
Benthic response to shellfish farming in Thau lagoon: Pore water signature
14
Benthic response to shellfish farming in Thau lagoon: Pore water signature E. Metzger * , C. Simonucci, E. Viollier, G. Sarazin, F. Pre ´vot, D. Je ´ze ´quel Laboratoire de Ge ´ochimie des Eaux, Universite ´ Denis Diderot-Paris 7 & IPGP, Case postale 7052, 75251 Paris Cedex 05, France Received 18 November 2006; accepted 23 November 2006 Available online 18 January 2007 Abstract Vertical distributions of dissolved species across the sedimentewater interface (SWI), including major cations (sodium, potassium, magne- sium, calcium), minor cations (lithium, strontium, barium), redox sensitive species (dissolved manganese, iron, sulfate, sulfide, ammonium) and other chemical parameters (pH, alkalinity, soluble reactive phosphorous, dissolved silica) were studied in a Mediterranean lagoon used for in- tensive shellfish farming. In order to quantify the impact of this activity on diagenetic processes and the influence of seasonal changes, two stations contrasted with respect to organic carbon fluxes were sampled in Thau lagoon from March 2001 to August 2002 during four field cam- paigns in winter, spring, summer and fall. Well-defined layers enriched with redox sensitive species were observed following the conventional sequence of early diagenetic reactions. However, differences were observed between both stations in depths and thickness layers. Concentration gradients extended down to more than 92 cm depth at the central position of the lagoon (station C4 e 8 m depth) and down to 40 cm depth inside shellfish farming zones (station C5 e 9 m depth). Station C4 showed an unusual diagenetic signature: sharp dissolved oxygen, iron, nitrate and manganese gradients existed at the SWI but gradients of dissolved sulfide and alkalinity as well as other parameters (dissolved silica, Ba, etc.) were recorded only from 25 to 30 cm depth downward. Seasonal changes were observed in pore water composition as deep as 30e50 cm in station C4 (only 15 cm in station C5). The center of the lagoon is not directly subjected to biodeposits deriving from shellfish activity. Isotopic and bioturbation data allowed to rule out a reworking of the sediment deeper than a few centimeters. In addition to organic content of the sediment, physical parameters were likely to induce the 10e20 cm gap between dissolved iron and sulfide profile as well as the higher vertical extent of diagenetic sequence observed at station C4. Conversely to station C5, station C4 underwent stronger currents and wave effect probably generating advective transport of water through the sediment, but no permeability data were available to confirm this hypothesis. During summer, climatic conditions generated vertical stratification of the water column and transient suboxic conditions at the bottom. Such conditions drove the upward shift of redox fronts, compacting the diagenetic sequence. These effects were reinforced at station C5 by shellfish and its farm structures (mainly attenuation of current and increased heat absorption). Ó 2006 Published by Elsevier Ltd. Keywords: early diagenesis; redox; sulfide; pore water; advection; shellfish farming; Thau lagoon 1. Introduction Trophic equilibrium in coastal lagoons is very fragile. Such semi-confined aquatic systems tend to become eutrophic due to nutrients and contaminant input related to human activities (i.e., bank urbanization, industrial emission, poor waste-water treatment and intensive agriculture in the watershed) and low water renewal. Eutrophication leads to biological productivity increase in surface waters and modifies the response of benthic organisms to settling organic matter (Aller et al., 1998; Lopez, 2003). De- pending on the renewal rate of dissolved O 2 in the overlying waters and the settling rate of organic rich particles which can be metabolized, the oxic/anoxic boundary can shift down- ward (low organic matter sedimentation) or upward (high organic matter sedimentation). In coastal environments, where * Corresponding author. E-mail address: [email protected] (E. Metzger). 0272-7714/$ - see front matter Ó 2006 Published by Elsevier Ltd. doi:10.1016/j.ecss.2006.11.011 Estuarine, Coastal and Shelf Science 72 (2007) 406e419 www.elsevier.com/locate/ecss
-
Upload
cnrs-bellevue -
Category
Documents
-
view
1 -
download
0
Transcript of Benthic response to shellfish farming in Thau lagoon: Pore water signature

Estuarine, Coastal and Shelf Science 72 (2007) 406e419www.elsevier.com/locate/ecss
Benthic response to shellfish farming in Thau lagoon: Porewater signature
E. Metzger*, C. Simonucci, E. Viollier, G. Sarazin, F. Prevot, D. Jezequel
Laboratoire de Geochimie des Eaux, Universite Denis Diderot-Paris 7 & IPGP, Case postale 7052, 75251 Paris Cedex 05, France
Received 18 November 2006; accepted 23 November 2006
Available online 18 January 2007
Abstract
Vertical distributions of dissolved species across the sedimentewater interface (SWI), including major cations (sodium, potassium, magne-sium, calcium), minor cations (lithium, strontium, barium), redox sensitive species (dissolved manganese, iron, sulfate, sulfide, ammonium) andother chemical parameters (pH, alkalinity, soluble reactive phosphorous, dissolved silica) were studied in a Mediterranean lagoon used for in-tensive shellfish farming. In order to quantify the impact of this activity on diagenetic processes and the influence of seasonal changes, twostations contrasted with respect to organic carbon fluxes were sampled in Thau lagoon from March 2001 to August 2002 during four field cam-paigns in winter, spring, summer and fall. Well-defined layers enriched with redox sensitive species were observed following the conventionalsequence of early diagenetic reactions. However, differences were observed between both stations in depths and thickness layers. Concentrationgradients extended down to more than 92 cm depth at the central position of the lagoon (station C4 e 8 m depth) and down to 40 cm depth insideshellfish farming zones (station C5 e 9 m depth). Station C4 showed an unusual diagenetic signature: sharp dissolved oxygen, iron, nitrate andmanganese gradients existed at the SWI but gradients of dissolved sulfide and alkalinity as well as other parameters (dissolved silica, Ba, etc.)were recorded only from 25 to 30 cm depth downward. Seasonal changes were observed in pore water composition as deep as 30e50 cm instation C4 (only 15 cm in station C5). The center of the lagoon is not directly subjected to biodeposits deriving from shellfish activity. Isotopicand bioturbation data allowed to rule out a reworking of the sediment deeper than a few centimeters. In addition to organic content of thesediment, physical parameters were likely to induce the 10e20 cm gap between dissolved iron and sulfide profile as well as the higher verticalextent of diagenetic sequence observed at station C4. Conversely to station C5, station C4 underwent stronger currents and wave effect probablygenerating advective transport of water through the sediment, but no permeability data were available to confirm this hypothesis. Duringsummer, climatic conditions generated vertical stratification of the water column and transient suboxic conditions at the bottom. Such conditionsdrove the upward shift of redox fronts, compacting the diagenetic sequence. These effects were reinforced at station C5 by shellfish and its farmstructures (mainly attenuation of current and increased heat absorption).� 2006 Published by Elsevier Ltd.
Keywords: early diagenesis; redox; sulfide; pore water; advection; shellfish farming; Thau lagoon
1. Introduction
Trophic equilibrium in coastal lagoons is very fragile. Suchsemi-confined aquatic systems tend to become eutrophic dueto nutrients and contaminant input related to human activities(i.e., bank urbanization, industrial emission, poor waste-water
* Corresponding author.
E-mail address: [email protected] (E. Metzger).
0272-7714/$ - see front matter � 2006 Published by Elsevier Ltd.
doi:10.1016/j.ecss.2006.11.011
treatment and intensive agriculture in the watershed) and lowwater renewal.
Eutrophication leads to biological productivity increase insurface waters and modifies the response of benthic organismsto settling organic matter (Aller et al., 1998; Lopez, 2003). De-pending on the renewal rate of dissolved O2 in the overlyingwaters and the settling rate of organic rich particles whichcan be metabolized, the oxic/anoxic boundary can shift down-ward (low organic matter sedimentation) or upward (highorganic matter sedimentation). In coastal environments, where

407E. Metzger et al. / Estuarine, Coastal and Shelf Science 72 (2007) 406e419
particulate organic carbon fluxes are high, the oxygen penetra-tion depth generally settles a few millimeters below the sedi-mentewater interface (SWI) or can reach the water column,if oxygen availability is more.
In Thau lagoon, in addition to nutrient input from the wa-tershed, permanent shellfish production (mussels and oysterswhich represent 15 000 t y�1 or 10% of the French production;oyster mean density: 40 individuals m�2) significantly impactsecosystem functioning (Souchu et al., 1998b). This activityleads to a local overload of organic matter due to high fecesfluxes, especially during summer when the growth rate ofshellfish is at its maximum (Mazouni et al., 1998; Gangneryet al., 2003). Shellfish farming generates high seasonal varia-tions of oxygen demand in the lagoon. Due to a slow but irre-versible (up to now) increase in eutrophication, the watercolumn becomes partially or totally anoxic, causing sulfidesto diffuse from the sediment and inducing high shellfishmortality, which has important consequences on the localeconomy (Deslous-Paoli et al., 1998). Summer anoxia crises,locally called ‘‘mala€ıgues’’, usually start along the north coastof the lagoon (harbor places) or in bottom waters in the shell-fish farming area (Souchu et al., 1998a).
The aim of this paper is to characterize the response of sed-iment pore waters to intensive shellfish breeding by comparingpore water compositions within and out of the farming zone,and their seasonal evolution. Intensive shellfish breedinggenerates high seasonal feces fluxes which contribute to theoccurrence of water column anoxia. When oxygen vanishesfrom pore waters, anaerobic respiration of nitrate and metaloxyhydroxides takes over. This biogeochemical process leadsto the dissolution of manganese oxides (MnOx) and ironoxyhydroxides (e.g., FeOOH). Contaminants adsorbed ontooxides are subsequently released into pore waters and may dif-fuse into the water column. Data presented here provided thegeneral support for determining redox conditions that prevailin sediments and for the analysis of key parameters of mobilityof trace metals. Different redox sensitive compounds such asdissolved iron, manganese, sulfate and hydrogen sulfidewere analyzed, together with other metabolites of microbialrespiration such as ammonia and soluble reactive phosphorous(SRP), biogenic compounds such as dissolved silica, and con-servative tracers such as sodium, lithium and potassium. Cal-cium, strontium, magnesium and barium were studied sincethey constitute reactants or products of dissolution/precipita-tion processes. Finally, alkalinity and pH were determined asintegrative chemical parameters of diagenetic reactions.
2. Material and methods
2.1. Study area and sampling stations
Thau lagoon is located on the French Mediterranean coastin the vicinity of the city of Sete (Fig. 1). It is a shallow lagoonwith a mean depth of 4 m (maximum 10 m and locally 32 m,La Vise) and covers an area of 70 km2 (Chapelle et al., 2000).The lagoon is a semi-confined ecosystem, with two seawaterinlets, and several riverine inputs. Only one of them is
permanent (La Vene River). Thermal and karstic groundwater(e.g., the submarine resurgence La Vise) also contributes tofreshwater input (Aquilina et al., 2002). The catchment areais about 300 km2 and the average water residence time is threemonths (Millet, 1989). Tidal range is mostly less than halfa meter. Currents are principally driven by dominant windsfrom north and northwest regularly blowing at 10 m s�1 andrising several times per year to more than 30 m s�1. Windspeeds of 10 m s�1 generate current velocities close to10 cm s�1 in the entire water column (Lazure, 1992). Shellfishfarming areas cover about one-fifth of the lagoon area (Fig. 1).Density of shellfish tables strongly limits water circulation (bya factor of 3) and is clearly one of the triggering factors ofanoxia events (Chapelle et al., 2000). Shellfish activity gener-ates important biodeposits (about 500 kg ha�1 yr�1, DeCasabianca et al., 1997).
Two biogeochemically contrasted sites were chosen for thisprocess study. The contrast is due to the distance of the sam-pling sites from the shellfish farming ‘‘racks’’, which createcontrasted conditions of biodeposition and benthic fluxes(Mesnage, 1994; Plante-Cuny et al., 1998). One station (C5)is located close to a mussel farming table (Mytilus galloprovin-cialis) (N 43�25.9940, E 3�39.6570) and is under the influenceof biodeposits. The working area of station C5 is a circle ofabout 10 m in diameter and the water column height is about9 m. Station C4 is considered as a reference station (without‘‘racks’’) and is located at the center of the lagoon (N43�24.0180, E 3�36.7030), at least 2 km away from the shellfishfarms. The working area of station C4 is a circle of about100 m in diameter with a depth of about 8 m. In both stations,
43°20’N
SHELLFISH FARMING ZONES
0 2 Km
3°40’E
C4
C5
SETE
MEDITERRANEANSEA
THAU Lagoon
PALL
AS R
IVER VÈ
NE
RIV
ER
1
Fig. 1. Thau lagoon map with location of sites C4 and C5 (from Gilbert et al.,
1997).

408 E. Metzger et al. / Estuarine, Coastal and Shelf Science 72 (2007) 406e419
sediment topography is relatively flat, with an absence ofvisible channels or ripple marks. In the top 45 cm, sedimentporosity remains roughly homogeneous in station C4 (0.85)while at station C5, porosity decreases from 0.95 to 0.7 at�10 cm depth (Dedieu et al., 2007). Mean grain size doesnot show large variation: at station C4, it ranges from 10 to20 mm, while at station C5 it varies from 19 mm, at the SWI,to 12 mm at 15 cm depth, and remains fairly constant below.The sediment of station C4 consists of 55% fine silts and clays,35% silts and 10% sand from all over the profile. At stationC5, near the SWI, the sediment consists of 40% fine siltsand clays, 45% silts and 15% sand. Here, the percentage offine silts and clays increases with depth while that of sand de-creases (Schmidt et al., 2007). We observed the presence ofworms (mainly polychaetes) and small crabs as well as well-preserved shells in the sediment cores. Marine plants werenot present at the sampled stations (although they could beabundant in some other areas of Thau lagoon).
Field campaigns were conducted between March 2001 andAugust 2002 to investigate station differences and seasonalvariations. Station C4 was studied in MarcheApril 2001, De-cember 2001eJanuary 2002, and JulyeAugust 2002, and sta-tion C5 during December 2001eJanuary 2002, AprileMay2002, and JulyeAugust 2002.
2.2. Water column measurements
WTW sensors (temperature, pH and dissolved oxygen) anda multiparameter YSI sensor (depth, salinity, temperature, pHand dissolved oxygen) were used to obtain vertical profiles andchronological records. The pH was calibrated against NISTstandards and oxygen against water-vapor saturated air withsalinity, temperature, and pressure compensation. Salinityprobe was calibrated using KCl 0.1 mol L�1.
2.3. Interstitial water sampling
Pore water samples were collected with dialysis devices(the so-called ‘‘peepers’’) (Hesslein, 1976; Carignan et al.,1985). Two types were utilized with different pore volumesand vertical resolutions (respectively, 13 mL/2.5 cm and3 mL/1 cm close to the SWI, then 20 mL/2.2 cm). Peeperswere acid-cleaned with HCl Normapur� 2 mol L�1, thenrinsed three times with deionized water (Milli-Q� Millipore,18 MU cm). The dialysis cells were filled with deionizedwater and covered with a sheet of nylon filtration membrane(Biodyne� A, Pall; 0.2 mm pore size), then covered witha Plexiglas� lid with windows corresponding to each cell.Peepers were stored in Milli-Q water and degassed withdinitrogen for 12 h before their introduction into thesediments.
The peepers were deployed by scuba divers and retrieved 3weeks later. The time requirement for complete equilibrationwas largely fulfilled (Webster et al., 1998). Replicate peeperswere deployed at each station during each campaign in orderto check for lateral heterogeneity. Distance between peepersnever exceeded 20 m. Recovered peepers were kept under
nitrogen atmosphere during transport and sampling. Plastictips of micropipettes were adapted to Luer� fitting polypropyl-ene syringes (20 mL) to facilitate the piercing of the nylonmembrane and to avoid metal contamination during pore watercollection. Subsamples were transferred in pre-acidifiedFalcon� tubes (a drop of HNO3 suprapur�) to be preservedfor further total concentration analysis. Additionally, tubescontaining appropriate reagent (see below) for field analysisreceived aliquots of each sample. Peeper sampling, distribu-tion, and preservation of subsamples for each analysis andpH measurements were performed within 1 h after peeperretrieval.
2.4. Field analyses
A 1-mL subsample was used to measure pH with a glasselectrode and a double junction reference electrode to avoidelectrolyte poisoning from sulfide species. Spectrophotomet-ric (l¼ 590 nm) determination of alkalinity was carried outusing 500 mL of pore water dispensed into tubes containingformic acid/bromophenol blue reagent (Podda and Michard,1994; Sarazin et al., 1999). The sample (200 mL) was alsodistributed in pre-acidified tubes (HCl Normapur, 10�2 molL�1) for sulfate analysis. BaCl2 plus gelatin reagent wasadded to precipitate barium sulfate and maintain bariteprecipitate in suspension. Turbidity was recorded at 470 nm(Tabatabai, 1974). Sample fraction for
PH2S analysis
(200 mL) was immediately transferred to a tube containing3 mL of a ‘‘trap’’ solution made of zinc chloride(1.5� 10�2 mol L�1) and gelatin (3 g L�1) to precipitateZnS (preventing sulfide oxidation and gas loss). When neces-sary, samples were diluted with NaCl stock solution, thecomposition of which matches surface seawater salinity. Totalhydrogen sulfide concentration was measured by methyleneblue method (Merck Spectroquant� 14779; l¼ 660 nm).Standards were prepared with Na2S (w10 mmol L�1) stocksolution made daily. NH4
þ and SRP (soluble reactive phospho-rous) analyses were carried out using Merck Spectroquant�
14752 and 14848 methods after acidification with hydro-chloric acid, degassing H2S with dinitrogen, and neutraliza-tion with sodium hydroxide.
2.5. Laboratory analyses
Concentrations of Fe, Mn, Si, Ba, Sr, Na, Ca, Mg, and Liwere determined by ICP-AES (Optima 3000 Perkin Elmer).Calibration standards were prepared with commerciallyavailable solutions (SPEX� plasma standards). Samples werediluted 100 times. Dilutions were made with acidifiedMilli-Q� deionized water (HNO3 sp., 10�2 mol L�1; Merck).Accuracy of ICP-AES measurements was checked with differ-ent international standards (SLRS-4 for metals and CASS-4for minor and major components). Precision was within1e6% when signal was one order of magnitude higher thanblank one.

409E. Metzger et al. / Estuarine, Coastal and Shelf Science 72 (2007) 406e419
3. Results
3.1. Bottom waters
Surface water salinity and temperature measured nearstation C5 (N 43�26.170, E 3�39.940) by IFREMER duringthe program period are reported in Fig. 2. Contrasted resultswere observed for field campaign periods according to the sea-son. In winter 2001, temperature was below 5 �C, and reached23 �C in summer. Salinity ranged from 33 in spring 2001 to 39in summer 2002. Data from Dedieu et al. (2007), who used insitu microprofilers, showed that in December 2001 and April2002, bottom water was saturated in dissolved oxygen forboth sites. However, oxygen flux was the same for both sta-tions in December 2001, while in April 2002 flux was higherand more heterogeneous at station C5, indicating a higheroxygen demand in the sediment in this site. In August 2002,the authors showed that oxygen demand increased in both sitesand observed that saturation index of dissolved oxygenchanged in bottom waters in agreement with our results. Theseshowed that at the beginning of the campaign (Fig. 3) oxygenconcentration in bottom water corresponded to only 30% ofsaturation and rapidly rose to 80% within 3 days. Duringthis period pHNBS ranged from 7.8 to 8.3.
3.2. Pore waters
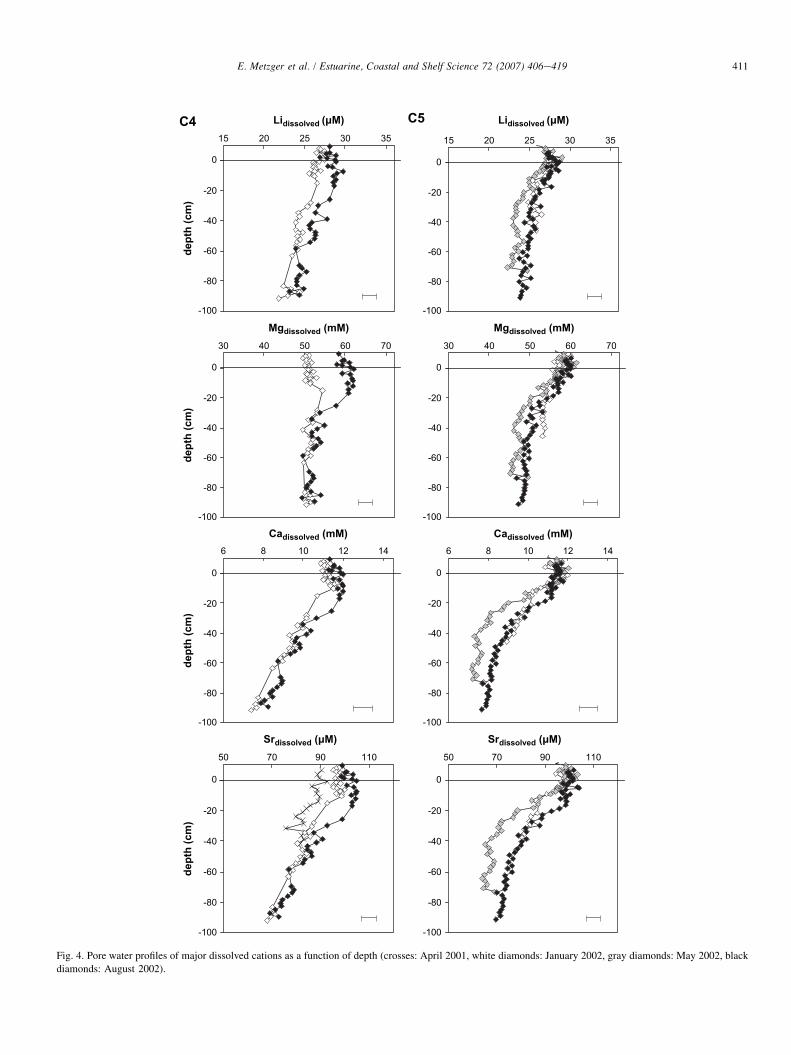
3.2.1. Alkaline and alkali earth elementsConcentration profiles for dissolved Li, Mg, Ca and Sr are
shown in Fig. 4. Na and K have similar patterns than Li andare not shown. Barium is described in Section 3.2.4 becauseof its special ‘‘nutrient-like’’ behavior compared to other alkaliearth elements. For both stations, the same general patternswere observed: Li and Mg concentrations were higher in theoverlying water and within the first 10e20 cm of pore watersthan within the underneath profile. These cations did not showsignificant differences at depth between station C4 and stationC5. Their concentrations corresponded to seawater with meansalinity of 35. Ca and Sr concentrations decreased strongly;their concentrations were highly correlated (R2> 0.99).However, sharper gradients occurred at station C5 with
04/0
1
12/0
1
04/0
2 08/0
2
25
27
29
31
33
35
37
39
41
11/00 05/01 11/01 05/02 11/02 05/03
Salin
ity
051015202530354045
50
Tem
p (°C
)
Fig. 2. Temperature (dotted line) and salinity (black line) of surface waters as
a function of time measured by IFREMER near Bouzigues (www.ifremer.fr).
quasi-asymptotic values reached at about 30 cm depth. Onthe other hand, a continuous decrease was observed at stationC4. It should be noticed that at station C4 cation concentra-tions were roughly the same, whatever the season, from about50 cm depth to the bottom. Above this limit, concentrationsvaried according to the sampling period, August 2002 showingthe highest values, and April 2001 the lowest (Sr profiles, theonly ones for which three periods are available, show charac-teristic trends).
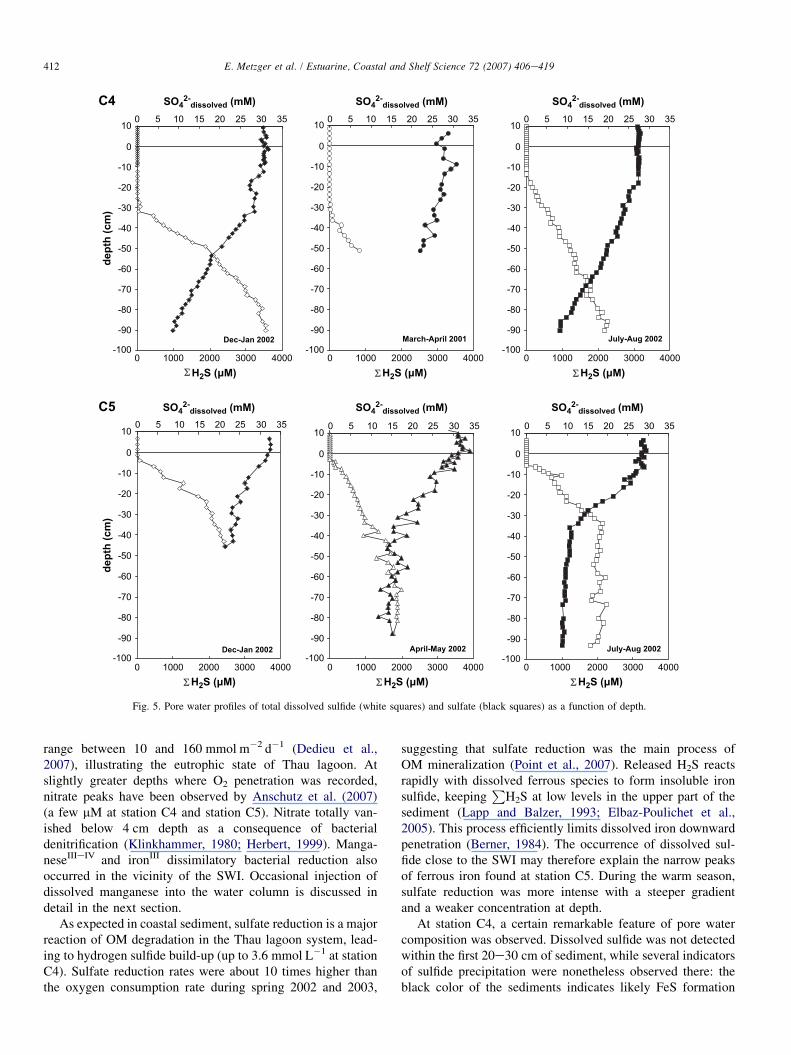
3.2.2. Sulfur speciesSulfate and hydrogen sulfide (
PH2S) profiles are shown in
Fig. 5. For both stations, sulfate concentration decreased inpore water at roughly the same depth at which an increaseof sulfide concentration was observed. The main differencebetween station C4 and station C5 was the depth at whichsulfide began to appear. This occurred between 18 and35 cm at station C4, depending on the season, and at only5 cm at station C5. Another important difference concernedSO4
2� in depth concentration, which reached quasi-asymptoticvalues at station C5, and decreased steadily, to 8 mmol L�1
(90 cm depth) at station C4.Seasonal variations of the appearance depth of sulfides
were observed at station C4.P
H2S appeared at about 35 cmin April 2001, at 32 cm in January 2002 and at 18 cm inAugust 2002. Like cations, sulfate concentrations from50 cm and below did not vary with time. However, betweenJanuary and August 2002, one observed different chemicalgradients at the top of the core. In August 2002, sulfate anddissolved sulfide gradients were almost constant from 18 cmto the bottom, whereas in January 2002, a slight slope changeoccurred at 50 cm depth. If one extrapolates a straight linefrom deep gradients, the line intercepts the depth axis closeto 18 cm (i.e., the characteristic depth observed in August).
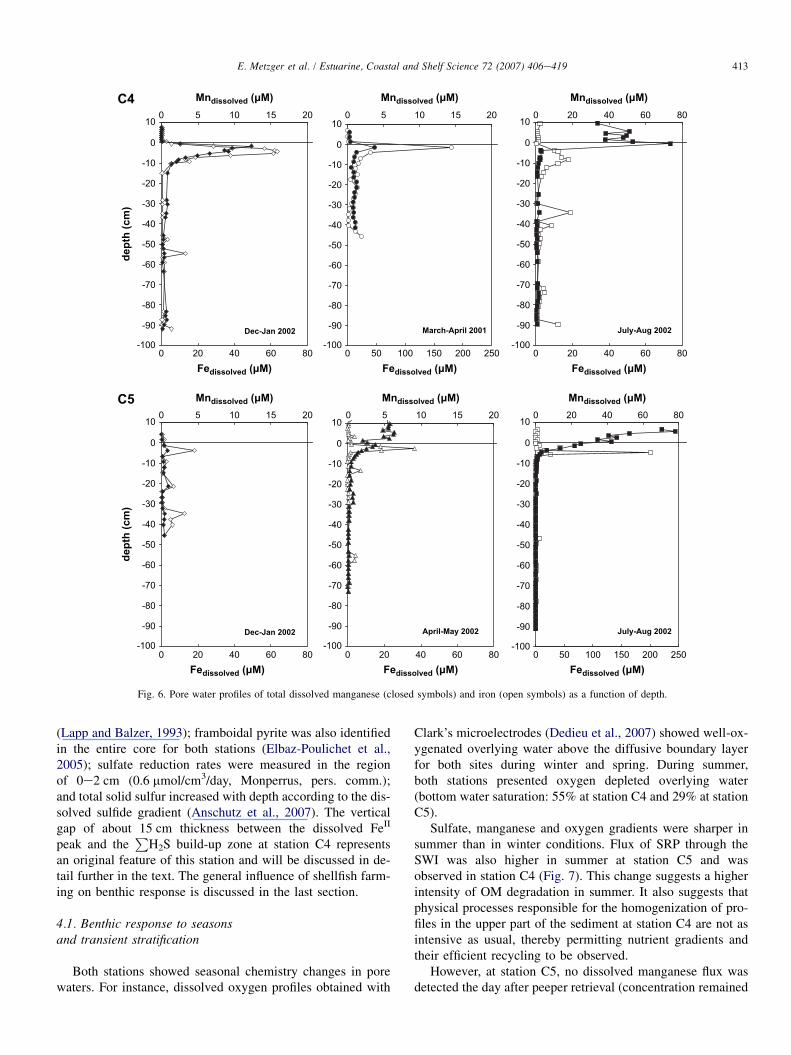
3.2.3. Mn, FeProfiles of total dissolved manganese and iron are shown in
Fig. 6. At station C4, Mn profiles displayed seasonal variablemaxima a few centimeters below the SWI. Maximum concen-trations ranged from 4 mmol L�1 in April 2001 to 80 mmol L�1
(at the SWI) in August 2002. In winter and spring seasons, Mnconcentrations in the overlying water were close to the detec-tion limit (less than 1 mmol L�1). In contrast, during summer,Mn concentrations in the overlying water ranged between 30and 55 mmol L�1, while the maximum (80 mmol L�1) was at-tained at the SWI. At depth, Mn concentrations were closeto the analytical limit.
At station C5, a different pattern was observed. At the SWI,Mn concentration increased from winter to summer. There wasno apparent maximum close to the SWI in winter, while inMay and August 2002, concentrations were higher in the over-lying waters than in the superficial pore waters (6 and80 mmol L�1, respectively).
Profiles of dissolved Fe concentrations were similar forboth stations, the most important feature to observe beingthe occurrence of a concentration maximum below the SWI
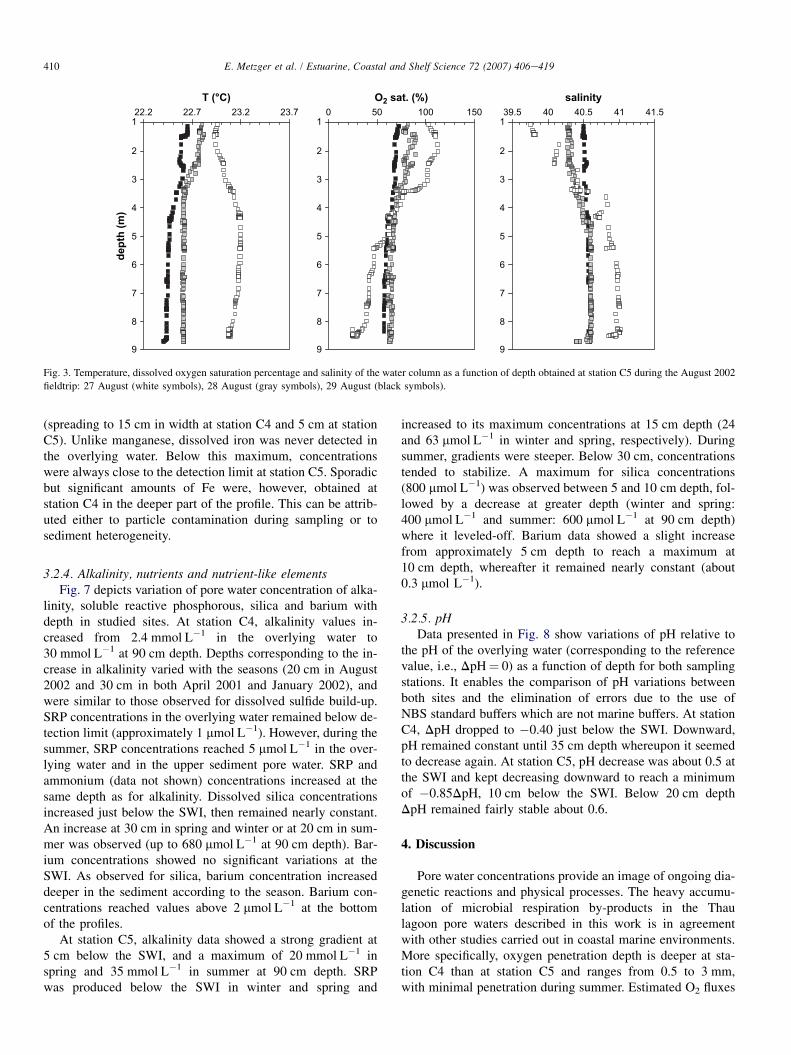
410 E. Metzger et al. / Estuarine, Coastal and Shelf Science 72 (2007) 406e419
1
2
3
4
5
6
7
8
9
22.2 22.7 23.2 23.7T (°C)
dep
th
(m
)1
2
3
4
5
6
7
8
9
0 50 100 150O
2 sat. (%)
1
2
3
4
5
6
7
8
9
39.5 40 40.5 41 41.5salinity
Fig. 3. Temperature, dissolved oxygen saturation percentage and salinity of the water column as a function of depth obtained at station C5 during the August 2002
fieldtrip: 27 August (white symbols), 28 August (gray symbols), 29 August (black symbols).
(spreading to 15 cm in width at station C4 and 5 cm at stationC5). Unlike manganese, dissolved iron was never detected inthe overlying water. Below this maximum, concentrationswere always close to the detection limit at station C5. Sporadicbut significant amounts of Fe were, however, obtained atstation C4 in the deeper part of the profile. This can be attrib-uted either to particle contamination during sampling or tosediment heterogeneity.
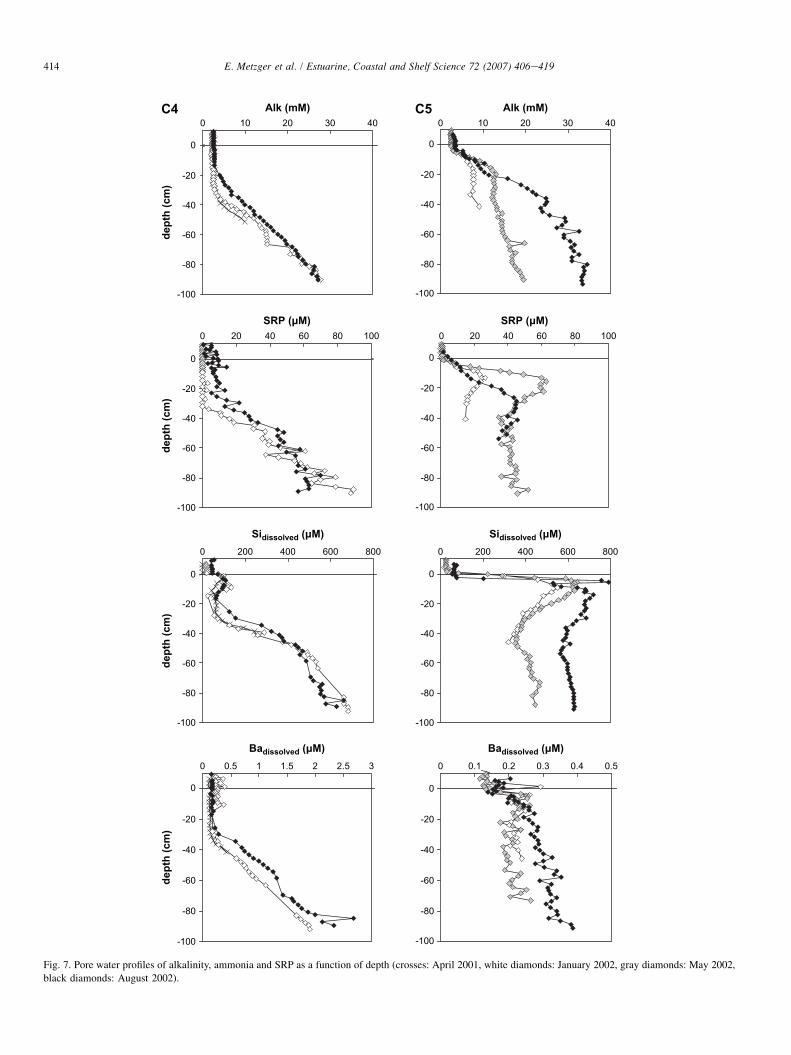
3.2.4. Alkalinity, nutrients and nutrient-like elementsFig. 7 depicts variation of pore water concentration of alka-
linity, soluble reactive phosphorous, silica and barium withdepth in studied sites. At station C4, alkalinity values in-creased from 2.4 mmol L�1 in the overlying water to30 mmol L�1 at 90 cm depth. Depths corresponding to the in-crease in alkalinity varied with the seasons (20 cm in August2002 and 30 cm in both April 2001 and January 2002), andwere similar to those observed for dissolved sulfide build-up.SRP concentrations in the overlying water remained below de-tection limit (approximately 1 mmol L�1). However, during thesummer, SRP concentrations reached 5 mmol L�1 in the over-lying water and in the upper sediment pore water. SRP andammonium (data not shown) concentrations increased at thesame depth as for alkalinity. Dissolved silica concentrationsincreased just below the SWI, then remained nearly constant.An increase at 30 cm in spring and winter or at 20 cm in sum-mer was observed (up to 680 mmol L�1 at 90 cm depth). Bar-ium concentrations showed no significant variations at theSWI. As observed for silica, barium concentration increaseddeeper in the sediment according to the season. Barium con-centrations reached values above 2 mmol L�1 at the bottomof the profiles.
At station C5, alkalinity data showed a strong gradient at5 cm below the SWI, and a maximum of 20 mmol L�1 inspring and 35 mmol L�1 in summer at 90 cm depth. SRPwas produced below the SWI in winter and spring and
increased to its maximum concentrations at 15 cm depth (24and 63 mmol L�1 in winter and spring, respectively). Duringsummer, gradients were steeper. Below 30 cm, concentrationstended to stabilize. A maximum for silica concentrations(800 mmol L�1) was observed between 5 and 10 cm depth, fol-lowed by a decrease at greater depth (winter and spring:400 mmol L�1 and summer: 600 mmol L�1 at 90 cm depth)where it leveled-off. Barium data showed a slight increasefrom approximately 5 cm depth to reach a maximum at10 cm depth, whereafter it remained nearly constant (about0.3 mmol L�1).
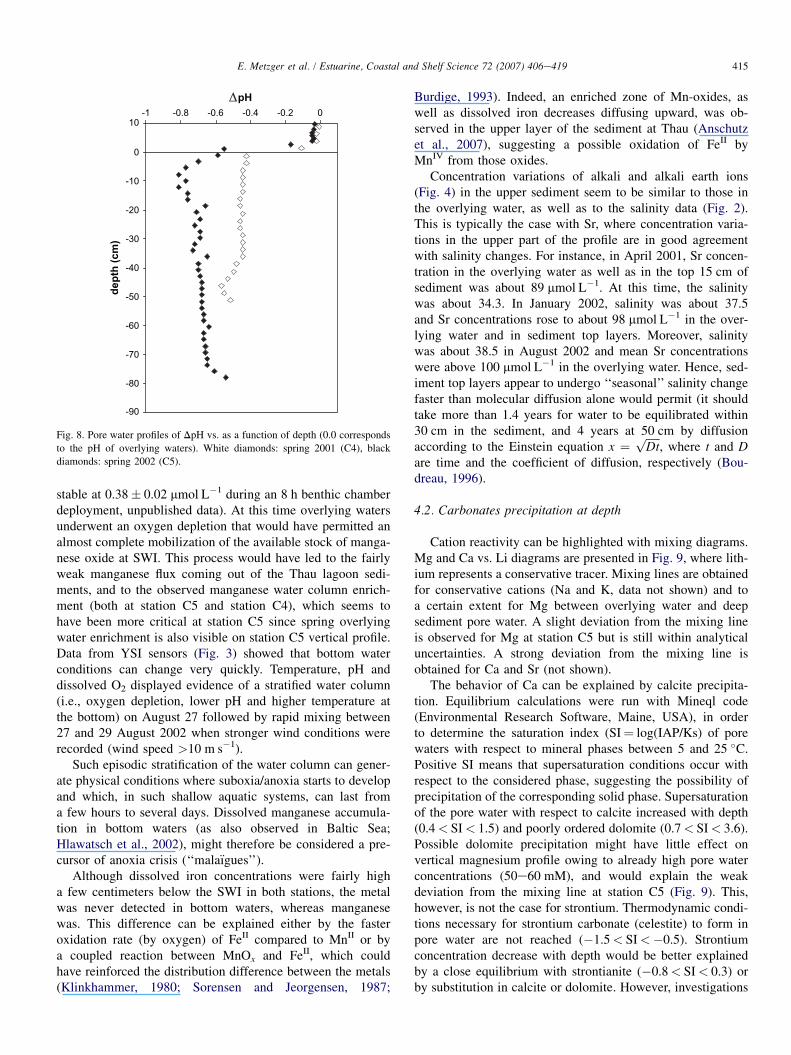
3.2.5. pHData presented in Fig. 8 show variations of pH relative to
the pH of the overlying water (corresponding to the referencevalue, i.e., DpH¼ 0) as a function of depth for both samplingstations. It enables the comparison of pH variations betweenboth sites and the elimination of errors due to the use ofNBS standard buffers which are not marine buffers. At stationC4, DpH dropped to �0.40 just below the SWI. Downward,pH remained constant until 35 cm depth whereupon it seemedto decrease again. At station C5, pH decrease was about 0.5 atthe SWI and kept decreasing downward to reach a minimumof �0.85DpH, 10 cm below the SWI. Below 20 cm depthDpH remained fairly stable about 0.6.
4. Discussion
Pore water concentrations provide an image of ongoing dia-genetic reactions and physical processes. The heavy accumu-lation of microbial respiration by-products in the Thaulagoon pore waters described in this work is in agreementwith other studies carried out in coastal marine environments.More specifically, oxygen penetration depth is deeper at sta-tion C4 than at station C5 and ranges from 0.5 to 3 mm,with minimal penetration during summer. Estimated O2 fluxes
411E. Metzger et al. / Estuarine, Coastal and Shelf Science 72 (2007) 406e419
-100
-80
-60
-40
-20
0
15 20 25 30 35
Lidissolved
(µM) Lidissolved
(µM)
dep
th
(cm
)
C4
-100
-80
-60
-40
-20
0
6 8 10 12 14
Cadissolved
(mM) Cadissolved
(mM)
dep
th
(cm
)
-100
-80
-60
-40
-20
0
30 40 50 60 70
Mgdissolved
(mM) Mgdissolved
(mM)
dep
th
(cm
)
-100
-80
-60
-40
-20
0
50 70 90 110
Srdissolved
(µM)
dep
th
(cm
)
-100
-80
-60
-40
-20
0
15 20 25 30 35
C5
-100
-80
-60
-40
-20
0
6 8 10 12 14
-100
-80
-60
-40
-20
0
30 40 50 60 70
-100
-80
-60
-40
-20
0
50 70 90 110
Srdissolved
(µM)
Fig. 4. Pore water profiles of major dissolved cations as a function of depth (crosses: April 2001, white diamonds: January 2002, gray diamonds: May 2002, black
diamonds: August 2002).
412 E. Metzger et al. / Estuarine, Coastal and Shelf Science 72 (2007) 406e419
March-April 2001
-100
-90
-80
-70
-60
-50
-40
-30
-20
-10
0
10
0 1000 2000 3000 4000H
2S (µM)
0 5 10 15 20 25 30 35
Dec-Jan 2002
-100
-90
-80
-70
-60
-50
-40
-30
-20
-10
0
10
0 1000 2000 3000 4000H
2S (µM)
dep
th
(cm
)
0 5 10 15 20 25 30 35
SO4
2-
dissolved (mM)
SO4
2-
dissolved (mM) SO
4
2-
dissolved (mM) SO
4
2-
dissolved (mM)
SO4
2-
dissolved (mM) SO
4
2-
dissolved (mM)C4
July-Aug 2002
-100
-90
-80
-70
-60
-50
-40
-30
-20
-10
0
10
0 1000 2000 3000 4000H
2S (µM)
0 5 10 15 20 25 30 35
Dec-Jan 2002
-100
-90
-80
-70
-60
-50
-40
-30
-20
-10
0
10
0 1000 2000 3000 4000H
2S (µM)
dep
th
(cm
)
0 5 10 15 20 25 30 35
C5
April-May 2002
-100
-90
-80
-70
-60
-50
-40
-30
-20
-10
0
10
0 1000 10002000 3000 4000H
2S (µM)
0 5 10 15 20 25 30 35
July-Aug 2002
-100
-90
-80
-70
-60
-50
-40
-30
-20
-10
0
10
0 2000 3000 4000H
2S (µM)
0 5 10 15 20 25 30 35
Fig. 5. Pore water profiles of total dissolved sulfide (white squares) and sulfate (black squares) as a function of depth.
range between 10 and 160 mmol m�2 d�1 (Dedieu et al.,2007), illustrating the eutrophic state of Thau lagoon. Atslightly greater depths where O2 penetration was recorded,nitrate peaks have been observed by Anschutz et al. (2007)(a few mM at station C4 and station C5). Nitrate totally van-ished below 4 cm depth as a consequence of bacterialdenitrification (Klinkhammer, 1980; Herbert, 1999). Manga-neseIIIeIV and ironIII dissimilatory bacterial reduction alsooccurred in the vicinity of the SWI. Occasional injection ofdissolved manganese into the water column is discussed indetail in the next section.
As expected in coastal sediment, sulfate reduction is a majorreaction of OM degradation in the Thau lagoon system, lead-ing to hydrogen sulfide build-up (up to 3.6 mmol L�1 at stationC4). Sulfate reduction rates were about 10 times higher thanthe oxygen consumption rate during spring 2002 and 2003,
suggesting that sulfate reduction was the main process ofOM mineralization (Point et al., 2007). Released H2S reactsrapidly with dissolved ferrous species to form insoluble ironsulfide, keeping
PH2S at low levels in the upper part of the
sediment (Lapp and Balzer, 1993; Elbaz-Poulichet et al.,2005). This process efficiently limits dissolved iron downwardpenetration (Berner, 1984). The occurrence of dissolved sul-fide close to the SWI may therefore explain the narrow peaksof ferrous iron found at station C5. During the warm season,sulfate reduction was more intense with a steeper gradientand a weaker concentration at depth.
At station C4, a certain remarkable feature of pore watercomposition was observed. Dissolved sulfide was not detectedwithin the first 20e30 cm of sediment, while several indicatorsof sulfide precipitation were nonetheless observed there: theblack color of the sediments indicates likely FeS formation
413E. Metzger et al. / Estuarine, Coastal and Shelf Science 72 (2007) 406e419
March-April 2001
-100
-90
-80
-70
-60
-50
-40
-30
-20
-10
0
10
0 50 100 150 200 250
0 5 10 15 20
Dec-Jan 2002
-100
-90
-80
-70
-60
-50
-40
-30
-20
-10
0
10
0 20 40 60 80
dep
th
(cm
)
0 5 10 15 20
Mndissolved
(µM)
Mndissolved
(µM) Mndissolved
(µM) Mndissolved
(µM)
Mndissolved
(µM) Mndissolved
(µM)C4
July-Aug 2002
-100
-90
-80
-70
-60
-50
-40
-30
-20
-10
0
10
0 20 40 60 80Fe
dissolved (µM)Fe
dissolved (µM)Fe
dissolved (µM)
Fedissolved
(µM) Fedissolved
(µM) Fedissolved
(µM)
0 20 40 60 80
C5
Dec-Jan 2002
-100
-90
-80
-70
-60
-50
-40
-30
-20
-10
0
10
0 20 40 60 80
dep
th
(cm
)
0 5 10 15 20
April-May 2002
-100
-90
-80
-70
-60
-50
-40
-30
-20
-10
0
10
0 20 40 60 80
0 5 10 15 20
July-Aug 2002
-100
-90
-80
-70
-60
-50
-40
-30
-20
-10
0
10
0 50 100 150 200 250
0 20 40 60 80
Fig. 6. Pore water profiles of total dissolved manganese (closed symbols) and iron (open symbols) as a function of depth.
(Lapp and Balzer, 1993); framboidal pyrite was also identifiedin the entire core for both stations (Elbaz-Poulichet et al.,2005); sulfate reduction rates were measured in the regionof 0e2 cm (0.6 mmol/cm3/day, Monperrus, pers. comm.);and total solid sulfur increased with depth according to the dis-solved sulfide gradient (Anschutz et al., 2007). The verticalgap of about 15 cm thickness between the dissolved FeII
peak and theP
H2S build-up zone at station C4 representsan original feature of this station and will be discussed in de-tail further in the text. The general influence of shellfish farm-ing on benthic response is discussed in the last section.
4.1. Benthic response to seasonsand transient stratification
Both stations showed seasonal chemistry changes in porewaters. For instance, dissolved oxygen profiles obtained with
Clark’s microelectrodes (Dedieu et al., 2007) showed well-ox-ygenated overlying water above the diffusive boundary layerfor both sites during winter and spring. During summer,both stations presented oxygen depleted overlying water(bottom water saturation: 55% at station C4 and 29% at stationC5).
Sulfate, manganese and oxygen gradients were sharper insummer than in winter conditions. Flux of SRP through theSWI was also higher in summer at station C5 and wasobserved in station C4 (Fig. 7). This change suggests a higherintensity of OM degradation in summer. It also suggests thatphysical processes responsible for the homogenization of pro-files in the upper part of the sediment at station C4 are not asintensive as usual, thereby permitting nutrient gradients andtheir efficient recycling to be observed.
However, at station C5, no dissolved manganese flux wasdetected the day after peeper retrieval (concentration remained
414 E. Metzger et al. / Estuarine, Coastal and Shelf Science 72 (2007) 406e419
-100
-80
-60
-40
-20
0
0 10 20 30 40Alk (mM)C5
-100
-80
-60
-40
-20
0
0 200 400 600 800
Sidissolved
(µM)
-100
-80
-60
-40
-20
0
0 0.1 0.2 0.3 0.4 0.5
Badissolved
(µM)
-100
-80
-60
-40
-20
0
0 10 20 30 40Alk (mM)
dep
th
(cm
)
C4
-100
-80
-60
-40
-20
0
0 200 400 600 800
Sidissolved
(µM)
dep
th
(cm
)
-100
-80
-60
-40
-20
0
0 0.5 1 1.5 2 2.5 3
Badissolved
(µM)
dep
th
(cm
)
-100
-80
-60
-40
-20
0
0 20 40 60 80 100SRP (µM)
dep
th
(cm
)
-100
-80
-60
-40
-20
0
0 20 40 60 80 100SRP (µM)
Fig. 7. Pore water profiles of alkalinity, ammonia and SRP as a function of depth (crosses: April 2001, white diamonds: January 2002, gray diamonds: May 2002,
black diamonds: August 2002).
415E. Metzger et al. / Estuarine, Coastal and Shelf Science 72 (2007) 406e419
stable at 0.38� 0.02 mmol L�1 during an 8 h benthic chamberdeployment, unpublished data). At this time overlying watersunderwent an oxygen depletion that would have permitted analmost complete mobilization of the available stock of manga-nese oxide at SWI. This process would have led to the fairlyweak manganese flux coming out of the Thau lagoon sedi-ments, and to the observed manganese water column enrich-ment (both at station C5 and station C4), which seems tohave been more critical at station C5 since spring overlyingwater enrichment is also visible on station C5 vertical profile.Data from YSI sensors (Fig. 3) showed that bottom waterconditions can change very quickly. Temperature, pH anddissolved O2 displayed evidence of a stratified water column(i.e., oxygen depletion, lower pH and higher temperature atthe bottom) on August 27 followed by rapid mixing between27 and 29 August 2002 when stronger wind conditions wererecorded (wind speed >10 m s�1).
Such episodic stratification of the water column can gener-ate physical conditions where suboxia/anoxia starts to developand which, in such shallow aquatic systems, can last froma few hours to several days. Dissolved manganese accumula-tion in bottom waters (as also observed in Baltic Sea;Hlawatsch et al., 2002), might therefore be considered a pre-cursor of anoxia crisis (‘‘mala€ıgues’’).
Although dissolved iron concentrations were fairly higha few centimeters below the SWI in both stations, the metalwas never detected in bottom waters, whereas manganesewas. This difference can be explained either by the fasteroxidation rate (by oxygen) of FeII compared to MnII or bya coupled reaction between MnOx and FeII, which couldhave reinforced the distribution difference between the metals(Klinkhammer, 1980; Sorensen and Jeorgensen, 1987;
-90
-80
-70
-60
-50
-40
-30
-20
-10
0
10-1 -0.8 -0.6 -0.4 -0.2 0
pH
dep
th
(cm
)
Fig. 8. Pore water profiles of DpH vs. as a function of depth (0.0 corresponds
to the pH of overlying waters). White diamonds: spring 2001 (C4), black
diamonds: spring 2002 (C5).
Burdige, 1993). Indeed, an enriched zone of Mn-oxides, aswell as dissolved iron decreases diffusing upward, was ob-served in the upper layer of the sediment at Thau (Anschutzet al., 2007), suggesting a possible oxidation of FeII byMnIV from those oxides.
Concentration variations of alkali and alkali earth ions(Fig. 4) in the upper sediment seem to be similar to those inthe overlying water, as well as to the salinity data (Fig. 2).This is typically the case with Sr, where concentration varia-tions in the upper part of the profile are in good agreementwith salinity changes. For instance, in April 2001, Sr concen-tration in the overlying water as well as in the top 15 cm ofsediment was about 89 mmol L�1. At this time, the salinitywas about 34.3. In January 2002, salinity was about 37.5and Sr concentrations rose to about 98 mmol L�1 in the over-lying water and in sediment top layers. Moreover, salinitywas about 38.5 in August 2002 and mean Sr concentrationswere above 100 mmol L�1 in the overlying water. Hence, sed-iment top layers appear to undergo ‘‘seasonal’’ salinity changefaster than molecular diffusion alone would permit (it shouldtake more than 1.4 years for water to be equilibrated within30 cm in the sediment, and 4 years at 50 cm by diffusionaccording to the Einstein equation x ¼
ffiffiffiffiffiDtp
, where t and Dare time and the coefficient of diffusion, respectively (Bou-dreau, 1996).
4.2. Carbonates precipitation at depth
Cation reactivity can be highlighted with mixing diagrams.Mg and Ca vs. Li diagrams are presented in Fig. 9, where lith-ium represents a conservative tracer. Mixing lines are obtainedfor conservative cations (Na and K, data not shown) and toa certain extent for Mg between overlying water and deepsediment pore water. A slight deviation from the mixing lineis observed for Mg at station C5 but is still within analyticaluncertainties. A strong deviation from the mixing line isobtained for Ca and Sr (not shown).
The behavior of Ca can be explained by calcite precipita-tion. Equilibrium calculations were run with Mineql code(Environmental Research Software, Maine, USA), in orderto determine the saturation index (SI¼ log(IAP/Ks) of porewaters with respect to mineral phases between 5 and 25 �C.Positive SI means that supersaturation conditions occur withrespect to the considered phase, suggesting the possibility ofprecipitation of the corresponding solid phase. Supersaturationof the pore water with respect to calcite increased with depth(0.4< SI< 1.5) and poorly ordered dolomite (0.7< SI< 3.6).Possible dolomite precipitation might have little effect onvertical magnesium profile owing to already high pore waterconcentrations (50e60 mM), and would explain the weakdeviation from the mixing line at station C5 (Fig. 9). This,however, is not the case for strontium. Thermodynamic condi-tions necessary for strontium carbonate (celestite) to form inpore water are not reached (�1.5< SI<�0.5). Strontiumconcentration decrease with depth would be better explainedby a close equilibrium with strontianite (�0.8< SI< 0.3) orby substitution in calcite or dolomite. However, investigations

416 E. Metzger et al. / Estuarine, Coastal and Shelf Science 72 (2007) 406e419
on solids (Elbaz-Poulichet et al., 2005) did not detect dolo-mite. Therefore, calcite precipitation remains the best meansof explaining the calcium and strontium decreases. Carbonateprecipitation is tightly linked to sulfate reduction, whichgenerates HS� and HCO3
� and causes alkalinity andP
CO2
increase. Alkalinity increase is stoichiometrically more impor-tant, however, than proton production (related to
PCO2 in-
crease and FeS precipitation). These processes increasesolution SI and support CaCO3 precipitation (Mucci et al.,2000). It would therefore explain the depth correspondenceof supersaturation conditions and the observed dissolved sul-fide increase for both stations.
4.3. Downward shift of chemical gradientsin station C4 pore waters
In this section, the striking feature of pore water composi-tion of diagenetic by-products at station C4 is discussed. Incomparison to station C5, pore water data at station C4 (refer-ence station located at the central zone of the lagoon) clearlyshow a downward shift of some by-products of OM diagenesis(alkalinity, nutrients,
PH2S, etc.). This unusual shift creates
a large gap between the zone of oxygen, nitrate, manganeseand iron oxide reduction and hydrogen sulfide build-up.
7
8
9
10
11
12
18 20 22 24 26 28 30Li
Ca
37
42
47
52
57
62
18 20 22 24 26 28 30Li
Mg
Fig. 9. Correlation of Ca and Mg vs. Li in pore waters sampled in August 2002
at station C4 (white diamonds) and station C5 (black diamonds). The line
corresponds to the mixing line between seawater and karstic water (from Aqui-
lina et al., 2002).
Additionally, alkaline and alkali earth cation concentrationprofiles, as well as sulfate profiles, show quasi-steady stateconcentrations below 50 cm, whereas upper layers seem toundergo seasonal salinity or metabolic activity variations.
The gap would therefore appear to be the consequence ofphysical processes, which decrease the pore water renewaltime and prevent slow reactions from being expressed (i.e.,there would seem to be a competition between kinetic andtransport processes). Fluid flow into sediment can have severalorigins: wave induced advection, lateral transport of karsticwater, physical sediment reworking (bottom current inducedresuspension/deposition, human activity such as dredging),and biological sediment reworking.
The possibility that biological processes or a general re-working of the upper sediment might be responsible for thegap can be eliminated at station C4 for the following reasons:(1) 210Pb measurements performed in sediment cores of stationC4 show that the mixed/homogenous sediment layer never ex-ceeded 5 cm (Schmidt et al., 2007). (2) Isotopic tracer resultsare consistent with luminophore incubation experiments. Thelatter enables an estimation of maximum macrofauna penetra-tion that does not exceed 10 cm depth at the period of higheractivity (warm season). Average value throughout the year is5 cm (Duport et al., 2007). Advection of bottom water throughthe upper layer of sediment remains the most likely explanationof the downward shift of gradients as suggested by Huettel andWebster (2000). Unfortunately, direct permeability data arenot available, but they can be estimated roughly using theKozenyeCarman empiric equation established for sandy sedi-ments as a first approximation of the permeability (Huettel andWebster, 2000). Station C4 sediments appear to be one order ofmagnitude more permeable than those of station C5 (Table 1).Wind and wave effects should be more discernable at stationC4 than at station (where farming structures tend to stronglylimit currents and water column mixing, Harzallah andChapelle, 2002). Silty clay sediment like that in Thau lagooncannot be classified as ‘‘permeable’’ (Huettel and Webster,2000; Janssen et al., 2005) and would hardly allow efficientpore water advection. However, shell debris, which are nottaken into account for granulometry determination but are fre-quently observed in sediment cores, could increase sedimentpermeability.
4.4. Influence of intensive shellfish farming
The main source of energy for heterotrophic bacteria aswell as benthic fauna is the organic matter (OM) accumulated
Table 1
Calculated permeability of sediment of Thau lagoon estimated from Kozenye
Carman equation (k¼ 5.6� 10�3F3d2/(1�F)2) (Huettel and Webster, 2000),
where k is the permeability (m2), F is the porosity and d is the mean grain
diameter (mm)
Station Porosity Mean grain size Estimated permeability
C4 0.85 15� 10�6 3� 10�11
C5 0.70 12� 10�6 3� 10�12

417E. Metzger et al. / Estuarine, Coastal and Shelf Science 72 (2007) 406e419
in the sediment. The concentration and lability (freshness) oforganic matter determine the efficiency of its mineralizationand the concentrations of its by-products. Metabolite accumu-lation in pore waters results from the balance between reactionrates and transport processes. TOC measurements at the top ofthe sedimentary column show that the organic content atstation C5 is higher than at station C4 (Anschutz et al.,2007): about 6% at the station C5 SWI, decreasing sharplyto 3.4% at 10 cm depth, and then remaining almost constant,whereas at station C4 TOC was about 4% at the SWI anddecreased steadily to 2.4% down to 75 cm depth (Elbaz-Poulichet et al., 2005). Benthic macrofauna differences werealso found between studied sites. Organism counting and iden-tification in station C5 sediments showed characteristic popu-lations of organic rich sediments (Deslous-Paoli et al., 1998;Duport et al., 2007). The quality of OM that reaches SWI atstation C4 is also more refractory (Mesnage et al., pers.comm.).
As a consequence, the diagenetic sequence at station C5 iscompressed in a thin sediment layer, as illustrated by the largeincrease of mineralization by-products at 5 cm depth down-ward. Below 35 cm, steady state seems to be achieved forseveral compounds (SO4
2�,P
H2S,P
NH3, SRP, dissolvedsilica). Anoxia or sub-anoxia crises may affect this stationperiodically, as is also suggested by the temporal variabilityof macrofauna populations (Duport et al., 2007). These condi-tions are generated by diffusion of sulfide and other reducedcompounds into the overlying waters as observed in May2003 (Jezequel et al., 2007; Metzger et al., 2007). In contrastto this, station C4 showed a relative stability of macrofaunapopulations with time. Due to lower labile POC input and pos-sible advective inflow, the diagenetic sequence is much morespread out at station C4.
Discrepancies of dissolved barium distribution (Fig. 7) be-tween the two stations illustrate the influence of shellfish farm-ing. Dissolved barium increases sharply with depth at stationC4 (20e30 cm depth), while at station C5 dissolved bariumshows only a very weak increase. This difference could beexplained by a higher barium regeneration rate at station C5SWI, a richer barium biodeposit at station C4 (feces wouldcontain less barium than diatom aggregates), or both. Bariumis mostly associated with clays and also with deposited organicmatter particles and would produce profiles similar to those ofnutrients (SRP, ammonia, dissolved silica) (Froelich et al.,1979; von Breymann et al., 1992; Torres et al., 1996; Nahrand Bohrmann, 1999). At both stations, dissolved barium con-centrations in the top 50 cm can be controlled by barite precip-itation/dissolution where SI values range from �0.3 to 0.2. SIvalues were calculated using Rushdi et al.’s (2000) apparentsolubility constant computed at salinity 37 and 15 �C forpure barite and strontian barite (slightly more soluble). Below50 cm, a shift towards supersaturated SI values is observed(from 0.4 to 0.7 at station C4 and from �0.3 to 0.4 at stationC5). Barium enrichment in pore waters must therefore be dueto sulfate depletion. Barium concentrations appear to bemainly controlled by barite formation. Although station C5 en-counters undersaturation conditions with respect to barium
sulfate, barium does not accumulate significantly in porewaters. A lower stock of ‘‘reactive’’ barium in sediments be-low 50 cm depth would explain this observation and supportthe hypotheses of different settling particles and regenerationrates between stations.
Differences in pore water composition between the sites aremainly due to the establishment of intensive shellfish farming.The presence of shellfish tables has changed hydrodynamicconditions in the lagoon (Chapelle et al., 2000), and therefore,the sediment deposition conditions. Physical perturbationsmay include the weakening of currents, winds and swells, aswell as increased temperatures due to lower evaporation ratesand better absorption of sun light by farm structures and thehigh density of animals suspended at Intensive shellfish farm-ing in Thau lagoon started in 1920 (Deslous-Paoli et al., 1998),which corresponds to about 27 cm of sediment at station C4and 13 cm at station C5 according to sedimentation ratesfrom Elbaz-Poulichet et al. (2005) (respectively, 0.33 and0.16 cm y�1). At station C5, this thickness corresponds tothe increase in mean grain size (from about 10 mm at 15 cmdepth to 19 mm at the SWI; Schmidt et al., 2007). No signifi-cant changes are observed in mean grain size at station C4 butdata are reliable only for the first 45 cm. Detritic contributionto the sediment deposition in the lagoon lessened after the es-tablishment of shellfish farming as shown by the decrease ofthorium in the upper part of station C4 sediment from 30 cmdepth (Elbaz-Poulichet et al., 2005). Roughly, at the samedepth, trace element data (U, Cu, Cd, Pb, Ag, Mo, Ba, Sr,not shown) display clear concentration discontinuities in thesolid phase (Elbaz-Poulichet, pers. commun.). Before shellfishfarming, plankton was probably uniformly deposited in thewhole lagoon and supplied labile organic matter to the sedi-ment. In contrast, after the establishment of shellfish farming,the distribution of organic carbon was concentrated within thefarming zones. Bivalves are probably efficient filteringorganisms at the lagoon scale and may clear the lagoon whichfavor primary production, according to Deslous-Paoli et al.(1998). This process generated differences in the benthic faunaand flora observed between studied sites and probably alteredthe composition of organic matter (mainly in terms of lability)as discussed and/or the OM fluxes.
5. Conclusion
Pore water compositions at two stations, one located in thevicinity of a shellfish farming site (C5) and the other awayfrom the shellfish farming area (C4), were studied in winter,spring and summer. In the shellfish production zone, the earlydiagenetic sequence is condensed within the top 5 cm and thewhole sequence seems to be achieved within 30 cm. Indeed,higher input of labile organic matter generates sharp gradientsof redox sensitive species in the upper sediment and sulfidebuild-up starts at 5 cm below the SWI. Conversely, at the ref-erence station, dissolved iron and manganese appeared withinthe first 10e15 cm and sharp increase of other OM minerali-zation by-product concentrations are found deeper, from 20to 30 cm in depth. The diagenetic sequence was extended

418 E. Metzger et al. / Estuarine, Coastal and Shelf Science 72 (2007) 406e419
over more than 90 cm (the depth limit of this study). Theunusual shift of hydrogen sulfide gradient (and many othercompounds) possibly originates from advective flow that‘‘sweeps’’ dissolved species or induces precipitation (e.g., bar-ite). Even if permeability appears to be low, other physicalprocesses such as diffusion, bioturbation or sediment resus-pension were eliminated as possible explanations for this shift.
High inputs of POC at station C5 sediment from shellfishexcreta are also responsible for partial anoxia in the overlyingwater, especially during the warm season when oxygenrenewal is lower. This results in the occasional release ofdissolved manganese in bottom waters, especially at stationC5. This situation is reinforced by low wind conditions andstabilization effects due to farm structures (waves and currentsweakening and consequent temperature increasing). Neverthe-less, it should be emphasized that rapid water column evolu-tions were observed in a 1-day period, as demonstrated bybottom water re-oxygenation. In conclusion, shellfish farminghas physical implications as well as biological and diageneticeffects, both of which favor anoxia crises (‘‘mala€ıgues’’).
Due to permanent reducing conditions prevailing below theSWI, the sedimentary column of Thau lagoon potentially con-stitutes an efficient trap for trace metals. For example, owingto higher pore water sulfide build-up in the shellfish farmingzone, divalent metals such as cadmium can accumulate effi-ciently. This role is greatly discussed in other articles of thepresent volume (Metzger et al., 2007; Muresan et al., 2007;Point et al., 2007).
Acknowledgements
This study was supported by PNEC (MICROBENT pro-gram) and IFREMER. The authors thank J.-L. Guillou(IFREMER Sete) for the field support; M. Evrard, D. Lav-ergne, M. Pepe (LGE, University of Paris 7) for analyses assis-tance and J.-J. Bourrand (LGIS, Universite Paris 7) for divingassistance. Special thanks to the anonymous reviewers fortheir very precise and helpful remarks and suggestions andto Richard Anker for English reviewing.
References
Aller, R.C., Hall, P.O.J., Rude, P.D., Aller, J.Y., 1998. Biogeochemical hetero-
geneity and suboxic diagenesis in hemipelagic sediments of the Panama
Basin. Deep Sea Research Part I: Oceanographic Research Papers 45,
133e165.
Anschutz, P., Chaillou, G., Lecroart, P., 2007. Phosphorous diagenesis in the
sediment of the Thau Lagoon. Estuarine, Coastal and Shelf Science 72
(3), 447e456.
Aquilina, L., Ladouche, B., Doerfliger, N., Seidel, J.L., Bakalowicz, M.,
Dupuy, C., Le Strat, P., 2002. Origin, evolution and residence time of sa-
line thermal fluids (Balaruc springs, southern France): implications for
fluid transfer across the continental shelf. Chemical Geology 192, 1e21.
Berner, R.A., 1984. Sedimentary pyrite formation: an update. Geochimica et
Cosmochimica Acta 48, 605e615.
Boudreau, B.P., 1996. The diffusive tortuosity of fine-grained unlithified
sediments. Geochimica et Cosmochimica Acta 60, 3139e3142.
Burdige, D.J., 1993. The biogeochemistry of manganese and iron reduction in
marine sediments. Earth Science Reviews 35, 249e284.
von Breymann, M.T., Brumsack, H., Emeis, K.C., 1992. Depositional and
diagenitic behavior of barium in the Japan sea. In: Pisciotto, K.A., Ingle
Jr., J.C., von Breymann, M.T., Barron, J., et al. (Eds.), Proceedings of
the ODP, Scientific Results, Ocean Drilling Program, College Station,
Texas, pp. 651e665.
Carignan, R., Rapin, F., Tessier, A., 1985. Sediment pore water sampling for
metal analysis. Geochimica et Cosmochimica Acta 49, 2493e2497.
Chapelle, A., Menesguen, A., Deslous-Paoli, J.-M., Souchu, P., Mazouni, N.,
Vaquer, A., Millet, B., 2000. Modelling nitrogen, primary production
and oxygen in a Mediterranean lagoon. Impact of oysters farming and
inputs from the watershed. Ecological Modelling 127, 161e181.
de Casabianca, M.-L., Laugier, T., Marinho-Soriano, E., 1997. Seasonal
changes of nutrients in water and sediment in a Mediterranean lagoon
with shellfish farming activity (Thau Lagoon, France). ICES Journal of
Marine Science 54, 905e916.
Dedieu, K., Thouzeau, G., Chavaud, L., Clavier, J., Jean, F., Rabouille, C.,
2007. Benthic O2 distribution and dynamics in a Mediterranean lagoon
(Thau, France): An in situ microelectrode study. Estuarine, Coastal and
Shelf Science 72 (3), 393e405.
Deslous-Paoli, J.-M., Souchu, P., Mazouni, N., Juge, C., Dagault, F., 1998.
Relationship between environment and resources: impact of shellfish farm-
ing on a Mediterranean lagoon (Thau, France). Oceanologica Acta 21,
831e843.
Duport, E., Gilbert, F., Poggiale, J.-C., Dedieu, K., Rabouille, C., Stora, G.,
2007. Benthic macrofauna and sediment reworking quantification in
contrasted environments in the Thau Lagoon. Estuarine, Coastal and Shelf
Science 72 (3), 522e533.
Elbaz-Poulichet, F., Seidel, J.L., Jezequel, D., Metzger, E., Prevot, F.,
Simonucci, C., Sarazin, G., Viollier, E., Etcheber, H., Jouanneau, J.-M.,
2005. Sedimentary record of redox-sensitive elements (U, Mn, Mo) in a transi-
tory anoxic basin (the Thau lagoon, France). Marine Chemistry 95, 271e281.
Froelich, P.N., Klinkhammer, G.P., Bender, M.L., Luedtke, N.A., Heath, G.R.,
Cullen, D., Dauphin, P., Hammond, D., Hartman, B., Maynard, V., 1979.
Early oxidation of organic matter in pelagic sediments of the eastern
equatorial Atlantic: suboxic diagenesis. Geochimica et Cosmochimica
Acta 43, 1075e1090.
Gangnery, A., Chabirand, J.-M., Lagarde, F., Le Gall, P., Oheix, J., Bacher, C.,
Buestel, D., 2003. Growth model of the Pacific oyster, Crassostrea gigas, cul-
tured in Thau Lagoon (Mediterranee, France). Aquaculture 215, 267e290.
Gilbert, F., Souchu, P., Bianchi, M., Bonin, P., 1997. Influence of shellfish
farming activities on nitrification, nitrate reduction to ammonium and
denitrification at the wateresediment interface of the Thau lagoon, France.
Marine Ecology Progress Series 151, 143e153.
Harzallah, A., Chapelle, A., 2002. Contribution of climate variability to occur-
rences of anoxic crises ‘malaigues’ in the Thau lagoon (southern France).
Oceanologica Acta 25, 79e86.
Herbert, R.A., 1999. Nitrogen cycling in coastal marine ecosystems. FEMS
Microbiology Reviews 23, 563e590.
Hesslein, R.H., 1976. An in-situ sampler for close interval pore water studies.
Limnology and Oceanography 21, 912e914.
Hlawatsch, S., Neumann, T., van den Berg, C.M.G., Kersten, M., Harff, J.,
Suess, E., 2002. Fast-growing, shallow-water ferro-manganese nodules
from the western Baltic Sea: origin and modes of trace element incorpora-
tion. Marine Geology 182, 373e387.
Huettel, M., Webster, I.T., 2000. Porewater flow in permeable sediments. In:
Boudreau, B., Jorgensen, B.B. (Eds.), The Benthic Boundary Layer.
Oxford University Press, Oxford, pp. 144e179.
Janssen, F., Huettel, M., Witte, U., 2005. Pore-water advection and solute
fluxes in permeable marine sediments (II): benthic respiration at three
sandy sites with different permeabilities (German Bight, North Sea). Lim-
nology and Oceanography 50, 779e792.
Jezequel, D., Brayner, R., Viollier, E., Metzger, E., Prevot, F., Fievet, F., 2007.
Two-dimensional determination of dissolved iron and sulfur species in ma-
rine sediment pore-waters by thin-films based imaging. Thau lagoon
(France). Estuarine, Coastal and Shelf Science 72 (3), 420e431.
Klinkhammer, G.P., 1980. Early diagenesis in sediments from the eastern
equatorial Pacific, II. Pore water metal results. Earth and Planetary Science
Letters 49, 81e101.

419E. Metzger et al. / Estuarine, Coastal and Shelf Science 72 (2007) 406e419
Lapp, B., Balzer, W., 1993. Early diagenesis of trace metals used as an
indicator of past productivity changes in coastal sediments. Geochimica
et Cosmochimica Acta 57, 4639e4652.
Lazure, P., 1992. Etude de la dynamique de l’Etang de Thau par modele
numerique tridimentionnel. Vie et Milieu 42, 137e145.
Lopez, P., 2003. Effect of changes in water salinity on ammonium, calcium,
dissolved inorganic carbon and influence on water/sediment dynamics.
Estuarine, Coastal and Shelf Science 56, 943e956.
Mazouni, N., Gaertner, J.-C., Deslous-Paoli, J.-M., 1998. Influence of oyster
culture on water column characteristics in a coastal lagoon (Thau, France).
Hydrobiologia 373e374, 149e156.
Mesnage, V., 1994. L’etude de la mobilite des formes de phosphate a l’inter-
face eau-sediment dans des ecosystemes lagunaires. PhD thesis, Universite
de Montpellier I, Montpellier, France, unpublished.
Metzger, E., Simonucci, C., Viollier, E., Sarazin, G., Prevot, F., Elbaz-
Poulichet, F., Seidel, J.L., Jezequel, D., 2007. Influence of diagenetic
processes in Thau lagoon on cadmium behavior and benthic fluxes.
Estuarine, Coastal and Shelf Science 72 (3), 497e510.
Millet, B., 1989. Fonctionnement hydrodynamique du bassin de Thau. Valida-
tion d’un modele numerique de circulation (programme ECOTHAU).
Oceanologica Acta 12, 37e46.
Mucci, A., Sundby, B., Gehlen, M., Arakaki, T., Zhong, S., Silverberg, N.,
2000. The fate of carbon in continental shelf sediments of eastern Canada:
a case study. Deep Sea Research Part II: Topical Studies in Oceanography
47, 733e760.
Muresan, B., Cossa, D., Jezequel, D., Prevot, F., Kerbellec, S., 2007. The bio-
geochemistry of mercury at the sedimentewater interface in the Thau la-
goon. 1. Partition and speciation. Estuarine, Coastal and Shelf Science 72
(3), 472e484.
Nahr, T.H., Bohrmann, G., 1999. Barium-rich authigenic clinoptilolite in
sediments from the Japan Sea e a sink for dissolved barium? Chemical
Geology 158, 227e244.
Plante-Cuny, M.-R., Plante, R., Mazouni, N., Fontaine, M.-F., Souchu, P., De-
slous-Paoli, J.-M., Grenz, C., 1998. Oxygen fluxes involving the benthic
micro- and macrophytic components in the Thau Lagoon under pre-anoxic
conditions. Oceanologica Acta 21, 819e829.
Podda, F., Michard, G., 1994. Mesure colorimetrique de l’alcalinite.
Comptes Rendus de l’Academie des Sciences de Paris. Serie II 319,
651e657.
Point, D., Monperrus, M., Tesssier, E., Amouroux, D., Donard, O.F.X.,
Guyonneaud, R., Leynaert, A., Chavaud, L., Thouzeau, G., Graille, R.,
2007. Biological control of trace metal and organometal benthic fluxes
in a eutrophic lagoon (Thau lagoon, Mediterranean sea, France). Estuarine,
Coastal and Shelf Science 72 (3), 457e471.
Rushdi, A., McManus, J., Collier, R.W., 2000. Marine barite and celestite
saturation in seawater. Marine Chemistry 69, 19e31.
Sarazin, G., Michard, G., Prevot, F., 1999. A rapid and accurate spectroscopic
method for alkalinity measurements in sea water samples. Water Research
33, 290e294.
Schmidt, S., Jouanneau, J.-M., Weber, O., Lecroart, P., Radakovitch, O.,
Gilbert, F., Jezequel, D., 2007. Sedimentary processes in the Thau Lagoon
(France): from seasonal to century time scales. Estuarine, Coastal and
Shelf Science 72 (3), 534e542.
Sorensen, J., Jeorgensen, B.B., 1987. Early diagenesis in sediments from
Danish coastal waters: microbial activity and MneFeeS geochemistry.
Geochimica et Cosmochimica Acta 51, 1583e1590.
Souchu, P., Abadie, E., Vercelli, C., Buestel, D., Sauvagnargues, J.C., 1998a.
La crise anoxique du bassin de Thau de l’ete 1997. Bilan du phenomene et
perspectives. IFREMER, Sete, France, unpublished, pp. 33.
Souchu, P., Gasc, A., Collos, Y., Vaquer, A., Tournier, H., Bibent, B.,
Deslous-Paoli, J.M., 1998b. Biogeochemical aspects of bottom anoxia in
a Mediterranean lagoon (Thau, France). Marine Ecology Progress Series
164, 135e146.
Tabatabai, M.A., 1974. A rapid method for determination of sulfate in water
samples. Environmental Letters 7, 237e242.
Torres, M.E., Brumsack, H.J., Bohrmann, G., Emeis, K.C., 1996. Barite fronts
in continental margin sediments: a new look at barium remobilization in
the zone of sulfate reduction and formation of heavy barites in diagenetic
fronts. Chemical Geology 127, 125e139.
Webster, I.T., Teasdale, P.R., Grigg, N.J., 1998. Theoretical and experimental
analysis of peeper equilibration dynamics. Environmental Science and
Technology 32, 1727e1733.




















