
Application of RNAi to confirm theobromine as the major intermediate for caffeine biosynthesis in...
11
Application of RNAi to confirm theobromine as the major intermediate for caffeine biosynthesis in coffee plants with potential for construction of decaffeinated varieties Shinjiro Ogita 1,2 , Hirotaka Uefuji 1 , Masayuki Morimoto 1 and Hiroshi Sano 1, * 1 Research and Education Center for Genetic Information, Nara Institute of Science and Technology, Nara 630-0192, Japan (*author for correspondence; e-mail: [email protected]); 2 Biotechnology Research Center, Toyama Prefectural University, Toyama 939-0398, Japan Received 3 April 2004; accepted in revised form 24 May 2004 Key words: Agrobacterium, caffeine, coffee, RNAi, somatic embryogenesis, theobromine Abstract The caffeine biosynthetic pathway in coffee plants has been proposed to involve three distinct N-meth- yltransferases, xanthosine methyltransferase (XMT), 7-N-methylxanthine methyltransferase (MXMT; theobromine synthase), and 3,7-dimethylxanthine methyltransferase (DXMT; caffeine synthase). We pre- viously isolated all corresponding cDNAs designated as CaXMT1, CaMXMT1, CaMXMT2 and CaD- XMT1, respectively, and showed that caffeine was indeed synthesized in vitro by the combination of their gene products. In order to regulate caffeine biosynthesis in planta, we suppressed expression of CaMXMT1 by the double stranded RNA interference (RNAi) method. For this purpose, we first established a protocol for efficient somatic embryogenesis of Coffea arabica and C. canephora, and then Agrobacterium-mediated transformation techniques. The RNAi transgenic lines of embryogenic tissues derived from C. arabica and transgenic plantlets of C. canephora demonstrated a clear reduction in transcripts for CaMXMT1 in comparison with the control plants. Transcripts for CaXMT1 and CaDXMT1 were also reduced in the most cases. Both embryonic tissues and plantlets exhibited a concomitant reduction of theobromine and caffeine contents to a range between 30% and 50% of that of the control. These results suggest that the CaMXMT1-RNAi sequence affected expression of not only CaMXMT1 itself, but also CaXMT1 and CaDXMT1, and that, since the reduction in theobromine content was proportional to that for caffeine, it is involved in the major synthetic pathway in coffee plants. The results also indicate that the method can be practically applied to produce decaffeinated coffee plants. Introduction To date, approximately 12 000 alkaloids have been identified, mostly from plants (Hashimoto and Yamada, 2003). Purine alkaloids are derivatives of purine nucleotides, among which caffeine is one of the most well known as an essential ingredient of beverages, including coffee and tea (Ashihara and Crozier, 2001). Caffeine is also used as central nervous system stimulant, cardiac and diuretic due to its pharmacological effects. Because of the commercial importance of this alkaloid, its biosynthetic pathway in coffee plants has long been the focus of intensive studies. Based on recent knowledge of enzymology and molecular biology, it was proposed that caffeine is synthesized through multiple methylation of xanthine deriva- tives (Figure 1) (Ashihara and Crozier, 2001). The first step is methylation of xanthosine by xantho- sine methyltransferase (XMT), yielding 7-meth- ylxanthosine. After removal of its ribose residue, the resulting 7-methylxanthine is methylated by 7-N-methylxanthine methyltransferase (MXMT or theobromine synthase), with production of 3,7-dimethylxanthine (theobromine). Theobro- mine is then methylated by 3,7-dimethylxanthine Plant Molecular Biology 54: 931–941, 2004. ȑ 2004 Kluwer Academic Publishers. Printed in the Netherlands. 931
Transcript of Application of RNAi to confirm theobromine as the major intermediate for caffeine biosynthesis in...

Application of RNAi to confirm theobromine as the major intermediatefor caffeine biosynthesis in coffee plants with potential for constructionof decaffeinated varieties
Shinjiro Ogita1,2, Hirotaka Uefuji1, Masayuki Morimoto1 and Hiroshi Sano1,*1Research and Education Center for Genetic Information, Nara Institute of Science and Technology,Nara 630-0192, Japan (*author for correspondence; e-mail: [email protected]);2 Biotechnology Research Center, Toyama Prefectural University, Toyama 939-0398, Japan
Received 3 April 2004; accepted in revised form 24 May 2004
Key words: Agrobacterium, caffeine, coffee, RNAi, somatic embryogenesis, theobromine
Abstract
The caffeine biosynthetic pathway in coffee plants has been proposed to involve three distinct N-meth-yltransferases, xanthosine methyltransferase (XMT), 7-N-methylxanthine methyltransferase (MXMT;theobromine synthase), and 3,7-dimethylxanthine methyltransferase (DXMT; caffeine synthase). We pre-viously isolated all corresponding cDNAs designated as CaXMT1, CaMXMT1, CaMXMT2 and CaD-XMT1, respectively, and showed that caffeine was indeed synthesized in vitro by the combination of theirgene products. In order to regulate caffeine biosynthesis in planta, we suppressed expression of CaMXMT1by the double stranded RNA interference (RNAi) method. For this purpose, we first established a protocolfor efficient somatic embryogenesis of Coffea arabica and C. canephora, and then Agrobacterium-mediatedtransformation techniques. The RNAi transgenic lines of embryogenic tissues derived from C. arabica andtransgenic plantlets of C. canephora demonstrated a clear reduction in transcripts for CaMXMT1 incomparison with the control plants. Transcripts for CaXMT1 and CaDXMT1 were also reduced in themost cases. Both embryonic tissues and plantlets exhibited a concomitant reduction of theobromine andcaffeine contents to a range between 30% and 50% of that of the control. These results suggest that theCaMXMT1-RNAi sequence affected expression of not only CaMXMT1 itself, but also CaXMT1 andCaDXMT1, and that, since the reduction in theobromine content was proportional to that for caffeine, it isinvolved in the major synthetic pathway in coffee plants. The results also indicate that the method can bepractically applied to produce decaffeinated coffee plants.
Introduction
To date, approximately 12 000 alkaloids have beenidentified, mostly from plants (Hashimoto andYamada, 2003). Purine alkaloids are derivatives ofpurine nucleotides, among which caffeine is one ofthe most well known as an essential ingredient ofbeverages, including coffee and tea (Ashihara andCrozier, 2001). Caffeine is also used as centralnervous system stimulant, cardiac and diuretic dueto its pharmacological effects.
Because of the commercial importance of thisalkaloid, its biosynthetic pathway in coffee plants
has long been the focus of intensive studies. Basedon recent knowledge of enzymology and molecularbiology, it was proposed that caffeine is synthesizedthrough multiple methylation of xanthine deriva-tives (Figure 1) (Ashihara and Crozier, 2001). Thefirst step is methylation of xanthosine by xantho-sine methyltransferase (XMT), yielding 7-meth-ylxanthosine. After removal of its ribose residue,the resulting 7-methylxanthine is methylatedby 7-N-methylxanthine methyltransferase (MXMTor theobromine synthase), with production of3,7-dimethylxanthine (theobromine). Theobro-mine is then methylated by 3,7-dimethylxanthine
Plant Molecular Biology 54: 931–941, 2004.� 2004 Kluwer Academic Publishers. Printed in the Netherlands.
931

methyltransferase (DXMT or caffeine synthase) togive caffeine (1,3,7-trimethylxanthine) (Figure 1).
We cloned cDNAs encoding these three en-zymes, and designated as CaXMT1 (accessionnumber AB048793), CaMXMT1 (AB048794),CaMXMT2 (AB084126) and CaDXMT1(AB084125) (Ogawa et al., 2001; Uefuji et al.,2003). The cDNAs for MXMT designated asCTS1 (AB034700) and CTS2 (AB054841), and forDXMT designated as CCS1 (AB086414) were alsoreported (Mizuno et al., 2003). We showed that acombination of these three bacterially expressedproteins (CaXMT1, CaMXMT2 and CaDXMT1)could successfully convert xanthosine into caffeinein vitro, suggesting that caffeine is also synthesizedin vivo via this pathway (Uefuji et al., 2003).However, whether this is the major route of caf-feine synthesis in planta remained unclear, sincebypass pathways involving 1,7-dimethylxanthine(paraxanthine) and 3-methylxanthine have alsobeen predicted to contribute to the caffeine syn-thesis (Figure 1) (Ashihara and Crozier, 1999).
We describe here the production of transgeniccoffee plants, both Coffea arabica and C. caneph-ola, featuring suppression of CaMXMT1expression by the double-stranded RNA
(dsRNA)-mediated interference method (RNAi).Resulting transgenic tissues and plantlets could beshown to exhibit a marked reduction of transcriptsnot only for CaMXMT1 but also for CaXMT1and CaDXMT1, and concomitantly, of theobro-mine and caffeine contents. The reduction rate ofcaffeine was proportional to that of theobromine.The conclusion that, in coffee plants, the majorpathway of caffeine synthesis is mediated bythe theobromine synthase, CaMXMT1, was thussubstantiated. Furthermore, the RNAi andregeneration system developed in this study toproduce genetically modified coffee plants with alow level of caffeine, the prototype of which wepreliminarily reported earlier (Ogita et al., 2003),appears commercially promising.
Materials and methods
Plant materials
Newly flushed young leaves approximately 5–9 cmlong were collected from coffee plants of bothC. arabica L. var. Caturra and C. canephora var.Conilon, growing under a natural photoperiod in a
Figure 1. Putative caffeine biosynthetic pathway in coffee plants. Solid arrows indicate major routes, and broken arrows indicate
minor or predicted routes.
932

greenhouse. Collected leaves were surface steril-ized with 70% ethyl alcohol for 1 min followed by2% NaClO solution for 10 min. They were thenwashed three times with sterile water, dried on asterile paper, and used as explants,
Preparation of embryogenic cultures
Embryogenic cultures of both C. arabica andC. canephora were performed on modifiedhalf-strength MS medium (modified 1/2MS),containing 3% sucrose, half concentrations ofinorganic elements and the same concentrationsof other elements as with the original MS med-ium (Murashige and Skoog, 1962). The pH of allmedia used was adjusted to 5.7 with several dropsof 1 N KOH and/or 1 N HCl. The completemodified 1/2MS solution containing 0.3% gellangum was autoclaved for 20 min at 121 �C, and a20-ml aliquot was poured into a petri dish with90-mm diameter. Modified 1/2MS medium sup-plemented with 1 lMM of N-(2-chloro-4-pyridyl)-
N¢-phenylurea (4-CPPU) and 5 lMM of 2,4-dichlorophenoxyacetic acid (2,4-D) was appliedto induce embryogenic tissues of C. arabica.Sterilized explants were cut into pieces ofapproximately 5–7 mm2 and placed on modified1/2MS medium positioning the leaf upper sidedown and cultures were incubated at 25 �C in thedark. For further maintenance of subcultures,initiated embryogenic tissues were transferred tothe same medium at 3-week intervals. Twentymicro molars of 2-isopentenyladenine (2ip) wasadded to modified 1/2MS for direct somaticembryogenesis of C. arabica and C. canephora.Sterilization of explants, preparation of medium,and other procedures were the same as describedabove. Somatic embryos were directly formedwithout producing calli on the cut edges ofleaves. For further induction of development,initiated somatic embryos were transferred tomodified 1/2MS medium with or withoutN6-benzyladenine (BA). The cultures were incu-bated at 25 �C in the dark for induction and
Figure 2. Somatic embryogenesis of coffee plants. (A) Embryogenic tissues of C. arabica derived from leaf sections on modified 1/2MS
medium containing 1 lMM of 4-CPPU and 5 lMM of 2,4-D. (B) Somatic embryos of C. canephora directly induced from cut edges of leaf
section on modified 1/2MS medium containing 20 lMM of 2ip. (C) Germination of C. canephora somatic embryos. (D) One-year-old
coffee plants grown in a greenhouse (C. canephora). Scale bars in (A) and (B) indicate 1 cm, respectively.
933
maintenance of somatic embryos, and at 25 �Cwith a 16-h/8-h light/dark cycle under fluorescentillumination at 30–35 mol m)2 s)1 for develop-ment and germination, respectively.
Construction of transformation vectors
The 30-untranslated region (UTR) and codingregion of CaMXMT1 cDNA (AB048794) wasselected for design of double stranded RNAinterference (RNAi) constructs. RNAi-S (short)was constructed using 139-bp (positions1139–1277 of the cDNA) and 161-bp (positions1117–1277) sections. As the spacer, a 517-bp DNA
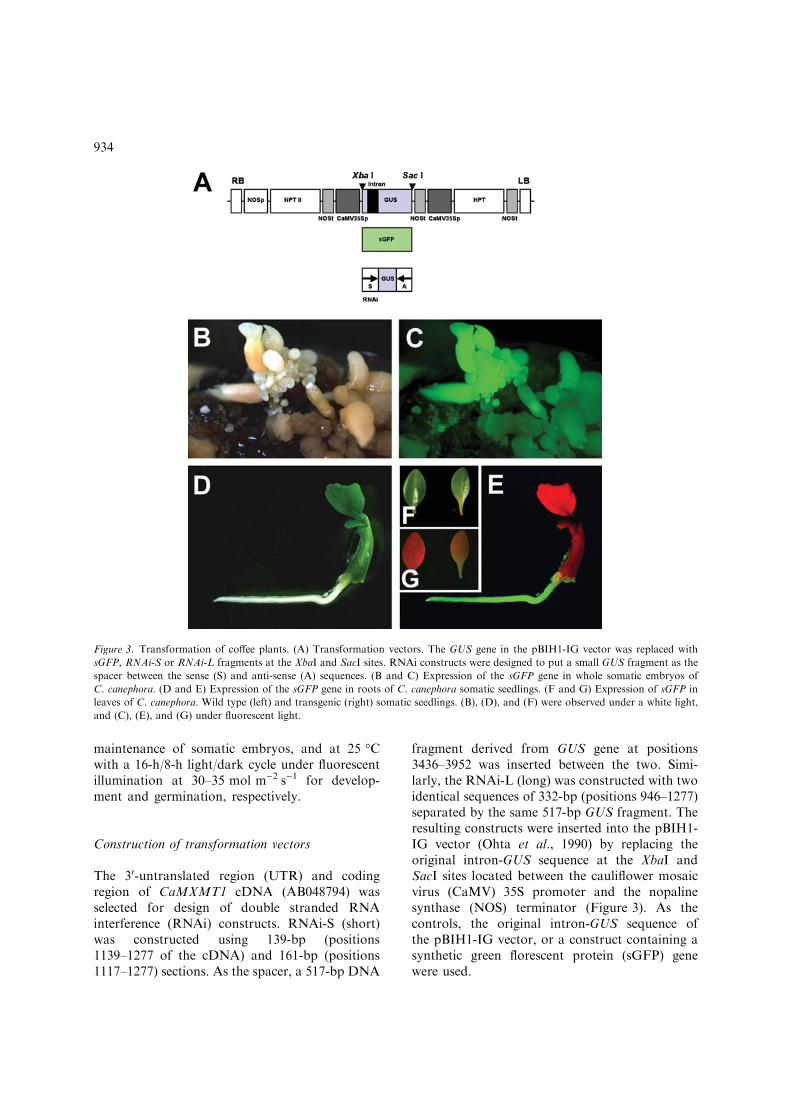
fragment derived from GUS gene at positions3436–3952 was inserted between the two. Simi-larly, the RNAi-L (long) was constructed with twoidentical sequences of 332-bp (positions 946–1277)separated by the same 517-bp GUS fragment. Theresulting constructs were inserted into the pBIH1-IG vector (Ohta et al., 1990) by replacing theoriginal intron-GUS sequence at the XbaI andSacI sites located between the cauliflower mosaicvirus (CaMV) 35S promoter and the nopalinesynthase (NOS) terminator (Figure 3). As thecontrols, the original intron-GUS sequence ofthe pBIH1-IG vector, or a construct containing asynthetic green florescent protein (sGFP) genewere used.
Figure 3. Transformation of coffee plants. (A) Transformation vectors. The GUS gene in the pBIH1-IG vector was replaced with
sGFP, RNAi-S or RNAi-L fragments at the XbaI and SacI sites. RNAi constructs were designed to put a small GUS fragment as the
spacer between the sense (S) and anti-sense (A) sequences. (B and C) Expression of the sGFP gene in whole somatic embryos of
C. canephora. (D and E) Expression of the sGFP gene in roots of C. canephora somatic seedlings. (F and G) Expression of sGFP in
leaves of C. canephora. Wild type (left) and transgenic (right) somatic seedlings. (B), (D), and (F) were observed under a white light,
and (C), (E), and (G) under fluorescent light.
934

Agrobacterium-mediated transformation
Agrobacterium tumefaciens strain EHA101 (Hoodet al., 1986) was transfected with the pBIH1-IGvector containing RNAi-S, RNAi-L and sGFPconstructs and resulting cells were used to trans-form coffee plants according to the method(Hatanaka et al., 1999) with modifications. Briefly,embryogenic tissues of C. arabica and somaticembryos of C. arabica and C. canephora were co-cultured with a bacterial suspension having anabsorbancy of approximately 0.5 at 600 nm for10 min in liquid modified 1/2MS medium con-taining plant growth regulator(s) and 50 mg l)1 ofacetosyringon. After aspiration of the bacterialsuspension, the infected tissues were transferredto induction-maintenance media containing300 mg l)1 of cefotaxim and 50 mg l)1 of hygro-mycin. They were then subcultured at 3-weekintervals onto fresh medium with the same com-position for approximately 2–4 months. Duringsubcultures, most infected tissues gradually turnedbrown and necrotic in part. However, transformedtissues survived and newly regenerated. Theresulting transformed coffee tissues and somaticembryos were finally selected on modified 1/2MSmedium containing 100 mg l)1 of hygromycin.Genomic DNA was extracted from hygromycinresistant material (cultured tissues, somatic em-bryos or leaves of somatic seedlings), and sub-jected to PCR analysis of the hygromycinphosphotransferase (HPT) gene as described(Hatanaka et al., 1999).
RT-PCR
Total RNA was prepared from embryogenic tis-sues or leaves of plantlets with an RNeasy PlantMini Kit (Qiagen, Germany) and subjected to RT-PCR using an RNA PCR Kit (AMV) Ver.2.1(Takara, Japan) with five sets of gene specificprimers; for CaXMT1, 50-ATCAACTGGTTCTCGCCAAG-30 and 50-CTGCTCTAACGGAAGATGCA-30; for CaMXMT1, 50-TCCTACAATC- TGGCTCTTGC-30 and 50-TTCATCGCCGTATACTTGGA-30; for CaMXMT2, 50-CCTACAATCTGGCTCTTGCC-30 and 50-TTCATCGCCGTATACTTGGA-30; for CaDXMT1, 50-TCATTCTACAATCTGTTTCTCATCAG-30 and50-TATGGAATTCGGGTTCTCGA-30; and fora-tubulin (AF363630), 50-GAGAGTGCATCTC
CATCCAT-30 and 50-CTGCTCAGGGTGGAAGAGT-30. PCR was carried out under the condi-tions of up to 35-cycle of denaturation at 94 �C for30 s, annealing at 58 �C for 30 s and extension at72 �C for 1 min. Aliquots were sampled at appro-priate cycles and, after fractionation on agarose gelelectrophoresis, visualized with ethidium bromidestaining. The amount of products was densito-metrically estimated and the optimal PCR cycle forexponential phase of amplification was determinedfor each gene.
Quantification of caffeine and theobromine
Purine alkaloids were extracted according to theprocedure developed by Ashihara et al. (1996a)with modification. Approximately 100 mg ofembryogenic cell tissues of C. arabica which weremaintained at 3-weeks intervals, and approxi-mately 10–20 mg of young leaves which werecollected 2–3 weeks after flushing from 1-year-oldplantlets of C. canephora, were boiled at 80 �Ctwice for 20 min each in a total of 2 ml ofultrapure water with homogenization. Afterresulting homogenates were centrifuged at10 000 · g for 10 min, supernatants were filteredthrough a 0.2 lm mesh filter (DISMIC-25cs;ADVANTEC, Japan) and diluted with ultrapurewater. The resulting 20-ll aliquots were analyzedby HPLC using a Puresil C18 column (4.5 mm-i.d. · 250 mm, Waters, USA) at 35 �C. The col-umn was developed at a flow-rate of 1 ml min)1
with 20% methanol solution using the multisol-vent delivery system (Waters). Caffeine andtheobromine in the samples were densitometri-cally monitored at 270 nm with a tunable absor-bance detector (Waters). Mean values werecalculated from at least six independent mea-surements per sample.
Results
Establishment of the tissue culture systemfor transformation
Rapid and efficient transformation of higherplants depends on an efficient tissue culture sys-tem, which must be individually optimized forspecies of interest. In order to establish such asystem for coffee plants, we first examined the
935

culture conditions for somatic embryogenesis.When a leaf section of C. arabica was incubatedon a modified half-strength Murashige and Skoogmedium (modified 1/2MS) containing 1 lMM N-(2-chloro-4-pyridyl)-N’-phenylurea (4-CPPU), newyellow-whitish tissues emerged around leaf cutedges after 10 days. After 30 days culture, how-ever, most initiated tissues with 1 lMM of 4-CPPUturned brown and necrotic in part. To avoid suchnecrosis, we examined the effects of 2,4-dichlo-rophenoxyacetic acid (2,4-D) and found that acombination of 5 lMM 2,4-D and 1 lMM 4-CPPUwas best for proliferation of yellow-whitishcompact tissue (Figure 2A). Upon maintenance3 weeks after transfer to this medium, theembryogenic tissues actively proliferated in asynchronous fashion. This culture condition wastherefore used throughout the present study. Forefficient plant regeneration, a direct somaticembryogenesis method was also adopted. Whenleaf cuttings were cultured on modified 1/2MSmedium containing 20 lMM 2ip, sections graduallyturned brown within 2–4 months, but somaticembryos were directly formed without producingcalli on the cut edges. The critical point for directsomatic embryogenesis of both C. arabica and C.canephora was transfer of browned leaf explantsto fresh medium at 3-week intervals. Secondarysomatic embryos were then directly formed fromthe epidermal tissues of the initial embryos,without callus formation, on medium containing20 lMM of 2ip (Figure 2B). These somatic embryoswere allowed to further develop by transfer ontomodified 1/2MS medium containing 5 lM of BAfor C. arabica and hormone-free for C. canepho-ra. They germinated frequently and developedinto somatic seedlings within an additional3–6 months (Figure 2C). In our experiments, C.canephora grew faster than C. arabica. Theregenerated small plantlets were transferred topots and naturally cultivated in a greenhouse(Figure 2D). Results clearly indicated that ourmethod is advantageous over so far publishedmethods (Hatanaka et al., 1991; van Boxtel andBerthouly, 1996) in terms of efficiency andreproducibility.
Transformation
In order to establish a method to stably transformand regenerate coffee plants, we performed a pilot
experiment with sGFP as the reporter gene, drivenby the cauliflower mosaic virus (CaMV) 35S pro-moter (Figure 3A). Somatic embryos from C.canephora prepared as described above weretransformed with a vector containing sGFP andHPT genes through Agrobacterium tumefaciensinfection, and allowed to grow to maturity.Expression of sGFP could be detected in all partsof regenerated somatic embryos (Figure 3B, C)and also in whole regenerated plants (Figure 3D–G), indicated that the introduced gene was stablyexpressed throughout the tissue.
Based on these findings, we next transformed asingle embryogenic cell line derived from a leafsection of one mother tree of C. arabica with thedsRNA sequence of CaMXMT1. Two constructswhich contained either a short RNA fragmentwith 139 and 161 nucleotides (designated RNAi-S)or a long RNA fragment with two identical se-quences of 332 nucleotides (designated RNAi-L)were prepared (Figure 3A). Both were driven bythe CaMV35S promoter. A preliminary transfor-mation using intron-GUS (iGUS) and RNAi-Linto C. arabica embryogenic tissues yielded fiveindependent cell lines (two for iGUS and three forRNAi-L) which showed similar growth and mor-phological characteristics as the non-transformedwild-type. After selection with hygromicin andconfirmation of the introduced HPT gene by PCR(data not shown), the tissues were allowed to ac-tively proliferate on fresh modified 1/2MS mediumcontaining 1 lMM 4-CPPU and 5 lMM 2,4-D.Transformation of C. canephora somatic embryoswith RNAi-L and RNAi-S was similarly per-formed.
Quantitative RT-PCR
For quantification of transcripts, the exponentialamplification phase of RT-PCR was determinedby drawing samples every few cycles using RNAsamples from wild-type and transgenic lines ofC. canephora, RNAi-L #4 and RNAi-S #4. Theamount of products was densitomerically deter-mined after gel electrophoresis and staining withethidium bromide (Figure 4A). Results clearlyindicated that amplification reached exponentialphase between 30 and 32 cycles for both controland transgenic samples, and that transcript levelsof both CaXMT1 and CaMXMT1 were lower intransgenic lines than in the control (Figure 4A).
936
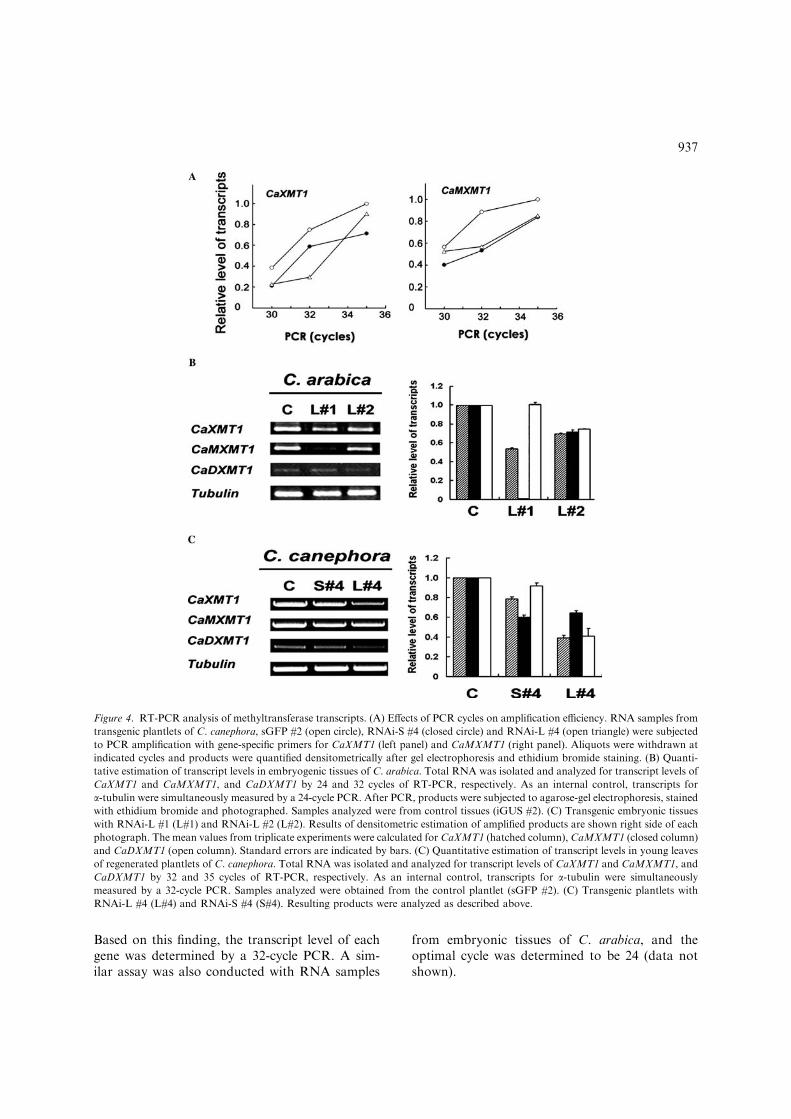
Based on this finding, the transcript level of eachgene was determined by a 32-cycle PCR. A sim-ilar assay was also conducted with RNA samples
from embryonic tissues of C. arabica, and theoptimal cycle was determined to be 24 (data notshown).
Figure 4. RT-PCR analysis of methyltransferase transcripts. (A) Effects of PCR cycles on amplification efficiency. RNA samples from
transgenic plantlets of C. canephora, sGFP #2 (open circle), RNAi-S #4 (closed circle) and RNAi-L #4 (open triangle) were subjected
to PCR amplification with gene-specific primers for CaXMT1 (left panel) and CaMXMT1 (right panel). Aliquots were withdrawn at
indicated cycles and products were quantified densitometrically after gel electrophoresis and ethidium bromide staining. (B) Quanti-
tative estimation of transcript levels in embryogenic tissues of C. arabica. Total RNA was isolated and analyzed for transcript levels of
CaXMT1 and CaMXMT1, and CaDXMT1 by 24 and 32 cycles of RT-PCR, respectively. As an internal control, transcripts for
a-tubulin were simultaneously measured by a 24-cycle PCR. After PCR, products were subjected to agarose-gel electrophoresis, stained
with ethidium bromide and photographed. Samples analyzed were from control tissues (iGUS #2). (C) Transgenic embryonic tissues
with RNAi-L #1 (L#1) and RNAi-L #2 (L#2). Results of densitometric estimation of amplified products are shown right side of each
photograph. The mean values from triplicate experiments were calculated for CaXMT1 (hatched column), CaMXMT1 (closed column)
and CaDXMT1 (open column). Standard errors are indicated by bars. (C) Quantitative estimation of transcript levels in young leaves
of regenerated plantlets of C. canephora. Total RNA was isolated and analyzed for transcript levels of CaXMT1 and CaMXMT1, and
CaDXMT1 by 32 and 35 cycles of RT-PCR, respectively. As an internal control, transcripts for a-tubulin were simultaneously
measured by a 32-cycle PCR. Samples analyzed were obtained from the control plantlet (sGFP #2). (C) Transgenic plantlets with
RNAi-L #4 (L#4) and RNAi-S #4 (S#4). Resulting products were analyzed as described above.
937

RNA interference in embryogenic tissuesof C. arabica
Among four stable transformed lines obtainedfrom C. arabica embryogenic tissues, two lineswere examined for the efficiency of RNAi by esti-mating the levels of CaMXMT1 transcripts (Fig-ure 4B). The RNAi-L #1 transformant displayed aclear reduction in CaMXMT1 transcripts, showingalmost no accumulation as estimated by densito-metric tracing. In addition, transcripts for Ca-XMT1 were reduced to 50% of the control level.The other line, RNAi-L #2 also exhibited decreasein both CaMXMT1 and CaXMT1 transcripts. Thetranscript level of CaDXMT1 was apparently notaffected in the RNAi-L #1 line but was reduced inthe RNAi-L #2 case. Transcripts for CaMXMT2were not detected in these lines even after 35 cyclesof PCR (data not shown).
The endogenous levels of theobromine andcaffeine in four lines were then determined usingHPLC. Control wild-type and iGUS tissues con-tained theobromine at approximately 90 lg g)1
fresh tissue, whereas values for transgenic linesRNAi-L #1 and RNAi-L #2 were only 13 and32 lg, respectively (Table 1), with reduction ratesof 85% and 65%, respectively. The caffeine con-tent in the controls (both non-transformed wild-type and transgenic iGUS lines) was 18 lg g)1
fresh tissue in average, but almost 0 and 3.7 lg inthe RNAi-L #1 and RNAi-L #2 lines, the reduc-tion being 100% and 80%, respectively (Table1).These results clearly indicated that the RNAisequence of CaMXMT1 sufficiently suppressed thecaffeine biosynthesis pathway, probably byaffecting not only the target CaMXMT1 but alsothe closely related CaXMT1. The results alsosuggested that the major pathway of caffeine syn-thesis involves theobromine.
Reduction of caffeine in plantlets
The same polypeptide (AF494414) as CaMXMT1was found to be present in C. canephora. Subse-quently, the effects of introduction of CaMXMT1-RNAi constructs into C. canephora were examined
Figure 5. Plantlets of C. canephora. Control (wild type, left),
and transgenic (RNAi-S #2, right) plantlets were grown in a
greenhouse for approximately 2 years (ca. 30 cm in height).
Table 1. Purine alkaloid contents in embryogenic tissues from
C. arabica. Endogenous theobromine and caffeine levels were
estimated in embryogenic tissues of wild type, transgenic
intron-GUS (iGUS), and transgenic RNAi C. arabica.
Lines Theobromine* Caffeine*
Control #1 (wild type #1) 91.5 � 4.3 27.3 � 2.8
Control #2 (wild type #2) 85.5 � 2.1 14.1 � 2.4
Control #3 (iGUS #1) 91.5 � 4.5 19.2 � 1.5
Control #4 (iGUS #2) 82.1 � 1.8 12.6 � 1.4
Average 87.7 � 3.2 18.3 � 2.0
RNAi-L #1 13.5 � 2.0 0.0 � 0.0
RNAi-L #2 32.0 � 2.7 3.7 � 0.3
RNAi-L #3 44.3 � 2.1 8.1 � 2.1
RNAi-L #4 73.1 � 1.9 10.6 � 0.4
Average 40.7 � 2.2 5.6 � 0.7
*Results are expressed as lg g)1 fresh tissue.
Table 2. Purine alkaloid contents in transgenic plantlets.
Endogenous theobromine and caffeine levels were estimated
in leaves of wild type, transgenic sGFP, and transgenic RNAi
plantlets of C. canephora.
Lines Theobromine* Caffeine*
Control #1 (wild type #1) 0.97 � 0.11 7.54 � 0.04
Control #2 (wild type #2) 0.93 � 0.04 8.82 � 0.22
Control #3 (sGFP #1) 1.11 � 0.02 7.73 � 0.05
Control #4 (sGFP #2) 0.90 � 0.09 9.66 � 0.21
Average 0.98 � 0.07 8.44 � 0.12
RNAi-S #1 0.30 � 0.05 4.07 � 0.11
RNAi-S #2 0.22 � 0.01 2.53 � 0.17
RNAi-S #3 0.29 � 0.01 4.88 � 0.02
RNAi-S #4 0.25 � 0.01 4.63 � 0.10
RNAi-S #5 0.31 � 0.01 4.35 � 0.06
Average 0.27 � 0.02 4.09 � 0.09
RNAi-L #1 0.66 � 0.01 2.77 � 0.06
RNAi-L #2 0.59 � 0.12 3.94 � 0.04
RNAi-L #3 0.58 � 0.08 4.70 � 0.23
RNAi-L #4 0.54 � 0.03 3.88 � 0.03
RNAi-L #5 0.47 � 0.03 3.94 � 0.04
Average 0.57 � 0.05 3.85 � 0.07
*Results are expressed as mg g)1 fresh tissue.
938

with five independent transgenic lines for RNAi-S,five for RNAi-L and two for sGFP. They weregrown to plantlets, and young leaves were col-lected 2–3 weeks after flushing. Two representativelines were selected and the transcript levels of thethree methyltransferase genes were examined byRT-PCR (Figure 4C). Results were essentially thesame as seen in embryogenic tissues of C. arabica,showing a clear reduction in transcripts not onlyfor CaMXMT1 but also for CaXMT1. Transcriptsfor CaDXMT1 were also reduced in intact leaftissues. In the case of RNAi-L #4 seedlings, forexample, the relative levels of CaXMT1, CaM-XMT1 and CaDXMT1 were 39%, 64% and 41%,respectively, of those of the control (sGFP #2) line(Figure 4C). These results suggested that a repre-sentative RNAi sequence for one key gene candown-regulate three distinct methylation steps ofthe caffeine biosynthetic pathway.
The contents of purine alkaloids were thenmeasured by HPLC. Wild-type and transgeniclines expressing sGFP equally contained endoge-nous theobromine at approximately 1 mg g)1 freshleaf tissue (Table 2). In contrast, five transgeniclines with RNAi-S contained an average of only0.27 mg (70% reduction), and those with RNAi-L0.57 mg (40% reduction), although the amountvaried depending on the individual line (Table 2).The caffeine content in the controls was approxi-mately 8.4 mg g)1 fresh leaf tissue, while that inboth RNAi-S and RNAi-L was 4.0 mg, showingan average of 50% reduction (Table 2). Theamount was again variable depending on the line,with a notable example (RNAi-S #2) containingonly 2.5 mg (70% reduction). The results showedthe RNAi method to also be effective in planta,although the suppression was not as complete asobserved in embryogenic tissues of C. arabica.
Discussion
In order to construct transgenic coffee plants withsuppressed caffeine synthesis, two problems had tobe solved: isolation of genes of interest; andestablishment of an efficient transformation andregeneration system. After intensive survey, wesuccessfully accomplished the former (Uefuji et al.,2003). For the latter, we focused on direct for-mation of somatic embryos without producingembryogenic tissues for transformation in the
present study, and established the method withhigh transformation efficiency and ubiquitousexpression of the marker gene (sGFP) in somaticembryos and whole regenerated plants. Usingthis regeneration method, we transformed bothC. arabica and C. canephora with CaMXMT1RNAi constructs. Resulting transgenic linesshowed a 50–85% reduction in theobromine con-tent in comparison with the control, indicatingefficient interference by RNAi method.
The present study cast light on two aspects ofcaffeine synthesis in coffee plants: first, the majorroute of biosynthesis; second, methodology forproduction of decaffeinated beans. The silencing ofthe second methylation step by RNAi was distinct.In randomly selected transgenic lines of bothC. arabica and C. canephora, expression of CaM-XMT1 transcripts was suppressed, although theeffect was variable with reduction to nearly 0%and to 80% the control value. An intriguing fea-ture is that expression of CaXMT1 was sup-pressed, showing a reduction to 50–80% that ofthe control. A similar suppression of CaDXMT1was also observed in the most cases, possibly dueto either direct interference by the CaMXMT1-RNAi construct or indirect effects on concertedregulation of caffeine synthesis. We currently favorthe former idea, since recent studies pointed toeffects of RNAi spreading from the initiator regioninto adjacent regions of the target gene, andamong genes whose sequences are closely related(Vaistij et al., 2002). The similarity among CaM-XMT1, CaXMT1 and CaDXMT1 is over 90% inthe coding region (Uefuji et al., 2003), suggestingthat the primary CaMXMT1 small double-stran-ded RNA progressively produces many secondarysmall double-stranded RNAs spanning its codingregion to the adjacent sequence of the initiatorregion, which in turn destroyed mRNAs forCaXMT1 and CaDXMT1.
The contents of theobromine and caffeine weremarkedly reduced in transgenic lines, by 50% and30%, respectively, in embryogenic tissues, and30–50% and 50% in plantlets. This reduction rateslightly differs from that expected from the tran-script levels for relevant genes. For example, inone line of embryogenic tissue (RNAi-L #1)CaMXMT1 transcripts proved undetectable, buttheobromine was still produced at 15% of thecontrol level. In contrast, two lines of plantlets(RNAi-S #4 and RNAi-L #4) exhibited rather
939

weak suppression in CaMXMT1 expression, toapproximately 60% of that in the controls, whiletheobromine contents were reduced by 75% and50 %, respectively. Several explanations are con-ceivable. For the former case, bypass pathwaysmight be activated. Since kinetic analyses sug-gested that caffeine synthase (DXMT) catalyzesboth theobromine and caffeine synthesis (Mizunoet al., 2003; Uefuji et al., 2003), and since thepresent RNAi construct did not interfere withCaDXMT1 in this particular line, it is possible thatCaDXMT1 substituted for silenced CaMXMT1,resulting in a production of theobromine at a lowlevel. For the latter case, it is highly possible thatRNAi synergistically suppressed theobrominesynthesis by reducing CaXMT1 that produces adirect precursor of CaMXMT1. A scarcity of7-methylxanthosine due to a reduction of Ca-XMT1 might greatly affect the production oftheobromine, even if CaMXMT1 was available toa certain extent.
Taken the results together, the present studywith both transgenic embryogenic tissues andregenerated plantlets clarified several importantfeatures of caffeine biosynthesis in coffee plants.First, RNAi appears to confer a global effect onexpression of relevant genes, although the primarytarget was clearly CaMXMT1. Second, reductionof the final product, caffeine, was consequently notdue to the suppression of a single enzyme, butrather due to a disturbance of the whole biosyn-thetic pathway. Third, it can safely be concludedthat theobromine is the major intermediate incaffeine biosynthesis, as its reduction was generallyproportional to a reduction of caffeine contents.
The present results provide evidence of suc-cessful production of a low-caffeine phenotype inboth ‘Arabica’ (C. arabica) and ‘Robusta’(C. canephora) coffee species, with 100% decaffe-ination in embryogenic tissues and 70% in plant-lets, respectively. At maturity, they may produceessentially normal coffee beans except for lowcaffeine content. However, technical improvementis certainly required before practical application soas to express the RNAi only in beans, and allowtargeting of multiple genes. In this context, con-struction of a limited expression system might beuseful, employing, for example, the promoter ofthe 11S seed storage protein gene of C. arabica(Acuna et al., 1999; Marraccini et al., 1999; Rogerset al., 1999).
Acknowledgements
The authors thank Mr. S. Kurokawa (The Botani-cal Gardens of Toyama, Japan) for the generousgift of plant materials, Drs. N. Koizumi and Y.Yamaguchi (Nara Institute of Science and Tech-nology), and Dr. T. Kusano (Tohoku University)for valuable suggestions, and Dr Malcolm Moore(Intermal, Nagoya) for critical reading of themanuscript, respectively. This work was supportedby grants from the Research for the Future Pro-gram (JSPS-RFTF00L01604) of the Japan Societyfor the Promotion of Science, and from the NewEnergy and Industrial Technology DevelopmentOrganization (NEDO), Japan.
References
Acuna, R., Bassuner, R., Beilinson, V., Cortina, H.,
Cadena-Gomez, G., Montes, V. and Nielsen, N.C. 1999.
Coffee seeds contain 11S storage proteins. Physiol. Plant 105:
122–131.
Ashihara, H., Monterio, A.M., Moritz, T., Gillies, F.M. and
Crozier, A. 1996a. Catabolism of caffeine and related purine
alkaloids in leaves of Coffea arabica L. Planta 198: 334–339.
Ashihara, H., Monterio, A.M., Gillies, F.M. and Crozier,
A. 1996b. Biosynthesis of caffeine in leaves of coffee. Plant
Physiol. 111: 747–753.
Ashihara, H. and Crozier, A. 2001. Caffeine: A well known but
little mentioned compound in plant science. Trends Plant Sci.
6: 407–413.
Ashihara, H. and Crozier, A. 1999. Biosynthesis and metabo-
lism of caffeine and related purine alkaloids in plants. Adv.
Bot. Res. 30: 118–205.
Hashimoto, T. and Yamada, Y. 2003. New genes in alkaloid
metabolism and transport. Curr. Opin. Biotech. 14: 163–168.
Hatanaka, T., Arakawa, O., Yasuda, T., Uchida, N. and
Yamaguchi, T. 1991. Effect of plant growth regulators on
somatic embryogenesis in leaf cultures of Coffea canephora.
Plant Cell Rep. 10: 179–182.
Hatanaka, T., Choi, Y.E., Kusano, T. and Sano, H. 1999.
Transgenic plants of coffee Coffea canephora from embryo-
genic callus via Agrobacterium tumefaciens-mediated trans-
formation. Plant Cell Rep. 19: 106–110.
Hood, E.E., Halmer, G.L., Fraley, R.T. and Chilton, M.D.
1986. The hypervirulence analysis of the Agrobacterium
tumefacience A281 is encoded in a region pTIB0542 outside
of T-DNA. J. Bacteriol. 168: 1291–1301.
Marraccini, P., Deshayes, A., Petiard, V. and Rogers, W.J.
1999. Molecular cloning of the complete 11S seed storage
protein gene of Coffea arabica and promoter analysis in
transgenic tobacco plants. Plant Physiol. Biochem. 37: 273–
282.
Mizuno, K., Okuda, A., Kato, M., Yoneyama, N.,
Tanaka, H., Ashihara, H. and Fujimura, T. 2003. Isola-
tion of a new dual-functional caffeine synthase gene
940

encoding an enzyme for the conversion of 7-methylxan-
thine to caffeine from coffee (Coffea arabica L.). FEBS
Lett. 534: 75–81.
Murashige, T. and Skoog, F. 1962. A revised medium for rapid
growth and bioassays with tabacco tissue cultures. Physiol.
Plant 15: 473–479.
Ogawa, M., Herai, Y., Koizumi, K., Kusano, T. and Sano,
H. 2001. 7-Methylxanthine methyltransferase of coffee
plants. J. Biol. Chem. 276: 8213–8218.
Ogita, S., Uefuji, H., Yamaguchi, Y., Koizumi, N. and Sano, H.
2003. Producing decaffeinated coffee plants. Nature 423: 823.
Ohta, S., Mita, S., Hattori, T. and Nakamura, K. 1990.
Construction and expression in tobacco of a b-glucuronidase(GUS) reporter gene containing an intron within the coding
sequence. Plant Cell Physiol. 31: 805–813.
Rogers, W.J., Bezard, G., Deshayes, A., Meyer, I., Petiard,
V. and Marraccini, P. 1999. Biochemical and molecular
characterization and expression of the 11S-type storage
protein from Coffea arabica endosperm. Plant Physiol.
Biochem. 37: 261–272.
Uefuji, H., Ogita, S., Yamaguchi, Y., Koizumi, N. and Sano,
H. 2003. Molecular cloning and functional characterization
of three distinct N-methyltransferases involved in the caffeine
biosynthetic pathway in coffee plants. Plant Physiol. 132:
372–380.
Vaistij, F.E., Jones, L. and Baulcombe, D.C. 2002. Spread-
ing of RNA targeting and DNA methylation in RNA
silencing requires transcription of the target gene and a
putative RNA-dependent RNA polymerase. Plant Cell 14:
857–867.
van Boxtel, J. and Berthouly, M. 1996. High frequency somatic
embryogenesis from coffee leaves. Plant Cell Tiss. Org. Cult.
44: 7–17.
941















![[A brief "update" on renal effects of caffeine.]](https://static.fdokumen.com/doc/165x107/630bdf00dffd330585081b05/a-brief-update-on-renal-effects-of-caffeine.jpg)




