDGEMENTS
This work was supported by the Hungar-ian National Science Foundation (grantnos. OTKA T042927 and T032690). The au-thors wish to thank Prof. Emeritus IstvánRusznák and Johanna Reicher CSc. fortheir help and advice.
Literature Cited
1. Akin, D. E., Dodd, R. B., Perkins, W.,Henriksson, G., and Eriksson, K.-E. L.,Spray Enzymatic Retting: A NewMethod for Processing Flax Fibers, Tex-tile Res. J. 70(6), 486-494 (2000).
2. Bailey, M. J., Biely, P., and Poutanen, K.J., Interlaboratory Testing and Methodsfor Assay of Xylanase Activity, J.Biotechnol. 23, 257-270 (1992).
3. Bailey, M. J., and Nevalainen, K. M. H.,Induction, Isolation and Testing of Sta-ble Trichoderma reesei Mutants, EnzymeMicrob. Technol. 3, 153-157 (1981).
4. Carmona, E. C., Brochetto-Braga, M.R., Pizzirani-Kleiner, A. A., and Jorge,J. A., Purification and BiochemicalCharacterization of an Endoxylanasefrom Aspergillus versicolor, FEMSMicrobiol. Lett., 166, 311-315 (1998).
5. Csiszár, E., Losonczi, A., Szakács, G.,Rusznák, I., Bezúr, L., and Reicher, J.,Enzymes and Chelating Agent in Cot-ton Pretreatment, J. Biotechnol., 89,271-279 (2001).
7. Csiszár, E., Rusznák, I., and Reicher, J.,Wirksamere alkalishe Beuche derBaumwolle, Melliand Textilber., 6, 411-415 (1987).
8. Csiszár, E., Szakács, G., and Rusznák, I.,Combining Traditional Cotton Scour-ing with Cellulase Enzymatic Treat-ment, Textile Res. J. 68(3), 163-167(1998).
9. Demarty, M., Morvan, C., and Thellier,M., Calcium and the Cell Wall, Plant,Cell and Environment, 7, 441-448(1984).
10.Ghose, T. K., Measurement of CellulaseActivities, Pure and Appl. Chem. 59,257-268 (1987).
11.Grant, J. S., Carbohydrate Metabolism:Structural Carbohydrates, in “ Plant Bio-chemistry”, Academic Press, New York,1997. pp. 216-222.
12.Grant, G. T., Morris, E. R., Rees, D. A.,Smith P., and Thomas, D., Biological In-teraction between Polysaccharides andDivalent Cations: The Egg Box Model,FEBS Microbiol. Lett., 32, 195-198(1973).
13.Jarvis, M. C., Structure and Properties ofPectin Gels in Plant Cell Walls, Plant,Cell and Environment, 7, 153-164(1984).
14.Kobayashi, T., Hatada, Y., Higaki, N.,Lusterio, D. D., Ozawa T., Koike, K.,Kawai, S., and Ito, S., Enzymatic Proper-ties and Deduced Amino Acid Se-quence of a High-alkaline PectateLyase from an Alkaliphilic Bacillus Iso-late, Biochim. Biophys. Acta, 1427,145-154 (1999)
15.Kobayashi, T., Higaki, N., Suzumatsu,A., Sawada, K., Hagihara, H., Kawai, S.,and Ito, S., Purification and Propertiesof a High-molecular–weight, AlkalineExopolygalacturonase from a Strain ofBacillus, Enzyme Microb. Technol., 29,70-75 (2001).
16.Kubicek, C. P., �-Glucosidase Excretionby Trichoderma pseudokoningii Correla-tion with Cell Wall Bound�-1,3-glucanase Activities, Arch.Microbiol. 132, 349-354 (1982).
17.Miller, G. L., Use of Dinitro-Salicylic asReagent for the Determination of Re-ducing Sugars, Anal. Chem., 31, 426-428(1959)
75
Role of EDTA Chelating Agent in Bioscouring of Cotton

18.Nierstrasz, V. A., and Warmoeskerken,M. M. C. G., Process Engineering andIndustrial Enzyme Applications, in
“Textile processing with enzymes”,Woodhead Publishing Ltd., Cambridge,England, 2003, pp. 129-130.
19.Sakamoto, T., Hours, R. A., and Sakai,T., Purification, Characterization andProduction of Pectic Transeliminaseswith Protopectinase Activity from Bacil-lus subtilis, Biosci. Biotech. Biochem.,58(2), 353-358 (1994).
20.Sharma, H. S. S., Effects of the Applica-tion of Chemical Additives to Desic-cated Flax on Retting, Biotechnol. Lett.,8, 219-223 (1986).
21.Sharma, H. S. S., and Van Sumere, C. F.,Enzyme Treatment of Flax, Genet, Eng.Biotechnol. 12, 19-23 (1992).
22.Spurway, T. D., Morland, C., Coopers,A., Sumner, I., Hazlewood, G. P.,O’Donell, A. G., Pickersgill, R. W., andGilbert, H. J., Calcium Protects aMesophilic Xylanase from ProteinaseInactivation and Thermal Unfolding, J.Biol. Chem., 272(28), 17523-17530(1997).
23.Wood, W. A. and Kellogg, S. T., Biomass(Part B Lignin, Pectin, and Chitin),Methods in Enzymology, Vol. 161,366-373, Academic Press, London,1988.
76
Appendix III.

Appendix IV .
77
Biocatalysis and Biotransformation (accepted)

Appendix IV .
78

Influence of EDTA Complexing Agent on Biopreparation of Linen Fabric
79

Appendix IV .
80

Influence of EDTA Complexing Agent on Biopreparation of Linen Fabric
81

Appendix IV .
82




















