Aspects Récents de la Biologie des Crustacés - Archimer
235
ACTES de COLLOQUES n° 8 -1988 CONCARNEAU, 6-9 juin 1987 ASPECTS RÉCENTS de la BlOLOGIE des CRUSTACÉS Sous la direction de Yves Le GAL - Alain VAN WORMHOUDT Laboratoire Biologie Marine Collège de France-CONCARNEAU IFREMER
-
Upload
khangminh22 -
Category
Documents
-
view
0 -
download
0
Transcript of Aspects Récents de la Biologie des Crustacés - Archimer

ACTES de COLLOQUES n° 8 -1988
CONCARNEAU, 6-9 juin 1987
ASPECTS RÉCENTS
de la
BlOLOGIE des CRUSTACÉS
Sous la direction de Yves Le GAL - Alain VAN WORMHOUDT
Laboratoire Biologie Marine Collège de France-CONCARNEAU
IFREMER

2 L
ACTES de COLLOQUES
8-1988
ASPECTS RÉCENTS
de la
BlOLOGIE des CRUSTACÉS
10e réunion des Carcinologistes de langue française Concarneau, 6-9 juin 1987
Sous la direction de Yves Le GAL - Alain VAN WORMHOUDT
Laboratoire Biologie Marine du Collège de France-CONCARNEAU
IFREMER

LA 10ème RÉUNION DES CARCINOLOGISTES DE LANGUE FRANÇAISE s'est tenue au
Laboratoire de Biologie Marine de Concarneau du 6 au 9 juin 1987
LE COMITÉ D'ORGANISATION était composé de
Chantal BELLON Brest (29) Auguste LE ROUX Bailleron (56) Geneviève PAYEN Paris
Yves LE GAL Concarneau (29) Alain VAN WORMHOUDT Concarneau (29)
Ouvrage édité par L'IFREMER
avec le concours financier du COLLEGE DE FRANCE
Service de la Documentation et des Publications (S D P)
IFREMER - Centre de Brest B.P. 70 - 29263 PLOUZANÉ
Tél. : 98.22.40.13 - Telex 940 627 F
ISSN 0761.3962 © IFREMER Institut Français de recherche pour l'exploitation de la mer, 1988

AVANT PROPOS
Les développements récents de la biologie nous ont accoutumés à ne plus
considérer le monde vivant qu'à travers quelques procaryotes et rares
vertébrés. Un peu de recul nous permet, cependant, de prendre rapidement
conscience de la diversité réelle des situations évolutives ou écologiques
d'un monde où, à tout prendre, les vertébrés supérieurs que nous sommes
apparaissent relativement marginaux.
L'observation d'autres groupes zoologiques conduit à analyser en détail des
particularités physiologiques, des types d'organisation, des stratégies
adaptatives, des répartitions écologiques qui enrichissent singulièrement
notre connaissance du vivant.
L'apport des recherches portant sur la biologie des Crustacés est, à cet
égard, très significatif, qu'il s'agisse des problèmes de morphogénèse, de
régulations hormonales ou nerveuses, d'adaptation à l'environnement, le monde
des Crustacés nous offre des exemples de ces modèles particulièrement
pertinents dont sont friands les biologistes modernes.
Gardons nous cependant d'une vision trop morcelée du fonctionnement d'un
système vivant. La connaissance que nous avons d'un organisme ou d'un type
d'organisation doit être globale car nous devrons être en mesure d'en
apprécier toutes les caractéristiques du fonctionnement, y compris dans ses
interactions avec son environnement. De la molêcule à l'écosystème, il y a
une continuité, ou un gradient régulier, d'enchaînements du métabolisme,
des régulations de communications. Comprendre le vivant, c'est prendre la
mesure de l'ensemble.
Cette approche globale qui n'exclut pas une analyse fouillée des phénomènes,
est bien celle des Carcinologistes de Langue Française dont la Xème réunion a
eu lieu en Juin 1987 au Laboratoire de Biologie Marine du Collège de France à
Concarneau. C'est tout à l'honneur du professeur DRACH et des collègues
d'avoir su, voici vingt ans, créer une dynamique assurant une cohésion de
fait dans des études aussi diverses. Les discussions qui ont enrichi cette
réunion sont l’illustration du bien fondé de cette idée.
Cinq grands thèmes ont été abordés au cours de cette réunion : Morphogénèse
et développement, Hormones et métabolisme, contrôles et physiologie
adaptative, populations et enfin exploitation, chaque thème étant introduit
par une conférence destinée à faire le point sur la question ou sur une
avancée particulière dans le domaine considéré.
Y. LE GAL
A. VAN WORMHOUDT
3

HOMMAGE AU PROFESSEUK A. BAUCHAU ( t 2novembre 1987)
C'est au retour d'un voyage d'étude en Polynésie française (Antenne de Moorea dépendant du Museum National d'Histoire Naturelle de Paris) que le Professeur Adrien Bauchau, biologiste et carcinologue aux Facultés Universitaires Notre-Dame de la Paix de Namur (Belgique), nous a quittés brusquement.
Il était né à Anvers le 11 mars 1918. Ses notes de lycéen témoignent très tôt d'un esprit observateur et curieux de tout. II entre à 19 ans chez les Jésuites et, après une licence en Philosophie (1942), il prépare brillamment la licence en Zoologie (1946) et un doctorat en Sciences (1948) à l'Université de Louvain. Sa thèse réalisée sous la direction du Professeur Koch porte sur le rôle physiologique de la glande sinusaire chez Eriocheir sinensis H.M. Edw. Les travaux de cette période sont couronnés par 3 distinctions : le Prix Empain (1949), le Prix de l'Académie des Sciences de Belgique (concours annuel, 1949) et le Prix du concours des Bourses de voyage du gouvernement Belge (1951). Après une licence en Théologie (1952), il séjourne deux années aux U.S.A. dont une comme Research Fellow dans le laboratoire du Professeur William de l'Université de Harvard. Là, en collaboration avec le Docteur Bliss, il effectue des recherches sur la physiologie de la croissance des Crustacés. Le Docteur Gabe (Paris) identifie à cette époque l’organe Y, ce qui ouvre des perspectives quant au contrôle hormonal de ce phénomène. De retour à Namur où il devient professeur de Biologie et Endocrinologie comparée (1954), il poursuit des recherches sur le contrôle neuroendocrinien, via l'organe X et la glande sinusaire, de diverses activités enzymatiques associées aux phénomènes de croissance et de mue.
Des travaux ultérieurs porteront sur le contrôle endocrinien de la glycémie, sur la structure et les fonctions des hémocytes. Plus récemment, il entreprit des recherches sur la chémoreception (phéromones) en liaison avec la reproduction, sur le contrôle du mimétisme des crabes du genre Hippa et sur les structures adaptatives des crabes "éboueurs" du genre Scopimera.
Ceux et celles qui ont vécu avec lui ou l'ont rencontré resteront marqués par ce scientifique de haut niveau doublé d’une personnalité attachante, constamment en éveil et à la recherche de réponses aux questions de notre temps.
4

PARTICIPANTS
ANSENNE A. GAUDY R. LIGNON J.
BAUCHAU A. GERVASI E. MOCQUARD J.P.
BELLAN-SANTINI D. GOFFINET G. MOULINS M.
BOUCHON D. GUILLAUME J. NAGY F.
BOULIGAND Y. HAFIDI N. PEQUEUX A.
CASANOVA B. HERNANDORENA A. PROULX D.
CASTEL J. HUGUET D. RAIMON R.
CAZALETS J.R. JEUNIAUX Ch. SECRETAN S.
CHARFI-CHEIKHROUHA F. JUCHAULT P. SOUTY-GROSSET C.
CHENTOUFI A. JUGAN P. THOREZ A.
COMPERE Ph. KALLEN J. TOULEMONT A.
COUTURE P. KERAMBRUN P. TURQUIN-BUFFIERE M.J.
DAUBY P. LANGIS R. VAN HERP F.
DAUVIN J.C. LAUBIER A. VAN WORMHOUDT A.
DE LA NOUE J. LAUBIER L. VASSEROT J.
FEURTET A. LE ROUX A. WANSON S.
5

Cet ouvrage illustre certains des développements récents de la biologie des Crustacés tels qu'ils ont été abordés lors de la Xème réunion des Carcinologistes en Juin 1987, au Laboratoire de Biologie Marine de Concarneau.
Des articles de fond servent d'ossature aux communications présentées dans cinq domaines : Morphogénèse et développement, hormones et métabolisme, contrôles et physiologie, population et exploitation des espèces.
L'ensemble vise à donner les éléments d'une vision d'ensemble de la biologie d'un groupe aussi homogène dans son organisation que divers dans ses adaptations que l'est celui des crustacés.
Cet ouvrage s'adresse à tous ceux dont l'intérêt se porte vers la biologie, l'écologie des espèces mais aussi à ceux dont les objectifs sont plus directement associés à la gestion des ressources vivantes.
This volume aims to survey some of the recent advances in the Biology of Crustacea presented during the 10th meeting of Carcinology in June 87 at the Marine Station of the Collège de France, in Concarneau.
Background papers and reports account with five main thema : Morphogenesis and development, hormones and metabolism, controls and physiology, populations and exploitation of living ressources.
Throughout, the emphasis is on giving a global insight on a zoological group homogenous at the organization level whereas adaptatively diversified.
This volume should appeal to those interested in biology, ecology of the crustacean species and also to those more directly involved in the management of living ressources.
6

THEME 1. MORPHOGENESE 11
- Problèmes de morphogénèse cuticulaire chez les Crustacés.
Bouligand Y. 13
- Production de chitine par les Crustacés du zooplancton de la baie de Calvi (Corse) .
Gervasi E., Jeuniaux Ch. et Dauby P. 33
- Organisation, sclérotisation et composition minérale des sclérites d'une espèce d'Isopode marin et de quatre espèces d'Isopodes terrestres oniscoïdes.
Ansenne A., Compère Ph. et Goffinet G. 39
- Mise en place de l'épicuticule chez le Crabe Carcinus maenas
Compère Ph. 47
- Aspects ultrastructuraux de la biodégradation de l'ancienne cuticule de Carcinus maenas durant la période préecdysiale.
Thorez A., Compère Ph. et Goffinet G. 55
- Action des chocs thermiques sur le développement d' Artemia .
Hernandorena A. 65
- Progression de l'organisation phragmale du métamère thoracique de quelques malacostracés.
Casanova B., Huguet D. et Secrétan S. 69
THEME 2. HORMONES ET METABOLISME 83
- La neuroendocrinologie des Crustacés : Evaluation des progrès obtenus ces dernières années.
Van Herp F. 85
- Isolement et caractérisation d'une protéine membranaire ovocytaire capable de fixer spécifiquement la vitellogénine.
Jugan P. et Van Herp F. 101
7

- Quelques aspects de la régulation du système neuroendocrine produisant la CHH et de la relation entre le rythme circadien et la glycémie.
Kallen J. 105
- Effets dè l'ablation des pédoncules oculaires, au cours des stades zoe III et IV sur la raue de métamorphose de Palaemonetes varians (Leach) (Crustacea, Decapoda).
Le Roux A. 109
- Purification des peptides apparentés à la calcitonine de saumon chez les Crustacés. Résultats préliminaires.
Van Wormhoudt A. 113
THEME 3. ECOPHYSIOLOGIE. ADAPTATION 121
- Contrôle neuromodulateur et flexibilité d'expression d'un réseaux neuronal central : le système pylorique des crustacés décapodes.
Nagy F. , Cazalets J.R. et Moulins M. 123
- Approche écophysiologique et ultrastructurale de l'osmorégulation chez les Crustacés décapodes.
Pequeux A. et Lignon J. 137
- Flux unidirectionnels d'ions Na et C1 dans les branchies isolées et perfusées du crabe vert Carcinus maenas .
Wanson S. et Pequeux A. 149
- Variation de la consommation d'oxygène au cours du nycthémère chez Armadillidium vulgare
Chentoufi A. et Mocquard J.P. 153
- Etude des facteurs de l'environnement (température et photopériode) sur la reproduction de différentes souches d' Armadillidium vulgare .
Souty-Grosset C., Mocquard J.P. ,Juchault P. et Hafidi N. 159
- Ecophysiologie de la reproduction de la crevette des marais Palaemonetes varians
Bouchon D. 165
- Structures adaptatives à la filtration des aliments chez le crabe des sables Scopimera gordonae
Bauchau A. 171
8

9
THEME 4. POPULATIONS 175
- Biologie et génétique des populations de Crustacés : stratégies mises en jeu. Essai de synthèse.
Gaudy R. et Kerambrun P. 177
- Etude du polymorphisme enzymatique dans cinq populations naturelles d' Armadillidium vulgare
Raimon R. 191
- Les âges de Niphargus virei Turquin-Buffière M.J. 195
- Complément à la définition " Idotea chelipes , espèce polytypique" à partir des fractions protéiques et particulièrement de l'hémocyanine. Comparaison avec Idotea balthica basteri .
Charfi-Cheikhrouha F. 199
- Reproduction et croissance chez les femelles du Crustacé oniscoïde Armadillidium vulgare après inoculation d'une bactérie symbiotique féminisante.
Juchault P. et Mocquard J.P. 203
- Actualisation des données sur l'écologie, la biogéographie et la phylogénie des Ampeliscidae (Crustacés, Amphipodes) atlantiques après la revision des collections d' E. chevreux .
Bellan-Santini D. et Dauvin J.C. 207
- Données préliminaires sur les Amphipodes de l'Iroise et de ses abords, leurs affinités géographiques.
Dauvin J.C. et Toulemont A. 217
- Biologie du Copepode Eurytemora affinis hirundoides dans la Gironde : données morphométriques.
Feurtet A. et Castel J. 223

THEME 5. EXPLOITATION 229
- L'exploitation mondiale des Crustacés : Bilan et perspectives.
Laubier A. et Laubier L. 231
- L'aquaculture des crevettes, rôle de la nutrition.
Guillaume J. 241
- Perspectives d 'accroissement de la production de l'océan Atlantique par l'introduction d'espèces de Crustacés comblant des niches écologiques vides.
Vasserot J. 247
- Contribution d'un film bactérien à la culture intensive de Daphnia magna .
Langis R., Proulx D., de la Noüe J. et Couture P. 251
10

THÈME 1 - MORPHOGÉNÈSE
- Problèmes de morphogénèse cuticulaire chez les Crustacés.
Bouligand Y.
- Production de chitine par les Crustacés du zooplancton de la baie de Calvi (Corse).
Gervasi E., Jeuniaux Ch. et Dauby P.
- Organisation, sclérotisation et composition minérale des sclérites d'une espèce d'Isopode marin et de quatre espèces d'Isopodes terrestres oniscoïdes.
Ansenne A. , Compère Ph. et Goffinet G.
- Mise en place de l'épicuticule chez le Crabe Carcinus maenas
Compère Ph.
- Aspects ultrastructuraux de la biodégradation de l'ancienne cuticule de Carcinus maenas durant la période préecdysiale.
Thorez A., Compère Ph. et Goffinet G.
- Action des chocs thermiques sur le développement d' Artemia .
Hernandorena A.
- Progression de l'organisation phragmale du métamère thoracique de quelques malacostracés.
Casanova B., Huguet D. et Secrétan S.
11

ASPECTS RÉCENTS de la Biologie des CRUSTACÉS Actes de Colloques - 8 - 1988 - IFREMER, pages 13 à 32
PROBLEMES DE MORPHOGENESE CUTICULAIRE CHEZ LES CRUSTACES
BOULIGAND Y.
RESUME - Le tégument des Crustacés est un excellent modèle pour la morphogenèse cuticulaire, aux divers niveaux d'organisation. A l'échelle supramoléculaire, les sous-unités chitino-protéiques s'assemblent selon une organisation spécifique de type cristal liquide stabilisé. A l'échelle des cellules et des populations cellulaires, interviennent divers contrôles, celui de l'architecture fine de l'épicuticule, celui de la distribution du minéral et celui des caractères globaux tels que les isométries et les allométries de croissance.
MOTS CLES - Métrique de la cuticule, Listes de priorité, Cristaux liquides stabilisés, Minéralisation, Artéfacts.
ABSTRACT - The crustacean integument is an excellent model for the study of cuticular morphogenesis, at different organization levels. At the supermolecular level, subunits of chitin and protein assemble and show the organization of a stabilized liquid crystal. At higher levels corresponding to cells and cells populations, several controls are involved in the fine architecture of the epicuticle, in the mineral distribution and in characters which concern the whole animal, such as isometric or allometric growth.
KEY WORDS - Cuticle metrics, Priority lists, Stabilized liquid crystals, Mineralization, Artifacts.
INTRODUCTION - Les progrès en morphogenèse depuis une dizaine d'années doivent beaucoup à la renaissance de la génétique de la Drosophile et, en particulier, à la découverte de gènes impliqués dans la segmentation et dans la différenciation des métamères (Revues dans Spierer et Goldschmidt-Clermont, 1985, ainsi que dans Gehring, 1987). Les caractères étudiés sont repérables dans la morphologie externe et comportent une part importante de leur expression au niveau cuticulaire. Il faut donc s'attendre à un renouvellement de nos connaissances sur la cuticule des Arthropodes au cours de la prochaine décennie. Les Crustacés tiendront une place originale au sein de ce courant et il convient de faire le point des problèmes de morphogenèse cuticulaire tels qu'ils se posent aujoud'hui. Nous les examinerons à divers niveaux d'organisation, en soulignant les principaux écueils à surmonter, notamment les variations insoupçonnées de l'épiderme et des structures cuticulaires, ainsi que les artéfacts.
Laboratoire d'Histophysique et de Cytophysique de l'E.P.H.E., Centre de Biologie Cellulaire du C.N.R.S., 67, rue Maurice-Günsbourg, 94200 Ivry-sur-Seine (F.)
13

MORPHOLOGIE GENERALE - Chez les Crustacés Décapodes, après le développement larvaire, les jeunes accèdent à leur forme presque définitive, en dépit de certaines allométries très sensibles, comme
l'écart relatif des yeux plus prononcé en début de croissance. Pour
d'autres régions cependant, il y a presque similitude. Le crabe qui se
défait d'une ancienne carapace en a élaboré une nouvelle de forme presque identique, la différence portant principalement sur la taille. Avant la
mue, les cellules épidermiques se divisent et leur effectif est pratiquement doublé. Cette "crise mitotique” s'accompagne d'un décollement entre l'ancienne cuticule et l'épiderme; celui-ci se plisse et produit une nouvelle cuticule souple et plus étendue. La cytologie de l'épiderme qui sécrète cette nouvelle cuticule est d'autant plus complexe, que vient se surajouter à l'activité de synthèse du système chitinoprotéique, la production, avant la mue, de particules chargées de la résorption de l'ancienne cuticule. Nous appelons "exolysomes" ces corpuscules extracellulaires qui permettent la lyse plus ou moins prononcée des matrices extracellulaires. Dans le cas qui nous intéresse, ces particules comportent probablement des chitinases et des enzymes protéolytiques et ont l'allure de minuscules liposomes de 120 nm de diamètre environ, à une seule bicouche. Leur production paralt assurée par certaines cellules épidermiques. Quelques micrographies illustrent les principes de ce renouvellement cuticulaire (Fig. 1 et 2).
Lors de l'exuviation, le crabe absorbe une grande quantité d'eau qui passe dans la cavité générale et gonfle le corps, en déplissant progressivement la jeune cuticule; ceci aide aussi à rejeter l'ancienne carapace. La minéralisation commence quand le déplissement est achevé (Drach, 1939) . Le gonflement hydrique ne produit aucune extension de la cuticule nouvellement élaborée, parce que les forces mises en jeu sont loin d'atteindre le seuil à partir duquel une telle extension serait possible. La carapace a donc été sécrétée sous forme plissée, en ayant d'emblée sa forme définitive, aux plis près. Les crabes savent se préparer un habit plus grand sous un vêtement ancien, devenu trop petit, et donnent ainsi une illustration des talents de géomètre de leurs cellules épidermiques. La sécrétion d'une cuticule à l'état plissé, avec la métrique correcte, n'est pas un mince problème.
Figure 1 - Coupe de cuticule de Carclnus maenas en cours de renouvellement (D2) . Un spécimen de 1,2 cm de large au niveau du céphalothorax est examiné dans sa région sub-branchiale latérale. La couche pigmentaire (pp) est en cours de sécrétion. Des pinnules sont observées en section (ps) ou
en vue longitudinale (pl) . Le liquide exuvial renferme des exolysomes
(exl). L'épicuticule comporte deux couches, l’une externe très mince (ee)
et l'autre interne (ei), plus épaisse et munie de racines (re) s'enfonçant
plus ou moins dans l'épaisseur de la procuticule. Les strates de la
cuticule pigmentaire présentent une zone de forte distorsion D. A la base
de la procuticule, on observe un niveau d'assemblage des fibrilles (na).
Le cytoplasme épidermique renferme divers corpuscules et vésicules (c et
v), des épaississements (e) de la membrane plasmique, des configurations
d' exocytose (ex), des microtubules (mt) et leur regroupement en
tonofibrilles (tf). Un paysage cytologique très différent est observé lors
de la sécrétion de la couche principale. Fixation: glutaraldéhyde, acide
osmique; contraste: acide phospho-tungstique; barre: l µm. La préparation
du matériel est la même dans les deux planches suivantes (Fig. 2 et 4).
14

15

Imaginons deux feuilles de papier, l'une ordinaire, s'appliquant sur un plan, et l'autre de même matière et même épaisseur, que l'on aurait obtenue (par un procécé peu usuel) sur un moule sphérique. Si nous chiffonnons ces deux feuilles, rien de leur aspect ne les distingue plus. Au contraire, si nous les déplissons, l'une d'elles s'applique sur un plan et non l'autre, parce que les deux feuilles ont conservé leur métrique.
De même, une carapace de crabe présente des régions relativement planes et d'autres qui ne sont pas applicables sur le plan. Quand la nouvelle carapace se déplisse au cours de la mue, elle conserve sa métrique. Par conséquent, la cuticule est sécrétée sur un moule épidermique plissé, avec une métrique définie. Pour revenir à nos feuilles de papier, on aurait pu imaginer de les fabriquer elles-aussi sur des moules complexes figurant des plis. Dans une telle situation, il serait très difficile d'obtenir des papiers qui se développent sans problème sur une surface plane ou sur une surface sphérique. C'est pourtant ce que sait faire l'épiderme d’un crabe, lorsqu'il sécrète une nouvelle carapace en prévision de la mue. Les choses peuvent être examinées au microscope et on contate que les plis ne sont pas aléatoires. Dans la région dorsale, la carapace est ornée de nombreuses petites intumescences ou tubercules et, chez Carcinus maenas par exemple, la plupart des plis sont orientés en formant des lignes plus ou moins concentriques autour des axes des tubercules. Entre les tubercules eux-mêmes, la distribution de plis paraît souvent aléatoire.
Chez les Insectes, des plis analogues existent, notamment chez la punaise Rhodnius prolixus. Ils sont visibles au microscope avant le repas de sang et disparaissent avec la réplétion. Leur distribution est pratiquement aléatoire et c'est dans l'état plissé que la cuticule est secrétée (Wigglesworth, 1945). Au contraire, lorsqu'on observe l'éclosion d'un papillon, les ailes sont très finement plissées selon un ordre régulier, avec des plis équidistants se recoupant souvent à angle droit.
Le problème posé est de comprendre comment la métrique de la cuticule est maintenue à travers ce processus de croissance par mue, qui exige un plissement de l'épiderme, selon des orientations aléatoires ou non. L'exemple des crabes est utile, parce que l'on observe facilement dans l'ancienne et la nouvelle carapaces les traces des limites des cellules épidermiques qui les ont sécrétées. On peut comparer des régions homologues de l'ancienne et de la nouvelle cuticule, mais la mise en correspondance précise des cellules-mères et cellules-filles est souvent
Figure 2 - Quelques aspects des processus d'exolyse. Spécimens et préparation comme dans la Fig.1. a) Le liquide exuvial renferme des exolysomes (exl) et, sous l'épicuticule (ep) avec ses pinnules (p) et la couche pigmentaire (pp), certaines cellules épidermiques renferment des corps à exolysomes ou supposés tels (ce). b) Grandissement plus élevé des deux corps à exolysomes (exl) de la figure précédente et la membrane enfermant ces corps (mce). c) Espace exuvial entre l'ancienne et la nouvelle cuticule (ac, nc) de la partie latérale de la cavité branchiale. La cuticule dilacérée (cd) forme une masse importante et on observe un pli P de la nouvelle cuticule encore très mince. d) Espace exuvial dans la région sub-branchiale, entre l'ancienne cuticule (ac) et la nouvelle (nc). Les exolysomes (exl) en présence de la couche membraneuse intacte (mi) ou dilacérée (md) . Chaque barre correspond à un µm.
16

17

difficile ou même impossible et il faudra trouver des marqueurs cellulaires. Dans l'épiderme, qui est un épithélium simple à cellules presque isodiamétriques, il suffit, pour que la métrique de la cuticule soit conservée d'un stade au suivant, que la distribution des fuseaux mitotiques soit isotrope dans le plan même de l'épiderme. L'orientation d'un fuseau s'identifie à celle du segment qui joint les centres de deux cellules soeurs. On repère parfois assez bien les couples de cellules issus d'une même cellule mère. L'examen comparé des mosaïques cellulaires de l'épiderme devrait, grâce aux traces laissées dans les cuticules successives, permettre d'analyser l'origine des allométries. Plusieurs facteurs peuvent être impliqués et, en particulier, la perte de l'isodiamétrie des cellules et l'anisotropie de distribution des fuseaux mitotiques, mais rien n'est encore démontré. Les méthodes automatiques d'analyse des textures et des réseaux seront de précieux auxiliaires dans l'étude des mécanismes de l'isométrie ou de l'allométrie et des comportements des populations cellulaires qui en sont à l'origine.
MORPHOLOGIE DE SURFACE - Le dépôt de l'épicuticule intervient juste après l'apolyse, lors des premières étapes de la phase D chez les Crustacés, et coïncide avec la mise en place de la morphologie. II y a maintien de certaines structures du stade précédent, à l'échelle près, avec parfois introduction de nouveautés importantes ou, au contraire, disparition de structures antérieures. Il faut souligner que c'est la mince couche de surface de l'épicuticule qui est la première dépositaire de la morphologie externe. Les principaux caractères différenciés à ce niveau concernent la segmentation du corps et celle des appendices, la distribution des soies et l'élaboration des structures ornementales de surface, examinées notamment par Bocquet et al. (1976) et par Renaud (1977) . Cette couche externe de l'épicuticule est élaborée initialement sous forme de plaques qui deviennent rapidement jointives. Ces plaques apparaissent au sommet de replis ou de microvillosités de la membrane plasmique externe des cellules épidermiques. La membrane présente un épaississement assez semblable à un hémidesmosome sous chaque plaque épicuticulaire en différenciation (Bazin, 1977). L'intervention du cytosquelette est manifeste dans cette initiation de la morphogenèse cuticulaire et mérite une étude immunocytochimique, aussi bien à l'échelle ultrastructurale qu'à celle de la microscopie photonique, afin d'avoir une vue globale des phénomènes. Ces structures proches d'hémidesmosomes furent observées initialement chez les Insectes par Rinterknecht et Lévi (1966) et analysées plus en détail par Locke et Huie (1979).
Peut-on espérer, dans un avenir pas trop éloigné, relier ces processus morphogénétiques intervenant au niveau du cytoplasme épidermique le plus externe et les mécanismes génétiques du développement tels que l'étude de la Drosophile permet de les envisager aujourd'hui? Une amorce de réponse vient de certaines convergences apparaissant entre les résultats génétiques récents, les concepts classiques de l'embryologie causale et les travaux de Grandjean sur la segmentation et les phanères à caractère aléatoire dans un groupe d'Acariens du sol. L'évocation de ces travaux nous éloignerait des Crustacés, mais indiquons seulement que plusieurs méthodes furent conçues autrefois par Grandjean (1942, 1943) pour établir
des listes de priorité entre organes cuticulaires parfaitement identifiés, dont la variation se fait par présence-absence. Elles sont basées sur la
morphologie comparée soit des espèces, soit des stades du développement, en appliquant des calculs statistiques d'un genre inédit. Ces listes
18

mériteraient d'être appelées "arbres de priorité", parce que les calculs permettent de définir un graphe de hiérarchies entre organes qui a la forme d'un arbre. Ce graphe rappelle celui des inductions hiérachisées qui interviennent dans le développement embryonnaire.
Les hiérarchies mises en évidence par Grandjean à partir des seules données du phénotype cuticulaire sont également du même type que celles découvertes aujourd'hui entre les gènes de morphogenèse chez la Drosophile. Cette concordance doit faire espérer des progrès importants dans l'analyse des mécanismes morphogénétiques très en aval du génôme lui-même. Je ne présenterai pas plus en détail cette contribution de Grandjean, analysée dans un récent travail (Bouligand, 1988).
Les Arthropodes présentent une morphologie définie avec une grande rigueur géométrique et tout indique qu'ils constituent de nos jours l'un des meilleurs matériels d'analyse expérimentale de la morphogenèse. Le tégument des Crustacés paraît être un bon candidat dans cette perspective. Les méthodes introduites par Grandjean leur sont parfaitement applicables. Toutefois, les cycles ne sont pas aisément reproductibles au laboratoire et c'est un handicap pour les expériences de génétique. De nos jours, cependant, les méthodes de clonage permettent de manipuler les gènes et une partie importante des processus morphogénétiques peut être examinée en culture organotypique. L'épiderme et la cuticule des Décapodes ont donc toutes leurs chances dans la compétition.
LA STRUCTURE STRATIFIEE ET SES CONSTANTES - Malgré des variations déconcertantes, qui seront examinées plus loin, la structure de la matrice organique de la cuticule offre deux constantes au niveau supramoléculaire. II s'agit d'abord de l'ultrastructure du complexe chitine-protéine (Fig.3, a-d) , sous forme de baguettes claires réunies par un ciment sombre, structure commune aux trois couches pigmentaire, principale et membraneuse, en dépit de très différentes proportions de la chitine (Giraud-Guille, 1984a). Cette structure en nid d'abeilles est horizontale, contrairement à celle de l'épicuticule, puisque les fibrilles de la procuticule sont, pour la plupart, parallèles au plan des strates. Des recherches en contraste de diffraction électronique confirment la présence de la chitine au niveau des éléments clairs aux électrons (Chanzy et Giraud-Guille, 1988), comme l'avait proposé autrefois Rudall (1965). L'origine de la structure en nid d'abeille n'est pas connue et diverses hypothèses ont été discutées (Giraud-Guille et Bouligand, 1986).
La deuxième constante structurale est la présence d'une structure torsadée, déterminant un aspect caractéristique en coupe oblique, avec des séries superposées d'arceaux emboîtés et de même orientation (Bouligand, 1965). L'étude stéréologique de ces motifs obtenus en section a montré que les fibrilles chitino-protéiques sont horizontales, c'est à dire parallèles au plan de la cuticule, alignées parallèlement entre elles à chaque niveau horizontal, avec une orientation qui tourne progressivement quand on change de niveau (Fig.3, e-h). L'empilement hélicoïdal obtenu est gauche, c'est à dire que les fibrilles vues de l'extérieur tournent dans le sens des aiguilles d'une montre, quand on change progressivement de niveau, en allant de l'intérieur vers l'extérieur de la cuticule (Bouligand, 1965). Les cônes d'incertitude dessinés autour des directions moyennes indiquent que l'ordre n'est pas rigoureux et qu'il existe certaine déviations aléatoires. Schmidt avait proposé ce modèle hélicoïdal
19

dès 1924, à partir d'observations au microscope polarisant, mais avait attribué les structures en arceaux à des fibrilles supplémentaires reliant des niveaux différents et assurant un surcrolt de cohésion. II existe bien des fibrilles verticales plus ou moins développées au sein des canalicules verticaux, qui jouent ce rôle, en n'ayant cependant rien à voir avec les séries d'arceaux. La structure torsadée est différente chez les Copépodes (Fig.3,f,g) et la rotation des fibrilles y est discontinue, avec de minces couches différenciées, entre lesquelles on observe des angles de 20° environ (Bouligand, 1965); les détails ultrastucturaux sont indiqués sur la Fig.3,h. Un ciment sombre contient les sous-unités chitino-protéiques entre des baguettes claires plus larges.
Les deux constantes d'organisation supramoléculaire des Crustacés, le nid d'abeilles du complexe chitine-protéine et la torsion, se retrouvent chez presque tous les autres Arthropodes. La structure torsadée est comparable géométriquement à la disposition mutuelle des molécules ou des polymères dans les cristaux liquides cholestériques, ce qui suggère l'intervention d'un autoassemblage de type cristallin liquide (Bouligand, 1967-1972) . Rappelons que le terme cholestérique vient du fait que les premiers cristaux liquides de ce type furent observés avec des dérivés du cholestérol. De nos jours, les exemples de cristaux liquides cholestériques sont connus des composés chimiques les plus variés. Alors que de nombreux analogues biologiques de cristaux liquides ont pu être reproduits par autoassemblage in vitro, (Revue dans Livolant, 1984 et Bouligand et al., 1985), aucun progrès n'a pu être accompli dans ce sens pour la cuticule et ce sont vraisemblablement des difficultés biochimiques très réelles qui doivent être incriminées.
La chitine est un polysaccharide azoté, dont on commence à connaltre des solvants (Rutherford, 1978). Certains dérivés comme l'hydrohypropyl-chitosane forment des phases cristallines liquides (Ogura et al., 1982). Les recherches sont plus avancées sur la cellulose, un autre polysaccharide fondamental de la biosphère, qui forme souvent des séries d'arceaux d'origine cholestérique dans les parois de jeunes cellules
Figure 3 - Le complexe chitine-protéine forme un tube dont la partie axiale, claire aux électrons, correspondrait à la chitine et la périphérie dense aux protéines. Ces sous-unités de 5 nm de diamètre s ' alignent dans la cuticule de Carcinus maenas en ménageant certains espaces accessibles à la minéralisation. a) Dans la couche pigmentaire externe, elles se groupent en fibrilles séparées, de section souvent complexe. b) Dans la couche pigmentaire profonde, les sous-unités chitino-protéiques s'alignent en formant une structure réticulée en section. c) Dans la couche principale, elles donnent des fibrilles séparées, isodiamétriques. d) Dans la couche membraneuse, l'alignement est dense et ne ménage pas d'espaces suffisants pour la nucléation cristalline (a-d: d'après Giraud-Guille, 1984). e) Distribution des directions fibrillaires à plus grande échelle. L'orientation fibrillaire tourne de manière sénestre le long d'une normale
à la cuticule. Les cônes indiquent la dispersion des fibrilles autour de
leur direction moyenne et la rotation est continue. f,g,h) Chez les
Copépodes, la rotation se fait par niveaux discrets, selon des angles B,
B'... variables, de 20' environ, de même sens. Des baguettes claires sont
séparées par un ciment sombre. Celui-ci apparaît, à fort grandissement,
constitué de sous-unités chitino-protéiques jointives, ménageant des
espaces libres trop exigus en général pour être accessibles à la calcite.
20

21

végétales, ou dans la tunique de certaines Ascidies (Bouligand, 1972). Parallèlement, in vitro, certains dérivés de la cellulose comme l'hydroxypropyl-cellulose sont connus pour former des phases cholestériques (Werbowyj et Gray, 1976). De toute façon, la découverte de ces polymères cristallins liquides ne résoud pas entièrement le problème de l 'autoassemblage, tel qu'il se produit à la base de la cuticule au cours de sa croissance en épaisseur. La formation des fibrilles chitinoprotéiques comporte une série d'étapes complexes, visualisées par les ultrastructures (Bouligand, 1975) et il est probable que les filaments les plus récemment déposés, non encore agrégés, correspondent à la sécrétion d'un polymère cristallin liquide, mais le caractère fluide est rapidement aboli par le regroupement en fibrilles, favorisé par la cristallisation de la chitine et la formation de liaisons covalentes, processus qui stabilisent la matrice ainsi constituée. Rappelons l'existence sous la cuticule en cours de sécrétion d'un niveau d'assemblage des fibrilles chitino-protéiques, simple pour la couche pigmentaire (Fig. 1) et différencié en quatre stades distincts pour la couche principale (Fig.4d et Bouligand, 1975).
Des microvésicules baptisées chitosomes, et contenant des chitine synthétases, ont été découvertes chez certains champignons (Bracker et al., 1976). La croissance de fibrilles de chitine est obtenue à partir de ces chitosomes isolés in vitro mals, à notre connaissance, ces travaux n'ont pas été reproduits chez les Crustacés et les Arthropodes, même si le rôle des membranes paraît démontré (Horst, 1983) . La synthèse des fibrilles de chitine se produit à l'extérieur des cellules, au contact de la membrane épidermique, souvent au niveau de différenciations denses, semblables aux hémidesmosomes évoqués plus haut à propos de l'épicuticule, ce qui suggère une localisation membranaire des synthétases, à préciser.
VARIATIONS DES STRUCTURES TEGUMENTAIRES - La difficulté essentielle de l'étude du tégument est sa diversité insoupçonnée. On observe déjà en surface, au niveau épicuticulaire, en microscopie à balayage, des ornements très différenciés selon les régions du corps. II existe plusieurs types de cuticule, notamment la cuticule dure et minéralisée, par opposition à celle des articulations. Dans la cuticule minéralisée, il faut distinguer celle où sont insérés les muscles, cette cuticule étant soit externe, soit interne, sous forme d'apodème ou d'endophragme. II y a également la cuticule externe minéralisée sans insertions musculaires. Même dans ce type de cuticule apparemment homogène, les différenciations sont importantes, selon la région et l'âge. Pour la trame organique, elles portent sur plusieurs points.
1. Le mode d’agrégation des fibrilles chitino-protéiques en faisceaux dans la couche pigmentaire. (Les diverses textures qui en résultent sont représentées sur la Fig. 3). 2. La proportion des fibrilles verticales présentes au sein des canalicules verticaux, entre les microvillosités et les fibrilles horizontales. 3. Le développement des cloisons interprismatiques, sortes d'impressions laissées par les limites des cellules épidermiques dans les strates les plus externes de la couche pigmentaire.
II est à noter que l'apolyse et la formation des divers niveaux de la trame organique de la carapace peuvent être légèrement décalés les uns par
22

rapport aux autres suivant les régions où on les considère. De même, pour la minéralisation, Drach (1939) avait montré les variations de texture de la mosaïque calcitique, selon le niveau dans la cuticule et la région du corps. II en avait précisé la chronologie, en mettant en évidence des gradients. Les régions antérieures du corps, notamment celles en saillie et les extrémités distales des appendices sont les premières calcifiées.
Les observations sont aisément reproductibles à condition de préciser l'espèce, bien entendu, la région du corps examinée, le stade dans le cycle d'intermue et la largeur du céphalothorax, donnant une indication sur l'âge. On observe en effet, dans des régions comparables, des différences sensibles entre de très jeunes individus et des adultes pris au même stade du cycle d'intermue. Quelques exemples vont souligner cette diversité.
L'épicuticule interne est constituée d'une structure en nid d'abeilles, comprenant des baguettes claires aux électrons, orientées normalement au plan cuticulaire et réunies par un ciment plus opaque. Des images bien résolues en ont été obtenues par Compère et Goffinet (1987 a) , chez Carcinus maenas. Je pense utile de préciser que cette structure est observée chez les individus de grande taille, dont le céphalothorax excède 3 cm par exemple, et n'apparaît pas jusqu'ici chez les jeunes où la largeur est de 1 cm environ. Elle n'y est pas nécessairement absente mais, dans les mêmes conditions de préparation, le contraste obtenu n'est pas le même. Cela correspond probablement à des différences biochimiques, liées au développement.
Les canaux et canalicules qui traversent verticalement la carapace des Arthropodes furent observés par les premiers histologistes, notamment Vitzou (1882) chez les Crustacés Décapodes. Leur caractère rubanné hélicoïdal fut reconnu par Schmidt (1924) et rappelé par Drach (1939). Les canalicules renferment des prolongements cytoplasmiques des cellules épidermiques, comme en témoignent de nombreuses micrographies ultrastructurales (Bouligand, 1966 a, 1971). Quelques illustrations en sont proposées sur la Fig.4. Les membranes qui limitent ces microvillosités présentent des différenciations denses aux électrons (Fig.4a), distribuées de manière assez variée, à l'instar des épaississements denses de la membrane plasmique, où s'élaborent les premiers filaments chitino-protéiques. Les canalicules sont absents en général de la cuticule des petits Crustacés et, en particulier, de celle des Copépodes (Bouligand, 1966 b), mais il y a des exceptions.
Les estimations données par les auteurs sur la densité, par unité de surface, des canalicules présents au sein de la cuticule sont contradictoires et Compère et Goffinet (1987 b) signalent que la raison en
est le manque d'indications précises sur les régions étudiées. II faut mentionner aussi l'importance de l'âge. En effet, lorsque des études suffisantes auront été accomplies, ces différences seront probablement interprétées en recourant à des arguments dimensionnels concernant les
rapports surface sur volume, très diminués chez les grands individus, et
corrigés par le plus grand développement des microvillosités
intracanaliculaires. Compère et Goffinet montrent sur de beaux clichés
l'existence de branches horizontales des microvillosités cytoplasmiques
(1987 a), mais cette structure présente à ma connaissance des variations
avec l’âge. En effet, on l'observe chez les grands individus et rarement
23

chez les petits, ce qui suggère que ces expansions horizontales des microvillosités interviennent chez les jeunes de manière brève par rapport à la durée de la phase de formation de la couche pigmentaire. Compère et Goffinet (1987 a, Fig.30) suggèrent également que ces branches horizontales permettent la ramification des canalicules, mais la démonstration reste à faire. Un point essentiel est que ces expansions horizontales déterminent la texture de la couche pigmentaire, décrite en détail par Giraud-Guille (1984) . Les fibrilles chitino-protéiques sont déposées entre ces expansions cytoplasmiques horizontales. Celles-ci se retirent et laissent libre un espace, qui sera occupé ultérieurement par le minéral, lors de sa mise en place après la mue.
Les variations tégumentaires les plus saisissantes sont celles des cellules épidermiques. II en existe de nombreux types et leur cytologie présente des changements appréciables, selon les régions, le stade dans le cycle d'intermue et l'âge. Lorque je préparais ma thèse, j'avais constaté, par extrapolation, qu'il me fallait probablement deux siècles pour explorer de manière simplement descriptive l'ensemble des structures tégumentaires de Carcinus maenas, avec le rythme et le degré de précision que j'avais adoptés. En termes plus réalistes, une équipe de vingt chercheurs est nécessaire pendant dix ans. Je ne connais pas de biologistes cellulaires conscients du travail qui les attend. L'épiderme des Insectes m'a toujours semblé moins diversifié, mais ce n'est peut-être qu'une apparence. La cuticule des Insectes paraît elle aussi homogène dans sa structure, mais cela tient souvent à la grande densité de la matière organique et donc à une difficile pénétration des fixateurs et des contrastants. Quand l'examen est plus approfondi, on découvre également chez les Insectes de nombreux thèmes de variations.
MORPHOGENESE DE LA MOSAIQUE CALCITIQUE - La minéralisation de la carapace des Décapodes est initiée en des régions et à des niveaux déterminés de la cuticule (Drach, 1939; Bouligand, 1970). Elle est contrôlée par une série de mécanismes très différents dans leurs principes, puisqu'ils résultent de l'intervention d'enzymes telles que les phosphatases alcalines (Travis, 1965) et l'anhydrase carbonique, dosée et localisée par Giraud-Guille (1977 a,b, 1981, 1984 b). Le rôle des phosphatases serait de couper les ions phosphates qui, en se liant au réseau de calcite, peuvent inhiber sa croissance (Simkiss, 1964) . De plus, divers processus électrochimiques
Figure 4 - Variations ultrastructurales des canalicules verticaux de la carapace de Carcinus maenas. a) Aspect classique des sections rectangulaires des prolongements cytoplasmiques, où la membrane présente une forte densité aux électrons, en formant souvent deux zones opposées (flèche). b) Le clivage local déterminé par la présence d'un prolongement cytoplasmique, disparu ici, peut contenir des fibrilles chitinoprotéiques verticales (fv) et on observe en (hv) un passage continu entre fibrilles horizontales et fibrilles verticales. c) Dans une ancienne cuticule en phase D2, on observe parfois, au niveau de la couche principale, la présence de très fins microcanalicules (mc). Certaines microvillosités de dimensions usuelles (cvd) sont chargées d'un matériel dense. d) Coupe longitudinale de la microvillosité d'un canalicule vertical au niveau de son lien avec la cellule épidermique. Quatre stades distincts sont observés dans l'assemblage des fibrilles (1,2,3,4). Des vésicules (v) sont présentes sous la membrane plasmique externe de l'épiderme. Les barres représentent l mµ.
24

25

jouent un rôle déterminant (Digby, 1967), ainsi que certaines particularités de la trame organique qui conduisent à la formation de dislocations-vis dans le réseau cristallin de calcite, ce qui accélère probablement la croissance du minéral (Giraud-Guille, 1984 c) . Ces mécanismes sont souvent débattus comme s'il s'agissait d'hypothèses pouvant s'exclure, alors qu'ils semblent plutôt coexister et oeuvrer en synergie (Revue dans Giraud-Guille, 1980) .
L ' un des facteurs déterminants de la minéralisation est l'espace disponible à la croissance cristalline. La matrice organique doit ménager des espaces dont les dimensions dépassent 300 Â (Bouligand, 1970). Cela résulte du principe selon lequel un germe cristallin n'a de chance de grandir que si ses dimensions excèdent certaines valeurs critiques et s'il y a sursaturation. Cela exclut la calcite de l'épicuticule et de la couche membraneuse, contrairement à ce qui est lu dans Roer et Dillaman (1984) et Dennell (1960). L'absence de calcite dans l'épicuticule est visible dans l'une de mes micrographies publiées, mais on l'observe dans les interstices de procuticule situés entre les racines de l'épicuticule (Bouligand, 1970) et dans certains espaces libres conduisant aux épines épicuticulaire (Hegdahl et al., 1977c). La croissance de la mosaïque calcitique ne modifie pas l'espace entre les fibrilles et ne provoque pas de changement de volume appréciable sur coupe. Le front de minéralisation ne détermine aucune distorsion des strates (Bouligand, 1970).
LA QUESTION DES ARTEFACTS - La cuticule est un matériau dur, difficile à bien fixer et à inclure, présentant des propriétés mécaniques originales. II en résulte des artéfacts aussi variés que méconnus. A ce jour, on peut en reconnaltre quatre types liés à la fixation, à la déminéralisation, à la déshydratation et à la microtomie. Même avec une grande rigueur sur l'âge, le stade dans le cycle d'intermue et le choix de la région, les prolongements cytoplasmiques des canalicules sont plus ou moins conservés, selon la pénétration des fixateurs. II faut souligner la qualité de la fixation dans les travaux récents de Compère et Goffinet (1987,a,b) et le progrès parait venir de ce qu'ils fixent longtemps, avant et après la déminéralisation. Pour celle-ci, les moyens violents doivent être proscrits et j’ai vérifié que les "macrofibres" décrites par Dennell, qui utilise du HCl (1973), sont en fait des tubes artéfactuels créés par le dégagement du gaz carbonique.
L'alcool éthylique utilisé pour les déshydratations introduit des cristallisations artéfactuelles, étudiées autrefois par Prenant (1927) . Dans les cuticules en début de calcification, la présence des carbonates est reconnue par l'effervescence aux acides; l’état cristallisé l'est par la polarisation chromatique observée au microscope polarisant. Chez Carcinus maenas, la mosaïque est initiée sous forme de disques de calcite sous l'épicuticule et est bien reconnaissable au microscope polarisant, sur un tégument fraîchement prélevé. La déshydratation par l'alcool éthylique fait apparaitre, autour de chaque disque, des auréoles de minuscules cristaux constitués probablement de calcite. Ces microcristaux sont également reconnaissables dans les ultrastructures. Des formations comparables ont été décrites comme représentant les premières phases de la minéralisation (Travis 1963, Yano 1975, Hegdahl, 1977b) et méritent une discussion plus approfondie. En fait, chez Carcinus maenas, toutes les observations au microscope polarisant sur matériel frais montrent l'existence de fronts de minéralisation bien tranchés. De plus, on
26

comprendrait mal comment des microcristaux naissant séparément,
formeraient ultérieurement de grands domaines avec des distributions
continues des axes cristallographiques de la calcite. Ces mosaïques sont
bien définies au microscope polarisant, sur les cuticules en début de
minéralisation, comme nous l'avons déjà indiqué, mais aussi dans la
carapace entièrement calcifiée, sur coupe, comme en témoignent nos micrographies reproduites dans le livre de Neville (1975). Pour obtenir
une distribution continue des axes de la calcite sur de grands domaines,
il faudrait que les microcristaux originels suivent les orientations de la
trame organique. II se trouve au contraire que la mosaïque calcitique est relativement indépendante du réseau organique. Les axes isotropes ou axes c de la calcite sont généralement parallèles aux couches torsadées et sont
donc horizontaux comme les fibrilles, sans leur être parallèles pour
autant.
Des images d'agrégats cristallins à la surface ou à l'intérieur des
villosités des canalicules furent publiées par Travis (1970) sans se poser
la moindre question sur leur origine éventuellement artéfactuelle.
Celle-ci est probable puisque, dans la cuticule entièrement minéralisée en
C4, la calcite est déposée entre les fibrilles verticales des canalicules,
et non au sein des villosités elles-mêmes (Bouligand 1970, Hegdahl 1977
a) . De plus, en C4, on peut creuser la carapace d'un crabe, sans la
percer, avec une fraise de dentiste, en formant un alvéole qui atteint
seulement les strates externes de la couche principale. Après quinze
jours, les coupes histologiques montrent la présence d'un matériel qui
couvre partiellement le fond de l'alvéole et dont les affinités tinctoriales sont semblables à celles de l'épicuticule. Ce matériel est
présent dans certains des canalicules qui ont dû le véhiculer. II est donc difficile d'imaginer la minéralisation des structures cytoplasmiques intracuticulaires, si elles demeurent ainsi fonctionnelles au sein de la carapace achevée et calcifiée.
De nombreux artéfacts sont liés à la microtomie. Pour les coupes de matériels non minéralisés ou déminéralisés, des bandes sombres courant parallèlement aux séries d'arceaux ont été interprétées comme des
structures physiquement ou chimiquement différenciées, non seulement chez
les Crustacés, mais chez bien d'autres Arthropodes. En fait il s'agit d'un
artéfact qui tient fondamentalement à des variations d'épaisseur de coupe.
Le couteau rencontre des fibrilles dont j'orientation varie périodiquement
en offrant une résistance également périodique. Cela détermine des sortes
de marches d'escalier à la surface des coupes. Ces marches étant décalées
entre les deux faces des coupes, il en résulte des bandes épaisses
alternant avec des bandes minces. Plusieurs études détaillées de cet
atéfact ont été réalisées (Bouligand 1972, 1986; Giraud-Guille 1986).
Pour ce qui concerne le matériel calcifié, sa fragilité est évidente et,
en coupe, on obtient le plus souvent de fines baguettes parallèles
distribuées entre les fibrilles. Ainsi, les axes longitudinaux de calcite,
parallèles aux fibrilles, décrits par Hegdahl (1977 a) , ne correspondent
en rien à des axes cristallographiques. Cet aspect résulte des fractures
innombrables, introduites par le couteau du microtome au sein du minéral,
entre les canaux occupés par les fibrilles. Cependant, les meilleures
coupes montrent de vastes régions avec continuité du minéral entre les
f ibrilles. II y a lieu aussi de rappeler une propriété connue, et non
exclusive, de la calcite, qui est de former des mâcles mécaniques
27

(Friedel, 1910, p.226, 1964, p.423), ce qui change l'orientation des axes cristallographiques après coupe, détail qui n'a jamais inquiété les spécialistes de la biominéralisation! Les diagrammes de diffraction électronique donnent par conséquent des indications probablement erronées. Ces diagrammes demeurent utiles en révélant l'existence de domaines où les axes varient sans discontinuités. Par contre, les discontinuités mises en évidence proviennent soit d'authentiques discontinuités de la texture originale au sein de la cuticule, soit de discontinuités surajoutées par les effets mécaniques de la microtomie.
PERSPECTIVES - L'intérêt des Crustacés Supérieurs dans l'étude biochimique est évident, ne serait-ce que par la masse de la cuticule de certains Décapodes, comparée à celle des Insectes. Parmi les problèmes actuels, soulignons les difficultés rencontrées par les biochimistes dans l'étude des protéines cuticulaires, dues aux liaisons covalentes qui les lient de manière intime à la chitine, aussi bien chez les Insectes que chez les Crustacés. La voie la plus prometteuse est de cloner les gènes des protéines cuticulaires et ce travail est possible dès aujourd'hui chez les Décapodes. Des recherches considérables devraient donc être développées dans cette voie, au cours des prochaines années, parce que les complexes chitino-protéiques ont un grand avenir industriel. Les populations cellulaires épidermiques peuvent être suivies de stade en stade et des informations devraient en être retirées sur les mécanismes de l'isométrie et de l'allométrie. Les architectures dessinées au niveau de l'épicuticule méritent une étude spéciale dans le cadre des conceptions de Grandjean, qui permettent d'entrevoir un lien avec les données récentes de la génétique du développement. Bien des expériences sont à envisager sur des cultures organotypiques. La cuticule des Crustacés est également l'un des modèles biologiques fondamentaux de structure minéralisée et la formation de la mosaïque calcitique est un aspect rarement évoqué de la morphogenèse.
Bazin F. Action inhibitrice de l'ecdystérone sur la régénération chez le crabe Carcinus maenas (L.). Comptes Rendus Acad. Sci. Paris, 284, D : 1211-1214 (1977).
Bocquet C., Cals P. et Renaud B. Ordonnance et variations locales des populations cellulaires épidermiques de Carcinus maenas (L.) (Crustacé Décapode) . Comptes Rendus Acad. Sc. Paris, 283, D: 349-352 (1976) .
Bouligand Y. Sur une architecture torsadée répandue dans de nombreuses cuticules d'Arthropodes. Comptes Rendus Acad. Sci. Paris, 261, 3665-3668 (1965) .
Bouligand Y. La géométrie des constituants cuticulaires chez les Crabes à l'échelle des ultrastructures. 6th Internat. Congr. Electr. Microsc., Kyoto, 2: 577-578 (1966 a).
Bouligand Y. Le tégument de quelques Copépodes et ses dépendances musculaires et sensorielles. Mém. Muséum Hist. Nat., Nouvelle Série, A, Zoologie, 40, (4), 189-206 (1966 b).
Bouligand Y. Comparaison de certains matériels biologiques à la structure des cristaux liquides cholestériques. J. Microscopie, 6: 41 a (1967).
Bouligand Y. Aspects ultrastructuraux de la calcification chez les crabes. 7ème Congr. Internat. Microsc. Electr., Grenoble, 3:105-106 (1970).
Bouligand Y. Les orientations fibrillaires dans le squelette des Arthropodes. I. L'exemple des Crabes, l'arrangement torsadé des strates. J. de Microscopie, 5: 305-322 (1971).
28

Bouligand Y. Twisted fibrous arrangements in biological materials and cholesteric mesophases. Tissue 4 Cell, 4: 189-217 (1972).
Bouligand Y. Defects and textures in cholesteric analogues given by some biological systems. J. de Physique, 36 C1: 331-336 (1975).
Bouligand Y. Theory of microtomy artefacts in arthropod cuticle. Tissue 4 Cell, 18: 621-643 (1986).
Bouligand Y. La priorité des organes selon François Grandjean: une articulation précise entre ontogenèse et phylogenèse. Ontogenèse et Evolution. Coll. Internat. C.N.R.S. Editions du C.N.R.S. (sous presse) .
Bouligand Y., Denèfle J.-P., Lechaire M. et Maillard M. Twisted architectures in cell-free assembled collagen gels: study of collagen substrates used for cultures. Biol. Cell, 54: 143-162 (1985).
Bracker C.E., Ruiz-Herrera J. et Bartnicki-Garcia S. Structure and transformation of chitin synthetase particles (chitosomes) during microfibril synthesis in vitro. Proc. Nat. Acad. Sci., 73: 4570-4574 (1976) .
Chanzy H et Giraud-Guille M.-M. Communication personnelle (1988). Compère P. et Goffinet G. Ultrastructural shape and three-dimensional
organization of the intracuticular canal systems in the mineralized cuticle of the green crab Carcinus maenas. Tissue 4 Cell, 19: 839-857 (1987 a) .
Compère P. et Goffinet G. Elaboration and ultrastuctural changes in the pore canal system of the mineralized cuticle of Carcinus maenas during the moulting cycle. Tissue 4 Cell, 19: 859-875 (1987).
Dennell R. Integument and exoskeleton. The Physiology of Crustacea. Waterman T.H. ed., Acad. Press, N.-Y. et Londres, 1: 455-463 (1960).
Dennell R. The structure of the cuticle of the shore-crab Carcinus maenas (L.). Zool. J. Linn. Soc., 52: 159-163 (1973).
Digby P.S.B. Calcification and its mechanism in the shore crab Carcinus maenas (L.). Proc. Linn. Soc. Lond., 178: 129-146 (1967).
Drach P. Mue et Cycle d'intermue chez les Crustacés Décapodes, Ann. Inst. Océanogr., Paris (N. S.) 19: 103-391 (1939).
Friedel G. Leçons de cristallographie, 1 vol. 310 p. Hermann, Paris (1911) et nouveau tirage de la 2ème édition. 1 vol. 600 p. A. Blanchard, Paris (1964) .
Gehring W.J. Homeotic genes, the homeobox, and the spatial organization of the embryo. The Harvey Lectures. Series. 81: 153-172 (1987) .
Giraud M.-M. L'anhydrase carbonique et la calcification tégumentaire chez le crabe Carcinus maenas Linné. Comptes Rendus Acad. Sci. Paris, 284: 453-456 (1977 a).
Giraud M.-M. Histochimie des premières étapes de la minéralisation de la cuticule du Crabe Carcinus maenas: tentative de localisation de l'anhydrase carbonique. Comptes Rendus Acad. Sci. Paris, 284: 1541-1544 (1977 b).
Giraud M.-M. Aspects coopératifs de la minéralisation de la carapace des Crustacés. La Morphogenèse de la Biologie aux Mathématiques, Bouligand Y., éd., Recherches Interdisciplinaires, Maloine, Paris: 147-150 (1980).
Giraud M.-M. Carbonic anhydrase activity in the integument of the crab Carcinus maenas during the intermolt cycle. Comp. Biochem. Physiol., 69 A: 381-387 (1981)
Giraud-Guille M.-M. Fine structure of the chitin-protein system in the crab cuticle. Tissue 4 Cell, 16: 75-92 (1984 a).
Giraud-Guille M.-M. Calcification initiation sites in the crab cuticle. The interprismatic septa. An ultrastructura1l oytochemical study. Cell Tissue Res., 236: 413-420 (1984 b).
29

Giraud-Guille, Les matrices extracellulaires analogues de cristaux liquides - structure et biominéralisation - exemple de la cuticule du crabe Carcinus maenas. Thèse d'Etat, Université Paris VI, (1984 c).
Giraud-Guille M.-M. Direct visualization of microtomy artefacts in sections of twisted fibrous extracellular matrices. Tissue & Cell, 18: 603-620 (1986).
Giraud-Guille M.-M. et Bouligand Y. Chitin protein molecular organisation in Arthropods, Chitin in Nature and Technology, Muzzarelli et al. ed., Plenum Publ. Co, 29-35 (1986).
Grandjean F. Les méthodes pour établir les listes de priorité et la concordance de leurs résultats. Comptes Rendus Acad. Sci., Paris, 214: 729-733 (1942).
Grandjean F. Priorité absolue et statistique en biologie. Comptes Rendus Séanc. Soc. Phys. Hist. Nat.Genève, 60: 135-139 (1943).
Hegdahl T., Gustavsen F., Silness J. The structure and mineralization of the carapace of the crab (Cancer pagurus L.) 1. The endocuticle; 2. The exocuticle; 3. The epicuticle. Zoologica Scripta, 6: 89-99; 101-105; 215-220 (1977 a; b; c).
Horst M.N. The biosynthesis of chitin. Isolation and characterization of Polyphenol-linked intermediates from brine shrimps microsomes. Arch. Biochem. Biophysics, 223: 254-263 (1983).
Livolant F. Cholesteric organisation of DNA in vivo and in vitro. European J. Cell Biology, 33: 300-311 (1984).
Locke M. et Huie P. Apolysis and the turnover of plasma membrane plaques during cuticle formation in an insect.Tissue & Cell, 11: 277-291 (1979).
Neville A.C. Biology of the Arthropod Cuticle, Coll. Zoophysiology and Ecology, 4/5: 1 Vol. 448 p., Springer Vlg. (1975).
Ogura K., Kanamoto T, Takanori S., Tanaka K. et Iwakura Y. Liquid crystalline phases based on chitosan and its derivatives, Chitin & Chitosan, Proc. 2nd Internat Conf. : 39-44 (1982) .
Prenant M. Les formes minéralogiques du calcaire chez les êtres vivants et le problème de leur déterminisme. Biol. Rev., 2: 364-393 (1927).
Renaud B. Les régions et les limites de la carapace d'un Crustacé décapode Carcinus maenas (L.) [Brachyrhyncha Portunidae]; leurs rapports avec les systèmes fonctionnels principaux. Bull. Muséum Nat. Hist. Nat. 3e Série, n° 454, Zoologie 317: 569-594 (1977).
Rinterknecht E. et Lévy C. Etude au microscope électronique du cycle cuticulaire au cours du 4ème stade larvaire chez Locusta migratoria. Z. Zellforsch. mikrosk. Anat., 72: 390-407 (1966).
Roer R et Dillaman R. The structure and calcification of crustacean cuticle. Amer. Zool., 24: 893-909 (1984).
Rudall K.M. Skeletal structure in insects. Aspects of Insect Biochemistry (T. W. Goodwin ed.), Acad. Press 83-92 (1965).
Rutherford F.A. Characterisation of marine chitins using a unique inert solvent. Thesis, Univ. of Delaware. Proc. lst Internat . Conf. Chitin/Chitosan. MITSG 78-7, 182.
Schmidt W.J. Die Bausteine des Tierkörpers in polarisierten Lichte. Fr. Cohen Verlag, Bonn, Cf. 247-257 (1924).
Simkiss K. Phosphates as crystal poisons of calcification. Biol. Rev., 39: 487-505 (1964) .
Spierer P. et Goldschmidt-Clermont M. La génétique du développement de la mouche. La Recherche, 16: 452-461 (1985).
Travis D.F. Structural features of mineralization from tissue to macromolecular levels of organization in the decapod crustacea. Ann. N.-Y. Acad. Sci.; 109: 177-245 (1963).
30

Travis D.F. The deposition of skeletal structures in the crustacea. 5. The histomorphological and histochemical changes associated with the development and calcification of the branchial exoskeleton in the crayfish, Orconectes virilis Hagen, Acta histochem., 20: 193-222 (1965).
Travis D.F. The comparative ultrastructure and organization of five calcified tissues. Biological Calcification, Cellular and Molecular Aspects. Schraer H. ed., North Holl. Publ. Co.: 203-311 (1970).
Vitzou A. N., Recherches sur la structure et la formation des téguments chez les Crustacés Décapodes. Arch. Zool. Exptl. et Gén., Sér. I, 10: 451-476 (1882).
Werbowyj R.S.et Gray D. Liquid crystalline structure in hydroxypropyl cellulose solutions. Mol. Cryst. Liq. Cryst. (Letters), 34: 97-103 (1976).
Wigglesworth V.B. Growth and form in an Insect. Essays on Growth and Form presented to d'Arcy Wentworth Thompson. Oxford, at Clarendon Press: 23-40 (1945).
Yano Y. An electron microscope study on the calcification of the exoskeleton in a shore crab. Bull. Japanese Soc. Scient. Fisheries, 41: 1079-1082 (1975).
31

ASPECTS RÉCENTS de la Biologie des CRUSTACÉS Actes de Colloques - 8 - 1988 - IFREMER, pages 33 à 38
PRODUCTION DE CHITINE PAR LES CRUSTACES DU ZOOPLANCTON DE LA BAIE DE CALVI (CORSE).
GERVASI E.(1) JEUNIAUX Ch.(1) OAUBY P. (2)
RESUME - La teneur en chitine a été mesurée dans des échantillons de Crustacés dominant dans le zooplancton de la baie de Calvi. Les pourcentages de chitine, exprimés par rapport au poids sec, s'étagent entre 3,10 % chez Clausocalanus spp. et 12,22 % chez les Cladocères (appartenant principalement au genre Evadne). Nous avons vérifié, dans ie cas d'une espèce de Copépode particulièrement bien représentée, Calanus helgolandicus, que le pourcentage de chitine (3.26 %) est relativement constant au cours des divers stades de développement. Sur base de ces résultats et des valeurs de biomasse et de production globale estimées par Dauby (1985) dans le même golfe de Calvi pendant un cycle annuel (1983-1984), nous avons calculé les biomasses et la production de chitine pour chaque espèce et pour l'en-semble des espèces dominantes des Crustacés du zooplancton. La production jour-nalière de chitine présente un maximum pendant le mois de mai (20.06 mg.m j 1). Pour l'ensemble de l'année (mars 1983 à février 1984), la production de chitine
par le zooplancton a été de 1.0014 g.m
Mots-clés : plancton, production, chitine, Copépodes.
ABSTRACT - Chitin was estimated in samples of planktonic Crustaceans living in
Calvi Bay (Corsica). The percentages of chitin, with respect to dry weight, are comprised between 3.10 % in Clausocalanus spp. and 12.22 % in Cladocera (mainly Evadne). The percentage of chitin remained approximatively constant during larval development in Calanus helgolandicus (3.26 %). Using these results and the values of total biomass and production estimated by Dauby (1985) at the same place during a complete annual cycle (1983-1984), chitin biomass and production were calculated for every planktonic crustacean species (or group of species) and for the whole crustacean plankton. Daily chitin production was maximum during may (20.06 mg.m d_1). As far as the whole year is considered (march 1983 to february 1984), chi-tin planktonic production was estimated to 1.0014 g.m
Key-words : plankton, production, chitin, copepods.
INTR0DUCTI0N - Ce travail s'inscrit dans un programme de recherches sur l'écologie de la chitine, c'est-à-dire sur la place de ce polysaccharide aminé dans les cycles
biogéochimiques du Carbone et de l'Azote. Cette étude se justifie d'autant plus
que la chitine et ses dérivés ont pris récemment un intérêt particulier depuis
qu'on leur a reconnu toute une série de propriétés qui ouvrent des perspectives
d'application tant dans le domaine pharmaceutique et médical qu'en agronomie, dans
l'industrie des textiles et dans celle des films photographiques ou dans le trai-
tement des eaux polluées (Muzzarelli, 1977), comme l'ont confirmé récemment les
travaux de la 3me conférence internationale sur la chitine et le chitosane
(1) : laboratoire de Morphologie, Systématique et Ecologie animales, Institut de Zoologie, Université de Liège, B-4020 LIEGE (Belgique) et Station de Recher-ches Océanographiques STARESO , f-20260 Calvi (Corse, France).
(2) : GHER, Institut de Physique, Bât. B5, Université de Liège, B-4000 Sart: lilman
33

(Muzzarelli, Jeuniaux et Gooday, 1986).
Dans le but d'inventorier diverses sources potentielles de chitine, nous avons mesuré la biomasse et la production de ce polysaccharide dans divers compartiments d'un écosystème marin méditerranéen, en Corse, dans la baie de Calvi (Jeuniaux et al., 1986).
Le présent travail a pour but de calculer la production de chitine par les crustacés planctoniques de la baie de Calvi à partir des données publiées par Dauby (1982, 1985) sur la biomasse et la production zooplanctoniques de ce même écosystème au cours d'un cycle annuel de mars 1983 à mars 1984. Cette utilisa-tion des résultats de Dauby nécessitait la détermination préalable de la teneur en chitine des principales espèces constituant ce zooplancton, et des variations éventuelles au cours du développement larvaire.
SITE ETUDIE, MATERIEL ET METHODES
Les mesures de biomasse et de production globales du zooplancton ont été effectuées (Dauby, 1985) du 1er mars 1983 au 28 février 1984, dans une zone située au nord de la citadelle de Calvi, sur des fonds d'environ 120 mètres. La colonne d'eau prospectée s'étendait de -100 mètres à la surface; les prélèvements ont été réalisés au moyen d'un filet triple de type WP2.
Parmi les espèces dominantes à tégument chitineux, on compte les Copé-podes Claanus heigolandicus, Centropaqes typicus, Acartia clausi, 0ithona spp., Clausocalanus spp., Paracalanus spp. et Ctenocalanus vanus, des Cladocères du genre Evadne et les larves de diverses espèces de Crustacés Décapodes.
Le dosage de la chitine a été réalisé sur des lots de Crustacés (adul-tes ou larves), récoltés au même endroit en mai 1986, triés, rincés à l'eau dis-tillée et séchés sur place à l'étuve à 60 °C. pendant 18 heures puis en exsicca-teur sous vide pendant 1 heure, avant détermination du poids sec. Afin de dispo-ser d'une quantité de matériel suffisante pour permettre une mesure précise du poids sec, nous avons réuni en un seul lot les individus appartenant aux genres Clausocalanus, Paracalanus et Ctenocalanus, en raison de leur grande similitude 'morphologique. Pour la même raison, les deux espèces du genre Clausocalanus (C. arcuicornis et C. furcatus) ont été regroupées, de même que les deux espèces du genre 0ithona (0. nana et 0. helgolandica)■
La chitine a été dosée par une méthode enzymatique spécifique (Jeuniaux, 1963, 1965) adaptée à du matériel planctonique. Après pesée (Balance Mettler M5; sensibilité : 1 pg), le matériel étudié est traité pendant 2 heures par HCl 0.5 N à 100 °C., sous vide, puis, après lavages, par NaOH 0.5N à 100 °C. pendant 6 h. Après lavages à l ' eau distillée, le matériel est incubé pendant 10 h. à 37 °C. dans 2 ml d ' une solution tamponnée (pH 5.2) de chitinase purifiée à 0.5 mg (Chitinase U.S. Biochemical Corporation). Cette chitinase commerciale contenant des N-acétyl-glucosaminidases, l ' hydrolyse enzymatique complète est obtenue au cours d ' une seule incubation enzymatique suivie d'une incubation du culot de centrifugation dans une solution tamponnée. L'acétylglucosamine libérée est mesurée par la méthode colorimétrique de Reissig et al. (1955).
Des essais préliminaires ont montré qu'il est indispensable d'appliquer cette méthode à du matériel fraîchement récolté, lavé et séché. On obtient des résultats aberrants si on utilise du matériel planctonique congelé, ou du matériel formolé et conservé dans le formol depuis plusieurs mois.
34

RESULTATS ET DISCUSSION
1. Importance quantitative de la chitine dans les principaux taxons de Crustacés planctoniques
Le tableau I rassemble les résultats du dosage de la chitine sur des lots fraîchement récoltés et triés de Crustacés planctoniques présents dans la baie de Calvi. Ces résultats concernent des adultes de Copépodes et de Clado-cères, ainsi que des larves de Crustacés Décapodes. On voit que la proportion de chitine, ramenée au poids sec total, varie sensiblement d'un taxon (ou d'un groupe de taxons) à l'autre. Elle va de 3.10 % chez Clausocalanus spp. à 12.22 % chez les Cladocères du genre Evadne. La quantité de chitine par individu adulte varie considérablement d'un taxon à l'autre, notamment en fonction de leur taille.
2. Variation au cours des stades de développement
Pour une mesure précise de la production de chitine, il importe de connaître la proportion de chitine en fonction du poids sec au cours de dévelop-pement larvaire. Cette mesure a été réalisée dans le cas d'une espèce de Copé-pode de taille appréciable, Calanus hegolandicus, dont les cinq stades copépo-dites étaient relativement abondants dans nos échantillons de plancton. Les résultats de ces analyses montrent que, alors que la quantité de chitine par individu double approximativement d'un stade au stade suivant, la proportion de chitine par unité de poids sec reste à peu près constante entre le premier stade copépodite et le stade adulte (3.26 mg ?ô). Nous admettrons, pour une première évaluation de la production planctonique de chitine, qu'il en va de même pour les autres espèces et les autres stades larvaires de Copépodes.
3. Calcul de la production planctonique de chitine
Le calcul de la production de chitine a été effectué en postulant que le pourcentage de chitine en fonction du poids sec ne varie pas au cours de l'année pour une espèce donnée, que celui-ci est constant pendant le développement larvaire, comme l'indiquent nos observations sur Calanus helgolandicus, que la chitine de la vieille cuticule est totalement récupérée avant l'exuviation, et que les oeufs sont dépourvus de chitine.
Sur base des résultats de Dauby (1985), la production de chitine par chaque espèce planctonique au cours du cycle annuel a été calculée par l'équa-
tion p _ Nn.AWn.C
■‘ÏÏT1 tn
où i = le nombre de classes de taille jusqu'au stade adulte, Nn = le nombre d'individus de la classe de taille n,AWn = l ' accroissement moyen de biomasse (poids sec) des individus de la classe n, tn = la durée pendant laquelle un individu reste dans la classe de taille n , et C = le pourcentage de chitine par rapport au poids sec chez l'espèce considérée.
Le tableau I rassembe les valeurs calculées de production annueiie pour les principaux taxons. La figure 1 exprime l'évolution annuelle de la pro-duction journalière de chitine pour l'ensemble des principaux taxons de Crustacés planctoniques dont nous avons déterminé la teneur en chitine.
CONCLUSION
tion 1 z: n=1
La biomasse de chitine zooplanctonique est maximale en avril et en mai,
avec un pic de 414 mg.m-2 au milieu du mois de mai. Pendant le reste de l'année,
les valeurs sont comprises entre 10 et 100 mg.m
La production de chitine présente également un maximum pendant le mois
de mai (20.061 mg par m2 et par jour). Ces valeurs de biomasse et de production
35

sont principalement dûes à l'espèce Centropaqes typicus, de loin la plus abon-dante et dont la teneur en chitine est élevée.
Pour un cycle annuel complet, la production de chitine par les Crustacés planctoniques dans le golfe de Calvi peut être évaluée à 1 g. par m , soit, pour la superficie de 2 200 hectares que représente ce golfe, une produc-tion totale de 22 tonnes de chitine.
Comparée à la production de chitine par les biocénoses benthiques peuplant la côte rocheuse du même golfe de Calvi (Jeuniaux et al., 1986), la production zooplanctonique apparaît donc 10 fois plus élevée.
La forte production printannière de chitine au mois d'avril et au mois de mai permet de proposer des limites précises aux périodes d'exploitation éventuelle du zooplancton méditerranéen pour l ' approvisionnement en chitine à des fins industrielles.
Taxon Chitine en % du poids sec
Chitine µg/individu (adultes)
Production Chitine -2 -1
mg.m an
Calanus helqolandicus 3.26 1.73 22.5
Centropages typicus 7.56 0.59 392.2
Acartia clausi 8.58 0.30 87.8
Clausocalanus spp. 3.10 0.06 184.5
Oithona spp. (5.0)(1) - 50.9
autres Copépodes (5.0)(1) - 168.0
larves de Décapodes 4.44 - 95.5
Cladocères 12.22 0.16 0.0(2)
Tab. 1 : Pourcentage de chitine, quantité de chitine par individu adulte, et production annuelle de chitine par les principaux taxons de Crustacés planctoniques de la baie de Calvi (Corse). (1) : les chiffres entre parenthèses sont des valeurs moyennes, calcu-lées sur base des résultats obtenus pour d'autres espèces. (2) : les populations de Cladocères de la baie de Calvi étant proba-blement d'origine exogène (adultes âgés), la production est considérée comme nulle (Dauby, 1982, 1985).
36

Fig. 1 : Variation annuelle de la production de chitine par les espèces dominantes de crustacés planctoniques dans la baie de Calvi (Corse).
DAUBY P., 1982. Quelques aspects d'un cycle annuel du zooplancton en baie de Calvi (Corse). Ann. 5oc. r. Zool. Belg., 112, 69-77. DAUBY P., 1985. Plankton productivity in the gulf of Calvi (Corsica). Progr. Belgian Oceanogr. Res., Brussels, 442-450. JEUNIAUX Ch., 1963. Chitine et chitinolyse, Paris, Masson éd., 181 p. JEUNIAUX Ch., 1965. Chitine et phylogénie : application d'une méthode enzymatique de dosage de la chitine. Bull. Soc. Chim. Biol., 47, 2267-2278. JEUNIAUX Ch., BUSSERS J.C., VOSS-FOUCART M.F., POULICEK M., 1986. Chitin produc-tion by animals and natural communities in marine environment. in Chitin in Natu-re and Technology, MUZZARELLI, JEUNIAUX and GOODAY editors. Plenum Press, N.Y., 515-522. MUZZARELLI R.A.A., 1977. Chitin, Pergamon Press, Oxford, 309 p. MUZZARELLI R.A.A., JEUNIAUX Ch., GOODAY G., 1986. Chitin in Nature and Technoloqy, Plenum Press, N.Y., 583 p. REISSIG J.L., STROMINGER J.L., LELOIR L.E., 1955. A modified colorimetric method for estimation of N-acetylamino sugars. J. Biol. Chem. , 217, 959.
37

ASPECTS RÉCENTS de la Biologie des CRUSTACÉS Actes de Colloques - 8 - 1988 - IFREMER, pages 39 à 46
ORGANISATION, SCLEROTISATION ET COMPOSITION MINERALE DES SCLERITES D'UNE ESPECE D'ISOPODE MARIN ET DE QUATRE ESPECES D'ISOPODES TERRESTRES ONISCOÏDES.
ANSENNE,A., COMPERE,Ph.(1}
et GOEFINET,G.
RESUME - La cuticule minéralisée de 4 espèces d'Isopodes terrestres (Liqia oceanica, Oniscus asellus, Porcellio scaber et Armadiliidium vuiqare) et d'une espèce d'Isopode marin (Sphaeroma serratum) présente une orqanisation générale répondant au plan fondamental de la cuticule des Crustacés décapodes. De plus, comme chez ces derniers, les couches procuticulaires ne sont pas tannées et les teneurs en sels minéraux, de 60 à 80% du poids sec total, y sont du même ordre de grandeur. Toutefois, la distribution des microfibrilles chitino-protéiques selon un système homogène lâche au niveau de la couche proximale de la procuticule de toutes les espèces étudiées ne permet1aucune comparaison avec la couche principale, ni avec la couche membraneuse non minéralisée des décapodes. D'autre part, la cuticule de S.serratum se singularise de celle des espèces terrestres par son épaisseur (45 µm contre 15 pm environ), une teneur plus élevée en calcium et magnésium, un système fibrillaire associé à la procuticule distale nettement plus lâche, un réseau canaliculaire peu défini et la présence de protéines tannées au niveau des couches de l'épicuticule.
mots-clés : cuticule, minéralisation, sclérotisation, Isopodes.
AB5TRACT - The mineralized tergite cuticle of four terrestrial isopods (Oniscoidea) (Ligia oceanica, Oniscus asellus, Porcellio scaber and Armadillidium vulqare) and of the marine isopod Sphaercma serratum shows a general organization comparable to the fundamental plan of the decapod sclerite cuticle. Moreover, as in decapods, the procuticular layers are not tanned and mineralization rates, from 60 to 80% of the total cuticular dry weight, are of the same importance. However, the chitin protein microfibril distribution according to a homogeneous system in the mineralized procuticular proximal layer in all isopod species so far studied does not allow any compa-rison of this layer with the non mineralized membranous layer, nor with the principal layer of decapods. On the other hand, in comparison with that of terrestrial species, the cuticle of the marine isopod S.serratum exhibits some pecularities with regard to its higher thickness, higher calcium and magnesium contents, a looser fibrillar system in the procuticle distal layer, a comparatively less organized pore canal system and the presence of tanned proteins in the epicuticle layers.
key-words : cuticle, mineralization, sclerotisation, isopods.
(1) Aspirant au Fonds National de la Recherche Scientifique. - Laboratoire de Morphologie animale, Université de Liège, Institut de Zoologie, 22, quai Van Beneden, B-4020 LIEGE (Belgique). - Ce travail a été réalisé grâce au Fonds de la Recherche Fondamentale Collective (convention n°2.4506.83).
39

INTRODUCTION
Avec plus de 4.000 espèces, les Isopodes représentent un des plus grands ordres de Crustacés. Ils constituent un groupe zoologique en quête de milieux très divers : on les rencontre depuis les milieux marins benthiques profonds jusqu'aux milieux dulcicoles et terrestres, sans oublier les formes parasites.
Par rapport aux Décapodes (Bliss, 1968) et aux Amphipodes (Hurley, 1968), les Isopodes Onisco'ides présentent un plus haut niveau d'adaptation à la vie sur terre, mais ils ont poussé cette conquête moins loin, et peut-être plus tard, que les autres groupes d'arthropodes tels les Insectes, les Myriapodes et les Arachnides.
Face aux problèmes que pose la vie en milieu terrestre, les Oniscoides présentent une série de caractères morphologiques, physiologiques et comportementaux qui ont fait l'objet de nombreuses recherches. Par contre, leur tégument est mal connu. Or, il est évident, et les études récentes vont toutes dans ce sens, que la cuticule est appelée à jouer un rôle de première importance dans les échanges entre le milieu externe et l'organisme, s'opposant notamment à la perte d'eau comme c'est le cas chez les Arachnides et la plupart des Insectes.
C'est dans cette optique que la présente étude a été entreprise. En fait, elle tente de répondre aux 3 questions suivantes : a) existe-t-il au niveau de la cuticule minéralisée des Isopodes terrestres certains caractères parti-culiers ultrastructuraux par rapport aux systèmes cuticulaires mieux connus que sont la cuticule minéralisée et peu tannée des Crustacés décapodes et celle, essentiellement tannée, des Insectes; b) existe-t-il chez les Oniscoïdes des couches épicuticulaires spécialisées assurant l'imperméabilisatiqn du tégument et c) en raison du problème de l'approvisionnement en calcium lié à un envi-ronnement terrestre et de la nécessité de posséder une cuticule durcie, leur cuticule serait-elle plus sclérotisée et moins minéralisée que celle des Crustacés décapodes ?
CHOIX DES ESPECES
L'étude porte sur une espèce marine, Sphaeroma serratum, espèce de la zone intertidale récoltée à Roscoff (France), et quatre espèces terrestres. Celles-ci , dans l'ordre décroissant de leur indice de perte d'eau sont respectivement : Liqia oceanica, provenant de l'horizon inférieur de l'étage supralittoral de la Pointe-aux-Oies à Wimereux (Erance), Oniscus asellus et Porcellio scaber, récoltée à Liège (Belgique) et Armadillidium vulgare, récolté à Duren (Rép.Eéd.Allemagne). Toutes les observations ont été réalisées au niveau des 7 premiers tergites sur des individus en intermue.
ORGANISATION ULTRASTRUCTURALE
L'ensemble des observations au microscope électronique à transmission révèle que la cuticule des Isopodes marins et terrestres présente un plan général d'organisation commun répondant au plan général de la cuticule des arthropodes (Fig.l). Elle se compose d'une mince épicuticule superficielle hérissée de microstructures diverses (écailles, épines, ... cfr. Figs.l, 8) La zone procuticulaire sous-jacente, relativement pauvre en éléments
40

fibrillaires (Fig .5, 7) paraît fortement minéralisée, le traitement par les
solutions de décalcification entraînant fréquemment le décollement de l'épicu-
ticule (Fig. . 1, 2, 3). La procuticule lamellaire est subdivisée en deux
couches minéralisées que nous désignerons respectivement procuticule distale et
procuticule proximale de manière à ne pas préjuger de leur homologie avec les
couches procuticulaires des Crustacés décapodes et des Insectes. D'une manière
générale, couche distale et couche proximale sont traversées par un réseau
complexe de canalicules verticaux (Fig .1,2) tel que l'ont récemment décrit
Compère et Goffinet (1987) chez Carcinus maenas. L'épicuticule de toutes
les espèces examinées comprend une épicuticule interne et une épicuticule
externe (Fig . 3, 4). Cette dernière présente une stratiTication nette telle
qu'on peut l'observer chez les décapodes. Elle contient un nombre variable de
strates, 7 chez Oniscus et Liqia, 3 à 4 seulement chez Armadillidium et
Sphaeroma, sans qu'il soit possible de mettre telle ou telle autre organisation
en relation avec les conditions du milieu auquel ces espèces sont inféodées.
L1épicuticule interne présente une structure en tous points comparable : aspect
finement granuleux et homogène et présence de canalicules s1interrompant au
niveau de la première couche de l'épicuticule externe (cuticuline, Fig.4).
Au niveau de la couche procuticulaire distale, d'une épaisseur de 15 jjm
environ, des espèces terrestres, les fibres chitino-protéiques s'associent en
un système réticulé semblable à celui qui caractérise la couche pigmentaire de
Carcinus maenas (Giraud-Guille, 1984). Par contre, dans la couche correspondante
de Sphaeroma, deux à trois fois plus épaisse (45 µm), les fibres forment un
système lâche, traduisant ainsi un taux de minéralisation élevé. De plus, et
contrairement aux espèces terrestres, cette couche paraît dépourvue de canali-
cules (Fig.6). Enfin, au niveau de la couche proximale de toutes les espèces
étudiées, les fibres se distribuent selon un système homogène et lâche. Cette
couche se différencie ainsi de la couche membraneuse non minéralisée des
Crustacés décapodes et de la couche principale où, chez ces derniers, les
éléments microfibrillaires s'associent en macrofibres (Giraud-Guille, 1984).
DISTRIBUTION DU TANNAGE PROTEIQUE (SCLEROTISATION)
A notre connaissance, la littérature ne fournit aucune donnée expérimen-
tale indiquant la présence ou l'absence de sclérotisation chez les Isopodes.
Seuls quelques auteurs (Bocquet-Vedrine, 1976, 1979; Price et Holdich,1980)
en supposent l'existence sans la démontrer.
L'application de divers tests histologiques et histochimiques (réactions
argentaffine de Lillie et Burtner ,1970; mise en évidence des résidus tyrosine
d'origine protéique selon Glenner et Lillie (1970); coloration à l'Azan),révèle
que pour l'ensemble des espèces, seules les couches épicuticulaires sont concer-
nées par le phénomène de sclérotisation. Chez S.serratum, les couches épicuticu-
laires ainsi que les microstructures qui leur sont associées manifestent en
effet une réaction nettement positive, comme on peut l'observer chez les
Crustacés décapodes (Dennell, 1947; Krishnan, 1951; Travis, 1965), alors que
chez les Oniscoïdes, les microstructures paraissent les seules structures
tannées. II ressort de cette analyse qu'on ne peut attribuer une certaine
"imperméabilisation" de la cuticule des Isopodes terrestres au seul phénomène
de sclérotisation. D'autre part, la rigidité et la dureté de leur sclérites
n'apparaissent pas comme étant le résultat d'un tannage de la procuticule.
41

Fiq-1 : Vue générale du tégument de O.asellus. Fiq. 2 : Procuticule distale de L.oceanica. Fig.3 : Epicuticule de O.asellus. Fig.4 : Epicuticule de L■oceanica. ce, canalicule épicuticulaire. cp, canalicule procuticulaire. cu, cuticuline. Ec, écaille d'origine épicuticulaire. Ee, épicuticule externe. Ei, épicuticule interne. Ep, épicuticule. Pd, procuticule distale. Pp, procuticule proximale. Astérisques, zone de décollement entre épicuticule et procuticule.
42

Fig.5 : Procuticule distale de A.vulqare. Fiq. 6 : Procuticule distale de S.serratum. A remarquer l'absence de canalicules. Fiq, 7 : A.vulqare. Epaississement local de la procuticule sous-épicuticulaire au niveau du bord Jatéral d'un tergite.riq.8 : Epine d'origine épicuticulaire de O.asellus. Ep, épicuticule. Pd, procuticule distale. Pp, procuticule proximale.
43

TAUX DE MINERALISATION
L'examen des valeurs du dosage du calcium et de magnesium cuticulaires, exprimées en pourcents du poids sec de cuticule (Tableau 1), révèle que trois espèces d'Isopodes terrestres, L.oceanica, O.asellus et P.scaber, se caracté-risent par des teneurs relatives en calcium identiques (24%).
Tableau I1 : Teneurs en Ca et en Mg dans la cuticule des 7 premiers tergites au stade de l 'intermue chez 5 espèces d'Isopodes(n = 4)(Méthode de dosage par absorption atomique.
Espèce
Teneurs exprimées en % du poids sec total de cuticule
Rapport Ca/Mg (x 100)
Degré de minéralisa-tion (1)
Sphaeroma serratum 31.2 ± 1.2 1.23 ± 0.09 4.0 82.2 i 3.4
Ligia oceanica 24.0 ± 0.8 0.20 ± 0.02 0.8 62.1 ± 2.2
Oniscus asellus 24.0 ± 0.8 0.31 ± 0.03 1.3 61.0 ± 2.1
Porcellio scaber 24.4 ± 1.1 0.46 - 0.08 1.9 62.6 ± 3.3
Arma dillid. vulgare 29.4 ± 1.2 0.44 ± 0.03 1.5 75.1 ± 3.1
(1) Valeurs calculées en supposant que le calcium et le magnésium sont libérés à partir de carbonates.
Ces valeurs sont plus élevées chez l'espèce terrestre à faible indice de perte d'eau, A.vulgare (29.4%) ainsi que chez l'espèce marine, S.serratum (31%). Les teneurs relatives en magnésium sont d'autre part nettement plus faibles chez les 3 espèces terrestres (0.2 à 0.5?ô) par rapport à l'espèce marine (1.2%). Chez cette dernière, le rapport Ca/Mg y est de 2 à 5 fois plus élevé. Sachant d'autre part que la presque totalité du calcium et du magnésium cuticulaires se présentent sous la forme de calcite magnésienne, l'importance de la minéra-lisation de la cuticule des espèces étudiées se situe entre 60 et 80% environ du poids total cuticulaire, soit un taux de minéralisation du même ordre de grandeur que celui mentionné pour la plupart des Crustacés décapodes dulcicoles (Huner et al., 1976; Mills et al, 1976; Huner et Lindqvist, 1985) et marins (Welinder, 1974; Goffinet et Compère, 1986).
44

CONCLUSION
Si la cuticule tergale des Isopodes marins et terrestres se distingue
par des caractères originaux (absence de couche membraneuse, distribution
homogène et lâche des fibres chitino-protéigues de la couche procuticulaire
minéralisée proximale, réseau canaliculaire peu défini dans la couche procuticu-
laire distale de Sphaeroma ) ,son organisation générale ainsi que son haut degré
de minéralisation sont autant de caractères rappelant incontestablement celle
des sclérites de Crustacés décapodes. D'autre part, une sclérotisation limitée
aux seules microstructures épicuticulaires ainsi que l'absence apparente
d'une couche de cires chez les Oniscoïdes suggèrent que les Isopodes terrestres
possèdent un tégument perméable , corroborant ainsi certaines observations
selon lesquelles ces organismes semblent présenter une faible résistance à
la dessication.
BLISS,D.E., 1968. Transition from water to land in decapod crustaceans.
Am.Zool., 8, 355-392. BOCQUET-VEDRINE, J., 1976. Les voies d'absorption de l'eau au cours de l'acquisi-
tion de la forme adulte chez le Crustacé Isopode Epicaride Crinoniscus equitans
Perez. Ext■Arch,Zool.Exper,et Gén,, 117, 3, 422-433. BOCQUET-VEDRINE, J., 1979. La croissance tégumentaire et ses rapports avec la
mue chez Crinoniscus equitans (Crustacé Isopode Cryptoniscien). Arch,Zool■
ēxp.gén. , 120, 45-63. COMPERE ,Ph. et GOFFINET, G., 1987. Ultrastructural shape and three-dimensional
organization of the intracuticular canal systems in the mineralized cuticle of
the green crab Carcinus maenas. Tissue Cell, 19 (6), sous presse.
DENNEL,R., 1947. The occurrence and significance of phenolic hardening in the
newly formed cuticle of Crustacea Decapods. Proc.Roy.Soc.(London), B 134,
485-507. GIRAUD-GUILLE,M.M., 1984. Fine structure of the chitin-protein system in the
crab cuticle. Tissue Cell, 16, 75-92. GLENNER.G.G. et LILLIE,R.D., 1970. In : Histochimie normale et pathologigue,
vol.2. Ed. Yanker, P. , et Jolles, G., Gauthier-Villars éd“., Paris.
GOFFINET,G. et.COMPERE, Ph., 1986. Pore canals and ultrastructural organization
of chitinoproteins in the euticle of the crab Carcinus maenas. In : Chitin in
Nature and Technoloqy. Ed. R.Muzzarelli, Ch.Jeuniaux andG.W.Gooday. Plenum
Publ.Corp., London-N.Y., pp.37-43.
HUNER, J.V. et LINDQVIST, O.V., 1985. Exoskeleton mineralization in astacid and
cambarid Crayfishes (Decapoda, Crustacea). Comp.Bioch.Physiol., 80A, 515-521. HUNER, J.V., KOWALCZUK, J.G. et VAULT, J.W., 1976. Calcium and magnesium
levels in the intermolt (C4) carapaces of three species of freshwater Crayfish :
(Cambaridae; Decapoda). Comp.Biochem.Physiol., 55A, 183-185. HURLEY,D.E., 1968. Transition from water to land in amphipod crustaceans. Am.Zool., 8, 327-353. KRISHNAN, G., 1951. Phenolic tanning and pigmentation of the cuticle in Carcinus maenas. Quart.J.Microscop.Si., 92, 333-342. LILLIE,R.D. et BURTNER, H.J., 1970. In : Histochimie normale et pathologique,
vol.2. Ed. Ganter, P. et Jolles, G., Gauthier-Villars éd., Paris.
MILLS, B.J., SUTER, P. et LAKE, P.S., 1976. The amount and distribution of calcium in the exoskeleton of intermolt crayfish of the genera Enqaqeus and Geocharax.Austr.J. Mar.Freshwater Res, , 27, 517-523. PRICE, J.B. et HOLDICH, D.M., 1980. The formation of the epicuticle and associated structures in Oniscus asellus (Crustacea, Isopoda). Zoomorphologie,
94, 321-332. TRAVIS,D.F., 1965. The deposition of skeletal structures in Crustacea. 5 :
The histomorphological and histochemical changes associated with the development
and calcification of the branchial exoskeleton in the crayfish Qrconectes
virilis Hagen. Acta Histochem., 20, 193-222. WELINDER, B.S., 1974. The crustacean cuticle. I. Studies on the composition
of the cuticle. Comp.Biochem.Physiol., 47A, 779-787.
45

ASPECTS RÉCENTS de la Biologie des CRUSTACÉS Actes de Colloques - 8 - 1988 - IFREMER, pages 47 à 54
MISE EN PLACE DE L’EPICUTICULE CHEZ LE CRABE Carcinus maenas
COMPERE, Ph.(1)
RESUME - L'épicuticule du crabe Carcinus maenas se compose toujours de trois couches distinctes : le recouvrement fibreux superficiel, la cuticuline et I'épicuticule interne. Comme chez les Insectes, la cuticuline est la première couche mise en place sous forme de courts segments au niveau des plaques denses de l'apex des cellules ectodermiques. La fusion de ces segments aboutit à la formation d’une couche pentalamellaire continue de 37 nm d'épaisseur. Au cours d'une seconde étape, l'épicuticule interne est secrétée au contact immédiat de la face interne de la cuticuline sans 1'intervention de plaques denses,. absentes de l'apex des cellules ectodermiques. Le dépôt du recouvrement fibreux superficiel n'a lieu qu'au cours des stades tardifs de la période préecdysiale (D2-D3). Au niveau des sclérites, cette organisation caractéristi-que de 1'épicuticule persiste au cours de l 'intermue. Elle paraît subir des modifications plus ou moins importantes au niveau des zones cuticulaires perméables (branchies).
mots-clés : épicuticule, cuticuline, Carcinus maenas.
ABSTRACT - The epicuticle of the crab Carcinus maenas consists in three distinct layers : the fibrous surface coat, the cuticulin layer and the inner epicuticle. As in Insects, short segments of the cuticulin layer are first deposited at the top of dense plaques of the ectodermal cell membrane. Theirassembling gives rise to a five-layered continuous structure of about 37 nm thick. Afterwards, the inner epicuticle is added inside close to the cuticulin layer but dense plaques are not concerned with the phenomenon. The deposition of the fibrous surface coat outside to the cuticulin layer takes place only during late postmolt stages (D2-D3). In mineralized cuticles, this typical epicuticle structure persists during all intermolt long. In contrast, it seems to be more or less greatly modified in permeable cuticular regions (gill cuticle).
key-words : epicuticle, cuticulin, Carcinus maenas.
(1) Aspirant au fonds National de la Recherche Scientifique (Belgique).
- Laboratoire de Morphologie animale, Université de Liège, Institut de
Zoologie, 22, quai Van Beneden, B-A020 LIEGE (Belgique).
- Ce travail a été réalisé grâce au Fonds de la Recherche Fondamentale
Collective (conventions n°s 2.4506.83 et 2.4516.86).
47

INTRODUCTION
L'épicuticule des Arthropodes apparaît le plus souvent comme une couche
cuticulaire superficielle pluristratifiée. Malgré sa faible épaisseur (0.1 à
3 >jm), son importance est considérable. Mise en place dès le début de la
période préecdysiale, elle constitue à tout instant du cycle de mue l'interface
entre le milieu externe et l'organisme. Continue, elle limite les régions
tégumentaires les plus diverses et montre une structure variable en rapport
avec la spécialisation mécanique ou physiologique du tégument. On lui reconnaît
ainsi un rôle primordial dans la perméabilité cuticulaire.
Chez les Insectes, la structure, l'origine et la formation des couches
épicuticulaires sont relativement bien connues pour plusieurs espèces grâce aux
travaux de Locke (1966, 1969, 1976), Filshie et Waterhouse (1969),
Delachambre (1970), Filshie (1970, 1982) et Gnatzy et Romer (1984). Chez les
Crustacés par contre, la plupart des auteurs se limitent à une simple descrip-
tion des couehes successives telles qu'elles apparaissent en intermue.
L'épicuticule des sclérites est généralement subdivisée en deux parties :
l'épicuticule externe l'épicuticule interne (Green et Neff, 1972;
Christiansen et Costlow, 1982). L'épicuticule externe est une mince couche
superficielle apparemment composée de plusieurs strates dont une au moins
correspondrait à la cuticuline décrite par Locke (1966) chez les Insectes.
L'épicuticule interne, plus épaisse, est formée d'une matrice dense aux
électrons. Elle émet vers le bas des prolongements côniques, ou racines épicu-
ticulaires, pénétrant la pro'cuticule. Elle est traversée par un grand nombre de
minces canalicules verticaux de 25 nm de diamètre, les canalicules épicuticu-
laires, en relation directe avec les canaux de la procuticule (Compère et
Goffinet, 1987b et c).
Le but de ce travail est d'établir avec certitude quels éléments ou strates
interviennent dans la formation de 1'épicuticule du crabe Carcinus maenas.
Dans cette optique, nous nous sommes intéressés au problème de la mise en place
de ces strates ainsi qu'à leur évolution au cours du cycle de mue. Ces informa-
tions nous ont permis d'établir des correspondances entre plusieurs strates
épicuticulaires des Insectes et des Crustacés, et d'aborder ainsi le problème de leur homologie.
MATERIEL ET METHODES
Les crabes Carcinus maenas, de 2 à 3 cm de long, ont été récoltés à la Station Marine de Wimereux (France). Les stades du cycle de mue ont été déter-minés selon la méthode de Drach et Tchernigovtzeff,(1967), complétée par Vilotte (1982) et Anger (1983). Des fragments de différentes zones cuticulaires miné-ralisées (sclérites) telles que le bouclier céphalique, la lame branchiostège interne et l'abdomen, ont été isolés et préparés selon la technique classique pour la microscopie électronique à transmission (milieu d'enrobage, Epon 812). Nos observations ont été réalisées à l'aide de microscopes électroniques à transmission de types Siemens 101 et Jeol 100 SX sous une tension d'accéléra-tion de 80 kV.
48

RESULTATS
Quel que soit le sclérite examiné, la mise en place des premières strates épicuticulaires s'effectue au stade Dl",' après le creusement de l'espace ecdysial, par dégradation des strates de la couche membraneuse. Les soies en formation font toutefois exception à la règle (fig.l). Ces formations sont en effet toujours plus précoces de telle sorte que 1'observation depuis la base de leur matrice jusqu'à leur sommet permet de suivre la progression de la mise en place des composantes épicuticulaires.
Après l'apparition de l'espace ecdysial, l'ectoderme montre une surface ondulée, préfigurant déjà les futurs reliefs cuticulaires tels que les épines épicuticulaires (Fig.2). La membrane plasmique est garnie de plaques denses régulièrement espacées et au sommet desquelles un matériel dense aux électrons semble s'accumuler. A la base des soies en formation, les plaques denses sont déjà coiffées de courts segments d'apparence pentalamellaire (Fig.3). Vers l'extrémité distale des soiès, on assiste à la croissance puis à la fusion de ces segments en plaques plus étendues, de manière à former finaiement une couche continue de 37 nm d'épaisseur (Fig.A) que nous avons appelée cuticuline par analogie avec celle décrite chez les Insectes (Locke, 1966). Cette dernière montre une structure pentalamellaire symétrique (Fig.5). On y distingue aisément deux strates denses aux électrons de 12 nm d'épaisseur, encadrant deux strates claires plus minces (5 nm) et séparées par une fine strate de densité électroni-que élevée. Au niveau des barbules proches de l'extrémité distale des soies, l'épicuticule interne est en cours d'élaboration au contact direct de la face de la cuticuline (Fig.6). A ce niveau, la membrane plasmique est dépourvue de plaques denses.
Au début du stade D2, la nouvelle cuticule du feuillet externe de la lame branchiostège (Fig.7), comprend les premières lamelles de la couche pigmentaire en cours de formation, l 'épicuticule interne, épaisse de 2 à 3 µm, ainsi que la cuticuline qui se trouve en contact direct avec le liquide ecdysial. Durant la seconde moitié de la période préecdysiale, la cuticuline est progressivement recouverte d'un revêtement fibreux dense aux électrons (Figs.8 et 9). Après 1'exuviation, 1'épicuticule ne semble plus subir de modification structurale importante. A partir de cet instant, la cuticuline se distingue difficilement des autres couches épicuticulaires auxquelles elle est partiellement intégrée (Figs.10 et 11). La strate inférieure dense aux électrons semble en effet fusionner avec la matrice de 1'épicuticule interne alors que la strate supérieu-re est étroitement accolée au recouvrement fibreux superficiel. Les deux strates claires ainsi que la strate médiane demeurent bien visibles.
49

Fiqs.1-5. Lame branchiostège du stade Dl'". Fiq.1. Matrice sétigère en section transversale.b,barbules de la soie; ds.portion distale de la future soie; ps; portion proximale. Fiq.2. Apex des cellules ectodermiques(F) dont la membrane plasmique porte de nombreuses plaques denses (pl.).ep, épines épicuticulaires en Formation; es, espace ecdysial; mb, couche membraneuse en cours de déqradation. Fiq.3. Base d'une soie en formation montrant de courts segments de cuticuline (ç) au sommet des plaques denses (pl) de la membrane plasmique. Fiq.A.Cuticuiine (c) formant une couche continue au niveau de la portion distale d'une future soie à proximité de la surface de l'ectoderme (E).pl, plaques denses.Fiq.3.Fort qrossissement montrant la structure pentalamellaire de la cuticuline nouvellement mise ert place.sd, strates denses aux électrons;sc,strates claires aux électrons; t, fine strate médiane.
50

Fig.6. Barbule d'une soie au stade D1 montrant la mise en place de l'épicuti-
cule interne (ei) au contact direct de la cuticuline (c).m, membrane plasmique.
Figs. 7 à 10. Lames branchiostèges au début du stade D2 (7), au stade D2 moyen
(8) en fin de période préecdysiale (9) et en intermue (10). c, cuticuline;
ei, épicuticule interne; es, espace ecdysial; pg, couche pigmentaire; r f, recou-
vrement fibreux superficiel. Fig.11. Lame branchiostège en intermue. Détail
de la cuticuline (c) partiellement intégrée au recouvrement fibreux superficiel
(rf) et à l'épicuticule interne (ci).
51

ÜISCUSSION
Nos observations montrent que l'épicuticule des sclérites du crabe Carcinus maenas comprend trois couches distinctes : le recouvrement fibreux superficiel, la cuticuline et 1'épicuticuline interne. Les deux premières forment l'épicuti-cule externe. Ce schéma structural fondamental est généralement d'application l'épicuticule associée à d'autres régions tégumentaires non minéralisées
telles que les membranes articulaires et les branchies (Compère et Goffinet, 1987a). De plus, il correspond parfaitement à celui proposé par Kümmel et al. (1970) chez l'écrevisse Orconectes limosus.
Comme chez Insectes (Locke, 1966, 1976; Filshie et Waterhouse, 1969; Delachambre, 1970; Filshie, 1970, 1982; Gnatzy et Romer, 1984) et les Arachnides (Filshie, 1976), la cuticuline est la première couche cuticulaire mise en place en début de période préecdysiale. Plusieurs arguments plaident en faveur d'une homologie entre la cuticuline des Crustacés et celles des Insectes. En effet, dans les deux groupes, elle est mise en place sous la forme de courts segments grâce à l'intervention directe de plaques denses de l'apex des cellules ecto-dermiques. La fusion de ces segments, par croissance latérale de chacun d'eux, aboutit d'autre part à la formation d'une couche continue d'apparence pentala-mellaire. Celle de Carcinus maenas së distingue cependant par son épaisseur, 37 nm au lieu de 17.5 nm chez les Insectes Calpodes ethlius (Locke, 1966) et Tenebrio molitor (Delachambre 1970), 8.5 nm chez Neqaria viridula (Filshie et Waterhouse, 1969), 10 à 12 nm chez Lucilia cuprina (Filshie, 1970) et 16 nm chez l'acarien Boophilus microplus (Filshie, 1976). De plus, contrairement à celle des Insectes, elle ne semble subir aucune modification structurale importante depuis sa formation jusque et y compris le stade d'intermue. II n'est est pas de même au niveau des cuticules perméables impliquées dans des échanges ioniques et/ou respiratoires (branchies et feuillet interne de la lame branchiostège) où des remaniements secondaires conduisent à la disparition d'une ou plusieurs de ses strates (Compère et Goffinet, 1987a; Compère, résultats non publiés).
Au cours d'une seconde étape, l'épicuticule interne est sécrétée au contact immédiat de la cuticuline. Son mode de formation, sans l'intervention de .plaques denses, suggère qu'elle est homologue de celle des Insectes ou "protein epicuticle" décrite par Locke (1969) chez Calpodes ethlius.
Enfin, par sa localisation et sa mise en place tardive après la formation de la cuticuline, il est tentant de faire correspondre le recouvrement fibreux
52

superficiel de la cuticule des Crustacés avec la couche de cire des Insectes. Tous deux seraient en outre mis en place par l'intermédiaire des canalicules épicuticulaires. En effet, chez les Crustacés (Jeuniaux et al., 1986) comme chez les Insectes (Locke et Krishnan, 1971; Delachambre, 1971) des composés argentaffines ont été mis en évidence au niveau de canalicules épicuticulaires ainsi qu'à la face externe de la cuticuline peu avant l'exuviation. Nos connaissances sont toutefois insuffisantes pour appuyer une telle hypothèse. De plus, contrairement au recouvrement superficiel déposé durant la seconde moitié de la période préecdysiale, la couche de cire des Insectes est le plus souvent sécrétée après l'exuviation.
En conclusion, cette étude permet non seulement de confirmer le caractère universel, admis par de nombreux auteurs, de la cuticuline au sein des Arthropodes, mais encore de proposer son homologie, du moins chez les Insectes et les Crustacés, sur la base d'une argumentation ultrastructurale et morpho-génétique. Des hypothèses quant à l'analogie ou la correspondance d'autres strates épicuticulaires ent-re ces deux groupes ont également pu être formulées.
ANGER K., 1983. Moult cycle and morphogenesis in Hyas araneus larvae (Decapoda, Majidae), reared in the laboratory. Helgolander Meeresuntersuchunoen 36, 285-302. ’ CHRISTIANSEN M.E. et COSTLOW T.D., 1982. Ultrastructural study of the estuarine crab Rhithropanopeus harisii : Effect of the insect growth regulator dimilin (diflubenzuron) on the formation of the larval cuticle. Mar.Biol., 66. 217-266. COMPERE Ph. et GOFFINET G., 1987a. Aspects ultrastructuraux et fonctionnels de diverses régions cuticulaires non minéralisées d'un crustacé décapode, Carcinus maenas. Annls. Soc.R.Zool,de Belqique, 117 (2), 177-191. COMPERE Ph. et GOFFINET G., 1987b. Ultrastructural shape and three dimensional organization of the intracuticular canal systems in the mineralized cuticle of the green crab, Carcinus maenas. Tissue & Cell, sous presse. COMPERE Ph. et GOFFINET G., 1987c. Elaboration and ultrastructural changes of the pore system in the mineralized cuticle of Carcinus maenas during the molting cycle. Tissue & Cell, sous presse. DELACHAMBRE J., 1970. Etude de l'épicuticule des Insectes. I. Le développement de 1'épicuticule chez l'adulte de Tenebrio molitor. Z. Zellforsch.microsk. Anat., 108, 380-396. : = DELACHAMBRE J., 1971. La formation des canaux cuticulaires chez l'adulte de Tenebrio molitor. Etude ultrastructurale et remarques histochimiques. Tissue & Cell, 3 (3), 499-520. DRACH P. et TCHERNIGOVTZEFF C., 1967. Sur la méthode de détermination des stades d'intermue et son application générale aux Crustacés. Vie et Milieu Série A, Biol.Mar., 18, 595-609. FILSHIE B.K., 1970. The fine structure and deposition of the larval cuticle of the sheep blowfly (Lucilia cuprina). Tissue & Cell, 2 (3), 479-498. FILSHIE B.K., 1976. The structure and deposition of the epicuticle of the adult female cattle tick (Boophilus microplus). In : The Insect Integument, édité par H.R.Hepburn, Elsevier, Amsterdam, 193-206. FILSHIE B.K., 1982. Fine structure of the cuticle of Insects and others Arthropods. In : Insect Ultrastructure, vol.I, édité par R.C.King et H.Akai, Plenum Press, New-York London, 281-312.
53

FILSHIE B.K. et WATERHOUSE D.E., 1969. The structure and development of a surface pattern on the cuticle of the qreen vegetable buq Nezara viridula. Tissue & Cell, 1(2), 367-385. GNATZV W. et ROMER E., 1984. Arthropods : Cuticle : Formation, Moulting and Control. In : Bioloqy of the Intequment, 1. Invertebrates, édité par J.Bereiter-Hahn, A.G. Matoltsy et K.S.Richards, Springer Verlag, Berlin, Heidelberg, New Vork, Tokyo, 638-684. GREEN J.P. et NEFF M.R., 1972. A survey of the fine structure of the intequment of the fiddler crab Uca puqilator. Tissue 4 Cell, 4 (1), 137-171. JEUNIAUX Ch., COMPERE Ph. et GOFFINET G., 1986. Structure, synthèse et dégradation des chitinoprotéines de la cuticule des Crustacés décapodes. Boll,Zool., 53, 183-196. KUMMEL G., CLAASSEN H. et KELLER R., 1970. Zur Feinstruktur von Cuticula und Epidermis beim Flusskrebs Orconectes limosus wahrend eines Hautungszyklus. Z.Zellforsch.mikrosk.Anat., 109, 517-551. LOCKE M., 1966. The structure and formation of the cuticulin layer in the epicuticle of an insect, Calpodes ethlius (Lepidoptera, Hesperiidae). J.Morph,, 118, 461-494. LOCKE M., 1969. The structure of epidermal cell during the development of the protein epicuticle and the uptake of molting fluid in an insect. J.Morph., 127, 7-40. LOCKE M., 1976. The role of the plasma membrane plaques and Golgi complex vesicles in cuticle deposition during the moult/intermoult cycle. In : the Insect Intequment, édité par H.R.Hepburn, Elsevier, Amsterdam, 237-258. LOCKE M. et KRISHNAN N., 1971. The distribution of phenoloxydases and polyphenols during cuticle formation. Tissue &'Cell, 3 (1), 103-126. VILOTTE 0., 1982. Recherches sur la biologie et l'élevage de la cigale de mer Scyllarus arctus (L., 1758) Crustacea, Decapoda, Scyllaridae). Thèse Doct.Sci., Univ.Sci. & Techn. Languedoc, Montpellier.
54

ASPECTS RÉCENTS de la Biologie des CRUSTACÉS Actes de Colloques - 8 - 1988 • IFREMER, pages 55 à 64
ASPECTS UL TRASTRUCTURAUX DE LA BIODEGRADATION DE L'ANCIENNE CUTICULE DE Carcinus maenas DURANT LA PERIODE PREECDYSIALE.
THOREZ, A. , COMPERE, Ph.(1)
el GOFFINET, G.
RESUME - L'obseruation ultrastructurale de diverses régions tégumentaires du crabe Carcinus maenas pendant la période préecdysiale montre l'existence d'une biodégradation cuticulaire importante au niveau des sclérites. Au moment de 1'exuviation, ce phénomène est tel qu'il affecte la totalité de la couche mem-braneuse ainsi que les deux tiers inférieurs de la couche principale. II se tra-duit par une désorganisation suivie de la disparition des éléments microfibril-laires chitino-protéiques. De nombreuses gouttelettes ecdysiales, de même aspect que celles décrites chez les Insectes, et des formations sphériques à double paroi semblent participer activement au processus de dégradation. Des phénomènes comparables s'observent au niveau des régions cuticulaires non minéralisées sou-mises aux déformations mécaniques (membrane articulaire). Par contre, au niveau des zones tégumentaires impliquées dans les échanges respiratoires et/ou ioni-niques (branchies, feuillet interne des lames branchiostèges), ils sont nette-ment plus discrets et n'apparaissent que très tardivement.
mots-clés : cuticule, Carcinus maenas, mue (pré), biodégradation.
A3STRACT - Ultrastructural observations of various integumental regions of the crab Carcinus maenas show an important biodegradation of the old sclerites du-ring premolt. At the ecdysis, this biodegradation extends from the basis of the membranous layer up the upper third of the principal layer. The first biodegra-dation features are a disorganization followed by a complete dissappearance of the microfibrils. Numerous ecdysial droplets, like those described in Insects and double membrane bounded microstructures probably take part in biodegrada-tion processes. Similar phenomenons are seen in unmineralized cuticles, except for permeable tegumental regions (gill and inner branchiostegite cuticles), Where they are delayed and by far less developped.
key-words : cuticle, integument, Carcinus maenas, moult (pre), biodegradation.
INTRODUCTIQN
Chez les Crustacés décapodes, la cuticule est divisée en deux régions d'importance très inégale : 1'épicuticule, mince couche superficielle d'appa-rence pluristratifiée, et la procuticule, plus épaisse et d'aspect lamellaire. Au niveau des sclérites, la procuticule comprend trois couches distinctes : la couche pigmentaire, la couche principale et la couche membraneuse. Celle-ci se différencie des autres par sa faible épaisseur, l'absence de minéralisation et une distribution homogène et compacte des microfibrilles chitino-protéiques En contact direct avec l'ectoderme, c'est la dernière strate procuticulaire à
Laboratoire de Morphologie animale, Quai Van Beneden, 22, B-4020 LIEGE (Belgique)
(1) Aspirant au Fonds National de la Recherche Scientifique.
- Ce travail a été réalisé grâce au Fonds de la Recherche Fondamentale Collec-tive (convention n° 2.4506.83 et 2.4516.86).
55

être mise en place, en fin de période post-ecdysiale (stade selon la nomen-clature de Drach et Tchernigoutzeff, 1967).
Des ëtudes biochimiques réalisées chez Carcinus maenas montrent que la couche membraneuse se transforme rapidement en un gel hygroscopique de nature glycoprotéique (Jeuniaux, 1959, 1963). Cette gélification s'accompagne d'une libération d'acides aminés et de N-acétyl-glucosamine et se poursuit jusqu'à 1'exuviation, entre la nouvelle cuticule en voie de formation et l'ancienne en cours de dégradation. On admet généralement que ce gel joue un rôle de "lubri-fiant" au moment de l'exuviation et qu'il constitue un réservoir d'enzymes hy-drolytiques susceptibles de diffuser vers les strates de l'ancienne cuticule, les produits résultant de cette activité chitino et protéolytique étant récu-pérés par l'ectoderme et incorporés dans les matériaux de la nouvelle cuticule (Freeman, 1980). Chez Carcinus maenas en particulier, quelque 50 % environ de la chitine et des protéines acido-solubles de l'ancienne cuticule seraient mobi-lisées (Jeuniaux et al., 1986).
A partir d'observations en microscopie électronique à transmission, le présent travail considère d'abord les modifications et altérations progressives de la couche membraneuse et des strates proximales de la couche principale du tégument minéralisé de C. maenas (lame branchiostège externe, feuillet externe de l'abdomen, bouclier céphalothoracique), au cours des stades préecdysiaux. Les modifications de l'organisation de la procuticule proximale seront ensuite envisagées et comparées au niveau des régions tégumentaires non minéralisées, à savoir au niveau de la membrane articulaire, ainsi que du feuillet interne de la lame branchiostège et de la branchie, structures impliquées dans des phéno-mènes d'échanges hydro-minéraux et respiratoires.
Les observations ont été réalisées sur des crabes de petite taille (4 à 5 cm), respectivement aux stades Do, D|, D^', D"| et D., ainsi que sur des exu-vies fraîchement prélevées. La détermination des stades est basée sur la métho-de de Drach et Tchernigovtzeff (1967), complétée par Buchholz (1982) et Anqer (1983).
8I0DEGRADATI0N DES ZDNES CUTICULAIRES MINERALI5EE5 (5CLFRITES)
D'un point de vue morphologique, la cuticule des sclérites de C. maenas se caractérise, au cours de la phase préecdysiale, par une série de phénomènes complexes. On retiendra en particulier le décollement entre l'épiderme et 1 ' an-cienne cuticule ainsi que les remaniements de la zone apicale de l'épiderme, l'apparition des gouttelettes ecdysiales et des microstructures arrondies à double paroi, la désorganisation suivie de la disparition des éléments micro-fibrillaires. Ces phénomènes se déroulent selon une séquence bien déterminée.
a) Le phénomène de_l'apolyse et le stade Do
On admet classiquement d'après les travaux de Drach et Tchernigovtzeff (1967), que la phase préecdysiale d'un cycle de mue de Crustacé décapode se manifeste dès le stade Do par un décollement de l'ectoderme et de la cuticule (apolyse), ménageant de ce fait un espace exuvial où s'édifiera la nouvelle cuticule.
56

Un tel décollement n'a jamais été observé à ce stade chez C. maenas, quelle que soit la région tégumentaire examinée (fig. -1). Contrairement à l'opinion généralement répandue, l'apolyse ne peut donc être considérée comme le critère morphologique type définissant le stade Do. Un véritable espace exuvial, tel qu'il est décrit dans la littérature (Jungreis, 1978a et b; Gnatzy et Romer, 1980) n'apparaît que tardivement, à la fin du stade ' (fig. 4) et de façon synchrone dans toutes les régions cuticulaires étudiées. Cet espace semble par ailleurs résulter plus d'un creusement de la couche membraneuse suite à la dé-gradation des fibrilles chitino-protéiques que d'un véritable décollement de la cuticule et de l'ectoderme.
Cependant, si l'apolyse ne constitue pas un critère morphologique caracté-ristique du stade Do, certains remaniements de la membrane plasmique apicale de l'épiderme des sclérites témoignent des premières manifestations de l'entrée en cycle de mue. En effet, dès le stade Do, outre une intense vacuolisation de l'apex des cellules (fig. 1), les plaques denses, organisateurs membranaires impliqués dans le dépôt des ‘éléments fibrillaires de l'ancienne cuticule, sont les premières structures à subir une dégradation. A certains endroits, on assis-te à leur internalisation dans le cytoplasme apical où elles sont ensuite in-cluses dans des corps multivésiculaires, probablement d'origine lysosomiale. De nouvelles plaques denses apparaîtront plus tard, juste avant l'apolyse, dès la mise en place des premières strates (cuticuline) de la nouvelle cuticule (stade D
1"' ).
b) Les gouttelettes_ecdysiales
Une autre manifestation morphologique précoce de l'entrée en cycle de mue consiste en 1'apparition, à la base de la couche membraneuse, de petites forma-tions granulaires denses aux électrons (fig. 1). Très rapidement, ces formations, non limitées par une membrane, paraissent fusionner pour constituer de grands massifs polymorphes dont l'aspect et la structure sont comparables aux goutte-lettes ecdysiales décrites chez les Insectes (Filshie, 1970; Locke et Krishnan, 1973; Gnatzy et Romer, 1980). Le mode de formation et l'évolution de ces goutte-lettes chez C. maenas, corroborent les hypothèses de Filshie (1970) et Locke (1976) selon lesquelles ces structures, chez les Insectes, contiendraient des précurseurs d'enzymes actifs du fluide de mue et joueraient un rôle primordial dans les processus de dégradation cuticulaire.
57

Fiq. 1. Feuillet externe de la lame branchiostège au stade Do. Plaques denses associées à des invaginations de la membrane plasmique apicale (têtes de flèche) ou incluses dans de petites vésicules. Flèches; gouttelettes ecdysiales. v; va-cuoles. Fiq. 2. Feuillet externe de la lame branchiostège au stade D^'". Micros-tructures à double paroi (têtes de flèche) à l'apex des cellules ectodermiques et au niveau de la zone dégradée de la couche membraneuse (mb). E; ectoderme. Fiq. 3. Feuillet externe de la lame branchiostège au stade D1'". La zone dégra-dée de la couche membraneuse couvre sept à huit lamelles. En bordure du front de dégradation, ies gouttelettes ecdysiales (ed) sont entourées d'un grand nombre de petits grains d'aspect identique (flèches). mb; couche membraneuse intacte. E: ectoderme•astérisque; lacune. Tête de flèche; microstructures à double paroi.
58

Fig. 4. Feuillet externe de la lame branchiostège au stade D1
"' ' tardif. Apparition
de l'espace ecdysial (es). La couche membraneuse (mb) est dégradée sur une épais-seur de treize lamelles environ. E; ectoderme. ed; gouttelettes ecdysiales. asté-risque; lacune. Fiq. 5. Feuillet externe de la lame branchiostège au stade avancé. La totalité de la couche membraneuse ainsi que les lamelles proximales de la couche principale (pr) sont dégradées. ep; épicuticule. es; espace ecdysial. pg; couche pigmentaire. tête de flèche; microstructures à double paroi. Fiq.6. Exuvie du feuillet externe de la lame branchiostège. La couche membraneuse (mb) subsiste sous une forme filamenteuse désorganisée. La couche principale (pr) appa-raîL profondément dégradée. Flèche; macrofibres altérées. astérisque ; fantôines de canalicules intracuticulaires. Fiq. 7. Lamelles branchiales (D^) . ar; ancien-ne cuticule. E; ectoderme. es; espace ecdysial. nc; nouvelle cuticule.
59

Le fait que certaines régions tégumentaires qui, comme nous le verrons par
la suite, ne subissent pas ou peu d'altération sont dépourvues de ces gouttelet-
tes, plaide en faveur de cette hypothèse.
Toutefois, contrairement aux Insectes où elle ne serait que très progressi-
ve (Locke, 1976), la sécrétion des gouttelettes ecdysiales chez C. maenas semble
beaucoup plus limitée dans le temps, s'effectuant par un largage massif en début
de période préecdysiale (stades Do et Dj). Dès le stade D1", ces structures font
en effet défaut ou n'existent plus que sous la forme de quelques amas de petite
taille au niveau de la zone proximale de l'épiderme, où de nombreuses lacunes
évidées de tout contenu microfibrillaire font progressivement leur apparition
(fig. 3). Par contre, dans la région dégradée supérieure, en particulier au ni-
veau de la zone correspondant au front de dégradation, ces gouttelettes ecdysia-
les présentent les figures classiques de gros massifs denses aux électrons.
L'observation à haute résolution de ces massifs montre qu'ils semblent subir une
désintégration progressive de leur contenu par émission d'amas minuscules. La
présence de ces derniers dans les canaux cuticulaires suggère d'autre part que
les canalieules verticaux constituent des voies d'accès privilégiées aux zones
cuticulaires supérieures non encore affectées par les phénomènes de dégradation
(fig. 3).
La disparition des gouttelettes ecdysiales (fig. 3) dès le stade D2 coïnci-
de avec le moment où, selon Jeuniaux (1959, 1963), la couche membraneuse se trans
forme en un gel hygroscopique caractéristique. De là à supposer que la désorgani-
sation des gouttelettes entraîne la libération d'enzymes confirme ainsi les obser
vations de cet auteur sur Maia squinado et Cancer paqurus et pour qui la couche
membraneuse gélifiée constitue, dès le stade D2, le réservoir d'enzymes hydroly-
tiques assurant la dégradation des couches procuticulaires proximales jusqu'au
moment de 1'exuviation. Notons que chez les Insectes, Noble-Nesbitt (1963), Lo-
cke (1964), Locke et Krishnan (1973) ont également établi un parallélisme entre
la disparition des goüttelettes ecdysiales et la digestion de l'ancienne cuticule
par le fluide de mue.
c) Les microstructures arrondies à double_paroi
Très tôt, dès le stade D', de petites formations sphériques à contenu dense
aux électrons, de 100 nm de diamètre et limitées par une double membrane, font
leur apparition dans le cytoplasme apical des cellules épidermiques et au niveau
des lamelles proximales de la couche membraneuse en cours de dégradation (fig.2)
60

Leur nombre s'accroît considérablement au cours des stades ultérieurs, leur pré-sence étant étroitement associée à la progression centrifuge du processus de dégradation. Parallèlement, elles semblent se vider de leur contenu mais, au contraire des gouttelettes ecdysiales, leur présence est notée dans le liquide exuvial jusqu'aux stades ultimes précédant directement l'exuviation.
L'application de tests histochimiques (comme la réaction de Feulgen), cou-plée à des observations à haute résolution, révèlent qu'il ne s'agit pas d'élé-ments bactériens, ni de particules virales.
Si la nature exacte de ces microstructures demeure énigmatique, leur ori-gine, leur évolution et leur localisaiton au cours de la phase préecdysiale, ainsi que leur absence dans les cuticules non affectées par les phénomènes de dégradation, suggèrent néanmoins leur participation active dans le processus de biodégradation.
d) La dégradation du système fibrillaire
Les premiers signes de dégradation des fibres chitino-protéiques se mani-festent dès le stade Do au niveau des strates proximales de la couche membra-neuse par une profonde altération de l'organisation spatiale du système fibril-laire. L'orientation originelle des microfibrilles selon une architecture tri-dimentionnelle hélicoîdale caractéristique (- système universel en "contrepla-qué" décrit par Bouligand, 1965, 1971, 1972), devient en effet complètement anar-chique.
Cette désorganisation est bientôt suivie par la disparition locale des élé- . ments microfibrillaires, faisant ainsi place à des espaces lacunaires. Le déve-loppement accru du nombre et de l'importance de ces lacunes, parallèlement à la progression centrifuge du processus de biodégradation, traduit une résorption massive des matériaux chitino-protéiques hydrolysés (figs. 4 et 5). Témoignant de l'intense résorption de matériaux cuticulaires, on observe, comme chez les Insectes (Locke et Collins, 1967; Delachambre, 1971), de nombreuses "coated vesicles" dès le stade D1' dans la région apicale des cellules de l'ectoderme.
Au moment de l'exuviation, la dégradation des matériaux organiques dl'an-cienne cuticule est telle qu'elle affecte la totalité de la couche membraneuse ainsi que les deux tiers inférieurs de la couche principale (figs.5,6). Les ca-nalicules intracuticulaires verticaux, initialement limités par un manchon de fibres verticales ne subsistent que sous l'aspect de "fantômes", 1'organisation caractéristique des microfibrilles horizontales en macrofibres ayant d'autre part complètement disparu (fig. 6).
BIODEGRADATION DES ZONES TEGUHENTAIRES NON HINFRALISEES
A l'inverse des régions minéralisées où les phénomènes de dégradation cuti-culaire se déroulent selon un même schéma, la comparaison des zones tégumentai-res non minéralisées montre des figures de dégradation très variables. Rappe-lons toutefois que, quel que soit le niveau considéré, l'apolyse a toujours lieu à la fin du stade D^"'.
61

a) Membrane articulaire
Si l'on excepte quelques particularités d'importance secondaire, les figu-res de dégradation de la membrane articulaire montrent de grandes ressemblances auec celles observées dans les zones minéralisées (ampleur des phénomènes de dé-gradation, microstructures à double paroi, gouttelettes ecdysiales).
II est vrai que, anatomiquement, la membrane articulaire ne constitue qu' un simple prolongement des sclérites et qu'elle présente, à l'exclusion de la présence des sels de calcium, des caractéristiques fort proches de celles des régions minéralisées quand on considère son épaisseur, le nombre de lamelles procuticulaires et son organisation générale.
b) Feuillet interne de la lame branchiostège et branchie
Par rapport aux régions minéralisées et à la membrane articulaire, les phé-nomènes de dégradation cuticulaire dans ces deux régions se manifestent d'une manière beaucoup plus discrète.
Les premiers signes morphologiques de l'entrée en mue n'apparaissent que très tardivement, lors de la mise en place de la nouvelle cuticule (stade D"I). L ' internalisation des plaques denses qui constituait le premier signe morpholo-gique de biodégradation dans le cas des sclérites, n'a pas été observée ici. De plus, ces phénomènes de dégradation se réduisent le plus souvent à une dis-parition de l ' organisation lamellaire et de l'association compacte et homogène des éléments microfibrillaires des trois ou quatre lamelles proximales de la cuticule. Ces phénomènes ne s'accompagnent d'ailleurs jamais d'une formation de gouttelettes ecdysiales,ni de microstructures à double paroi (fig. 7).
Des observations récentes sur la morphologie (Goffinet et Compère, 1986) et sur la physiologie (Péqueux, communication personnelle) des cuticules impliquées dans des phénomènes respiratoires et d'échanges ioniques, montrent une organisa-tion et des propriétés particulières de ces cuticules. Faut-il voir là une rela-tion entre l'apparition tardive et discrète des phénomènes de biodégradation, ne prenant place que lorsque la sécrétion de la nouvelle cuticule a débuté, et la nécessité pour l'organisme de maintenir constantes ses conditions internes d'ho-méostasie au cours du cycle de mue ? Seule une étude plus poussée sur la nature chimique de ces cuticules serait susceptible d'apporter une réponse.
CONCLUSION
Les phénomènes de biodégradation cuticulaire revêtent des aspects très dif-férents selon les régions envisagées. D'une manière générale, les régions à vc-cation essentiellement mécanique, telles que les sclérites et les membranes ar-ticulaires, subissent une dégradation précoce et très marquée. Au contraire, les zones cuticulaires impliquées dans des phénomènes respiratoires (feuillet inter-ne de la lame branchiostège et branchie) et/ou d'échanges hydro-minéraux (bran-chie) ne sont pas ou peu affectées. Le fait que la dégradation de ces zones cu-ticulaires perméables est peu poussée et tardive par rapport aux zones à rôle mécanique, suggère que l'ectoderme présente deux pics d'activité cellulaire dis-tincts durant la période préecdysiale. Ils correspondraient respectivement à la biosynthèse dcs enzymes hydrolytiques suivie de la résorption des matériaux de
62

l'ancienne cuticule et à la biosynthèse de matériaux du nouueau tégument. Seul
le second type d'activité se manifesterait dans les zones cuticulaires perméa-
bles.
ANGER, K., 1983. Moult cycle and morphogenesis in Hyas araneus larvae (Decapoda
Majidae), reared in the laboratory, Helgoländer Meeresunters, 36, 285-302.
BOULIGAND, Y., 1965a. Sur une architecture torsadée répandue dans de nombreuses
cuticules d'Arthropodes, C.R. hebd. Séanc. Acad. Sc. Paris, 261, 3665.
BOULIGAND, Y., 1971. Les orientations fibrillaires dans le squelette des Arthro-
podes. I. L'exemple des crabes, 1'arrangement torsadé des strates, J. Micros.
Paris, 11, 441-472. BOULIGAND, Y., 1972. Twisted fibrous arrangements in biological materials and
cholesteric mesophases, Tissue and Cell, 4, 2, 189-217. BUCHHOLZ, F. , 1982. Drach's molt staging system adapted for Euphausiids, Marine
Biol., 66, 301-305. DELACHAMBRE, F., 1971a. Modifications de l'épiderme au cours de la sécrétion de
1'épicuticule imaginale chez Tenebrio molitor, Z. Zellforsch. Mikrosk. Anat.,
112, 97-119. DRACH, P. et TCHERNIGOVTZEFF, C., 1967. Sur la méthode de détermination des stra-
tes d'intermue et son application générale aux Crustacés, Vie et Milieu, sér.A,
Biol. Marine, 18, 595-609. FILSHIE, B.K., 1970. The fine structure and deposition of the larval cuticle of
the sheep blowfly (Lucilia cuprina), Tissue and Cell, 2, 479-489.
FREEMAN, J.A., 1980. Hormonal control of chitinolytic activity in the integument
of Balanus amphitrite in vitro, Comp. Biochem. Physiol., 65A, 1, 13-17.
GNATZY, W., R0MER, F., 1980. Morphogenesis of mechanoreceptor and epidermal cells
of crickets during the last instar, and its relation to molting-hormone level,
Cell Tissue Res., 213, 369-391. G0FFINET, G. , C0MPERE, Ph., 1986. Aspects ultrastructuraux et fonctionnels de di-
verses régions cuticulaires non minéralisées d'un Crustacé décapode Carcinus
maenas, Annls. Soc.r. Zool. Belq., 117 (2), 177-191.
JEUNIAUX, Ch. , 1959. Sur la gélification de la couche membraneuse chez les Crabes
en mue, Arch. Intern. Physiol. Bioch., 67, 516-517. JEUNIAUX, Ch. , 1963. Chitine et chitinolyse. Synthèse d’enzymes chitinolytiques
et dégradation de la chitine cuticulaire chez les Crustacés décapodes, édité
par Masson et Cie, Paris, 136-144. JEUNIAUX, Ch. , C0MPERE, Ph., G0FFINET, G., 1986. Structure, synthèse et dégrada-
tion des chitinoprotéines de la cuticule des Crustacés décapodes, Boll. Zool.,
53, 183-196. JUNGREIS, A.M., 1978a. The composition of larval-pupal moulting fluid in the to-
bacco horn worm Manduca sexta, J. Insect Physiol., 24, 65-73.
JUNGREIS, A.M., 1978b. Physiology of molting in insects, in : TREHERNE, J.E.,
BERRIDGE, M.J., WIGGLESWORTH, V.B. (Eds.), Advancer of Insect Physiology, 14,
109-183. L0CKE, M. , 1964. The structure and formation of the integument in Insects, in :
R0CKSTEIN, the Physiology of Insecta, vol. III, Academic Press, New-York, 379-
470. L0CKE, M. , 1976. The role of plasma membrane plaques and Golgi complex vesicles
in cuticle deposition during the moult/intermoult cycle, in : HEPBURN, M.R.
(ed.), the insect intequment, Elsevier, Amsterdam, 237-258.
63

LOCKE, M., COLLINS, J.U., 1967. Protein uptake in multivesicular bodies in the molt/intermolt cycle of an insect, Science, N.Y., 155, 467-469.
LOCKE, M., KRISHNAN, N., 1973. The formation of the ecdysial droplets and the ec-dysial membrane in an insect, Tissue and Cell, 5, 3, 441-450.
NOBLE-NESBITT, J., 1963b. The cuticle and associated structures of Podura aquati-ca at the moult, Q.J. microsc. Sci., 104, 369-391.
64

ASPECTS RÉCENTS de la Biologie des CRUSTACÉS Actes de Colloques - 8 - 1988 - IFREMER, pages 65 à 68
ACTION DES CHOCS THERMIQUES SUR LE DEVELOPPEMENT D'ARTEMIA.
A. HERNANDORENA (1)
RESUME - Artemia a été soumis à des chocs thermiques à 40' à la fin de la vie embryonnaire et au début de la vie postembryonnaire et les effets à long terme de ces chocs sur le développement ultérieur examinés. Les chocs thermiques infligés pendant la période du développement embryonnaire comprise entre l'hydratation des oeufs et l'éclosion des nauplii n'ont aucune répercussion sur le développement ultérieur. Par contre les chocs thermiques infligés dès l'éclosion alors que les larves ne s'alimentent pas encore mais que leurs cellules se multiplient, induisent des anomalies au niveau des appendices thoraciques et génitaux qui pourraient correspon-dre à des transformations homéotiques. Ces résultats permettent de mieux situer les effets morphogénètiques obtenus antérieurement en modifiant la composition du milieu nutritif dans lequel les larves sont élevées.
mots-clés : Artemia. chocs thermiques, homéoses.
ABSTRACT - Heat shocks at 40‘ were delivered to Artemia at the end of embryonic development and at the beginning of postembryonic development and their effects on subsequent development examined. Heat shocks delivered during preemergence development have no effect on subsequent development. Heat shocks delivered to newly hatched nauplii induce anomalies on the morphogenesis of thoracic and genital appendages. These anomalies might correspond to homeotic transformations. The induction of morphological anomalies by thermal conditions prevailing during early larval life makes it easier to integrate, the morphological effects previously induced by nutritional conditions imposed to older feeding stages.
Key-words : Artemia. heat shocks, homeoses.
La qualité l'alimentation offerte aux larves d'Artemia permet de remettre en cause le caractère normalement appendiculé des segments thoraciques et le caractère normalement apode des segments abdominaux (Hernandorena 1970, 1974, 1979). Le début de la vie larvaire échappe à ce type d'expérimentation puisque la larve nauplius ne s'alimente pas et que les trois premières mues larvaires s’effectuent aux dépens des réserves. Or avant même que la larve n'ait commencé à s'alimenter, le mesoderme correspondant à la formation des sept premiers segments thoraciques est délimité (Anderson, 1967). II était donc intéressant d'envisager une intervention plus précoce, antérieure au stade auquel les larves deviennent sensibles à la composition du milieu nutritif. Pour ce faire, Artemia (race Utah) a été soumis à des chocs thermiques à 40' et les effets à long terme de ces chocs sur le développement ultérieur examinés. La température de 40' a été choisie car les effets à court terme des chocs à 40' sur la biologie moléculaire d'Artemia sont connus (Miller et McLennan 1986). Les larves choquées ou non sont élevées dans des condi-tions axéniques sur un nouveau milieu nutritif simplifié (Hernandorena, non publié). Les oeufs sont incubés et les larves élevées à 25°+0.5° Les animaux arrivés au terme de leur développement sont anesthésiés et examinés au microscope.
(1) Laboratoire MNHN, Plateau de l'Atalaye, 64200 Biarritz.
65

Photo 1-5. Anomalies induites par un choc thermique à 40° d'une durée de 4 heures infligé le jo.
66

Effets des chocs thermiques à 40' sur les larves. Les résultats les plus spectaculaires concernent l'action des chocs infligés le jo dès l'éclosion des nauplii, 24 heures après 1'hydratation des oeufs. Les individus ayant subi le jo un choc à 40' d'une durée de 1 ou 2 heures ne manifestent aucune anomalie évidente. Sur les 314 individus ayant survécu à un choc thermique à 40' d'une durée de 4 heures, 117 soit 37 % manifestent des anomalies (Tableau I).
nombre pourcentaae : : Fusion des appendices thoraciques 74 63 : : Appendices génitaux anormaux 34 29 : : Antennes tronquées 6 5 : : Abdomen spiralé 3 2 : Total 117 '
Tableau I. Anomalies induites par un choc thermique à 40' d'une durée de 4 heures infligé le jo.
Chez un nombre réduit d'adultes, l'anomalie porte sur l'une ou l'autre des antennes ou les deux à la fois, qui se trouvent réduites à un moignon plus ou moins important ou sur l'abdomen dont les segments sont organisés en spirale (photo 1). L'anomalie la plus fréquente est la fusion, souvent unilatérale, des appendices thoraciques adjacents (photo 2). Le phénotype de ces individus correspond à celui qui résulte de la mutation "swimmerette" décrite chez Artemia par Squire et Grosch (1967). La fusion des ébauches appendiculaires des segments adjacents, anormale dans le cas des segments thoraciques, est normale dans le cas des segments génitaux G1 et G
2. Chez certains individus, la partie antérieure du gonopode qui prend
naissance sur le segment génital G1 présente une morphologie qui ressemble à celle d'un appendice thoracique alors que la partie postérieure qui prend naissance sur le segment génital G2 présente la structure d'un appendice génital (photo 3). La mise au point sur un plan focal supérieur montre que chez le même individu, la structure du 2e gonopode est normale (photo 4). La fusion des ébauches appendiculaires des segments G1 et G2 peut ne pas se produire. Dans ce cas, l'appendice du segment Gj ressemble davantage à un appendice thoracique qu'à un appendice génital et le segment abdominal A1 normalement apode, porte une ébauche appendiculaire surnuméraire (photo 5). Tout se passe comme s'il était possible de transformer le segment G1 en segment thoracique T12, le segment G2 en segment G1 et le segment A1 en segment G
2. Ces effets pourraient correspondre à des transformations
homéotiques. Sachant que chez la Drosophile ce sont les chocs thermiques infligés à l'embryon (Maas 1948, Santamaria 1979) et non à la larve qui produisent des phénocopies des mutations homéotiques, il paraissait intéressant de vérifier chez Artemia, les effets sur le développement ultérieur des chocs thermiques infligés aux embryons.
Effet des chocs thermiques à 40' sur les embrvons. Des chocs thermiques à 40' d'une durée de lh, 2h ou 4h infligés lh, 6h ou 18h après 1'hydratation des oeufs n'ont aucune influence sur le développement ultérieur. Une particularité du développement embryonnaire d'Artemia pourrait expliquer ces résultats négatifs. La période comprise entre l'hydratation des oeufs et l'éclosion des nauplii se caractérise par l'absence de multiplications cellulaires et l'absence de biosynthèse d'ADN (Nakanishi et al 1962, Olson et Clegg 1978). Les multiplications cellulaires redémarrent à l'éclosion (stade Ej) moment à partir duquel les chocs thermiques ont une influence sur le développement. ultérieur. Les cellules qui se divisent au moment du choc seraient les seules capables de répercuter sur le développement
67

ultérieur les traces laissées par les chocs thermiques.
Conclusion - L'étude de l'action des chocs thermiques montre que le développement d'Artemia met en oeuvre des processus qui déterminent longtemps à l'avance l'identité des segments : le passage des segments thoraciques aux segments génitaux est décidé dès le jo alors que le corps de la larve nauplius n'est pas encore segmenté. Chez la Drosophile, ces processus correspondent à la mise en oeuvre de l'activité des gènes homéotiques (Lewis 1978). L'existence de tels gènes n'est pas démontrée chez les Crustacés. Si les effets obtenus chez Artemia correspondent bien à des transformations homéotiques, le choc thermique infligé le jo, induirait des phénocopies des mutations homéotiques et la description des copies aura précédé la description des originaux ! Mais alors que le passage des segments thoraciques aux segments abdominaux est remis en cause par des chocs thermiques infligés pendant la vie embryonnaire chez la Drosophile, le passage des segments thoraciques aux segments génitaux est remis en cause par un choc thermique infligé pendant la vie postembryonnaire chez Artemia. Cette donnée est fondamentale car elle permet de comprendre que les conditions nutritionnelles subies par les larves d'Artemia jusqu'au stade IX puissent avoir un effet à retardement sur la morphogenèse d'appendices surnuméraires sur les segments abdominaux normalement apodes (Hernandorena 1970, 1974, 1979). L'expression des gènes qui codent la formation de ces appendices est déréprimée et comme le souli-gne Sang (1984) cette dérépression correspond à la production d'homéoses.
ANDERSON D.T., 1967. Larval development and segment formation in the branchiopod crustaceans Limnadia stanleyena King (Conchostraca) and Artemia salina (L.) (Anostroca). Aust. J. Zool.. 15, 47-91. HERNANDORENA A., 1970. Obtention de morphogenèses appendiculaires abortives et surnuméraires chez Artemia salina (L.) (Crustacé branchiopode) par carences alimentaires de base pyrimidique et de nucléotide purique. C.R Acad. Sci. Paris, 271, 1406-1409. HERNANDORENA A-, 1974. Besoin alimentaire en acide adénylique, croissance et morphogenèse chez Artemia salina (L.) (Crustacé branchiopode). Ann. Nutr. Alim., 28, 65-82. HERNANDORENA A., 1979. Relationships between purine and pyrimidine dietary requirements and Artemia salina morphogenesis. Comp. Biochem. Phvsiol.. 62 B, 7-12. LEWIS E.B. 1978. A gene complex controlling segmentation in Drosophila. Nature. 276, 565-570. MAAS A.H., 1948. Uber die Auslôbarkeit von temperatur-modifikationen wâhrend der Embryonalent-wieklung von Drosophila melanogaster. Wilhelm Roux's Arch., 143, 315-372. MILLER D., McLENNAN A.G., 1986. Changes in intracellular levels of AP3 A and Ap4 A in cysts and larvae of Artemia do not correlate with changes in protein synthesis after heat shocks. Nucleic Acid. Res.. 14, 6031-6040. NAKANISHI Y.H., IWASAKI T., OKIGATI T., KATO H., 1962. Cytological studies of Artemia salina I. Embryonic development without cell multiplication after the blastula stage in encysted dry eggs. Annot. Zool. Jap., 35, 223-228. OLSON C.S., CLEGG J.S., 1978. Cell division during development of Artemia salina■ Wilhelm Roux's Arch.. 184, 1-13. SANG J.H., 1984. Genetics and development. Longman, London, New York, 398 p SANTAMARIA P., 1979. Heat shock induced phenocopies of dominant mutants of the bithorax complex in Drosophila melanoqaster. Molec. Gen. Genet.. 172, 161-163. SQUIRE R.D., GROCSH, D.S., 1967. "Swimmerette" a new sex-linked recessive mutant in the brine shrimp Artemia salina L. Biol. Bull., 133, 487.
68

ASPECTS RÉCENTS de la Biologie des CRUSTACÉS Actes de Colloques - 8 - 1988 - IFREMER, pages 69 à 82
PROGRESSION DE L'ORGANISATION PHRAGMALE DU METAMERE THORACIQUE
DE QUELQUES MALACOSTRACES
B. CASANOVA(1)D. HUGUET(2)S. SECRETAN(3)
RESUME : En comparant le squelette d’un raétamère thoracique répétitif
chez divers Malacostracés, on assiste au développement, puis à la complexité
croissante, de la structure phragmale. Celle-ci se manifeste tout d'abord
chez certains Mysidacés et Euphausiacés sous forrae d’une lame
interarthrodiale issue des sternites. Chez les Decapodes Penaeidae, Atyidae,
Processidae , Palaeraonidae, cette larae se diversifie et il s’y adjoint des
éléments d’origine pleurale. L’araorce de cloison ainsi constituée
s'accroît et se ramifie chez les Nephropidae.
Mots-clés : endophragme, évolution, Mysidacés, Euphausiacés, Décapodes,
Penaeidae, Atyidae , Processidae , Palaeraonidae , Nephropidae.
ABSTRACT : A comparison of the skeletton of a successive thoracic segment
in various Malacostraca shows the developraent and growing coraplexity of
phragmal structure. It first appears in some Mysicacea and in Euphausiacea
by means of an interarthrodial laraina issued from sternite. In Decapoda
Penaidae, Atyidae, Processidae, Palaeraonidae, this lamina is diversified and
some eleraents frora pleural origin join it. The beginning of the so
constituted wall considerably increases in Nephropidae and becomes ramified.
Key-words : endophragrae, evolution, Mysidacea, Euphausiacea, Decapoda,
Penaeidae, Atyidae, Processidae, Palaemonidae, Nephropidae.
Les éléraents structuraux faisant l'objet du présent travail
comparatif participent à l’enserable du squelette cuticulaire métamérisé
noraraé squelette axial. Celui-ci ne coraporte ni les appendices, ni la
carapace et l'épimère qui la double et la relie au squelette axial, puisque
l'une et l’autre sont insegraentées (Secretan, 1977). Le squelette axial
céphalothoracique est seul considéré ici. L’armature de soutien des métamères se corapose de deux catégories de
pièces squelettiques. - les sclérites simples , constitués d'un seul feuillet, qui
isolent le corps du milieu extérieur : sternites, pleurites et tergites (là
où la carapace n'"absorbe" pas ces derniers).
(1) Université de Provence, Centre St-Charles. Biologie animale, Plancton.
Place Victor Hugo, 13331 Marseille Cedex 3.
(2) Faculté des Sciences. Bd Lavoisier. Belle-Beille. 41045 Angers Cédex.
(3) Institut de Paléontologie, Museura National d'Histoire Naturelle.
8, Rue Buffon. 75005 Paris.
69

- Les phragmes , qui sont des invaginations internes à doubles feuillets émanant des sternites et des pleurites de deux métamètres à leur jonction : intérosternites, intéropleurites. Ceux-là sont dits "intersegmentaires" puisque chaque feuillet émane d'un segment différent. Secondairement, il arrive que l'un des feuillets d'un phragme émette à son tour une invagination. Cela donne alors un phragme dit "intrasegmentaire" puisque ses deux feuillets se rapportent à un mêrae segment : intrasternites, intrapleurites (Secretan, 1980).
Selon les groupes, la structure phragmale du squelette axial céphalothoracique peut être quasiment inexistante ou, au contraire, atteindre une grande complexité. A l'intérieur d'un même animal, elle présente des variations d'un métamère à l'autre, à partir d'un type de base spécifique de chaque groupe. Pour établir et comparer ces types de base à travers les principales divisions des Malacostracés, nous avons choisi un métamètre aussi peu spécialisé que possible, d'un modèle répétitif dans le corps, non modifié ni par une fonction masticatrice ni par une fonction reproductrice. Nous l'avons étudié chez des Mysidacés et des Euphausiacés et chez des Décapodes Penaeidae, Atyidae, Processidae, Palaemonidae et Nephropidae, qui sont considérés comme représentant des stades de plus en plus évolués dans l'organisation des Malacostracés et nous avons cherché à savoir comment se concrétise cette évolution au niveau de la structure squelettique de ce métamère type.
Chez les Mysidacés Lophogastrida, l'organisation phragmale est très peu développée, en particulier chez Gnathophausia zoea W. Suhm, 1873 (fig. 1) où la jonction des sternites se fait suivant un repli tégumentaire transversal. Les pleurites portent de petites crêtes internes limitant une zone d'aspect réniforrae où s'insèrent les muscles ; ils se continuent dorsalement par les tergites reliés les uns aux autres par une membrane articulaire. Une zone membraneuse sépare les pleurites des cavités arthrodiales où s'articulent les appendices ; cette zone, d'apparence insegmentée porte des branchies. Les cavités arthrodiales successives sont séparées par les prolongements latéraux des deux sternites juxtaposés, unis par un étroit repli. Chez les Mysida, tels Gastrosaccus spinifer Goës, 1864, apparaissent des phragmes caractérisés : d'une part intérosternaux, résultant de l'invagination plus accentuée du repli inter-sternal formant ainsi une cloison interarthrodxale ; d'autre part interopleuraux, contigus aux précédents (fg. 2).
Cette structure phragmale observée chez les Mysida va se retrouver magnifiée chez les Euphausiacés. En effet, Meganyctiphanes norvegica M. Sars, 1857 , choisie comme espèce type, présente des sternites qui émettent à leur jonction de grandes lames aliformes latérales séparant les cavités arthrodiales (fig. 3). Invaginés vers l'intérieur et incurvés vers l'avant, ces phragmes intérosternaux en position interarthrodiale constituent l'élément endosquelettique majeur des Euphausiacés. En effet, les replis intérospleuraux plus ou moins réduits selon les genres, n'ont pas réellement valeur de phragmes, mais ils sont cependant en contiguité avec les extrêmités distales des phragmes intérosternaux (fig. 4). Chez les Euphausiacés apparaissent des intrasternites tubulaires, à raison d'une paire par sternite, en bordure des cavités arthrodiales.
Chez les Décapodes Penaeidae comme Bentheogennema intermedia Bate, 1888, Gennadas elegans Smith, 1882, et Penaeus trisulcatus Leach, 1815, l’importance du système phragmal est très variable et va régulièrement croissant de la première à la dernière espèce (fig. 5-6). Cependant on peut remarquer que c'est toujours latéralement et entre les cavités arthrodiales,
70

comme dans les ordres précédents, que s'observent des pointes et des laraes inter et intrasegraentaires. Par comparaison avec les Euphausiacés on notera : le grand développement des phragines inter et intra pleuraux, l'agrandissement des cavités arthrodiales qui réduisent ainsi la largeur des sternites, particulièrement net chez P. trisulcatus (fig. 6), et la constitution des cloisons interarthrodiales, auxquelles participent à la fois les intérosternites et les intéropleurites.
Chez les Atyidae, on retrouve une structure qui n'est pas sans rappeler celle des Euphausiacés. Dugastella marocana Bouvier, 1912 prêsente des phragmes en lame séparant les cavités arthrodiales (fig. 7). Leur forme est un peu différente : elle est triangulaire et leur base, dans la région centrale, prend naissance sur le bord du sternite. Les cloisons phragmales d' Atyaephyra desmaresti Millet ont le même aspect et occupent le même emplacement. Au niveau interpleural, un repli en forme de bourrelet prolonge ces cloisons. Enfin, des phragmes interpleuraux sont visibles (fig. 8).
Chez les Processidae, Processa edulis présente un phragme interosternal toujours en position interarthrodiale ; il est plus allongé et plus lobé que chez les Atyidae (fig. 9). Le sternite possède un phragme intrasternal qui n'existe pas chez les atyidae. Un intéropleurite bien individualisé vient rejoindre l'intérosternite dont il est séparé par une mince échancrure et avec lequel il forme conjointement la cloison interarthrodiale. Ceci se rapproche de ce que l'on observe chez les Penaeidae où les interopleurites et les intérosternites sont partie constitutive de cette cloison.
Chez les palaemonidae, Palaemonetes varians possède des sternites très élargis. Les phragmes intérosternaux affectent une forme de pilier dans leur région proximale (fig. 10). Ce caractère s'accentue chez Palaemon serratus où les piliers interosternaux sont plus développés et découpés (fig. 11) ; ils prennent naissance plus près de l'axe médian de l'animal et sont inclinés obliquement vers l'avant. Ces deux espèces présentent aussi des intrasternites et des intrapleurites.
Chez les Nephropidae, l'orientation des cloisons est beaucoup plus verticale. Les intêrosternites de Nephrops norvegicus Linné, 1753, étant en piliers verticaux, on ne les voit pas en vue dorso ventrale (fig. 12) ; seules, leurs extrêmités en plateaux sont visibles. Ces plateaux, ou processus aliformes, participent à la liaison des phragraes qui forment ainsi une cloison continue séparant un métamère de son voisin (fig. 13). Cette intercloison comporte, d'une part des intérosternites dont la base des piliers est jointive sur la ligne médiane du sternite, d'autre part, des intëropleurites qui se sont beaucoup développés par rapport à ceux des familles précédentes et dont les extrêmités convergent vers l'axe médian du corps. Intérosternites et intéropleurites s'accrochent entre eux à leur sommet par un système d'engrenage de leurs extrêmités folliacées (fig. 17).
Un autre phragme, très grand, participe à cet engrenage, un phragme intrasegmentaire, repli émis par le seul feuillet postérieur de
1'intéropleurite de l'intercloison antérieure du métamère. Cet intrapleurite
s'étend vers l'arrière et vient rejoindre, dans l'axe médian, les phragmes
de l'intercloison postérieure du métamère dont il subdivise ainsi le volume
en le cloisonnant obliquement de part et d'autre du sternite. En conséquence, la cavité arthrodiale paraît dédoublée (fig. 14).
De nouveaux intrasternites, lamelleux et folliacés, apparaissent,
issus de feuillet antêrieur du pilier intérosternal de la cloison
71

postérieure. Les autres intrasternites, qui naissent d'un repli des bords du sternite lui-même, et qui existaient déjà, tubulaires, chez les
Euphausiacés s'étalent ici de chaque côté du sternite où ils forment, de
part et d'autre de celui-ci, les condyles articulaires des appendices , d'où leur appellation d'"arthrosternites".
La succession des piliers intérosternaux est bien visible sur une
coupe sagittale du céphalothorax (fig. 15) montrant l'importance des
intercloisons. Sur le segment isolé en coupe sagittale (fig. 16), on voit
nettement le rôle de verrouillage que joue l'intrapleurite qui relie les
deux intercloisons entre elles et les différents niveaux d'accrochage des
phragmes entre eux.
CONCLUSION
Chez les Mysidacés les plus primitifs, les Lophogastrida, on ne
décèle aucun phragme caractérisé. Le point de départ se situe donc chez les
Mysida, et surtout chez les Euphausiacés, où les premiers phragmes apparaissent sous forme de lames qui s'élèvent entre les cavités
arthrodiales tout en prenant naissance sur le bord des sternites, les
intéropleurites étant réduits à de minces replis.
Avec les Décapodes, chez les Penaeidae d'abord, des amorces
d'intéropleurites distincts se joignent à ces lames intérosternales pour
constituer la séparation entre deux métamères. Les Atyidae montrent une
structure voisine de celle des Penaeidae, non sans rappeler toutefois celle
des Euphausiacés. Le développement des phragmes intéropleuraux s'accentue
chez les Processidae. Chez les Palaemonidae, ce sont les intérosternites qui
s'individualisent et qui, pour la première fois, s'élèvent en piliers situés
à la limite latérale du sternite. Chez les Nephropidae, ces piliers se
renforcent et se rejoignent sur l'axe médian sternal et on assiste au grand
développement des éléments pleuraux inter et intrasegmentaires. La novation
des Nephropidae se concrétise essentiellement par la remarquable extension
de l'intrapleurite qui relie obliquement entre elles les deux cloisons qui
limitent, antérieurement et postérieurement, le métamère.
En résumé, la naissance et l'évolution de l'intercloison phragmale
peut être schématisée de la façon suivante : elle apparait, entre les cavités arthrodiales, sous forme d'une lame (fig. 18, hachures obliques) qui va se développer (fig. 18, hachures horizontales), d'une part dans sa région
proximale en s'allongeant en un étroit pilier, d'autre part dans sa région
latérale, à la base de la limite interpleurale. Ces formations vont s'agrandir (fig. 18, pointillé). • Le pilier va s'évaser tout en se
rapprochant de l'axe médian où il devient, à sa base, jointif de son symétrique. Latéralemet, l'intéropleurite s'élève le long des pleurites et
s'étend en largeur et son extrêmitê proximale vient s'accrocher à celle de
son symétrique. La flèche verticale indique la région commune à ces
différents stades, région qui est aussi le point de départ du développement
que soulignent les deux flèches incurvées, dans la région sternale d'une
part, dans la région pleurale d'autre part. On peut considérer que la zone
qui relie l'intérosternite à l'intéropleurite chez les Nephropidae (en
grisé sur la fig. 17) est le vestige de la lame sterno-interappendiculaire
initiale.
SECRETAN S. (1977). Epimère et connexions épiméro-endophragmales chez
les Crustacés Décapodes macroures. Bull. Soc. Zool. Fr. , 102 (4) : 345-374.
SECRETAN S. (1980). Le plan de base du "squelette axial" d'un Crustacé Décapode macroure. C.r. hebd. Séanc. Acad. Sci. , sér. D, 291 : 877-880.
72

PLANCHES
Légendes des photos :
Squelette axial des métamères thoraciques, en vue dorso-ventrale chez :
Fig. 1 : Gnathophausia zoea (Mysidacé Lophogastrida). Partie gauche du squelette. Fig. 2 : Gastrosaccus spinifer (Mysidacé Mysida). Fig. 3-4 : Meganyctiphanes norvegica (Euphausiacé) 3-région sternale ; 4-région pleurale. Fig. 5 : Bentheogennema intermedia (Decapoda Penaeidae) Fig. 6 : Penaeus trisulcatus (Decapoda Penaeidae). Fig. 7 : Dugastella marocana (Decapoda Atyidae) Fig. 8 : Atyaephyra desmaresti (Decapoda Atyidae). Partie gauche du squelette. Fig. 9-10 : Processa edulis (Decapoda Processidae). 9-cavités arthrodiales ; 10-détail des phragmes. Fig. 11 : Palaemonetes varians (Decapoda Palaemonidae). Fig. 12 : Palaemon serratus (Decapoda Palaemonidae). Fig. 13-14 : Nephrops norvegicus (Decapoda Nephropidae) 13-Squelette axial cephalothoracique complet ; 14-métamère isolé, A/photographié ; B/dessiné à la chambre claire.
Squelette axial en vue latérale (coupe sagittale) chez :
Fig. 15-16 : Nephrops norvegicus (Decapoda Nephropidae) 15-squelette axial cephalothoracique complet ; 16-métamère isolé, A/photographie ; B/dessiné à la chambre claire.
Intercloison isolée en vue antéro-postérieure chez :
Fig. 17 : Nephrops norvegicus (Decapoda Nephropidae). Vue antérieure.
Fig. 18 : Schéma récapitulatif, en vues superposées, de la progression des phragmes intersegmentaires de 1'intercloison thoracique dont les flèches soulignent le point de départ et 1'orientation du développement. Les différentes figures représentent les stades concrétisés par : A/ les Euphausiacés et les Décapodes Atyidae ; B/ Les Décapodes Penaeidae, processidae et Palaemonidae ; C/ les Decapodes Nephropidae.
Abréviations : c.a. : cavité arthrodiale ; f.br. : fenêtre branchiale ; i.a. P1 : intrapleurite ; i.a. St : intra-sternite ; i.o. pl : intéropleurite ; i.o. St : intérosternite ; M.a. : membrane articulaire ; pi. : pilier ; P1 : pleurite ; pr. al. : processus aliforme ; r.i.o. P1 : repli intéropleural ; St : sternite ; T : tergite.
Fig. 1 à 6 : clichés J.J. Roccabianca Fig. 7 à 12 : clichés G. Morin Fig. 13 à 17 : clichés D. Serrette
73

74

75

76

77

78
■'ig. ii-j
Fig.13 DECAPODA Nephrops norvegicua
Nephropidae
79

| DECAPODA iNephronidael I Mephrops norvegicusI
80

iFig-J-T]
81

THÈME 2 - HORMONES ET MÉTABOLISME
- La neuroendocrinologie des Crustacés : Evaluation des progrès obtenus ces dernières années.
Van Herp F.
- Isoleraent et caractérisation d'une protéine merabranaire ovocytaire capable de fixer spécifiqueraent la vitellogénine.
Jugan P. et Van Herp F.
- Quelques aspects de la régulation du systèrae neuroendocrine produisant la CHH et de la relation entre le rythme circadien et la glycémie.
Kallen J.
- Effets de l'ablation des pédoncules oculaires, au cours des stades zoe III et IV sur la mue de raétaraorphose de Palaeraonetes varians (Leach) (Crustacea, Decapoda) .
Le Roux A.
- Purification des peptides apparentés à la calcitonine de sauraon chez les Crustacés. Résultats préliminaires.
Van Wormhoudt A.
83

ASPECTS RÉCENTS de la Biologie des CRUSTACÉS Actes de Colloques - 8 -1988 - IFREMER, pages 85 à 100
LA NEUROENDOCRINOLOGIE DES CRUSTACES: EVALUATION DES PROGRES OBTENUS CES DERNIERES ANNEES
VAN HERP F.
RESUME - Dans les recherches neuroendocrines chez les crustacés, l'applica-tion des méthodes modernes (chimie microanalytique, immunocytochimie, élec-trophysiologie et récemment aussi la biologie moléculaire) est de plus en plus fréquente et contribue à éclaircir la complexité du contrôle neuroen-docrine de différents processus physiologiques. Une attention particulière est accordée à l'hormone hyperglycémiante (CHH), à l'hormone inhibitrice (VIH) et stimulatrice (VSH) de la vitellogenèse, à l'hormone inhibitrice (MIH) de la mue et aux hormones peptidiques apparentées à certaines hor-mones de Vertébrés et d'Insectes. Les interactions réciproques de tous ces facteurs et la coordination avec des amines biogènes et des stéroldes sont signalées en relation avec le contrôle du métabolisme, de la reproduction et de la mue.
mots-clés : neuroendocrinologie, crustacés, progrès récent
ABSTRACT - The application of modern methods (microanalytical chemistry, immunocytochemistry, electrophysiology and recently also molecular biology) becomes more and more frequent in the study of the crustacean neuroendocri-nology and contributes to the elucidation of the complexity of the neuro-endocrinological control of different physiological processes. Particular attention is given to the crustacean hyperglycemic hormone (CHH), to the gonad inhibiting (GIH) and stimulating hormone (GSH), to the moult inhibit-ing hormone (MIH) and to the peptidergic factors related to some hormones from Vertebrates and Insects. The interactions of these factors as well as the coordination with biogenic amines and steroids in the control of the metabolism, reproduction and moulting are postulated.
key-words : neuroendocrinology, crustaceans, recent progress
De nombreuses recherches morphologiques et physiologiques sur le sys-tème neuroendocrine du pédoncule oculaire ont mis en évidence sa fonction régulatrice sur plusieurs processus vitaux (Cooke & Sullivan, 1982; Keller, 1983; Kleinholz, 1985; Quackenbush, 1986). Déjà en 1928, Hanström étudiait pour la première fois la morphologie et la cytologie du pédoncule oculaire des Crustacés décapodes.
En général, le système neuroendocrine du pédoncule oculaire contient plusieurs régions de cellules neuroendocrines, les organes X et un organe neurohémal, la glande du sinus où les terminaisons axonales de ces cellules accumulent et libèrent les neurohormones.
Figure 1 donne une impression de la composition de l'axe nerveux avec ses quatre ganglions optiques du pédoncule oculaire indiquant ces centres neuroendocrines (MTGX, MEX, X et SG).
Laboratoire de Zoologie, Faculté des Sciences, Université Catholique, Toernooiveld, 6525 ED Nimègue, Pays-Bas
85

Malgré plusieurs études en microscopie photonique et électronique de
ces centres neuroendocrines, la localisation exacte des sites de synthêse
des neurohormones, ne pouvait être donnée (Andrew, 1983; Chaigneau, 1983).
ABBREVIATIONS
HO = organe hémiellipsoidale
IN = neurons LG = lamina ganglionaris
ME = medulla externa
MEX = organe X de la ME
Ml = medulla interna
MT = medulla terminalis
MTGX = organe X de la MT
NO = nerf optique OC = chiasma optique
OM = ommatides SG = glande du sinus
X = organe X
Fig. 1. Schéma des ganglions optiques du pêdoncule oculaire de l'ëcrevisse indiquant les centres neuroendocrines
Grâce à deux antisérums du lapin, l'un développé par Jaros et Keller
(Jaros & Keller, 1979) contre l'hormone hyperglycémiante (CHH) du crabe,
Careinus maenas, et l'autre produit dans notre laboratoire (Van Herp & Van Buggenum, 1979; Kallen, 1985) contre la CHH de l'ëcrevisse, Astacue Zepto-daetylus, le système neuroendocrine synthétisant ce neuropeptide pouvait être localisé.
Des études combinêes en immunocytochimie et microscopie électronique complétaient l'identification de ce système chez l'écrevisse (Gorgels-Kallen & Van Herp, 1981; Gorgels-Kallen & Voorter, 1984). Un groupe de cel-lules, caractérisées par une accumulation du matériel immunoréactif, est situé dans la partie distale de l'MTGX. Les complexes de Golgi sont entou-
rés par des granules positives. Les fibres nerveuses immunopositives peuvent être suivies dans le tractus de l'MTGX vers la glande du sinus.
Dans cette région neurohêmale des gouttelettes immunoréactives sont disper-
sées de facon homogène et abondante. En ultrastructure, l'immunoréaction
est seulement concentrée dans le type de granule le plus grand (type 5-143 nm) .
En outre, les cellules neuroendocrines, produisant l'hormone hypergly-cémiante, sont injectées avec le "Jaune Lucifer" pour étudier la possibili-té de communication de ces cellules de CHH entre elles-mêmes où avec les autres cellules du système nerveux du pêdoncule oculaire. La micro-injec-tion de "Jaune Lucifer" dans les péricaryons des cellules de CHH chez l'écrevisse a montrê que les axones transportant le facteur hormonal, sont ramifiées dans la médulla terminalis (Gorgels-Kallen, 1985).
86

L'étude en microscopie électronique a révélé des figures synaptiques sur ces ramifications. Les terminaisons sont abondamment pourvues de gra-nules CHH et montrent des contactes synaptiques. Les axones présynaptiques sont en contact avec plusieurs embranchements de l'axone CHH. Dans les terminaisons de ces axones présynaptiques se trouvent deux types d'inclu-sions: a) des corps denses aux électrons d'environ 67 nm; b) des vésicules transparentes avec un diamètre de 32 nm.
N'ayant jamais dêtecté des figures synaptiques dans les autres régi-ons, nous supposons que ces ramifications sont responsables de la communi-cation entre les cellules nerveuses et les cellules de CHH.
A la suite d'analyses immunocytochimiques, en utilisant des anticorps contre des neuropeptides et des amines biogènes, une réaction immunopositi-ve était obtenue pour deux anticorps contre le Met-enképhaline et un anti-corps contre la sérotonine au niveau de la microscopie électronique. En microscopie photonique, des produits apparentés à la sérotonine (Elofsson, 1983) et à 1'enképhaline (cf. références dans le Tab. 1) étaient déjà loca-lisés dans le pédoncule oculaire d'autres Crustacés en utilisant des anti-corps spécifiques.
Les résultats de cette étude immunocytochimique et des effets physio-logiques de la sérotonine et de l'enképhaline sur l'activitê du système CHH et sur 1'hyperglycémie sont illustrés dans le résumé de ma collègue Kallen (cf. communication dans cette édition). Notre hypothèse de travail est que l'activité des cellules CHH est contrôlée par un facteur inhibi-toire: 1'enképhaline, d'une part et un facteur stimulatoire: la sérotonine, d'autre part.
En conclusion, par les travaux immunocytochimiques entrepris ces der-nières années par plusieurs chercheurs (Gorgels-Kallen et al., 1982; Mar-tin et al., 1984; Van Herp et al., 1984; Keller et al., 1985) en utilisant l'anti-CHH du crabe et l'anti-CHH de l'écrevisse, il a été possible d'étu-dier en détail la localisation du système neuroendocrine responsable de la synthèse de l'hormone hyperglycémiante chez différentes espèces de Crusta-cés comme généralisé dans le schéma de la localisation (Fig. 2).
87

Fig. 2. Schéma des ganglions optiques du pédoncule oculaire et la localisation du système hyperglycémiante
88

Ces dernières années, l'emploi d'antisérums contre des neuropeptides
de Vertébrés a mis en évidence chez les Invertébrés la présence de sub-
stances antigéniques apparentées, ouvrant ainsi de nouvelles perspectives
de recherche sur les relations entre ces produits et leur fonction.
Le pédoncule oculaire assure différents contrôles hormonaux chez les
Crustacés: c'est dans cet organe qu'on a principalement recherché la pré-
sence éventuelle de neuropeptides apparentés aux hormones de Vertébrés et
d'Invertébrés. Un résumé de ces études est donné dans le Tableau 1.
Quelques résultats que notre laboratoire a obtenus dans le pédoncule
oculaire de l'écrevisse (Van Deijnen et al., 1985) sont décrits pour il-
lustrer les implications possibles pour le fonctionnement neuronale des
ganglions optiques.
Fig. 3 a,b,c. Schéma du pédoncule oculaire de l'écrevisse indiquant la localisation des substances apparentées à l'a-MSH, CCK, sécrétine, vasotonine et FMRF-amide
Des substances apparentées à l'a-MSH, CCK et sécrétine sont locali-
sées en abondance dans les organes X. Quelques cellules nerveuses, qui ne
furent pas identifiées jusqu'ici comme peptidergiques sont mises en évi-
dence avec les mêmes anticorps dans le lamina ganglionaris, le chiasme op-
tique et les interneurones. Des fibres axonales semblent constituer la
connexion entre les cellules neuroendocrines et la glande du sinus, étant
également positive pour les trois anticorps (Fig. 3a). L'antivasotocine
reconnaît approximativement les mêmes sites, mais le grand nombre de fi-
bres est surprenant. La glande du sinus est aussi immunopositive (Fig. 3b) .
Les immunoréactions après l'usage de 1'anti-FMRF-amide donnent la même im-
pression. Des cellules et des axones peptidergiques sont localisées dans
les différentes régions. La glande du sinus reste négative (Fig. 3c).
En conclusion: l'utilisation des anticorps contre différents peptides
est favorable pour étudier la complexité du système peptidergique dans le
pédoncule oculaire des Crustacés. En plus, cette approximation donne aussi
89

Tableau 1 : Hormones peptidiques apparentées aux hormones (1e Vertébrés et d'Invertébrés.
peptide "like" activité biologique localisation méthode de détection
calcitonine métabolisme
du calcium?
pédoncule oculaire
intestin/hépato-pancréas
cyto
RIA/recept.
(1) *
• <S)
enkephalin neuromodulations? pédoncule oculaire cyto/phys (4,7,9)
gastrin/CCK cycle de mue?
l'estomac
pédoncule oculaire
cyto/
RIA/
isolation
(3,8,17
GIP ? pédoncule oculaire cyto (17)
glucagon ? pédoncule oculaire cyto (17)
insuline glycogenèse hépatopancréas RIA (13)
ocMSH ? pédoncule oculaire cyto (17)
neurophysine ? pédoncule oculaire cyto (18)
oxytocine ? pédoncule oculaire cyto (17)
90

Tableau 1 : (Suite)
peptide "like" activité biologique localisation méthode de détection
secrétine ? pédoncule oculaire cyto (17)
substance P ? pédoncule oculaire cyto (3)
somatostatine ? cerveaux cyto (11)
vasotocine ? pédoncule oculaire cyto (17)
vasopressine ? pédoncule oculaire cyto (18)
AKH (hormone adipokinétique)
activité "RPCH"
neuromodulateur? pédoncule oculaire
cyto/
phys/
isolation
(2,10, 12,14)
FMRF neuromodulateur? pédoncule oculaire cyto (4,6,1
proctoline effets musculaires organe péricardiale RIAAsolatian
(15flG cyto
* Références : (1) Bellon et al, 1984 ; (2) Bellon et al., 1986 ; (3) Favrel
et al., 1987 ; (4) Fingerman et al., 1985 ; (5) Fouchereau-Peron et al-, 1987
(6) Jacobs et Van Herp, 1984 ; (7) Jaros et al., 1985 ; (8) Larson et Vigna,
1983 ; (9) Mancillas et al-, 1981 ; (10) Mangerich et al., 1986 ; (11) Martin
et Dubois, 1981 ; (12) Mordue et Stone, 1976 ; (13) Sanders, 1983 ; (14)
Schooneveld et al., 1987 ; (15) Stangier et al., 1986 ; (16) Sullivan, 1979 ;
(17) Van Deijnen et al., 1985 ; (18) Van Herp et Bellon, 1982.
91

l'impression qu'il existe une diffêrence fonctionnelle entre ces cellules neuroendocrines des organes X et les cellules, principalement localisées dans les régions des interneurones. Nous supposons que ces dernières cel-lules ont une fonction dans la neuromodulation ou la neurotransmission.
Des résultats comparables sont obtenus chez d'autres espèces de Crus-tacés (références dans le Tab. 1).
On a récemment trouvé, que la communication entre les cellules pep-tidergiques se déroule par différents mécanismes de relais. II paraît que les messagers ne sont pas seulement transfêrés par des synapses ou par le sang, mais aussi par les espaces intercellulaires.
Les trajets qui peuvent être suivis par les messagers dans le pédon-cule oculaire des Crustacés, sont actuellement inconnus.
II y a quelques années que Van Deijnen étudiait la vascularisation des ganglions optiques chez l'écrevisse par perfusion avec l'encre de Chine. En combinaison avec quelques résultats de l'ultrastructure du pé-doncule oculaire, il pouvait postuler que les trois mécanismes de relais ne peuvent pas être exclus chez les Crustacés (Van Deijnen et al., 1984).
Dans le neuropile du medulla externa, par exemple, plusieurs régions contiennent des terminaisons axonales plein de granules et montrent régu-lièrement des figures d'exocytose, supposant une communication synaptique.
D'autre part, dans la région des interneurones du même medulla, on retrouve des terminaisons axonales entourées d'espaces dont on suppose que ce sont des espaces sanguins. Par immunocytochimie, une réaction positive est détectée avec l'anti-FMRF dans les mêmes sites.
De plus, l'arrangement du système vasculaire, comme illustré dans la Figure 4, indique qu'une partie des terminaisons axonales dans la région supêrieure des ganglions optiques est connectée avec la glande du sinus de manière neuro-hémo-neurale et que la libération de neuropeptides accumulés dans ce centre, peut être contrôlée par ce système.
Par contre, la perfusion du système sanguin du pédoncule oculaire et une étude microscopique de la région du medulla externa ont montré des es-paces intercellulaires non pénétrés par l'encre de Chine (Van Deijnen, 1986).
Fig. 4. Arrangement du système vas-culaire dans le pédoncule oculaire de l'écrevisse
92

La signification de ces dernières structures pour l'intégration neu-roendocrine du pédoncule oculaire est inconnue, mais elle entraîne plusi-eurs remarques d'ordre morphologique et fonctionnelle. C'est intéressant de signaler que ces structures sont localisées dans les mêmes régions où nous retrouvons les réactions immunocytochimiques pour différents anti-sérums de peptides. Ceci indique que plusieurs de ces peptides pourraient être libérés de manière non-synaptique dans l'espace intercellulaire.
Comme fut signalé dans l'introduction, plusieurs démarches sont actu-ellement suivies pour étudier les hormones peptidiques chez les Crustacés. Outre les études immunologiques, plusieurs laboratoires ont introduit les techniques de chromatographie liquide à haute pression (l'HPLC) pour iso-ler les peptides de la glande du sinus (Keller & Kegel, 1984) . Une atten-tion particulière a êté accordée, entre autres, à quelques hormones typi-ques de Crustacés comme l'hormone hyperglycémiante (CHH), à l'hormone in-hibitrice de la mue (MIH) et à l'hormone inhibitrice de la vitellogenèse (VIH). Un autre point d'intêrêt est l'étude des substances apparentées aux hormones de Vertébrés comme la calcitonine et les gastrines, par des tech-niques biochimiques (références dans le Tab. 1).
L'hormone hyperglycémiante des Crustacés a fait l'objet d'un certain nombre de travaux concernant sa séparation et sa purification comme ceux de Kleinholz (Kleinholz, 1976), Keller (Keller, 1981; et al.,1985), Huberman (Huberman & Aguilar, 1986), Newcomb (Newcomb, 1983) et Martin (Martin et al., 1984). Ainsi on dispose, actuellement, d'informations précises sur l'isolement et la composition en acides aminés.
L'hormone inhibitrice de la mue a été récemment étudiée par Webster et Keller (1986) chez Carcinus et par Chang et al. (1987) chez Homarus en utilisant l'HPLC.
Avec la même technique, l'isolement de l'hormone inhibitrice de la vitellogenèse est entrepris par deux équipes francaises, celle de Poitiers et celle de Paris. En collaboration avec la dernière, notre laboratoire a mis au point une méthode d'HPLC en phase inverse pour la séparation simul-tanée de plusieurs activités biologiques comme la MIH, RPCH, CHH et VIH à partir du même matériel de glande du sinus de homard (Soyez et al., 1982; Van Deijnen et al., 1984).
Récemment une approche nouvelle a êté développée dans notre groupe de recherche pour purifier et étudier chez le homard, la CHH, la VIH et le facteur VSH, dont on présume qu'il stimule la vitellogenèse. La séparation en HPLC est basée sur les caractères d'ionicité des molécules. Les essais "classiques" sont utilisés pour détecter les activités biologiques. Des essais hétérologues sont effectués en utilisant l'écrevisse Orconectes pour l'activité hyperglycémiante et la crevette Palaemonetes pour les ac-tivités contrôlant la vitellogenèse secondaire (VIH et VSH). En outre des essais nouveaux sont développés. Les fractions sont analysées en ELISA par usage des antisérums: l'anti-CHH de l'écrevisse et l'anti-VIH de homard. Ce dernier anticorps fut préparé par Meusy et collègues (Meusy et al., sous presse).
Les résultats après la première phase de séparation, basée sur l'io-nicite, sont montrés dans la Figure 5. Les fractions les plus importantes au point de vue de l'activité biologique sont localisées dans la deuxième partie du chromatogramme. L'activité hyperglycémiante est concentrée dans le double pic I et les pics VI/VII. Ces deux zones ont aussi une activité immunologique pour l'anti-CHH. Le polymorphisme de la CHH n'est pas sur-prenant, puisqu'il est décrit par plusieurs auteurs. L'existence d'une activité stimulatrice de la vitellogenèse est, par contre, remarquable parce que cette activité est localisée dans la même zone que le deuxième pic hyperglycémiant. Cet effet, direct ou indirect, reste à étudier.
93

difftrtncttfldiam. VIH/VSH
Fig. 5. Séparation des neuropeptides de la glande du sinus de
homard et analyse des activités biologiques et immuno-
logiques (A: séparation en HPLC/colonne TSK—SP-5PW; B:
activité hyperglycémique (CHH); C: activité ovocytaire
(VIH/VSH) ; D: activité immunologique (anti-CHH/anti-VIH)
94

Les résultats différents dans le bio-essai VIH/VSH pour les extraits bruts
de la glande du sinus peuvent être expliqués par l'interaction de l'effet stimulatrice/inhibitrice ainsi que par la variabilité du matériel, inclus
le lot des glandes et les animaux utilisés dans l'expérience. L'activité
VIH est plutôt retrouvée dans deux zones dont une possède aussi une immu-
noréactivitê pour l'anti-VIH. Pour le moment, on ne peut pas expliquer exactement pourquoi les zones VIII et IX réagissent avec l'anticorps. Pour
quelle raison la zone III ne réagit pas avec l'anti-VIH, reste à étudier. C'est important de noter ici que cette zone inhibe aussi la liaison de la
vitellogénine aux récepteurs membranaires de l'ovocyte vue l'essai "in
vitro", décrit par Jugan et Soyez (1985). Pour le moment des expériences additives pour séparer les produits
actifs, p.ex. basées sur les caractères d'hydrophobicité des molécules
sont en cours. Les plus amples informations sur ces dernières recherches
seront publiées dans un journal spécialisé. Pour le moment, on peut dire que le matériel du double pic de la pre-
mière phase (Fig. 5) a les mêmes caractères d'hydrophobocité, mais que
l'effet de température pendant l'extraction est remarquable. A 4°C, par
exemple, le matêriel se sépare en deux pics majeurs. Les deux ont une ac-
tivité immunologique, mais l'activité hyperglycémiante est seulement ac-
cordée au produit de poids moléculaire de 7.5 kDa après une électrophorèse
en SDS-urée. Jusqu'à maintenant nous ne pouvons pas accorder aucune acti-
vité biologique à l'autre pic. Que ce produit de 6.7 kDa ait une signifi-
cation biologique ou soit plutôt un artéfacte, reste à étudier. L'influ-
ence de la méthode d'extraction est minimale pour les zones VI/VII de la
première colonne. Les deux contiennent un produit prédominant ayant une
hydrophobicité identique. Ce matériel est immunoréactif pour l'anti-CHH,
hyperglycémiante et a aussi un effet stimulateur sur la vitellogenèse. La
pureté de ce matériel reste à vérifier, car après électrophorèse nous re-
trouvons deux fractions très voisines avec un poids moléculaire de 8.4 et
7.4 kDa. L'étude de leur rôle dans les deux activités biologiques est en
cours. II est frappant de constater que Chang et collègues (Chang et al.,
1987) ont récemment correlé une activité inhibitrice de la mue à un pepti-
de de 8.7 kDa chez le homard et que Webster et Keller (1986) signalent que
la MIH et la CHH de crabe sont très voisines. La zone IX de la première
colonne, ayant une activité biologique et immunologique pour la VIH, ne
montre aucune différence par rapport à la méthode d'extraction et corres-
pond à un peptide de poids moléculaire de 7 kDa. Soyez et Van Deijnen
(Soyez et al., 1982; Van Deijnen et al., 1984) ont rapporté environ le
même poids moléculaire pour la VIH du même matériel, mais séparé sur une
colonne en phase inverse. Le matériel du pic VIII, qui est immunoréactif
pour 1'anti-VIH, ressemble au pic IX par la fraction à poids moléculaire de 29.8 kDa. La possibilité que les fractions de 7 et de 29.8 kDa aient
des relations peptide-précurseur, reste à étudier.
Pour consolider les rêsultats présentés ici et pour étudier les in-
teractions de VIH et de CHH/VSH en dêtail, il nous semblait nécessaire de
développer des essais plus précis que l'essai "in vivo". Ainsi nous sommes
actuellement en train de développer des méthodes qui devront nous per-
mettre d'éviter la variabilité entre les animaux expérimentaux, d'examiner
dans des systèmes homologues et d'exploiter les interactions possibles de
ces facteurs hormonaux dans le cycle complet de l'ovocyte. Puisque la VIH inhibe la liaison de la vitellogénine aux récepteurs
membranaires de l'ovocyte, bloquant ainsi la première phase de l'endocyto-
se de cette lipoprotéine, nous avons développé deux essais basês sur ce
résultat. Le premier essai utilise la vitellogénine marquée à la peroxida-
se pour étudier le "UPTAKE". Le deuxième est mis au point à partir d'un
95

anticorps contre le récepteur membranaire. Les détails de l'antisérum pro-duit et les perspectives concernant son utilisation sont notés dans le ré-sumé de Jugan et Van Herp (cf. communication dans cette édition).
Comme conclusion générale, les données concernant les quatre neuro-peptides qui retiennent l'attention de plusieurs laboratoires et qui sont très proches par rapport à leurs caractères biochimiques sont rassemblêes dans la Figure 6. Ce schéma résume les interactions réciproques de ces neuropeptides et la coordination avec d'autres facteurs dans le contrôle de la mue et de la reproduction.
Fig. 6. Schéma résumant les interactions réciproques de MIH, CHH/"VSH" et de VIH et la coordination avec d'autres facteurs dans le contrôle de la mue et de la repro-duction
96

ANDREW R.D., 1983. Neurosecretory pathways supplying the neurohemal organs in Crustacea, in : Neurohemal ovgans of Arthropods, édité par A.P. Gupta, Charles C. Thomas Publishers, Springfield, 90-117. BELLON-HUMBERT C., VAN HERP F., SCHOONEVELD H., 1986. Immunocytochemical study of the red pigment concentrating material in the eyestalk of the prawn Palaemon servatus, Pennant, using rabbit antisera against the insect adipokinetic hormone, Biol. Bull., 171, 647-659. BELLON C-, VAN HERP F., VAN WORMHOUDT A., 1984. Localisation immunocyto-chimique de neuropeptides et d'amines biogènes dans le pédoncule oculaire de la crevette Palaemon serratus (Pennant), Ann. Soe. Roy. Zool. Belg., 114, 164. CHAIGNEAU J., 1983. Neurohemal organs in Crustacea, in : Neurohemal organs of Arthropods, éditê par A.P. Gupta, Charles C. Thomas Publishers, Spring-field, 53-89. CHANG E.S., BRUCE M.J., NEWCOMB R.W., 1987. Purification and amino acid composition of a peptide with molt-inhibiting activity from the lobster, Homarus amerioanus, Gen. Comp. Endoor., 65, 56-64. COOKE E.M., SULLIVAN R.E., 1982. Hormones and neurosecretion, in : The Biology of Crustaoea, édité par D.E. Blios, H.L. Atwood, D.C. Sandeman, Academic Press, Inc., New York, Vol. 3, 205-290. ELOFSSON R., 1983. 5-HT-like immunoreactivity in the central nervous sys-tem of the crayfish, Pacifastacus leniusoulus, Cell Tissue Res., 232, 221-236. FAVREL P., VAN WORMHOUDT A., STUDLER J.M., BELLON C., 1987. Immunochemical and biochemical characterization of gastrin/cholecystokinin-like peptides in Palaemon serratus (Crustacea Decapoda): intermoult variations, Gen. Comp. Endoor., 65, 363-372. FINGERMAN M., HANUMANTE M.M., KULKARNI G.K., IKEDA R., VACCA L.L., 1985. Localization of substance P-like, leucine-enkephalin-like, methionine-enkephalin-like, and FMRF amide-like immunoreactivity in the eyestalk of the fiddler crab, Voa pugilator, Cell Tissue Res., 241, 473-477. FOUCHEREAU-PERON M., ARLOT-BONNEMAINS Y., MILHAUD G., MOUKHTAR M.S., 1987. Immunoreactive salmon calcitonin-like molecule in Crustaceans: high con-centrations in Nephrops norvegious, Gen. Comp. Endoor., 65, 179-183. GORGELS-KALLEN J.L., 1985. Appearance and innervation of CHH-producing cells in the eyestalk of the crayfish Astaous leptodaotylus examined after tracing with Lucifer Yellow, Cell Tissue Res., 240, 385-391. GORGELS-KALLEN J.L., VAN HERP F., 1981. Localization of crustacean hyper-glycemic hormone (CHH) in the X-organ sinus gland complex in the eyestalk of the crayfish Astacus leptodactylus (Nordmann, 1842), J. Morphot., 50, 347-355. GORGELS-KALLEN J.L., VAN HERP F., LEUVEN R.S.E.W., 1982. A comparative im-munocytochemical investigation of the crustacean hyperglycemic hormone (CHH) in the eyestalks of some decapod crustacea, J. Morphol., 174, 161-168. GORGELS-KALLEN J.L., VOORTER C.E.M., 1984. Secretory stages of individual CHH-producing cells in the eyestalk of the crayfish Astacus leptodactylus, determined by means of immunocytochemistry, Cell Tissue Res., 237, 291-298. HANSTRÔM B., 1928. Vergleichende Anatomie des Nervensystems der Wirbel-losen Tiere, Springer, Berlin. HUBERMAN A., ANGUILAR M.B., 1986. A neurosecretory hyperglycemic hormone from the sinus gland of the Mexican crayfish Procambarus bouvieri (Ortmann) I. Purification and biochemical characterization of the most abundant form of the hormone, Comp. Biochem. Physiol., 85B, 197-203. JACOBS A.A.C., VAN HERP F., 1984. Immunocytochemical localization of a substance in the eyestalk of the prawn Palaemon serratus, reactive with an anti-FMRF-amide rabbit serum, Cell Tissue Res., 235, 601-605.
97

JAROS P.P., DIRCKSEN H., KELLER R., 1985. Occurrence of immunoreactive en-kephalins in a neurohemal organ and other nervous structures in the eye-stalk of the shore crab, Carcinus maenas L. (Crustacea, Decapoda), Celï Tissue Res., 241, 111-117. JAROS P.P., KELLER R., 1979. Immunocytochemical identification of hyper-glycemic hormone-producing cells in the eyestalk of Carcinus maenas, Cell Tissue Res., 204, 379-385. JUGAN P., SOYEZ D., 1985. Démonstration "in vitro" de 1'inhibition de la croissance ovocytaire par un extrait de glande du sinus chez la crevette, Maevobraehium rosenbergii, C.R. Aead. Se., 300, 705-709. KALLEN J.L., 1985. The hyperglycemic hormone producing system in the eye-stalk of the crayfish Astacus leptodactylus, Thèse Doct. Sei., Univ. Nijmegen, 127 p. KELLER R., 1981. Purification and amino acid composition of the hypergly-cemic neurohormone from the sinus gland of Creonectes limosus and compari-son with the hormone from Carcinus maenas, J. Comp. Physiol., 141, 445-450. KELLER R., 1983. Biochemistry and specificity of the neurohemal hormones in Crustacea, in : Neurohemal organs of Arthropods, édité par A.P. Gupta, Charles C. Thomas Publishers, Springfield, 118-148. KELLER R., JAROS P.P., KEGEL G., 1985. Crustacean hyperglycemic neuropep-tides, Amer. Zool., 25, 207-221. KELLER R., KEGEL G., 1984. Studies on crustacean eyestalk neuropeptides by use of high performance liquid chromatography, in : Biosynthesis, Metabol-ism and Mode of Aetion of Invertebrate Hormones, édité par J. Hofmann, M. Porchet, Springer-Verlag, Berlin, 146-154. KLEINHOLZ L.H., 1976. Purified hormones from the crustacean eyestalk and their physiological specificity, Nature, 258, 256-257. KLEINHOLZ L.H., 1985. Biochemistry of Crustacean Hormones, in : The Biolo-gy of Crustaaea, éditë par D.E. Bliss, L.H. Montel, Academic Press, Inc., New-York, Vol. 5, 464-522. LARSON B.A., VIGNA S.R., 1983. Gastrin/cholecystokinin-like immunoreactive peptides in the Dungeness crab, Cancer magister (Dana): immunochemical and biochemical characterization, Regul. Pept., 7, 155-170. MANCILLAS J.R., Mc GINTY J.F., SELVERSTON A.I., KARTEN H., BLOOM F.E., 1981. Immunocytochemical localization of enkephalin and substance P in re-tina and eyestalk neurones of lobster, Nature, 293, 576-578. MANGERICH S., KELLER R., DIRCKSEN H., 1986. Immunocytochemical identifica-tion of structures containing putative red pigment-concentrating hormone in two species of decapod crustaceans, CelZ Tissue Res., 245, 377-386. MARTIN G., DUBOIS P., 1981. A somatostatin-like antigen in the nervous system of an isopod, Poreellio dilatatus, Gen. Comp. Endocr., 45, 125-130. MARTIN G., KELLER R., KEGEL G., BESSE G., JAROS P.P., 1984. The hypergly-cemic neuropeptide of the terrestrial isopod, Porcellio dilatatus. I. Iso-lation and characterization, Gen. Comp. Endoer., 55, 208-216. MARTIN G., JAROS P.P., BESSE G., KELLER R., 1984. The hyperglycemic neuro-peptide of the terrestrial isopod, PoreeZlio dilatatus. II. Immunocytoche-mical demonstration in neurosecretory structures of the nervous system, Gen. Comp. Endoer., 55, 217-226. MEUSY J.J., MARTIN G., VAN DEIJNEN J.E., SOYEZ D., GALLO J.M., 1987. Immunochemical and immunocytochemical study of the crustacean vitello-genine-inhibiting hormone (VIH), Gen. Comp. Endoer., sous presse. MORDUE W., STONE J.V., 1976. Comparison of the biological activities of an insect and a crustacean neurohormone that are structurally similar, Nature, 264, 287-289. NEWCOMB R.w., 1983. Peptides in the sinus gland of Cardisoma earnifex: iso-lation and amino acid analysis, J. Comp. PhysioZ., 153, 207-221. QUACKENBUSH L.S., 1986. Crustacean endocrinology, a review, Can. J. Fish. Aquat. Soi., 43, 2271-2282.
98

SANDERS B., 1983. Insulin-like peptides in the lobster, Homarus america-nus. I. Insulin immunoreactivity, Gen. Comp. Endocr. , 50, 366-373. SCHOONEVELD H., VAN HERP F., VAN MINNEN J., 1987. Demonstration of sub-stances immunologically related to the identified arthropod neuropeptides AKH/RPCH in the CNS of several invertebrate species, Brain Research, 406, 224-232. SOYEZ D., FEYEN J., VAN DEIJNEN J.E., 1982. Preliminary results on the separation of Homarus americanus (L.) neuropeptides by high performance liquid chromatography using four bioassays, J. Endocr., 94, 23. STANGIER J., DIRCKSEN H., KELLER R., 1986. Identification and immunocyto-chemical localization of proctolin in pericardial organs of the shore crab, Carcinus maenas, Peptides, 7, 67-72. SULLIVAN R.E., 1979. A proctolin-like peptide in crab pericardial organs,
J. exp. Zool., 210, 543-552. VAN DEIJNEN J.E.M., 1986. Structural and biochemical investigations into the neuroendocrine system of the optic ganglia of decapod Crustacea, Thèse Doct. Sci., Univ. Nijmegen, 201 p. VAN DEIJNEN J.E., SOYEZ D., VAN HERP F., 1984. Purification simultanée et caractérisation préliminaire de quelques hormones: CHH, RPCH, GIH et MIH de la glande du sinus du homard, Homarus americanus (Edwards), Ann. Soc.
Roy. Zool. Belg., 114, 176-177. VAN DEIJNEN J.E., VAN DER STAPPEN J., RULAND M., 1984. Une étude ultra-structurale des ganglions optiques chez l'écrevisse Astacus leptodactyius:
nouvelles données concernant le système neuroendocrine, Ann. Soa. Roy.
Zool. Belg., 114, 177. VAN DEIJNEN J.E., VEK F., VAN HERP F., 1985. An immunocytochemical study of the optic ganglia of the crayfish Astacus leptodactylus (Nordmann 1842)
with antisera against biologically active peptides of vertebrates and in-vertebrates, Cett Tissue Res., 240, 175-183. VAN HERP F., BELLON-HUMBERT C-, 1982. Localisation immunocytochimique de substances apparentées à la neurophysine et la vasopressine dans le pédon-cule oculaire de Palaemon serratus (Pennant) (Crustacé Décapode Natantia), C.R. Acad. Sci., 295, 97-102. VAN HERP F., VAN BUGGENUM H.J.M., 1979. Immunocytochemical localization of hyperglycemic hormone (HGH) in the neurosecretory system of the eyestalk
of the crayfish Astacus leptodactylus, Experientia, 35, 1527-1528. VAN HERP F., VAN WORMHOUDT A., VAN VENROOY W.A.J., BELLON-HUMBERT C., 1984. Immunocytochemical study of crustacean hyperglycemic hormone (CHH) in the eyestalks of the prawn Pataemon serratus (Pennant) and some other Palaemo-nidae, in relation to variations in blood glucose level, J. Morphot., 182, 85-94. WEBSTER S.G., KELLER R., 1986. Purification, characterization and amino acid composition of the putative moult-inhibiting hormone (MIH) of Carci-nus maenas (Crustacea Decapoda), J. Comp. Physiol., 156B, 617-624.
99

ASPECTS RÉCENTS de la Biologie des CRUSTACÉS Actes de Colloques - 8 - 1988 - IFREMER, pages 101 à 104
ISOLEMENT ET CARACTERISATION D'UNE PROTEINE MEMBRANAIRE OVOCYTAIRE CAPABLE DE FIXER SPECIFIQUEMENT LA VITELLOGENINE
JUGAN P. VAN HERP F.
RESUME - La solubilisation de membranes ovocytaires chez l'écrevisse Ovco-nectes limosus a permis de mettre en évidence par SDS-PAGE et par êlectro-blotting un composant de poids moléculaire de 30 kDa, capable de fixer spécifiquement la vitellogénine. La nature protéique de ce site de liaison nous est suggérêe par sa sensibilité à l'action d'enzymes protéolytiques comme la pronase. Un anticorps polyclonal a été produit contre le composé membranaire de 30 kDa et la spécificité tissulaire de cet anticorps a été éprouvée par ELISA.
mots-clés : isolement, caractérisation, protéine membranaire ovocytaire, vitellogénine, écrevisse
ABSTRACT - After solubilization of ovocyte-membranes from the crayfish Ovconectes limosus, a component with a molecular weight of 30 kDa and cap-able to fix vitellogenin specifically, could be characterized by SDS-PAGE and by electroblotting. Because this isolated component is sensible to the digestion of proteolytic enzymes such as pronase, we assume that we are dealing with a protein. A polyclonal antiserum is produced against this component and its tissue-specificity is verified by ELISA.
key-words : isolation, characterization, membrane-protein, vitellogenin, crayfish
L'endocytose est un mêcanisme général utilisé par les cellules pour prélever de manière spécifique des molécules présentes dans le milieu ex-tracellulaire (Roth & Porter, 1964; Goldstein et al., 1976). l'un des sys-tèmes les plus connus est 1'internalisation de la vitellogénine par les ovocytes en vitellogenèse (Roth et al., 1976). La fixation de la vitello-génine sur des rêcepteurs membranaires ovocytaires est à l'origine de cette sélectivité (Yusko et al., 1981; Köning S Lanzrein, 1985; Rôhrkasten S Ferenz, 1986 a et b; Opresko S Wiley, 1987). La caractérisation du récep-teur ovocytaire à la vitellogénine a été effectuée chez la poule (Wood S Roth, 1980).
Chez les Crustacés, la croissance ovarienne est sous le contrôle d'une hormone inhibitrice de la vitellogenèse (VIH) (Kleinholz, 1985; Payen, 1986). Cette hormone régule le processus d'endocytose ovocytaire de la vitellogénine (Jugan S Soyez, 1985). Le mécanisme d’action cellulaire de cette hormone n'est pas connu, mais il est possible qua la VIH affecte la distribution et le recyclage des récepteurs ovocytaires à la vitello-génine.
Dans le but d'êtudier cette interaction avec plus de précision, nous avons isolé et partiellement caractérisé, chez l'écrevisse Ovconectos li-mosus, une protéine membranaire ovocytaire de 30 kDa et capable de fixer
Laboratoire de Zoologie, Faculté des Sciences, Université Catholique, Toernooiveld, 6525 ED Nimègue, Pays-Bas
101

spécifiquement la vitellogénine (VTG). Un anticorps polyclonal a été pro-
duit contre cette protéine et la spécificitê tissulaire de cet anticorps a ëtê ëprouvëe par microtitration (ELISA - Enzyme Linked Immunosorbent Assay).
Matériel et méthodes
Des ovocytes en vitellogenëse, provenant d'un ovaire de l'écrevisse Ovconectes limosus, sont homogénëisës dans un tampon phosphate de sodium (300 ovocytes/1 ml de tampon). Les membranes sont séparées du cytoplasme par centrifugation (10 000 g) et sont solubilisées dans un tampon Tris-HCl contenant du SDS (2%) et du g-mercaptoethanol (5%). Après une centrifuga-tion (40 000 g) le surnageant contenant les protéines membranaires solubi-lisées est soumis à une électrophorèse en condition dénaturante (Laemmli, 1970).
Les protéines membranaires sêparêes par êlectrophorêse sont transfé-rêes du gel de polyacrylamide à une feuille de nitrocellulose suivant la technique de 1 ' électroblotting.
Les bandes de nitrocelluloses sont incubées en présence de vitello-génine marquée à la peroxydase. La détection de la peroxydase est effec-tuée à l'aide d'un substrat, le 4-chloro-l-naphthol. Différentes conditi-ons ont été testêes: compétition par la vitellogénine native, par l'hémo-cyanine, prétraitement par la pronase.
Après localisation de la protêine membranaire fixant spécifiquement la vitellogénine, une électrophorèse prêparative a été rêalisée. Cent yg de protéine ont étê isolés par élution et injectés à un lapin. L'anticorps produit a étê testé en immunoblotting et par microtitration ELISA.
Rêsultats et conclusion
Parmi de nombreux constituants protéiques extraits des membranes ovo-cytaires et sêparês par êlectrophorèse (Pl. I, Fig. a), un composé de poids moléculaire 30 kDa est capable de fixer la vitellogénine marquée à la peroxydase (P1. I, Fig. b). La spécificité de la liaison a été verifiée par compétition avec de la vitellogénine non marquée (Pl. I, Fig. c) et avec de 1'hémocyanine (Pl. I, Fig. d): seule la vitellogénine native dé-place la liaison de la vitellogénine marquée, l'hémocyanine est sans effet. La dégradation préalable des protéines membranaires par la pronase (P1. I, Fig. e) inhibe la liaison de la vitellogénine sur le site de 30 kDa.
Un anticorps de lapin a été produit contre cette protéine de 30 kDa. Cet anticorps a été testé en immunoblotting (Pl. I, Fig. f). Des membranes solubilisées provenant d'ovaire, de testicule, de muscle et d'hépatopan-créas ont été testées par ELISA en utilisant les anticorps purifiés à par-tir de l'antisérum total et marqués à la peroxydase (Fig. 2 a-d). Seules les membranes ovariennes réagissent avec l'anticorps.
Fig. 2. Immunoréactivitê des membranes d'ovaire (a),
de testicule (b) , de muscle (c) et d'hépato-
pancréas (d) estimée par un lecteur ELISA à
492 nm
102

PLANCHE I
Fig. a-f. Des membranes ovocytaires solubilisées soumises à une électro-phorèse SDS-PAGE et à une électroblotting
Légendes
a. Séparation des protéines membranaires ovocytaires par électrophorèse b-d. Incubation du "ruban nitrocellulose" en présence:
(b) de la vitellogénine marquée à la peroxydase (VTG-HRP); (c) de VTG-HRP et d'un excès de vitellogénine native; (d) de VTG-HRP et d'un excès de l'hémocyanine.
e. Préincubation du "ruban nitrocellulose" dans la pronase suivie d'une incubation en présence de VTG-HRP.
f. Incubation avec le sérum de lapin anti-protéine 30 kDa suivie d'une incubation avec du sérum de chèvre anti-lapin marqué à la peroxydase
103

En conclusion, il a été possible de caractériser et d'isoler à partir
de membranes ovocytaires, une protëine de 30 kDa capable de fixer spécifi-
quement la vitellogénine. La production d'un anticorps contre cette proté-
ine nous permet maintenant d'envisager, par une méthode immunoenzymatique,
le dosage de cette protéine au cours des cycles de vitellogenèse primaire
et secondaire. La biosynthêse de cette protéine ainsi que sa mise en place
dans la membrane plasmique ovocytaire seront êtudiées à l'aide de techni-
ques biochimiques, immunocytochimiques et de biologie moléculaire.
GOLDSTEIN J.L., BASU S.K., BRUNSCHED G.Y., BROWN M.S., 1976. Release of
low-density lipoprotein from its cell surface receptor by sulfated gly-cosaminoglycans, Cell, 7, 85-95. JUGAN P., SOYEZ D., 1985. Démonstration in vitvo de l'inhibition de l’en-
docytose ovocytaire par un extrait de glande de sinus chez la crevette Macrobraehium rosenbergii, C.R. Acad. Sci., 20, 705-709. KLEINHOLZ L.H., 1985. Biochemistry of crustacean hormones, in : The Biology of Crustacea, édité par D.E. Bliss, L.H. Mantel, Academic Press, New York,
9, 463-522. KÔNING R., LANZREIN B., 1985. Binding of vitellogenin to receptors in
oocyte membrane preparations of the ovoviviparous cockroach Nauphoeta cinenerea, Inseot Biochem., 15, 735-737. LAEMMLI U.K., 1970. Cleavage of structural proteins during the assembly of
the head of bacteriophage, Nature, 221, 680-685. OPRESKO L.K., WILEY H.S., 1987. Receptor-mediated endocytosis in Xenopus
oocytes. I. Characterization of the vitellogenin receptor system, J. Biol. Chem., 262, 4109-4115. PAYEN G.G., 1986. Endocrine regulation of male and female genital activity in crustaceans. A retrospect and perspectives, Adv. Invert. Repr., 4,
125-134. RÔHRKASTEN A., FERENZ H.J., 1985a. In vitro study of selective endocytosis of vitellogenin by locust oocytes, Roux's Arch. Dev. Biol., 194, 411-416. RÔHRKASTEN A. , FERENZ H.J. , 1985b. Solubilization of the locust vitello-genin receptor, Biochim. Biophys. Acta, 860, 577-582. ROTH T.F., CUTTING J.A., ATLAS S.B., 1976. Protein transport: a selective
membrane mechanism, J. Supramol. Struct., 4, 527(487)-548(508). ROTH T.F., PORTER K.R., 1964. Yolk protein uptake in the oocyte of the mosquito Aedes aegypti L., J. Ceï-Z Biot. , 20, 313-332. WOOD J.W., ROTH T.F., 1984. A specific subunit of vitellogenin that medi-ates receptor binding, Biochem. J., 23, 5774-5780. YUSKO S., ROTH T.F., 1976. Binding to specific receptors on oocyte plasma membranes by serum phosvitin-lipovitellin, J. Supramol. Struct., 4, 89-97.
104

ASPECTS RÉCENTS de la Biologie des CRUSTACÉS Actes de Colloques - 8 -1988 - IFREMER, pages 105 à 108
QUELQUES ASPECTS DE LA REGULATION DU SYSTEME NEUROENDOCRINE PRODUISANT LA CHH ET DE LA RELATION ENTRE LE RYTHME CIRCADIEN ET LA GLYCEMIE
KALLEN J.
RESUME - La teneur en glucose dans l'hémolymphe de l'écrevisse montre un rythme circadien se traduisant par une augmentation du glucose pendant la nuit. Les résultats d'une étude consacrée à la régulation de ce phénomène montrent que ce rythme est synchronisé par la photopériode, détecté ni par les yeux ni par le photorécepteur caudal, mais par un autre récepteur si-tué dans le pédoncule oculaire. Le rythme est transmis dans l'organisme par une horloge interne, probablement localisée dans les ganglions opti-ques, qui peut maintenir indépendamment le rythme durant plusieurs cycles. Les informations entre le milieu externe et interne doivent être trans-mises aux cellules neuroendocrines produisant l'hormone hyperglycémiante. Notre étude a montré que les injections de sérotonine et dopamine pro-voquent une hyperglycémie et que l'injection de met-enképhaline réduit la quantité de glucose dans l'hémolymphe de même que l'augmentation de son rythme circadien. Ces neurotransmetteurs ont été localisés par immunocyto-chimie dans le pédoncule oculaire, en relation avec le système neuroendo-crine produisant la CHH.
mots-clés : écrevisse, CHH, rythme circadien, neurotransmetteurs, immuno-cytochimie
ABSTRACT - In crayfish, the hemolymph glucose level shows an endogenous circadian rhythmicity which is expressfed by a nocturnal glucose rise. The results of a study on the regulation of this phenomenon indicate that this rhythm is entrained by the light/dark schedule. These light stimuli are not detected by the compound eyes nor by the caudal photoreceptor but by a receptor located elsewhere in the eyestalk. The endogenous clock modulat-ing this rhythmicity most probably is located within the optic lobes. In-formations about the external light/dark situation have to be transmitted to the CHH cell system. This study shows that injections of serotonin and dopamin result in hyperglycemia and that injections of met-enkephalin de-crease hemolymph glucose level as well as the nocturnal circadian glucose peak. Using immunocytochemistry, these neurotransmitters have been located in the eyestalk and their relationship with the CHH cell system is des-cribed.
key-words : crayfish, CHH, circadian rhythm, neurotransmitters, immuno-cytochemistry
Chez l'écrevisse, le système neuroendocrine produisant l'hormone hy-perglycémiante (CHH = Crustacean Hyperglycemic Hormone) est localisé dans le pédoncule oculaire. Ce système est constitué de cellules neurosécrétri-ces qui font partie de la MTGX (Medulla Terminalis Ganglionic X-organ).
Laboratoire de Zoologie, Faculté des Sciences, Université Catholique, Toernooiveld, 6525 ED Nimègue, Pays-Bas
105

L' application de techniques immunocytochimiques nous a permis de localiser les cellules dans la partie latéro-ventrale et la partie distale de la MTGX. La CHH est transportée par un tractus vers la glande du sinus (Van Herp & Van Buggenum, 1979; Gorgels-Kallen & Van Herp, 1981). Une première étude a montré que, dans des conditions photopériodiques constantes (LD 12 : 12), le système hyperglycémiante a un rythme circadien en relation avec l'activité synthétique des péricaryons, le transport des granules neurosécrétoires vers la glande du sinus et la libération de CHH dans 1'hémolymphe. Ces phénomènes se traduisent par une augmentation du glucose dans le sang environ quatre à six heures après le début de la nuit (Gorgels-Kallen & Voorter, 1985).
Le but du travail présenté ici est d'obtenir des informations plus détaillées sur le rôle de la photopériode dans la régulation du rythme hyperglycémiant circadien et sur la localisation du (des) photorécepteur(s) et du (des) oscillateur(s) chez l'écrevisse Astacus leptodactylus.
En plus, nous avons étudié la présence dans le pédoncule oculaire et l'effet biologique de substances connues comme étant des neurotransmet-teurs/neuromodulateurs, pouvant avoir un rôle dans la transmission des in-formations du milieu externe et interne vers les cellules à CHH, chez les écrevisses Astacus leptodactylus et Orconectes limosus.
Méthodes
Le rythme circadien du taux de glucose dans l'hémolymphe est déter-miné sur une période de 24 heures sous les conditions suivantes: 1. une photopériode normale (LD 12 : 12); 2. l'obscurité continue pour 6, 11 et 35 jours; 3. une phase-shift de 12 heures; l'hémolymphe a été collectée après 6, 12 et 18 jours; 4. les ommatides masqués pour 35 jours; 5. les pédon-cules oculaires masqués pour 35 jours; 6. les pédoncules oculaires y com-pris la zone rostrale du céphalothorax masqués pour 35 jours; 7. la dis-ruption du nerf optique; l'hémolymphe a été collectée après 35 jours.
La prêsence des amines biogènes et des enképhalines est examinée im-munocytochimiquement (méthode du PAP) aq» çjiveau de la microscopie photo-nique en utilisant des antisérums spécifiques contre la sérotonine, la dopamine (les deux antisérums nous sont mis en disponibilité par Dr. H.W.M. Steinbusch, Université Libre d'Amsterdam) et la met-enképhaline (cet antisérum nous est mis en disponibilité par J.H.H.M. van Daal, Labo-ratoire de Physiologie Animale, Nimègue). Pour étudier leurs effets bio-
logiques sur la glycémie, ces substances ont également été injectées.
Résultats
Les résultats montrent que le rythme circadien de la teneur en gluco-se dans l'hémolymphe de l' écrevisse peut persister pendant plusieurs cy-cles en l'absence de toute photopériodicité (même après 11 jours d'obscu-rité continue). Pour une durée plus importante (35 jours), l'absence des signaux photoniques induit la disparition du rythme glycémique et provoque une diminution de la glycémie. Le rétablissement d'une phase-shift se tra-duit par une adaptation très lente du rythme glycêmique à la nouvelle pho-topériode. Même 18 jours après la phase-shift, l'adaptation n'est pas en-core complète. L'absence de perception de la lumière par les ommatides ne réduit pas le rythme circadien. Par contre, après masquage des pédoncules oculaires, les mesures montrent une augmentation très faible du glucose pendant la nuit. Le masquage des pédoncules oculaires, y compris le rostre, donne des résultats identiques aux mesures obtenues dans une obscurité continue de 35 jours. La section du nerf optique ne modifie pas le rythme circadien du glucose dans 1'hémolymphe.
106

Par immunocytochimie on peut visualiser la présence de sérotonine, de
dopamine et de met-enképhaline dans le pédoncule oculaire. Notamment dans
le neuropile de la MTGX, dans la région des ramifications des cellules à
CHH, on distingue des axones immunopositives pour les trois substances.
Les injections de sérotonine mais aussi de dopamine provoquent des augmen-
tations de teneur en glucose dans le sang. L'injection de met-enképhaline
diminue la teneur en glucose dans l'hémolymphe et diminue de plus l'hyper-
glycémie nocturne.
Conclusion
Le rythme circadien de la teneur en glucose d'Astaous leptodactylus
est synchronisé par la photopériode et transmis dans l'organisme par une
horloge interne qui peut maintenir, indêpendamment du milieu, le rythme
pendant plusieurs cycles. Par consëquent ce rythme peut être considêré
comme endogène. La périodicité n'est détectée ni par les yeux ni par le photorécep-
teur caudal, mais probablement par un autre récepteur situé dans le pédon-
cule oculaire. Des sections sélectives au niveau du nerf optique ont montré que
cette horloge interne est localisée dans les ganglions optiques.
La présence des axones immunopositives pour la sérotonine, la dopa-
mine et la met-enképhaline et, de plus, leurs effets biologiques sur la
glycémie de l'écrevisse, portent à croire que ces substances peuvent être
impliquées dans la régulation du rythme circadien neuroendocrine des cel-
lules à CHH et la glycémie.
GORGELS-KALLEN J.L., 1985. Appearance and innervation of CHH-producing
cells in the eyestalk of the crayfish Astacus leptodactylus examined after
tracing with Lucifer Yellow, Cell Tissue Res., 240, 385-391. GORGELS-KALLEN J.L., VAN HERP F., 1981. Localization of Crustacean Hyper-
glycemic Hormone (CHH) in the X-organ sinus gland complex in the eyestalk
of the crayfish, Astacus leptodactylus (Nordmann, 1842), J. Morphol., 170,
347-355. GORGELS-KALLEN J.L., VOORTER C.E.M., 1985. The secretory dynamics of the
CHH-producing cell group in the eyestalk of the crayfish, Astacus lepto-
dactylus, in the course of the day/night cycle, Cell Tissue Res., 241,
361-366. VAN HERP F. , VAN BUGGENUM H.J.M., 1979. Immunocytochemical localization of
hyperglycemic hormone (HGH) in the neurosecretory system of the eyestalk
of the crayfish Astacus leptodactylus, Experientia, 35, 1527-1528.
107

ASPECTS RÉCENTS de la Biologie des CRUSTACÉS Actes de Colloques - 8 - 1988 - IFREMER, pages 109 à 112
EFFETS DE L'ABLATXON DES PEDONCULES OCULAIRES , AU COURS DES STADES ZOE III ET ZOE IV . SUR LA MUE DE METAMORPHOSE DE PALAEMONETES VARIANS (LEACH) (CRUSTACEA, DECAPODA) .
LE ROUX A.(1)
RESUME - La présence de deux (ou au moins d'un) pêdoncules oculaires est nécessaire à l'induction d'une métamorphose complète, monoecdysiale de Palaemonetes varians . A 20°C, cette induction est réalisée durant un laps de temps de 20 heures environ, situé de part et d'autre de la troisième mue, dans le cas d'un développement en 4 stades zoé- Le mécanisme de l'induction est discuté.
Mots-clés . métamorphose, mue, pédoncule oculaire, Palaemonetes varians .
ABSTRACT - The presence of two (or at least one) eyestalks is required for inducing a complete, monoecdysial, metamorphosis in Palaemonetes varians . This induction is performed, at 20°C, in about 20 hours situated on either side of the third larval moult when metamorphosis occurs at the fourth one. The mecanism of induction is discussed.
Key-words : Metamorphosis, molting, eyestalks, Palaemonetes varians .
Costlow (cf Costlow, 1968) a montré que l'ablation des pédoncules oculaires (P.O.) retarde mais ne supprime pas la métamorphose des Brachyoures Sesarma reticulatum et Rhithropanopeus harrisii . Cet effet retardateur de l'ablation des P.O. sur Ta métamorphose à également été observé chez les Caridea (Little, 1969 ; Le Roux, 1984) et le horaard américain (Charmantier et coll., 1985 ; Snyder et Chang, 1986). Les mécanismes endocriniens responsables de cet effet, et ceux qui induisent la métamorphose elle-même, n'ont pas encore été élucidés.
Costlow (1968) suggère que les P.O. libèrent une hormone responsable de la transformation morphologique des zoés et des mégalopes. Cette hormone serait élaborée dans un site extra-pédonculaire puisque la métamorphose n'est pas supprimée par la pédonculectomie. Freeman et Costlow (1983) émettent l'hypothèse de l'intervention d'un facteur inhibiteur de la métamorphose (M.I.F. = Metamorphosis-Inhibiting Factor) qui s'opposerait aux transformations morphologiques en fin de phase zoé. Ce facteur à effets négatifs serait antagoniste de facteurs positifs (inducteurs) dont l'intervention serait programmée par l'intermédiaire de la crustecdysone.
Dans nos élevages, le développement larvaire de Palaemonetes varians s'accomplit normalement en quatre stades zoé (développement standard). Les deux épines dorsales postérieures de la larve se lysent au cours du stade DQ (Le Roux, 1984). Ceci est l'indicateur d'une induction de la larve dans une direction juvénile.
(1) Université de RENNES I Station de Biologie Marine de Bailleron, 56860 SENE
109

Nous avons tenté de savoir :
1) Si l'induction est alors totale, étant donné que des organes larvaires (les exopodites des péréiopodes en particulier) restent Eonctionnels jusqu'à la fin du stade larvaire.
2) A quel moment commence cette induction.
3) Quel est l'effet de l'ablation des P.O. sur le phénomène.
Chez des larves de P. varians élevées dans des conditions standard et donc destinées à se métamorphoser à la fin du stade Zoé IV, nous avons procédé à l'ablation des P.O. à différents moments des stades Zoé III et Zoé IV.
RESULTATS
Le moment où l'opération a été réalisée et les résultats obtenus sont indiqués sur la figure 1.
Z' 3° 4° Mue ' Stade III Mue Stade IV Mue Stade I
D0' D0" D1' D1" °2 D0' D0' D1' D
1" D2
.. larves.
1.1 larves.
1.2 1 1
2.1 t ► laret.
t , 1.3 Exo.Po : fonctionnels Ep. : larvaires
2.2 Chimères.cf. ci.dessus
t -6 t
Chimères. Pt.1.5 soies t t Exo.Pe : début
t régression
» t Ep : ± lar..juv.
t t
A1.2 ► Chimèrcs . cf. ci.dessus
c , , t
► Juvéniles.
INDUCTION de le
METAMORPHOSE (20h env. i 20°C)
Fig. 1 - Palaemonetes varians . Moment de l'ablation des P.O. dans les différents lots et résultats obtenus. Ep. : épines, Exo. : exopodites, Juv. : juvéniles, Lar. : larvaires, Pl. : pléopodes, Pe. : péréiopodes.
110

Il apparaît que lorsque l' ablation est réalisée avant l'étape D1"
du stade zoé II (lotsA,3-4
, 1-1, 1-2 et 2-1) on obtient, au stade V1des
individus larvaires, chez Iesquels la métamorphose a été retardée.
Entre l'étape D1" et la mue des zoés III (lots 1-3 et 2-2), l'ablation conduit, au stade V, à des animaux qui combinent des caractères larvaires et des caractères juvéniles. Nous les désignons du nom de chimères. Les pléopodes 1 à 4 possèdent un nombre variable de soies, les exopodites des péréiopodes demeurent fonctionnels, les deux épines médio-dorsales du céphalothorax et les épines latérales du pléonite n° 5 restent du type larvaire.
Les larves opérées entre la 3ème mue et l'étape D0
" du stade zoé IV (lots 2-3 et Al-2) produisent, au stade V, des chimères chez lesquelles les caractères juvéniles sont plus marqués que ci-dessus et les caractères larvaires plus régressés.
Les animaux opérés en D0
" et en début de D1 du stade zoé IV (lot C), chez lesquels on peut observer in vivo la lyse des deux épines médio-dorsales de la carapace, donnent au stade V des juvéniles normaux.
On peut donc considérer que la métamorphose est totalement induite dès l'étape DQ„ du stade zoé IV. Cette induction commence à partir de D „ du stade zoé III et s'accomplit en 20 heures environ à 20°C.
Ce créneau d'induction se retrouve de part et d'autre de la 4ème mue (avec des résultats légèrement moins nets) chez des animaux destinés à se métamorphoser à l'issue du stade zoé V.
Par ailleurs, en présence d'un seul P.O., l'induction est soit totale soit partielle (production de chimères).
DISCUSSION ET CONCLUSION
1 - Notre expérimentation confirme que la présence de deux (ou du moins d'un) P.O. est indispensable pour que la métamorphose s'accomplisse totalement en une seule mue (métamorphose monoecdysiale).
2 - La position de la période d'induction perraet d'avancer plusieurs hypothèses la concernant :
Elle pourrait résulter d'une dépression particulièrement importante du taux d'ecdystérone, au cours de la mue inductrice, sous l'action d'une forte production de M.I.H. (Molt Inhibiting Hormone) au niveau des P.O. . Le quatrième intermue est en effet fréquemment plus long que les précédents et l'ablation des P.O. supprime cet allongement.
- Elle pourrait être due à la levée d'une inhibition d'origine extra-pédonculaire (M.I.F. de Freeman et Costlow)sous l'effet des P.O. au voisinage de la 3ème mue.
- On peut envisager qu'elle est attribuable à un facteur inducteur d'origine extra-pédonculaire, libéré sous contrôle des P.O. .
Enfin 1 ' intervention des organes mandibulaires (0. Md) que Laufer et coll. (1987) ont identifié comme étant la source d'une hormone juvénile chez les Décapodes doit être envisagée.
Les 0. Md sont présents à l'état paucicellulaire au cours de la phase zoé, leur activation par ablation des P.O. pourrait entraîner une
111

libération d'hormone juvénile, hormone du statu quo chez les Insectes (Willis, 1981) et donc inhibitrice de la métamorphose. Cependant, comment expliquer alors que la métamorphose puisse se réaliser progressivement au cours des raues suivantes, alors que l'hyperfonctionnement (ou du moins l' hypertrophie) des 0. Md se maintient ou se développe (cf Le Roux, 1983).
La même hypertrophie des 0. Md après pédonculectomie rendrait très improbable la possibilité qu'ils soient à l'origine d'un facteur inducteur de la métaraorphose. En outre, on s'explique mal qu'ils puissent secréter un facteur inhibiteur (M.I.F.) puisque lorsque la pédonculectoraie est réalisée précocement (stade zoé II) la métamorphose n'est pas définitivemnt supprimée.
La nature et le site de production du (ou des ?) facteur responsable de la raétaraorphose des Décapodes deraeure encore très énigmatique. La connaissance précise de la période au cours de laquelle s'accomplit l'induction du phénoraène peut contribuer à apporter des réponses aux questions qui se posent à son sujet.
CHARMANTIER G., CHARMANTIER-DAURES M. et AIKEN D.E., 1985. Intervention des pédoncules oculaires dans le contrôle de la métamorphose chez Homarus americanus H. Milne Edwards, 1987 (Crustacea, Decapoda). C.R. Acad. Sci. 300, n° 7, 271-276.
C0STL0W J.D. Jr, 1968. Metamorphosis in crustaceans. In Metamorphosis,ETKIN and GILBERT, L.I. ed., North Holland pub.lishing C°, Amsterdam, pp. 3-41.
FREEMAN A.J. and COSTLOW J.D., 1983. Endocrine control of spine epidermis resorption during metamorphosis in crah larvae. Roux's Arch. Dev. Biol. 19.2, 362-365.
LAUFER H., BORST D., BAKER F.C., CARRASCO C., SINKUS M., REUTER C.C., TSAI L.W., SCH00LEY D.A., 1987. Identification of a juvenile-like compound in a crustacean. Science' 235, 202-205.
LE ROUX A., 1983. Réactions de 1'organe mandib.ulaire à l'ablation des pédon-cules oculaires. chez les larves et les juvéniles de Palaemonetes varians (Leach) (Decapoda, Natantia). C.R. Acad. Sci. Paris. 296, 697-700.
LE ROUX A., 1984. Quelques effets de l'ablation des pédoncules oculaires sur les larves et les premiers stades juvéniles de Palaemonetes varians. Dull Soc. Zool. Fr., 109, 1, 43-60.
RathhE G., 1969.Thelarval development of the shrimp, Palaemon macrodacti,lus
Rathbun, reared in the laboratory, and the effect of eyestalk extirpation on
molting rates and CHANG E.S., 1986 Effects of eyestalk ablation on larval Biol. ZT° of the american WILLIS J.H., 1981 . Juvenile hormone : the status of "status quo". Amer. Zool. ,
112

ASPECTS RÉCENTS de la Biologie des CRUSTACÉS Actes de Colloques - 8 - 1988 - IFREMER, pages 113 à 120
PURIFICATION DE PEPTIDES APPARENTES A LA CALCITONINE DE SAUMON CHEZ LES CRUSTACES RESULTATS PRELIMINAIRES-
A- VAN WORMHOUDT(1)
RESUME - Des peptides réagissant avec un anticorps anti-calcitonine de saumon ont été mis en évidence par immunocytochimie dans le pédoncule
oculaire et par radioimmunoessai dans l'hémolymphe et le tube digestif. 2 types de molécules ont été purifiées des molécules de masse
moléculaire comprises entre 3 et 5 kDa et des molécules de masse moléculaire
22 kDa pouvant correspondre à des précurseurs. Leur caractérisation est en
cours.
Mots clés . peptides hormonaux, calcitonine, crustacés.
ABSTRACT - In Crustacean, immunoreactive peptides were detected with salmon-calcitonin antibodies by immunocytochemistry in its eyestalks and by
radioimmunoassay in the haemolymph and the digestive tract. Two different molecules were purified : the first with their molecular
weight coraprised between 3-5 kDa, the second having a molecular weight of 22
kDa which could correspond to a precursor of the first one. Their
characterization is on progress.
Key-words . hormonal peptides, calcitonin, crustaceans.
L'utilisation de sondes immunologiques a perrais , ces dernières années,
la mise en évidence de nombreux peptides apparentés aux horraones de vertébrés
chez les invertébrés et plus particulièrement chez les Crustacés (Van-Deynen
et al. , 1985). Nous nous sommes intéressés à la calcitonine-like étant donné l'importance du métabolisrae du calcium chez les Crustacés. Chez les vertébrés
la calcitonine est un petit peptide de 3500 Da, purifié initialement à partir
de glandes thyroïdes de rat en 1963 (Coop, 1979), puis chez les poissons à
partir de glandes ultimobranchiales. Son action principale chez les vertébrés
supérieurs est hypocalcéraiante mais elle peut agir également sur les
secrétions du tube digestif (Holz et al , 1973 ; Drack et al , 1976). Chez
le poisson son rôle est moins certain. Son existence a été démontrée dans
le tube digestif de la Cione (Fritsch et al. ,1980).
1. LOCALISATION IMMUNOCYTOCHIMIQUE CHEZ LES CRUSTACES
Dès 1984, en collaboration avec C. Bellon et F. Van-Herp des molécules
apparentées à la calcitonine de saumon ont été mises en évidence dans les
cellules neurosecrétrices du pédoncule oculaire et la glande du sinus de
Palaemon serratus . Ces résultats ont été confirmés par la suite chez
Homarus americanus (voir photo) avec le même anticorps spécifique de la
calcitonine de saumon, et ne reconnait pas la partie N-terminal commune à
toutes les calcitonines, situées au niveau du site actif de la molécule.
(I) Laboratoire de Biologie Marine du Collège de France, 29110 CONCARNEAU, Francc.
113

Planche
Localisation immunocytochimique des cellules apparentées à la calcitonine de saumon chez Palaemon serratus (A-B-C) et Homarus americanus (D.E.F.).
L'anticorps contre la calcitonine de saumon est dilué au 1/500. A - partie supérieure de la glande du sinus (G.S) immunoréactive contre la Medulla externa (M.E). B - quelques cellules de la Medulla terminalis (organe X1) C - cellules de la Medulla externa D - quelques axones terminaux disséminés dans toute la glande du sinus. E - F - Cellules et son tractus localisés dans la Medulla terminalis : notez
la longueur du tractus qui rejoint la glande du sinus.
114

115

2. CARACTERISATION BIOCHIMIQUE
Chez Palaemon serratus les molécules ont été mesurées dans l'hémolymphe par radioimmunoessai en utilisant le même anticorps, dans l'hémolymphe en collaboration avec Arlot-Bonnemains et Fouchereau-Péron. Leur caractérisation a été poursuivie chez la crevette et également le homard et la langoustine où des quantités très importantes ont été mesurées (Fouchereau-Péron et al ., 1985 , Van-Wormhoudt et al. , 1987).
^ • Peptides apparentés immunologiquement à la calcitonine de saumon chez la crevette Palaemon serratus - Filtration : A) G 50 SF hémolymphe : 0,5 ml , B) G 50 SF Meduila externa : 50 glandes ; C) G 50 SF Medulla terminalis : 50 glandes. (colonne 140 x 7 cm : acétate MH, 10 mM pH 8.6) ; D) HPLC molécules 3-5000 Da isolées par filtration de l'hemolymphe. (Colonne ultrtuphère ODS Beckman). hCT . Calcitonine humaine ; sCT = calcitonine de saumon ; Alb = albumine bovine ; Rase = Ribonucléase pancréatique ; Ins : insuline.
Chez P. serratus , dans l'hémolymphe, séparée sur G 50 SF des molécules
de 3 à 5000 Da coexistent avec des molécules plus grosses traduisant une
certaine hétérogénéité. On retrouve les molécules de petite masse moléculaire
dans le pédoncule oculaire au niveau de la Medulla terminalis et de la
Medulla externa (Fig. 1 A-B-C). C'est cependant dans l'hépatopancréas que
116

les quantités les plus importantes ont été mesurées (130 + - 40 ng/g PF). Par HPLC, en phase reverse (ODS) , les petites molécules de l'hépatopancréas sont éluées par 18 % de n-propanol, à un niveau différent de la calcitonine de saumon traduisant une moins grande hydrophobicitê et des différences de structure (Fig. 1D).
Chez Homarus americanus des quantités de 3,9 +- 1,5 ug/g PF ont été mesurées dans le pédoncule oculaire. Cependant, une filtration d'extraits acide acétique 0,1 M montre que 90 % des molécules sont de grosses molécules de taille voisine de 20 kDa (Fig. 2A). Par HPLC des molécules migrant au même niveau que la calcitonine de crevette ont été retrouvées en quantités très faibles et correspondent sans doute aux petites molécules (Fig. 2B).
Fig- 2 : Peptides apparentés immunologiquement à la calcitonine de saumon chez le Homard. A) filtration sur colonne G 50 SF d'un extrait de pédoncule oculaire (1 g. PF) ; B) HPLC d'un extrait de glande du sinus repris dans du n-propanol 16 %, centrifugé puis passé sur colonne ODS (Beckman). Les valeurs de CT correspondent à 1 équivalent de glande du sinus.
Fig. 3 : Peptides apparentés immunologiquement à la calcitonine de saumon
chez la langoustine. Filtration - A) G 50 F Hépatopancréas (23 g) ; B) G 50 F
hémolymphe (50 ml équivalent) ; C) G 50 F estomac (10 g) (colonne 140 x 2,5 cm
acétate NH^ 10 mM, pH 8.6) ; D) Desoxyribonucléase ; R = ribonucléase ; L
= Insuline ; sCT = calcitonine de saumon ; G 17 = gastrine 17.
117

Chez Nephrops norvegicus (Fig. 3) , des quantités importantes sont extraites en milieu acide acétique 0.1 M et correspondent essentiellement à de grosses molécules dans les différents tissus étudiés, hémolymphe (1 ug/ml), hépatopancrêas (44,6 +- 10,7 ug/g PF) et estomac (4,5 +- 2,1 ug/g PF).
Dans le cas de l 'hépatopancréas et de l'hémolymphe, les molécules de masse moléculaire comprises entre 22 et 24 kDa, ont été purifiées et caractérisées par leur composition en acides aminés (Van Wormhoudt et Fouchereau-Péron, 1987). Un anticorps a pu être obtenu qui permettra peut être de mieux déterminer l'origine de ces molécules.
DISCUSSION
La nature hormonale de certaines molécules apparentées à la calcitonine est mise en évidence dans le cas des molécules de petite masse moléculaire localisées dans les cellules neurosecrétrices du pédoncule oculaire. Ces petites molécules extraites à partir d'un extrait HCl de langoustine (Fouchereau-Péron et al ., 1987) réagissent corapétitivement au niveau des raembranes de foie de rat avec la calcitonine de saumon.
La nature hormonale des grosses molécules (22-24 kDa), isolées à partir du tube digestif de langoustine, en milieu acide acétique 0.1 M, reste à préciser même si des molécules identiques sont retrouvées dans l'hémolymphe. Dans ce cas, en effet, en tenant compte des résultats de la composition en acides aminés il faudrait admettre des concentrations circulantes d'hormone—like de 50 ug/ml jamais trouvées chez les vertébrés, même si, dans le cas de malades atteints de tumeurs épidermo-bronchique, le taux de calcitonine augmente fortement et en particulier celui des grosses molécules de prohormone de 12 à 25 kDa. Chez les crustacés, l'hypothèse d'une prohormone circulante peut être faite dans la mesure ou des expériences réalisées au laboratoire par Y. Arlot-Bonnemains montrent que la précipitation par un anticorps contre la calcitonine de sauraon des produits de la traduction in vitro des ARNm extraits de l'oesophage de langoustine donne essentiellement des molécules de 24 kDa. L'hypothèse de l'autoaggrégation de petites molécules où de leur liaison à des grosses molécules de transport peut être écartée ; aucune dissociation n'étant possible par électrophorèse SDS.
ARLOT-BONNEMAINS Y. , VAN-WORMHOUDT A., FAVREL P. et M. FOUCHEREAU-PERON, 1986. Calcitonin-like peptide in the shrimp Palaemon ie.AAaXui , during the intermolt cycle .ExpeAÀeuitLa., 42, 419-420. ARL0T-B0NNEMAINS Y., PER0N M. , FAVREL P. et A. VAN-WORMHOUDT ,1985. Caractérization d'un peptide apparenté à la calcitonine chez Palaemon serratus : Variation au cours du cycle de mue et analyse chromatographique. Cali. Biol Marine. (sous presse). BELLON C., VAN-HERP F. et A. VAN-WORMHOUDT, 1984. localisation immunocytochimique de neuropeptides et d'amines b.iogènes dans le pédoncule oculaire de la crevette Palaemon serratus ■ Ann. Soc. Row. Zool. Belg. , 114, suppl. 1, p. 164. a g COOP D.H.,1979 . Calcitonin comparative endocrinology in EndocAÀnoZogij V. 2 ed. L. de Groet Grune et Stratton Inc. N.Y., 130 p. 637-651. DRACH G.I. , KOELZ H.R. and A.L. BLUM, 1976. Human calcitonin stimulates salivary amylase output in man. Gut . 7, 620-623.
118

FOUCHEREAU-PERON M. , ARLOT-BONNEMAINS T. , MILHAUD G. et M.S. MOUKHTAR,1987. Immunoreactive salmon calcitonin-like molecule in Crustacean : high concentrations in Nephrops norvegicus . Gcn. i Comp. Endocrinol. > 65, 179-183. FRITSCH M.A.R., VAN NOORDEN S. and A.G.E. PEARSE,1980. Calcitonin-like immunochemical staining in the alimentary stract of Ciona intestinalis '■
CeXZ & Tissue Res. 205, 439-444. HOLZ. J., HINNE H. and R. ZIEGLER, 1973. The influences of acute hyper and hypocalcemia and of calcitonin on exocrine pancreatic fonction in man. Résearch in Exp. Med., 160, 152-165. VAN-DEYNEN J.E., VEK F. and F. VAN-HERP,1985. An immunocytochemical study of the optic ganglia of the crayfish AbXacub leptodactylus with antisera against biologically active peptides of vertebrates and invertebrates. Cett. Tissue Res. , 240, 175-183.
VAN-WORMHOUDT A. et M. FOUCHEREAU-PERON,1987. Partial characterization and amino acid composition of a high molecular weight peptide with salmon calci tonin immunoreacti vi ty in a crustacean. Wephrops norvegicus • Biockem. Biophys . Res. Comm., 148 (1) 463-470.
Ce travail a bénéficié d'une subvention CNRS CR L 72
119

THÈME 3 - ÉCOPHYSIOLOGIE - ADAPTATION
- Contrôle neuromodulateur et flexibilité d'expression d'un réseaux neuronal central : le système pylorique des crustacés décapodes.
Nagy F., Cazalets J.R. et Moulins M.
- Approche écophysiologique et ultrastructurale de l'osmorégulation chez les Crustacés décapodes.
Pequeux A. et Lignon J.
- Flux unidirectionnels d'ions Na et C1 dans les branchies isolées et perfusées du crabe vert Carcinus maenas .
Wanson S. et Pequeux A.
- Variation de la consommation d'oxygène au cours du nycthémère chez Armadillidium vulgare
Chentoufi A. et Mocquard J.P.
- Etude des facteurs de l'environnement (température et photopériode) sur la reproduction de différentes souches d' Armadillidium vulgare .
Souty-Grosset C., Mocquard J.P.,Juchault P. et Hafidi N.
- Ecophysiologie de la reproduction de la crevette des marais Palaemonetes varians
Bouchon D.
- Structures adaptatives à la filtration des aliments chez le crabe des sables Scopimera gordonae
Bauchau A.
121

ASPECTS RÉCENTS de la Biologie des CRUSTACÉS Actes de Colloques - 8 - 1988 - IFREMER, pages 123 à 136
CONTROLE NEOROMODOLATEUR ET FLEXIBILITE D'EXPRESSION D'UN RESEAUX NEURONAL CENTRAL : LE SYSTEME PYLORIQUE DES CRUSTACES
DECAPODES.
NAGY F., CAZALETS J.R. ET MOULINS M.O)
RESUME - L'activité rythmique du filtre pylorique des crustacés décapodes est programmée dans un réseau de 14 neurones du système nerveux stomatogastrique . L'activité du réseau pylorique repose sur des propriétés rythmogènes endogènes de ses neurones et sur l'arrangement de leurs relations synaptiques. Ces deux facteurs sont sournis au contrôle de neurones modulateurs centraux. Deux de ces neurones ont pu être identifiés, le neurone APM et le neurone PS, qui l'un induit et l'autre supprime les propriétés régénératives des neurones pyloriques. APM peut ainsi démarrer et PS arrêter l'activité rythmique pylorique, mais tous deux peuvent également modifier son expression. Grâce à ce contrôle, le réseau pylorique peut élaborer plusieurs types de patterns moteurs.
mots-clés: crustacés, stomatogastrique, oscillations, modulation.
AJBS..TEAC1 - In decapod crustacea, the rhythmic activity of the pyloric filter is prograrnmed within a network of 14 neurons in the stomatogastrlc nervous system. The pyloric network activity is underlied by both the endogenous regenerative properties of its neurons and the organization of their synaptic connections. Both factors are controlled by central modulatory neurons. Two of these neurons were identified, the APM neuron which induces and the PS neuron which suppresses the regenerative properties of pyloric neurons. APM can start and PS can stop the rhythmic pyloric activity, but both of them can also change its expression. Because of this control, the pyloric network can produce several types of rnotor pattern.
key-words: crustacea, stomatogastric, oscillations, modulation.
1. Introduction.
Savoir comment une séquence motrice est organisée dans un centre nerveux est une des questions rnajeures qui intéressent les neurobiologistes. C'est grâce à l'utilisation d'un certain nombre de préparations nerveuses d'Invertébrés et en particulier de Crustacés, qu'on possède maintenant des éléments de réponse à
(1) Laboratoire de Neurobiologie et Physiologie Comparées, CNRS et Université de Bordeaux I, Place Dr Peyneau, 33120 ARCACHON, France.
123

Figure 1: Le système nerveux stomatogastrique et le réseau pylorique. A. Représentation schématique de l'intestin antérieur des grands crustacés en vue latérale gauche; seule la musculature pylorique a été représentée. B. Diagramme de la préparation isolée_in_vitro C. Relations synaptiques inhibitrices au sein du réseau pylorique. D. Enregistrements intracellulaires des trois types de neurones pyloriques. Barres verticales : 10 mV; barre horizontale : 1 sec. APM, neurone modulateur pylorique antérieur; CG, OG, STG, ganglions commissuraux, oesophagien et stomatogastrique; vlvn, nerf moteur pylorique; stn, nerf stomatogastrique; D, Cl, C2, muscles pyloriques innervés respectivement par les neurones PD, LP, PY.
124

cette question. On sait ainsi depuis un certain temps qu'une activité motrice rythmique est programmée au sein d'un réseau de neurones, et que l'activité de ce réseau repose à la fois 1) sur l'arrangement des relations synaptiques entre ses neurones; 2) sur des propriétés bioélectriques particulières de ces neurones, comrne par exemple la faculté de décharger rythmiquement de façon endogène (Selverston et Moulins, 1985).
On a pu montrer plus récemment qu'en fait ni l'un ni l'autre de ces facteurs déterminant n'est fixe. Par exempie, certaines propriétés des neurones peuvent être induites, amplifiées, réduites ou supprimées. Ces variations peuvent conduire au démarrage ou à l'arrêt de l'activité rythmique d'un réseau. Cela peut également conduire à ce que les neurones d'un réseau ne participent pas forcément tous à l'élaboration d'un pattern moteur donné, ou à ce que, quand ils y participent, les relations synaptiques qu'ils échangent voient leur efficacité varier. C'est à dire qu'un réseau sur le plan structural peut correspondre à plusieurs réseaux sur le plan fonctionnel. En d'autres termes un réseau peut produire plusieurs types de pattem rnoteur.
Une des préparations qui a précisément permis de dégager ces concepts est le système nerveux stomatogastrique des Crustacés Décapodes (Selverston et Moulins, 1987).
2. Le système nerveux stomatogastrique et son réseau pylorique.
Le système nerveux stomatogastrique contrôle les mouvements rythmiques de l'intestin antérieur (oesophage, estomac). L'estomac est, chez les crustacés, subdivisé en une poche gastrique et une chambre pylorique. C'est dans la poche gastrique (ou moulinet gastrique) que se fait le broyage des aliments par trois puissantes dents cuticulaires, tandis que dans la chambre pylorique les particules alimentaires sont filtrées vers l'intestin moyen. La musculature (de type strié) des dents gastriques et celle du filtre pylorique sont cornmandées par deux réseaux de neurones (respectivement 11 et 14 neurones) situés dans le ganglion stomatogastrique (STG). Ce ganglion est placé sur la face dorsale de l'estomac dans la lumière de l'aorte dorsale. Le STG est relié à la chaîne nerveuse ventrale de l'anirnal au niveau des ganglions commissuraux et ceci via le rrerf stornatogastrique (stn) qui est le seul nerf afférent au STG. C'est également via le stn qu'il est relié au ganglion oesophagien (Fig.1A).
Une particularité du système nerveux stomatogastrique est qu'une fois isoié ia vitro, il reste actif et continue à produire des activités rythmiques tout à falt semblables à celles qu'on peut enregistrer sur l'anirnal intact par électromyographie. Sur ce type de préparatiorr isolée, (Fig.lB), il a donc été possible d'étudier en détail l'organisation des réseaux gastrique et pylorique par ies méthodes de l'électrophysiologie intracellulaire. Pour cela, le STG est au préalable dégainé et les corps cellulaires de ses neurorres sont empalés, sous contrôle dune loupe binoculaire, par des microélectrodes de verre rernplies d'uri
125

électroiyte. Simultariérnent les patterns moteurs élaborés par ies réseaux du STG sont enregistres sur différents nerfs rnoteurs â l'aide d électrodes extracellulaires en piatine.
Le réseau pylorique qui est de loin ie mieux connu, rnanifeste une activité triphasique rythrnique avec mise en jeu successive, au cours de chaque cycle, de 3 neurones (PD-AB) commandant ies muscles dilatateurs (D) puis du neurone LP qui commande les rnuscies constricteurs antérieurs (Cl), et, enfin des 8 neurones PY qui cornmandent les muscles constricteurs postérieurs (C2). Dans chaque groupe les neurones sont électriquement couplés; les divers groupes de neurones sont associés par des inhibitions réciproques (Fig.lC) (Miller, 1987).
3. Modulation des propriétés neuronales dans le réseau pylorique: Démarrage, contrôle et arrêt d'un générateur de pattem moteur.
3.1. Les neurones pyloriques manifestent des propriétés ry thmogène s.
Le caractère rythmique de l'activité du réseau pylorique est lié au fait que ses neurones manifestent des propriétés bioélectriques particuüères qui leur confèrent des capacités rythmogènes. C'est en particulier le cas des neurones PD-AB qui sont des oscillateurs endogènes. Leur potentiel de membrane oscille régulièrement et ceci corxduit le neurone à émettre sporxtanément et rythmiquement une salve de potentiels d'action chaque fois qu'il franchit le seuil de décharge. La fréquence est dépendante du voltage et augmente régulièrement lorsqu'orx dépolarise la cellule. Dans ia rnesure où les neurones PD-AB (neurones dilatateurs) inhibent les neurones constricteurs (en particulier LP et PY, cf. Fig. 1D), ces derniers sont rnis au silence par chacune de ces salves. Les neurones constricteurs ne sont, pas eux-mêmes des neurones passifs. En fait leur potentiel de membrane manifeste deux niveaux de stabilité, l'un (hyperpolarisé) où le neurone est silencieux, l'autre (dépolarisé) où il décharge (neurones dit à "plateaux"). Le passage d'un niveau à l'autre est un phénomène actif, régénératif, qui peut étre déclanché par une brève dépolarisation (passage du niveau de repos au niveau de décharge) ou une brève hyperpolarisation (passage du niveau de décharge au niveau de repos). Les propriétés de plateaux influencent la durée et l'intensité des salves de décharge des neurones constricteurs ainsi que leur phase dans la séquence rythmique pylorique (Russell et Hartline, 1982).
Les diverses propriétés bioélectriques des neurones pyloriques qui jouent un rôle prirnordial dans l'organisation de l'activité du réseau présentent une autre particularité fondamentale. Elles ne peuvent s'exprimer que dans certaines conditions
126

3.2. L'activité du réseau dépend d'afférences centraies: Les propriétés rythmogènes des neurones pyloriques sont conditionneile s.
Sur les préparations in Vitro comme celi.es de la Figure 1B, le réseau pylorique est toujours actif. Mais ceci ne reste vrai que si le ganglion stomatogastrique est relié aux centres antérieurs (gangiions oesophagien et commissuraux). En effet, si on sectionne i.e nerf stomatogastrique qui assure cette liaison ou si l'on bloque la conduction dans ce nerf en intercaliant sur son trajet un compartirnent dépourvu d'ions (soiution isotonique de saccharose), i'activité pylorique cesse. Tous les neurones deviennent silencieux ou déchargent de manière tonique. Si la conduction dans le nerf storaatogastrique est restaurée, l'activité rythmique du réseau pylorique réapparaît (Mouiins et Cournil, 1982). On doit donc en conclure que certaines afférences spontanément actives et empruntant le nerf stornatogastrique sont nécessaires à l'expression de l'activité rythmique du réseau. On peut d'ailleurs confirmer cette manière de voir en stimulant le nerf à l'arrière du compartimerit utiüsé pour bloquer la conduction: Dans ces conditions le réseau redevient rythmiquement actif. Par ailleurs, une stimulation tonique est suffisante pour obtenir ce résultat, ce qui montre que les afférences assurant l'activité du réseau ne lui apportent pas de données temporelles rnais seulement une activation.
L'inactivation du réseau après déafférentation du STG est due au fait que, dans ces conditions, tous les neurones pyloriques perdent la capacité d'exprimer leurs propriétés rythmogènes. Ainsi pour les neurones dilatateurs (PD-AB) qui sont des oscillateurs endogènes, le blocage de conduction dans le nerf stornatogastrique se traduit non seulernent par un arrét des osciilations, mais également par une totale incapacité d'osciller. Dépoiarisés, ces neurones ne manifestent plus qu'une décharge tonique, caractéristique d'un neurone "passif". Après blocage de ia conduction dans le nerf stomatogastrique tous les autres neurones pyloriques, qui sont des neurones à "plateaux", perdent eux aussi leurs propriétés régénératives et se comportent en neurones "passifs" à décharge tonique.
On doit donc en conclure que l'arrét de l'activité du réseau après blocage du nerf stomatogastrique est lié à cette perte de propriétés régénératives. Ces propriétés des neurones pyloriques sont dites coriditionnelies, leur expressiori étant sourrdse à des afférences provenant des centres aritérieurs. Comme nous allons ie voir, une de ces afférences conditionnarite s a pu être identifiée. il s'agit de l'interneurone modulateur pylorique antérieur, APM.
3.3. Contrôle permiss.if des propriétés rythmogènes pyloriques par ie neurone moduiateur APM.
Le neurone modulateur pyiorique antérieur (APM) est un interneurone unique dont ia décharge peut. modifier profondément et durablement 1 activité du réseau pyiorique en modifiant les
127

Figure 2: Contrôle modulateur permissif du réseau pylorique par le neurone APM. A. La décharge du neurone APM déclenche l'activité rythmique du réseau pylorique. Al) Le réseau pylorique est silencieux; A2) Le neurone APM décharge de façon tonique et le rythme pylorique démarre. B. Une décharge phasique brève (5 sec) du neurone APM active fortement et durablement le rythme pylorique. Barres horizontales : 2 sec; barres verticales : 20 mV.
128

propriétés rythmogènes des neurones de ce réseau. Le corps cellulaire du neurone APM est situé dans ie gangiion oesophagien. De ce corps ceiiulaire partent deux axones qui traversent chacun un autre centre prémoteur (ganglion commissural) avant de se projeter dans ie gangiion stomatogastrique par le neri' stomatogastrique (Fig.lB).
3.3.1. Induction dea propriétés rythmogènes des neurones pyloriques par le. neurone APM.
Comme nous l'avons déjà indiqué, ies neurones pyloriques perdent leurs propriétés régénératives lorsque le gangiion stomatogastrique ne reçoit plus d'afférence provenant des centres antérieurs. Chez les langoustes Jasus lalandii et Palinurus vulgaris il est heureusement possible de supprimer toutes les afférences permissives d'origine commissurale, qui sous-tendent normalernent les capacités rythmogènes endogènes des neurones pyloriques, sans avoir à bloquer la conduction axonale dans le nerf afférent stomatogastrique. II suffit pour cela de bloquer l'activité synaptique dans les ganglions commissuraux par un liquide physiologique dépourvu d'ions calcium et additionné d'ions cobalt. Dans ces conditions l'intemeurone APM peut toujours influencer le réseau pylorique (sa conduction axonale étant préservée daris le nerf afférent) alors que les neurones pyloriques n'élaborent plus d'activité rythmique et se comportent comme des neurones passifs. Une décharge relativement brève d'APM (2 à 5 sec) à basse fréquence (inférieure à 30 Hz) rétablit alors, pour tous les neurones pyloriques, la capacité de produire des dépolarisations régénératives. C'est à dire qu'elle rétablit les propriétés osciilantes des neurones pacemakers et les propriétées de plateaux des neurones constricteurs. Tous les neurones pyloriques redeviennent donc, sous l'influence du neurone APM, capable de produire rythmiquement, et de façon endogène, des salves de potentiel d'action (Dickinson et Nagy, 1983).
L'induction par le neurone APM des propriétés de tous les neurones du réseau pylorique entraîne pour ce dernier une conséquence fonctionnelle fondamentale: la seule décharge du neurone APM peut déclencher l'activité rythmique d'un réseau pylorique préalablement silencieux (Fig.2A).
Un certains nombre d'arguments expérimentaux montrent qu'APM infiuence directement ("monosynaptiquement j chaque neurone pylorique. D'autre part les effets d'APM présentent des caractéristiques temporelles particulières. Premièrement ces effets s'établissent avec une latence relativement importante (une à plusieurs secondes), et ceci bien que les potentiels d'action d'APM atteignent le ganglion stomatogastrique avec un délai proche de 100 rasec. Deuxièmement, ces effets sont durables. Ainsi la faculté pour les neurones pyloriques de produire des dépolarisations régénératives se prolonge plusieurs dizaines de secondes après une décharge de quelques secondes du neurone APM. Par conséquent l'action du neurone APM, dont la décharge peut induire durablement la faculté d'exprimer des conductances
129

voltage-dépendantes pour tous les neurones du réseau pylorique, s'apparente à un processus de neurornodulation.
Eri fait les effets d'APM ne sont pas uniquement de type "tout ou rien", c'est à dire qu'APM peut également moduler les propriétés régénératives des neurones pyloriques lorsque ceux-ci les exprirnent déjà spontanérnent. APM riest par conséquent pas seulement capable de faire démarrer l'activité rythmique dun réseau pylorique silencieux, rnais peut également, corame nous allons le voir, contrôler finement l'expression d'une activité rnotrice pylorique spontanée.
3.3.2. Modulation des propriétés rythmogènes des neurones pyloriques. par le neurone APM.
Bien que la décharge d'APM soit suffisante pour induire la facuité des neurones pyloriques à produire rythmiquement des dépolarisations régénératives, cette décharge n'est pas indispensable pour que le rythme pylorique s'exprime. Bans la plupart des préparations isolées du système nerveux stornatogastrique le neurone APM est silencieux, or le réseau pylorique continue à élaborer son activité rythrnique normale, pour autant que le ganglion stornatogastrique reste connecté aux ganglions commissuraux. Cependant, dans ces conditions, la mise en jeu expérimentale du neurone APM provoque toujours une forte augrnentation des propriétés oscillatoires des neurones pyloriques. Cet effet se traduit par l'augmentation de tous les pararnètres du rythme pylorique qui dépendent des propriétés oscillatoires Cela provoque une augrnentation de la fréquence d'oscillation (et donc des salves de décharge) des neurones pyloriques, de l'arnplitude de ces oscillations et de la fréquence des potentiels d'action dans les salves (Fig.2B). II est à noter que tout comme pour l'induction des propriétés oscinatoires des neurones pyloriques, l'arnplifications de ces propriétés par la décharge d'APM est un phénomène durable qui se prolonge plusieurs dizaines de secondes après urie décharge d'APM ne durant. elle-même que quelques secondes (Fig.2B) (Nagy et Dickinson, 1983).
Le contrôle exercé par ie neurone APM ne se résume cependant pas seulernent à une activation générale du rythme pylorique, mais lui confère égalernent une grande flexibilité d'expression. En effet les différents neurones pyloriques rnanifestent des sensibilités différentes à sa décharge , les neurones ies pius sensibles étant les neurones constricteurs PY. En fonction de la frequence et de ia durée de la décharge d'APM, les propriétés régénératives peuvent ètre amplifiées pour un nombre variable de neurories dans le réseau, incluant ou non les neurories du groupe pacemaker. En conséquence la déeharge du neurone APM peut provoquer soit une augmentation de l'intensité des décharges des neurones constricteurs sans modifier le rythne pylorique, soit une augmentation à la fois de l'intensité de décharge de t.ous les neurones pyloriques et une accélération de la fréquence du rythme, so.it rnème arréter momentanérnent le rythrne pylorique et induire une interise décharge tonique des
130

neurones constricteurs. Gette flexibilité dans l'élaboration du
pattem moteur pylorique, consécutive à la décharge du neurone
APM, doit vraisemblablement se traduire chez l' animal par une
plus grande richesse du registre comportemental du fiitre
pylorique.
Or cette flexibilité est encore accrue par le fait que la
décharge d'APM entraine également une rnodification de la
sensibiiité des neurones pyloriques aux influences synaptiques.
Sans entrer dans le détail, nous pouvons dire que cette
modification de sensibilité conduit à augmenter le contraste des effets synaptiques sur les neurones pyloriques en augmentant la réponse de ces neurones aux influences fortes et en diminuant
leur réponse aux influences faibles. L'interneurone APM permet ainsi aux neurones pyloriques de filtrer les influences synaptiques qu'ils subissent (Nagy et Dickinson, 1983).
En résuraé, l'interneurone APM modifie l'efficacité relative des relations synaptiques dans le réseau pylorique, ce qui entraîne un recablage fonctionnel de ce réseau. Etant donné que tous les éléments du réseau pylorique (à l'exception d'un seul) sont des motoneurones, le recablage fonctionnel provoqué par APM est un moyen efficace de moduler le comportement moteur du filtre pylorique.
Les effets du neurone APM ont été relativement détaillés ici d'une part parce qu'ils sont exemplaires et d'autre part parce qu'APM est actuellement, dans le système nerveux stomatogastrique, le seul neurone exerçant de tels effets a être véritablement identifié . On sait cependant que le STG est également sournis au contrôle d'un grand nombre de fibres neuromodulatrices à action similaire mais dont l'origine n'a pas encore pu être précisée. One expérimentation combinant des techniques immunohistochimiques et pharmacologiques a ainsi révélé que les neurones pyloriques sont soumis a un contrôle modulateur permissif de la part, entre autres, de fibres aminergiques (serotonine, dopamine, octopamine), peptidergiques (proctoline, FMRFamide, substance P) et cholinergiques (Flamm et Harris-Warrick, 1986; Marder, 1987).
3.4. Controle suppressif des propriétés rythmogènes pyloriques par le neurone modulateur PS.
Nous venons de voir qu 'un intemeurone unique (APM) exerçant un contôle rnodulateur de type permissif sur les propriétés cellulaires des neurones du réseau pyloriques, pouvait faire démarrer l'activité rythmique de ce réseau, l'accélérer ou la modifier. En fait, ce réseau est également soumis à l'influence de neurones exerçant un contrôle modulateur diamétralement opposé à celui du neurone APM. Ainsi chez Homarusgammarus, un interneurone unique, le neurone pylorique suppresseur (PS), peut interrompre durablement ou diminuer fortement le rythme pylorique en supprimant ou en atténuant les propriétés régénératives de certains neurones du réseau. Le corps cellulaire de ce neurone est également situé dans le ganglion oesophagien.
131

Figure 3: Contrôle modulateur suppressif du réseau pylorique par le neurone PS. A. Diagramme de la préparation. B. Une décharge phasique brève (6 sec) du neurone PS arrête durablement l'activité rythmique du réseau pylorique. C. Une décharge tonique du neurone PS modifie le nornbre de neurones participant au rythme pylorique. Barres horizontales : 2 sec en A, 0.5 sec en B; barres verticales : 10 mV.
132

Son axone se projette directernent dans le STG par le nerf stomatogastrique (Fig.3A) (Cazalets, 1987; Cazalets et al., 1987).
3.4.1. Mise au silence . du réseau pylorigue par ie neurone PS.
Lorsque le réseau pylorique produit un rythme vigoureux et stable, une décharge brève (2 à 5 sec) du neurone PS induit une série de modifications profondes et durables (de l'ordre de la minute) de l'activité de l'ensemble des neurones pyloriques. La plus frappante de ces rnodifications est un arrêt plus ou moins long de toute activité rythmique (Fig.3B). L'arrét du rythme pylorique se traduit par une hyperpolarisation et un arrêt durable des osciilations des neurones pacemakers dilatateurs (PD-AB) et du neurone constricteur LP, ainsi que par la dépolarisation et la décharge tonique des neurones constricteurs PY. Là encore, une série d'arguments expérimentaux a rnontré que le neurone PS agissait monosynaptiquement sur les différents neurones pyloriques.
Les effets du neurones PS aboutissant à l'arrêt du rythme pylorique, semblant être différents pour chaque catégorie de neurone du réseau, il s'est avéré nécessaire pour comprendre l'impact exact de PS sur chacun de ces neurones d'analyser ces effets dans des conditions où le neurones considéré était soustrait à l'influences des autres éléments du réseau. Pour ce faire le neurone considéré est isolé in situ par élimination des neurones qui lui sont présynaptiques dans le réseau, cette élimination étant réalisée par la technique de photoinactivation. Mise au point sur le système nerveux stomatogastrique (Miller et Selverston, 1979), cette technique performante s'effectue en deux étapes: - la première consiste à injecter dans le corps cellulaire d'un neurone, par l'intermédiaire d'une micropipette de verre, un colorant fluorescent: le Jaune Lucifer ou la 6-carboxy fluoresceine. Ces colorants sont chargés négativement. Placés dans une microélectrode, ils peuvent donc d'une part permettre l'enregistrement et donc l'identification du neurone empalé, d'autre part être injectés sous l'effet d'un courant. Le colorant se répartit par diffusion jusque dans les parties du neurone impliquées dans les interactions synaptiques. Après injection le colorant reste vital tant que la préparation est éclairée en lumière blanche. - dans un deuxième temps, la préparation est éclairée en lumière bleue intense (450-490 nm, lampe à vapeur de mercure montée sur la loupe binoculaire). Le colorant rnanifeste alors une fluorescence jaune qui s'accompagne de réactions d'oxydation très rapidement léthales pour le neurone injecté. Un neurone est considéré comme inactivé lorsque: 1) son potentiel de membrane est nul; 2) il ne produit plus de potentiels d'action enregistrables sur les nerfs moteurs; 3) ses effets postsynaptiques ont disparu.
La possibilité d'isoler in situ un neurone pylorique par photoinactivation des neurones qui lui sont présynaptiques dans
133

le réseau a perrnis de montrer que l'arrêt prolongé du rythme pylorique provoqué par la décharge du neurone PS est dû essentiellement à J.a suppression des propriétés régénératives de trois neurones dans le réseau, les deux neurones dilatateurs PD et le neurone constricteur LP. Ainsi, une décharge brève de PS (20 Hz pendant 2 sec) supprirne la capacité d'osciiler des neurones PD pour une durée de 10 à 20 sec, et supprime les propriétés de plateau du neurone LP pendant près d'une minute. Dans ce laps de temps ces neurones deviennent passifs et, spontanément silencieux, ils déchargent toniquement s'il sont dépolarisés (Cazalets, 1987). Remarquons que cette suppression pour les neurones PD et LP de la faculté d'exprimer des conductances voltage-dépendantes est durable. Par conséquent, l'action du neurone PS, tout comme celle du neurone APM, s'apparente à un processus de neuromodulation.
Cependant, si la seule décharge du neurone PS peut arréter durablement l'activité du générateur pylorique, son action, tout corome celle du neurone APM, n'est pas uniquement du type tout ou rien. Lorsqu'il décharge à basse fréquence, PS peut égalernent modifier l'expression du pattern pylorique.
3.4.2. Modification du...pattern pylorique par le. neurone E.S.
Comme nous l'avons déjà mentionné, tous les neurones pyloriques ne manifestent pas la même sensibilité vis à vis de la décharge du neurone PS (l'arrêt d'activité du neurone LP est par exemple beaucoup plus prolongé que celui des neurones PD). Cette différence de sensibilité apparait particulièrement clairement lorsque la fréquence de décharge de PS n'est pas suffisante pour arrêter le rythme pylorique, et aboutit a des modifications plus ou moins profondes de la structure du pattem raoteur pylorique. Ces modifications concement particulièrement le nombre de neurones participant au rythme pylorique, et la phase à laquelle les neurones constricteurs déchargent dans le cycle des neurones pacemakers dilatateurs.
Ainsi lorsque le neurone PS décharge toniquement à des fréquences croissantes inférieures à 10 Hz, les neurones constricteurs déchargent de plus en plus tôt dans le cycle des dilatateurs. D'autre part, la durée des salves du neurone LP devient de plus en plus courte alors que celle des neurones PY s'allonge. Si la fréquence de décharge de PS s'accroît encore, le neurone LP devient silencieux, c'est à dire que le pattern pyiorique normalement triphasique devient brusquement biphasique. Enfin lorsque la fréquence de PS dépasse 10 Hz, i'activité des neurones PD et PY cesse d'être rythmique. Cependant même dans ce cas, deux autres neurones normalement associés au réseau pyloriques, les neurones VD et IC qui commandent les rnuscles respectivement dilatateurs et constricteurs controlant l'entrée du filtre pylorique, continuent à osciller et à décharger rythmiquement. Par conséquent, ces deux neurones qui sont normalement inféodés aux neurones pacemakers pyloriques, s'avèrent capables d'osciller indépendamrnent lorsque le neurone P3 décharge (Fig.3C) (Cazalets, 1987).
134

En résumé, il apparait que le neurone rnodulateur PS peut à
lui seul non seulement arrêter durablement le rythme pylorique,
mais également en fonction de sa fréquence de décharge peut
provoquer des reconfigurations fonctiormelles du réseau
pyloriques qui se traduisent par l'élaboration de patterns
rnoteurs différents.
4. Conclusion.
Le système nerveux stomatogastrique des crustacés
décapodes, en particulier à travers son réseau pylorique,
constitue un modèle performant qui permet d'ébaucher, de tester
et d'affiner des concepts fondarnentaux de ia neurobiologie
modeme. Il a en particulier perrnis d'établir qu'un réseau neuronal
apparamrnent cablé rigidement est capable d'élaborer une activité
motrice extrêmement flexible et adaptable. Il a également permis
de montrer que les nombreux "phénotypes" de cette activité sont
sélectionnés par des neurones localisés dans des centres
hiérarchiquement supérieurs, et qui exercent un contrôle de type
neuromodulateur sur les propriétés bioélectriques des neurones
du réseau générateur de pattern moteur. Il a enfin permis
d'établir que ce contrôle neuromodulateur d'origine central repose
sur deux grand types d'influences, d'une part des influences
permissives qui induisent ou amplifient les propriétés
régénératives des neurones cibles, et d'autre part des influences
suppressives qui diminuent ou font disparaître ces propriétés.
CAZALETS J.H., 1987. Contrôle de l'activité du réseau neuronique
pylorique du homard par des afférences modulatrices
suppressives: étude pharmacologique et électrophysiologique.
Thèse de doct. Univ. Bordeaux I, 111 p. CAZALETS J.R., NAGY E., MOOLINS M., 1987. Suppressive control of
a rhythmic central pattem generator by an identified
modulatory neuron in crustacea. Neurosci. Let. 81, 267-272.
DICKINSON P.S., NAGY F. 1983. Control of a central pattern
generator by an identified modulatory intemeurone in
Crustacea. II. Induction and modification of plateau
properties in pyloric neurones. J. exp. Biol. 105, 59-82. FLAMM R.E., HARRIS-WARRICK R.M., 1986. Aminergic modulation in
lobster stomatogastric ganglion. 1. Effects on motor pattem
and activity of neurons within the pyloric circuit. J. Neurophysiol. 55, 847-865.
MARDER E., 1987. Neurotransmitters, neuromodulators. In
Selverston A.I., Moulins M. (eds) The crustacean stomatogastric system. A model for the study of central
nervous systems. Springer, Heidelberg pp 263-300.
MILLER J.P., 1987. Pyloric mechanisms. In: Selverston A.I., Moulins
M. (eds) The crustacean stomatogastric system. A model for
the study of central nervous systems. Springer, Heidelberg
pp 109-136. MILLER J.P., SELVERSTON A.I., 1979. Rapid killing of single neurons
by irradiation of intracellularly injected dye. Science 206,
702-704.
135

MOULINS M., COURNIL I., 1982. All-or-none control of the bursting properties of the pacemaker neurons of the lobster pyloric pattem generator. J. Neurobiol. 13, 447-458.
NAGY F., DICKINSON P.S., 1983. Control of a central pattem generator by an identified modulatory intemeurone in crustacea. I. Modulation of the pyloric motor output. J. exp. Biol. 105, 33-58.
RUSSELL D.F., HARTLINE D.K., 1982. Slow active potentials and bursting motor pattems in pyloric network of the lobster, Panulirus interruptus. J. Neurophysiol. 48, 914-937.
SELVERSTON A.I., MOULINS M., 1985. Oscillatory neural networks. Ann. Rev. Physiol. 47, 29-48.
SELVERSTON A.I., MOULINS M., 1987. The crustacean stomatogastric system. A model for the study of central nervous systems. Springer, Heidelberg 338 p.
136

ASPECTS RÉCENTS de la Biologie des CRUSTACÉS Actes de Colloques - 8 - 1988 - IFREMER, pages 137 à 148
APPROCHE ECOPHYSIOLOGIQUE ET ULTRASTRUCTURALE DE L'OSMOREGULATION CHEZ LES CRUSTACES DECAPODES
PEQUEUX A.(1)(2) LIGNON J.(2)
RESUME - L'euryhalinité et la distribution écologique des Crustacés déca-podes sont étroitement liées à la plus ou moins grande aptitude de certains tissus spécialisés à transporter les ions inorganiques Na+ et Cl-. C'est l'analyse de la nature de ces mouvements ioniques, ainsi que des mécanismes qui les régissent, considérés dans un contexte écophysiologique, qui fera l'objet principal de cet article. Une attention toute particulière sera réservée à la branchie du crabe chinois Eriocheir sinensis, véritable modèle pour étudier les mouvements transépithéliaux d'ions inorganiques en relation avec le contrôle de l'osmolalité sanguine chez les Crustacés. A l'appui de l'approche écophysiologique du transport transépithélial bran-chial de NaCl, ce travail comprendra un important volet morphologique et ultrastructural ainsi qu'une analyse des propriétés de perméabilité de la cuticule qui borde l'épithélium et quelques considérations sur leurs impli-cations possibles dans les propriétés du tissu branchial.
mots-clés: osmorégulation, Crustacés décapodes, salinité, écophysiologie, Eriocheir sinensis, Carcinus maenas, Maia squinado, Cancer pagurus, cuticule, branchie, Na+, Cl-
ABSTRACT - Euryhalinity and ecological distribution of decapod Crustacea are primarily dependent on the ability of some specialized boundary epithelia to transport Na+ and Cl-. This paper deals mostly with recent physiological data on ion transport across the gill tissue and an attempt is made to correlate them with blood osmoregulation and ecological distribution in euryhaline Decapods. A particular attention is paid to the gills of the fully euryhaline Chinese crab Eriocheir sinensis. Supporting this ecophysiological approach, there is an important ultrastructural analysis of the gill epithelium under various salinity conditions. The possible part played by the cuticle in ions raovements is also considered.
key-words: osmoregulation, decapod Crustacea, salinity, ecophysiology, Eriocheir sinensis, Carcinus maenas, Maia squinado, Cancer pagurus, cuticle, gill, Na+, Cl-
(1) Université de Liège, Laboratoire de Physiologie Animale, 22 quai Van Beneden, B-4020 Liège, Belgique.
(2) Laboratoire d'Etude des Régulations Physiologiques, CNRS, 23 rue Becquerel, F-67087 Strasbourg, France.
137

1. Introduction
La plus ou moins grande capacité de tolérance à un stress osmotique constitue l'un des facteurs conditionnant largement l'établissement d'une population animale dans un milieu à salinité variable comme la zone intertidale. II s'agit très certainement du facteur essentiel de leur réussite dans les écotones tels que les estuaires, les lagons côtiers et les mangroves.
En première analyse, la salinité peut affecter un organisme aquatique de manières directe et indirecte. Les effets directs sont à mettre en relation avec la capacité des espèces considérées d'assurer les balances hydrique et ionique de leurs fluides internes, intra et extracellulaires.
Les effets indirects résultent d'interactions plus complexes avec un ou plusieurs autres facteurs de stress tels que la température, la disponibilité en oxygène ou l'action de substances toxiques. Dans le con-texte de la survie d'une population dans un milieu donné, ils revêtent donc une importance considérable. il est malheureusement très malaisé de les appréhender du fait, notamment, du grand nombre de paramètres qui peuvent interréagir. II s'agit probablement de la raison pour laquelle les études écophysiologiques menées en laboratoire n'ont jusqu'à ce jour été orientées pratiquement que vers la seule compréhension des divers mécanismes utilisés par les animaux pour assurer leur balance en eau et en ions.
Les mouvements d'eau étant déterminés par des différences d'osmolarité et l'osmolarité des fluides biologiques étant, chez les Invertébrés, essen-tiellement déterminée par les concentrations en ions inorganiques, ces études ont donc surtout porté sur la nature et les modalités de mouvements ioniques.
2. Osmorégulation des fluides intra et extracellulaires chez les Crustacés décapodes
La relation qui existe entre les osmolalités interne et externe s'ex-prime habituellement en termes d'osmoconformité et d'osmorégulation.
La plupart des espèces de Crustacés décapodes marins sont osmocon-formes et la concentration de leur hémolymphe en ions inorganiques est, dans la grande majorité des cas, essentiellement identique à celle de l'eau de mer (EM) environnante. S'il arrive que ces concentrations diffèrent, l'osmolarité sanguine totale reste néanmoins la même que celle de l'envi-ronnement marin, et l'expérimentation dans ce domaine a conclu à l'absence virtuelle de tout gradient électrochimique de NaCl et d'eau au niveau de la surface du corps, impliquant l'absence de tout flux diffusionnel net. Les animaux relevant de tels groupes ne rencontrent donc pratiquement aucun problème d'osmorégulation et peu de problèmes d'iono-régulation tant qu'ils restent confinés au milieu marin puisque l'exigence fondamentale à un fonctionnement cellulaire optimal, à savoir le maintien d'un environnement cellulaire stable et constant, est assurée. Cette exigence est en outre assurée sans dépense énergétique puisque, pour chaque substance, l'énergie potentielle est identique dans l'hémolymphe et dans le milieu ambiant. Il n'en est plus de même quand la salinité de ce milieu varie. II en résulte en effet une différence d'énergie potentielle entre les deux milieux, ce qui génère un flux diffusionnel d'ions inorganiques et d'eau responsable à plus ou moins court terme d'un changement de composition de l'hémolymphe. Dans ce cas, le maintien d'un environnement cellulaire stable et constant ne peut plus être assuré que si l'organisme produit en sens inverse un flux
138

exactement égal aux pertes diffusionnelles. Une telle réaction consomme nécessairement de l’énergie métabolique.
Chez les espèces osmoconformes, l'expérience a démontré qu'il se rétablit rapidement un équilibre de concentrations entre l'hémolymphe et le milieu extérieur lorsque la salinité de ce dernier varie. Les espèces appartenant à ce groupe semblent donc bien dépourvues de tout mécanisme de contrôle de la composition de leur milieu extracellulaire et leur aptitude à faire face à une modification de composition du milieu environnant ne dépend que de l'aptitude de leurs cellules à supporter les variations de composition des fluides extracellulaires.
Chez les espèces régulatrices, un travail est effectué par l'animal qui maintient sa concentration osmotique interne à un niveau relativement constant. Elles ne représentent, chez les Crustacés décapodes, que quelques groupes plus restreints d'espèces. II s'agit toutefois des espèces qui ont réussi, avec plus ou moins de succès, la conquête de la plus grande variété des habitats et qui ont développé les mécanismes adaptatifs les plus intéressants.
Quelle que soit l'efficacité des mécanismes de régulation mis en oeuvre, aucune de ces espèces n'est cependant capable de maintenir l'osmo-larité de son sang â un niveau rigoureusement constant, ce qui implique que, lors de leur acclimatation à un milieu de salinité différente, leurs cellules ont à surmonter un choc osmotique dont l'amplitude dépend de l'efficacité des mécanismes de contrôle au niveau sanguin.
De ce qui précède, il ressort que l'on peut difficilement dissocier l'étude des mécanismes de contrôle de la composition de 1'environnement cellulaire, de l'examen des processus directement impliqués dans le main-tien du volume cellulaire.
L'expérience a montré, tant in vivo que in vitro, que la durée de la phase de régulation du volume cellulaire consécutive à un choc osmotique peut être très différente d’une espèce à l'autre; il est en effet possible de dégager un certain degré de corrélation entre l'efficacité des méca-nismes de régulation du volume cellulaire et le profil écophysiologique de l'espèce considérée. En effet, la régulation du volume cellulaire, consécu-tive à l'application d'un stress hypoosmotique, est environ trois fois plus rapide chez le crabe chinois Eriocheir sinensis, une espèce euryhaline aux performances osmorégulatrices très élevées, que chez le crabe vert Carcinus maenas qui ne peut tolérer que des dilutions plus limitées (Gilles et Jeuniaux, 1979; Harris, 1976). Les espèces osmoconformes, tel le tourteau Cancer pagurus, semblent, elles, incapables de réguler leur volume cellu-laire dans des périodes de temps comparables (Wanson et al., 1983). L'effi-cacité de ces processus qui relèvent du contrôle de l'osmolarité du liquide intracellulaire prend donc ici toute sa signification. II s'agit sans aucun doute des mécanismes essentiels sinon déterminants de la distribution des espèces dans les milieux à salinité variable. Plusieurs revues leur ont été consacrées (Gilles, 1979, 1980a,b); le lecteur intéressé est invité à s'y référer.
En bref, la différence entre espèces conformes et espèces régulatrices se situe, de ce point de vue, à deux niveaux: (1) un niveau cellulaire mettant en oeuvre des mécanismes fondamentalement semblables qualitative-ment chez les espèces conformes et régulatrices, mais quantitativement plus puissants chez les régulateurs, (2) un niveau périphérique limitant les chocs encourus par les cellules et exclusivement rencontré chez les espèces
139

régulatrices.
Le diagramme de la Fig. 1 résume, de manière très schématique, l'en-semble des mécanismes engagés dans le contrôle et la régulation des ba-lances hydrique et ionique d'un Crustacé décapode régulateur "typique".
Fig. 1. Représentation schéma-tigue des mécanismes et organes engagés dans le contrôle et la régulation des balances hgdrique et ionique d'un Crustacé décapode régulateur "typique". Les flèches schématisent les flux nets (con-tenant des composantes passive et/ou active). Cx: concentration du soluté x
dans les fluides interne (i), externe (o) et uri-naire (u).
Jx: flux du solutë x aux ni-veaux des branchies (B) (flux net) et du système excréteur tubulaire (t).
Jeau: flux d'eau au niveau des branchies (B) (flux net) et du système excréteur tubu-laire (t).
Jf: vitesse d'ultrafiltration. Ju: vitesse de production de
l'urine (Ju = Jeau B).
3. Régulation anisosmotique des fluides extracellulaires
Si les mécanismes de régulation des fluides extracellulaires, éven-tuellement doublés d'adaptations comportementales (vie dans des terriers ou fait de se soustraire temporairement aux fluctuations de salinité) déter-minent dans une large mesure les possibilités de colonisation des écotones à salinité variable, il est évident que l'acquisition de mécanismes de
contrôle et de régulation de l'osmolalité sanguine contribue à assurer le succès de nombreuses espèces dans leur conquête des milieux dilués.
Chez les Crustacés décapodes, le rôle déterminant joué par les bran-
chies dans ces mécanismes de régulation a été reconnu depuis longtemps.
Dans les années '50, Koch suggérait déjà que les diverses branchies pou-
vaient être morphologiquement et fonctionnellement différentes et que le flux entrant de Na+ était, chez le crabe chinois, nettement plus important
dans les branchies situées postérieurement que dans les branchies anté-
rieures (Koch et al., 1954) Le développement plus récent de préparations
perfusées de branchies isolées a permis de conclure à l'existence, chez le
crabe chinois E. sinensis mais également chez la plupart des espèces hyper-
régulatrices en milieu dilué, à une véritable spécialisation physiologique
de certaines branchies ou tout au moins de certaines régions des lamelles
branchiales (pour revue: Gilles et Péqueux, 1981, 1983, 1985, 1986). A cet
égard, il est apparu que la branchie du crabe chinois pouvait etre considé-
rée comme un véritable modèle pour étudier les mouvements transépithéliaux
d'ions inorganiques en relation avec le contrôle de l'osmolalité sanguine
chez les Crustacés.
140

3.1. Etude structurale et fonctionnelle de la branchie d'E. sinensis
3.1.1. En milieu dilué (ED)
Chez le crabe chinois d'eau douce, l'épithélium des trois paires de branchies les plus postérieures apparaît comme beaucoup plus épais (10 pm et plus) que l'épithélium des branchies situées antérieurement (2-4 yra). II est bordé extérieurement d'une cuticule qui tend, par contre, à être plus mince (0,3 µm contre 1 µm). L'épithélium des branchies postérieures est caractérisé par un important système de plissements et d'invaginations des membranes plasmatiques situées aux poles apical et basolatéral des cel-lules. L'espace cytoplasmique est occupé d'un nombre considérable de mito-chondries (Péqueux et Barra, 1980; Péqueux et al., 1980; Gilles et Péqueux, 1981; Barra et al., 1983). Ce type de structure est tout à fait caractéris-tique des "épithélia transporteurs de sels".
L'étude fonctionnelle sur préparation perfusée a permis de confirmer, en parfait accord avec les conclusions de l'étude ultrastructurale, que seules les branchies postérieures sont le siège des mécanismes de transport actif transépithélial d'ions Na+ et Cl_ (Péqueux et Gilles, 1981; Gocha et al., 1987) (Fig. 2). Les branchies antérieures ne semblent être le siège
Fig. 2. Modèle fonctionnel des branchies antérieure et postérieure du crabe chinois E. sinensis d'eau douce. Les divers systèmes de transport schématisès ont été placès en accord avec les mesures de flux ioniques et de potentiels èlectriques effectués sur des branchies isolées et perfusêes. Les deux faces de 1'épithèlium sont baignées de solutions physiologiques identiques (d'après Péqueux et Gilles, 1987).
141

d'aucun transport transépithélial d'ions Cl_ (Gocha et al., 1987) et les mouvements de Na+ y restent de nature essentiellement passive (Péqueux et Gilles, 1981) (Fig. 2). Ces observations concordent parfaitement avec les résultats de l'étude ultrastructurale et semblent ne réserver à ces bran-chies qu'une signification essentiellement respiratoire. Toutefois, il a été établi que la "perméabilité apparente" de cet épithélium aux ions Na+
dépend de la concentration de cet ion dans le milieu environnant (Péqueux et Gilles, 1981). En effet, une diminution de la concentration externe en Na+ entraîne une diminution immédiate de "perméabilité apparente" au Na+. Chez une espèce régulatrice comme le crabe chinois qui est confronté à des milieux très dilués, un tel réajustement présente un intérêt physiologique considérable puisque la perte de sels se trouve être diminuée en dépit de L'augmentation du gradient de concentration ionique transépithélial. Le maintien des balances ionique et osmotique sanguines s'en trouve donc être facilité. Nous verrons plus loin qu'en plus de tels "réajustements immé-diats" de la "perméabilité épithéliale apparente" au Na+, il est également possible de déceler des réajustements "à plus long terme" dont la signifi-cation est identique. Ces mécanismes, de même d'ailleurs que 1'extrêmement bas niveau de la "perméabilité apparente" au Na+ des branchies posté-rieures, peuvent être considérés comme une catégorie de "processus limi-tants" contribuant à assurer, aux cotés des "processus compensatoires" comme les transports actifs, le contrôle de l'osmolalité sanguine.
3.1.2. En eau de mer (EM)
L'ultrastructure de l'épithélium des branchies antérieures reste, après acclimatation à l'eau de mer, fondamentalement semblable à ce qu'elle était en eau douce (Péqueux et al., 1987). L'amplitude des fiux unidirec-tionnels de Na+ a, par contre, considérablement augmenté (Péqueux et Gilles, 1981). Elle atteint en outre un niveau largement supérieur à celui mesuré dans les mêmes conditions expérimentales sur les branchies d'un crabe d'eau douce, suggérant que la perméabilité au Na+ a dû diminuer de façon significative lors de l'acclimatation prolongée à l'eau douce. Ce réajustement "à long terme" de la perméabilité au Na+ semble différent du réajustement immédiat rapporté précédemment. Bien que contribuant tous deux à limiter l'importance de la perte de sels en milieu dilué, il est probable que les deux systèmes aient des bases moléculaires différentes et répondent à des messagers différents.
Dans les branchies postèrieures, l'acclimatation à l'eau de mer s'ac-compagne d'une régression considérable, voire de la disparition complète, du système d'invaginations apicales (Péqueux et al., 1987). Parallèlement, les flux transépithéliaux de Na+ tombent à un niveau indétectable. L'expé-rience a prouvé que l'activité du/des mécanisme(s) de transport actif est sensible à la concentration en Na+ de l'hémolymphe et que l'élévation de concentration sanguine en Na+ qui se produit en EM suffit à elle seule à interrompre de manière durable son/leur activité (Péqueux et Gilles, 1981).
3.1.3. Lors du passage EM-ED
Du point de vue ultrastructural, l'épithélium des branchies posté-rieures évolue progressivement vers le schéma caractéristique d'un animal d'ED par apparition de "blebs" à la base des quelques rares replis apicaux, le creusement de citernes à ce niveau ainsi que la formation de replis de plus en plus profonds. Le processus nécessite environ cinq jours (Gilles et Péqueux, 1985; Péqueux et al., 1987) (Fig. 3).
Parallèlement, l'activité de transport actif ne se restaure que très
142

progressivement. Le transport actif de Na+, à en juger par la mesure des flux unidirectionnels et l'activité de la Na+K+ATPase, ne retrouve en effet son niveau maximum qu'après huit jours d'acclimatation, bien qu'un niveau significatif soit déjà atteint après trois jours (Péqueux et Gilles, résultats non publiés).
Fig. 3. Représentation schèmatique des change-ments ultrastructuraux qui se produisent dans 1 'épithélium des bran-chies postêrieures du crabe chinois E. sinen-sis lors du passage de l'eau de mer (EH) à l'eau douce (ED). C: cuticule, M: mitochon-drie, MA: membrane api-cale, MBL: membrane baso-latérale.
3.2. Applicabilité du_modèle Erioçheir aux aut.res_Crustacés_décapodes
De ce qui précède, il resosrt que la branchie d'Eriocheir sinensis apparaît comme un matériel particulièrement intéressant pour étudier les mouvements d'ions transépithéliaux en relation avec l'osmorégulation san-guine chez les Crustacés et progresser vers une meilleure compréhension des relations structure-fonction à ce niveau.
Dans quelle mesure ce modèle est-il toutefois applicable aux autres espèces de Crustacés ?
D'une étude comparée des branchies de trois crabes appartenant à des catégories écophysiologiques différentes, à savoir l'osmoconforme euryhalin
Cancer pagurus, l'hyperrégulateur médiocre Carcinus maenas et l'hyperrégu-lateur puissant Eriocheir sinensis, il ressort que 1'organisation structu-rale de ces branchies et leurs propriétés physiologiques peuvent être mises en relation directe avec les capacités d'osmorégulation des espèces consi-dérées.
Cette relation est, en effet, particulièrement nette chez le crabe chinois E. sinensis.
Chez le tourteau Cancer pagurus, une espèce osmoconforme incapable de contrôler son osmolalité sanguine, toutes les branchies ne sont limitées que par un épithélium peu différencié, de type respiratoire, et l'étude des propriétés de leur différence de potentiel électrique transépithéliale ne permet d'identifier aucune activité de transport (Péqueux et al., 1984).
Chez 1'hyperrégulateur modéré Carcinus maenas, les deux types d'épi-thélia existent: respiratoire et de type transporteur. L'épithélium trans-porteur est également limité aux branchies postérieures mais son étendue ne
143

correspond qu'à 30% maximum de la surface lamellaire totale (Compère et al., 1985). D'expériences réalisées sur branchies isolées et perfusées, il est apparu que les propriétés physiologiques de la branchie entière corres-pondent à la résultante des propriétés spécifiques des deux types d'épithé-lia, à savoir un épithélium respiratoire très perméable aux ions Na+ et un épithélium transporteur peu perméable au Na+ comme chez le crabe chinois E. sinensis. Contrairement à la situation qui prévaut chez le crabe chi-nois, les branchies postérieures de C. maenas sont en effet le siège de mouvements passifs de Na+ considérables (Wanson et Péqueux, ce volume). II est donc très vraisemblable que les performances écophysiologiques modérées du crabe vert soient en relation directe avec le développement restreint de tissu spécialisé dans la récupération active et la limitation des pertes de sels.
De ce qui précède, il apparaît donc clairement que le modèle d'organi-sation structurale et fonctionnelle de la branchie d'E. sinensis peut être parfaitement appliqué, moyennant quelques adaptations spécifiques, aux autres crabes euryhalins.
4. Signification physiologique de la cuticule
Jusqu'à ce jour, les physiologistes de la branchie ont presque systé-matiquement ignoré, dans leurs modèles fonctionnels, la cuticule chitineuse qui borde l'épithélium à sa face apicale. Des travaux récents tendent pourtant à démontrer que cette cuticule pourrait jouer un rôle non négli-geable dans les propriétés de perméabilité du tissu branchial (Avenet et Lignon, 1985; Lignon, 1987b) et donc revêtir une signification écophysiolo-gique importante (Lignon, 1987a).
-2
A STACl
H
JS NE
F
OMAR EPHRO
JS C.MA
PS MAIA
L T
ENAS
C.PAG URUS
2 |
? O
-3 _ i - -3 □
S) \ £ (C -4 - -4
5 1 - -5 O o
-6 C - -6 C
3
-7 . L Na* ▪NH;
O CI- HCOj-
■ -7
Fig. 4. Valeurs comparées de perméabilité cuticulaire pour quelques espèces ioniques chez divers Crustacès dècapodes; F: filaments branchiaux, L: lame branchiale (chez ies astacidés).
144
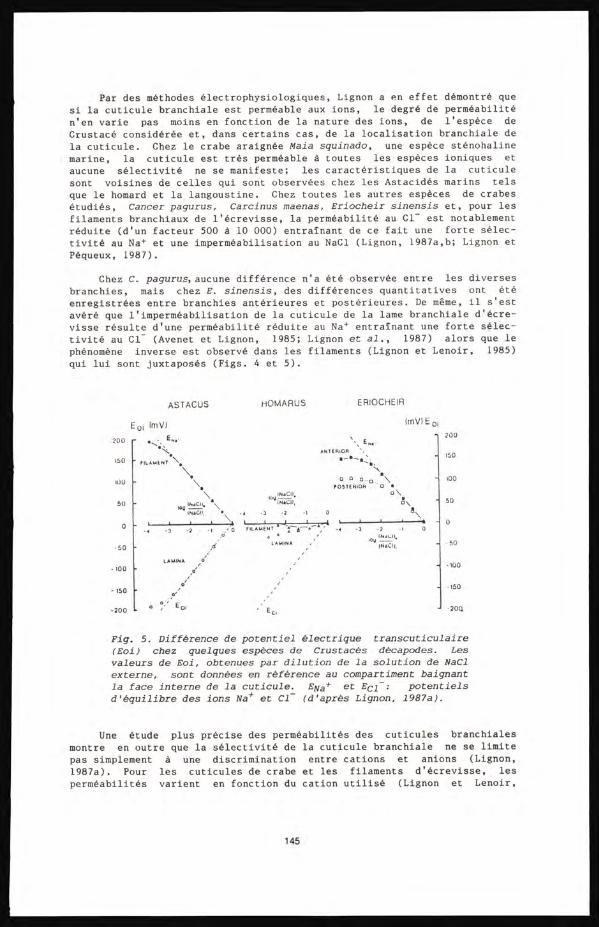
Par des méthodes électrophysiologiques, Lignon a en effet démontré que
si la cuticule branchiale est perméable aux ions, le degré de perméabilité
n'en varie pas moins en fonction de la nature des ions, de l'espèce de
Crustacé considérée et, dans certains cas, de la localisation branchiale de
la cuticule. Chez le crabe araignée Maia squinado, une espèce sténohaline
marine, la cuticule est très perméable à toutes les espèces ioniques et
aucune sélectivité ne se manifeste; les caractéristiques de la cuticule
sont voisines de celles qui sont observées chez les Astacidés marins tels
que le homard et la langoustine. Chez toutes les autres espèces de crabes
étudiés, Cancer pagurus, Carcinus maenas, Eriocheir sinensis et, pour les filaments branchiaux de l'écrevisse, la perméabilité au Cl_ est notablement
réduite (d'un facteur 500 à 10 000) entraînant de ce fait une forte sélec-
tivité au Na+ et une imperméabilisation au NaCl (Lignon, 1987a,b; Lignon et
Péqueux, 1987).
Chez C. pagurus, aucune différence n'a été observée entre les diverses
branchies, mais chez E. sinensis, des différences quantitatives ont été
enregistrées entre branchies antérieures et postérieures. De même, il s'est
avéré que 1'imperméabilisation de la cuticule de la lame branchiale d'écre-
visse résulte d'une perméabilité réduite au Na+ entraînant une forte sélec-
tivité au Cl_ (Avenet et Lignon, 1985; Lignon et al., 1987) alors que le
phénomène inverse est observé dans les filaments (Lignon et Lenoir, 1985)
qui lui sont juxtaposés (Figs. 4 et 5).
Fig. 5. Diffèrence de potentiel èlectrique transcuticulaire (Eoi) chez quelques espèces de Crustacés décapodes. Les valeurs de Eoi, obtenues par dilution de la solution de NaCl externe, sont données en référence au compartiment baignant la face interne de la cuticule. ENa
+ et EC1-: potentiels
d'èquilibre des ions Na+ et Cl~ (d'après Lignon, 1987a).
Une étude plus précise des perméabilités des cuticules branchiales
montre en outre que la sélectivité de la cuticule branchiale ne se limite
pas simplement à une discrimination entre cations et anions (Lignon,
1987a). Pour les cuticules de crabe et les filaments d'écrevisse, les
perméabilités varient en fonction du cation utilisé (Lignon et Lenoir,
145

1985; Lignon, 1987b). II en va de même pour la cuticule de la lame d'écre-visse vis-à-vis des anions (Avenet et Lignon, 1985; Aubert et Lignon, 1986).
Par ailleurs, il a également été montré que les cuticules branchiales sont fonctionnellement asymétriques: les potentiels transcuticulaires obte-nus en effectuant des changements ioniques, soit sur une face, soit sur l'autre, ne changent pas uniquement de signe mais changent aussi d'ampli-tude (Avenet et Lignon, 1985; Lignon et Lenoir, 1985; Lignon, 1987b). Cette asymétrie fonctionnelle a été plus largement étudiée sur la lame d'écre-
visse et montre très clairement des propriétés non ohmiques de la cuticule
qui présente une forte rectification au Cl_ (Lignon et al., 1987). L'en-
semble de ces résultats ne paralt pouvoir s'expliquer qu'en attribuant la sélectivité cuticulaire à une des couches les plus externes de la cuticule,
vraisemblablement l'épicuticule, alors que 1'endocuticule agirait comme une
simple couche de diffusion.
De ces travaux, il ressort donc clairement que les propriétés de perméabilité de la cuticule branchiale des Crustacés décapodes peuvent
varier nettement d'une espèce à l'autre ainsi qu'en fonction de la locali-sation dans la branchie. La cuticule des espèces osmoconformes ne présente
pas de sélectivité ionique marquée et est très nettement plus perméable que
celle des espèces régulatrices. Ceci laisse supposer que l'espace sous-
cuticulaire, particulièrement développé chez les espèces acclimatées à des
milieux dilués, est un compartiment physiologique important dont la signi-
fication et le rôle restent encore à préciser. Le rôle de la cuticule
pourrait être triple. En fonction de son degré de perméabilité, la cuticule
pourrait réduire les effets de variations brusques de la composition du
milieu ambiant sur ce compartiment sous-cuticulaire. Par ailleurs, la
cuticule pourrait réduire les gradients osmotiques importants auxquels
serait soumise en son absence la raembrane apicale de l'épithélium. Enfin,
la permsélectivité cuticulaire en fonction de sa localisation branchiale
pourrait correspondre à des voies de transport spécifique de l'espèce
ionique correspondante par l'épithélium sous-jacent.
5. Conclusion et perspectives
De l'analyse qui précède, il ressort que le profil des réponses d'un
organisme, voire d'une population, aux sollicitations de l'environnement
est largement, sinon principalement, déterminé par quelques particularités
adaptatives de sa/leur physiologie. Ces particularités adaptatives sont
pratiquement impossibles à dégager de travaux menés sur le terrain et ont
nécessairement échappé aux trop nombreuses investigations écologiques cher-
chant à comprendre la distribution d'une espèce animale en abordant l'envi-
ronnement dans son ensemble. Seule 1'expérimentation en laboratoire devrait
permettre de les révéler et les résultats obtenus à ce jour sont, à cet
égard, particulièrement encourageants.
L'expérimentation sur préparation perfusée de branchie isolée a par
exemple débouché sur une caractérisation plus claire des mécanismes et des
forces mis en oeuvre dans le contrôle et la régulation de la composition
des fluides intra et extracellulaires chez les Crustacés décapodes, que
n'avaient pu le faire les travaux "classiques" réalisés sur l'animal en-
tier. Au-delà de préoccupations purement écophysiologiques, le modèle d'or-
ganisation structurale et fonctionnelle de la branchie de Crustacés déca-
podes est en outre apparu comme un outil performant pour aborder l'étude
des relations structure-fonction en général, et dans les épithélia trans-
porteurs de sels en particulier.
146

Si certains des résultats rapportés dans cet article sont encore
fragmentaires, l'intérêt de cette approche justifie largement l'analyse qui
en a été faite. Elle a notamment permis d'identifier, parmi les diverses
branchies ou au sein d'un même organe parfois, une certaine forme de spé-
cialisation physiologique. Elle a aussi et surtout révélé le tissu bran-
chial comme une structure composite opposant plusieurs barrières succes-
sives au passage de diverses substances: une cuticule, un espace sous-
cuticulaire et un épithélium pour lequel il faut de plus considérer les
voies transcellulaire (membranes apicale et basolatérale) et paracellu-
laire. Leur étude approfondie mérite d'être poursuivie et étendue à la plus
grande variété d'espèces possibles de manière à en préciser l'importance
relative en fonction du profil écophysiologique de l'espèce considérée.
C'est de l'intégration des diverses informations qui en résulteront que
l'on pourra aboutir à une meilleure compréhension des stratégies dévelop-
pées pour surmonter les contraintes opposées par les milieux à salinité
variable.
AUBERT A., LIGNON J.M., 1986. Cuticular anionic selectivity in the gill
lamina of the crayfish. Europ. Soc. for Comp. Physiol. Biochem., 8th
Conf., Strasbourg, August 31-September 2, 1986.
AVENET P., LIGNON J.M., 1985. Ionic permeabilities of the gill lamina
cuticle of the crayfish, Astacus leptodactylus (E), J. Physiol. (Lond.), 363, 377-401
BARRA J.A., PEQUEUX A., HUMBERT W., 1983. A morphological study on gills of
a crab acclimated to fresh water. Tissue and Cell , 15, 583-596.
COMPERE Ph., PEQUEUX A., GOFFINET G., GILLES R., 1985. Ultrastructure of
the branchial epithelium of the green crab, Carcinus maenas, as a
function of the salinity of the environment. Biol. Cell, 54, 7a.
GILLES R., 1979. Mechanisms of Osmoregulation in Animals. Wiley-Inter-
science, London, New York, 667 p.
GILLES R., 1980a. Cell volume control as studied in crustacean tissues. In:
Gilles R. (ed.). Animals and Environmental Fitness, vol. 2. Pergamon
Press, Oxford, New York, pp. 3-24.
GILLES R., 1980b. Mechanisms of volume regulation and control of the level
of intracellular osmotic effectors. In: Engell H.C. (ed.). Metabolic
Response to Surgery and Injury. Am. J. Par. Nutr. GILLES R., JEUNIAUX Ch., 1979. Osmoregulation and ecology in media of
fluctuating salinity. In: Gilles R. (ed.). Mechanisms of Osmoregulation
in Animals. Wiley-Interscience, London, New York, pp. 581-608. GILLES R., PEQUEUX A.J.R., 1981. Cell volume regulation in crustaceans:
relationship between mechanisms for controlling the osmolality of
extracellular and intracellular fluids. J. Exp. Zool., 215, 351-362.
GILLES R., PEQUEUX A., 1983. Interactions of chemical and osmotic
regulation with the environment. In: Bliss D. (ed). The Biology of
Crustacea, vol. 8. Academic Press, London, pp. 109-177.
GILLES R., PEQUEUX A., 1985. Ion transport in crustacean gills:
Physiological and ultrastructural approaches. In: Gilles R. et Gilles-
Baillien (eds). Transport Processes, Iono- and Osmoregulation.
Springer-Verlag, Berlin, Heidelberg, pp. 136-158.
GILLES R., PEQUEUX A., 1986. Physiological and ultrastructural studies of
NaCl transport in crustacean gills. Boll. Zool. 53: 173-182.
G0CHA N., PEQUEUX A., WANS0N S., GILLES R., 1987. Cl" fluxes across
isolated, perfused gills of the Chinese crab Eriocheir sinensis (M. Edw.) acclimated to fresh water. Comp. Biochem. Physiol. (sous presse).
147

HARRIS R.R., 1976. Extracellular space changes in Carcinus maenas during
adaptation to low environmental salinity. J. Physiol. (Lond.) 258: 31-
32.
KOCH H.J., EVANS J., SCHICKS E., 1954. The active absorption of ions by the
isolated gills of the crab Eriocheir sinensis (M. Edw.). Meded. K. Vlaam Acad. Wet K1 . Wet., 16, 1-16.
LIGNON J.M., 1987a. Structure and permeability of decapod crustacea
cuticle. In: Kirsch R., Lahlou B. (eds). Comparative Physiology of
Environmental Adaptations. Part I: Adaptations to Salinity and
Dehydration. Karger, Bâle, pp. 178-187.
LIGNON J.M., 1987b. Ionic permeabilities of the isolated gill cuticle of
the shore crab Carcinus maenas. J. Exp. Biol. (sous presse).
LIGNON J.M., AUBERT A., AVENET P., 1987. Rectification retardée aux ions
Cl_ dans la lame branchiale de l'Ecrevisse, Astacus leptodactylus (E).
C.R. Acad. Sci. B, 304: 473-476.
LIGNON J., LENOIR P., 1985. Perméabilités ioniques de la cuticule des
filaments branchiaux de l'Ecrevisse Astacus leptodactylus (E). C.R. Acad. Sci. B, 301, 443-446.
LIGNON J.M., PEQUEUX A., 1987. Perméabilités ioniques comparées de la
cuticule branchiale de quelques Crustacés décapodes. Ass. Physiol.
Paris, 12-13 mai 1987.
PEQUEUX A., BARRA J.A., 1980. Fine structure of the gill epithelium of the
euryhaline crab Eriocheir sinensis as a function of the salinity of the acclimation medium. Eur. J. Cell. Biol. 22, 470.
PEQUEUX A., BARRA J.A., GILLES R., 1987. Sub-cuticular spaces and osmotic
adaptations. In: Kirsch R. et Lahlou B. (eds). Comparative Physiology
of Environmental Adaptations. Part I: Adaptations to Salinity and
Dehydration. Karger, Bale, pp. 188-197.
PEQUEUX A., BARRA J.A., GOESSENS G., WANSON S., GILLES R., 1984. Fine
structure of gill epithelium in three crabs, as related to their
ionoregulation performances. Abstract volume, First Congress of
Comparative Physiology and Biochemistry (Section of IUBS), Liège
(Belgium), Aug. 27-31, 1984, p. B-135.
PEQUEUX A.J.R., BARRA J.A., N0NN0TTE L., KIRSCH R., GILLES R., 1980. Fine
structure of salt-absorbing epithelium in gills of the crab Eriocheir
sinensis acclimated to freshwater. In: Gilles R. (ed.). Animals and
Environmental Fitness. Comp. Biochem. Physiol., suppl. 2, Abstract vol.
Pergamon, Oxford, pp. 13-14.
PEQUEUX A.J.R., GILLES R., 1981. Na+ fluxes across isolated perfused gills
of the chinese crab Eriocheir sinensis. J. Exp. Biol., 92, 173-186.
PEQUEUX A., GILLES R., 1987. The transepithelial potential difference of
isolated perfused gills of the chinese crab Eriocheir sinensis
acclimated to freshwater. Comp. Biochem. Physiol. (sous presse).
WANS0N S., PEQUEUX A., GILLES R., 1983. Osmoregulation in the stone crab
Cancer pagurus. Mar. Biol. Lett. 4: 321-330.
148

ASPECTS RÉCENTS de la Biologie des CRUSTACÉS Actes de Colloques - 8 - 1988 - IFREMER, pages 149 à 152
FLUX UNIDIRECTIONNELS D'IONS Na+ ET Cl* DANS LES BRANCHIES ISOLEES ET PERFUSEES DU CRABE VERT CARCINUS MAENAS
WANSON S.(1) PEQUEUX A.(1)(2)
RESUME - Les mouvements transépithéliaux branchiaux d'ions Na+ et C1 ont
été mesurés et caractérisés chez le crabe euryhalin C. maenas au moyen de
préparations perfusées de branchies prélevées sur des crabes d eau de mer
et sur des crabes acclimatés à de l'eau de mer diluée. II est démontré que
seules les trois paires de branchies postérieures sont capables de générer
un flux net entrant d'ions Na+ et C1 capable de contrebalancer les pertes
qui se produisent en milieu dilué.
mots-clés: Carcinus maenas, branchie, flux, Na+, C1
ABSTRACT - Transepithelial movements of Na+ and C1 were measured in
isolated perfused gills of the shore-crab C. maenas at two different sal-
inities. Posterior gills are shown to be the only ones able to generate net
Na+ and Cl_ influxes counterbalancing the salt loss occurring when in
dilute media.
key-words: Carcinus maenas, gill, flux, Na+, C1
L'état hyper-osmotique observé chez les espèces dulcicoles de crusta-
cés décapodes ou chez les espèces euryhalines acclimatées à un milieu dilué
est le résultat de phénomènes de charge et de décharge d'ions contrôlés par
des mécanismes de transport actif et des mouvements passifs. Certaines
espèces de crabes ont acquis la faculté de s'acclimater à des milieux dont
la gamme des salinités peut s'étendre de l'eau douce à l'eau de mer (Erio-
cheir sinensis), tandis que d'autres comme le crabe vert Carcinus maenas ne
peuvent survivre dans de bonnes conditions qu'à des dilutions plus limitées
(30% d'eau de mer). Si les mécanismes responsables du contrôle et de la
régulation de la balance ionique sanguine chez le crabe chinois sont ac-
tuellement relativement bien connus, il n'en est pas de même du crabe vert
C. maenas. II n'a notamment pas encore été possible de préciser les raisons
de ses facultés limitées de régulation anisosmotique sanguine, ni d'aboutir
à une caractérisation claire des mécanismes et forces responsables du
transport d'ions au niveau branchial.
(1) Université de Liège, Laboratoire de Physiologie Animale, 22 quai Van
Beneden, B-4020 Liège, Belgique.
(2) jusqu'à mars 1988: Laboratoire d'Etude des Régulations Physiologiques,
CNRS, 23 rue Becquerel, F-67087 Strasbourg, France.
149

Le présent travail s'est donc fixé pour objectif de mesurer et de
caractériser les mouvements transépithéliaux d'ions Na+ et C1 au moyen de
préparations perfusées de branchies prélevées sur des crabes d'eau de mer
(EM) et sur des crabes acclimatés à de l'eau de mer diluée (EM 30%).
C. m a en a s
f I u x Na* (jjEq/gP. F. /h.)
Fig. 1: Flux unidirectionnels de radiosodium (22Na+) dans des prêparations perfusées de bran-chies antérieures et posté-rieures de crabes verts C. mae-nas d'EM et d'EM 30%. Moyenne de (n) expériences ± DS.
Les détails techniques relatifs à la dissection et à la perfusion des
branchies ainsi qu'à l'estimation des flux unidirectionnels d'ions Na+ et
Cl" sont essentiellement comparables à ceux décrits dans les travaux effec-
tués sur le crabe chinois (Péqueux et Gilles, 1981; Gocha et al., 1987).
Lorsque les branchies de crabes d'EM sont perfusées par et incubées
dans de l'EM artificielle, les flux unidirectionnels entrant et sortant de
Na+ atteignent des valeurs de l'ordre de 2.500-3.000 pmoles Na+-g .h-1 , ne
générant aucun flux net significatif de Na+ (Fig. 1).
A l'issue d'une période d'acclimatement de plusieurs semaines à de
l'EM 30%, les flux entrant et sortant de Na+ tombent respectivement à 1.834
t 429 (5) et 770 ± 62 (3) µmoles•g_1-h-1 dans les trois paires de bran-
chies les plus postérieures incubées dans et perfusées par le même milieu
physiologique correspondant au plasma sanguin des crabes en EM 30%.
En milieu dilué, les branchies postérieures deviennent donc le siège
d'un flux net entrant d'ions Na+. Par contre, dans les paires de branchies
situées plus antérieurement, les flux entrant et sortant de Na+, bien que
réduits d'environ 25% par rapport à leur niveau mesuré en EM, restent
élevés mais semblables; il n'en résulte donc aucun flux net significatif de
Na+.
150

Les résultats expérimentaux repris à la Fig. 2 démontrent, qu'en
milieu dilué, les deux types de branchies présentent également des facultés
différentes de transporter les ions C1 . Si le flux sortant de C1 reste
dans tous les cas indépendant de la concentration en C1 du milieu d'incu-
C. maenas EM/3
Fig. 2: Effet de la con-centration externe en ions Cl- sur les flux unidirectionnels de Cl-
dans des préparations perfusées de branchies antérieures et posté-rieures de crabes verts C. maenas d'EM 30%. Mo-yenne de (n) expériences ± DS.
bation, l'amplitude du flux entrant augmente par contre avec le niveau de
C1‘ externe. Cette amplitude reste toutefois nettement supérieure dans les
branchies postérieures qui sont dès lors les seules à générer un flux net
entrant très important de cet anion.
II ressort de ce travail que les branchies antérieures sont beaucoup
plus perméables aux ions Na+ qu'aux anions C1 et que la perméabilité au
Na+ diminue légèrement à l'issue de l'acclimatement à un milieu dilué.
Elles semblent toutefois incapables de générer un flux net entrant de l'une
ou l'autre espèce ionique. Chez C. maenas, les branchies postérieures
semblent donc aussi être les seules directement impliquées dans le contrôle
de la balance sanguine en ions Na+ et C1 lors de l'acclimatement à un
milieu dilué. Seules les branchies postérieures sont en effet capables de
générer un flux net entrant de Na+ et de Cl- capable de contrebalancer les
pertes qui se produisent dans le sens du gradient de concentration ionique.
GOCHA N., PEQUEUX A., WANSON S., GILLES R., 1987. Cl- fluxes across iso-
lated, perfused gills of the Chinese crab Eriocheir sinensis (M. Edw.)
acclimated to freshwater, Comp. Biochem. Physiol. (sous presse).
PEQUEUX A., GILLES R., 1981. Na+ fluxes across isolated perfused gills of
the Chinese crab Eriocheir sinensis, J. Exp. Biol., 92, 173-186.
151

ASPECTS RÉCENTS de la Biologie des CRUSTACÉS Actes de Colloques - 8 - 1988 - IFREMER, pages 153 à 158
VARIATION DE LA CONSOMMATION D'OXYGENE AU COURS DU NYCTHEMERE CHEZ ARMADILLIDIUM VULGARE (CRUSTACE ISOPODE TERRESTRE).
CHENTOUFI A. (1) , MOCQUARD J.P. (1)
RESUME - La plupart des isopodes terrestres présentent un rythme nychthemeral d'activité locomotrice avec un maximum au cours de la nuit. L'activité respira-toire d'Armadillidium vulgare suit également un rythme nychthéméral avec deux maximum d'activité, au début et en fin de nuit.
Les variations de ce rythme d'activité respiratoire ont été étudiées sous différentes conditions : régime photopériodique naturel, conditionsconstantes de lumière, séjour photopériodique artificiel, décérébration. Les résultats obtenus laissent supposer l'existence de deux oscillateurs circadiens plus ou moins autonomes.
Mots clés : respiration, rythme nycthéméral, photopériode, Armadillidium vulgare
ABSTRACT - Most of terrestrial isopodes exhibit a nycthemeral rhythmof locomo-tor activity with a peak during the nigHtC The respiration activity of Armadillidium vulgare also follows a nychthemeral pattern with two maxima of activity at the begining and at the end of the night.
The variations of the respiratory activity have been studied under dif-ferent conditions : natural photoperiodicity, constant illumination, articifial photoperiodicity, decerebration.
The results are in favour of the existence of two more or less autonomous circadian oscillators.
Key words : Respiration, nychthemeral rhythm photoperiodism, Armadillidium.
INTRODUCTION -
La majorité des isopodes terrestres présente un rythme nycthéméral d'activi-té locomotrice avec un maximum au cours de la nuit (Wieser et al., 1969). Edney (1960) suggère que l'intensité respiratoire est liée à l'activité locomotrice. Dans le but d'étudier comment l'activité d'A. vulgare pouvait être conditionnée Par l alternance jour nuit nous avons examiné les variations de l'intensité res— piratoire sous différentes conditions.
Matériel et Méthodes -
Nous avons choisi des mâles ayant une masse voisine de 100 mg et au stade C de leur cycle d'intermue. Les femelles ont été écartées, leur intensité respira-toire variant notablement en fonction de leur état sexuel.
La température a été fixée à 20°C, elle.a été associée à des régimes photo-périodiques différents. Le métabolisme respiratoire a été étudié au moyen d'un respiromètre différentiel Gilson. Les mesures ont été effectuées sur des individus îsolés en relevant la consommation d'oxygène horaire.
(1) Laboratoire de Biologie Animale de l'Université de Poitiers et UA CNRS n° 673
"Physiologie et Génétique des Crustacés", 40, Avenue Pineau, 86022 Poitiers Cedex, France.
153

154
Figure 1 : Taux de consommation d'oxygèneCen microlitres par heu.-e)
4 20°C,
A : sous photopèriode naturelle (ΦN)
B, C, 0 : sous lumière continue CLL)
C1 jour sous LL)
C30 jours sous LL)
Fiqure 2 : Situation des pics représentant les maximums de la
consommation d'oxygène, sous photopériode naturelle tÿgxÿj/
et sous lumière continue I ~l
C : coucher du soleil, L : lever du soleil
J : nombre de jours sous lumière continue
T.U. : temps universel
•: pic du soir ; 0: pic du matin C50 jours sous LL)

Résultats -
1. Respiration sous régime photopériodique naturel.
L'activité respiratoire d' A. vulgare suit un rythme nycthéméral (Fig. l.A).
Pour 36 mâles d'une masse moyenne de 98,91 (±2,96)mg, la consommation d'oxygène
d’un individu est de 33,96 (±2,38)pl/h en moyenne la nuit, et de 24,35 (±3,42)pl/
h en moyenne le jour. Ces 2 valeurs sont significativement différentes (F = 5,64
pour 1 et 22 d.d.l.).
Cette activité est diphasique, c'est-à-dire qu'au cours du nycthémère deux
maximums alternent avec deux périodes de repos. Ces deux maximums d'activité se
situent, 1'un le plus important à la tombée de la nuit (46,16 µl/h) et l'autre
au lever du jour (30,33 µl/h).
2. Respiration sous conditions constantes de lumière.
Même en l'absence du cycle photopériodique naturel le rythme de la consomma-
tion d'oxygène se maintient environ 50 jours (Fig. l.B.C.D). Au bout de 50 jours,
la consommation d'oxygène se répartit de façon homogène; elle est de 20,50pl/h en
moyenne pendant le jour et de 18,24 µl/h pendant la nuit. Ces deux valeurs ne sont
pas significativement différentes (F = 0,54 pour 1 et 22 d.d.l.).
En lumière continue, tant que le rythme persiste le pic du soir reste le plus
important (Fig. 2). II se déplace régulièrement vers le milieu de la nuit (envi-
ron 1 heure par 15 jours, soit 3 minutes par jour), tandiâ, que le pic matinal
reste toujours à Droximité de l'heure du lever du soleil.
3. Respiration sous régime photopériodique artificiel LD 18:6, LD 6:18.
Pour 30 animaux utilisés de masse moyenne 117,16 (± 7,01)mg, et pendant le premier jour sous LD 18:6 et LD 6:18, la consommation d'oxygène présente les mêmes variations que celles vues sous photopériode naturelle (Fig. 3a, 3a'). Au bout d'un mois elle suit le rythme du nouveau cycle photopériodique imposé (Fig. 3b, 3b ' ), elle est monophasique, il existe un seul pic après extinction qui pourrait être l'homologue du pic du soir mais décalé dans le temps, cependant le pic matinal disparait.
4. Effet de la décérébration et du vernissage des yeux composés sur la consomma-tion d'oxygène. a) La décérébration à la pince (Fig. 4.A) de 10 individus d'une masse moyenne de 138,20 (±7,36)mg ou par électrocoagulation de 10 autres de masse moyenne de 80,22 (±2,32)mg provoque une diminution globale de l'activité respiratoire sous photopériode naturelle et une disparition de la rythmicité. b) Les 8 individus aux yeux vernis de masse moyenne de 97,25 (±2,13) mg et élevés sous photopériode naturelle présentent le même rythme que celui des 4 témoins de masse proche 95,50 (±2,60)mg. Nous n'avons pas relevé de différences significa-tives ni entre les valeurs obtenues au coucher du soleil (F = 0,03 pour 1 et 10 d.d.l), ni entre les valeurs obtenues au lever du soleil(F = 0,21 pour 1 et 10 d. d. 1.). Cependant sous conditions artif icielles» ils ne réagissent ni à l'allu— mage, ni à l'extinction de la lumière.
155

figure 3 : Taux de consommation d’oxygène d'A. vulgare (en microlitres par heure) sous cycle
photopériodique artificiel : L.D. 18:6 (a, b) et L.D. 6:18 (a‘,b‘).
»,a' : 1er jour sous L.D. E : extinction ; A : allumage ; T.U. : temps universel b,b' : 30 jours sous 1.0.
Figure 4 : Taux de consommation d'oxygène d’A. vulgare (en microlitrespar heure).
•—• Pour les témoins
0—0 et les Décérébrés A : depuis un jour à la pince
B : depuis une semaine au
coagulasem
L • lever du soleil
C : coucher du soleil
T.U. : temps universel
156

Figure 3 : Taux de consommation d'oxygène d’A. vulgare (en microlitres par heure) sous cycle
photopériodique artificiel : L.D. 18:6 (a, b) et L.D. 6:18 (a',b').
a,a' : 1er jour sous L.D. E : extinction ; A : allumage ; T.U. : temps universel
b,b’ : 30 jours sous 1.0.
Fiqure 4 : Taux de consommation d’oxygène d’A. Vulyare
(en microlitres par heure).
#—« Pour les témoins
0—0 et les Décérébrés A : depuis un jour à la pince
^ B : depuis une semaine au
coagulasem
L : lever du soleil
C : coucher du soleil
T.U. : temps universel
157

Conclusion et Discussion -
Nous retrouvons pour la respiration d'A. vulgare des faits analogues à ceux qui ont été signalés pour l'activite locomotrice.
En 1956, Cloudsley-Thompson a indiqué que l'activité locomotrice d'A. vulgare a une distribution bimodale avec aeux pics d'activité, l'un aux environs du crépuscule et l'autre aux environs de l'aube. Paris (1963) attribue cette acti-vité de l'aube au fait que les animaux cherchent un abri après leur activité noc-turne. L'existence d'une activité circadienne a été confirmée par Smith et al.
(1979) chez la même espèce.
Nos résultats permettent une approche des modalités de la synchronisation de ces rythmes par la photopériode.
La persistance pendant une longue durée d'une rythmicité circadienne sous lumière continue montre l'implication d'une ou plusieurs horloges internes. Le fait que les deux pics d'activité observés en conditions naturelles évoluent dif-féremment après mise en conditions constantes, laisse supposer qu'ils reflètent l'existence de deux oscillateurs circadiens plus ou moins autonomes.
L'extinction rapide du rythme après décérébration montre que le protocéré-bron est le siège de tout ou partie de ces systèmes, et que la destruction de cet organe ne se résume pas à une interruption des voies de perception des stimulus photopériodiques. En effet l'apposition de vernis sur les yeux n'abolit pas immédiatement la rythmicité. Ceci pourrait être le résultat d'une occultation imparfaite du système visuel. En fait il semble bien que l'aveuglement soit effectif puisque les animaux aux yeux vernis ne présentent plus de réponse immé-diate à l'extinction ou à l'allumage en lumière artificielle.
Les résultats ci-dessus sont à rapprocher de ceux obtenus récemment dans nutre laboratoire concernant la reproduction de la même espèce. L'étude de 1 ' in-duction photopériodique de l'entrée en reproduction laisse également supposer l'existence de deux oscillateurs circadiens.
CXCUDSLEY-ThCM’SCN J.L., 1956. Studies in diumal rtiythms. VII. Hunidiiy responses and noctumal activity in woodlice (Isopoda). J. exp. Biol., 33-, 576-582. EDNEY E.B., 1960. Terrestrial adaptation (in : Waterman T.H., ed. ). The Physiology of Crustacea, 1. 367-393.
PARIS D.H., 1963. Ihe ecology of Arrrndillidim vulgare (Iscpoda : Oniscoïdea) in California grassland : food, ennemies and weather. Ecol. Monogr., 33, 1-22. M J.T.F., LARIMER J.L., 1979. Circadian wheel-running behaviour in the Iscpod, Arrrndillidiwi vulgare. J. exp. Zool., 209, n°l, 73-80. WIESER W., SCHWEIZER G., HARTEHBSTEIN R., 1969. Pattem of reJ ease of Gaseous ammonia by terrestrial arthropods. Oeoologia (Berl.), 3 , 390-400.
158

ASPECTS RÉCENTS de la Biologie des CRUSTACÉS Actes de Colloques - 8 - 1988 - IFREMER, pages 159 à 164
ETUDE DES FACTEURS DE L'ENVIRONNEMENT (TEMPERATURE ET PHOTOPERIODE) SUR LA
REPRODUCTION DE DIFFERENTES SOUCHES D'Armadillidium vulgare Latreille (Crustacé,
Isopode, Oniscoïde).
SOUTY—GROSSET C.(1) MOCQUARD J.P.(1) JUCHAULT P.(1) HAFIDI N.(1)
(1) (1)
RESUME - Les modalités de la reproduction d'Armadillidium vulgare issues de
latitudes variées ont été étudiées dans différentes conditions de photopériode
et tempêrature. Les résultats obtenus montrent que les femelles des basses
latitudes se reproduisent plus rapidement que celles des hautes latitudes.
Cependant, d'autres caractéristiques du lieu d'origine semblent intervenir.
Mots-clés : Facteurs de 1'environnement, reproduction, variation géographique ,
Armadillidium vulgare, Crustacé.
ABSTRACT - The main characteristics of the reproduction of several strains of
Armadillidium vulgare from different latitudes have been studied in various con-
ditions of photoperiod and temperature. The results show that females from low
latitudes undergo the reproduction process more rapidly than those of higher
iatitudes. However other biogeographic characteristics should be considered.
Key-words : Environmental factors, reproduction, geographic variation,
Armadillidium vuigare, Crustacea.
Introduction -
A. vulgare est une espèce d'origine méditerranéenne qui se serait répandue dans diverses régions extratropicales de l'Asie et de l'Amérique après y avoir
été introduite par l'homme. Les croisements entre des souches éloignées sont
féconds et les variations étudiées sont donc intraspécifiques. Nous avons
étudié au laboratoire, dans différentes conditions photopériodiques,1'entrée
puis le maintien en reproduction de dix souches issues de latitudes variées :
Sucre (SUC, Bolivie, 19°02'S, altitude 2815m), Teneriffe (TEN, Canaries 28°27'N)
Riverside (RIV, Californie, 33°58'N), Rabat (RAB, Maroc 34°02'N), Mostaganem
(MOS, Algérie 35°51'N), Sete (SET, France, 43°24'N), St. Georges les B. (GEO,
France 46°40'N), Santec (SAN, France, 48°42'N), Northampton (NOR, Angleterre,
52°14'N), Helsingjir (HEL, Danemark, 56°02'N).
1. Variation géographique de l' entrée en reproduction chez des femelles
accouplées soumises à la photopériode de Poitiers. (Fig. I)
Les animaux sont nés au iaboratoire et n'ont donc pas été soumis aux
conditions climatiques originelles de leurs parents. Ils sont élevés à 20°C,
sous la photopériode naturelle de Poitiers (46°40'N). Chaque femelle, initiale-ment élevée sans mâle, a été accouplée en décembre. Le délai d'entrée en
reproduction a ézé exprimé en nombre de jours à partir du 1er Janvier. La date d'entrée en reproduction de chaque souche est d'autant plus tardive que la
latitude d'origine est la plus haute (avec environ deux jours de retard pour
chaque degré de latitude supérieur), à l'exception de la souche de Sucre qui se
comporte comme celle de Santec (48°42'N).
(1) Laboratoire de Biologie Animale de l'Université de Poitiers et UA CNRS
n° 673 "Physiologie et Génétique des Crustacés", 40, Avenue du Recteur
Pineau, 86022 Poitiers Cédex, France.
159

Fig. I : Délai moyen d'entrée an reproduction en fcncticn de la latitude du lieu d'origine de la soudie ; = délai exprimé an nombre moyen de jours à partir du 1er Janvier (les écarts-types étant toujours inférieurs à 4 jcurs ne scnt pas représentés) ; RD = durée du jour (photcphase) à Poitiers pour la date ocnsidérée.
Fig. II : Induction de la reproducticn à 20°C, LD 18:6 pourcentage de femelles ayant effectué une première mue de nature parturielle en fanction de la latitude du lieu d'origine de la souche.
160

Fig. m : "Maintien de la reproduction" : pour chaque souche, pourcentEge de femelles ayant effectué une mue parturielie en foncticn des différentes ccnditicns de température et de photo-période : 15°C (1,2) ; 20°C (3,4) ; 25°C (5,6) ; LD 6:18 (1,3,5) ; ID 18:6 (2,4,6).
161

2. Variation géographique de l'entrée en reproduction chez des femelles isolées
élevées en jours longs. (Fig. II)
La ponte intervenant même en l'absence d'accouplement, chaque femelle est
élevée isolément (à 20oC, LD 18:6) car la présence du mâle accélère l'entrée
en reproduction de la femelle et induit un allongement de la durée de la période
de la reproduction (Jassem et al., 1982). Chaque souche a été caractérisée par
le pourcentage de femelles dont la première mue après la mise en condition est
parturielle. Les résultats montrent que les femelles de basses latitudes
entrent en reproduction plus rapidement que celles provenant de latitudes plus
élevées, à l'exception de la souche de Sucre qui se comporte encore comme
celle de Santec.
3. Variation géographique du "maintien en reproduction" dans différentes
conditions de température et de photopériode. (Fig. III)
Juchault et al. (1980) ont montré que les différences entre souches
portent aussi sur la durée de la période de reproduction, fonction du nombre de
mues parturielles successives. En ce qui concerne les souches étudiées nous
avons constaté que les femelles des Canaries élevées sous la photopériode de
Poitiers effectuent jusqu'à 5 pontes successives, alors que celles du Danemark
n'en font qu'une. Dans cette expérience, nous avons provoqué une première mue
parturielle à 20°C LD 18:6 chez des femelles isolées, issues des dix souches,
puis nous avons étudié le maintien en reproduction de ces femelles soumises à différentes conditions de température e.tfde photopériode à partir du jour de leur mue parturielle. Six combinaisons ont été réalisées avec trois températures
(15°C, 20°C, 25°C) et deux photopériodes (LD 6:18 et LD 18:6). Chaque population,
a été caractérisée par le pourcentage de mues parturielles lors de la première mue suivant le changement de température (T) et de photopériode (P).
D'une manière générale (Fig. III) le maintien en reproduction est d'autant
plus difficile que la souche provient d'une latitude élevée. Hormis la souche la
plus nordique (HEL) où la reproduction s'arrête toujours, on observe deux cons-tantes : maintien de la reproduction à température élevée et sous longue photo-
phase (6), à l'opposé arrêt absolu à basse température et sous courte photophase
(1). Par contre les réponses aux autres conditions permettent de différencier
les souches. II est ainsi possible de caractériser un premier groupe, nordique
ou européen (SET, GEO, SAN, NOR) : pas ou très peu de reproduction sous les conditions 2 et 3, un nombre de femelles parturielles toujours inférieur à 100% et variant dans le sens inverse de la latitude sous les conditions 4 et 5. Le
deuxième groupe est caractérisé par un maintien absolu de la reproduction sous les conditions 4 et 5 et un nombre plus ou moins élevé de mues parturielles à
basse température mais photophase longue (2). L'action de l'éclairement semble
primer celle de la température (sauf pour la souche TEN). Les deux souches américaines (SUC et RIV) semblent les plus sensibles aux conditions défavorables.
162

CONCLUSION -
Chez les femelles dM. vulgare les modalités de la reproduction sont fonc-
tions de la latitude du lieu d'origine de la souche : les femelles des basses
latitudes se reproduisent plus rapidement que celles des hautes latitudes et
se maintiennent plus facilement en reproduction. D'autres caractéristiques du
lieu d'origine que la latitude semblent intervenir : la reproduction tardive de
la souche de Sucre pourrait être une adaptation à un climat plus rude dû à
l'altitude. Par contre les différences entre des souches issues de régions de
latitude et d'altitude analogues (Northampton et Helsingor, Rabat et Riverside)
semblent montrer que la différenciation entre populations n'est pas seulement
en relation avec les facteurs climatiques généraux. II faut souligner que les
expériences ont été menées sur des animaux nés au laboratoire et donc que les
caractères observés sont bien génétiquement fixés. Des résultats, non relatés
ici, ont montré un déterminisme génétique des modalités de la reproduction,
probablement multifactoriel avec intervention de gènes autosomaux et de gènes
liés au sexe.
JASSEM W., JUCHAULT P., MOCQUARD J.P., 1982. Déterminisme de la reprodueticn saisonnière des femelles
dM. vulgare Latr. (Crustacé, Iscpode, Oiiscoïde). V. Rôle du mâle dans le cycle de reproductiœ des
femelles (induction et durée de la période de reproductiœ). Ann. So. Nat. Zool. Paris, 4, 195-201.
JUCHAULT P., PAVESE A., MXQUARD J.P., 1980. Déterminisme de la reproducticn saisonnière des
femelles d'A. mlgare Latr. (Crustacé, Iscpode, Oniscoîde). II. Etude en conditions expérimentales
de femelles d'origines géographiques différentes. Artn. So. Nat. Zoot., Paris, 2, 99-108.
163

ASPECTS RÉCENTS de la Biologie des CRUSTACÉS Actesde Colloques - 8 - 1988 - IFREMER, pages 165 à 170
ECOPHVSIOLOGIE DE LA REPRODUCTION DE LA CREVETTE DES MARAIS, PALAEMON'ETES VARIANS ( Leach 1811)
D.BOUCHON °
Résumé - L’application de photophases fragmentées montre que la perception de la longueur du jour chez Palaemonetes varians est sous la dépendance d’un système circadien. Ce système serait formé de deux oscillateurs, l’un synchronisé par 1’aube, l’autre par le crépuscule. La valeur absolue de la durée d’éclairement intervient néanmoins dans l’obtention d’un effet de type jours longs.
mots-clés : Palaemonetes varians. photopériodisme, reproduction, écophysiologie
Abstract - Squeleton photoperiods experiments show that a circadian system is involved in the perception of the day length in Palaemonetes varians. This system consists of two oscillators, one synchronized at dawn and the other at dusk. Nevertheless, the illumination length must be taken into account for a long day effect.
key-words : Palaemonetes varians. photoperiodism, reproduction, ecophysiology
1. INTRODUCTION
Palaemonetes varians est une espèce très bien adaptée aux milieux littoraux saumâtres. Sa durée de vie maximale est proche de 24 mois avec une période de reproduction s’étendant de Mai à Septembre (Laurent-Bizot-Espiard 1984).
Au laboratoire, les crevettes sont élevées dans un aquarium à circuit fermé, qui comprend une salle en conditions naturelles et 12 caissons indépondants, programmables en température et photopériode. Nous retiendrons deux critères de comparaison entre les différentes séries expérimentales : le pourcentage de femelles ovigères d’une part, et les dates d’entrée en reproduction, d’autre part.
° Laboratoire de Biologie Animale, Université de Poitiers, U.A. C.N.R.S. 673, 40 avenue du Recteur Pineau, 86022 Poitiers cedex
165

Fig. 1 : Activité reproductrice
Fig. 2 : Pélai d'entrée en reproduction
166

2. RESULTATS
Dans un preraier temps, nous avons étudié l’action corabinée de la température et de la photopériode sur l’induction de la reproduction de P. varians. Les résultats obtenus (Bouchon et al■. 1985) montrent que la température est un facteur limitant l’action de la photopériode. A température hivernale, quel que soit le régime photopériodique, les femelles ne pondent pas. A température pius élevée une courte photophase ne suffit pas à maintenir les femelles en repos sexuel, la ponte est retardée mais inéluctable. L’association d’une température estivale et d’une longue photophase provoque l’entrée en reproduction et ce, d’autant plus rapidement que la durée d’éclairement est proche d’un optimum de 18 heures (*).
Pour préciser l’action du photopériodisme, nous avons effectué 2 groupes d’expérimentations utilisant des photophases fragmentées. Dans chaque groupe, nous avons pris comme référence une série d’animaux soumis à une photophase unique de 18 heures, c-onsidérée comme optimale. Les dates d’entrée en reproduction de toutes les séries ont été calculées en prenant pour point 0, les dates obtenues dans les 2 séries références.
Le premier groupe d’expérimentations comporte 7 séries (figure 1, groupe 1). La série 1 est la série référence. Dans la série 2 les crevettes sont élevées sous une photopériode LD 18:30. Elles ne subissent donc une longue photophase qu’un jour sur deux. La série 3 regroupe des individus soumis à une photopériode LD 12:12, admise comme non stimulante. Dans les séries 4 et 5 nous appliquons une photophase unique de 9 heures, soit dans la première partie de la phase diurne (série 4), soit dans la seconde partie (série 5). Enfin, les séries 6 et 7 reprennent le même protocole expérimental, mais en ajoutant un flash lumineux d’une heure, soit à 1’aube (série 6), soit au crépuscule (série 7) d’un jour théorique de 18 heures.
En comparant les pourcentages de femelles ovigères dans les différentes séries, nous relevons une activité reproductrice maximale pour la série 2, supérieure à la série 1 référence. Les séries 4 et 5 montrent un très faible taux de femelles parturielles, quel que soit la position, durant le jour, de la phase éclairée. Elle est inférieure à celle obtenue dans la série 3, sous 12 heures de lumière. Les résultats des séries 6 et 7 sont comparables à ceux de la série 3 et dénotent une activité reproductrice moyenne.
Les séries 1 et 2 ont même délai d’apparition des premières mues parturielles (figure 2, groupe 1). Les femelles des séries 4 et 5 ont un comportement identique et entrent en reproduction très tardivement (57 jours +-20). Les femelles des séries 6 et 7 font une mue parturielle de façon synchrone. Elles ont un retard de 5 jours sur les femelles de la série 1.
(* : cette Sud-Vilaine l’auteur au
étude a fait l’objet dont le rapport
C.R.E.M.A L’Houmeau
d’un contrat G.I.S de la région scientifique a été présenté par en Mars 1986)
167

Ces résultats montrent que la mesure du jour, ches P . varians nécessite la présence conjointe de 2 phases éclairées, l’une à l’aube et l’autre au crépuscule. II semble toutefois que le simple "bornage" aube/crépuscule ne soit pas optimal, comme le montrent les séries 6 et 7, puisque qu’un jour "subjectif" de 18 heures ne produit pas le même effet qu’une photophase unique de même durée. On ne peut distinguer de prépondérance particulière de 1’aube ou du crépuscule puisque les séries 4 et 5 puis 6 et 7 produisent respectivement les mêmes effets. De plus la série 2 montre qu’une longue photophase, appliquée toutes les 48 heures, a un effet de jour long. A 1’évidence, il existe un rythme circadien endogène impliqué dans la mesure du temps d’éclairement.
Dans un deuxième temps nous avons mis au point 8 nouveaux protocoles (figure 1, groupe 2). Les individus de la série 1 ont servis de référence. Les séries 2, 3 et 4 sont des photophases fragmentées symétriques dont les flashes lumineux sont aux bornes d’un jour subjectif de 18 heures. La durée de ces flashes suit une progression arithmétique de la série 2 à la série 4. Les séries 5, 6, 7 et 8 simulent le parcours d’une photophase entière de 15 heures au long du nycthémère.
Les taux d’activité reproductrice relevés dans les séries
2, 3 et 4 évoluent dans le même sens que la valeur absolue de la durée d’éclairement. La série 4, soumise à 14 heures absolues de lumière, a un pourcentage de femelles ovigères comparable à celui obtenu sous une photophase unique de 18 heures (39% contre 42%). Les séries 5, 6, 7 et 8 ont des taux semblables, indiquant une activité reproductrice moyenne.
Les dates d’entrée en reproduction relevées (figure 2, groupe 2) confirment les résultats obtenus pour les taux d’activité reproductrice. Les séries 2, 3 et 4 ont un délai moyen d’apparition des preraières mues parturielles qui augmente avec la quantité de lumière reçue. Les femelles de la série 4 se détachent nettement et entrent en reproduction plus tôt que
les femelles de la série 1. Les femelles des séries 5 à 8 ont un comportement similaire, avec un délai moyen d’entrée en reproduction supérieur à celui obtenu dans la série 1.
Les résultats des séries 2 à 4 montrent que des photophases fragmentées symétriques ont un effet semblable à
une photophase unique de même durée bien qu’intervienne, en synergie, la valeur absolue du temps d illumination. Les dates
d’entrée en reproduction similaires obtenues dans les séries o
à 8 montrent que des photophases uniques de même durée
appliquées en quelque point du nycthémcre produisent les mèmes
effets. II semble donc que le système circadien endogène de P._ varians ne présente pas de phase photosensible paiticulière.
3. CONCLUSION
La perception de la longueur du jour chez P. varians est
sous 1a dépendance d’uri système circadien endogène.
L’application d’une photophase matinale ou crépusculaire
unique, ne suffit pas à provoquer un effet de .iour long.
168

II semble donc que la simple coïncidence d’un point
particulier de l’oscillation circadienne interne et de la photophase ne permette pas la mesure du jour. L’on doit
supposer l’existence de 2 oscillateurs, l’un synchronisé par
l’aurore, l’autre par le crépuscule. L’induction
photopériodique serait alors fonction de la relation de phase
de ces 2 oscillateurs ("coïncidence interne" de Pittendrigh
1972). Les résultats obtenus ici, avec les photophases fragmentées
symétriques, semblent prouver un tel fonctionnement chez P_i. varians ■ N'ous avons montré, cependant, que la synchronisation aube/crépuscule n’était pas suffisante en elle-même, mais
qu’intervenait la valeur absolue de la durée d’éclairement.
BOUCHON D., REMOISSENET G., MOCQUARD J.P., 1985. Influence de la photopériode et de la température sur l’entrée en reproduction de Palaemonetes varians Leach (Crustacé décapode natantia). Bull. Soc. Zool. Fr., 110(4), p. 439-445.
LAURENT-BIZOT-ESPIARD A., 1983. Ecologie, dynamique de population et valeur nutritive de la crevette Palaemonetes varians dans les marais du littoral charentais. Convention entre l’E.P.R. Poitou-Charentes, l’Université de Poitiers, et l’A.D.A.C.O., rapport d’activité, 50p.
PITTENDRIGH C.S., 1972. Circadian surfaces and the diversity of possible roles of circadian organization in photoperiodic induction. Proceedings of the National Academy of Sciences U.S.A, 69, p. 2734-2737
169

ASPECTS RÉCENTS de la Biologie des CRUSTACÉS Actes de Colloques - 8 - 1988 - IFREMER, pages 171 à 174
STRUCTURES ADAPTATIVES A LA FILTRATION DES ALIHENTS CHEZ LE CRABE DES SABLES, SCOPIMERA GORDONAE SERENE AND MOOSA 1984.
A- BAUCHAU
RESUME - Chez le crabe des sables, Scopimera gordonae , on observe une
adaptation très particulière des pièces buccales, assurant une mobilisation
du sable et son brassage en vue de détacher le matériel nutritif. Ces
observations indiquent que Scopimera gordonae joue un r61e d'éboueur de
la pellicule superficielle du sable.
Mots-clés . pièces buccales, filtration, adaptation, Scopimera gordonae
ABSTRACT - In the sand crab, Scopimera gordonae the buccal appendages
are adapted to an efficient mobilization and washing of the sand in order to
collect the nutritive elements. These observations confirm the role of
Scopimera gordonae in the recycling of the organic material associated with
the sand particles.
Key-words : buccal pieces, filtration, adaptation, Scopimera gordonae .
En 1981, Serene et Moosa décrivaient sur matériel indonésien une
nouvelle espèce de Scopimera (famille des ocypédidés), le Scopimera
gordonae . Nous l'avons retrouvée en Papouasie-Nouvelle Guinée (1984-1985) et
avons pu préciser son mode d'alimentat,ion assez particulier, étroitement lié
à la nature du sable des plages.
La Papouasie-Nouvelle Guinée présente un relief très tourmenté. II
s'agit en effet du front Nord de la plaque indo-australienne soumise aux
poussées de la grande plaque Pacifique et de celle des Philippines. Des
tremblements de terre y sont fréquents. Ce front présente une ceinture de
volcans soit en activité (Manam), soit effondrés et envahis par la mer
(Hansabay). On trouve sur les plages de cette région un sable noir aux grains
très fins qui convient particulièrement à l'alimentation des Scopimera. Ce
sable contraste de manière frappante avec le sable corallien clair et plus
grossier de l'ile de Laing et des baies du sud de Hansabay où Scopimera est
absent. Nos observations ont été faites à la station de recherche de
l'Université Libre de Bruxelles, située dans la baie de Hansa.
Sur la moitié supérieure de l'estran de sable noir (en deçà des limites
de marées hautes), Scopimera gordonae laisse de nombreuses traces très
artistiques de son activité alimentaire. II racle la pellicule superficielle
(+-2 mm de profondeur) avec ses chélipèdes et trie rapidement les matières
organiques avec ses appendices buccaux, tandis que la partie minérale
indigeste est rejetée en arrière sous forme de boulettes (2-4 mm)
régulièrement alignées au rythme de 8 par minute. II avance latéralement
(1) Département de Biologie, Facultés Universitaires N-D de la Paix, Namur, Belgique.
171

jusqu'à une trentaine de centimètres de son terrier, fait alors une volte-face de +- 180°, progresse en direction centripède et pivote à nouveau, ce qui donne un parcours en zig-zag autour de celui-ci. Ce petit terrier
consiste en un simple puits vertical de 20 à 25 cm de profondeur qui atteint
la couche sablonneuse fortement imbibée d'eau. II s'y enferme pendant la
marée haute, s'y réfugie en cas d'alerte et y trouve l'eau nécessaire pour
maintenir une hydratation suffisante de ses branchies.
Son travail est rapide et efficace grâce à une adaptation très
particulière des pièces buccales : maxillipèdes, maxilles et mandibules. Le
Scopimera gordonae utilise ses deux chélipèdes pour amener le sable à la
base inférieure des 3ème maxillipèdes. Ces pièces buccales ont un merus et un
ischion suturés et élargis qui forment un bouclier couvrant tout l'épistome.
Le sable est ensuite acheminé entre les 2ème et 1er maxillipèdes où le
tri se fera gràce à deux mécanismes concomitants. D'une part, un courant
d'eau venant de la chambre branchiale passe entre les deuxième et premier
maxillipèdes libérant les particules organiques légères d'avec le sable plus
gros. D'autre part, la face interne des 2ème et la face externe des 1er
maxillipèdes sont tapissées de soies spécialisées en forme de cuillère au
bord distal muni de trois dents. Entre ces "cuillères" sont réparties des
soies cylindriques et rigides qui les soutiennent et qui empêchent
l'encrassement de leur base. Les maxillipèdes 2 et 1 sont mobiles et brossent
le sable en détachant ainsi le matériel nutritif tandis que les grains de
sable sont chassés à leur sommet où ils s'accumulent en boulettes.
Les aliments sont, eux, entrainés vers les maxilles. La face externe du
lobe supérieur des maxilles 2 est couverte de soies en cuillère finement
dentelée. Celle des maxilles 1 est couverte de soies en lancette armée. Les
maxilles 1 et 2 fonctionnent comme deuxième écran filtrant.
II existe un gradient dans la structure des soies des mandibules par
rapport à l'ouverture de la bouche : les plus distales sont courtes et
coniques : ensuite elles s'allongent et leur extrémité se recourbe. Les plus
proximales s'allongent encore et leur extrémité est concave et dentelée. La
surface lisse des maxilles et mandibules est couverte de bactéries
probablement collectées par les brosses de soies voisines pour être ingérées. En fin de parcours, la matière alimentaire est dirigée vers la
bouche par les rangées de longues soies plumeuses implantées sur les bords
supérieurs des mandibules bilobées.
Ces observations indiquent que le Scopimera gordonae joue le rôle
d'éboueur de la pellicule superficielle du sable et intervient ainsi dans le
recyclage des matériaux organiques qui y sont présents.
BAUCHAU A.G. and E. PASSELECQ-GERIN, 1987. Adaptative structure to filter
feeding in the sand crab Scopimera gordonea . Serene and Moosa, 1981.
Indo-Malayan Zoology (sous presse).
FIELDER D.R., 1970. The feeding behaviour of the sand crab Scopimera
inflata (Decapoda Ocypodidae). J. Zool. , London, 160 : 35-49.
SERENE R. and M.K. M00SA, 1981. Description de Scopimera gordonae
sp.nov. (Crustacea Decapoda Brachyura) , une espèce des eaux orientales
172

d'Indonésie. Bull- Inst. r. Sci. Natur. Belgique . Biologie 53 : 1-9.
SXLAS E.G. and C. SANKARANKUTTY, 1967. Field investigations on the shore
crabs of the Gulf of Mannar and Palk Bay with special reference to the
ecology and behaviour of the pellet crab Scopimera proxima Kemp. Symp.
Series Mar. Biol. Assoc. india . Serie 2 : 1008-1025.
TWEEDIE M-W.F., 1950. Notes on grapsoid crabs from the Raffles Museum.
Bull. Raffles Mus. 23 : 310-324.
V0GEL F. , 1984. Comparative and functional morphology of the spoon-tipped
setae on the second maxillipeds in Dotilla Stimpson, 1858 (Decapoda
Brachyura Ocypodidae). Crustaceana , 47 : 225-234.
173

THÈME 4 - POPULATIONS
- Biologie et génétique des populations de
Crustacés : stratégies mises en jeu.
Essai de synthèse.
Gaudy R. et Kerambrun P.
- Etude du polymorphisme enzymatique
dans cinq populations naturelles
d' Armadillidium vulgare
Raimon R.
- Les âges de Niphargus virei
Turquin-Buffière M.J.
- Complément à la définition " Idotea chelipes ,
espèce polytypique" à partir des fractions
protéiques et particulièrement de 1'hémocyanine.
Comparaison avec Idotea balthica basteri .
Charfi-Cheilchrouha F.
- Reproduction et croissance chez les femelles
du Crustacé oniscoïde Armadillidium
vulgare après inoculation d'une bactérie
symbiotique féminisante.
Juchault P. et Mocquard J.P.
- Actualisation des données sur l'écologie, la
biogéographie et la phylogénie des Ampeliscidae
(Crustacés, Amphipodes) atlantiques après la
revision des collections d' E. chevreux . Bellan-Santini D. et Dauvin J.C.
- Données préliminaires sur les Amphipodes
de l'Iroise et de ses abords, leurs
affinités géographiques.
Dauvin J.C. et Toulemont A.
- Biologie du Copepode Eurytemora affinis
hirundoides dans la Gironde :
données morphométriques.
Feurtet A. et Castel J.
175

ASPECTS RÉCENTS de la Biologie des CRUSTACÉS Actes de Colloques - 8 - 1988 - IFREMER, pages 177 à 190
BIOLOGIE ET GENETIQUE DES POPULATIONS DE CRUSTACES STRATEGIES MISES EN JEU, ESSAI DE SYNTHESE-
GAUDY R.(1) KERAMBRUN P. (1)
RESUME - La population, située à un niveau interraédiaire entre l'individu
et l'espèce, constitue une structure dynamique caractérisée par différents
flux.
Un flux somatique correspondant à l'arrivée et au départ
d'individus par les processus de natalité et mortalité. Cet aspect,
essentiellement de nature quantitative, fait l'objet principal de la Biologie
des Populations (aspect démographique). Dans la nature, ce flux est
contrôlé par les interactions abiotiques et biotiques se manifestant au
niveau écophysiologique ou dans les relations de compétition et de prédation.
- Un flux génique, de nature plus spécifiquement qualitative, qui est
à la base du polymorphisme des populations.
Une synthèse des principales données de la littérature concernant ces
deux approches du concept de population chez les crustacés est proposée.
La Biologie des Populations, par le biais du flux somatique, analyse
les variations quantitatives matérialisées par le potentiel biotique r
(taux intrinsèque de croissance), q,ui indique les capacités théonques
maximales d'expansion de la population et par le taux de croissance
instantanée r, réellement observé dans la nature, qui résulte de
1'antagonisme des taux de natalité et de mortalité.
La Génétique des Populations , par le biais du flux somatique, analyse
les variations quantitatives matérialisées par le potentiel biotique r
(taux intrinsèque de croissance) qui indique les capacités théonques
maximales d'expansion de la population et par le taux de croissance
instantanée r, réellement observé dans la nature, qui résulte de
l'antagonisme des taux de natalité et de mortalité.
La Génétique des populations , qui analyse la structure génétique des populations naturelles, a pour objectif une meilleure compréhension des
mécanismes de maintien du polymorphisme et des relations entre celui-ci et
les conditions physiques, chimiques et trophiques de 1'environnement. Dans
cette double finalité, elle rejoint la Biologie des Populations dont les
aspects démographiques peuvent être perturbés par une baisse de
polymorphisme (dépression consanguine) ou par des modifications des
conditions écologiques.
Mots-clés : Crustacés, Biologie des Populations , Génétique des Populations.
(1) Centre d'Océanographie de Marseille ; Faculté des Sciences de Luminy 13288 Marseille Cedex 9, France.
177

ABSTRACT - The population, which is situated at an intermediary level
between the individual and the species, constitutes a dynamic structure
characterised by different flows.
- A somatic flow corresponding to the arrival and the departure of
individuals by the processes of birth and mortality. This aspect, essentially
of a quantitative nature, is the main object of the Biology of Populations
(demographic aspect). In nature, this flow is controlled by biotic and
abiotic interactions which are manifested at the ecophysiological level or in
the relations of competition and predation.
A genic flow, of more specifically qualitative nature , which is at
the base of the polymorphism of populations.
A synthesis of the main published data concerning these two
approaches of the concept of population is proposed for crustaceans. The
Biology of Populations, via the somatic flow, analyses the quantitative
variations materialised by the biotic potential r (endogenic rate of
growth) which indicates the maxima theoretical capaci?ies of expansion of the
population and by the instantaneous rate of growth r, really observed in
nature, which results from the antagonism of the rates of birth and
mortality.
Population Genetics , which analyses the genetic structure of natural
populations, has as an objective a better understanding of the mecanisms of
support of the polymorphism and of the relations between this one and
physical, chemical and trophic conditions of the environment. In this double
objectif, Population Genetics meet the Biology of Populations whose
demographic aspects can be disturbed by a decrease of polymorphism
(inbreeding) or by changes in ecological conditions.
Key-words . Crustaceans, Populations Biology, Populations Genetics.
178

1. INTRODUCTION
Se situant à un niveau de concept intermédiaire entre celui d'individu
et celui d'espèce, la notion de population est d'une approche moins directe
car elle sous-tend un ensemble de processus en interactions avec le milieu
ou les autres organismes, qui n'apparaissent souvent qu'à l'issue d'une
analyse plus ou moins laborieuse. La population au sens où nous l'entendrons
ici peut être définie par sa structure: ensemble des individus d'une même
espèce qui occupent, en panmixie, un espace donné à un moment donné
("dème") ou par son fonctionnement, comme une unité dotée de propriétés
particulières au sein de 1'écosystème, et non comme une simple juxtaposition
d'individus indépendants. En particulier, les notions de capacités physio-
logiques (tolérance aux facteurs du milieu) définies au niveau de
l'individu, ne peuvent être extrapolées au niveau populationnel, qu'en
ajoutant aux critères de survie de l'individu, ceux de survie de la popula-
tion, ces derniers étant basés en grande partie sur les processus de
natalité. On peut ainsi concevoir pour un organisme donné deux niches écolo-
giques emboitées, une niche "somatique" définissant l'étendue de variation
des facteurs permettant la survie de l'individu, et une niche "germinale",
plus restrictive et donc incluse dans la précédente, dans laquelle est
précisée l'étendue des mêmes facteurs permettant un processus de
reproduction suffisant pour assurer la pérennité de l'espèce d'une
génération à l'autre, en tenant compte de la mortalité d'origine diverse
affectant les individus de tous âges composant la population.
La population, bien qu'isolant une collection d'individus de même
espèce, ne constitue pas une entité figée, mais représente au contraire une
structure dynamique caractérisée par un ensemble de flux de différentes
natures: en dehors de l'ensemble de relations avec le milieu, de nature
physique ou chimique, dans le cadre du bilan d'énergie (nutrition, rejet
d'excrétats) ou dans celui de 1'interaction avec les autres organismes de
l'écosystème (compétition, prédation) la population est essentiellement
traversée par un flux propre à sa matière vivante; la population est
concernée continuellement par un apport d'individus (natalité ou immigra-
tion) et par une perte due à la mortalité ou à 1'émigration, ce courant
étant plus ou moins rapide selon les caractéristiques abiotiques ou
biotiques du milieu (hydrodynamisme, température, ressources trophiques,
abondance des prédateurs, etc...) ou selon les modalités du cycle biologique
propre à l'espèce.
Ce flux de matière somatique s'accompagne d'un flux génique dont
l'essentiel est interne à la population étudiée (panmixie), mais dont une
partie peut concerner un autre dème isolé jusque là et pouvant être atteint
pour des raisons diverses (introduction volontaire ou accidentelle,
suppression de barrières physiques ou chimiques, etc...). Ce dernier
processus est d'une importance considérable pour le maintien du polymorphis-
me génétique donnant à l'espèce les potentialités nécessaires pour survivre
à une modification de son environnement.
Ainsi, l'étude des variations spatio-temporelles des populations peut
être abordée par deux voies différentes: la Biologie et la Génétique des
Populations. Notre propos n'est pas de fournir un exposé exhaustif de ces
deux approches chez les crustacés, mais, par l'analyse d'un certain nombre
d'exemples, d'essayer de montrer quelques stratégies mises en jeu par les
populations et d'en dégager les aspects complémentaires.
Historiquement, cependant, le lien entre biologie et génétique ne s'est
resserré que récemment, à la suite notamment des importants progrès de la
génétique moderne, alors que la sensibilisation aux problèmes de biologie
des populations est plus ancienne puisqu'elle remonte aux idées de Malthus,
largement prises en compte par la suite dans la théorie évolutionniste de
Darwin. Depuis Malthus, la Biologie des Populations animales a surtout
179

progressé en prenant pour objet d'étude l'homme lui-même, pour qui on dispose traditionnellement de statistiques détaillées sur une longue période (état civil), et sur ce qui touche à son alimentation, à son bien être ou à la protection de son environnement: Xes ravageurs de récoltes, les vecteurs de maladies, le gibier et ses prédateurs, etc...
Les plus gros progrès dans l'analyse théorique des phénomènes ont ainsi été obtenus chez les rongeurs et leur prédateurs, et surtout chez les insectes propices à l' élevage en cycle rapide d'individus nombreux. C'est également chez les insectes (la drosophile en particulier) et pour les mêmes raisons que la Génétique des Populations animales a fait le plus de progrés et amené les hypothèses les plus fécondes au plan théorique.
Chez les crustacés, l'étude de la Biologie des Populations a surtout été développée chez les entomostracés (cladocères et copépodes en particulier) qui constituent un matériel favorable en raison de la fiabilité relative des stratégies d'échantillonnage utilisables dans la nature, et dans la mesure où la rapidité de leur cycle biologique permet d'envisager l' élevage en cycle complet d'individus nombreux sous des conditions aisément contrôlables. En Génétique des Populations, l'éventail des groupes zoologiques pris en compte est plus étendu en raison de la diversité des critères quantitatifs ou qualitatifs étudiés.
2. BIOLOGIE DES POPULATIONS
Sous ce terme, nous regroupons les concepts de démographie et d'écolo-gie des populations que nous considérons inséparables, puisque, par opposition à la biologie de l'individu, qui sous-entend surtout l'étude des grandes fonctions physiologiques (nutrition, métabolisme, relation, repro-duction), la Biologie des Populations fait essentiellement appel à la fonction de reproduction et à son efficacité à long terme, matérialisée par le maintien de générations successives (faisant l'objet de l'étude démographique), ceci en fonction des conditions biotiques ou abiotiques du milieu (analysées dans une optique écologique).
Les fluctuations temporelles des populations sont la résultante de leurs capacités physiologiques de croissance et des contraintes de l'envi-ronnement. La capacité maximale d'expansion numérique est caractérisée par le paramètres r , appelé selon les auteurs taux intrinsèque d'accroissement naturel, capacTté innée d'accroissement en nombre, paramètre de Malthus, ou potentiel biotique. II caractérise une population se développant exponen-tiellement dans un environnement idéal, sans limitation trophique ou spatiale et sans prédateur. Ce paramètre, qui est une grandeur sans dimension, permet des comparaisons de potentiels d'expansion dans l'ensemble du règne animal; il est d'autant plus élevé que la taille de l'espèce est petite, car il dépend beaucoup de la rapidité du cycle biologique. Par ailleurs, il peut varier selon les conditions du milieu, la température notamment, si bien que les différentes valeurs trouvées dans la littérature ne sont pas forcément comparables si les conditions optimales du milieu d'élevage n'ont pas été déterminées au préalable.
En fait, r n'est approché dans la nature que dans de rares cas (colonisation d'un milieu neuf, par exemple), et pour un temps limité, l'un des exemples classiques étant celui de l'expansion de la population de lapins après leur introduction en Australie. En général, même pour des conditions écologiques idéales et sans prédation, les contraintes du milieu (nourriture, espace disponible) deviennent rapidement limitantes au fur et à mesure que la population croit en nombre. On aboutit ainsi à la courbe logistique dans laquelle on fait intervenir le concept de K, charge écologique maximale, valeur limite de la possibilité de croissance numérique.
Ces deux modèles ne sont pleinement applicables que sous certaines conditions (stabilité de la structure de population avec des taux de recrutement et de mortalité constants) rarement réalisées dans la nature. En
180

particulier, ils sont inutilisables quand l'effectif et la structure des populations fluctuent en raison de la succession de cohortes ou du fait d'une prédation sélective ou non.
On arrive ainsi au troisième type de courbe de croissance dans laquelle les oscillations numériques résultent des variations des taux de natalité b ("birth rate") et de mortalité d ("death rate"). Le taux de croissance r de la population est positif ou négatif selon l'importance respective de b et de d . Alors que r représente la capacité maximale théorique (calculée sous conditions expérimentales optimales), le calcul de r permet de quantifier le phénomène réellement observé dans la nature en tenant compte de l'impact des contraintes de 1'environnement et des interrelations avec les autres orga-nismes.
Les notion de r et de r constituent un cadre commode pour analyser la Biologie des Populations car leur calcul intègre plusieurs éléments relatifs aux tables de vie ou au cycle biologique des espèces, dont chacun, considéré isolément, peut réagir à l'action d'un facteur du milieu dans un sens plus ou moins favorable au succès de la population. Les principaux paramètres intervenant dans le taux de croissance des populations sont : - la fécondité (1 ), - la mortalité au cours du développement embryonnaire ou larvaire (m ), - la rapidité du cycle biologique, et en particulier la séquence temporelle du phénomène de ponte (âge d'apparition de la maturité et durée de la période fertile des feraelles.
Le calcul de r tient corapte de ces différents éléments en fonction du temps x selon l'équaïion de base
£ 1 m e-rmXdX- 1 1 et m sont calculés d'après des données d'observations journa-
lières d'élevages de lots d'individus ayant commencé leur développeraent en même temps.
Cette procédure a été utilisée chez les copépodes calanoides Diaptomus (Gehrs et Robertson, 1975) mais elle a été surtout développée chez les harpacticoides, principalement ceux du genre Tisbe (cf. Parise et Lazzaretto, 1966; Gaudy et Guérin, 1977; Bergmans, 1981, 1984).
La fécondité, considérée au niveau populationnel, dépend en premier lieu de la proportion de femelles: le sex-ratio est en général voisin de 1, mais des écarts à cette règle ne spnt pas rares, d'autant plus que la salinité, la température ou même la nature de la nourriture peuvent affecter ces taux (Gaudy et al., 1982). La fertilité des femelles peut présenter des variations individuelles: chez les copépodes du genre Tisbe, par exemple, 5 à 10% des femelles demeurent stériles. Par ailleurs, l'âge des femelles joue un rôle sur l'abondance des pontes journalières, les plus prolifiques se situant chez les copépodes dans le premier tiers de la vie adulte, alors que les dernières pontes produites par les femelles sénescentes ne produisent généraleraent que peu d'oeufs (Gaudy et Guérin, 1977).
La mortalité s'exerce à partir du stade oeuf; chez Tisbe holothuriae, jusqu'à 20% d'oeufs n'éclosent pas (Gaudy et Guérin, 1977). Par la suite, la courbe de mortalité affecte en général une forme caractéristique. Chez les entomostracés, la mortalité larvaire, d'abord très forte chez les jeunes stades, diminue ensuite progressivement jusqu'à des valeurs très réduites au début de la vie adulte, avant de s'intensifier à nouveau lors de la sénes-cence (Gehrs et Robertson, 1975; Bergmans, 1981) .
En dehors de leurs variations intrinsèques en fonction de l'âge, la fécondité et la mortalité dépendent en outre des facteurs du milieu. Chez les copépodes, le taux de production d'oeufs apparait directement lié à l'abondance de la ressource trophique: expérimentalement, on observe le plus souvent une proportionnalité directe entre la ration alimentaire ingérée et la production d'oeufs. La température agit essentiellement sur la cinétique des processus physiologiques, et donc sur la vitesse du cycle biologique, ceci dans les limites des tolérances spécifiques, saisonnières ou géogra-phiques (cf. revue de Vernberg et Vernberg, 1972 ). Elle semble agir égale-ment, dans une certaine mesure, sur la production d'oeufs et sur le sex-
181

ratio. Sur le plan démographique, l'augmentation de la température aboutit à un accroissement du taux de croissance instantanée de la population . Cet accroissement peut être continu dans toute la gamme des températures testées (Heip et Smol, 1976) ou présenter un maximum ou un plateau ayant probable-ment une signification écophysiologique (adaptation de la population à un optimum thermique) comme chez le copépode Eurytemora hirundoides (Castel et al., 1983).
La salinité influe également sur la dynamique des populations selon le degré de tolérance des espèces ou de certains de leurs stades de développe-ment, plus sensibles. Lazzaretto Colombera (1970), montre par, exemple une réduction de la survie, de la fertilité et du taux net de reproduction de Tisbe en milieu dessalé, la modification observée étant plus ou moins ac-cusée selon l'origine géographique de la souche utilisée, ce qui semble indiquer un probable mécanisme génétique agissant sur les processus de régulation physiologique.
Dans la nature, les facteurs du milieu agissent rarement isolément. II est donc utile d'analyser expérimentalement l'effet simultané de plusieurs facteurs. Nous avons par exemple considéré, chez Tisbe holothuriae, l'impact des variations conjuguées de température et de salinité sur la démographie de populations élevées à partir de deux sources alimentaires différentes (Gaudy et al., 1982). La longévité, le temps moyen de génération, le nombre de descendants, le sex-ratio et les valeurs de r sont modifiées selon les combinaisons de facteurs considérés. Le taux de croissance maximal se situe à la température la plus élevée (24°C) et dans les conditions de salinité (38°/
00) les plus proches de celles du milieu naturel habituel. En traçant
les courbes d'iso-valeurs de r (en pourcentage du r maximal), on constate que le potentiel d'expansion 9e l'espèce se réduit au fur et à mesure que les conditions physico-chimiques s'éloignent de l'optimum, mais la nature de la nourriture joue aussi un rôle : avec la Germaline, aliment dérivé de céréales, le r est proportionnellement moins affecté par les variations de facteurs qu'avec le Renutryl, préparation polyvitaminée à usage médical. Le r indique ainsi, au niveau populationnel des différences dans le degré de sïénothermie-eurythermie des populations en fonction du type de nourriture disponible. Ceci laisse entrevoir le degré de complexité des interrelations pouvant, dans la nature, agir sur le potentiel d'expansion des populations de différents dèmes.
Une étape intermédiaire entre les études démographiques basées sur la filiation d'individus isolés et celles traitant de la Biologie des Popula-tions dans la nature consiste à étudier les fluctuations des paramètres de dynamique dans des populations de masse élevées sous conditions, au moins partiellement, contrôlées. Ce type d'approche permet d'analyser les mécanis-mes régulant le maintien des populations face aux contraintes intrinsèques ou extrinsèques dues à la limitation de l'espace, par exemple, l'effet du surpeuplement (avec ses conséquences liées à 1'accumulation des métabolites et, au plan génétique, à l'inbreeding), ou l'impact d'une exploitation régulière, simulant une prédation non sélective.
Dans ce type d'élevage, les variations de sex-ratio et de structure de la population jouent un rôle important dans la régulation quantitative des populations. Katona (1970) indique, par exemple, que le sex-ratio de deux espèces de copépodes du genre Eurytemora diffère selon la densité d'orga-nismes dans l'élevage. De même, Heinle (1970) montre sur des cultures d'Acartia tonsa et d'Eurytemora affinis que la densité, la structure de la population et le sex-ratio varient en fonction du taux journalier d'exploi-tation, et avance l'existence de deux mécanismes homéostatiques de régula-tion: le cannibalisme, plus élevé quand la population est nombreuse (rareté des nauplii) et, dans le cas contraire, 1'augmentation du taux de femelles (avec pour corollaire celui du taux de reproduction), ces femelles semblant provenir de mâles génotypiques se transformant en femelles phénotypiques au cours de leur croissance.
182

Des phénomènes assez semblables se rencontrent également chez les
daphnies (Marshall, 1967) et chez des harpacticoïdes, dont Tisbe holothuriae
(Hoppenheit, 1975, 1976). Nous avons pu ainsi démontrer chez cette dernière
espèce que le nombre d'oeufs par femelles, le taux de femelles ovigères, le
sex-ratio et la production de nauplii fluctuaient en fonction des conditions
d'élevage, avec une tendance à 1'homéostasie indiquée par les relations
inverses entre nombre d'oeufs par sac, sex-ratio et abondance globale (Gaudy
et Guérin, 1982). Le mécanisme de ces régulations est encore mal connu. II
ferait appel à l'effet d'un complexe chimique de nature encore indéfinie,
émis par la population lorsqu'elle atteint des densités élevées (Hoppenheit,
1976; Fava et Crotti, 1979).
Si les études expérimentales sont nécessaires pour analyser l'impact de
facteurs isolés ou peu nombreux sur la démographie, elles ne peuvent rendre
compte de la complexité des situations présentées par l'action simultanée de
l'ensemble des facteurs du milieu physique ou biotique en conditions
naturelles. Les analyses de populations dans leur milieu visent en premier
lieu à décrire leurs fluctuations numériques, à rechercher les facteurs
présentant le plus grand pouvoir explicatif, enfin à tenter de modéliser ces
variations par une approche déterministe ou stochastique. Chez les crustacés zooplanctoniques, on observe généralement, au cours
du cycle annuel, une succession de périodes d'abondance et de pauvreté. Celles-ci peuvent résulter de perturbations hydrodynamiques ou de variations de la pression de prédation, mais elles correspondent souvent à la succes-sion de générations saisonnières plus ou moins bien individualisées. Chez les copépodes, il est possible de discriminer les générations par le suivi
de plusieurs critères: abondance des oeufs par femelle, pourcentage des stades larvaires, variations du sex-ratio (les mâles d'une cohorte détermi-née se développant plus vite et vivant moins longtemps que les femelles),
taux des femelles à spermatophores, taille des adultes reflétant les dif-férentes conditions thermiques ou trophiques saisonnières régnant au cours de leur développement larvaire. On peut, par exemple, distinguer chez les copépodes pélagiques des régions tempérées, la succession de 5 à 7 généra-tions annuelles (Gaudy, 1984).
Ces fluctuations d'abondance peuvent être décrites en termes démogra-phiques par une valeur positive ou négative du taux de croissance instan-tanée de la population, r, celui-ci traduisant l'antagonisme entre la nata-lité (b) et la mortalité (d).
Le taux de natalité des crustacés zooplanctoniques portant leurs oeufs (ou dont on connaît expérimentalement le taux de ponte journalier), est calculé par la méthode du "egg ratio" développée chez les rotifères et
étendue chez les copépodes et les daphnies (Edmonsson et al., 1954; Hall, 1964). On calcule d'abord le taux de natalité finale ("per capita") B =* E/D.l/N où E est le nombre d'oeufs, D la durée du développement embryon-naire et N l'effectif de la population. On en tire le taux de natalité instantanée, b, d'après la formule b — ln (1 + B). Comme r — b - d, il est possible, en déterminant r par comptage des effectifs entre deux prélève-ments successifs de la population, d'en déduire d.
Une méthode récente de modélisation de la population consiste à évaluer directement r par le calcul de b et d (Argentesi et al., 1974; Green, 1976; Castel et Feurtet, 1985). d peut être calculé, non seulement pour la popula-tion dans son ensemble, mais pour ses compartiments d'âge, par exemple, chez les copépodes: oeufs, nauplii, copépodites et adultes. L'effectif de chaque compartiment dépend, en effet, du flux d'individus y entrant ou en sortant et de la mortalité des individus à l'intérieur de cette catégorie. Les différents flux peuvent être alors calculés par un système d'équations différentielles en examinant les effectifs des compartiments à intervalles réguliers, en tenant compte du temps de résidence des individus dans chaque catégorie, évalué en général sur des élevages. L'intérêt de cette méthode est de pouvoir situer à quel niveau d'âge se situe l'impact des prédateurs
183

ou des facteurs de létalité. Dans la nature, en effet, l'action sélective de la prédation semble l'une des principales causes de modification de la structure des populations.
Ce bref examen de quelques aspects de la Biologie des Populations laisse apparaitre que , si les connaissances théoriques en démographie obtenues sur des élevages sont assez développées chez les crustacés (harpacticoides en particulier), nous n'en sommes encore, à l'échelle de la nature, qu'à une étape de description des phénomènes. Beaucoup de mécanismes explicatifs des fluctuations numériques demeurent encore inconnus, en raison de la multiplicité des facteurs et de la latitude d'adaptation écophysiolo-gique des espèces, celle-ci étant en partie contrôlée par la génétique.
GENETIQUE DES POPULATIONS
Comme en ce qui concerne la Biologie des Populations, notre propos n'est pas de fournir un exposé exhaustif des données de la littérature dans le domaine de la Génétique des Populations de Crustacés. L'excellente syn-thèse de Hedgecock et al. (1982) sur la Génétique des Crustacés pourra être consultée à cet effet. Nous nous attacherons plutôt ici à mettre en relief certains aspects de la Génétique des Populations qui conduisent à souligner les relations entre la variabilité génétique des populations et les condi-tions de 1'environnement et montrent, par conséquent, l'importance de la Génétique des Populations dans le développement de la Biologie des Popula-tions.
Chez les Crustacés, la Génétique des Populations a été abordée en considérant trois critères principaux de polymorphisme : la coloration, à l'origine de polychromatisme ; les remaniements chromosomiques, à l'origine du polymorphisme chromosomique ; les enzymes, dont la variabilité est à l'origine du polymorphisme enzymatique.
Du point de vue systématique, ces études ont concerné : - parmi les Entomostracés : des Copépodes (polychromatisme et polymorphisme enzymatique), des Branchiopodes et des Cirripèdes (polymorphisme enzymatique), - parmi les Malacostracés : des Péracarides, Isopodes (polychromatisme, polymorphisme chromosomique et enzymatique) et Amphipodes (polymorphisme enzymatique) et des Eucarides, Euphausiacés et Décapodes (polymorphisme enzymatique) .
La variabilité enzymatique représente le critère le plus étudié, de manière plus ou moins approfondie, chez un grand nombre d'espèces de Crustacés. C'est la raison pour laquelle nous considérerons ici uniquement cet aspect de la Génétique des Populations de Crustacés. A partir de quel-ques exemples, nous essaierons d'analyser les relations qui existent entre l'Ecologie, la Biologie et la Génétique des Populations, de manière à déga-ger les différentes stratégies possibles. Voyons donc comment se présente le poljrmorphisme enzjrmatique chez les Crustacés.
Hedgecock et al. (1982) ont rassemblé les données du polymorphisme biochimique concernant 97 espèces de Crustacés. Ces données indiquent un taux moyen de polymorphisme de 30,5 % et un taux moyen d'hétérozygotie de 7,3 %. Elles font apparaitre un taux d'hétérozygotie plus élevé chez les Copépodes, les Cirripèdes et les Euphausiacés (H > 0,12) que chez les Déca-podes (H < 0,07). Toutefois, au sein d'un groupe taxonomique donné, et en particulier au sein d'un même genre, il existe de grandes différences entre espèces : chez les Copépodes harpacticoïdes des genres Tigriopus et Tisbe, le taux d'hétérozygotie varie du simple au triple (Battaglia et al., 1978a); chez les Cirripèdes, le genre Balanus présente une hétérozygotie moyenne plus élevée que le genre Chthamalus (Hedgecock, 1979) ; chez 4 espèces de krill (Euphausia), l'hétérozygotie varie du simple au quadruple (Valentine et Ayala, 1976 ; Fevolden et Ayala, 1981). Si l'on considère une enzyme donnée, ces différences sont encore plus nettes. Ainsi, la mannose-phosphate-isomérase est polymorphe chez toutes les espèces de Cirripèdes
184

chez lesquelles elle a été testée (11), chez 8 espèces de Penaeides sur 9 ;
par contre, elle n'est polymorphe que chez 4 espèces d'Anomoures sur 11 et
chez une seule espèce de Brachyoure sur 16 (Hedgecock et al., 1982).
Quelles relations peut-on établir entre ce polymorphisme et les
conditions de l'environnement ? Battaglia et al. (1978 a) ont comparé la variabilité génétique de
plusieurs espèces de Crustacés des genres Tisbe, Tigriopus et Gammarus
vivant dans des biotopes marins, saumâtres ou des flaques supralittorales.
La variabilité est la plus élevée dans les populations marines ; elle est la
plus faible dans les flaques supralittorales et intermédiaire dans les eaux
saumâtres. Tigriopus, qui peuple les flaques supralittorales, apparait comme
le genre dans lequel on a mis en évidence l'un des niveaux de variabilité
génétique les plus bas parmi les organismes marins étudiés sous cet aspect.
Les auteurs ont émis l'hypothèse d'une stratégie adaptative fondée sur la
fixation d'allèles capables de conférer à leurs porteurs une plasticité
physiologique élevée. On peut, néanmoins, semble-t-il, s interroger sur le
degré de consanguinité des populations qui peuplent les flaques supralitto-
rales : il pourrait être responsable, au moins en partie, du faible taux
d'hétérozygotie observé. Tisbe holothuriae, étudiée comparativement dans des
biotopes marins et saumâtres (Battaglia et al., 1978 b), est, de toutes les
espèces étudiées par ces auteurs, celle qui présente le plus grand nombre
d'allèles par locus, le plus grand nombre de locus polymorphes et le plus
haut degré d'hétérozygotie moyenne. Toutefois, la population marine est plus
variable que celle des eaux saumâtres. L'instabilité du milieu saumâtre,
plus que la faible salinité, semble être responsable de cette baisse
d'hétérozygotie. Cependant, des résultats inverses ont été obtenus en étudiant des
Cirripèdes : l'hétérozygotie moyenne est 2 fois plus élevée chez Chthamalus
montagui (0,179) qui se rencontre à des niveaux tidaux plus exposés, que
chez C. stellatus (0,087) qui se trouve à des niveaux tidaux plus bas (Dando
et al., 1979). Considérant, d'une part, ces disparités de comportement génétique en
fonction de 1'instabilité des conditions physiques et chimiques de l'envi-
ronnement et , d'autre part, les taux de variabilité génétique de divers
invertébrés marins, Ayala et al. (1975), Valentine et Ayala (1978) et Ayala
et Valentine (1979) mettent l'accent sur le rôle des ressources trophiques
dans cette variabilité. Ils proposent un modèle trophique selon lequel la
variabilité temporelle (le caractère saisonnier) des ressources trophiques
serait un facteur de sélection en faveur des allèles les plus "flexibles".
La variabilité temporelle des ressources trophiques serait donc corrélée à
des niveaux plus bas d'hétérozygotie que les milieux trophiquement stables
qui favoriseraient le maintien d'allèles spécialisés. Cette conception re-
joint celle de Battaglia et al. (1978 a et b), mais le facteur de variabi-
lité est différent. Considérant le pélagos, ils prévoient ainsi une diminu-
tion du taux d'hétérozygotie pour des latitudes croissantes, ce qu'ils
observent effectivement pour 4 espèces de krill (Valentine et Ayala, 1976 ;
Fevolden et Ayala, 1981): Euphausia distinguenda, espèce intertropicale,
présente l'hétérozygotie la plus grande (0,213), E. mucronata, en zone
tempérée, présente une hétérozygotie intermédiaire (0,141), alors que dans
l'océan arctique, 1'hétérozygotie est seulement de 0,091 chez E.
crystallorophias et de 0,057 chez E. superba. II convient toutefois de remarquer que le régime alimentaire de ces
espèces est différent, Euphausia superba étant phytoplanctonophage. Or,
la diversité du régime alimentaire est souvent à l'origine d'une adaptation
des enzymes digestives (Van Wormhoudt et al., 1980) ; celles-ci peuvent présenter des variations saisonnières (Van Wormhoudt et al., 1973 ; Barnes et Blackstock, 1975). Des cultures de Tisbe holothuriae sur différents régimes alimentaires montrent que cette adaptation met en jeu des isoenzymes (Guérin et Kerambrun, 1982), ce qui s'observe également dans l'adaptation
185

d'Acartia clausi et de Centropages typicus à la pollution urbaine (Rivière
et Kerambrun, 1983). Des variations cycliques de certains alloenzymes sont
connues dans des populations naturelles de Drosophila pseudoobscura et D.
persimilis (Dobzhansky et Ayala, 1973). Chez Balanus balanoïdes, des dif-
férences dans le polymorphisme d'une estérase ont été décrites en fonction
du niveau de fixation sur le rivage et des saisons (Flowerdew et Crisp,
1976). Chez Asellus aquaticus, le génotype de l'amylase apparait corrélé
avec la nourriture ou le choix de l'habitat (Christensen, 1977) ; de grandes
variations des fréquences de l'allèle Amy-2 s'observent selon les popula-
tions. Gooch et Hetrick (1979) observent des faits analogues pour 3 locus
enzymatiques chez Gammarus minus et Borowsky (1984) montre, chez 3 espèces
de Gammares, l'existence de 2 zones d'activité, Amy-1 et Amy-2, tandis que
certains individus présentent une troisième bande, Amy-X. Expérimentalement,
Amy-X est induite spécifiquement par un régime riche en amylose ou glyco-
gène et par abaissement de la température d'élevage. L'auteur conclue que
l'expression de Amy-X et Amy-1 varie selon la saison et constitue une répon-
se adaptative à la variation saisonnière du régime alimentaire. Par ail-
leurs, chez Gammarus palustris, espèce dont le régime alimentaire est varié,
les préférences alimentaires diffèrent selon le génotype (Borowsky et al.,
1985).
II apparait donc nécessaire, pour comprendre les stratégies mises en
jeu, de prendre en considération, d'une part, le rôle métabolique des
enzymes étudiées et, d'autre part, l'éthologie alimentaire des espèces.
En ce qui concerne le rôle des enzymes dans le métabolisme, les enzymes
couramment testées par électrophorèse peuvent être classées en 2 groupes
dont le niveau d'hétérozygotie est différent dans les populations de Droso-
philes (Gillespie et Kojima, 1968 ; Kojima et al., 1970). La distinction se
situe entre les enzymes métabolisant le glucose (Groupe I) et les autres
(Groupe II). Les enzymes du groupe I, à spécificité restreinte, sont beau-
coup moins variables que les enzymes du groupe II, susceptibles d'agir sur
différents substrats offerts par 1'environnement. D'où l'hypothèse d'une
stratégie d'adaptation mettant en jeu une hétérozygotie plus grande des
enzymes du groupe II dans les milieux variables.
Chez les Crustacés, néanmoins, cette différence est beaucoup moins
nette que chez la Drosophile. Mais il faut souligner que la comparaison
n'est pas toujours aisée du fait du choix des enzymes testées par les différents auteurs qui est assez variable. Ainsi, les différences d'hétérozygotie décrites par Dando et al. (1979) entre 2 espèces de
Chthamalus en fonction du niveau tidal concernent 13 enzymes du groupe I,
alors que la variation en fonction de 1'environnement décrite par Nevo et
al. (1977, 1978) chez Balanus amphitrite porte presque exclusivement sur des
enzymes du groupe II.
Chez les Décapodes, Nelson et Hedgecock (1980) ont étudié 44 espèces et
comparé 51 espèces en prenant en considération divers paramètres de l'envi-
ronnement et du comportement (trophique notamment). Ils font ainsi app-
araitre une corrélation entre l'hétérozygotie des enzymes du groupe I et la
variation environnementale, tandis que l'hétérozygotie des enzymes du groupe
II est corrélée avec le généralisme trophique. Les chiffres fournis par ces
auteurs indiquent des pourcentages de corrélations positives qui diffèrent
de manière très hautement significative selon le groupe d'enzymes. D'une
manière générale, ils observent une corrélation positive entre le taux moyen
d'hétérozygotie et l'instabilité des ressources trophiques (et la latitude),
ce qui va à l'encontre de l'hypothèse de Ayala et Valentine et est en
contradiction avec le comportement génétique du krill. Néanmoins, les va-
leurs de l'hétérozygotie moyenne ne rendent pas compte de celles de
l'hétérozygotie des enzymes des groupes I et II qui peuvent varier en sens
inverse. En effet, la variation de l'hétérozygotie moyenne en fonction de la
variabilité environnementale rend bien compte du comportement des enzymes du
186

groupe I, dont elle tend plutôt à minimiser les variations, mais ne rend
absolument pas compte des valeurs de 1'hétérozygotie des enzymes du groupe
II. Ainsi, chez les Décapodes, les taux relatifs d'hétérozygotie observés
pour les enzymes des groupes I et II ne semblent pas dépendre de l'instabi-
lité des ressources trophiques, mais du comportement trophique des espèces.
Considérant le régime alimentaire des espèces étudiées, Nelson et Hedgecock
montrent, d'une part, que l'hétérozygotie moyenne la plus faible s'observe
chez les phytophages qui présentent les taux d'hétérozygotie les plus bas à
la fois pour les enzymes des groupes I et II, tandis que, d'autre part, les suspensivores et les détritivores ont les taux d'hétérozygotie les plus élevés. C'est pourquoi, ils proposent un modèle de stratégie tenant compte à
la fois de la variabilité environnementale et des caractéristiques trophi-ques des espèces.
CONCLUSION
Les Crustacés offrent une immense diversité : dans l'échelle des tailles, dans la durée de vie, les modalités et le temps de développement, le mode de vie (pélagiques, benthiques, fixés, parasites), le régime trophique (phytophages, carnivores, omnivores, suspensivores, détritivores),... Cette diversité soulève un certain nombre de problèmes qui se posent à la fois en Biologie des Populations et en Génétique des Popula-tions.
La Blologie des Populations doit évidemment prendre en compte cette diversité et considérer les caractéristiques propres à chaque espèce. L'étude expérimentale implique la réalisation de cultures dans des condi-tions physiques, chimiques et nutritionnelles compatibles avec l'accomplis-sement satisfaisant du cycle biologique des individus de l'espèce concernée. C'est là une première difficulté que connaissent bien tous ceux qui se heurtent au délicat problème des élevages. Les paramètres physiques et chimiques sont généralement assez bien maîtrisés pour les espèces marines ou d'eau douce, mais gardent un caractère arbitraire en ce qui concerne les espèces saumâtres pour lesquelles la variabilité du milieu qui leur est propre peut difficilement être reproduite en laboratoire. Par contre, le choix de tel ou tel régime alimentaire revêt un aspect beaucoup plus délicat qui peut conduire à des résultats dont l'incidence peut être très différente chez des espèces pourtant faciles à élever. De plus, les conditions expéri-mentales (espace limité, nourriture qualitativement peu variée,...) peuvent ne pas correspondre au comportement trophique naturel des espèces (qui ne sont plus contraintes de rechercher leur nourriture). Or, la détermination des potentialités démographiques maximales, qui constitue l'un des buts de la Biologie des Populations, ne peut se faire que dans des conditions physico-chimiques et nutritionnelles optimales. Leur reconstitution expéri-mentale représente une seconde difficulté et n'est réalisable que pour un nombre très limité d'espèces : d'une manière générale, les plus tolérantes et les moins spécialisées au plan trophique.
En ce qui concerne la Génétique des Populations, l'un des objectifs majeurs est de parvenir à une meilleure compréhension des mécanismes de maintien du polymorphisme des populations naturelles et des relations entre le polymorphisme et les conditions de l'environnement. La Génétique des Populations de Crustacés est donc, elle aussi, liée à leur diversité, en particulier en ce qul concerne le mode de vie. La variabilité génétique des populations est influencée par les paramètres physiques et chimiques du milieu, leur fluctuation temporelle et spatiale et par le comportement trophique des espèces. Ces facteurs de sélection, qui contrôlent la structure génétique des populations, influent aussi sur les paramètres démographiques. La Génétique des Populations apparait ainsi étroitement liée à la Biologie des Populations dont les aspects dynamiques peuvent être considérablement perturbés non seulement par une baisse du polymorphisme
187

(consanguinité), mais aussi par les modifications des conditions écologi-
ques. Cette liaison est conditionnée par l'Ecologie qui contrôle, d'une
part, le développement démographique des populations, et d'autre part, leur
diversité génétique. Cette dernière est essentielle à l'adaptation des
populations à l'hétérogénéité de l'environnement, mais aussi au maintien des
performances physiologiques et démographiques. C'est pourquoi, par exemple
en ce qui concerne les Décapodes d'intérêt économique, Dynamique et Géné-
tique des Populations sont complémentaires pour envisager une meilleure
exploitation des stocks. Le maintien de cette diversité constitue un pro-
blème majeur qui se pose à l'expérimentateur : les cultures à partir d'un
faible échantillon conduisent inévitablement à une baisse de polymorphisme
et à une dérive par rapport aux populations naturelles. Cet aspect prend
toute son importance en aquiculture où la consanguinité est inévitable et où
le contrôle de la variabilité génétique apparait nécessaire.
Mais il convient d'évoquer un dernier problème, tout à fait fondamental
en Biologie des Populations et en Génétique des Populations, qui est celui
de la perception de la variabilité spatio-temporelle de leur environnement
par les organismes des différentes espèces. Cette perception est
certainement différente selon le mode de vie (benthique, pélagique, fixé),
les caractéristiques écologiques (latitude, proximité du rivage,
euryhalinité, ...) et l'éthologie alimentaire des espèces. Dans le cadre de
leurs réponses écophysiologiques, diverses adaptations non génétiques ont
été décrites ; il ne faut peut être pas en exclure trop définitivement toute
composante génétique.
Les quelques aspects de la Biologie des Populations et de la Génétique
des Populations qui ont été exposés permettent d'envisager un certain nombre
de stratégies : par exemple, variations du sex-ratio, cannibalisme, compor-
tement trophique conduisant à un optimum de reproduction, variations du taux
d'hétérozygotie, variabilité des différentes enzymes en fonction du degré de
stabilité de 1'environnement et du comportement alimentaire. Nous rappelle-
rons seulement pour mémoire les stratégies r et k bien connues pour être
adoptées la première en milieu instable, la seconde en milieu stable (Bar-
bault, 1983). Ces diverses stratégies, dont résultent, pour chaque popula-
tion, une certaine structure et une certaine organisation dans le temps et
dans l'espace, font clairement ressortir l'importance des données de la
Génétique des Populations dans le développement de la Biologie des Popula-
tions. L'immense diversité des Crustacés offre un choix quasi-illimité de
possibilités et fait de ce groupe systématique un matériel de choix pour
appréhender le fonctionnement des populations naturelles.
ARGENTESI F., DE BERNARDI R., DI COIA G., 1974. Mathematical models for the
analysis of populations dynamics in species with continuous recruitment,
Mem. Ist. ital. Idrobiol., 31, 245-275.
AYALA F.J., VALENTINE J.W., 1979. Genetic variability in the pelagic envi-
ronnement : A paradox ?, Ecology, 60, 24-29.
AYALA F.J., VALENTINE J.W., ZUMWALT G.S., 1975. An electrophoretic study of
the antarctic zooplankter Euphausia superba, Limnol. Oceanogr. , 20, 4, 635-
640. BARBAULT R., 1983. Ecologie générale, Masson éd., Paris.
BARNES H., BLACKSTOCK J., 1975. Seasonal changes on the activity of certains
enzymes in crude extracts of two common cirripedes, Balanus balanoides (L.)
and B. balanus (L.), J. exp. mar. Biol. Ecol., 19, 81-96.
BATTAGLIA B., BISOL P.M., FAVA, G., 1978 a. Genetic variability in relation
to the environment in some marine invertebrates, in : Marine organisms,
Genetics, Ecology and Evolution, édité par B. Battaglia et J.A. Beardmore,
Plenum, New York, 53-70. BATTAGLIA B., BISOL P.M., VAROTTO V., 1978 b. Variabilité génétique dans des
188

populations raarines et lagunaires de Tisbe holothuriae (Copepoda,
Harpacticoïda), Arch. Zool. exp. gén., 119, 251-264.
BERGMANS M., 1981. A demographic study of the life of Tisbe furcata (Baird
1837) (Copepoda, Harpacticoïda). J. mar. Biol. Ass. U.K., 61, 691-705.
B0R0WSKY R., 1984. Environmental control of amylase phenotype in Amphipods
of the genus Gammarus, Biol. Bull., 167, 647-657.
BOROWSKY R., B0R0WSKY B., MILANI H., GREENBERG P., 1985. Amylase variation
in the salt marsh Amphipod, Gammarus palustris, Genetics, 111, 311-323.
CASTEL J., CANTIES C., POLI J.M., 1983. Dynamique du copépode Eurytemora
hirundoides dans l'estuaire de la Gironde : effet de la température,
Oceanologica Acta, N. SP., Proceedings 17th European marine Biology Sympo-
sium, Brest, France, 57-61. CASTEL J., FEURTET A., 1985. Dynamique du copépode Eurytemora hirundoides
dans l'estuaire de la Gironde : utilisation d'un modèle à compartiments. J.
Rech. Océanogr., 10, 4, 134-136. CHRISTENSEN B., 1977. Habitat preference among amylase genotypes in Asellus
aquaticus (Isopoda, Crustacea)., Hereditas, 87, 21-26.
DANDO P.R., SOUTHWARD A.J., CRISP D.J., 1979. Enzyme variation in Chthamalus
stellatus and C. montagui (Crustacea, Cirripeda). Evidence for the presence
of C. montagui in the Adriatic, J. Mar. Biol. Ass. U.K., 59, 307-320.
DOBZHANSKY T., AYALA F., 1973. Temporal frequency changes of enzyme and
chromosomal polymorphisms in natural populations of Drosophila, Proc. Natl
Acad. Sci. USA, 70, 3, 680-683. EDMONSON W.T., COMITA G.F., ANDERSON G.C., 1954. Reproductive rate of
copepods in nature and its relation to phytoplankton populations, Ecology,
43, 625-634. FAVA G., CROTTIE., 1979, Effect of crowding on nauplii production during
mating time in Tisbe clodiensis and T. holothuriae (Copepoda, Harpacticoi-
da), Helgolànder wiss. Meeresunters, 32, 466-475. FEVOLDEN S.E., AYALA F.J., 1981. Enzyme polymorphism in Antarctic krill
(Euphausiacea); genetic variation between populations and species, Sarsia,
66, 167-181. FLOWERDEW M.W., CRISP D.J., 1976. Allelic esterase isozymes, their variation
with season, position on the shore and stage of development in the Cirripeda
Balanus balanoides, Mar. Biol., 35, 319-325. GAUDY R. , 1984. Biological cycle of Centropages typicus in the northwestern
Mediterranean neritic waters, Crustaceana, suppl. 7, 199-213.
GAUDY R., GUERIN J.P., 1977. Dynamique des populations de Tisbe holothuriae
(Crustacea : Copepoda) en élevage sur trois régimes artificiels différents,
Mar. Biol., 39, 137-145. GAUDY R., GUERIN J.P., 1982. Population dynamics of Tisbe holothuriae
(Copepoda : Harpacticoida) in exploited mass cultures, Neth. J. Sea Res., 16, 208-216. GAUDY R., GUERIN J.P., MORAITOU-APOSTOLOPOULOU M., 1982. Effect of tempera-ture and salinity on the populations dynamics of Tisbe holothuriae Humes
(Copepoda : Harpacticoida) fed on two different diets, J. exp. Mar. Biol.
Ecol., 57, 257-271. GEHRS C.W., ROBERTSON A., 1975. Use of life tables in analysing the dynamics
of Copepod populations, Ecology, 56, 665-672. GILLESPIE J.H., K0JIMA K., 1968. The degree of polymorphism in enzymes
involved in energy production compared to that in nonspecific enzymes in two Drosophila ananassae populations, Proc. Natl. Acad. Sci. USA, 61, 582-585.
GOOCH J.L., HETRICK S.W., 1979. The relation of genetic structure to
environmental structure : Gammarus minus in a karst area, Evolution, 33, 1, 192-206. GREEN J.D., 1976. Population dynamics and production of the calanoïd copepod Calamoecia lucasi in a northern New Zealand lake, Arch. Hydrobiol., (suppl.) 50, 313-400. GUERIN J.P., KERAMBRUN P., 1982. Effects of diet on esterases, alkaline phosphatase, malate dehydrogenase and phosphoglucomutase activity observed
189

by polyacrylamide gel electrophoresis in Tisbe holothuriae (Harpacticoïd Copepod), Comp. Biochem. Physiol., 73 B, 4, 761-770. HALL D.J., 1964. An experimental approach to the dynamics of a natural population of Daphnia galeata mendotae, Ecology, 45, 94-112. HEDGECOCK D., 1979. Biochemical genetic variation and evidence of speciation in Chthamalus barnacles of the tropical eastern Pacific, Har. Biol., 54, 207-214. HEDGECOCK D., TRACEY M.L., NELSON K., 1982. Genetics, in : The Biology of Crustacea, édité par L.G. Abele, Acad. Press, New York, vol.2, 283-403. HEINLE D.R., 1970. Population dynamics of exploited cultures of Calanoïd Copepods, Helgolânder wiss. Meeresunters, 20, 366-372. HEY C., SMOL N., 1976. Influence of temperature on the reproductive potential of two brackish-water harpacticoïds (Crustacea : Copepoda), Mar. Biol., 35, 327-334. HOPFENHEIT M., 1975. Zur Dynamik exploitierten Population von Tisbe holothuriae (Copepoda : Harpacticoïda), II Populationdichte, Alterzusammen-setzung, Wachstum und Ausbente, Helgolànder wiss. Meeresunters, 27, 377-395. HOPFENHEIT M. , 1976. Zur Dynamik exploitierten Population von Tisbe holothuriae (Copepoda : Harpacticoïda), III Reproduktion, Geschlechtsver-hâltnis, Entwicklungsdauer und Überlebenszeit, Helgoländer wiss. Meeres-unters, 28, 109-137. KATONA S.K., 1970. Growth characteristics of the copepod Eurytemora affinis and E. herdmani in laboratory cultures, Helgolànder wiss. Meeresunters, 20, 373-384. K0JIMA K., GILLESPIE J., TOBARI Y.N., 1970. A profile of Drosphila species enzymes assayed by electrophoresis. I Number of alleles, heterozygosities, and linkage disequilibrium in glucose-metabolizing systems and some other ensymes, Biochem. genet., 4, 627-637. LAZARETTO-COLOMBERA, 1970. Effetti della diluzione su alcuni parametri di popolazione in razze geografiche del copepode Tisbe holothuriae Humes (Har-pacticoïda), Archo. Oceanogr. Limnol., 16, 263-274. MARSHALL J.B., 1967. Radiation stress in exploited Daphnia population, Limnol. Oceanogr., 12, 154-158. NELSON K., HEDGECOCK D., 1980. Enzyme polymorphism and adaptive strategy in the Decapod Crustacea, Am. Nat., 116, 2, 238-280. NEVO E., SHIMONY T., LIBNI M., 1977. Thermal selection of allozyme polymor-phisms in barnacles, Nature, (London), 267, 699-701. NEVO E., SHIMONY T., LIBNI M., 1978. Pollution selection of allozyme poly-morphisms in barnacles, Experientia, 34, 1562-1564. PARISE A., LAZZARETTO I., 1966. Misure di popolazione sul copepode Tisbe furcata Baird (Harpacticoïda), Mem. Accad. Patavina Sci. Let. Atti, 79, 3-11. RIVIERE D., KERAMBRUN P., 1983. Impact d'une pollution d'origine urbaine sur les activités enzymatiques de deux copépodes planctoniques (Acartia clausi et Centropages typicus), Mar. Biol., 75, 25-35. VALENTINE J.W., AYALA F.J., 1976. Genetic variability in krill, Proc. Natl Acad. Sci. USA, 73, 2, 658-660. VALENTINE J.W., AYALA F.J., 1978. Adaptative strategies in the sea, in : Marine organisms, Genetics, Ecology and Evolution, édité par B. Battaglia et J.A. Beardmore, Plenum, New York, 323-345. VAN WORMHOUDT A., CECCALDI H.J., MARTIN B.J., 1980. Adaptation de la teneur en enzymes digestives de l' hépatopancréas de Palaemon serratus (Crustacea, Decapoda) à la composition d'aliments expérimentaux, Aquaculture, 21, 63-78. VAN WORMHOUDT A., CECCALDI H.J., LE GAL Y., 1973. Variations aux différentes saisons des activités enzymatiques digestives chez Palaemon serratus ayant subi l'ablation des pédoncules oculaires, C. R. Acad. Sc. Paris, 277, 369-372. VERNBERG W.B., VERNBERG F.J., 1972. Environmental physiology of marine animals, Springer Verlag, Berlin, 346 p.
190

ASPECTS RÉCENTS de la Biologie des CRUSTACÉS Actes de Colloques - 8 -1988 - IFREMER, pages 191 à 194
ETUDE Dü POLYMORPHISME ENZYMATIQUE DANS CINQ POPULATIONS NATURELLES D'
ARMADILLIDIUM VULGARE Latreille (Crustacea Isopoda Oniscidea).
RAIMON R.
RESUME - L'hétérozygotie moyenne (11,4 %)observée chez Armadillidium vulgare est plus élevée que chez les autres crustacés aquatiques (4-6 %). Ces diffé-rences de structure génétique seraient plus en rapport avec la biogéographie de l'espèce qu'avec la diversité des modes de détermination du sexe.
Mots clés: Polymorphisme enzymatique, population, Armadillidium vulgare, Crustacés.
ABSTRACTS - The mean heterozygoty observed (11,4 %) in ‘Armadillidium vulgare appears to be higher than in other aquatic crustaceans (4,6 %). These diffe-rences in the genetic structure could be related to the biogeography of the species and not with the diversity in the modalities of sex determination.
Key words : Enzymatic polymorphism, population, Armadillidium vulgare, Crustaceans.
Chez les Crustacés Isopodes terrestres quelques travaux ont été consacrés à l'étude du polymorphisme enzymatique (Sassaman 1978, Beck et Price 1981 et 1985, Garthwaite et Sassaman 1985, Sassaman et Garthwaite 1983, Legrand et al. 1986). Le but de notre étude a été de comparer les populations européennes d'Armadilli-dium vulgare aux populations américaines étudiées par Beck et Price 1981 ; nous avons également examiné s'il existaitunê relation entre le polymorphisme enzyma-tique et le mode de détermination du sexe chez cette espèce cosmopolite.
En effet l'existence de populations où s'observent des phénomènes de monogé-nie (descendances unisexuées) (Vandel 1941, Juchault et Legrand 1986) a conduit Howard (1942) a émettre l'hypothèse que l'inbreeding serait plus faible dans ces populations que dans les populations où les descendances offrent un sex ratio normal (descendances amphogènes). Cette hypothèse a cependant été critiquée par Johnson (1977).
Tous les animaux étudiés bnt été prélevés dans la nature au sein de cinq populations. Selon le mode de détermination du sexe, on peut distinguer 3 types de femelles fonctionnelles : des femelles génétiques (type ZW) dont les descendances sont amphogènes, des néo-femelles ZZF, (= mâles féminisés par une bactérie symbiotique F) amphogènes, des néo-femelles ZZf. (= mâles génétiques féminisés par un facteur f) (Legrand et Juchault 1984) dont les descendances sont monogènes. Ces femelles sont en proportions variables dans les cinq populations étudiées.
St. Georges les Baillargeaux (Vienne) (nombreuses femelles ZZf et quelques femelles ZZF.)
Santec (Finistère) (femelles ZZf et ZW) Sète (Hérault) (femelles ZZf) Northampton (Angleterre) (femelles ZZf.) Mostaganem (Algérie) (femelles ZW)
(1) Laboratoire de Biologie Animale de l'Université de Poitiers et UA CNRS n° 673 "Physiologie et Génétique des Crustacés", 40, Avenue du Recteur Pineau, 86022 POITIERS CEDEX, France.
191

TABLEAU 1 : FREQUENCE DES ALLELES ET HETEROZYGOTIES
St.-Georges Sète Santec North. Mosta.
PGH 100 0,71 0,45 0,63 0,37* 0,98
102 0,29 0,55 0,67 0,63 0,02
H1 0.41 0.49 0,46 0,46 0,03
GOT 96 0,20 0,32 0,41 0,32 0,25
100 0,80 0,68 0,59 0,68 0,75
H1 0.03 0.03 0,08 0,08 0,38
ODH 92 0 0 0 0 0,29
96 0 0 0 0 0,12
98 0,02 0,02 0,05 0,05 0
100 0,98 0,98 0,95 0,95 0,59
H1 0.03 0,03 0.08 0.08 0,55
PH.Ac 92 0 0 0 0 0,05
96 0 0 0 0 0,17
100 0,98 0,80 1,00 0,96 0,63
104 0,02 0,20 0 0,04 0,14
108 0 0 0 0 0,01
H1 0.03 0,31 / 0,07 0,55
A03
98 0,06 0 0,02 0,14* 0,04
100 0,94 1,00 0,98 0,86 0,96
H1 0.12 / 0.04 0.23 0,07
AO4
96 0,02 / 0,05 0,02* 0,01
100 0,98 1,00 0,95 0,98 0,99
H1 0.04 / 0.09 0.02 0.02
LAP 100 1,00 1,00 1,00 1,00 0,78
102 0 0 0 0 H1 0,22
H 0,08 0,11 0,10 0,11 0,17
(*) valeurs s'écartant significativement des proportions
théoriques
192

Nous avons employé la technique d'électrophorèse verticaie sur gel de
polyacrylamide. Les enzymes ont été révélées selon la technique de Harns (1976).
7 énzymes ont été étudiées : représentant 11 systèmes : LDH, GOT1
, GOT2
, AO1
,
A02, A03, A04, PGM, ODH, LAP, Phos. ac.
Résultats
Le Tableau 1 montre les fréquences des allèles pour les 7 systèmes qui sont
polymorphes. La loi de Hardy-Weinberg a été vérifiée pour chaque système et à
quelques rares exceptions les proportions réelles ne diffèrent pas significative
ment des proportions théoriques. L'hétérozygotie moyenne est de 11,4 %, mais si
l'on considère les 4 populations les plus homogènes et géographiquement regrou-
pées (St.-Georges, Sète, Santec et Northamptol'hétérozygotie moyenne est
alors de 10 %. Le dendrogramme construit à partir des distances de NEI (fig. 1)
montre l'isolement de la population de Mostaganem.
North.
Sète
Santec
St. Georges
Mostaganem
Distances de NEI
Figure 1
Discussion
L'hétérozygotie moyenne de.11,4 % est voisine de celle observée chez les
invertébrés (13 à 15 %). Chez les crustacés aquatiques, différents travaux
montrent des hétérozygoties très variables : 4,7 % sur 7 espèces d’ecrevisses
(Nemeth 1979), 4 % chez le Homard (Hedgecock et al. 1977) et 5,8 % chez 11 autres
espèces de Décapodes (Hedgecock et al. 1976). Armadillidium vulgare offre par
contre une hétérozygotie plus forte que ces espèces aquatiques ; cependant chez
cette même espèce, Beck et Price (1981) ont trouvé une hétérozygotie de 3 %. Ces
auteurs expliquent ce très faible taux par l'effet de fondation : Armadilhdium_
vulgare ayant pour lieu d'origine le pourtour méditerranéen, l'espece aurait ete
introduite par l’homme aux Etats-ünis et se serait alors répandue à partir d'un
faible nombre de fondateurs. Le chiffre de 10 % pour les populations europeennes
(et de 17 % à Mostaganem) pourraient être en rapport avec l'ancienneté de ces
populations. Si 1'on admet avec Howard que l’inbreeding est plus faible dans
ces popula-
tions à descendances monogènes, on peut penser que 1 hétérozygotie y est donc
plus élevée que dans une population où les descendances sont amphogènes. Les
résultats se rapportant à la population de Mostaganem ne confirment pas cette
hypothèse. Par ailleurs, une population de type amphogène (Danemark) non étudiée
ici, est très proche des 4 populations européennes (monogènes) aussi bien pour le
niveau d'hétérozygotie que pour les fréquences alléliques des systèmes étudiés.
Dans l'état préliminaire de nos recherches il semble que les différences de
structure génétique soient plus en rapport avec la biogéographie qu avec la
diversité des modes de détermination du sexe.
193

Beck M.L. et Price J.O., 1981. Genetic variation in the terrestrial isopod Armadillidium vulgare. J. Hered. T2, 15-18. Garthwaite R.L. et Sassaman C., 1985.Porcellionides floria, new species from north america ; provinciality in the cosmopolitan isopod Poroellionides pruino-sus ('Brandtj. J. Crust. Biol. 5 (3), 539-555-Harris H. et Hopkinson D.A., 1976. Kand book of enzyme electrophoresis in human genetics, Horth-holland publishinc company, Amsterdnm : Oxford american Elsevier publishing company, INC, New York. Hedgecock D., Shleser R.A. et Helson K., 1976. Applications of biochemical genetics to aquaculture. Proceedings of the XIII pacific Science congress. J. Fish. Res. Boad Can. 33, 1108-1119. Hedgecock D., Nelson K., Simons J. et Shleser R., 1977. Genic simularity of Ame-rican and European species of the lobster Homarus. Piol. Bull. , 152, M-50.
Howard H.W., 19L2. The genetic of Armadillidiutn vulgare Latreille. II. Studies on the inheritance of monogeny and amphogeny. J. Genetics 44, 143-159.
Johnson C., 1977. Evolution of sex ratios in the isopod, Venezilla evergladensis,
Evolution, 31, n° 3, 603-610. _ ... Juchault P. et Legrand J.J., 1986. Monogeny in Armadillidium vulgare Latreille
(Crustacea Isopoda Oniscidea). Advanoes in Invertebrates Reproductuon U, Elsevier
Sc. Publ. BV, Porchet et al. ed., U17-U2U. . . . Legrand J.J. et Juchault P., 1984. Nouvelles données sur le determimsme geneti-
que et épigénétique de la monogénie chez le crustacé isopode terrestre Armadilli
dium vulgare Latr.. Génét. Evol. 16, 1.57-84.
Nemeth S.T. et Tracey M.L., 1979. Allozyme variability and relatedness m six
crayfish species. J. Hered., 70, 37-U3. . . Sassaman C. et Garthwaite R.L., 1985. Independent assortment of four loci in
the
isopod, Porcellio dilatatus. J. Hered. 76, U70-U71. . Sassaman C., 1978. Dynamics of a lactate dehydrogenase polymorphismin
the wood
lanse Porcellio scaber Latr. : Evidence for partial assortative mating and hete-
rosis in natural populations. Genetics 88, 591-609. .
Vandel A., 19U1. Recherches sur la génétique et la sexualite des Isopodes terres-
tres. VI. Les phénomènes de monogénie chez Les Oniscoïdes. Bull. Biol. Fr. Beig.,
75, 3, 316-363.
194

ASPECTS RÉCENTS de la Biologie des CRUSTACÉS Acles de Colloques - 8 - 1988 - IFREMER, pages 195 à 198
LESAOESDE Niphargusvirei(CRUSTACE STYGOBIE).
par M.J.TURQUIN-BUFFIERE (1)
RFSIJME- On détermine l'âge absolu de l'amphipode stygobie Niphargus virei par sa discrémination à partir d’histogrammes annuels de taille; il est validé par les valeurs obtenues lors de récoltes printanières, non plus estivales, et par la croissance observée en élevage. La croissance pondérale est utilisée pour confirmer le taux daccroissement très faible de cette espèce. L’âge des populations subit des fluctuations corrélées au facteur "pluviométrie" qui agit, par le biais de la chasse d'une partie du stock hors des biotopes souterrains, à la façon de Veffort de pêche" des populations exploitées.
mots-clès: Niphargus v/rei, longévité, mortalité, taux d'accroissement, âge.
ABSTRACT- The life-span of Niphargus virei is estimated with the length histogramms of annual summer samples; it fits in with the data of rare spring distributions and with the growth observed in the laboratory. Increase in body wet weight confirms the low growth rate of this species (maximum: 0.06 mg. day-1 ); the mean age of populations changes on account of rainfall, which determines the drift of part of the population out of its natural habitat.
key-words: Niphargus virei, longevity, mortality, growth-rate, age.
I- INTRODUCTION
Niphargus virei, crustacé stygobie, possède des caractéristiques uniques pour un amphipode d'eau douce puisque certains individus ont été maintenus en vie durant 14 ans. Vivant dans un milteu où les saisons sont excessivement atténuées, cet anlmal ne subit pas d'anecdysis hivernel, et constitue un bon matériel pour étudier les modalités de sa croissance.
2- L'AGE BIOLOGIQUE
Ginet( 1960,p.208. ) a montré que de la première à la cinquième année le nombre des mues passe de 5,5± 1,1 par an à 1,2± 0,3; ce rythme très lent de Niphargus virei par rapport aux gammaridés épigés ne retentit pas sur le nombre de mues nécessaires à atteindre la maturité aexuelle: 9 à 13 pour le premier, 8 à 14 pour les seconds. Chez les individus âgés ie durée de l'intermue doit dépasser un an et plus si l'allongement observé de la naissance à 5 ans se poursuit.
3- L'AGE ABSOLU
a) croissance linéaire
L'exploitation de la dérive lors des crues à l'exutoire de Drom, puis ctens d'autres sites
U.A. CNRS 367 Ecologie des Eaux Douces Equipe d'Hydrobiologie et Ecologie Souterraines. Université LYON 1,43 Boulevard du 11 Novembre, 69622 Villeurbanne cédex, France
195

permet de tracer des histogrammes de taille. Le choix du pas de taille est très important pour l’exploitation correcte de ces histogrammes, nous utilisons des classes de 0,1 ou 0,2 mm de péréïopode 7. La ponte saisonnière induit une vague annuelle de recrutement se traduisant par un pic d'abondance sur les histogrammes. Les méthodes classigues de discrimination de Vâge ont permis de transformer les hlstogrammes plurimodaux en pyramides des âges. La loi de croissance de Niphargus virei, selon la formule de Von Bertalanffy, s'écrit donc.
LP7t(min.an-i)= 6,5 ( 1-e-0.097(t+0.l934) ) La validité des âges- au moins pour les premières années- est corroborée par la croissance
linéaireen élevageet par Vétude d'histogrammesobtenus entredeux périodes de recrutement c'est-à-direoù les animaux ont 6 mois, 1,5 an, 2,5 ans, etc.
b) croissance pondérale
•méthode Sutcliffe et al ( 1 981 ) ont pu élever de leur naissance à la mort des lots de gammere dressant
ainst la courbe de crolssance ( logistique) masse/temps. Ce travail est impossible pour Niphargus à cause de sa trop grande longévité. C'est pourquoi, j'ai mis en élevage des animaux prélevés dans le milieu naturel de taille aussi variée que possible. La relation taille, masse fraiche, a été établieMmg=6,73.P7mm2.43 . r=0,97
Les Niphargus dont une partie de la croissance s'est effectuée dans la nature, sont alors pesés tous les qulnze jours ; certains subissent 5 muesen un an; d'autres, les plus grands, donc les plus âgés n'ont pas mué après 450 joursà la température de 10±0,5*C.
Figurel: Taux d'accroissement moyen absolu (ML2-MH) de lots de Niphargus de taille croissante calculé sur une année, en mg par jour.
•résultats - taux d'accroissement
La figure 1 montre le taux d'accroissement absolu en fonction de la masse fraiche de l'amphlpode. La grande incertitude pour les animsux de plus de 100 mg est dûe au falble nombre des mues, donc au manque relatif de données, mais aussi à la diversité des accroissements individuels. Quoiqu'il en soit 1e taux d'accroissement décroit avec la masse. Comparé au gammare (0,7mg à la naissance jusqu'à 55-60mg, durée de vie 350-400 jours, taux d'accroissement absolu meximum: 0,5mg. jour-1), Niphargus devient 10 fois plus gros, mais avec un taux d'accroissement maximum de 0,06 mg.jour-1: il faudrait au minimum 8 500 jours, soit plus de 23 ans à Niphargus pour atteindre 1e poids de 500 mg.
196

- courbe de croissance la croissance pondérale de lots de masse variée , etudiée pendant 450 jours a permis d élaborer
une série de courbes de croissance de la forme (tableaul).
)a figure 2 regroupe, en les raccordant à leur point de jonction, 5 de ces courbes. I) n'a pas été
possible d'estimer la croissance au delà de 188 mg faute d'exuviation des animaux de cet âge
Momg k Cs t jours
4,5 0,0044 0,44
0
15,5 320 0,0027 0,27
23,5 0,0018 0,18
470
47 0,08
843 0,0008
130 2070 0,0003 0,03
200
Tableau 1: Expressions exponentielles de la croissance de Niphargus au début de sa vie: Mt=Mo.e Mo. masse au début d'une période t„ k: taux de croissance instantané ou spécifique; Cs: taux instantané en % de la masse humide par jour; les observations commencent pour des animaux de 4 mg ( t=0, différent de la naissance).
• conciusion Ce poids correspond à 26,30 mm de longueur du corps et 4,60mm de P7 : la lecture de la
courbe age/ taille (Turquin, 1984) indique que ces animaux sont des 8+ Or la courbe de
croissance en poids donne 8 ans et demi pour passer de 4 à 188 mg auquels il faut ajouter plusieurs mois grandir de la naissance à 4 mg . 11 existe donc une assez bonne corrélation entre ces
deux méthodes d'évaluation de l'âge, totalement différente dans leur principe
Figure 2:
Croissance pondérale de Niphargus virei d'après le tableau t
4- L'AOE ECOLOGIQUE
a) méthode
Les histogrammes annuels de taille sont transformêsen pyramides des ages, pyramides oü 80% des individus peuvent avoir moins de un an. Seules les cinq premières classes sont utilisées dans
les calculs ultérieurs des mortalités. En effet la courbe des prises est toujours rectiligne pour ces
cinq classes, dénotant un bon échantillonnage, une mortalité constante. et la validité de la discrimination de l'âge.
• mortalité totale La structure démographique d'une population naturelle suivie depuis quinze ans a subi 2
transltions démographiques (Turquin,l984). Le coefficient inst8ntanédemortal1té totale est
corrélé au facteur " pluviométrie", ce dernier agissant par le biais des crues et du drift à la façon
de l'"effort de pêche " des populations exploitées.
197

• âge moyen de la population En conséquence, en période d années sèches I age moyen de la population , représentée par une
pyramide è base étroite, peut atteindre 10 ans; après quelques annnées pluvleuses en été, l'ège moyen diminue à 2-3 ans, c'est-à-dire en dessous de l'âge à la première reproduction (v’ers 4 ans) On conçoit que certains systèmes karstiques soient impropres è la vie de cette espèce du simple fait de leur fonctionnement hydrologique (Turquin.l 987).
5 CONCLUSION Niphargus virei fait preuve d' une longévité remarquable, longévité pressentie pour d'autres péracarides ( Magniez, 1976) et prouvée pour les décapodes stygobies américains (Cooper, 1975) La saisonnalité des pontes, surprenante dans ce milieu souvent qualifié de stable au regard des biotopes aquatiques superficiels, s'explique par la nécessité du repeuplement en altitude et à l'amont du drainage du réseau karstique, repeuplement compensant l'égouttement estival des conduits aidé par le fort rhéotropisme de Niphargus. Plus qu'è la qualité des eaux, la survie de l'espèce est lièe à la structure géologique et au fonctionnement hydrologique du massif calcaire: si Niphargus viret abonde dans les karsts de plateaux où l'eau chemine pendant plusieurs decennies depuis la surface jusqu'à l'exutoire , il est absent en Chartreuse, dans )e Vercors où prédominent les circulations verticales rapides.
COOPEfT J.E.,1975. Ecdogtcal and behaMoral sludies in Shelta Cave, /4abama, with emphais on Decapod Crustaceans PhilosophyDissertation, Kentucky Univ. GINET R,1960- Ecologie, Ethologie et Biologie de Niphargus (Amphipode, Gemmaridé hypogél Ann Spéléol, 24, 387-397.
MAGNIEZ G.,1976. Contribution àlaconnaisaance de la biologie des Stenascellidae {Qvsteceaisopoât Asetoia de3 eaux souterrames) Thèse Doc. ès Sc Nat , A.O. 10;238,Dijon ; 1-228. SUTCUFFE D W ,CAHfaCK TJFt. à L.G. WILLOUGHBY,1981. Effects of diet, bodysize, ageand temperature on grovàhratesintheamphipod Gammarvsputex .fresinealerBiaiogy\ 1,183-214. TURQUM U.J., 1981- Profil démographique et enwormement chez une population de Niphargus nrei (Amphipode trogfobée). Bull. Soc.Zoli. de Fer. 106,4,457-466. TURQUIN M.J.,1984. Un cas de transition démographique dans le mieu souterram Yerh mtemai ferein. Limnol, 22,1751-1754. TURQUIN BUFFIERE M -J ,1986. Factors controing poplation size of tvo species of perermM ampNpoda ( cannibalsm, or floods dedmafion). in: .Adunees in mvertetnie Reprodudion Y édité par M.Porchet, J-C. Andries, et A. Dhainaut, Ets evier, 548. TURQUM-BUFFIBIÉ M.-J.,1987. Les crues, dans Técosystème souterrain, sont-eHes neutres (ressources ou perturbafion) ? But. mens Soc. Lim. Oe Lyon, accepté le 21 octobre 1986.
198

ASPECTS RÉCENTS de la Biologie des CRUSTACÉS Actes de Colloques - 8 - 1988 - IFREMER, pages 199 à 202
COMPLEMENT A LA DEFINITION " IDOTEA CHELIPES . ESPECE POLYTYPIQUE" A PARTIR DES FRACTIONS PROTEIQUES ET PARTICULIEREMENT DE L'HEMOCYANINE. COMPARAISON AVEC IDOTEA BALTHICA BASTERI .
CHARFI-CHEIKHROUHA F-
RESUME - Les électrophorégrammes des protéines sur gel de polyacrylamide à gradient de concentration (2,5 - 15 %) ont permis de mettre en évidence 24 fractions protéiques chez l'espèce polytypique I. chelipes dont 4, à activité plus intense, correspondent à l'hémocyanine. Une nette ressemblance est observée au niveau des protéinogrammes des diverses sous-espèces et populations d' I. chelipes , alors que ceux d' I.b. basteri semblent bien différents. Une différence est également notée entre les sexes au sein. de la même population. Elle touche la fraction la plus lente dont l'activité est plus intense chez les femelles.
mots-clés . Idotea chelipes chelipes , Idotea chelipes bocqueti , Idotea chelipes mediterranea . Idotea balthica basteri , Tunisie, Mediterranée , Atlantique. protéinogramme . hémocyanine.
ABSTRACT -Electrophoregrams of proteins on polyacrylamid gel concentrated at 2.5 - 15% made it possible to separate 24 protein fractions in polytypic I. chelipes species, 4 of which having a higher activity and corresponding to hemocyanin. A clear ressemblance is evident in proteinograms of various sub-species and in I. chelipes pqpplations whereas those of I.b. basteri seem to be quite different. A variation also exists between sexes within the same population. It corresponds to the slowest fraction whose activity is intenser with females.
Key-words : Idotea chelipes chelipes , Idotea chelipes bocqueti , Idotea chelipes mediterranea , Idotea bacthica basteri , Tunisia, Mediterranean , Atlantic, proteinogram , hemocyanin.
INTRODUCTION
Les protéines, présentant soit une structure multimoléculaire soit une activité physiologique particulière ou bien ces 2 propriétés à la fois, constituent un matériel de choix pour les recherches moléculaires à but taxinomique. Ainsi nous avons choisi d'étudier 1’hémocyanine qui présente une nette hétérogénéité selon les espèces. En effet, plusieurs auteurs dont Fine et coll. (1975), Maguire et Fielder (1975), Sevilla (1977) et Laulier (1984) ont mis en évidence la valeur spécifique de cette protéine chez les Crustacés. Dans le même ordre d'idées, nous avons essayé les techniques électrophorétiques sur Idotea chelipes et Idotea bocqueti des côtes
(1) Laboratoire de Biologie Animale, Faculté des Sciences de Tunis, Campus Universitaire, 1060 Tunis Belvédère, Tunisie.
199

Fig. 1 Lieux de récolte d' Idotea chelipess. I. ch. chelipes : raarais salans du Croisic ; bassin d’Arcachon.
I. ch. mediterranea : étangs français de Gruissan, Leucate et Canet ; lagunes tuni-siennes de l'Ichkeul, Bizerte, Char-El-Melh et Tunis.
* I. ch. bocnacU : Ksibet-El-Mediouni ; mer de Boughrara. I. bolthica bosteri en syrapatrie avec I .ch.mediterranea dans le lac de Bizerte.
Fig. 2
Protéinogrammes totaux après électrophorèse sur gel de poly-acrylamide à gradient linéaire de concentration 2,5-15%. A. photo du gel : 1, I.b.basteri <£ 2-3, I .ch.mediterranea ♂ et ♀ ; 4, I.ch.chelipes ♀ ; S.l.ch. bocquett J . B. Schéraa du gel : 1,1.b. basteri ♀ ; 2, I.chelipes ♀ . H: fractions hémocyaniques L(lentes), I(inter-médiaires) et R(rapides). V:pro-téinc spécifique femelle.
200

tunisiennes , considérées jusqu'alors par nos études classiques comme étant 2 sous-espèces de la même espèce polytypique Idotea chelipes (Charfi-Cheikhrouha , 1980). Notre étude a été par la suite étendue à d'autres populations méditerranéennes et atlantiques des côtes françaises et à une autre espèce bien distincte I. balthica représentée par sa forme I.b. basteri .
MATERIEL ET METHODES
La répartition des stations de récolte est représentée sur la figure 1. Les échantillons sont formés de surnageant provenant de la centrifugation à 17.300 g et à 4°C pendant 10' de broyats de 5 individus dans 500 ul de saccharose 40 X et de tampon de migration tris-glycine à pH 8.6 dans les proportions 1/1. Afin d'éviter les variations dues au cycle de mue, à la congélation et au rythme circadien, nous avons opéré des animaux frais au stade C d'intermue et toujours à la même heure.
L'électrophorèse est réalisée sur un gel d'acrylamide de 1 mra d'épaisseur. Le gel de séparation, formé d'un gradient linéaire 2,5 - 15%, est surmonté d'un gel de concentration à 2,5 % d'acrylamide. La migration est effectuge à 4°C pendant 4 h sous une tension de 100 V et une intensité de 35 mA dans un tampon tris-glycine â pH 8.6 dans les proportions 0,6 g et 2,88 g/1. Quelques gouttes de bleu de bromophénol à 1 % additionnées à ce tampon, marquent le front de migration.
Les fractions protéiques sont détectées par leur coloration au bleu de Coomassie R 250 (Sigma). Quant aux fractions protéiques correspondantes à l'hémocyanine, elles sont révélées par leur activité spécifique : révélation péroxydasique selon la méthode de Shrauwen et réaction à l'acide rubéanique détectant la présence de cuivre (Horn et Kerr, 1969).
RESULTATS
Sur les 24 bandes protéiques Jo'bservées sur le protéinogramme d' I. chelipes (Fig. 2), 4 sont de nature cuprique et à activité péroxydasique ; elles correspondent donc à l'hémocyanine. La plus lente, mais la plus concentrée, paraît formée de 2 fractions juxtoposées de mobilités relatives MR égales à 34 et 35. Elle constitue l'hémocyanine principale HL .. Les 2 autres fraction, de même intensité mais de mobilités relatives différentes, constituent respectivement 1'hémocyanine intermédiaire HI à MR = 55 et 1'hémocyanine rapide HR à MR =67. Ce profil électrophorétique, observé chez I. chelipes , est identique à celui d' I. bocqueti . Quant à celui d' I. balthica , il présente également les 4 fractions hémocyaniques de même aspect que celles observées chez I. chelipes mais avec des mobilités relatives plus élevées H'L (MR = 35 et 36), H'I (MR = 57) et H'R (MR = 69). 1
Concernant 1'hémocyanine, il n'existe pas de dimorphisme sexuel apparent sur le gel. Cependant, nous remarquons, pour les différentes espèces étudiées, la présence d'une bande protéique très lente, de nature cuprique et qui paraît beaucoup plus intense chez les femelles.
DISCUSSION ET CONCLUSION
L'existence d'au moins 3 fractions hémocyaniques sur le gel de polyacrylamide à gradient de concentration a été déjà retrouvée chez certains Isopodes (Sevilla, 1977 ; Picaud, 1983). Nos observations coïncident bien avec celles de Kaim-Malka et coll.(1983) sur I. bacthica basteri . Nos
201

résultats, sur la structure biochimique et le poids moléculaire de ces 3 fractions, feront l'objet d'une note ultérieure. La protéine la plus lente, se retrouvant en forte proportion chez les femelles et en très faible dose chez les mâles, semble avoir une origine extraovarienne et constitue selon Souty et Picaud (1981) la vitellogénine synthétisée au niveau du tissu adipeux.
Nous avons remarqué qu' I. balthica est génétiquement isolée, aussi bien dans la nature qu'au laboratoire, d' I. bocqueti et des 2 formes atlantique et méditerranéenne d' I. chelipes . Elle s'éloigne également par des différences morphologiques notables des 3 entités déjà citées. Très proches sur le plan morphologique, ces entités ont des exigences écologiques différentes et ne présentent pas de barrière génétique au laboratoire. Ainsi des hybrides, obenus entre I. bocqueti et I. chelipes de nos côtes, sont féconds, ont une croissance intermédiaire entre celle des parents et présentent même une vigueur hybride. Ceux, provenant de la forme atlantique avec bocqueti et chelipes de la Méditerranée, sont morts, par excès de chaleur au laboratoire, avant la puberté.
Les résultats protéiques illustrent bien la valeur taxinomique spécifique de l'hémocyanine. En effet, I. balthica se distingue des 3 formes d' I. chelipes par ses fractions hémocyaniques plus rapides. Quant au protéinogramme d' I. chelipes , quelle que soit l'origine des populations étudiées, il présente un profil similaire avec des hémocyanines analogues. Cette stabilité structurale de l'hémocyanine, longtemps considérée comme une protéine spécifique, ne nous permet pas d'élever au rang d'espèce les 3 entités étudiées et nous porte donc à les considérer comme 3 sous-espèces : Idotea chelipes chelipes , Idotea chelipes bocqueti et Idotea chelipes mediterranea , appartenant à une même espèce polytypique Idotea chelipes .
Remerciements : Nous remercions vivement J.P. Labourg pour l'envoi des échantillons.
CHARFI-CHEIKHROUHA F. 1980. Recherches syscémaciques, biologiques eC expérimencales sur les Idotées de Tunisie (Isopodes Valvifères). Thëse 3ême cycle Unlv. Tunis , 193 p. FINE J.M. , MARNEÜX M. , LAMBIN P. , ROCHU D. , BARON J.C. et CHIDÀLIA W. 1975. Structure imraunochimique de 1'hémocyanine. Etude chez dix espèces de Crustacés Décapodes. Cah. Biol. Mar. , 16, 246-253. HORN E.C., KERR M.S., 1969. The hemolymph proteins of the blue crab Callinectes sapidus I. Hemocyanins and certain other major protein constituents. Comp. Biochem. Physiol. , 29, 493-508. KAIM-MALKA R.A., MUNOZ P., CECCALDI H.J. 1983. Electrophoresis study of heraolymph proteins of Idotea balthica basteri (Crustacea : Isopoda). Comp. Biochem. Physiol. , 74 B, 433-440. LAULIER M. 1984. Valeur taxonomique spécifique des sous-unités hémocyaniques chez cinq espèces du genre Sphaeroma (Isopodes Flabellifères). Bull. Soc. Zool. France, 109, 4, 365-376. MAGUIRE G.B. , FIELDER D.R. 1975. Disc electrophoresis of the haeraolymph protelns of sorae Portunid crabs (Decapoda : Portunidae) II. Physiologica1 and taxonoraic aspects. Comp. Biochem. Physiol. , 52 A, 43-47. PICAUD J.L. 1983. Les protéines spécifiques femelles des Crustacés Isopodes. Constitution, synthèse et certains aspects du contrôle de leur synthèse et de leur libération. Thèse Doct. Sci. Poitiers, 214 p.
SEVILLÀ C. 1977. Les hémocaynines chez les Oniscoïdes. Dissociation sous l'influence du pH et données structurales. Arch. int. Physiol. Biochem. , 85, 125-131. SOUTY C. , PICAUD J.L. 1981. Vitellogenin synthesis in the fat body of the marine Crustacean Isopoda, Idotea balthica basteri , during vitellogenesis. Reprod. Nutr. Develop. , 21, 95-101.
202

ASPECTS RÉCENTS de la Biologie des CRUSTACÉS Actes de Colloques - 8 - 1988 - IFREMER, pages 203 à 206
REPRODUCTION ET CROISSANCE CHEZ LES FEMELLES DU CRUSTACE ONISCOIDE ARMADILLIDIUM VULGARE Latr. APRES INOCULATION D'UNE BACTERIE SYMBIOTIQUE FEMINISANTE.
JUCHAULT P.(1) MOCQUARD J.P. (1)
RESUME - La présence dans certaines populations d'Armadillidium vulgare d'une bactérie symbiotique (bactérie F) intracytoplasmique est transmissible aux descendants par les ovocytes plasmiques. Ce micro-organisme induit alors une féminisation des mâles qui évoluent en néo-femelles fonctionnelles.
Le rôle de la bactérie F sur l'entrée en reproduction, la croissance et la fertilité des femelles asymbiotiq.ues a été étudié. Une baisse de la ferti-lité, ralentissant l'apparition des néo-femelles est observée. Ce processus pourrait faciliter la mise en place par sélection de mécanismes destinés à lutter contre la féminisation de tous les individus contaminés.
Mots-clés : reproduction, croissance, sex-ratio, bactéries féminisante, Armadillidium, crustacés.
ABSTRACT - In some populations of Armadillidium vulgare a symbiotic bacteria (F bacteria) is transmitted to the progency via the ovocytes and induces a feminization of the males.
The effect of the F bacteria on the begining of the reproduction, the fertility of the non-symbiotic females has been studied. A drop in fertility is observed. This process- could be related to the selection of mechanisms counteracting the feminization of all the contaminated males.
Key-words : reproduction, sex-ratio, F bacteria, Armadiilidium, crustacea.
Dans certaines populations d'Armadillidium vulgare, on observe des femelles dont les organes hébergent une petite bactérie symbiotique intracytoplasmique qui semble appartenir au groupe des Chlamydiales. Transmis aux descendants par les ovocytes, ce microorganisme (F) féminise les mâles qui évoluent alors en néo-femelles fonctionnelles. Tout individu porteur de cette bactérie devient réfractaire à l'action masculinisante de l'hormone androgène (Juchault et Legrand, 1985). II est possible d'introduire expérimentalement cette bactérie chez une femelle génétique (femelles asymbiotique, de type WZ) : au lieu d'engendrer des descendances à sex ratio normal, les femelles génétiques inoculées donnent naissance à des descendances comportant quelques mâles et de nombreuses femelles symbiotiques (femeiles génétiques WZF et néo-femelles ZZp) (Legrand et Juchault, 1970). Dans certaines populations, les femelles génétiques n'existent plus et tous les individus sont donc des mâles génétiques ZZ. Dans de tels cas, on observe, à côté des néo-femelles ZZ„ , d'autres néo-femelles ZZ dont la féminisation est le résultat de l'action d'un autre facteur (f) plus instable et incapable de s'opposer chez un adulte à l'action masculini-sante de l'hormone androgène (Juchault et Legrand, 1986). Avant d'élaborer un modèle susceptible d'expliquer la mise en place et les évolutions possibles des
(1) Laboratoire de Physiologie et Génétique des Crustacés, L'A CNRS 673, Université de Poitiers, 86022 POITIERS Cedex, Francc.
203

divers mécanismes du déterminisme du sexe dans les populations dM, vulgare, nous avons pensé qu'il était nécessaire d'acquérir diverses données permettant d'évaluer quantitativement l'impact de certains paramètres agissant directement ou indirectement, sur la reproduction. Dans cette étude, nous avons recherché si la bactérie F pouvait modifier l'entrée en reproduction, la croissance et la fertilité des femelles asymbiotiques.
RESULTATS -
1. Cas des femelles (néo-femelles ZZF. et ZZF.) récoltées dans la nature (Niort).
Aucune différence n'a été observée entre les néo-femelles ZZ (20 ? ) et-ZZ (52 ?) en ce qui concerne la date d'entrée en reproduction, la fertilité et laf
croissance pendant les 250 jours d'élevage au laboratoire.
2. Cas des femelles génétiques WZ et des néo-femelles ZZ. , après inoculation de la bactérie F. 1
Les résultats expérimentaux (tableau) montrent que l'entrée en reproduction des deux catégories de femelle n'est pas modifiée par la présence de la bacté-rie. Par contre, le taux de croissance des femelles génétiques inoculées est presque deux fois plus faible que celui des femelles génétiques témoins. Cet effect ne s'observe pas chez les néo-femelles ZZ . La diminution du taux de croissance des femelles génétiques inoculées agit indirectement sur la fertilité car cette dernière est fonction de la masse des génitrices.
Valencia : WZ (Espagne)
1 Rabat : ZZ (Maroc)
Bactérie + - + -
Masse initiale en mg
n = 19 n = 20 86,4 14,2 91,013,8
F = 0,66
n = 17 n = 19 94,6 15,5 97,2 15,6
F = 0,11 Délai entrée en 114,6± 5,6 117,5 ±3,7 96,4± 5,2 97,0± 4,5 | reproduction,en jours F = 0,19 F = 0,01
Taux de croissance mf - mi 10000 tc = : X mi d
32,91 3,2 58,11 4,6 F = 19,48*
57,414,3 57,516,1 F = 0,0
Fertilité Y = nombre de
descendants
Y = 74,9 Y = 107,3 X = 119,5 X = 150,9
Y = 62,9 Y = 68,3 X = 146,7 X = 149,0
X = masse lors de la ponte
Y = 0,758 X - 11,1 Y = 0,156 X + 42,6 |
Tableau : Entrée en reproduction, croissance et fertilité chsz des fcmelles génétiques WZ (populaticn de Valence, Espagne) et des néo-fcmelles ZZf. (population de Rabat, Maroc) après inoculation de la bactérie fcminisante (+) ; carparaison avec des femelles témoins WZ et ZZ indames (-). Le délai d'entrée en reproduction a été calculé à partir dj 1er janvier ; taux de croissance tc : mi = masse initiale, mf = masse finale, d = délai entre mi et mf.
204

DISCUSSION
Comme cela a été proposé pour les bactéries symbiotiques d'Insectes, on peut penser que la bactérie féminisante dM. vulgare dérive d 'une forme qui, à l'origine, a pu avoir un effet pathogène. Un tel effet a d'ailleurs été observé chez certaines espèces d'Oniscoïdes après inoculation de cette bactérie (Juchault et al. , 1974). Dans les populations d'/l, vulgare, on peut donc considérer la présence d'individus hébergeant des bactéries comme le résultat d'une sélection ayant favorisé les génotypes permettant 1'établissement de cette association. En fait, cet état d'équilibre pourrait être envisagé comme le résultat d'une co-évolution de l'hôte et du microorganisme. L'hypothèse que les néo-femelles ZZf. dériveraient des femelles ZZF. (le facteur f corres-pondant à une partie de l'ADN bactérien) ayant été proposée (Juchault et Legrand, 1986) on peut penser que les souches à facteur f ont conservé cette adaptation à la bactérie F ; 1'introduction expérimentale de ce microorganisme n'aurait donc aucun effet pathogène. Par contre il n'en est pas de même chez des femelles génétiques dont, vraisemblablement, les ascendants n'ont jamais été en contact avec cette bactérie. Chez ces individus, le ralentissement de la croissance après inoculation de la bactérie peut être attribué à un effet pathogène. On peut donc penser que l'arrivée de la bactérie F dans une population indemne puisse entrainer dans un premier temps une diminution de la vitalité - et donc de la fertilité - chez les femelles contaminées, que cette contamination intervienne par voie traumatique ou génétique. Cette baisse de la fertilité, en ralentissant dans un premier temps l'apparition des néo—femelles, pourrait faciliter la mise en place, par sélection, de mécanismes destinés à lutter confere la féminisation de tous les individus dans une population contaminée. De tels mécanismes ont été mis en évidence dans des populations où la bactérie est présente ; l'un d'eux consiste en la sélection de gènes masculi-nisants capables de s'opposer à l'action féminisante de cette bactérie (Legrand et Juchault, 1984).
JU31AULT P., IHEAND J.J., MAKTIN G., 1974. Action interspécifique du facteur épigénétique féminisant responsable de la thélygénie et de 1 ' intersexualité du crustacé Armadillidim vulgare (ïsqpode Oiiscoïde), Am. Ehbr. Marph., 7, 265-276. JICHAULT P., LEtîiAND J.J., 1985. Contribution à l'étude du mécaniane de l'état réfractaire à l^hormone androgène chez les Anradillidùm vulgare Latr. (Crustacé, Iscpode, Oiiscoîde) hébergeant un bactéroïde féninisant, Gen. Ccnp. Enfocrmdl., 60, 463-467. JUCHAULT P., IEGRAND J.J., 1986. Ihermosensibilité de l'expressicn ou de la transmission d'un facteur fénimsant responsable de certaines formes de monogénie chez le Crustacé Oniscoïde ArtmdïUlidim vulgare, Gén. Sél. Evol., 18, 393-404. LEGRAND J.J., JICHAULT P., 1970. Modification expérimentale de la proportion des sexes chez les Crustacés Isopodes terrestres : induction de la thélygénie chez Armadillidùm vulqare Latr, C.R. Acxd. Sa. Fr., 270, 706-706. LEGRAND J.J., JUCHAULT P., 1984. Nouvelles données sur le déterminisme génétique et épigénétique de la monogénie chez le Crustacé isopode terrestre Anradillidiun vulgare Latr., Génét Sél Evol., 16, 57-84.
205

ASPECTS RÉCENTS de la Biologie des CRUSTACÉS Actes de Colloques - 8 - 1988 - IFREMER, pages 207 à 216
ACTUALISATION DES DONNEES SUR L’ECOLOGIE, LA BIOGEOGRAPHIE ET LA PHYLOGENIE DES AMPELISCIDAE (CRUSTACES - AMPHIPODES) ATLANTIQUES APRES LA REVISION DES COLLECTIONS D'E. CHEVREUX.
BELLAN-SANTINI D.(*) DAUVIN J.C.(**)
RESUME - Le genre Ampelisca comprend près de 140 espèces dans le monde, il est très largement répemdu dans tous les milieux de 0 à 4000 m ; son intérêt est de trois ordres : phylogénétique, biologique et écologique. L'analyse d'un abondant matériel nouvellement récolté ainsi que la révision d'anciennes collections (E. Chevreux), nous ont conduits, depuis une décennie à décrire 11 nouvelles espèces, ce qui porte à 41 le nombre d'espèces dans l'Atlantique nord-est.
La grande spéciation du genre Ampelisca, ainsi que sa forte homogénéité suggèrent qu'il s'agit d'un taxon en cours de spéciation. Les relations phénétiques mettent en évidence 2 grands groupes d'espèces, l'un très apomorphe, l'autre plus hétérogène.
Plus du tiers des espèces sont endémiques ; plus de la moitié des espèces n'ont été récoltées que dans une seule des 5 provinces considérées.
Du point de vue bathymétrique, 3 espèces sont limitées au plateau continental, 34 ne dépassent pas l'isobathe 1000 m, 4 sont bathyales.
Deux espèces sont inféodées aux substrats durs, 7 sont vasicoles, les autres semblent avoir des exigences écologiques moins strictes.
Les études de biologie ont montré que le cycle de développement d'une espèce est fortement lié à la latitude.
mots-clés : Amphipodes, Ampelisca, Atlantique nord-est, phylogénie, écologie, biogéographie.
ABSTRACT - In the world, the Ampelisca genus is constituted by more than 140 species which are found between 0 to 4000 m depth. Phylogeny, biology émd ecology are the most important interest in this genus. The present analysis of new and older collections (E. Chevreux) have permitted to describe 11 new species. 41 species are listed in the north-eastern Atlantic fauna. These are different arguments to think that Ampelisca groups was actuéilly in speciation. Two groups issued from a phenetic analysis are discriminated by their apomorphic levels. More them of the third are endemic species. 21 species are present only in one of the five biogeographic region. 23 species are limited to the continental shelf, 34 live up to the first 1000 meters and 4 are bathyal. 2 species are strictly distributed on hard-bottoms, 7 are found in muddy bottoms and the other have no typical soft-bottom distribution. The biology of the species is correlated with the latitude.
key-words : Amphipoda, Ampelisca, north-eastern Atlantic, phylogeny, ecology, biogeography.
(*) Station Marine d'Endoume, UA CNRS 41, 13007 Marseille. («*) CNRS LP 4601 et Univ. P. & M. Curie, Paris VI, Station Biologique, 29211 Roscoff.
207

INTRODUCTION
Le genre Ampelisca est un genre de Crustacés Amphipodes parmi les plus vastes, il comprend actuellement plus de 140 espèces décrites dans l'Océan Mondial. Ce genre a une très large distribution mondiale mais un fort taux d'endémisme local ; on récolte des représentants du genre dans tous les milieux de 0 à 4000 m. Certaines espèces représentent des éléments essentiels dans le macrobenthos des mers froides et tempérées où ils peuvent constituer un maillon important dans l'alimentation de certains poissons et mammifères marins. Les espèces les mieux connues du point de vue biologique montrent une dépendance étroite aux facteurs du milieu : température, nature du sédiment.
Une vaste révision du genre, actuellement en cours, a permis pour la Méditerranée et l'Atlantique Nord-est à partir de nombreuses collections actuelles et anciennes de dresser le bilan des espèces présentes dans cette partie du monde : 20 espèces nouvelles ont été décrites et un certain nombre redéfinies.
L'apport de la collection d'E. Chevreux du Museum National d'Histoire Naturelle de Paris a été capital dans cette étude, elle contenait 32 espèces parmi lesquelles 17 avaient été décrites par lui (Dauvin & Bellan-Santini, 1986). Un certain mélange et quelques confusions existent dans les données publiées et auxquelles les auteurs actuels se réfèrent souvent ; nous les avons rectifié et nous avons pu ainsi tracer les limites de nouvelles distributions à la lumière des résultats les plus récents. Cette note réactualise et complète les données mentionnées dans Bellan-Santini & Dauvin (sous presse), d'après nos dernières investigations. On trouvera l'ensemble des références bibliographiques dépouillées dans cet article.
1. ETAT DU GENRE AMPELISCA EN ATLANTIQUE NORD-EST
Dans l'Alantique nord-est dont nous excluons ici la Méditerranée s. str., le recensement global donne 41 espèces (la signalisation d'A. natalensis mentionnée par Reid (1951) n'a pas été considérée comme valable). Sur les 41 espèces que nous avons rencensé, nous en avons retrouvé 31 dans les différentes collections étudiées, parmi les 10 restantes pour lesquelles nous ne possédons que des données puisées dans la littérature, nous en excluons 2 de notre étude ; les diagnoses les concernant étant insuffisantes : A. incerta Reid (1951) et A. jarli Reid (1951), cette dernière étant de plus un mâle.
Notre mise au point concerne donc 39 espèces actuellement bien caractérisées.
2. PHYLOGENIE DU GROUPE
Afin d'essayer de comprendre le sens de l'évolution au sein de ce groupe, nous avons recherché les caractères susceptibles de permettre une analyse phylogénétique. 58 caractères significatifs ont été recherchés, dans les descriptions des espèces ; ces caractères sont reconnus par différents auteurs (Barnard 1960 ; Dickinson 1982) comme possédant une signification phylogénétique chez les Amphipodes. Sur les 58 caractères étudiés, 51 ont été conservés comme significatifs dans le groupe considéré. Une matrice de similarité phénétique rnontre l'existence de deux groupes d'espèces : le premier groupe correspondant à des espèces très apomorphes et le second plus diversifié (Bellan-Santini 8c Dauvin, sous presse). Le calcul du degré d'apomorphie (codage de 40 des caractères considérés en caractère apomorphe 1 ; plésiomorphe 0) exprimé en pourcentage a permis d'effectuer un classement des espèces depuis la plus évoluée A. odontoplax (67,5) jusqu'aux moins évoluées : A. rubella (25) et A^ latifrons (22,5 à 25) (fig. 1).
208

degré d'apomorphie
Figure 1. - Espèces d'Ampelisca classées suivant leur degré d'apomorphie croissant. Pour les espèces insuffisamment décrites, le sous-lignage indique le degré minimum d'apomorphie obtenu à partir de critères mentionnés ; la flèche signale le degré d'apomorphie maximum possible.
Pour certaines espèces dont nous ne possédons que les descriptions fournies dans la littérature (certains caractères étant inconnus) nous avons calculé les valeurs extrêmes du leur degré d'apomorphie, c'est le cas de A. bidentata qui peut être l'espèce la moins évoluée avec un degré d'apomorphie de 20 mais qui peut aussi se retrouver beaucoup plus haut jusqu'à un degré de 40. En tenant compte de ces incertitudes, on remarque que le groupe des 6 espèces les plus évoluées se détache au-delà de 50 : A. odontoplax, A. spooneri, A. eschrichtii, A. macrocephala, A. brevicornis et A. spinifer ; les autres espèces se répartissent ensuite suivant un gradient décroissant assez régulier. On observe que les espèces abyssales présentent des degrés d'apomorphie parmi les plus élevés suggérant une colonisation de ces milieux profonds à partir d'espèces primitives néritiques.
209

Arctique Boréale Lusita-nienne
Mauréta-nienne
Sénéga- Bathyale lienne
A. eschrichtii t.c. t.c. i A. spinifer + r. P.c. ? r. A. spooneri + p.c. p.c. ? r. A. tenuicornis t.c. t.c. 1 r. A. typica t.c. t.c. ? p.c. A. diadema t.c. t.c. ? 1 A. spinipes + t.c. t.c. t.c. r A. brevicornis s.l. t.c. t.c. t.c. t.c. A. aequicornis t.c. p.c. r 1 A. macrocephala t.c. A. pusilla p.c. A. toulemonti + r. r. A. amblyops r. i A. anomala r. r. A. gibba c p.c. i A. lusitanica + r. A. remora + r. A. calypsonis + r. A. massiliensis + i A. pseudosarsi + r A. pseudospinimana + r A. spinimana t.c. ? p.c. A. armoricana + c. ? c. A. rubella + p.c. ? r. A. sarsi + c ? r. A. serricaudata + r i 1 A. latifrons p.c A. hupferi + r. A. heterodactyla + p.c A. ctenopus + p.c A. bidentata + 1 A. palmata 1 A. monoculata + 1 A. senegalensis + r. A. verga + P.c A. abyssicola r. A. uncinata r. A. odontoplax p.c. A. compacta + r.
Tableau 1. Distribution géographique des 39 espèces d'Ampelisca (d'après les limites biogéographiques définies par Fredj, 1974). L'abondance des récoltes est indiquée suivant une cotation en 5 points : 1 : une signalisation ; r : rare, 2 à 4 signalisations ; p.c. : peu commune, 5 à 10 signalisations ; c : commune, 11 à 20 signalisations ; t.c. : très commune, plus de 20 signalisations. ? : absence de données mais présence probable ; + : espèces endémiques de l'Atlantique Nord-Est.
210

A. biàentata Schellenberg, 1925 A. eenegalenoia Chevreux, 1925 A. hupferi Schellenberg, 1925 /. ctcnopua Schellenberg, 1925 A. keterodactyla Schellenberg, 1925 A. lueitanica Bellan-Santini & Marques A. toulemonti Dauvin & Bellan-Santini, 1982 A. verga Reid, 1951 A. eerraticaudata Chevreux, 1888 A. remora Bellan-Santini & Dauvin, 1986 A. armcricana Bellan-Santini & Dauvin, 1981 A. nonoculata Dauvin & Bellan-Santini, 1985 A. latifrons Schellenberg, 1925 A. spooneri Dauvin & Bellan-Santini, 1982 A. sarsi Chevreux, 1887 A. pseudcspinimana Bellan-Santini & Kaïm-Malka, 1977 A. spinifer Reid, 1951 A. pseudosarsi Bellan-Santini & Kaïm-Maika, 1977 A. rubella Costa, 1864 A. macrocephala (Liljeborg, 1852) A. t-ypica (Bate, 1856) A. palmata Barnard, 1916 A. brevicovnia (Costa, 1853) A. calypsonis Belian-Santini & Kaîm-Malka, 1977 A. diaderra (Costa, 1853) A. ma8silicnsi8 Bellan-Santini & Kaïm-Malka, 1977 A. anomala Sars, 1882 A. pu8illa Sars, 1895 A. tenuicomis Liljeborg, 1855 A. spinimana Chevreux, 1887 A. spinipes Boeck, 1860 A. aequicomis Bruzelius, 1859 A. uncinata Chevreux, 1857 A. compacta Norman, 1882 A. anblycps Sars, 1895 A. eschrichtii Krôyer, 1852 A. oàontoplax Sars, 1895 A. gibba Sars, 1882 A. abusi'iccla Stebbing, 1888
Figure 3. - Distribution bathymétrique des 30 espèces d'Ampelisca selon une échelle logarithmique. Le trait épais indique les profondeurs effectives de récolte et le trait fin l'étendue possible des bathymétries.
ro

Roche Détritique Sable Vase
A.lusitanica A.rubella
A.spooneri
A.serraticaudata
A.verga
A.brevicornis s.l.
A.heterodactyla
A.senegalensis 907
A.sarsi
A.spinipes
A.typica
A.armoricana
A.spinimana
A.toulemonti
A.diadema
A.pseudospinimana
A.tenuicornis
A.aequicornis
A.eschrichtii
A.qibba
A.latifrons
A.spinif er A.remora
A.calypsonis
A.massiiiensis
A.pseudosarsi
A.uncinata
A.ctenopus
A.macrocephala
A.abyssicola A.amblyops
A. anomala
A.hupferi
A.monoculata
A.odontoplax
A.pusilla
Figure 2. - Distribution écologique des espèces d'Ampelisca en fonction du substrat^ . trait plein : préférence écologique et tireté : présence occasionnelle (aucune donnée n'est disponible pour compacta, A. bidentata et A_^ palmata).
212

5. BIOLOGIE
Les données sur la biologie des espèces de l'Atlantique nord-est sont fragraentaires et ne concernent qu'un nombre restreint d'espèces (voir revue de Dauvin, 1984 et Bellan-Santini & Dauvin, sous presse). Les espèces A. brevicornis et A^ tenuicornis ont fait l'objet de plusieurs études en diverses régions du nord au sud de l'Europe permettant une étude comparée de leur stratégie de reproduction (tableau 2).
On observe une nette corrélation entre la latitude et la période de reproduction. Ainsi, A. brevicornis se reproduit toute l'année en Méditerranée, de mars à décembre à Arcachon et seulement de mai à septembre en mer d'Irlande. II en est de même pour A^ tenuicornis. Typiquement bivoltin à Marseille et à Arcachon (deux générations par période de reproduction annuelle) le cycle reproducteur d'A. brevicornis est univoltin en mer d’Irlande et du Nord et intermédiaire en Manche.
Des températures voisines de 10°C semble d'ailleurs nécessaires pour que ces deux espèces commencent leur vitellogenèse (Sheader, 1977 ; Hastings, 1981 et Dauvin, 1984).
6. CONCLUSION ET PERSPECTIVES
Le genre Ampelisca représente dans l'Océan mondial un important groupe d'invertébrés marins par le nombre élevé de ses espèces lesquelles montrent une étendue de spéciation avec de grands écarts entre les espèces plus plésiomorphes et la plus apomorphe. II occupe une vaste distribution depuis l'intertidal jusqu'à 4000 m. Son importance quantitative dans les communautés macrobenthiques est également a souligner tout comme sa très grande sensibilité aux facteurs écologiques faisant de ces espèces d'excellents indicateurs des conditions naturelles ou perturbées du milieu.
L'étude et la révision de collections aussi bien récentes qu'anciennes permettent actuellement pour 39 des 41 espèces recensées dans l'Atlantique nord-est (Méditerannée exclue) d'émettre avec une bonne marge de sécurité quelques conclusions quant à l'évolution du genre qui comprend actuellement plus de 50 % d'endémiques locales, sa distribution biogéographique et sa répartition suivant les types de substrat et la profondeur. II subsiste malgré tout d'importantes lacunes qui nécessiteraient un réel programme d'étude. Une bonne étude phylogénique ne sera possible que lorsqu'on aura pu retrouver d'autres exemplaires des espèces mal ou insuffisamment décrites.
Si on possède un réseau de données relativement serré dans les zones arctique et boréale, plus réduit mais en cours d'étude de la zone lusitanienne, il n'en est pas de même pour les zones maurétaniennes et sénégaliennes où seules des données fragmentaires ou anciennes sont disponibles. II résulte de cette disproportion des données ,un biais très important dans certaines estimations sur l'étendue et la densité de répartition des espèces.
Les mêmes hétérogénéités existent entre les données du plateau continental et celles du talus et des fonds bathyaux et abyssaux. II est remarquable de noter la dispersion des références existant au-delà de 200 m.
Les données que l'on possède sur les espèces du plateau continental permettent de cerner l'importance des espèces qui constituent des populations denses telles A. cu-moriceuia, A. spinipes, A. sarsi, A. tenuicornis et à un moindre degré A. brevicornis et A. typica pour la zone qui nous intéresse. II est fort regrettable que des études de biologie d'espèces n'aient été réalisées que sur trop peu d'espèces, ne permettant des conclusions générales que pour deux d'entre elles : A. brevicornis et A. tenuicornis.
Les études sur les Ampeliscidés commencées depuis plus d'un siècle et intensifiées au cours des deux dernières décenies montrent qu'il s'agit d'un matériel de choix au carrefour de plusieurs disciplines dont la phylogénie, l'écologie, la biologie et l'Océanographie.
213

Période de reproduction Cycle reproducteur
Régions Seuil printanier des 10°C
A. brevicornis A. tenuicornis A. brevicorms A. tenuicomis
Mer d' lrlande, Ile de Man. (Hastings, 1981)
juin mai-septembre - univoltisme -
Mer du Nord, Northumberland. (Sheader, 1977)
juin - mai-novembre -univoltisme
(très rares cas de bivoltisme)
Mer du Nord, Helgoland Bight. (Klein et ah, 1975)
juin mai-octobre univoltisme
Manche, Rance Maritiroe. (Dauvin, 1984)
mai avril-octobre mai-décembre intermédiaire univoltisme-bivoltisme
bivoltisme
Manche, Baie de Morlaix. (Dauvin, 1979, 1984)
avril avril-décembre mai-décembre intermédiaire univoltisme-bivoltisme
bivoltisme
Atleuitique, Concarneau. (Mesnesguen, 1980)
mars-avril avril-octobre bivoltisme -
Atlantique, Arcachon. (Salvat, 1967)
mars mars-décembre bivoltisrae -
Méditerranée, Marseille. (Kaîm-Malka, 1969)
- toute l'année bivoltisme -
Tableau 2. Période de reproduction et cycle reproducteur d'A. brevicornis et A. tenuicornis en fonction de la latitude.
214

BARNARD J.L., 1960. New bathyal and sublittoral ampeliscid amphipods from California, with an illustrated key to Ampelisca. Pacif. Nat., 1, 16, 1-36. BELLAN-SANTINI D., DAUVIN, J.C., sous presse. Eléments de synthèse sur les Ampelisca du nord-est Atlantique. Crustaceana, suppl. 13. DAUVIN J.C., 1979. Recherches quantitatives sur le peuplement des sables fins de la Pierre Noire et sur sa perturbation par les hydrocarbures de l'Amoco Cadiz. Thèse Doct. 3ème cycle, Univ. P. & M. Curie, Paris, 251 p. DAUVIN J.C., 1984. Dynamique d'écosystèmes macrobenthiques des fonds sédimentaires de la baie de Morlaix et leur perturbation par les hydrocarbures de l'Amoco Cadiz. Thèse Doct. Etat, Univ. P. & M. Curie, Paris, 468 p + annexes 193 p. DAUVIN J.C., BELLAN-SANTINI, D., 1986. Révision de la collection des Ampeliscidés d'Edouard Chevreux du Muséum d'Histoire Naturelle de Paris : remise à jour taxonomique et conséquences sur la distribution bathymétrique et biogéographique de ce groupe. Bull. Mus. natn. Hist. nat., Paris, 4° sér., 8, section A, 4, 867-891. DICKINSON J.J., 1982. The systematics and distributional ecology of the family Ampeliscidae (Amphipoda : GammEu-idae) in the Northeastern Pacific region. 1. The genus Ampelisca. Publ. Oceanogr. Biol., 10, 1-39. FREDJ G., 1974. Essai de stockage et d'exploitation des données en écologie marine. Application à l'étude biogéographique du benthos méditerranéen et perspectives. Thèse Doct. Etat, Univ. Nice, 176 p. HASTTNGS M.H., 1981. The life cycle and productivity of an intertidal population of the amphipod Ampelisca brevicornis. Estuar. Coast Shelf Sci., 12, 665-677. KAIM-MALKA R., 1969. Biologie et écologie de quelques Ampelisca (Crustacea, Amphipoda) de la région de Marseille. Tethys, 1, 4, 977-1022. KLEIN G., RACHOR E., GERLACH S.A., 1975. Dynamics and productivity of two populations of the benthic tube-dwelling amphipod Ampelisca brevicornis (Costa) in Helgoland bight. Ophelia, 14, 139-159. MENESGUEN A., 1980. La macrofaune benthique de la baie de Concarneau. Thèse Doct. 3ème cycle, Univ. Brest, 127 p. NORMAN A.M., 1882. Report on the Crustacea. Exploration of the Faroe Channel during the summer of 1880, in H.M.'s hired ship" Knight Errant." Proc. R. Soc. Edinh.. 11, 683-690. REID D.M., 1951. Report on the Amphipoda (Gammeuridea and Caprellidea) of the coast of Tropical West Africa. Atlantide Rep.. 2, 189-291. SALVAT B., 1967. La macrofaune carcinologique endogée des sédiments meubles intertidaux. Ethologie, bionomie et cycle biologique. Mém. Mus. Hist. Nat.. 45, 1-275. SHEADER M., 1977. Production and population dynamics of Ampelisca tenuicornis (Amphipoda) with notes on the biology of its parasite Sphaeronella longipes (Copepoda). J. mar, biol, Ass. U.K., 57, 955-968.
)
215

ASPECTS RÉCENTS de la Biologie des CRUSTACÉS Actes de Colloques - 8 - 1988 - IFREMER, pages 217 à 222
DONNEES PRELIMINAIRES SUR LES AMPHIPODES DE L'IROISE ET DE SES ABORDS , LEURS
AFFINITES BIOGEOGRAPHIQUES.
DAUVIN J.C.*. TOULEMONT A.**
RESUME - Le dépouilleraent des dragages effectués de 1963 à 1968 en mer d ' Iroise a conduit à l 'identification de 76 espèces d'Amphipodes Gammariens et Caprelliens. Les affinités biogéographiques des différentes espèces présentes ont été déterminées.
Mots-clés : Amphipodes, Iroise, biogéographie.
ABSTRACT - From 1963 to 1968 a series of dredging in the sea of Iroise has led to the identification of 76 species of Amphipoda, Gammarian and Caprellians. The biogeographic affinities of the different species have been determined.
Key-words : Amphipoda, Iroise, biogeography. - r
INTRODUCTION Depuis les travaux de Chevreux à la fin du siècle dernier, les études spécialisées sur
les Amphipodes des côtes atlantiques françaises sont pratiquement inexistantes. Grâce aux travaux d'exploration des peuplements benthiques, entrepris à partir des années 1960, des données sur les répartitions écologiques et bathymétriques de ce groupe ont été recueillies. Le dépouillement des dragages effectués de 1963 à 1968 en mer dlroise et dans les baies adjacentes est achevé en ce qui concerne les Amphipodes. 76 espèces d'Amphipodes Gamraariens et Caprelliens ont été identifiées à partir du matériel présent dans près de 500 stations sur les 1000 stations qui contenaient de la macrofaune. Cette étude complétée des quelques autres concernant cette région porte à 94 espèces Tinventaire connu de la faune des Amphipodes de llroise et ses abords. Les affinités biogéographiques de cette faime avec celles des mers voisines ont été recherchées.
1. INVENTAIRE FAUNISTIQUE, ECOLOGIE ET BATHYMETRIE
Les 76 espèces recensées appartiennent à 21 familles dont 20 Gammaridea et 1 Caprellidea. (Tableau 1).
II convient de mentionner les signalisations méridionales d'Hippomedon propinquus et Tryphosella horingi, les signalisations septentrionales de Leucothoë richiardii et Stenothoë tergestina, la description de deux nouvelles espèces d'Ampelisca à partir de ce matériel et la présence d'Ampelisca spinifer et d'Haploops nirae dans ces eaux. 30 espèces sont présentes en une station, 29 entre 2 et 5 stations, 12 entre 6 et 20 ; 5 espèces seulement ont été récoltées en plus de 21 stations : Corophium volutator (22 stations), Ampelisca brevicornis (34), Hippomedon denticulatus (45), Maera othonis (56) et A. spinipes (281).
* CNRS, LP 4601 et Univ. P. & M. Curie, Paris VI, Station Biologique, 29211 ROSCOFF. **Institut Océanographique, 195 rue Saint Jacques, 75005 PARIS.
217

LYSIANASSIDAE Hippomecbn denticulatue (Bate, 1857) Hippomedon propinquus (Sars, 1890) Ichnopus epinicomis Boeck, 1861 Lepidepecrum longicome (Bate & Westwood, 1861) Lyeianaeea ceratina (Walker, 1899) Lyaeianaeea plumoqa Boeck, 1871 Tmetonyx eimilis (Sars, 1891) Tryphoeella horingi (Boeck, 1871) Tryphoeitee longipee (Bate&Westwood, 1861)
AMPELISCIDAE Ampelieca aequicomie Bruzelius, 1859 Ampelieca armoricana Bellan-Santini & Dauvin 1981 Ampelieca brevicomie (Costa, 1853) Ampelieca diadema (Costa, 1853) Ampelieca gibba Sars 1882 Ampelieca earei Chevreux, 1887 Ampelieca epinimana Chevreux, 1887 Ampelieca epinifer Reid, 1951 Ampelieca spinipee Boeck, 1861 Ampelieca epooneri Dauvin 4 Be 11 an-Santini 1982 Ampelieca tenuicomie Liljeborg, 1855 Ampelieca toulemonti Dauvin 4 Bellan-San.1982 Ampelieca typica (Bate, 1856) haploope nirae Kaïm-Malka, 1976
ACANTHONOTOZOMATIDAE Panoplea eblanae (Bate, 1857) Panoplea minuta (Sars. 1882)
COLOMASTIGIDAE Colomaetix pueilla Grube, 1861
LEUCOTHOIDAE Leucothoë inciea Robertson, 1892 Leucothoë richiardii Lessona, 1865 Leucothoë epinicarpa (Abildgaard, 1789)
STENOTHOIDAE Stenothoë tergeetina (Nebeski, 1881)
GAMMARIDAE Gammarue locusta (Linné, 1758)
MELITIDAE Ceradocue eemieerratue (Bate, 1862) Cheirocratue aseimilie (Liljeborg, 1852) Elasmopus rapax Costa, 1853 Ganmarella fucicola (Leach, 1814) Maera othonie (Milne-Edwards, 1930) Melita gladioea Bate, 1862 Melita obtusata (Montagu, 1813)
HAUSTORIIDAE Bathyporeia guilliamsoniana (Bate, 1856) Bathyporeia sarsi Watkin, 1938 Bathyporeia tenuipes Meinert, 1877 Urothoë brevicomis Bate, 1862 Urothoë elegans (Bate, 1856) Urothoë pulchella (Costa, 1853) Urothoë marina (Bate, 1856)
OEDICEROTIDAE Monoculodee carinatue (Bate, 1856) Monoculodea subnudus Nnrnun iQflo
EUSIRIDAE Eueirue longipes Boeck, 1861
CALLIOPIIDAE Apherusa jurinei (Milne-Edwards, 1830) Apherusa ovalipes Norman & Scott, 1906 Garrmarellus angulosus (Rathke, 1843)
LTYLIDAE
Atylus guttatue (Costa, 1851) Atylus swarmerdami (Milne-Edwards, 1830) Atylue vediomensie (Bate 4 Westwood, 1862)
DEXAMINIDAE Dexamine spinosa (Montagu, 1813) Tritaeta gibbosa (Bate, 1862)
^MPITHOIDAE Ampithoe ramondi Audouin, 1826
[SAEIDAE Chmmaropeie macutata (Johnston, 1828) Photie iongicaudata (Bate 4 Westwood, 1862)
^ORIDAE Aora typica Krôyer, 1845 Leptocheirue hirsutimanus (Bate, 1862) Leptocheirus pectinatus (Norman, 1869)
^ODOCERIDAE Podoceru8 variegatus Leach, 1814
'OROPHIIDAE Corophium acherusicum (Costa, 1851) Corophium acutum Chevreux, 1908) Corophium sextonae Crawford, 1937 Corophium voiutator (Pallas, 1766) Uncioia crenatipaima (Bate, 1862)
;SCHYROCERIDAF Erichtoniue punctatus (Bate, 1852) Jassa faicata (Montagu, 1808)
:APRELLIDAE
Capreiia acanthifera Leach, 1814 Capreüa acutifrons Latreille, 1816 Capretia tinearie (Linné, 1767) Capretla tubercutata Bate 4 Westwood, 1868 Phtisica marina Slabber, 1769 Paeudoprotetia phasma (Montagu, 1804)
Tableau 1. Liste faunistique des espèces d'Amphipodes récoltées.
218

Les principales espèces rencensées affectionnent les fonds sableux et sablo-vaseux (Fig. 1) ; les fonds grossiers très répandus dans cette région sont relativemeut pauvres en Amphipodes. Par rapport à la littérature et à nos données, 13 espèces ne dépassent pas 20 m de profondeur, 22 ne sont pas connues au-delà de .50 m. 41 espèces soit 54 % n'ont été récoltées que dans les 120 premiers mètres (profondeur maximale d'échantillonnage en Iroise). 55 espèces (72 %) ne fréquentent que le plateau continental; 12 espèces atteignent 500 m et 9 dépassent 500 m.
2. AFFINITES BIOGEOGRAPHIQUES
214 espèces d'Amphipodes Gammaridae de substrat ineuble du plateau continental ont été recensées dans la région atlantique s'étendant de la mer d'Iroise au Cap Breton et en Manche. 7 zones ont été délimitées (Tableau ci-dessous). 126 espèces sont communes à 2, 3, 4, 5 ou 6 zones.
Zone 1 Zone 2 Zone 3 Zone 4 Zone 5 Zone 6 Zone 7
Mer d'Irlande
Mer Celtique
Manche Ouest
Manche Est
Mer dlroise
et abords
Nord-Gascogne
Sud Gascogne
Lincoln, 1979
Lincoln, 1979
invest. pers.
ChewreuxChevreux Chevreux & Fage, & Fage, & Fage,
1925 1925 1925 Toulemond Gentil, Quiniou,
Truchot, 1976 1978 1964 et comm. cette
invest. pers. étude Lincoln, Glaçon,
1979 1977
Chevreux & Fage,
1925 Lagardère,
1976
Chevreux & Fage,
1925 Sorbe, 1984
invest. pers.
Nombre total d'espèces
135 92 153 107 94 126 127
Nombre d'espèces en commun avec 2,3,4,5 et 6 zones
86 49 104 67 54 80 71
Des similitudes de point fortes ou très fortes apparaissent entre la zone 5 et les zones limitrophes 2, 3, 4 et 6 (mer Celtique, Manche et Nord-Gascogne) dans l'analyse sur les 214 espèces ; par contre, dams l'analyse sur les 126 espèces présentes en 2, 3, 4, 5 ou 6 zones, la zone 5 présente de très faibles affinités avec l'ensemble des 6 autres zones.
Dans l'espace factoriel des deux premiers axes de l'analyse des correspondances effectuée à partir de la matrice présence-absence des 126 espèces (n° 1 à 214) et des 7 zones (Z1 à Z7) la zone 5 se sépare nettement de l'ensemble des 6 autres zones (Fig. 2). Une classification hiérarchique permet de séparer 5 groupes spécifiques : A à D. Le groupe A composé de 37 espèces permet de distinguer la zone 5 (mer d'Iroise) qui semble, par conséquent, présenter une faune amphipodologique purticulière. Les distributions de certaines espèces permettent de donner des éléments d'interprétation. On rencontre en Iroise à la fois des espèces dont la limite Nord ne s'étend pas au-delà de la Manche orientale (Corophium acherusicum), de la Manche occidentale (Ampelisca sarsi, A, spinimana) de llroise (Westwoodilla rectirostris). D'autres espèces méridionales
219

n'atteignent pas l'Iroise (Microdeutopus armatus, Monoculodes gibbosus). A l'inverse, on rencontre d'autres espèces dont la limite sud ne s'étend pas au-delà de la zone nord-Gascogne (Gammarellus angulosus, Panoplea eblanae), de l'Iroise (Bathyporeia sarsi, Cheirocratus assimilis, Urothoë poseidonis). Enfin, des espèces boréales n'atteignent pas l'Iroise (Ampelisca macrocephala, Paraphoxus oculatus, Tmetonyx cicada). Ainsi, la mer Iroise apparaît comme un carrefour privilégié d'espèces d'affinités boréales et tempérées ou tempérées chaudes.
CONCLUSION
Cette étude montre l'intérêt des recherches bionomiques pour faire progresser la connaissance sur un groupe. Dans de tels travaux il est très utile de fournir des listes faunistiques complètes avec dans la mesure du possible les positionnements précis des stations dans lesquelles les espèces sont présentes. La faune amphipodologique de la mer Iroise semble particulière mais les investigations dans cette zone demeurent insuffisantes. Toutefois, l'Iroise apparaît comme une zone de mélange de faune avec des limites biogéographiques d'espèces méridionales ou septentrionales. A cet égard, Fischer (1881) et Fredj (1974) placent la frontière entre les provinces celtique ou boréale au Nord et la province lusitanienne au sud au niveau de l'Iroise.
CHEVREUX E., FAGE, L., 1925. Amphipodes. Faune de France, 9, 488 p. FISCHER P., 1881. Manuel de conchyliologie et de paléontologie conchyliologique. F. Savy, édit., Paris. FREDJ G., 1974. Essai de stockage et d'exploitation des données en écologie majine. Application à l’étude biogéographique du benthos méditerranéen et perspectives. Thèse doct. Etat, Univ. Nice, 176 p. GENTIL F., 1976. Distribution des peuplements benthiques en baie de Seine. Thèse Doct. 3ème cycle, Univ. Paris VI, 70 p. GLACON R., 1977. Faune et flore du littoral du Pas de Calais et de la Manche orientale. Ed. Inst. Biol. Mar. Reg. Wimereux, 51 p. LAGARDERE J.P., 1976. Recherches sur la distribution verticale et sur l’alimentation des Crustacés Décapodes de la pente continentale de l'Atlantique Nord-orientale. Thèse Doct. Etat, Univ. Marseille, 188 p. LINCOLN R.J., 1979. British Marine Amphipoda. Gammaridea. Bristish Museum (Natural History), 658 p. QUINIOU L., 1978. Les Poissons démersaux de la baie de Douarnenez. Alimentation et écologie. Thèse Doct. 3ème cycle, Univ. Brest, 222 p. SORBE J.C., 1984. Contribution à la connaissance des peuplements suprabenthiques néritiques sud-gascogne. Thèse Doct. Etat, Univ. Bordeaux, 301 p. TOULMOND A., TRUCHOT J.P., 1964. Inventaire de la faune marine de Roscoff. Amphipodes. Cumacés. Trav. Stat. Biol. Roscoff, n.s., 15, 42 p.
220

Fig. 1. Préférences écologiques
Fig. 2. Analyse des correspondances
221

ASPECTS RÉCENTS de la Biologie des CRUSTACÉS Actes de Colloques - 8 - 1988 - IFREMER, pages 223 à 228
BIOLOGIE DU COPEPODE Eurytemora affinls hirundoxdes DANS LA GIRONDE :
DONNEES MORPHOMETRIQUES.
FEURTET A. et CASTEL J.
RESUME Eurytemora affinis hirundoides est le copépode planctonique dominant dans l'estuaire de Gironde. soumis à de très forts gradients de salinité. La réponse des populations à ces gradients est évaluée par les variations des caractéristiques biométriques simples typiques à l'espèce. Les variations observées traduisent l'influence de deux facteurs importants : la salinité et la turbidité.
Mots-clés : Morphométrie, estuaire, salinité, copepode, Eurytemora affinis hirundoides .
ABSTRACT : Eurytemora affinis hirundoides is the most abundant Copepod in the Gironde estuary which is submitted to strong salinity gradients. The response of the populations to these gradients is mesured by following the variation of simple biometric characteristics. The observed variations reflect the influence of two important factors : salinity and turbidity.
Key-words : Morphometry, estuary, salinity, Copepoda, Eurytemora affinis hirundoides .
Les Copépodes planctoniques sont particulièrement abondants dans les estuaires. pourtant, dans de tels milieux ils doivent faire face à des conditions défavorables résultant de deux facteurs principaux : la dilution de l'eau de mer et la quantité de matières en suspension (M.E.S.). Les variations de salinité entraînent des efforts d'osmorégulation et modifient la sustentation des organismes planctoniques. Les importantes concentrations en M.E.S., en réduisant la pénétration de la lumière, limitent la production de phytoplancton et donc les ressources alimentaires. Ces deux facteurs, salinité et turbidité, vont donc intervenir sur la biologie des populations de Copépodes et, en particulier, sur leurs caractéristiqes morphométriques.
En effet, les variations de taille des Copépodes reflètent l'influence de facteurs externes tels que la température, la nourriture, la prédation taille-sélective (Crawford et Daborn, 1986). Parallèlement, des variations de poids traduisent des modifications d'état nutritionnel ou d'état de reproduction. De plus, différentes caractéristiques biométriques permettent parfois de séparer des populations ou des générations soumises à des conditions de milieu différentes (Gaudy, 1971). Toutefois, il est difficile de relier les caractéristiques morphométriques des Copépodes à un seul facteur.
(1) Institut de Biologie Marine, Université de Bordeaux I - 33120 ARCACHON
223

Eurytemora affinis hirundoides est le Copépode planctonique dominant dans l'estuaire de Gironde, long d'une centaine de Km entre Bordeaux et Royan. II est particulièrement abondant dans la zone oligo-mésohaline (0.5-10 %.) où sont également notées les plus fortes concentrations en M.E.S. dépassant parfois 1 g/1 (Castel, 1981 ; Castel et Feurtet, 1986). L'existence de forts gradients de salinité et de turbidité font que les populations sont soumises à des conditions de milieu très différentes depuis l'amont vers l'aval. La réponse des populations est évaluée par les variations de caractéristiques biométriques simples, typiques à l'espèce (Fig. 1). Les mesures sont pratiquées sur le prosome :
Fi-g. 1 : E.a. hirundoides. Mesures pratiquées sur les adultes.
longueur et largeur au niveau de la limite premier-deuxième segment thoracique, sur l'abdomen, sur une rame furcale et sur les expansions alaires pour les femelles. A chaque point de prélèvement 100 mâles et 100 femelles adultes sont mesurés et pesés (micro-balance sensible à 0,1 ug) .
L'évolution de la taille des adultes, représentée par la longueur du prosome, a été suivie le long de l'estuaire du pk 5 au pk 78 (respectivement 5 et 78 km en aval de Bordeaux). Les tailles minimales des adultes sont observées entre le pK 35 et le pk 52, qu'il s'agisse des mâles ou des femelles (Fig. 2). C'est également dans cette zone que les concentrations en
Fig. 2 : E.a. hirundoides. Longueur du prosome (L) et poids sec (P) des adultes en relation avec la turbidité (en grisé), le long de l'estuaire de Gironde (juillet 1980). pk = point kilométrique dont l'origine est Bordeaux.
224

M.E.S. sont les plus élevées. Corrélativement à la taille, le poids individuel raoyen dirainue du pk 5 au pk 35 puis augraente du pk 52 à 1'embouchure. La perte de poids peut atteindre 49 % pour les raâles et 63 % pour les femelles. En première approximation une dirainution de taille et donc de poids peut être interprétée comrae une accélération de la vitesse de développement de mue chez l'adulte). L’évolution du facteur de condition (FC = P/L . 10 ), représentatif de "1'embonpoint" des aniraaux contredit cette hypothèse (Fig. 3).
Fig. 3 : E.a. hirundoides. Evolution du facteur de condition des adultes en relation avec la turbidité (en grisé), le long de l'estuaire de Gironde (Juillet 1980). pk = point kilométrique dont l'origine est à Bordeaux.
En effet, les valeurs de FC, à peu près similaires pour les deux sexes (1,40 pour les raâles et 1,42 pour les feraelles), présentent égaleraent un minimura dans la zone de forte turbidité. On peut donc en conclure que la population est dans un état de déficit nutritionnel entre le pk 35 et le pk 52.
L'évolution des rapports dimentionnels concernant la furca et les expansions alaires (Fig. 4) serable indépendante des concentrations en M.E.S. On constate une augraentation significative du rapport L/1 de l’amont vers l'aval. Cette accroisseraent est fort entre le pk 52 et le pk 67 au niveau du raaximum de turbidité. Une telle augmentation correspond en fait à une diminution de la surface furcale. Pour les feraelles, le rapport H/B des expansions alaires raontre une décroissance à peu près continue de l'amont vers l’aval, sans particularité au niveau du maxiraum de turbidité.
225

Fig. 4. : E. a. hirundoides. Evolution de deux caractéristiques biométriques et valeurs de turbidité (en mg/1) le long de l'estuaire de Gironde (mars 1979). pk = point kilométrique dont l'origine est à Bordeaux.
La diminution des surfaces portantes de l'amont vers l'aval semble pouvoir être reliée au gradient de salinité, c'est à dire de densité et de viscosité de l'eau (Fig. 5). En effet, la densité de l'eau augmente beaucoup à partir du pk 52. Parallèlement on constate une très nette diminution de la surface corporelle des individus. En aval, les fortes densités de l'eau favorisent la flottabilité des organismes dont la surface portante décroît. En amont, l'eau douce, de densité proche de l'unité, nécessite une augmentation de surface corporelle.
itg- 5 : E. a. hirundoides. Evolution de la surface corporelle totale des adultes et valeurs de la densité de l'eau le long de l'estuaire de Gironde (mars 1979). pk = point kilométrique dont l'origine est à Bordeaux.
226

E. a- hirundoides présente donc de fortes variations morphologiques, de taille et de poids le long de l'estuaire de Gironde. Le facteur température pourrait expliquer en partie les variations de taille des adultes car il intervient sur la durée du développement. L'influence de ce facteur est ici éliminée puisque les prélèvements ont été réalisés en moins d'une semaine aux différents points situés le long de l'estuaire. L'influence de la prédation taille-sélective semble pouvoir être négligée. Les principaux prédateurs sont en effet des alevins de poissons migrateurs, anadromes ou catadromes ; il est peu probable que la prédation soit plus intense à un endroit particulier de l'estuaire plutôt qu'à un autre.
Les faibles valeurs de taille, de poids et surtout du facteur de condition au niveau du maximum de turbidité résultent d'un déficit nutritionnel, les fortes turbidités limitant la production phytoplanctonique. Ceci est confirmé par une diminution de la fécondité observée dans cette zone (Castel et Feurtet, 1986) et par des élevages en laboratoire montrant une meilleure croissance des animaux quand les milieux de culture sont supplémentés en algues unicellulaires (Poli et Castel, 1983).
Le facteur salinité induit également des variations morphologiques chez Eurytemora , variations qui se traduisent par des modifications des surfaces portantes. La salinité, par 1'intermédiaire de la densité et de la viscosité de l'eau, conditionne la flottabilité. Cette dernière est optimale vers l'aval mais s'accompagne de conditions d'osmoticité précaires dues à de fortes salinités. En effet, Eurytemora est isosmotique à 6 %. et osmorégule ente 3 et 6 Z. (Gyllenberg et Lunqvist, 1979). En amont, la faible portance du milieu est compensée par un développement furcal et alaire.
En définitive, les variations morphométriques constatées chez E. a. hirundoides traduisent la réponse des populations à deux facteurs importants en estuaire : la salinité et la turbidité. XI s'agit pour l'espèce de trouver un compromis entre deux nécessités : (1) assurer sa sustentation tout en réduisant les efforts d'osmorégulation ; (2) satisfaire ses besoins énergétiques dans des conditions nutritionnelles souvent sub-optimales.
CASTEL J. , 1981. Aspects de l'étude écologique du plancton de l'estuaire de la Gironde. Océanis , 6^ , 535-577. CASTEL J., FEURTET A., 1986. Influence des matières en suspension sur la biologie d'un Copépode estuarien : Eurytemora hirundoides (Nordquist, 1888). Coll. Nat. CNRS "Biologie des populations" , 391-396. CRAWFORD P., DAGORN G., 1986. Seasonal variations in body size and fecundity in a Copepod of turbid estuaries. Estuaries , _9 , 133-141. GAUDY R., 1971. L'allongement antennulaire chez Centropages typicus ; signification adaptative et utilité biométrique. Rapp. P.-v. Comm. int. Explor. scient. Mer Méditer. , , 363-365. GYLLENBERG G. , LUNDQVIST G. , 1979. The effects of temperature and salinity on the oxygen consumption of Eurytemora hirundoides (Crustacea, Copepoda). Ann. Zool. Fennici 16 , 205-208. POLI J.M., CASTEL J., 1983. Cycle biologique en laboratoire d'un Copépode planctonique de l'estuaire de la Gironde : Eurytemora hirundoides (Nordquist, 1888), Yi^J!
111 Milieu, 3.3, 79-86.
227

THEME 5 - EXPLOITATION
- L'exploitation mondiale des Crustacés : Bilan et perspectives.
Laubier A. et Laubier L.
- L'aquaculture des crevettes, rô1e de la nutrition.
Guillaume J.
- Perspectives d'accroissement de la production de l'océan Atlantique par 1'introduction d'espèces de Crustacés comblant des niches écologiques vides.
Vasserot J.
- Contribution d'un film bactérien à la culture intensive de Daphnia magna .
Langis R., Proulx D., de la Noüe J. et Couture P.
229

ASPECTS RÉCENTS de la Biologie des CRUSTACÉS Actes de Colloques - 8 - 1988 - IFREMER, pages 231 à 240
L'EXPLOITATION MONDIALE DES CRUSTACES : BILAN ET PERSPECTIVES
LAUBIER Annie <]> LAUBIER Lucien (2)
RESUME - Depuis une vingtaine d'années, l'exploitation mondiale de Crustacés est passée de 1.263.000 tonnes en 1968 à 3.419.000 tonnes en 1985, dépassant de plus de un million de tonnes les évaluations de 1970. Les crevettes Penaeides ont connu un développement très important. Le krill s'est révélé pratiquement non exploitable commercialement. A l'heure actuelle, les principaux stocks paraissent pleinement exploités. Au cours de la même période, I'élevage des crevettes Penaeides a connu une expansion rapidc, qui devrait se poursuivre au cours des prochaincs années. Mots-clés : Crustacés, exploitation, bilan mondial, perspectives.
ABSTRACT - Since nearly twenty years, world exploitation of Crustaceans increases from 1.263.000 tons in 1968 to 3.419.000 tons in 1985, overcoming the predictions of 1970 by more than one million tons. Penaeid prawns fisheries experienced a very fast development. The krill resource proved to be practically commercially unworkable. By now, the major Crustaceans stocks appear fully exploited. During the same period, Penaeid aquaculture experienced a fast growth, which should continue in the near future. Key-words : Crustaceans, exploitation, world balance, prospects.
Avec les Mollusques, les Crustacés constituent l’un des deux principaux groupes d'Invertébrés aquatiques traditionnellement exploités par la pêche et l'élevage. Les Poissons représentent à eux seuls un peu plus des trois quarts de la production mondiale en tonnage : en 1985, la production mondiale de ressources vivantes aquatiques s'est élevée à 84.945.300 tonnes, dont 74.825.200 tonnes provenant du milieu marin; dans ce dernier total, Ies Poissons marins interviennent pour 64.086.000 tonnes contre 5.840.800 tonnes de Mollusques et 3.419.000 tonnes de Crustacés. En revanche, en valeur, les Crustacés représentent une proportion plus élevée et leur exploitation progresse depuis une vingtaine d'années à une vitesse supérieure à la moyenne des pêches mondiales.
Conformément à la demande des organisateurs de la Xème réunion des Carci-nologistes de langue française, cette communication à caractère très général a pour objet de présenter l'exploitation actuelle des Crustacés à l'échelle mondiale, en cher-chant à dégager les tendances récentes et, dans une moindre mesure, les perspectives.
A l'exception des pêches artisanales de Cirripèdes - au Portugal et dans certaines régions françaises de l'Ouest comme les côtes bretonnes et les îles du Ponant, il s'agit du pouce-pied, Pollicipes cornucopiae, au Chili de la grande balane Balanus (Megabalanus) psittacus dont l'exploitation représente plusieurs milliers de tonnes par an - et des récoltes de cystes d'Artemia salina ou d'adultes pour l’aquaculture et l'aquariologie, les crustacés exploités appartiennent tous aux Malacostracés et, pour l'essentiel, aux Décapodes. Les Stomatopodes n'interviennent dans les captures que pour quelques milliers de tonnes et les Euphausiacés, avec le cas particulier du krill antarctique.
Laboratoire de Physiologie des Etres marins, Institut Occanographique, 195, rue Saint-Jacques, 75005 PARIS, France. (2) Institut Français de Recherche pour l'Exploitation de la Mer (IFREMER), 66, avenue d'Iéna, 75116 PARIS, France.
231

Euphausia superba, ne semblent pas devoir prendre à moyen terme l'importance considérable qu’on leur accordait il y a une quinzaine d’années, bien au contraire. D'autre part, plus de 90 pour cent de la production totale enregistrée au cours de l'année 1985 résulte de la pêche. L'aquaculture de Crustacés, qui se développe d'ailleurs rapidement dans la mesure où elle était à peu près inexistante il y a une trentaine d'années, est encore limitée en 1985 à un tonnage compris entre 7 et 8 pour ccnt de la production totale.
Comme pour les Poissons, les pêches de Crustacés ont atteint au cours des cinq dernières années leur niveau optimal : la phase d'expansion des pêcheries doit être considérée comme achevée (à l'exception du cas du krill antarctique, où les limitations actuellement rencontrées sont d'ordre technique et économique et ne se situent pas au niveau de la ressource biologique). II faut donc admettre qu'il n'existe plus dans l'océan mondial, en l'élat actuel des connaissances, de ressources potentielles importantes de Crustacés inconnues ou non encorc exploitées
TABLEAU 1. EVALUATION DE LA PRODUCTION MONDIALE POTENTIELLE DE CRUSTACÉS EN MILLIERS DETONNES ("THE FISH RESOURCES OFTHE OCEANS", J. GULLAND EDIT., F.A.O., 1971)
(homards et langoustes = hom.lan ; crabes et araignées = cr.ar ; crevettes = crev)
PRODUCTION POUR 1968 POTENTIEL ESTIME
HOM.LAN CR,AR CREV HOM.LAN CR,AR CREV ATLANT. N.O. 31,7 2,2 8,6 32 10 27 ATLANT. N.E. 29,9 13,3 65,4 65,3 20,5 82,8 MEDITERR. 4 - 23 4 - 50 PACIF. N. - 208 41,1 - 250 131,2 ATLANT. C.O. 13,3 76,9 125 21 80 160 ATLANT. C.E. 2,5 - 4,6 4 10 63 OC. IND. OCC. 0,4 - 121,3 3,1 - 144 OC. IND. OR. 8,6 - 58,9 10 - 98,1 PACIF. C.O. 1,7 39,3 304 5 150 544,1 PACIF. C.E. - 1,5 65 5 32,5 75 ATLANT. S.O. 3,5 15,8 37,1 4 21 86 ATLANT. S.E. 10,7 - 2 11,5 - 7 PACIF. S.O. 12.3 1 4,5 14,5 1 4,9 PACIF. S.E. 0,1 14,2 11,5 0,1 66,5 20 TOTAL 118,7 372,2 772,1 179,5 641,5 1492,1
II y a près d'une vingtaine d'années, la F. A. 0. prenait l'initiative de publier un Atlas des ressources de Crustacés préparé sous la direction éditoriale de J.A. GULLAND (F.A.O., 1971), donnant notamment, à la lumière des informations disponibles à cette époque, les perspectives de production, c'est-à-dire le potentiel biologique. Le tableau 1 synthétise l'ensemblc des données concernant les Crustacés et confronte, région par région et pour les trois principaux groupements qui pouvaient alors être reconnus avec quelques certitudes dans Ies statistiques (homards, langoustes et cigales, crabes et araignées de mer, crevettes), la production effective pour l’année 1968 et le potentiel estimé pour chacun de ces groupements dans chacune des régions. Le découpage régional de I'océan mondial adopté pour cette étude est relativement succinct, puisqu'il se borne à reconnaître quatorze grandes régions, l'océan Antarctique notamment n'ctanl pas identifié.
232

Selon les zones considérées, les progressions qui pouvaient ctrc escomptccs d'après le potentiel indentifié par les experts reposent pour l'essentiel sur les espèces ou groupes d'espèces qui suivent :
I. Atlantinue Nord-Ouest : Pandalus borealis, P montagui et Cancer irroratus paraissent pouvoir supporter un taux d'exploitation nettement plus élevé. En revanche, le homard américain est considéré comme largemcnt exploité.
?.. Atlantique Nord-Est : Pandalus borealis et Nephrops norvegicus, encore incomplètement exploitées, sont supposées pouvoir fournir 12 pour cent du total des prises mondiales de Crustacés.
3. Méditerranée : l'accroissement (x 2) est principalement fondé sur les crevettes Penaeides littorales ct profondes, cette estimation était considérée comme optimiste par plusieurs experts.
4. Pacifique Nord : progression principalement fondée sur une meillcure gestion des stocks de Pandalides, Pandalus borealis, P. jordani et P platyceros, Pandalopsis dispar, rendue possible par les progrès des connaissances sur la biologie de ces espèces hermaphrodites protandres, ainsi que des crabes Chionoecetes opilio ct Cancer magister.
5. Atlantique centre-Ouest : accroissement fondé pour l'essentiel sur une meilleure gestion des Penaeides littorales et des stocks de langoustes el de cigales du golfe des Caraïbes.
6. At.lantique centre-Est : accroissement des captures de Penaeides littorales et création de pêcheries nouvelles sur les crabes et sur les Penaeides profondes de la pente continentale, conduisant à un développement très important (xl4).
7. Océan Indien occidental : accroissement réduit (x 1,3) sur les Penaeides et les Caridea littoraux et lagunaires, aux Indes en particulier (certains experts envisagent le doublement des captures).
8. Océan Indien oriental : accroissement de la production de crevettes Penaeides littorales ( x 1,8).
9. Pacifiaue centre-Ouest : forte augmentation des captures de crabes ( x 4), principalement Scylla serrata et Portunus pelagicus et accroissement des prises de Penaeides littorales (x 1,9, soit 240.000 tonnes en plus!). Les stocks profonds encore inconnus dans cette zone ne sont pas retenus. En 1968, cette région représente plus de 45 pour cent du total mondial, proportion qui devrait légèrement diminuer.
10. Pacifiaue centre-Est : la progression est fondée sur l'exploitation des stocks de Galathéides, Pleuroncodes monodon et P. planipes, Munida gregaria. Certaines sont pélagiques et abondent en zone d'upwelling, d'où cette évaluation relativement optimiste.
II. Atlantiaue Sud-Ouest : accroissement portant sur les Penaeides littorales du Brésil (x 2,2) et du reste du secteur.
12. Atlantique Sud-Est : augmentation importante (x 3) fondée sur le déve-loppement des pêcheries de Penaeides de profondeur.
13. Pacifiaue Sud-Ouest : pratiquement pas d'augmentation escomptée, les stocks paraissant déjà totalement exploités.
14. Pacifiaue Sud-Est : forte augmentation de l'exploitation des Galathéides Pleu-roncodes monodon et Cervimunida johni (x 5), ouverture d'une pêcherie de Penaeides de profondeur.
La production totale pourrait ainsi passer de 1.263.000 tonnes à 2.313.000 tonnes, ce qui représente un peu moins du double. II est apparu intéressant d'analyser ces données par rapport aux captures effectivement constatées pour 1985, à partir des mêmes statistiques de la F.A.O. Le tableau 2 présente les chiffres statistiques de production mondiale de Crustacés, toutes zones confondues, depuis 1958, selon un découpage zoologique plus fin que celui utilisé précédemment. II subsiste d'assez
233

importames incertitudes dans les statistiques nationales, où toutes les espèces ne sont ni identifiées, ni même rattachées à un groupe zoologique déterminé ; d'où les valeurs élevées des Crustacés divers hors crevettes et des Natantia indéterminés, qui ont été regroupés avec les Penaeides iittorales, ce groupement n'étant guère satisfaisant. De même, les statistiques concernant les pêches en eau douce sont assez difficiles à utiliser, étant donné la présence d'un pourcentage élevé de Crustacés divers. En revanche, les Crustacés dits nobles (homards, langoustes et cigales, langoustines), certaines formes d'eau douce, écrevisses et crevettes (qui appartiennent principalement au genre Macrobrachium) sont identifiés avec une bonne précision.
TABLEAU 2. PRODUCTION MONDIALE DE CRUSTACÉS EXPRIMÉE EN MILLIERS DE TONNE ENTRE 1958 ET 1985 ( N.S. = NON SÉPARÉS DANS LES STATISTIQUES DE LA F.A.O.)
1958 1966 1973 1978 1985 EAU DOUCE Astacidés N.S. N.S. 7 8,8 Total 91,3 Caridés et divers N.S. N.S. 50 143 EAU DE MER Crabes et araignées 197 365 450 760 887 Homards 36 32 31 40,5 55.5 Langoustes et cigales 43 59 63 68 83,8 Langoustines 19 32 52 46,5 61,4 Galathées 13 13 25 23,4 10,3 Penaeides littorales et Natantia indéterminés 440 616 1.000 1.369 1.556 Penaeides profondes 13 21 32 75 62 Pandalidés 35 60 80 176 226 Crangonidés 49 60 45 47 55,8 Palaemonidés N.S. N.S. N.S. 2,1 2,8 Stomatopodes (squilles) N.S. N.S. N.S. 4 5,3 Euphausiacés (krill) - - - 132 191 (les plus fortes captures de krill ont été enregistrées en 1982 avec 529.000 tonnes) Divers hors crevettes 50 77 85 60 69,9
TOTAL 860 1.275 1.785 2.895 3.419
Le total mondial pour 1985 de 3.419.000 t montre tout d'abord que les prévisions faites en 1970 s’avèrent globalement très prudentes. Par ailleurs, la seule ressource qui, en 1970, pouvait apparaître comme susceptible de foumir à terme 100.000.000 t, c'est-à-dire le krill antarctique, ne représente que 191.400 t et l'exploitation apparaît déjà en phase de recul après une production maximale en 1981. En 1985, deux pays occupent une place privilégiée dans les captures de krill : l'URSS, avec 82 pour cent des prises et ie Japon avec 17 pour cent.
L'analyse par enscmbles zoologiques révèle de fortes différences dans la précision dcs estimations.
Pour I'ensemble des Crustacés nobles, homards, langoustes et cigales, langoustines, l'évaluation est très proche de la production actuelle quinze ans plus tard (potentiel de 179.500 t apprécié en 1970, pcche de 200.700 t en 1985).
234

Pour le vaste ensemble des crabes et des araignées, la prévision déjà forte de
641 500 t a été sensiblement dépassée par la production qui attcint 886.800 t. L’exploitation des galathées est en regression sensible, contrairement aux
prévisions ce sont surtout les crevettes dans leur ensemble (Penaeides et Caridés) qui
subissent la plus forte augmentation en passant d'une estimation de 1.492.000 ne
production réelle de 1.902.800 t, progression qui constitue un record en valeur absolue.
Ces résultats peuvent être expliqués par des facteurs differents
- Chez les Penaeides littorales, un accroissement substantiel de l effort de pêche,
en particulier dans l’Indo-Pacifique, est allé de pair avec une amélioration des methodes
8 - Dans le Pacifique Nord, la gestion des stocks de Pandalidés a été renouvelée par
la connaissance précise du cycle biologique et de la croissance de ces hermaphrodites
Pf0ta - Le développement dans l’Atlantique oriental des pêchenes de Penaeides dc
profondeur (entre 150 et 600 mètres).
TABLEAU 3. LES PRINCIPALES ESPECES (OU GROUPES D’ESPECES) DE CRUSTACÉS DONT LA
PRODUCTION POUR L’ANNÉE 1985 EST ÉGALE OU SUPÉRIEURE A 10.000 TONNES.
Espèces figurant parmi les soixante-dix principales espèces i nrnrinrtinn tntale de 47.012,200 tonnes) ■
Acetes japonicus 222.608 T Pandalus borealis 188.964 T Euphausia superba 191.472 T
Espèces dont la production est comprise entre 10.000 et—100.000 tonnes_
Macrobrachium sp p. 18.400 T renaeus azittu*
Cancer magister 14.029 T P merguiensis 54.147 T
Cancer pagurus 30.480 T P. duorarum 15.512 T
Portunus trituberculatus 33.783 T P monodon 14.3/2 1
Callinectes sapidus 93.829 T P. setiferus 44.596 T
Paralithodes spp. 40.054 T P chinensis 2U./30 1
Chionoecetes opilio 44.650 T P kerathurus 16.063 T
Panulirus argus 40.926 T Metapenaeus spp. 35.536 T
Panulirus cygnus 11.264 T Pleoticus muelleri 16.226 T
Jasus spp. 16.399 T Xvpho- Plesiopenaeus 15.462 T
Nenhrops norvegicus 61.092 T Crangon crangon 54.203 T
Homarus americanus 53.436 T
Malgré cette progression globalement satisfaisante, pratiquement aucune espèce
de Crustacé n’occupe par rapport à l’ensemble des espèces exploitées dans les milieux
aquatiques de position privilégiée : ainsi, parmi les soixante dix espèces foumissant
chacune une production de 100.000 t ou plus, on ne trouve qu'un petit sergest.de bentho-pélagique de la mer de Chine et du Pacifique Nord—Ouest, Acetes japonicus,
(encore faut-il préciser que sous ce nom unique utilisé dans les statistiques se
dissimulent sans doute dautres espèces, comme Acetes chinensis), la crevette rose du
Groenland Pandalus borealis et le krill Euphausia superba, pour un total de 603.044 t.
Une trentaine d'autres espèces dont la production est compr.se entre 10.000 t et 100.000 t
totalisent plus de 815.000 t, soit un peu moins du quart du total.
235

La progression rapide des pêches de crevettes s'explique, au plan économique, par augmentation des prix sur le marché international et la demande élevée des pavs hautement industrialisés, comme les Etats-Unis et le Japon. L'introduction de la chaîne du troid a permis de lever les contraintes techniques qui s'opposaient au développement de ]a consommation de ces produits de conservation délicate. Parmi les principaux pavs producteurs de crevettes, on trouve aussi bien des pays industrialisés qui disposent à interieur de leur Zone Economique Exclusive de ressources considérables (par exemple es Etats Unis; quatrième producteur mondial) que des pays en voie de développement pour esquels les crevettes Penaeides constituent une source appréciable de devises (par exemple la Thailande ou le Mexique).
. Alors quelle etait a peu près inexistante, tout au moins dans les statistiques otncielles, il y a à peine plus d'une trentaine d'années, l'aquaculture des crevettes principalement des crevettes Penaeides et de Macrobrachium, se développe réguliè’-rement depuis l’obtention d'un certain nombre de résultats scientifiques qui ont fait I objet de recentes mises au point.
Le tableau 4 foumit la Iiste des dix premiers pays producteurs.
TABLEAU 4. LISTE DES DIX PREMIERS PAYS PRODUCTEURS DE CREVETTES POUR L'ANNÉE 1985.
I. CHINE 229.150 T 2. INDE 198.050 T 3. THAILANDE 161.570 T 4. ETATS UNIS 155.805 T 5. INDONESIE 114.630 T 6. NORVEGE 91.123 T 7. MEXIQUE 77.938 T 8. MALAISIE 69.004 T 9. BRESIL 67.460 T 10. JAPON 59.807 T TOTAL 1.224.537 T
,Dans le cadre de ce travail, il est nécessaire de rappeler de façon très succincte les principales filières d'élevage actuellement utilisées et surtout de situer Ia production aquacole actuelle et ses perspectives à court et moyen terme par rapport à l’exploitation des stocks sauvages de crevettes Penaeides..
Les élevages de crevettes Penaeides ont débuté il y a plusieurs centaines d'années dans des lagunes aménagées d'Indonésie et d'autres pays d'Extrême Orient. Tirant parti des tropismes migratoires des jeunes crevettes nées en mer, l'homme a appris très tôt à rctemr les animaux dans les lagunes saumâtres où ils pénétrent, puis à les pêcher au moment ou, devenus adultes, ils cherchent à regagner la mer. Ces techniques fondées sur le recrutement naturel sont encore pratiquées avec succès dans différents pays et sont meme à l'origine de développements récents : ainsi, la pêche spécialisée de post-larves de Penaeides (Penaeus vannamei et P. stylirostris essentiellement) le Iong des cotes de l'Equateur mobilise près de 100.000 "larveros".
La découverte du cycle biologique des Penaeides quelques années avant la deuxieme guerre mondiale et les progrès scientifiques et techniques ultérieurs ont conduit à une méthode d’obtention des post-larves fondée sur la capture de femelles sauvages parvenues à maturité. Lorsque l'espèce désirée n'existe pas naturellement dans la région d'élevage, il faut évidemment contrôler la totalité du cycle biologique en captivité. Cela a été obtenu de façon fiable il y a une douzaine d'années environ chez Penaeus. japonicus , puis appliqué à d'autres espèces. La méthode fait appel, séparément ou en combinaison, au contrôle de la maturation par les facteurs externes et à 1 épédonculation. Selon les techniques, plus ou moins stressantes, les rendements
236

obtenus varient dans d'importantes limites et peuvent atteindre 100 pour cent par l'emploi de la méthode écophysiologique (A. LAUBIER, 1986).
TABLEAU 5. PRODUCTION MONDIALE DE CREVETTES D'ÉLEVAGE, PROGRESSION ENTRE 1975 ET 1983 ET PERSPECTIVES.
1975 1983 1990 PENAEIDES 17.000 T 116.110 T 200-300.000 T MACROBRACHIUM 1.000 T 3.000 T 10.000 T
Le tableau 5 indique la production totale de l'aquaculture de crevettes Penaeides et Caridés et ses perspectives à moyen terme. L'élevage des crevettes Caridés est facilité, en ce qui concerne l'obtention des jeunes individus, par les particularités biologiques de ce groupe (oeufs portés par les femelles, absence de stade nauplien libre). En revanche, d'autres difficultés liées au cannibalisme plus ou moins élevé de ces crevettes ont limité le développement à quelques espèces, parmi lesquelles Macrobrachium rosenbergi domine Iargement.
L’analyse des principaux pays producteurs, pour l'année 1983, montre l'importance prise par quelques espèces (Penaeus monodon, P. vannamei et P . stylirostris) ainsi que les progrès extrêmement rapides enregistrés dans des pays qui ne disposaient d’aucune tradition ancienne (l'Equateur en est le meilleur exemple).
TABLEAU 6. LES PRINCIPAUX PAYS PRODUCTEURS DE CREVETTES PENAEIDES D'ÉLEVAGE EN 1983.
EOUATEUR 35.600 T Penaeus vannamei, P. stylirostris INDONESIE 11.900 T Penaeus monodon, P. merguiensis INDE 15.000 T Penaeus indicus, P. monodon VIETNAM 15.000 T Penaeus monodon THAILANDE 14.200 T Penaeus monodon, P. merguiensis FORMOSE 10.941 T Penaeus monodon PHILIPPINES 4.250 T Penaeus monodon BANGLADESH 3.000 T Penaeus monodon JAPON 2.000 T Penaeus iaponicus PANAMA 2.000 T Penaeus vannamei, P. stylirostris CHINE 1.600 T Penaeus orientalis BRESIL 1.000 T Penaeus schmitti, P iaponicus PEROU 1.000 T Penaeus vannamei, P. stylirostris SRI LANKA 500 T Macrobrachium rosenbergi BIRMANIE 500 T Penaeus monodon HONDURAS 500 T Penaeus vannamei, P. stylirostris MALAISIE 200 T Penaeus monodon FRANCE (DomTom) 70 T Penaeus stylirostris. P. monodon COREE 50 T Penaeus orientalis DIVERS TOTAL 1
500 T 16.110 T
Diverses espèces
Les perspectives à moyen terme (1990) indiquées dans le tableau 5 peuvent paraître élevées. Cependant, certains spécialistes comme l'américain C. MOCK, connu pour la mise au point de la technique d'élevage larvaire en eau claire - dite technique de Galveston - se montrent encore plus optimistes et n'hésitent pas à envisager une
237

progression de plus de 13 fois la production de 1985 en moins de dix ans, dans le cas des pays d'Asie où toutes les conditions techniques, écologiques, foncières, économiques et humaines paraissent réunies. C. MOCK est en revanche plus réservé en ce qui conceme l'Amérique du Sud où il prévoit durant la même période un accroissement de une fois et demi seulement par rapport à la production actuelle. En Amérique centrale et en Amérique du Nord, on prévoit un développement assez marqué, la production pouvant dépasser en 1990 le cap des 50.000 tonnes.
Concernant Macrobrachium rosenbergi, les progrès seront moins rapides, ne serait-ce que par suite de la limitation de la fécondité des femelles, qui accroît sensiblement les manipulations d'animaux dans les écloseries et conduit à des stocks de géniteurs importants.
Pour illustrer ces quelques considérations, le développement récent de l'aquaculture de crevetles Penaeides en Equateur fournit un exemple particulièrement démonstratif des potentialités de cet élevage, lorsque les conditions écologiques, techniques et économiques sont réunies. Alors que ce pays n'avait aucune production aquacole il y a moins de quinze ans, les élevages se sont très rapidement développés, d'abord à partir de la production naturelle de post-larves collectées dans la nature, puis avec l'appoint d'écloseries de grande capacité. Le tableau 7 illustre la véritable explosion qu’a connue l’aquaculture de crevettes dans ce pays.
En octobre 1986, sur un total de 105 écloseries autorisées par le gouvemement, 30 étaient effectivement en fonctionnement et 15 autres prêtes à entrer en fonction-nement ; 22 installations étaient en cours de constmction. La production annuelle de post-larves en écloserie atteignait alors 500 millions de post-larves, soit à peu près 5 pour cent de la quantité totale utilisée par les éleveurs. En ce qui conceme les bassins, la surface autorisée représentait finl985 92300 hectares, c'est-à-dire plus du double de la surface réellement utilisée. Avec les techniques actuellement disponibles, c'est 100.000 hectares au moins de terres qui peuvent être aménagés en bassins d'élevage. II faut enfin noter qu’au plan socio-économique, les chiffres de production masquent de grandes disparités et de nombreux échecs. A l'heure actuelle, la production annuelle par l'aquaculture atteindrait les 50.000 tonnes de crevettes, ce qui confirmerait les tendances constatées au cours des demières années.
TABLEAU 7. LE DÉVELOPPEMENT DE L'AQUACULTURE DE CREVETTES PENAEIDES EN EQUATEUR
PRODUCTION PECHE ELEVAGE SURFACE DE BASSINS 1972 6.800 T 6.800 T - -
1974 6.500 T 6.500 T - -
1975 7.500 T 7.500 T - -
1976 9.000 T 7.830 T 1.170 T 363 Ha (Autorisés) 1978 9.700 T 4.958 T 4.215 T 5.800 Ha (Utilisés) 1980 17.500 T 7.800 T 9.180 T 12.600 Ha " " 1982 29.500 T 8.000 T 21.500 T 29.573 Ha " ” 1983 44.600 T 8.900 T 35.600 T 49.000 Ha " " 1984 39.900 T 6.300 T 33.600 T 46.200 Ha " " 1985 36.228 T 6.000 T 30.205 T 41.547 Ha ” "
On peut donc prévoir que l'aquaculture de crevettes Penaeides poursuivra son développement et atteindra avant la fin du siècle le dixième de la pêche mondiale de
238

Crustacés. II s'agit d'un produit de haute valeur marchande, très recherché par les pays industrialisés et qui correspond bien à l'évolution du goût du consommateur.
En ce qui conceme les pêches, on doit souligner le niveau d exploitation très élevé de la plupart des stocks connus. Certes, des mesures d'aménagement, en particulier concemant les stocks de crevettes Penaeides pêchées à la fois en mer à l’état adulte et en lagunes lorsqu'elles sont jeunes, peuvent avoir des conséquences importantes sur ces formes dont le cycle de vie est court. En revanche, la plupart des stocks de Crustacés des mers tempérées et froides ont des vitesses de croissance relativement lentes et paraissent totalemcnt exploités. II ne faut cependant pas oublier la tendance des experts réunis il y a une vingtaine d'années à sous-estimer les ressourccs potentielles.
Par contre, les perspectives attrayantes offertes par le krill antarctique il y a une quinzaine d’années ont été progressivement réduites à néant : il est vrai que la biomasse de ce stock, à la suite de la destruction des populations de grands Cétacés et malgré la prédation accrue exercée par d'autres groupes animaux (Oiseaux, Poissons, Céphalopodes), représente plus de 500.000.000 de tonnes (résultats du programme intemational BIOMASS) ; il n'en reste pas moins que d’importantes contraintes grèvent exagérément l'exploitation accessibilité et disponibilité des stocks de krill, dont la répartition en bancs est encore mal comprise, difficultés de traitement, de conservation et de transformation du produit, valeur marchande faible. Ainsi, l'exploitation de l une des rares ressources potentielles dont l'importance avait été soulignée par de nombreux spécialistes apparaît aujourd'hui et sans doute pour de nombreuses années comme une véritable impasse.
F. A. O., 1971. The fish resources of the ocean, R: Crustacean resources, J.A. GULLAND edit., Fishing News (Books) Ltd publ., 206-245.
F. A. O., 1981. Atlas des ressources biologiques des mers. Collection F.A.O. : Pêches, i-x, 1-23, 73 cartes.
F. A. O., 1987 Annuaire statistique des pêches, captures et quantités débarquées,60, i-viii, 1-461.
LAUBIER, A., 1986. Les crevettes Penaeides. In L'aquaculture, G. BARNABE édit., 1, Les crevettes Penaeides, 459-491.
MC PADDEN, Ch. A., 1985. Breve estudio de la industria camaronera del Escuador. Boletin Cientifico y Technico, Instituto Nacional de Pesca, 8, 1, 1-68.
239

ASPECTS RÉCENTS de la Biologie des CRUSTACÉS Actes de Colloques - 8 - 1988 - IFREMER, pages 241 à 246
L'AQUACULTURE DES CREVETTES, ROLE DE LA NUTRITION
J. GUILLAUME
RESUME - L1 aquaculture des crevettes se développe rapideraent et représente deja 80 % de la consomraation mondiale. Elle porte principalement sur des especes variees de peneides et se pratique selon des techniques tres diverses où 1 ' alimentation joue un role tres inégal. Ce rôle est discute dans le cas des elevages plus ou moins extensifs et des elevages intensifs. L'accent est mis sur les carences dont souffrent nos connaissances sur la nutrition proprement dite des crustacés et sur 1 ' importance de la technologie alimentaire pour la fabrication des granules pour juveniles et des micropartiucles ou microcapsules pour larves.
mots-clés : crevette, nutrition, alimentation
SUMMARY - Shrimp on prawn aquaculture is developping fast and amounts presently to 8 % of world consumption. It concerns mainly various species of penaeids and is done with different techniques where feeding plays a variable role. This role is discussed both in the case of more or less extensive farming and in the case of intensive farming. The lack of knowledge in the field of nutritjon itself is emphasized as well as the importance of feed technology both for juvenile foods and larval microcapsules or microparticules.
key words : shrimp, prawn, nutrition, feeding
La production mondiale de crevettes d'aquaculture n'est pas connue avec precision, les statistiques de plusieurs pays etant dans ce domaine soit fantaisistes soit inexistantes. On evalue néanmoins le tonnage total a 120-150 000 T (1986) soit 8 % environ de la consommation et on prévoit que ce tonnage devrait doubler d'ici 1990.
Cette production porte essentiellement sur trois genres : Macrobrachium, Metapenaeus et surtout Penaeus represente par une demi-douzaine d'especes principales, élevees le plus souvent dans leur aire d'origine (ex. P. monodon dans la région indo-pacifique, P. vannamei et P. stylirostris en Amerique, P. orientalis en Chine, etc...) mais parfois aussi transplantées (ex. P. japonicus en Europe, P. vannamei a Tahiti ou en Nouvelle Calédonie).
Laboratoire Nutrition et Elevage, IFREMER Centre de Brest, BP 337, 29273 BREST Cedex, France.
241

Ces diverses espèces font l'objet d'élevages de nature très variée : en Asie du Sud-Est (Malaisie, Indonésie, Philippines, Thaïlande) les elevages traditionnels, à caractère très extensif dorainent, mais évoluent assez rapidement de nos jours ; en Amérique latine on crée sur des milliers d'hectares de vastes bassins destinés à l'élevage semi-intensif ; en France, sur une echelle encore tres modeste, on aménage d'anciens marais salants ou 1 ' on pratique un élevage que 1 ' on essaie d ' intensifier progressivement. Au Japon et a Taiwan on ne trouve guère que des élevages carrement intensifs ou la production dépasse parfois 5 T/ha au lieu d'une ou quelques centaines de kg dans les "tambaks" indonésiens ; signalons enfin que l'on experimente dans plusieurs pays des systèmes ultra-intensifs dits "moulinettes" ou l'on peut obtenir l'équivalent de 30 T par ha.
L'alimentation des crevettes pose des problèmes très différents selon l'espece elevee et surtout selon le mode de culture. En effet les besoins nutritionnels, et en particulier le besoin en protéines varient fortement d ' une espece a l'autre ; de plus et surtout la nourriture peut très bien servir d'abord a fertiliser le bassin ou nourrir les proies dans les elevages extensifs alors qu'elle doit imperativement fournir directement à la crevette tout ce dont elle a besoin dans les élevages intensifs.
Pour repondre a la demande d'un élevage récent mais en plein developpement une industrie de fabrication d'aliments est née, qui s'appuie sur des recherches fondamentales ou appliquées qui ont fait l'objet de plusieurs revues récentes (New, 1980 ; Guillaume, 1986*) sans essayer de resumer ces syntheses nous donnerons un aperçu des problèmes rencontrés dans le domaine de la nutrition des crevettes d'aquaculture en insistant sur quelques axes de recherches en cours.
1. L'ALIMENTATION DANS LES ELEVAGES SEMI-INTENSIFS
Quand les crevettes disposent a la fois de nourriture naturelle, proies vivantes, végétation, dérivés organiques divers, et de granulés, il est difficile d'estimer la quantité de granulés qu'il convient de distribuer (d'autant plus que 1'AD connaît souvent mal la biomasse des animaux d'elevage) ; il est plus difficile encore de déterminer la composition théorique idéale de l'aliment qui sert de complément à la nourriture naturelle. Dans la pratique on a recours soit à des aliments couvrants aussi bien que possible des besoins de l'espèce (aliments destinés à l'élevage intensif) soit à des aliments simplifiés, meilleur marche, fabriqués le plus souvent selon des normes voisines mais avec des matieres premieres de qualite inférieure. L'expérience montre que la distribution de ces aliments augmente bien la croissance des crevettes, mais sans que 1 ' on sache si les granulés sont ingérés directement par les crevette ou s'ils le sont d'abord par d'autres organismes voire s'il sont degrades par les bacteries.
Pour réaliser des progrès dans ce domaine plusieurs tentatives sont faites actuel lement, visant une quantification de la part respective des nourritures naturelle et artificie lle . La méthode classique des contenus stomacaux est bien entendu utilisée mais elle est laborieuse chez les crevettes peneides ou tous les aliments sont finement broyés et ne donne que des resultats instantanés. A l ' inverse la méthode dite du Delta carbone, reposant sur les variations des teneurs en C12 et C13, offre l'avantage de permettre une quantification dès la matière assimilée
* Nous renvoyons egalement et surtout à la''Nutrition des Crustacés et des Insectesf Act.es du Colloque CNERNA-CNRS (en cours de publication).
242

provenant de divers maillons de la chaine alimentaire d'une part, de la
nourriture artificielle d'autre part (à condition que l’on fabrique des
aliments ayant un rapport isotopique différent de celui de la nourriture naturelle, bien entendu). Cette méthode initiée par Schroeder (1983) en Israël est actuellement appliquée au suivi d'élevages de crevettes aux Etats Unis et en France. Elle devrait permettre une meilleure vision de l'alimentation et de la nutrition des crevettes dans les bassins d'élevage.
2. LA NUTRITION EN ELEVAGE INTENSIF
La nutrition des crevettes ne recevant aucune nourriture naturelle pose des problemes d'ordre scientifique mais aussi d'ordre technique et même technologique.
E n premier lieu, avant d ' aborder l'étude de la nature et du pourcentage des différents nutriments qu'il convient d'apporter à la crevette, il faut résoudre le problème de l'appétence et surtout de la stabilité à 1 ' eau des particules alimentaires que l'on fournit à l'animal. En effet ces derniers ne sont consommés que lentement, ils subissent une dissolution plus ou moins intense selon la nature des composants et ce "lessivage" peut rendre totalement illusoire la composition théorique du régime, affectant considérablement sa valeur nutritionnelle. Pour minimiser ces pertes deux approches sont possibles : on peut tout d'abord employer des liants ayant ou non une valeur nutritionnelle propre (par ex. gelatine, amidons transformés pour la premiere categorie, carboxymethylcellulose, colle formol-uree* pour la seconde) ; on peut aussi utiliser un procede particulier d'agglomération des particules : la cuisson-extrusion. La maitrise de ces procedes est un prealable a toute fabrication d'aliment "crevette" de qualite (Cuzon et Guillaume, 1987).
Au plan nutritionnel strict les crevettes posent des problèmes d'une toute autre ampleur. Les besoins de ces animaux sont, certes, connus dans leurs grandes lignes. On peut également dire que, pour l'essentiel, les grands principes de la nutrition connus chez les vertebres sont applicables aux crustacés. Mais il existe UIJ certain nombre de particularites : la plupart des crevettes ont, dans leur tube digestif, une activité chitinasique et meme parfois cellulasique encore mal quantifiees ; toutes semblent capables de synthétiser aisément les vitamines A et D à partir de carotenoides varies et de cholesterol respectivement ; mais toutes sont incapables de synthétiser le cholestérol ; les phospholipides alimentaires exercent un effet bénéfique sur l'absorption des lipides et en particulier du cholestérol ; la glucosamine, precurseur de la chitine, aurait egalement une action favorable ; l'absence d'estomac vrai et de digestion acide
modifie 1'utilisation de certains élèments et en particulier des minéraux. On pourrait citer un certain nombre d'autres particularites dont les conséquences sont encore souvent mal perçues.
Quelques besoins quantitatifs ont été estimés chez les espèces les plus courantes et en particulier le besoin en protéines qui est du meme ordre de grandeur que celui des poissons mais varie notablement d'une espèce à l'autre, les espèces carnivores ayant tendance a etie plus exigeantes que les especes omnivores. Les crevettes ont egalement une faible tolérance vis à vis des glucides, tendance "diabétique" rappelant
* produit interdit en alimentation animale dans la CEE mais utilise en Extrême-Orient.
243

celle des poissons, et elles ont a peu pres les mêmes besoins en acides gras essentiels que ces derniers, mais, à l'inverse des téléostéens, elles paraissent incapables de tirer parti de régimes rihces en lipides car elles n'ont que des reserves lipidiques très limitées et leur métabolisme energetique repose avant tout sur le catabolisme proteique.
On pourrait suposer que ces connaissances suffisent à l 'élaboration d'aliments couvrant les besoins des crevettes pourvu que l 'on prenne soin, comme il est d'usage en pareil cas, d'ajouter au régime des prémélanges vitaminiques et mineraux apportant à l'animal les composés "mineurs" dont on connait mal les besoins. L ' expérience montre que l'on peut à partir de matieres premieres distinctes formuler des régimes ayant la même composition theorique en protéines, lipides ou glucides et des marges de securite confortables en nutriments "mineurs" qui présentent des efficacites tres differentes. Ce constat démontre l'existence de lacunes dans nos connaissances et nous a amené à rechercher s'il n'existait pas des "facteurs nutritionnels" particuliers aux crevettes. L'efficacité de la farine de calmar ne s'expliquant pas par sa teneur en acides aminés ou en autres nutriments connus , ni par son rôle appétent nous avons émis l'hypothese de l'existence d ' un "facteur de croissance inconnu" Xocalisé dans la fraction protéique (Cruz et Guillaume, 1983). Des recherches ulterieures ont montré que ce facteur stimulait de 30 à 50 % (et même parfois davantage) la croissance de japonicus, P. vannamei, P. monodon et surtout P. stylirostris, mais non P. indicus (Cruz et al., 1987). Ce facteur a ete fortement concentre, des fractions actives à 200 et même 100 ppm ayant été obtenues récemment. Toutefois le mode d'action et la nature chimique exacte de ce facteur demeurent inconnus et de nouvelles recherches sont necessaires avant que l ' on puisse soit recenser les matieres premieres qui en contiennent soit synthétiser la molécule pour l'ajouter aux aliments commerciaux.
3. ALIMENTS POUR LARVES
L'élevage des larves de crevettes se fait encore presque sy stematiquement a partir d'aljg.ues unicellulaires puis d'artémies. Cependant depuis quelques années de grands efforts ont été faits pour elaborer des aliments inertes susceptibles de remplacer cette nourriture vivante et partant d'abaisser les coûts de production des postlarves. Ici encore on se heurte a un double problème : celui de la composition elle-meme des aliments et celui de la technologie des microparticules dont le diametre doit etre de quelques dizaines de micromètres et qui doivent malgre cela resister au lessivage. Deux techniques ont fait l'objet d ' experimentations a ce jour : la microencapsulation et 1 ' agglomération à l'aide de polyholosides tels que les carraghénanes.
II semble qu ' aujourd 'hui aucune de ces microparticules ne donne d'aussi bons resultats que la nourriture vivante : elles permettent, certes, dans des elevages commerciaux, un developpement aussi rapide et une survie aussi bonne que celles que l'on obtient avec la nourriture vivante, mais le poids des postlarves est fortement reduit, ce qui pourrait resulter d'une carence nutritionnelle qui reste a elucider. D'ores et deja on peut cependant supprimer totalement les algues dans les écloseries commerciales et l'economie qui en résulte est notable.
En conclusion, la nutrition constitue encore souvent un facteur limitant dans les elevages de crevettes, surtout dans les élevages intensifs, mais les recherches en cours, tant au niveau appliqué qu'au niveau fondamental , permettent des progres que se répercutent rapidement sur 1'aquaculture. On peut considerer que le developpement de cette
244

production actuellement en plein "décolage" et surtout son intensification seront étroitement lies a ces progres.
CRUZ-RICQUE L.E., 1987. Recherches sur la nature et le mode d'action d'un facteur de croissance extrait du calmar dans la nutrition des crevettes pénéides (Crustacea decapoda). These d'Universite, UBO.
CRUZ-SUAREZ L.E., GUILLAUME J.C., 1983. Facteur de croissance inconnu de la farine de calmar pour la crevette japonaise : localisation de ce facteur. CIEM CM 1983/F:14.
CRUZ-RICQUE L.E., CUZON G., GUILLAUME J. and AQUACOP, 1987. Squid protein effect on growth of four penaeid shrimp. J. World Aqac. Soc. (sous presse).
CUZON G., GUILLAUME J., 1987. Choix des matières premieres et fabrication des aliments destinés aux crevettes d'élevage. in La nutrition des crustacés et des insectes (sous presse).
GUILLAUME J., 1987. Besoins énergétiques, protéiques, vitaminiques et miné-raux chez les crustacés. in La nutrition des crustaces et des insectes (sous presse).
NEW M.B., 1980. A bibliography of shrimp and prawn nutrition. Aquaculture 21, 101-128.
SCHROEDER G.L., 1983. Stable isotopic ratios as naturally occuring tracers in the aquaculture food web. Aquaculture 30, 203-210.
245

ASPECTS RÉCENTS de la Biologie des CRUSTACÉS Actes de Colloques - 8 - 1988 - IFREMER, pages 247 à 250
PERSPECTIVES D'ACCROISSEMENT DE LA PRODUCTION DE L'OCEAN ATLANTIQUE PAR L'INTRODUCTION D'ESPECES DE CRUSTACES COMBLANT DES NICHES ECOLOGIQUES VIDES.
VASSEROT J.(1)
RESUME : Un certain nombre de biotopes particuliers de nos côtes seraient susceptibles d'accueillir des espèces importées et combler ainsi des niches écologiques vides. Plusieurs exemples d'introductions possibles sont présentés.
Mots-clés : Crustacés, production, introduction d'espèces.
ABSTRACT : It should be possible to increase the production of some areas of the east atlantic by introducing well adapted foreign marine species. Some example concerning crustaceans are given.
Key-words : Crustaceans, production.
En raison de son caractère beaucoup plus récent l'océan Atlantique présente une faune relativement pauvre. Cette pauvreté est plus marquée sur sa rive orientale, en Afrique et surtout en Europe. II paraît cependant possible d'accroître plus ou moins directement la production raarine consommable par l'homme en comblant des "niches écologiques" vides par des espèces judicieusement choisies dans d'autres zones biogéographiques de climat comparable.
Dans certains biotopes particuliers, très pauvres sur nos côtes, comme par exemple les plages fortement battues par les vagues pourraient s'établir notamment les Hippidés du genre Emerita , consommés par l'homme dans l'Inde et en Amérique centrale et constituant une ressource alimentaire très importante pour les oiseaux et certains poissons. Leurs antennes plumeuses filtrant les particules entraînées par les vagues sont éventuellement susceptibles, en diminuant l'hydrodynamisme au voisinage immédiat du fond, de freiner l'érosion par les vagues, la houle et les courants qui menace certaines côtes comme celles du Togo.
II peut s'agir également d'adaptations écologiques n'ayant pas d'équivalents exacts ailleurs : c'est le cas sur les côtes européennes (sauf dans le sud du Portugal) des crabes "écumeurs de sédiments" qui à mer basse râclent la surface des plages comme les Ucas des côtes nord-américaines et dans 1'Indo-pacifique les genres Macrophtalmus et surtout Scopimera , ce dernier étant particulièrement adapté à récolter des bactéries à la surface du sable, purifiant ainsi l'estran. Sur nos rivages pourraient s'établir bon nombre de ces espèces, notamment Scopimera globosa atteignant Hokkaido vers le nord.
(1) Station Biologique de Roscoff, Université Pierre et Marie Curie, 29211 ROSCOFF, France.
247

Dans le doraaine subtidal de petits crabes filtreurs sont susceptibles de rendre de grands services. Ainsi Pinnixa rathbuni , nourriture iraportante surtout pour les poissons plats. Certes les fonds de sables fins européens sont déjà pourvus de crustacés filtreurs du genre Ampelisca , raais ces Amphipodes sont très vulnérables aux pollutions pétrolières qui ont pour effet de diminuer considérablement cette ressource pour des années (Dauvin, 1986). Le "complément" aliraentaire constitué par des Décapodes serait très probablement moins sensible aux effets nocifs des "marées noires".
Dans le domaine terrestre Birgus latro représenterait au long des rivages intertropicaux une nouvelle ressource alimentaire (très riche en lipides), ce qui assurerait en même temps de meilleures chances de survie à cette espèce menacée par la surexploitation dans son habitat naturel. II faudrait toutefois s'assurer qu'il n'y a pas de risques de compétition nuisible pour les Crabes terrestres endémiques de l'océan Atlantique. En produisant en écloseries des juvéniles déjà parvenus au stade terrestre on alimenterait éventuellement des élevages familiaux de ce Crustacé savoureux, en utilisant les déchets domestiques et ceux de la culture et de l'élevage.
Les lacunes de la faune atlantique concernent aussi le domaine de la taille des animaux : ainsi les Balanes de nos cStes sont bien trop petites pour la consommation humaine, mais il n'en est pas de même pour les espèces géantes Balanus nubilis du nord-ouest de l'Amérique et surtout Balanus psittacus du Pérou et du Chili qui sont très recherchées corame nourriture, et pourraient se prêter éventuellement à l'aquaculture. Ces Cirrhipèdes coexistant déjà dans leur milieu naturel avec des Balanes plus petites de la même taille que celles vivant dans l'Atlantique, il n'y aurait pas à craindre une compétition nuisible pour celles-ci. D'ailleurs les murailes de ces espèces géantes offriraient de nouveaux supports et abris à la faune indigène (notamment aux jeunes crustacés s'abritant à l'intérieur des Balanes mortes).
Ce problème de taille se pose aussi dans le domaine pélagique : ainsi les Galathéides du genre Pleuroncodes forment au large des cStes du Mexique au Pérou, des essaims de crustacés nageurs plus avantageux par leur taille (4,5 centimètres) que les Euphausiacés et surtout les Copépodes, pour de grands prédateurs pélagiques comme les thons et les baleines. Leur introduction serait donc utile dans l'Atlantique s'il n'y existe pas déjà de crustacés présentant les mêmes caractéristiques écologiques, car elle raccourcirait les chaînes alimentaires planctoniques, diminuant ainsi très avantageusement les pertes de matière vivante qui se produisent d'un échelon à l'autre : de plus
( les prédateurs gaspilleront moins d'énergie à la
recherche de leur nourriture, leur appétit étant satisfait plus rapidement avec un plus petit nombre de proies de plus grandes dimensions.
Des perspectives particulièrement intéressantes concernent la récupération de "culs-de-sac" à différents niveaux des chaînes alimentaires. Ces introductions présentent un autre avantage : en transformant les Ulves et les Sargasses en nourriture pour des animaux coraestibles des crustacés du nord du Japon et de l'Amérique (Amphipodes essentiellement) permettraient de contrôler la pullulation envahissante de ces algues.
A des échelons supérieurs de la chaîne alimentaire le très grand anomoure Paralithodes camtschactica constitue, sur les fonds meubles
248

baignés par les eaux froides de l'Atlantique boréal, une nouvelle ressource
alimentaire de grande importance économique en récupérant des "culs-de-sac
alimentaires" comme le clypéastride Echinarachnius parma , les Ophiures
(notamment Ophiura sarsi au large du Canada oriental) et les Ascidies
Pelonaia corrugata et Boltenia echinata . Plus au sud pourrait
s'établir, sur les fonds rocheux, l'espèce japonaise Paralithodes
brevipes , plus eurytherme et susceptible éventuellement de contrôler les
pullulations de l'oursin Strongylocentrotus drobachiensis qui ravage
souvent les peuplements d'algues ; il faudrait faire des recherches pour
savoir s'il peut aussi "récupérer" les énormes populations constituées par
les Ophiothrix au nord de la zone occupée par les Langoustes. Et en limite
sud il peut constituer une nouvelle ressource alimentaire pour le Homard.
En zone tempérée (où l'eau ne dépasse guère +20°C) la grosse crevette
japonaise littorale Pandalus kessleri peut accroltre considérablement
la production, insuffisante pour la consommation nationale en France et aux
Etats-Unis, en utilisant certaines espèces d'algues, de Spongiaires, de
Cnidaires et d'Ascidies qui ne consomme guère Palaemon serratus , la
"Crevette rose" qui vit, elle aussi, dans les herbiers de Zostères et les
peuplements de Laminaires : il n'y a donc pas de concurrence alimentaire à
redouter (une récupération plus indirecte des Hydraires, et aussi de
certains Bryozoaires éventuellement, serait possible par la très grande
Caprelle japonaise Caprella kroyeri , si du moins poissons et crevettes
la consomment volontiers).
Une crevette encore plus grande que P. kessleri , Penaeus
notialis (anciennement appelé : " Penaeus duorarum forme A") de l'est
des Etats-Unis, peut peupler les lagunes du sud de l'Europe, car cette
espèce d'après son extension géographique vers le nord est plus résistante
au froid que les autres pénéides. C'est aussi en milieu lagunaire et
estuarien que vit le savoureux crabe Callinectes sapidus de l'est de
l'Amérique (qui d'ailleurs est déjà présent en Méditerranée orientale...) :
malheureusement c'est un prédateur des jeunes huîtres... Ce problème ne se
poserait guère pour de grands Portunidés indo-pacifiques vivant sur fonds
meubles et littoraux en milieu franchement marin, et sans équivalent au nord
de la Mauritanie, bien que Portunus sanguinolentus passant par le Canal
de Suez se soit déjà implanté en Méditerranée orientale ; or P.
trituberculatus , absent des zones équatoriales mais atteignant le Sud de
Hokkaido, peut prospérer dans les eaux tempérées assez chaudes en été en
créant une nouvelle ressource pour la pêche européenne côtière. Et
justement les écloseries japonaises produisent beaucoup de juvéniles
exportables pour peupler l'Atlantique.
Dans les mangroves et estuaires intertropicaux peut vivre le très gros
crabe indo-pacifique Scylla serrata : il n'existe pas d'espèce d'aussi
grande taille peuplant ces biotopes dans l'Atlantique ; et, point très
important à noter, il est inoffensif pour les Peneides fréquentant les
mêmes milieux, d'après des observations australiennes.
Mais on peut assigner à ces introductions un autre but, plus
désintéressé : étendre artificiellement la répartition géographique
d'espèces d'un grand intérêt scientifique, mais naturellement très
localisées, pour mieux garantir leur survie. Tel est le cas de Linuparus
trigonus , Langouste primitive peuplant les fonds de sable et graviers en
zone subtropicale au Japon et surtout, sur la côte pacifique de l'archipel
nippon, du plus grand crabe du monde, Macrocheira kaempferi endémique de
249

ce pays : il faut des fonds en majorité raeubles mais présentant des profondeurs assez grandes proches de la côte pour que cette espèce puisse effectuer au cours du cycle annuel les migrations nécessaires à son développement. Une des régions les plus favorables paraît être notamment le sud-ouest de la péninsule ibérique, ainsi que, probablement, l'ouest de la Méditerranée. Un autre crabe géant, le plus lourd du monde puisqu'il peut atteindre une quinzaine de kilogrammes, vit dans les eaux plus tempérées entre 90 et 950 mètres de profondeur au sud de l'Australie et notamment dans le détroit de Bass : c'est Pseudocarcinus gigas , dont au moins les jeunes individus peuvent parfois remonter dans la zone des grandes algues brunes. C'est sur les côtes orientales de l'Atlantique, au long de la Péninsule Ibérique (et peut-être aussi en Mer d'Alboran...), ainsi qu'au sud-ouest et à l'extrême sud de l'Afrique qu'il me paraît avoir les plus grandes chances de s'implanter. Ces deux crabes géants, dont il faudrait étudier en détail le régime alimentaire, ont une chair très savoureuse et constitueraient une nouvelle ressource gastronomique s'ils parvenaient à constituer des populations assez importantes.
Deux exemples choisis dans le milieu terrestre sont très encourageants pour de telles entreprises : le premier se situe justement dans le détroit de Bass, domaine favori de Pseudocarcinus gigas ; dans une île on a introduit jadis des Koalas qui y faisaient défaut et y ont prospéré. Or, en Australie proprement dite, ces charmants marsupiaux sont maintenant menacés par une maladie sexuellement transmissible provoquée par des Chlamydias, et la population "artificielle" de cette ile n'étant pas contaminée, elle permet d'assurer l'avenir de cette espèce précieuse.
Le second exemple concerne le règne végétal : le "Cyprès de Monterey" Cupressus lambertiana est une espèce relique qui après les glaciations ne subsistait que sur une petite presqu'ile de Californie ; et maintenant il prospère sur les côtes occidentales de l'Europe : en Bretagne, ses effectifs dépassent de beaucoup ceux de son aire géographique originelle.
Evidemment chaque espèce représente un cas particulier nécessitant des études écologiques spécifiques pour déterminer ses potentialités, ses avantages et ses éventuels inconvénients. On peut éviter un des principaux risques, l'introduction de parasites, en ne lâchant en milieu naturel que des jeunes nés en écloserie, ou des larves si leur métamorphose est trop difficile à obtenir en captivité. Dans le cas des Péracarides dont les jeunes semblables aux parents mènent le même genre de vie, il faudrait séparer les nouveaux-nés de la mère dès leur sortie du marsupium.
DAUVIN J.C. (1987). Evolution à long terme (1978-1986) des populations d'Amphipodes des sables fins de la Pierre Noire (Baie de Morlaix, Manche Occidentale) après la catastrophe de l'Amoco Cadiz. Marine Environmental Research , 247-273.
250

ASPECTS RÉCENTS de la Biologie des CRUSTACÉS Actes de Colloques - 8 - 1988 - IFREMER, pages 251 à 255
CONTRIBUTION D'UN FILM BACTERIEN A LA CULTURE INTENSIVE DE DAPHNIA MAGNA
R.LANGIS(1,2) .D.PROULX(1,3) ,J.DE LA NOUE(1,3)et P.COUTURE 1,2)
RESUME . L'addition de parois à l'intérieur d'un bassin de culture de Daphnies favorise la colonisation bactérienne et conduit à un accroissement de la productivité de Daphnia .
Mots-clés : Bactéries, biofilm, production, Daphnia .
ABSTRACT : Laboratory scale experiments show that can increase of the biofilm in cultivation tanks improves significatively the productivity of Daphnia .
Key-words : Bacteria, biofilm, production, Daphnia .
INTRODUCTION
Le rô1e des bactéries dans 1'alimentation des daphnies est relativement bien établi (1-8) ; toutefois, les interactions qui s'établissent entre les différentes composantes (bactéries-microalgues-zooplancton) d'un système intensif d'élevage des daphnies sont loin d'être toutes connues. Comme les bactéries tendent à se concentrer à l'interface solide-liquide dans les systèmes aquatiques (9) , nous avons voulu vérifier si l'ajout de parois à l'intérieur d'un bassin de culture de daphnies, favorisant la colonisation bactérienne, pouvait conduire à un .accroissement de la productivité de Daphnia .
MATERIEL ET METHODES
Des cultures de Daphnia magna ont été menées en bassins de plexiglass (13 L), de surface intérieure de 2 476 cm2 ou de 7 616 cm2 (Fig. 1) , nettoyés quotidiennement ou non. Les bassins étaient maintenus sous un éclairage continu (0.3 klux à la surface) et à la température de 20°C. Les daphnies (population initiale de 500 individus/L) étaient alimentées quotidiennement en remplaçant la moitié du volume de culture (excluant les daphnies) avec une suspension de Scenedesmus obliquus (910 +- 148 mg poids sec). S. obliquus a été cultivée (20 L) en mode semi-continu à partir d'un effluent secondaire urbain (9-20 mg N-NH4+/L , 3-8 mg P-PO4 /L) sous des conditions contrôlées d'éclairage (30 klux ; 16 h lumière : 8 h noirceur) et de température (20°C). Une aération continue de 300 L/h pour les algues et de 900 ml/min) pour les daphnies assurait l'agitation des populations respectives. Le suivi des populations de daphnies (densité, biomasse) a été fait au moyen d'échantillons prélevés tous les 3-4 jours.
(1) Groupe de recherche en recyclage biologique et aquiculture (2) INRS-EAU, 2700 rue Einstein, Ste-Foy (Québec) Canada GIV 4C7 (3) Faculté des sciences et de génie, Université Laval, Ste Foy (Québec) GIK 7P4
251

Figure 1. Bassins d'élevage des daphnies. Figure 2. Production cumulée de Daohnia magna en cutture intensive. Bassin Q non colonisé; 0 non colonisé avec parois; + coionisé; ■ colonisé avec parois.
Figure 3. Récotte moyenne (112 d; n- 58) de Daphnia maona en cutture intensive. (Les bassins portant le mdme numéro ne ditfèrent pas significativement)
Figure 4. Poids unitaire moyen (112 d) de Daphnia magna en cuiture intensive.
252

Figure 5. Pourcentage d'azote inorganique sous forme NH+4 durant la culture intensive de Daphnia magna.
253

RESULTATS ET DISCUSION
Les résultats (Fig. 2-3) montrent que la présence des parois et du biofilm augmentent la densité, la biomasse et la récolte de D. magna par un facteur de quatre, alors que le pourcentage de daphnies ovigères (17 %) , le poids moyen des daphnies (Fig. 4) et la composition chimique (protéines, lipides, sucres et acides aminés) des biomasses de daphnies ne sont pas modifiées par la présence d'un biofilm important.
La productivité accrue dans les bassins comportant des parois additionnelles est explicable à la fois par un biofilm plus important et par un effet de paroi qui agit sur la distribution spatiale des microorganismes, des algues et des daphnies dans le bassin, permettant ainsi une meilleure utilisation des nutriments. Le rôle précis des microorganismes semble plutôt qualitatif et porte à la fois sur 1'alimentation de Daphnia et la détoxification du milieu de culture par nitrification.
Sur le plan de 1'alimentation, les bactéries procureraient à la daphnie principalement des substances chimiques (acides aminés, acides gras et vitamines) nécessaires à sa croissance et à sa reproduction (2-4). Hadas et al (7) ont de plus montré que les microorganismes jouent un rôle important dans la digestion des particules par Daphnia . Sur le plan de la détoxification du milieu, la présence du biofilm contribue significativement à l'élimination de l'ammoniaque produit par les daphnies (Fig. 5) ; or, on sait que ce déchet métabolique est toxique pour les daphnies lorsque la concentration approche 1.89-2.08 mg NH3/L (10), niveau rapidement atteint en culture intenkive.
CONCLUSION
Les différences significatives obtenues à l'échelle du laboratoire (13 L) montrent que la productivité de systèmes de culture de daphnies à plus grande échelle pourrait être accrue par une augmentation du biofilm présent sur des surfaces additionnelles dans les bassins.
1. TAUB F.B. et DOLLAR A.M. (1968). The nutritional inadequacy of Chlorella and Chlamydomonas as food for Daphnia magna . Limnol. Oceanogr. , 13 . 607-17. 2. D'AGOSTINO A.S. et PROVASOLI L. (1970). Dixenic culture of Daphnia magna Straus. Biol. Bull. , 139 : 485-94. 3. TEZUKA Y. (1971). Feeding of Daphnia on planktonic bacteria. Jpn J. Ecol. 21 (3-4) : 127-34. 4. KRYUCHKOVA N.M. et RYBAK V.Kh. (1976). Growth od Daphnia magna Straus in a medium enriched with dissolved organic matter. Hydrobiol. , 12 (2) : 48-52. 5. COVENEY M.F. , CRONBERG G. , ENEL M. , LARSSON K. et OLOFSSON L. ( 1977). Phytoplankton, zooplankton and bacteria - standing crop and production relationships in a eutrophic lake. Oikos. , 29 : 5-21. 6. MYRAND B. et DE LA NOUE J. (1982). Croissance individuelle et dynamique de population de Daphnia magna en culture dans les eaux usées traitées. Hydrobiologia , 97 167-77. 7. HADAS 0., BACHRACH U., KOTT Y. et CAVARI B.Z. (1983). Assimilation of E. coli cells by Daphnia magna on the whole organism level. Hydrobiologia , 102 : 163-9.
254

Reproduit par INSTAPRINT S.A. 1-2-3, levée de la Loire - LA RICHE - B.P. 5927 - 37059 TOURS Cedex
Tél. 47 38 16 04
Dépôt légal 3*™ trimestre 1988

Cet ouvrage illustre certains des développements récents de la biologie des crustacés, tels qu’ils ont été abordés lors de la Xème réunion des carcinologistes en juin 1987, au laboratoire de biologie marine de Concarneau.
Des articles de fond servent d’ossature aux communications présentées dans cinq domaines : morphogénèse et développement, hormones et métabolisme, contrôles et physiolo-gie, population et exploitation des espèces.
L’ensemble vise à donner les éléments d’une vision d'ensemble de la biologie d’un groupe aussi homogène dans son organisation que divers dans ses adaptations que l’est celui des crustacés.
Cet ouvrage s’adresse à tous ceux dont l’intérêt se porte vers la biologie, l’écologie des espèces mais aussi à ceux dont les objectifs sont plus directement associés à la gestion des ressources vivantes.
This volume aims to survey some of the recent advances in the Biology of Crustacea presented during the 10th meeting of Carcinology in June 87 at the Marine Station of the Collège de France, in Concarneau.
Background papers and reports account with five main thema : Morphogenesis and development, hormones and metabolism, controls and physiology, populations and exploitation of living ressources.
Throughout, the emphasis is on giving a global insight on a zoological group homogenous at the organization level whereas adaptatively diversified.
This volume should appeal to those interested in biology, ecology of the crustacean species and also to those more directly involved in the management of living ressources.
Service de la Documentation et des Publications (SDP)
IFREMER - Centre de Brest B.P. 70 - 29263 PLOUZANÉ
Tél. 98.22.40.13 - Télex 940 627 F
IFREMER ISSN : 0761 3961




















