
Behavioral effects of phencyclidine and ketamine alone and in combination with other drugs
11
European Journal of Pharmacology, 144 (1987) 331-341 331 Elsevier EJP 50052 Behavioral effects of phencyclidine and ketamine alone and in combination with other drugs Larry D. Byrd 1,,, Leanna J. Standish and Leonard L. Howell Yerkes Regional Primate Research Center and i Departments of Pharmacology and Psychology, Emory University, Atlanta, Georgia 30322, U.S.A. Received 15 September 1987, accepted 29 September 1987 The behavioral effects of phencyclidine (PCP) and ketamine administered alone and in combination with naloxone, atropine, methyl atropine, chlorpromazine and d-amphetamine were studied in squirrel monkeys trained to press a response lever under a fixed-ratio 30 schedule maintained by the termination of a stimulus associated with electric shock presentation. Under non-drug conditions, a period of high-rate responding in the presence of the stimulus associated with shock presentation was followed by a period of no responding during a 40-s timeout scheduled between fixed-ratio components. Mean rates of responding during fixed-ratio components decreased monotonically as PCP dose increased from 0.1 to 0.56 mg/kg, and doses of 3.0 and 5.6 mg/kg ketamine produced decreases in mean response rate comparable to doses of 0.3 and 0.56 mg/kg PCP. The dose-effect functions revealed that ketamine was approximately one-tenth as potent as PCP. The present data also characterized the time-course effects of PCP and ketamine, with the former having effects that were slower in onset yet more persistent in time. None of the drugs studied in combination with PCP and ketamine provided evidence of a pharmacological antagonism of the behavioral effects of the latter two drugs. Rather, the data indicated an enhancement of behavioral effects when certain drug combinations were studied. Phencyclidine; Ketamine; Naloxone; Atropine; Schedule-controlled behavior; (Squirrel monkey) 1. Introduction Previous work has suggested that the behav- ioral effects of PCP and ketamine are qualitatively similar, although PCP is more potent and its dura- tion of action is greater (Balster, 1980). PCP has discriminative properties which generalize to ketamine (Holtzman, 1980), both compounds have been shown to disrupt performance on simulta- neous brightness discriminations (Tang and Franklin, 1983), and both drugs increase respond- * To whom all correspondence should be addressed: Yerkes Regional Primate Research Center, Emory University, Atlanta, Georgia 30322, U.S.A. ing under fixed-interval schedules of food presen- tation in the mouse (Wenger and Dews, 1976), rat (Meliska and Trevor, 1978) and pigeon (Wenger, 1976). These reports have shown that sub- anesthetic doses of PCP and ketamine can have similar effects on schedule-controlled behavior. Several neurotransmitter systems have been im- plicated in the actions of PCP and its structural analogues. Smith et al. (1980) reported that an analgesic effect of ketamine was inhibited in rats by the opiate antagonist, naloxone, and that ketamine displaced [3H]naloxone in an opiate re- ceptor-binding assay. Vincent et al. (1978) re- ported that PCP can also compete for binding to opiate receptors in the brain. Moreover, certain 0014-2999/87/$03.50 © 1987 Elsevier Science Publishers B.V. (Biomedical Division)
-
Upload
independent -
Category
Documents
-
view
1 -
download
0
Transcript of Behavioral effects of phencyclidine and ketamine alone and in combination with other drugs

European Journal of Pharmacology, 144 (1987) 331-341 331 Elsevier
EJP 50052
Behavioral effects of phencyclidine and ketamine alone and in combination with other drugs
L a r r y D. Byrd 1,,, L e a n n a J. S tand ish and L e o n a r d L. Howel l
Yerkes Regional Primate Research Center and i Departments of Pharmacology and Psychology, Emory University, Atlanta, Georgia 30322, U.S.A.
Received 15 September 1987, accepted 29 September 1987
The behavioral effects of phencyclidine (PCP) and ketamine administered alone and in combination with naloxone, atropine, methyl atropine, chlorpromazine and d-amphetamine were studied in squirrel monkeys trained to press a response lever under a fixed-ratio 30 schedule maintained by the termination of a stimulus associated with electric shock presentation. Under non-drug conditions, a period of high-rate responding in the presence of the stimulus associated with shock presentation was followed by a period of no responding during a 40-s timeout scheduled between fixed-ratio components. Mean rates of responding during fixed-ratio components decreased monotonically as PCP dose increased from 0.1 to 0.56 mg/kg, and doses of 3.0 and 5.6 mg/kg ketamine produced decreases in mean response rate comparable to doses of 0.3 and 0.56 mg/kg PCP. The dose-effect functions revealed that ketamine was approximately one-tenth as potent as PCP. The present data also characterized the time-course effects of PCP and ketamine, with the former having effects that were slower in onset yet more persistent in time. None of the drugs studied in combination with PCP and ketamine provided evidence of a pharmacological antagonism of the behavioral effects of the latter two drugs. Rather, the data indicated an enhancement of behavioral effects when certain drug combinations were studied.
Phencyclidine; Ketamine; Naloxone; Atropine; Schedule-controlled behavior; (Squirrel monkey)
1. Introduction
Previous work has suggested that the behav- ioral effects of PCP and ketamine are qualitatively similar, although PCP is more potent and its dura- tion of action is greater (Balster, 1980). PCP has discriminative properties which generalize to ketamine (Holtzman, 1980), both compounds have been shown to disrupt performance on simulta- neous brightness discriminations (Tang and Franklin, 1983), and both drugs increase respond-
* To whom all correspondence should be addressed: Yerkes Regional Primate Research Center, Emory University, Atlanta, Georgia 30322, U.S.A.
ing under fixed-interval schedules of food presen- tation in the mouse (Wenger and Dews, 1976), rat (Meliska and Trevor, 1978) and pigeon (Wenger, 1976). These reports have shown that sub- anesthetic doses of PCP and ketamine can have similar effects on schedule-controlled behavior.
Several neurotransmitter systems have been im- plicated in the actions of PCP and its structural analogues. Smith et al. (1980) reported that an analgesic effect of ketamine was inhibited in rats by the opiate antagonist, naloxone, and that ketamine displaced [3H]naloxone in an opiate re- ceptor-binding assay. Vincent et al. (1978) re- ported that PCP can also compete for binding to opiate receptors in the brain. Moreover, certain
0014-2999/87/$03.50 © 1987 Elsevier Science Publishers B.V. (Biomedical Division)

332
opioids can produce discriminative stimulus ef- fects similar to those produced by PCP and ketamine. Dose-related increases in PCP-ap- propriate responding have been occasioned by opioids in the rat (Holtzman, 1980) and the squir- rel monkey (Brady and Balster, 1982; Holtzman, 1982), and ketamine-appropriate responding has been occasioned by opioids in the pigeon (Herling et al., 1981). Studies have also implicated cholinergic mechanisms in the pharmacology of PCP. Adams (1980) and Glick et al. (1979) re- ported that PCP mimicked and potentiated the behavioral effects of muscarinic anticholinergics and that cholinergic agonists antagonized the ef- fects of PCP. These data are consistent with other reports that PCP binds to muscarinic receptors (Vincent et al., 1978). PCP has also been reported to inhibit dopamine uptake in vivo (Vickroy and Johnson, 1980) and to release dopamine in vitro (Vickroy and Johnson, 1982). PCP can enhance amphetamine-induced stereotypy in the rat (Bal- ster and Chait, 1978), and neuroleptics can osten- sibly inhibit PCP-induced stereotypy (Castellani and Adams, 1981). PCP can also affect schedule- controlled behavior in a manner very similar to that of d-amphetamine in the rat (Murray, 1978) and the squirrel monkey (Byrd, 1982). These stud- ies suggest that catecholaminergic mechanisms may also be involved in the pharmacology of PCP.
Given the number of reports implicating several neurotransmitter systems in the mediation of the effects of PCP and ketamine, the present study was undertaken (1) to characterize the effects of PCP and ketamine on aversively maintained, con- ditioned behavior in the squirrel monkey and to describe the time course of the effects, and (2) to determine whether the behavioral effects of the two drugs might be modulated by the administra- tion of certain other drugs known to have behav- ioral effects in the squirrel monkey. A fixed-ratio schedule of stimulus-shock termination was cho- sen because little is known about the effects of PCP and ketamine on aversively maintained be- havior, and because this schedule is known to provide excellent, stable support of fixed-ratio performance in the squirrel monkey (Byrd, 1977; Kelleher and Morse, 1964; Morse and Kelleher, 1966).
2. Materials and methods
2.1. Subjects
Three adult male squirrel monkeys (Saimiri sciureus) served as subjects. All were housed indi- vidually with continuous access to water and were maintained at free-feeding body weight.
2.2. Apparatus
During daily test sessions, each monkey was seated in a Plexiglas chair of the type described by Kelleher and Morse (1964) with modifications (Byrd, 1979), and enclosed within a ventilated, sound-attenuating chamber equipped with white noise to mask extraneous sounds. A response lever (Coulbourn Inst ruments Co., No. E21-03) mounted on the wall in front of the monkey registered a lever press (response) when operated by a downward force greater than .19 N. Illumina- tion was provided by pairs of 7.5-W a.c. colored lights mounted on the front wall just above eye level. The monkey's tail was held motionless in a small stock, and two brass electrodes rested on a shaved portion near the end of the appendage. Electrode paste (EKG Sol) minimized changes in impedance between the tail, the electrodes and a 6-mA stimulus of 200 ms duration. A Radio Shack TRS-80:I microcomputer controlled experimental events on-line and recorded data.
2.3. Procedure
Monkeys were trained to press a response lever to terminate a light associated with the occasional delivery of electric shock (Morse and Kelleher, 1966). Initially, shock was delivered every 15 s in the presence of a red light, and a response terminated the red light and initiated a brief (2-s) period of white light that was followed by a time- out when no lights were present. Responses had no consequences during the white light or the timeout. As response rate increased, the duration of the timeout, the interval between shocks and the number of responses required to terminate the red light were increased. During the final stage of training, 30 responses were required to terminate

333
the red fight [fixed-ratio 30], shock was scheduled every 30 s following onset of the red light, and the timeout period was 40 s. Test sessions were con- ducted five days per week, each session compris- ing 30 fixed-ratio components or a maximum of 60 min, and each component was followed by a 40-s timeout. To preclude the presentation of ex- cessive shocks due to drug-induced impairment of responding, the shock source was disconnected on days when drugs were administered. Absence of the shock on drug days presented no problem or effect because the squirrel monkeys seldom re- ceived a shock under the fixed-ratio 30 schedule when the shock unit was connected and, therefore, its absence was not discriminated. We have used this procedure with success in numerous studies, and we have no evidence that performance de- teriorates due to the absence of the shock source on drug days.
Drug administration and testing began once characteristic fixed-ratio responding was en- gendered and performance during daily sessions had stabilized for each monkey. Drugs were ad- ministered immediately prior to sessions on Tues- day and Fridays, and saline was administered on Thursdays as a control. Drugs were dissolved in 0.9% saline, doses were determined in terms of the salt, and injections were made intramuscularly in the thigh in a volume of 0.4-0.6 ml. Phencyclidine HC1 (PCP) in doses of 0.1-0.56 mg/kg and keta- mine HC1 in doses of 0.3-5.6 mg/kg were admin- istered alone and in combination with naloxone (1.0 and 3.0 mg/kg), atropine (1.0 mg/kg), methyl atropine (1.0 mg/kg), chlorpromazine (0.1 and 0.3 mg/kg) and d-amphetamine (0.1 and 0.3 mg/kg). The chronological order in which the drugs were studied was ketamine (alone and with naloxone), PCP (alone and with atropine), chlorpromazine (alone and with ketamine), d-amphetamine (alone and with ketamine or PCP), chlorpromazine and PCP, atropine (alone and with ketamine), nalox- one (alone and with PCP), and methyl atropine (alone and with ketamine or PCP). Each drug dose or drug combination was studied at least twice and, in most cases, three times in each monkey, and means ( + S.E.M.) for the group were derived from individual subject means. Control rates of responding were determined for each drug experi-
ment based on individual mean rates obtained on six or more occasions when saline was injected in each monkey. Mean control response rate (+S.E.M.) for the group was then derived for each drug experiment and used as the basis for calculating percent of control values. The signifi- cance of the differences between mean drug values was determined using Student's two-tailed t-test. Time-course effects for ketamine and PCP were derived by determining response rate during indi- vidual fixed-ratio components occurring at 10-min intervals following drug injection.
3. Results
3.1. Control performance
Responding under the stimulus-shock termina- tion schedule was characteristic of fixed-ratio per- formance as described previously (Byrd, 1977; Kelleher and Morse, 1964). The pattern of re- sponding comprised a period of responding at a high rate in the presence of the red light and little or no responding in the presence of the 2-s white light that appeared upon completion of the fixed- ratio 30 requirement. Responding did not occur during the 40-s timeout period between fixed-ratio components. Individual mean rates of responding per session in the presence of the red light were 1.92 (S-53), 1.17 (S-54) and 2.17 (S-59) responses/s for each of the three monkeys and, at these rates, sessions were usually completed within 30-40 min. A cumulative record, typical of fixed-ratio perfor- mance during saline and non-drug sessions, is shown in fig. 1 (top).
3.2. Effects of phencyclidine
Mean rates of fixed-ratio responding decreased in a dose-dependent manner following PCP ad- ministration. The rates obtained after 0.1 mg /kg did not differ from response rates after saline administration, and rates decreased monotonically as PCP dose increased to a maximum of 0.56 mg/kg. Response rates decreased to 40% of saline control rates after 0.56 mg/kg, and the pattern of responding was not characteristic of fixed-ratio

334
SALINE
,~ y J r- J i--
S - 5 3
0 . 3 m g / k g PCP
W 0~ w "I o S J Q. (/I UJ nr
0 . 5 6 mg /kg PCP
0
H
3 . 0 m g / k g KET
5 .6 m g / k g KET j -~
I I
5 M INUTES
Fig. 1. Effect of phencychdine and ketamine dose on pattern of responding under a fixed-ratio 30 stimulus-termination schedule in a squirrel monkey. Completion of each fixed-ratio 30 component was followed by a 40-s timeout. Drug or saline was injected i.m. immediately prior to a session. Each cumulative record shows the complete session or, at the highest doses, the first 40-50 min of the session. The response pen reset to baseline after five fixed-ratio components were completed. The event pen was displaced downwards during presentation of the red light associated with shock. Vertical marks on the event line indicate when shock would
have been delivered.
performance. Fixed-ratio responding after the two higher doses of PCP is i l lustrated in the cumula- tive records of fig. 1. As can be seen, responding was main ta ined at a decreased rate after 0.3 m g / k g PCP, but the response pat tern reflected a gradual acceleration in responding dur ing the red light rather than an abrup t change to a sustained, high rate of responding characteristic of fixed-ratio per- formance in the squirrel monkey. The gradual acceleration in responding and the generally lower
rate of sustained responding dur ing the red light cont r ibuted to lower mean rates after 0.3 m g / k g PCP.
3.3. Effects of ketamine
In contrast to PCP, a dose of 0.3 m g / k g keta- mine had no effect on mean response rate, and higher doses (3.0 and 5.6 m g / k g ) decreased re- sponding (fig. 1, bottom). A slightly enhanced rate

335
after 1.0 m g / k g was due primarily to the perfor- mance of one monkey, and the mean rate for the group after 1.0 m g / k g did not differ significantly f rom that obtained after saline. In comparison with phencyclidine, ketamine was approximately one-tenth as potent in disrupting responding. Do- ses of 3.0 and 5.6 m g / k g ketamine reduced mean response rate about as effectively as doses of 0.3 and 0.56 m g / k g PCP.
3. 4. Time-course effects of phencycfidine and keta- mine
A comparison of the time-course effects of a range of doses of PCP and ketamine characterized
KETAMINE
2.0
1.0
0
i .o t,o,.o Q:: 3,0] ~ ~ 3.0 mg/kg
2 0 ¢
5.6 ~ / k g
S-59 PHENCYCLIDINE
i~ ,17 mglkg
0.3 en~/k~
2.0
l 0
o , T Y T , , 0 I0 20 30 40 50 ~0 0 I0 ZO 30 40 50 60
TIME (rain)
Fig. 2. Time-course effects of ketamine (left) and saline or phencychdine (right) on mean response rate maintained under a fixed-ratio 30 stimulus-termination schedule• A data point at 0 s represents response rate during the first fixed-ratio compo- nent in a session; data points at other post-injection times were derived from individual fixed-ratio components corresponding to those times. Each data point is the mean resulting from three administrations in one squirrel monkey. Vertical lines
through the symbols indicate the S.E.M.
more definitively differences in the behavioral ef- fects of these two drugs. Phencyclidine had little or no effect on fixed-ratio performance at the lowest dose (0.1 mg /kg ) , but response rates de- creased at higher doses. The time to onset of the decreasing effect was less and the durat ion of the decreasing effect was greater as dose increased. The durat ion of the rate-decreasing action of the highest dose of PCP was sufficiently long that responding did not return to baseline or control levels even after 60 min post-injection. In contrast to PCP, the rate-decreasing effect of ketamine was more rapid in onset and the durat ion of the effect was briefer. Response rates returned to baseline levels within 40 min after the highest dose of ketamine, but after none of the doses of PCP that decreased responding was recovery so rapid. Time-course data representative of individual monkeys and illustrating differences in potency and in onset and durat ion of action of the two drugs are shown in fig. 2.
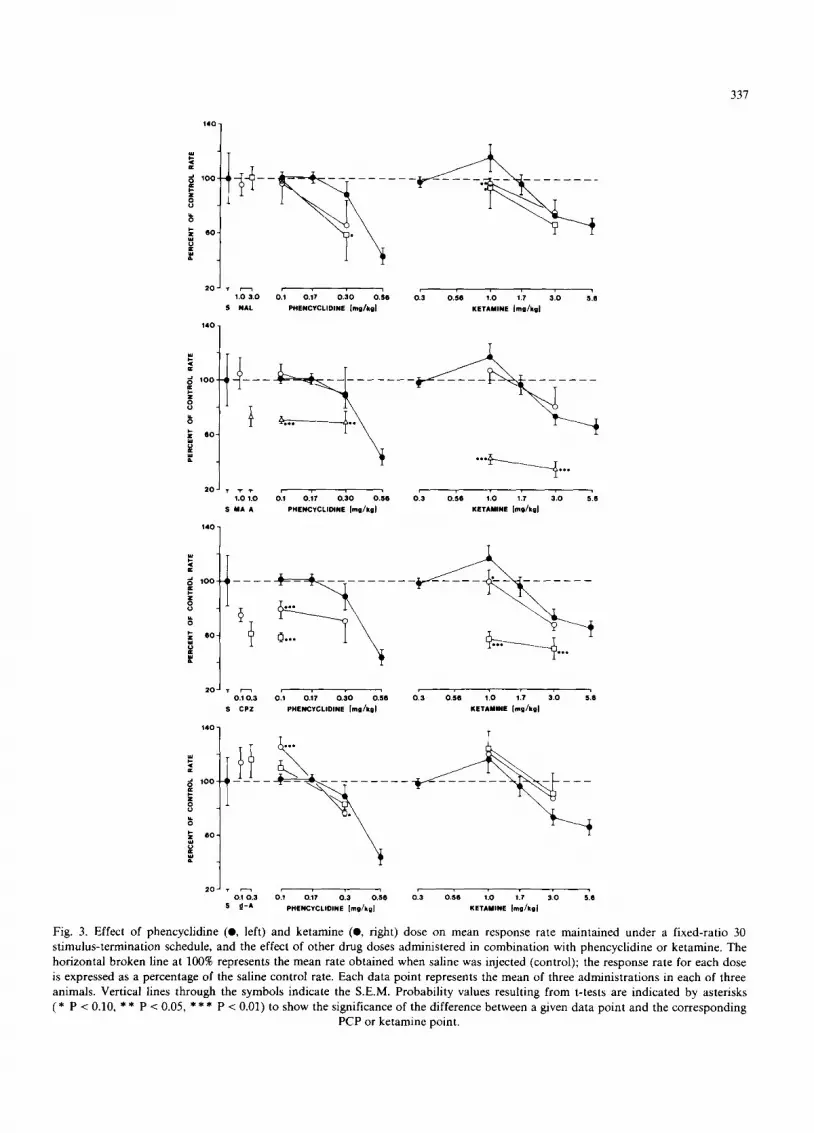
3.5. Effects of phencyclidine and ketamine in combi- nation with naloxone
Naloxone (1.0 or 3.0 m g / k g ) had little or no behavioral effect at 1.0 or 3.0 m g / k g when admin- istered alone or in combinat ion with a behavior- ally ineffective dose of PCP (0.1 mg /kg ) . Mean rates for the group of monkeys were approxi- mately the same whether 0.1 m g / k g PCP was administered alone or in combina t ion with either dose of naloxone (fig. 3, top left). However, both doses of naloxone enhanced the rate-decreasing effect of a higher dose of PCP (0.3 mg /kg ) , a dose of PCP that decreased rates to 88% of control values when administered alone. A consequence of administering either dose of naloxone with 0.3 m g / k g PCP was a failure of two of the three subjects to complete 30 fixed-ratio components within a 60-min session, an outcome that never occurred when 0.3 m g / k g PCP was administered alone (fig. 1). Moreover, when naloxone was ad- ministered in combinat ion with 1.0 m g / k g keta- mine, response rates were also lower than after either drug alone (fig. 3, top right). Combined with 1.0 m g / k g ketamine, both doses of naloxone reduced response rates substantially below those

336
observed after ketamine alone. However, naloxone had less effect on the rate-decreasing effect of a higher dose of ketamine (3.0 mg/kg) . A disruption of responding at the beginning of the session was typical after 3.0 m g / k g ketamine alone (fig. 1), and naloxone served to increase slightly the dura- tion of the disruption when combined with keta- mine. All subjects completed 30 fixed-ratio com- ponents within the 60-min limit whether ketamine was administered alone or with naloxone.
3. 6. Effects of phemyclidine and ketamine in combi- nation with atropine
Atropine (1.0 mg/kg) had a rate-decreasing effect whether administered alone or in combina- tion with PCP. A dose of 1.0 m g / k g atropine alone decreased overall mean rate to approxi- mately 75% of saline control rate and, when atro- pine was administered in combination with either 0.1 or 0.3 m g / k g PCP, response rates were signifi- cantly lower than those obtained with PCP alone (fig. 3, left). However, after none of the combina- tions of PCP and atropine did any subject fail to complete 30 fixed-ratio components within the 60-min session limit.
When 1.0 m g / k g atropine was administered in combination with ketamine, the interaction was more dramatic and response rates were signifi- cantly lower than those obtained with ketamine or atropine alone (fig. 3, right). Response rates fol- lowing the combination of atropine and ketamine were as low as 35% of saline control rates. Whereas all of the subjects completed 30 fixed-ratio com- ponents within 60 min after either atropine or ketamine alone, none did so after atropine com- bined with 3.0 m g / k g ketamine, and only one subject did so after atropine and 1.0 m g / k g ketamine.
3.7. Effects of phencyclidine and ketamine in combi- nation with methyl atropine
Methyl atropine (1.0 mg/kg) had little or no effect when administered alone or in combination with either PCP (0.1 and 0.3 mg/kg) or ketamine (1.0 and 3.0 mg /kg ) (fig. 3). The dose-effect curves for PCP and ketamine were unaltered by methyl
atropine, and statistical tests revealed no dif- ferences between group response rates as a conse- quence of methyl atropine administration.
3.8. Effects of phencyclidine and ketamine in combi- nation with chlorpromazine
Chlorpromazine decreased overall mean re- sponse rates when administered alone and in com- bination with PCP (fig. 3, left) or ketamine (fig. 3, right). The higher dose of chlorpromazine (0.3 mg/kg) alone decreased response rate and also disrupted the fixed-ratio pattern of responding: none of the subjects completed the session within the 60-min limit. The lower dose (0.1 mg/kg) decreased rate less, the fixed-ratio pattern re- mained relatively intact, and all subjects com- pleted the session. When either dose of chlor- promazine was administered in combination with a behaviorally ineffective dose of PCP (0.1 mg/kg) , response rates were significantly lower than those obtained with PCP alone. Whereas fixed-ratio per- formance was maintained at decreased rates after either 0.3 m g / k g PCP or 0.1 m g / k g chlorproma- zine alone, responding was interrupted by periods of no responding when the two were combined. Moreover, only two subjects completed the session in the time allotted when 0.1 m g / k g chlorproma- zine was administered in combination with 0.3 m g / k g PCP, and none completed the session when the higher dose of chlorpromazine was combined with 0.1 m g / k g PCP.
When chlorpromazine was administered in combination with ketamine, overall mean re- sponse rates were also lower than the rates ob- tained after ketamine alone (fig. 3, right), and the degree of response rate decrease covaried with the dose of chlorpromazine. Response rates after 1.0 m g / k g ketamine were reduced significantly, for example, when combined with 0.1 m g / k g chlor- promazine, and the rates were reduced further when the higher dose of chlorpromazine was com- bined with ketamine. The combined effect of 0.1 m g / k g chlorpromazine and 3.0 m g / k g ketamine was a marked disruption of the sustained high rate of responding typical of fixed-ratio performance. Instead of an abrupt change to a high response rate, the response pattern resembled more closely
337
i 100
o
20 j "r 1.O 3.0 OJ1 O.;7 O.iO 0.50
$ NAL PHENCYCLIDINE Img/kgJ
140
i 100
8
~ ao
20
140
100
~ ec
1OO
o
6o
,.~,.'o o'., o.~7 o.~o o.~ S MA A PHENCYCLIDINE [mg/kgl
o13 o.se 1'.o 1'.7 31o s',e KETAMINE Img/kgl
O'.3 O.56 1~O 117 3'.0 5'.8 KETAMINE [ms/kgJ
Z. X
T r-~ o.o.3 o:1 o.;7 o.;o o.sa
S CPZ PHENCYCLIDINE [mg/kgl O.3 O.56 1.O 1.7 3.0 5.6
KETAMINE (mg/kgJ
T O.1 0.3 O.1 O.17 0.3 0.56 0.3 0.56 1.O 1.7 3.0 5.6 S ~-A PHENCYCLIOINE [mg/kgJ KETAMINE Img/kgl
Fig. 3. Effect of phencyclidine (O, left) and ketamine (O, right) dose on mean response rate maintained under a fixed-ratio 30 stimulus-termination schedule, and the effect of other drug doses administered in combination with phencyclidine or ketamine. The horizontal broken line at 100% represents the mean rate obtained when saline was injected (control); the response rate for each dose is expressed as a percentage of the saline control rate. Each data point represents the mean of three administrations in each of three animals. Vertical lines through the symbols indicate the S.E.M. Probability values resulting from t-tests are indicated by asterisks ( * P < 0.10, * * P < 0.05, * * * P < 0.01) to show the significance of the difference between a given data point and the corresponding
PCP or ketamine point.

33g
the positively accelerated responding typical of fixed-interval schedules, and there were periods of no responding. When the higher dose of chlor- promazine (0.3 mg/kg) was administered in com- bination with either dose of ketamine (1.0 or 3.0 mg/kg), rates were decreased significantly, the response pattern was disrupted severely and there were extended periods of no responding.
3.9. Effects of phency'clidine and ketamine in combi- nation with d-amphetamine
d-Amphetamine (0.1 or 0.3 mg/kg) had a slight rate-increasing effect on fixed-ratio responding when administered alone, but the rates were not significantly different from saline control rates. When the lower dose of d-amphetamine (0.1 mg/kg) was administered in combination with a behaviorally ineffective dose of PCP (0.1 mg/kg), however, response rates were significantly higher than those obtained with PCP alone (fig. 3, left), although not significantly different from d- amphetamine alone. Both doses of d-amphetamine also enhanced the rate-decreasing effect of the higher dose of PCP (0.3 mg/kg). Whereas dis- cernible fixed-ratio patterns were maintained when either 0.1 mg /kg d-amphetamine or 0.3 mg/kg PCP was administered alone, patterns were dis- rupted and periods of no responding were ob- served when the two were administered together. In contrast, neither dose of d-amphetamine sig- nificantly altered the effect of either of two doses of ketamine (1.0 or 3.0 mg/kg), although rates tended to be higher (fig. 3, right). Similarly, the patterns of responding that were observed after d-amphetamine in combination with ketamine were not markedly different from those observed after ketamine alone. All subjects completed ses- sions in the time allotted when d-amphetamine was combined with ketamine, but only two of the subjects completed 30 fixed-ratio components when 0.3 mg/kg d-amphetamine was adminis- tered with 0.3 mg/kg PCP.
4. Discussion
The data reported here extend our knowledge of the behavioral pharmacology of phencyclidine
(PCP) and ketamine in the squirrel monkey. Previ- ous reports have described the effects of PCP on conditioned behavior in this species maintained under schedules of food presentation (Chait and Balster, 1978), but there are few data describing effects on aversively maintained behavior. In the present experiment, fixed-ratio responding was maintained by the termination of a stimulus asso- ciated with electric shock (i,e., fixed-ratio stimulus termination), and the effects of PCP were found to be qualitatively and quantitatively similar to the effects obtained when fixed-ratio responding re- sulted in food presentation (Chait and Balster, 1978). In the latter experiment and in the present one, increasing doses of PCP caused a monotonic decrease in mean rate of responding under a fixed-ratio schedule. Other reports have described similar effects of PCP on fixed-ratio responding in pigeons (Wenger, 1976) and mice (Wenger and Dews, 1976). These results provide increasing evi- dence that the behavioral effects of PCP have a generality that transcends the type of consequent event maintaining the behavior.
Comparison of the dose-effect curves for PCP and ketamine in the present experiment confirmed earlier indications of the relative potencies of these two drugs. The dose-effect curve for ketamine was displaced approximately one log unit to the right of the PCP curve, indicating that the former was one-tenth as potent as PCP in decreasing respond- ing under the fixed-ratio schedule. Brady et al. (1980) reported a comparable difference in potency when PCP and ketamine were studied in rhesus monkeys maintained under a fixed-interval sched- ule of food presentation, and similar differences have been observed in pigeons (Wenger, 1976) and mice (Wenger and Dews, 1976). In addition to a difference in potency, the present data also de- scribed differences in the duration of action of PCP and ketamine, with the former having effects that were slower in onset yet more persistent in time. Mean response rates returned to baseline levels typically within 40 min after the highest dose of ketamine, whereas responding did not return to baseline rates after as long as 60 min post-injection when the highest dose of PCP was studied. Previous studies have provided indirect evidence of differences in the duration of the

339
behavioral effects of PCP and ketamine (Byrd, 1982; Wenger, 1976; Wenger and Dews, 1976), and the present data provide confirming evidence of the differences in time-course effects.
None of the drugs studied in combination with PCP and ketamine provided evidence of a pharmacological antagonism of behavioral effects in that dose-effect curves were not shifted to the right. Indeed, the data indicated an enhancement of effect when certain drug combinations were studied. For example, pretreatment with atropine enhanced the rate-decreasing effects of PCP and ketamine. Enhancement of PCP's effects by anti- cholinergics has been described by others and has been attributed to antimuscarinic actions (Adams, 1980; Glick et al., 1979). However, the rate-de- creasing effects of atropine administered alone in the present study make interpretation of the re- sults difficult. A pretreatment dose of naloxone that had no effect when administered alone also enhanced the rate-decreasing effects of PCP, but inhibition of PCP metabolism and PCP elimina- tion by naloxone may have been responsible for the latter results. For example, Nabeshima et al. (1983) reported that the acute administration of morphine enhances a variety of pharmacological effects of PCP, and the authors suggested the inhibition of PCP metabolism and elimination by morphone as a possible mechanism.
The role of catecholamines in the mediation of the behavioral effects of PCP and ketamine is relatively ill-defined. Balster and Chait (1978) re- ported that PCP enhanced the behavioral effects of amphetamine, and they suggested that PCP may have a dopaminergic action. However, there was little evidence for enhancement of the behav- ioral effects of PCP and ketamine by ampheta- mine in the present study. Others have reported that the neuroleptics haloperidol and chlor- promazine can antagonize the effects of PCP, another indication that dopaminergic mechanisms are involved (Garey et al., 1980; Murray and Horita, 1979). In the present study, however, pre- treatment with chlorpromazine only enhanced the rate-decreasing effects of PCP and ketamine, pro- viding no evidence for antagonism. The latter findings may have resulted from chlorpromazine- induced inhibition of PCP and ketamine metabo-
lism. Oxidative biotransformation in the liver is the major route of metabolism of PCP (Wong and Biemann, 1976) and ketamine (Chang and Glazko, 1974). Drugs, such as chlorpromazine, that also undergo oxidative biotransformation by the hepatic system may compete for common meta- bolic pathways and, thereby, enhance the effects of PCP or ketamine. For example, diazepam or secobarbital can increase ketamine-induced sleep time as well as the plasma half-life of ketamine in humans (Lo and Cumming, 1975). Also, diazepam can prolong the biological half-life of ketamine and reduce the concentration of ketamine required to achieve adequate anesthesia (Idvall et al., 1983). Given the propensity of chlorpromazine to inhibit oxidative drug metabolism (Rubin et al., 1964), the interactions observed in the present study may have resulted from metabolic factors.
Previous reports have demonstrated similarities in the pharmacology of PCP and ketamine, and in the present study, the interactions between PCP and a variety of drugs were qualitatively similar to those observed for ketamine. These results suggest that PCP and ketamine may have similar pharmacological mechanisms of action. However, it is not evident from the present study whether the actions of PCP and ketamine are mediated through an interaction with PCP-specific receptor sites or whether these sites are involved in the actions of other pharmacological agents. PCP binding sites in the brain, which may be PCP receptors, have been identified by several inves- tigators (Quirion et al., 1982; Zukin and Zukin, 1979). The probability exists, however, that the effects of PCP on the central nervous system are mediated through PCP-specific receptors which interact with several neurotransmitter systems. Blocking the interaction of PCP with one neuro- transmitter system may not prevent it from pro- ducing a behavioral effect through another system. In addition, the contribution of metabolic effects and nonspecific interactions with neurotransmitter systems remains to be delineated. Experiments designed to elucidate the relative contribution of metabolic factors and receptor-mediated effects will help distinguish further the pharmacology of these two compounds.

340
Acknowledgements
This research was supported by U.S. Public Health Service grants DA-01161 and RR-00165 to the Yerkes Regional Primate Research Center from the Division of Research Resources, National Institutes of Health. The Yerkes Center is fully accredited by the American Association for Accreditation of Laboratory Animal Care. R. Penrod, M. Arthur, P. Plant, H. Wells and F. Kiernan assisted in preparing the manuscript and illustrations. Drugs were provided graciously by Endo Labora- tories (naloxone HCI), Lilly Laboratories (atropine sulfate), Pennwah Corporation (methyl atropine nitrate), Smith, Kline and French Laboratories (chlorpromazine HCI and d- amphetamine sulfate), Parke-Davis (ketamine HCI) and the National Institute on Drug Abuse (phencyclidine HCI).
References
Adams, P.M., 1980, Interaction of phencyclidine with drugs affecting cholinergic neurotransmission, Neuropharmacol- ogy 19, 151.
Balster, R.L., 1980, The effects of phencyclidine and three analogues on motor performance in mice, Pharmacology 20, 46.
Balster, R.L. and L.D. Chait, 1978, The effects of phencycli- dine on amphetamine stereotypy in rats, European J. Pharmacol, 48, 445.
Brady, K.T. and R.L. Balster, 1982, Discriminative stimulus properties of stereoisomers of cyclazocine in phencyclidine- trained squirrel monkeys, Life Sci. 31, 541.
Brady, K.T., R.L. Balster, L.T. Meltzer and D. Schwertz, 1980, Comparison of phencyclidine and three analogues on fixed-interval performance in rhesus monkeys, Pharmacol. Biochem. Behav. 12, 67.
Byrd, L.D., 1977, Responding in the squirrel monkey under fixed-ratio schedules of stimulus termination, J. Exp. Anal. Behav. 27, 495.
Byrd, L.D., 1979, The behavioral effects of cocaine: rate de- pendency or rate constancy, European J. Pharmacol. 56, 355.
Byrd, L.D.. 1982, Comparison of the behavioral effects of phencyclidine, ketamine, d-amphetamine and morphine in the squirrel monkey, J. Pharmacol. Exp. Ther. 220, 139.
Castellani, S. and P.M. Adams, 1981, Effects of dopaminergic drugs on phencyclidine-induced behavior in the rat, Neuro- pharmacology 20, 371.
Chait, L.D. and R.L. Balster, 1978, The effects of acute and chronic phencyclidine on schedule-controlled behavior in the squirrel monkey, J. Pharmacol. Exp. Ther. 204, 77.
Chang, T. and A.J. Glazko, 1974, Biotransformation and dis- position of ketamine, Int. Anesthesiol. Clin. 12, 157.
Garey, R.E., S. McQuilty, D. Tootle and R.G. Heath, 1980, The effects of apomorphine and Haldol on PCP-induced behavioral and motor abnormalities in the rat, Life Sci. 26, 277.
Glick, S.D., R.D. Cox, S. Maayani and R.C. Meibach, 1979. Anticholinergic behavioral effect of phencyclidine, European J. Pharmacol. 59, 103.
Herling, S., E,H. Coale Jr., D.W. Hein, G. Winger and J.H. Woods, 1981, Similarity of the discriminative stimulus ef- fects of ketamine, cyclazocine, and dextrophan in the pi- geon, Psychopharmacology 73, 286.
Holtzman, S.G., 1980, Phencyclidine-like discriminative effects of opioids in the rat, J. Pharmacol. Exp. Ther. 214, 614.
Holtzman, S.G., 1982, Phencyclidine-like discriminative stimu- lus properties of opioids in the squirrel monkey, Psycho- pharmacology 77, 295.
Idvall, J., K.F. Aronsen, P. Stenberg and L, Paalzow, 1983, Pharmacodynamic and pharmacokinetic interactions be- tween ketamine and diazepam, European J. Clin. Pharma- col. 24, 337.
Kelleher, R.T. and W.H. Morse, 1964, Escape behavior and punished behavior, Fed. Proc. 23, 808.
Lo, N.J. and J.F. Cumming, 1975, Interaction between sedative premedicants and ketamine in man and in isolated perfused rat livers, Anesthesiology 43, 307.
Meliska, C.J. and A.J. Trevor, 1978, Differential effects of ketamine on schedule-controlled responding and motility, Pharmacol. Biochem. Behav. 8, 679.
Morse, W.H. and R.T. Kelleher, 1966, Schedules using noxious stimuli. 1. Multiple fixed-ratio and fixed-interval termina- tion of schedule complexes, J. Exp. Anal. Behav. 9, 267.
Murray, T.F., 1978, The effects of phencyclidine on operant behavior in the rat: biphasic effect and tolerance develop- ment, Life Sci. 22, 195.
Murray, T.F. and A. Horita, 1979, Phencyclidine-induced stereotyped behavior in rats: dose response effects and antagonism by neuroleptics, Life Sci. 24, 2217,
Nabeshima, T., S.P. Sivam and I.K. Ho, 1983, Effect of morphine on the responses to and disposition of phen- cyclidine in mice. I. Enhancement of phencyclidine effects by acute morphine administration, J. Pharmacol. Exp. Ther. 225, 325.
Quirion, R., R.P. Hammer Jr., M. Herkenham and C.B. Pert, 1982, Autoradiographic localization of the phencyclidine/ sigma 'opiate' receptor in rat brain, in: Problems of Drug Dependence 1981, ed. L.S. Harris (National Institute on Drug Abuse Monograph 41, Washington, D.C.), p. 178.
Rubin, A., T.R. Tephly and G.J. Mannering, 1964, Kinetics of drug metabolism by hepatic microsomes, Biochem. Pharmacol. 13, 1007.
Smith, D.J., G.M. Pekoe, L i . Martin and B. Coalgate, 1980, The interaction of ketamine with the opiate receptor, Life Sci. 26, 789.
Tang, A.H. and S.R. Franklin, 1983, Disruption of brightness discrimination in a shock avoidance task by phencyclidine and its antagonism in rats, J. Pharmacol. Exp. Ther. 225, 503.
Vickroy, T.W. and K.M. Johnson, 1980, In vivo administration of phencyclidine inhibits 3H-dopamine accumulation by rat brain striatal slices, Subst. Ale. Actions/Misuse 1, 351.

Vickroy, T.W. and K.M. Johnson, 1982, Similar dopamine-re- leasing effects of phencyclidine and noramphetamine stimulants in striatal slices, J. Pharmacol. Exp. Ther. 223, 669.
Vincent, J.P., D. Cavey, J.M. Kamenka, P. Geneste and M. Lazdunski, 1978, Interaction of phencyclidine with the muscarinic and opiate receptors in the central nervous system, Brain Res. 152, 176.
Wenger, G.R., 1976, The effect of phencyclidine and ketamine
341
on schedule-controlled behavior in the pigeon, J. Pharma- col. Exp. Ther. 196, 172.
Wenger, G.R. and P.B. Dews, 1976, The effects of phencycli- dine, ketamine, d-amphetamine and pentobarbital on schedule-controlled behavior in the mouse, J. Pharmacol. Exp. Ther. 196, 616.
Wong, I.K. and K. Biemann, 1976, Metabolites of phencycli- dine, Clin. Toxicol. 9, 583.
Zukin, S.R. and R.S. Zukin, 1979, Specific [3H]phencyclidine binding in rat central nervous system, Proc. Natl. Acad. Sci. U.S.A. 76, 5372.




















