
Effect of body composition methodology on heritability estimation of body fatness
Upload
independentCategory
view
0download
0
www.elsevier.com/locate/meatsci
Meat Science 73 (2006) 42–47
MEATSCIENCE
Association of MC4R gene variants with growth, fatness,carcass composition and meat and fat quality traits in heavy pigs
C. Ovilo a,*, A. Fernandez a, M.C. Rodrıguez a, M. Nieto b, L. Silio a
a Departamento de Mejora Genetica Animal, INIA, Ctra Coruna Km 7.5, 28040 Madrid, Spainb COPESE, Conde de Sepulveda 24, 40300 Sepulveda, Spain
Received 30 May 2005; received in revised form 19 September 2005; accepted 25 October 2005
Abstract
The melanocortin receptor 4 (MC4R) gene is implicated in the regulation of feeding behaviour and body weight in humans and mice.A missense mutation (Asp298Asn) located in a highly conserved region of this gene has clearly been associated with backfat depth, feedintake and growth rate in different porcine lines. In this work the complete coding region of the gene was sequenced in samples from sixpigs of a commercial hybrid line and two polymorphisms were detected at positions 709 (C/T) and 1426 (G/A). The last one correspondsto the missense mutation, and has been genotyped in 333 animals with phenotypic records and 68 out of their 81 parents. An associationstudy of these genotypes with several performance and quality traits was performed within the statistical animal model framework. Theresults confirmed the effect of the missense mutation on growth and fat deposition traits (live weight at 140d and backfat depth), andsupported new effects on carcass composition (loin and shoulder weights) and traits related to fat and meat quality (profile of fatty acids;muscle Minolta L*, a* and Ho colour parameters). A transmission-disequilibrium test provided no evidence of spurious association dueto population stratification.� 2005 Elsevier Ltd. All rights reserved.
Keywords: MC4R; Heavy pigs; Carcass composition; Quality traits
1. Introduction
Research on mutations in targeted functional genes(candidate genes) and their association with economictraits is performed to ascertain the genetic basis of produc-tion traits and to develop DNA tests as selection tools inpig breeding schemes (De Vries, Sosnicki, Garnier, & Pla-stow, 1998). Many of these discoveries are being used bythe pig industry in combination with performance informa-tion to improve pig production (Rothschild, 2004). Thisapproach is a very promising route for improvement ofmeat quality, since direct meat quality records are notavailable for potential breeding animals. Some well knownexamples could be the gene tests used to remove HAL orRN mutations with undesirable effects on meat quality
0309-1740/$ - see front matter � 2005 Elsevier Ltd. All rights reserved.
doi:10.1016/j.meatsci.2005.10.016
* Corresponding author. Tel.: +34 91 3471492; fax: +34 91 3572293.E-mail address: [email protected] (C. Ovilo).
(Fujii et al., 1991; Le Roy et al., 2000). These tests haveallowed a clear improvement of the technological qualityof pig meat produced in many countries. New applicationsof genomics would be required to adapt technological andsensory qualities to the requirements of processors andconsumers, in order to deal with growing market segmen-tation (Monin, 2003).
Dry-cured meat products represent a large part of thepig meat products on the European market. Among thecomponents of raw material, lipids in muscle and adiposetissue play a key role in the final quality of these products(Gandemer, 2002). Heavy carcass types are required for theelite industries (Iberian or Parma hams) to guarantee ade-quate weight of cuts, sufficient thickness of subcutaneousfat, and optimal intramuscular fat content and fatty acidcomposition (Lo Fiego, Santoro, Macchioni, & De Leoni-bus, 2005; Lopez-Bote, 1998). Specialized pig lines are nec-essary with breeding goals adapted to these singular

C. Ovilo et al. / Meat Science 73 (2006) 42–47 43
production-marketing sytems, and lower emphasis on effi-ciency of lean deposition. In this context, possibilities ofgene assisted selection should be carefully evaluated.Recent studies reported associations of a missense muta-tion in Calpastatin gene (CAST) with fresh meat textureand water holding capacity (Ciobanu et al., 2004) and withdry-cured ham moisture content (Stalder, Rothschild, &Lonergan, 2005). Moreover, some of the candidate genesassociated with growth and fat deposition in pigs (MC4R
or LEPR) play an important role in the control of energyhomeostasis and fat mobilization (Ovilo et al., 2005;Rothschild, 2004), and their effects on lipid compositionof muscle and fat tissues should be investigated.
The interaction between melanocortins and their recep-tors in the hypothalamus is one of the main neuroendocr-inological pathways controlling energy balance (Wardlaw,2001). In pigs a missense mutation has been described intheMC4R locus (Asp298Asn) that is included in its seventhtransmembrane region, which is a highly conserved regionacross different types of melanocortin receptors and acrossspecies (Kim, Larsen, & Rothschild, 1999). This mutationhas been associated with fatter and faster growing pigs,having significant effects on backfat, days to 110 kg, testdaily gain and daily food intake in different commercialpig lines (Hernandez-Sanchez, Visscher, Plastow, & Haley,2003; Houston, Cameron, & Rance, 2004; Kim, Larsen,Short, Plastow, & Rothschild, 2000; Kim et al., 2004).Functional analyses of this mutation indicate thatAsp298 is required for normal G protein coupling in theMC4R signaling to adenylyl cyclase (Kim, Reecy, Hsu,Anderson, & Rothschild, 2004).
The objectives of this work were: (a) the identification ofthe polymorphisms present in the MC4R gene in a com-mercial hybrid line used to produce heavy pigs for process-ing of dry-cured meat products, and (b) the study of itsassociation with several previously studied productivetraits (growth and fatness), and with other traits relatedto carcass composition and meat and fat quality. A trans-mission disequilibrium test (TDT) was applied to avoidspurious associations as a consequence of population strat-ification (Hernandez-Sanchez et al., 2003).
2. Material and methods
2.1. Animals and phenotypic records
Animals studied were 333 male pigs from a hybrid com-mercial line obtained by the breeder company Gene + Ibe-rica. These pigs proceed from 9 Landrace sires and 72Taihu x Large-White dams. The animals were reared onthe same farm in five different batches from approximately6 kg of live weight up to slaughter, which took place atabout 127 kg with an average age of 198 days. Animalswere fed ad libitum with a diet containing 2090 kcal ofdigestible energy and 175 g of crude protein per kg of drymatter up to 60 kg of live weight, and thereafter with a dietcontaining 2415 kcal of digestible energy and 140 g of
crude protein per kg. Live weights and measures of backfatthickness were recorded at 160 and 187 days of age. Back-fat thickness was measured on live pigs with a Renco LeanMeter ultrasonic equipment, at three locations (shoulder,last rib and ham) on the right and left side of each pig,and the average of the six measurements was used for anal-ysis. Average daily gain was calculated as the weight gaindivided by the number of days elapsed between the twomeasurements. A sample of 83 animals was also weighedat 140 days of age.
After slaughter, individual carcass weight was recorded,and backfat thickness was measured with a Fat-O-Meter atthree different locations (P2, Gluteus medius and Scapula)on the left side of each animal. Measurements of pH at45 min were taken on the semimembranosus muscle usinga HANNA (HI 8424) pH meter. The weight of the pre-mium cuts (trimmed hams, shoulders and one of the loins)were individually recorded.
A sample of Longissimus dorsi (LD) muscle at the levelof the fourth rib was taken from each animal and usedfor the determination of intramuscular fat, protein andwater content, as well as for the instrumental analysis ofcolour. A Meat Analyzer, Infratec 1265 (Tecator, Switzer-land), working in the wavelength range 850–1050 nm bythe Near Infrared Transmission (NIT) principle was usedfor the chemical analysis of intramuscular fat, proteinand water content. The colour parameters (lightness, L*;redness, a*, and yellowness, b*) were determined using aMinolta CM-2002 (Minolta Camera Co., Osaka, Japan)on loin samples bloomed for 24 h at 4 �C. In addition,the parameters chroma (C*) and hue angle (Ho) were calcu-lated from a* and b* measures using the following equa-tions C* = ((a*)2 + (b*)2)1/2 and Ho = arctg(b*/a*).Subcutaneous fat samples were taken from the tail inser-tion area in the coxal region of the carcasses. Determina-tion of the main fatty acids (palmitic C16:0, stearicC18:0, oleic C18:1 and linoleic C18:2) on these sampleswas carried out by gas chromatography using an Autosys-tem XL Perkin Elmer. Fatty acids were separated on silicacapillary column Omega Wax320 (Supelco, Bellafonte, PA,USA) (30 m length; 0.25 mm internal diameter; 0.25 mmfilm thickness). The fatty acid methyl esters (FAME) wereidentified by comparison of each retention time with thoseof standards (Supelco 37 Component FAME mix andPUFA standard no. 2, animal source, Supelco, Bellafonte,PA, USA).
Information of the recorded traits is included in Table 1.
2.2. MC4R gene sequencing and mutation detection
The MC4R gene was sequenced from genomic DNA ofsix pigs obtained from blood samples. Two primer pairswere designed from the porcine sequence available (Gen-bank AB021664) to amplify two overlapping fragmentsof 633 and 666 bp, respectively. The first fragment wasamplified with primers of sequence: forward primer, 5 0-TGG AGA AAA TCG CTG AGG CTA CC-3 0, and
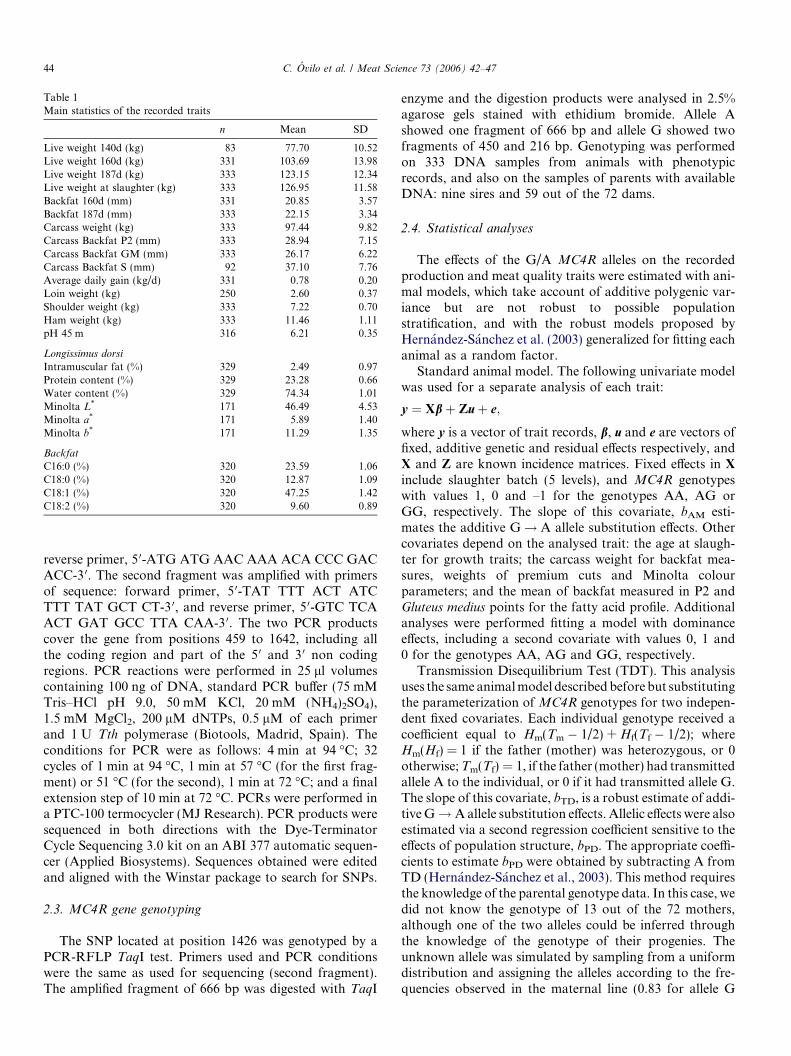
Table 1Main statistics of the recorded traits
n Mean SD
Live weight 140d (kg) 83 77.70 10.52Live weight 160d (kg) 331 103.69 13.98Live weight 187d (kg) 333 123.15 12.34Live weight at slaughter (kg) 333 126.95 11.58Backfat 160d (mm) 331 20.85 3.57Backfat 187d (mm) 333 22.15 3.34Carcass weight (kg) 333 97.44 9.82Carcass Backfat P2 (mm) 333 28.94 7.15Carcass Backfat GM (mm) 333 26.17 6.22Carcass Backfat S (mm) 92 37.10 7.76Average daily gain (kg/d) 331 0.78 0.20Loin weight (kg) 250 2.60 0.37Shoulder weight (kg) 333 7.22 0.70Ham weight (kg) 333 11.46 1.11pH 45 m 316 6.21 0.35
Longissimus dorsi
Intramuscular fat (%) 329 2.49 0.97Protein content (%) 329 23.28 0.66Water content (%) 329 74.34 1.01Minolta L* 171 46.49 4.53Minolta a* 171 5.89 1.40Minolta b* 171 11.29 1.35
Backfat
C16:0 (%) 320 23.59 1.06C18:0 (%) 320 12.87 1.09C18:1 (%) 320 47.25 1.42C18:2 (%) 320 9.60 0.89
44 C. Ovilo et al. / Meat Science 73 (2006) 42–47
reverse primer, 5 0-ATG ATG AAC AAA ACA CCC GACACC-3 0. The second fragment was amplified with primersof sequence: forward primer, 5 0-TAT TTT ACT ATCTTT TAT GCT CT-3 0, and reverse primer, 5 0-GTC TCAACT GAT GCC TTA CAA-3 0. The two PCR productscover the gene from positions 459 to 1642, including allthe coding region and part of the 5 0 and 3 0 non codingregions. PCR reactions were performed in 25 ll volumescontaining 100 ng of DNA, standard PCR buffer (75 mMTris–HCl pH 9.0, 50 mM KCl, 20 mM (NH4)2SO4),1.5 mM MgCl2, 200 lM dNTPs, 0.5 lM of each primerand 1 U Tth polymerase (Biotools, Madrid, Spain). Theconditions for PCR were as follows: 4 min at 94 �C; 32cycles of 1 min at 94 �C, 1 min at 57 �C (for the first frag-ment) or 51 �C (for the second), 1 min at 72 �C; and a finalextension step of 10 min at 72 �C. PCRs were performed ina PTC-100 termocycler (MJ Research). PCR products weresequenced in both directions with the Dye-TerminatorCycle Sequencing 3.0 kit on an ABI 377 automatic sequen-cer (Applied Biosystems). Sequences obtained were editedand aligned with the Winstar package to search for SNPs.
2.3. MC4R gene genotyping
The SNP located at position 1426 was genotyped by aPCR-RFLP TaqI test. Primers used and PCR conditionswere the same as used for sequencing (second fragment).The amplified fragment of 666 bp was digested with TaqI
enzyme and the digestion products were analysed in 2.5%agarose gels stained with ethidium bromide. Allele Ashowed one fragment of 666 bp and allele G showed twofragments of 450 and 216 bp. Genotyping was performedon 333 DNA samples from animals with phenotypicrecords, and also on the samples of parents with availableDNA: nine sires and 59 out of the 72 dams.
2.4. Statistical analyses
The effects of the G/A MC4R alleles on the recordedproduction and meat quality traits were estimated with ani-mal models, which take account of additive polygenic var-iance but are not robust to possible populationstratification, and with the robust models proposed byHernandez-Sanchez et al. (2003) generalized for fitting eachanimal as a random factor.
Standard animal model. The following univariate modelwas used for a separate analysis of each trait:
y ¼ Xbþ Zuþ e;
where y is a vector of trait records, b, u and e are vectors offixed, additive genetic and residual effects respectively, andX and Z are known incidence matrices. Fixed effects in X
include slaughter batch (5 levels), and MC4R genotypeswith values 1, 0 and –1 for the genotypes AA, AG orGG, respectively. The slope of this covariate, bAM esti-mates the additive G ! A allele substitution effects. Othercovariates depend on the analysed trait: the age at slaugh-ter for growth traits; the carcass weight for backfat mea-sures, weights of premium cuts and Minolta colourparameters; and the mean of backfat measured in P2 andGluteus medius points for the fatty acid profile. Additionalanalyses were performed fitting a model with dominanceeffects, including a second covariate with values 0, 1 and0 for the genotypes AA, AG and GG, respectively.
Transmission Disequilibrium Test (TDT). This analysisuses the same animalmodel described before but substitutingthe parameterization of MC4R genotypes for two indepen-dent fixed covariates. Each individual genotype received acoefficient equal to Hm(Tm � 1/2) + Hf(Tf � 1/2); whereHm(Hf) = 1 if the father (mother) was heterozygous, or 0otherwise;Tm(Tf) = 1, if the father (mother) had transmittedallele A to the individual, or 0 if it had transmitted allele G.The slope of this covariate, bTD, is a robust estimate of addi-tiveG ! Aallele substitution effects. Allelic effects were alsoestimated via a second regression coefficient sensitive to theeffects of population structure, bPD. The appropriate coeffi-cients to estimate bPD were obtained by subtracting A fromTD (Hernandez-Sanchez et al., 2003). This method requiresthe knowledge of the parental genotype data. In this case, wedid not know the genotype of 13 out of the 72 mothers,although one of the two alleles could be inferred throughthe knowledge of the genotype of their progenies. Theunknown allele was simulated by sampling from a uniformdistribution and assigning the alleles according to the fre-quencies observed in the maternal line (0.83 for allele G

Table 2Mean effect of G (Asp298)! A (Asn298) allele substitution (b) for theanalyzed traits using standard animal model
b SE(b) t P value
Live weight 140d (kg) 5.451 1.995 2.732 <0.01Live weight 160d (kg) 1.322 0.981 1.348 <0.20Live weight 187d (kg) 1.398 1.057 1.323 <0.20Backfat 160d (mm) 0.374 0.276 1.355 <0.20Backfat 187d (mm) 0.295 0.295 0.998 <0.40Carcass Backfat P2 (mm) 1.464 0.643 2.277 <0.05Carcass Backfat GM (mm) 0.741 0.534 1.388 <0.20Carcass Backfat S (mm) 1.145 1.173 3.534 <0.001Average daily gain (kg/d) 0.004 0.016 0.250 >0.50Loin weight (kg) �0.081 0.025 3.240 <0.005Shoulder weight (kg) �0.105 0.028 3.750 <0.001Ham weight (kg) �0.065 0.054 1.204 <0.40pH 45 m �0.069 0.064 1.078 <0.40
Longissimus dorsi
Intramuscular fat (%) 0.021 0.085 0.247 >0.50Protein content (%) �0.331 0.199 1.663 <0.10Water content (%) �0.909 0.625 1.454 <0.20Minolta L* 1.788 0.552 3.263 <0.005Minolta a* �0.373 0.174 2.138 <0.05Minolta b* �0.297 0.171 1.744 <0.10C* �0.417 0.213 1.958 <0.10Ho 1.338 0.617 2.169 <0.05
Backfat
C16:0 (%) 0.233 0.084 2.774 <0.01C18:0 (%) 0.283 0.090 3.144 <0.005C18:1(%) �0.263 0.118 2.229 <0.05C18:2 (%) �0.236 0.059 4.000 <0.001
t, t-statistic from testing b = 0 vs. b 6¼ 0.
C. Ovilo et al. / Meat Science 73 (2006) 42–47 45
and 0.17 for alleleA).We performed 1000 replicates to createdifferent populations of maternal genotypes and the coeffi-cients bTD and bPDwere estimated for each replicate. The dif-ference between the means of the bTD and bPDestimates wastestedwith a Student�s t-test. A significant difference betweenthese two parameters could be interpreted as an evidence forspurious association. The QxPak software (Perez-Enciso &Misztal, 2004) was used to run the different models to esti-mate variance components and fixed effects.
3. Results
The MC4R gene contains a single exon of approxi-mately 1 kb. A total of 1184 bp were PCR amplified andsequenced from genomic DNA. Multiple alignments ofthe sequences of six individual samples of the same hybridline allowed the identification of two SNPs. The first one isa synonymous C/T substitution at position 709(AB021664). The second is a G/A missense mutation atposition 1426 that replaces aspartic acid (GAU) withasparagine (AAU), and that has been described previously(Kim et al., 2000). A TaqI PCR-RFLP protocol wasapplied to genotype this second mutation on 333 animalsof the studied hybrid line with phenotypic records, andthe frequencies observed for the genotypes AA, AG andGG were 0.08, 0.36 and 0.56, respectively.
The analyses using the standard animal model revealedsignificant effects of MC4R alleles on growth and fatnesssummarized in Table 2. The AA animals were heavierand fatter than GG ones. Nevertheless, out of the severalweight and fat measures analysed, only the effect on liveweight at 140 days, and carcass backfat at P2 and scapulawere statistically significant. Mean effects of allele substitu-tion are equivalent to approximately one half standarddeviation for live weight at 140d and carcass backfat atthe scapula, and to 0.2 standard deviations for carcassbackfat at P2. The missense mutation also influenced thecarcass composition (Table 2). Allele A significantlydecreased the weights of loin and trimmed shoulderadjusted for a common carcass weight, these effects beingequivalent to 0.2 S.D. for both traits. However, a signifi-cant effect of the allele A on the weight of trimmed hamwas not estimated (�0.065 ± 0.054; P < 0.40).
Significant effects of MC4R alleles were also evidencedon LD muscle colour and fatty acid composition of subcu-taneous backfat (Table 2). The loins of AA animals exhib-ited higher values for lightness (L*) and lower ones forredness (a*) than those of GG animals, which lead to a lessintense colour (higher hue angle Ho values). The backfat ofAA pigs showed higher content of saturated fatty acids(C16:0 and C18:0) and lower content of unsaturated fattyacids (C18:1 and C18:2) than GG animals. The percentagesof all the measured fatty acids are influenced by this poly-morphism, with effects ranging from 0.20 to 0.26 S.D.However, other important meat quality traits, such asmoisture, protein and intramuscular fat content (IMF) orpH 45 m, were scarcely affected by MC4R alleles.
Only additive effects of MC4R alleles were observed.Significant dominant effects on the analysed traits werenot detected in the additional analyses performed (resultsnot shown).
The results obtained with the transmission disequilib-rium test may be summarized as follows: the bTD estimatewas significant for the same traits as the animal model,excepting the measure P2 of carcass backfat; the bTD esti-mates were not statistically different from bPD estimatesfor all the analysed traits, discarding the risk of populationstratification, and hence discarding false positive results.Thus, the estimates of gene effects obtained with the animalmodel can be considered valid. When we do not find evi-dence for stratification/admixture in the population, theanimal model is a more powerful analysis to estimate alleleeffects, showing smaller standard errors.
4. Discussion
This study confirms the association previously reportedfor the Asp298Asn mutation of MC4R gene with severalproductive traits in pigs slaugthered at 110 kg of liveweight, and provides new information regarding the traitsand populations involved. The statistical methods usedhere (animal model and TDT) allow us to reduce the riskof false positives that usually threatens this type of candi-date gene association analysis, due to possible stratification

46 C. Ovilo et al. / Meat Science 73 (2006) 42–47
of the analysed population. Considering together the worksof Kim et al. (2000) and Hernandez-Sanchez et al. (2003),both based on the same animal material, the previous evi-dence indicated an effect of the mentionedMC4R polymor-phism on backfat thickness and growth (test daily gain andlifetime daily gain), on four different commercial pig lines(A, Landrace; B, Large-White; C, Large-White x Duroc;and D, synthetic line). In Kim et al. (2000), a fifth commer-cial line (E) coming from the cross of Meishan and LargeWhite pigs did not yield significant association results forany trait, but moreover it showed a trend for backfat inthe direction opposite to that observed in the other lines.
The present work is based on the analysis of a commer-cial hybrid line derived from a synthetic population comingfrom the intercross of Chinese Taihu and Large-White pigsselected for lean growth during eight generations, and afinal cross with Landrace pigs. This population would beequivalent to a cross between lines A and E of the previousstudies. Our results show an association of the MC4R geneSNP with backfat and growth measured as live weight at140 days. As this one is the only growth measure that yieldssignificant association results, it can be postulated that theeffect on growth of MC4R polymorphism is only producedin earlier growth stages. This interpretation agrees withprevious results that report significant effects of theMC4R missense mutation on average daily gain but inwhich the test period comprised 70–160 days of age (Kimet al., 2000). The sense of the estimated effects is the samethat was observed in previous work; allele A increasing fat-ness and growth, supporting the causality of the analysedpolymorphism. Differences in association results betweenthe two similar populations of Chinese-european origin(line E and our hybrid line) may indicate interaction withthe genetic background, although confused resultsobtained for line E must be interpreted cautiously as thenumber of observations was low for this line. In fact, neg-ative association results have been only obtained for thismutation in studies with a reduced number of animals thatcould limit the ability to detect small sized effects, as occursin line E analysed by Kim et al. (2000), or in the small WildBoar x Large White intercross population analysed byPark, Carlborg, Marklund, and Andersson (2002).
The study of associations of candidate genes is a step forthe knowledge of the genetic basis of productive traits, andcompared to other genomic approaches (QTL detection) ispotentially more easily and efficiently implemented inbreeding programmes. Based on this knowledge, new strat-egies of gene assisted selection or introgression could bedesigned to modify pig meat and fat characteristics. Forthis purpose, allele variants with favourable effects on eco-nomic traits must be previously tested in diverse pig popu-lations and production systems. These analyses areessential to confirm the effects previously found and to con-trast the gene effects on different meat and fat quality traitsto take this information into account for its possible prac-tical use in selective breeding. In this sense the present workprovide important information regarding the possible use-
fulness of the Asp298Asn mutation on MC4R gene for itsuse in pig breeding programmes. Several carcass composi-tion and meat and fat quality traits have been analysed forassociation with MC4R gene, which have not been studiedpreviously. In relation to carcass composition traits, theweights of loin and trimmed shoulder were significantlyaffected by MC4R SNP, allele G being the favourable forboth traits. Regarding meat quality and composition, outof the eight traits analysed only the effects on MinoltaL*, a* and Ho parameters were significant. Finally, withrespect to fatty acid composition traits, all the four fattyacids recorded in this work are affected by the analysedpolymorphism.
The results presented here give information about theusefulness of this marker for gene assisted selection inbreeding nucleus. The desirable genotype will depend onthe interests that base the selection decisions accordingto the production system and its specific market. Noneof the alleles would have a clear advantage, as selectionof the A allele will lead to fatter and heavier pigs, butwith a lower yield in premium cuts, meat of lighter colourand a modified fatty acid profile which will tend to satu-ration. Nevertheless, positive selection of allele A in damlines could be favourable to help its reproductive aptitude,and selection for increasing allele G frequency in sire lineswould be adequate to improve the premium cuts yield andthe meat colour. These tactics would increase the propor-tion of heterozygous individuals for the MC4R gene inthe commercial crossbred pigs, leading to a higher unifor-mity of the carcasses in relation to the affected productiveand quality traits, well appreciated by the meatmanufacturers.
Acknowledgements
The authors are grateful to the collaboration of AlfonsoFuentetaja, Fernando Flamarique and Gene + Iberica,Nines Lopez for technical assistance and Malena Serranofor the statistical analysis software. This research wasfunded by grant FIT-010000-2001-5.
References
Ciobanu, D. C., Bastiaansen, J. W. M., Lonergan, S. M., Thomsen, H.,Dekkers, J. C. M., & Plastow, G. S. (2004). New alleles in calpastatingene are associated with meat quality traits in pigs. Journal of Animal
Science, 82, 2829–2839.De Vries, A. G., Sosnicki, A., Garnier, J. P., & Plastow, G. S. (1998). The
role of major genes and DNA technology in selection for meat qualityin pigs. Meat Science, 49(1), S245–S255.
Fujii, J., Kinya, O., Zorzato, F., De Leon, S., Khanna, V. K., &Weiler, J. E.(1991). Identification of a mutation in porcine ryanodine receptorassociated with malignant hyperthermia. Science, 253, 448–451.
Gandemer, G. (2002). Lipids in muscles and adipose tissues, changesduring processing and sensory properties of meat products. Meat
Science, 62(3), 309–321.Hernandez-Sanchez, J., Visscher, P., Plastow, G., & Haley, C. (2003).
Candidate gene analysis for quantitative traits using the transmissiondisequilibrium test: the example of the melanocortin four receptor inpigs. Genetics, 164, 637–644.

C. Ovilo et al. / Meat Science 73 (2006) 42–47 47
Houston, R. D., Cameron, N. D., & Rance, K. A. (2004). A melanocortinfour receptor (MC4R) polymorphism is associated with performancetraits in divergently selected large white pig populations. Animal
Genetics, 35(5), 386–390.Kim, K. S., Larsen, H. J., & Rothschild, M. F. (1999). Rapid commu-
nication: linkage and physical mapping of the porcine melanocortinfour receptor (MC4R) gene. Journal of Animal Science, 78, 791.
Kim, K. S., Larsen, N., Short, T., Plastow, G., & Rothschild, M. F.(2000). A missense variant of the porcine melanocortin four receptor(MC4R) gene is sassociated with fatness, growth and feed intake traits.Mammalian Genome, 11, 131–135.
Kim, K. S., Thomsen, H., Bastiaansen, J., Nguyen, N. T., Dekkers, J. C.M., Plastow, G. S., et al. (2004). Investigation of obesity candidategenes on porcine fat deposition quantitative trait loci regions. Obesity
Research, 12(12), 1981–1994.Kim, K. S., Reecy, J. M., Hsu, W. H., Anderson, L. L., & Rothschild, M.
F. (2004). Functional and phylogenetic analyses of a melanocortin fourreceptor in domestic pigs. Domestic Animal Endocrinology, 26, 75–86.
Le Roy, P., Elsen, J. M., Caritez, J. C., Talmant, A., Juin, H., & Sellier, P.(2000). Comparison between the three porcine genotypes for growth,carcass composition and meat quality traits. Genetics, Selection and
Evolution, 32, 165–186.Lo Fiego, D. P., Santoro, P., Macchioni, P., & De Leonibus, E. (2005).
Influence of genetic type, live weight at slaughter and carcass fatnesson fatty acid composition of subcutaneous adipose tissue of raw hamin the heavy pig. Meat Science, 69, 107–114.
Lopez-Bote, C. (1998). Sustained utilization of Iberian pig breed. Meat
Science, 49(1), S17–S27.Monin, G. (2003). Genomics: improving qualitative characteristics and
value of meat for pigs. Outlook on Agriculture, 32(4), 227–233.Ovilo, C., Fernandez, A., Noguera, J. L., Barragan, C., Leton, R.,
Rodrıguez, C., et al. (2005). Fine mapping of porcine chromosome6 QTL and LEPR effects on body composition in multiplegenerations of an Iberian by Landrace intercross. Genetical
Research, 85(1), 57–67.Park, H. B., Carlborg, O., Marklund, S., & Andersson, L. (2002).
Melanocortin four receptor (MC4R) genotypes have no major effect onfatness in a Large White x Wild Boar intercross. Animal Genetics, 33,155–157.
Perez-Enciso, M., & Misztal, I. (2004). Qxpak: a versatile mixed modelapplication for genetical genomics and QTL analyses. Bioinformatics,
20, 2792–2798.Rothschild, M. F. (2004). Porcine genomics delivers new tools and results:
this little piggy did more than just go to market. Genetical Research,83, 1–6.
Stalder, K. J., Rothschild, M. F., & Lonergan, S. M. (2005). Associationsbetween two gene markers and indicator traits affecting fresh and dry-cured ham processing quality. Meat Science, 69, 451–457.
Wardlaw, S. L. (2001). Obesity as a neuroendocrine disease: lessons to belearned from proopiomelanocortin and melanocortin receptor muta-tions in mice and men. Journal of Clinical Endocrinology and
Metabolism, 86, 1442–1446.
Copyright © 2022 FDOKUMEN




















