
BIOLOGIE - Kaya Print
108
Colegiul Național „Sfântul Sava” Gerard Enache BIOLOGIE CLASA A IX-A SEMESTRUL I CAIETUL ELEVULUI_______________________________ 2014-2015
-
Upload
khangminh22 -
Category
Documents
-
view
6 -
download
0
Transcript of BIOLOGIE - Kaya Print

C o l e g i u l N a ț i o n a l „ S f â n t u l S a v a ”
Gerard Enache
BIOLOGIE CLASA A IX-A
SEMESTRUL I
CAIETUL ELEVULUI_______________________________
2014-2015
2 / 108
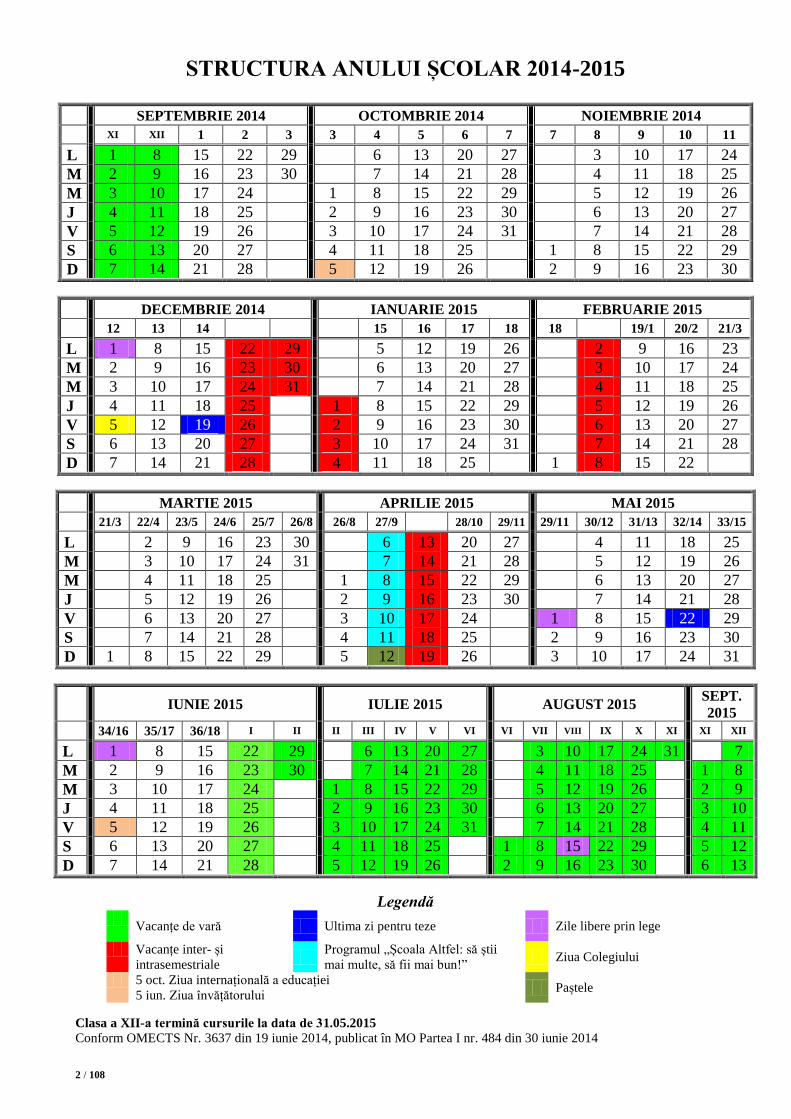
STRUCTURA ANULUI ȘCOLAR 2014-2015
SEPTEMBRIE 2014 OCTOMBRIE 2014 NOIEMBRIE 2014
XI XII 1 2 3 3 4 5 6 7 7 8 9 10 11
L 1 8 15 22 29 6 13 20 27 3 10 17 24
M 2 9 16 23 30 7 14 21 28 4 11 18 25
M 3 10 17 24 1 8 15 22 29 5 12 19 26
J 4 11 18 25 2 9 16 23 30 6 13 20 27
V 5 12 19 26 3 10 17 24 31 7 14 21 28
S 6 13 20 27 4 11 18 25 1 8 15 22 29
D 7 14 21 28 5 12 19 26 2 9 16 23 30
DECEMBRIE 2014 IANUARIE 2015 FEBRUARIE 2015
12 13 14 15 16 17 18 18 19/1 20/2 21/3
L 1 8 15 22 29 5 12 19 26 2 9 16 23
M 2 9 16 23 30 6 13 20 27 3 10 17 24
M 3 10 17 24 31 7 14 21 28 4 11 18 25
J 4 11 18 25 1 8 15 22 29 5 12 19 26
V 5 12 19 26 2 9 16 23 30 6 13 20 27
S 6 13 20 27 3 10 17 24 31 7 14 21 28
D 7 14 21 28 4 11 18 25 1 8 15 22
MARTIE 2015 APRILIE 2015 MAI 2015
21/3 22/4 23/5 24/6 25/7 26/8 26/8 27/9 28/10 29/11 29/11 30/12 31/13 32/14 33/15
L 2 9 16 23 30 6 13 20 27 4 11 18 25
M 3 10 17 24 31 7 14 21 28 5 12 19 26
M 4 11 18 25 1 8 15 22 29 6 13 20 27
J 5 12 19 26 2 9 16 23 30 7 14 21 28
V 6 13 20 27 3 10 17 24 1 8 15 22 29
S 7 14 21 28 4 11 18 25 2 9 16 23 30
D 1 8 15 22 29 5 12 19 26 3 10 17 24 31
IUNIE 2015 IULIE 2015 AUGUST 2015
SEPT.
2015
34/16 35/17 36/18 I II II III IV V VI VI VII VIII IX X XI XI XII
L 1 8 15 22 29 6 13 20 27 3 10 17 24 31 7
M 2 9 16 23 30 7 14 21 28 4 11 18 25 1 8
M 3 10 17 24 1 8 15 22 29 5 12 19 26 2 9
J 4 11 18 25 2 9 16 23 30 6 13 20 27 3 10
V 5 12 19 26 3 10 17 24 31 7 14 21 28 4 11
S 6 13 20 27 4 11 18 25 1 8 15 22 29 5 12
D 7 14 21 28 5 12 19 26 2 9 16 23 30 6 13
Legendă
Vacanțe de vară Ultima zi pentru teze Zile libere prin lege
Vacanțe inter- și
intrasemestriale
Programul „Școala Altfel: să știi
mai multe, să fii mai bun!” Ziua Colegiului
5 oct. Ziua internațională a educației
5 iun. Ziua învățătorului Paștele
Clasa a XII-a termină cursurile la data de 31.05.2015
Conform OMECTS Nr. 3637 din 19 iunie 2014, publicat în MO Partea I nr. 484 din 30 iunie 2014

3 / 108
Organizarea clasei – evaluarea An școlar 2014-2015
Evaluarea scrisă anunțată poate fi completată prin evaluare orală, referate, proiecte, portofolii, studii de caz, eseuri.
Indiferent de forma de evaluare, notarea începe întotdeauna de la nota 2 (doi) – notă care reprezintă punctele acordate din
oficiu și cunostințe nule. Nota 1 (unu) va fi acordată pentru fraudă, tentativă de fraudă sau intenție de fraudă la evaluarea scrisă (copiat la
lucrări din cărți, caiete, de la colegi sau folosind telefonul ori alte mijloace) sau/și orală sau/și la realizarea altor sarcini de lucru finalizate
prin notă.
Evaluarea scrisă se va desfășura conform programării de mai jos (care urmează a fi completată cu datele exacte consecutiv
stabilizării orarului). În cazul claselor cu două ore pe săptămână, testele și teza se susțin în prima sau a doua oră din săptămână, conform
programării.
În cazul evaluării scrise elevii se vor așeza în bănci în ordine alfabetică, începând cu rândul de la perete. Dacă există absenți,
locul acestora rămâne neocupat. Elevii vor primi, de regulă, instrument de scris, foaie de test și ciornă. Redactarea testului se va face doar
cu culoare albastră, respectând indicațiile de pe subiect sau/și ale profesorului.
Elevii nu se vor ridica din bănci până când nu au fost strânse, conform regulilor precizate, toate testele și instrumentele de scris
(eventual și ciornele). Dacă durata evaluării scrise este mai mică de 50 de minute, elevii vor rămâne în bănci, în aceeași ordine în care au
susținut testul, până la finalul orei/în momentul părăsirii sălii de clasă de către profesor.
Modul de predare a testelor (eventual și a ciornelor) și a instrumentelor de scris este precizat de către profesor. De regulă, cele
două teste de la o bancă se pun unul peste altul și se predau băncii din față; testele vin, sub formă de seturi de câte două, din spate în față,
la prima bancă, astfel aranjate încât setul corespunzător bănci din față să fie deasupra celorlalte seturi.
În timpul orelor de evaluare scrisă, elevii nu vor păstra asupra lor și în/pe/sub/lângă bănci, pe/sub scaun nici un obiect care ar
putea fi asociat cu frauda – penar, ciorne proprii, alte instrumente de scris, ghiozdan/geantă, caiete de orice fel, foi scrise, telefoane și alte
dispozitive electronice (și/sau accesoriile acestora) etc. Nerespectarea acestei reguli se sancționează direct cu nota 1 (unu).
În cazul clasei a XII-a, semestrul al II-lea are doar 15 săptămâni (9 februarie – 31 mai 2015).
La nota finală se va ține seama de prezența la cursuri, de existența și starea caietului de notițe, de participarea activă la oră,
de comportament și atitudine etc.
Programare evaluare scrisă
Tipul de
evaluare Semestrul I Semestrul al II-lea
Pentru clasele IX-XI cu 2 ore/săptămână, profil real
Test I S7-1 S6-1
Test II S14-1 S14-1
TEZĂ S14-2 S14-2
Pentru clasele cu 1 oră/săptămână – profil uman (a XI-a și a XII-a)
Test I S7 S6
Test II S14 S14 (a XI-a) S12 (a XII-a)
Clasa a XII-a – 2 ore/săptămână, profil real, cu sau fără teză
Test I S7-1 S6-1
Test II S14-1 S12-1
TEZĂ S14-2 S12-2
Studiul biologiei vizează:
pregătirea școlară temeinică pentru integrarea optimă în viața activă
educația pentru sănătate a generației tinere
formarea unui comportament ecologic la elevi
inițierea în specialitate, ca bază de orientare școlară și profesională
Din această perspectivă, predarea biologiei în ciclul inferior al liceului se realizează prin curriculum-nucleu (trunchiul comun),
obligatoriu de parcurs de către toți elevii, cât și prin curriculum diferențiat cuprinzând, în programă, conținuturi marcate cu scris italic și
asterisc, obligatoriu numai pentru filiera teoretică, profilul real.
Trunchiul comun reprezintă oferta educațională constând din aceleași conținuturi și sarcini de învățare pentru același număr de ore la
toate liceele și urmărește atingerea finalităților propuse pentru educația de bază, condiție pentru asigurarea egalității șanselor pentru toți elevii.
Curriculum-ul diferențiat reprezintă oferta educațională constând din disciplinele ariei curriculare corespunzătoare profilului liceului cu
alocări orare asociate acestora. În acest mod elevii pot beneficia, pe lângă studii comune, și de un parcurs diferențiat de învățare corespunzător
profilului pentru care au optat. Orele atribuite prin curriculum diferențiat sunt obligatorii, iar notele obținute de elevi sunt consemnate în unica
rubrică, din catalog, pentru disciplina studiată.
Prezentul curriculum este conceput într-o manieră flexibilă, care permite profesorului libertatea:
să aleagă singur succesiunea capitolelor sau a temelor, cu condiția ca aceasta să respecte logica internă a domeniului, să asigure în
final parcurgerea integrală a materiei și realizarea, la elevi, a competențelor propuse.
să construiască demersul didactic, în cadrul fiecărei lecții, în funcție de propria personalitate, de particularitățile clasei și de
caracterul experimental al biologiei.

4 / 108
COMPETENŢE GENERALE VALORI ŞI ATITUDINI
1. Receptarea informaţiilor despre lumea vie
2. Explorarea sistemelor biologice
3. Utilizarea şi construirea de modele şi algoritmi în scopul
demonstrării principiilor lumii vii
4. Comunicarea orală și scrisă utilizând corect terminologia
specifică biologiei
5. Transferarea și integrarea cunoştinţelor și metodelor de lucru
specifice biologiei în contexte noi
Interes pentru realizările şi descoperirile din domeniul ştiinţelor
Motivaţia pentru informarea şi documentarea ştiinţifică
Dezvoltarea curiozităţii şi respectului faţă de orice formă de viaţă
Grija faţă de propria persoană, faţă de ceilalţi şi faţă de mediul înconjurător
Dezvoltarea toleranţei faţă de opiniile altora
Cultivarea sensibilităţii
Interes pentru aplicarea cunoştinţelor de biologie în viaţa cotidiană
Implicarea în rezolvarea unor probleme de interes global
Listă de conținuturi
Capitolul I. CELULA – UNITATEA STRUCTURALĂ ŞI FUNCŢIONALĂ A SISTEMELOR BIOLOGICE (A VIEȚII)
Noţiuni introductive/Introducere în biologie
Compoziţia chimică a materiei vii*
Tipuri fundamentale de celule: celule procariote și celule eucariote
Structura, ultrastructura şi rolul (enunţarea funcţiei fără descrierea mecanismelor) componentelor celulare
Celula procariotă: structură, ultrastructură*
Celula eucariotă (animală, vegetală; structură și ultrastructură; procese celulare) a) înveliş celular:
- plasmalemă / membrană celulară (modelul mozaicului fluid) transport transmembranar: semipermeabilitate, transport pasiv (incl. osmoza și consecințele acesteia –
plasmoliza), transport activ
- perete celular - capsulă*
b) citoplasmă:
- fundamentală (citosol/hialoplasmă)
- structurală – organite celulare (+ rol):
ribosomi (sinteza proteinelor)
centrosom (formarea fusului de diviziune) reticul endoplasmatic (transport intra- și intercelular al substanțelor)
aparat Golgi (secreția substanțelor)
lizosomi (digestie intracelulară, fagocitoză) mitocondrii (cu criste, cu tubuli; respirație celulară)
miofibrile*
neurofibrile* corpusculi Nissl*
cili*
flageli* vacuole
plastide (fotosintetizante: cloroplaste, rodoplaste, feoplaste; nefotosintetizante: cromoplaste, leucoplaste –
amiloplaste, oleoplaste, proteoplaste; roluri: fotosinteza, depozitarea substanețelor de rezervă) - incluziuni (incl. incluziuni de săruri organice – oxalat de Ca)
- mișcări citoplasmatice
c) nucleu: - înveliș nuclear
- nucleoli - carioplasmă/nucleoplasmă:
cromatină (acizii nucleici – tipuri şi rol, cromosomi interfazici și metafazici)
Diviziunea celulară
Importanţă
Ciclu celular
Clasificare
a) diviziune directă (amitoză)* – modalități de realizare b) diviziune indirectă (cariochinetică)
- cromosomi şi fus de diviziune – număr, alcătuire şi rol
- mitoză (faze, importanţă) - meioză (etape, faze, importanţă; recombinarea intracromosomală și intercromosomală)
Capitolul II. EREDITATEA ŞI VARIABILITATEA LUMII VII Concepte: ereditate, variabilitate
Mecanismele transmiterii caracterelor ereditare
Legile mendeliene ale eredităţii Legea purităţii gameţilor
Legea segregării independente a perechilor de caractere
Importanţa legilor mendeliene*
Abateri de la segregarea mendeliană (semidominanţă*, supradominanţă*, gene letale*, codominanţă)
Teoria cromosomală a eredității Plasarea lineară a genelor în cromosomi*
Transmiterea înlănțuită a genelor* Schimbul reciproc de gene între cromosomii omologi
Recombinare genetică
Recombinare intracromozomală
Recombinare intercromozomală*

5 / 108
COMPETENŢE SPECIFICE ŞI CONŢINUTURI
Competenţe specifice Conţinuturi
Trunchi comun Curriculum diferenţiat
1. Receptarea informaţiilor despre lumea vie 1.1. Culegerea de date din surse
variate de informare/ documentare
despre organizarea lumii vii
Celula – unitatea structurală şi funcţională a lumii vii
Compoziţia chimică a materiei vii
1.2. Clasificarea indivizilor
biologici pe baza caracterelor
generale
Celula procariotă, celula eucariotă (vegetală, animală)
1.3. Identificarea structurilor
biologice microscopice în vederea
caracterizării lor
Structura celulei procariote
Structura şi ultrastructura celulei eucariote:
perete celular
membrană celulară/plasmalemă
citoplasmă
nucleu – membrană nucleară, nucleoli, carioplasmă, cromatină,
cromosomi (acizi nucleici – tipuri şi rol)
reticul endoplasmatic (neted, rugos)
aparat Golgi (dictiosomi)
ribosomi
lizosomi
centrosom
mitocondrii (cu criste, cu tubuli)
plastide (fotosintetizatoare: cloroplaste, rodoplaste, feoplaste;
nefotosintetizatoare: leucoplaste, amiloplaste)
vacuole
incluziuni
Miofibrile, neurofibrile, corpusculi Nissl, cili, flageli, capsulă
Ultrastructura celulei procariote
Oleoplaste, proteoplaste, incluziuni de săruri organice – oxalat de
calciu.
2. Explorarea sistemelor biologice 2.1. Utilizarea investigaţiei pentru
identificarea unor caractere generale
ale organismelor şi evidenţierea
componentelor şi proceselor
celulare
Celula – unitatea structurală şi funcţională a lumii vii
Cloroplaste, amiloplaste
Mișcări citoplasmatice
Mitoza
Osmoza, plasmoliza
Cromoplaste
Cromosomi metafazici
Meioză (etapele reducţională şi ecvaţională)
2.2. Prelucrarea rezultatelor obţinute
din investigaţii şi formularea
concluziilor
Celula – unitatea structurală şi funcţională a lumii vii: rolul
componentelor celulare:
transport transmembranar
fotosinteză
respiraţie
diviziune
digestie intracelulară – fagocitoză
sinteza proteinelor
transport intra- şi intercelular al substanţelor
secreţia substanţelor
formarea fusului de diviziune
depozitarea substanţelor de rezervă
Transport transmembranar activ
3. Utilizarea şi construirea de modele şi algoritmi în scopul demonstrării principiilor lumii vii 3.1. Reprezentarea lumii vii pe
baza modelelor
Celula – unitatea structurală şi funcţională a lumii vii
Amitoză
Recombinare intercromosomală
3.2 Aplicarea unor algoritmi de
identificare şi rezolvare de
probleme
Structura celulei
Procese celulare
Cromoplaste la morcov şi tomate
Evidenţierea incluziunilor ergastice: grăsimi, cristale de oxalat de
calciu
Evidenţierea diviziunii prin înmugurire la drojdia de bere
Evidenţierea cromosomilor metafazici la Allium, Vicia
Evidenţierea diviziunii meiotice la Secale sp., Triticum sp.
4. Comunicarea orală şi scrisă utilizând corect terminologia specifică biologiei 4.1 Utilizarea corectă a
terminologiei specifice biologiei în
diferite situaţii de comunicare
Diviziunea celulară indirectă:
mitoză (importanţă, structuri implicate – fus de diviziune,
cromosomi – număr, alcătuire şi rol, faze ale diviziunii:
profază, metafază, anafază, telofază, citochineză)
meioză (importanţă, etapa reducţională – profază I, metafază I,
anafază I, telofază I, etapa ecvaţională)
Recombinare genetică intracromosomală
Terminologie: celulă, procariot, eucariot, denumirile componentelor
celulare, semipermeabilitate, osmoză, plasmoliză, fagocitoză,
interfază, mitoză, meioză, haploid, diploid, ciclu celular,
cromosomi
Unitate structură-funcţie (componente celulare)
Unitate-diversitate (celulă)
Evoluţie de la simplu la complex (tipuri fundamentale de celule:
procariote, eucariote)
Amitoză – modalităţi de realizare
Recombinare genetică intercromosomală
Transport transmembranar activ
4.2 Prezentarea informaţiilor
folosind diverse metode de
comunicare
Structura şi ultrastructura celulei eucariote
Rolul componentelor celulare
Transport transmembranar activ
Lista lucrărilor practice*
observaţii microscopice asupra structurii celulei procariote şi eucariote
cloroplastele de la Elodea, mişcarea de rotaţie şi circulaţie a plasmei şi
cloroplastelor
cromoplastele de la morcov* şi tomate*
evidenţierea incluziunilor ergastice: amidon, grăsimi*, cristale de oxalat de
calciu*
evidenţierea celulelor stomatice
osmoza
plasmoliza şi deplasmoliza
evidenţierea diviziunii prin înmugurire la drojdia de bere*
evidenţierea diviziunii celulare mitotice la Allium cepa
evidenţierea cromozomilor metafazici la Allium sp.*, Vicia sp.*
evidenţierea diviziunii meiotice la Secale sp.*, Triticum sp.*
*Vor fi efectuate în limita timpului și a materialelor disponibile

6 / 108
CUPRINS Capitolul I. CELULA ............................................................................................................................................................................................... 9
Introducere în biologie ...................................................................................................................................................................... 11
Compoziția chimică a materiei vii* ................................................................................................................................................... 13 Generalități .................................................................................................................................................................................. 13
Elementele și compușii chimici esențiali ai materiei vii .............................................................................................................. 13
Compuși anorganici ..................................................................................................................................................................... 14 Compuși bioorganici ................................................................................................................................................................... 15
Generalități ........................................................................................................................................................................... 15
Glucide ................................................................................................................................................................................. 16 Lipide ................................................................................................................................................................................... 18
Proteine ................................................................................................................................................................................ 19 Acizi nucleici ........................................................................................................................................................................ 21
Structura ARN. Tipuri de ARN ..................................................................................................................................... 23
Celula – generalități ......................................................................................................................................................................... 24 Teoria/doctrina celulară .................................................................................................................................................................. 24
Definiția celulei ............................................................................................................................................................................... 24
Forma celulelor ............................................................................................................................................................................... 24 Dimensiunea celulelor ..................................................................................................................................................................... 24
Tipuri fundamentale de celule ...................................................................................................................................................... 24
Celula procariotă (PK) ..................................................................................................................................................................... 27 Generalități .................................................................................................................................................................................. 27
Elementele de (ultra)structură ...................................................................................................................................................... 27
Peretele celular bacterian ...................................................................................................................................................... 28 Plasmalema (membrana plasmatică) ..................................................................................................................................... 28
Spațiul periplasmic ............................................................................................................................................................... 28
Glicocalixul bacterian ........................................................................................................................................................... 29 Fimbriile ............................................................................................................................................................................... 29
Pilul ...................................................................................................................................................................................... 29
Flagelul ................................................................................................................................................................................. 29 Filamentele axiale ................................................................................................................................................................. 29
Citoplasma ............................................................................................................................................................................ 29
Ribosomii ............................................................................................................................................................................. 29 Materialul/aparatul genetic ................................................................................................................................................... 30
Incluziuni citoplasmatice ...................................................................................................................................................... 30
Lectură. Biofilme bacteriene ................................................................................................................................................ 30
Celula eucariotă (EK) ....................................................................................................................................................................... 35
Celula animală ............................................................................................................................................................................... 35
Plasmalema ............................................................................................................................................................................. 35 Ultrastructura plasmalemei. Modelul mozaicului fluid ..................................................................................................... 36
Funcțiile plasmalemei ....................................................................................................................................................... 37
Transport (trans)membranar .......................................................................................................................................... 37 Transportul pasiv .................................................................................................................................................... 38
Difuzia (difuzia simplă/termică, difuzia facilitată, osmoza) ................................................................................. 38
Filtrarea ............................................................................................................................................................... 41 Transportul activ ..................................................................................................................................................... 41
Transportul activ propriu-zis (transport primar, transport secundar/contransport) ............................................... 41
Transportul vezicular (exocitoză, endocitoză, trafic intracelular, transcitoză) ..................................................... 42 Transport (trans)membranar – rezumat ................................................................................................................... 44
Citoplasma .............................................................................................................................................................................. 45
Citosolul ............................................................................................................................................................................... 45 Organitele celulare ................................................................................................................................................................ 45
Organite comune ........................................................................................................................................................... 45
Organite corpusculare ............................................................................................................................................. 45 Ribosomii (granulele lui Palade) .................................................................................................................. 45
Centrosomul ................................................................................................................................................. 46
Citoscheletul ................................................................................................................................................. 46 Microtubulii .......................................................................................................................................... 46
Cilii și flagelii ................................................................................................................................. 46
Filamentele intermediare ....................................................................................................................... 47 Microfilamentele (filamentele de actină) ............................................................................................... 47
Microvilii ....................................................................................................................................... 48 Pseudopodele .................................................................................................................................. 48
Organite veziculare ................................................................................................................................................. 48
Reticulul endoplasmatic (RE) ....................................................................................................................... 48 REN ...................................................................................................................................................... 48
RER ....................................................................................................................................................... 48
Aparatul Golgi (dictiosomul) ........................................................................................................................ 49 Lizosomii ..................................................................................................................................................... 50
Peroxisomii .................................................................................................................................................. 50
Mitocondria .................................................................................................................................................. 50 Organite specifice .......................................................................................................................................................... 54
Miofibrilele ............................................................................................................................................................. 54
Neurofibrilele ......................................................................................................................................................... 54 Corpii Nissl ............................................................................................................................................................. 54
*Pseudopodele ........................................................................................................................................................ 54
*Cilii și flagelii ....................................................................................................................................................... 54 *Microvilii .............................................................................................................................................................. 54
Incluziunile citoplasmatice ................................................................................................................................................... 54
Nucleul .................................................................................................................................................................................... 55

7 / 108
Celula vegetală .............................................................................................................................................................................. 58 Vacuola ................................................................................................................................................................................... 58
Peretele celular ....................................................................................................................................................................... 59
Plastidele .................................................................................................................................................................................. 61
Tipuri de plastide .............................................................................................................................................................. 61
Structură generală ............................................................................................................................................................. 61
Plastidele algelor – cromatofori ......................................................................................................................................... 61 Proplastide ........................................................................................................................................................................ 61
Leucoplaste ....................................................................................................................................................................... 61
Cromoplaste ...................................................................................................................................................................... 61 Cloroplaste ........................................................................................................................................................................ 62
Reproducerea celulară ..................................................................................................................................................................... 63
Fisiunea binară la bacterii ............................................................................................................................................................ 63 Ciclul celular la eucariote ............................................................................................................................................................ 63
Organizarea celulară a materialului genetic ............................................................................................................... 63 Interfaza ................................................................................................................................................................................ 63
Faza M (diviziunea celulară – mitoza și citokineza) ............................................................................................................. 64
Rolurile/importanța diviziunii celulare ....................................................................................................................... 64 Tipuri de diviziune celulară ........................................................................................................................................ 64
Faza M mitotică ............................................................................................................................................................. 65
Diviziunea celulară și reproducerea sexuată ................................................................................................................................ 65
Reproducere asexuată și reproducere sexuată ....................................................................................................................... 66
Bazele citologice ale eredității și variabilității ...................................................................................................................... 66
Seturi de cromosomi în celulele umane ................................................................................................................................ 66 Faza M meiotică ................................................................................................................................................................... 66
Meioza I .............................................................................................................................................................. 67
Meioza II ............................................................................................................................................................. 67 Comparație între mitoză și meioză ........................................................................................................................................ 68
Sursele variației genetice ...................................................................................................................................................... 68
Capitolul II. ELEMENTE DE GENETICĂ ............................................................................................................................................................ 71 Introducere ........................................................................................................................................................................................ 73
Genetică factorială (mendeliană, clasică). Legile mendeliene ale eredității (Teoria factorilor ereditari) ................................... 73
Generalități .................................................................................................................................................................................. 73 Concepte și noțiuni ...................................................................................................................................................................... 74
Organizarea florii bisexuate/perfecte de la angiosperme ............................................................................................................. 74
Monohibridarea și primele două legi ale eredității ....................................................................................................................... 75 Dihibridarea și legea a treia a eredității ........................................................................................................................................ 80
Stabilirea numărului și genotipurilor gameților ............................................................................................................................ 83
Legile probabilităților și interpretarea matematică a rezultatelor lui Mendel ................................................................................ 83 Importanța legilor lui Mendel ....................................................................................................................................................... 84
Abateri de la segregarea mendeliană ............................................................................................................................................ 85
Dominanța completă sau ereditate de tip Pisum ............................................................................................................. 85 Interacțiuni alelice ................................................................................................................................................................ 86
Semindominanță (dominanța incompletă, ereditate de tip Zea) ..................................................................................... 86
Supradominanță (vigoare hibridă, heterozis) ................................................................................................................. 88 Codominanță ................................................................................................................................................................. 88
Gene letale ..................................................................................................................................................................... 93
Polialelie ........................................................................................................................................................................ 93 Interacțiuni genice (non-alelice) ........................................................................................................................................... 94
Epistazie ........................................................................................................................................................................ 94
Apariția unui fenotip nou cu păstrarea raportului de segregare ...................................................................................... 94 Poligenie ........................................................................................................................................................................ 94
Pleiotropie ............................................................................................................................................................................. 95
Parametri cantitativi ai fenomenului ereditar ............................................................................................................................... 95 Penetranță și expresivitate .................................................................................................................................................... 95
Heritabilitate ......................................................................................................................................................................... 95
Teoria cromosomală a eredității ...................................................................................................................................................... 99 Etape premergătoare .................................................................................................................................................................... 99
Tezele teoriei cromosomale a eredității ........................................................................................................................................ 100
Plasarea liniară a genelor în cromosomi ................................................................................................................................ 100 Transmiterea înlănțuită a genelor din același cromosom – teza linkage-ului ......................................................................... 100
Teza crossing-over-ului ........................................................................................................................................................ 100
Hărți genetice ................................................................................................................................................................. 102 Recombinarea genetică ..................................................................................................................................................................... 105
Recombinarea genetică la procariote ........................................................................................................................................... 105 Recombinarea genetică la eucariote ............................................................................................................................................. 105
Recombinarea intercromosomală (segregarea independentă a perechilor de omologi) ......................................................... 105
Recombinarea intracromosomală (crossing-over) ................................................................................................................ 105 Fecundația aleatorie .............................................................................................................................................................. 106
Conversia genică (schimb nereciproc de material genetic) ................................................................................................... 106
Recombinarea de tip V(D)J .................................................................................................................................................. 106 Recombinarea nespecifică/transpozițională .......................................................................................................................... 106

8 / 108

9 / 108
Capitolul I
CELULA

10 / 108

11 / 108
INTRODUCERE ÎN BIOLOGIE
Biologia (gr. bios – viață, logos – cuvânt, știință, legein – a selecta, a aduna) este știința fundamentală a naturii
care studiază legile generale ale apariției, organizării, dezvoltării și transformării (evoluției) materiei vii, cât și relațiile care
se stabilesc între organisme, între organisme și mediu, între ontogenie și filogenie; este știința care se ocupă cu studiul
originii, organizării și evoluției (transformării adaptative) materiei vii.
François Jacob (genetician, 17 iun. 1920 – 19 apr. 2013) considera că „viul se reduce la
o grămăjoară de materie pe care viața vine să o mângâie o clipă”, oferindu-ne astfel una dintre
cele mai metaforice și pline de conținut definiții ale materiei vii, subliniind în același timp și
relația dintre concretul material al viului și abstractul metafizic al vieții.
Termenul „biologie” a intrat în limbajul științific relativ recent, fiind utilizat pentru
prima dată de către Michael Christoph Hanov/Hanow (1695-1773, meteorolog, istoric, profesor
de matematică german) în titlul volumului al III-lea al lucrării Philosophiae naturalis sive
Physicae dogmaticae: Geologia, biologia, phytologia generalis et dendrologia, publicată în 1766.
Cu sensul modern, actual, termenul a fost utilizat independent de Thomas Beddoes (1760-1808, medic și profesor
englez) în 1799, Karl Friedrich Burdach (1776-1847, fiziolog german) în 1800 și de Gottfried Reinhold Treviranus
(1776-1837, naturalist german) în Biologie oder Philosophie der lebenden Natur și Jean-Baptiste Lamarck (Jean-Baptiste
Pierre Antoine de Monet, Chevalier de la Marck, 1744-1829, naturalist, profesor francez) în Hydrogéologie, în anul 1802.
Înainte de aceste momente, cunoștințele despre lumea vie, despre natură în general, erau reunite sub denumirea de
Istorie naturală, Filosofie naturală sau chiar Teologie naturală.
Curiozitatea și setea de cunoaștere l-au motivat și determinat pe om să cerceteze viul și viața. Și-a pus întrebări și a
căutat răspunsurile. Din strădaniile sale s-au acumulat, de-a lungul timpului, foarte multe cunoștințe din domeniul biologiei
care, pentru a fi mai ușor gestionate, utilizate și transmise, au fost grupate, în funcție de obiectul sau fenomenul studiat, pe
domenii. Au apărut astfel domenii și ramuri ale biologiei, dar și științe de graniță care reflectă strânsa legătură a științei
viului cu fizica, chimia, matematica, ingineria etc.
Dintre domeniile principale (fundamentale) ale biologiei fac parte:
Morfologia (gr. morfi, μοπυή – formă) se ocupă cu studiul formei, aspectului organismelor vii.
Anatomia (gr. ana, ανά – prin, de; tomē, τομή – secțiune) se ocupă cu studiul macro- si microstructurii
organismelor vii.
Fiziologia (gr. physis, υύσηρ – natură, fel de a fi) se ocupă cu studiul funcțiilor și funcționării organismelor
vii.
Zoologia (Biologia animală, gr. zoon, ζώων – animale) se ocupă cu studiul animalelor, mai puțin al omului.
Fitologia (Botanica, Biologia vegetală, gr. phyton, υςτων – plante, botanē, bοτανα/ε – plante furajere,
ierburi) se ocupă cu studiul plantelor.
Microbiologia (gr. micros, μικπόρ – mic, bios – viață) se ocupă cu studiul bacteriilor – bacteriologia – și al
microorganismelor eucariote, de care se ocupă algologia p.p., protozoologia etc.; din considerente istorice, uneori
include și vir(us)ologia, cu toate că virusurile nu sunt considerate vii.
Micologia (gr. mykes, μύκηρ – ciuperci) se ocupă cu studiul ciupercilor.
Genetica (gr. gennao, γεννάω – a ridica, génnisis, γέννησηρ – a da naștere) studiază ereditatea si
variabilitatea organismelor vii.
Ecologia (gr. oikos, οίκορ – casă, gospodărie) cercetează relațiile dintre organisme între ele și dintre
organisme și mediul lor de viață.
Sistematica studiază diversitatea organismelor, relațiile de înrudire (relații filetice/filogenetice) dintre
acestea și istoria lor evoluționară. Sistematica reunește taxonomia și filogenetica.
Taxonomia biologică (gr. táxi/s, τάξη/ρ – ordine, aranjare, clasă; nómos, νόμορ – lege, regulă, normă,
drept) este știința care elaborează și aplică principii și reguli de clasificare a organismelor și include
patru componente majore: descrierea, identificarea, nomenclatura/denumirea și clasificarea.
Filogenetica/filogeneza este domeniul biologic care are drept scop să descopere istoria evoluției unui
taxon de la apariția acestuia până în prezent, cât și relațiile de înrudire ale acestuia (filiația – strămoșii
și legăturile de rudenie dintre diverși taxoni).
Biologia celulară (citologia, gr. kytos – vas, celulă) – se ocupă cu studiul structurii, ultrastructurii,
funcțiilor și funcționării celulelor.
Antropologia (gr. anthropos, ανθπωπορ – om) – se ocupă cu studiul originii, evoluției, organizării și
culturii societăților omenești și ale oamenilor de la apariția lor până în prezent.
Ca ramuri ale domeniilor de bază (domenii devenite aproape științe autonome, unele chiar din momentul
delimitării obiectului de studiu, cum ar fi zoologia și botanica, unificate târziu prin teoria celulară a lui Schleiden,
Schwann și Virchow) mai pot fi enumerate: histologia (studiul țesuturilor), biologia dezvoltării (cu subdomeniul
embriologie), fitopatologia (studiul bolilor plantelor), algologia/ficologia (studiul algelor), prot(oct)istologia (studiul
prot(oct)istelor), parazitologia, neurobiologia, imunobiologia, virusologia, evoluționismul, zoopsihologia, etologia (gr.
ēthos, ήθορ – caracter; studiază comportamentul animalelor) etc.

12 / 108
Dintre științele de graniță mai cunoscute și mai importante sunt biochimia și biologia moleculară, biofizica,
biomatematica și biostatistica, biogeografia, paleontologia, biotehnologiile și ingineria genetică etc. Acestea asigură
corelații între științele biologice și științele medicale, farmacologie, industria alimentară ș.a.
Un domeniu recent apărut este bionica (biomimetică, bioimitare, bioinspirație, biognozie, inginerie bionică
creativă) – stiința care studiază structura și principiile de funcționare ale sistemelor biologice pentru a le putea aplica în
realizarea de mașini, aparate, mecanisme și procese asemănătoare acestora (sisteme inginerești și tehnologie). Natura este o
sursă inepuizabilă de idei și de modele care aplicate îmbunătățesc tehnica și cresc în consecință gradul de confort al omului.
Termenul „bionică” a fost introdus în vocabularul științific în 1958 de către Jack Ellwood Steel (medic militar american).
Exemple ale imitării în tehnică a sistemelor biologice:
Rețelele neuronale din encefal au fost model pentru rețelele cibernetice;
Orientarea delfinilor și a liliecilor prin intermediul ultrasunetelor a stat la baza funcționării radarului și a sonarului;
Forma hidrodinamică a peștilor și a mamiferelor acvatice a constituit model pentru construirea submarinelor și a
ambarcațiunilor plutitoare (vapoare, corăbii, bărci etc.);
Forma aerodinamică și zborul păsărilor se regăsesc în structura și funcționarea avioanelor;
Ochii telescopici ai peștilor abisali au fost imitați în relizarea telescoapelor (sisteme optice) și a altor mecanisme
care funcționează principiul extensiei mecanice (macarale, chiar și banala undiță de pescuit);
Ochii animalelor nocturne au stat la baza oglinzii reflectorizante;
Pielea delfinului a reprezentat model de înveliș pentru submarine (folosirea principiilor structurale ale
tegumentului delfinului se materializează prin scăderea forței de frecare la înaintare a submarinului);
Arhitectura paiului, a oaselor și fagurii au fost modele pentru structurile de rezistență ale construcțiilor;
Mișcarea prin reacție a cefalopodelor, a meduzelor, a unor insecte a stat la baza realizării motorului cu reacție etc.
*** Pentru alte asemenea exemple, consultați cărțile lui Tudor Opriș.***

13 / 108
COMPOZIȚIA CHIMICĂ A MATERIEI VII*
Generalități Universul (care include și sistemele biologice) este constituit din două/trei componente – materie, energie (și
informație sau principiu organizator).
Materia este caracterizată prin faptul că ocupă spațiu (adică are volum) și are masă; materia există într-o
multitudine de forme (roci, gaze, metale, uleiuri, sisteme biologice etc.), fiecare cu specificul ei.
Materia este alcătuită din elemente chimice prezente în formă pură sau în combinații numite compuși chimici și
există în patru stări de agregare – solidă (are formă și volum bine definite), lichidă (are volum bine definit, dar adoptă
forma spațiului în care se află), gazoasă (lipsită de formă și volum bine definite) și plasmă (majoritară la scala întregului
Univers, dar rară pe Pământ unde este reprezentată de fulgere, aurore polare, flăcări foarte fierbinți).
Energia este definită drept capacitatea de a realiza lucru, adică de a pune materia în mișcare; nu are masă și
volum și poate fi pusă în evidență și măsurată numai prin efectele asupra materiei.
Energia există în două forme sau capacități de lucru – energie cinetică (energie în acțiune) și energie potențială
(energie depozitată, inactivă, dar care are capacitatea sau potențialul de a se manifesta). Există mai multe tipuri de energie
– chimică (energia legăturilor chimice), electrică, mecanică, radiantă (electromagnetică), calorică – în general,
interconvertibile.
Elementele și compușii chimici esențiali ai materiei vii Un element chimic este o substanță care nu mai
poate fi desfăcută în substanțe mai simple prin reacții/metode
chimice obișnuite. În prezent, din cele 112 elemente chimice
cunoscute (la care se mai adaugă alte câteva elemente în curs
de cercetare), 92 sunt prezente în natură, iar celelalte au fost
obținute artificial în laborator. Aceste elemente sunt
prezentate sintetic sub forma sistemului periodic, studiat la
chimie.
Din cele 92 de elemente naturale, aproximativ 25
sunt considerate esențiale în organizarea și funcționarea
materiei vii. Dintre acestea însă, patru – C (carbon), H
(hidrogen), O (oxigen), N (azot) – constituie aproximativ
96% din masa sistemelor biologice, urmate de P (fosfor), S
(sulf), Ca (calciu), K (potasiu) (împreună reprezintă restul de
4%) și de alte elemente în cantități foarte mici (urme).
Elementele „în urme” sunt acelea necesare
organismelor doar în cantități foarte mici; unele, precum Fe,
sunt necesare tuturor formelor de viață; altele sunt necesare doar anumitor specii (precum iodul la vertebrate). Compușii chimici sunt substanțe care rezultă în urma combinării a cel puțin două elemente chimice identice sau diferite, într-un raport
cantitativ/numeric constant și caracteristic fiecărui compus. De exemplu, sarea de bucătărie, clorura de sodiu NaCl, este un compus alcătuit din elementele Na și Cl în raport de 1:1. Compusul rezultat prin interacția dintre elemente are proprietăți noi (proprietăți emergente), diferite de ale elementelor care o
compun luate separat (exemplu: 2Na + Cl2 = 2NaCl, în care Na este metal, Cl2 este gaz toxic, iar compusul rezultat, clorura de sodiu, este sarea de
bucătărie, nu numai netoxic, dar și necesar sistemelor biologice).
Tabel 1. Elemente esențiale pentru plante
Element Simbol Număr
atomic
Masă
atomică
Cantitate necesară în
țesuturi raportată la Mo
Hidrogen H 1 1 60.000.000
Carbon C 6 12 35.000.000
Oxigen O 8 16 30.000.000
Azot N 7 14 1.000.000
Potasiu K 19 39,1 250.000
Calciu Ca 20 40,1 125.000
Magneziu Mg 12 24,3 80.000
Fosfor P 15 31 60.000
Sulf S 16 32 30.000
Clor Cl 17 35,4 3000
Bor B 5 10,8 2000
Fier Fe 26 55,8 2000
Mangan Mn 25 54,9 1000
Zinc Zn 30 65,4 300
Cupru Cu 29 63,5 100
Molibden Mo 42 95,9 1
(Sodiu Na 11 23) Neesențial pt. plante
Cobalt Co 27 58,9 N/A1
1 N/A – abreviere folosită în tabele pentru a specifica lipsa unor informații; provine din engl. not applicable, not available, no answer

14 / 108
În urma analizei chimice concrete a organismelor vii, s-au identificat în compoziția acestora mai multe categorii de
compuși chimici: compuși anorganici (apă, săruri, acizi, baze, oxizi) și compuși bioorganici (glucide, lipide, proteine, acizi
nucleici).
COMPUȘI ANORGANICI
Compușii anorganici sunt reprezentați în principal de apă și săruri minerale, dar și de oxizi, acizi și baze
anorganice.
Apa este un compus chimic simplu, de natură moleculară (între atomii componenți se stabilesc legături
covalente), alcătuit din H și O, în raport numeric de 2 la 1, având deci formula H2O; ca urmare a diferenței mari de
electronegativitate dintre O și H, molecula apei este polară (polul negativ fiind reprezentat de atomul de O – elementul mai
electronegativ, iar cel pozitiv este împărțit în mod egal între cei doi atomi de H), însă neutră din punct de vedere electric.
Constituie aproximativ 60–70% din masa organismelor vii, având deci rol structural în edificarea acestora; alte
roluri, la fel de importante, sunt acelea de solvent polar în cazul soluțiilor (amestecuri omogene de cel puțin două substanțe
diferite), de fază de dispersie în cazul amestecurilor coloidale (neomogene sau heterogene), fiind mediu de transport
(sângele de la animale, sevele brută și elaborată de la plante) pentru diferite substanțe și mediu de reacție pentru aproape
toate procesele (bio)chimice care se desfășoară în sistemele biologice, dar și reactant în multe asemenea reacții; are un rol
important în termoreglare, prin căldura specifică și căldura de vaporizare foarte mari (are capacitea de a înmagazina sau
ceda cantități foarte mari de căldură, fără a-și modifica semnificativ temperatura), și de protecție mecanică, intrând în
compoziția păturilor de lichid din jurul unor organe.
Prin toate aceste proprietăți, apa, considerată „solvent universal”, este un compus indispensabil pentru existența
vieții.
Între moleculele de apă se stabilesc forțe de atracție electrostatică numite legături de hidrogen. Aceste legături
asigură coeziunea puternică între moleculele de apă și explică urcarea sevei brute prin corpul plantei, la înălțimi mari, sub
forma unei coloane de apă neîntrerupte (vă imaginați că frunza trage de moleculele din vârful coloanei de apă, iar celelalte
molecule, fiind atașate de cele din vârf și între ele, vor fi la rândul lor trase în sus). Apa are proprietatea de adeziune –
atașarea moleculelor de apă la diverse suprafețe sau substanțe (cum ar fi atașarea la pereții vaselor conducătoare lemnoase
prin care circulă ascendent prin plantă seva brută). Coeziunea și adeziunea sunt forțe de atracție, care se manifestă evident
în fenomenul de capilaritate (TEMĂ – căutați informații despre acest fenomen).
Sărurile minerale sunt compuși ionici constituiți dintr-un ion metalic, deci pozitiv, și un ion sau grupare
ionică negativă; în stare anhidră sunt substanțe solide cu structură cristalină. Cristalele anorganice sunt caracterizate prin raporturi spațiale și numerice foarte precise si rigide între ionii componenți, rezultatul fiind o
structură înalt ordonată, cu geometrie riguroasă, infinit repetată, dimensiunea rețelei cristaline tridimensionale fiind nedeterminată. Sărurile minerale există în două forme în sistemele biologice: solidă și dizolvată.
Săruri solide constituie sau fac parte din învelișurile externe de protecție (cochilii, tece, exoschelet – la unele
protozoare, alge, moluște, artropode, echinoderme etc.) sau structurile interne de rezistență (endoschelet – oase, spiculi)
ale organismelor și sunt reprezentate de CaCO3, CaPO4, MgCO3, Mg3(PO4)2, SrSO4, CaF2 etc.
În solvent apos se dizolvă, adică se disociază în ionii componenți, care sunt de două categorii: ioni simpli
(monoatomici), rezultați prin cedare sau acceptare de electroni de către un singur element chimic/atom (Na+, K
+, Ca
2+,
Mg2+
, Fe2+
/Fe3+
, Mn2+
, Zn2+
, Cu+/Cu
2+, Ni
2+, Se
6+,4+,2–, Mo
6+→2+, Cr
6+/Cr
3+, Co
2+/Co
3+, Si
4+, Sn
4+, F
–, Cl
–, I
– etc.), și radicali
ionici (ioni poliatomici), rezultați prin combinarea covalentă a mai multor elemente chimice diferite, dar care ca întreg
prezintă sarcină electrică, ionizarea fiind consecința cedării sau acceptării de electroni doar de către unul din atomii
componenți (CO32–
, HCO3–, SO4
2–, PO4
3–, HPO4
2–, NO3
–, NO2
–, SiO4
4–, NH4
+ etc.).
Sărurile au rol mecanic, structural (de protecție și de susținere) când sunt în formă solidă și diverse roluri
funcționale esențiale sub formă de ioni (fac parte din unele vitamine – Co, din enzime – Zn, Mn, din pigmenți – Mg, Cu,
Fe; contribuie la generarea tăriei ionice și a presiunii osmotice, la generarea potențialului membranar, la transmiterea
informației prin neuron, la contracția musculară etc.).
Acizii anorganici sunt compuși chimici care au în compoziție atomi de H legați covalent de alte elemente
chimice mult mai electronegative (O, Cl, F etc.). Acest fapt face ca legătura covalentă la care participă H să fie polară și

15 / 108
deci labilă, ușor de desfăcut heterolitic (inegal), iar în mediu apos H să fie pus în libertate ca proton (H+). Ceea ce rămâne
în urma disocierii protonului este încărcat electric negativ și reprezintă baza conjugată (A–) a acidului respectiv, care poate
fi anion nemetalic simplu (Cl–) sau radical anionic (anion poliatomic), aceiași ca și la săruri.
Conform teoriei protolitice, acizii sunt compuși donori de protoni. Aciditatea unei soluții este reprezentată de concentrația de ioni de H (mai exact, de ioni hidroniu – H3O
+) și se exprimă cantitativ prin pH
(potentia Hydrogenum – puterea Hidrogenului). Acesta are formula pH = – log10[H+] (cologaritmul zecimal din concentrația de protoni dintr-o soluție).
pH 7 – soluții neutre, pH<7 – soluții acide (concentrație mare de H+), pH>7, max. 14 – soluții bazice/alcaline (concentrație mică de H+, mare de OH–).
În sistemele biologice, cei mai frecvenți acizi minerali sunt HCl (rol funcțional în sucul gastric, facilitând
digestia), H3PO4 (rol structural, fiind prezent în compoziția acizilor nucleici, a fosfolipidelor, a nucleotidelor etc.), H2CO3
(rol funcțional în sucurile digestive și în lichidele corpului, ca parte a unui sistem tampon acido-bazic).
Bazele sunt compuși chimici care au în compoziție un cation metalic și un radical anionic specific – (OH)– –
denumit hidroxil sau oxidril.
Conform teoriei protolitice, bazele sunt compuși acceptori de protoni. Ca urmare a acestei definiții, se consideră
baze și compuși care nu conțin radicalul hidroxil, precum NH3 (amoniac) și HCO3– (ion bicarbonat) care au mare afinitate
pentru H+
formând, în mediu apos, NH4+
(ion amoniu), respectiv H2CO3 (acid carbonic) care este instabil și se descompune
în H2O și CO2.
(Atenție! A nu se confunda radicalul ionic hidroxil cu radicalul covalent hidroxil, grupare funcțională neîncărcată
electric, foarte reactivă și specifică celei de-a doua categorii de compuși din materia vie – compușii organici.)
Oxizii sunt compuși binari, ionici sau covalenți, care au în compoziție oxigen și un alt element chimic, metal sau
nemetal. Dacă la formarea oxidului participă un metal, legătura dintre acesta și oxigen este de natură ionică (oxigenul are
electrovalența –2, iar metalul, electrovalență pozitivă); asemenea oxizi sunt oxizi bazici, deoarece în urma reacției lor cu
apa rezultă baze. Dacă la formarea oxidului participă un nemetal, acesta va fi legat de oxigen prin legături covalente, în
general polare, în care oxigenul are covalența maxim 6; asemenea oxizi sunt oxizi acizi, deoarece în urma reacției cu apa se
formează acizi minerali.
În sistemele biologice SiO2 (oxid de siliciu, silice), oxid acid, reprezintă componenta principală a spiculilor
silicioși ale unor spongieri, spicule care constituie un tip de endoschelet; SiO2 întră în componența sau formează căsuțele
sau/și spiculii, scleritele, țepii, eventualele forme de „schelet” ale unor protozoare; de asemenea, același oxid constituie
elementul compozițional principal al solzilor silicioși ai multor crisofite (alge unicelulare aurii) și al frustulei (căsuței)
diatomeelor (alt grup de alge unicelulare). SiO2 realizează mineralizarea pereților celulari ai celulelor tulpinale și foliare
ale unor plante erbacee, conferindu-le rigiditate și rezistență; silicificarea (impregnarea cu silice) este frecventă si la
nivelul pereților celulari ai perilor protectori sau glandulari de la graminee și urzici.
Un alt oxid, mult mai frecvent și mai important este CO2 (di/bioxid de carbon), oxid acid gazos (de aceea nu are
rol structural, ci doar funcțional), prezent în atmosferă în proporție de 0,039% (procente volumetrice) și vital pentru
organismele fototrofe (plante, alge), fiind unul dintre reactanții principali în procesul de fotosinteză.
De asemenea, CO2 este prezent dizolvat în mediul intern al animalelor, reprezentând produs al metabolismului
final și destinat eliminării din organism. La toate categoriile de organisme CO2 este important prin participarea sa la
menținerea pH-ului mediului intern și al soluției intracelulare la valori specifice și relativ constante, prin intermediul
sistemului tampon carbonat – bicarbonat (CO32–
– HCO3– ).
COMPUȘI BIOORGANICI Generalități
Compușii organici sunt reprezentați de substanțe alcătuite aproape exclusiv din elementele chimice vitale,
fundamentale – C, H, O, N, P, S.
Cu studiul compușilor organici în general se ocupă chimia organică. Particularitatea acestor compuși este dată de
proprietatea remarcabilă a atomilor de carbon de a forma, prin legare covalentă între ei sau cu atomii altor elemente (N, O,
S, P etc.), catene/lanțuri liniare, ramificate sau ciclice/inelare de lungimi variate – proprietatea de catenare. În funcție de numărul de atomi care constituie catena, deci de mărimea moleculei, compușii organici se împart în două categorii: molecule mici
(care uneori au rol de monomeri) și molecule mari (macromolecule, unele fiind polimere). Monomerii reacționează între ei, se asociază covalent cap la cap (eliminând câte o moleculă de apă pentru fiecare legătură covalentă formată),
rezultând astfel catene foarte lungi, reprezentate de polimeri (care, implicit, au dimensiuni mari, fiind deci macromolecule). Procesul chimic prin care un
număr mare de molecule similare sau identice – monomeri – se leagă între ele formând o moleculă unică de dimensiuni mari – o macromoleculă polimeră – se numește polimerizare.
Formarea unei legături covalente noi între două molecule (care pot fi monomeri sau nu) prin eliminarea unei molecule de apă se
numește condensare. Se poate spune deci că moleculele monomere
sunt unități structurale care formează prin condensare/condensări repetate compuși chimici macromoleculari.
În cazul anumitor compuși organici, monomerii nu mai
sunt molecule simple, de dimensiuni mici, ci chiar macromolecule care se asamblează necovalent, constituind structuri sau agregate
supramoleculare, forțele stabilizatoare fiind reprezentate de interacții
ionice, hidrofobe, legături de hidrogen și legături van der Waals între unitățile structurale componente (această situație se întâlnește, în special, în cazul proteinelor și acizilor nucleici).
Procesul antagonic polimerizării este depolimerizarea și se realizează prin hidroliză, scindarea secvențială, cu ajutorul apei, a polimerului în
unitățile sale componente, monomerii. Însă nu toate macromoleculele sunt polimeri, unele fiind constituite în mod primar din numeroși atomi, neprovenind printr-o reacție de
polimerizare (chiar dacă aceste macromolecule au dimensiuni mari, nu ajung totuși la acelea caracteristice polimerilor, de mii de atomi în catenă), așa cum
este cazul lipidelor.

16 / 108
Nu toate substanțele/compușii care constituie obiectul de studiu al chimiei organice se regăsesc în compoziția
organismelor vii, ci doar o parte dintre acestea. Fiind atât de specifice sistemelor biologice prin prezența/localizarea
exclusivistă și prin funcțiile lor, a apărut, la granița dintre chimie și biologie, un nou domeniu (considerat de către unii chiar
știință de sine stătatoare) – biochimia. Aceasta are ca obiect de studiu structura (prin aceasta înțelegându-se compoziția
chimică, dimensiunea, geometria moleculelor organice), proprietățile in vitro, funcțiile și funcționarea in vivo a patru mari
categorii de compuși bioorganici: glucide, proteine, lipide și acizi nucleici.
Glucidele, numite și zaharuri, carbohidrați, oze/ozide, sunt cei mai simpli compuși bioorganici, alcătuiți doar din
C, H și O.
Din punct de vedere al dimensiunii moleculei (și implicit, al
numărului de atomi de C) și al solubilității lor, glucidele se împart în
trei clase principale: monozaharide (oze), oligozaharide și polizaharide.
Prima categorie, monozaharidele, reprezintă clasa principală
de zaharuri, deoarece compușii din celelalte clase rezultă prin
polimerizarea acestora.
Din punct de vedere chimic monozaharidele sunt derivați
carbonilici – aldehide și cetone – polihidroxilați (polihidroxialdehide și
polihidroxicetone) și pot avea ca schelet hidrocarbonat catene cu trei
până la șapte (zece) atomi de carbon. Denumirea lor categorială, generică
(de clasă), dată pe baza formulei moleculare și a grupării funcționale
principale, indică atât gruparea funcțională carbonilică, cât și numărul de
atomi de carbon din moleculă; de exemplu, un monozaharid cu trei atomi
de C și o grupare aldehidică se va numi aldotrioză; glucoza, manoza și
galactoza sunt aldohexoze, iar fructoza este o cetohexoză; riboza –
monozaharid esențial din structura acizilor nucleici – este o aldopentoză;
ribuloza – monozaharid implicat în faza a doua a fotosintezei – este o
cetopentoză ș.a. Dar, ca urmare a fenomenului de izomerie, aceeași formulă moleculară este valabilă pentru mai multe monozaharide care se deosebesc prin
gruparea funcțională, prin formula structurală și, implicit, prin proprietățile (fizico-)chimice – de exemplu, formula moleculară C6H12O6 corespunde atât fructozei, o cetohexoză, cât și glucozei, o aldohexoză, pe de o parte, dar și altor aldohexoze precum aloza, galactoza, manoza, guloza, taloza etc., pe de
altă parte.
În sistemele biologice monozaharidele se
pot găsi și cu structura prezentată mai sus, dar sunt
destul de frecvente și oze
modificate prin oxidare sau reducere, prin adăugare
reversibilă sau ireversibilă,
substitutivă sau aditivă, de alte molecule sau grupări
funcționale precum metil, etil,
acetil, fosfat, amino, sulfat (procesele chimice corespunzătoare fiind metilare, etilare, acetilare, fosforilare, aminare, sulfatare), prin deshidratare intramoleculară (cu formare de lactone), prin condensare cu acid lactic sau acid piruvic etc.
Formulele structurale reprezintă modele geometrice, bi- sau tridimensionale, de dispunere a atomilor în molecule. Pentru explicarea
proprietăților monozaharidelor au fost imaginate mai multe categorii de formule structurale, în plan [formula de proiecție (liniară și inelară) Fischer] și tridimensionale (reprezentare cu „pene și puncte”, formulele perspectivice Haworth – inelele furanozice și piranozice, având cinci, respectiv șase atomi,
reprezentările de tip „scaun” și „baie”), ca și modelele de anomerie α și β.
Prin polimerizarea sau, mai exact, conform precizărilor de mai sus, prin policondensarea monozaharidelor rezultă,
în funcție de numărul de molecule care participă la această reacție, oligozaharide și polizaharide (aceste categorii de
zaharuri mai sunt denumite și ozide).
Oligozaharidele sunt glucide alcătuite dintr-un număr redus (2 – 10) de unități monomere (monozaharide). Cele
mai frecvente oze care se regăsesc în structura oligozaharidelor sunt α- și β-glucoza, galactoza, fructoza, manoza ș.a.
Acestea pot participa la polimerizare (se poate utiliza, în cazul oligozaharidelor, termenul de oligomerizare) în forma de
bază sau modificate prin oxidare, reducere, acetilare, aminare, sulfatare, fosforilare, condensare cu alte molecule [precum acidul piruvic, în cazul
acizilor neuraminici (sialici)], prin eterificare cu acid lactic (ca în cazul acidului muramic din componența unității dizaharidice a componentei polizaharidice a mureinei – macromoleculă
structurală din peretele celular bacterian).
În funcție de numărul de oze care constituie oligozaharidele, acestea
por fi dizaharide, trizaharide, tetra-, penta-, hexazaharide etc. Cele mai
importante și mai frecvente/răspândite glucide din categoria oligozaharidelor
sunt dizaharidele dintre care sunt de amintit zaharoza sau sucroza (zahărul
alimentar), alcătuită din α-glucoză și β-fructoză, maltoza și izomaltoza
(ambele rezultă din hidroliza amidonului și a glicogenului și sunt alcătuite din
molecule de α-glucoză), celobioza (rezultă din hidroliza celulozei și este
alcătuită din două molecule de β-glucoză), lactoza (alcătuită din glucoză și β-
galactoză), trehaloza, soforoza, nigeroza, gențiobioza.
Dintre dizaharidele alcătuite din oze modificate (ozele modificate se mai numesc și derivați de monozaharide),
mai importante sunt acelea cu rol de unități (motive) structurale pentru macromolecule precum chitina, polizaharidul

17 / 108
mureinei, componentele glucidice ale matricei extracelulare (glicozaminoglicanii – acizii hialuronici (hialuronan = hialuronat),
dermatan-sulfat și condroitin-sulfat, heparan-sulfat, keratan-sulfat). Oligozaharide alcătuite din mai mult de două oze – simple sau derivate – se
găsesc în componența glicosfingolipidelor neutre (glicozil-ceramide), a gangliozidelor
(glicosfingolipide cu acid sialic în componența oligoglucidului) și a lipopolizaharidelor
(LPS) din membrana externă a bacteriilor Gram-negative. Fragmente oligoglucidice
se pot lega covalent, permanent sau temporar, de anumite proteine
formând glicoproteine.
Ultima clasă de glucide, polizaharidele, este reprezentată de
macromolecule polimere rezultate prin policondensarea unui număr mare
de unități (motive) structurale monozaharidice sau dizaharidice, simple
sau derivate (amintite mai sus). Polizaharidele cu cea mai mare pondere,
calitativă și cantitativă, în sistemele biologice sunt: celuloza, amidonul,
glicogenul, chitina, polizaharidul mureinei, glicozaminoglicanii, deja
menționate anterior.
În organismele vii, glucidele
îndeplinesc trei categorii de roluri: energetic,
structural și într-o mai mică măsură,
funcțional.
Rolul energetic este îndeplinit în
mod direct de către monozaharide (dintre
care glucoza reprezintă combustibilul
principal, fiind preferat de aproape toate
celulele), acestea fiind degradate până la
H2O și CO2, iar o parte din energia eliberată
(conținută în legăturile covalente ale moleculelor de oze) în acest proces este utilizată de către celulă pentru desfășurarea
activităților specifice. Dizaharidele și polizaharidele nu au rol energetic direct, dar reprezintă rezerve de monozaharide, care
rezultă din hidroliza acestora, participând, deci, indirect la metabolismul energetic al sistemelor biologice. Dintre
dizaharide, cele mai importante din punct de vedere energetic sunt zaharoza și lactoza, deoarece sunt surse alimentare de
glucoză, fructoză și galactoză. Zaharoza este și forma în care sunt transportate, prin vasele liberiene, glucidele rezultate în
procesul de fotosinteză, de la sediul producerii lor – țesuturile asimilatoare clorofiliene, în general localizate în frunze – la
toate organele nefotosintetizatoare (vegetative și reproducătoare) ale plantei.
Fiind alcătuite dintr-un număr mic de oze, iar prezența lor
în organism, dependentă, în general, de aportul alimentar/de
nutrienți, dizaharidele nu constituie rezerve consistente de
monozaharide. Asemenea stocuri/rezerve de combustibil metabolic
sunt reprezentate de anumite polizaharide, precum glicogenul (la
animale, ciuperci, unele protoctiste și unele bacterii) și amidonul
(specific celulelor vegetale, dar și, în forme modificate, unor
protoctiste, mai ales alge). Aceste două glucide polimere reprezintă
forma în care este depozitată, în celule, o parte a excesului alimentar
de glucoză (și de alte oze, dar de obicei transformate în prealabil tot
în glucoză), fiind constituite exclusiv din acest monozaharid.
Glicogenul este localizat în citoplasmă, iar amidonul, în cloroplaste
și leucoplaste caracteristice – amiloplaste. Prin hidroliza
secvențială/treptată a acestor glucide polimere rezultă, în final,
cantităti mari de glucoză, degradată mai departe specific cu eliberare
de energie (proces complex, numit respirație celulară).
Rol structural au în principal polizaharidele, precum
celuloza, chitina, poliglucidul mureinei, glicozaminoglicanii, care
formează sau participă la constituirea structurilor de rezistență
celulare, tisulare sau ale organismului întreg.
Astfel, celuloza și derivații ei formează peretele celular al
celulei vegetale (fiind o caracteristică specifică și definitorie a
regnului Plantae) și al unor celule algale, dar se găsește și în
compoziția tunicii urocordatelor (animale cordate).
Chitina, alcătuită din glucoză aminoacetilată, formează peretele celular al celulelor fungale (fiind, de asemenea,
o trăsătură specifică a reprezentanților regnului Fungi), dar, prin mineralizare cu CaCO3, și exoscheletul multor artropode
(crustacei, păienjeni și scorpioni, miriapode, insecte).
Mureina este elementul structural principal al peretelui celular al celulelor bacteriene având drept component
de bază un polizaharid.
Glicozaminoglicanii constituie, alături de proteine, matricea extracelulară – substanță care umple spațiul dintre
celulele animale, le asigură acestora suport de fixare sau de deplasare, reprezintă, în cazul țesuturilor conjunctive, substanța
fundamentală a acestora (putând fi mineralizată, ca în cazul țesuturilor osos și cartilaginos, conferindu-le rezistență
mecanică).

18 / 108
Alte oligo- și polizaharide constituie „învelișul dulce” –
capsula, glicocalixul – al unor celule (animale, bacteriene, fungale,
algale etc.), structură cu multiple funcții, precum susținere și protecție
mecanică și antiimună, aderență și recunoaștere intercelulară ș.a.
Dintre monozaharide, rol structural au glucoza (componentă
polizaharidelor cu rol structural), riboza și deoxiriboza (participă la
edificarea acizilor nucleici).
Rol funcțional au unele oligoglucide, precum cele alcătuite
din molecule de manoză, care sunt atașate de proteine, indicând
destinația acestora în celulă. Alte glucide, oligo- sau polimere, au rol de
factori de virulență, intervin în reglarea presiunii osmotice
intracelulare (fiind substanțe higrofile și osmotic-active), protejează proteinele împotriva digestiei enzimatice (ca în cazul
imunoglobulinelor și a proteinelor membranare, aceste din urmă fiind protejate de glicocalix sau de capsulă).
Uneori, prezența lor are semnificație negativă, capsula bacteriană fiind cauza, in vivo, și dovada în laborator a
caracterului patogen și a virulenței unei speciei, polizaharidele capsulare mascând anumite structuri moleculare
membranare care ar putea fi recunoscute de celulele sistemului imunitar care astfel activate, ar neutraliza agentul patogen
respectiv.
Lipidele sunt compuși bioorganici în general insolubili în apă (deci hidrofobi), dar solubili în solvenți organici
(eter, benzen, cloroform etc). Moleculele de lipide sunt, de obicei, de mari dimensiuni, fără însă a fi polimere; conțin în
mod constant C, O, H la care se adaugă P, N, S în cazul anumitor tipuri de lipide.
În funcție de structura moleculară și de proprietățile fizico-chimice, lipidele sunt de mai multe categorii:
Acizi grași (AG). Sunt molecule formate în mod obișnuit din catene liniare (neramificate) constituite dintr-un
număr mare și, în general, par de atomi de carbon; au grupare funcțională –COOH (carboxil), care are caracter acid punând
în libertate H+ și deci este hidrofilă. Atașată de gruparea carboxil este o catenă hidrofobă (nepolară) de atomi de C legați
covalent fie numai prin legături simple (acid gras saturat), fie și prin legături duble (dacă este doar o asemnea legătură în
toată catena – acid gras mononesaturat, dacă sunt mai multe – acid gras polinesaturat).
Roluri: rol structural/plastic – intră în alcătuirea altor clase de lipide (trigliceride, fosfolipide); rol energetic
semnificativ, conținând o cantitate mai mare de energie decât glucidele, dar care poate fi extrasă numai pe cale
oxidativă/aerobă (1g de lipide – 9,3 kcal); rol funcțional redus – unii acizi grași (numiți acizi grași esențiali, acidul α-
linolenic (ω-3) și acidul linoleic (ω-6), la care se adaugă unii condiționat esențiali – acidul γ-linolenic, acidul lauric și
acidul palmitoleic) constituie vitamina F (denumire dată pentru șapte ani, 1923-
1930, acizilor grași esențiali care au fost ulterior clasificați drept lipide); unii
acizi grași (acidul arahidonic) sunt implicați în semnalizarea intracelulară.
Trigliceride (TG)/triacilgliceroli (TAG). Sunt molecule formate prin
esterificarea glicerolului/glicerinei cu trei acizi grași. Sunt total hidrofobe și deci
insolubile în apă (cu toate că atât glicerolul, cât și acizii grași sunt ±solubili în
apă). Se mai numesc popular grăsimi neutre.
În funcție de tipul de acizi grași legați la glicerol, TAG sunt de două
tipuri – lichide (grăsimile vegetale, uleiurile; acizii grași sunt cu catenă
nesaturată) și solide (grăsimile animale; acizii grași sunt cu catenă saturată).
Roluri: rol structural – prin acumulare în
celule edifică țesutul adipos la animale și
parenchimul de înmagazinare a substanțelor de
rezervă la plante, fiind principala formă în care se
depozitează lipidele în organism; rol funcțional –
prin țesutul pe care în formează, au rol de protecție
mecanică și termică (limitează termoliza – pierderea
de căldură din organism); rol energetic semnificativ
– sunt forma de depozit a acizilor grași.
Fosfolipide (FL). Sunt lipide complexe
formate din glicerol, doi acizi grași și o grupare
polară (deci hidrofilă) care conține în mod obligatoriu radical fosfat, alături de
alte componente – colină, inozitol, serină, etanolamină etc.
Moleculele de FL au forma unor acadele: prezintă un cap globular
polar hidrofil, care conține fosfatul și glicerolul, și două cozi nepolare
hidrofobe, reprezentate de cele două molecule de acizi grași.
Roluri: în principal, rol structural – participă la edificarea
membranelor celulare.
Ceruri. Sunt lipide complexe formate din lanțuri lungi de AG unite cu
lanțuri lungi de alcooli sau cu steroli. Sunt solide la temperatura camerei, sunt
hidrofobe și deci impermeabile pentru apă (proprietate din care derivă localizarea și rolurile rol).
Roluri: rol structural – albinele le folosesc la edificarea fagurilor; rol funcțional – acoperă și protejează unele
organe ale plantelor, prevenind pierderea de apă (deshidratarea); la mamifere, glandele ceruminoase din peretele

19 / 108
conductului auditiv extern produc cerumen, care are rol protector pentru timpan, împiedicând impuritățile să ajungă la
acesta.
***Tot în această categorie pot fi incluse unele ceruri complexe precum cutina (componentă a cuticulei – peliculă
lipidică, cu grosime variată, care acoperă organele supraterane ale plantelor, prevenind deshidratarea) și suberina
(substanță care impregnează și rigidizează pereții celulari ai unor țesuturi care astfel capătă proprietăți mecanice – susținere,
protecție, dar și de izolare termică și prevenirea deshidratării)***.
Steroli (steroizi). Sunt lipide cu structură complexă (mai multe catene de
atomi de carbon ciclice/inelare condensate, de care se atașează grupări funcționale
și catene liniare neramificate și ramificate), care au ca principal reprezentant
colesterolul, de la care derivă ceilalți reprezentanți ai clasei. Sunt molecule
amfipolare – au o regiune polară și hidrofilă și o regiune nepolară și hidrofobă, ca
și FL.
Roluri: rol structural – colesterolul (dar și alți steroli) participă la formarea membranelor celulare; rol funcțional
– din colesterol se sintetizează în organismul animal vitamina D (rol în absorbția Ca și fixarea acestuia în oase), acizii
biliari (din care se formează sărurile biliare cu rol în digestia și absorbția lipidelor), unii hormoni (cei sexuali și cei
produși de corticosuprarenală); la plante sterolii sunt reprezentați de unii hormoni – brasinosteroizii.
Terpene și terpenoide. Sunt molecule lipidice care conțin, în principal, numai C și H și care derivă de la izopren.
Se găsesc într-o mare varietate de forme – pigmenți (caroten), latex (din care se formează cauciucul), uleiuri
volatile/eterice (care sunt responsabile de parfumul unor plante – trandafir, mușcată etc.), vitamine (vitamina A este o
substanță derivată din caroten, vitamina K, vitamina E), scualen (prezent în
ficatul rechinilor asigurându-le flotabilitatea; este precursorul colesterolului, de
aceea steroizii sunt considerați derivați de terpene) etc.
Sfingolipidele. Sunt lipide structurale, ca și FL, prezente în cantități
mari în țesutul nervos, unde participă la formarea mielinei (rol de izolator
electric, de protecție mecanică, de hrănire pentru axonii neuronilor).
Proteinele sunt compuși bioorganici macromoleculari polimeri.
Monomerii din care se formează, prin policondensare, proteinele se numesc
aminoacizi.
Aminoacizii sunt molecule organice mici, cu două
grupări funcționale obligatorii – carboxil (–COOH), care
conferă moleculei caracterul acid, și amino (–NH2), care conferă
caracter bazic. Se deosebesc între ei prin restul moleculei, notat
R(adical), și care variază de la un aminoacid la altul.
În alcătuirea proteinelor întră 20 (sau 21, după unii
autori) de aminoacizi (numiți uzuali sau proteinogeni), din care
9(10) sunt aminoacizi esențiali (organismele heterotrofe, cum
sunt animalele, au nevoie de toți cei 21 de aminoacizi pentru a-
și sintetiza proteinele proprii, dar nu pot sintetiza în cantități
suficiente decât 11 dintre ei, iar pe ceilalți – numiți esențiali –
trebuie să-i preia din exterior – din dietă): metionină, valină,
treonină, fenilalanină, leucină, izoleucină, triptofan, lizină,
histidină și arginină (ultimii doi fiind esențiali doar la sugari).
Sunt considerați aminoacizi neesențiali alanina,
asparagina, acidul aspartic, acidul glutamic și condiționat
esențiali (adică este necesar un aport crescut în anumite situații
– stres, boală), cisteina (poate înlocui metionina), glutamina,
tirozina (poate înlocui fenilalanina), glicina, ornitina, prolina și
serina.
Aminoacizii au rol structural, fiind componente ale
peptidelor și ale proteinelor, dar și rol funcțional participând la
reglarea presiunii osmotice și a pH-ului lichidelor corpului, și
rol energetic redus, fiind folosiți drept combustibil numai în
condiții extreme, după epuizarea glucidelor și a lipidelor.
Aminoacizii se pot lega covalent între ei prin eliminrea
unei molecule de apă între gruparea carboxil a unui aminoacid
și gruparea amino a următorului aminoacid, legătura rezultată
numindu-se legătură peptidică.
Prin policondensare (numită eronat și polimerizare) se
formează catene/lanțuri de aminoacizi, reprezentate de peptide
și de proteine.
Peptidele sunt de două categorii, în funcție de numărul
de aminoacizi care participă la formarea lor:
A. oligopeptide – conțin între 2 și 10 aminoacizi
B. polipeptide – conțin între 10 și 50 aminoacizi

20 / 108
Unele polipeptide au rol funcțional foarte important, fiind reprezentate de unii hormoni (ocitocina, ADH,
bradikinină etc.).
Peptidele care conțin peste 50 de aminoacizi (numărul lor ajungând până la 1000, chiar și mai mult) sau, după un
alt criteriu, care au masă moleculară peste 10.000 Da sunt considerate proteine (cu toate că delimitarea este arbitrară).
Clasificarea proteinelor
În funcție de forma generală a moleculei și de relația cu apa, proteinele sunt
globulare (de obicei ușor solubile și cu localizare intracelulară) și fibrilare (de obicei
insolubile, cu localizare extracelulară, cu rol de susținere).
În funcție prezența sau absența în molecula proteică a unor grupări diferite
de catenele peptidice/proteice, proteinele pot fi proteine simple sau homoproteine
(formate numai din aminoacizi) și proteine conjugate sau heteroproteine (formate
dintr-o grupare proteică, numită apoproteină, și o grupare neproteică, numită
grupare prostetică; împreună, cele două grupări – proteică și prostetică, formează
holoproteina).
După natura grupării prostetice, există mai multe clase de heteroproteine:
fosfoproteine, glicoproteine, lipoproteine, cromoproteine (proteine colorate, cum ar fi
hemoglobina – roșie, mioglobina – roșie, hemocianina – albastră, flavoproteinele –
galbene), metaloproteine (feritina are Fe, ceruloplasmina conține Cu), nucleoproteine
(asociere între acizi nucleici și proteine bazice).
Nivelurile structurale ale proteinelor
Proteinele sunt molecule de mari dimensiuni, care adoptă conformații
spațiale particulare, care le permit să-și îndeplinească funcțiile specifice. Se descriu
astfel, în funcție de particularitățile spațiale, patru niveluri structurale.
1. Structura primară este reprezentată de numărul, tipul și ordinea în
moleculă a celor 20(21) de aminoacizi. Proteina are, în general, formă
liniară.
2. Structura secundară este reprezentată de două conformații spațiale
locale pe care le poate forma lanțul proteic – α-helix (conformație
spiralată) și β-pliată (conformație rezultată prin plierea pe ea însăși a
catenei proteice, fiecare pliu fiind paralel cu celelalte). 3. Structura terțiară este reprezentată de împachetarea proteinei care este
deja organizată în structură secundară. Se formează domenii structural-funcționale mari ale proteinei.
4. Structura cuaternară este întâlnită numai la proteinele care, formă funcțională, sunt constituite din mai multe
catene polipeptidice distincte, care se asociază, de obicei, prin alte tipuri de legături decât cele covalente.
Catenele polipeptidice individuale se numesc protomeri, iar proteina funcțională este un multimer. Exemplu
– hemoglobina (vezi structura hemoglobinei).
Rolurile proteinelor
Rol structural esențial – proteinele reprezintă componenta bioorganică majoritară din sistemele biologice,
constituind scheletul molecular organic al acestora. Exemple – colagen, elastină sunt prezente în tendoane, oase, cartilaje,
ligamente, matricea celulară în general; keratina se găsește în piele, coarne, copite, fulgi, unghii, păr etc.
Rol energetic redus – aminoacizii sunt folosiți drept combustibil numai după epuizarea glucidelor și a lipidelor.
Roluri funcționale numeroase și importante:
a. Enzime. Enzimele sunt biocatalizatori – participă calitativ, nu cantitativ la reacțiile biochimice din organism.
Enzimele cresc viteza de reacție și determină sensul reacțiilor și succesiunea lor în cadrul căilor metabolice
(calea meatabolică este un lanț de reacții biochimice). Toate reacțiile biochimice din organism sunt catalizate
Denaturarea și
renaturarea
proteinelor

21 / 108
de enzime. Una dintre cele mai abundente enzime din natură este RuBisCO (ribulozo-bisfosfat-carboxilaza-
oxigenaza) – este enzima principală a procesului de fotosinteză.
b. Depozit/rezervă de aminoacizi. Aminoacizii vor fi extrași din proteine și utilizați în alte procese metabolice.
Exemple – aleurona, zeina (din semințele plantelor), albuminele (din sânge – serumalbumina, din ou –
ovalbumină), cazeina din lapte.
c. Transport. Unele proteine transportă diverse alte substanțe – hemoglobina și hemocianina trasportă O2,
unele proteine plasmatice (din sânge) transportă ioni minerali și hormoni, prin membrana celulelor sunt
transportate diverse substanțe de către canale, pompe și translocatori etc.
d. Mișcare. Deplasarea diverselor componente în celulă, a celulelor și chiar a organismelor în întregime este
realizată prin intervenția, la diferite niveluri, a două proteine – actină și miozină (proteine contractile prezente
în toate tipurile de celule, dar deosebit de abundente în celulele musculare).
e. Apărarea organismului. În sângele animalelor se găsesc anticorpi (imunoglobuline), proteine implicare în
coagularea sângelui (previne hemoragia) și proteinele sistemului complement (atacă celulele străine
organismului), toxine și veninuri.
f. Mesageri chimici. Unele proteine conțin și transmit mesaje de la celulă la celulă și chiar altor organisme.
Exemple – unii hormoni (insulina, hormonul de creștere etc.), feromoni, unii neurotransmițători (asigură
comunicarea dintre neuroni și alte celule), factori de creștere etc.
g. Alte roluri – reglarea presiunii osmotice și a pH-ului lichidelor corpului etc.
Acizii nucleici sunt cele mai complexe substanțe de pe Terra. Ei reprezintă substratul chimic al eredității
deoarece depozitează, transmit și operează cu informația genetică, responsabilă de toate trăsăturile unui organism.
Acizii nucleici sunt compuși bioorganici macromoleculari polimeri. Unitățile de bază (monomerii) din care sunt
alcătuiți acizii nucleici se numesc nucleotide => acizii nucleici sunt compuși
polinucleotidici.
Un nucleotid la rândul său este alcătuit din trei componente:
1. Radical/i fosfat (1-3)
2. Glucid cu 5 atomi de C – pentoză
3. Bază azotată
Radicalul fosfat – este comun tuturor nucleotidelor, atât celor din ARN, cât și
celor din ADN. Într-un nucleotid se poate găsi un singur radical fosfat (în cazul
nucleotidelor incluse în macromolecula polimeră) sau se pot găsi doi sau trei radicali fosfat (nucleotidele libere).
Pentoza este riboza în nucleotidele ARN (ribonucleotide) și deoxiriboza în nucleotidele ADN
(deoxiribonucleotide).
Baza azotată – există două tipuri de baze azotate: pirimidinice și purinice.
Bazele azotate pirimidinice sunt: (1) Uracilul U – se găsește exclusiv în ARN. (2) Timina T – există doar în
ADN. (3) Citozina C – se găsește atât în ARN, cât și în ADN.
Bazele azotate purinice sunt: (1) Adenina A – se găsește atât în ARN, cât și în ADN. (2) Guanina G – se găsește
atât în ARN, cât și în ADN.
Legarea covalentă a celor trei componente între ele – vezi desenul de mai sus.
Baza azotată împreună cu pentoza formează un/o nucleozid/ă. Nucleozidul împreună cu 1-3 radical/i fosfat
formează un/o nucleotid/ă (denumit nucleozid-mono/di/tri-fosfat).
Nucleozide din ARN: adenozină A-R, guanozină G-R, citidină C-R, uridină U-R; din
ADN: deoxiadenozină A-dR, deoxiguanozină G-dR, deoxicitidină C-dR, (deoxi)timidină T-dR. R
– riboză, dR – deoxiriboză, A, G, C, T, U – baze azotate.
Nucleotidele din ARN sunt: AMP – adenozin-monofosfat, GMP – guanozin-monofosfat,
CMP – citidin-monofosfat, UMP – uridin-monofosfat.
Nucleotidele din ADN sunt: dAMP – deoxiadenozin-monofosfat, dGMP, dCMP, dTMP.
În stare liberă se găsesc și formele di- și trifosfat ale acestor nucleotide: ADP – adenozin-
difosfat, GDP, CDP, UDP, ATP, GTP, CTP, UTP, respectiv dADP, dGDP, dCDP, dTDP, dATP,
dGTP, dCTP, dTTP.
ATP – moleculă macroergică: depozitează, transportă, transferă energie în sistemele
biologice; ATP = energie metabolică. Energia este stocată în legături covalente speciale.
Nivelurile structurale ale acizilor nucleici Structura primară este reprezentată de numărul, tipul și succesiunea/secvența
nucleozid-monofosfaților legați covalent într-o monocatenă de acid
nucleic.
Legătura covalentă între două nucleozide-monofosfat
succesive se realizează prin intermediul radicalului fosfat al unui
nucleozid-monofosfat și pentoza nucleozidului următor, constituind o punte covalentă
fosfodiesterică.
Ca urmare a acestui mod de legare covalentă prin punți fosfodiesterice monocatena
de acid nucleic va fi polarizată deoarece la un capăt va rămâne liber carbonul 5’-fosfat
(capătul 5’-fosfat) al pentozei nucleotidului terminal, iar la celălalt capăt al monocatenei va
rămâne liber carbonul 3’-OH (capătul 3’-OH) al ribozei acestui nucleotid terminal.

22 / 108
Partea nespecifică a monocatenelor de acizi nucleici este coloana glucido-fosforică/pentozo-fosforică (alcătuită
din pentoze legate covalent prin punți fosfodiesterice). Componenta specifică a fiecărei monocatene de acid nucleic este
reprezentată de succesiunea (ordinea) caracteristică (unică) pentru fiecare catenă în parte a bazelor azotate în cadrul
catenei.
Structura secundară este reprezentată de structura bicatenară a acizilor nucleici. În
general, moleculele de ARN au doar structură primară, fiind deci monocatenare (există și excepții).
Deoarece ADN este o moleculă bicatenară, de obicei se vorbește despre structura secundară
a ADN.
ADN este o macromoleculă alcătuită din două monocatene atașate una de cealaltă prin
punți de hidrogen formate între bazele azotate aparținând nucleotidelor din cele două catene; cele
două monocatene sunt răsucite spiral (helical/elicoidal) spre dreapta – răsucire dextrogiră în jurul
unui ax virtual comun. ADN – macromoleculă dublucatenară helicală dextrogiră.
Cele două monocatene ale dublului helix ADN sunt geometric paralele, dar biochimic
antiparalele, deoarece fiecare monocatenă este polarizată 5’-3’, dar ele sunt dispuse în molecula
bicatenară invers orientate una față de cealaltă – capătului 5’ al unei catene îi corespunde
capătul 3’ al celeilalte catene și invers.
Cele două catene ale dublului helix sunt complementare, deoarece fiecărei baze
azotate aparținând unui nucleotid dintr-o catenă îi corespunde doar o anumită bază azotată a
unui nucleotid din cealaltă catenă, cele două baze corespondente, numite baze complementare,
legându-se prin punți de hidrogen. Există doar patru împerecheri posibile de baze azotate, în
fiecare pereche una fiind purinică, iar cealaltă pirimidinică. Deoarece structura ADN a fost
descoperită de Watson și Crick, aceste împerecheri posibile de baze azotate se numesc perechi
Watson-Crick: A=T, T=A, G≡C și C≡G.
Legătura dintre cele două monocatene se realizează prin punțile de hidrogen care se formează între bazele azotate
împerecheate pe criterii de complementaritate astfel: între A și T se formează două punți de hidrogen, iar între G și C se
formează trei punți de hidrogen.
Perechile de baze Watson-Crick sunt perechi de baze complementare.
Deoarece perechile Watson-Crick conțin baze complementare rezultă că și și
monocatenele de ADN vor fi în ansamblul lor complementare și codeterminate
(prezența unei baze azotate pe una dintre catene impune prezența pe cealaltă catenă a
bazei azotate complementare). Acest fapt ne permite să deducem imediat secvența
unei monocatene de ADN, cunoscând-o pe cea a catenei complementare.
Se poate realiza o analogie între structura secundară a ADN și o scară în
spirală: balustradele scării sunt cele două coloane glucido-fosforice ale celor două
monocatene de ADN, iar treptele sunt perechile de baze azotate dintre coloane, care sunt dispuse perpendicular pe axul
virtual comun al macromoleculei și deci paralele între ele.
Moleculele de ADN pot fi circulare (nucleoidul și plasmidele de la procariote) sau liniare (ca în cromosomii de la
eucariote).
La eucariote, moleculele linare de ADN se asociază cu proteine bazice (histone) și constituie cromatina (complex
supramolecular fibrilar nucleo-histonic). O fibră de cromatină este un cromosom. Acesta devine aparent prin
supracondensarea fibrei de cromatină (ca un fular răsucit puternic) – se scurtează și se îngroașă. În alcătuirea cromosomilor
mai intră ARN cromosomal, lipide, ioni de Ca și Mg, alte proteine (proteine non-histonice).
Din complementaritatea celor două monocatene ale moleculei de ADN rezultă imediat codeterminarea acestora și,
mult mai important, mecanismul prin care se realizează copierea cu fidelitatea a moleculei de ADN – proces numit
re(du)plicare semiconservativă: dintr-o moleculă inițială de ADN (= ADN parental), rezultă două (reduplicare)
molecule-fiice identice între ele și identice cu molecula parentală, care dispare ca entitate distinctă, însă se regăsește pe
jumătate în fiecare din cele două molecule-fiice de ADN (se conservă
pe jumătate – semiconservativ). Acest mecanism este foarte
important, dat fiind rolul ADN-ului – substrat biochimic al
informației genetice, depozitar și transmițător în descendență al
acesteia. Astfel, dintr-o moleculă de ADN care conține o anumită
informație genetică, prin reduplicare vor rezulta două molecule de
ADN care vor conține aceeași informație și care vor fi transmise în
descendență, care astfel va avea și va folosi aceeași informație ca și
părinții.
Cromosomii pot fi bicromatidici (formați din două cromatide-surori identice, paralele și atașate între ele pe
întreaga lungime), monocromatidici (o singură cromatidă) sau bivalenți (tetrade cromatidiale, doi cromosomi
bicromatidici omologi atașați lateral unul de altul). O cromatidă este un filament de cromatină puternic condensat,
conținând deci o singură moleculă de ADN. Din punct de vedere morfo-structural, cromatida prezintă o constricție primară
– centromer, două brațe egale sau inegale ca lungime separate prin centromer (notate cu p – brațul scurt și q – brațul lung)
și două extremități numite telomere. Unii cromosomi prezintă și o constricție secundară la nivelul brațului scurt, care
delimitează un segment de cromatidă numit satelit (asemenea cromosomi se numesc cromosomi sateliferi).

23 / 108
Structura ARN. Tipuri de ARN ARN este o moleculă monocatenară poliribonucleotidică, care conține ribonucleotide (nucleotide care au în
alcătuire riboza, iar baza azotată este una dintre A, G, C și U). Anumite tipuri de ARN au o structură secundară.
În toate sistemele biologice și la unele virusuri, singura moleculă care conține și transmite informația
genetică este ADN, iar ARN operează cu această informație.
În funcție de rol și de structură, există mai multe tipuri de ARN:
I. ARN-uri virale
1. ARNv (ARN viral) – se întâlnește la unele virusuri – la ribovirusuri precum virusul gripal, virusul
poliomielitei, HIV etc. La aceste virusuri ARNv este depozitarul și transmițătorul de informație genetică.
II. ARN-uri celulare
2. ARNr (ARN ribosomal) – prezent în toate celulele în alcătuirea subunităților ribosomale, în care are rol atât
structural, cât și funcțional catalitic. Este sintetizat pe baza informației din ADN și diferitele tipuri de ARNr sunt asamblate
cu proteine ribosomale, formându-se subunitățile ribosomale. Reprezintă 80-90% din cantitatea totală de ARN dintr-o
celulă.
3. ARNm (ARN mesager) – reprezintă 2-5% din ARN celular. Are dimensiuni variate, deoarece dimensiunea lui
corespunde cu lungimea genei ADN a cărei informație o copiază și o transportă în citoplasmă. Genele ADN conțin
informația genetică, dar nu părăsesc nucleul; însă utilizarea informației genetice se realizează în citoplasmă. De aceea
trebuie să existe o moleculă cu valoare de mesager care să preia informația din genele nucleare și să o transporte în
citoplasmă la aparatul de biosinteză proteică, unde va fi folosită. Astfel, ARNm aduce la nivelul ribosomilor mesajul
genetic copiat din ADN, mesaj care va fi folosit la sinteza unei proteine specifice.
Dogma centrală a geneticii și biologiei moleculare
4. ARNt (ARN de transport sau de transfer) – este o moleculă monocatenară
mică, de cca. 80 nucleotide, care formează și regiuni bicatenare ca urmare a plierii pe ea
însăși; are astfel o structură bidimensională în formă de frunză de trifoi, fiind alcătuit din
patru brațe bicatenare, trei dintre ele prezentând terminal câte o buclă monocatenară, iar
al patrulea conținând capetele libere 3’-OH și 5’-P ale monocatenei.
Porțiunile bicatenare sunt stabilizate prin punți de hidrogen formate între A și
U, respectiv G și C. La capătul 3’-OH există întotdeauna 3 nucleotide ACC, iar la
capătul 5’ există întotdeauna o G.
La capătul 3’-OH, ARNt va lega un aminoacid și îl va transporta în ribosom în
vederea includerii într-o proteină în procesul de biosinteză proteică.
Deci, moleculele de ARNt sunt molecule adaptor, care transportă aminoacizii în ribosomi și corelează mesajul
genetic din ARNm cu sinteza proteinei.
Rezumat
ARN ADN Riboză Deoxiriboză
A, G, C, U A, G, C, T
Monocatenar – catena este polarizată: capăt
5'-fosfat și capăt 3'-OH
Bicatenar helical dextrogir; catenele sunt
polarizate, antiparalele, complementare, codeterminate
Sinteză prin transcriere pe baza genelor ADN Reduplicare semiconservativă
Operează cu informația genetică
(excepție – ribovirusurile – ARN depozitează și transmite informația genetică, adică are rol
de moleculă genomică)
Depozitează și transmite în descendență
informația genetică
Alte tipuri de ARN
5. ARNsn (ARN nuclear mic) – are rol structural și catalitic în spliceosom = complex ARN-proteine care prelucrează pre-ARNm.
6. ARNsno (ARN nucleolar mic) – ajută la procesarea pre-ARNr în vederea formării subunităților ribozomale în nucleol.
7. SRP-ARN – este component al SRP (particula de recunoaștere a semnalului) = complex ARN-proteine care recunoaște secvențele semnal
ale proteinelor cu destinație spre reticulul endoplasmic.
8. ARNsi (interferent mic) 9. ARNmi (microARN) Atât ARNsi cât și ARNmi sunt implicați în reglarea expresiei genice.
10. XISTARN – este implicat în procesul de inactivare prin heterocromatinizare a cromozomilor X suplimentari din celulă. Întotdeauna într-o celulă trebuie să existe un singur cromozom X funcțional, celălalt sau ceilalți fiind inactivați, rezultând cromatina sexuală sau corpusculul Barr.
Toate tipurile de ARN de la 5 la 10 se numesc colectiv ARNhn (ARN heterogen).
ADN TRANSCRIERE
ARN TRADUCERE
PROTEINĂ
REVERSTRANSCRIERE
RE
PL
ICA
RE

24 / 108
CELULA – GENERALITĂȚI
Toate organismele vii sunt alcătuite din celule. Celula este cea mai mică entitate vie (care posedă toate însușirile
distinctive și definitorii ale viului).
Primele celule au fost observate de către Robert Hooke (1635-1703, naturalist, arhitect, polimat englez) în 1665.
El a folosit un microscop rudimentar, de construcție proprie, pentru a analiza o secțiune grosieră prin stejarul de plută
(Quercus suber). A văzut cămăruțe (compartimente) cu pereți rigizi și le-a denumit celule. În limba latină cella înseamnă
cameră, chilie, cămară, iar diminutivul este cellula. A considerat că pereții erau vii, iar celulele erau goale2.
Teoria/doctrina celulară După aproape 200 de ani de la observarea primelor celule, în 1838, Matthias Jakob Schleiden (1804-1881,
botanist german) studiază mai îndeaproape celulele vegetale, iar Theodor Schwann (1810-1882, zoolog și fiziolog
german), în 1839, pe cele animale. Cei doi cercetători formulează independent primele două postulate ale teoriei celulare.
Această teorie va fi completată cu al treilea postulat în 1855(58) de către medicul anatomo-patolog Rudolf Ludwig Karl
Virchow (1821-1902), care o preia, la rândul lui, de la Robert Remak (1815-1865, embriolog, fiziolog, neurolog prusac).
Completarea Virchow-Remak este o extindere a postulatului lui Francesco Redi (1626-1697, medic, naturalist, poet
italian): Omne vivum ex ovo (Orice ființă vie provine dintr-un ou, adică dintr-o celulă inițială).
Teoria celulară a lui Schleiden, Schwann și Virchow susține:
1. Toate organismele vii sunt alcătuite din una sau mai multe celule nucleate; nu există viață în afara celulei.
2. Celula este unitatea elementară structurală și funcțională a lumii vii.
3. Omnis cellula e cellula. Celulele provin numai din alte celule vii, prin procesul de diviziune celulară; acest proces
asigură înmulțirea celulelor, iar multiplicarea numărului de celule are drept consecințe creșterea și dezvoltarea
organismelor.
În prezent s-au mai adăugat și alte postulate:
4. Celula este un tot unitar – un ansamblu coerent de părți diferite aflate în interacțiune.
5. Celula este un sistem biologic deschis – un ansamblu de elemente diferite interconectate structural, spațial,
funcțional și temporal, care schimbă permanent cu mediul materie, energie și informație.
Definiția celulei Celula este unitatea elementară (fundamentală, de bază) structurală, funcțională și genetică (informațională)
a tuturor organismelor vii; prezintă integralitate (elementele constitutive formează un tot unitar), metabolism propriu
(schimb de substanță și energie cu mediul), capacitate de autoreglare (schimb de informație cu mediul), de
autoconservare și de autoreproducere.
Celulele pot exista și funcționa singure (ca entități autonome, independente, de sine stătătoare), îndeplinind toate
funcțiile specifice viului (nutriție, relație, reproducere) ca în cazul organismelor unicelulare: protozoare, unele alge, unele
ciuperci, sau se pot asocia constituind grupări sau agregate cu niveluri crescânde de interdependență: cenobii, colonii și
țesuturi. Țesuturile alcătuiesc organe, iar organele se asociază în sisteme de organe, a căror totalitate armonios integrată
structural și funcțional formează organismul pluricelular.
Forma celulelor Este corelată în principal cu funcția îndeplinită, dar și cu poziția în țesut, adică depinde de presiunea celulelor
din jur. Inițial este sferică, apoi poate deveni ovală, cilindrică, prismatică, stelată, piramidală (unii neuroni), fusiformă
(fibre sclerenchimatice, celule musculare netede), reniformă (celule stomatice), poliedrică (celulele vegetale
parenchimatice), cubică, piriformă sau rămâne sferică (adipocit, ovul, condrocit etc.). Unele celule au formă variabilă,
schimbându-și permanent conturul deoarece emit pseudopode. În general, forma variază mai puțin la plante decât la
animale.
Dimensiunea celulelor În medie, dimensiunea celulelor este cuprinsă între 1 și 100 m (1m = 10
-6 m = 10
-3 mm); celulele pot avea și
dimensiune de fracțiuni de micron, dar și centimetri și metri. Dimensiunea depinde de programul genetic, de funcția
îndeplinită, de starea fiziologică a organismului și a celulei, de disponibilitatea nutrienților etc.
Cele mai multe celule nu se văd cu ochiul liber, dar există si excepții: celulele de portocală, de pepene verde,
fibrele de in, celulele musculare striate, oul de pasăre etc. Alte exemple: hematia – 7,5 m, ovulul uman – 150-200 m,
fibra musculară striată scheletică – 1-30 cm.
Cele mai multe celule nu au dimensiuni mari din motive practice, cele mai importante fiind comunicarea
intracelulară și schimbul de substanțe (necesitățile metabolice). Pe de o parte, pentru a asigura funcțioarea unitară a
celulei, diferitele zone ale acesteia trebuie să comunice rapid între ele. Comunicarea intracelulară și coordonarea activității
celulare presupun transferul de substanțe (molecule cu diverse roluri – energetic, structural, enzime, tipuri de ARN,
mesageri chimici etc.) între diverse părți ale celulei, iar acest transfer se realizează, în multe cazuri, prin difuzie, un tip de
transport eficient numai pe distanțe mici (de aceea celulele de mari dimensiuni au mai mulți nuclei, fiecare responsabil de
2 R. Hooke a greşit în două privinţe – pereţii celulari pe care i-a văzut nu sunt vii (reprezintă o componentă neprotoplasmatică a celulei), iar celulele nu
sunt goale, de obicei. Cu toate acestea, în al doilea caz, greşeala o reprezintă generalizarea pe care a făcut-o, pentru că, de fapt, celulele de suber pe care
le-a observat Hooke sunt moarte şi nu au conţinut.
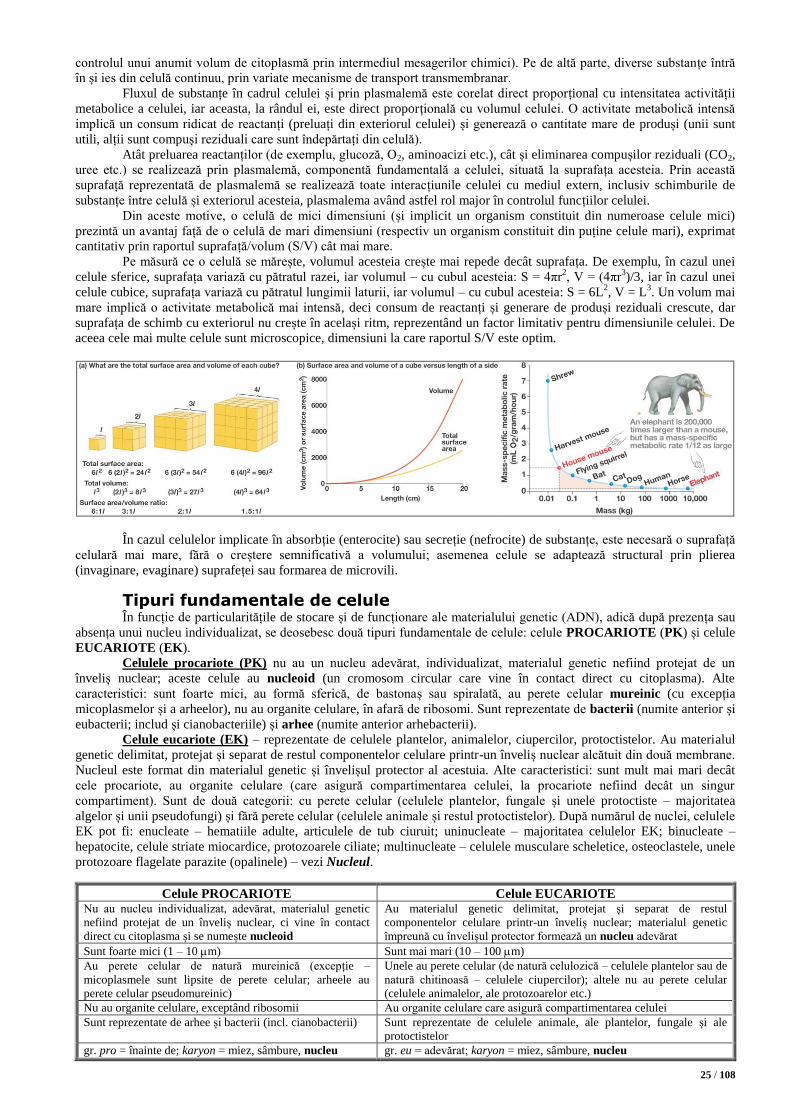
25 / 108
controlul unui anumit volum de citoplasmă prin intermediul mesagerilor chimici). Pe de altă parte, diverse substanțe întră
în și ies din celulă continuu, prin variate mecanisme de transport transmembranar.
Fluxul de substanțe în cadrul celulei și prin plasmalemă este corelat direct proporțional cu intensitatea activității
metabolice a celulei, iar aceasta, la rândul ei, este direct proporțională cu volumul celulei. O activitate metabolică intensă
implică un consum ridicat de reactanți (preluați din exteriorul celulei) și generează o cantitate mare de produși (unii sunt
utili, alții sunt compuși reziduali care sunt îndepărtați din celulă).
Atât preluarea reactanților (de exemplu, glucoză, O2, aminoacizi etc.), cât și eliminarea compușilor reziduali (CO2,
uree etc.) se realizează prin plasmalemă, componentă fundamentală a celulei, situată la suprafața acesteia. Prin această
suprafață reprezentată de plasmalemă se realizează toate interacțiunile celulei cu mediul extern, inclusiv schimburile de
substanțe între celulă și exteriorul acesteia, plasmalema având astfel rol major în controlul funcțiilor celulei.
Din aceste motive, o celulă de mici dimensiuni (și implicit un organism constituit din numeroase celule mici)
prezintă un avantaj față de o celulă de mari dimensiuni (respectiv un organism constituit din puține celule mari), exprimat
cantitativ prin raportul suprafață/volum (S/V) cât mai mare.
Pe măsură ce o celulă se mărește, volumul acesteia crește mai repede decât suprafața. De exemplu, în cazul unei
celule sferice, suprafața variază cu pătratul razei, iar volumul – cu cubul acesteia: S = 4πr2, V = (4πr
3)/3, iar în cazul unei
celule cubice, suprafața variază cu pătratul lungimii laturii, iar volumul – cu cubul acesteia: S = 6L2, V = L
3. Un volum mai
mare implică o activitate metabolică mai intensă, deci consum de reactanți și generare de produși reziduali crescute, dar
suprafața de schimb cu exteriorul nu crește în același ritm, reprezentând un factor limitativ pentru dimensiunile celulei. De
aceea cele mai multe celule sunt microscopice, dimensiuni la care raportul S/V este optim.
În cazul celulelor implicate în absorbție (enterocite) sau secreție (nefrocite) de substanțe, este necesară o suprafață
celulară mai mare, fără o creștere semnificativă a volumului; asemenea celule se adaptează structural prin plierea
(invaginare, evaginare) suprafeței sau formarea de microvili.
Tipuri fundamentale de celule În funcție de particularitățile de stocare și de funcționare ale materialului genetic (ADN), adică după prezența sau
absența unui nucleu individualizat, se deosebesc două tipuri fundamentale de celule: celule PROCARIOTE (PK) și celule
EUCARIOTE (EK).
Celulele procariote (PK) nu au un nucleu adevărat, individualizat, materialul genetic nefiind protejat de un
înveliș nuclear; aceste celule au nucleoid (un cromosom circular care vine în contact direct cu citoplasma). Alte
caracteristici: sunt foarte mici, au formă sferică, de bastonaș sau spiralată, au perete celular mureinic (cu excepția
micoplasmelor și a arheelor), nu au organite celulare, în afară de ribosomi. Sunt reprezentate de bacterii (numite anterior și
eubacterii; includ și cianobacteriile) și arhee (numite anterior arhebacterii).
Celule eucariote (EK) – reprezentate de celulele plantelor, animalelor, ciupercilor, protoctistelor. Au materialul
genetic delimitat, protejat și separat de restul componentelor celulare printr-un înveliș nuclear alcătuit din două membrane.
Nucleul este format din materialul genetic și învelișul protector al acestuia. Alte caracteristici: sunt mult mai mari decât
cele procariote, au organite celulare (care asigură compartimentarea celulei, la procariote nefiind decât un singur
compartiment). Sunt de două categorii: cu perete celular (celulele plantelor, fungale și unele protoctiste – majoritatea
algelor și unii pseudofungi) și fără perete celular (celulele animale și restul protoctistelor). După numărul de nuclei, celulele
EK pot fi: enucleate – hematiile adulte, articulele de tub ciuruit; uninucleate – majoritatea celulelor EK; binucleate –
hepatocite, celule striate miocardice, protozoarele ciliate; multinucleate – celulele musculare scheletice, osteoclastele, unele
protozoare flagelate parazite (opalinele) – vezi Nucleul.
Celule PROCARIOTE Celule EUCARIOTE Nu au nucleu individualizat, adevărat, materialul genetic
nefiind protejat de un înveliș nuclear, ci vine în contact
direct cu citoplasma și se numește nucleoid
Au materialul genetic delimitat, protejat și separat de restul
componentelor celulare printr-un înveliș nuclear; materialul genetic
împreună cu învelișul protector formează un nucleu adevărat
Sunt foarte mici (1 – 10 m) Sunt mai mari (10 – 100 m)
Au perete celular de natură mureinică (excepție –
micoplasmele sunt lipsite de perete celular; arheele au
perete celular pseudomureinic)
Unele au perete celular (de natură celulozică – celulele plantelor sau de
natură chitinoasă – celulele ciupercilor); altele nu au perete celular
(celulele animalelor, ale protozoarelor etc.)
Nu au organite celulare, exceptând ribosomii Au organite celulare care asigură compartimentarea celulei
Sunt reprezentate de arhee și bacterii (incl. cianobacterii) Sunt reprezentate de celulele animale, ale plantelor, fungale și ale
protoctistelor
gr. pro = înainte de; karyon = miez, sâmbure, nucleu gr. eu = adevărat; karyon = miez, sâmbure, nucleu

26 / 108

27 / 108
CELULA PROCARIOTĂ (PK)
Generalități Procariotele, reprezentate în special de bacterii, sunt primele organisme care au apărut pe Terra, acum cca. 3,5
miliarde de ani. În prezent ele se găsesc aproape pretutindeni. Larga lor răspândire se datorează marii lor capacități de a se
adapta la diferite condiții de mediu și de a consuma o gamă foarte diversă de substanțe (exploatează eficient mediul).
Pot metaboliza unele substanțe pe care alte organisme nu le pot metaboliza (cauciuc, mase plastice, detergenți,
substanțe toxice etc.). Au metabolismul foarte ridicat datorită raportului suprafață/volum foarte mare și a faptului că pot
cataboliza o gamă foarte largă de substanțe.
Au durata de generație (de viață) foarte scurtă, divizându-se cu o rată înaltă (de câteva ori pe zi.
În condiții optime de temperatură, umiditate, pH și cu nutrienți adecvați cantitativ și calitativ, bacteria Escherichia
coli are durata de generație de doar 20 de minute.
SARCINĂ DE LUCRU. Calculați numărul de celule care rezultă în asemenea condiții într-o singură zi, ce masă
totalizează, știind că o singură celulă are m = 0,4 fg (1 fg = 10–15
g; fg – femtogram), și ce volum ocupă, știind că o singură
celulă are un volum mediu de 1,12 μm3.
Bacteriile populează cele mai neospitaliere medii, în care alte organisme (eucariotele) nu pot rezista: locuri cu
aciditate, salinitate, temperaturi extreme sau în roci aflate la câțiva (cel puțin doi) km adâncime sub suprafața Terrei.
Cele mai multe PK sunt celule solitare libere, dar există și specii care agregă tranzient sau permanent formând
colonii. Dar toate procariotele sunt unicelulare.
Celula tipică de PK are diametrul de 1-5(10) m (față de 10-100 m cât au celulele eucariote). Excepție face
Thiomargarita namibiensis3 care poate ajunge la diametrul de 750 m.
TEMĂ – căutați pe Internet imagini cu Thiomargarita namibiensis.
Celulele bacteriene aparțin la trei categorii principale de forme,
controlate genetic și determinate, în principal, de peretele celular rigid
(care este însă și suplu, elastic, deci deformabil într-o oarecare măsură):
Formă sferică/sferoidală: bacteriile de formă sferică se
numesc coci (pot fi solitare – coci, se pot grupa câte doi –
diplococi sau mai mulți aranjați liniar – streptococi – sau în
grupuri amorfe, cu aspect de ciorchini de struguri –
stafilococi); tot în această categorie se încadrează și tetradele
(grupuri de patru celule coplanare) și sarcinile (grupuri de opt
celule aranjate tridimensional în forma unui cub).
Formă bacilară/de baston: bacteriile cu această formă se
numesc bacili; extremitățile pot fi rotunjite sau truncate; pot fi
solitare – bacil, grupate câte două – diplobacil sau mai multe,
dispuse liniar – streptobacil, ca șipcile într-un gard – palisadă,
sau ca o rozetă/stea; formele intermediare între coc și bacil se
numesc cocobacili.
Formă spiralată: bacteriile de formă spiralată se numesc spirili (rigizi și cu puține spire) și spirochete (flexibile
și cu multe spire); tot în această categorie întră și vibrionii – bacterii curbate, cu aspect de bumerang sau de
virgulă.
Există și bacterii a căror formă este foarte variată – bacterii pleomorfe/polimorfe, condiție determinată genetic,
cum ar fi speciile din genurile Rhizobium și Corynebacterium.
Elemente de (ultra)structură Structura de referință a celulei PK este peretele celular. În funcție de
localizarea acestora în raport cu peretele celular, componentele celulei PK se
împart în două categorii:
A. Structuri intraparietale – constituie protoplastul și sunt reprezentate
de plasmalemă și derivatele acesteia, citoplasmă, ribosomi, nucleoid
și structuri genetice accesorii, incluziuni, spațiu periplasmic, spor
etc.
B. Structuri extraparietale – glicocalix, flageli, pili, fimbrii, spini etc.
3 Domeniu Bacteria, filum Proteobacteria, clasa γ-Proteobacteria; bacterie Gram-negativă, cocoidă, se grupează în lanţuri de celule cu aspect de şirag de perle ca urmare a prezenţei în celulă a granulelor microscopice de S care reflectă lumina incidentă; chemolitotrofă, acceptorul final de electroni este
nitratul; oxidează H2S la S elemental care se acumulează sub formă de granule microscopice, citoplasma devenind înalt refractilă şi opalescentă, celula
asemănându-se cu o perlă; descoperită în 1997 în sedimentele platoului continental al Namibiei, Golful Walvis şi în 2005 în Golful Mexic

28 / 108
1. Peretele celular bacterian Peretele celular este un înveliș rigid, situat
deasupra plasmalemei, care înconjoară complet celula
bacteriană. Lipsește la micoplasme (PK saprotrofe
din sol și ape menajere sau comensale la om și
animale; au plasmalemă rigidă care le asigură
protecție față de șocul osmotic) și la bacteriile
halofile extreme (PK care trăiesc în medii saline
foarte concentrate). Prin eliminarea experimentală a
conținutului celular (protoplast), peretele rămâne sub
forma unui sac gol și rigid, care păstrează forma
inițială a celulei, având deci rol foarte important în
determinarea și menținerea formei celulei bacteriene.
Are grosime cuprinsă între 15 și 30 (80) nm.
În funcție de grosimea și de
structura/arhitectura peretelui celular există două categorii de bacterii: Gram4-pozitive [Gram (+)] – au perete celular gros,
cu structură relativ simplă, și Gram-negative [Gram (–)] – au perete celular subțire, dar mai complex organizat.
Componenta chimică esențială a peretelui celular este mureina (peptidoglican). Mureina este alcătuită dintr-o
componentă glucidică (predominantă) și una peptidică.
La bacteriile Gram-negative, mureina reprezintă numai 2,5-10% din masa uscată a peretelui (față de 80% la
bacteriile Gram-pozitive), care este însă mult mai complex, datorită prezenței unei structuri caracteristice – membrana
externă, o replică structurală a plasmalemei, care poate fi astfel considerată membrană internă. Membrana externă a
peretelui conține fosfolipide (35%), proteine (l5%) și lipopolizaharide (50%).
La arhee peretele celular prezintă diferențe structurale majore față de eubacterii. Peretele lor nu conține mureină,
ci pseudomureină.
Rolurile peretelui celular
Fiind rigid, determină și menține forma celulei și previne (cito)liza osmotică în mediu hipotonic. Asigură
protecția fizică a celulei. Intervine pasiv (la bacteriile Gram-pozitive) sau activ și selectiv, prin porinele și transportorii
membranei externe (la bacteriile Gram-negative) în permeabilitatea învelișului celular. La bacteriile Gram-pozitive
participă la formarea septului de diviziune. Prin unele componente, amplifică virulența bacteriană și are proprietăți
imunogenice.
2. Plasmalema (membrana plasmatică) Din punct de vedere biochimic este alcătuită din fosfolipide și proteine; sterolii sunt prezenți numai la micoplasme
și la unele arhee, conferind un plus de rigiditate plasmalemei acestora.
Arhitectura și funcțiile sunt foarte asemănătoare cu cele ale plasmalemei celulei EK, care va fi prezentată detaliat
în capitolul dedicat celulei EK.
Plasmalema celulei bacteriene poate prezenta invaginări tubulare, veziculare sau lamelare în citoplasmă: mezosom
și tilacoide.
2.1. Mezosomul („corpul din mijloc”) este o invaginare plasmalemală frecventă și cu aspect lamelar la
bacteriile Gram-pozitive, rară și cu aspect tubular la bacteriile Gram-negative. Mezosomul are numeroase roluri: este
implicat în procesele de biosinteză și de creștere celulară; constituie suport pentru atașarea cromosomului și a plasmidelor;
este implicat în replicarea cromosomului, în segregarea celor doi cromosomi-fii rezultați în urma replicării între cele două
celule-fiice; împreună cu plasmalema participă la procesele energogenetice/respiratorii ale celulei (conține sinergonul respirator),
fiind echivalentul structural și funcțional al mitocondriei.
2.2. Tilacoidele (cromatofori) sunt invaginări tubulare, veziculare sau lamelare care, în general, păstrează
legătura cu plasmalema, prezente la PK fotosintetizante (cianobacterii ș.a.). La nivelul lor este localizat sinergonul
fotosintetizant (moleculele – enzime, pigmenți asimilatori, citocromi etc. – implicate în fotosinteză).
În general, invaginările plasmalemale au rolul de a crește suprafața de reacție a membranei plasmatice.
2.3. Plasmalema arheelor are o ultrastructură tipică (două suprafețe hidrofile, miez hidrofob), dar este
unică prin natura lipidelor.
3. Spațiul periplasmic Spațiul periplasmic este un compartiment celular al bacteriilor Gram-negative, situat între plasmalemă și
membrana externă a peretelui celular (sau între plasmalemă și stratul mureinic al peretelui); bacteriile Gram-pozitive nu au
un spațiu periplasmic propriu-zis, ci un spațiu echivalent, mai îngust.
Acest spațiu conține apă, proteine (enzime hidrolitice/degradative, transportori etc.), oligozaharide și nutrienți.
În spațiul periplasmic se acumulează și se depozitează nutrienți înainte de a intra în citoplasmă (rol adaptativ
foarte important, deoarece bacteriile trăiesc, de obicei, în medii oligotrofe, sărace în nutrienți); la acest nivel, nutrienții sunt
degradați enzimatic parțial, iar produșii rezultați sunt transferați prin plasmalemă în citoplasmă.
4 Hans Christian Joachim Gram (1850-1938), bacteriolog, farmacolog şi medic danez.

29 / 108
4. Glicocalixul bacterian Glicocalixul este stratul gelatinos, vâscos/lipicios, de polizaharide sau/și polipeptide (compoziția chimică variază
cu specia), situat deasupra peretelui celular la multe PK. Este reprezentat de:
o Polizaharidul extracelular (PZE) este alcătuit exclusiv din polizaharide:
dacă este ferm aderent la peretele celular și prezintă organizare omogenă, se numește capsulă.
dacă este slab atașat de peretele celular, cu grosime neuniformă, consistență fluidă și distribuție
dezordonată/neorganizată în jurul celulei, se numește strat mucos.
zooglea este o masă de material polizaharidic în care este inclus un număr mare de celule bacteriene.
o Glicocalixul comportamental este alcătuit dintr-o rețea de polizaharide și glicoproteine, care formează o
structură pericelulară pâsloasă, dezordonată, prin intermediul căreia celula aderă la alte celule sau la substrat
inert.
Glicocalixul bacterian are două roluri principale: (a) ajută PK să adere la substrat sau la alți indivizi cu formare de
colonii și biofilme; (b) participă la inițierea procesului infecțios de către PK patogene5 (asigurând ancorarea acestora de
celulele mucoaselor) și le protejează împotriva sistemului imunitar al gazdei infectate.
5. Fimbriile Fimbriile sunt apendici filiformi, în general scurți și inegali ca lungime (1-20 μm), rigizi sau flexibili, numeroși,
uneori spiralați, prezenți în special la bacteriile Gram-negative, rar la cele Gram-pozitive. Au rol în aderarea PK la substrat
inert sau la alte celule bacteriene în formarea de colonii. Sunt formate din proteina fimbrilină.
De exemplu, Neisseria gonorrhoeae (agentul etiologic al gonoreei – ITS; este o bacterie Gram-negativă, având
forma de bob de cafea; este colonială – diplococ) se atașează la și colonizează mucoasele uretrale și vaginală cu ajutorul
fimbriilor și al filamentelor glicocalixului, cauzând boala.
TEMĂ – căutați informații despre gonoree – etiologie, simptomatologie, tratament, prevenire etc.
6. Pilul Pilul este un apendice filiform, de obicei unic, lung (~20 μm), canaliculat; are rol în fenomenul de conjugare =
transfer de material genetic de o celulă donoare la o altă celulă – celulă acceptoare (conjugarea bacteriană este considerată
un fenomen de parasexualitate). Pilul există la bacteria donoare și are rolul de fixare a bacteriei acceptoare; prin scurtarea
treptată a pilului, cele două celule vor fi aduse în vecinătate permițându-se realizarea transferului de ADN; se mai numește
și pil conjugativ. Este format din proteina pilină.
7. Flagelul Flagelii sunt structuri filiforme lungi, ondulate, semirigide, cu rol în propulsarea celulei bacteriene în mediu lichid.
Lungimea variază între 4 și 70 μm, iar diametrul este relativ constant de 20 nm. Diferă structural și compozițional de
flagelii celulelor eucariote, care sunt mult mai complecși.
Bacteriile lipsite de flageli se numesc atrihe; cele care prezintă flageli sunt de patru tipuri, în funcție numărul și de
modul de dispunere a acestora: monotrihe – un singur flagel polar; amfitrihe – câte un flagel la fiecare pol/extremitate al/a
celulei; lofotrihe – doi sau mai mulți flageli la un pol sau la ambii poli ai celulei; peritrihe – mai mulți flageli distribuiți pe
întrega suprafață a celulei.
Flagelul este constituit din trei componente de bază: filamentul extern, cârligul și corpu(sculu)l bazal.
8. Filamentele axiale Filamentele axiale sunt prezente la spirochete6.
9. Citoplasma Citoplasma este masa celulară apoasă care umple spațiul delimitat de plasmalemă; este densă, vâscoasă (gel fluid),
semitransparentă, elastică, fin granulară, omogenă (la celulele îmbătrânite capătă aspect granular neomogen), fără curenți
citoplasmatici și fără citoschelet tipic.
Citoplasma conține apă (70-80%), proteine (în special enzime și factori implicați în biosinteza de acizi nucleici și de
proteine, dar și proteine structurale), glucide, lipide, acizi nucleici, ioni anorganici (în concentrație mai mare decât în
mediul extern) și numeroși compuși cu masă moleculară mică, fiind astfel un complex sistem coloidal.
În citoplasmă se găsesc materialul genetic (nucleoidul și plasmidele), ribosomi de 70S și incluziuni.
10. Ribosomii Ribosomii de 70S sunt singurele organite celulare comune tuturor PK. Sunt organite corpusculare submicroscopice,
sferoidale, cu diametru de cca. 20 nm, formate din ARNr și proteine ribosomale care se asamblează în două subunități –
mare de 50S și mică de 30S. Aceste subunități se asociază la rândul lor și formează ribosomul funcțional.
Ribosomii sunt localizați în citoplasmă și atașați la nivelul feței interne a plasmalemei. Există în trei stări – ribosomi
solitari liberi în citoplasmă, polisomi (mai mulți ribosomi atașați liniar de o moleculă de ARNm) și subunități ribosmale.
5 De exemplu, Bacillus anthracis produce o capsulă formată din acid D-glutamic (izomer atipic de aminoacid) şi numai tulpinile încapsulate produc boala – antrax (cărbune). 6 Un foarte cunoscut exemplu de spirochetă este Treponema pallidum (agentul etiologic al sifilisului); o altă spirochetă importantă este Borrelia
burgdorferi, agentul etiologic al bolii Lyme.

30 / 108
Ribosmii sunt sediul sintezei proteinelor specifice celulei PK, de aceea numărul lor este corelat cu starea de
activitate fiziologică – este mic în celulele în repaus, dar crește foarte mult în celulele cu activitate intensă, precum cele în
creștere, și este cuprins între 15.000 și 100.000. Biosinteza proteinelor se realizează pe baza mesajului genetic din ARNm.
11. Materialul/aparatul genetic Materialul genetic al bacteriilor este reprezentat de două categorii de structuri: nucleoidul și plasmidele.
11.1. Nucleoidul (cromosom bacterian, material genetic esențial) este liber în citoplasmă și conține gene
esențiale existenței (arhitectură celulară, metabolism, creștere, reglare și coordonare a activităților) și reproducerii
bacteriei. Este o structură omoloagă nucleului celulei eucariote și reprezintă unicul cromosom bacterian. Bacteriile sunt
haploide deoarece într-o celulă există un singur cromosom.
Este reprezentat de o macromoleculă de ADN dublu catenară, circulară, închisă covalent, localizată direct în
citoplasmă, fără a fi protejată sau delimitată de un înveliș nuclear. Acesta se supraspiralizează, se pliază și ocupă centrul
celulei, fiind protejat mecanic de citoplasma densă.
11.2. Structuri genetice accesorii/extracromosomale sunt reprezentate de plasmide =
molecule mici de ADN bicatenar circular, atașate de proteine membranare și care se replică independent de nucleoid.
Plasmidele conțin între cinci și 100 de gene, numite gene accesorii, în general neesențiale pentru existența bacteriei în
medii de viață normale (câștigarea sau pierderea lor nu are efect asupra existenței celulei) și care conțin informație genetică
„de confort” care permite celulei să se adapteze la condiții de mediu noi sau modificate. Asemenea gene sunt implicate în
fenomenul de conjugare bacteriană – plasmidele F (sau plasmide de sex) sau care conțin, de exemplu, gene pentru
rezistența la antibiotice (plasmidele R, plasmidele Col), pentru producerea de toxine, pentru sinteza de enzime. Unele
plasmide pot fi transferate de la o bacterie la alta, iar ADN-ul plasmidial este intens utilizat în tehnicile de inginerie
genetică.
12. Incluziuni citoplasmatice Incluziunile sunt reprezentate de depozite de diverse substanțe (nutrienți, enzime, compuși osmotic activi etc.).
Celulele bacteriene acumulează anumiți nutrienți atunci când aceștia sunt abundenți în mediu și îi folosesc când sunt
deficienți în mediu.
12.1. Granulele metacromatice (de volutină) sunt o rezervă de fosfat anorganic care poate fi folosită în
sinteza de ATP.
12.2. Granulele de polizaharide sunt formate din glicogen sau amidon. Reprezintă rezerve de glucoză,
utilizată în scop energetic.
12.3. Incluziuni lipidice se întâlnesc la numeroase specii de bacterii. 12.4. Granulele de sulf 7
12.5. Carboxisomii sunt incluziuni care conțin enzima RuBisCO – foarte importantă în fotosinteză.
Carboxisomii se întâlnesc la bacteriile fotosintetizante (cianobacterii).
12.6. Vacuolele cu gaz (aerosomi) sunt structuri cavitare prezente la multe bacterii acvatice (în special la cele
imobile), inclusiv cianobacterii, bacterii fotosintetizante anoxigenice și halobacterii.
Rolul principal al vacuolelor este de a asigura celulelor flotabilitatea necesară menținerii lor la adâncimea adecvată
la care au acces la oxigen, lumină și nutrienți, funcționând pe post de corpi de flotație – prin umplere cu gaz, densitatea
specifică a celulei scade, iar celula se deplasează ascendent; prin eliminarea gazului din vacuolă, celula devine mai densă și
se deplasează spre straturile profunde ale apei.
12.7. Magnetosomii sunt incluziuni de oxid de fier (Fe3O4, magnetit) sau/și sulfură de fier (Fe3S4, greigit), sub
formă de cristale, formate de unele bacterii Gram-negative8 (bacterii magnetotactice) conferindu-le capacitatea de a se
orienta și de a migra în lungul liniilor câmpului geomagnetic.
Lectură Biofilme bacteriene În natură microorganismele rar trăiesc în colonii izolate, monospecifice. De obicei trăiesc în comunități multispecifice, multistratificate, vâscoase,
dezvoltate pe suprafețe sau la interfețe și denumite biofilme. Acest fapt a fost serios luat în considerare odată cu dezvoltarea microscopiei confocale care a făcut
posibilă decelarea structurii tridimensionale a biofilmelor in situ, nemodificate prin tehnicile de prelucrare specifice microscopiei electronice. Comunicarea celulă-celulă prin mecanismul de quorum sensing permite bacteriilor să-și coordoneze activitatea și să se grupeze în comunități care le asigură beneficii, în mod similar cu
organismele pluricelulare. Ca urmare, biofilmele nu sunt doar straturi bacteriene vâscoase, ci sisteme biologice dotate cu integralitate și considerate în prezent o
formă primitivă de diferențiere celulară; bacteriile sunt organizate în comunități funcționale, înalt coordonate. Biofilmele sunt de obicei atașate de o suprafață, de un
substrat, care poate fi piatră, lemn, cochilie din mediul acvatic, dinte uman (sub formă de placă dentară sau tartru) sau o membrană mucoasă. Cu toate acestea, unele
comunități de tip biofilm iau forma unor grupări bacteriene particulate plutitoare sau în suspensie. Un exemplu este masa floculentă (formată prin precipitarea sau
agregarea particulelor suspendate) care se formează în anumite tipuri de bazine de epurare a apelor reziduale. În cadrul unei comunități biofilmice bacteriile sunt
capabile să utilizeze în comun nutrienții și sunt mai protejate față de factorii nocivi din mediu, precum desicarea, substanțele antimicrobiene, antibioticele și efectorii
sistemului imunitar al organismului, comparativ cu forma lor alternativă de existență, respectiv cea de celule plutitoare, libere. Strânsa apropiere/proximitate a microorganismelor în cadrul biofilmului facilitează transferul de informație genetică prin, de exemplu, conjugare.
O problemă serioasă a fost de-a lungul timpului definirea conceptului de biofilm; după mai multe încercări, în care erau subliniate diverse criterii
(considerate de autorii definiției drept cele mai semnificative), s-a ajus la o definiție relativ satisfăcătoare, cel puțin pentru acest stadiu al cunoașterii:
7 Sunt specifice bacteriilor sulfuroase chemosintetizante (Beggiatoa, Thiobacillus thiooxidans), care obţin energia necesară desfăşurării proceselor
metabolice prin oxidarea unor compuşi ai S (în special H2S) până la S, care se acumulează în celule sub formă de particule alb-gălbui strălucitoare, având
rol de rezervă energetică deoarece, după epuizarea H2S din mediu, S din celule este oxidat la sulfat (forma cea mai oxidată a S), cu eliberare de energie. Granule de S se întâlnesc şi la bacteriile sulfuroase purpurii (familia Chromatiaceae); acestea fac fotosinteză anoxigenică, folosind H2S (şi nu H2O) ca
donor de electroni, rezultând S ca produs rezidual care se acumulează în celule (CO2 + H2S → substanţe organice + S). 8 Magnetit biogen, similar celui din bacterii, a fost găsit şi la organisme eucariote – euglenoide, somon, porumbei, oameni.

31 / 108
Prin biofilm se înțelege o comunitate microbiană sesilă compusă din celule care sunt în mod ireversibil atașate la un substrat, la o interfață sau unele de altele, care sunt înglobate într-o matrice de substanțe polimerice extracelulare produse de către aceste celule și care prezintă un fenotip modificat, în privința
ratei de creștere și a transcrierii genelor.
Un biofilm începe să se formeze când bacteriile libere (planctonice) încep să adere la o suprafață. Dacă biofilmul ar crește uniform, devenind
multistratificat și compact, nutrienții nu ar fi disponibili în profunzimea acestei structuri; în același timp s-ar acumula metaboliți toxici. Bacteriile din aceste
comunități evită aceste neajunsuri sintetizând substanțe polimerice extracelulare ce formează matricea biofilmului și se diferențiază în structuri columnare între care
se găsesc canale prin care apa poate circula și transporta nutrienții și reziduurile. Această structură reticulară se constituie într-un sistem circulator primitiv. Bacterii
individuale și agregate de celule înglobate în matricea polizaharidică uneori părăsesc biofilmul matur și diseminează, constituind centre de nucleație pentru noi
biofilme, ceea ce se soldează cu extinderea biofilmului inițial. Un biofilm este alcătuit în general dintr-un strat superficial de aproximativ 10 μm grosime, cu coloane care extind până la 200 μm deasupra acestui strat. În apele rapid curgătoare, biofilmele pot lua forma unor filamente unduitoare atașate cu un capăt de substrat.
Substratul, prin proprietățile fizico-chimice, influențează semnificativ adsorbția inițială a microorganismelor, acumularea acestora și agregarea lor în
comunități biofilmice. Dintre aceste proprietăți, cele mai importante ar fi: mărimea suprafeței disponibile, hidrofobicitatea, energia de suprafață, adsorbția
substanțelor ce formează așa-numitul film condiționant, prezența materialelor humice (în mediul extern) și a fibronectinei (în organismul animal), defecte fizice
(structura granulară a suprafeței, fisuri, excrescențe), gradul de coroziune, reactivitatea fotochimică a stratului de oxidare superficială etc.
Dezvoltarea biofilmului până la stadiul de maturitate (caracterizat prin juxtapoziție stabilă și congruență fiziologică între celulele componente) se
realizează treptat, aderența microorganismelor fiind un fenomen lent, stadial și condiționat de numeroși factori. Spre exemplu, microorganismele trebuie să ajungă la o distanță suficientă de substrat încât să permită forțelor van der Waals, interacțiunilor electrostatice și punților de hidrogen să se manifeste și să fie eficiente; iar
această proximitate necesară este asigurată de gravitație, termo- și chimiotactism și de forța și direcția curenților acvatici.
Bacteriile din biofilme pot coopera în îndeplinirea unor sarcini complexe. De exemplu, sistemele digestive ale rumegătoarelor necesită cel puțin cinci
tipuri diferite de specii microbiene care să degradeze celuloza. Acești microbi din sistemul digestiv al rumegătoarelor sunt localizați în cea mai mare parte în
comunități biofilmice. Biofilmele sunt de asemenea elemente esențiale în funcționarea adecvată a sistemelor de epurare a apelor reziduale, dar pot deveni o problemă
în conducte și tuburi, pe care le pot chiar bloca fizic prin dezvoltare exagerată; de asemenea, din biofilme de pot desprinde bacterii libere sau agregate bacteriene
care contaminează fluidul.
Biofilmele sunt un factor important în sănătatea oamenilor. De exemplu, microbii din biofilme sunt probabil de 1000 de ori mai rezistenți la microbicide. Experți de la CDC Atlanta estimează că 65% din infecțiile bacteriene la oameni sunt consecutive formării de biofilme. Cele mai multe infecții nosocomiale sunt
probabil legate de biofilme formate pe catetere. De fapt, biofilmele se formează pe aproape toate dispozitivele medicale utilizate în interiorul corpului, printre care și
valve cardiace mecanice. Biofilmele, care pot include și pe acelea formate de fungi precum Candida, sunt întâlnite în numeroase stări patologice, precum infecțiile
legate de utilizarea lentilelor de contact, cariile dentare si boala periodontala, ca și infecțiile cronice cu Pseudomonas sp. la bolnavii cu fibroză chistică.
Una dintre abordările privind prevenirea formării biofilmelor este încorporarea de substanțe antimicrobiene în suprafețele pe care s-ar putea forma
biofilme. Deoarece semnalele chimice care permit comunicarea prin mecanismul de quorum sensing sunt esențiale în formarea biofimelor, se desfășoară cercetări
pentru elucidarea naturii acestor substanțe și pentru găsirea de modalități de blocare a lor. O altă abordare este descoperirea că formarea biofilmelor poate fi inhibată
de lactoferină, substanță antimicrobiană prezentă în multe secreții ale omului. Lactoferina chelatează fierul, iar lipsa fierului inhibă motilitatea pe suprafață, esențială pentru agregarea bacteriilor în biofilme.
Formarea biofilmelor pe suprafețe pune probleme serioase în numeroase sisteme acvatice, reprezentate în special de afectarea suprafețelor colonizate.
Astfel biofilmele formate reprezintă rezervoare potențial patogene, cauzează coroziunea (principalele incriminate fiind bacteriile sulfat-reducătoare) și deteriorarea
metalelor, pierderi energetice și reducerea eficienței suprafețelor. De exemplu, acumularea de biofilme în conductele de apă din instalațiile de răcire are ca efect
scăderea eficienței transferului termic; biofilmele pot reduce diametrul conductelor până la colmatare; uneori metaboliții biofilmelor pot afecta negativ diverse medii
lichide circulante prin conducte sau depozitate în rezervoare, cum ar fi acidifierea petrolului; împiedică funcționarea eficientă a stațiilor de tratare a apelor reziduale
compromițând filtrele biologice și dispozitivele de aerare. Quorum sensing este un tip de proces decizional utilizat de grupurile descentralizate în vederea coordonării comportamentului. Numeroase specii de
bacterii utilizează acest proces să-și coordoneze expresia genelor în conformitate cu densitatea populațională locală. În mod similar, insectele sociale utilizează
quorum sensing în luarea deciziilor colective privind locul de cuibărire. În plus față de rolurile biologice, quorum sensing are aplicații în informatică și robotică.
Quorum sensing poate funcționa ca proces de luare a deciziilor în orice sistem descentralizat atâta vreme cât componentele individuale:
(a) au mijloace de apreciere a numărului de componente care interacționează
(b) manifestă un răspuns standard odată ce pragul numeric al componentelor a fost atins și detectat
Unele dintre cele mai bune exemple de quorum sensing provin din studii efectuate pe bacterii. Acestea folosesc quorum sensing să-și coordoneze
anumite comportamente în funcție de densitatea locală a populațiilor bacteriene. Quorum sensing se poate realiza atât între indivizi din aceeași specie, ca și între indivizi din specii diferite și poate interveni în reglarea unei game de procese diferite, în esență având rol de rețea de comunicare. Ca semnale pot fi și sunt folosite o
varietate de molecule diferite. Frecvent sunt folosite oligopeptide la bacteriile Gram (+), N-acil-homoserin-lactone (AHL) la bacteriile Gram (–) și o familie de
autoinductori cunoscuți drept AI-2, atât la Gram (+), cât și la Gram (–).
Bacteriile care utilizează constant quorum sensing produc și secretă anumite molecule de semnalizare (numite autoinductori sau feromoni; de aceea,
fenomenul de quorum sensing s-a numit inițial autoinducție). Aceste bacterii au de asemenea un receptor care detectează specific molecula semnalizatoare
(inductorul). Când inductorul se leagă la receptor, activează transcrierea anumitor gene, inclusiv pe cele pentru sinteza inductorului. Probabilitatea ca o bacterie să
întâlnească propriul inductor este foarte mică. De aceea, pentru a fi activată transcrierea, celula trebuie să intâlnească molecule de semnalizare secretate de alte
celule în mediul în care se găsește. Când doar puține alte bacterii de același tip ajung în proximitate, fenomenul de difuzie reduce concentrația inductorului în mediul înconjurător până la aproape zero, astfel că bacteriile produc în continuare puțin inductor. Totuși, pe măsură ce populația crește, concentrația de inductor depășește
un anumit prag, determinând sinteza de și mai mult inductor. Aceasta se constituie într-o buclă de feedback pozitiv și receptorul devine activat maximal. Activarea
receptorului induce reglarea prin stimulare a altor gene specifice, determinând ca toate celulele să înceapă transcrierea aproximativ simultan; acest comportament
coordonat al ceulelor bacteriene poate fi util în diverse situații, precum sinteza enzimei luciferază de către Vibrio fischeri doar atunci când populația este suficient de
mare pentru ca efectul de bioluminiscență să fie evident; sinteza enzimei în condițiile unei populații reduse ar fi consum inutil de energie, atâta timp cât nu s-ar
atinge scopul – bioluminiscența.
Una dintre proprietățile remarcabile ale biofilmelor este faptul că bacteriile integrate în biofilm prezintă fenotipuri total diferite față de aceleași specii planctonice, fenotipuri care explică integralitatea, cooperarea metabolică, rezistența sau recalcitranța sau, mai nou, toleranța la substanțe antimicrobiene naturale sau
artificiale, rezistența la infecții virale/fagice și la atacul protozoarelor. Acestea și multe altele constituie un exemplu elocvent de adaptare, care poate fi chiar premisă
pentru evoluție. În acest sens, pe baza structurii biofilmelor, corelată cu particularitățile funcționale, se vorbește de o formă primitivă de homeostazie a comunității
bacteriene respective.
Biofilmele bacteriene au importantă semnificație medicală, nu în puține cazuri negativă. Omul (dar și alte animale) este în relație de comensalism cu
numeroase specii de microorganisme (bacterii, fungi, protozoare), care constituie așa-numita microbiotă normală a diferitelor regiuni, segmente ale corpului (tub
digestiv, căi respiratorii, căi urinare, tegument, căi genitale etc.). De multe ori prezența acestor organisme, în condiții homeostatice, de sănătate, este benefică,
ajutând organismul gazdă prin activitățile lor (de exemplu, sinteza unor vitamine). Însă dacă se modifică parametrii fizico-chimici din regiunile populate de microorganismele comensale, acestea fie se înmulțesc exagerat, iar activitățile lor sunt depreciante pentru gazdă, fie ajung să fie înlocuite de germeni oportuniști,
potențial patogeni care formează populații mari când întâlnesc condițiile favorabile, fizico-chimice, dar și ecologice (lipsa competitorilor, reprezentați în acest caz de
microbiota normală). În ambele cazuri, microorganismele au preferință pentru forma de organizare în biofilme, ceea ce face foarte dificil de tratat și de readus la
situația normală, din cauza recalcitranței crescute a biofilmelor la substanțele antimicrobiene.
Una dintre cele mai mari probleme pe care le pun biofilmele este impactul negativ pe care îl au asupra activității medicale și sănătății oamenilor – infecțiile
nosocomiale, contaminarea biomaterialelor folosite în medicină sau pur și simplu adaptarea în timp a unor specii la acțiunea substanțelor antimicrobiene, alături de
intrarea în scena patogenității a unor specii care până acum erau fie comensale, fie externe și relativ indiferente în raport cu omul (infecții emergente).

32 / 108
SCRIEȚI LEGENDA SCHEMEI URMĂTOARE

33 / 108
SPAȚIU PENTRU REZOLVAREA TEMELOR ȘI A SARCINILOR DE LUCRU

34 / 108

35 / 108
CELULA EUCARIOTĂ (EK) Celula animală
Celula eucariotă are trei componente de bază: plasmalemă (membrană plasmatică), citoplasmă și nucleu.
Plasmalema Plasmalema sau membrana plasmatică este o foiță fină (7-10 nm), flexibilă, situată la periferia celulei, definind
granițele acesteia.
Din localizarea periferică a plasmalemei rezultă mai multe consecințe/roluri:
participă la determinarea și menținerea formei celulei
protejează celula
separă structurile interne de mediul extracelular => asigură individualitatea celulei, dar nu o izolează, ci ...
... integrează celula în mediu, prin localizare și organizarea sa particulară participând în mod activ/dinamic și
reglator la numeroase activități celulare:
controlează schimburile de substanțe și particule dintre celulă și mediul extern – are proprietatea de
permeabilitate selectivă/semipermeabilitate (adică permite pătrunderea în celulă doar a substanțelor
utile, nu și a celor nocive și nefolositoare)
participă în mod direct la atașarea celulă-celulă (la formarea de țesuturi) și celulă-substrat
este implicată în comunicarea celulară – reprezintă sediul recepției semnalelor din exterior separă cele două mari compartimente lichidiene ale organismului pluricelular – fluidul intracelular (citoplasma) și fluidul extracelular.
Membrana plasmatică este polarizată electric – pozitiv pe fața externă și negativ pe fața internă.
Plasmalema poate prezenta deformări (care implică sau nu și alte componente celulare) permanente (cili,
microvili, stereocili, flageli) sau temporare (pseudopode).
Din punct de vedere biochimic este constituită în principal din (fosfo)lipide și proteine.
Membrana plasmatică se mai numește și membrană celulară, dar este preferată sintagma „membrană
plasmatică”, deoarece multe dintre structurile celulare sunt delimitate de membrane cu arhitectură și compoziție similare
celei care delimitează celula și toate aceste membrane intră în categoria membranelor celulare.

36 / 108
Ultrastructura plasmalemei Modelul mozaicului fluid (Singer & Nicolson, 1972)
Plasmalema este o structură fluidă dinamică, aflată în continuă mișcare. Conform modelului mozaicului fluid9
(elaborat de Seymour Jonathan Singer (1924- ) și Garth L. Nicolson (1943- ) în 1972), plasmalema este o foiță foarte
subțire (7-10 nm) alcătuită dintr-un dublu strat = bistrat de molecule lipidice, de consistența uleiului de măsline, în care
sunt incluse/inclavate proteine. Moleculele lipidice ale bistratului se mișcă liber, executând mișcări laterale și mișcări de
rotație în jurul propriului ax. Unele proteine plutesc în bistratul lipidic fluid, conferind aspectul de mozaic în permanentă
schimbare; altele, în special cele periferice, au mișcări limitate deoarece sunt ancorate de structuri intracelulare care intră în
alcătuirea citoscheletului.
Lipidele membranare constituie componenta structurală de bază a membranei și sunt reprezentate de fosfolipide
(majoritatea) și cantități mici de colesterol și glicolipide (acestea din urmă doar pe fața externă a plasmalemei).
O moleculă de fosfolipid are formă de acadea fiind alcătuită dintr-un cap globular încărcat electric = polar și deci
hidrofil (aceste capete polare, fiind „iubitoare de apă”, vin în contact cu cele două medii apoase care scaldă fețele externă și
internă ale membranei – mediul extracelular și citosolul) și din două „cozi” nepolare, deci hidrofobe și lipofile, reprezentate
de molecule de acizi grași. Cozile nepolare, fiind hidrofobe, evită apa și se dispun în centrul membranei (al bistratului
fosfolipidic). Rezultă astfel o structură de tip „sandwich” (comună tuturor membranelor biologice) reprezentată de două
foițe paralele de molecule de fosfolipide orientate „în oglindă” sau „coadă la coadă”, capetele polare fiind expuse mediilor
apoase din interiorul și din afara celulei. Moleculele de tipul fosfolipidelor care au o regiune polară și hidrofilă și o regiune
nepolară și hidrofobă se numesc molecule amfipatice/amfipolare. Cele două monostraturi de fosfolipide ale plasmalemei diferă prin tipurile și cantitatea de lipide pe care le conțin. Cele mai multe fosfolipide
membranare sunt nesaturate (precum fosfatidil-colina), o stare în care au cozile mai puțin flexibile și curbate
(crescând spațiul dintre ele), fapt care determină creșterea fluidității membranei. Glicolipide (fosfolipide care
au grupări glucidice atașate covalent) sunt prezente doar pe fața externă a plasmalemei și reprezintă aproximativ 5% din totalul lipidelor membranare. Resturile glucidice ale glicolipidelor, ca și grupările fosfat
ale fosfolipidelor, conferă polaritate capetelor glicolipidelor, în timp ce cozile de acizi grași sunt nepolare.
Colesterolul reprezintă aproximativ 20% din lipidele membranare, moleculele sale reprezentate de inele plane
fiind dispuse printre cozile fosfolipidelor. Aproximativ 20% din suprafața externă a plasmalemei conține domenii lipidice, ansambluri
dinamice de fosfolipide saturate, strâns împachetate, asociate cu sfingolipide și cu mari cantități de colesterol. Aceste regiuni lipidice dense sunt mai stabile și ordonate și mai puțin fluide decât restul membranei și pot
conține diverse tipuri de proteine în diverse cantități. Ca urmare a acestor particularități compoziționale și
arhitecturale, domeniile lipidice sunt considerate regiuni membranare în care se concentrează moleculele implicate în semnalizarea celulară.
*Fosfolipidele au proprietatea de a se
auto-orienta unele față de altele și față de apă; ca
urmare a acestei proprietăți, fosfolipidele se pot
autoasambla în structuri închise, sferice (cu zona
centrală hidrofobă, constituită din regiunile
nepolare – cozile de AG – ale moleculelor de FL,
din care a fost îndepărtată apa, și suprafața
hidrofilă, constituită prin juxtapunerea regiunilor
polare ale FL – capetele globulare ale acestora),
numite micele, care se refac rapid în cazul în care sunt perturbate mecanic.
9 În 1915 s-a stabilit că membranele celulare sunt alcătuite din lipide şi proteine. În 1925 olandezii E. Gorter şi F. Grendel propun organizarea
fosfolipidelor sub formă de bistrat, singura care putea să explice proprietatea de graniţă stabilă între două medii apoase. În 1935, Hugh Davson şi James Danielli completează modelul de membrană sugerând că proteinele, de tip globular, sunt localizate pe cele două feţe ale bistratului fosfolipidic, imaginând
astfel modelul trilaminar de tip „sandwich” – un bistrat fosfolipidic între două straturi continue de proteine. Acest model a fost susţinut şi dezvoltat şi de J.
David Robertson, care a propus în 1959 modelul membranei unitare (unit membrane).

37 / 108
Proteinele membranare – care constituie aproximativ 50% din componența membranei – sunt de două mari
categorii: integrale și periferice (unii autori disting și o a treia categorie – proteine ancorate de membrană, dar pot fi asimilate proteinelor
integrale). Sunt responsabile de majoritatea funcțiilor particulare ale plasmalemei.
Proteinele integrale sunt ferm atașate (prin legături covalente) și în general scufundate în bistratul lipidic. Unele
proemină doar pe o față a membranei, însă cele mai multe sunt transmembranare, adică străbat întreaga grosime a
bistratului, proeminând pe ambele fețe ale membranei. Ca și lipidele, prezintă regiuni hidrofobe (care interacționează/vin în
contact cu cozile fosfolipidelor din partea/zona centrală a membranei) și regiuni hidrofile (care vin în contact cu capetele
fosfolipidelor și cu mediile apoase intra- și extracelular).
Proteinele transmembranare sunt implicate în principal în transport. Unele formează canale sau pori prin care
trec molecule mici hidrosolubile sau ioni, evitând astfel miezul hidrofob al bistratului fosfolipidic. Altele acționează drept
cărăuși/carrier care leagă o substanță pe o față a membranei și apoi o translocă prin membrană fără a forma un por, ci prin
modificarea conformațională a proteinei transportoare. Alte proteine sunt receptori pentru hormoni sau pentru alți
mesageri chimici și conduc mesajele din exterior în celulă – proces numit transducția semnalului.
Proteinele periferice nu sunt scufundate în bistratul lipidic și sunt slab atașate (nu prin legături covalente, ci
ionice, van der Waals sau de hidrogen) de lipidele membranare sau de proteinele integrale. În această categorie este inclusă și o
rețea de filamente de pe fața internă a membranei (rețeaua de spectrină, prezentă doar la unele tipuri celulare) și care participă la susținerea mecanică a
acesteia. Unele proteine periferice sunt enzime, altele sunt implicate în realizarea
rolurilor mecanice ale membranei – schimbarea formei celulei în timpul diviziunii
celulare și în contracția musculară sau atașarea celulă-celulă.
Plasmalema este o structură fluidă dinamică aflată în mișcare continuă. Are
consistența uleiului de măsline. Moleculele lipidice ale bistratului se mișcă liber – rotație în jurul
propriului ax și lateral, paralel cu suprafața membranei; caracterul amfipatic/amfipolar le împiedică să să dea peste cap și să treacă dintr-un monostrat în celălalt, deci monostraturile își păstrează compoziția relativ
constantă și asimetrică (există și asemenea mișcări – de flip-flop – dar sunt foarte rare și catalizate
enzimatic). Unele proteine plutesc liber în bistrat. Altele, în special cele periferice, au mișcări limitate deoarece sunt fixate de structuri intracelulare care intră în alcătuirea citoscheletului.
Funcțiile proteinelor membranare
1. Transportori de diferite substanțe în și din celulă; transportorii sunt selectivi
și specifici pentru o anumită substanță; unii sunt saturabili – au o capacitate
maximă de transport pe unitatea de timp; sunt reprezentați de canale ionice,
translocatori (proteine-carrier = cărăuși) și pompe.
2. Enzime – sunt biocatalizatori ai multor reacții biochimice/metabolice din
celulă.
3. Receptori – leagă la suprafața externă a membranei mesageri chimici și
transmit mesajul acestora în celulă.
4. Unele proteine au rol de marker-i celulari – sunt recunoscuți de alte celule;
funcționează ca un fel de amprente sau semne particulare de identificare.
5. Rol structural – unele proteine realizează conectarea structurală a două
celule învecinate (formând joncțiuni de ancorare de tipul desmosomilor, al
joncțiunilor înguste etc.); alte proteine se atașează la elementele moleculare
proteice ale matricei extracelulare și la elementele de citoschelet,
stabilizând astfel poziția membranei și implicit forma celulei și asigurând
coordonarea mișcărilor celulare.
Glicocalixul reprezintă învelișul lipicios și „dulce” (gr. glykys – dulce, calima
– înveliș) de la suprafața externă a membranei celulare; este alcătuit din resturile
oligoglucidice ramificate, atașate covalent de lipidele și proteinele membranare
(formând glicolipide și glicoproteine). Aceste glicolipide și glicoproteine sunt
sintetizate de către celulă.
Deoarece fiecare celulă are un alt tip de glucide în alcătuirea glicocalixului,
acesta constituie un marker biologic înalt specific prin intermediul căruia celulele
care vin în contact se recunosc reciproc. De exemplu, spermatozoidul recunoaște ovocitul prin intermediul glicocalixului
unic al ovocitului și celulele sistemului imunitar identifică o bacterie ca fiind străină corpului legându-se la anumite
glicoproteine membranare ale glicocalixului bacterian; de asemenea, cele patru grupe sangvine din sistemul ABO/ABH – O, A, B și AB –
sunt determinate de diversitatea resturilor oligoglucidice din alcătuirea glicocalixului.
Funcțiile plasmalemei
Transport (trans)membranar Celulele organismelor pluricelulare (ale animalelor) sunt scăldate de un fluid – fluidul interstițial sau intercelular
– care derivă din sânge și poate fi descris drept o „supă” foarte bogată în substanțe nutritive (are mii de ingrediente –
aminoacizi, glucide, acizi grași, vitamine, substanțe reglatorii precum hormonii, neurotransmițătorii, factorii de creștere,
săruri) și produși reziduali. Pentru a funcționa optim, celulele trebuie să extragă din acest amestec tipurile de substanțe și în
cantitățile precise necesare la un moment dat. Traficul de substanțe prin membrană este selectiv sau diferențial, ca urmare a
proprietății de barieră selectiv permeabilă a plasmalemei – permite trecerea unor substanțe și o împiedică pe a altora.

38 / 108
Astfel, permite nutrienților să intre în celulă, dar reține la exterior substanțele nedorite. În același timp, reține proteinele
celulare și alte substanțe utile în celulă și permite reziduurilor metabolice să iasă.
Substanțele traversează plasmalema în două moduri de bază – pasiv sau activ, clasificare care are la bază
implicarea energetică a celulei în realizarea transportului transmembranar. În transportul pasiv substanțele traversează
membrana fără consum energetic din partea celulei. În transportul activ celula asigură energia metabolică (ATP)
necesară deplasării substanțelor prin membrană.
I. Transportul pasiv Este reprezentat de două tipuri principale – difuzie și filtrare. Difuzia este o modalitate de transport membranar
pasiv întâlnită la toate celulele corpului. Filtrarea are aplicabilitate mai restrânsă, petrecându-se doar la nivelul pereților
capilarelor (membrană tisulară poroasă).
A. Difuzia Difuzia este tendința moleculelor sau a ionilor de a se distribui uniform în mediul
pe care îl au la dispoziție. Toate moleculele au energie cinetică și sunt în continuă mișcare și
de aceea se îndepărtează din zonele în care se găsesc în concentrație mare (energia cinetică
totală, rezultată prin însumarea energiilor cinetice ale tuturor moleculelor din zona
respectivă, este mare) în zonele în care concentrația lor este mică (energia cinetică totală
este mai mică), adică difuzează în sensul gradientului de concentrație sau, exprimat plastic,
se deplasează „la vale”. Viteza difuziei nete este direct proporțională cu mărimea diferenței
de concentrație, adică a gradientului.
Deoarece forța propulsoare pentru difuzie este energia cinetică a moleculelor însele, viteza difuziei este
influențată de mărimea moleculei (molecule mici → viteză mare) și de temperatură (temperatură mai ridicată → viteză
mai mare).
Trebuie precizat că simpla concentrație, care poate fi echivalată cu energia cinetică totală a moleculelor dintr-o
anumită zonă a spațiului, nu presupune în mod obligatoriu transport (difuzie) și are mai degrabă valoare de energie
potențială a unui transport. Această energie se va manifesta doar dacă
va exista o a doua zonă a spațiului în care concentrația moleculelor este
mai mică, adică dacă există un gradient de concentrație și, evident, dacă
mediul permite deplasarea moleculelor (dacă este permeabil). Acest
gradient are valoarea unei forțe (chimice) care asigură transportul din
zona cu concentrație mare în zona cu concentrație mică. Rezumând,
concentrația singură este o sursă de energie, iar gradientul de
concentrație este forța care determină transportul. Într-un volum închis, efectul final al difuziei va fi un amestec omogen de molecule, situație în care sistemul a
atins echilibrul – moleculele se deplasează în mod egal în toate direcțiile posibile, ceea ce echivalează cu absența unei
mișcări nete (în aceste condiții, nu mai există gradient de concentrație care, se spune, s-a descărcat/epuizat treptat prin
transportul pe care l-a susținut).
Prin miezul său hidrofob, plasmalema este o barieră fizică împotriva difuziei libere. Prin plasmalemă vor difuza
totuși anumite molecule: (1) cele liposolubile, indiferent de dimensiunea lor; (2) cele hidrofile, dar suficient de mici (ioni
minerali) pentru a putea trece prin canale proteice membranare, numite și canale ionice; (3) cele hidrofile mari, care vor fi
asistate/ajutate de molecule cărăuși.
În funcție de particularitățile substanțelor difuzibile, se deosebesc trei tipuri majore de difuzie:
1. Difuzia simplă/termică – difuzia neasistată a particulelor liposolubile;
2. Difuzia facilitată – difuzia asistată de proteine membranare;
3. Osmoza – difuzia neasistată a solventului (de obicei apa) printr-o membrană selectiv permeabilă.
Difuzia simplă. Substanțele nepolare și liposolubile (precum oxigenul, dioxidul de carbon, vitaminele
liposolubile, ureea, etanolul) difuzează direct prin bistratul lipidic (miezul hidrofob nu opune rezistență). Oxigenul, fiind
permanent în concentrație mai mare în sânge decât în celule, va difuza continuu din sânge în celule, în timp ce dioxidul de carbon, care este în concentrație
mare în celule, va difuza din celule în sânge.
Difuzia facilitată. Anumite molecule, în special glucoză și alte monoglucide, aminoacizi, și ioni, sunt
transportate pasiv chiar dacă, fiind hidrofile, nu pot străbate direct bistratul fosfolipidic. Asemenea molecule trec prin
membrană printr-un tip de transport pasiv numit difuzie facilitată/asistată. În acest tip de difuzie, substanța transportată fie
se leagă la proteine cărăuși din membrană (precum cei din familia transportorilor de glucoză – GLUT) și este translocată
prin intervenția acestora, fie se deplasează prin proteine-canal cu por apos.
Cărăușii. Un cărăuș este o proteină integrală transmembranară care prezintă specificitate pentru moleculele
anumitor substanțe organice polare (deci hidrofile) care sunt prea mari pentru a trece prin canalele ionice
membranare, precum glucidele și aminoacizii. Cel mai acceptat model funcțional al procesului de
translocare a acestor substanțe presupune o modificare în forma cărăușului care îi pemite mai întâi să lege și
să acopere și apoi să elibereze substanța transportată, protejând-o pe traseu de regiunile nepolare ale
membranei. Situsul de legare a substanței transportate se deplasează de pe o față a membranei pe cealaltă
prin modificarea conformațională a proteinei-cărăuș. Ca și în difuzia simplă, substanța transportată cu
ajutorul cărăușului, precum glucoza, se deplasează în sensul gradientului de concentrație. Glucoza este, în
mod normal, în concentrație mai mare în sânge decât în celule, unde este rapid consumată pentru sinteza de

39 / 108
ATP. Astfel, transportul glucozei este de obicei unidirecțional – intră în celulă. Transportul mediat de cărăuși
este limitat de numărul de proteine transportoare prezente în membrană. De exemplu, când toți cărăușii
pentru glucoză sunt ocupați, se spune că sunt saturați și transportul glucozei se petrece cu rată maximă
(Tmax). Se spune că difuzia facilitată prin intermediul cărăușilor este saturabilă.
Canalele. Canalele sunt proteine transmembranare cu rol de transport de substanțe, de obicei ioni (sau apă, în
cazul unor canale speciale), prin canale apoase, dintr-o parte în alta a membranei. Canalele sunt selective
prin mărimea porului și prin sarcina aminoacizilor care mărginesc canalul/porul și pot fi permanent deschise
(numite canale de scurgere) și permit fluxul neîntrerupt de ioni sau apă în concordanță cu gradienții de
concentrație sau prezintă poartă și sunt controlate (închise sau deschise) de diferite semnale (chimice,
electrice, mecanice, luminoase etc.). Ca și cărăușii, multe canale pot fi inhibate de anumite molecule,
prezintă saturație și tind să fie specifice. Deplasarea prin canal a substanțelor se face în sensul gradientului
de concentrație. În cazul traversării membranei prin difuzie simplă, rata de difuzie nu este controlabilă deoarece nu se poate modifica rapid solubilitatea în
lipide a substanței transportate. În cazul difuziei facilitate, rata de difuzie este controlabilă, deoarece permeabilitatea membranei pentru substanțele
transportate poate fi modificată prin reglarea activității cărăușilor și a canalelor.
Oxigenul, apa, glucoza și diverși ioni sunt vitali pentru homeostazia celulară. De aceea, transportul lor pasiv prin
difuzie – simplă sau facilitată – reprezintă o economisire semnificativă a energiei celulare, al cărei consum al crește
exponențial dacă ar fi transportate activ.
Osmoza (gr. osmos – împingere). Reprezintă difuzia solventului, de obicei apa, printr-o membrană selectiv
permeabilă.
Osmoza are loc ori de câte ori concentrația soluților în cele două compartimente separate de o membrană selectiv
permeabilă (care nu permite trecerea soluților) este diferită, implicit va fi diferită și „concentrația” apei (concentrație de
soluți mare → concentrația apei mică).
Gradul în care concentrația apei este influențată de soluți depinde de numărul și nu de tipul particulelor de solut,
deoarece o moleculă sau un ion de solut teoretic înlocuiește o moleculă de apă. De fapt, cu cât molecula sau particula este
mai mare, cu atât numărul de molecule de apă pe care le înlocuiește în soluție este mai mare.
Osmolaritatea = numărul de osmoli per litru de soluție (concentrația totală a particulelor de solut per litru de
soluție); se măsoară în osmoli/l; plasma sangvină, fluidul interstițial și citoplasma au osmolaritatea de cca. 300 mosm/l.
Osmolalitatea = numărul de osmoli de solut per kilogram de apă.
Un osmol este definit drept un mol de particule dizolvate. Dacă particulele respective nu disociază ionic în
solvent, atunci molul și osmolul au aceeași valoare (ex., o soluție de glucoză 1M are osmolaritatea 1 osm/l). Dacă solutul
disociază ionic, va genera două sau mai multe particule dizolvate în soluție, iar concentrația molară și osmolaritatea nu vor
mai fi egale (ex., o soluție de NaCl 1M va avea osmolaritatea 2 osm/l, deoarece din fiecare particulă de NaCl rezultă, prin
disociere în apă, două particule – Na+ și Cl
–, deci numărul total de soluți va fi dublu; o soluție de CaCl2 1M va avea
osmolaritatea 3 osm/l, deoarece dintr-o particulă de CaCl2 vor rezulta, prin disociere în apă, trei particule – Ca2+
și 2Cl–).
Atunci când volume egale de soluții apoase cu osmolarități diferite sunt separate de o membrană permeabilă
pentru toate moleculele din sistem, se va petrece difuzia netă atât a apei cât și a solutului, fiecare deplasându-se în sensul
propriului gradient de concentrație. În final, se atinge echilibrul când atât concentrația solutului, cât și a apei sunt egale în
cele două compartimente separate de membrană.
Dacă analizăm același sistem, dar membrana este impermeabilă pentru moleculele de solut, rezultatul va fi
diferit. Apa va difuza rapid din compartimentul în care osmolaritatea este mai mică (concentrație mică de soluți,

40 / 108
deci concentrație mare a apei) în cel cu osmolaritate mai mare (concentrație mare de soluți, deci concentrație mică a
apei) până când concentrațiile (a apei și a soluților) de pe cele două fețe ale membranei devin egale – echilibrul în
acest caz se atinge doar prin deplasarea apei, nu și a solutului. Apa se va deplasa de la concentrație mică de soluți, deci
mare de apă, la concentrație mare de soluți, deci mică de apă. Deplasarea doar a apei va determina modificări, uneori
semnificative, ale volumului celor două compartimente separate de membrană. În condiții experimentale, volumele celor două
compartimente separate de membrana semipermeabilă sunt infinit expandabile și de aceea modificarea volumului nu are nici un efect asupra presiunii lichidului.
Cu toate că moleculele de apă sunt polare, acestea traversează direct bistratul lipidic prin osmoză. Acest fapt este
surprinzător, deoarece ne așteptăm ca moleculele de apă să fie respinse de miezul hidrofob al membranei. O posibilă
explicație este că moleculele de apă au dimensiuni foarte mici raportat la cele ale cozilor hidrofobe ale FL și de aceea se pot
strecura printre ele (cozile hidrofobe nu tolerează cantități mari de apă în jurul lor, dar în acest caz moleculele individuale
de apă nu staționează și nu se acumulează în vecinătatea cozilor). O altă posibilă explicație este că mișcările randomice ale
lipidelor membranare determină formarea unor mici goluri/spații între cozile lor oscilante, permițând apei să se strecoare
prin membrană deplasându-se de la un „buzunar” la altul.
Apa se deplasează liber și reversibil și prin canale specifice pentru apă, formate de proteine transmembranare,
numite acvaporine (AQP), foarte numeroase în plasmalema eritrocitelor și a celulelor implicate în echilibrul hidric al
organismului, precum nefrocitele (celule de la nivelul nefronului – unitatea structurală și funcțională a rinichiului, sediul
formării urinei).
Pentru caracterizarea cantitativă a osmozei se folosește ca parametru presiunea osmotică (proprietate coligativă
a unui amestec; proprietate coligativă – care depinde numai de numărul și de natura moleculelor substanței). Aceasta este
definită drept presiunea care trebuie aplicată pe membrana semipermeabilă, dinspre compartimentul mai concentrat
în soluți, pentru a preveni osmoza; este deci o contrapresiune ipotetică la adresa presiunii hidrostatice generată de apa
care tinde să intre în compartimentul mai concentrat. Presiunea osmotică este direct proporțională cu concentrația
compartimentului în care intră apa (π~c, π = cRT).
Două compartimente sau soluții cu aceeași osmolaritate sunt izoosmotice; dacă au osmolarități diferite, soluția
mai concentrată este hiperosmotică, iar cea mai diluată este hipoosmotică.
Tonicitatea este capacitatea unei soluții de a schimba forma și volumul sau tonusul celulelor prin modificarea
volumului apei interne (tono – tensiune).
Soluțiile cu aceeași concentrație de soluți nedifuzibili precum cea din celule (soluție salină 0,9% sau glucoză 5%)
sunt izoton(ic)e. Celulele aflate într-un asemenea mediu își păstrează forma și nici nu câștigă, nici nu pierd apă. Fluidele
extracelulare și majoritatea soluțiilor administrate intravenos sunt izotonice.
Soluțiile cu o concentrație de soluți nedifuzibili mai mare decât a citosolului sunt hipertonice. Celulele imersate
în asemenea soluții pierd apă prin osmoză și se micșorează și se zbârcesc – crenează.
Soluțiile care sunt mai diluate decât citosolul se numesc hipotonice, iar imersarea celulelor în asemenea soluție
determină preluarea rapidă de apă prin osmoză, umflarea și chiar spargerea celulei (citoliză), dacă volumul citosolului
crește peste un anumit prag. Apa distilată este un exemplu extrem de hipotonicitate – nu conține soluți și apa va intra în
celulă până când aceasta se va sparge (citoliză/liză osmotică). Riscul de citoliză este cu atât mai mare cu cât diferența de
osmolaritate între citosol și mediul extracelular este mai mare.
Turgescență, plasmoliză, deplasmoliză. În cazul celulelor vegetale vii, care au perete celular rezistent, ± rigid,
deasupra plasmalemei, plasarea în soluții cu diferite tonicități are alte efecte comparativ cu celula animală. La celulele
vegetale, membranele selectiv permeabile sunt plasmalema și tonoplastul/membrana vacuolară (sunt, cu mici excepții,
liber permeabile doar pentru apă); peretele celular este permeabil atât pentru apă, cât și pentru majoritatea soluților, de
aceea nu intervine în osmoză.
O celulă vegetală imersată într-o soluție izotonică (cu aceeași osmolaritate ca și sucul celular/vacuolar),
indiferent de starea din momentul imersării, își echilibrează osmolaritatea cu cea a mediului extern => celulă flacidă
(presiunea netă care acționează la nivelul plasmalemei este nulă).
Imersarea unei celule vegetale într-o soluție hipertonică determină efluxul de apă din celulă (în special din sucul
vacuolar) prin osmoză => scade treptat volumul protoplastului (celula vegetală fără perete celular) => începe să se
desprindă plasmalema de peretele celular, deoarece peretele, fiind relativ rigid, nu se poate deforma la fel ca și protoplastul
=> stadiul de plasmoliză concavă: protoplastul are aspect ± stelat, deoarece plasmalema s-a desprins doar pe anumite
porțiuni și este încă atașată în mai multe puncte de peretele celular; între punctele de atașare protoplastul are aspect concav.
Prin efluxul în continuare al apei, volumul protoplastului scade și mai mult => stadiul de plasmoliză convexă: protoplastul

41 / 108
capătă aspect sferoidal, elipsoidal sau lenticular, deoarece plasmalema s-a desprins complet de peretele celular. Între
peretele celular și protoplastul contractat se găsește lichid hipertonic.
Dacă o celulă vegetală plasmolizată (parțial – plasmoliză concavă sau total – plasmoliză convexă) este
imersată într-o soluție hipotonică (sau se înlocuiește soluția hipertonică inițială, care a determinat plasmoliza, cu apă
distilată sau cu o soluție hipotonică), se petrece deplasmoliza: protoplastul va prelua apă prin osmoză => crește volumul
protoplastului => protoplastul se dilată => plasmalema contactează peretele celular, protoplastul revine treptat la forma
inițială, ocupând tot spațiul delimitat de perete.
Dacă o celulă vegetală flacidă este imersată într-o soluție hipotonică => influx de apă în protoplast (în special în
vacuolă) prin endosmoză => protoplastul se dilată => volumul de lichid intracelular apasă asupra plasmalemei (implicit și
asupra peretelui celular) => plasmalema și peretele se tensionează => în celulă se dezvoltă o presiune hidrostatică internă
– presiune de turgescență/de membrană – care se manifestă
asupra plasmalemei și peretelui și se opune influxului de apă
prin osmoză => celulă rigidă. Dezvoltarea și mai ales menținerea
presiunii de turgescență sunt posibile datorită peretelui celular
rezistent care limitează expansiunea rapidă a protoplastului și nu
permite spargerea celulei. O asemenea celulă umflată și rigidă
se numește celulă turgidă, iar starea care o caracterizează –
stare de turgescență.
Cu cât osmolaritatea intracelulară (cantitatea de soluți
nedifuzibili din celulă) este mai mare, cu atât presiunea osmotică intracelulară este mai mare și cu atât mai mare trebuie să
fie presiunea hidrostatică de turgescență care trebuie să se dezvolte la nivelul plasmalemei și peretelui pentru a se opune
influxului net de apă în celulă. Asemenea modificări majore în presiunea hidrostatică (și osmotică) nu se petrec în celulele animale, lipsite de pereți celulari rigizi.
Dezechilibrul osmotic determină celula animală să se umfle sau să se micșoreze (prin câștig, respectiv prin pierdere de apă), fie până când se echilibrează
osmotic cu mediul extracelular, fie până când membrana este tensionată până la sau chiar dincolo de punctul de rupere.
Osmolaritatea și tonicitatea sunt proprietăți diferite ale unei soluții. Osmolaritatea descrie pe concentrația
totală a soluților, iar tonicitatea descrie efectele soluției asupra volumului celulei, care depinde de concentrația de
soluți și de permeabilitatea pentru soluți a plasmalemei. Osmoza este extrem de importantă în determinarea distribuției apei în diverse compartimente lichidiene ale organismului (celule, sânge ș.a.).
Osmoza continuă până când presiunile osmotică și hidrostatică de la nivelul membranei sunt egale. De exemplu, apa este scoasă din capilarele sangvine de
presiunea hidrostatică a sângelui asupra peretelui capilarului, dar prezența în sânge a soluților care sunt prea mari să traverseze peretele capilarului
determină atragerea apei înapoi în sânge și ca urmare pierderea netă de plasmă este foarte mică.
Difuzia simplă și osmoza se petrec direct prin plasmalemă și nu sunt procese selective. În aceste procese, trecerea unei
molecule prin membrană depinde de mărimea sa sau de solubilitatea în lipide și nu de particularitățile structurale. Difuzia facilitată pe de altă parte
este înalt selectivă. Transportorul pentru glucoză se combină specific cu glucoza, în mod similar interacției unei enzime cu substratul
său, iar canalele ionice permit trecerea doar a anumitor ioni, fiind deci selective.
B. Filtrarea Filtrarea este procesul prin care apa și soluții sunt forțați de către presiunea hidrostatică să treacă printr-o
membrană poroasă (perete capilar). Ca și difuzia, filtrarea este un transport pasiv și implică o sursă de energie (presiunea
hidrostatică) și un gradient (o forță), care, în acest caz, este gradientul presional care împinge fluidul cu soluți – filtratul –
dintr-o zonă cu presiune mare într-o zonă cu presiune mică.
În organism, presiunea hidrostatică a sângelui determină ieșirea fluidului din capilar, fluid care conține soluți
esențiali pentru țesuturi. Filtrarea este responsabilă și de formarea urinei la nivelul nefronilor. Filtrarea este selectivă doar
relativ la dimensiunea particulelor, nu și la natura lor chimică – în cazul sângelui, sunt reținute în capilar doar elementele
figurate și proteinele prea mari pentru a trece prin porii peretelui.
II. Transportul activ Atunci când o celulă utilizează energia legăturilor covalente macroergice ale ATP pentru a deplasa soluți prin
membrană, procesul de transport este considerat activ. Substanțele care traversează activ plasmalema nu pot să treacă în
direcția necesară prin transport pasiv. Aceste substanțe sunt fie prea mai mari pentru a trece prin canale, fie incapabile să
se dizolve în bistratul lipidic (sunt deci hidrofile), fie nu trebuie să se deplaseze în sensul gradientului lor de
concentrație (numit și gradient chimic), electric sau electrochimic (adică transportul lor trebuie făcut „la deal”,
împotriva gradientului).
Se descriu două mecanisme majore de transport membranar activ – transportul activ propriu-zis și transportul
vezicular.
A. Transportul activ propriu-zis Ca și difuzia facilitată mediată de cărăuși, necesită proteine membranare translocatoare care să se combine
specific și reversibil cu substanța de transportat. Totuși, difuzia facilitată se realizează întotdeauna în sensul gradienților de
concentrație, care îi asigură și energia cinetică și forța necesare transportului. Transportorii activi, numiți și pompe de
soluți, deplasează soluții, în principal ioni (Na+, K
+, Ca
2+) „la deal”, împotriva gradientului de concentrație sau
electrochimic și pentru a realiza acest lucru celulele trebuie să consume energie de la ATP.
Transportul activ propriu-zis este de două tipuri, în funcție de sursa directă de energie folosită:
1. Transport activ primar – se realizează prin pompe ionice (pompa de Na+-K
+, pompa de Ca
2+, pompa de H
+
etc.), iar energia provine direct din ATP și este eliberată prin hidroliza acestuia chiar de către pompă (ATP + H2O → ADP
+ Pi + Energie).

42 / 108
2. Transport activ secundar (cotransport) – transportul este realizat cu
ajutorul energiei stocate în gradienți ionici creați și menținuți prin operarea unei
pompe de transport primar (este deci folosită indirect tot energia eliberată prin hidroliza
ATP). Pompa consumă direct ATP pentru a crea și menține un gradient ionic, deci o
parte din energia ATP-ului se transferă și se depozitează în respectivul gradient. Prin
descărcarea pasivă a acestui gradient (susținută de propria sa energie, provenită din
ATP) este susținut transportul altei substanțe (glucoză, galactoză, zaharoză, aminoacizi,
alți ioni etc.), de obicei împotriva gradientului ei chimic sau electrochimic. Sistemele de
transport activ secundar sunt sisteme cuplate, adică asigură trecerea simultană prin
membrană a două substanțe diferite. Dacă cele două substanțe sunt transportate în
același sens, sistemul este simport; dacă cele două substanțe sunt transportate în sensuri
opuse prin membrană, sistemul de transport este antiport. Pentru a înțelege acest
transport, vă gândiți la următoarea situație: apa trebuie pompată la deal (și pompa consumă energie electrică, dar care nu se
pierde, ci se stochează ca energie potențială chiar în apa transportată la înălțime) și va curge singură la vale (eventual
punând în mișcare o turbină sau roată de apă), pe seama energiei sale potențiale care devine astfel cinetică. Energia de
mișcare a apei în curgerea ei la vale este, de fapt, energia electrică folosită de pompă pentru a deplasa apa „la deal”. Transport activ primar. În acest tip de transport, prin hidroliza ATP se eliberează o grupare
fosfat care va fi atașată proteinei transportoare (care este astfel fosforilată); consecutiv fosforilării, proteina
transportoare suferă o modificare conformațională în așa fel încât împinge prin membrană solutul legat de ea.
Cel mai studiat și cunoscut sistem de transport activ primar este pompa de sodiu-potasiu, care este un transportor proteic cu funcție enzimatică, numit de aceea și Na+-K+ ATP-ază. Concentrația ionilor de
K în celulă este de aproximativ 10-40 ori mai mare decât în mediul extracelular, iar în cazul ionilor de Na,
situația este exact invers. Aceste diferențe de concentrații ionice sunt esențiale pentru ca celulele excitabile precum celulele musculare și neuronii să funcționeze normal, dar și pentru toate celelalte celule ale corpului
pentru a-și menține volumul citosolic în limite normale. Deoarece atât Na+, cât și K+ se deplasează lent, dar
continuu prin canalele pasive (canale de scurgere) din plasmalemă, în sensul gradienților lor de concentrație, pompa de Na+-K+ funcționează ± continuu ca un sistem antiport scoțând Na+ din celulă și introducând K+,
ambii ioni fiind astfel transportați împotriva gradienților lor de concentrație. Alt exemplu de transport activ
primar este reprezentat de pompele de calciu care asigură sechestrarea activă a calciului ionic din citosol în organite specifice (precum REN) sau îl elimină din celulă.
Transport activ secundar. O singură pompă energizată de ATP, precum pompa de Na+-K+,
poate indirect să asigure transportul activ secundar al mai multor soluți. Prin scoaterea din celulă prin membrană a sodiului împotriva gradientului său de concentrație, pompa depozitează energie în gradientul ionic
creat și menținut. Atunci, ca și apa care la deal trebuie pompată, dar va curge singură la vale (eventual punând
în mișcare o turbină sau roată de apă), o substanță pompată prin membrană poate realiza lucru util când revine pasiv, în gradient de concentrație, în compartimentul în care este în concentrație mai mică. Astfel, odată cu
sodiul care revine în celulă cu ajutorul unei proteine cărăuș (difuzie facilitată) sunt antrenate sau
contransportate în același cărăuș și alte substanțe. De exemplu, unele glucide, aminoacizi și numeroși ioni sunt contransportați în acest mod din lumenul intestinal în enterocite. Cu toate că ambele substanțele
contransportate se deplasează pasiv, Na+ trebuie să fie pompat în lumenul intestinului pentru a se menține
gradientul de difuzie. Gradienții ionici pot fi folosiți și pentru energizarea sistemelor antiport precum cele care ajută la reglarea pH-ului citosolic care utilizează gradientul de Na+ pentru a elimina din celulă ioni de
hidrogen.
Indiferent dacă energia este asigurată direct (transport activ primar) sau indirect (transport activ secundar),
fiecare pompă membranară sau contransportor transportă doar substanțe specifice. Astfel, sistemele de transport activ
asigură o cale prin care celula să fie foarte selectivă în cazul în care substanțele nu pot trece prin difuzie – dacă nu există
pompă, nu se realizează transport.
B. Transportul vezicular Particule mari, macromolecule și fluide sunt transportate transmembranar prin transport vezicular, realizat cu
participarea de regiuni mari de membrană care constituie vezicule în care se găsesc substanțele de transportat. Transportul
vezicular este mai multe tipuri.
I. Exocitoză – externalizarea de diferite substanțe din celulă, împachetate în prealabil în vezicule delimitate de o
membrană tipică. Dacă substanțele sunt produse special de către celulă pentru a fi eliminate, având diverse roluri în mediul
extracelular, procesul de externalizare se numește secreție (hormoni, neurotransmițători, mucus etc.). Dacă substanțele
eliminate sunt reziduuri metabolice, procesul de externalizare se numește excreție. Mecanismul este același pentru ambele
procese: substanța este închisă într-o veziculă membranară, care fuzionează cu plasmalema și apoi se rup ambele în punctul
de contact, conținutul fiind eliberat la exteriorul celulei; membrana veziculei se adaugă plasmalemei.
II. Endocitoza – internalizarea de particule (bacterii, fragmente de celule, virusuri, fire de praf etc.) și de
substanțe (utile sau inutile, chiar dăunătoare – toxine) cu ajutorul unor vezicule delimitate de membrană. Este de trei tipuri:
fagocitoză, pinocitoză și endocitoză mediată de receptori.

43 / 108
II.1. Fagocitoza (endocitoză de fază solidă, phagein – a mânca). O particulă externă de dimensiuni mari
(proteine foarte mari, bacterii, resturi de celule moarte, virusuri, fire de praf) este înconjurată de un pseudopod (evaginare
plasmalemală) și este închisă într-o veziculă = fagosom. Cu acest fagosom eventual fuzionează câțiva lizosomi (organite
celulare care conțin enzime digestive hidrolitice care funcționează la pH acid => hidrolaze acide) => fagolizosom în care
este realizată degradarea/digestia particulei; compușii utili sunt absorbiți în citoplasmă, resturile sunt, de cele mai multe ori,
eliminate prin exocitoză. Fagocitoza a apărut în evoluție ca proces de hrănire (amibe) și a căpătat rol de apărare (este
folosită de unele leucocite și de macrofagele de la animale pentru a apăra organismul de agenți particulați nocivi – bacterii,
virusuri, resturi de celule etc.).
II.2. Pinocitoza (endocitoză neselectivă de fază fluidă, pinein – a bea). Plasmalema se scufundă/invaginează în
dreptul unei picături de fluid extern care conține soluți de mici dimensiuni; picătura (solventul cu soluții) se scurge în
invaginare, marginile acesteia fuzionează, formându-se o veziculă plină cu lichid – pinosom – care fuzionează eventual cu un endosom.
Este întâlnită la majoritatea celulelor; este importantă în internalizarea soluților dizolvați de către celulele absorbtive din
rinichi și intestin.
II.3. Endocitoza mediată de receptori (pinocitoză înalt selectivă). Unele substanțe externe se leagă de receptori
membranari și se formează în aceste zone ale plasmalemei invaginări, apoi vezicule – endosomi. Este o modalitate de a
crește probabilitatea internalizării unor substanțe necesare, dar care sunt în concentrație foarte mică în mediul extracelular.
În absența receptorilor membranari pentru aceste substanțe, probabilitatea de a fi internalizate respectivele substanțe, prin
pinocitoză, ar fi direct proporțională cu concentrația lor. Este mecanism de preluare (internalizare) a unor hormoni,
colesterol, Fe și a majorității macromoleculelor de către celule.
III. Trafic intracelular – unele substanțe circulă între diversele compartimente (organite) ale celulei
împachetate în vezicule (învelite într-o proteină – coatomer) – se desprind prin înmugurire dintr-un organit (ex., RER), se
deplasează prin citoplasmă până la alt compartiment (AG), fuzionează cu membrana acestuia, iar conținutul veziculei se
descarcă la acest nivel.
IV. Transcitoză = endocitoză + trafic intracelular + exocitoză, desfășurate succesiv (ca în cazul unor celule
endoteliale prin care trec substanțe din sânge în fluidul intercelular și invers).

44 / 108
TRANSPORT TRANSMEMBRANAR – REZUMAT
Proces Sursă de
energie Descriere Exemple
Dif
uzie
Difuzie termică
(simplă)
Energia
cinetică a
particulelor
Deplasarea netă a particulelor (ioni, molecule etc.) dintr-o zonă cu
concentrație mare într-o zonă cu concentrație mică, în sensul
gradientului de concentrație
Traversarea regiunilor lipidice ale
plasmalemei de către lipide, O2, CO2,
uree, etanol
Difuzie facilitată La fel ca și difuzia simplă, dar substanța care difuzează este atașată
de o proteină-carrier membranară sau se deplasează printr-o
proteină-canal din membrană
Intrarea în celulă a glucozei, fructozei
și a unor ioni
Osmoză
Difuzia simplă a apei printr-o membrană selectiv-permeabilă (cu
permeabilitate selectivă) de la concentrație mică de soluți (soluție
diluată) la concentrație mare de soluți (soluție concentrată)
Mișcarea (fluxul) apei în și din celulă
direct prin faza lipidică a
plasmalemei sau prin pori
membranari (acvaporine)
Filtrare Presiune
hidrostatică
Mișcarea apei și a soluților printr-o membrană semipermeabilă
(prin plasmalemă sau printre celule) dintr-un compartiment cu
presiune hidrostatică mare într-un compartiment cu presiune
hidrostatică mai mică, adică în sensul gradientului presional
Fluxul de apă, nutrienți și gaze prin
pereții capilarelor; ultrafiltrarea
glomerulară de la nivel renal
(formarea urinei primare)
Proces Sursă de
energie Descriere Exemple
Tra
nsp
ort
acti
v Transport activ
primar ATP
Transport de substanțe împotriva gradientului de concentrație sau a
gradientului electrochimic; presupune traversarea plasmalemei de
către soluți cu ajutorul unor pompe care utilizează direct energia
rezultată din hidroliza ATP
Ioni (Na+, K+, H+, Ca2+ ș.a.)
Transport activ
secundar
(cotransport)
Gradientul
concentrației
ionice
menținut cu
ATP
Cotransportul (transportul cuplat) a doi soluți prin membrană;
energia este asigurată de către gradientul ionic creat de o pompă de
soluți activă primar (adică energia este asigurată indirect);
transportul simport asigură deplasarea celor doi soluți în același
sens; transportul antiport asigură deplasarea celor doi soluți în
sensuri opuse prin membrană
Mișcarea soluților polari sau
încărcați electric: intrarea în celulă a
aminoacizilor prin transport simport;
ieșirea din celulă a Ca2+, H+ prin
transport antiport
Transport vezicular Sursă de
energie Descriere Exemple
Exocitoză
ATP
Eliminarea (secreție sau excreție) de substanţe din celulă;
substanţa este închisă într-o veziculă membranară, care fuzionează
cu plasmalema şi apoi se rupe, eliberând conţinutul la exteriorul
celulei
Secreţia neurotransmiţătorilor, a
hormonilor, a mucusului; eliminarea
din celulă a reziduurilor metabolice
Endocitoză (prin vezicule
calimate în
clatrină)
Fagocitoză (endocitoză
de fază
solidă)
O particulă externă de dimensiuni mari (proteine, bacterii, resturi
de celule moarte) este înconjurată de pseudopode (evaginări
plasmalemale) şi este închisă într-o veziculă (= fagosom) învelită
cu o proteină – clatrină
La om, este întâlnită la fagocitele cu
rol imunitar (unele leucocite şi
macrofagele). La organismele
unicelulare (amibe) are rol în hrănirea
acestora
Pinocitoză (endocitoză
de fază
fluidă)
Plasmalema se invaginează în dreptul unei/sub o picătură de fluid
extern alcătuit din soluţi de mici dimensiuni; marginile membranei
fuzionează, formându-se o veziculă plină cu lichid (= pinosom);
vezicula este învelită cu o proteină – clatrină
Este întâlnită la majoritatea celulelor;
este importantă în preluarea soluţilor
dizolvaţi de către celulele absorbtive
din rinichi şi intestin
Endocitoză
mediată de
receptori
Endo- şi transcitoză selective; substanţele externe se leagă de
receptori membranari şi se formează în aceste zone ale
plasmalemei invaginări (apoi vezicule) învelite în clatrină
Mecanism de preluare (internalizare)
a unor hormoni, colesterol, Fe şi a
majorităţii macromoleculelor de către
celule
Trafic vezicular
intracelular (prin vezicule
calimate în coatomer)
Vezicule învelite într-o proteină – coatomer – înmuguresc şi se
desprind din unele organite şi se deplasează către alte organite
unde îşi descarcă conţinutul
Mecanism responsabil de aproape tot
traficul intracelular de molecule
TIP DE
TRANSPORT
CRITERII
DIFUZIE
SIMPLĂ
TRANSPORT MEDIAT DE PROTEINE
TRANSPORT PASIV TRANSPORT ACTIV
CANAL DIFUZIE
FACILITATĂ PRIMAR SECUNDAR
Sensul fluxului net
În sensul
gradientului
electrochimic
În sensul gradientului
electrochimic
În sensul gradientului
electrochimic
Împotriva gradientului
electrochimic
Împotriva gradientului
electrochimic
Necesitatea
proteinelor transportoare
Nu Da – canal ionic Da – translocator Da – pompă Da – pompă și
translocator
Necesitatea
ATP
Nu Nu Nu Da Da
Sursa de energie
Energia cinetică
a particulelor
transportate
Energia cinetică a
particulelor transportate
Energia cinetică a
particulelor transportate
ATP sau altă sursă
chimică
Gradientul electrochimic
al altui solut
Saturație Nu Uneori Da Da Da
Specificitate Nu Da Da Da Da
Caracterul substanței
transportate
Hidrofob
(nepolar)
Hidrofil (ionizat sau
polar)
Hidrofil (ionizat sau
polar)
Hidrofil (ionizat sau
polar)
Hidrofil (ionizat sau
polar)
Exemple Acizi grași, O2,
CO2
Ioni anorganici (Na+, K+,
Cl-, Ca2+)
Molecule organice
(glucoză)
Ioni anorganici (Na+, K+,
H+, Ca2+)
Molecule organice și ioni
anorganici (glucoză,
aminoacizi, H+, Ca2+)

45 / 108
Citoplasma
Citoplasma este materialul celular dintre plasmalemă și nucleu. La microscopul optic citoplasma are aspectul unui
material gelatinos, omogen, nestructurat. Însă microscopul electronic a relevat că citoplasma, în funcție de gradul de
organizare, este constituită din trei elemente majore: citosolul, organitele și incluzi(un)ile.
1. CITOSOLUL (hialoplasma, citoplasma fundamentală, citoplasma
nestructurală, matricea citoplasmatică) este fluidul ± vâscos, semitransparent în care sunt
localizate („suspendate”) celelalte componente ale citoplasmei și nucleul.
Este un amestec complex cu proprietăți atât de amestec coloidal, cât și de soluție.
Soluția citosolică este formată în cea mai mare parte din apă în care sunt dizolvate glucide,
aminoacizi, săruri minerale și o diversitate de alți soluți organici și anorganici; această soluție
reprezintă mediu de dispersie în care se găsesc coloizi organici (în special proteine; particule hidrofile cu diametrul de 1-
1000 nm, care nu ionizează în apă și care nu sedimentează), numiți micele, care reprezintă faza dispersată.
În hialoplasmă există o rețea de filamente proteice, numită citoschelet, care asigură forma celulei și mișcarea
ordonată a celorlalte componente celulare (unii consideră citoscheletul parte a citosolului deoarece este reprezentat de o
rețea de proteine dispersate în acesta, alții îl consideră organit celular).
Citosolul este sediul multor procese metabolice anaerobe (ex., glicoliza – descompunerea glucozei, în 10 etape,
până la două molecule de acid piruvic, cu producere de doi moli de ATP/mol de glucoză; fermentația lactică;
fermentația alcoolică etc.).
2. ORGANITELE/ORGANELELE CELULARE sau citoplasmatice sunt structuri diferențiate ale
citoplasmei (compartimente celulare specializate) care îndeplinesc diverse funcții în cadrul celulei. Compartimentele
celulare asigură medii locale care permit desfășurarea unor funcții metabolice specifice, astfel încât procese metabolice
altfel incompatibile se pot desfășura simultan în aceeași celulă. Această compartimentare este crucială pentru funcționarea
celulei. Fără ea, mii de enzime ar fi amestecate randomic și activitatea biochimică ar fi haotică.
I. În funcție de prezența în celule și de importanța lor în ansamblul activităților celulare, organitele celulei animale
sunt de două categorii:
A. Comune tuturor celulelor și vitale pentru existența acestora: ribosomi, centrosom, citoschelet, RE (REN și
RER), aparat Golgi (dictiosom), lizosomi, peroxisomi, mitocondrii.
B. Specifice – prezente în anumite celule în care sunt esențiale pentru anumite funcții: miofibrile (în celulele
musculare striate), neurofibrilele și corpii Nissl/tigroizi (în neuron), organite/structuri locomotorii
[pseudopode, cili, flageli (pulselă – situat în partea posterioară a celulei; prin mișcare împinge celula; tractelă
– situat în partea anterioră a celulei; prin mișcare trage celula), cirri, membrane ondulante], alte structuri –
microvili (rol de absorbție).
II. Din punct de vedere structural, organitele celulare ale celulei animale sunt de două categorii:
corpusculare/particulate și veziculare/membranare.
A. Organitele corpusculare/particulate sunt lipsite de un înveliș membranar – ribosomii, centrosomul
(centrul celular), citoscheletul.
B. Organite veziculare/membranare: prezintă un înveliș membranar (cu compoziție și organizare similare
plasmalemei; învelișul membranar permite acestor organite să mențină un mediu intern diferit de citosolul
care le înconjoară) care delimitează o cavitate/lumen; sunt de două categorii:
a) cu înveliș unimembranar – reticul endoplasm(at)ic (RE) – neted (REN) și rugos/granular (RER/REG),
aparat Golgi, lizosomi, peroxisomi.
b) cu înveliș dublu-membranar – mitocondriile.
Celula vegetală prezintă o serie de particularități:
1. Centrosomul are o organizare particulară – este lipsit de centrioli
2. Prezintă perete celular celulozic
3. Prezintă o vacuolă centrală mare permanentă, delimitată de un înveliș unimembranar (numit tonoplast)
4. Prezintă organite specifice veziculare, cu înveliș dublu-membranar – plastidele (dintre care cele mai
răspândite și cunoscute sunt cloroplastele – sediul fotosintezei)
II. ORGANITE COMUNE
A. ORGANITE CORPUSCULARE
1. Ribosomii (granulele lui Palade10)
Sunt corpusculi (particule, granule) sferici denși foarte mici (submicroscopici) alcătuiți din proteine ribosomale și
un tip de ARN – ARNr. Fiecare ribosom (cu mărimea totală de 80S) este alcătuit din două subunități globulare inegale –
una mare, de 60S (μ = 2800 kDa) și una mică de 40S (μ = 1400 kDa). Cele două subunități sunt asamblate din componentele
moleculare (ARNr și proteine ribosomale) în nucleol, părăsesc nucleul și abia în citoplasmă cele două subunități se
asamblează formând ribosomul funcțional.
10 George Emil Palade (19 noi. 1912 – 8 oct. 2008) – savant american de origine română. A descoperit ribosomii împreună cu Albert Claude și Christian
de Duve, descoperire pentru care au primit Premiul Nobel pentru Medicină și Fiziologie în 1974.

46 / 108
Subunitatea mare conține trei tipuri de ARNr – ARNr 5S, ARNr 5,8S și ARNr 28S – și aproximativ 49 de proteine ribosomale; subunitatea mică este formată dintr-un singur tip de ARN – ARNr 18S – și aproximativ 33 de proteine ribosomale.
Există două categorii de ribosomi – liberi, în suspensie în citosol [uneori atașați mai mulți de o
singură moleculă de ARNm, formând poli(ribo)somi] și atașați/legați la suprafața unei membrane (a
reticulului endoplasmatic rugos (RER) și nucleare externe).
ROL Ribosomii sunt sediul sintezei de proteine prin corelarea mesajului genetic cu structura
primară a proteinelor.
2. Centrosomul (centrul celular) Este un organit celular unic (care însă se dedublează în vederea diviziunii celulare) localizat în vecinătatea
nucleului, spre partea anterioară a celulei.
Dpdv structural este alcătuit dintr-o pereche de corpi cilindrici numiți centrioli orientați spațial perpendicular unul
pe celălalt, înconjurați de o zonă mai densă de citoplasmă numită centrosferă/matrice centrosomală, bogată în proteine
numite tubuline (prin polimerizarea cărora se alungesc microtubulii). Un centriol este un cilindru gol pe interior al cărui
perete este alcătuit 9 triplete de microtubuli (localizate deci la periferia cilindrului) – model „9(3)+0” („nouă triplete
periferice și nici un microtubul central”, prin analogie cu modelul structural al cilului/flagelului).
Centriolii ajută la asamblarea și organizarea microtubulilor, dar nu sunt esențiali pentru această funcție la toate
eucariotele. Celula vegetală are microtubuli bine organizați, deși centrosomul este lipsit de centrioli.
ROLURI 1. Poziția lui în celulă indică polul anterior al acesteia.
2. Intervine în diviziunea celulară formând fusul de diviziune.
3. Este centrul de organizare a microtubulilor (COMT) al celulelor EK.
4. *Rădăcina flagelară (situată în citoplasmă și care ancorează flagelul) are structură similară unui centriol.
3. Citoscheletul La începuturile microscopiei electronice, biologii considerau că organitele celulei eucariote plutesc liber în citosol, sub formă de suspensii.
Perfecționarea tehnicilor de microscopie optică și electronică a dus la evidențierea citoscheletului – o rețea de structuri fibrilare proteice care se întinde
prin întreaga citoplasmă. Citoscheletul este alcătuit din trei tipuri de elemente fibrilare – microtubuli, filamente intermediare și microfilamente (filamente de actină)
și are rol important în organizarea structurală și a activității celulei.
3.1. Microtubulii Se găsesc în citoplasma tuturor celulelor eucariote. Sunt structuri cilindrice goale pe interior, cu
diametrul extern de 24-25 nm, diametrul intern de 14-15 nm și cu lungimea de la 200 nm la 25 μm. Peretele unui
microtubul este alcătuit din proteine globulare numite tubuline. Tubulina este un heterodimer alcătuit din două subunități polipeptidice ușor diferite – α- și β-tubulină. Tubulinele se asamblează în protofilamente prin așezare
„cap-la-coadă” [(αβ)(αβ)(αβ)(αβ)(αβ)......], iar 13 asemenea protofilamente dispuse circumferențial (după direcția
generatoarelor cilindrului), alipite, formează peretele microtubulului. Protofilamentele sunt astfel aranjate încât unei subunități de α-tubulină dintr-un protofilament îi corespunde câte o subunitate de β-tubulină în filamentele
învecinate, ceea ce conferă microtubulului aspect spiralat.
Un microtubul crește în lungime prin adăugarea de heterodimeri de tubulină la capetele tubulului (care diferă prin rata de polimerizare => microtubulul este polarizat: capăt + și capăt –). Microtubulii pot fi
dezasamblați/depolimerizați rapid, iar tubulina utilizată pentru construirea de microtubuli în altă parte a celulei, în
funcție de necesități.
ROLURI 1. Au rol structural
i. Susțin mecanic celula (sunt rezistenți la compresie) și participă la menținerea formei acesteia. ii. Intră în alcătuirea centriolilor (și a fusului de diviziune), cililor, flagelilor, corpilor bazali.
2. Sunt implicați în motilitate
i. Intracelulară 1. Deplasarea organitelor – constituie căile (similare unor șine de tren sau cabluri
de funicular) pe care se deplasează organitele echipate cu proteine motorii (ex.,
kinezina), orientând și limitând direcțiile de deplasare (de ex., microtubulii
orientează veziculele de secreție de la AG la plasmalemă).
2. Mișcarea cromosomilor în timpul diviziunii celulare, prin edificarea fusului de
diviziune (vezi Reproducerea celulară) ii. Celulară – realizate prin intervenția cililor și flagelilor.
3. Participă la formarea plăcii celulare în citokineza la celulele vegetale.
Cili și flageli La eucariote, un aranjament specializat de microtubuli este responsabil de structura și funcționarea flagelilor și
cililor – apendici locomotori prezenți la unele celule.

47 / 108
Multe organisme eucariote unicelulare se deplasează prin mediul apos cu ajutorul cililor sau al flagelilor; flageli se
întâlnesc și la unele celule (reproducătoare sau/și somatice/vegetative) de la animale, alge, plante și ciuperci. Când cilii sunt
prezenți la celule fixate într-un țesut, au rolul de a deplasa fluid și particule pe suprafața țesutului respectiv. De exemplu,
celulele ciliate care căptușesc traheea determină pe suprafața mucoasei un curent unidirecțional în sens ascendent
(spre căile respiratorii superioare) de mucus care a reținut diverse resturi din aerul inspirat, asigurând astfel îndepărtarea lor din plămâni. În tractul reproducător al femeii, celulele ciliate care căptușesc oviductele (trompele
falopiene/uterine) ajută la deplasarea ovocitului sau a proembrionului către uter.
Numărul cililor la suprafața unei celule este de obicei mare, spre deosebire de cel
al flagelilor, care este limitat la 1-câțiva/celulă. Cilii și flagelii au același diametru
(aproximativ 0,25 μm), dar diferă prin lungime – cilii au 2-20 μm, iar flagelii sunt de obicei
mult mai lungi (10-200 μm), putând uneori depăși lungimea celulei.
Flagelii și cilii diferă și prin modelul de bătaie. Flagelul prezintă o mișcare
ondulată care generează forța pe aceeași direcție cu axa flagelului. Cilii lucrează asemenea
vâslelor, efectuând bătăi alternative de avans și revenire care generează forță motrice pe o
direcție perpendiculară pe axa cilului.
Cu toate că diferă ca lungime, număr/celulă și model de bătaie, cilii și flagelii au
aceeași ultrastructură: un filament axial/central format din microtubuli, numit axonemă,
acoperit de un înveliș membranar reprezentat de o extensie a plasmalemei. Partea centrală a structurii flagelului – axonema – este alcătuită din 9 dublete de
microtubuli dispuși periferic și circular în jurul unui dublet central. Fiecare dublet periferic
este alcătuit din doi microtubuli alipiți care au zona de atașare comună, astfel un microtubul
este complet (13 molecule de tubulină pe circumferință, adică 13 protofilamente) și notat A,
iar celălalt este incomplet (doar 11 molecule de tubulină, adică 11 protofilamente) și notat
B. Perechea centrală este formată din doi microtubuli nealipiți și deci compleți. Acest
aranjament, numit modelul „9(2)+2”, se întâlnește la aproape toți flagelii și cilii celulelor
eucariote.
Amintiți-vă structura flagelului de la procariote. Comparați cu structura flagelului
de la eucariote. Cele 10 dublete de microtubuli (nouă periferice și unul central) sunt conectate
structural și solidarizate de proteine flexibile care fac axonema să semene, în aspect
transversal, cu o roată de căruță – dubletele periferice împreună cu proteinele care se extind
între ele (nexină și proteina motorie dineină axonemală) formează marginea roții, iar
spițele roții sunt reprezentate de alte proteine (numite spițe radiale) care unesc fiecare
dublet periferic cu teaca internă a dubletului central. Cei doi microtubuli centrali sunt uniți
printr-o punte – puntea microtubulilor centrali (central microtubule bridge). Acest
aranjament se repetă periodic în lungul axonemei.
Ansamblul de microtubuli dintr-un flagel/cil este ancorat în celulă la nivelul
corpului bazal – identic structural cu un centriol.
3.2. Filamentele intermediare Filamentele intermediare sunt structuri fibrilare proteice cu diametru de 8-12 nm, intermediar între microtubuli și microfilamente, și cu
compoziție chimică variată. Există mai multe tipuri de filamente intermediare, fiecare tip fiind construit din subunități moleculare diferite (clasificate în șase tipuri: I – keratine
acide; II – keratine bazice; III – desmină, periferină, vimentină, GFAP-proteină glială fibrilară acidă; IV – α-internexină, neurofilamente, sinemină/desmulină, sincoilină; V – lamine A și B; VI –
nestină (unii autori o includ în tipul IV); plus proteine asociate – filagrina, plectina etc.), care se asamblează prin supraîncolăcire în cabluri mai groase. Sunt elementele citoscheletale care constituie o armătură intracelulară stabilă (chiar după ce celulele mor, adesea rețeaua de filamente
intermediare persistă – exemplu: stratul cel mai extern al epidermului pielii este alcătuit din celule moarte pline cu keratină), specializată să reziste la
acțiunea forțelor mecanice de tracțiune (să suporte tensiunea, ca și microfilamentele). Diferitele tipuri de filamente intermediare funcționează ca eșafodaj
al întregului citoschelet.
ROLURI
1. Susțin mecanic celula (sunt rezistenți la tensiune/tracțiune) și participă la menținerea formei acesteia. 2. Fixarea poziției unor organite – nucleul este localizat într-o cușcă făcută dintr-o împletitură de filamente intermediare și menținut pe
poziție prin ramificații care se extind în citoplasmă.
3. Constituie lamina nucleară (formată din proteine numite lamine) care căptușește învelișului nuclear ranforsându-l; se dezorganizează la începutul diviziunii celulare și se reface la finalul acesteia.
3.3. Microfilamentele (filamentele de actină) Microfilamentele sunt structuri fibrilare solide, de aproximativ 7-8 nm în diametru, prezente în toate celulele
eucariote. Se mai numesc filamente de actină deoarece sunt constituite din molecule de actină – proteină globulară contractilă, actină G. Un microfilament (actină F) este alcătuit din două lanțuri simple, formate dintr-o succesiune liniară de
subunități/molecule de actină G, răsucite unul în jurul celuilalt.
În afară de forma de filamente lungi, microfilamentele se pot organiza în rețele structurale tridimensionale datorită prezenței unor proteine de legătură (actinină, filamină, profilină, gelsolină, fibrilină etc.) situate în lungul
filamentelor. Împreună cu miozina constituie aparatul contractil al celulelor eucariote. Un microfilament crește în lungime prin adăugarea de monomeri de actină G la ambele capetele (care diferă prin
rata de polimerizare => microfilamentul este polarizat: capăt + și capăt –). Microfilamentele pot fi dezasamblate/depolimerizate rapid, iar actina G
utilizată pentru construirea de filamente în altă parte a celulei, în funcție de necesități.
ROLURI 1. Au rol structural
i. Susțin mecanic celula (sunt rezistente la tensiune/forțe de tracțiune) și participă la menținerea formei acesteia. ii. Participă la ranforsarea plasmalemei (și implicit la menținerea formei celulei) formând o rețea corticală/subplasmalemală care
conferă cortexului celulei (ectoplasmei, strat citoplasmatic periferic) consistență semisolidă de gel (față de starea mai fluidă a
citoplasmei centrale – endoplasmă).

48 / 108
iii. Intră în alcătuirea microvililor – au în regiunea centrală un mănunchi de microfilamente. 2. Sunt implicate în motilitate
i. Intracelulară – cicloza (flux citoplasmatic) – un curent circular de citoplasmă
caracteristic celulelor vegetale mari, cu rol de accelerare a distribuirii
substanțelor și organitelor în celulă; este determinată atât de interacțiunea
actină-miozină, cât și de conversia sol-gel a citoplasmei (datorată asamblării
reversibile rapide actină G ↔ actină F ↔ rețele). ii. Celulară
1. Contracția celulelor musculare – determinată de interacțiunea dintre actină și miozină (proteine contractile); în fibrele musculare striate actina F și miozina se asociază permanent și constituie structuri înalt ordonate – miofibrile (organite
specifice) – responsabile de contracție.
2. Citokineza la celulele animale – actina și alte tipuri de miozină interacționează constituind agregate temporare, mai puțin elaborate față de celula musculară (inel/centură contractil/ă
subplasmalemal circumecuatorial) care determină contracții
localizate (strangulare a celulei) => șanțul de clivaj/citokineză care împarte o celulă animală în două celule-fiice (vezi Reproducerea
celulară).
3. Formare de pseudopode și deplasarea prin mișcări amiboidale – determinate atât de conversia sol-gel a citoplasmei (datorată
transformării reversibile rapide actină G ↔ actină F ↔ rețele), cât
și de contracții localizate rezultate prin interacțiunea actină-miozină.
§Microvilii sunt prelungiri digitiforme („în deget de mănușă”), rigide, egale și echidistante ale suprafeței libere a
plasmalemei. Conțin la interior un fascicul de filamente de actină (are rol mecanic de rigidizare), care constituie regiunea
centrală a microvililor. Cresc foarte mult suprafața plasmalemei. Sunt prezenți la suprafața celulelor absorbtive precum
enterocitele și unele nefrocite (celule ale nefronului) – celulele specializate în transportul transplasmalemal de substanțe.
§Pseudopodele sunt prelungiri temporare ale plasmalemei (care antrenează și citoplasma subiacentă), realizate
prin intervenția microfilamentelor.
Într-o zonă a celulei rețeaua de actină se dezorganizează (inclusiv rețeaua corticală) => plasmalema se deformează
local sub forma unei evaginări în care se scurge și citoplasma subiacentă = pseudopod. Acesta se extinde până când actina
se reasamblează într-o rețea => pseudopodul se stabilizează și se atașează la substrat. Microfilamentele din partea opusă
formării pseudopodului (și deci sensului deplasării) interacționează cu miozina determinând contracția celulei în această
regiune => citoplasma este împinsă („stoarsă”) în pseudopodul fixat de substrat => celula se trage în zona unde s-a atașat
pseudopodul.
Amibele nu sunt singurele celule care se deplasează prin târâre, ci și unele celule din corpul animalelor: unele
leucocite și alte tipuri celulare. Pseudopodele se formează și în procesul de fagocitoză.
B. ORGANITE VEZICULARE
1. Reticulul endoplasm(at)ic (RE) Reticulul endoplasmatic este o rețea intracitoplasmatică extinsă de membrane care formează tubuli și vezicule
aplatizate anastomozate/interconectate și care îndeplinește multiple roluri.
Etimologie: lat. reticulum – rețea mică; gr. endoplasmic – în interiorul citoplasmei.
Există două varietăți distincte, diferite structural și funcțional, dar interconectate, de RE: REN (Reticulul
Endoplasmatic Neted) și RER (Reticulul Endoplasmatic Rugos).
Rolul clasic atribuit RE este acela de sistem de transport al substanțelor prin celulă – un sistem circulator
intracitoplasmatic.
RETICULUL ENDOPLASM(AT)IC NETED (REN) REN este un sistem membranar format dintr-o rețea de tubuli anastomozați, fără ribozomi pe suprafață și
interconectat cu RER; este localizat spre periferia celulei.
ROLURI 1. Implicat în metabolismul lipidelor – sinteza colesterolului și a acizilor biliari (hepatocite); sinteza
hormonilor steroidici de către glanda corticosuprarenală și a hormononilor sexuali de către celulele
endocrine testiculare și ovariene (foarte bogate în REN); absorbția, sinteza și transportul trigliceridelor
(enterocite) etc.
2. Detoxificarea medicamentelor, a anumitor droguri, otrăvuri (pesticide) și agenți carcinogeni (în ficat și
rinichi).
3. Implicat în metabolismul glucidelor – degradarea/depolimerizarea glicogenului depozitat la glucoză liberă,
proces numit glicogenoliză (în special în hepatocite).
4. Depozitarea calciului ionic. Celulele musculare scheletice și cardiace au un REN foarte elaborat (numit
reticul sarcoplasmatic) care are un rol important în depozitarea de ioni de calciu și eliberarea lor în timpul
contracției musculare.
Cu excepția exemplelor de mai sus, cele mai multe celule conțin puțin sau chiar deloc REN adevărat.
RETICULUL ENDOPLASM(AT)IC RUGOS/GRANULAR (RER/REG) RER este un sistem membranar format din vezicule aplatizate (numite și cisterne) interconectate/anastomozate,
cu ribosomi pe suprafața externă/citosolică, care astfel în ME apare rugoasă (aspră, cu granulații). Este localizat
circumnuclear. Membrana RER continuă membrana nucleară externă, iar spațiul perinuclear se continuă cu lumenul RE.

49 / 108
ROLURI
1. Modificări postsinteză ale proteinelor sintetizate la nivelul ribosomilor atașați RER, în special ale
proteinelor destinate exportului din celulă (secreției). Ribosomii atașați RER produc toate proteinele
secretate de celule și de aceea RER este foarte dezvoltat în majoritatea celulelor secretoare (plasmocitele
secretă anticorpi, hepatocitele produc majoritatea proteinelor sângelui, anumite celule pancreatice secretă
în sânge hormonul proteic insulină etc.). Proteinele secretorii pleacă din RE, în principal spre aparatul
Golgi, împachetate în vezicule de transport.
2. Sinteza de fosfolipide și de membrane. RER este „fabrica de membrane” a celulei deoarece la acest nivel
sunt produse fosfolipidele și proteinele integrale care intră în alcătuirea tuturor membranelor celulare. De la
RER membranele ajung în alte părți ale celulei sub forma veziculelor de transport.
3. Proteinele și membranele produse sunt transferate altor compartimente celulare – aparat Golgi,
lizosomi, plasmalemă etc. – prin intermediul unor vezicule de transport care înmuguresc din RER.
2. Aparatul Golgi (dictiosomul) Aparatul Golgi (AG, „centru de recepție-expediere”) este reprezentat de un ansamblu de 4-6 vezicule aplatizate
(numite saculi) stivuite (suprapuse unele peste altele ca monedele în fișic) și cu marginile dilatate, tubuli anastomozați și
micro- și macrovezicule sferice desprinse prin înmugurire din saculi și care orbitează în jurul acestora. AG este situat în
apropierea nucleului, între RE și plasmalemă. Într-o celulă pot exista multe asemenea stive. O stivă se mai numește
dictiosom. Veziculele din vecinătatea AG sunt implicate în transferul de substanțe atât între saculii aceleiași stive Golgi,
cât și între AG și alte structuri.
În secțiune transversală, un sacul are aspect de halteră ușor curbată – o porțiune centrală mai îngustă și două capete
mai dilatate, de la nivelul cărora înmuguresc veziculele.
O stivă Golgi are polaritate – fața cis (față de intrare/regiune de recepție; orientată spre RE; ușor convexă) și fața
trans (față de ieșire/regiune de expediere; orientată spre plasmalemă; aspect concav; prezintă o rețea de tubuli), între care se
află regiunea mediană.
Activitatea AG este strâns corelată cu activitatea RE. Prin AG, produșii elaborați în RE sunt transferați și
distribuiți în alte regiuni celulare.
Vezicule de transport aduc diverse substanțe de la RE la AG. Veziculele de transport care înmuguresc din RER se
deplasează spre și fuzionează cu membrana saculului Golgi de la nivelul feței cis a acestuia. La nivelul feței trans se
formează prin înmugurire noi vezicule care se desprind și se deplasează către alte situsuri.
AG este principalul „controlor de trafic” pentru proteinele celulare. Funcția sa principală este de a modifica,
concentra și împacheta în vezicule proteine (și (fosfo)lipide) produse în RER și a le trimite astfel împachetate către alte
destinații.
Moleculele care tranzitează AG au o triplă destinație: unele sunt exportate ca produși celulari de secreție (sau
excreție), altele vor intra în structura plasmalemei (și a peretelui celular la celulele vegetale), iar altele vor ajunge în
membrana și lumenul lizosomilor, în general ca enzime hidrolitice.
Veziculele golgiene cu produși destinați exportului (ex., proteine) se desprind de la nivelul feței trans drept
vezicule de secreție care participă la exocitoză. Celulele specializate pentru secreție proteică (celulele diverselor tipuri de
glande, exocrine și endocrine) au un AG foarte dezvoltat.
Pe lângă rolul său de modificare a unor tipuri de molecule (mai ales proteine), AG și sintetizează anumite
macromolecule – multe polizaharide (pectine și unele polizaharide necelulozice) produse de celulele plantelor și
încorporate, alături de celuloză, în peretele celular, participând la formare și îngroșarea acestuia.
La plante, vezicule golgiene intervin și în diviziunea celulară, în citokineză, participând la formarea plăcii
celulare și a lamelei mijlocii.
ROLURI 1. Modificarea, împachetarea și distribuirea proteinelor în vederea secreției, includerii în lizosomi și
încorporării în plasmalemă.
2. Sinteza, împachetarea și distribuirea unor polizaharide necelulozice în vederea includerii în perete celular.
3. Participarea, în celulele plantelor, la citokineză prin formarea plăcii celulare și a lamelei mijlocii.

50 / 108
3. Lizosomii Lizosomii („corpi de dezintegrare”) sunt vezicule sferice delimitate de un înveliș unimembranar și care conțin
peste 40 de tipuri de enzime hidrolitice care acționează la pH acid (hidrolaze acide) care realizează digestia intracelulară a
particulelor exogene, dar și a componentelor celulare senescente sau deteriorate structural și funcțional.
Enzimele hidrolitice și membranele lizosomale sunt elaborate de către RER și apoi transferate aparatului Golgi
pentru prelucrări ulterioare. Lizosomii provin prin înmugurire din fața trans a AG.
Lizosomii realizează digestia intracelulară într-o varietate de circumstanțe. Amibele și multe alte protoctiste se
hrănesc prin internalizarea (ingestia prin fagocitoză) organismelor mai mici sau a altor particule alimentare. Vacuola
digestivă (fagosom) formată pe această cale fuzionează cu un lizosom ale cărui enzime digeră particula alimentară. Produșii
de digestie, incluzând glucide simple, aminoacizi și alți monomeri trec prin membrana lizosomală în citosol și devin
nutrienți pentru celulă (vezi Fagocitoza).
Unele celule animale (incl. umane) realizează fagocitoză în scop de apărare a corpului: de ex., neutrofilele și
macrofagele, un tip de celule sangvine albe care ajută la apărarea organismului prin internalizarea și distrugerea bacteriilor
și a altor invadatori/particule patogene. Aceste tipuri celulare se numesc fagocite și conțin numeroși lizosomi. Osteoclastele
sunt celule care degradează osul în scopul remodelării și ajustării structurii la funcția pe care trebuie să o îndeplinească și
conțin, de asemenea, foarte mulți lizosomi.
Lizosomii intervin și în reciclarea materialului organic celular – proces numit autofagie. Cu ajutorul lizosomilor
celula se reînnoiește continuu.
Ruperea lizosomilor are drept consecință autodigestia celulei – proces numit autoliză. Autoliza este baza
distrugerii controlate a celulelor (apoptoză).
ROLURI 1. Digestia particulelor preluate prin endocitoză (particule exogene), în special bacterii, virusuri și toxine -
heterofagie.
2. Degradarea organitelor senescente sau compromise structural și funcțional – autofagie.
3. Glicogenoliză – depolimerizarea glicogenului => glucoză.
4. Degradarea unor țesuturi inutile, precum membranele interdigitale de la fetus, mucoasa uterină în timpul fazei
hemoragice a ciclului endometrial/uterin.
5. Degradarea osului – osteoliză – pentru a elibera ioni de calciu în sânge și în scop de modelare.
6. Lizosomii sunt implicați în apoptoză.
4. Peroxisomii Peroxisomii (microcorpii) sunt vezicule sferice, puțin mai mici decât lizosomii și delimitate de un înveliș unimembranar, care conțin enzime
oxidoreducătoare puternice – oxidazele și catalazele. Enzimele peroxisomale au rol în detoxificarea unor substanțe chimice toxice de origine exogenă sau endogenă, dar și în degradarea unor molecule
precum aminoacizii și acizii grași. Cea mai importantă funcție a peroxisomilor este de a neutraliza radicalii liberi – compuși chimici periculoși, înalt reactivi, care pot altera
structura și implicit funcțiile moleculelor biologice.
Oxidazele utilizează O2 pentru a detoxifica substanțe nocive – convertesc radicalii liberi la peroxid de hidrogen, care este de asemenea reactiv
și periculos, dar este rapid descompus în apă și oxigen de către catalază: 2H2O2 → 2H2O + O2 Radicalii liberi și peroxidul de hidrogen sunt produși secundari normali ai metabolismului celular, dar au efect devastator asupra celulei dacă li se permite acumularea.
Peroxisomii nu se formează prin înmugurire din aparatul Golgi precum lizosomii, ci sunt organite autoreplicante formate prin simpla fisiune binară a peroxisomilor preexistenți.
Peroxisomii sunt numeroși în special în hepatocite și nefrocite, care sunt foarte active în detoxificarea organismului.
5. Mitocondria Mitocondriile sunt organite veziculare cu înveliș dublu-membranar, prezente în toate celulele eucariote, cu
excepția hematiilor adulte (lipsite de organite și de nucleu) și a unor protoctiste care au mitocondrii modificate structural și funcțional,
având chiar denumiri particulare – diplomonadele (Giardia intestinalis, parazit intestinal la mamifere) au mitosomi, iar parabasalidele (Trichomonas
vaginalis, prezent în mod obișnuit în vaginul femeilor) au hidrogenosmi. Totalitatea mitocondriilor dintr-o celulă constituie condriomul.
Localizare. Mitocondriile sunt situate, în general, în vecinătatea structurilor intracelulare care necesită aport
crescut de energie metabolică (ATP), adică în zonele cu activitate metabolică intensă (în jurul miofibrilelor în celula
musculară, în jurul axonemei cililor și flagelilor, în proximitatea RER și a AG, în jurul nucleului) sau în vecinătatea
surselor de combustibil celular (glicosomi = granule de glicogen – sursă de glucoză; picături lipidice – sursă de acizi
grași; amiloplaste – depozit de amidon, care este sursă de glucoză etc.).
Formă și dimensiune. Este destul de variată: sferică la drojdii, elipsoidală în hepatocite, filamentoasă în
fibroblaste, cilindrică, de bastonaș etc. În general, lungimea mitocondriilor este cuprinsă între 0,5 și 1 μm.
Număr. Variază între unu și 1.000.000, în funcție de tipul celular, de mărimea, de starea metabolică, de necesarul
energetic al celulei, de rolul energetic al acesteia în organism etc. Este relativ constant la un anumit tip celular și este
cuprins, în general, între 500 și 2000 per celulă. În celulele de drojdie de bere (ciuperci ascomicete unicelulare) se găsesc 2-
50 mitocondrii, spermatozoidul conține în jur de 25 de mitocondrii, amoebele (organisme unicelulare – protozoare) din
genul Chaos ating dimensiuni de 1-3 mm și pot conține până la 500.000 de mitocondrii. În hepatocit mitocondriile
constituie 22% din masa citoplasmei (1000-2000 de mitocondrii), iar în celula miocardică procentul ajunge la 50.
Structură. Mitocondria prezintă un înveliș dublu-membranar care delimitează două compartimente. Învelișul este
constituit din două membrane:
membrană mitocondrială externă netedă: are grosime de aproximativ 7,5 μ și compoziție chimică și
arhitectură similare plasmalemei; separă mitocondria de citosol; este foarte permeabilă pentru o gamă largă
de substanțe; prezintă numeroase canale ionice – porine mitocondriale

51 / 108
membrană mitocondrială internă cutată/pliată spre interiorul mitocondriei, fie sub formă de criste/creste
(cresc suprafața membranei interne de cca. 10x), fie de tubuli; conține un procent ridicat de proteine –
peste 70% – implicate în reacțiile redox din CTEM (catena transportoare de electroni mitocondrială), în
sinteza de ATP, în transportul înalt selectiv al substanțelor prin membrană; CTEM este formată din mai
mulți transportori de electroni, dintre care unii translocă H+ prin membrana internă; este impermeabilă
pentru majoritatea substanțelor (în special pentru ioni, inclusiv protoni); la nivelul feței dinspre matrice se
găsesc structuri particulate proeminente, numite clasic oxisomi; acestea reprezintă complexe enzimatice la
nivelul cărora se realizează fosforilarea ADP la ATP pe baza energiei eliberate în cursul reacțiilor redox
ale CTEM și se numesc ATP-sintaze (F0F1-ATP-aze)
Aceste două membrane delimitează două compartimente:
spațiul intermembranar este delimitat de cele două membrane și are compoziție similară cu a citosolului, cu
excepția proteinelor care sunt prea mari pentru a difuza liber prin membrana externă.
lumenul (cavitatea) mitocondriei este delimitat de membrana internă și conține un material vâscos, similar
unui gel (are concentrație foarte mare de proteine), numit matrix/matrice mitocondrial/ă, mitoplasmă
sau substanță fundamentală. În această matrice se găsesc: apă, ADNmt (ADN mitocondrial –
macromoleculă dublu catenară circulară, similară nucleoidului bacterian), ribosomi de 70S, ioni minerali
(Ca2+
, Mg2+
, K+, Na
+ etc.), ATP, ADP, Pi, enzime implicate în multe procese metabolice: ciclul Krebs
(ciclul acidului citric sau al acizilor tricarboxilici), biosinteza acizilor grași, β-oxidarea (degradarea)
acizilor grași, oxidarea aminoacizilor, biosinteza proteinelor mitocondriale etc.
ROLURI Mitocondria este sediul principal al respirației celulare, numită de aceea și respirație mitocondrială.
Respirația celulară este procesul metabolic (catabolic; biochimic și fiziologic) prin care substratul chimic cu rol
energetic (glucoză, acizi grași, mai rar aminoacizi sau alți compuși organici sau chiar anorganici) este
descompus/degradat oxidativ și este astfel eliberată energia legăturilor chimice ale acestuia și stocată concomitent în
molecule macroergice (ATP). Desfășurarea proceselor caracteristice sistemelor biologice (sinteze, transport transmembranar, intracelular și intercelular, locomoție, creștere,
dezvoltare, reproducere etc.) necesită un permanent aport de energie. Principalii compuși organici cu rol energetic sunt glucoza și acizii grași (au rol de
substrat energetic/respirator). Aceștia conțin, în legăturile lor covalente, cantități mari de energie (energie chimică) pe care celula trebuie să o extragă în
vederea susținerii proceselor metabolice consumatoare de energie (procese anabolice/de sinteză și transporturi). Eliberarea acestei energii presupune ruperea legăturilor covalente, fapt care are drept consecință degradarea respectivilor compuși organici. Procesele metabolice de degradare a compușilor
organici, (eventual) însoțită de eliberare de energie, sunt procese catabolice. Cea mai eficientă modalitate de rupere a legăturile covalente, cu eliberarea
treptată/etapizată de cantități mari de energie, este prin oxidare – degradare oxidativă/aerobă (catabolism oxidativ sau aerob). Prin oxidarea substratului respirator la nivel celular energia legăturilor covalente ale acestuia este eliberată treptat, „în porții mici”, și în paralel stocată în compuși organici
macroergici, precum ATP-ul, care o transportă și o transferă proceselor metabolice consumatoare de energie. Scopul (și rolul) mitocondriei, prin ciclul Krebs și fosforilarea
oxidativă, este de a sintetiza ATP – ribonucleotid care depozitează,
transportă în cadrul celulei și al organismului și transferă proceselor
endergonice (consumatoare de energie) energie chimică. Ca urmare a
implicării în fluxul energetic din cadrul metabolismului, ATP-ul se mai
numește și „energie metabolică” sau „monedă energetică”, iar mitocondria
care îl produce – „uzină/centrală energetică” a celulei.

52 / 108
Studiu de caz. Respirația celulară a glucozei – substratul respirator preferat și utilizat de majoritatea sistemelor
energetice – se desfășoară în patru etape:
1. Glicoliza (în citosol, anaerob)
2. Activarea piruvatului (în mitocondrie)
3. Ciclul Krebs (în mitocondrie – în matricea mitocondrială; aerob)
4. Fosforilarea oxidativă (în mitocondrie – în membrana mitocondrială internă; aerob).
Ecuația degradării totale a glucozei este: C6H12O6 + 6O2 + 38ADP + 38Pi → 6CO2 + nH2O + 38ATP sau,
simplificată: C6H12O6 + 6O2 → 6CO2 + 6H2O + energie (ATP + căldură).
În urma acestei reacții rezultă 686 kcal/mol (în condiții standard – pH 7 și 25ºC), din care numai aproximativ 40%
(~300 kcal) va fi stocată în legăturile fosfat-macroergice ale celor 38 de molecule de ATP, restul pierzându-se sub formă de
căldură. O legătură fosfat-macroergică a ATP depozitează 7,3 kcal/mol.
Glucoza este oxidată de către O2 la CO2 și H2O, cu eliberare de energie; o parte din această energie este folosită
în fosforilare – adăugarea unei grupări fosfat la un substrat (în acest caz – ADP) => ATP. Astfel, fosforilarea ADP este
corelată cu oxidarea glucozei => fosforilare oxidativă.
Definiția respirației celulare poate fi particularizată: procesul de degradare, în prezența oxigenului, a
monozaharidelor până la CO2, apă și energie.
*Teoria chemiosmotică a lui Peter Dennis Mitchell (Premiul
Nobel în 1978)
O parte din energia eliberată prin mecanism oxidativ din
legăturile covalente ale substratului respirator (glucoza), este folosită (și
implicit depozitată) pentru transportul activ de H+ prin membrana
mitocondrială internă din matrice în spațiul intermembranar => este
generat și menținut un gradient electrochimic între cele două
compartimente. Acest gradient conține energie. Prin descărcarea
controlată a acestui gradient (printr-o enzimă particulară – ATP-
sintaza/F0F1-ATP-aza), energia cinetică a H+ care se deplasează pasiv (în gradient
electrochimic) prin membrana mitocondrială internă, din spațiul intermembranar în
matrice, este folosită pentru fosforilarea ADP la ATP de către ATP-sintază =>
energia cinetică a gradientului este depozitată în legătura fosfat-macroergică a ATP-
ului nou-format. Astfel, prin intermediul gradientului electrochimic, energia
legăturilor covalente ale substratului respirator, extrasă prin oxidare, este depozitată
în legătura macroergică a ATP-ului rezultat prin fosforilare => fosforilare
oxidativă.
§Alte roluri: biosinteza unor proteine mitocondriale, sinteza acizilor grași,
degradarea oxidativă (β-oxidarea) a acizilor grași, depozitare de calciu ionic și
implicare în semnalizarea (intra)celulară, reglarea morții celulare programate
(apoptoză – engl. apoptosis), diferențiere celulară, controlul ciclului celular și al creșterii celulare, implicare în procesul de
îmbătrânire/senescență etc.
§Biogeneză. Mitocondriile sunt organite semiautonome și autodivizibile, având genom și aparat de biosinteză
proteică proprii și capacitate de diviziune prin fisiune, modalitate prin care se și multiplică. Mitocondriile se transmit de la
o generație de indivizi la următoarea numai pe cale maternă, prin gametul feminin, ovulul.
TEMĂ. Stabiliți legătura dintre mitocondrii și celulele procariote. Cine a elaborat teoria endosimbiozei și ce
presupune aceasta? Ce este apoptoza?

53 / 108

54 / 108
II. ORGANITE SPECIFICE
1. Miofibrilele Miofibrilele constituie elementele contractile ale celulei musculare striate (scheletice și cardiace). Fiecare fibră
musculară conține numeroase miofibrile cilindrice, cu diametrul de 1-2 µm, împachetate dens și dispuse paralel cu axul
lung al fibrei.
În lungul fiecărei miofibrile sunt evidente striații – reprezentate de succesiunea repetată de benzi transversale
(discuri) întunecate A și clare I. Într-o fibră musculară intactă, benzile A și I din toate miofibrilele sunt aproape perfect
aliniate unele cu altele, dând aspectul striat celulei ca întreg.
Fiecare bandă A are la jumătate o zonă mai deschisă la culoare – zona/banda H (gr. helle – luminos), bisectată de o
membrană – membrana M. Banda I prezintă de asemenea la jumătate o membrană zigzagată – membrana Z (stria Amici),
care solidarizează miofibrilele între ele și se atașează și la sarcolemă, menținându-se aliniate.
Un sarcomer este regiunea dintr-o miofibrilă
cuprinsă între două membrane Z succesive și conține la
fiecare capăt câte o ½ de bandă I și în centru o bandă A.
Având în medie 2 µm lungime, sarcomerul este cea mai
mică unitate contractilă a fibrei musculare =>
sarcomerele sunt unitățile funcționale ale mușchiului
scheletic; acestea sunt aliniate coadă-la-cap (ca
vagoanele) într-o miofibrilă.
Examinarea la nivel molecular a modelului de
bandare a miofibrilelor arată că acesta provine din
aranjarea ordonată în cadrul sarcomerului a două tipuri
de structuri filamentoase numite miofilamente:
miofilamente groase și miofilamente subțiri.
Miofilamentele groase se întind pe întreaga lungime a benzii A. Filamentele subțiri se întind în banda I și pătrund
parțial și în banda A, printre cele groase.
Filamentele groase sunt alcătuite în principal din proteina contractilă miozină. Filamentele subțiri sunt alcătuite în
principal din proteina contractilă actină, alături de care sunt prezente și câteva proteine reglatorii: tropomiozina și
troponina (ajută la controlul interacției actinei cu miozina din timpul contracției).
În ultimii zece ani s-au decoperit și alte tipuri de filamente musculare. Unul dintre aceste tipuri de filamente nou
descoperite – miofilamentul elastic – este alcătuit dintr-o proteină gigantică numită titină.
2. Neurofibrilele Neurofibrilele sunt organite specifice neuronilor. Sunt elemente de citoschelet care se extind în corpul celular și în
prelungirile neuronului (dendrite și axon). Au rol mecanic de susținere și rol în traficul de vezicule prin celulă – prin axon,
de la corpul celular la butonii terminali și invers.
3. Corpii Nissl Corpii Nissl (substanța bazofilă, corpii tigroizi), de asemenea organite specifice neuronilor, sunt mase dense de
RER, având rol în sintezele proteice specifice neuronului. Sunt prezente la nivelul corpului celular al neuronului.
4. §Pseudopodele – vezi Citoscheletul
5. Cilii și flagelii – vezi Citoscheletul
6. §Microvilii – vezi Citoscheletul
3. INCLUZIUNILE CITOPLASMATICE sunt componente nevii ale celulei, reprezentate de diverse
substanțe chimice sau particule, delimitate (în celulele vegetale) sau nedelimitate (în celulele animale) de membrane. Pot fi
prezente sau pot lipsi din celulă, în funcție de tipul celular și de starea de activitate metabolică a acesteia. Sunt lipsite de
activitate metabolică proprie, dar pot fi mobilizate de către sistemele enzimatice celulare. Au origine endogenă (rezultate
din metabolismul celular – unele sunt reintroduse în metabolism, altele sunt scoase din circuitul metabolic) sau exogenă
(preluate din exterior; de obicei sunt inerte).
Exemple 1. Depozite de nutrienți
a) Glicosomi – granule de glicogen, localizate, de obicei, lângă REN; sunt abundenți în hepatocite și
miocitele striate, dar și în celulele epiteliului vaginal și uterin; reprezintă o importantă sursă de energie
prin glucoza din care este format glicogenul și care este eliberată enzimatic prin glicogenoliză.
b) Granule de amidon – formă și dimensiuni variate; sintetizat și depozitat în leucoplaste (temporar și în
cloroplaste).
c) Inulină – polizaharid specific anumitor grupe de plante; depozitat în vacuolă.
d) Aleuronă – incluziune proteică prezentă în celulele unor semințe și fructe; depozitată în vacuole sau în
proteoplaste.
e) Picături lipidice – trigliceride depozitate în special în adipocite și unele celule vegetale (din semințele
oleaginoase), dar și în alte tipuri de celule (hepatocite); sunt fluide la temperatura corpului și au aspect
de picături sferice refractile în celulele vii; reprezintă rezerve locale de energie și de material plastic
utilizat în sinteza de membrane și a altor componente structurale lipidice sau a unor produși de secreție
Filament subţire
(de actină)
Filament gros
(de miozină)
Membrana M
Membrana Z
Banda luminoasă (H)
Disc întunecat A Disc clar I
Sarcomer

55 / 108
2. Diverse tipuri de cristale
a) Cristale organice
(i) Proteice – normale pentru aumite tipuri de celule (Sertoli, Leydig și, ocazional, macrofage);
prezente în oricare regiune a celulei – nucleu, mitocondrie, RE, AG și în matricea citoplasmatică.
(ii) Oxalat de Ca – caracteristic plantelor; acidul oxalic este un produs toxic al metabolismului celular;
se depozitează în vacuole unde este detoxificat prin combinarea cu ioni de calciu și prin cristalizarea
consecutivă => cristale insolubile și netoxice, solitare sau agregate.
b) Cristale anorganice de CaCO3, SiO2, CaSO4, CuSO4, NaCl, ioduri; rare și întâlnite mai ales în celulele
vegetale, formate și depozitate în vacuole.
3. Pigmenți a) Granule de melanină (melanosomi) – produsă de melanocitele din piele și păr, de celulele pigmentare din
retină și de neuroni din substanța neagră; are rol protector în piele (ecranarea radiațiilor UV) și participă
la fotorecepție; rolul în neuroni nu este complet înțeles; sunt delimitate de membrană.
b) Lipofuscină – pigment galben până la brun, prezent în miocitele cardiace și în neuronii din SNC; se
consideră că ar avea activitate lizosomală.
c) Antocianine – specifice plantelor; depozitate în vacuole (vezi Celula vegetală).
d) Hemoglobina – heteroproteină prezentă în hematii; rol de transportor de gaze respiratorii (O2 și CO2).
4. Apă – depozitată în vacuole la plante.
5. Reziduuri metabolice – scoase din circuitul metabolic prin depozitare, de obicei în formă precipitată sau
cristalizată, în vacuole, vezicule sau direct în citosol.
6. Produși de secreție (mucină, hormoni, sebum etc.) – depozitați temporar în vezicule sau direct în citosol, până
la eliminarea din celulă
7. Virusuri etc.
TEST I
Nucleul
Nucleul este cel mai mare și mai evident organit din celula eucariotă. A fost descoperit în anul 1831 de Robert
Brown11
în celulele orhideelor, ulterior generalizându-se prezența sa în celulele vegetale și animale.
Localizare. Nucleul este situat în general central, dar, în funcție de tipul, forma și de vârsta celulei, poate fi
situat și excentric/periferic; de exemplu, la celulele vegetale adulte și la adipocite este împins la periferia celulei de către
vacuola centrală, respectiv de picătura de lipide; la celula musculară striată este tot periferic, centrul celulei fiind ocupat de
miofibrile.
Formă și dimensiune. Frecvent are formă sferică sau eliptică, dar poate fi
și fusiform, lenticular, semilunar, filamentos, stelat, lobat, amorf etc.; de multe ori
forma nucleului este corelată cu forma celulei (de exemplu, în celulele cubice
nucleul este sferic, în cele cilindrice este ușor alungit). Mărimea nucleului este relativă și se apreciază prin raportul nucleo-(cito)plasmatic (RNP,
RNC). RNP este supraunitar la celulele de tip blast, nediferențiate sau slab diferențiate și foarte active metabolic, și subunitar la celulele de tip cit, mature, diferențiate, a căror activitate metabolică este la un
nivel scăzut, de întreținere.
În general, la plantele superioare nucleul are o mărime cuprinsă între 5 și 15 μm; aceasta crește cu gradul de ploidie, dar și în cursul ciclului celular.
Număr. În general, în fiecare celulă există un singur nucleu. Însă, în
funcție de particularitățile structurale și funcționale ale celulei, există și abateri de la
această regulă:
Celule anulcleate/enucleate: hematiile adulte și
articulele/elementele mature de tub ciuruit (vas liberian prin care
circulă seva elaborată la plante)
Celule binucleate: unele hepatocite, unele celule miocardice, unele protozoare (ciliatele precum
parameciul), celulele miceliului secundar al ciupercilor asco- și bazidiomicete;
Celule multinucleate: celulele/fibrele musculare striate scheletice, protozoarele opaline, algele verzi din
ordinele Siphonales și Siphonocladales, osteoclastele.
Numărul de nuclei este corelat:
cu intensitatea proceselor metabolice din celulă (celulele foarte active metabolic au mai mulți nuclei care
să asigure coordonarea)
cu dimensiunea celulei (celulele mari au nevoie de mai mulți nuclei, fiecare responsabil de coordonarea
activității unei părți a respectivei celule)
cu originea celulei (celulele musculare striate scheletice au dimensiuni mari și s-au format prin fuziunea
mai multor celule mici uninucleate).
ROLURI. Nucleul are trei roluri principale:
1. Stocarea și protejarea materialului genetic.
2. Transmiterea materialului genetic în cursul diviziunii celulare.
3. Controlul și coordonarea activității celulare.
11 Botanist şi paleobotanist scoţian, 1773-1858; a realizat prima descriere detaliată a nucleului celular şi a fluxului citoplasmatic; a introdus conceptul de
mişcare browniană pe baza studiilor asupra polenizării şi fecundaţiei.

56 / 108
Structură – nucleul este constituit din trei componente:
1. Învelișul nuclear dublu membranar, numit și anvelopă nucleară, este format din două membrane elementare
– membrană nucleară internă și membrană nucleară externă – fiecare cu grosime de 7,5-10 nm, dar cu compoziție
chimică diferită, între care se află spațiul perinuclear, larg de 25-100 nm. Membrana externă prezintă ribosomi pe
suprafață și se continuă cu membrana RER, iar spațiul perinuclear se continuă cu lumenul RER.
Imediat sub învelișul nuclear se găsește lamina nucleară – structură proteică formată din filamente intermediare
(alcătuite din proteine fibrilare numite lamine), cu rol de ranforsare a învelișului nuclear; lamina nucleară se
dezorganizează la începutul diviziunii celulare și se reorganizează la finalul acesteia.
Învelișul nuclear prezintă din loc în loc întreruperi, numite pori nucleari, la nivelul cărora cele două membrane
nucleare, internă și externă, fuzionează, continuându-se una cu cealaltă. Numărul porilor variază în funcție de tipul celular,
de starea de activitate metabolică, de etapa din interfază în care se află celula; este cuprins între
câteva zeci și câteva mii per nucleu (în medie, 2000-4000). Porii asigură tot schimbul selectiv
de substanțe și particule cu citoplasma. Porii nucleari au compoziție și arhitectură moleculare foarte complexe, care nu au fost stabilite cu
certitudine12, dar au fost propuse mai multe modele posibile pe baza imaginilor obținute prin diferite tehnici de
microscopie electronică. Sunt formați din mai multe tipuri de proteine, numite nucleoporine, fiecare por conținând sute de asemenea molecule care constituie o structură cu simetrie octogonală (cu aspect de butoi, după un model) –
complexul poral nuclear sau complexul nucleoporal, cu diametrul extern de cca 120 nm, iar cel intern (al porului
propriu-zis) este de cca. 40 nm.
Învelișul nuclear separă nucleoplasma de citoplasmă și reglează traficul de substanțe
și particule între cele două compartimente celulare.
2. Nucleoplasma (carioplasma) – materialul pe care îl delimitează învelișul nuclear. Este format din două
componente: cariolimfa și cromatina.
a) Cariolimfa este o soluție apoasă care conține săruri minerale sub formă de ioni, nucleotide libere, enzime,
proteine reglatoare și structurale (matricea nucleară de care se atașează cromosomii, echivalentă citoscheletului prezent în
citosol); este sediul cromatinei și al nucleolului.
b) Cromatina este componenta nucleoplasmică principală, reprezentată de filamente nucleo-proteice, cu aspect de
șirag de mărgele. Un asemenea filament cu granule pe lungime este format dintr-o macromoleculă liniară de ADN (acid
nucleic, „nucleo-”) asociat cu proteine structurale numite histone (componenta proteică a cromatinei), formând un complex
supramolecular nucleo-proteic sau nucleo-histonic. Granulele din alcătuirea filamentului de cromatină se numesc
nucleosomi. ADN-ul este compusul biochimic care deține, în formă codificată, informația genetică/ereditară sub forma
genelor. Totalitatea materialului genetic din celulă constituie genomul. Genomul celular este format din genomul nuclear,
genomul mitocondrial și genomul cloroplastic.
Un asemenea filament de cromatină este un cromosom interfazic, pliat mai dens sau mai lax în nucleoplasmă.
Regiunile în care filamentele de cromatină sunt împachetate mai dens formează heterocromatina nucleară, iar regiunile în
care filamentele sunt împachetate mai lax formează eucromatina nucleară.
La începutul diviziunii celulare, filamentele de cromatină se supraspiralizează și se condensează, scurtându-se și
îngroșându-se; devin astfel cromosomi de diviziune sau mitotici, cu aspectul tipic de cromosom.
Un organism are în nucleul celulelor din care este format corpul un număr caracteristic și constant de
cromosomi. Dacă este vorba de celulele sexuale – gameții – ale organismului, numărul de cromosomi este notat cu „n”, iar
celulele sunt haploide („n” este simbolul numărului haploid de cromosomi); celulele corpului, numite celule somatice, au
un număr dublu de cromosomi față de gameți, număr notat cu 2n și numit număr diploid de cromosomi.
Deci gameții sunt haploizi (n), iar celulele somatice sunt diploide (2n). Exemplu: la om, spermatozoidul și ovulul
au fiecare câte n = 23 de cromosomi, iar celulele somatice au câte 2n = 46 de cromosomi.
Toți indivizii unei specii au același număr de cromosomi în celulele corpului lor, numărul de cromosomi fiind un
important criteriu de delimitare și diferențiere a speciilor. Exemplu: cimpanzeii au 2n = 48, iar oamenii au 2n = 46,
musculița de oțet are 2n = 8, ceapa are 2n = 16 etc.
Cromosomii de diviziune pot fi bicromatidici (formați din două cromatide-surori identice, paralele și atașate între
ele pe întreaga lungime, mai strâns la nivelul centromerului), monocromatidici (o singură cromatidă) sau bivalenți
(tetrade cromatidiale, doi cromosomi bicromatidici omologi atașați lateral unul de altul prin intermediul a câte o cromatidă)
– vezi Acizii nucleici și Reproducerea celulară.
O cromatidă este un filament de cromatină puternic condensat, conținând deci o singură moleculă de ADN. Din
punct de vedere morfo-structural, cromatida prezintă o constricție primară – centromer, două brațe egale sau inegale ca
lungime separate prin centromer (notate cu p – brațul scurt și q – brațul lung) și două extremități numite telomere. Unii
cromosomi prezintă și o constricție secundară la nivelul brațului scurt, care delimitează un segment de cromatidă numit
satelit (asemenea cromosomi se numesc cromosomi sateliferi), la nivelul căruia se găsește RON (vezi mai jos).
Fiecare filament de cromatină este ancorat de membrana nucleară internă și ocupă un anumit teritoriu din
nucleoplasmă numit domeniu cromosomal, între care se găsesc domenii intercromosomale.
3. Nucleolul este o regiune de obicei sferică/sferoidală, mai densă/refringentă a nucleoplasmei, organizată în
asociere cu anumite zone ale anumitor cromosomi – zonele care conțin genele ADNr, adică genele pe baza cărora se va
sintetiza ARNr. Aceste zone ale cromosomilor se numesc RON – regiuni organizator nucleolare. Nucleolul nu este
delimitat de membrană.
Structură. Nucleolul prezintă trei regiuni decelabile la ME: centri fibrilari, componentele fibrilare dense (pars
fibrosa) și componentele granulare (pars granulosa).
12 Complexul nucleoporal de la plante este foarte puţin cunoscut, atât din puncte de vedere compoziţional şi arhitectural, cât şi funcţional. Cele mai multe
studii au fost făcute la animale şi la drojdii.

57 / 108
Centrul fibrilar conține buclele ADN care conțin genele ADNr. Acestea constituie organizatorul nucleolar și codifică pentru diferitele tipuri de ARNr. Componenta fibrilară conține în principal transcripte
primare de ARNr (pre-ARNr), care încep să se complexeze cu proteine ribosomale. Componenta granulară
conține subunități ribosomale în diferite stadii de asamblare, formate prin complexarea ARNr-urilor mature cu
proteine ribosomale. Organizarea tripartită este valabilă numai la vertebratele superioare (amniote – reptile,
păsări, mamifere); la alte grupe de organisme (anamniote – pești, amfibieni) s-a observat o organizare bipartită
(componentă fibrilară și componentă granulară). La alte grupe de oganisme, în special la plante, nucleolul prezintă central o zonă clară, numită vacuolă nucleolară.
Rolul nucleolului. La nivelul nucleolului se sintetizează ARNr (ARN ribosomal) și tot aici ARNr este asamblat cu
proteine ribosomale importate din citoplasmă pentru a forma subunitățile ribosomale. Aceste subunități vor ajunge, prin
porii nucleari, în citoplasmă și vor forma ribosomii funcționali. Numărul de nucleoli dintr-o celulă este corelat cu
activitatea metabolică a celulei – celulele foarte active metabolic necesită un număr mare de proteine, adică este necesar un
număr mare de ribosomi funcționali și implicit de subunități ribosomale. Ca urmare, în nucleul unor asemenea celule se
găsesc mai mulți nucleoli – doi, trei, patru etc.

58 / 108
Celula vegetală
Celula vegetală este constituită din aceleași componente ca și celula animală plus trei structuri caracteristice și
definitorii:
1. Vacuola centrală mare permanentă
2. Peretele celular celulozic
3. Plastidele
1. Vacuola Împreună cu plastidele și cu peretele celular, vacuola este una din cele trei structuri caracteristice care diferențiază
celulele vegetale de cele animale. Vacuola este un constituent neprotoplasmatic (neviu) al celulei vegetale.
Celulele tinere conțin de obicei numeroase vacuole mici care cresc în dimensiuni și confluează într-o singură
vacuolă pe măsură ce celula se mărește și maturează. Totalitatea vacuolelor dintr-o celulă constituie vacuomul.
Vacuolele sunt organite veziculare mărginite de un înveliș unimembranar numit tonoplast sau membrană
vacuolară.
Vacuola derivă de obicei direct din aparatul Golgi, dar se poate forma și din RE.
Vacuolele conțin un lichid numit suc celular/vacuolar. Principalul component al sucului celular este apa, care
conține și alte componente care variază în funcție de specie, de tipul plantei, al organului și tipul celular și de starea lor
fiziologică și gradul de dezvoltare, de condițiile ecologice în care trăiește planta etc.
Sucul vacuolar conține săruri minerale dizolvate (ionii anorganici precum Ca2+
, K+, Cl
–, Na
+ și HPO4
2–, CO3
2–,
NO3–
etc.) și compuși organici precum glucide (glucoză, zaharoză, substanțe pectice care prin hidratare formează
mucilagii etc.), acizi organici și aminoacizi, taninuri, alcaloizi, uneori și pigmenți antocianici hidrosolubili. Uneori, o
substanță particulară este prezentă într-o concentrație atât de mare încât formează cristale. Cristalele de oxalat de calciu,
care au forme variate, sunt cele mai comune.

59 / 108
În majoritatea cazurilor, vacuolele nu sintetizează moleculele pe care le conțin, ci le acumulează din alte părți ale
citoplasmei.
Diferite tipuri de vacuole cu funcții distincte pot fi găsite într-o singură celulă matură.
ROLURI
1. Rol structural. În celula matură, aproximativ 90% din volumul celulei poate fi ocupat de vacuolă, iar restul
citoplasmei este reprezentat de un strat fin periferic, lângă plasmalemă. Cea mai mare parte a creșterii în
dimensiuni a celulei rezultă din mărirea vacuolei care determină, prin acumulare de lichid (apă), apariția unei
presiuni hidrostatice interne (stare de turgescență – stare umflată și rigidă) și implicit menținerea rigidității
țesutului format din celulele vegetale – unul din principalele roluri ale vacuolei.
2. Depozitare de metaboliți primari (cu rol nutritiv) precum glucide (zaharuri), acizi organici și proteine de
rezervă.
3. Îndepărtează (prin sechestrare permanentă) metaboliții secundari toxici, precum nicotina și taninul, din restul
citoplasmei. Metaboliții secundari conținuți în vacuole sunt toxici nu doar pentru planta însăși, dar și pentru
patogeni, paraziți și/sau erbivori și de aceea joacă un rol important în apărarea plantei.
4. Degradarea/digestia macromoleculelor și a organitelor celulare îmbătrânite sau deteriorate. Datorită acestor
activități digestive, vacuolele sunt comparabile ca funcție cu lizosomii din celulele animale.
5. Depozitare de pigmenți – culorile albastru, violet, purpuriu, cărămiziu și roșu ale celulelor și implicit ale
anumitor organe ale plantelor sunt determinate de un grup de pigmenți numiți antocianine (solubile în apă și
dizolvate în sucul celular).
§Antocianinele sunt responsabile de culorile roșu și albastru ale unor legume (ridichi, varză), fructe (struguri,
prune, cireșe) și o multitudine de flori (albăstriță, ciocul-berzei, nemțișor, trandafir, bujor). Uneori pigmenții sunt atât de
intenși încât maschează clorofila (verde) în frunze – ca la arțarul roșu ornamental. Antocianinele sunt responsabile și de
culoarea roșu aprins a unor frunze toamna (vița canadiană). Acești pigmenți se formează în zilele reci, dar însorite, când
frunzele încetează să mai producă clorofilă. Pe măsură ce clorofila se degradează, antocianinele formate în vacuole nu mai
sunt mascate. În frunzele care nu formează pigmenți antocianici, degradarea clorofilei în toamnă demaschează pigmenții
carotenoizi, galbeni până la portocalii, mult mai stabili, deja prezenți în cloroplaste.
2. Peretele celular (PC) Peretele celular este o structură neprotoplasmatică (nevie) rigidă, specifică celulei vegetale, celulei fungale și
majorității celulelor algale, situată deasupra plasmalemei și mult mai groasă decât aceasta – 0,1-câțiva μ.
În continuare vom discuta despre peretele celular (PC) al celulelor plantelor.
ROLURI 1. Protejează celula și îi menține forma.
2. Fiind rezistent și rigid previne spargerea celulei prin absorbție excesivă de apă.
3. Conferă rigiditate plantei ca întreg, asigurându-i susținere și conferindu-i sedentaritate.
4. Reprezintă o barieră poroasă pentru circulația și distribuția apei, mineralelor și a altor nutrienți.
5. Adăpostește molecule specializate care reglează creșterea și protejează planta de boli și patogeni
bacterieni și fungali. Planta se protejează devenind rezistentă la atac fie prin producerea de fitoalexine (substanțe toxice pentru
patogeni), fie prin sinteza și depozitarea de lignină în PC (formând o barieră mecanică în calea patogenilor).
Tipuri de PC
La celulele plantelor există trei tipuri de pereți celulari: perete celular primordial, perete celular primar și perete
celular secundar.
PC primordial (lamelă mijlocie, substanță intercelulară) – comun tuturor celulelor dintr-un țesut, fiind
situat între celule. Dpdv biochimic este format din substanțe pectice și apă – se formează un mucilagiu adeziv care
menține celulele lipite în țesut – asigură astfel solidarizarea structurală a celulelor în cadrul țesutului. Este primul perete
care se formează în timpul citokinezei, din conținutul veziculelor golgiene care fuzionează pentru a forma placa celulară.
PC primar – propriu fiecărei celule, situat deasupra plasmalemei. Dpdv biochimic, este format din apă (~65%),
celuloză, hemiceluloză, substanțe pectice, glicoproteine (prezente obligatoriu în toți pereții primari) și substanțe
incrustante care împregnează peretele determinând creștere în grosime și conferind rigiditate și rezistență suplimentare
pereților anumitor celule – suberină, cutină, ceruri, lignină, substanțe minerale (CaCO3, SiO2). În PC primar se găsesc
și enzime (peroxidaze, fosfataze, celulaze, pectinaze), unele implicate în reacțiile prin care se formează, remodelează sau
lizează rețeaua structurală a peretelui. Asemenea schimbări sunt importante mai ales pentru coacerea fructelor și pentru
căderea frunzelor toamna.
Peretele primar își adaptatează dimensiunile, crescând odată cu celula – este plastic. Peretele celular primar este
mai subțire și mai flexibil decât peretele celular secundar și este uneori păstrat de celulă într-o stare nemodificată (fără
substanțe incrustante) sau puțin modificată fară adăugarea unui perete secundar chiar după ce procesele de creștere s-au
oprit.
Arhitectură moleculară: celuloza formează fibrile care se organizează sub forma unei rețele; fibrilele sunt
solidarizate prin punți transversale de hemiceluloză și glicoproteine. Rețeaua de fibrile celulozice constituie o armătură
sau un schelet care este inclus într-o matrice amorfă reprezentată de substanțe pectice hidratate (analogie cu betonul armat

60 / 108
– armătura metalică și cimentul). Armătura celulozică este
rezistentă la tracțiune/întindere, iar matricea amorfă este
rezistentă la compresiune și conferă peretelui plasticitate.
PC secundar – propriu fiecărei celule; este
depus/format de celulă între peretele primar și plasmalemă după
încetarea creșterii celulare. Dpdv biochimic este format din
celuloză, hemiceluloză, (substanțele pectice pot lipsi) și
frecvent lignină (care conferă rigiditate și rezistență la
compresie), rar cutină și suberină, fără glicoproteine și
enzime. Este caracteristic doar anumitor celule – celulelor
specializate pentru conducerea rapidă a sevei brute (apă cu săruri
minerale dizolvate) prin corpul plantei (vase conducătoare
lemnoase) și celulelor specializate pentru susținerea mecanică a
organelor plantei.
Arhitectură moleculară: schelet de microfibrile de
celuloză inclus în matrice de hemiceluloză; frecvent peretele este
impregnat cu lignină. De obicei sunt depuse succesiv trei straturi
distincte – S1, S2 și S3 – care diferă prin orientarea microfibrilelor
de celuloză.
Din loc în loc, pereții celulari ai celulelor învecinate sunt
întrerupți la același nivel în ambele celule, formându-se un por
care conectează celulele, numit punctuațiune. Acest por sau
canal este căptușit de plasmalemă, astfel plasmalemele celor două celule învecinate se continuă una cu cealaltă. Prin
punctuațiune trec dintr-o celulă în cealaltă tubuli – plasmodesme sau desmotubuli – care derivă din și conectează REN din
cele două celule. Prin intermediul acestor plasmodesme, prin care sunt transportate diverse substanțe, se face legătura între
citoplasmele a două celule învecinate – se realizează astfel solidarizarea funcțională a celulelor în țesut.
*Lignina, cutina, suberina, cerurile fac pereții celulari mai puțin vulnerabili la atacul fungilor și bacteriilor. Cutina, suberina
și cerurile sunt substanțe lipidice întâlnite de obicei în pereții țesuturilor externe, protectoare ale corpului plantei și au rolul în principal
de a reduce pierderea de apă din plantă.

61 / 108
3. Plastidele Plastidele sunt componente caracteristice ale celulelor vegetale și sunt implicate în procese precum fotosinteza și
depozitarea de diverse substanțe. Plastidele sunt organite semiautonome și autodivizibile (se reproduc prin fisiune binară
– proces de divizare în două jumătăți egale, caracteristic bacteriilor). Totalitatea plastidelor dintr-o celulă constituie
plastidomul.
Tipuri de plastide 1. După structură și compoziție chimică, sunt trei tipuri de plastide:
leucoplastele – plastide incolore
cromoplastele – plastide colorate (altfel decât verde)
cloroplastele – plastide verzi
2. După rol, plastidele sunt de două mari categorii:
fotosintetizante (cloroplastele de la plante și echivalentele lor de la alge – cromatoforii)
nefotosintetizante (leuco- și cromoplaste)
3. După gradul de maturitate (vârstă), plastidele sunt de trei categorii, interconvertibile într-o oarecare măsură:
proplast(id)e – plastide tinere, imature structural și funcțional; din ele se formează plastidele mature
plastide mature structural și funcțional
gerontoplaste (plastide îmbătrânite/senescente)
Structură generală Plastidul este un organit vezicular cu înveliș dublu-membranar – membrană externă și membrană internă,
ambele netede, între care există un spațiu intermembranar. Învelișul se mai numește peristromiu.
Lumenul plastidului, delimitat de membrana internă a învelișului, conține un fluid – substanță fundamentală sau
matrice, mai mult sau mai puțin omogenă, numită stromă, cu o compoziței chimică variată și variabilă și în care poate fi
prezentă a treia membrană a plastidului – membrana tilacoidală – care formeză un sistem de structuri membranare cu grad
de dezvoltare variat, în funcție de tipul de plastid.
1.1. Plastidele algelor – cromatofori Plastidele algelor se numesc cromatofori și sunt exclusiv fotosintetizante. Au forme foarte variate – lenticulară,
discoidală, bandiformă (panglică dreaptă sau spiralată), reticulată, stelată, de clopot, de cupă, placă perforată, granulații etc.
Culoarea cromatoforilor este și ea variată – clorofila (care are culoare verde) este prezentă în toate tipurile de cromatofori,
însă uneori alți pigmenți sunt în cantitate mai mare, culoarea lor devenind dominantă – roșu, brun, galben-auriu, brun-
gălbui, verde-gălbui etc.
Din punct de vedere structural, cromatoforii sunt mai variați decât cloroplastele plantelor, având înveliș format din
(1)2-4 membrane și un sistem tilacoidal cu organizare foarte variată. La unele grupe de alge, cromatoforii au denumiri
particulare – rodoplaste (la algele roșii = rodofite), feoplaste (la algele brune = feofite) etc.
1.2. Proplastidele Proplastidele sunt plastide mici, incolore sau verzi pal, nediferențiate (fără sistem tilacoidal), care se întâlnesc în
celulele meristematice (care se divid) din rădăcini și lăstari. Ele sunt precursorii tuturor celorlalte plastide diferențiate,
precum cloroplastele, cromoplastele și leucoplastele.
1.3. Leucoplastele Leucoplastele (plastidele incolore) sunt lipsite de pigmenți și de un sistem elaborat de membrane interne, fiind cele
mai puțin diferențiate, din punct de vedere structural, dintre plastidele mature. Leucoplastele au, de obicei, formă sferică,
eliptică, discoidală. Sunt plastide care depozitează diverse substanțe de rezervă, care sunt sintetizate din precursori chiar la
acest nivel – polizaharide (amidon), proteine (aleuronă), uleiuri, rășini, taninuri etc. În funcție de tipul substanței depozitate,
leucoplastele au denumiri particulare sugestive – amiloplaste (amidon), proteoplaste (proteine), oleoplaste (uleiuri),
rezinoplaste (rășini) etc.
1.4. Cromoplastele Cromoplastele (chroma gr. – culoare), ca și cloroplastele, sunt plastide pigmentate. Cu formă variată (sferică,
ovală, lenticulară, aciculară etc.), cromoplastele sunt lipsite de clorofilă, dar sintetizează și rețin pigmenți carotenoizi, care
sunt responsabili de culorile galbenă, portocalie și roșie ale multor flori, frunze senescente, unele fructe și unele rădăcini, ca
la morcov. Cromoplastele se pot dezvolta din cloroplaste preexistente printr-o transformare în care clorofila și sistemul
membranar intern ale cloroplastului dispar și se acumulează mase de carotenoizi, așa cum se întâmplă în timpul coacerii
multor fructe (tomate, de exemplu). Au un sistem tilacoidal slab dezvoltat, la nivelul căruia să găsesc pigmenții carotenoizi.
Cromoplastele au rol ecologic – prin colorarea anumitor organe ale plantelor acționează ca atractanți pentru
insecte și alte animale, având un rol esențial în polenizarea încrucișată a plantelor cu flori și la dispersia fructelor și a
semințelor.
Carotenoizii sunt de două categorii majore: caroteni (nu conțin oxigen în moleculă; α-, β-, γ-caroten – portocalii
și licopen/licopină – roșie) și xantofile (conțin oxigen în moleculă; numeroase – luteină, neoxantină, zeaxantină etc., de
culoare galbenă, unele sunt brune – fucoxantina).

62 / 108
1.5. Cloroplastele Cloroplastele sunt organite veziculare cu înveliș dublu-membranar, prezente doar în unele celule ale plantelor – în
cele expuse radiației luminoase. Sunt cele mai răspândite plastide și sunt responsabile de culoarea verde caracteristică a
plantelor. Cloroplastele au formă discoidală sau lenticulară. O singură celulă mezofilară conține cca. 40 de cloroplaste, iar
un mm2 de frunză conține aproximativ 500.000 de cloroplaste.
Structură. Cloroplastul are o structură complexă, edificată de trei membrane (1, 2, 7), care delimitează trei
compartimente (A, B, C).
Învelișul cloroplastului (4), numit și anvelopă plastidială sau peristromiu, este format din două membrane –
cloroplastică externă (1) și cloroplastică internă (2), ambele netede, între care se află un spațiu intermembranar – spațiul
periplastidial (A).
Membrana internă delimitează un spațiu (B) în care se găsește un fluid vâscos, gelatinos, cu aspect granular, numit
stromă (5) sau substanță fundamentală. Stroma conține apă, (plasto)ribosomi de 70S (6), granule de amidon (13),
plastoglobuli (12, particule sferice formate în principal din lipide), glucide, acizi organici, aminoacizi, Mg2+
, PO43–
,
diverse tipuri de ARN, ADNclp (11), ATP, ADP, NADPH+H+, NADP
+, diverse tipuri de proteine structurale și
funcționale, dintre care cele mai importante sunt enzimele implicate în faza a doua (fotoindependentă, ciclul Calvin) a
fotosintezei.
ADNclp (11, ADN cloroplastic, ADN plastidial)
este reprezentat de o moleculă bicatenară circulară,
închisă covalent; într-un cloroplast există ~20 de còpii ale
unei singure molecule de ADNclp.
Tot în stromă se găsește a treia membrană a
cloroplastului, membrana tilacoidală (7), independentă
de celelalte două; din punct de vedere biochimic, pe lângă
fosfolipide și diverse tipuri de proteine, conține și
pigmenți asimilatori (clorofile, carotenoizi – caroteni și
xantofile), organizați în fotosisteme. Această membrană
constituie un sistem membranar complex, sistemul
tilacoidal, format din două categorii de structuri:
a) Vezicule aplatizate, cu aspect
discoidal/sacular, numite vezicule
tilacoidale (9) sau vezicule granale, dispuse în stive/teancuri ca monedele în fișic. O asemenea stivă se
numește granum (10), iar totalitatea stivelor dintr-un cloroplast constituie grana.
b) Vezicule alungite, cu aspect lamelar sau tubular, dispuse paralel cu axa longitudinală a cloroplastului și care
trec de la un granum la altul conectându-le; se numesc lamele stromatice (8), tilacoide/lamele intergranale.
Atât veziculele tilacoidale, cât și lamelele stromatice prezintă cavități, toate fiind interconectate rezultând un
sistem membranar tridimensional cu o cavitate unică, spațiul tilacoidal (C).
Rol principal. Cloroplastele reprezintă sediul unuia dintre cele mai importante procese metabolice de pe Pământ –
fotosinteza.
Definiție. Fotosinteza este procesul metabolic și fiziologic prin care organismele fotoautotrofe (majoritatea
plantelor și alte câteva grupe de organisme) își sintetizează compușii organici specifici din compuși anorganici (CO2 și
H2O), folosind lumina (hν) ca sursă de energie, săruri minerale și pigmenți asimilatori drept catalizatori, rezultând și O2 ca
produs rezidual.
Ecuație
CO2 + H2O substanțe organice + O2↑
6CO2 + 6H2O C6H12O6 + 6O2↑
Fotosinteza este un complex proces redox (de oxido-reducere) în care CO2 este redus la compuși organici
(convențional glucoză, C6H12O6) de către H cedat de apă, care astfel se oxidează la O2.
Acest proces se desfășoară în două faze – faza I, fotodependentă („de lumină”) și faza a II-a, fotoindependentă
(„de întuneric”), care se petrec în compartimente diferite ale cloroplastului.
Faza fotodependentă a fotosintezei
Faza I se petrece în membranale tilacoidale și în spațiul tilacoidal și implică două procese care se desfășoară
concomitent: conversia energetică fotochimică și fotoliza apei.
Conversia energetică fotochimică presupune captarea și depozitarea de energie luminoasă în compuși chimici
macroergici – ATP și NADPH (energia luminoasă devine energie chimică), care vor fi folosiți în faza a II-a a fotosintezei.
Fotoliza apei presupune scindarea oxidativă a apei în H+, e
– și O2 (apa este deci sursa oxigenului din fotosinteză!!)
Faza fotoindependentă a fotosintezei În a doua serie de reacții fotosintetice, care se petrece în stromă, ATP și NADPH, generate în faza I, sunt
utilizate/consumate pentru sintetizarea glucidelor simple din CO2.
Fixarea și reducerea C anorganic din CO2 se petrec în stroma cloroplastelor prin intermediul unei serii de reacții
anabolice numită ciclul Calvin-Benson-Bassham.
săruri minerale
pigmenţi asimilatori
hν
pigmenţi asimilatori
hν săruri minerale
glucoză

63 / 108
REPRODUCEREA CELULARĂ
FISIUNEA BINARĂ LA BACTERII
Procariotele (bacteriile) se reproduc printr-un tip de diviziune celulară numit fisiune binară (sciziparitate), care
literal înseamnă „împărțire în două părți (divizare în
jumătăți)”.
Majoritatea genelor bacteriene este localizată într-un
singur cromosom bacterian reprezentat de o moleculă de
ADN circular + proteine asociate = nucleoid. Când replicarea
cromosomului este completă și bacteria a ajuns la aproximaiv
dublul dimensiunilor inițiale, cei doi cromosomi-fii se
îndepărtează unul de celălalt spre poli opuși ai celulei, iar în
zona ecuatorială se formează fie un sept transversal care
crește centripet (la bacteriile Gram-(+)), fie un inel
contractil din proteine FtsZ (similar inelului de clivaj de la
EK) care determină invaginarea circumecuatorială a
plasmalemei și a peretelui celular (la bacteriile Gram-(–)),
împărțind celula parentală în două celule-fiice. Fiecare celulă-
fiică moștenește un genom complet.
CICLUL CELULAR LA EUCARIOTE
Ciclul celular reprezintă viața unei celule din momentul apariției ei prin diviziunea celulei parentale până la
finalul propriei ei diviziuni. Ciclul celular este împărțit în două faze – INTERFAZA și FAZA M/DIVIZIUNEA
CELULARĂ.
Diviziunea celulară reprezintă procesul prin
care celulele se reproduc și presupune împărțirea
conținutului unei celule (celulă-mamă) între/în două
celule-fiice. Diviziunea celulară implică distribuirea de
material genetic identic – ADN – celor două celule-
fiice. O celulă care urmează să se dividă își duplică mai
întâi ADN-ul și trimite două copii la capete opuse ale
celulei și ulterior se scindează în două celule-fiice.
Celulele noi sunt similare atât ca structură cât și ca
funcție cu celula parentală și între ele.
Organizarea celulară a materialului genetic Patrimoniul genetic al celulei, ADN-ul, se numește genom. Genomul celulelor procariote, de obicei, este alcătuit
dintr-o singură moleculă lungă de ADN dublu catenar și circular care constituie singurul cromozom al celulei procariote,
numit și nucleoid.
Genomul nuclear al celulelor eucariote este alcătuit din mai multe molecule de ADN liniar, fiecare moleculă fiind
un cromosom. Înainte ca celula să se dividă, întregul ADN trebuie să fie copiat și apoi cele două copii să fie separate prin
diviziune astfel încât fiecare celulă-fiică să aibă câte un genom complet.
Replicarea și distribuirea unei asemenea cantități enorme de ADN este posibilă deoarece moleculele de ADN sunt
împachetate sub formă de cromosomi (gr. chroma = culoare; soma = corp; corpusculi colorați; se mai numesc și genofori –
purtători de gene). Fiecare specie de eucariote are un număr caracteristic și constant de cromosomi în nucleul fiecărei
celule a corpului.
Fiecare cromosom conține o singură foarte lungă moleculă liniară de ADN care conține de la câteva sute până la
câteva mii de gene – unitățile care codifică trăsăturile ereditare ale unui organism. (vezi Acizii nucleici și Nucleul)
INTERFAZA
Înainte ca o celulă să înceapă să se dividă efectiv, trebuie să se replice ADN-ul și să se sintetizeze mai multe dintre
proteinele asociate cu ADN-ul în cromosom, trebuie de asemenea să producă un supliment de organite și de alte
componente citoplasmatice adecvat pentru două celule-fiice și să asambleze structurile necesare desfășurării fazei M.
Aceste procese de pregătire se petrec în timpul interfazei ciclului celular, împărțită la rândul ei în trei faze: G1, S și G2.
Faza G1 („gol” presintetic) este o perioada de intensă activitate biochimică. Pe măsură ce celula crește în
dimensiuni, își sintetizează mai multe enzime (ADN-polimeraze, ARN-polimeraze), ribosomi, organite, membrane și alte
molecule și structuri citoplasmatice. Cei doi centrioli ai unicului centrosom încep să se separe unul de celălalt și să se
duplice, rezultând doi centrosomi.

64 / 108
În timpul fazei S (faza de sinteză) a ciclului celular se realizează dublarea cantității de informație genetică prin
replicarea ADN (2C → 4C) => cromosomii devin din monocromatidici, bicromatidici; tot în această fază sunt de
asemenea sintetizate multe dintre proteinele asociate ADN-ului, precum histonele.
Rolul principal al fazei G2 („gol” postsintetic/premitotic) este de a verifica dacă replicarea cromosomilor este
completă și corectă și dacă eventualele erori au fost reparate. Duplicarea perechii de centrioli este completă, iar cele două
noi perechi de centrioli compleți sunt situate în apropierea nucleului.
Astfel, celula crește (în G1), continuă să crească pe măsură ce își copiază cromosomii (în S), crește în continuare
pe măsură ce finalizează pregatirile pentru diviziunea celulară (în G2) și se divide (în M).
O celulă umană tipică poate suferi o diviziune la 24 ore. Din acest durată, faza M va ocupa mai puțin de o oră, în
timp ce faza S – 10-12 ore, faza G2 de obicei durează 4-6 ore, iar G1 va dura aproximativ 5-6 ore. Faza G1 este cea mai
variată ca lungime la diferite tipuri de celule.
FAZA M (DIVIZIUNEA CELULARĂ – MITOZA ȘI CITOKINEZA)
MITOZA – diviziunea nucleului – este un proces continuu, dar este împărțită convențional în 4 sau 5 faze:
profaza, (prometafaza), metafaza, anafaza și telofaza. Aceste patru/cinci faze constituie procesul prin care materialul
genetic reduplicat în timpul fazei S este împărțit în mod egal între doi nuclei-fii.
CITOKINEZA – diviziunea citoplasmei – se suprapune ultimei faze a mitozei și încheie faza M a ciclului
celular; în timpul citokinezei citoplasma celulei parentale se separă în cele două celule-fiice.
Rolurile/Importanța diviziunii celulare
La organismele unicelulare precum bacteriile și multe protiste, diviziunea celulară determină creșterea
numărului de indivizi dintr-o populație.
La organismele pluricelulare, precum plantele și animalele, diviziunea celulară (împreună cu creșterea în
dimensiuni a celulei) este mijlocul prin care cresc și se dezvoltă organismele.
Diviziunea celulară este de asemenea mijlocul prin care sunt reparate sau înlocuite țesuturile lezate sau
epuizate (senescente), rol mult mai evident la animale.
Un tip particular de diviziune – meioza – este implicat în reproducerea sexuată și în generarea variabilității
în descendență.
Diviziunea celulară este un proces foarte riguros controlat – celulele se divid doar în anumite momente și
regiuni ale organismului pluricelular și cu anumite rate. Există situații în care controlul diviziunii nu
funcționează corespunzător, iar celulele ajung să se dividă haotic și cu rată foarte înaltă, generând tumori
canceroase – un exemplu de importanță negativă a diviziunii celulare.
Tipuri de diviziune celulară La sistemele biologice se întâlnesc mai multe tipuri de diviziune celulară, clasificate după cum urmează:
1. Diviziune multiplă – caracteristică unor protozoare; celula-mamă se împarte în numeroase celule-fiice
2. Diviziune binară – celula-mamă se împarte în două celule-fiice. Aceasta poate fi:
A. Egală – cele două celule-fiice sunt identice sau aproape identice între ele; la rândul ei, poate fi:
a. Amitoză = diviziune directă: nu se individualizează cromosomii, nu se formează fusul de diviziune,
nu se dezorganizează învelișul nuclear; citoplasma și nucleul se divid sincron; este o diviziune celulară
simplă, întâlnită, în anumite condiții, la plante, la animale și, după unii autori, la procariote(?); se
întâlnește în țesuturile în curs de regenerare, în cele canceroase, la gale, la leucocite, în culturi de
celule pe medii toxice etc.; în funcție de mecanismul de realizare, este de două tipuri:
a.1. Prin strangulare/gâtuire – la jumătatea celulei-mamă apare un inel contractil format
din filamente de actină și miozină, care se strânge treptat și determină invaginarea progresivă
a plasmalemei (celula capătă aspectul unei clepsidre); invaginarea avansează centripet, ca
diafragma unui aparat fotografic, ajungând și la nucleu, care suferă același fenomen de
strangulare; în final, celula-mamă este împărțită în două celule-fiice, fiecare cu nucleul ei; este
întâlnită la celulele lipsite de perete celular.
a.2. Prin formarea unui sept/perete transversal (sau longitudinal) – în zona
circumecuatorială a celulei-mamă se formează un perete despărțitor în alcătuirea căruia intră
atât perete celular, cât și plasmalemă și care crește centripet – spre centrul celulei-mamă,
obligând și nucleul să se dividă; în final, rezultă două celule-fiice mai mult sau mai puțin
egale; este întâlnită la celulele care prezintă perete celular.
b. Cariokineză = diviziune indirectă: se individualizează cromosomii, se formează fusul de diviziune,
se dezorganizează învelișul nuclear; este o diviziune asincronă; este de două tipuri principale:
b.1. Diviziune mitotică (discutată mai jos)
b.2. Diviziune meiotică/reducțională (discutată mai jos)
B. Inegală – cele două celule-fiice au dimensiuni foarte diferite; este caracteristică unor ciuperci (drojdii); se
numește diviziune prin înmugurire: nucleul celulei-mamă se apropie de plasmalemă care, în acea zonă, se evaginează;
nucleul se fragmentează, unul dintre fragmente pătrunde în evaginare, împreună cu o cantitate de citoplasmă; evaginarea se
strangulează bazal și se închide, rezultând o celulă-fiică de mici dimensiuni – un mugure – care rămâne adesea atașată de
cealaltă celulă-fiică, de mari dimensiuni; celula mică în timp va crește, ajungând la dimensiunile celeilalte și a celulei-
mamă.

65 / 108
FAZA M MITOTICĂ
Faza G2 a INTERFAZEI
Anvelopa nucleară este prezentă, delimitând nucleul.
Nucleul conține unul sau mai mulți nucleoli.
Cromosomii, duplicați pe parcursul fazei S a interfazei, încă nu s-au condensat, fiind sub forma filamentelor de cromatină.
S-au format doi centrosomi prin replicarea celui parental. [La celulele animale, fiecare centrosom conține câte doi centrioli.]
A. MITOZA
Profaza Metafaza Anafaza Telofaza Individualizarea cromosomilor prin
supraspiralizarea și condesarea
fibrelor de cromatină. Cromosomii
sunt bicromatidici (replicați).
Dezorganizarea învelișului nuclear.
Dezorganizarea nucleolilor.
Formarea fusului de diviziune prin
deplasarea celor doi centrosomi la poli opuși ai celulei și extinderea
între ei a filamentelor de fus.
Filamentele de fus se prind de cromosomii bicromatidici – un
cromosom este prind de două filamente de fus de la poli opuși.
În celulă există 2n cromosomi bicromatidici => 2n x 2 = 4n
cromatide.
Fusul este complet format – centrosomii
sunt la poli opuși ai
celulei.
Cromosomii, trași de
filamentele de fus, se
așează în placa
metafazică – plan
imaginar echidistant între cei doi poli ai
fusului de diviziune,
deci situat la ecuatorul celulei.
În placa metafazică există 2n cromosomi
bicromatidici => 2n x
2 = 4n cromatide.
Filamentele de fus încep să se contracte/scurteze => fiecare cromosom
bicromatidic este clivat longitudinal la
nivelul centromerului => doi cromosomi monocromatidici (identici ca informație
genetică): unul este prins de un filament
de la un pol la celulei, iar celălalt este prins de filamentul de la polul opus.
Prin scurtarea progresivă a filamentelor de fus, cromosomii monocromatidici
sunt trași la polii celulei-mamă.
La finalul anafazei, la cei doi poli ale celulei se găsesc seturi echivalente și
complete de cromosomi.
În celula-mamă există 2 x 2n = 4n
cromosomi monocromatidici, 2n la un
pol și 2n la polul opus; deoarece cromosomii sunt monocromatidici =>
celula conține 4n cromatide în total.
Se formează cei doi nuclei-fii în celula parentală.
Se dezorganizează fusul de diviziune
Se formează învelișurile
nucleare.
Se reformează nucleolii.
Se decondensează cromosomii => redevin filamente de
cromatină.
Nucleul recapătă aspectul
filamentos reticulat-granular caracteristic interfazei.
În fiecare nucleu există câte 2n
cromosomi monocromatidici.
2n crs bicromat 2n crs bicromat
2n + 2n crs monocromat 2n crs
monocromat
2n crs
monocromat 2n crs
monocromat
2n crs
monocromat
MITOZA – diviziunea unui nucleu în doi nuclei-fii identici din punct de vedere genetic – este acum completă.
B. CITOKINEZA
Diviziunea citoplasmei = citokineza se suprapune parțial telofazei => cele două celule-fiice se individualizează la scurtă vreme după
încheierea mitozei.
La celulele animale, citokineza implică formareă
unui șanț de clivare (format
prin intervenția unui inel
contractil circumecuatorial
alcătuit din microfilamente
și miozină), care strangulează treptat și
împarte celula-mamă în
două celule-fiice.
La celulele plantelor citokineza implică formarea unui sept
transversal, numit placă celulară
(format prin fuziunea de vezicule
Golgi pline cu substanțe pectice
care vor constitui lamela mijlocie),
care apare între cei doi nuclei-fii și crește centrifug, atingând în final
pereții celulei parentale, care este
astfel împărțită în două celule-fiice.
DIVIZIUNEA CELULARĂ ȘI REPRODUCEREA SEXUATĂ
Reproducerea este o proprietate definitorie a sistemelor biologice, care le asigură persistența spațio-temporală.
Alte două proprietăți definitorii sunt ereditatea și variabilitatea. Ereditatea asigură transmiterea cu fidelitate a
caracterelor de la părinți la descendenți și astfel determină asemănarea dintre aceștia. Variabilitatea stă la baza apariției
unor diferențe prin care descendenții nu sunt identici părinților, dar nici între ei, fiecare individ fiind cu adevărat unic din
punct de vedere genetic.

66 / 108
Reproducere asexuată și reproducere sexuată Organismele care se reproduc asexuat (fără fecundație, uniparental) produc descendenți care sunt còpii exacte ale
organismelor parentale. În reproducerea asexuată părinte este un singur individ și transmite descendenților còpii ale
tuturor genelor sale. Un individ care se reproduce asexuat dă naștere unei clone – un individ sau un grup de indivizi genetic
identici între ei și cu părintele care i-a produs.
Prin reproducere sexuată, doi părinți generează un descendent care are o combinație unică de gene moștenite de la
cei doi părinți. Spre deosebire de o clonă, descendenții rezultați prin reproducere sexuată diferă genetic față de frații lor și
de ambii părinți. Diversitatea/variabilitatea genetică este o consecință importantă a reproducerii sexuate.
Bazele citologice ale eredității și variabilității Părinții își înzestrează descendenții cu informație codificată sub forma unităților ereditare numite gene. La animale
și plante, celulele reproducătoare, numite gameți, sunt vehiculele care transmit genele de la o generație la următoarea. Prin
fecundație, la animale, gametul masculin (spermatozoidul) și feminin (ovulul) se unesc formând zigotul/celula-ou – prima
celulă a noii generații – care conține gene de la ambii părinți.
Meioza, un tip particular de diviziune celulară, la animale se desfășoară doar în gonade13
(ovare și testicule) și
duce la formarea gameților. În fiecare generație, meioza reduce/înjumătățește numărul de cromosomi de la 2n (două
seturi), caracteristic speciei, la n (un singur set), compensându-se astfel dublarea care se petrece în fecundație.
Fecundația presupune fuziunea a doi gameți de sex opus, fiecare cu câte n cromosomi, și deci restabilește
numărul de cromosomi la 2n, iar mitoza conservă acest număr în celulele somatice ale noului individ.
Examinarea celor 2n cromosomi dintr-o singură celulă în mitoză relevă existența a câte doi cromosomi din fiecare
tip. Cei doi cromosomi care formează o pereche au origini diferite – unul provine de la mama individului (cromosom
matern), celălalt – de la tatăl individului (cromosom patern), au aceeași lungime, aceeași poziție a centromerului și
același model de bandare și se numesc cromosomi omologi.
Ambii cromosomi din fiecare pereche conțin genele care codifică pentru aceleași caractere ereditare, nu
neapărat pentru aceleași variante ale caracterelor. De exemplu, dacă gena pentru culoarea ochilor este situată într-un anumit
locus dintr-un anumit cromosom, atunci omologul acelui cromosom va conține în locusul echivalent aceeași genă (cea care
specifică pentru culoarea ochilor), dar una poate să codifice pentru varianta albastră, iar cealaltă pentru varianta căpruie a
culorii ochilor (vezi Capitolul II, Elemente de genetică).
Astfel, cei 2n cromosomi dintr-o celulă diploidă pot fi grupați în n perechi de omologi, fiecare pereche conținând
un cromosom de origine maternă și unul de origine paternă.
Regrupând cei 2n cromosomi după originea lor, rezultă două seturi a câte n cromosomi fiecare – un set de n
cromosomi materni (contribuția mamei la formarea individului respectiv) și un set de n cromosomi paterni (contribuția
tatălui la formarea aceluiași individ); fiecărui cromosom dintr-un anumit set n îi corespunde un cromosom – numit
cromosom omolog – din celălalt set n.
Seturi de cromosomi în celulele umane La oameni, fiecare celulă somatică – oricare altă celulă a corpului în afară de gameți – conține 46 cromosomi.
Cei doi cromosomi distincți, numiți X și Y, reprezintă o importantă excepție de la tiparul general al cromosomilor
omologi în celulele somatice umane. Femeile au o pereche de cromosomi omologi X (XX), dar bărbații au un cromosom X
și un cromosom Y, neomolog lui X (XY). Deoarece acești cromosomi se corelează cu și determină sexul individului,
cromosomii X și Y se numesc cromosomii sexului = heterosomi. Ceilalți cromosomi se numesc autosomi.
Moștenim un cromosom din fiecare pereche de la fiecare părinte. Deci cei 46 de cromosomi din celulele noastre
somatice sunt de fapt două seturi de câte 23 cromosomi – un set matern (de la mamă) și un set de proveniență paternă (de la
tată). Numărul de cromosomi dintr-un singur set se notează cu n. Orice celulă cu două seturi de cromosomi se numește
celulă diploidă și are un număr diploid de cromosomi, abreviat 2n. Pentru oameni, numărul diploid este 46 (2n=46) –
numărul de cromosomi din celulele noastre somatice.
Spre deosebire de celulele somatice, gameții (spermatozoid și ovul) conțin un singur set de cromosomi. Asemenea
celule se numesc celule haploide și fiecare conține un număr haploid de cromosomi (n). Pentru oameni numărul haploid
este 23 (n = 23) – numărul de cromosomi existent în gameți.
FAZA M MEIOTICĂ Meioza, ca și mitoza, este precedată de replicarea cromosomilor. Totuși această unică replicare este urmată de
două diviziuni celulare consecutive, numite meioza I și meioza II. În urma acestor două diviziuni rezultă patru celule-fiice
(în loc de două celule-fiice în mitoză), fiecare cu doar jumătate din numărul de cromosomi al celulei parentale.
Interfaza
Cromosomii se replică pe parcursul fazei S, dar rămân necondensați (sub forma filamentelor de cromatină).
Fiecare cromosom replicat constă în două cromatide-surori identice din punct de vedere genetic, conectate pe întreaga
lungime, cu maxim de atașare la nivelul centromerului. Centrosomul se duplică, formându-se doi centrosomi.
13 La plante meioza se petrece în organele reproducătoare masculine – antere – și feminine – ovule, rezultând spori din care se vor forma, în final, gameții
masculini – spermatiile – și feminini – oosferele.

67 / 108
MEIOZA I (etapa reducțională)
Profaza I Metafaza I Anafaza I Telofaza I și
citokineza I
Reprezintă mai mult de 90% din durata meiozei.
Se individualizează cromosomii bicromatidici (2n).
Cromosomii omologi se caută, se găsesc și se împerechează,
formându-se perechi de omologi = bivalenți = tetrade
cromatidiale (grup de patru cromatide) => cei 2n crs bicromat formează n perechi de omologi
Sinapsa cromosomală: în fiecare bivalent cei doi omologi sunt aliniați cu mare precizie genă la genă, centromer la centromer și
se alipesc pe întreaga lungime prin intermediul unei structuri
proteice cu aspect de fermoar (numită complex sinaptonemal).
Sinapsa favorizează producerea crossing-over-ul –
recombinarea intracromosomală
Fiecare tetradă prezintă 1-n chiasme – regiuni de contact între
cromatidele nesurori unde s-a petrecut crossing-over; acestea
mențin împreună omologii până la finalul metafazei I.
Se dezorganizează învelișul nuclear și nucleolii.
Se formează un aparat fusorial/fus de diviziune (ca și în mitoză).
Cele n perechi de omologi bicromatidici, prinse de
microtubuli de la poli opuși, se deplasează spre placa metafazică I trase de filamentele de fus.
Se petrece și recombinarea intercromosomală, „dansul
cromosomilor” – alt mecanism de generare a variabilității în
descendență și în populație.
Perechile de omologi sunt aranjate în placa
metafazică cu câte un cromosom bicromatidic
din fiecare pereche
orientat spre un pol, iar celălalt spre polul opus,
atașați de microtubuli
de la poli opuși.
Prin scurtarea progresivă a
microtubulilor se desfac bivalenții în cei
doi omologi
bicromatidici.
Cromosomii
bicromatidici omologi se deplasează spre poli
opuși, trași de
filamentele de fus.
Spre fiecare pol se
deplasează câte n cromosomi
bicromatidici
Cromatidele-surori
rămân atașate între ele
– cromosomul
bicromatidic nu clivează longitudinal
Cei n cromosomi bicromatidici de la
fiecare pol al celulei-mamă se
decondensează.
Se reformează învelișul nuclear și nucleolii =>
doi nuclei-fii haploizi.
Citokineza se desfășoară de obicei
simultan cu telofaza I => două celule-fiice
haploide.
La celula animală se
formează un șanț de
clivare
La celulele plantelor se formează placă
celulară.
2n crs bicromat
inițial distincți, apoi grupați în
n perechi de omologi
2n crs bicromat
grupați în
n perechi de omologi
n + n crs bicromat n crs
bicromat
n crs
bicromat n crs
bicromat
n crs
bicromat
Meioza I se numește și etapă reducțională deoarece înjumătățește (reduce) numărul de seturi de cromosomi per celulă – o reducere de la două seturi
(starea diploidă) la un set (starea haploidă).
INTERKINEZA (interfaza II) Cele două celule-fiice haploide intră într-o scurtă (de obicei) interfază II = interkineză, perioadă în care nu se produce replicarea cromosomilor, deoarece aceștia sunt deja replicați (sunt bicromatidici).
MEIOZA II (etapa ecvațională)
Profaza II Metafaza II Anafaza II Telofaza II și citokineza II
Se individualizează cromosomii
bicromatidici (n).
Se dezorganizează învelișul
nuclear și nucleolii.
Se formează un aparat fusorial.
Cei n cromosomi bicromatidici, prinși de microtubuli de la poli
opuși, se deplasează spre placa
metafazică II.
Cromosomii bicromatidici sunt
poziționați în placa metafazică la fel ca în mitoză, atașați de
microtubuli care se extind de la
poli opuși.
Ca urmare a crossing-over-ului
din meioza I, cele două
cromatide-surori ale fiecărui cromosom nu sunt identice
genetic.
Cei n cromosomi bicromatidici,
prin scurtarea/contracția progresivă a microtubulilor,
clivează longitudinal și
cromatidele-surori se despart,
devenind cromosomi
monocromatidici
Din n cromosomi bicromatidici rezultă n+n cromosomi
monocromatidici – n cromosomi
monocromatidici spre un pol și n cromosomi monocromatidici spre
celălalt pol.
Cei n cromosomi
monocromatidici de la fiecare pol al celulei-mamă se
decondensează
Se reformează învelișul nuclear și
nucleolii => doi nuclei-fii
haploizi.
În paralel se petrece citokineza.
n crs bicromat n crs bicromat
n + n crs monocromat n crs
monocromat
n crs
monocromat n crs
monocromat
n crs
monocromat
Meioza II se numește și etapă ecvațională deoarece numărul de cromosomi per celulă rămâne constant (lat. aequalis – egal) de la o generație celulară la
următoarea (din celula-mamă cu n cromosomi rezultă două celule-fiice fiecare cu câte n cromosomi), ca și în cazul mitozei, cu diferența că aceasta din
urmă se desfășoară pe celule diploide 2n.
Diviziunea meiotică a unei celule parentale produce patru celule, fiecare cu un set haploid, nereplicat și recombinat, de cromosomi. Fiecare din cele patru
celule-fiice este diferită genetic de celelalte și de celula parentală. Cele patru celule sunt gameți la animale și spori la plante.

68 / 108
Comparație între mitoză și meioză Meioza reduce numărul seturilor de
cromosomi de la două (diploid) la unul (haploid), în
timp ce mitoza conservă numărul de seturi
cromosomale. Ca urmare, mitoza produce celule-fiice
identice din punct de vedere genetic cu celula parentală,
în timp ce meioza produce celule care diferă genetic
față de celula parentală și una față de celelalte.
Trei evenimente sunt unice pentru meioză,
toate trei petrecându-se în timpul meiozei I:
1. Sinapsa și crossing-over-ul. În timpul
profazei I, cromosomii omologi duplicați
(bicromatidici) se aliniază și se conectează fizic pe
întreaga lor lungime prin intermediul unei structuri
proteice în formă de fermoar – complexul
sinaptonemal, rezultând un bivalent sau tetradă
cromatidială. Această legătură este fundalul pe care se
realizează, în timpul profazei I, crossing-over-ul
(recombinarea intracromosomală) – schimbul reciproc
și egal de fragmente cromatidiale între cromosomii
omologi. Fiecare tetradă conține în mod normal cel
puțin o regiune în formă de „X” numită chiasmă – dovada fizică a crossing-over-ului.
2. Tetrade în placa metafazică. În metafaza I a meiozei se poziționează în placa metafazică cromosomii omologi
sinapsați (tetrade) și nu cromosomi replicați (bicromatidici) individuali ca în mitoză.
3. Separarea omologilor. În anafaza I a meiozei se separă perechea de omologi, adică cromosomii duplicați din
fiecare bivalent se deplasează spre polii opuși, dar fiecare cromosom rămâne bicromatidic, neproducându-se clivarea
longitudinală a centromerului ca în mitoză.
§Sursele variației genetice Mutațiile reprezintă sursa primară a diversității genetice. Aceste modificări în ADN-ul unui organism creează
diferitele variante ale genelor. Odată aceste diferențe apărute, reamestecarea variantelor în timpul reproducerii sexuate
generează variația pe care o manifestă fiecare membru al unei specii având propria combinație de trăsături.
La speciile care se reproduc sexuat, comportamentul cromosomilor în timpul meiozei și fecundației este
responsabil pentru majoritatea variabilității care apare la fiecare generație. Există trei mecanisme principale care
contribuie la variația genetică apărută prin reproducerea sexuată: segregarea independentă a perechilor de omologi
(recombinarea intercromosomală), crossing-over-ul (recombinarea intracromosomală) și fecundația aleatorie/randomizată.
1. Segregarea independentă a cromosomilor
În metafaza I, perechile de omologi, fiecare alcătuită dintr-un cromosom matern și unul patern, sunt situate în
placa metafazică. Fiecare pereche se poate orienta fie cu omologul matern, fie cu cel patern mai aproape de un pol dat.
Astfel există 50% șansa ca o anumită celulă-fiică a meiozei I să primească cromosomul matern dintr-o anumită pereche de
omologi și 50% șanse că va primi cromosomul patern din aceeași pereche.
Deoarece fiecare pereche de cromosomi omologi se orientează independent de celelalte perechi în metafaza I, în
urma meiozei I rezultă o separare și distribuire a omologilor materni și paterni în celulele-fiice, independente pentru
fiecare pereche în parte raportat la toate celelalte perechi. Aceasta se numește segregare independentă.
Exemplu. Fiecare celulă-fiică reprezintă una din toate combinațiile posibile de cromosomi materni și paterni.
Numărul de combinații posibile pentru celulele-fiice formate prin meioza unei celule diploide cu două perechi de
cromosomi omologi (n = 2, 2n = 4) este 4 (patru), adică 22. De remarcat că doar două din aceste patru combinații de celule-
fiice vor rezulta din meioza unei singure celule diploide, deoarece o singură celulă parentală va avea doar una sau cealaltă
posibilitate de aranjare a cromosomilor în metafaza I, dar nu pe ambele. Totuși, populația de celule-fiice care rezultă din
meioza unui număr mare de celule diploide conține toate cele patru tipuri în proporții aproximativ egale. În cazul în care n
= 3, 2n = 6, sunt posibile 8, adică 23 combinații de
celule-fiice. Generalizând, numărul de combinații
posibile când cromosomii segregă independent în
timpul meiozei este 2n
, unde n este numărul
haploid al organismului.
În cazul oamenilor, numărul haploid (n)
din formulă este 23. Astfel, numărul de combinații
posibile de cromosomi materni și paterni în gameți
este 223
, adică aproximativ 8 milioane (8.388.608).
Fiecare gamet pe care îl produce un individ
conține doar una din cele aproximativ 8 milioane
de combinații posibile de cromosomi moșteniți de
la mamă și de la tată.

69 / 108
2. Crossing-over-ul
Crossing-over-ul începe foarte devreme în profaza I, când cromosomii omologi se împerechează lax pe toată
lungimea lor. Fiecare genă dintr-un omolog este precis aliniată cu gena corespondentă din celălalt omolog. Într-un singur
eveniment de crossing-over, moleculele de ADN din două cromatide nesurori – o cromatidă maternă și una paternă din
perechea de omologi – se rup în același loc și apoi fragmentele sunt reunite fiecare la cealaltă moleculă de ADN. Ca efect,
două segmente omoloage schimbă locul sau „se încrucișează” (cross over), producând cromosomi cu o nouă combinație de
gene, atât materne, cât și paterne.
Segregarea independentă a acestor cromatide surori neidentice în timpul meiozei II amplifică și mai mult numărul
de tipuri genetice ale celulelor-fiice care rezultă din meioză.
3. Fecundația aleatorie – vezi premisa/ipoteza a 4-a a lui Mendel
Exemplu. La oameni, spre exemplu, fiecare gamet masculin și fiecare gamet feminin reprezintă una din
aproximativ 8 milioane de combinații posibile de cromosomi ca urmare a segregării independente din timpul meiozei.
Fuziunea unui singur gamet masculin cu un singur gamet feminin în timpul fecundației va produce un zigot cu oricare din
aproximativ 64 trilioane (8 milioane x 8 milioane) de combinații diploide posibile. Dacă se calculează 223
x 223
exact, se va
obține un total care este de fapt de peste 70 trilioane (70.368.744.177.664). Adăugând această variație la cea generată prin
crossing-over, numărul de posibilități este cu adevărat astronomic.
Evenimentele fazei M meiotice
SARCINĂ DE LUCRU. Ordonați și schematizați etapele fazei M mitotice
1 2 3
4 5 6

70 / 108
A B

71 / 108
Capitolul II
ELEMENTE DE GENETICĂ

72 / 108

73 / 108
INTRODUCERE
Genetica este știința care studiază ereditatea și variabilitatea organismelor.
Termenul „genetică” a fost introdus de englezul William Bateson (1861-1926), în 1906, la a treia Conferință
Internațională de Hibridarea Plantelor.
În 1909, Wilhelm Ludwig Johannsen (1857-1927, botanist danez) a dat denumirea de „gene” unităților de
informație ereditară și a introdus termenii „genotip” și „fenotip”.
Ereditatea (lat. hereditas – moștenire) este proprietatea fundamentală a ființelor vii prin care se asigură
transmiterea cu fidelitate a caracterelor de la părinți la descendenți și prin care este astfel determinată asemănarea dintre
aceștia. Reprezintă aspectul continuității fenomenului ereditar.
Caracterele ereditare sunt acele trăsături morfo-anatomice, fiziologice, biochimice și comportamentale care se
transmit constant (de fiecare dată) și cu mare fidelitate (în aceeași formă), de la părinți la descendenți, de-a lungul
generațiilor.
Dar descendenții doar seamănă, nu sunt identici cu părinți și nici între ei, având și trăsături proprii, adică există
și deosebiri între părinți și descendenți, denumite variații – expresia discontinuității fenomenului ereditar. Variațiile de
care se ocupă genetica sunt variații ereditare – odată apărute într-o generație se vor transmite și generațiilor următoare.
Variabilitatea este o latură a fenomenului ereditar și o lege universală a lumii vii care stă la baza apariției unor
diferențe prin care descendenții nu sunt identici părinților, dar nici între ei, fiecare individ fiind cu adevărat unic din punct
de vedere genetic.
Există și variații neereditare (numite modificații) – deosebiri aparente, exterioare, apărute în urma acțiunii unor
condiții particulare de mediu – unele maladii (virozele, bacteriozele, micozele, parazitozele etc.), bronzarea pielii, părul
vopsit, membre amputate, informația acumulată în cursul vieții etc.
Etapele dezvoltării geneticii ca știință
1. Genetica clasică/factorială – 1865, Johann Gregor Mendel; 1900, Carl Correns, Erich von Tschermak-
Seysenegg, Hugo de Vries
2. Citogenetica – 1902-1903, Walter Sutton, Theodor Boveri; 1906-1913, Thomas Hunt Morgan și colab. (Alfred
Sturtevant, Calvin Blackman Bridges, Hermann Joseph Muller)
3. Genetica populațiilor – 1906, William Ernest Castle, Charles W. Woodworth
4. Genetica moleculară – 1944, Oswald Theodore Avery, Colin Munro MacLeod, Maclyn McCarty; 1953, James
Dewey Watson, Francis Harry Compton Crick, Maurice Hugh Frederick Wilkins, Rosalind Elsie Franklin
Primele cunoștințe legate de ereditate se conturează odată cu sedentarizarea omului. Acesta renunță la viața
nomadă și începe să crească animale și să cultive plante (adică să practice agricultura). În cadrul acestor îndeletniciri, omul,
în vederea păstrării și amplificării unor proprietăți ale animalelor domestice și ale plantelor cultivate, recurge la selecție
artificială – alegerea și păstrarea pentru perpetuare a acelor indivizi care prezintă calitățile dorite. Selecția artificială se
bazează pe cunoașterea faptului că trăsăturile organismelor se moștenesc în mare măsură în formă identică sau foarte
asemănătoare de la o generație la alta, care înseamnă chiar fenomenul ereditar. Sub forma selecției artificiale a fost
înțeles și s-a operat cu fenomenul ereditar, în mod empiric, până în 1865, an în care au fost puse bazele științifice ale
științei care urma să se numească genetică.
În 1865 Johan Gregor Mendel, prin experimentele sale ale căror rezultate s-au materializat în lucrarea „Cercetări
asupra hibridării plantelor”, pune bazele geneticii ca știință; acest an este considerat anul nașterii geneticii, iar Mendel –
părintele acesteia.
GENETICĂ FACTORIALĂ (MENDELIANĂ, CLASICĂ)
Legile mendeliene ale eredității
(Teoria factorilor ereditari)
GENERALITĂȚI
Johann Mendel (20 iulie1822 – 6 ianuarie 1884) s-a născut într-o familie de etnici germani dintr-o localitate din
fostul Imperiu Habsburgic – Hynčice (Vražné) – care astăzi face parte din Republica Cehă și care, la recensământul din
2001, totaliza 58 de locuințe și 232 de rezidenți.
În copilărie a lucrat la ferma familiei, practicând grădinărit și apicultură. Între 1840 și 1843 a studiat filosofie
teoretică și practică și fizică la Universitatea Palacký din Olomouc, Moravia, Republica Cehă. Aici îl are ca profesor pe
Johann Karl Nestler, șeful Departamentului de Istorie Naturală și Agricultură, care îi va inspira cercetările de mai târziu
prin propriile sale preocupări în domeniul trasmiterii ereditare a caracterelor, realizând numeroase experimente la plante și
animale, în special la oi.
La îndemnul profesorului său de fizică și matematici aplicate, Friedrich Franz, Mendel începe pregătirea
clericală în 1843 la Abația augustiniană „Sfântul Thomas” din Brno, unde își ia numele Gregor.

74 / 108
În perioada 1851-1853, susținut de abatele mânăstirii, aprofundează fizica la Universitatea din Viena, avându-l
ca profesor pe Christian Doppler, revenind în Brno ca profesor în special de fizică. În 1867 devine/preia funcția de abate
al mânăstirii.
În perioada 1856/57-1963/65 (7-8 ani) Mendel realizează experimente de hibridare la/cu mai multe specii de
plante (Phaseolus sp. – fasole, Mirabilis sp. – barba-împăratului/frumoasa nopții, Cirsium sp. – pălămidă, ciuline, Linaria
sp. – linariță, Hieracium sp. – vulturică, Calceolaria sp. – papucel, pantofior), în special la Pisum sativum – mazăre, în cele
două hectare ale grădinii mânăstirii. Timp de doi ani, Mendel a analizat cca. 29.000 de plante de mazăre din 34 de soiuri,
din care a selectat 22 de soiuri, anume pe acelea care prezentau caractere constante și distincte și produceau o descendență
omogenă/uniformă, adică erau soiuri pure.
Pe lângă preocupările botanice, Mendel a crescut și albine în stupi de concepție proprie și a studiat astronomia și
meteorologia (în 1865 a înființat Societatea Meteorologică Austriacă), cele mai multe dintre articolele publicate fiind din
domeniul meteorologiei.
Pisum sativum este model ideal de studiu al fenomenului ereditar deoarece, pe de o parte, (1) se reproduce
sexuat prin autogamie/autofecundație consecutivă autopolenizării, ceea ce înseamnă că, în absența mutațiilor, își păstrează
puritatea, adică structura genetică și constanța caracterelor de-a lungul generațiilor; pe de altă parte, (2) se pretează la
polenizare artificială încrucișată consecutiv castrării florilor (din floarea bisexuată se îndepărtează, înainte de polenizare,
elementele sexuale bărbătești – staminele), adică la hibrid(iz)are.
Hibrid(iz)area reprezintă încrucișarea (pe cale naturală sau artificială) a două organisme (de obicei din aceeași
specie, dar și din specii diferite) care diferă prin unul sau mai multe caractere (monohibridare, dihibridare, polihibridare).
Individul rezultat, numit hibrid, este impur din punct de vedere genetic, deci heterozigot.
CONCEPTE ȘI NOȚIUNI Genotipul reprezintă natura constituției genetice a unui organism (factorii ereditari/genele + calitatea lor de a fi
dominanți sau recesivi)
Fenotipul reprezintă totalitatea caracterelor morfo-anatomice, fiziologice, biochimice și de comportament care
rezultă din interacțiunea genotipului cu mediul și care pot fi cumva observate, măsurate, apreciate cantitativ
și/sau calitativ
Genotipul stabilește norma de reacție a fenotipului (adică limitele între care poate să varieze fenotipul)
Organisme pure = homozigote – acele organisme care păstrează constant nemodificate caracterele de-a lungul
generațiilor; nu segregă în descendență
Organisme impure = heterozigote – de la generație la generație apar variații în genotip și în fenotip; ele segregă
în descendență
Gameții, indiferent de genotipul (homo- sau heterozigot) organismului care i-a produs sunt întotdeauna puri din
punct de vedere genetic
ORGANIZAREA FLORII BISEXUATE/PERFECTE DE LA ANGIOSPERME Floarea este un complex de organe al plantelor superioare al cărui rol este reproducerea sexuată. Clasic, se
consideră că au flori pinofitele (gimnospermele) și magnoliofitele
(angiospermele), dar în prezent sunt considerate flori doar complexele de
organe cu rol reproducător de la magnoliofite (numite de aceea și antofite,
din gr. anthos – floare, phyton – plantă).
O floare completă este alcătuită din:
Peduncul/pedicel floral – la extremitatea superioară poartă
celelalte elemente ale florii; poate lipsi => floare sesilă
Receptacul (axă florală) – extremitatea superioară (cu
morfologie variată – lățită, alungită sau scobită) a
pedunculului floral, pe care se inseră celelalte elemente ale
florii
Înveliș floral – protejează organele reproducătoare, atrage
polenizatorii; poate fi periant – diferențiat în sepale (a
căror totalitate constituie caliciul, K) și petale (a căror
totalitate constituie corola, C) – sau perigon – format din
tepale (poate fi sepaloid, dacă tepalele seamănă cu sepalele,
sau petaloid, dacă tepalele seamănă cu petalele).
Organe reproducătoare masculine – stamine (a căror
totalitate constituie androceul, A) – și feminine – carpele (a
căror totalitate constituie gineceul/pistilul, G)
Stamina are două componente – filament (parte sterilă) în vârful
căreia se găsește antera (partea fertilă); filamentul se prelungește în anteră
sub forma conectivului. La nivelul anterei se găsește țesutul sporogen, format din celule-mamă de spori diploide. Din
aceste celule, prin meioză, se vor forma microsporii n (celulele-mamă ale granulelor de polen). Microsporii suferă o
mitoză haploidă și devin granule de polen; una dintre celulele granulei se mai divide o dată mitotic și va forma două
spermatii – gameții masculini.

75 / 108
Gineceul (pistilul) are trei componente – ovar (partea inferioară,
mai dilatată), stil (regiune conductivă pentru tubul polenic) și stigmat (partea
superioară, lățită și lipicioasă a stilului; regiunea de recepție a polenului). În
interiorul ovarului se găsesc 1-n ovule, atașate de peretele acestuia la nivelul
unor zone specializate, numite placente. Ovulul este o structură pluricelulară
complexă formată din funicul și corp. Corpul este format din nucelă și 1-2
integumente care acoperă nucela, lăsând la partea apicală un orificiu numit
micropil. La nivelul nucelei se găsește țesutul sporogen 2n dintr-o celulă a
căruia se vor forma, prin meioză, 4 macrospori n, dintre care numai unul este
funcțional și reprezintă celula-mamă a sacului embrionar. Acest macrospor
funcțional se divide mitotic de trei ori, fără citokineză, rezultând 8 nuclei n
dintre care, prin celularizare, unul va intra în alcătuirea oosferei – gametul
feminin propriu-zis, iar alți doi (nucleii polari) vor fuziona => un nucleu 2n care va intra în alcătuirea celulei centrale –
gametul feminin secundar/accesoriu.
MONOHIBRIDAREA ȘI PRIMELE DOUĂ LEGI ALE EREDITĂȚII
Monohibridarea reprezintă încrucișarea a două organisme care se deosebesc printr-un singur caracter (sau, cum
se mai spune în mod eronat, printr-o singură pereche de caractere).
Într-o primă serie de experimente, Mendel a urmărit transmiterea ereditară a câte unui caracter exprimat prin două
variante fenotipice alelomorfe/contrastante. Caracterele urmărite de către Mendel și variantele lor alelomorfe (contrastante)
pereche sunt prezentate în tabelul 1.
Tabelul 1. Caractere urmărite de Mendel în experimentele de monohibridare
Caracter Variante fenotipice alelomorfe
Talia plantei Înaltă Scundă
Culoarea seminței/cotiledoanelor Galbenă Verde
Forma seminței (bobului) Netedă Zbârcită
Culoarea florii Roșie Albă
Culoarea fructului (păstăii) Verde Galbenă
Forma fructului (păstăii) Dreaptă Strangulată
Poziția florilor pe tulpină Axilară Axială
În primul experiment de monohibridare Mendel a urmărit transmiterea ereditară a caracterului „talia plantei”. În al
doilea experiment a urmărit transmiterea ereditară a caracterului „forma seminței/bobului”, acesta devenind clasic în
prezentarea descoperirilor lui Mendel.
Astfel, Mendel a ales două soiuri pure de mazăre care se deosebeau prin forma seminței (bobului) – un soi făcea
semințe netede (bogate în amidon), iar celălalt soi făcea semințe zbârcite (conțin dextrină). Nu a luat în calcul celelalte
diferențe între cele două soiuri, concentrându-și atenția doar asupra formei seminței. Mendel știa că soiurile sunt pure cu
privire la forma seminței, deoarece plantele care rezultau din semințe netede făceau, la rândul lor, numai semințe netede și
acest lucru se repeta anual; similar și pentru soiul cu semințe zbârcite. Deci caracterul era constant de-a lungul generațiilor,
ceea ce însemna că erau soiuri pure.
Anul I
În primăvara primului an de experimente, Mendel a semănat cele două tipuri de semințe, care au germinat și au
rezultat două populații de plante de mazăre, aparținând la soiuri diferite.
În perioada mai-iunie, plantele au făcut flori; dacă Mendel nu ar fi intervenit, fiecare floare ar fi ajuns la
maturitate, ar fi produs polen care ar fi căzut pe stigmatul gineceului aceleiași flori – autopolenizare, iar în urma fecundației
– autofecundație, s-ar fi format fructe – păstăi, cu semințe netede sau zbârcite, în conformitate cu tipul de bob din care
provenea planta.
Mendel însă a deschis florile soiului cu semințe zbârcite înainte ca partea bărbătească (staminele) a acestora să
ajungă la maturitate și să producă polen și a îndepărtat, prin tăiere, staminele (a realizat castrarea florilor). Astfel,
respectivele flori (și plante în ansamblu) au rămas unisexuate femele și, în absența granulelor de polen, aceste flori nu ar
forma semințe, reproducerea sexuată fiind compromisă. NU UITAȚI, este vorba despre soiul pur cu semințe zbârcite, care
ar fi produs tot semințe zbârcite dacă Mendel nu intervenea și nu ar fi produs semințe dacă intevenția lui se oprea la
castrare.
Când florile soiului cu sămânță netedă au ajuns la maturitate și staminele acestora au produs polen, a prelevat cu o
pensulă fină granule de polen, care reprezintă sexul masculin, și le-a adus pe stigmatul gineceului florilor castrate ale
soiului cu sămânță zbârcită – a realizat polenizare încrucișată artificială. Din granulele de polen s-au format gameții
masculini (spermatii) care au fecundat gameții feminini (oosfere). NU UITAȚI, cele două tipuri de gameți proveneau de la
soiuri diferite, fiecare contribuind la formarea noii generații cu informație specifică soiului de origine. În urma fecundației
se formează zigotul, iar acesta se divide mitotic și formează embrionul, inclus în sămânță.
Acest embrion, împreună cu sămânța din care face parte, reprezintă primul stadiu al noii generații, numită F1
(prima filiație), iar cele două plante/soiuri care au contribuit, prin gameții lor, la formarea lui reprezintă genitorii (G) sau
părinții (P). În cazul autopolenizării, părintele este unic, în cazul polenizării încrucișate există doi părinți.
Dacă s-a realizat fecundație, florile soiului cu sămânță zbârcită au format fructe cu semințe. Este vorba de semințe
formate pe plantele din soiul cu bob zbârcit, care însă au fost castrate înainte de autopolenizare și polenizate artificial de

76 / 108
către soiul cu sămânță netedă. Ce fel de semințe a găsit Mendel în păstăile formate de plantele din soiul cu sămânță
zbârcită? Numai zbârcite? Numai netede? Zbârcite și netede?
Plantele din soiul cu bob zbârcit, polenizate artificial cu polen de la soiul cu bob neted, au format NUMAI boabe
NETEDE!
Aceste boabe reprezintă primul stadiu al generației F1 și, deoarece au rezultat prin hibridare, sunt hibride/impure
(ca și generația în sine).
Pe baza acestor rezultate, Mendel a formulat prima lege a eredității – LEGEA UNIFORMITĂȚII
HIBRIZILOR ÎN/DIN PRIMA GENERAȚIE.
Caracterul vizat a fost forma bobului, iar variantele fenotipice – neted și zbârcit. Observând comportarea la
hibridare a celor două variante fenotipice ale caracterului, Mendel a tras concluzia că ceea ce determină forma netedă este
mai „tare” decât ceea ce determină forma „zbârcită” și că atunci când acești determinanți se întâlnesc se va manifesta doar
cel care determină forma netedă, pe care l-a numit DOMINANT. De asemenea, este dominantă și varianta fenotipică
determinată, adică forma netedă a bobului. Determinantul celeilalte variante, care nu s-a mai manifestat în F1, a fost numit
RECESIV, iar varianta fenotipică zbârcită – recesivă.
Mendel a păstrat aceste boabe hibride F1 pentru a le cultiva anul următor.
Anul al II-lea
În primăvara celui de-al doilea an de experimente, Mendel însămânțează boabele netede hibide F1; acestea
germinează și rezultă o populație de plante F1 hibride.
În perioada mai-iunie, plantele au făcut flori; de data aceasta, Mendel nu a mai intervenit în desfășurarea
reproducerii, iar în flori s-a petrecut autopolenizare, urmată de autofecundație.
După fecundație s-au format zigoții – primele celule ale generației a doua, F2 – care se divid mitotic formând
embrioni incluși în semințe. Aceste semințe/boabe reprezintă stadiul inițial al generației F2 și se formează în păstăile
rezultate din florile plantelor hibride F1.
Mendel a așteptat ca fructele și semințele să ajungă la maturitate, după care le-a cules și contabilizat ceea ce a
obținut. Ce fel de boabe credeți că au rezultat?
Mendel a numărat 5474 de boabe NETEDE și 1850 de boabe ZBÂRCITE!!! Varianta „bob zbârcit” a
REAPĂRUT în F2! Pe baza acestor rezultate, Mendel a tras concluzia că determinantul variantei recesive NU DISPARE în
F1 (când nu se mai manifestă), ci rămâne ASCUNS până când revin condițiile favorabile manifestării lui.
Mendel a numit acest lucru SEGREGARE (DISJUNCȚIE) = apariția, în generația F2, din plante hibride F1
(provenite deci din boabe hibride de un singur tip – netede) a celor două tipuri de boabe – netede și zbârcite –
caracteristice nu părinților (F1), ci genitorilor (G), adică „bunicilor” lui F2.
De ce SEGREGARE? Segregare înseamnă împărțire, separare, iar Mendel a folosit acest termen pentru a explica
faptul că s-a plecat de la o singură variantă fenotipică – boabele netede F1 – și acestea au dat în descendență două
variante fenotipice, ca și cum varianta netedă F1 s-a separat/împărțit în cele două variante fenotipice din F2, netedă și
zbârcită.
Mendel a făcut raportul între numărul de boabe netede (variantă DOMINANTĂ) și cel de boabe zbârcite (variantă
RECESIVĂ) obținute în F2: 5474:1850 = 2,9589
Mendel nu s-a mulțumit cu acest rezultat, pregătirea lui de matematician spunându-i că, pentru a putea face o
generalizare, trebuie să obțină mai multe date din experimente similare. Ca urmare, a realizat alte câteva monohibridări în
care a urmărit transmiterea în descendență și a altor caractere, obținând următoarele rezultate:
Tabelul 2. Rezultatele obținute de Mendel în experimentele de monohibridare
Caracter Variante fenotipice alelomorfe Raport
D/r DOMINANTĂ recesivă Talia plantei Înaltă 787 Pitică 277 2,8411
Culoarea cotiledoanelor (bobului) Galbenă 6022 Verde 2001 3,0094
Forma seminței (bobului) Netedă 5474 Zbârcită 1850 2,9589
Culoarea florii (culoarea tegumentului seminal) Roșie (cenușie) 705 Albă (albă) 224 3,1473
Culoarea fructului (păstăii) Verde 428 Galbenă 152 2,8158
Forma fructului (păstăii) Dreaptă 882 Strangulată 299 2,9498
Poziția florilor pe tulpină Axilară 651 Axială (terminală) 207 3,1449
TOTAL 14949 5010 2,9838
Pe baza acestor rezultate, Mendel a observat că în F2 raportul cantitativ între varianta dominantă și cea recesivă
este de aproximativ 3/1 (3D/1r, 3 dominant la 1 recesiv).
Astfel, Mendel enunță a doua lege a eredității – LEGEA SEGREGĂRII FENOTIPICE, ÎN
MONOHIBRIDARE, ÎN GENERAȚIA A DOUA, ÎN RAPORT DE 3 DOMINANT LA 1 RECESIV.
Cum a explicat Mendel rezultatele obținute Abordarea revoluționară a lui Mendel pentru explicarea fenomenelor observate a constat în: (1) introducerea
discontinuității în urmărirea transmiterii caracterelor (urmărire analitică – doar un caracter, apoi două.... ); (2) folosirea și
prelucrarea populațiilor de organisme și nu a indivizilor izolați => interpretare statistică; (3) modelarea – introducerea
de simboluri simple pentru desemnarea determinanților de caractere, numiți de el factori ereditari corpusculari – „A”
pentru factorul ereditar dominant, „a” pentru factorul ereditar recesiv.

77 / 108
Pentru a explica rezultatele obținute și pentru a putea trage concluzii de maximă generalitate, cu valoare de legi,
Mendel a plecat de la mai multe premise/ipoteze:
1. Mendel postulează că fiecare trăsătură a unui organism este controlată/determinată de câte o pereche de
factori ereditari corpusculari discreți, distincți din punct de vedere fizico-chimic. Mendel contrazice astfel
concepția anaxagoreană14
de până atunci conform căreia transmiterea caracterelor ereditare se face DIRECT și-i
dovedește falsitatea, dovedind că transmiterea caracterelor ereditare se face INDIRECT – prin intermediul
factorilor ereditari corpusculari.
2. Factorii ereditari se găsesc în nucleul tuturor celulelor. În celulele somatice (ale corpului) există câte o pereche de
factori ereditari pentru fiecare trăsătură (DE CE? – orice organism care se reproduce sexuat provine din doi
genitori care participă în mod egal la realizarea descendentului). În gameți există doar un factor ereditar din
fiecare pereche (adică gameții sunt întotdeauna puri din punct de vedere genetic). Ca urmare, Mendel a
simbolizat cu AA soiul pur cu boabe netede și cu aa soiul pur cu boabe zbârcite, adică cele două soiuri genitoare
pe care a experimentat, iar gameții produși de acestea – A, respectiv a.
3. La formarea gameților (din celulele-mamă de gameți, care conțin perechi de factori ereditari), factorii ereditari din
fiecare pereche se separă în gameți diferiți => fiecare gamet are doar câte un factor ereditar din fiecare
pereche. 4. În fecundație se realizează unirea pe bază de hazard/probabilitate a gameților de sex opus; teoretic, oricare
gamet de un sex are șanse egale de a se uni cu oricare gamet de sex opus, dar odată fecundația realizată, dispare
orice posibilitate ca acei gameți să mai participe la altă fecundație.
Mendel a simbolizat soiul pur cu boabe netede AA, iar pe cel cu boabe zbârcite aa. Unul dintre ele va reprezenta
sexul masculin (AA), iar celălalt – sexul feminin (aa). A realizat și încrucișarea reciprocă (masculin aa și feminin AA), dar
a obținut aceleași rezultate, deci transmiterea caracterului „forma bobului” nu este influențată de sex.
În cazul plantelor din soiul pur AA, în toate celulele somatice ale acestora există perechea de factori AA,
responsabilă de determinarea variantei fenotipice dominante „bob neted”; în celulele-mamă de gameți (cmg, situate în ovul
și în antere) se găsește aceeași pereche AA.
Aceste celule-mamă de gameți (cmg) suferă meioză (R!) și formează gameți15
care conțin doar UN SINGUR
FACTOR, A, din perechea AA prezentă în celula-mamă, deoarece această pereche se împarte/segregă în gameți diferiți.
Deoarece cei doi factori ai perechii sunt identici (AA reprezintă genotip homozigot), va rezulta un singur tip de gameți –
gameți cu A (chiar dacă un gamet primește un factor din pereche, iar un alt gamet, pe celălalt, factorii sunt identici și deci
nu există diferențe între gameți cu privire la factorul care determină forma bobului).
În cazul plantelor din soiul pur aa, judecata este identică și deci acestea vor forma, de asemenea, un singur tip de
gameți – gameți cu a.
Consecutiv polenizării încrucișate realizate de Mendel în primul an de experimente, la fecundație vor participa
gameți conținând factori ereditari diferiți, A și a, ambii responsabili de același caracter, forma bobului, dar de variante
fenotipice diferite, alelomorfe – netedă și zbârcită. În zigot (prima celulă a noii generații, F1), cei doi factori din gameți
reformează perechea caracteristică celulelelor somatice, responsabilă de caracterul „forma bobului”, dar această pereche
este heterozigotă – conține doi factori diferiți, Aa, din care DOAR UNUL SINGUR, cel DOMINANT A, se va manifesta,
determinând varianta netedă a formei bobului pentru toate boabele rezultate – deci generația F1 este omogenă/uniformă
fenotipic, dar și genotipic, toți indivizii conținând perechea de factori Aa. Celălalt factor, a, este prezent, dar
nemanifest/inactiv în prezența celui dominant și de aceea a fost numit RECESIV.
Mendel a simbolizat cu o singură literă caracterul analizat și a folosit majuscula pentru varianta fenotipică
dominantă, respectiv pentru factorul ereditar care a determinat-o, și minuscula pentru varianta fenotipică recesivă (aceea
care nu mai apărut în F1), respectiv pentru factorul ereditar determinant. Folosind aceste simboluri, Mendel și-a transpus
observațiile într-o schemă de hibridare, prezentată mai jos.
Mendel a continuat experimentul și anul următor, pornind de la boabele hibride netede care reprezintă F1 și pe
care le-a însămânțat, iar acestea au germinat, apărând plantule care au crescut și s-au dezvoltat, rezultând, în final,
generația F1 de plante hibride. Aceste plante au fost simbolizate de Mendel tot Aa, provenind din boabe Aa; în florile lor,
la nivelul structurilor de reproducere, există celule-mamă de gameți ♂ (în anterele staminelor) și celule-mamă de gameți ♀
(în ovulele din ovarul gineceului), toate conținând perechea de factori Aa responsabilă de forma bobului.
Celulele-mamă de gameți vor suferi meioză (R!) și vor forma gameți (!Vezi mai sus!). La formarea gameților,
perechea Aa din celula-mamă de gameți se împarte în gameți diferiți, deci vor rezulta două tipuri de gameți, unii cu A și
ceilalți cu a, în proporții egale.
Astfel, în aceeași floare, deoarece Mendel nu a mai intervenit în desfășurarea reproducerii sexuate, se formează
două tipuri de gameți ♂, jumătate cu A și cealaltă jumătate cu a, și două tipuri de gameți ♀, jumătate cu A și cealaltă
jumătate cu a. Acești gameți pot participa la autofecundație, proces care se realizează probabilistic – oricare gamet de un
sex are șanse egale de a se uni cu oricare gamet de sex opus.
În schema de hibridare se figurează fecundațiile posibile/potențiale, nu cele efective, și de aceea rezultatele au
caracter și valoare statistică.
În urma autofecundației rezultă generația a doua, F2, reprezentată inițial de zigoți, apoi de boabe/semințe în
păstăile formate pe plantele din F1. Prin unirea probabilistică a celor celor două tipuri de gameți ♂ cu cele două tipuri de
14 Anaxagora din Clazomenae (Asia Mică), cca. 500-428 î.e.n., filosof grec presocratic, membru al Şcolii Ioniene de filosofie 15 Procesul de formare a gameţilor la plante – gametogeneza – este puţin mai complicat, deoarece prin meioză, la plante, se formează spori care sunt
haploizi şi se divid mitotic rezultând structuri pluricelulare, gametofiţi, din ale căror celule se vor diferenţia gametangi cu gameţi. Aşadar, până la gameţi,
se petrec sporogeneza, gametofitogeneza şi gametangiogeneza.

78 / 108
gameți ♀, rezultă patru variante posibile de perechi de factori ereditari – AA, Aa, Aa și aa, dintre care primele trei vor
determina fenotipul dominant „bob neted” (simbolizat A), iar ultima va determina fenotipul recesiv „bob zbârcit”
(simbolizat a).
Astfel, din totalul boabelor F2 numărate de Mendel, 5474, adică ¾, au fost netede, iar 1850, adică ¼, au fost
zbârcite, rezultând un raport de 3D:1r (3 dominant la 1 recesiv sau, exprimat procentual, 75% dominant la 25% recesiv).
Acesta este raportul de segregare după fenotip.
Din punct de vedere genotipic, generația hibridă F1 este, de asemenea, uniformă/omogenă, fiind reprezentată doar
de genotipul heterozigot Aa; acesta SEGREGĂ în descendență rezultând cele trei genotipuri diferite ale generației F2 –
AA (homozigot dominant), Aa (heterozigot) și aa (homozigot recesiv), în raport de 1:2:1 (raport de segregare după genotip,
exprimat și procentual 25% AA : 50% Aa : 25% aa sau ¼ AA: 2/4 Aa : ¼ aa).
La prima vedere, cele două raporturi de segregare, fenotipică și genotipică, sunt total diferite (fenotipic – 3:1,
genotipic – 1:2:1), dar, deoarece, în condiție heterozigotă, factorul A se manifestă total dominant față de factorul a,
nepermițând acestuia să se manifeste în fenotip, se poate stabili corespondența între cele două raporturi. Genotipurile AA și
Aa reprezintă împreună ¾ din totalul genotipurilor rezultate în F2 și ambele determină același fenotip dominant A, „bob
neted”, exact ca și proporția obținută în segregarea fenotipică.
Se constată că fenotipului recesiv a, „bob zbârcit”, îi corespunde un singur genotip posibil, aa (ceea ce implică
faptul că, știind că un anumit fenotip este recesiv, putem scrie cu siguranță genotipul care îl determină), iar fenotipul
dominant A, „bob neted”, poate fi determinat de două genotipuri diferite, AA și Aa.
Pentru un individ dat care prezintă fenotipul dominant, nu se poate ști cu certitudine dacă are genotip AA
(homozigot dominant) sau Aa (heterozigot), dar se poate stabili, fie prin observarea descendenței (dacă nu segregă, este soi
pur, deci homozigot; dacă segregă, este soi impur, hibrid, deci heterozigot), fie prin experimente de retroîncrucișare
(backcross) sau de test cross.
Experimentul de retroîncrucișare, care presupune încrucișarea fie a unui individ cu fenotip dominant, dar cu
genotip necunoscut, fie a unui hibrid cu unul dintre părinți sau cu un individ cu genotip identic cu al unuia dintre părinți,
este utilizat în două scopuri:
a. Determinarea genotipului unui individ cu fenotip dominant, caz în care este sinonim cu test cross.
b. Obținerea de descendenți care să semene mai mult cu unul dintre părinți.
Experimentul test cross presupune încrucișarea unui individ cu fenotip dominant și genotip necunoscut (AA sau
Aa) cu un individ cu genotip cunoscut, de preferat homozigot recesiv aa sau heterozigot Aa. Folosindu-se un genotip în
care există cel puțin un factor recesiv a, poate fi pusă în evidență existența un eventual factor recesiv în genotipul
necunoscut; în cazul utilizării unui genotip cunoscut AA, toți descendenți vor manifesta fenotipul dominant și nu se va
putea stabili dacă genotipul testat conține vreun factor recesiv.
Dacă individul cu genotip cunoscut este unul dintre părinți sau are genotip identic cu al unuia dintre părinții
individului al cărui genotip vrem să-l stabilim, atunci experimentul se mai numește retroîncrucișare sau backcross (vezi
mai sus).
În urma analizei descendenței se poate stabili dacă genotipul verificat este homozigot sau heterozigot.
EXPLICAȚI.
Anul al III-lea
Prin însămânțarea și germinarea boabelor F2 (netede și zbârcite), rezultă populația de plante F2. Acestea s-au
autopolenizat și autofecundat, rezultând, în final, semințele care reprezintă stadiul inițial al generației F3.
Astfel, din semințele zbârcite F2 (care reprezintă 25% din totalul de semințe din F2), despre care știm că au
genotip homozigot aa, în anul al III-lea vor rezulta plante care vor produce NUMAI semințe zbârcite (F3), fiind soi pur.
Din cele 25% semințe netede F2 cu genotip homozigot AA, în anul al III-lea vor rezulta plante care vor produce
NUMAI semințe netede (F3), fiind, de asemenea, soi pur.
Din cele 50% semințe netede F2 cu genotip heterozigot Aa, în anul al III-lea vor rezulta plante care vor produce
două tipuri de semințe F3 – 75% netede și 25% zbârcite, adică se petrece segregare fenotipică mendeliană. Dintre cele 75%
semințe F3 netede, 25% sunt homozigote AA și 50% sunt heterozigote Aa.
Astfel, prin raportare la procentele din F2, în F3 vor fi contabilizate:
37,5% organisme AA – 25% provenite direct din cele 25% AA din F2, la care se adaugă 25% AA rezultate prin
segregare din cele 50% Aa din F2, adică 25/100 x 50/100 = ¼ x ½ = 1/8 =12,5%. 25% + 12,5% = 37,5%
organisme pure AA.
37,5% organisme aa – 25% provenite direct din cele 25% aa din F2, la care se adaugă 25% aa rezultate prin
segregare din cele 50% Aa din F2, adică 25/100 x 50/100 = ¼ x ½ = 1/8 =12,5%. 25% + 12,5% = 37,5%
organisme pure aa.
25% organisme Aa – 50% din cele 50% Aa din F2, adică ½ x ½ = ¼ = 25% organisme hibride Aa.
Se constată că procentul de organisme heterozigote în F3 a scăzut la jumătate față de F2.
Anul al IV-lea, al V-lea....
În F4, numai cele 25% organisme Aa vor segrega, rezultând 12,5% organisme pure AA și aa (adică 50% din 25%)
și 12,5% organisme hibride Aa (adică 50% din 25%). În F5 va scădea și mai mult procentul de organisme heterozigote Aa –
6,25%, în F6 – 3,125% etc., înjumătățindu-se cu fiecare generație, tinzând la zero după un număr mare de generații. (Vezi
Importanța legilor lui Mendel)

79 / 108
g ♂
g ♀ A a
A AA Aa
a Aa aa
AA aa
AA AA aa aa
A A a a
cmg ♀
R!
cmg ♀ cmg ♂ cmg ♂
R! R! R!
g ♀ g ♀ g ♂ g ♂
Castrare
G: x
A
Polenizare
încrucişată
Aa
F
F1:
cmg ♀ cmg ♂
R! g ♀
A a
Aa Aa
A a
R!
Aa
g ♀ g ♂ g ♂
AA Aa Aa aa F2:
Boabe
netede
Boabe
netede
Boabe
netede
Boabe
zbârcite
100% boabe netede hibride, heterozigote –
uniformitate fenotipică şi genotipică
Plante hibride An II
An I
Semănare, germinare, creştere, dezvoltare
Segregare
genotipică
Segregare
fenotipică
¼ (25%) ¼ (25%)
¼ (25%) r ¾ (75%) D
2/4 (50%)

80 / 108
DIHIBRIDAREA ȘI LEGEA A TREIA A EREDITĂȚII
Mendel nu s-a oprit cu experimentele după ce a constatat și legiferat modul în care se transmite în descendență un
singur caracter. S-a gândit să analizeze transmiterea ereditară simultană a două caractere, cum ar fi forma și culoarea
seminței, prin dihibridare.
Dihibridarea reprezintă încrucișarea a două organisme care se deosebesc prin două caractere (sau, cum se mai
spune în mod eronat, prin două perechi de caractere).
În acest scop a folosit două soiuri pure pentru cele două caractere, unul cu semințe netede A și galbene B (variante
dominante ale celor două caractere, fapt stabilit prin monohibridări), fenotip simbolizat AB și cu genotip dublu-homozigot
dominant AABB, și celălalt cu semințe zbârcite a și verzi b (variante recesive), fenotip simbolizat ab și cu genotip dublu-
homozigot recesiv aabb.
Anul I
Mendel a procedat ca și în cazul monohibridării: a castrat genitorul aabb înainte de autopolenizare și astfel acesta
a participat la hibridare în calitate de sex femel; a adus polen de la genitorul AABB, care deci a participat la hibridare în
calitate de sex mascul.
Individul mascul AABB va produce la nivelul granulelor sale de polen un singur tip de gameți (spermatii), având
genotipul AB, iar individul femel va produce, de asemenea, la nivelul ovulelor din ovarului gineceului, un singur tip de
gameți (oosfere) – cu genotipul ab. În urma fecundației la care participă cele două tipuri de gameți produși de genitori,
rezultă F1, reprezentată de zigoți, apoi de embrioni în semințele din păstăile formate pe genitorul femel aabb. Aceste
semințe, care reprezintă stadiul inițial al generației dublu-hibride F1, sunt toate netede și galbene (fenotip AB), dar cu
genotip dublu-heterozigot AaBb.
În urma fecundației dintre gameții masculin AB și feminin ab, în zigot se refac perechile de factori ereditari
caracteristice celulelor somatice, dar în ambele perechi un factor este dominant – cel de la tată – și unul este recesiv – cel de
la mamă, adică perechile sunt heterozigote (Aa, respectiv Bb).
Se constată că F1 este uniformă, atât din punct de vedere fenotipic AB, cât și din punct de vedere genotipic AaBb,
deci legea I a eredității este valabilă și în cazul dihibridării.
Mendel păstrează semințele dublu-hibride netede și galbene (AB) pentru a le semăna anul următor în scopul
obținerii generației a doua, F2.
Anul al II-lea
Mendel însămânțează boabele dublu-hibride netede și galbene, cu genotip AaBb, reprezentând F1, iar acestea
germinează, formând plantule care cresc și se dezvoltă rezultând o populație de plante F1 dublu-hibride.
În perioada mai-iunie, plantele au făcut flori; de data aceasta, Mendel nu a mai intervenit în desfășurarea
reproducerii, iar în flori s-a petrecut autopolenizare, urmată de autofecundație (aceeași floare a produs și gameții masculini
și gameții feminini).
După fecundație s-au format zigoții – primele celule ale generației a doua, F2 – care se divid mitotic formând
embrioni incluși în semințe. Aceste semințe reprezintă stadiul inițial al generației F2 și se formează în păstăile rezultate din
florile plantelor hibride F1.
Mendel a așteptat ca fructele și semințele să ajungă la maturitate, după care le-a cules și a contabilizat ceea ce a
obținut. Ce fel de boabe credeți că au rezultat?
La nivelul anterelor florilor plantelor hibride F1 se găsesc celulele-mamă de gameți masculini, care au genotipul
AaBb (precum toate celulele somatice). Din aceste cmg ♂ se vor forma, prin meioză (R!), patru tipuri de gameți
masculini – AB, Ab, aB și ab. Formarea acestora are la bază LEGEA SEGREGĂRII INDEPENDENTE A
PERECHILOR DE FACTORI EREDITARI DIN CELULELE-MAMĂ DE GAMEȚI LA FORMAREA
GAMEȚILOR – fiecare pereche de factori ereditari din celula-mamă de gameți se împarte în gameți diferiți independent
de celelalte perechi.
În mod obligatoriu, în gameți ajunge câte un factor din fiecare pereche prezentă în celula-mamă, dar nu există
nici o regulă care să stabilească vreo asociere particulară a respectivilor factori. Astfel, în cazul celulei-mamă de gameți
masculini cu genotipul AaBb, perechea Aa se va împărți în gameți diferiți, deci jumătate din numărul total vor conține
factorul A, iar cealaltă jumătate – factorul a; dar, deoarece în celula-mamă există două perechi de factori, gameții trebuie să
conțină un factor și din perechea a doua, Bb, care, de asemenea, se va împărți în gameți diferiți: jumătate cu B și cealaltă
jumătate cu b.
Deci cele două perechi de factori segregă independent una de cealaltă și astfel, în gametul care a primit factorul A
din perechea Aa, poate să ajungă, cu aceeași probabilitate, factorul B (în acest caz gametul având genotipul AB) sau
factorul b (în acest caz gametul având genotipul Ab) din perechea Bb. În mod similar se formează gameții aB și ab.
Gameții AB și ab se numesc gameți parentali, iar Ab și aB – gameți recombinați.
Aceeași judecată se aplică și în cazul formării celor patru tipuri de gameți feminini – AB, Ab, aB, ab – la nivelul
ovulelor din gineceu.
Așadar, în aceeași floare a plantelor F1 hibride se formează patru tipuri de gameți masculini și patru tipuri de
gameți feminini, care au șanse egale de a participa la autofecundație, rezultând deci 16 combinații posibile de zigoți
(respectiv semințe), adică diversitatea generației F2.
Rezultatul fecundațiilor posibile poate fi reprezentat printr-o schemă de hibridare sau, mai simplu, cu ajutorul
pătratului lui Punnett – un tabel în care se trec, pe prima coloană, respectiv pe prima linie, genotipurile celor două tipuri
de gameți – masculini, respectiv feminini – și în celelalte căsuțe genotipurile și fenotipurile posibile rezultate prin
fecundație.

81 / 108
Pătratul lui Punnett16
g ♂
g♀ AB Ab aB ab
AB AABB
AB
AABb
AB
AaBB
AB
AaBb
AB
Ab AABb
AB
AAbb
Ab
AaBb
AB
Aabb
Ab
aB AaBB
AB
AaBb
AB
aaBB
aB
aaBb
aB
ab AaBb
AB
Aabb
Ab
aaBb
aB
aabb
ab
În urma contabilizării semințelor F2, Mendel a obținut următoarele rezultate: 315 semințe17
netede și galbene AB,
108 semințe netede și verzi Ab, 101 semințe zbârcite și galbene aB și 32 semințe zbârcite și verzi ab, în total 556 semințe.
16 Reginald Crundall Punnett (1875-1967) – genetician britanic, cofondator în 1910 al Jurnalului de Genetică, alături de William Bateson. Pătratul lui
Punnett este o diagramă cu ajutorul căreia poate fi prezisă descendenţa posibilă a unei încrucişări. 17 În unele lucrări de specialitate se consideră că ar fi vorba de indivizi (plante), nu de semințe.
AABB aabb
AABB AABB aabb aabb
AB AB ab ab
cmg ♀
R!
cmg ♀ cmg ♂ cmg ♂
R! R! R!
g ♀ g ♀ g ♂ g ♂
Castrare
G: x
AB
Polenizare
încrucişată
AaBb
F
F1:
cmg ♀ cmg ♂
R!
g ♀
Ab ab
AaBb AaBb
Ab ab
R!
AaBb
g ♂
AABB aaBb F2:
100% boabe netede şi galbene, dublu-hibride,
dublu-heterozigote – uniformitate fenotipică şi
genotipică
Plante hibride An II
An I
Semănare, germinare, creştere, dezvoltare
Continuare – vezi Pătratul lui Punnett
AB aB aB AB

82 / 108
Dacă analizăm această descendență separat pentru fiecare caracter se observă că se respectă legea a II-a eredității:
pentru forma bobului au rezultat 423 (315 + 108) semințe netede A și 133 (101 + 32) semințe zbârcite a, adică
în raport fenotipic de 3A:1a.
pentru culoarea bobului au rezultat 416 (315 +
101) semințe galbene B și 140 (108 + 32) semințe
verzi b, adică în raport fenotipic 3B:1b.
În urma analizei globale a descendenței din F2 se
constată că rezultă patru variante fenotipice: semințe netede
și galbene AB, semințe netede și verzi Ab, semințe zbârcite și
galbene aB și semințe zbârcite și verzi ab; dintre acestea, două
sunt variantele prezente la genitori (variante parentale – AB,
respectiv ab), iar celelalte două sunt variante recombinate între
cele parentale (o variantă fenotipică a unui caracter de la un
părinte și o variantă fenotipică a celuilalt caracter de la celălalt
părinte) – semințe netede și verzi Ab și semințe zbârcite și
galbene aB.
Raportul se segregare fenotipică pentru cele patru
variante este 9/16 AB : 3/16 Ab : 3/16 aB : 1/16 ab, adică
9:3:3:1. Cum s-a obținut acest raport?
Prin unirea probabilistică a celor patru tipuri de
gameți masculini cu cele patru tipuri de gameți feminini rezultă
16 combinații posibile de factori ereditari, adică 16 genotipuri,
distribuite după cum urmează, în funcție de relația dominanță-
recesivitate din cadrul fiecărei perechi de factori:
9 (din 16) sunt de tip A_B_ (AABB, AABb, AaBB, AaBb) – corespunde fenotipului AB
3 (din 16) sunt de tip A_bb (AAbb, Aabb) – corespunde fenotipului Ab
3 (din 16) sunt de tip aaB_ (aaBB, aaBb) – corespunde fenotipului aB
1 (din 16) este de tip aabb – corespunde fenotipului ab
Ca urmare a diversității mari de genotipuri, în acest caz nu mai poate fi scris și un raport de segregare după
genotip care să aibă relevanță statistică.
Explicarea rezultatelor Rezultatele obținute de Mendel în dihibridare nu pot fi explicate decât dacă se acceptă faptul că, la formarea
gameților prin meioză, perechile de factori ereditari responsabile de determinarea celor două caractere analizate – forma și
culoarea seminței – segregă independent una de cealaltă în gameți diferiți, rezultând astfel, în proporții egale, cele patru
tipuri de gameți masculini, respectiv feminini – AB, Ab, aB, ab.
Segregarea perechilor de factori ereditari se corelează cu o segregare a caracterelor și o combinare a
variantelor fenotipice ale acestora. Astfel, dacă analizăm separat cele două caractere urmărite de Mendel, caracterul
„forma seminței”, reprezentat la F1 doar prin varianta netedă (dominantă), se împarte în F2 în cele două variante ale
genitorilor – netedă și zbârcită; caracterul „culoarea seminței”, reprezentat la F1 doar prin varianta galbenă (dominantă), se
împarte în F2 în cele două variante ale genitorilor – galbenă și verde.

83 / 108
Dar cele două caractere au fost analizate simultan, astfel se constată că variantele dominante din F1 ale celor două
caractere urmărite, în F2 au segregat și, în același timp, s-au combinat în patru variante fenotipice – AB, Ab, aB, ab – dintre
care numai una (AB) corespunde generației F1, respectiv două (AB și ab) corespund genitorilor.
Cunoscând comportamentul cromosomilor în meioză, se poate explica cum și de ce se realizează disjuncția
independentă a perechilor de factori Aa și Bb. Această segregare este posibilă (și cu raportul reieșit din experiment)
deoarece perechile de factori implicate sunt plasate în perechi diferite de cromosomi omologi (A și a sunt într-o pereche,
iar B și b sunt într-o altă pereche de omologi).
Stabilirea numărului și genotipurilor gameților Un organism homozigot de tip AA, BB, aa, bb etc. produce, prin meioză, un singur tip de gameți – A, B, a, b etc.
Un organism dublu-homozigot de tip AABB, AAbb, aaBB, aabb, AABBCC, AAbbCC etc. produce, de
asemenea, un singur tip de gameți – AB, Ab, aB, ab, ABC, AbC etc., conform legii a III-a a eredității (gameții conțin câte
un factor ereditar din fiecare pereche de factori din celulele-mamă de gameți).
Un organism simplu-heterozigot de tip Aa sau Bb etc. produce două tipuri de gameți – A și a, respectiv B și b etc.
Un organism dublu-hetrozigot de tip AaBb produce patru tipuri de gameți – AB, Ab, aB, ab.
Un organism de tipul AaBB sau AABb va produce două tipuri de gameți – AB și aB, respectiv AB și Ab.
Un organism triplu-heterozigot AaBbCc va produce opt tipuri de gameți – ABC, ABc, AbC, Abc, aBC, aBc,
abC, abc.
Un organism de tipul AaBBCC va produce două tipuri de gameți – ABC și aBC.
Un organism de tipul AABbCc va produce patru tipuri de gameți – ABC, ABc, AbC, Abc.
Ați identificat regula matematică prin care se poate stabili numărul de tipuri de gameți pe care îi produce un
organism cu un anumit genotip?
Pentru scrierea corectă a tuturor tipurilor de gameți se folosește metoda dichotomiei.
Exemplu. Care sunt gameții formați de un organism triplu-heterozigot AaBbCc? Dar AaBBCcDD?
A
B C → ABC
A B
C D → ABCD c → ABc
b C → AbC
c D → ABcD c → Abc
a
B C → aBC
a B
C D → aBCD c → aBc
b C → abC
c D → aBcD c → abc
Legile probabilităților și interpretarea matematică a rezultatelor lui Mendel 1. Probabilitatea de manifestare/apariție a unui anumit eveniment este egală cu raportul dintre numărul de
cazuri favorabile și numărul de cazuri posibile.
Exemplul 1. Care este probabilitatea ca, aruncând un zar, să cadă cu numărul 1 în sus? Există șase cazuri posibile,
zarul putând să cadă cu oricare din cele șase fețe în sus, dar numai una singură este favorabilă => probabilitatea
evenimentului este 1/6.
Exemplul 2. Care este probabilitatea ca într-o monohibridare, în F2, să rezulte un organism cu fenotip dominant?
Există patru cazuri posibile de genotipuri (responsabile de fenotipuri) – AA, Aa, Aa și aa – și trei dintre acestea
determină un fenotip dominant – cazul favorabil => probabilitatea evenimentului este ¾.
Se poate face astfel o generalizare – întotdeauna probabilitatea de apariție a unui fenotip dominant în F2 în
monohibridare este ¾, iar a unui fenotip recesiv este ¼.
2. Probabilitatea apariției/manifestării simultane a două evenimente distincte/independente este egală cu
produsul dintre probabilitățile lor de apariție/manifestare separată.
Exemplul 1. Care este probabilitatea ca, aruncând două zaruri, să cadă ambele cu numărul 1 în sus? Pentru fiecare
zar separat probabilitatea evenimentului este 1/6, iar pentru manifestarea simultană probabilitatea este de 1/36, adică 1/6 x
1/6.
Exemplul 2. Care va fi probabilitatea apariției unui fenotip de tipul AB, PAB? Fenotipul A independent are
probabilitatea de apariție PA = ¾, iar fenotipul B independent are aceeași probabilitate de apariție PB = ¾. Probabilitatea de
apariție simultană a lui A și B este dată de produsul dintre probabilitățile de apariție separată a celor două fenotipuri, deci
PAB = PA x PB = ¾ x ¾ = 9/16.
În mod similar se calculează probabilitatea de apariție a fenotipurilor:
Ab → PAb = PA x Pb = ¾ x ¼ = 3/16
aB → PaB = Pa x PB = ¼ x ¾ = 3/16
ab → Pab = Pa x Pb = ¼ x ¼ = 1/16.
Vă duc cu gândul la ceva aceste rezultate? Calculați probabilitatea de apariție a fenotipurilor determinate de
următoarele genotipuri: AabbCcdd, aaBbCCDdee, aabbcc, AaBbCc.

84 / 108
Legile lui Mendel (principiile eredității) 1. Legea uniformității hibrizilor în prima generație
2. Legea segregării fenotipice, în F2, în monohibridare, în raport de 3 dominant la 1 recesiv
3. Legea segregării independente a perechilor de factori ereditari la formarea gameților și obținerea în F2, în
dihibridare, a unui raport de segregare fenotipică de 9:3:3:1
***În unele lucrări de specialitate sunt enunțate doar două legi ale lui Mendel și nu trei, deoarece primele sale două legi sunt
reunite în una singură – LEGEA PURITĂȚII GAMEȚILOR.
Legile lui Mendel (principiile eredității) – variantă alternativă 1. Legea purității gameților – gameții sunt întotdeauna puri din punct de vedere genetic deoarece conțin doar un
singur factor ereditar din fiecare pereche de factori din celula-mamă de gameți
2. Legea segregării independente a perechilor de factori ereditari la formarea gameților și obținerea în F2, în
dihibridare, a unui raport de segregare fenotipică de 9:3:3:1
IMPORTANȚA LEGILOR LUI MENDEL*
Importanță teoretică
Elaborarea primelor legi ale eredității a însemnat o etapă foarte importantă în cunoașterea mecanismelor prin care
se asigură transmiterea caracterelor în descendență de-a lungul generațiilor și cum se realizează segregarea caracterelor la
hibrizi.
Marea importanță a legilor mendeliene constă, în primul rând, în demonstrarea faptului că transmiterea ereditară a
caracterelor se realizează prin intermediul unor factori ereditari corpusculari, prezenți în toate celulele organismului. Prin
combinarea probabilistică a acestor factori, de origine maternă și paternă, rezultă segregarea caracterelor în descendență.
Mendel a fost primul cercetător care și-a imaginat existența atât în celulele somatice (sub formă de pereche), cât și
în celulele sexuale (în formă simplă) a unor particule materiale, numite de el factori ereditari, răspunzătoare de
transmiterea caracterelor ereditare la descendenți.
Cercetări ulterioare efectuate la nivel celular au demonstrat că celulele somatice au un număr dublu de cromosomi,
2n, comparativ cu cele sexuale, n. Genele dispuse în cromosomii omologi se recombină în cazul hibridării sexuate pe baza
legilor lui Mendel. În felul acesta s-a demonstrat că factorii ereditari mendelieni au existență reală, materială, ei fiind situați
în cromosomi și prezentând independență în procesul de recombinare (este vorba de recombinarea intercromosomală).
Prin aceste descoperiri sunt infirmate ipotezele și teoriile transmiterii directe a caracterelor – teoria pangenezei (C.
Darwin), teoria plastidulelor (E. Haeckel), teoria pangenelor (H. de Vries), teoria plasmei germinale (A. Weismann), teoria
micelară (K. W. Nageli)
În al doilea rând, studiile lui Mendel au dovedit că factorii ereditari recesivi pot să nu se manifeste la descendenți,
rămânând în stare ascunsă. Aceasta înseamnă că transmiterea factorilor ereditari nu este afectată de prezența caracterului
respectiv, ei moștenindu-se la urmași numai pe baza combinării lor probabilistice.
Legile mendeliene constituie baza înțelegerii modului în care se realizează combinarea probabilistică a genelor și
transmiterea lor de la o celulă la celulele-fiice și astfel de la o generație la alta și, implicit, baza teoretică și practică a
cercetărilor de ameliorare a plantelor și animalelor
Importanță practică
Cunoașterea legilor mendeliene a dat posibilitatea realizării de organisme hibride care prezintă noi combinații de
gene, diferite de cele ale genitorilor.
Prin încrucișarea unor organisme homozigote, AA x aa, prima generație este heterozigotă Aa în proporție de
100%, iar în F2 se produce segregarea, astfel că 50% dintre descendenți sunt homozigoți (AA și aa) și 50% sunt
heterozigoți. În generația a treia F3, obținută prin autofecundație, numai 25% dintre descendenți sunt heterozigoți Aa; în
generația a patra F4, obținută tot prin autofecundație, numai 12,5% dintre descendenți etc. Aceasta înseamnă că în fiecare
generație de după hibridare crește frecvența organismelor homozigote și se reduce cea a organismelor heterozigote/hibride.
Cunoașterea acestui fenomen prezintă importanță practică în ameliorare deoarece soiurile de plante și rasele de
animale trebuie să prezinte un anumit grad de homozigoție care le dă posibilitatea să-și transmită caracterele cât mai fidel la
urmași. Ce implicație are acest lucru asupra heterozisului?
De asemenea, prin cunoașterea modului în care se combină și segregă caracterele la hibrizi, se pot realiza noi
combinații de gene, utile pentru practică. Astfel, dacă se hibridează două linii homozigote care se deosebesc prin două
caractere (AABB x aabb), în F1 descendența va fi dublu heterozigotă AaBb, iar în generațiile următoare se reduce
heterozigoția și va crește gradul de homozigoție. Homozigoții apăruți vor fi de patru tipuri: doi de tip parental (AABB,
aabb) și doi de tip recombinat (Aabb, aaBB). Ca urmare, prin încrucișarea a două linii homozigote, rezultă în cele din urmă
patru linii homozigote, din care jumătate reprezintă noi combinații de gene (linii ameliorate).
Cunoașterea acestui fenomen dă posibilitatea amelioratorului să combine, prin încrucișare, caractere de la două
sau mai multe soiuri/rase.
În afara importanței teoretice, legile lui Mendel prezintă o foarte mare importanță practică. Întreaga activitate de
obținere de noi soiuri de plante sau rase de animale sau ameliorarea celor aflate în producție se bazează pe mecanismul de
transmitere și combinare a genelor la descendenți. În orice experiență de încrucișare prin care se urmărește transferarea
unui caracter avantajos (rezistență la boli, la ger, conținut mare de proteine, glucide, valorificarea nutrețurilor etc.) de la
un/o soi/rasă la altul/alta trebuie să se țină seama de numărul de gene/factori ereditari implicate, de puritatea genetică și de
modul de transmitere a caracterelor – dominant sau recesiv.
În genetica umană, cunoscând modul de transmitere a unor caractere normale sau patologice, se poate interveni
prin sfaturi genetice pentru reducerea frecvenței unor maladii ereditare.
TEST II

85 / 108
ABATERI DE LA SEGREGAREA MENDELIANĂ Dominanță completă sau ereditate de tip Pisum Interacțiunea dintre factori ereditari descoperită și descrisă de J. Mendel (și redescoperită și confirmată ulterior de
C. Correns, E. von Tschermak și H. de Vries) a fost numită dominanță completă deoarece, în condiție heterozigotă,
doar un singur factor ereditar din pereche – cel dominant – funcționează ca determinant genetic (manifestându-se sub forma
unui fenotip), celălalt – cel recesiv – este prezent, dar nemanifest, fiind dominat complet de perechea sa (recesivitate
completă). Deoarece acest model de transmitere în descendență a caracterelor a fost descoperit prin experimente realizate
pe mazăre, se mai numește și ereditate de tip Pisum.
Terminologia introdusă și folosită de către Mendel cu referire la determinanții genetici s-a modificat după 1900.
Conform Mendel, un caracter este determinat de o pereche de factori ereditari, prezentă invariabil la toți indivizii care
prezintă respectivul caracter. Diferitele variante fenotipice ale caracterului sunt determinate de variații ale factorilor
ereditari (A și a) din pereche și de modul în care aceștia se grupează și interacționează.
În prezent, se consideră că, în general, un caracter este determinat de o genă; indivizii care prezintă respectivul
caracter înseamnă că au și gena care îl determină, aceeași genă fiind prezentă la toți. Dar gena există, la fiecare individ în
parte, prin două variante fizice (adică printr-o pereche) ale ei, numite alele, care ocupă același locus în cromosomii
omologi. Alela este echivalentă factorului ereditar, iar gena – perechii de factori responsabile de un anumit caracter. Astfel,
diferitele variante fenotipice ale caracterului sunt determinate de variații ale alelelor din pereche și de modul în care acestea
se grupează și interacționează.
Se consideră că, din punct de vedere evolutiv, un caracter apare la început sub forma unei singure variante
fenotipice prezentă la toți indivizii unei populații/specii și determinată de o genă reprezentată de o singură alelă (A); toți
indivizii populației respective vor avea unica alelă A sub forma perechii de alele AA corespunzătoare genei.
Ulterior, alela A (una sau ambele – mai rar – din pereche) de la unii indivizi va suferi mutații (modificări
structural-funcționale) și va apărea astfel o a doua alelă, a1, în populație, alături de A. Prin fluxul genic din cadrul
reproducerii sexuate, cele două alele (ale aceleiași gene!) existente în populație se vor împerechea în trei moduri posibile
– AA, Aa1, a1a1. În oricare individ al populației poate fi găsit doar una din cele trei perechi posibile (gena este reprezentată
în individ de o pereche de alele).
Dacă se mai produce o altă mutație a alelei A sau a alelei a1, va apărea o a treia alelă în populație, a2, iar numărul
de perechi de alele va crește – AA, Aa1, Aa2, a1a1, a1a2, a2a2 ș.a.m.d.
CALCULAȚI câte perechi de alele vor exista într-o populație după cinci evenimente mutaționale. Scrieți
aceste perechi.
Cu fiecare eveniment mutațional, crește numărul de alele ale aceleiași gene și deci numărul de variante fenotipice
ale aceluiași caracter, determinate, pe de o parte, de alelele distincte și, pe de altă pare, de modul în care acestea se
împerechează și interacționează în cadrul perechii (ex., dacă alela a2 este dominantă față de a1, atunci varianta fenotipică
determinată de a1 va avea șansa să se manifeste doar când a1 este în condiție homozigotă a1a1; în celalalte combinații fie se
manifestă A, fie a2). Important de reținut că toate alelele unei gene afectează unul și același caracter.
De obicei, alela sălbatică sau normală, prima apărută evolutiv, este dominantă față de toate celelalte alele
apărute prin mutație din ea sau unele din altele.
Experimente realizate post-Mendel au evidențiat și alte tipuri de interacțiuni alelice și chiar genice, în care
raporturile de segregare se modifică, aparent sau în mod real, față de cele descoperite de Mendel sub forma dominanței
complete în mono- și dihibridare.

86 / 108
I. INTERACȚIUNI ALELICE Această categorie de abateri de la segregarea mendeliană are la bază variații al interacțiunii între alelele (cel puțin
două) aceleiași gene.
1. Semidominanță (dominanță incompletă, ereditate de tip Zea)* Este interacțiunea alelică în care genotipul heterozigot Aa determină un fenotip intermediar între fenotipurile
parentale (ale genitorilor homozigoți AA, aa), deoarece cele două alele se manifestă una față de alta parțial dominant,
respectiv parțial recesiv (prezența alelei recesive împiedică manifestarea plenară a alelei dominante); de aceea se numește
semidominanță sau dominanță incompletă – dominanță „pe jumătate”.
În acest caz, în F1 rămâne valabilă legea I a lui Mendel, toți hibrizii Aa fiind uniformi fenotipic (dar cu fenotipul
intermediar), iar în F2 raportul de segregare fenotipică va coincide cu cel de segregare genotipică, 1:2:1. Astfel,
fenotipul reflectă clar genotipul, fiecare genotip determinând un alt fenotip.
Se mai numește ereditate de tip Zea deoarece a fost descrisă și la porumb (Zea mays).
Exemple a) Carl Correns, în 1912, a încrucișat un soi de Mirabilis jalapa (barba-împăratului sau frumoasa nopții) cu flori
roșii RR cu unul cu flori albe rr. În F1 au rezultat numai hibrizi Rr cu flori roz (fenotip intermediar între roșu și alb). Prin
autopolenizarea hibrizilor din F1 a rezultat F2 caracterizată de raportul de segregare fenotipică și genotipică 1 Roșu/RR : 2
Roz/Rr : 1 Albe/rr. Rezultate identice se obțin cu Antirrhinum majus (gura leului) – soi cu flori roșii x soi cu flori albe.
b) prin încrucișarea unei găini cu penaj negru AA cu una cu penaj alb aa, în F1 rezultă numai găini hibride Aa cu
penaj albastru-cenușiu (găini de Andaluzia). Prin încrucișarea a două găini de Andaluzia, în F2 rezultă un raport de
segregare genotipică și fenotipică de 1 AA/găini negre : 2 Aa/găini de Andaluzia : 1 aa/ găini albe
c) prin încrucișarea unui soi de porumb cu boabe albastre AA cu un soi cu boabe galbene aa, în F1 rezultă numai
porumb hibrid Aa cu boabe violet. Prin autopolenizarea hibrizilor din F1 rezultă în F2 un raport de segregare genotipică și
fenotipică de 1 AA/boabe albastre : 2 Aa/boabe violet : 1 aa/boabe galbene
d) prin încrucișarea unui individ cu păr cârlionțat (Hcr
Hcr
) cu unul cu păr drept (HstH
st), în F1 rezultă numai
indivizi hibrizi cu păr ondulat (HstH
cr).
e) bas/altistă y1y
1 x tenor/soprană y
2y
2 => bariton/mezzosoprană y
1y
2
REALIZAȚI SCHEMELE ACESTOR ÎNCRUCIȘĂRI

87 / 108

88 / 108
2. Supradominanță (vigoare hibridă, heterozis)* Este interacțiunea alelică în care genotipul heterozigot Aa determină o sporire a însușirilor biologice (vitalitate,
fertilitate, talie, productivitate, vigurozitate, rezistență la boli/dăunători etc.) comparativ cu fenotipurile genitorilor
homozigoți => AA<Aa>aa. Acest fenomen manifestat la indivizii hibrizi din F1 stă la baza vigorii hibride (fenomenul
heterozis).
Acest fenomen este întâlnit la plante și la animale. Poate fi somatic (afectează dimensiunile), reproductiv
(dezvoltarea mai mare a organelor reproducătoare – semințe, fructe), adaptativ (rezistență sporită la condiții de mediu
nefavorabile, boli și dăunători).
Are importanță practică deosebită în ameliorarea speciilor de plante cultivate și animale domestice – floarea
soarelui, secară, stejar, plop, oi, iepuri de casă, găini pentru carne și ouă etc. Ex., creșterea productivității porumbului prin
cultivarea de hibrizi simpli și dubli.
Exemplu La drosofilă, gena care determină culoarea roșie-cărămizie a ochilor este dominantă w
+, iar cea care determină
culoarea albă w este recesivă. La heterozigoți w+w se constată o creștere semnificativă a cantității de pigment față de ambii
părinți homozigoți. (Exemplul este preluat din literatură, dar nu este relevant pentru fenomenul de vigoare hibridă.)
3. Codominanță Este interacțiunea alelică în care genotipul heterozigot Aa determină un fenotip nou față de fenotipurile parentale
(ale genitorilor homozigoți AA, aa), deoarece cele două alele contribuie/se manifestă în mod egal sau aproximativ egal în
determinare fenotipului – colaborează de pe poziții identice sau similare => codominanță.
Exemple a) în populația umană există trei alele responsabile de determinarea grupelor sangvine
în sistem ABO/ABH: LA, L
B și l (denumiri alternative – I
A, I
B, i). Alela l este total recesivă față
de LA și L
B. În tabelul următor sunt prezentate variantele de genotipuri și fenotipurile
determinate:
Fenotipul grup sangvin AB este cu totul nou față de fenotipurile parentale grup
sangvin A și grup sangvin B (deoarece este o variabilă calitativă, nu cantitativă ca în cazul
culorii florilor din semidominanță, care depinde de cantitatea de pigment). În determinarea
fenotipului grup sangvin AB se manifestă în mod egal ambele alele LA și L
B, care funcționează
astfel codominant.
b) anemia falciformă (drepanocitoză, siclemie) este o hemoglobinopatie care se manifestă prin defecte structural-
funcționale ale hemoglobinei. La om, Hb este localizată în hematii (globule roșii), având rolul de transportor de O2 de la
plămâni la țesuturi (și, într-o mai mică măsură, de CO2 de la țesuturi la plămâni).
Hemoglobina este o heteroproteină tetrameră formată, la adult, din patru catene polipeptidice de globină, două α și
două β, și simbolizată în consecință α2β2; fiecare catenă are atașată o grupare prostetică, numită hem, care coordinează
central un ion bivalent de Fe. Hemoglobina normală se notează HbA – cele patru catene polipeptidice sunt normale.
Pentru fiecare din cele două catene diferite din alcătuirea Hb există câte o genă – gena pentru catena α, respectiv
gena pentru catena β. Ca urmare a unei mutații punctiforme a genei care codifică pentru catena β (localizată în cromosomul
11), aminoacidul acid glutamic din poziția a 6-a este înlocuit cu aminoacidul valină; această modificare a catenei β
determină asamblarea unei hemoglobine anormale, HbS, care, în anumite condiții, polimerizează/agregă necovalent, cu
formarea de lanțuri lungi care determină forma de seceră a hematiilor (lat. falx – seceră).
Condiția homozigotă HbSHbS determină anemia falciformă – hematii în formă de seceră, friabile și mai puțin
flexibile/elastice ca urmare a prezenței în hematii a unor lanțuri de Hb anormală (HbS – formată din două catene α și două
catene β modificate); asemenea hematii rezistă în circulație doar 10-20 de zile, după care suferă hemoliză. Maladia este
foarte gravă, determinând moartea indivizilor înainte de 50 de ani?.
Condiția heterozigotă HbAHbS nu determină boala, indivizii fiind doar purtători (boala se manifestă doar în
condiții de hipoxie sau de deshidratare puternică), deoarece cele două alele care codifică pentru catena β funcționează
codominant (50-60% HbA și 40-50% HbS), determinând producerea atât de molecule de hemoglobină normală (HbA), cât
și de hemoglobină anormală (HbS), în proporții aproximativ egale; în aceast caz, moleculele de hemoglobină normală sunt
suficiente, în condiții de viață obișnuite, pentru ca individul să nu manifeste simptome de boală.
Genotip Fenotip
(grup sangvin)
LALA A
LAl A
LBLB B
LBl B
ll O/H
LALB AB

89 / 108
APLICAȚII

90 / 108

91 / 108

92 / 108

93 / 108
4. Gene letale* Este interacțiunea alelică în care un anumit genotip determină moartea indivizilor (are efect letal). Experimentul
clasic prin care a fost evidențiat efectul letal al unor alele în anumite combinații a fost realizat în 1902-1905 de Lucien
Cuénot (1866–1951, biolog francez), care a studiat transmiterea culorii blănii la șoareci. Acesta a stabilit că șoarecii
galbeni sunt întotdeauna heterozigoți.
Cuénot a încrucișat doi șoareci cu blana galbenă și a rezultat o descendență
neuniformă – șoareci galbeni (2/3) și șoareci de altă culoare (albă, 1/3), în raport de 2:1, cu
toate că se aștepta să obțină în descendență o segregare mendeliană: 3 șoareci galbeni la 1
șoarece de altă culoare (alb). Deoarece au segregat în descendență, înseamnă că genitorii
galbeni sunt heterozigoți pentru culoarea blănii. De ce se poate afirma cu certitudine că
ambii genitori sunt heterozigoți și nu doar unul, iar celălalt homozigot? Concluzia a fost că alela care determină culoarea galbenă a blănii este dominantă,
iar șoarecii galbeni sunt întotdeauna heterozigoți, condiția homozigotă fiind letală (fapt
dovedit în 1910 de către William Ernest Castle și Clarence Cook Little: prin sacrificarea
femelelor gestante înainte de naștere au constatat că o parte (¼) din embrionii de culoare
galbenă mor în etape timpurii ale dezvoltării embrio-fetale).
Genele letale pot fi homozigote dominante sau recesive, mai rar heterozigote.
Alte exemple – crapii oglindă, oile Karakul brumării. La porumb și la alte specii de plante se cunosc numeroase
gene care în stare homozigotă determină apariția unor plante albinotice (fără clorofilă), care nu pot supraviețui (gene letale)
deoarece nu pot face fotosinteză sau a unor plante la care există o rată oarecare de supraviețuire (supravețuiesc numai
unele), având o cantitate redusă de clorofilă (gene semiletale).
5. §Polialelie În cazul polialeliei, gena care codifică pentru un anumit caracter este reprezentată în populație de cel puțin trei
alele situate în același locus și care determină variații ale caracterului respectiv, iar în fiecare individ – de o pereche de
alele, identice sau diferite, din cele prezente în populație. În cadrul fiecărei perechi de alele, interacțiunile pot fi de tipul
dominanței complete, dominanței incomplete, supradominanței, codominanței. În general, alela sălbatică este total
dominantă față de toate celelalte.
Exemple a) gena pentru grupele sangvine în sistem ABO/ABH este reprezentată în populație de trei alele (vezi
Codominanța)
b) gena pentru culoarea ochilor la Drosophila este reprezentată în populație de o serie polialelă:
w+→w
co→w
bl→w
e→w
ch→w
h→w
bf→w
t→w
p→w
i→w
c) gena pentru culoarea ochilor la om: Eb/br
→Egr
→Ebl
d) gena pentru culoarea părului la om: MBk
→MBw
→MBd
e) gena pentru forma firului de păr la om: Hst→H
cr→H
wo
f) gena pentru culoarea blănii la rozătoare: C→cch
→ch→c

94 / 108
II. §INTERACȚIUNI GENICE (NON-ALELICE)
Această categorie de abateri de la segregarea mendeliană are la bază variații al interacțiunii între gene diferite,
fiecare cu cel puțin o alelă. Aceste interacțiuni se pot manifesta prin apariția unor fenotipuri intermediare sau noi și/sau prin
modificarea raporturilor de segregare mendeliene.
1. Epistazie (dominantă și recesivă) Este fenomenul de interacțiune între gene distincte (fiecare cu alelele ei) prin care exprimarea unei gene este
inhibată de cealaltă genă. Gena inhibată se numește genă hipostatică, iar gena inhibitoare se numește genă epistatică.
Dacă gena epistatică are efect numai în condiție homozigotă dominantă sau heterozigotă, fenomenul se numește epistazie
dominantă; dacă are efect numai în condiție homozigotă recesivă, fenomenul se numește epistazie recesivă.
Exemple a) se încrucișează două rase de găini cu penaj alb – Wyandotte (ccii) și Leghorn (CCII). În F1 rezultă numai găini
albe, iar prin încrucișarea între ei a hibrizilor din F1, rezultă în F2 un raport de segregare 13 găini albe : 3 găini negre. Gena
C codifică pentru sinteza de pigment (condițiile CC și Cc determină producerea de pigment; cc nu determină sinteza de
pigment); gena I este genă epistatică dominantă (în condiții II și Ii va inhiba expresia genei C, iar în condiție ii nu va avea
efect asupra genei C). În acest caz se constată atât modificarea raportului de segregare, cât și apariția unui fenotip nou. De
unde știm că în acest caz sunt implicate numai două gene, C și I? Realizați schema de încrucișare. b) fenotipul Bombay (fenotipul de grup sangvin ascuns)
c) epistazie dominantă: se încrucișează ovăzul sălbatic (Avena fatua) cu boabe negre (AABB) cu ovăzul cultivat
(Avena sativa) cu boabe galbene (aabb). În F1 rezultă numai boabe negre, iar prin autopolenizarea hibrizilor rezultă în F2
următorul raport de segregare: 12 boabe negre : 3 boabe cenușii : 1 boabe galbene. A determină culoarea neagră, B
determină culoarea cenușie. În acest caz se constată atât modificarea raportului de segregare, cât și apariția unui fenotip
nou. Realizați schema de încrucișare.
d) epistazie recesivă: se încrucișează două rase de câini, una homozigotă cu blana albă (BBcc) și una homozigotă
cu blana maro (bbCC). În F1 rezultă numai hibrizi de culoare neagră (BbCc), iar prin încrucișarea între aceștia rezultă în F2
următorul raport de segregare: 9 negri : 4 albi : 3 maro. B_C_ – negri; bbC_ – maro; B_cc și bbcc – albi; gena c este
epistatică în condiție homozigotă recesivă. În acest caz se constată atât modificarea raportului de segregare, cât și apariția
unui fenotip nou. Realizați schema de încrucișare.
e) albinismul
2. Apariția unui fenotip nou cu păstrarea raportului de segregare W. Bateson și R. C. Punnett au studiat determinismul genetic al formei crestei la
găini. Astfel, găinile din rasa Leghorn au creastă simplă (rrpp), cele din rasa Brahmas au
creastă tip „mazăre” (rrPP), iar cele din rasa Wyandotte au creastă de tip „trandafir”
(RRpp).
Realizați următoarele încrucișări: Leghorn x Brahmas, Leghorn x Wyandotte
Prin încrucișarea raselor Brahmas și Wyandotte, prima generație a fost uniformă,
dar cu un fenotip nou – creastă de tip „nucă”. Prin încrucișarea între ei a hibrizilor din F1 a
rezultat în F2 următorul raport de segregare: 9 creastă „nucă” : 3 creastă „mazăre” : 3
creastă „trandafir” : 1 creastă simplă. În acest caz se constată apariția unor fenotipuri noi,
fără modificarea raportului de segregare mendelian. Realizați schema de încrucișare.
3. Poligenie Este fenomenul genetic caracterizat prin faptul că un singur fenotip/caracter este determinat prin acțiunea
conjugată a mai multor gene distincte (mai multor perechi de alele), care ocupă loci diferiți. A fost observat la plante și
animale, inclusiv la om, având mare importanță în moștenirea caracterelor cantitative (talie, greutate, productivitate –
semințe, fructe, ouă, lapte etc.)
Exemple a) talia
b) epistazia
c) forma crestei la găini
d) culoarea pielii la om: cel puțin două gene; alelele au efect aditiv (P1P1P2P2 –
negru; p1p1p2p2 – alb)
e) inteligența – caracter multifactorial (poligenic + ambiental)
f) culoarea bobului de grâu: Herman Nilsson-Ehle a încrucișat un soi de grâu
cu bob roșu foarte intens cu un soi cu bob alb. În F1 descendența a fost uniformă, cu
bobul roșu intermediar. În F2 s-a obținut următoarea segregare: 1 roșu foarte intens : 4
roșu intens : 6 roșu intermediar : 4 roz : 1 alb sau 15 nuanțe de roșu : 1 alb. Din acest
raport de segregare reiese că în determinismul culorii bobului de grâu sunt implicate
două gene distincte și că alelele lor au efect cumulativ/aditiv.

95 / 108
III. §PLEIOTROPIE
Pleiotropia reprezintă fenomenul genetic reprezentat de capacitatea unei singure perechi de alele (unei singure
gene) de a afecta mai multe caractere diferite.
Exemple a) maladia fenilcetonurie de la om (determinată de mutații ale unei singure gene – gena care codifică pentru
enzima fenilalanin-hidroxilază, care catalizează transformarea Phe în Tyr) cauzează retard mintal, reducerea pigmentării
părului și a pielii etc.
b) anemia falciformă – efectul primar al mutației genei care codifică pentru catena β este sinteza unei
hemoglobine anormale HbS; efecte pleiotrope – aspect falciform al hematiilor, friabilitate mare a acestora, anemie,
hiperactivitatea măduvei osoase hematogene, ischemie la nivelul tuturor organelor și alterarea fucțională a acestora etc.
c) gena vg de la drosofilă determină, pe lângă aripi vestigiale, și modificarea perișorilor de pe partea dorsală a
corpului, a spermatecii, reducerea fertilității și a vitalității etc.
§Parametri cantitativi ai fenomenului ereditar I. PENETRANȚĂ ȘI EXPRESIVITATE Penetranța reprezintă gradul în care se manifestă fenotipic în populație un anumit genotip (de obicei dominant).
Poate fi completă atunci când toți indivizii cu un anumit genotip prezintă fenotipul respectiv, sau incompletă – când numai
unii dintre cei care au genotipul prezintă fenotipul respectiv. Pentru individ, penetranța este un fenomen de tip „tot sau
nimic”.
Expresivitatea este o măsură a gradului în care se manifestă fenotipic la individ un anumit genotip (exemplu –
polidactilia poate să afecteze toate cele patru membre sau doar pe unele).
II. HERITABILITATE Heritabilitatea este un parametru prin care se exprimă contribuția genetică la determinarea unui fenotip.
Heritatbilitatea măsoară și exprimă contribuția relativă a diferențelor genetice și non-genetice (determinate de factori de
mediu sau de hazard) responsabile de variația fenotipică totală dintr-o populație. Măsoară cât din variabilitatea fenotipică
este determinată de variația genetică. Spre exemplu, studii mai vechi considerau că inteligența are o heritabilitate de 0.5-
0.8, deci este în mare parte determinată genetic, restul fiind determinat de factorii ambientali. Alte exemple: alcoolismul are
heritabilitate de 0,5-0,6, iar maladia Alzheimer – 0,58-0,79.
SPAȚIU PENTRU REZOLVAREA TEMELOR ȘI A SARCINILOR DE LUCRU

96 / 108

97 / 108

98 / 108

99 / 108
TEORIA CROMOSOMALĂ A EREDITĂȚII
Etape premergătoare În 1875, Wacław Mayzel (1847-1916, medic și histolog polonez) realizează prima descriere a mitozei, pe baza
observațiilor făcute, începând din 1873, pe celule de broască, iepure și de cornee de pisică. De obicei însă, descrierea
mitozei și elucidarea comportamentului cromosomilor îi sunt atribuite lui Walther Flemming (1843-1905), cel care a și
denumit procesul în articolele sale din 1878 și 1882. Tot Flemming este cel care a descoperit cromosomii, denumiți astfel
de Heinrich Wilhelm Gottfried von Waldeyer-Hartz (1836-1921, anatomist german).
Meioza a fost descoperită și descrisă pentru prima dată în 1876 de către biologul german Oscar Hertwig, prin
studii pe ovulele de arici de mare. Comportamentul cromosomilor în meioză a fost descris în 1883 de către zoologul
belgian Edouard Von Beneden, dar semnificația procesului în reproducere și ereditate a fost stabilită în 1890 de către
biologul german August Weismann.
În 1900, Carl Correns, Erich von Tschermak-Seysenegg, Hugo de Vries redescoperă independent legile lui
Mendel.
În această perioadă, oamenii de știință au manifestat preocupări privind localizarea materialului genetic (a
factorilor ereditari). Cunoscându-se importanța procesului sexual în reproducere și în transmiterea caracterelor în
descendență, cei mai buni candidați la rolul de vehicul al factorilor ereditari au fost gameții (celulele sexuale), care asigură
legătura între generații. Deoarece s-a constatat totuși că, în general, gameții de sexe diferite au dimensiuni diferite (gametul
♀ este, de obicei, mai mare, gametul ♂ este, de obicei, mai mic), s-a considerat inițial
că, la formarea descendentului, cele două sexe participă în mod inegal; Mendel însă
postulase o contribuție egală a celor doi părinți la înzestrarea descendenților, ca și
localizarea factorilor în nucleul celulelor, iar constatarea că totuși nucleii gameților sunt
de aceeași dimensiune susține aceste postulate. Astfel, nucleii celulelor reprezintă sediul
materialului genetic.
În 1902, Walter S. Sutton și Theodor Boveri, independent unul de celălalt,
pun bazele teoriei cromosomale a eredității18
prin experimente pe lăcuste – Brachystola
magna (Sutton) – și arici de mare – Paracentrotus lividus (Boveri); Sutton introduce
termenii „autosom” și „heterosom”; ei emit ipoteza plasării factorilor ereditari în
cromosomi.
În 1903 W. S. Sutton constată că numărul de caractere ale unui organism (și
deci numărul de perechi de factori ereditari care le determină) este mult mai mare decât
numărul de cromosomi, dar factorii ereditari și cromosomii se comportă similar în
meioză și fecundație => trage concluzia că un cromosom conține mai mulți factori/gene
dispuși liniar, în locuri bine precizate.
Comparație între comportamentul factorilor ereditari și al cromosomilor în
meioză și fecundație
1. Factorii ereditari/cromosomii sunt în perechi, unul de origine ♂, celălalt de
origine ♀, în nucleul celulelor somatice; cromosomii pereche sunt cromosomi omologi.
2. La formarea gameților, prin meioză, perechea de factori/cromosomi omologi se separă în gameți diferiți => un
gamet primește un singur factor/cromosom din fiecare pereche, deci se transmit în descendență ca entități discrete.
3. Prin fecundație, în zigot se refac perechile de factori ereditari/cromosomi omologi.
În 1909 W. Johannsen redenumește factorii ereditari gene și introduce conceptele de „genotip” și „fenotip”.
În 1906 geneticianul William Ernest Castle și entomologul Charles W. Woodworth, care pun bazele geneticii
populațiilor constatând constanța frecvenței genelor în populațiile de indivizi, fac cadou colecția lor de Drosophila
melanogaster (musculița de oțet, musculița de fructe) zoologului englez Thomas Hunt Morgan.
Drosophila melanogaster – model ideal de studiu al fenomenului ereditar
1. Se crește ușor în laborator, pe medii de cultură simple
2. Are timp de generație scurt – (10)12-14(15) zile, în funcție de temperatură
3. Prezintă prolificitate mare – număr mare de descendenți/generație => intrepretare statistică
4. Prezintă un număr mare (peste 500) de mutante naturale (echivalente soiurilor de la mazăre)
5. Are un număr mic de cromosomi mari – 2n = 8, ușor de observat
6. Prezintă în celulele glandelor salivare ale stadiului larvar cromosomi gigantici – cromosomi politeni
În perioada 1906-1913(5) T. H. Morgan (1866-1945) și colaboratorii săi (Alfred Sturtevant, Calvin Blackman
Bridges, Hermann Joseph Muller) realizează experimente de hibridare (mono-, di-, trifactoriale etc.) la Drosophila
melanogaster și, pe baza rezultatelor, elaborează și extind teoria cromosomală a eredității (pentru care T. H. Morgan a
primit premiul Nobel în 1933).
Morgan schimbă sistemul de denumire a alelelor: b, b+, w, w
+, vg, vg
+ etc. Astfel, denumirea alelei este
abrevierea cu o literă sau două a denumirii fenotipului mutant, determinat deci de alela mutantă, de obicei recesivă (ochi
18
Teoria cromosomală Sutton-Boveri, teoria Sutton-Boveri – teorie unificatoare a geneticii, care identifică cromosomii drept sediul materialului
genetic, pe baza similitudinilor între comportamentul factorilor ereditari mendelieni și cel al cromosomilor.

100 / 108
albi – white – w, corp negru – black – b, aripi vestigiale – vestigial – vg etc.), iar alela sălbatică, normală, de obicei
dominantă, este denumită cu aceeași literă sau grup de litere la care se superscrie în partea dreaptă semnul „+”.
Tezele teoriei cromosomale a eredității 1. Plasarea liniară a genelor în cromosomi*
Pe de o parte, în diviziune cromosomii se comportă precum factorii ereditari, dar își păstrează integritatea morfo-
structurală, adică se transmit întregi (Boveri a numit aceste proprietăți ale cromosomilor continuitate și individualitate).
*Dovezi ale corelației între cromosomi și fenotip la Drosophila (Sophophora) melanogaster – lipsa unui
cromosom din perechea IV (deci 2n = 7) determină fenotipul eyeless; determinismul sexelor (XX – individ ♀, XY – individ
♂).
Pe de altă parte, numărul de caractere fenotipice ereditare (corelat direct proporțional cu numărul de determinanți
genetici – gene/perechi de factori ereditari) este mult mai mare decât numărul de cromosomi din complementul
cromosomal => mai mulți determinanți genetici (gene/factori ereditari) sunt plasați, în succesiune liniară (precum
perlele/mărgelele pe ață), într-un singur cromosom, care funcționează ca un depozitar și vehicul pentru aceștia. Poziția
genei în cromosom este, în mod normal, specifică și se numește locus genic (pl. loci genici).
2. Transmiterea înlănțuită (în bloc, lincată) a genelor din același cromosom în cursul diviziunii celulare – teza
linkage-ului/lincajului*
[4. Corespondența între numărul perechilor de cromosomi și numărul grupelor de linkage]
În diviziune cromosomii se comportă precum factorii ereditari, dar își păstrează integritatea morfo-structurală,
adică se transmit întregi => genele conținute se transmit împreună, în bloc, de la celula-mamă la celulele-fiice. Fenomenul
de transmitere în bloc a genelor din același cromosom = linkage/lincaj.
Exemple I. W. Bateson și R. C. Punnett au încrucișat două soiuri de Lathyrus odoratus (măzăriche parfumată), unul cu
flori purpurii și polen alungit (AABB) cu unul cu flori roșii și polen rotund (aabb). În F1 toți hibrizii AaBa aveau flori
purpurii și polen alungit. În F2 s-a obținut un alt raport de segregare decât cel mendelian așteptat pentru dihibridare: 9:1:1:3.
Cei doi au subliniat tendința variantei fenotipice A de a se transmite în descendență împreună cu B, respectiv a variantei
fenotipice a cu b, adică de a se transmite lincate. Explicația – genele A și B sunt situate în același cromosom, iar acesta se
transmite integral în descendență, deci cele două gene se transmit în tandem; a și b sunt localizate în cromosomul omolog și
se transmit, de asemenea, împreună. În acest caz este mai corectă scrierea genotipurilor astfel AB/AB, respectiv ab/ab, în
loc de AABB, respectiv aabb. Bateson și Punnett nu au dat însă o explicație pentru apariția fenotipurilor recombinate Ab
și aB.
Există două variante posibile de plasare a genelor lincate ale organismului heterozigot:
a) AB/ab – justifică proporția mare a fenotipurilor AB (9) și ab (3), față de Ab (1) și aB (1) în segregare 9:1:1:3
b) Ab/aB – rezultă prin crossing-over; justifică proporția mică a fenotipurilor Ab (1) și aB (1) în segregarea
9:1:1:3
Cele două variante dau raporturi de segregare diferite. Lincajul este baza pentru cartarea genelor și are rol important în ameliorare (lincaj între gene pentru caractere favorabile). Lincajul ajută la
urmărirea transmiterii unor gene ale căror efecte fenotipice sunt greu de decelat într-o anumită fază a ontogeniei.
II. T. Morgan a realizat o primă încrucișare între două musculițe de oțet (genitorii, generația parentală): un
mascul cu corp de culoare normală (gri-cenușiu) și aripi normale, b+b
+vg
+vg
+, cu o femelă dublu-mutantă – corp negru și
aripi vestigiale, bbvgvg. Descendenții (F1) au fost toți hibrizi, dar dublu-normali fenotipic, b+bvg
+vg (sau rescris
b+vg
+/bvg). A continuat experimentul încrucișând un mascul dublu-hibrid b
+bvg
+vg din F1 cu o femelă dublu-mutantă
bbvgvg (a realizat astfel un test cross/backcross). Fiind vorba de o dihibridare, se aștepta ca raportul de segregare din F2 să
fie unul mendelian, 1:1:1:1, însă a rezultat altceva – 50% b+vg
+ și 50% bvg, adică un raport de 1:1. Singura explicație și
concluzie în același timp este că b se transmite împreună cu vg, respectiv b+ cu vg
+ (adică b și vg, respectiv b
+ și vg
+ sunt
lincate), deoarece cromosomii în care se află își păstrează integritatea structurală în timpul meiozei – lincaj total.
Organismul dublu-heterozigot formează deci doar două tipuri de gameți – b+vg
+ și bvg, iar prin unirea probabilistică cu
unicul tip de gamet feminin (bvg) rezultă descendența de mai sus.
S-a stabilit că la drosofilă există patru grupe de
lincaj, număr care corespunde numărului de perechi de
cromosomi ai setului diploid, 2n = 8 cromosomi; la Zea mays
– 10 grupe de lincaj, 2n = 20; la Mus musculus – 20 de grupe
de lincaj, 2n = 40. La organismele haploide numărul grupelor
de lincaj este egal cu numărul de cromosomi.
3. Schimbul reciproc și egal de gene (fragmente
cromatidiale) între cromatidele nesurori ale
cromosomilor omologi sinapsați în profaza I – teza
crossing-over-ului.
Când T. Morgan a realizat încrucișarea reciprocă a
unui hibrid din F1, o femelă dublu-hibridă b+bvg
+vg cu un
mascul dublu-mutant bbvgvg (a realizat un tot test
cross/backcross), rezultatul a fost surprinzător – nici
segregare mendeliană 1:1:1:1, nici segregarea obținută
anterior, 1:1, ca urmare a lincajului. În F2 segregarea

101 / 108
fenotipică a fost următoarea: 41,5%
b+vg
+, 41,5% bvg (adică 83%
fenotipuri parentale; manifestă
lincaj), 8,5% b+vg și 8,5% bvg
+
(adică 17% organisme recombinate;
reprezintă o abatere de la lincaj) =>
lincaj parțial. O segregare atipică
au obținut și Bateson și Punnett la
Lathyrus odoratus (vezi mai sus).
Explicarea rezultatelor
Procentul foarte mare
(83%) de organisme cu fenotip
parental (b+vg
+ și bvg) susține
lincajul celor două gene, deci alelele
b+ și vg
+ sunt localizate în același
cromosom, la o oarecare distanță una
față de alta, iar b și vg sunt localizate
în cromosomul omolog, în aceeași
loci și deci la aceeași distanță una
față de alta ca și variantele lor
dominante. În meioză cei mai mulți
cromosomi se transmit nemodificați
de la celula-mamă la celulele-fiice.
Abaterea de la lincaj se
explică prin faptul că femelele
dublu-heterozigote b+bvg
+vg
produc, nu două, ci patru tipuri de
gameți (ovule) – cei mai mulți (83%)
sunt gameți nerecombinați, de tip
parental, la care se manifestă lincajul
(b+vg
+ și bvg), dar și câțiva (17%)
gameți recombinați prin crossing-
over (b+vg și bvg
+). Procentul mic al
acestor gameți recombinați susține
apariția lor prin alt mecanism decât
segregarea independentă
mendeliană, mai ales că respectivele gene sunt plasate în același cromosom. Prin unirea probabilistică a celor patru tipuri de
gameți feminini cu unicul tip de gamet masculin (bvg), rezultă descendența și segregarea obținute de Morgan.
Din punct de vedere citologic, în timpul meiozei, în profaza I, cromosomii omologi se împerechează aliniindu-se
perfect, genă la genă, centromer la centromer. Atașarea, numită sinapsă cromosomală, se realizează prin intermediul unui
complex proteic – complexul sinaptonemal – care se formează între cromatidele nesurori ale celor doi omologi. Această
atașare permite ruperea a două cromatide nesurori (una maternă și una paternă) la același nivel, iar fragmentele
cromatidiale (care conțin aceleași gene, dar nu neapărat aceleași alele ale lor) rezultate își schimbă reciproc poziția („se
încrucișează”) și apoi se realipesc la cromatidele de origine opusă (fragmentul desprins din cromatida de origine maternă se
alipește la cromatida paternă și invers). Rezultă astfel cromatide și cromosomi recombinați, care se vor transmite astfel în
gameți și apoi în descendență. Locul unde se realizează efectiv încrucișarea celor două cromatide nesurori se numește
chiasmă. Aceste chiasme sunt vizibile la microscopul optic într-un preparat cu celule în profaza I a meiozei, fiecare
chiasmă indicând un eveniment de crossing-over. Cu cât cromosomii sunt mai lungi, cu atât numărul de chiasme este mai
mare, deci și numărul de evenimente de recombinare – crossing-over simplu, dublu, triplu etc.
Crossing-over-ul se poate realiza numai între cromosomii omologi (perechile de autosomi și perechea XX), nu și
între cromosomii X și Y (nu sunt omologi).

102 / 108
§Hărți genetice Frecvența cu care apare crossing-over-ul (exprimată prin procentul de descendenți recombinați, ex., 17% în cazul experimentului de mai sus)
depinde de distanța la care se găsesc cei doi loci genici în cromosom – cu cât distanța este mai mare, cu atât este mai mare probabilitatea ca respectivele
cromatide nesurori, în timpul sinapsei cromosomale, să se rupă și să facă schimb reciproc de fragmente, deci să se realizeze recombinarea intracromosomală. De ce? Pentru că în cazul unei distanțe mai mari, din punct de vedere statistic/probabilistic, există mai multe puncte fizice în care să se
formeze chiasme, deci să se realizeze crossing-over-ul.
Astfel, frecvența cu care apar în descendență organisme recombinate este un indicator direct al poziției genelor în cromosom, fiind direct
proporțională cu distanța dintre ele, plecând și de la premisa că genele sunt plasate liniar în cromosmi. Această constatare stă la baza realizării unor hărți
genetice19 – hărțile cromosomale. O asemenea hartă este o reprezentare grafică, schematică, a localizării genelor în lungul unui cromosom și reflectă
poziția relativă a genelor unele față de altele în cromosom, stabilită pe baza frecvenței crossing-over-erelor, deci a procentelor de recombinare. Unitatea
de măsură a distanței relative între gene este chiar acest procent de recombinare, numit și morgan/morganon/centimorgan. Realizarea hărților cromosomale implică folosirea analizei genetice – efectuarea de hibridări și studierea descendenței de-a lungul mai multor
generații; se determină astfel grupele de lincaj și frecvența crossing-over-erelor.
Exemple
1. Prin analiza genetică s-a determinat că genele ipotetice A, B și C sunt lincate și că, prin încrucișarea unor indivizi triplu heterozigoți
AaBbCb, dar normali fenotipic (ABC), cu indivizi triplu mutanți aabbcc (fenotip abc), descendența este formată în principal din organisme asemănătoare părinților, ABC, respectiv abc, dar apar și organisme recombinate prin crossing-over în următoarele proporții:
a. Abc și aBC – 15% b. ABc și abC – 8%
c. AbC și aBc – 23%
Stabiliți poziția celor trei gene în cromosom. Prima recombinare: ABC și abc s-au rupt între A și B, respectiv între a și b și au schimbat poziția => fragmentul cu A s-a alipit la bc, iar
fragmentul cu a s-a alipit la BC => Abc și aBC; aceste organisme recombinate au reprezentat 15% din descendență => între A și B, respectiv între a și b
este o distanță de 15 centimorgani.
A doua recombinare: ABC și abc s-au rupt între B și C, respectiv între b și c și au schimbat poziția => fragmentul cu c s-a alipit la AB, iar
fragmentul cu C s-a alipit la ab => ABc și abC; aceste organisme recombinate au reprezentat 8% din descendență => între B și C, respectiv între b și c este
o distanță de 8 centimorgani. A treia recombinare: ABC și abc s-au rupt în două puncte între A și C, respectiv între a și c și au schimbat poziția => fragmentul cu B s-a alipit
la ac, între a și c, iar fragmentul cu b s-a alipit la AC, între A și C => AbC și aBc (s-a realizat un crossing-over dublu); aceste organisme recombinate au
reprezentat 23% din descendență => între A și C, respectiv între a și c este o distanță de 23 centimorgani. Din rezultatele de mai sus se poate deduce că A, B și C sunt situate în această ordine în cromosom, cu B mai aproape de C – distanța dintre A și
B plus distanța dintre B și C este egală cu distanța dintre A și C.
2. În tabelul și imaginea de mai jos sunt precizate procentele de recombinare prin crossing-over între unele gene din cromosomul X de la
Drosophila melanogaster. Pe baza acestor valori și a relațiilor dintre ele se poate stabili poziția genelor respective în cromosomul X (succesiunea lor și
distanța dintre ele).
Gena pentru
aripi miniaturale
Gena pentru ochi
rubinii
Gena pentru
corp galben
Gena pentru ochi
albi
Gena pentru corp galben 36,1% 7,5% - 1,4%
Gena pentru ochi albi 34,7% 6,1% 1,4% -
Gena pentru peri pârliți 15,1% 13,5% 21% 19,6%
Gena pentru aripi miniaturale - 28,6% 36,1% 34,7%
Prin hibridări experimentale s-a reușit localizarea unor gene în cromosomi la unele specii de eucariote – Drosophila melanogaster, Zea mays,
Neurospora crassa, Chlamydomonas reinhardtii, Solanum lycopersicon, Mus musculus, Homo sapiens => s-au realizat hărți cromosomale.
Elaborarea hărților cromosomale este o importantă realizare a geneticii moderne. La specia umană creează premisele prevenirii și tratării unor
boli cu transmitere ereditară.
19 Alte tipuri de hărți genetice sunt hărțile citologice și hărțile fizice.

103 / 108
APLICAȚII

104 / 108

105 / 108
RECOMBINAREA GENETICĂ
Recombinarea genetică este procesul prin care apar noi combinații ale genelor și alelelor provenite din surse
diferite (ex., alele materne și paterne), care se transmit în descendență și contribuie la generarea variabilității, alături de
mutații.
Recombinarea genetică la procariote
Se realizează prin multe mecanisme/procese de transfer de gene: transformare, conjugare, sexducție,
transducție fagică. Este un fenomen larg răspândit în natură. Stă la baza biotehnologiilor moderne. Implică cromosomul
bacterian, plasmidele și chiar virusuri.
Recombinarea genetică la eucariote
La speciile care se reproduc sexuat, comportamentul cromosomilor în timpul meiozei și fecundației este
responsabil pentru majoritatea variabilității care apare la fiecare generație.
Există trei mecanisme principale care contribuie la variația genetică apărută prin reproducerea sexuată: segregarea
independentă a cromosomilor (asocierea independentă/aleatorie a cromosomilor), crossing-over-ul și fecundația aleatorie.
1. Recombinare intercromosomală (segregarea independentă a perechilor de omologi)*
Un aspect al reproducerii sexuate care
generează variație genetică este orientarea
randomică a perechilor de cromosomi omologi în
metafaza meiozei I. În metafaza I, perechile de
omologi, fiecare alcătuită dintr-un cromosom
matern și unul patern, sunt situate în placa
metafazică (de observat că termenul matern și
patern se referă la mama, respectiv tatăl individului
ale cărui celule suferă meioza). Fiecare pereche se
poate orienta fie cu omologul matern, fie cu cel
patern mai aproape de un pol dat – orientarea lor
este aleatorie, ca și în cazul unei monede care,
aruncată, poate să cadă cu o față sau cu cealaltă în
sus. Astfel există 50% șansa ca o anumită celulă-
fiică a rezultată din meioza I să primească
cromosomul matern dintr-o anumită pereche de omologi și 50% șanse să primească cromosomul patern din aceeași
pereche.
Deoarece în metafaza I fiecare pereche de cromosomi omologi se orientează independent de celelalte perechi, în
urma primei diviziuni meiotice rezultă o separare și distribuire a omologilor materni și paterni în celulele-fiice,
independente pentru fiecare pereche în parte raportat la toate celelalte perechi. Acest fenomen/proces se numește segregare
independentă și a fost numit de Hermann Joseph Muller „dansul cromosomilor”. Fiecare celulă-fiică conține una din toate
combinațiile posibile de cromosomi materni și paterni. Numărul de combinații posibile pentru celulele-fiice formate prin
meioza unei celule diploide cu două perechi de cromosomi omologi (n = 2, 2n = 4) este 4 (patru), adică 22. De remarcat că
doar două din aceste patru combinații de celule-fiice vor rezulta din meioza unei singure celule diploide, deoarece o singură
celulă parentală va avea doar una sau cealaltă posibilitate de aranjare a cromosomilor în metafaza I, dar nu pe ambele.
Totuși, populația de celule-fiice care rezultă din meioza unui număr mare de celule diploide conține toate cele patru tipuri
în proporții aproximativ egale. În cazul în care n = 3, 2n = 6, sunt posibile 8, adică 23 combinații de celule-fiice.
Generalizând, numărul de combinații posibile rezultate prin segregare independentă a perechilor de omologi în timpul
meiozei este 2n , unde n este numărul haploid al organismului.
În cazul oamenilor, numărul haploid (n) din formulă este 23. Astfel, numărul de combinații posibile de cromosomi
materni și paterni în gameți este 223
, adică aproximativ 8 milioane (8.388.608). Fiecare gamet pe care îl produce un individ
conține doar una din cele aproximativ opt milioane de combinații posibile de cromosomi moșteniți de la mamă și de la tată.
2. Recombinare intracromosomală (crossing-over)
Ca o consecință a segregării independente a cromosomilor în timpul meiozei, fiecare individ produce o colecție de
gameți care diferă mult în combinația lor de cromosomi pe care i-a moștenit de la cei doi părinți – gameții conțin seturi de
cromosomi recombinate, rezultate prin segregarea independentă a perechilor de omologi. În explicarea segregării
independente, am considerat că fiecare cromosom individual ajuns într-un gamet este exclusiv și în întregime matern sau
patern ca origine. De fapt, această premisă este eronată (convenabilă însă în explicarea segregării independente), deoarece
crossing-over-ul produce cromosomi recombinați – cromosomi individuali care poartă gene (ADN) provenite de la ambii
părinți.
Crossing-over-ul începe foarte devreme în profaza I, când cromosomii omologi bicromatidici/duplicați se aliniază
și se conectează fizic lax pe întreaga lungime prin intermediul/intervenția unei structuri proteice cu aspect de fermoar –
complexul sinaptonemal; acest proces se numește sinapsă cromosomală. Fiecare genă dintr-un omolog este precis
aliniată cu gena corespondentă din celălalt omolog, ca și centromerii celor doi omologi. Într-un singur eveniment de
crossing-over, moleculele de ADN din două cromatide nesurori – o cromatidă maternă și una paternă din perechea de
omologi – se rup în același loc și apoi fragmentele sunt realipite încrucișat, fiecare la molecula de ADN cu origine opusă

106 / 108
(segmentul din cromatida paternă începând de la punctul
ruperii spre capăt este reatașat la cromatida maternă și
invers). Ca efect, două segmente omoloage schimbă locul
sau „se încrucișează” (cross over), producând cromosomi cu
o nouă combinație de gene (recombinați), atât materne, cât
și paterne – se realizează astfel o rearanjare genetică între
cromatidele nesurori.
Consecutiv dezasamblării complexului
sinaptonemal în profaza I târzie, cele patru cromatide ale
perechii de omologi sunt vizibile la microscopul optic sub
formă de tetradă cromatidială. Fiecare tetradă conține în
mod normal cel puțin o regiune în formă de „X” numită
chiasmă – dovada fizică a crossing-over-ului. Sinapsa și
crossing-over-ul în mod normal nu se produc în timpul
mitozei.
La oameni, se produc în medie 1-3 evenimente de
crossing-over per pereche de omologi, în funcție de
mărimea cromosomilor și de poziția centromerului.
În metafaza II, cromosomii care conțin una sau
ambele cromatide recombinate pot fi orientate în două
variante neechivalente față de ceilalți cromosomi, deoarece
cromatidele lor surori nu mai sunt identice. Segregarea
independentă a acestor cromatide surori non-identice în
timpul meiozei II amplifică și mai mult numărul de tipuri
genetice ale celulelor-fiice care rezultă din meioză.
Crossing-over-ul, prin combinarea ADN-ului
moștenit de la doi părinți într-un singur cromosom, este o
sursă importantă de variație genetică în ciclurile de viață
sexuale.
Crossing-over-ul poate fi intergenic (cel mai adesea) sau intragenic.
3. Fecundație aleatorie
Natura aleatorie/randomică a fecundației/singamiei se adaugă la variația genetică apărută prin meioză. La oameni,
spre exemplu, fiecare gamet masculin, respectiv feminin reprezintă una din aproximativ 8 milioane de combinații posibile
de cromosomi ca urmare a segregării independente din timpul meiozei. Fuziunea unui singur gamet masculin cu un singur
gamet feminin în timpul fecundației va produce un zigot cu oricare din aproximativ 64 trilioane (8 milioane x 8 milioane)
de combinații diploide posibile. Dacă se calculează 223
x 223
exact, se va obține un total care este de fapt de peste 70
trilioane. Adăugând această variație la cea generată prin crossing-over, numărul de posibilități este cu adevărat astronomic.
Nu-i de mirare că frații și surorile sunt atât de diferiți. Fiecare individ este într-adevăr unic.
4. Conversie genică (schimb nereciproc de material genetic)
Este o recombinare genetică nereciprocă și care se produce tot în meioză. Termenul a fost introdus de Winkler în
1930 și reluat de Lindegren în 1949. A fost evidențiată prin analiza tetradei la drojdii din genul Saccharomyces unde
uneori, în 1-10% din asce (organ de reproducere caracteristic unui grup de ciuperci – ascomicete), apare o abatere de la
raportul normal de segregare alelică de 2A:2a, apărând raporturi aberante de 3A:1a, 4A:0a, 1A:3a, 0A:4a.
La ascomicetele cu 8 ascospori în ască (Neurospora crassa, Aspergillus sp. etc.) apar abateri de la raportul normal
4A:4a de segregare alelică. Zigotul acestor ciuperci este 2n și heterozigot Aa; suferă meioză și o mitoză postmeiotică,
rezultând asca cu 8 celule n – ascospori; în cazul în care nu se realizează conversie, patru ascospori vor conține alela A, iar
ceilalți patru – alela a, deci raportul de segregare va fi de 4A:4a. Uneori raportul de segregare este atipic – 5A:3a, 6A:2a
etc.
La nivel molecular, conversia genică presupune un transfer unidirecțional/nereciproc al unui segment de ADN de
cca. 100-200 de nucleotide de la o cromatidă la cromatida cromosomului omolog. Segmentul transferat înlocuiește
segmentul omolog și astfel se trece de la starea heterozigotă la cea homozigotă. Din Aa, A trece în locul lui a => perechea
devine AA.
Dacă se transferă un segment dintr-o cromatidă în alta, cromatida donoare nu rămâne cu o lipsă? Cum se
rezolvă această problemă?
5. Recombinare de tip V(D)J
6. Recombinarea nespecifică/transpozițională
Acordul părintelui pentru utilizarea prezentului material ca suport de curs la orele de biologie
Subsemnatul, _______________________________________, părinte al elevului _____________________________, din
clasa a IX-a ___ de la Colegiul Național „Sfântul Sava”, București, sunt / nu sunt de acord cu utilizarea prezentului
material ca suport de curs/auxiliar didactic la orele de biologie.
Data_____________________ Semnătura________________________

107 / 108

108 / 108
APLICAȚII




















