
Apoptosis-Inducing Galactolipids from a Cultured Marine Diatom, Phaeodactylum tricornutum

Assessing the vulnerability of endemic diatomspecies in Lake Baikal to predicted future climate change:a multivariate approach
A N S O N W. M A C K AY *, D . B . R Y V E S w , D . W. M O R L E Y *, D . H . J E W S O N z and P. R I O U A L §
*Department of Geography, Environmental Change Research Centre, Pearson Building, University College London, Gower Street,
London WC1E 6BT, UK, wDepartment of Geography, Loughborough University, Loughborough LE11 3TU, zMagherafelt, Co.,
96 Desertmartin Road, Magherafelt, Co. Derry, BT45 5HE, N. Ireland, UK, §Institute of Geology and Geophysics, Chinese Academy
of Sciences, PO Box 9825, Beijing 100029, China
Abstract
Diatoms in Lake Baikal exhibit significant spatial variation, related to prevailing climate,
lake morphology and fluvial input into the lake. Here we have assessed the threats to
endemic planktonic diatom species (through the development of empirical models),
which form a major component of primary production within the lake. Multivariate
techniques employed include redundancy analysis (RDA) and Huisman–Olff–Fresco
(HOF) models. Our analyses suggest that eight environmental variables were significant
in explaining diatom distribution across the lake, and in order of importance these are
snow thickness on the ice, water depth, duration of days with white ice, suspended
matter in the lake, days of total ice duration, temperature of the water surface in July,
concentration of zooplankton and suspended organic matter. Impacts on dominant
phytoplankton diatom species are highlighted using t-value biplots. Predictions of
future climate change on Lake Baikal are likely to result in shorter periods of ice cover,
decreased snow cover across the lake in spring, increased fluvial input into the lake, and
an increase in the intensification of surface water stratification during summer months.
All these factors are likely to impact negatively on the slow-growing, cold-water
endemics such as Aulacoseira baicalensis and Cyclotella minuta, which currently
dominate diatom assemblages. Instead, taxa that are only intermittently abundant, at
present, in offshore areas (e.g. Stephanodiscus meyerii) are likely to become more
frequent. However, given the climatic gradient across the lake, the timing and extent
of changes in community structure are likely to vary. Moreover, palaeolimnological
records show that Lake Baikal diatom assemblages have been dynamic throughout the
Holocene, with both endemic and cosmopolitan species exhibiting periods of domi-
nance. Effects of climate change on the entire lake ecosystem may yet be profound as the
structure of the pelagic food web may change from one based on endemic diatom taxa
to one dominated by nondiatom picoplankton, and as limnological functioning (e.g.
stratification and mixing) affects deepwater oxygen availability, nutrient cycling and
trophic linkages.
Keywords: endemic diatoms, environmental controls, future climate change, Lake Baikal, multivariate
analysis, multivariate ordination, surface sediments
Received 3 February 2006; revised version received 17 April 2006 and accepted 26 June 2006
Introduction
The magnitude and impact of future climate variability
on freshwater ecosystems is uncertain and this is espe-
cially the case in central Asia, where there is a paucity of
studies compared with, for example, European and
North American regions (e.g. Schindler, 2001; Elliott
et al., 2005). Gaining more knowledge becomes impera-
tive because in recent decades wintertime warming in
central Asia has been greater than any other region ofCorrespondence: Anson Mackay, tel. 1 44 20 7679 0588,
fax 1 44 20 7679 0565, e-mail: [email protected]
Global Change Biology (2006) 12, 2297–2315, doi: 10.1111/j.1365-2486.2006.01270.x
r 2006 The AuthorsJournal compilation r 2006 Blackwell Publishing Ltd 2297

the world and this trend is expected to continue, with
winter temperatures in central Asia predicted to
increase by between 2 and 5 1C over the next 50 years
(IPCC, 2001). Uncertainty especially exists with regard
to the impacts of changing climate on planktonic
communities and the pelagic food web. For example,
shifts in dominance between phytoplankton com-
munities are likely to result in shifts between food
sources for zooplankton with unknown impacts on
other organisms higher up the food chain. This may
be due to a number of factors, including changes in
autotroph size (e.g. Moline et al., 2004) or disruption
to trophic linkages during key limnological periods
such as thermal stratification and overturn (Winder &
Schindler, 2004).
Lake Baikal in central Asia has long been recognized
as one of the world’s most remarkable freshwater
ecosystems, especially in terms of its age, depth,
volume, continental setting, long sedimentary archive,
and the high degree of biological endemism that exists
within the lake (Kozhova & Izmest’eva, 1998). Diatoms
contribute a major part of primary production (Fietz
et al., 2005) especially to the spring crop, which is
thought to determine annual productivity in the pelagic
lake (Popovskaya, 2000). Monitoring of diatom crops
has been carried out for many decades now, but these
have been largely restricted to the south basin where
pollution impact is greatest. One study suggests that
during the 1950s, productive years in the lake were
dominated by large celled, endemic species belonging
to the genera Aulacoseria (5Melosira) and Cyclotella
(Popovskaya, 2000), although Synedra tends to bloom
following years of peak Aulacoseira production. In the
1990s, small-celled, cosmopolitan species belonging to
the genus Nitzschia were observed in higher numbers in
some years, which has been attributed to both increased
nutrient input into the lake from economic develop-
ment in the catchment (Bondarenko, 1999) and to global
warming (Popovskaya, 2000). However, diatom-moni-
toring studies over the last decade have shown that
increases in Nitzschia cells have not been maintained in
recent years. Moreover, these monitoring studies show
that trends in the south basin are different from trends
in the north basin, and that species’ responses across the
whole lake are complex. Considerable uncertainty,
therefore, exists about possible drivers influencing
planktonic diatom communities in Lake Baikal.
Certainly in records of recently deposited sediments,
extracted from pelagic regions of the lake, there is little
or no response of diatoms to pollution (Mackay et al.,
1998) but there are lake-wide responses to shifts in
climate associated with the period coincident with
the ‘Little Ice Age’ and subsequent global warming
(Mackay et al., 1998, 2005).
Phytoplankton responses to predicted future climate
variability may be assessed using a variety of modelling
techniques. Recent work by Elliott et al. (2005) linked
together a phytoplankton model with a regional climate
model developed by the Hadley Centre, UK, to estimate
changes in phytoplankton communities and biomass in
an English lake up to 2100. However, given the uncer-
tainties of diatom successional trends across the length
of Lake Baikal and limited climate modelling in this
region of central Asia in general, we have adopted an
alternative approach, by developing empirical models
from spatially derived data of sedimentary diatom
assemblages and environmental data.
In recent years, multivariate techniques have been
used to explore the biogeography of diatom species,
and relationships with their changing environment in
a number of regions prone to intense winter cold (e.g.
Laing & Smol, 2000; Lim et al., 2001; Weckstrom &
Korhola, 2001; Ruhland & Smol, 2002; Ryves et al.,
2002; Bouchard et al., 2004). These studies are based
on multiple sites/lakes spread across gradients of in-
terest and, therefore, the assumption is made that most,
if not all, of the species are cosmopolitan. These data are
then used by palaeolimnologists to reconstruct past
environmental changes (e.g. in northern and Arctic
settings; McGowan et al., 2003; Antoniades et al., 2005;
Solovieva et al., 2005). In Lake Baikal, due to its size and
heterogenous sedimentary environments, the distribu-
tion of diatoms exhibits significant spatial variability
(e.g. Bondarenko et al., 1996; Mackay et al., 2003). How-
ever, many of the diatoms are endemic, and their
ecologies are poorly defined in comparison with some
cosmopolitan taxa.
The principal aim of this study is to assess the
potential threats from future climate change on endemic
diatoms in Lake Baikal, which form a principal compo-
nent at the base of the food chain (Yoshii et al., 1999).
However, this paper is distinctive from other diatom–
environment interaction studies in a number of re-
spects. Firstly, we include information derived from
remote sensing to expand the range of explanatory
variables initially used by Mackay et al. (2003), most
notably data on ice, which influences light transmission
into the epilimnion for photosynthesis. Secondly, many
of the canonical ordination techniques used in previous
studies (including Mackay et al., 2003) rely on w2 dis-
tances (e.g. canonical correspondence analysis; CCA),
which can give undue weight to rare species in the
dataset (Legendre & Gallagher, 2001). As diatom assem-
blages in Lake Baikal tend to be dominated by relatively
few species of abundant phytoplankton, the species
data are, therefore, transformed in order that Euclidean
distance methods, such as redundancy analysis (RDA),
can be used to explore relationships between species
2298 A . W. M A C K AY et al.
r 2006 The AuthorsJournal compilation r 2006 Blackwell Publishing Ltd, Global Change Biology, 12, 2297–2315
and environmental variables (Legendre & Gallagher,
2001). The data generated in this study will improve
our knowledge of interactions between Lake Baikal
diatom assemblages and environmental variables that
influence their growth, thus, providing information for
attempts to predict the impacts of future climate change
in an aquatic ecosystem whose biodiversity is of global
significance.
Study area
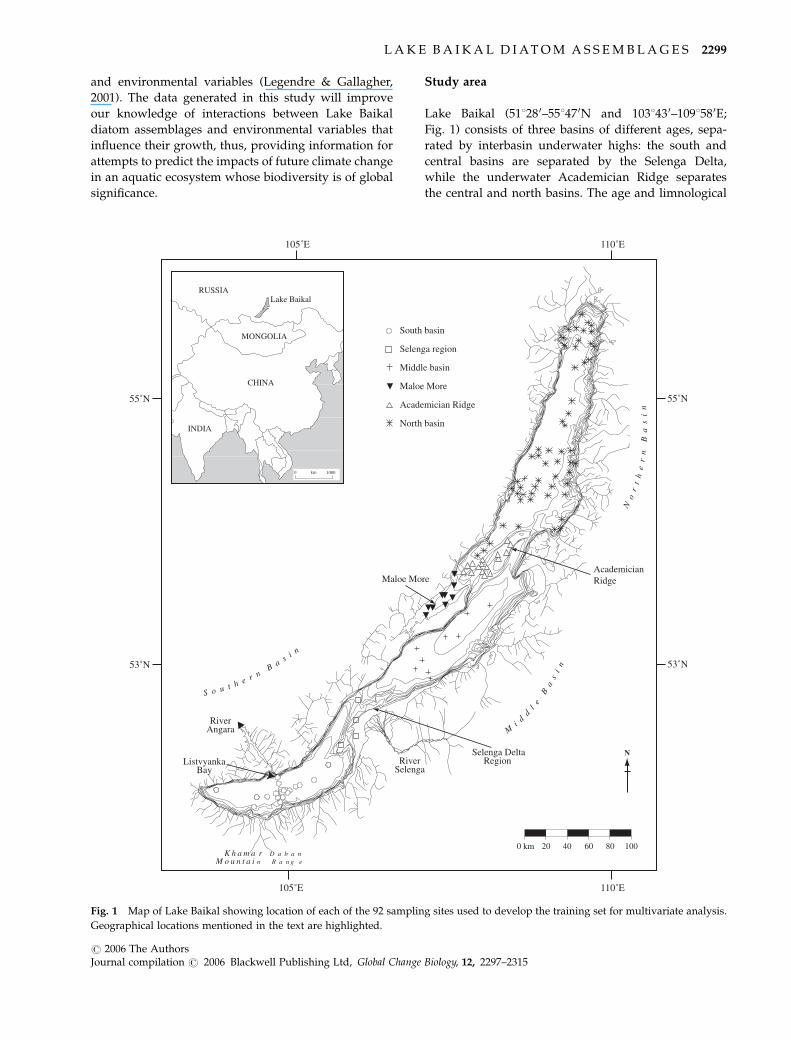
Lake Baikal (511280–551470N and 1031430–1091580E;
Fig. 1) consists of three basins of different ages, sepa-
rated by interbasin underwater highs: the south and
central basins are separated by the Selenga Delta,
while the underwater Academician Ridge separates
the central and north basins. The age and limnological
0 km 20 40 60 80 100
No
rt
he
rn
Ba
si
n
S o u t h e r nB
a s i n
Mi d
dl
eB
as
in
RiverAngara
RiverSelenga
Maloe More
Selenga DeltaRegion
AcademicianRidge
ListvyankaBay
K h a m a r D a b a nM o u n t a i n R a n g e
South basin
Selenga region
Middle basin
Maloe More
Academician Ridge
North basin
N
E011E501
E011E501
Lake Baikal
MONGOLIA
RUSSIA
CHINA
INDIA
Fig. 1 Map of Lake Baikal showing location of each of the 92 sampling sites used to develop the training set for multivariate analysis.
Geographical locations mentioned in the text are highlighted.
L A K E B A I K A L D I A T O M A S S E M B L A G E S 2299
r 2006 The AuthorsJournal compilation r 2006 Blackwell Publishing Ltd, Global Change Biology, 12, 2297–2315

characteristics of Lake Baikal has allowed an ecosystem
rich in species endemism to develop, and accordingly
it was designated a World Heritage Site in 1996. The
region around Lake Baikal is one of the most continental
in the world (Lydolph, 1977), with mean daily January
air temperatures of approximately �25 1C, rising
to 119 1C in July. Winters are long, dry and cold, while
summers are relatively short, warm and wet. Lake
Baikal freezes over every year and there is a distinct
north–south gradient in terms of ice duration and
thickness, with the south basin freezing later and thaw-
ing earlier than the north. The lake is distinct from other
deep lakes because its water column is saturated with
oxygen throughout, supporting an extensive, and
almost wholly endemic deep-water fauna. This is due
to both spring and autumnal overturn (i.e. the lake is
dimictic, despite strong stratification at 300 m) and
deep-water renewal from cold surface water intrusions
into deeper waters (Weiss et al., 1991; Wuest et al., 2005).
Methods
Diatom analysis of surface sediments
The dataset used in this study is based on 92 sites from a
training set of 126 surface sediment samples collected
from across the length of Lake Baikal, between 1992 and
1997 (Fig. 1; see Mackay et al., 2003 for full details). The
sedimentary environment of Lake Baikal is heteroge-
neous, and we have considered the impacts of sediment
redeposition, faunal bioturbation and diatom dissolu-
tion on the integrity of sites used in this study. Sites
were discarded if there was (i) visual evidence of
disturbance of the surface sediment–water interface,
(ii) visual evidence of turbidite layers present in the
upper sediments, when cores were opened longitudin-
ally or (iii) very few diatoms present in the surface
sediment (which results in relative abundances not
being representative of likely populations). This was
the case for one site, BAIK111, hence the number of sites
analysed is one less (92) than the 93 sites initially
investigated by Mackay et al. (2003). The surface sedi-
ments are oxygenated, allowing diverse faunal commu-
nities to live at the lake bottom (Martin et al., 2005).
Therefore, it follows that bioturbation will always
be an issue, and is one of the reasons why the strong
seasonal signals in sedimentation are not preserved in
annual laminations. Martin et al. (2005) present evi-
dence that concentrations of animal individuals (and
therefore bioturbation) are higher in shallow water
regions than in the abyssal plains. Moreover, bioturba-
tion probably plays a critical role in the preservation of
diatoms in surface sediments, through physical break-
age and irrigation of upper sediments, both of which
can be expected to enhance silica dissolution (Ryves
et al., 2003). However, we know from our own
high resolution analyses of surface cores that bioturba-
tion does not affect the overall stratigraphy of the
cores, as evidenced from radiometric 210Pb dating
and spheroidal carbonaceous particle analysis (Mackay
et al., 1998).
Given recent debates on the concept of ‘cosmopolitan-
ism’ within diatom research (e.g. Kociolek & Spaulding,
2000; Edlund & Jahn, 2001), diatom taxa were identified
according to a range of published papers and books,
most of which take account of the endemic diatom flora
found in Lake Baikal (e.g. Meyer, 1930; Skvortzov, 1937;
Zabelina et al., 1951; Skabichevskii, 1960; Gleser, et al.,
1988; Khursevich, 1989; Genkal et al., 1992; Flower, 1993;
Kozhova & Kobanova, 1995; Edlund et al., 1996).
Diatom counts were transformed to percentages before
all analyses.
Collation of environmental information
Environmental data in this study come from a number
of sources, including previous Russian studies, our own
measurements of water depth taken during each expe-
dition, and from remote sensing of snow and ice cover-
age across the lake. A description of the collation of
Russian and water depth data has already been detailed
in Mackay et al. (2003), so here we focus on the meth-
odology for the collection of data derived using remote
sensing techniques. Remote sensing is a useful tool in
the study of environmental variables at high spatial and
temporal resolution, especially over large and remote
regions such as Lake Baikal. Snow depth on ice has been
highlighted as one of the most important variables
influencing Lake Baikal diatom species (Mackay et al.,
2003). However, remote sensing of snow thickness/
depth is problematic. The best results are often obtained
from high resolution radar data, combined with an
extensive campaign of measuring actual snow depths
over the scene to calibrate the inferred snow depths (e.g.
Srivastan & Singh, 1991). Owing to the remoteness and
size of Lake Baikal and the dangers involved working
on the lake during the freeze–thaw cycle, it is too
dangerous to collect such ground-truth data. As a
result, this study attempts to investigate the seasonal
dynamics and duration of various ice-cover types on
Lake Baikal based on satellite images in the optical and
infrared spectra. Data for classification of Lake Baikal
ice cover were collated using the AVHRR meteorologi-
cal satellite (Morley, 2005). Similar data were also
collated in the studies of Lake Baikal by Bolgrien et al.
(1995), Le Core (1998), Semovski et al. (2000), Semovski
and Mogilev (2003), Kouraev et al. (in press). Classifica-
tion of snow and ice types on Lake Baikal has been
2300 A . W. M A C K AY et al.
r 2006 The AuthorsJournal compilation r 2006 Blackwell Publishing Ltd, Global Change Biology, 12, 2297–2315

previously carried out by Le Core (1998) using the
normalized difference snow index (NDSI) of Riggs
et al. (1994), although this study only exploits general
changes in ice cover characteristics and not, for exam-
ple, ice structure or relative wetness.
Images were chosen between 1996 and 1997, corre-
sponding to the final year in which surface sediments
were taken. AVHRR acquires data at a resolution of
approximately 1.1 km on the ground for each pixel, and
therefore, makes it an ideal tool for studying large sites
(such as Lake Baikal) that span several degrees of
latitude. Initially, the National Oceanographic Atmo-
spheric Administration’s (NOAA) Satellite Active
Archive (SAA) was screened for cloud-free images of
Lake Baikal for 1996–1997, and a total of 34 cloud-free
images of ice cover were obtained (between 10 Decem-
ber 1996 and 2 June 1997). The average time between
images is approximately 5 days, although during May
there are intervals of 10–15 days without cloud-free
images. The AVHRR data were processed using Erdas
Imagine 8.5, and the complete multispectral image was
classified using an unsupervised classification into
groups of pixels using the ISODATA clustering algo-
rithm, which is a centroid-based, iterative approach.
Classes according to coverage type were assigned using
comparisons with Le Core (1998) and by examining the
spectral signatures of certain classes, for example the
relatively higher reflectance of snow compared with
white ice in the visible channel. Pixels were classed into
five categories (open water, white ice, clear ice, wet ice
and snow) and recorded for each diatom sample loca-
tion (Morley, 2005). From these data, the total number of
days of the year when the lake was covered by white
ice, clear ice, wet ice and snow were calculated; a site
location was considered to be covered by a certain type
of ice up until this coverage changes in a subsequent
image. The total time of ice duration was also estimated,
as well as the total time of white ice and snow coverage
together as these are likely to be most limiting to light
penetration. Although only 1 year of data has been used
here, similar patterns have been demonstrated to occur
across the lake in other years covered by the dates of
diatom sample collection (e.g. Bolgrien et al., 1995;
Le Core, 1998; Semovski et al., 2000).
Data analyses
Multivariate datasets, especially those using composi-
tional data, are often analysed using w2 distance metrics
(such as correspondence analysis and the associated
technique of detrended correspondence analysis; DCA),
despite some misgivings with the approach (e.g. Faith
et al., 1987). An underlying issue with w2 distance
methods is that they are overly influenced by rare
species in the dataset, a problem not satisfactorily
addressed by often ad hoc methods of downweighting
included with some analysis programmes (P. Legendre,
personal communication). This is of particular concern
where the aim is to assess the significance of relation-
ships between species and environmental variables
(Legendre & Gallagher, 2001). An alternative approach,
using linear models (Euclidean distance) on suitably
transformed species data, has been recently proposed
(Legendre & Gallagher, 2001) and can be applied to
datasets where gradients of species turnover (as mea-
sured by DCA for example) would imply unimodal
responses, as here. In the absence of clear criteria for
choosing the optimal method a priori, Legendre &
Gallagher (2001) suggest selecting the method that
explains the highest fraction of the species data in direct
ordination. We found a linear model (using RDA) on
square-root transformed relative abundance data (the
Hellinger transformation; Legendre & Gallagher, 2001)
outperformed unimodal models (assessed with CCA),
whether or not species downweighting was applied,
and thus we have adopted this approach here.
All ordination methods were carried out using the
software package CANOCO v. 4.5 (ter Braak &
Smilauer, 2002). Additionally, samples and environ-
mental variables were screened within CANOCO v.
4.5 to determine samples with extreme environmental
values and to exclude those environmental variables
that exhibited high collinearity, which have the effect of
destabilizing the dataset. Relationships between diatom
distributions and environmental variables were initially
examined using the direct gradient technique of RDA,
which constrains ordination axes as linear combinations
of environmental variables. In order to identify a
minimum subset of variables that significantly explain
variation in the diatom data, redundant variables were
removed through a form of step-wise regression (for-
ward selection) together with Monte Carlo permutation
tests (P 5 0.0001; n 5 9999) and associated Bonferroni
corrections (ter Braak & Smilauer, 2002). Separate RDAs
were carried out to determine the explanatory power
of each significant environmental variable without
considering covariables to confirm each was significant
alone. The unique contribution of each of the forward-
selected variables was then assessed through a series of
partial RDAs with the remaining environmental vari-
ables as covariables (Table 1). Significance was again
tested using unrestricted Monte Carlo permutation
tests (P 5 0.0001, n 5 9999 permutations). Significant
positive and negative associations between dominant
planktonic taxa and forward selected environ-
mental variables were further explored using t-value
biplots and associated Van Dobben circles (ter Braak &
Looman, 1994).
L A K E B A I K A L D I A T O M A S S E M B L A G E S 2301
r 2006 The AuthorsJournal compilation r 2006 Blackwell Publishing Ltd, Global Change Biology, 12, 2297–2315

Tab
le1
Lim
no
log
ical
,cl
imat
ic,
chem
ical
and
bio
log
ical
dat
au
sed
asp
ote
nti
alex
pla
nat
ory
var
iab
les
for
the
dis
trib
uti
on
of
dia
tom
sin
surf
ace
sed
imen
ts
Cat
ego
ryE
xp
lan
ato
ryv
aria
ble
Co
de
Un
its
Dat
esm
easu
red
Min
imu
mM
axim
um
Mea
n
Sta
nd
ard
dev
iati
on
%ex
pla
ined
sin
gle
var
iab
le
RD
A
P (n5
9999
)
Lim
no
log
yW
ater
dep
th*
Dep
thm
1992
–199
720
.016
78.0
809.
648
0.3
10.1
0.00
01
Tra
nsp
aren
cyin
July
wT
ran
sju
lm
1961
9.0
25.0
21.4
4.6
——
Tra
nsp
aren
cyin
Sep
tem
ber
wT
ran
ssep
m19
612.
011
.06.
02.
0—
—
Alb
edo
inJu
lyw
Alb
edo
%n
/a
5.0
7.0
5.1
0.4
——
Tem
per
atu
reo
fJu
lyw
ater
surf
acez
Tem
pw
s1C
1896
–195
91.
013
.05.
92.
35.
70.
0003
Cli
mat
eA
nn
ual
sola
rra
dia
tio
nw
So
lar
mJm�
2n
/a
4100
.047
00.0
4478
.518
7.6
——
Ab
sorb
edra
dia
tio
nin
July
wA
bso
rbk
calc
m�
2n
/a
13.0
15.0
14.2
1.0
——
Mea
nJu
lyai
rte
mp
erat
ure
wJu
lair
1C
Lo
ng
term
ob
serv
atio
nsw
w9.
015
.012
.21.
7—
—
An
nu
alp
reci
pit
atio
nw
Pre
cip
mm
Lo
ng
term
ob
serv
atio
nsw
w17
5.0
550.
025
7.3
67.7
——
Sn
ow
thic
kn
ess
on
the
lak
ein
Mar
chw
Sn
ow
cm19
72–1
973
2.5
12.5
6.2
3.2
11.0
0.00
01
Ice
thic
kn
ess
inM
arch
§Ic
eth
ick
cm19
72–1
985
65.0
100.
082
.97.
2—
—
Len
gth
of
ice
cov
er§
Icel
eng
thd
ays
Lo
ng
term
ob
serv
atio
nsw
w12
6.0
168.
014
5.3
10.7
——
An
nu
ald
ura
tio
no
fw
hit
eic
eco
ver
age}
Day
sWI
day
s19
96–1
997
076
5317
.17.
80.
0001
An
nu
ald
ura
tio
no
fcl
ear
ice
cov
erag
e}T
clea
rd
ays
1996
–199
70
8516
.222
.6—
—
An
nu
ald
ura
tio
no
fsn
ow
cov
erag
e}T
sno
wd
ays
1996
–199
70
6332
.415
.7—
—
An
nu
ald
ura
tio
no
fw
etic
eco
ver
age}
Tw
etd
ays
1996
–199
710
5732
.011
.0—
—
Ice
cov
erd
ura
tio
n}
Day
sIce
day
s19
96–1
997
8316
813
1.0
24.7
5.7
0.00
05
An
nu
ald
ura
tio
no
fw
hit
eic
e
and
sno
wco
ver
age}
Tw
1s
day
s19
96–1
997
1413
185
.529
.8—
—
Ch
emis
try
N–
NO
3k
Nit
rate
mg
m�
3Ju
ly19
7815
.070
58.1
716
.4—
—
P–
PO
4k
Ph
osp
hat
em
gm�
3Ju
ly19
784.
012
.00
7.73
2.7
——
Su
spen
ded
mat
ter*
*S
usp
Mat
mg
L�
1Ju
ly19
780.
84.
002.
320.
65.
90.
0004
Su
spen
ded
org
anic
C**
Su
spO
rgC
mg
L�
1Ju
ly19
780.
31.
500.
380.
24.
10.
0054
Org
anic
C**
Org
Cm
gL�
1Ju
ly19
781.
11.
701.
330.
2—
—
Org
anic
N**
Org
Nmg
L�
1Ju
ly19
7870
.017
5.00
118.
3824
.2—
—
Org
anic
P**
Org
Pmg
L�
1Ju
ly19
787.
016
.50
9.96
2.1
——
Bio
log
yZ
oo
pla
nk
ton
bio
mas
sin
Sep
tem
ber
wZ
oo
bio
gm�
219
855.
030
.010
.08.
65.
50.
0009
Ori
gin
also
urc
ere
fere
nce
sin
clu
de:
* Dat
afr
om
this
stu
dy
;w A
tlas
Bai
kal
a(1
993)
;z V
erb
olo
vet
al.
(196
5);
§S
him
arae
vet
al.
(199
4);
}M
orl
ey(2
005)
;k V
oti
nse
vet
al.
(197
5);
**T
aras
ov
a&
Mes
cher
yak
ov
a(1
992)
;ww
Lo
ng
term
ob
serv
atio
ns
ov
erth
ela
stce
ntu
ry.
See
tex
tfo
rd
etai
ls.
n/
a,n
ot
avai
lab
le.
2302 A . W. M A C K AY et al.
r 2006 The AuthorsJournal compilation r 2006 Blackwell Publishing Ltd, Global Change Biology, 12, 2297–2315

The responses of eight of the main planktonic species
(as untransformed, percentage composition data pre-
sent in at least 10 sites) to the eight significant, modelled
environmental variables were determined by fitting
Huisman–Olff–Fresco (HOF) models (Huisman et al.,
1993) with the software HOF (Oksanen & Minchin,
2002; http://cc.oulu.fi/� jarioksa/). This program fits
the simplest response model from the five defined by
Huisman et al. (1993) to species data using maximum
likelihood estimation and a Poisson error distribution.
These models are a skewed unimodal response (model
V), a symmetrical or Gaussian model (model IV), a
monotonic model with a fixed plateau (model III), a
monotonically increasing or decreasing model (model
II) and finally the null model, which is flat with no
significant trend (model I). This method is particularly
useful when defining the type of species response to an
environmental variable (i.e. if there is a linear, unimodal
or no response). Here, we explore the responses of the
dominant planktonic taxa to each of the significant
modelled variables determined by ordination methods,
and assess the potential impacts of future climate
variability. Gaussian logit regression was used to test
the significance of optima of the HOF models that
exhibited unimodal responses (VI and V) for each of
the common planktonic diatom taxa (GLR version 1.1
Juggins 1994, unpublished program).
Results
Spatial distributions of dominant diatom taxa
Eighty-six diatom species were identified in the surface
sediments from 92 sites. Based on number of occur-
rences and N2 diversity (data not shown), the most
abundant taxa in our dataset were endemic plankton,
although some littoral species were also common,
including Staurosirella pinnata (Ehr.) Williams & Round
and Cocconeis placentula var. euglypta (Ehr.) Grun. Pro-
portional abundances of selected diatom species in the
training-set surface sediments have been mapped to
give an indication of the spatial distribution of species
across the lake (Figs 2a and f). These species have also
been fitted to HOF models: Aulacoseira baicalensis
(Meyer) Simonsen, A. skvortzowii Edlund, Stoermer &
Taylor, Cyclotella baicalensis Skv., Crateriportula inconspi-
cua (Mak. & Pom.) Flower & Hakansson, Cyclotella
minuta (Skvortzow) Antipova., Synedra acus var. radians
Kutz., S. acus var. radians (Kutz). Hust. fo. pusilla,
Stephanodiscus meyerii Genkal & Popovskaya. Mapping
in this way provides a useful, visual guide to species
distribution across the latitudinal gradient encom-
passed by the three basins. It is important to note here
that dissolution processes affect all taxa to different
extents (e.g. Ryves et al., 2003; Battarbee et al., 2005).
This is especially the case for Nitzschia acicularis W.
Smith, a cosmopolitan species commonly found in the
phytoplankton, but which is not recorded in surface
sediments due to dissolution processes that occur
throughout the water column (see Ryves et al., 2003
for full details).
Highest relative abundances of A. baicalensis are
found in the north basin (Fig. 2a), correlated with high
values for snow thickness and white ice formation (see
Fig. 3b), while the area of low abundance in the central
basin (Fig. 2a) occurs in an area of persistent clear ice
(Semovski et al., 2000; Morley, 2005). High values are
also recorded in the south basin, away from the shallow
waters of the Selenga Delta. A. skvortzowii can be found
in surface sediments throughout the lake; valves of both
vegetative cells and resting spores were found in 90 of
92 sites, but only with moderate N2 diversity; data not
shown. However, relative abundances are much greater
in the central region of the south basin than elsewhere
(i.e. relative abundances are very low in the central
region of the central basin, and throughout most of the
north basin; Fig. 2b). C. baicalensis and C. inconspicua
were found in only very small abundances, with no
clear patterns (Figs 2c and d), although it is important to
acknowledge that given the size of cells of the former
species, its contribution to biovolume will be signifi-
cant. C. minuta has a similar distribution to A. baicalensis
although relative abundances in the very north of the
lake are considerably lower (Fig. 2e). S. acus var. radians
fo. pusilla has a much more restricted distribution, only
occurring in the central region of the south basin, with
a couple of occurrences in the central basin (Fig. 2f).
Occurrence of the cosmopolitan species S. acus var.
radians is restricted to the clear ice region of the central
basin (Fig. 2g). There are other occasional occurrences,
mainly in shallower, coastal regions influenced by
fluvial input (e.g. at the Buguldeika Saddle; Fig. 2g).
Taken together, S. acus var. radians agg. is most abun-
dant in the south and central basins, with very little
presence at all in the north basin of Lake Baikal. More-
over, maximum abundance of S. acus coincides with
either circulation currents crossing between the western
shore to the opposite side of the lake in the central
basin, or with the convergence of circulation currents in
the south basin (see fig. 10.3.2 in Shimaraev et al., 1994).
These circulation patterns (which are responsible for the
transport of diatom species from shallow waters into
the pelagic regions of the lake) are dependent on both
whole-lake cyclonic circulation (Verbolov et al., 1965 in
Shimaraev et al., 1994) and local circulation patterns
influenced by fluvial input. S. meyerii (synonymous
with S. binderanus var. baicalensis Popovskaya & Genkal)
is most commonly found in shallow waters of the Maloe
L A K E B A I K A L D I A T O M A S S E M B L A G E S 2303
r 2006 The AuthorsJournal compilation r 2006 Blackwell Publishing Ltd, Global Change Biology, 12, 2297–2315
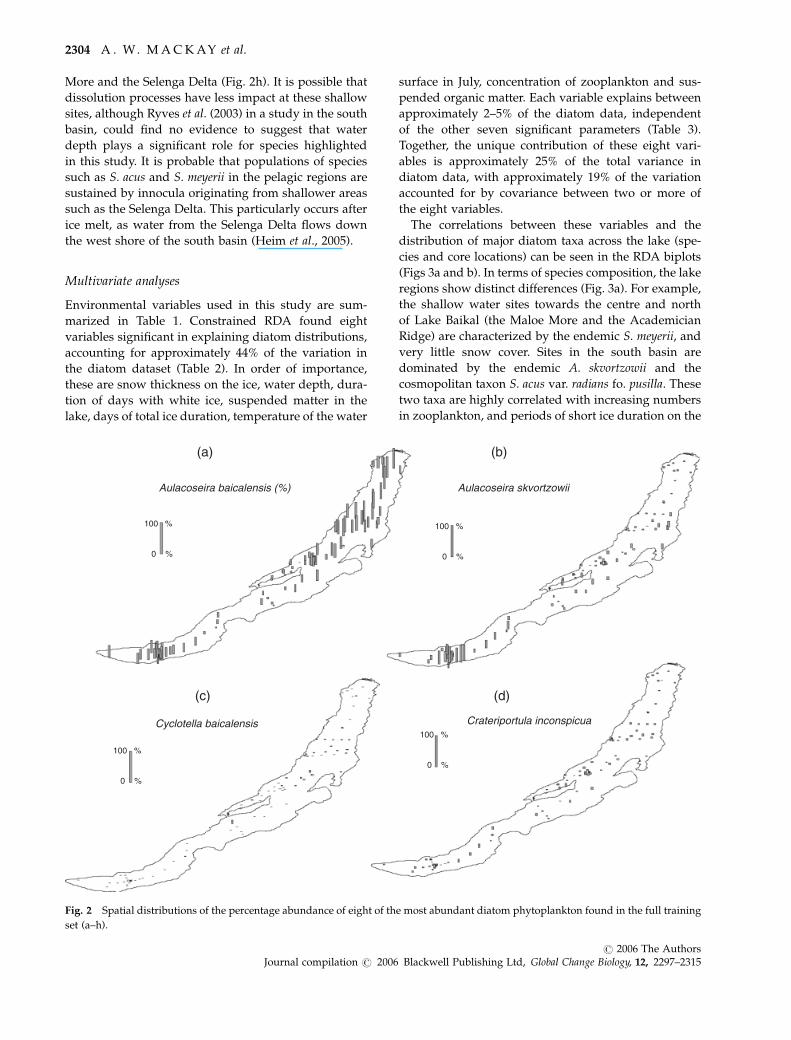
More and the Selenga Delta (Fig. 2h). It is possible that
dissolution processes have less impact at these shallow
sites, although Ryves et al. (2003) in a study in the south
basin, could find no evidence to suggest that water
depth plays a significant role for species highlighted
in this study. It is probable that populations of species
such as S. acus and S. meyerii in the pelagic regions are
sustained by innocula originating from shallower areas
such as the Selenga Delta. This particularly occurs after
ice melt, as water from the Selenga Delta flows down
the west shore of the south basin (Heim et al., 2005).
Multivariate analyses
Environmental variables used in this study are sum-
marized in Table 1. Constrained RDA found eight
variables significant in explaining diatom distributions,
accounting for approximately 44% of the variation in
the diatom dataset (Table 2). In order of importance,
these are snow thickness on the ice, water depth, dura-
tion of days with white ice, suspended matter in the
lake, days of total ice duration, temperature of the water
surface in July, concentration of zooplankton and sus-
pended organic matter. Each variable explains between
approximately 2–5% of the diatom data, independent
of the other seven significant parameters (Table 3).
Together, the unique contribution of these eight vari-
ables is approximately 25% of the total variance in
diatom data, with approximately 19% of the variation
accounted for by covariance between two or more of
the eight variables.
The correlations between these variables and the
distribution of major diatom taxa across the lake (spe-
cies and core locations) can be seen in the RDA biplots
(Figs 3a and b). In terms of species composition, the lake
regions show distinct differences (Fig. 3a). For example,
the shallow water sites towards the centre and north
of Lake Baikal (the Maloe More and the Academician
Ridge) are characterized by the endemic S. meyerii, and
very little snow cover. Sites in the south basin are
dominated by the endemic A. skvortzowii and the
cosmopolitan taxon S. acus var. radians fo. pusilla. These
two taxa are highly correlated with increasing numbers
in zooplankton, and periods of short ice duration on the
Aulacoseira baicalensis (%) Aulacoseira skvortzowii
Cyclotella baicalensis Crateriportula inconspicua
100 %
0 %
100 %
0 %
100 %
0 %
100 %
0 %
(a) (b)
(c) (d)
Fig. 2 Spatial distributions of the percentage abundance of eight of the most abundant diatom phytoplankton found in the full training
set (a–h).
2304 A . W. M A C K AY et al.
r 2006 The AuthorsJournal compilation r 2006 Blackwell Publishing Ltd, Global Change Biology, 12, 2297–2315

lake (Fig. 3b). The north basin is most characterized by
A. baicalensis and C. minuta. A. baicalensis is highly
correlated with increasing snow thickness estimates
on the lake, which in turn suggests that this taxon is
more tolerant of low irradiances than other taxa in the
dataset. Culture studies show that this species is also
intolerant of high irradiances (D. Jewson, unpublished
data), which contrasts with reports of Richardson et al.
(2000). However, their estimates of growth were based
on filament length; this can be misleading because with
A. baicalensis longer than normal vegetative cells can be
produced under light stress. Conversely, C. minuta is
highly correlated with ice duration on the lake – (i.e.
longer ice on the lake in spring promotes high abun-
dance of C. minuta during the following autumn turn-
over; Fig. 3b). The shallow waters of the Selenga Delta
are dominated by cosmopolitan plankton, including
S. acus var. radians, Stephanodiscus minutulus (Kutz.)
Cleve & Moller (not shown) and Stephanodiscus parvus
Stoermer & Hakansson (not shown). These latter two
taxa are more commonly found growing during the
summer in more nutrient-rich waters.
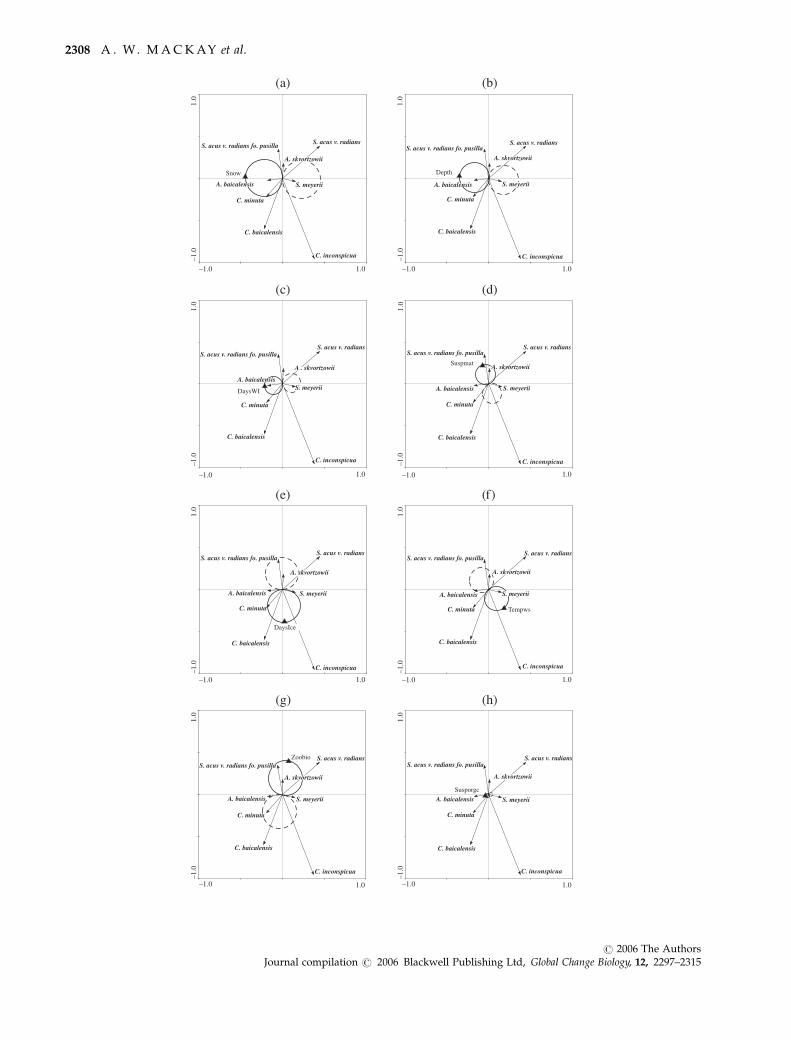
Results suggest that climatically controlled variables
(especially during spring, with the presence of ice on
the lake) are important in influencing diatom floristic
variation. Table 4 summarizes the distribution of HOF
models amongst the eight significant environmental
variables. More than half of the models show a signifi-
cant trend. Table 5 shows summary responses of the
eight main taxa to each of the significant environmental
variables, together with the corresponding fitted HOF
model. However, to assess which of these responses are
significant (both positively and negatively) t-value bi-
plots and Van Dobben circles have been constructed
and are shown in Figs 4a and h; ter Braak & Looman,
1994). Species that fall within the Van Dobben circles
are significantly (linearly) correlated (at P 5 0.05) to the
environmental variable, either positively (solid circles)
or negatively (dashed circles).
Snow depth on the lake has a positive, significant
effect on A. baicalensis and C. minuta, but a negative
impact on S. meyerii (Fig. 4a). Thus, we would expect
that, should future climate impacts result in a decline in
snow cover on the lake (see Discussion), then there is
Synedra acus v.radians fo. pusilla
Synedra acus v.radians
Cyclotella minuta (%)
Stephanodiscus meyerii (%)
100 %
0 %100 %
0 %
100 %
0 %
100 %
0 %
(e) (f)
(g) (h)
Fig. 2 (Continued)
L A K E B A I K A L D I A T O M A S S E M B L A G E S 2305
r 2006 The AuthorsJournal compilation r 2006 Blackwell Publishing Ltd, Global Change Biology, 12, 2297–2315

C. minuta
A. baicalensis
A. skvortzowii
C. inconspicua S. meyerii
S. acus v. radius
C. baicalensis
S. acus v. radians fo. pusilla
Depth
Snow
Suspmat
Zoobio
DaysWI
DaysIce
C. minuta
A. baicalensis
A. skvortzowii
C. inconspicua S. meyerii
S. acus v. radians
C. baicalensis
S. acus v. radians fo. pusilla
(a) (b)
−1.0−1.0
1.0
1.0−1.0
−1.0
1.0
1.0
Susporgc
Tempws
Fig. 3 Redundancy analysis (RDA) biplots showing (a) species and coded site locations and (b) species, abbreviated significant
explanatory variables and coded site locations. The coded site locations are previously outlined in Fig. 1. There are too many species to
identify individually in one diagram, and so in (b) all species other than the eight highlighted above are suppressed from view.
Table 2 Redundancy analysis (RDA) of species data (86 taxa) with eight forward selected variables using Hellinger’s distance
(Legendre & Gallagher 2001)
RDA Axes
1 2 3 4 Total variance
Eigenvalue 0.210 0.112 0.051 0.029 1.000
P 0.0001
Cumulative % variance of explanatory data 21.0 32.2 37.4 40.3
Sum of all eigenvalues 1.000
Sum of all canonical eigenvalues; P 5 0.0001 0.437
Table 3 Unique contributions to diatom variation of each of the eight significant environmental variables as determined using
RDA, with the remaining seven variables as co-variables, in order
No. Variable Eigenvalue % explained
P value for unique
explanation (n 5 9999)
1 Snow 0.048 4.8 0.0001
2 Depth 0.044 4.4 0.0001
3 DaysWI 0.034 3.4 0.0001
4 SuspMat 0.026 2.6 0.0004
5 DaysIce 0.025 2.5 0.0005
5 Tempws 0.025 2.5 0.0003
7 Zoobio 0.024 2.4 0.0009
8 SuspOrgC 0.018 1.8 0.0054
Total 8 vars 24.4 (combined)
Including covariances, % explained of eight variables 5 43.7% (P 5 0.0001, n 5 9999).
2306 A . W. M A C K AY et al.
r 2006 The AuthorsJournal compilation r 2006 Blackwell Publishing Ltd, Global Change Biology, 12, 2297–2315

likely to a be a decline in deep water, pelagic, endemic
diatoms. Depth of water in the lake elicits a similar
response to snow cover (Fig. 4b), but predictions of
future climate change are not likely to have a significant
effect on water depths across the lake in the time scale
of 100 years, as precipitation is greater than evaporation
in all models. Days of white ice duration on the lake
have a positive impact on A. baicalensis but negative
impact on S. meyerii (Fig. 4c; i.e. low irradiances under
the ice favour A. baicalensis). Suspended matter has a
significantly positive impact on A. skvortzowii only (Fig.
4d), which in the south basin is mainly derived from
Selenga River input (Heim et al., 2005). Ice duration (Fig.
4e) on the lake positively affects C. minuta, and this is
likely to be related to the fact that net increase in cells of
this species occurs during autumn overturn (see
’Discussion‘). The temperature of the water surface
during July also impacts significantly on several species
(Fig. 4f). Higher water temperatures have a negative
impact on diatom distribution on both the endemic
Aulacoseria species. However, warmer temperatures
are associated with significantly higher abundances of
S. meyerii, and it may be expected that predicted
changes in climate will result in higher abundances of
this species across the lake. Concentrations of zooplank-
ton, especially the endemic mesozooplankton Epischura
baicalensis Sars. are associated with lower abundances of
C. minuta, because these animals track populations of
C. minuta as a food source (Fig. 4g). Finally, within our
datasets, suspended organic matter in the lake does not
significantly impact on any of the eight species high-
lighted here (Fig. 4h), and only explains under 2% of
total diatom variation (Table 3).
Discussion
Ecological responses of dominant diatom taxa
A. baicalensis is most productive in areas of opaque ice
(e.g. because of snow cover or white ice) rather than
clear ice, which appears to be detrimental to growth.
This implies that increasing light penetration (e.g.
through clear ice) inhibits growth of this taxon. How-
ever, unlike C. minuta, there is only a weak correlation
with ice duration on the lake (Fig. 3b), which will have
implications for spatial responses within Lake Baikal to
future climate change (i.e. between the north and south
basins). In the north basin, ice break-up occurs later in
the spring, and subsequent warming of the lake water
delayed, compared with the south basin. In some years
in the north basin, spring turnover period can be con-
sequently shorter too (Shimaraev et al., 1994). Impor-
tantly, summer stratification is shorter in the north
basin, during which time there is a shift from diatom-
Table 4 Distribution of HOF models amongst the eight significant environmental variables
Model Snow Depth White ice SuspMat Ice dur Tempws Zoobio SuspOrgC Total %
V 1 6 0 1 4 3 0 0 17 4.5
IV 9 7 10 8 6 6 3 9 70 18.5
III 0 0 1 0 0 0 0 0 1 0.2
II 15 16 11 18 14 8 19 13 119 31.4
I 17 13 20 15 20 25 20 19 172 45.4
Note that the majority of the models are linear (II 1 I), 291/379 5 77%.
Table 5 HOF responses for each of the common planktonic taxa in Lake Baikal
Snow Depth White ice SuspMat Ice dur Tempws Zoobio Susp OrgC
Aulacoseira baicalensis II1 V* II1 IV* II1 IV* I IV*
A. skvortzowii IV* II1 IV* V II� V II1 I
Stephanodiscus meyerii II� IV* II� II� V* I II� II�Cyclotella minuta II1 V* II1 I II1 IV* II� I
Synedra acus v. radians II� I II� IV* IV* I I IV*
Crateriportula inconspicua I I I II1 I I II� I
S. acus v. radians f. pusilla I II1 IV* IV IV* II� II1 II�Cyclotella baicalensis I II1 I I II1 IV* I I
Where unimodal models are apparent (i.e. IV and V), significant optima determined using GLR are indicated with an asterisk.
L A K E B A I K A L D I A T O M A S S E M B L A G E S 2307
r 2006 The AuthorsJournal compilation r 2006 Blackwell Publishing Ltd, Global Change Biology, 12, 2297–2315
(a) (b)
(c) (d)
(e) (f)
(g) (h)
2308 A . W. M A C K AY et al.
r 2006 The AuthorsJournal compilation r 2006 Blackwell Publishing Ltd, Global Change Biology, 12, 2297–2315

dominated phytoplankton to dominance by autotrophic
picoplankton, especially the cyanobacteria Synechocystis
limnetica Popovskaya (Popovskaya, 2000; Fietz et al.,
2005).
A. skvortzowii populations can develop both in spring
in pelagic regions of the lake, and during autumn
turnover in shallow-water bays (e.g. Bolshiye Koty
and Barguzin Bay; Fig. 1) (Kozhova & Izmest’eva,
1998). According to the HOF models, growth of A.
skvortzowii under increasing snow and ice cover is
inhibited when compared with the deep-water ende-
mics, A. baicalensis and C. minuta. Thus a combination of
environmental factors, including earlier ice out time
and warmer waters in the south basin (as suggested
by RDA, Fig. 3b) are likely to be responsible for the
spatial distributions observed here.
Like A. baicalensis, C. minuta is exclusively pelagic.
Although this species can be found growing under the
ice in spring (when it usually auxosporulates; Kozhova
& Kobanova, 1995), the main period of net increase in
cell numbers occurs during autumn turnover when
nutrients become available again and concomitant
water transparency is low (confirmed by RDA and the
high correlation with increased suspended matter in
the water column; Fig. 3b). High relative abundance of
this species may be related to periods of warmer waters
and a dominance of autumnal over spring productivity,
due to either longer summers (Bradbury et al., 1994) or
restriction of the size of the spring crop relative to the
autumn crop, due to long, cold winters, with concomi-
tant increased ice duration on the lake. The latter reason
has been invoked by Mackay et al. (2005) to explain the
dominance of this species in the sediment record during
the period commonly referred to as the ‘Little Ice Age’
(approximately AD 1200–1840). At this time, there was a
weakening of the North Atlantic Oscillation (NAO;
Shindell et al., 2001) and the expansion of the Siberian
High (SH) across central Asia (Krenke & Chernavskaya,
2002). Our data suggest, therefore, that factors that
promote a decline in ice and snow cover during spring
(or variables that are linearly related to these factors),
will also have a negative impact on populations of
C. minuta the following autumn.
In palaeoecological studies, abundance of both forms
of S. acus is interpreted as being associated with in-
creased concentrations of silica, related to increased
catchment run-off and river discharge (Kilham &
Kilham, 1990; Bradbury et al., 1994). The distribution
of S. acus observed here reflects these parameters. For
example, biogeochemical silica mass balance (Callender
& Granina, 1995) shows that the production of diatoms
in Lake Baikal is supported mostly by remineralization
processes in the water column, accompanied by signifi-
cant contribution from fluvial inputs of dissolved silica,
especially from the Selenga River which flows into the
central and south basins. Very low relative abundances
of S. acus agg. in the north basin are likely, therefore, to
be related to increased snow and ice cover (as suggested
by RDA, Fig. 3b), together with lower concentrations of
silica input via rivers than in the other two basins
(Shimaraev et al., 1994). It is also worth noting that the
valves of S. acus suffer considerable breakage and
dissolution in the water column at the surface sedi-
ment–water interface (Ryves et al., 2003; Battarbee et al.,
2005). It is likely, therefore, that the distribution of this
species is under-represented in the sediment record.
In agreement with the Kilham & Kilham (1990)
hypothesis for Si : P ratios for cosmopolitan diatoms in
African Great Lakes (i.e. Synedra4Nitzschia4Stephano-
discus), Bradbury et al. (1994) suggested from palaeoe-
cological studies that phosphorus concentrations are
likely to be important for growing populations of
S. meyerii. This will occur when nutrient dynamics are
dominated by circulation, promoting a long period of
spring overturn and large release of nutrients from
bottom sediments. A recent study by Muller et al.
(2005) demonstrated that flux rates for phosphorus
(between the surface sediment–water interface) were
significantly higher in the Maloe More than elsewhere
in the lake during the period investigated (1994–2003),
agreeing with the assertion by Bradbury et al. (1994) that
high phosphorus supply is an important factor for this
species. S. meyerii exhibits a linear response to snow
depth over the gradient sampled, with highest abun-
dance at minimum snow depths (Fig. 4a; Table 5). RDA
also shows that S. meyerii is found in highest abun-
dances in shallow regions of Lake Baikal with low levels
of snow cover when the lake is frozen. The Maloe More
is subjected to high winds (Shimaraev et al., 1994), while
the region is also semi-arid, receiving less than 200 mm
of precipitation annually compared with the 300–
400 mm falling annually in the south basin (Atlas
Baikala, 1993). Both these factors result in low levels
of snow accumulation on the ice of frozen Maloe More
during spring. This fits with the assumption of
Bradbury et al. (1994) that this species is indicative of
Fig. 4 t-Value biplots, showing relationships between the eight highlighted species and each of the significant explanatory variables
(a 5 Snow; b 5 Depth; c 5 DaysWI; d 5 SuspMat; e 5 DaysIce; f 5 Tempws; g 5 Zoobio; h 5 SuspOrgC). Species that fall within the Van
Dobben circles are significantly (linearly) correlated (at P 5 0.05) to the environmental variable, either positively (solid circles) or
negatively (dashed circles).
L A K E B A I K A L D I A T O M A S S E M B L A G E S 2309
r 2006 The AuthorsJournal compilation r 2006 Blackwell Publishing Ltd, Global Change Biology, 12, 2297–2315

a dry climate (less snow cover) dominated by water
column circulation and nutrient regeneration.
Future impacts of climate change on Lake Baikal diatomassemblages
In continental regions in the northern hemisphere, ice
cover is an important variable in determining function-
ing of lake ecosystems. This is especially the case with
regard to the interactions between diatom communities
and ice and snow cover in locations vulnerable to global
warming, such as the Arctic (e.g. Douglas et al., 1994;
Sorvari et al., 2002; Smol et al., 2005), and high altitude
regions such as the European Alps (e.g. Lotter & Bigler,
2000; Ohlendorf et al., 2000) and in the Canadian Cor-
dillera (e.g. Karst-Riddoch et al., 2005). Ice cover plays a
major role in the stratification of the water column,
which in turn has an influence on lake mixing and
nutrient cycling processes (Shimaraev et al., 1994). A
recent study of 26 sites around the northern hemisphere
has shown that ice cover on lakes and rivers has been
responding to global warming since the mid 19th
century, with net reduction in ice duration through
combinations of later ice formation and earlier ice
break-up (Magnuson et al., 2000). One of the case
studies used in this investigation was an ice phenology
record from Lake Baikal, near the mouth of the Angara
River (Fig. 1). This record has been monitored continu-
ously since 1869, and its relationship to northern hemi-
sphere (NH) temperatures and large-scale climate
indices such as the NAO and the Arctic Oscillation
(AO) have been examined in detail (e.g. Livingstone,
1999; Shimaraev et al., 2002; Todd & Mackay, 2003).
These studies are important as they provide plausible,
mechanistic links between regional climate and its
impact on biology, especially the primary producers
within Lake Baikal. Todd & Mackay (2003) have argued
that, because some modes of NH variability are them-
selves predictable, then aspects of ice cover change on
Lake Baikal ought be predictable too and, therefore, it
should be possible to forecast some of the changes in the
biology that may occur. More recently, Panagiotopoulos
et al. (2005) detail a decline in the strength of the
Siberian High since the 1970s, which is linked to in-
creases in precipitation in central Asia. This decline is
projected to continue at least until approximately 2100,
and while it has not been possible to determine the
cause of the decline, future climate general circulation
model predictions suggest a negative correlation
between increases in greenhouse gases (GHGs) and
the intensity of the SH. Here, we will discuss some of
the possible impacts of future climate change on the
diatom populations in Lake Baikal as determined from
our multivariate approach.
Future climate impacts will result in:
(i) Shorter ice duration: Using scenarios of anthropo-
genic greenhouse gas emissions, model predictions of
climate variability over the next 100 years show pro-
nounced warming during winter in the Lake Baikal
region (IPCC, 2001). For example, under conditions of
a 1% annual increase in CO2, model simulations suggest
that, during the period 2071–2100, mean winter tem-
perature changes over the Northern Asian region range
from 1 4.8 to 1 9.3 1C, relative to the 1961–1990 period
(IPCC, 2001). Todd & Mackay (2003) calculated that,
assuming a linear dependence of Lake Baikal ice
duration on winter temperature, ice duration in the
south basin may decrease by a further 15–28 days
by 2071–2100. This decrease is rather less than that
estimated by Shimaraev et al. (2002) who calculated
that ice duration may decrease by as much as 2 months
in the south basin. Our analyses suggest that shorter
ice duration will favour growth of species which pre-
sently develop in shallow and coastal water regions
(e.g. A. skvortzowii, S. meyerii and S. acus; Fig. 4e),
whose cells are then transported into the open
water through circulation processes in the lake. Con-
versely, populations of the pelagic, endemic species
C. minuta are likely to decline, as ice duration gets
shorter (Fig. 4e).
(ii) Reduced snow cover extent: Snow cover extent in
central Asia is controlled by a number of factors, most
notably the availability of precipitation (Clarke et al.,
1999) linked to changes in e.g. Atlantic sea surface
anomalies and the associated NAO (Ye, 2001). Snow
cover on frozen Lake Baikal is the strongest single
variable in explaining variation in diatom assemblages
(Tables 1 and 3). Light is important because it has a
direct influence on algal photosynthesis (and primary
production) and provides the energy needed for con-
vective mixing to keep diatom cells suspended in the
photic zone (Granin et al., 2000). On Lake Baikal, depth
of surface snow often exceeds 10 cm, which results in
light becoming limiting for diatom cells in the water,
and mixing processes to weaken, causing heavier dia-
toms to sink out of the photic zone (Granin et al., 2000).
A. baicalensis is adapted to low light conditions and,
therefore, snow cover offers this species a competitive
advantage over other taxa, which partly explains its
dominance in the north basin of Lake Baikal. However,
should snow cover impede too much light, then growth
of this taxon too is inhibited (Mackay et al., 2005).
Regional analysis of snow cover extent indicates that
between the 1970s to the middle of the 1980s snow
cover over Eurasia was more extensive than between
late 1980s to late 1990s (Robinson & Frei, 2000), while
model predictions of future global warming suggests
that there will be less overall winter snow accumulation
2310 A . W. M A C K AY et al.
r 2006 The AuthorsJournal compilation r 2006 Blackwell Publishing Ltd, Global Change Biology, 12, 2297–2315

(Barnett et al., 2005). Our models suggest a decline in
snow cover associated with global warming will have a
negative impact on deep-water pelagic species such as
A. baicalensis and C. minuta, while promoting net growth
of more opportunistic, high-irradiance tolerant species
(both cosmopolitan and endemic), including S. meyerii
(Table 5; Figs 3b and 4a).
(iii) Increased amounts of precipitation: Shimaraev et al.
(2002) report that over the last 100 years, there has been
a positive relationship between increasing temperatures
and hydrological inflow into the lake, which also corre-
late to observed trends in increasing levels of precipita-
tion in central Asia (Aizen et al., 2001). As precipitation
levels at the latitude of Lake Baikal are estimated to
increase by 5–10%, Shimaraev et al. (2002) further esti-
mate that water inflow via Lake Baikal tributaries will
increase by 5–10% too. It is likely, therefore, that there
will be an increase in more generalist species with
wider tolerances being transported into the pelagic
regions of the lake. For example, t-value biplots (e.g.
Fig. 4d) highlight that increases in suspended matter are
significantly associated with the endemic A. skvortzowii,
although resuspension of this taxon from littoral
regions during autumn turnover will be important
too. Increased fluvial input into Lake Baikal will also
result in increased silica and nutrient input into the
lake, which in turn will encourage growth of species
such as S. acus even in the north basin, because of its
higher requirement for silica (Bradbury et al., 1994).
There is considerable palaeolimnological evidence that
S. acus was in the past a dominant component of the
diatom flora across the length of Lake Baikal, linked to
prevailing warmer and wetter climates (e.g. Bradbury
et al., 1994; Karabanov et al., 2000; Mackay et al., 2005;
Morley, 2005). Other cosmopolitan species, such as
Asterionella formosa Hassall, A. subarctica (O. Muller)
Haworth, A. granulata (Ehrenberg) Simonsen, which
can be relatively abundant locally in coastal areas,
would also likely benefit from such changes, and evi-
dence is growing that these increases may already be
underway (e.g. Kozhova & Izmest’eva, 1998; Mackay
et al., 1998).
(iv) An increased period of summer stratification: Pre-
dicted increases in temperature are also likely to have
an important impact on the length (i.e. longer, and
increased stability) and depth of summer and autumn
surface-water stratification. An increase in the stratifi-
cation period will have a negative impact on the success
of the resting stages produced by A. baicalensis and C.
minuta, as they slowly sink through the water column to
avoid warmer conditions and nutrient depletion (D.
Jewson & N. Granin, unpublished data). If turnover
during autumn is delayed, then the resting cells of these
species will sink further in the water column during
stratification, making their resuspension less likely,
resulting in a smaller number of cells being entrained
back into the photic zone for the following years in-
oculum. Thus, under a warmer climate, populations of
autumnal taxa such as the slow growing C. minuta will
decline.
We acknowledge, however, that there is a consider-
able amount of uncertainty, especially with respect to
the extent of increasing summer and autumnal stormi-
ness in the Lake Baikal region, which may actually
increase resuspension of cells from deeper waters or
from littoral regions. Another major impact resulting
from increased stratification will be a shift in balance
between large phytoplankton (i.e. diatoms) and auto-
trophic picoplankton (especially Synechocystis limnetica;
Fietz et al., 2005). Picoplankton dominate Lake Baikal
stratified waters, and because compared with diatoms
relative cell-size is small, mass growth occurs due to
high rates of nutrient assimilation and high rates of cell
division (Popovskaya, 2000). The distribution of pico-
plankton in the lake varies greatly with region, thereby
again ensuring a complex lake-wide response to global
warming (Fietz et al., 2005). Nevertheless, such shifts are
likely to have important implications for trophic inter-
actions in the Lake Baikal aquatic ecosystem, especially
between primary producers and consumers. Our ana-
lyses suggest that increases in zooplankton biomass are
linked to a decline in C. minuta, and a relative increase
in coastal species, including A. skvortzowii and S. acus v.
radians fo. pusilla. We are not at a stage yet to assess how
such changes will impact on complex consumers such
as the endemic gammarids (e.g. Macrohectopus branickii),
or how these changes will be propagated up the food
chain. However, the effects are likely to be important for
many endemic fish and may have consequences at all
trophic levels over time, even affecting the endemic
freshwater seal, Phoco sibirica (Yoshii et al., 1999). Thus,
although the magnitude and nature of the impacts of
such changes on Lake Baikal’s biodiversity and ecolo-
gical functioning are uncertain, there may be major
consequences for the conservation and economic
value of the lake. For example, a recent study on
another deep rift-lake rich in biodiversity, Lake
Tanganyika, has shown that recent climate warming
has strengthened lake stratification, which has had
a negative impact on nutrient recycling, causing a
decline in algal biomass and subsequent fish stocks
(O’Reilly et al., 2003).
Confounding factors and limitations of study
Linked to variations in diatom assemblages caused by
climate change, are confounding factors such as poten-
tial increases in pollutant input. Pollutant input occurs
L A K E B A I K A L D I A T O M A S S E M B L A G E S 2311
r 2006 The AuthorsJournal compilation r 2006 Blackwell Publishing Ltd, Global Change Biology, 12, 2297–2315

especially through poorly regulated land-use in the
catchment of Lake Baikal, most notably around the
southern basin and the Maloe More region, west of
the central basin (Fig. 1). For example, Popovskaya
(2000) highlights how in the shallow waters of the
Selenga Delta region, standing crops of phytoplankton
are approximately eight times higher in the 1980s than
they were in the 1960s, and that this figure is still
increasing. These large increases are attributed to
anthropogenic eutrophication of the Selenga River
and associated shallow waters of the Selenga Delta.
Evidence of eutrophication is also borne out by the
palaeolimnological record; increases in relative
abundances of small centric taxa more commonly asso-
ciated with nutrient rich waters (e.g. S. minutulus
and A. formosa) were found in a short core extracted
from the shallow waters of the Selenga Delta (fig. 11 in
Mackay et al., 1998). Recent attempts have been
made to model the impact of anthropogenic disturbance
on the lake’s biota using an ecosystem disturbance
model (Silow et al., 2001). Initial results suggest all
components of the planktonic communities become
disturbed, especially under-ice diatom communities,
together with an overall increase in nutrient concentra-
tion and increase in summer phytoplankton biomass.
These changes are similar to those predicted by
future climate variability and, therefore, it is now
important that refined models are developed to take
account of interactions between localized eutrophica-
tion and climate change impacts on the lake’s
ecosystem.
In Lake Baikal, diatom growth is only limited by Si
every third or fourth year, when there is a high abun-
dance of A. baicalensis (so-called Melosira years). While
the specific causes for Melosira years are still uncertain,
their occurrence is influenced by the interaction bet-
ween its life cycle and factors that influence population
growth, such as nutrients and grazing. We acknowl-
edge, therefore, the likelihood that our interpretations
underestimate the full complexity in the Lake Baikal
ecosystem. For example, we have not considered re-
sponses to Si in our training set, which other diatom-
based studies have found to be important (e.g. Ruhland
& Smol, 2002). Furthermore, phosphate and nitrate are
drawn down (by nondiatomaceous phytoplankton)
each year to undetectable concentrations in the upper
layers during summer stratification. Changes in future
climate will undoubtedly affect the availability and use
of nutrients, such as phosphate and nitrate, by aquatic
algae in the lake, and these responses have not as yet
been taken into account. This is partly because our
knowledge of the interactions between Lake Baikal
phytoplankton and nutrients is still relatively poor,
but also because we have not yet been able to compile
modern values for many of these nutrients, including
Si. Moreover, although diatoms responses will vary
spatially between the three lake basins, a single lake-
wide response is unlikely, confounded with the knowl-
edge that for some species in our dataset (e.g. cosmo-
politan taxa such as S. acus), we will not have captured
full environmental gradient responses.
As highlighted above, dissolution of diatom valves is
a significant process in both the water column and the
surface sediment–water interface (Ryves et al., 2003).
Dissolution causes differential preservation of diatom
species in Lake Baikal – for example, C. minuta is
relatively resistant to dissolution, whereas other taxa,
including N. acicularis, S. meyerii and S. acus are very
susceptible to dissolution (Battarbee et al., 2005).
N. acicularis is a common diatom found in Lake Baikal,
especially in the south basin. However, because
virtually no valves preserve in the surface sediments,
we are unable to model impacts of climate change on
this species, and we acknowledge that this is a potential
weakness in our methodology. Nevertheless, all other
taxa are represented, and although differential dissolu-
tion means we cannot provide absolute quantitative
estimates of how diatom assemblages will change
in the future, our qualitative interpretations remain
robust.
In this study, we have demonstrated that spatial
differences in limnological characteristics linked to pre-
vailing climate have a significant impact on the distri-
bution of both endemic and cosmopolitan diatoms in
Lake Baikal. We have modelled these relationships
using multivariate techniques. We have then used these
models to predict impacts of future climate change on
diatoms in this globally significant ecosystem. Our
analyses indicate that the responses to future climate
change are complex, both in terms of impacts on
individual species, but also on basin-wide differences
between the main regions of the lake. Overall, we
predict that by 2100, a decline in ice duration, decreased
snow cover on the lake, increased fluvial input and
increased stratification of the upper waters during
summer months will result in an increase in endemic
and cosmopolitan diatoms which at present dominate
shallow-water, offshore regions. However, these in-
creases will occur at the expense of slow-growing,
cold-water pelagic endemics. Ongoing monitoring of
phytoplankton communities across the lake will pro-
vide strong evidence to test our predictions from sur-
face sediment diatom assemblages and quantitative
data on population change. If our predictions are cor-
rect, such programmes will act as vital early warning
indicators of ecological change in Lake Baikal, and their
continuation needs to be supported by the international
scientific community.
2312 A . W. M A C K AY et al.
r 2006 The AuthorsJournal compilation r 2006 Blackwell Publishing Ltd, Global Change Biology, 12, 2297–2315

Acknowledgements
We would like to thank the various funding agencies that havehelped contribute to this work, especially UK NERC (GR3/10529; NER/S/A/2001/06430), EU FPV Continent (EVK2-CT-2000-0057) and The Royal Society. We thank Nick Granin at theLimnological Institute, Irktusk, with help in accessing relevantRussian information. We thank Pierre Legendre for advice on theuse of multivariate statistics. Finally, we are grateful for thecomments from three reviewers, which have helped to improvethis paper.
References
Aizen EM, Aizen VB, Melack JM et al. (2001) Precipitation and
atmospheric circulation patterns at mid-latitudes of Asia.
International Journal of Climatology, 21, 535–556.
Antoniades D, Douglas MSV, Smol JP (2005) Quantitative esti-
mates of recent environmental changes in the Canadian High
Arctic inferred from diatoms in lake and pond sediments.
Journal of Paleolimnology, 33, 349–360.
Atlas Baikala (1993) Baikal Atlas. Federal Service of Surveying
and Cartography, Moscow (in Russian).
Barnett TP, Adam JC, Lettenmaier DP (2005) Potential impacts of
a warming climate on water availability in snow-dominated
regions. Nature, 438, 303–309.
Battarbee RW, Mackay AW, Jewson D et al. (2005) Differential
dissolution of Lake Baikal diatoms: correction factors and
implications for palaeoclimatic reconstruction. Global and
Planetary Change, 46, 75–86.
Bolgrien DW, Granin NG, Levin L (1995) Surface temperature
dynamics of Lake Baikal observed from AVHRR images.
Photogrammetric Engineering and Remote Sensing, 61, 211–216.
Bondarenko NA (1999) Floral shift in the phytoplankton of Lake
Baikal, Siberia: recent dominance of Nitzschia acicularis.
Plankton Biology and Ecology, 46, 18–23.
Bondarenko NA, Guselnikova NE, Logacheva NF et al. (1996)
Spatial distribution of phytoplankton in Lake Baikal, Spring
1991. Freshwater Biology, 35, 517–523.
Bouchard G, Gajewski K, Hamilton PB (2004) Freshwater diatom
biogeography in the Canadian Arctic Archipelago. Journal of
Biogeography, 31, 1955–1973.
Bradbury JP, Bezrukova YV, Chernyaeva GP et al. (1994) A
synthesis of post-glacial diatom records from Lake Baikal.
Journal of Paleolimnology, 10, 213–252.
Callender E, Granina L (1995) Biogeochemical silica mass bal-
ance in Lake Baikal, Russia. In: Proceedings 8th International
Symposium of Water–Rock Interaction, pp. 341–344. Balkema,
Rotterdam.
Clarke MP, Serreze MC, Robinson DB (1999) Atmospheric con-
trols on Eurasian snow extent. International Journal of Climatol-
ogy, 19, 27–40.
Douglas MSV, Smol JP, Blake Jr W (1994) Marked post-18th
century environmental change in high-arctic ecosystems.
Science, 266, 416–419.
Edlund MB, Jahn R (2001) Report of a workshop on ‘Biogeogra-
phy and endemism of diatoms’. In: Proceedings 16th Interna-
tional Diatom Symposium (ed. Economou-Amilli A), pp.
575–587. Koeltz Scientific Books, Koenigstein.
Edlund MB, Stoermer EF, Taylor CM (1996) Aulacoseira skvortzo-
wii sp. nov. (Bacillariophyta), a poorly understood diatom
from Lake Baikal, Russia. Journal of Phycology, 32, 165–175.
Elliott JA, Thackeray SJ, Huntingford C et al. (2005) Combining
a regional climate model with a phytoplankton community
model to predict future changes in phytoplankton in lakes.
Freshwater Biology, 50, 1404–1411.
Faith DP, Minchin PR, Belbin L (1987) Composition dissimilarity
as a robust measure of ecological distance. Vegetatio, 69, 57–68.
Fietz S, Kobanova G, Izmest’eva L et al. (2005) Regional, vertical
and seasonal distribution of phytoplankton and photosyn-
thetic pigments in Lake Baikal. Journal Plankton Research, 27,
793–810.
Flower RJ (1993) A taxonomic re-evaluation of endemic
Cyclotella taxa in Lake Baikal, Siberia. Nova Hedwigia, 106,
203–220.
Genkal SI, Glezer ZI, Davydova NN et al. (1992) Diatom algae of
the USSR: relict and modern, Vol. II (ed. Makarova IV),
pp. 7–48. Nauka, St. Petersburg (in Russian).
Gleser SI, Makarova, Moisseeva AI et al. (eds) (1988) The Diatoms
of the USSR, Fossil and Recent, Vol. 2. Nauka, St. Petersburg (in
Russian).
Granin NG, Jewson DH, Gnatovsky RY et al. (2000) Turbulent
mixing under ice and the growth of diatoms in Lake Baikal.
Verhein International Vereinigung Limnologie, 27, 1–3.
Heim B, Oberhaensli H, Kaufmann H (2005) Variation in Lake
Baikal’s phytoplankton distribution and fluvial input assessed
by SeaWiFS satellite data. Global and Planetary Change, 46, 9–27.
Huisman J, Olff H, Fresco LFM (1993) A hierarchical set of
models for species response analysis. Journal of Vegetation
Science, 4, 37–46.
IPCC (2001) Climate Change 2001: The Scientific Basis. Working
Group 1 contribution to the IPCC Third Assessment Report
(Summary for Policy Makers) (http://www.ipcc.ch).
Karabanov EB, Prokopenko AA, Williams DF et al. (2000) A new
record of Holocene climate change from the bottom sediments
of Lake Baikal. Palaeogeography, Palaeoclimatology and Palaeoe-
cology, 156, 211–224.
Karst-Riddoch TL, Pisaric MFJ, Smol JP (2005) Diatom responses
to 20th century climate-related environmental changes in
high-elevation mountain lakes of the northern Canadian
Cordillera. Journal of Paleolimnology, 33, 265–282.
Khursevich GK (1989) Atlas of Stephanodiscus and Cyclostephanos,
Bacillariophyta, from the Upper Cenozoic deposits of the
USSR. (ed. Velichkevich FY), 167 pp. Nauka & Teknika, Minsk
(in Russian).
Kilham P, Kilham SS (1990) Endless summer, internal loading
processes dominate nutrient cycling in tropical lakes. Fresh-
water Biology, 23, 379–389.
Kociolek JP, Spaulding SA (2000) Freshwater diatom biogeogra-
phy. Nova Hedwigia, 71, 223–241.
Kouraev A, Semovski SS, Shimaraev MN et al. (in press)
Observations of Lake Baikal ice from satellite altimetry and
radiometry. Remote Sensing of Environment.
Kozhova OM, Izmest’eva LR (1998) Lake Baikal: Evolution and
Biodiversity. Backhuys Publishers, Leiden.
Kozhova OM, Kobanova GI (1995) On the morphology and
systematics of centric diatom algae of Lake Baikal. In: Ecologi-
L A K E B A I K A L D I A T O M A S S E M B L A G E S 2313
r 2006 The AuthorsJournal compilation r 2006 Blackwell Publishing Ltd, Global Change Biology, 12, 2297–2315

cal Problems, Vol. 2 (ed. Kozhova O.M.), pp. 51–52. Nauka,
Novosibirsk (in Russian).
Krenke AN, Chernavskaya MM (2002) Climate changes in the
preinstrumental period of the last millennium and their man-
ifestations over the Russian Plain. Isvestiya, Atmospheric and
Oceanic Physics, 38, S59–S79.
Laing TE, Smol JP (2000) Factors influencing diatom distribu-
tions in circumpolar treeline lakes of northern Russia. Journal
of Phycology, 36, 1035–1048.
Le Core HL (1998) Use of numerical models and satellite data to study
physical processes in Lake Baikal. Unpublished PhD Thesis,
University of Leicester, UK.
Legendre P, Gallagher ED (2001) Ecologically meaningful trans-
formations for ordination of species data. Oecologia, 129,
271–280.
Lim DSS, Kwan C, Douglas MSV (2001) Periphytic diatom
assemblages from Bathurst Island, Nunavut, Canadian High
Arctic: an examination of community relationships and habitat
preferences. Journal of Phycology, 37, 379–392.
Livingstone DM (1999) Ice break-up on southern Lake Baikal and
its relationship to local and regional air temperatures in Siberia
and to the North Atlantic Oscillation. Limnology and Oceano-
graphy, 44, 1486–1497.
Lotter AF, Bigler C (2000) Do diatoms in the Swiss Alps reflect
the length of ice cover? Aquatic Sciences, 62, 125–141.
Lydolph PE (1977) Climates of the Soviet Union. World Survey of
Climatology, Vol. 7. Elsevier, Amsterdam. pp. 91–115.
Mackay AW, Battarbee RW, Flower RJ et al. (2003) Assessing the
potential for developing internal diatom-based inference mod-
els in Lake Baikal. Limnology and Oceanography, 48, 1183–1192.
Mackay AW, Flower RJ, Kuzmina AE et al. (1998) Diatom
succession trends in recent sediments from Lake Baikal and
their relationship to atmospheric pollution and to climate
change. Philosophical Transactions of the Royal Society of London,
353, 1011–1055.
Mackay AW, Ryves DB, Battarbee RW et al. (2005) 1000 years of
climate variability in central Asia: assessing the evidence using
Lake Baikal diatom assemblages and the application of a
diatom-inferred model of snow thickness. Global and Planetary
Change, 46, 281–297.
Magnuson JJ, Robertson DM, Benson BJ et al. (2000) Historical
trends in lake and river ice cover in the northern hemisphere.
Science, 289, 1743–1747.
Martin P, Boes X, Goddeeris B et al. (2005) A qualitative assess-
ment of the influence of bioturbation in Lake Baikal sediments.
Global and Planetary Change, 46, 87–99.
McGowan S, Ryves DB, Anderson NJ (2003) Holocene records of
past effective precipitation in West Greenland. The Holocene,
13, 239–249.
Meyer KI (1930) Introduction into the algae flora of Lake Baikal.
Bulletin of the Moscow Society of Naturalists, 39, 179–396 (in
Russian).
Moline MA, Claustre H, Frazer TK et al. (2004) Alteration of the
food web along the Antarctic Peninsula in response to a
regional warming trend. Global Change Biology, 10, 1973–1980.
Morley DW (2005). Reconstructing past climate variability in
continental Eurasia. Unpublished PhD Thesis, University of
London, UK, 370 pp.
Muller B, Maerki M, Schmid M et al. (2005) Internal carbon
and nutrient cycling in Lake Baikal: sedimentation, upwelling
and early diagenesis. Global and Planetary Change, 46,
101–124.
Ohlendorf C, Bigler C, Goudsmit GH et al. (2000) Causes and
effects of long periods of ice cover on a remote high Alpine
lake. Journal of Limnology, 59, 65–80.
Oksanen J, Minchin PR (2002) Continuum theory revisited: what
shape are species responses along ecological gradients? Ecolo-
gical Modelling, 157, 119–129.
O’Reilly CM, Alin SR, Plisnier PD et al. (2003) Climate change
decreases aquatic ecosystem productivity of Lake Tanganyika,
Africa. Nature, 424, 766–768.
Panagiotopoulos F, Shahgedanova M, Hannachi A et al. (2005)
Observed trends and teleconnections of the Siberian High:
a recently declining center of action. Journal of Climate, 18,
1411–1422.
Popovskaya GI (2000) Ecological monitoring of phytoplankton
in Lake Baikal. Aquatic Ecosystem and Health Management, 3,
215–225.
Richardson TL, Gibson CE, Heaney SI (2000) Temperature
growth and seasonal succession of phytoplankton in Lake
Baikal, Siberia. Freshwater Biology, 44, 431–440.
Riggs G, Hall D, Salomonson V (1994) A snow index for the
Landsat Thematic Mapper and MODIS. In: Proceedings of
IGARSS ‘94 – International Geoscience and Remote Sensing Semi-
nar, pp. 1–10. ESA Publications, Noordwijk.
Robinson DA, Frei A (2000) Seasonal variability of northern
hemisphere snow extent using visible satellite data.
Professional Geography, 52, 307–315.
Ruhland KM, Smol JP (2002) Freshwater diatoms from the
Canadian Arctic treeline and development of paleolimnologi-
cal inference models. Journal of Phycology, 38, 249–264.
Ryves DB, Jewson DH, Sturm M et al. (2003) Quantitative and
qualitative relationships between planktonic diatom commu-
nities and diatom assemblages in sedimenting material and
surface sediments in Lake Baikal, Siberia. Limnology and
Oceanography, 48, 1643–1661.
Ryves DB, McGowan S, Anderson NJ (2002) Development and
evaluation of a diatom-conductivity model from lakes in West
Greenland. Freshwater Biology, 47, 995–1014.
Schindler DW (2001) The cumulative effects of climate warming
and other human stresses on Canadian freshwaters in the new
millennium. Canadian Journal of Fisheries and Aquatic Sciences,
58, 18–29.
Semovski SV, Mogilev NY (2003) The use of satellite imagery
in investigating the dynamics of the ice and snow cover Lake
Baikal. Nordic Hydrology, 34, 33–50.
Semovski SV, Mogilev NY, Sherstyankin PP (2000) Lake
Baikal ice: analysis of AVHRR imagery and simulation of
under-ice phytoplankton bloom. Journal of Marine Systems,
27, 117–130.
Shimaraev MN, Kuimova LN, Sinyukovich VN et al. (2002)
Manifestation of global climatic changes in Lake Baikal during
the 20th century. Doklady Earth Sciences, 383A, 288–291.
Shimaraev MN, Verbolov VI, Granin NG et al. (1994) Physical
Limnology of Lake Baikal. A Review (eds Shimaraev MN, Okuda
S), 80 pp. BICER Publishers, Irkutsk and Okayama.
2314 A . W. M A C K AY et al.
r 2006 The AuthorsJournal compilation r 2006 Blackwell Publishing Ltd, Global Change Biology, 12, 2297–2315

Shindell DT, Schmidt GA, Mann ME et al. (2001) Solar forcing
of regional climate change during the Maunder Minimum.
Science, 294, 2149–2152.
Silow EA, Baturin VA, Stom DJ (2001) Prediction of Lake
Baikal ecosystem behaviour using an ecosystem disturbance
model. Lakes and Reservoirs: Research and Management, 6,
33–36.
Skabichevskii AP (1960) Planktonic diatoms of the freshwaters of the
USSR: Systematics, Ecology & Distribution. MSU, Moscow (in
Russian).
Skvortzov BV (1937) Bottom diatoms from Olkhon Gate of Baikal
lake, Siberia. Philippine Journal of Science, 62, 293–377.
Smol JP, Wolfe AP, Birks HJB et al. (2005) Climate-driven regime
shifts in the biological communities of arctic lakes. Proceedings
of the National Academy of Sciences, 102, 4397–4402.
Solovieva N, Jones VJ, Nazarova L et al. (2005) Palaeolimnological
evidence for recent climatic change in lakes from the northern
Urals, arctic Russia. Journal of Paleolimnology, 33, 463–482.
Sorvari S, Korhola A, Thompson R (2002) Lake diatom response
to recent Arctic warming in Finnish Lapland. Global Change
Biology, 8, 153–163.
Srivastan KS, Singh RP (1991) Microwave radiometry of snow-
covered terrains. International Journal of Remote Sensing, 12,
2117–2131.
Tarasova EN, Mescheryakova AI (1992) Modern State of the
Hydrochemical Regime of Lake Baikal. Nauka, Novosibirsk (in
Russian).
ter Braak CJF, Looman CWN (1994) Biplots of reduced-rank
regression. Biometrical Journal, 36, 983–1003.
ter Braak CJF, Smilauer P (2002) Canoco for Windows 4.5.
Biometrics, the Netherlands.
Todd MC, Mackay AW (2003) Large-scale climate controls on
Lake Baikal ice cover. Journal of Climate, 16, 3186–3199.
Verbolov VI, Sokolınikov VN, Shimaraev MN (1965) Hydrome-
teorological Conditions and Heat Balance of Lake Baikal. Nauka,
Moscow.
Votinsev KK, Mescheryakova AI, Popovskaya GI (1975)
Organic Matter Turnover in Lake Baikal. Nauka, Novosibirsk
(in Russian).
Weckstrom J, Korhola A (2001) Patterns in the distribution,
composition and diversity of diatom assemblages in relation
to ecoclimatic factors in Arctic Lapland. Journal of Biogeography,
28, 31–45.
Weiss RF, Carmack EC, Koropalov VM (1991) Deep-water
renewal and biological production in Lake Baikal. Nature, 349,
665–669.
Winder M, Schindler DE (2004) Climate change uncouples
trophic interactions in an aquatic ecosystem. Ecology, 85,
2100–2106.
Wuest A, Ravens T, Granin NG et al. (2005) Cold intrusions
in Lake Baikal: direct observational evidence for deep-water
renewal. Limnology and Oceanography, 50, 184–196.
Ye HC (2001) Quasi-biennial and quasi-decadal variations in
snow accumulation over northern Eurasia and their connec-
tions to the Atlantic and Pacific Oceans. Journal of Climate, 14,
4573–4584.
Yoshii K, Melnik NG, Timoshkin OA et al. (1999) Stable isotope
analyses of the pelagic food web in Lake Baikal. Limnology and
Oceanography, 44, 502–511.
Zabelina MM, Kiselev IA, Proshkina-Lavrenko AI et al. (1951)
Diatom Algae (ed. Proshkina-Lavrenko AI), 108 pp. Sovetsya
Nauka, Moscow (in Russian).
L A K E B A I K A L D I A T O M A S S E M B L A G E S 2315
r 2006 The AuthorsJournal compilation r 2006 Blackwell Publishing Ltd, Global Change Biology, 12, 2297–2315
Copyright © 2022 FDOKUMEN




















