
Barrier characteristics of epithelial cultures modelling the airway and intestinal mucosa: A...
7
Barrier characteristics of epithelial cultures modelling the airway and intestinal mucosa: A comparison Driton Vllasaliu a,⇑,1 , Robyn Fowler a,1 , Martin Garnett a , Mike Eaton b , Snow Stolnik a a Division of Drug Delivery and Tissue Engineering, School of Pharmacy, Boots Science Building, University of Nottingham, University Park, Nottingham NG7 2RD, UK b European Technology Platform for Nanomedicine Secretariat, VDI/VDE Innovation + Technik GmbH, Department Innovation Europe, Steinplatz 1, 10623 Berlin, Germany article info Article history: Received 6 October 2011 Available online 2 November 2011 Keywords: Calu-3 Caco-2 Tight junctions Epithelial barrier Macromolecular permeability Mucosal delivery abstract The barrier characteristics of polarized layers of Calu-3 and Caco-2 cell lines, as commonly used in vitro models of intestinal and airway mucosa, respectively, were investigated by assessing the translocation of model macromolecules and nanoparticles. The barrier capacity of the cell layers towards the movement of macromolecules and nanoparticulates differed considerably between the cell lines. Permeability stud- ies revealed the existence of a notably larger solute molecular weight limit for paracellular diffusion in Caco-2 monolayers compared to Calu-3 cells. Removal of mucus in Calu-3 cells resulted in cell layers exhibiting a larger macromolecular permeability, in addition to improved nanoparticle translocation. Microscopic examination of the tight junctions, as cellular features that play a major role in preventing transepithelial movement of macromolecules, revealed that the appearance of cell–cell boundaries was notably different in the two cell lines, which could explain the differences in macromolecular permeabil- ity. The data overall showed that epithelial layers of airway Calu-3 and intestinal Caco-2 cell cultures in vitro exhibit a different level of restrictiveness and this is due to the cell morphology and the presence of mucus. Ó 2011 Elsevier Inc. All rights reserved. 1. Introduction The use of in vitro methods for investigation of mucosal drug absorption is commonplace in the field of drug delivery. A wealth of knowledge, especially mechanistic information, has been gained from these studies. In vitro investigations on mucosal drug absorp- tion often employ appropriately selected and cultured epithelial cell lines, providing models of mucosal tissue of interest. In this re- spect, the intestinal Caco-2 cell line is probably most extensively used to study intestinal drug absorption. The excellent in vitro–in vivo correlation observed with the Caco-2 cell line [1] has enabled its use as a tool not only to study mechanisms but also predict in vivo oral drug absorption. The bronchially derived Calu-3 cell line, on the other hand, is commonly used as a model of the upper airways in studies of drug absorption across the upper lung and the nose [2,3]. This cell line has also demonstrated good in vitro–in vivo correlation [4] and possesses the capacity to produce mucus [5], which is advantageous in studying the effect of mucus on drug absorption. Previous studies have reported the existence of differences in barrier characteristics between Calu-3 and Caco-2 cell lines, in addition to apparent morphological variations, which are expected considering their different mucosal origin. However, it is not en- tirely clear from the literature what cellular features give rise to this disparity in the capacity to act as a barrier to macromolecular translocation. Our work sought to clarify this by investigating the restrictiveness of polarized Calu-3 and Caco-2 cell layers. The bar- rier capacity of the cell layers towards the permeability of model macromolecules and proteins over a wide molecular weight range, in addition to nanoparticles, was investigated in detail. The impact of mucus in Calu-3 cells on macromolecular permeability was also studied, as were other cellular features that give rise to the barrier properties of the epithelium. 2. Materials and methods 2.1. Materials See Supplementary information. 2.2. Cell culture Calu-3 and Caco-2 cells were cultured using EMEM. Cells were seeded on Transwell Ò permeable supports at 10 5 –2 10 5 cells/ 0006-291X/$ - see front matter Ó 2011 Elsevier Inc. All rights reserved. doi:10.1016/j.bbrc.2011.10.108 ⇑ Corresponding author. Fax: +44 115 9515102. E-mail addresses: [email protected] (D. Vllasaliu), paxrf1@not tingham.ac.uk (R. Fowler), [email protected] (M. Garnett), snow. [email protected] (S. Stolnik). 1 These authors contributed equally to this work. Biochemical and Biophysical Research Communications 415 (2011) 579–585 Contents lists available at SciVerse ScienceDirect Biochemical and Biophysical Research Communications journal homepage: www.elsevier.com/locate/ybbrc
-
Upload
bridgehead -
Category
Documents
-
view
5 -
download
0
Transcript of Barrier characteristics of epithelial cultures modelling the airway and intestinal mucosa: A...

Biochemical and Biophysical Research Communications 415 (2011) 579–585
Contents lists available at SciVerse ScienceDirect
Biochemical and Biophysical Research Communications
journal homepage: www.elsevier .com/locate /ybbrc
Barrier characteristics of epithelial cultures modelling the airway andintestinal mucosa: A comparison
Driton Vllasaliu a,⇑,1, Robyn Fowler a,1, Martin Garnett a, Mike Eaton b, Snow Stolnik a
a Division of Drug Delivery and Tissue Engineering, School of Pharmacy, Boots Science Building, University of Nottingham, University Park, Nottingham NG7 2RD, UKb European Technology Platform for Nanomedicine Secretariat, VDI/VDE Innovation + Technik GmbH, Department Innovation Europe, Steinplatz 1, 10623 Berlin, Germany
a r t i c l e i n f o a b s t r a c t
Article history:Received 6 October 2011Available online 2 November 2011
Keywords:Calu-3Caco-2Tight junctionsEpithelial barrierMacromolecular permeabilityMucosal delivery
0006-291X/$ - see front matter � 2011 Elsevier Inc. Adoi:10.1016/j.bbrc.2011.10.108
⇑ Corresponding author. Fax: +44 115 9515102.E-mail addresses: [email protected].
tingham.ac.uk (R. Fowler), martin.garnett@[email protected] (S. Stolnik).
1 These authors contributed equally to this work.
The barrier characteristics of polarized layers of Calu-3 and Caco-2 cell lines, as commonly used in vitromodels of intestinal and airway mucosa, respectively, were investigated by assessing the translocation ofmodel macromolecules and nanoparticles. The barrier capacity of the cell layers towards the movementof macromolecules and nanoparticulates differed considerably between the cell lines. Permeability stud-ies revealed the existence of a notably larger solute molecular weight limit for paracellular diffusion inCaco-2 monolayers compared to Calu-3 cells. Removal of mucus in Calu-3 cells resulted in cell layersexhibiting a larger macromolecular permeability, in addition to improved nanoparticle translocation.Microscopic examination of the tight junctions, as cellular features that play a major role in preventingtransepithelial movement of macromolecules, revealed that the appearance of cell–cell boundaries wasnotably different in the two cell lines, which could explain the differences in macromolecular permeabil-ity. The data overall showed that epithelial layers of airway Calu-3 and intestinal Caco-2 cell culturesin vitro exhibit a different level of restrictiveness and this is due to the cell morphology and the presenceof mucus.
� 2011 Elsevier Inc. All rights reserved.
1. Introduction
The use of in vitro methods for investigation of mucosal drugabsorption is commonplace in the field of drug delivery. A wealthof knowledge, especially mechanistic information, has been gainedfrom these studies. In vitro investigations on mucosal drug absorp-tion often employ appropriately selected and cultured epithelialcell lines, providing models of mucosal tissue of interest. In this re-spect, the intestinal Caco-2 cell line is probably most extensivelyused to study intestinal drug absorption. The excellent in vitro–invivo correlation observed with the Caco-2 cell line [1] has enabledits use as a tool not only to study mechanisms but also predictin vivo oral drug absorption. The bronchially derived Calu-3 cellline, on the other hand, is commonly used as a model of the upperairways in studies of drug absorption across the upper lung and thenose [2,3]. This cell line has also demonstrated good in vitro–in vivocorrelation [4] and possesses the capacity to produce mucus [5],which is advantageous in studying the effect of mucus on drugabsorption.
ll rights reserved.
uk (D. Vllasaliu), [email protected] (M. Garnett), snow.
Previous studies have reported the existence of differences inbarrier characteristics between Calu-3 and Caco-2 cell lines, inaddition to apparent morphological variations, which are expectedconsidering their different mucosal origin. However, it is not en-tirely clear from the literature what cellular features give rise tothis disparity in the capacity to act as a barrier to macromoleculartranslocation. Our work sought to clarify this by investigating therestrictiveness of polarized Calu-3 and Caco-2 cell layers. The bar-rier capacity of the cell layers towards the permeability of modelmacromolecules and proteins over a wide molecular weight range,in addition to nanoparticles, was investigated in detail. The impactof mucus in Calu-3 cells on macromolecular permeability was alsostudied, as were other cellular features that give rise to the barrierproperties of the epithelium.
2. Materials and methods
2.1. Materials
See Supplementary information.
2.2. Cell culture
Calu-3 and Caco-2 cells were cultured using EMEM. Cells wereseeded on Transwell� permeable supports at 105–2 � 105 cells/

580 D. Vllasaliu et al. / Biochemical and Biophysical Research Communications 415 (2011) 579–585
cm2. Calu-3 cells were typically cultured using air-interfaced cul-ture (AIC), created on day two post-seeding. In some instances,which will be clearly specified, Calu-3 cells were also grown usingliquid-covered culture (LCC). Culture medium was replaced everyother day. Calu-3 and Caco-2 cells were typically cultured on per-meable supports for 9–14 and 21–23 days, respectively, prior totheir use as polarized cell layers. Cell layer integrity was routinelyscreened by measuring the transepithelial electrical resistance(TEER); only those displaying TEER P500 X cm2 were deemedappropriate for further use.
2.3. Cell barrier characterisation
2.3.1. Dextran permeabilityFITC-dextrans (FD) of varying molar mass were used as model
macromolecules. The permeability experiments were conductedin an identical way for both cell lines. Cells were initially equili-brated in HBSS for �45 min. HBSS was then replaced with FD4,FD10, FD20, FD40, FD70 or FD150 in HBSS (37 �C; 500 lg/ml). Cellswere placed in the incubator at 37 �C in-between sampling. FD per-meability was determined by regularly sampling the basolateralsolution (sampled volumes were replaced) and quantitation byfluorescence.
Solute permeability was expressed as apparent permeabilitycoefficient (Papp), calculated using the equation detailed in Supple-mentary information.
2.3.2. Permeability at 4 �CCalu-3 (cultured using LCC) and Caco-2 cell layers were incu-
bated with HBSS at 4 �C for �45 min. HBSS was then replaced withsolutions of FD4, FD10, FD20, FD40, FD70 and FD150 (in HBSS, 4 �C,500 lg/ml). Solute permeability was determined as detailed in theprevious section, with cells placed at 4 �C between samplingintervals.
2.3.3. Permeability enhancementCalu-3 and Caco-2 cell layers were equilibrated in HBSS. EDTA
(0.125% w/v) was then applied to the cells in combination withthe tested macromolecular solutes, FD20 or FD40 (500 lg/ml). FDpermeability was determined as before. The control experimentwas conducted by determining the permeability of FD20 andFD40 in the absence of EDTA.
2.3.4. Translocation of proteins across the cellsFITC-insulin and IgG were used as examples of proteins of mod-
est and large molecular weights (Mw � 5.8 and �150 kDa, respec-tively). These were applied to the cell layers in HBSS (pH 6.0) at80 lg/ml (FITC-insulin) and 1 lg/ml (IgG). Transport across thecells was determined by periodically sampling the basolateralsolution. FITC-insulin was quantified by fluorescence, and IgG byELISA (see Supplementary information).
2.4. Cell morphology
Transmission electron microscopy (TEM), scanning electronmicroscopy (SEM), immunostaining for mucins (MUC-5A andMUC-1). See Supplementary information.
2.5. Effect of mucus on permeability
Calu-3 cells were cultured using AIC or LCC for 10 days. Cellswere either equilibrated in HBSS for 1 h or incubated with NAC(0.3% w/v in HBSS) in 3 � 20 min intervals. FD4, FD10, FD40 orFD70 (500 lg/ml in HBSS) were then applied to the cell layersand permeability determined as before.
2.6. Nanoparticle translocation across the cell layers and effect ofmucus
Calu-3 and Caco-2 layers were cultured as described previously,with both AIC and LCC conditions employed for Calu-3 cells. Cellswere equilibrated in HBSS, followed by the application of fluores-cent polystyrene 50 nm nanoparticles (diluted 1:100 in HBSS). Api-cal-to-basolateral translocation was measured by periodicsampling of the basolateral solution. To measure the effect of mu-cus on nanoparticle transport, Calu-3 layers (cultured using AIC)were incubated with NAC (0.3% w/v in HBSS) in 3 � 20 min inter-vals to remove mucus prior to nanoparticle suspension application.Nanoparticles were quantified by fluorescence.
2.7. Statistical analysis
See Supplementary information.
3. Results and discussion
In drug delivery research the use of in vitro models of mucosaltissues to investigate drug absorption (and toxicity) is widely prac-ticed. These models are generally based on epithelial cells culturedas polarized layers on permeable supports. In this respect, the useof the Caco-2 and Calu-3 cell lines is popular in studies investigat-ing therapeutic absorption across the intestines and the airways,respectively. The importance and credibility of the data obtainedfrom drug transport experiments on epithelial cell monolayers ishighlighted by the fact that both the FDA and the European Medi-cines Agency [6] recommend the use of monolayers of suitable epi-thelial cells for classifying the permeability of drug compounds.
Studies comparing the biotherapeutic absorption potential ofthe airways with that of the intestines are surprisingly sparse.Studies investigating absorption of SK&F 110679 (a hexapeptidethat elicits growth hormone release) [7] and cAng-(1–7) (an angio-tensin-(1–7) analogue) [8] in animal models suggest a superiorabsorption of these peptides from the lung compared to that fromthe gastrointestinal tract. The in vitro experience with Calu-3 andCaco-2 cell lines seems to demonstrate the opposite phenomenon[9], although studies providing a direct comparison between thesetwo cell lines are limited. It must be noted that in vitro studiesbased on epithelial cell line models cannot accurately predictin vivo absorption and bioavailability as these models do not ac-count for many absorption-limiting factors associated with themucosal surfaces in vivo. Therefore our work does not seek to com-pare the potential capacity of the airway and intestinal mucosa formacromolecular (and nanoparticulate) absorption. Instead, we fo-cused on investigating the barrier properties of Calu-3 and Caco-2 cells as commonly used models of the airways and the intestinalepithelium, respectively. Morphological assessment of the cellcomponents giving rise to the barrier property of the epithelium,as well as macromolecular permeability was extensively studiedin the cell layers of both cell lines. Furthermore, the presence orabsence of mucus in Calu-3 and Caco-2 cell layers, respectively, en-abled us to evaluate the contribution of mucus in hindering trans-epithelial movement of macromolecules and nanoparticulates.
3.1. Cell barrier characterisation
Initial work characterising the barrier capacity of the Calu-3 andCaco-2 layers compared the permeabilities of FITC-dextrans of dif-ferent molecular weights. Up to a certain molecular mass, thesemacromolecules preferentially traverse the epithelium via theparacellular route, with active transport processes such as pinocy-tosis involved to a lesser extent. Transport across the cell layers is

D. Vllasaliu et al. / Biochemical and Biophysical Research Communications 415 (2011) 579–585 581
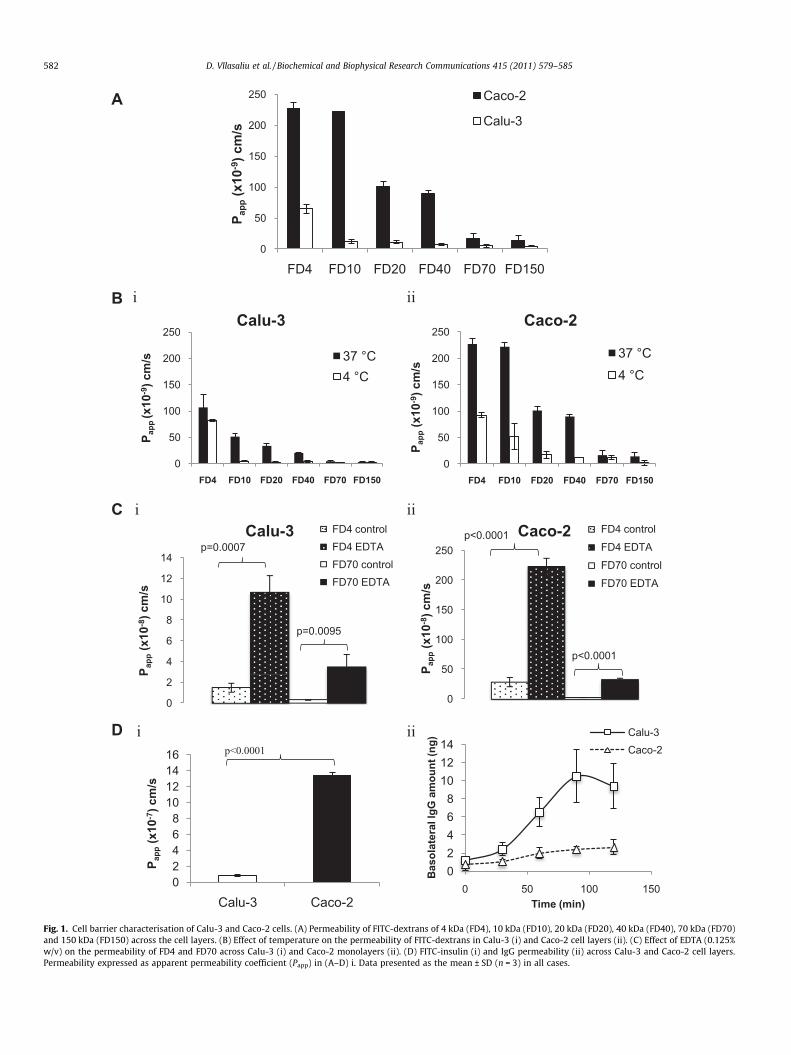
therefore expected to drop as solute exceeds a molecular size thatcan be accommodated by the paracellular corridor. Indeed, Fig. 1Ashows a pattern of decreased permeability with increasing solutemolecular mass from 4 kDa (FD4) to 150 kDa (FD150) in both celllines, which is in agreement with a previous study [10]. In this re-spect, Papp decreased from 227 � 10�9 to 14.1 � 10�9 cm/s in Caco-2 monolayers and from 65 � 10�9 to 4.17 � 10�9 cm/s in Calu-3layers. Importantly, FD permeability across the Calu-3 layers wasconsiderably lower compared to that across Caco-2 monolayers,the difference being especially prominent with FDs in the middleof the tested molecular mass range (up to 19-fold for FD10). It isalso interesting to note that the difference in permeability betweenFD4 (4 kDa) and FD10 (10 kDa) was greater than 2-fold in Calu-3layers, whilst in Caco-2 monolayers this difference was not statis-tically significant, suggesting that the paracellular route in Caco-2monolayers displays a larger solute molecular mass cut-off com-pared to Calu-3 layers. This is particularly noteworthy consideringthat Caco-2 monolayers displayed a markedly higher transepithe-lial electrical resistance (TEER), compared to Calu-3 cell layers(Supplementary information, Fig. S1).
This was further confirmed in the experiment assessing FD per-meability at 4 �C, whereby energy-requiring transport processesare inhibited and any observed permeability predominantly occurspassively, therefore enabling determination of the contribution ofthe paracellular route in the overall transport. FD permeabilitywas dramatically reduced at 4 �C, as compared to that at 37 �C(Fig. 1B), a finding that is in agreement with a previously-reportedstudy [11]. Comparing FD10 and FD20 permeabilities, the data re-veals a larger extent of permeability suppression in Calu-3 (11- and13-fold, respectively), compared to Caco-2 cell layers (4.3- and 6-fold, respectively). Our results hence indicate that the contributionof the paracellular route in Caco-2 monolayers extends to largermolecular mass solutes compared to Calu-3 layers and are inagreement with a study by Kamath et al., which utilised Calu-3and Caco-2 cell lines as cells with ‘tight’ and ‘loose’ tight junctions[9].
Treatment of cells with EDTA, as an absorption enhancer exert-ing its action by loosening the tight junctions, induced a dramaticincrease in the permeability of FD4 and FD70 compared to controlsin both Calu-3 (Fig. 1Ci) and Caco-2 (Fig. 1Cii) cell layers. The ex-tent of enhancement in dextran permeability was similar in Calu-3 and Caco-2 cell layers, amounting to 7.2- and 7.8-fold, respec-tively, for FD4, and 10.9- and 11.7-fold, respectively, for FD70.The data therefore shows that the tight junctions of both Calu-3and Caco-2 cell lines respond in a similar way to EDTA, despitetheir different baseline permeabilities.
The pattern of larger permeability across Caco-2 compared toCalu-3 cell layers was also noted for FITC-insulin (Fig. 2D), whichwe employed to establish whether the permeability trendsobserved with FDs are replicated with a therapeutic macromole-cule. Interestingly, FITC-insulin exhibited a larger permeabilityacross Calu-3 and Caco-2 cell layers (Papp 0.8 � 10�7 and13.4 � 10�7 cm/s, respectively) relative to FD4 (Papp 0.65 � 10�7
and 2.27 � 10�7, respectively), possessing a comparable molecularmass (4 kDa). Comparison of molecular sizes in solution betweeninsulin and FD4 is, however, problematic, given the globular struc-ture of insulin (as opposed to linear geometry of FD4) and its ten-dency to exist in aqueous environment as dimers, tetramers, andhexamers in equilibrium [12]. IgG followed the opposite trend ofsignificantly higher translocation across Calu-3 compared toCaco-2 cell layers, with maximal basolateral amounts reaching10.5 and 2.7 ng (significantly different, p = 0.011), respectively.While both paracellular and transcellular routes are expected tocontribute towards the overall transport of insulin across the celllayers, the considerably larger molar mass of IgG (�150 kDa) limitsits transport to the transcellular route only. In fact, IgG crosses the
epithelial barriers by transcytosis via the FcRn receptor [13,14], theexpression of which has been shown in Calu-3 [15] and Caco-2cells [16–18]. Our results therefore demonstrate that even thoughCaco-2 monolayers display a less restrictive paracellular barriercompared to Calu-3 layers, the extent of receptor-mediated trans-cellular transport of IgG appears to be notably inferior in theformer.
3.2. Cell morphology
Imaging of Calu-3 cells by electron microscopy demonstratedthe presence of apically located electron-translucent granules (ar-row, Fig. 2Ai). Having a diameter of approximately 1 lm theseround granules resemble mucin granules of airway goblet cells inorganization and size [19]. The expression of such structures inCalu-3 cells was also reported by other groups [5,20,21]. The mu-cus-producing capacity of Calu-3 cells was also confirmed byimmunostaining for two mucins, membrane-bound MUC-1, andsecretory MUC-5A (Fig. 2Bi and ii, respectively). The presence ofthese proteins was prominent and specific to the apical side ofthe cells, which is in agreement with a previous study by Kredaet al. [21].
SEM revealed that Calu-3 cells form a tightly packed layer ofcells following their culture on permeable supports (Fig. 2Ci). Fur-thermore the presence of microvilli can be observed from the fig-ure. The micrographs of Caco-2 cells revealed distinct cell–cellboundaries, with apparent apical surface microvilli (Fig. 2Cii).Interestingly, the undulations of the cell boundaries (arrows), canalso clearly be seen, highlighting the difference in morphology toCalu-3 cell layers, which comprise distinctly linear points of cell–cell contacts.
Investigation of the cell–cell boundaries in the cell layers, as de-picted by staining for ZO-1 tight junction protein also providedinteresting findings. ZO-1 distribution appeared as continuous‘belts’ at the points of cell–cell contact in both cell lines. However,whilst its pattern in Calu-3 cells was largely linear (Fig. 2Di), inCaco-2 cells a distinctly convoluted arrangement was clearly evi-dent (Fig. 2Dii). This observation, which is in agreement withSEM data, is important, as the obvious consequence of such organi-sation is a large surface area of the paracellular space in Caco-2cells. The larger total length of the paracellular space per area inCaco-2 cells would provide a larger capacity for the diffusion ofparacellular solutes, which may account for the observed differ-ence in permeability characteristics compared to Calu-3 cells.Interestingly, similar ZO-1 organisation in Caco-2 cells was also ob-served by Man et al. [22], Lindmark et al. [23] and Dickman et al.[24] though the authors of these studies did not discuss the impor-tance of this observation.
3.3. Effect of mucus on permeability
To determine the impact of mucus on macromolecular perme-ability we manipulated the culture conditions in Calu-3 cells toeither promote (AIC) or limit (LCC) the presence of mucus [5], inaddition to employing N-acetyl cysteine (NAC) to remove mucusprior to assessing macromolecular permeability. To confirm suc-cessful mucus removal by NAC, Calu-3 layers were imaged follow-ing their treatment with this material whereby considerably lowermucus staining was apparent compared to non-treated cells (Sup-porting material, Fig. S2). FD permeability was found to be 3.1-,3.9-, 4.4- and 1.6-fold lower across Calu-3 layers cultured usingAIC compared to LCC cells for FD4, FD10, FD40 and FD70, respec-tively (Table 1), suggesting that the presence of mucus in AIC cellsmay impede FD permeability. This observation is interesting, par-ticularly when considering the modest molecular mass of someof the tested FDs (e.g. FD4), although a similar phenomenon was
0
50
100
150
200
250
P app
(x10
-8) c
m/s
Caco-2 FD4 controlFD4 EDTAFD70 controlFD70 EDTA
0
2
4
6
8
10
12
14
P app
(x10
- 8) c
m/s
Calu-3 FD4 controlFD4 EDTAFD70 controlFD70 EDTA
0
50
100
150
200
250
FD4 FD10 FD20 FD40 FD70 FD150
P app
(x10
-9) c
m/s 37 °C
4 °C
0
50
100
150
200
250
FD4 FD10 FD20 FD40 FD70 FD150
P app
(x10
-9) c
m/s
37 °C4 °C
A
i ii
C
B
ii
D i ii
02468
101214
0 50 100 150
Bas
olat
eral
IgG
am
ount
(ng)
Time (min)
Calu-3Caco-2
02468
10121416
Calu-3 Caco-2
P app
(x10
- 7) c
m/s
Calu-3
p=0.0007
p=0.0095
p<0.0001
p<0.0001
Caco-2
p<0.0001
0
50
100
150
200
250
FD4 FD10 FD20 FD40 FD70 FD150
P app
(x10
-9) c
m/s
Caco-2
Calu-3
i
Fig. 1. Cell barrier characterisation of Calu-3 and Caco-2 cells. (A) Permeability of FITC-dextrans of 4 kDa (FD4), 10 kDa (FD10), 20 kDa (FD20), 40 kDa (FD40), 70 kDa (FD70)and 150 kDa (FD150) across the cell layers. (B) Effect of temperature on the permeability of FITC-dextrans in Calu-3 (i) and Caco-2 cell layers (ii). (C) Effect of EDTA (0.125%w/v) on the permeability of FD4 and FD70 across Calu-3 (i) and Caco-2 monolayers (ii). (D) FITC-insulin (i) and IgG permeability (ii) across Calu-3 and Caco-2 cell layers.Permeability expressed as apparent permeability coefficient (Papp) in (A–D) i. Data presented as the mean ± SD (n = 3) in all cases.
582 D. Vllasaliu et al. / Biochemical and Biophysical Research Communications 415 (2011) 579–585
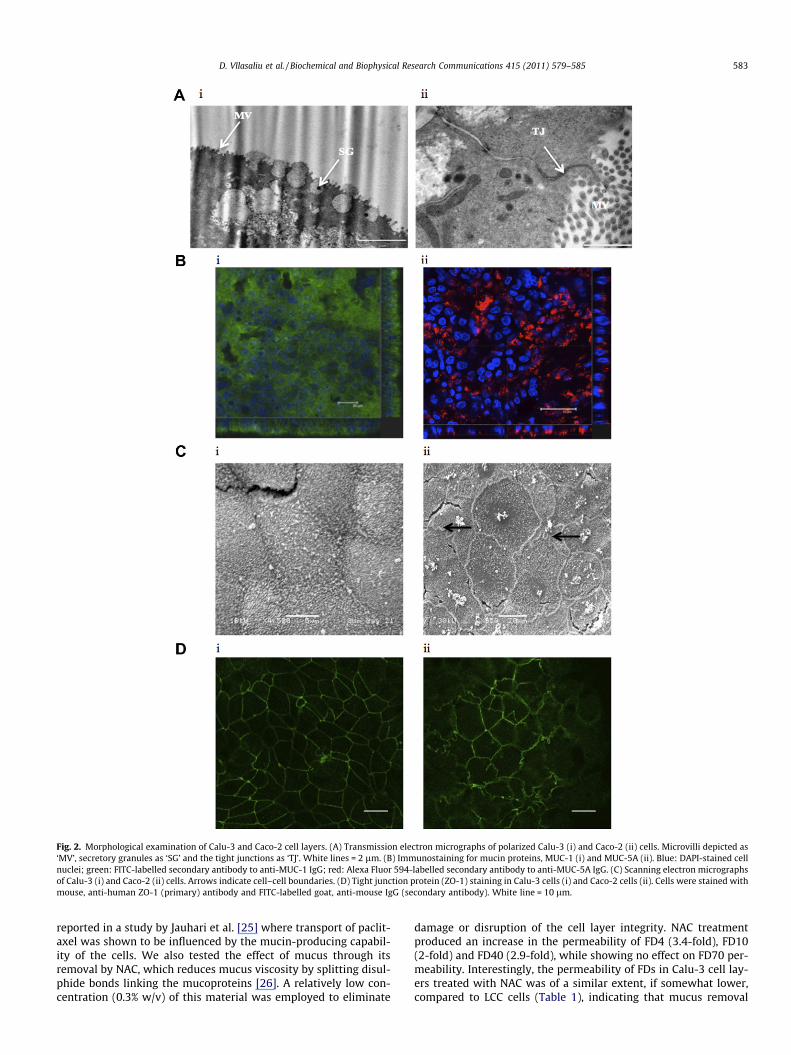
Fig. 2. Morphological examination of Calu-3 and Caco-2 cell layers. (A) Transmission electron micrographs of polarized Calu-3 (i) and Caco-2 (ii) cells. Microvilli depicted as‘MV’, secretory granules as ‘SG’ and the tight junctions as ‘TJ’. White lines = 2 lm. (B) Immunostaining for mucin proteins, MUC-1 (i) and MUC-5A (ii). Blue: DAPI-stained cellnuclei; green: FITC-labelled secondary antibody to anti-MUC-1 IgG; red: Alexa Fluor 594-labelled secondary antibody to anti-MUC-5A IgG. (C) Scanning electron micrographsof Calu-3 (i) and Caco-2 (ii) cells. Arrows indicate cell–cell boundaries. (D) Tight junction protein (ZO-1) staining in Calu-3 cells (i) and Caco-2 cells (ii). Cells were stained withmouse, anti-human ZO-1 (primary) antibody and FITC-labelled goat, anti-mouse IgG (secondary antibody). White line = 10 lm.
D. Vllasaliu et al. / Biochemical and Biophysical Research Communications 415 (2011) 579–585 583
reported in a study by Jauhari et al. [25] where transport of paclit-axel was shown to be influenced by the mucin-producing capabil-ity of the cells. We also tested the effect of mucus through itsremoval by NAC, which reduces mucus viscosity by splitting disul-phide bonds linking the mucoproteins [26]. A relatively low con-centration (0.3% w/v) of this material was employed to eliminate
damage or disruption of the cell layer integrity. NAC treatmentproduced an increase in the permeability of FD4 (3.4-fold), FD10(2-fold) and FD40 (2.9-fold), while showing no effect on FD70 per-meability. Interestingly, the permeability of FDs in Calu-3 cell lay-ers treated with NAC was of a similar extent, if somewhat lower,compared to LCC cells (Table 1), indicating that mucus removal
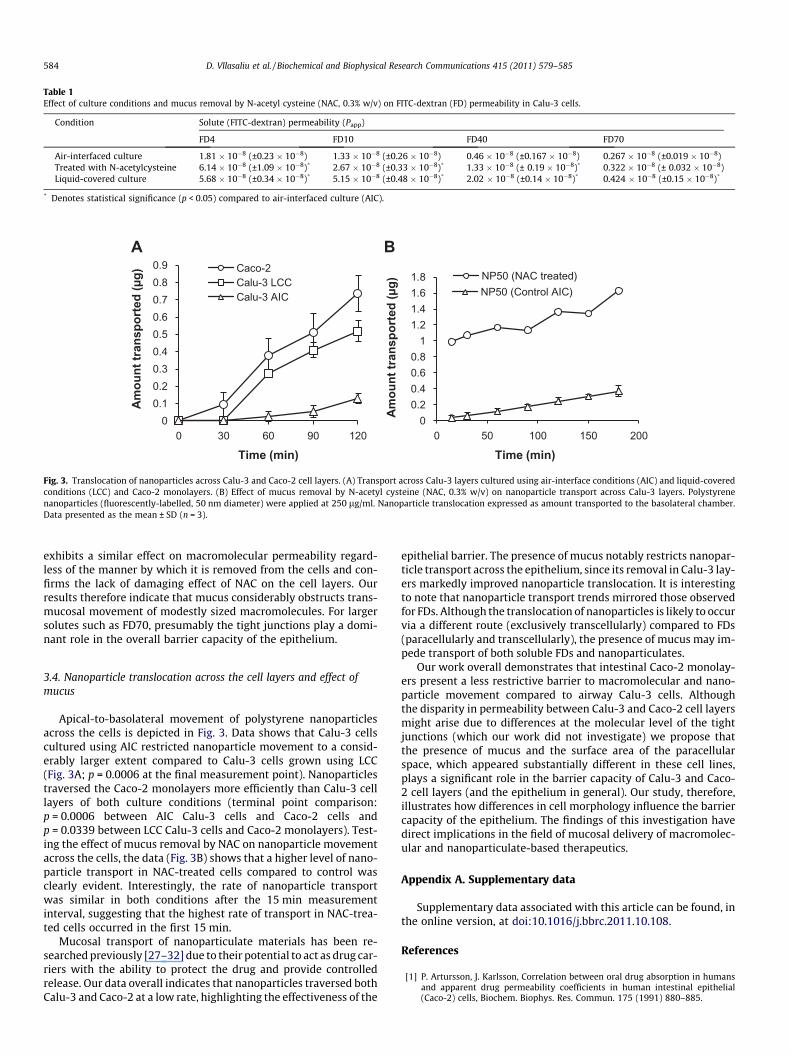
Table 1Effect of culture conditions and mucus removal by N-acetyl cysteine (NAC, 0.3% w/v) on FITC-dextran (FD) permeability in Calu-3 cells.
Condition Solute (FITC-dextran) permeability (Papp)
FD4 FD10 FD40 FD70
Air-interfaced culture 1.81 � 10�8 (±0.23 � 10�8) 1.33 � 10�8 (±0.26 � 10�8) 0.46 � 10�8 (±0.167 � 10�8) 0.267 � 10�8 (±0.019 � 10�8)Treated with N-acetylcysteine 6.14 � 10�8 (±1.09 � 10�8)* 2.67 � 10�8 (±0.33 � 10�8)* 1.33 � 10�8 (± 0.19 � 10�8)* 0.322 � 10�8 (± 0.032 � 10�8)Liquid-covered culture 5.68 � 10�8 (±0.34 � 10�8)* 5.15 � 10�8 (±0.48 � 10�8)* 2.02 � 10�8 (±0.14 � 10�8)* 0.424 � 10�8 (±0.15 � 10�8)*
* Denotes statistical significance (p < 0.05) compared to air-interfaced culture (AIC).
00.10.20.30.40.50.60.70.80.9
0 30 60 90 120
Amou
nt tr
ansp
orte
d (µ
g)
Time (min)
Caco-2Calu-3 LCCCalu-3 AIC
00.20.40.60.8
11.21.41.61.8
0 50 100 150 200
Amou
nt tr
ansp
orte
d (µ
g)
Time (min)
NP50 (NAC treated)NP50 (Control AIC)
A B
Fig. 3. Translocation of nanoparticles across Calu-3 and Caco-2 cell layers. (A) Transport across Calu-3 layers cultured using air-interface conditions (AIC) and liquid-coveredconditions (LCC) and Caco-2 monolayers. (B) Effect of mucus removal by N-acetyl cysteine (NAC, 0.3% w/v) on nanoparticle transport across Calu-3 layers. Polystyrenenanoparticles (fluorescently-labelled, 50 nm diameter) were applied at 250 lg/ml. Nanoparticle translocation expressed as amount transported to the basolateral chamber.Data presented as the mean ± SD (n = 3).
584 D. Vllasaliu et al. / Biochemical and Biophysical Research Communications 415 (2011) 579–585
exhibits a similar effect on macromolecular permeability regard-less of the manner by which it is removed from the cells and con-firms the lack of damaging effect of NAC on the cell layers. Ourresults therefore indicate that mucus considerably obstructs trans-mucosal movement of modestly sized macromolecules. For largersolutes such as FD70, presumably the tight junctions play a domi-nant role in the overall barrier capacity of the epithelium.
3.4. Nanoparticle translocation across the cell layers and effect ofmucus
Apical-to-basolateral movement of polystyrene nanoparticlesacross the cells is depicted in Fig. 3. Data shows that Calu-3 cellscultured using AIC restricted nanoparticle movement to a consid-erably larger extent compared to Calu-3 cells grown using LCC(Fig. 3A; p = 0.0006 at the final measurement point). Nanoparticlestraversed the Caco-2 monolayers more efficiently than Calu-3 celllayers of both culture conditions (terminal point comparison:p = 0.0006 between AIC Calu-3 cells and Caco-2 cells andp = 0.0339 between LCC Calu-3 cells and Caco-2 monolayers). Test-ing the effect of mucus removal by NAC on nanoparticle movementacross the cells, the data (Fig. 3B) shows that a higher level of nano-particle transport in NAC-treated cells compared to control wasclearly evident. Interestingly, the rate of nanoparticle transportwas similar in both conditions after the 15 min measurementinterval, suggesting that the highest rate of transport in NAC-trea-ted cells occurred in the first 15 min.
Mucosal transport of nanoparticulate materials has been re-searched previously [27–32] due to their potential to act as drug car-riers with the ability to protect the drug and provide controlledrelease. Our data overall indicates that nanoparticles traversed bothCalu-3 and Caco-2 at a low rate, highlighting the effectiveness of the
epithelial barrier. The presence of mucus notably restricts nanopar-ticle transport across the epithelium, since its removal in Calu-3 lay-ers markedly improved nanoparticle translocation. It is interestingto note that nanoparticle transport trends mirrored those observedfor FDs. Although the translocation of nanoparticles is likely to occurvia a different route (exclusively transcellularly) compared to FDs(paracellularly and transcellularly), the presence of mucus may im-pede transport of both soluble FDs and nanoparticulates.
Our work overall demonstrates that intestinal Caco-2 monolay-ers present a less restrictive barrier to macromolecular and nano-particle movement compared to airway Calu-3 cells. Althoughthe disparity in permeability between Calu-3 and Caco-2 cell layersmight arise due to differences at the molecular level of the tightjunctions (which our work did not investigate) we propose thatthe presence of mucus and the surface area of the paracellularspace, which appeared substantially different in these cell lines,plays a significant role in the barrier capacity of Calu-3 and Caco-2 cell layers (and the epithelium in general). Our study, therefore,illustrates how differences in cell morphology influence the barriercapacity of the epithelium. The findings of this investigation havedirect implications in the field of mucosal delivery of macromolec-ular and nanoparticulate-based therapeutics.
Appendix A. Supplementary data
Supplementary data associated with this article can be found, inthe online version, at doi:10.1016/j.bbrc.2011.10.108.
References
[1] P. Artursson, J. Karlsson, Correlation between oral drug absorption in humansand apparent drug permeability coefficients in human intestinal epithelial(Caco-2) cells, Biochem. Biophys. Res. Commun. 175 (1991) 880–885.

D. Vllasaliu et al. / Biochemical and Biophysical Research Communications 415 (2011) 579–585 585
[2] M. Amoako-Tuffour, P.K. Yeung, R.U. Agu, Permeation of losartan across humanrespiratory epithelium: an in vitro study with Calu-3 cells, Acta Pharm. 59(2009) 395–405.
[3] L. Li, N.R. Mathias, C.L. Heran, P. Moench, D.A. Wall, R.L. Smith, Carbopol-mediated paracellular transport enhancement in Calu-3 cell layers, J. Pharm.Sci. 95 (2006) 326–335.
[4] M. Sakagami, In vivo, in vitro and ex vivo models to assess pulmonaryabsorption and disposition of inhaled therapeutics for systemic delivery, Adv.Drug Deliv. Rev. 58 (2006) 1030–1060.
[5] C.I. Grainger, L.L. Greenwell, D.J. Lockley, G.P. Martin, B. Forbes, Culture of Calu-3 cells at the air interface provides a representative model of the airwayepithelial barrier, Pharm. Res. 23 (2006) 1482–1490.
[6] Bock. Validation of the Caco-2 cell monolayer system for determining thepermeability of drug substances according to the BiopharmaceuticsClassification System (BCS). <http://www.acrossbarriers.de/uploads/media/FCT02-I-0305_BCS.pdf>.
[7] P.L. Smith, S.E. Yeulet, D.R. Citerone, F. Drake, M. Cook, D.A. Wall, J. Marcello, SK& F 110679: comparison of absorption following oral or respiratoryadministration, J. Controlled Release 28 (1994) 67–77.
[8] L. de Vries, C.E. Reitzema-Klein, A. Meter-Arkema, A. van Dam, R. Rink, G.N.Moll, M.H. Akanbi, Oral and pulmonary delivery of thioether-bridgedangiotensin-(1–7), Peptides 31 893–8.
[9] A.V. Kamath, R.A. Morrison, N.R. Mathias, S.A. Dando, A.M. Marino, S. Chong,Modulation of tight junctions does not predict oral absorption of hydrophiliccompounds: use of Caco-2 and Calu-3 cells, Arch. Pharm. Res. 30 (2007) 1002–1007.
[10] B.I. Florea, M.L. Cassara, H.E. Junginger, G. Borchard, Drug transport andmetabolism characteristics of the human airway epithelial cell line Calu-3, J.Controlled Release 87 (2003) 131–138.
[11] Y. Matsukawa, V.H. Lee, E.D. Crandall, K.J. Kim, Size-dependent dextrantransport across rat alveolar epithelial cell monolayers, J. Pharm. Sci. 86 (1997)305–309.
[12] C.M. Yu, S. Mun, N.H. Wang, Phenomena of insulin peak fronting in sizeexclusion chromatography and strategies to reduce fronting, J. Chromatogr. A1192 (2008) 121–129.
[13] N.E. Simister, A.R. Rees, Isolation and characterization of an Fc receptor fromneonatal rat small intestine, Eur. J. Immunol. 15 (1985) 733–738.
[14] N.E. Simister, K.E. Mostov, An Fc receptor structurally related to MHC class Iantigens, Nature 337 (1989) 184–187.
[15] G.M. Spiekermann, P.W. Finn, E.S. Ward, J. Dumont, B.L. Dickinson, R.S.Blumberg, W.I. Lencer, Receptor-mediated immunoglobulin G transport acrossmucosal barriers in adult life: functional expression of FcRn in the mammalianlung, J. Exp. Med. 196 (2002) 303–310.
[16] S.M. Claypool, B.L. Dickinson, J.S. Wagner, F.E. Johansen, N. Venu, J.A. Borawski,W.I. Lencer, R.S. Blumberg, Bidirectional transepithelial IgG transport by astrongly polarized basolateral membrane Fcgamma-receptor, Mol. Biol. Cell 15(2004) 1746–1759.
[17] B.L. Dickinson, K. Badizadegan, Z. Wu, J.C. Ahouse, X. Zhu, N.E. Simister, R.S.Blumberg, W.I. Lencer, Bidirectional FcRn-dependent IgG transport in apolarized human intestinal epithelial cell line, J. Clin. Invest. 104 (1999)903–911.
[18] K. Sato, J. Nagai, N. Mitsui, Y. Ryoko, M. Takano, Effects of endocytosisinhibitors on internalization of human IgG by Caco-2 human intestinalepithelial cells, Life Sci. 85 (2009) 800–807.
[19] C.W. Davis, S.H. Randell, Airway goblet and mucous cells: identical, similar, ordifferent? in: S.M. (Ed.), Cilia and Mucus: from Development to RespiratoryDefense, Marcel Dekker, pp. 195–210.
[20] B.Q. Shen, W.E. Finkbeiner, J.J. Wine, R.J. Mrsny, J.H. Widdicombe, Calu-3: ahuman airway epithelial cell line that shows cAMP-dependent Cl-secretion,Am. J. Physiol. 266 (1994) L493–L501.
[21] S.M. Kreda, S.F. Okada, C.A. van Heusden, W. O’Neal, S. Gabriel, L. Abdullah,C.W. Davis, R.C. Boucher, E.R. Lazarowski, Coordinated release of nucleotidesand mucin from human airway epithelial Calu-3 cells, J. Physiol. 584 (2007)245–259.
[22] A.L. Man, F. Lodi, E. Bertelli, M. Regoli, C. Pin, F. Mulholland, A.R. Satoskar, M.J.Taussig, C. Nicoletti, Macrophage migration inhibitory factor plays a role in theregulation of microfold (M) cell-mediated transport in the gut, J. Immunol. 181(2008) 5673–5680.
[23] T. Lindmark, Y. Kimura, P. Artursson, Absorption enhancement throughintracellular regulation of tight junction permeability by medium chain fattyacids in Caco-2 cells, J. Pharmacol. Exp. Ther. 284 (1998) 362–369.
[24] K.G. Dickman, S.J. Hempson, J. Anderson, S. Lippe, L. Zhao, R. Burakoff, R.D.Shaw, Rotavirus alters paracellular permeability and energy metabolism inCaco-2 cells, Am. J. Physiol. Gastrointest. Liver Physiol. 279 (2000) G757–G766.
[25] S. Jauhari, A.K. Dash, A mucoadhesive in situ gel delivery system for paclitaxel,AAPS Pharm. Sci. Tech. 7 (2006) E53.
[26] F. Sun, S. Tai, T. Lim, U. Baumann, M. King, Additive effect of dornase alfa andNacystelyn on transportability and viscoelasticity of cystic fibrosis sputum,Can. Respir. J. 9 (2002) 401–406.
[27] G.P. Carino, J.S. Jacob, E. Mathiowitz, Nanosphere based oral insulin delivery, J.Controlled Release 65 (2000) 261–269.
[28] P. Jani, G.W. Halbert, J. Langridge, A.T. Florence, The uptake and translocationof latex nanospheres and microspheres after oral administration to rats, J.Pharm. Pharmacol. 41 (1989) 809–812.
[29] T. Jung, W. Kamm, A. Breitenbach, E. Kaiserling, J.X. Xiao, T. Kissel,Biodegradable nanoparticles for oral delivery of peptides: is there a role forpolymers to affect mucosal uptake?, Eur J. Pharm. Biopharm. 50 (2000) 147–160.
[30] A. Vila, A. Sanchez, M. Tobio, P. Calvo, M.J. Alonso, Design of biodegradableparticles for protein delivery, J. Controlled Release 78 (2002) 15–24.
[31] J. Brooking, S.S. Davis, L. Illum, Transport of nanoparticles across the rat nasalmucosa, J. Drug Target. 9 (2001) 267–279.
[32] M. de la Fuente, N. Csaba, M. Garcia-Fuentes, M.J. Alonso, Nanoparticles asprotein and gene carriers to mucosal surfaces, Nanomedicine (Lond.) 3 (2008)845–857.




















