Sensitizing protective tumor microenvironments to antibody-mediated therapy
Upload
khangminh22Category
view
0download
0
Antibody mediated immune response against
Flavivirus infection in West Australian
travellers
KRITU PANTA
Master of Science in Medical Microbiology
Tribhuvan University, Nepal
This thesis is presented for the degree of Doctor of Philosophy of The University of
Western Australia
School of Biomedical Sciences
2018

2
THESIS DECLARATION
I, Kritu Panta, certify that:
This thesis has been substantially accomplished during enrolment in the degree.
This thesis does not contain material which has been accepted for the award of any
other degree or diploma in my name, in any university or other tertiary institution.
No part of this work will, in the future, be used in a submission in my name, for any
other degree or diploma in any university or other tertiary institution without the prior
approval of The University of Western Australia and where applicable, any partner
institution responsible for the joint-award of this degree.
This thesis does not contain any material previously published or written by another
person, except where due reference has been made in the text.
The work(s) are not in any way a violation or infringement of any copyright, trademark,
patent, or other rights whatsoever of any person.
The research involving human data reported in this thesis was assessed and approved
by The University of Western Australia Human Research Ethics Committee.
Approval #: RA/4/1/5420
Written patient consent has been received and archived for the research involving
patient data reported in this thesis.
This thesis does not contain work that I have published, nor work under review for
publication.
Signature
Date: 22-06-2018

3
Abstract
This study assessed the immunological significance of Flavivirus diversity by analysing
longitudinal antibody-mediated immune responses in West Australian (WA) travellers
with dengue and Zika virus infection. The majority of study volunteers had never lived
in dengue and Zika endemic or epidemic countries and had well-defined monotypic
infection with DENV or ZIKV; some of them had received YFV vaccination. Dengue
and Zika do not occur in WA as the mosquito vectors are not present. Magnitude of
neutralising antibody, measured by focus reduction neutralisation test (FRNT); total
antibody, measured by haemagglutination inhibition (HI) test; as well as antibody
mediated enhancement (ADE) was assessed for antisera collected from individuals who
were sampled during acute phase (< 2months) and up to 6 years after presentation for
febrile illness. 35 antisera were tested against a collection of DENV isolates derived
from travellers presenting with febrile illness and who were diagnosed with DENV
infection; the virus collection includes representatives of all four serotypes collected
between 2010 – 2015. This collection is dependent on DENV circulating in the Asia
Pacific region at the time and thus is biased towards contemporaneous endemic or
epidemic isolates, predominantly DENV-1 and DENV-2. Phylogenetic analysis
identified genotypes and lineages, and representative isolates were used as targets.
Anti-DENV responses were highly cross-reactive during acute phase and neutralized
heterologous DENV at high magnitude. Over time and up to six years after infection
responses became increasingly serotype-specific - this could be visualised on antigenic
maps - however heterologous cross-neutralisation was maintained in all individuals to
some degree, even at 6 years, and magnitude of neutralisation was virus- and patient-
specific. Virus-specific neutralisation was not genotype or lineage dependent; individual
isolates were differentially neutralised by antisera from individuals collected at the

4
same time point after infection. Antigenic mapping showed serotype clustering could be
distinguished after 1 year. However, individual viruses were as close to heterologous
serotype clusters as they were to homologous clusters and this pattern persisted at 6
years. A second major observation was that the magnitude of responses to homologous
DENV in these monotypic travellers also declined over time, including responses to
autologous virus, and this was true for all 4 serotypes. The magnitude of ZIKV-specific
neutralising antibody responses declined over time in the small number (16) of study
volunteers with monotypic ZIKV infection. By 13 and 20 months after infection,
FRNT90 titres were below the limits of detection (<40 as per WHO criteria for
serodiagnosis of ZIKV infection). However, ZIKV-infected individuals with previous
flavivirus infection showed prolonged anti-ZIKV antibody responses up to 13 months
post onset of illness. This study demonstrated that antibody binding to ZIKV
capsid/prM protein can differentiate recent ZIKV infection in individuals with previous
DENV infection. Two ZIKV strains were assessed and showed a high degree of
differential cross-neutralisation, with high magnitude responses directed against the
prototype ZIKV MR766 isolate but not the contemporaneous ZIKV PRVABC59 in
acute phase monotypic DENV infection, highlighting the importance of strain selection
in ZIKV diagnostic and research approaches. ZIKV replication was enhanced by acute
phase (< 2months) antisera only.
Future work includes assessment of additional antisera against viruses which will
include those DENV that have circulated more recently in the region, including
serotypes 3 and 4 which have not circulated widely, and which were under-represented
in the present study; and assessment of the flavivirus-specific B cell repertoire.

5
Acknowledgements
I am incredibly grateful to my Principal Supervisor A/Prof Allison Imrie for her
guidance, patience and encouragement throughout my PhD. She has been a great
mentor constantly guiding me on how to progress my research and how to focus my
thinking so as to make logical and direct decisions. I am greatly indebted to her for the
efforts she has made to shaping me and my research throughout the past four years. I
sincerely thank all my co-supervisors: Clin/Prof. David Smith, Prof Nicholas De Klerk
and Dr. Alfred Tay for all their support and guidance during my PhD. I also want to
convey my gratitude to the late Professor Geoffrey Shellam for his support and
encouragement during the early stages of my PhD.
I am grateful to all my lab members for their constant support and encouragement
during times of success and failure in experiments. Thank you all for making my
Australian study experience wonderful. I am truly thankful to Suzi McCarthy for
teaching me laboratory skills and being there to listen. I want to thank Timo for his
constant support in the lab, and his family for all those Christmas and Easter lunches
making me feel like home. I wish to thank Alice and Harapan for all their help in the
lab. My list for lab friends would not complete without the late Peter Dunstan. I cherish
moments we had together, from first moment of learning lab skills, thanks Pete, you
were one of the most wonderful souls I have met. And finally Kara, my Aussie-
American best friend, thank you for our friendship and unflinching support and
encouragement during my PhD.
I thank Australia Awards Endeavour Postgraduate Fellowship for sponsoring my PhD
studies. I am also grateful to scholarship case managers for their constant support. I
want to thank all the study volunteers of this study for their gracious support. I wish to
thank the Serology department of PathWest Laboratory Medicine WA for supporting

6
laboratory procedures. I would also like to thank all the staff and PhD students of the
School of Biomedical Sciences for their support and little tips and tricks for
experiments.
Most importantly I am grateful to my hard-working mother for her undiminishing
support, love and prayers and for having faith in me. I still feel sorry I could not be
with you during the hardest times of the earthquake but I assure you all your sacrifice is
not wasted. I am also thankful to all my family members and friends back home in
Nepal and here in Perth. Words fail to express my gratitude to my husband Binit whose
constant encouragement, guidance and support throughout this journey has made it a lot
easier. Thank you Binit, for your faith in me, your constant support during all those ups
and downs during my PhD kept me going and thank you for making me smile even in
the worst situations.

7
Table of contents
Abstract ______________________________________________________________ 3
Acknowledgements _____________________________________________________ 5
List of Tables ________________________________________________________ 12
List of Figures _______________________________________________________ 14
List of abbreviations ___________________________________________________ 17
List of conference/meeting abstracts ______________________________________ 18
Chapter 1:- Thesis Introduction _________________________________________ 19
1.1. Overview of the study ___________________________________________ 19
1.2. Aims of study __________________________________________________ 20
1.3. Outline of chapters ______________________________________________ 21
Chapter 2: - Literature review ___________________________________________ 23
2.1.Preamble ______________________________________________________ 23
2.2. Flavivirus ______________________________________________________ 23 2.2.1. Virus structure ________________________________________________________ 25 2.2.2. Replication ___________________________________________________________ 29
2.3. Dengue Virus __________________________________________________ 32 2.3.1. Origin and distribution __________________________________________________ 32 2.3.2. Classification _________________________________________________________ 37 2.3.3. DENV Vector and Transmission __________________________________________ 42 2.3.4. Clinical spectrum of dengue disease _______________________________________ 45 2.3.5. Dengue case classification _______________________________________________ 47
2.4. Zika Virus ___________________________________________________ 49 2.4.1. Classification _________________________________________________________ 52 2.4.2. Vector borne ZIKV transmission __________________________________________ 53 2.4.3. Non- vector transmission of Zika _________________________________________ 54 2.4.4. Infection and complications ______________________________________________ 55
A) Guillian-Barre syndrome (GBS) ______________________________________________ 55 B) Neonatal ZIKV-related complications _________________________________________ 56
2.5. Flavivirus vaccine strains ______________________________________ 58 2.5.1. Yellow fever _________________________________________________________ 58 2.5.2. Japanese encephalitis ___________________________________________________ 59
2.6. Flavivirus immunopathogenesis _________________________________ 62 2.6.1. Flavivirus-specific innate immune responses ____________________________________ 62
A) Interferons _______________________________________________________________ 62 B) Complement ______________________________________________________________ 63
2.6.2. Acquired immune responses _________________________________________________ 65 A) Cellular immunity _________________________________________________________ 65 B) Humoral immunity _________________________________________________________ 67
i) Neutralising or protective antibody-mediated immunity ______________________ 70 ii) Cross reactive and enhancing antibody mediated immune response _____________ 71
2.7. Laboratory diagnosis of Flavivirus infection _______________________ 76 2.7.1. Virus isolation ________________________________________________________ 78 2.7.2. Genome detection _____________________________________________________ 79

8
2.7.3. Antigen detection ______________________________________________________ 80 2.7.4. Serology _____________________________________________________________ 81
2.8. Flavivirus infection in travellers _________________________________ 84 2.8.1. Dengue ______________________________________________________________ 84 2.8.2. Zika ________________________________________________________________ 85
Chapter 3:- Methods __________________________________________________ 87
3.1. Vero cells ______________________________________________________ 87
3.2. Virus amplification ______________________________________________ 87
3.3. Virus Titration _________________________________________________ 88 3.3.1. Determination of Tissue culture infectious dose __________________________________ 88 3.3.2. Determination of Focus forming units __________________________________________ 88
a. Preparation of monolayer ____________________________________________________ 88 b. Virus dilution _____________________________________________________________ 88 c. Test establishment __________________________________________________________ 88 d. Detection of FFU __________________________________________________________ 89
i. Optimisation of Fixative ________________________________________________ 89 ii. Optimisation of 4G2 monoclonal antibody concentration (primary antibody): _______ 89 iii. Optimising secondary antibody concentration ________________________________ 90 iv. Immunostaining _______________________________________________________ 90
3.3.3. Determination of Plaque forming units _________________________________________ 91
3.4. Methods for Chapters 5 and 6 _____________________________________ 92 3.4.1. Neutralising Antibody titre determination _______________________________________ 92
3.4.1.1. Preface ______________________________________________________________ 92 3.4.1.2. Establishment of neutralisation test ________________________________________ 94
a. Preparation of monolayer __________________________________________________ 94 b. Preparation of plasma sample ______________________________________________ 94 c. Focus reduction neutralisation test ___________________________________________ 94
i. Preparation of virus dilution ___________________________________________ 94 ii. Test establishment ___________________________________________________ 94 iii. Immunostaining and visualisation of FFU ________________________________ 95 iv. Plaque reduction neutralisation test ______________________________________ 95
d. Neutralisation titre determination ___________________________________________ 95 3.4.2. Hemagglutination inhibition antibody titre determination __________________________ 99
3.4.2.1 Principle _____________________________________________________________ 99 3.4.2.2. Methodology to establish HI test _________________________________________ 100
a. Preparation of viral antigen _____________________________________________ 100 b. Titration of inactivated virus ____________________________________________ 100 c. Preparation of plasma sample ___________________________________________ 100 d. Establishment of test __________________________________________________ 101
3.4.3. Mapping antigenic diversity using cartography _________________________________ 102 3.4.3.1. Preface _____________________________________________________________ 103 3.4.3.2. Cartography technique _________________________________________________ 103
3.4.4. Western blot ____________________________________________________________ 105 3.4.4.1. Preface _____________________________________________________________ 105 3.4.4.2. Methods ____________________________________________________________ 105
a. Optimisation of whole virus protein preparation _______________________________ 105 b. Quantification of Protein _________________________________________________ 107 c. Optimization of primary antibody concentration _______________________________ 107 d. SDS PAGE and Western blot ______________________________________________ 107 e. Detection of viral proteins ________________________________________________ 108
3.5. Method for chapter 7: - Antibody dependent enhancement ___________ 110 a. Cell lines ________________________________________________________________ 110 b. Virus stock and plasma samples ______________________________________________ 110

9
c. Establishment of test _______________________________________________________ 111 d. Detection of infectious units and determination of enhancement of infection. ___________ 111
Chapter 4: Study population and viruses _________________________________ 114
4.1. Preamble _____________________________________________________ 114
4.2. The cohort ____________________________________________________ 114 a. DENV-1 sera panel _____________________________________________________ 116 b. DENV-2 sera panel _____________________________________________________ 117 c. DENV-3 sera panel _____________________________________________________ 117 d. DENV-4 sera panel _____________________________________________________ 117 e. ZIKV sera panel ________________________________________________________ 118
4.2. Viruses used in this study _______________________________________ 124 4.2.1 DENV Panel _____________________________________________________________ 124
a. DENV-1 ________________________________________________________________ 127 b. DENV-2 ________________________________________________________________ 130 c. DENV-3 ________________________________________________________________ 133 d. DENV-4 ________________________________________________________________ 136 e. Autologous DENV ________________________________________________________ 139
4.2.2. ZIKV Panel _____________________________________________________________ 139 4.2.3. Flavivirus vaccine strains __________________________________________________ 142
a. Japanese encephalitis vaccine ______________________________________________ 142 b. Yellow fever vaccine strain _________________________________________________ 142
Chapter 5:- DENV-specific antibody responses in monotypic infection _________ 144
5.1. Preamble _____________________________________________________ 144
5.2. Introduction __________________________________________________ 144
5.3. Aims _________________________________________________________ 146
5.4. Results _______________________________________________________ 146 5.4.1. Neutralising antibody _____________________________________________________ 146
a. Specificity and cross-reactivity post DENV-1 infection _________________________ 148 i. Anti-DENV-1 antisera differentially neutralize DENV-1 ______________________ 156 ii. Neutralization among homologous DENV-1 decreases over time _______________ 158 iii. Cross-neutralisation against heterologous DENV ____________________________ 161 iv. Longitudinal decrease in heterologous DENV cross-neutralization by anti-DENV-1
antisera _________________________________________________________________ 164 v. Persistent neutralisation against autologous virus ____________________________ 166 vi. Cross-neutralization of ZIKV, YF17D and IMOJEV by DENV-1 antisera ________ 168
b. Specificity and cross-reactivity of anti-DENV-2 antibody _______________________ 170 i. Anti-DENV-2 neutralisation specificity against homologous serotype ____________ 177 ii. DENV-2 homologous NT decreased with time post infection___________________ 180 iii. Cross-neutralisation against heterologous DENV ____________________________ 182 iv. Decreased cross-neutralisation over time___________________________________ 185 v. Neutralisation against autologous DENV-2 _________________________________ 187 vi. Neutralisation against ZIKV, YF17D and IMOJEV. __________________________ 189
c. Neutralising responses post DENV-3 infection ________________________________ 191 i. Neutralisation among homologous DENV-3 ________________________________ 197 ii. DENV-3 neutralisation over time ________________________________________ 199 iii. Neutralisation against autologous serotype _________________________________ 205 iv. Neutralisation against related serogroups __________________________________ 205
d. Specificity of neutralising antibody response post DENV-4 infection _______________ 207 i. Longitudinal DENV-4 antisera against homologous virus _____________________ 214 ii. Heterologous cross-neutralisation by anti-DENV-4 antisera ____________________ 218 iii. Neutralisation against autologous serotype _________________________________ 218 iv. Neutralisation against related serogroups __________________________________ 218

10
5.4.2. Magnitude of neutralising antibody over time ________________________________ 221 5.4.3. Summary of neutralisation in DENV monotypic infection ______________________ 224 5.4.4. Total anti-DENV antibody: _______________________________________________ 225
a. Total antibody responses, DENV-1 infection _________________________________ 226 b. Total antibody responses, DENV-2 infection _________________________________ 230 c. Total antibody response post DENV-3 infection _______________________________ 235 d. Total antibody responses post DENV-4 infection ______________________________ 239
5.4.5. Summary of total antibody in DENV monotypic infection ______________________ 242 5.4.6. Longitudinal variation of neutralising and recognising anti-DENV antibody _______ 243
a. Cross-reactive anti-DENV responses within 2 months post infection _______________ 245 b. Anti-DENV antibody responses 2-12 months post infection ______________________ 247 c. Anti-DENV antibody responses 1-2 years post infection_________________________ 249 d. Anti-DENV antibody responses 2-4 years post infection_________________________ 251 e. Anti-DENV antibody responses 4 years post infection __________________________ 251
5.5. Summary discussion ____________________________________________ 254 5.5.1. Neutralisation endpoint stringency and increased specificity _______________________ 254 5.5.2. Anti-DENV antibody responses are virus- and time-dependent _____________________ 255
5.6. Research outcomes _____________________________________________ 256
Chapter 6: Antibody immune response post ZIKV infection __________________ 258
6.1 Introduction ___________________________________________________ 258
6.2. Aims of study _________________________________________________ 261
6.3. Results _______________________________________________________ 262 6.3.1. Neutralising antibody response ______________________________________________ 262 a. Within 1 month ___________________________________________________________ 267 b. 1 – 12 months post presentation ______________________________________________ 268 c. More than 12 months post presentation ________________________________________ 269
6.3.2. Total anti-ZIKV antibody _____________________________________ 274
6.3.3. Immunoblot analysis, ZIKV and DENV __________________________ 276 6.3.3.1. Positive and negative control ______________________________________________ 277 6.3.3.2. Anti-ZIKV antisera ______________________________________________________ 277 6.3.3.3. Anti-ZIKV/DENV antisera _______________________________________________ 278 6.3.3.4. Anti- ZIKV/YF17D _____________________________________________________ 278 6.3.3.5. Anti-DENV ___________________________________________________________ 278
6.4. Summary discussion ____________________________________________ 285 6.4.1. Anti-ZIKV specificity and cross-reactivity _____________________________________ 286 6.4.2. Anti-ZIKV antisera cross-neutralisation of DENV is time- and flavivirus background-
dependent ___________________________________________________________________ 288 6.4.3. Persistence of anti-ZIKV NAb in individuals with previous flavivirus infection ________ 289 6.4.4. Differential ZIKV MR766 and anti-ZIKV PRVABC59 neutralisation ________________ 289 6.4.5. Virus protein binding analysis by immunoblot __________________________________ 291
6.5. Research outcomes _____________________________________________ 293
Chapter 7: - Enhancement of ZIKV infection by monotypic anti-DENV antibody 294
7.1. Preamble _____________________________________________________ 294
7.2 Introduction ___________________________________________________ 294 7.2.1. Mechanism of ADE _______________________________________________________ 295 7.2.2. ADE in DENV and ZIKV infection __________________________________________ 295
7.3. Results _______________________________________________________ 299 7.3.1. Acute phase plasma: DENV plasma sample within two months post infection _________ 302

11
7.3.2. ZIKV enhancement by convalescent anti-DENV antisera collected 1-6 years post infection
____________________________________________________________________________ 303 7. 3.3. DENV-1 plasma samples __________________________________________________ 303 7.3.4. DENV-3 plasma samples __________________________________________________ 304 7.3.5. Pooled sera from DENV-1 and DENV-3 patients ________________________________ 304
a) At six months post infection _______________________________________________ 304 b) At 12 months post infection _______________________________________________ 304
7.4. Summary discussion ____________________________________________ 310 7.4.1. DENV and ZIKV in French Polynesia ________________________________________ 310 7.4.2. ZIKV enhancement by monotypic anti-DENV from travellers ______________________ 311
7.5. Research outcomes _____________________________________________ 313
Chapter 8:- Summary and conclusion ___________________________________ 314
Chapter 9:- References _______________________________________________ 317
Appendixes _________________________________________________________ 338

12
List of Tables
Table 2.1:- Assigning clades and clusters to flaviviruses reported by Kuno et al (8) .... 24
Table 2.2:- Flavivirus genes and their functions (12) ..................................................... 27
Table 2.3: Classification of DENV serotypes ................................................................. 40
Table 4. 1:- Demographic and laboratory findings of individuals with DENV-1
infection ........................................................................................................................ 119
Table 4. 2:- Demographic and laboratory findings of individuals with DENV -2
infection ........................................................................................................................ 120
Table 4. 3:- Demographic and laboratory findings of individuals with DENV -3
infection ........................................................................................................................ 121
Table 4. 4:- Demographic and laboratory findings of individuals with DENV -4
infection ........................................................................................................................ 122
Table 4. 5:- Demographic and Laboratory findings of individuals with ZIKV infection
....................................................................................................................................... 123
Table 4. 6:- Flaviviruses used in study ......................................................................... 126
Table 4. 7:- DENV-1 amino acid variation ................................................................... 129
Table 4. 8:- DENV-2 amino acid variation ................................................................... 132
Table 4. 9:- DENV-3 amino acid variation ................................................................... 135
Table 4. 10:- DENV-4 amino acid variation ................................................................. 138
Table 4. 11:- ZIKV amino acid variation ...................................................................... 141
Table 4. 1:- Demographic and laboratory findings of individuals with DENV-1
infection ........................................................................................................................ 150
Table 5. 1:- Anti-DENV-1 neutralising antibody titre at 50% reduction ..................... 152
Table 5. 2:- Anti-DENV-1 neutralising antibody titre at 75% reduction ..................... 153
Table 5. 3:-Anti-DENV-1 neutralising antibody titre at 90% ....................................... 154
Table 5. 4:- DENV-1-Sera panel statistical verification ............................................... 155
Table 4. 2:- Demographic and laboratory findings of individual with DENV -2
infection ........................................................................................................................ 171
Table 5. 5:- Anti-DENV-2 neutralising antibody titre at 50% reduction ..................... 173
Table 5. 6:- Anti-DENV-2 neutralising antibody titre at 75% reduction ..................... 174
Table 5. 7:- Anti-DENV-2 neutralising antibody titre at 90% reduction ..................... 175
Table 5. 8:- DENV-2-Sera panel statistical verification ............................................... 176
Table 4. 3:- Demographic and laboratory findings of individuals with DENV -3
infection ........................................................................................................................ 192
Table 5. 9:- Anti-DENV-3 neutralising antibody titre at 50% reduction..................... 193
Table 5. 10:-Anti-DENV-3 neutralising antibody titre at 75% reduction..................... 194
Table 5. 11:- Anti-DENV-3 neutralising antibody titre at 90% reduction.................... 195
Table 5. 12:- DENV-3-Sera panel statistical verification ............................................. 196
Table 4. 4:- Demographic and laboratory findings of individuals with DENV -4
infection ........................................................................................................................ 208
Table 5. 12:- Anti-DENV-4 neutralising antibody titre, 50% reduction ...................... 210
Table 5. 13:- Anti-DENV-4 neutralising antibody titre, 75% reduction ...................... 211
Table 5. 16:-DENV-4-Sera panel statistical verification .............................................. 212
Table 5. 14:- Anti-DENV-4 neutralising antibody titre, 90% reduction ...................... 212

13
Table 5. 15:- Total antibody response in DENV-1 sera panel ...................................... 227
Table 5. 18:- Total antibody response DENV-2 sera panel .......................................... 232
Table 5. 19:- Total antibody response of DENV-3 sera panel ...................................... 236
Table 5. 20:- Total antibody response in DENV-4 sera panel ...................................... 240
Table 4. 5: - Demographic and laboratory findings of individuals with ZIKV infection
....................................................................................................................................... 264
Table 6. 1:- Neutralising antibody response in ZIKV ................................................... 265
Table 6. 2: - Total antibody titre against ZIKV ............................................................ 275
Table 7. 1:- Description of plasma samples used in ADE ............................................ 300
Table 7. 2:- Virus output from U937 post exposure to respective anti-DENV - ZIKV
immune complex ........................................................................................................... 301

14
List of Figures
Figure 2.1:- Flavivirus Structure ..................................................................................... 28
Figure 2.2:- Flavivirus Replication ................................................................................. 31
Figure 2.3:-Dengue distribution in 2008 and 2016 ......................................................... 36
Figure 2.4:- DENV Phylogeny........................................................................................ 41
Figure 2.5:-DENV transmission cycle and mosquito-vectors (9) ................................... 42
Figure 2.6: Dengue clinical manifestations (World Health Organization 2009) ............ 46
Figure 2.8:- WHO Classification of dengue infection (2009) ........................................ 48
Figure 2.7:- WHO Classification of dengue infection (1997) ........................................ 48
Figure 2.9: -Zika virus spread from 1947-2016 ............................................................. 51
Figure 2.10:-The rise and fall of Zika ............................................................................. 51
Figure 2.11:- ZIKV Phylogeny ....................................................................................... 53
Figure 2.12- Immunological response during Flavivirus infection................................. 69
Figure 2.13:- Antibody threshold determining immune response in Flavivirus infection
......................................................................................................................................... 74
Figure 2. 14:- Adaptive immune responses in Flavivirus infection. ............................... 75
Figure 2.15:- Course of dengue infection and timing of diagnosis tests (7) .................. 77
Figure 2.16 :-Diagnosis of dengue. ................................................................................. 78
Figure 3.1: - Neutralisation ............................................................................................. 93
Figure 3. 2:- Establishment of neutralisation .................................................................. 96
Figure 3. 3:- Immunostaining and visualisation of FFU ................................................. 97
Figure 3. 4:- FRNT and PRNT ....................................................................................... 98
Figure 3. 5:- Hemagglutination of GRBC by flavivirus ................................................. 99
Figure 3. 6:- Hemagglutination inhibition by anti-flavivirus antibody ........................... 99
Figure 3. 7:- Measurement of total antibody by hemagglutination inhibition test ...... 102
Figure 3. 8:- Antigenic mapping strategy ..................................................................... 104
Figure 3. 9:- Comparison of protein lysate preparation methods ................................. 106
Figure 3. 10: Flow chart of Western blot analysis ........................................................ 109
Figure 3. 11:- Measurement of antibody-dependent enhancement of ZIKV infection by
anti-DENV antibody ..................................................................................................... 112
Figure 3. 12:- Viral load in the presence and absence of anti-DENV antibody ........... 113
Figure 4. 1:- Traveller cohort establishment and virus isolation. ................................. 115
Figure 4. 2:- Phylogenetic tree of DENV E-sequences including isolates from travellers
(11). ............................................................................................................................... 125
Figure 4. 3:- DENV-1 E- gene (1-394) alignment ........................................................ 128
Figure 4. 4:- DENV-2 E-gene (1-394) alignment ........................................................ 131
Figure 4. 5:- DENV-3 E-gene (1-394) alignment ......................................................... 134
Figure 4. 6:- DENV-4 E-gene (1-394) alignment ......................................................... 137
Figure 4. 7:- ZIKV E-gene alignment ........................................................................... 140
Figure 4. 8:- Phylogenetic tree showing the genetic relationship among the flaviviruses
strains used in this study. .............................................................................................. 143
Figure 5. 1:- Neutralising antibody response post DENV-1 infection, patient FLV-011
....................................................................................................................................... 151
Figure 5. 2:- Homologous strain neutralisation by DENV-1 antisera........................... 157
Figure 5. 3:- Differential neutralisation among DENV-1 homologous strains, patient
FLV-011 ........................................................................................................................ 159
Figure 5. 4:-Longitudinal variation of anti-DENV-1 neutralising antibody ................. 160

15
Figure 5. 5:- Heterologous cross-neutralisation by anti-DENV-1 antibody ................. 163
Figure 5. 6:- Longitudinal heterologous cross-neutralisation by anti-DENV-1 antibody
....................................................................................................................................... 165
Figure 5. 7:- Autologous, homologous and heterologous neutralisation ...................... 167
Figure 5. 8:- Cross-neutralisation across flavivirus serogroup ..................................... 169
Figure 5. 9:- Neutralising antibody response by anti-DENV-2 antisera, patient FLV -
014 ................................................................................................................................. 172
Figure 5. 10:- Homologous strain neutralisation by anti-DENV-2 antibody ............... 179
Figure 5. 11:- DENV-2-specific neutralisation over time, patient FLV 014 ................ 181
Figure 5. 12:- Variation of homologous strain neutralisation by anti-DENV-2. .......... 183
Figure 5. 13:- Heterologous cross-neutralisation by DENV-2 antisera ........................ 184
Figure 5. 14:- Decrease in heterologous neutralisation time post infection .................. 186
Figure 5.15:- Neutralisation against DENV-2 autologous strains ................................ 188
Figure 5. 16:- Longitudinal cross-neutralisation of flavivirus serogroups by anti-DENV-
2 antibody ...................................................................................................................... 190
Figure 5.17:- Homologous strain neutralisation by anti-DENV-3 antisera. ................. 198
Figure 5. 18:- Strain specificity over time by DENV-3 sera panel, patient FLV-038. . 200
Figure 5.19:- Change in homologous neutralisation over time, anti-DENV-3 antibody
....................................................................................................................................... 201
Figure 5. 20:- Heterologous cross-neutralisation by DENV-3 antisera ........................ 203
Figure 5. 21:- Heterologous neutralisation over time by DENV-3 antisera. ................ 204
Figure 5.22:- Autologous strain neutralisation by DENV-3 antisera ........................... 206
Figure 5.23:- Homologous and heterologous DENV neutralisation by anti-DENV-4
antibody, patient FLV 049 ............................................................................................ 209
Figure 5. 24:- Homologous virus neutralisation by DENV-4 antisera ........................ 215
Figure 5. 25:- Homologous strain neutralisation by DENV-4, patient FLV049........... 216
Figure 5. 26:- Variation in homologous strain neutralisation by DENV-4 antisera over
time. .............................................................................................................................. 217
Figure 5. 27: - Heterologous strain neutralisation by anti-DENV-4 antisera ............... 219
Figure 5. 28: - Heterologous neutralisation over time by anti-DENV-4 antisera. ........ 220
Figure 5. 29:- Magnitude of anti-DENV within 1 year of infection ............................. 223
Figure 5. 30:- Magnitude anti-DENV 1-2 years post infection .................................... 223
Figure 5.31:- Magnitude of anti-DENV 3-6 years post of infection ............................ 223
Figure 5.32:- Summary of homologous and heterologous NAb responses .................. 224
Figure 5. 33:- Total antibody responses against homologous DENV-1 ....................... 228
Figure 5. 34: - Total antibody responses, anti-DENV-1 antisera against heterologous
DENV ........................................................................................................................... 229
Figure 5. 35:- Total antibody responses, Anti-DENV-2 antisera against homologous
DENV-2 ........................................................................................................................ 233
Figure 5. 36:- Total antibody responses, anti-DENV-2 antisera against heterologous
DENV. .......................................................................................................................... 234
Figure 5. 37: - Total DENV-3 antibody response against homologous DENV ............ 237
Figure 5. 38:- Total antibody responses, anti-DENV-3 antisera against heterologous
DENV ........................................................................................................................... 238
Figure 5.39: - Total DENV-4 antibody against homologous DENV ............................ 241
Figure 5.40: - Total antibody responses, anti-DENV-4 antisera against heterologous
DENV ........................................................................................................................... 242

16
Figure 5. 41: - Summary of homologous and heterologous anti-DENV TAb .............. 243
Figure 5. 42:- Anti-DENV response and antigenic diversity at less than two months post
infection ........................................................................................................................ 246
Figure 5. 43:- Anti-DENV antibody response with a year of infection ........................ 248
Figure 5. 44:- Specificity of anti-DENV antibody 1-2-year post infection .................. 250
Figure 5. 45:- Anti-DENV-4 antibody at 2-4 years post infection ............................... 252
Figure 5. 46:- Anti-DENV-4 antibody 4 years post infection....................................... 253
Figure 6. 1:- Neutralising antibody response post ZIKV infection. ............................. 266
Figure 6. 2:- ZIKV neutralisation within 30 days post presentation of illness. ............ 270
Figure 6. 3:- ZIKV neutralisation 1-12 months post presentation of illness. ............... 271
Figure 6. 4:- ZIKV neutralisation more than 12 months post presentation of illness. .. 272
Figure 6. 5:- Variation in ZIKV neutralising antibody concentration over time based on
the flavivirus background ............................................................................................. 273
Figure 6. 6:- Coomassie stain. ....................................................................................... 280
Figure 6. 7:- Positive and Negative control .................................................................. 281
Figure 6. 8:- Monotypic anti-ZIKV antisera (FLV-064/FLV090) ................................ 282
Figure 6. 9:- Anti-ZIKV/DENV antisera (FLV-087).................................................... 283
Figure 6.10:- Anti-ZIKV/YFV antisera (FLV-032) ...................................................... 284
Figure 6. 11:- Anti-DENV antisera(FLV002/FLV030) ................................................ 285
Figure 7. 1:-ZIKV enhancement by monotypic anti-DENV pool at different stage post
infection. ....................................................................................................................... 306
Figure 7. 2:-ZIKV enhancement by anti-DENV-1 antisera .......................................... 307
Figure 7. 3:- ZIKV enhancement by anti-DENV-3 antisera ......................................... 308
Figure 7. 4 :-ZIKV enhancement based on French Polynesian co-circulation model. . 309

17
List of abbreviations
% percent
/ per
µl Microliter
ADE Antibody dependent enhancement
CPE Cytopathic effect
DENV Dengue Virus
DMEM Dulbecco's Modified Eagle's Medium
E Envelope protein
ELISA Enzyme linked immunosorbent assay
FBS Fetal bovine serum
FFU Focus forming units
FRNT Focus reduction neutralization test
HA Hemagglutination assay
HI Hemagglutination inhibition
IgG Immunoglobulin G
IgM Immunoglobulin M
JE IMOJEV Japanese encephalitis vaccine strain
mAb Monoclonal antibody
NAb Neutralizing antibody
NT Neutralization titer
PBS Phosphate buffer solution
PCR Polymerase chain reaction
PFU Plaque forming units
prM Pre-membrane protein
PRNT Plaque reduction neutralization test
RPMI 1640 Roswell Park Memorial Institute 1640 medium
SDS-PAGE Sodium dodecyl sulphate polyacrylamide gel electrophoresis
TAb Total antibody
WHO World Health Organization
YF17D Yellow fever 17D vaccine strain
ZIKV Zika Virus

18
List of conference/meeting abstracts
“Using cartography to define antigenic relationship among dengue viruses
(DENV) imported by travellers. American Society of Tropical Medicine and
Hygiene (ASTMH) 66th
Annual Meeting, Baltimore, United States of America,
2017.
“Using cartography to define antigenic relationship among dengue viruses
(DENV) imported by travellers”. Combined Biological Sciences Meeting
(CBSM), Perth, Australia, 2017.
“Antigenic mapping among imported Dengue virus in Western Australia”.
Centre for Research Excellence in Emerging Infectious Diseases (CREID)
Annual Colloquium, Sydney, Australia, 2017.
“Zika virus neutralizing antibodies in West Australian Travelers”. Australian
society of Microbiology (ASM) Annual Scientific Meeting, Perth, Australia,
2016.
“Illustrating antigen specific B-cell clonal expansion using RNA-seq”. West
Australian Society of Medical Research WA (ASMR-WA) symposium, Perth,
Australia, 2015.
“Illustrating antigen specific B-cell clonal expansion using RNA-seq” Science
on Swan Meeting, Perth, Australia, 2015.

19
Chapter 1:- Thesis Introduction
1.1. Overview of the study
This thesis is presented as a series of chapters that include the study of antibody
mediated immune responses elicited following dengue (DENV) and Zika (ZIKV) virus
infection.
Anti-DENV and anti-ZIKV humoral responses were studied in a group of travellers
who were infected during travel to endemic and epidemic countries. DENV and ZIKV
were strains isolated from viremic travellers; prototype strains; and representatives of
virulent and non-virulent lineages. Neutralising and total antibody responses were
studied. Cartographic maps were created to analyse antigenic relationships among
viruses and antisera. Furthermore cross-reactivity among flavivirus serogroups was also
studied.
Cross neutralisation among strains of ZIKV, cross-reactivity among DENV strains and
the flavivirus vaccine viruses YF17D and IMOJEV was studied. Identification of
antibody recognition site on viral protein was undertaken by Western immunoblot
analysis. Antibody-mediated enhancement of ZIKV in the presence of non-neutralising
concentrations of anti-DENV antibody was also studied.
Overall this thesis is an analysis of the different mechanism of antibody-mediated
immune response in monotypic and multitypic DENV and ZIKV infection among
travellers and how these responses change over time.

20
1.2. Aims of study
The main aim of the study is to understand of antibody mediated immune responses to
flavivirus infection in West Australian travellers, the great majority of whom present
with monotypic infection.
Specific Aims
1. To understand longitudinal variation of anti-DENV antibody responses against
contemporaneous DENV circulating in the Asia Pacific region, in a group of
travellers with monotypic infection
2. To determine specificity and cross-reactivity of anti-ZIKV antibody responses
against DENV and flavivirus vaccine viruses and to assess persistence of anti-ZIKV
immunity over time
3. To identify a marker that may be used to differentiate ZIKV and DENV infection
4. To determine ZIKV enhancement by anti-DENV antibody

21
1.3. Outline of chapters
Chapter 2 is a review of the literature on flaviviruses and immunopathogenesis of
flavivirus infection. The chapter begins with studies on flavivirus genus, structure and
replication and discusses dengue, Zika, and the yellow fever and Japanese encephalitis
vaccine viruses. The second section of the chapter reviews immunological responses
elicited by infection, with emphasis on the antibody-mediated immune response. The
final section reviews traveller-associated flavivirus infection and risk.
Chapter 3 details experimental methods used in the study. This chapter includes details
on optimization and establishment of tests used to derive DENV- and ZIKV-specific
antibody-mediated immune response.
The study population and viruses used in this study are described in Chapter 4. This
chapter includes demographic information and clinical history as well as genetic
characterisation of the viruses used in this study.
Chapter 5 defines the antibody-mediated immune response in monotypic dengue virus
infection. This chapter includes analysis of neutralising and total antibody responses
over time, against genetically diverse strains of DENV and other flaviviruses.
Furthermore, this chapter applies antigenic cartography to illustrate antigenic
relationships among cohort antisera and contemporaneous DENV.
In chapter 6, antibody-mediated immune responses among travellers with confirmed
and probable Zika virus infection was studied. This work identifies differential
neutralisation among patients with monotypic ZIKV infection, and patients with a
background of DENV infection and/or flavivirus vaccination. Differential neutralisation

22
among ZIKV strains and other flaviviruses is included in this chapter. Furthermore, this
chapter seeks to identify specific anti-ZIKV antibodies by westernblot.
Chapter 7 includes an assessment of enhancement of ZIKV in the presence of anti-
DENV antibody.
Chapter 8 summarize major findings of the study. In addition, this chapter outlines
implications of the work described in this thesis, and work that is being undertaken to
extend the findings of these studies.

23
Chapter 2: - Literature review
2.1. Preamble
This chapter provides an overview of the literature on flaviviruses and anti-flavivirus-
specific immune response. It provides background information on the Flavivirus genus
and focuses on dengue virus (DENV), Zika virus (ZIKV) and the vaccine strains of
yellow fever (YF17D) and Japanese encephalitis (IMOJEV). Available literature on
DENV and ZIKV with emphasis on viral genetic diversity and host immunological
responses was reviewed.
2.2. Flavivirus
The Flavivirus genus belongs to family Flaviviridae. It is a group that includes more
than 70 arthropod-borne viruses. Phylogenetically, these viruses are classified into
genotypes, clades, lineages and species; serological criteria divide the Flavivirus genus
into antigenic complexes (14).
Kuno et al. reported 14 clades based on conserved flavivirus genetic sequence and three
clusters based on vector associations (15). Gould et al described four
phylogenetic/ecological flavivirus groups: - two mosquito-borne, a tick-borne group,
and non-vectored viruses (16). In 1988, 68 then-recognised viruses were classified into
eight antigenic complexes: - tick-borne encephalitis (12 viruses), Rio Bravo (six),
Japanese encephalitis (10), Tyuleniy (three), Ntaya (five), Uganda S (four), dengue
(four) and Modoc (five) (15, 17). It is still uncertain which group is the oldest (18). The
most widely used classification of flaviviruses is clade and antigenic complex
classification by Kuno et al in 1998 as summarised in Table 2.1

24
Viruses in boldface belong to the corresponding antigenic complex to the right.
Viruses highlighted in grey are the flaviviruses used in this study
Refer Kuno et al for detail
Table 2.1:- Assigning clades and clusters to flaviviruses reported by Kuno et al (8)

25
2.2.1. Virus structure
Flaviviruses are ~500Å icosahedral asymmetric particles bearing ss-positive sense
RNA. Viral genomes code for single polypeptides that form structural and non-
structural proteins. Structure is formed by stoichiometric arrangement of three structural
proteins: core/capsid (C, 100AAs), membrane (M, 75AAs) and envelope (E, 495 AAs)
(2). The non-structural proteins NS1, NS2A, NS2B, NS3, NS4A, NS4B and NS5 are
involved in immune invasion, pathogenesis and viral replication (19-22).
Capsid (C) protein forms the nucleocapsid which encloses the viral genome. The
approximately 100-amino acid C protein shares low homology among flaviviruses but
shares a similar distribution of basic amino acids and hydrophobicity profiles (23). It
plays an important role in encapsidation of the viral genome and may also be involved
in regulation of viral replication (24). C-protein has also been reported to inhibit stress
granule formation thus facilitating propagation of virus in host cells (25).
The envelope (E) protein contains a transmembrane anchor and 3 globular domains
linked to the anchor by a stem region. Flaviviruses share the common envelope protein
with ~40% amino acid homology in their envelope protein. Studies in TBEV show two
domain form an elongated shape consisting of a central domain (DI) that connects an
Ig-like domain (DIII) to a dimerization domain (DII), the whole structure lying flat
along the surface of the virus lipid bilayer (26). Similarly in DENV, DI is described as a
β-barrel at the core connecting DII finger-like domain with a fusion loop at one end and
DIII at other end with short polypeptide, DI/III linker (27). DII is a long finger-like
protrusion from DI and contains a second N-linked glycan that binds to dendritic cell
surface receptors (28). DII has been shown to be involved in virus attachment to Vero
cells in culture (29, 30). These binding characteristics have been confirmed using
expressed DIII (31). As a Class II fusion protein, the E glycoprotein can undergo an

26
acid-catalysed oligomeric reorganization to a fusogenic homotrimer (32-35). This event
occurs in the endosome, allowing the viral nucleocapsid to escape into the cytoplasm
and initiate RNA and protein synthesis (36).
Dynamic interaction between the E-protein heteromeric units determines the virion
surface texture (smooth-rough) and hence affects availably of virus epitopes.(37).
Alternation of viral envelope protein configuration triggered by temperature has been
reported as “virus breathing” motion of particles (38).
The prM protein functions as a chaperone protein for the E glycoprotein during viral
maturation. It helps to maintain the E glycoprotein structure until viral morphogenesis is
complete and the virion escapes the acidic exocytic vesicles (39, 40). Cleavage of prM
to form M protein defines the transition from immature non-infectious virus particle to
infectious forms (40, 41). Description of structure and function of flavivirus genes is
shown in Table 2.2. Structure of flavivirus and mature and immature virion is given in
Figure 2.1.
27
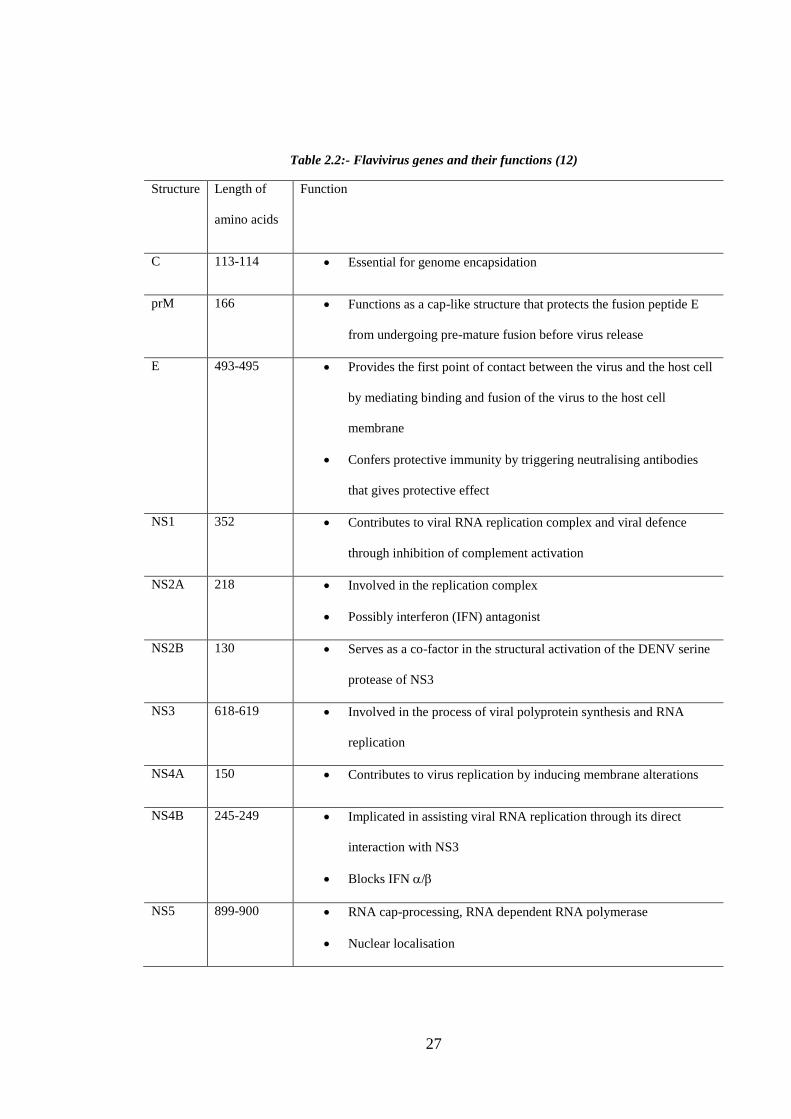
Table 2.2:- Flavivirus genes and their functions (12)
Structure Length of
amino acids
Function
C 113-114 Essential for genome encapsidation
prM 166 Functions as a cap-like structure that protects the fusion peptide E
from undergoing pre-mature fusion before virus release
E 493-495 Provides the first point of contact between the virus and the host cell
by mediating binding and fusion of the virus to the host cell
membrane
Confers protective immunity by triggering neutralising antibodies
that gives protective effect
NS1 352 Contributes to viral RNA replication complex and viral defence
through inhibition of complement activation
NS2A 218 Involved in the replication complex
Possibly interferon (IFN) antagonist
NS2B 130 Serves as a co-factor in the structural activation of the DENV serine
protease of NS3
NS3 618-619 Involved in the process of viral polyprotein synthesis and RNA
replication
NS4A 150 Contributes to virus replication by inducing membrane alterations
NS4B 245-249 Implicated in assisting viral RNA replication through its direct
interaction with NS3
Blocks IFN /
NS5 899-900 RNA cap-processing, RNA dependent RNA polymerase
Nuclear localisation

28
Figure 2.1:- Flavivirus Structure
A) Flavivirus genomic framework. Flavivirus RNA is ~11Kb, ss-RNA ORF genome that codes for
structural and non-structural proteins responsible for virion formation.
B) Mature and immature virion structure of dengue virus(2, 3)
Mature
virion Immature
virion
A
B

29
2.2.2. Replication
Virus enters host cells via inoculation by infected mosquito vectors across skin and is
introduced intradermally and intravenously, and dendritic cells enable virus migration
to lymph nodes (42, 43). Classic viral replication in all flavivirus includes attachment,
uncoating, RNA synthesis, assembly and release. Each step is illustrated in Figure 2.2.
Attachment of flavivirus to host is mediated by different cell surface receptors,
depending on virus and host cell type. C-lectin receptors DC-sign and mannose
receptors on dendritic cells and myeloid cells (36, 44), heat shock proteins (hsp),
heaparin sulphate (hs) (45-48) in mammalian and mosquito cell lines have been
identified as receptors. In other flaviviruses GAG glycosoaminoglycans have been
reported as the most common surface ligand. As the virion is attached receptor-
mediated endocytosis occurs, facilitating virus intake. The rate of virus intake by host
cells is host-dependent (40, 49). Antibody-dependent enhancement may also play a role
in enhancing virus uptake in the presence of sub-neutralising antibody induced in
previous homologous or heterologous flavivirus infection (50). Uncoating of the virion
is mediated by acidity in host cell compartments that triggers conformational change in
envelope protein, inducing fusion of viral and host membrane. Viral RNA is hence
released to mediate viral protein synthesis.
Synthesis of viral protein occurs in a series of process whereby structural and non-
structural proteins are synthesised using viral genome as template and host protein
machinery (36). Using host transcriptional systems viral RNA codes precursors of
structural and non-structural proteins. Structural protein form the virion while non-
structural proteins are involved in replication, assembly and modulation of host cell
responses (51). Non-structural proteins further play a role in viral particle movement

30
into cell organelles to complete the viral structure. Encapsidation of virion occurs in the
lumen of endoplasmic reticulum forming immature prM-C coated virion (52). Virus
utilises the host membrane as a scaffold for anchoring the viral replication complex
(53). Cleavage of prM to M to form mature infectious virion occurs in the trans-golgi
network mediated by furin proteases. This is followed by conformational change in E-
molecules forming mature smooth virion. Absence of transition of prM to M forms
immature non-infectious progeny (54). Release of infectious particles is pH mediated,
facilitating completion of the replication cycle and release of capsid coated RNA
particle in cytoplasm.

B
Figure 2.2:- Flavivirus Replication
Flavivirus replication utilizes host cellular mechanisms for formation of mature the virion, with cleavage of prM to M
(10).

32
2.3. Dengue Virus (DENV)
DENV is the causative agent of dengue fever (DF) and dengue haemorrhagic fever
(DHF). It emerged as human pathogen in the early 20th
century and is maintained in
circulation between human hosts and mosquito vectors. DENV comprises four
epidemiologically identical but genetically distinct serotypes. This section reviews
DENV classification, spectrum of disease and geographical distribution.
2.3.1. Origin and distribution
DENV may have originated in Africa or Asia. An African origin suggests DENV arose
in a sylvatic cycle in West Africa and disseminated Worldwide with the slave trade. An
Asian origin suggests evolution of DENV in the Malay Peninsula in a forest cycle
involving canopy-dwelling lower primates and mosquitoes (18, 55). Genetic analysis
suggests independent evolution of four serotypes of DENV from ancestral, sylvatic
viruses with the expansion of the sylvatic progenitors to new vectors and hosts. These
studies imply that sylvatic cycle in Asia and West Africa is the potential source of
endemic/epidemic strains of DENV (56). Phylogenetic analysis of sylvatic strains in
West Africa supports this hypothesis (57). Identification of all four sylvatic DENV
serotypes in canopy-dwelling non-human primates and Aedes mosquitos in Malaysia in
1978 supports a sylvatic lineage arising in the Asian-Oceanic region (58).
DENV was first isolated in 1943 during World War II by Hotta and Kimura (59). In
1944 Sabin and his colleagues isolated DENV (subsequently designated as the
prototype DENV-1Hawaii strain) from US soldiers stationed in India, New Guinea and
Hawaii (60). Significant spread of dengue occurred during this period of World War II
from 1941 to 1945. A dengue pandemic was reported among military personnel in
entire Pacific, from Australia to Hawaii and from New Guinea to Japan (61). However,
A

33
cases of haemorrhagic manifestations, shock and death associated with dengue were
reported as early as 1897 in Australia(62), 1915 in Taiwan (63) and 1928 in Greece(64).
Dengue has emerged as infectious disease that with widely distributed throughout the
tropic and subtropics. About 2.5 billion people, or 2/5th of the world’s population, live
in areas where there is a risk of dengue transmission. Dengue is endemic in more than
100 countries in Asia, the Pacific, the Americas, Africa, and the Caribbean. The World
Health Organization (WHO) estimates that 50 to 100 million infections occur yearly,
including 500,000 DHF cases and 22,000 deaths, mostly among children (65).
The history of dengue in Australia extends over more than 120 years. The earliest
reference described the import of eight cases by ship from Mauritius in 1873(66); the
first indigenous outbreaks likely occurred in Queensland at Townsville in 1879 and
Rockhampton in 1885. Several epidemics in northern Queensland were described in the
1890s and in the early part of the 20th century (67). The first cases in northern New
South Wales were reported in 1898, but epidemic activity did not extend southwards
until 1925-26 when cases were described as far south as Newcastle. Dengue was
reported in Western Australia in 1909-10 and was declared notifiable there in 1912.
Improvements in water storage systems and supply infrastructure lead to eradication of
Ae. aegypti and dengue by the 1960s. In 1914, dengue was reported from Darwin in the
Northern Territory (68). Recent Australian data from 2009 -2014 shows an average of
16.54 notifications annual crude rate per 100,000 population, with peak of 22.2 in 2010
and lowest 5.9 in 2009 (69).
Although dengue has not been present in WA since the 1940s (70), the state has
reported the highest number of annual notifications in the country (71). In recent years
new budget airlines began offering affordable package holidays and travel increased
300-fold between 2006 and 2010, coinciding with a sharp increase in dengue cases
notified to the Communicable Diseases Division of the WA Department of Heath (72).

34
Analysis of DENV derived from 6 febrile travellers entering WA after visiting seven
countries throughout Asia between 2010-2012 identified a diverse range of genotypes
and lineages within all four DENV serotypes (11). Most of the travellers had entered
from Bali, a popular holiday destination for residents of WA. Many of the imported
DENV were local regional variants with strong epidemic potential known to have
circulated in the region for some time. Other DENV were more recently introduced into
Bali from other countries. A new lineage of DENV-2 that appeared to be associated
with a major outbreak in Bali in 2012 was also identified and importation of this lineage
has continued up to 2017. Bali is a melting pot of substantial DENV diversity and
serves as a hub for DENV transmission and mixing. In October 2013 the first case of
locally acquired dengue fever occurred in a male with no history of travel outside WA
for many years and who was likely exposed in the Pilbara region in north-west WA
(73). The case may have been exposed to infected mosquitoes imported on international
cargo vessels docked at local iron ore export ports or via direct international flights into
Port Hedland or Perth airports. Between February 2014 and March 2016 Ae. aegypti
was frequently detected at Australian international airports (74, 75). The most regular
detections occurred at Perth International Airport but there is no evidence that Ae.
aegypti has become established. Dengue continues to be introduced into WA via
returning travellers (76).
In 2013, 390 million global DENV infections were estimated to have occurred of which
approximately one quarter - 96 million – were symptomatic, with varying severity
(77). Global incidence and prevalence continues to increase. In recent years there have
been outbreaks in countries where dengue has never been reported or has not occurred
for many years. In 2013, Japan had an outbreak in Tokyo after a lapse of 70 years (78).
Emergence of mosquito vectors in countries with no previous reports been identified
and movement of viruses, especially in travellers, is associated with increased disease

35
burden (79, 80). In 2015 India had its worst outbreak after 15 years. In 2016, large
outbreak of DENV was reported worldwide (78). Increased globalisation, travel and
trade are important determinants of dengue world-wide distribution (81, 82).
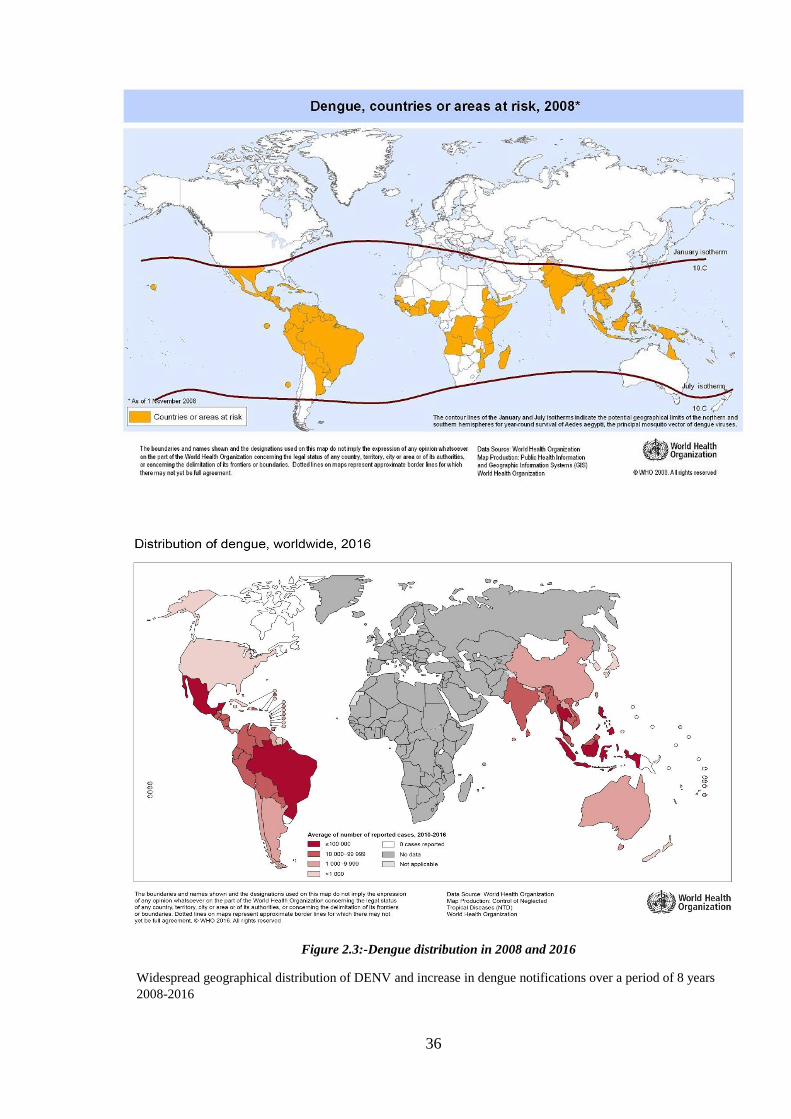
WHO dengue maps in Figure 2.3 show distribution of dengue and/or dengue
haemorrhagic fever, countries at risk of dengue in 2008 and the average number of
suspected or confirmed dengue cases reported to WHO from 2010-2016. These maps
illustrate the rapid geographic spread of DENV in an eight year period.
36
Figure 2.3:-Dengue distribution in 2008 and 2016
Widespread geographical distribution of DENV and increase in dengue notifications over a period of 8 years
2008-2016

37
2.3.2. Classification
DENV is classified as four distinct serotypes; DENV-1, DENV-2, DENV-3 and DENV-
4. DENV genetic diversity is attributed to interactions among virus, vector and host.
DENV has an error-prone RNA dependent RNA polymerase with no proof-reading
capacity, producing approximately one mutation per round of genome replication thus
leading to diversity among species and between serotypes (83, 84). Clonal evolution,
homologous recombination, intra-serotype recombination is also suggested due to co-
occurrence of different genotypes and serotypes in a single host or vector. However no
significant emergence due to recombination has been reported (85).Evolutionary forces
drive lineage replacement and clade turnover resulting in change in prevalent epidemic
strains. Vector migration, human movement and increased density of human living
conditions also impact generation of diversity (86, 87). Interaction between virus and
host immune mechanisms is also an important factor. Viruses associated with high
viremia and the capacity to escape immune responses are available to vectors for longer
periods and therefore are more likely to be transmitted during blood meals (88-90).
Phylogenetic analysis shows that emergence of modern serotypes of DENV occurred
after establishment of human populations large enough for urban transmission.
Emergence is believed to have been facilitated due to vectors switching from arboreal
Aedes to domestic and peri-domestic Aedes species. Diversity of serotypes is also
attributed to competition for susceptible hosts. Development of RNA sequencing
techniques allowed definition of genotypes within serotypes of DENV(57).
DENV serotypes, originally identified using serological approaches, are more precisely
classified phylogenetically into distinct genotypes representing clusters with nucleotide
sequence divergence of not more than 6%, and lineages within the genotypes may
represent strains with similar geographic origins. The definition of genotype and lineage

38
has varied in different studies; genotype distribution reviewed here is based on
definition by Chen et al (85).
Based on partial and whole genome sequences five distinct genotypes of DENV-1 have
been defined, named as Genotype I-V. The maximum nucleotide divergence is 6%
among genotypes (57, 91). The average nucleotide substitution rate is 6.56x10-4
substitution/site/year (92). Within genotypes there are distinct clades associated with
outbreaks sharing a specific spatiotemporal association. All the genotypes have
recorded human cycle transmission. Genotype III was reported to include sylvatic
isolates collected in Malaysia however recently E-gene analysis suggested possible
spill over from human to monkey, rather than true sylvatic DENV (56). Genotypes I
and IV are reported in recent epidemics in the Pacific while Genotype V has been
described in epidemics in Americas. Genotype II and Genotype III are not as widely
distributed as the other genotypes (24, 93). Spatiotemporal association of each genotype
is listed in table 2.3.
DENV-2 has six genotypes named after tentative topographical coverage: Asian
genotypes I and II, Cosmopolitan genotype, American genotype, Southeast Asia,
Asian/American genotype, in addition to Sylvatic genotype. Geographical distribution
of these genotypes is listed in Table 1. The sylvatic genotype of DENV-2 is the first
evidence of true sylvatic origin (94). Genotypic replacement of less viremic strains by
high viremic strains has been observed for DENV-2 (95, 96). Strain replacement is
attributed to greater fitness and longer viremia hence increased availability for the
vector. Distribution of DENV-2 genotypes is listed in table 2.3.
DENV-3 was delineated into four genotypes named Genotype I-IV by Lanciotti (97).
Genotype V was introduced subsequently (98). The prototype H87 strain used as a
WHO reference strain is included in this new genotype. DENV-3 genotypes evolved

39
and were maintained in circulation in particular regions and spilled over to other regions
with increased human and trade movement. However, in comparison to DENV-1 and
DENV-2, DENV-3 genotypes have shown more limited co-circulation. Topological
distribution of DENV-3 is described in table 2.3.
DENV-4 has four major genotypes, Genotype I-IV. Genotypes I and II show limited
topological distribution with the former circulating exclusively in Southeast Asia while
Genotype II circulates independently in Southeast Asia and Americas with limited
genetic exchange. Genotype I includes the prototype strain H241 isolated in the
Philippines, used as a WHO reference strain. Genotype III represents five distinct Thai
isolates circulating between 1997 and 2001 (99). Genotype IV is a sylvatic lineage and
is genetically distinct to the other genotypes (100) . The geographical topology of
DENV-4 is listed in table 2.3.
Figure 2.4. Shows phylogenetic relationships among DENV

40
Serotype Genotype Distribution
DENV-1
I
II
III
IV
V
Southeast Asia, China and East Africa
Isolated from Thailand between 1950s-1960s
Sylvatic strains originated from Malaysia with
possible spill over from human infection
Pacific Islands and Australia
Americas, West Africa, Asia, South East Asia
DENV-2
Asian genotype 1
Asian genotype 2
Cosmopolitan
American
Southeast Asia
Asian/American
Sylvatic genotype
Indonesia, Malaysia the Philippines and recent
isolates from South Pacific
Thailand Vietnam and Bangladesh
Sri Lanka, India, Africa and Samoa, two sub-clades
with global distribution
Indian subcontinent, Pacific Islands, Central and
South America.
Southeast Asia, South America likely introduced
from Vietnam, American strain.
Mixed strains from human and arboreal mosquitoes.
Genetically distinct from other strains representing
ancestral genotype.
DENV-3
I
II
III
IV
V
Strains from areas of Southeast Asia particularly
Indonesia.
Thailand
Sri Lanka, Taiwan, Singapore, Samoa, East Africa
Puerto Rico,Tahiti
Prototype H87-1956 Philippines isolates, China,
Japan and Brazil.
DENV-4
I
II
III
IV
Strains from Thailand, Philippines, Sri Lanka
Indonesia, Malaysia, Tahiti, Caribbean and
Americas
Thai strains that are distinct from other Thai strains
Sylvatic strains from Malaysia
Table 2.3: Classification of DENV serotypes

41
.
Figure 2.4:- DENV Phylogeny
Complete open reading frames of DENV sylvatic and endemic/epidemic strains ,
available in the GenBank library (Weaver and Vasilakis, 2009)(12)

42
2.3.3. DENV Vector and Transmission
Dengue is transmitted by the bite of an infected female mosquito. The predominant
mosquito vectors that maintain DENV in urban transmission cycles are from Aedes
genus of Culicidae family. Aedes aegypti, Ae albopictus and Ae polynesiensis are the
major human vectors. In 1906 Thomas Bancroft suggested Ae. aegypti as the carrier of
DENV, confirmed by John Burton in 1916 (101).
Aedes mosquitoes are day-biting vectors that breed mainly around urban environments
in clean fresh water and in manmade receptacles. Ae. aegypti is resilient to
environmental stress (e.g., droughts) or human interventions (e.g., control measures).
The ability of the eggs to withstand desiccation (drying) and to survive without water
for several months on the inner walls of containers allow greater adaptation to the
environment (102). The species of mosquito vector depends on the transmission cycle.
The widespread of DENV and its establishment in large human population was
Figure 2.5:-DENV transmission cycle and mosquito-vectors (9)

43
facilitated through vector switching from arboreal to peri-domestic mosquitoes (9).
DENV transmission between mosquitoes and humans does not need a sylvatic cycle for
maintenance. However, in forest settings sylvatic transmission exists between non-
human primates and Aedes mosquitoes. There are two distinct transmission cycles
described: sylvatic and human. The sylvatic cycle is hypothesised to be ancestral cycle
ecologically and is evolutionarily distinct from the human transmission cycle. This is
supported by phylogenetical studies showing sylvatic strains of West Africa are
genetically and evolutionarily distinct from all endemic and epidemic strains (103, 104).
The sylvatic transmission cycle is maintained between canopy-dwelling Aedes spp. and
non-human primates. The principle vectors responsible for this cycle are listed in Figure
2.5. West Africa and Southeast Asia are well-documented foci of sylvatic transmission
(9).
Establishment of human civilisation to a threshold that supported DENV transmission,
and adaptation of sylvatic DENV to both peri-domestic and urban mosquitoes resulted
in increased transmission via human cycles. DENV strains in human cycles are distinct
from sylvatic strains and circulate exclusively in domestic and peri-domestic
environments. Primary vector for transmission in human cycle is Ae. aegypti, Ae.
albopictus and Ae. polyenesiensis are secondary vectors (104-106).
Transovarial transmission (TOT) has been proposed as a mechanism of DENV
maintenance in both sylvatic and human cycles. Vertical transmission of DENV-1 and
DENV-4 was reported in Ae. albopictus in Brazil in 1990(107). Similarly, vertical
transmission of DENV-3 was reported in Ae. aegypti (108). A significant role for TOT
or vertical transmission in epidemiological persistence of DENV is yet to be confirmed.
A review of the literature of vertical transmission and its role in dengue epidemiology
by Grunnill and Boots concluded that a number of studies failed to establish evidence
for vertical transmission and maintenance of DENV circulation (109).

44
Other non-vector routes of DENV include needle stick injury, mother-to-child vertical
transmission, transfusion-related transmission and mucocutaneous exposure.
Transmission of dengue by accidental inoculation at a metropolitan hospital in France
was reported in 1990 (110). In 1996 a case of needle stick transmission of DENV-2
from an infected traveller to nurse was reported in France (111) and again in 2004, a
case of nosocomial transmission by needle stick injury was reported from an infected
traveller to a nurse in Germany (112). In 1994 vertical transmission of DENV-2 was
reported in a Thai woman; virus was isolated from her infant on the 6th
day of life(113).
Similarly, in a case of infant fatality with multi organ failure DENV-2 was isolated from
blood of the infant, borne to a mother with acute DENV infection (114). DENV-2 was
also detected in cord blood and infant’s blood in a case of vertical transmission from a
mother who was ill 5 days prior to birth, both mother and infant recovered and were
discharged 6 days after delivery (115). Boussemart et al confirmed two cases of
prenatal DENV transmission in 2001 (116). Intrapartum transmission of DENV-2 and
bone marrow transplant transmission of DENV-4 was reported during 1994-1995
DENV epidemic in Puerto Rico (117).

45
2.3.4. Clinical spectrum of dengue disease
Dengue clinical manifestations range from self-limiting to severe fatal disease. Dengue
fever (DF) is an acute febrile illness with two or more of the following symptoms and
signs: severe headache, severe eye pain, joint pain, muscle and/or bone pain, rash, mild
bleeding manifestation, low white cell count. Age, immune status, host genetic
background, infecting DENV strain are among the factors which contribute to infection
and disease severity (118). Severe dengue is characterised by fever that lasts from 2-7
days with general signs and symptoms consistent with dengue fever. Warning signs for
severe dengue develop with the decline in fever within 24-48 hrs. These include
capillary leakage leading to organ impairment, low platelet count, haemorrhagic
manifestations and possible fatality (30). The clinical spectrum of disease at different
day’s post-onset of acute illness is shown in Figure 2.6. There is no specific treatment
for dengue. Proper case characterisation and supportive treatment consisting of fluid and
electrolyte replacement therapy is the only available approach to management of severe
dengue. Successful management of dengue depends on early recognition of dengue
infection, recognition of warning signs for severe dengue, and access to medical care by
experienced health care workers (119).

46
Figure 2.6: Dengue clinical manifestations (World Health Organization 2009)

47
2.3.5. Dengue case classification
In 1997 WHO classified severe dengue cases as DF, DHF (Grades 1 and 2) and DSS
(DHF grades 3 and 4) (Figure 2.6) based on guidelines used in Thailand (120).
According to this classification, DF is characterised by high fever with two or more
clinical manifestation of headache, retroorbital pain, myalgia, arthralgia rash,
haemorrhagic manifestations. DHF fulfils four criteria: fever or history of fever lasting
2-7 days; a haemorrhage shown positive by tourniquet test or spontaneous bleeding,
thrombocytopenia and evidence of plasma leakage. DHF severity is classified based on
the presence or absence of spontaneous bleeding and severity of plasma leakage. (121,
122). However variations in clinical manifestations made application of this WHO
classification difficult and application of these criteria fails to detect severe dengue
(123). An assessment in Vietnam showed that the WHO classification resulted in a
considerable decrease in DHF cases and suggested that DF and DHF are part of a
continuous spectrum of dengue rather than being two separate clinical identities (124).
In 2009, WHO has introduced an algorithm to make clinical assessment of dengue
simple and intuitive. According to this new system dengue is now distinguished in three
categories: dengue without or with warning signs, and severe dengue. Dengue without
warning signs is an initial febrile phase with nonspecific symptoms but can be probable
dengue if there is history exposure to dengue in an epidemic/endemic area. Dengue with
warning signs occurs in the crucial phase soon after infection and may progress to
severe dengue. Dengue is classified as severe dengue if one or more of the following is
present: severe plasma leakage, severe haemorrhage and severe organ impairment.
Figure 2.7 and Figure 2.8 show the classical and recent classifications of dengue.
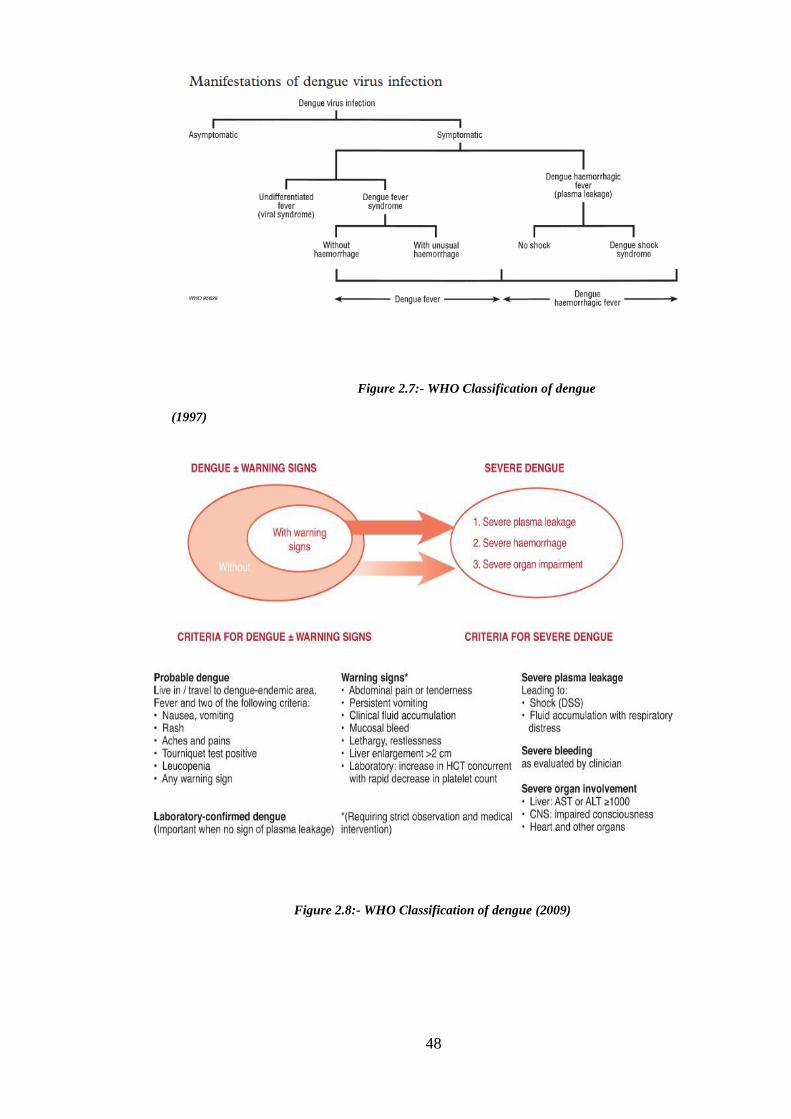
48
Figure 2.8:- WHO Classification of dengue (2009)
Figure 2.7:- WHO Classification of dengue
(1997)

49
2.4. Zika Virus
ZIKV is a flavivirus that belongs to the Spondweni serogroup, which includes Zika
(ZIKV) and Spondweni virus (SPONV). Both viruses show high level serological
cross-reactivity and have similar clinical presentations (125). ZIKV was first discovered
in 1947 in the Zika forest in Uganda following isolation from a sentinel monkey during
research on yellow fever virus. The first ZIKV isolate is the prototype strain MR766,
isolated from monkey no. 766 (126). In 1948, ZIKV was isolated from a pool of Aedes
africanus (ZIKV E/1 strain) (127). The first ZIKV isolate from humans was reported
during an outbreak of jaundice in Nigeria. Samples obtained from 3 cases were
investigated for yellow fever, ZIKV was isolated from one patient and two other
exhibited a rise in titre of serum antibodies against ZIKV(128).Subsequently, a strain of
ZIKV (P6-740) was isolated from Malaysia in 1966 (129) and first human case in
Asia was reported from Central Java, Indonesia in 1977-1978 (130). ZIKV re-emerged
in an outbreak mistakenly identified as dengue in 2007 in Yap state, Federated States of
Micronesia, and moved across the western and south Pacific to cause an outbreak in
French Polynesia in 2013 and subsequently, Brazil in 2015. This was the first known
introduction of ZIKV to the Americas.
More than 70% of the population in Yap were infected (131) during an epidemic where
the major symptoms were rash, arthralgia and conjunctivitis. Rapid diagnostic tests
suggested DENV as the causative agent. However samples from this outbreak when
analysed by CDC by RT-PCR demonstrated ~90% nucleotide identity with ZIKV,
confirming ZIKV as the causative agent of the ongoing epidemic. In 2013, 11.5% of the
population of French Polynesia was infected during an outbreak that lasted 21 weeks.
The first association between ZIKV infection and neurological complications (Guillian
Barre Syndrome) was also reported during this outbreak with a 20-fold increase in
incidence of GBS compared to the past four years. Phylogenetic analysis of ZIKV

50
isolated in French Polynesia showed homology with ZIKV isolated in Cambodia (132).
ZIKV outbreaks in the south Pacific followed, with transmission in New Caledonia,
Cook Islands and Easter Island in 2014 (126). In late 2014 and early 2015 cases of
exanthematic disease were reported in Brazil, with neurological syndromes, in regions
where ZIKV, DENV and CHIKV co-circulated (133). By the end of Sept 2015 cases of
Zika-associated neonatal microcephaly were reported and by the end of 2015, 440,000-
1,300,000 Zika cases in 18 states, with confirmed autochthonous transmission, were
estimated to have occurred in Brazil (134) and WHO declared a global emergency. In
October-November 2015 autochthonous ZIKV transmission was reported in Sincelejo,
Columbia. Virus isolated from this outbreak showed 99% sequence identity with FP
strain (135). ZIKV was then reported in 12 other countries and territories of the
Americas (126, 136). Systematic spread of ZIKV is illustrated in Figure 2.9 (137).With
global spread to more than 60 countries and the newly recognized association with
neurological complications Zika emerged as flavivirus with significant global disease
burden. However a steep fall in Zika cases has been noted (138). A single case was
confirmed locally transmitted in the United States to mid-August 2017. The decline in
incidence in countries with previous explosive transmission is proposed to result from
herd immunity and reduction in the number of susceptible people.
Figure 2.10 shows the rise and fall of Zika from late 2015 to 2017 in the Americas.

51
Figure 2.10:-The rise and fall of Zika
The rise and fall of Zika. Rapid increase in Zika cases in early 2016 and steep decline at the end of year
(Cohen, Science 2017)
Figure 2.9: Zika virus spread from 1947-2016
Zika virus spread 1947-2016. The chronological distribution of ZIKV from its earliest
discovery to neurological disorder-associated cases in 2016. (US Centres for Disease Control)

52
2.4.1. Classification
Phylogenetic analysis of flaviviruses has shown that ZIKV is placed in the mosquito-
borne cluster, based on the genomic sequence of 100-Kb genome segment at the 3′
terminus of the NS5 gene (8). As with other flaviviruses ZIKV is a positive sense ss-
RNA virus with a genome of approximately 11kb that codes for structural and non-
structural polyproteins. Among the flaviviruses of medical importance, ZIKV is closely
related to DENV with approximately 55.6 % amino acid homology across viral
polyprotein (139). ZIKV is reported to circulate as single serotype divided into strains
based on geographic origin as African and Asian strains. The African prototype MR766
1947 Uganda strain and the Asian prototype 2007 Yap strain share 96.5% amino acid
homology in the entire coding region (140).
Comparison of whole genome sequences shows 3 branches of ZIKV: East African
(Uganda cluster), West African (Senegal cluster) and Asian (131). The two African
lineages are believed to be due to independent introduction to these regions of prototype
MR766.
Phylogeny of ZIKV based on whole gene sequences is shown in Figure 2.11
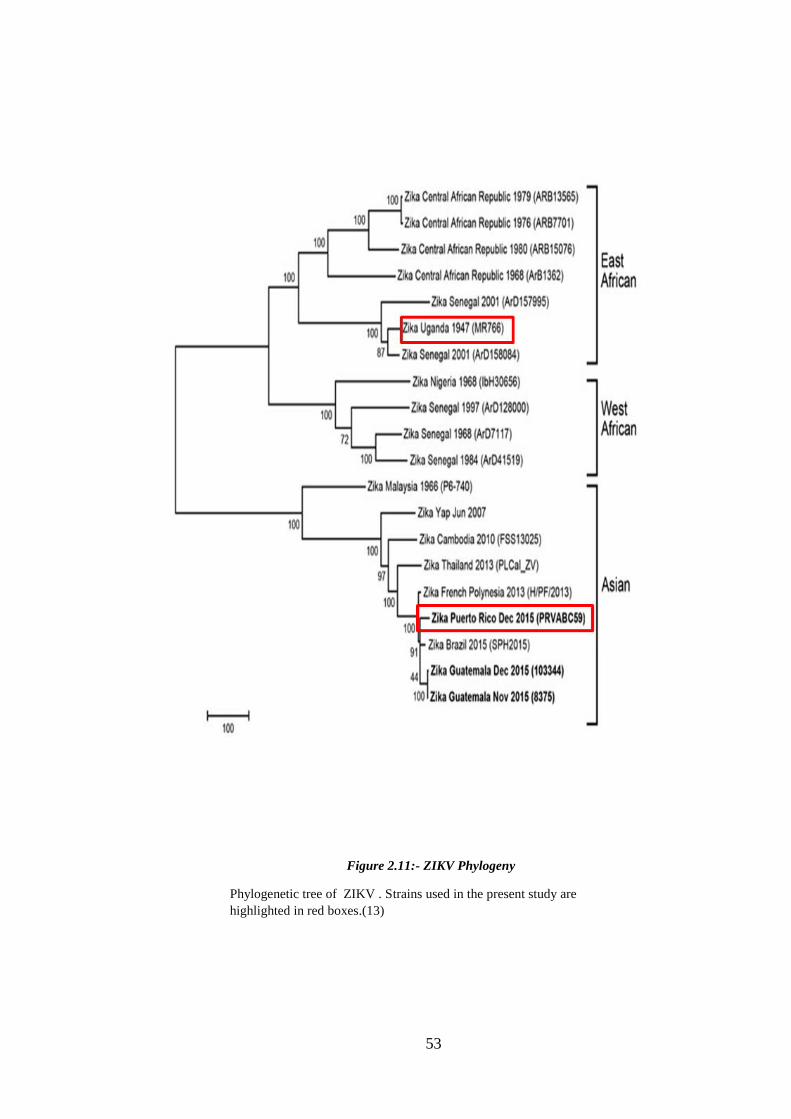
53
Figure 2.11:- ZIKV Phylogeny
Phylogenetic tree of ZIKV . Strains used in the present study are
highlighted in red boxes.(13)

54
2.4.2. Vector borne ZIKV transmission
As for DENV Aedes genus is the principle vector for ZIKV. The ZIKV African strain
MR766 was isolated from Ae. africanus and the ZIKV Asian (Yap) strain was first
isolated from Ae. aegpti and Ae hensilli in Yap (2007) (141, 142). Ae. albopictus was
the vector in Africa (143). The earliest ZIKV isolations from Asia were from Ae. aeypti
in Malaysia in 1966 (129). In southeastern Senegal, a study of 1700 mosquito pools of
11,247 mosquitoes showed that Ae. furcifer and Ae vittatus were involved in ZIKV
transmission to humans(144). The role of mosquitoes as a ZIKV vector has been
confirmed in animal models. Boorman et al reported successful transmission of ZIKV
by Ae. aegypti to rhesus monkeys and infant mice (145). ZIKV shows wide divergence
of mosquito species as potential vectors for transmission; vector selection
predominantly depends on the availability of mosquito species. ZIKV seems to be
adapted to a large number of mosquito species including those not commonly known to
transmit other flaviviruses. Isolation of ZIKV from diverse species of Aedes, Culex and
Mansonia mosquitoes suggests a wide range of potential ZIKV vectors (126).
2.4.3. Non- vector transmission of Zika
Vector-borne transmission is the most common route for spread of ZIKV however
maternofetal and sexual transmission has been confirmed (146, 147).
In October 2013, perinatal transmission of ZIKV was confirmed in two mothers in
French Polynesia. ZIKV infection in a newborn child was confirmed by RT-PCR on
serum collected four days post-delivery. Possible transplacental infection of the infant
during delivery suspecte (148). Maternofetal transmission was confirmed during the
Brazil outbreak in 2015 with reports of neonatal malformations including microcephaly
linked with ZIKV infection. Virus was also isolated from amniotic fluid and tissue
samples from an infected infant during this outbreak (149, 150).

55
In 2008, clinical and serological evidence indicated that two American scientists who
recently returned after working in southeastern Senegal had contracted ZIKV infection.
One of the scientists transmitted ZIKV to his wife after his return home, most likely as a
sexually transmitted infection. Symptoms in the two scientists included maculopapular
rash, fatigue, arthralgia, headache and hematospermia in one of the patients. The wife
developed muscle aches and pains, headache, photophobia chills and malaise. Anti-
ZIKV hemagglutination inhibition antibody titres and neutralising titres were elevated.
(147). ZIKV was isolated from semen of a patient in Tahiti who presented with
hematospermia(151). Similarly, in 2016 ZIKV infection was diagnosed in a previously
healthy 24-year old woman with no travel history to Zika endemic areas after sexual
contact with man who stayed in Brazil from December 11, 2015 to February 9, 2016;
ZIKV was isolated from semen of a man 18 and 24 days post-onset of symptoms (152).
Other non-vector routes of transmission include via blood transfusion and laboratory
contamination. In the 2013 FP outbreak 3% of 1,505 blood donors, asymptomatic at the
time of donation were found to be ZIKV-positive positive by PCR (148). ZIKV
infection has also been reported as a result of laboratory work with arboviruses in 1973
(153).
2.4.4. Infection and complications
Acute symptomatic ZIKV infection is a self-limiting mild flu-like illness. Symptoms are
similar to other flavivirus infections and include fever, rash, arthritits, arthralgia or
myalagia, conjunctivitis and fatigue. However recent outbreaks have reported an
association between neurological complications and Zika. Most documented cases are
of Guillian-Barre syndrome and of microcephaly in neonates.
A) Guillian-Barre syndrome (GBS)

56
GBS is a neurological disorder related to autoimmune responses in which the immune
response targets the peripheral nervous system. GBS includes weakness of the arms and
legs and in severe cases paralysis of muscles that support breathing. During the 2013
French Polynesia outbreak a 20-fold increase in GBS following ZIKV infection was
reported (154). In the 2015 outbreak in Brazil 62% c of 121 Zika cases were reported to
precede symptoms of GBS. Similarly, 54% of 22 ZIKV infections in El Salvador were
also reported to proceed to GBS. In Columbia 2015-16, 67% of neurological cases were
reported to correspond to GBS (155). From April 2015-March 2016, 1474 cases of GBS
and 164,4237 suspected and confirmed Zika cases were reported from Brazil, Columbia,
the Dominican Republic, El Salvador, Honduras, Suriname and Venezuela. Compared
to baseline (pre-ZIKV) there were 172%, 211%, 150%, 100%, 144%, 400% and 877%
increases in GBS in the respective countries, strongly suggesting a connection between
GBS and ZIKV infection(156). However proof of causality is yet to be established
(157).
B) Neonatal ZIKV-related complications
Congenital Zika syndrome and microcephaly are neonatal defects in ZIKV infection.
Partial skull collapse due to subcortical calcification, damage to back of eye, macular
scarring, focal pigmentary and congenital contractures are among the indicators for
congenital Zika syndrome listed by CDC. Microcephaly is malformation resulting in
smaller head size related to birth defects and neurological conditions. Symptoms vary
depending on extent of brain disruption. During the 2015 Brazil outbreak 97.5%
increase in microcephaly was reported (158). Similarly, the 2013 French Polynesia
outbreak was followed by increase in microcephaly in the following years. In Hawaii,
USA, the presence of IgM Ab was reported in a mother who gave birth to full term baby
with microcephaly (159). In Columbia 50 cases of microcephaly reported in a period of
three months in 2016, a higher rate than the previous record of 140 cases per year (160).

57
Spatiotemporal occurrence suggests an association between microcephaly and ZIKV
transmission; however more data to establish this association is required and
investigations are ongoing.

58
2.5. Flavivirus vaccine strains
Yellow fever 17D and Japanese encephalitis IMOJEV vaccine are the most successful
flavivirus vaccines produced and are used in endemic areas as well as by travellers from
non-endemic countries. Antibody-mediated immune responses in flavivirus infection
are highly cross-reactive and pre-existing antibodies induced by previous natural
infection or vaccination means that interpretation of serological test results may be
difficult. Analysis of vaccine-mediated immune responses is therefore integral to any
analysis of anti-flavivirus immunity, especially in travellers. The background of the
flavivirus vaccines YF17D and IMOJEV is reviewed in this section.
2.5.1. Yellow Fever
Yellow fever (YF) is a well-known acute haemorrhagic flavivirus infection. Yellow
fever initially originated in Africa and was introduced to the western hemisphere with
the slave trade. Infection was often associated with ports, as outbreaks were reported
with arrival of ships from known epidemic foci (161). YF was endemic in tropical
regions of Africa and South America, and YF epidemics occurred upon introduction of
YFV to North America, Caribbean and Europe (162). It was the major health threat
during the 20th
century. Development of a live-attenuated vaccine in the 1930s led to
decline of the disease. However epizootics and human transmission still continues
(163). Outbreaks have been recently reported in Uganda (2010), Sudan and Ethiopia
(2012-2013) (162). In 2017- early 2018, 35 confirmed human cases of yellow fever
were reported in Brazil, from the states of São Paulo (20 cases, including 11 deaths),
Rio de Janeiro (three cases, including one death), and Minas Gerais (11 cases, including
seven deaths), and the Federal District (one fatal case) (164).
Infection with YFV may cause an acute infection with symptoms including fever,
arthralgia, nausea and photophobia. Multiple organ systems are affected including the
liver, kidneys, gastrointestinal tract and brain with fever, jaundice, severe haemorrhage

59
and shock ensuing (165). Mortality is associated with jaundice and other severe liver
diseases. During the early 19th
century, yellow fever was mistaken as bacterial disease.
At the end of the 19th
century Ae. aegypti was shown to be the vector and that infection
was caused by a filterable agent (166). Max Thieler developed a vaccine for yellow
fever in 1937 by passaging the Asibi strain through laboratory animals including mice
and chickens; the attenuated, protective strain was named 17D (167).
YFV is classified into seven major genotypes: five African and two South American.
African genotypes have 10-23% nucleotide differences within genotypes while South
American genotypes have 16% differences in nucleotide sequences (168).
Wild type YFV is viscerotropic and affects a wide range of tissues and organs, with
liver being the predominant target .Virulence is highly dependent on virus strain and
host immune status (169).
YF17D was selected as the vaccine strain as it was believed to induce immune
responses similar to wild type infection. 17D escape variants are neutralised up to 10-
fold by monoclonal antibody derived from the sera of 17D vaccine recipients,
suggesting there is no risk of immune escape among mutant strains in the mouse model
(170). 17D has shown neurotrophic side effects in experimental lab animals but in
humans viremia induced by vaccination is very low and unlikely to invade the blood-
brain barrier however adverse effects have been reported in infants <- 6 months old.
17D-YFV is by far the most effective flavivirus vaccine ever produced.
2.5.2. Japanese encephalitis
Japanese encephalitis (JE) is an acute febrile illness caused by infection with Japanese
encephalitis virus (JEV).

60
JEV first emerged in Japan during the 1870s and spread throughout Asia. Five
genotypes of JEV have been described so far, each genotype has a different
geographical distribution. The five genotypes are thought to have evolved from an
ancestral Indonesian strain (171).
JE is transmitted by Culex spp mosquito (Culex tritaeniorhychus) (172). Domestic pigs
are the major reservoir for mosquito-human transmission. High magnitude, long-lasting
viremia in pigs and close proximity to human hosts means pigs are a potent reservoir for
transmission(173). Birds are another major reservoir host and enable wide geographic
distribution. Humans and non-avian vertebrates are dead-end host as they do not
produce viremia to sufficiently high levels to infect new mosquitos. Virus circulation is
maintained via zoonotic transmission and vertical transmission in mosquitoes (174).
JEV infection may result in severe disease with febrile symptoms, aseptic meningitis,
acute flaccid paralysis or classic meningoencephalitis. In the first outbreak in Japan,
casualty rates reached 30% with 30-50% of survivors suffering neurologic disability. JE
the major public health problem in Southeast Asia and China (175). There are two broad
epidemiological patterns: in sub-tropic countries disease is mostly prevalent during wet
monsoon season while in tropical countries JE is seen throughout the year. The first
outbreak in Australia occurred in the Torres strait in 1995, and one case was reported in
northern Queensland (176). Relocation of pigs away from homes and major drainage
works markedly reduced transmission and diminished cases of human infection.
JEV vaccination is the only preventive measure available, apart from vector control.
There are four types of JEV vaccine available derived from different JEV strains in
different countries: mouse brain-derived killed inactivated; cell culture-derived live-
attenuated; cell-culture derived killed inactivated; cell culture-derived live-attenuated
chimeric. In Australia, JE-Vax and IMOJEV are approved vaccines however
inactivated JE-Vax is no longer manufactured. Live-attenuated vaccine IMOJEV is used

61
for vaccination. JEV vaccination in Australia is recommended for travellers to epidemic
areas and laboratory workers who are at risk of exposure.

62
2.6. Flavivirus immunopathogenesis
Flavivirus immunopathogenesis is dependent on virus-host interactions. Strain-specific
modulation of host immune system has been reported in DENV. Infection with
pathogenic stains and high viremia in the host are correlated with availability of virus to
vector during a blood meal. Similarly, susceptibility of the vector to particular strains
and immune escape has been associated with clade replacement in DENV. Flavivirus
pathogenesis is reviewed in this section.
2.6.1. Flavivirus-specific innate immune responses
Innate immune responses are the first line of host defence against infection. Following
inoculation by infected mosquitoes, the innate immune system is activated and in turn
stimulates acquired immune responses. Major elements of the innate response include
initiation of type I interferon production, and activation of the complement pathway.
A) Interferons
Type I interferons (IFN) confer an anti-viral environment that inhibits the replication of
viral nucleic acid. Germline-encoded pattern recognition receptors (PRRs) present on
innate immune cells detect viral components. Most reported PRR sensors in flavivirus
infection are Toll-like receptors (TLRs), and cytoplasmic receptors such as retionic
acid-inducible gene-I (RIGI) or RIGI like receptors (RLRS)(177, 178). Binding of
these receptors to conserved pathogen associated molecular patterns (PAMPs) on
viruses activates the cascade resulting in production of IFNα and IFNβ and other
cytokines(179-181). These cytokines trigger activation of IFN-stimulating genes (ISGs)
containing IFN-stimulated response elements (ISRE). ISGs inhibit DENV infection by
inhibition of viral entry/fusion and production of exonuclease (181). Several ISGs
including OAS1 and OAS3, Oas-1b, IFITM and STAT are reported to initiate antiviral

63
activity (182, 183). IFITM proteins restrict direct and ADE-mediated DENV infection
(184). Although the innate immune system has several antiviral mechanisms, human
epidemic flaviviruses have developed several mechanisms to evade host innate
responses. DENV may inhibit IFN activity by inhibiting production and signalling of
type 1 IFN (185). DENV NS2B3 protease can cleave STING involved in
phosphorylation that results in production of Type 1 IFN (186). STING is an important
restriction factor for infection of DENV in mice. In addition DENV inhibits production
of type 1 IFN in antigen presenting cells including dendritic cells (DC), among the first
cells to encounter virus post-entry. DENV proteins NS2A, NS4A and NS4B attenuate
IFN-mediated activation of ISRE(187) and NS4B prevents phosphorylation of STAT
rendering the complex inactive during viral replication and thus downregulating
production of pro-inflammatory cytokines. Mutations in NS4B have also been linked
with increase in virulence of DENV strains(188). Other flavivirus proteins, such as
NS5, have been shown inhibit IFN signalling in TBEV, WNV and JEV(189-191). NS5
in DENV is reported to degrade STAT (189, 192). Ashour et al showed NS5 is involved
directly or indirectly in proteolytic processing of STAT-2. Understanding the role of
innate immunity in host protection, and evolution of viral evasive mechanisms, is
necessary to better understand the role of innate immune responses in
immunopathogenesis of flavivirus infection.
B) Complement
The complement pathway is a co-ordinated sequential enzymatic cascade formed by
non-covalent association between nine activated protein components and fragments
(193). Complement components are named as C1, C4, C3, C5, C6, C7, C8 and C9 in the
order of activation. Activation of complement includes three well defined pathways:
Classical pathway triggered C1q component binding to antigen-antibody complex on
the surface of pathogens; the lectin pathway initiated by mannose binding protein

64
(MBL) of carbohydrate structure on surface of microbes; and the alternative pathway
activated by hydrolysis of C3. These pathways activate the complement components
which assemble to form C5b-9 membrane attack complexes (MAC) that lyse infected
cells.
Complement is the heat labile component of plasma. Protective functions of
complement include priming of adaptive immune responses and complement-mediated
neutralisation (194). In flavivirus infection complement-mediated protective effects are
mainly by activation of B and T cells. All three complement activation pathways are
required for protection against WNV. Delay in priming adaptive immune response was
reported in mice lacking components of complement (195). Enhanced neutralising
effects of complement have been reported in DENV in the presence of complement
components (196). Another protective function of complement includes restriction of
antibody-dependent enhancement. Studies in WNV show that complement component
C1q restricts subclass-dependent antibody-dependent enhancement of flaviviruses in
cell culture and mice (197). However complement components C3 and C5 were
detected in DHF and DSS suggesting possible complement-mediated enhancement of
flavivirus infection (198, 199)
DENV, WNV and YFV can evade the complement activation pathway and thus protect
virus from complement-mediated neutralisation. WNV NS1 protein has been shown to
evade complement activation by recruiting complement regulatory protein H, which
results in attenuated deposition of MAC (200). Attenuation of the complement
activation pathway by DENV NS1 has also been reported. Avirutnan et al showed, by
binding C1s and C4 in a complex, NS1 promotes efficient degradation of C4 and thus
inhibits formation of convertase and activation of complement-dependent neutralisation.
Furthermore, binding of NS1 to C4b was shown to modulate complement activation by
enhancing C4b binding protein co-factor activity that attenuates the classical and lectin

65
pathways (201). NS1 binding to MBL has shown to protect virus from lectin-mediated
neutralisation by the lectin mediated pathway of complement (202).
Thus, host factors including genetic alleles that stimulate MBL concentration in
plasma, myeloid cells expressing complement receptors mediating antibody-mediated
enhancement, and viral interaction with complement proteins govern complement-
mediated immune responses in flavivirus infection.
2.6.2. Acquired immune responses
Flavivirus-specific acquired immune responses clears virus from the host and induces
immunological memory. Development of the acquired immune response in flavivirus
infection generally occurs after 4-7 days of primary infection. This includes genomic
rearrangement in B and T cells receptor genes forming a virus specific repertoire (203).
Acquired immune responses however may be protective or pathogenic. Sharing of
common viral epitopes among the flavivirus serocomplex confers immune responses
which may be protective or dysfunctional. Acquired immune responses elicited during
flavivirus infection are reviewed in this section.
A) Cellular immunity
DENV serotype-specific and cross-reactive T cell responses have been demonstrated
and include proliferative and cytotoxic responses. T-cell responses are activated by
recognition of viral peptide antigens bound to MHC class I or MHC class II molecules
on the surface of antigen presenting cells (APCs). These peptides are short fragment
derived from infecting virus. Peptides derived from processed intracellular virus are
restricted by MHC class I molecules and presented to CD8 T-cells, which differentiate
into cytotoxic T-cells. Peptide antigen from virus endocytosed by antigen presenting
cells are restricted by MHC class II molecules and presented to CD4 T-cells. Activated

66
CD4 T-cells differentiate to effector TH1 and TH2 T-cells. TH1 cells activate
macrophages and induce B-cells to produce IgG while TH2 cells initiate naïve humoral
immune response to stimulate B-cells to produce IgM, and subsequently neutralising
and opsonizing subtypes of IgG.
The earliest in vitro studies showed that peripheral mononuclear cells from dengue-
immune individuals contain CD4+ T-cells which proliferate and produce interferon
gamma (IFN-gamma) when stimulated by dengue antigens (204). Analysis of JEV-
specific memory T-cells using a synthetic peptide library showed an association
between polyfunctional CD4+ T-cell memory response and complete recovery from JE
(205). Similarly, high magnitude YFV-specific T-cell responses have been shown to
contribute to success of yellow fever vaccination (206, 207).
DENV-specific T-cell responses are mostly directed towards non-structural proteins
(208). CD8+ T cells most frequently recognise NS3, NS4B and NS5 proteins while
CD4+ T cells mostly target E, in addition to non-structural proteins including NS3
(209-211). Predominant recognition of non-structural proteins has been postulated to
contribute to suboptimal performance of the chimeric dengue vaccine, which includes
DENV E protein in a YFV backbone. Balanced proliferation of polyfunctional CD8+
and CD4+ targeted to structural and non-structural proteins contributed to generation of
high titres neutralising antibodies (212, 213). In the murine model, protective T-cell
responses induced by a conserved peptide pool, activated in the absence of antibodies,
suggested that a dengue T-cell vaccine would reduce risk of antibody mediated
enhancement of infection (214).
An association between DENV-specific T-cell responses and development of severe
dengue in secondary infection has been shown (215). Peptide variation due to amino
acid diversity among DENV serotypes is thought to induce responses targeted to

67
original infecting serotypes, resulting in biased cytokine production that leads to
exacerbation of infection. DENV-specific memory T-cell response in secondary
infection has been linked to original antigenic sin, resulting in low affinity cross-
reactive memory T-cells that are preferentially activated by infecting virus and
inducing dysfunctional responses (216-218). Furthermore, severe dengue is associated
with HLA type. In a large case control study of Vietnamese patients with DHF,
variation at the HLA-A locus was shown to influence CD8+ T-cell responses (219).
Comparison of cytokine profiles and T-cell degranulation in a cohort of children in
Southeast Asia with acute DENV infection showed HLA-A*24-restricted T cell
responses showed suboptimal degranulation but high cytokine production contributing
to vascular leakage in secondary dengue infection (220).
Advances in understanding T-cell mediated immune response have been made with
development of synthetic peptide antigen libraries, epitope peptide prediction
techniques and in vitro models to assess T-cell mediated anti-flavivirus responses.
However, gaps in our understanding of T-cell immunopathology in natural infection as
well as vaccine-mediated exposure to DENV and related flaviviruses still remain.
B) Humoral immunity
Humoral immunity is mediated by antigen-specific antibody-producing plasma and
memory B-cells. The high degree of homology among flaviviruses stimulates
production of highly cross-reactive but not necessarily cross-protective antibodies. Anti-
flavivirus-specific antibodies can be protective, cross-reacting or enhancing depending
on age, immune status, infecting virus strain and past exposure to other related
flaviviruses.
The human antibody response in dengue is complex as it involves polyclonal responses
to primary and secondary infections with four different DENV serotypes (221).The

68
adaptive immune responses to DENV is driven by the presence or absence of virus
neutralising antibodies and T-cell responses involved in initiating antibody synthesis.
Upon infection, DENV elicits IgM and IgG directly against viral membrane protein. A
primary dengue infection is characterized by a slow and low titre antibody response.
IgM antibody is the first immunoglobulin isotype to appear. Anti-dengue IgG is
detectable at low titre at the end of the first week of illness, and slowly increases. In
contrast, during a secondary infection, antibody titres rise rapidly and antibody reacts
broadly with heterologous serotypes. In secondary infection, high levels of IgG are
detectable even in the acute phase and they rise dramatically over the two weeks. IgM
levels are significantly lower in secondary dengue infections and false negative anti-
dengue IgM reactions are observed during secondary infection. According to the Pan
American Health Organization (PAHO) guidelines 80% of all dengue cases have
detectable IgM antibody by day five of illness, and 93-99% of cases have detectable
IgM by day six to ten of illness, which may then remain detectable for over 90 days.
(222, 223).
Figure 2.12 shows production of antibody at different stages of infection (6).

69
Figure 2.12- Immunological response during Flavivirus infection
Immunological response during Flavivirus infection. Response is based on phases of DENV infection.
Virus is detected for approximately one week in serum. Detection of IgM followed by IgG indicates
primary infection while detection of IgG with the onset of illness indicates secondary infection (6)
Primary infection Secondary infection

70
i) Neutralising or protective antibody-mediated immunity
Neutralising antibodies (NAb) prevent infection of cells by blocking attachment of virus
to the cell surface and preventing fusion to cell membrane; initiating complement or
antibody dependent cell cytotoxicity; and by opsonisation of virus. NAb produced
during primary dengue infection is thought to be life-long; anti-DENV-1 NAb has been
shown to persist for more than 60 years in individuals infected during the Second World
War in Hawaii, USA (224). Anti-DENV-2 NAb was reported to persist for 70 years
following an outbreak in Nagasaki, Japan, however DENV-1 was the epidemic virus in
original descriptions of the outbreak (225). NAb are used as marker protection and
indicator of vaccine efficacy. Mice lacking B cells are vulnerable to infection and
passive transfer of mAbs provides protection against viral infection (226-228).
Neutralization of infection by virus-specific antibodies can occur through several
mechanisms, including inhibition of binding to cell surface receptors or post-binding
inhibition of viral fusion. Neutralisation of infection is shown to be dependent on
availability of epitope and the number of antibodies binding to epitopes. There are two
proposed models of effective neutralisation; a single hit model proposed by Dulbecco et
al (1956) in which neutralisation requires binding of antibody to critical epitopes that
lock virus to a confirmation which prevents infection (229); and a multiple hit model
which proposes that neutralization requires a threshold number of neutralising
antibodies binding to viral epitopes to induce interference with attachment and fusion of
virus and hence prevent infection (230, 231). WNV monoclonal antibody (mAb-E16)
occupies ~25% of viral epitope sites per virion and gives effective neutralisation (218,
232). Pierson et al proposed a model of the stoichiometric requirements of for flavivirus
neutralisation based on analysis of WNV E-DIII specific mAbs Figure 2.13 shows that
if antibody can dock on to virion with stoichiometry sufficient to exceed threshold,

71
neutralisation occurs, while engagement of virion with stoichiometry below this
threshold may support antibody mediated enhancement of infection (4, 44).
Flavivirus envelope protein is the major epitope for neutralising antibodies. It has been
shown in DENV-2 that all 3-domains of E-protein have antigenic characteristics and
conformation of these domains plays a significant role in eliciting immune response.
Antigenic properties of E protein are studied in detail by isolating mAbs against
different regions of protein and show anti-E-protein antibodies elicit virus neutralising,
hemagglutination-inhibiting and fusion-blocking antibodies (233-235). Domain III of E
protein is recognised by several anti-DENV mAbs which effectively block virus
absorption (236-239). However neutralising antibody response elicited by E-protein is
highly cross reactive and not all E-protein-directed antibodies are neutralising (240,
241). Low neutralising but enhancing antibody was reported against prM and M protein
(242, 243).
ii) Cross reactive and enhancing antibody mediated immune response
DENV-specific antibody mediated immune response triggered by a particular serotype
provides life-long immunity to the infecting serotype but not to the other three
serotypes. In secondary dengue, antibody directed towards original primary serotypes is
shown to cross-react with heterologous secondary infecting serotypes, which is termed
original antigen sin. It was first described in 1983 by Halstead el al, in the study of
severe dengue in Thai children. The highest neutralising antibody responses were
directed towards the initial infecting DENV type during secondary infection (244). A
large repertoire of DENV-reactive memory B cells persist for years after infection in
dengue-immune individuals, and human E-specific as well as prM-specific mAbs
extensively cross-react with the four DENV serotypes (245). The degree of relatedness
among infecting serotypes and preferential activation of cross-reactive memory cells is
proposed to promote original antigenic sin (18). This phenomenon of cross reactivity

72
by past DENV is the proposed cause of the immunological cascade responsible for DHF
during secondary infection(246).
In secondary dengue infection with heterologous serotype, presence of non-neutralising
but cross-reacting antibody from past infection facilitates Fc - receptor mediated entry
of virus to mononuclear phagocytes. Fc receptor mediated entry triggers biased cytokine
responses thus switching off protective mechanism of macrophages and instead
enhances amplification of virus (247). This results in increase of viral load which then
drives an immunopathogenic cascade, exaggerating cytokine responses, that leads to a
transient increase in vascular permeability. This mechanism is termed antibody
dependent enhancement (ADE).The precise way in which vascular permeability is
altered is not clear but is more likely to be a functional change rather than structural
damage, as dengue shock is rapidly recoverable, and no inflammation is evident in the
leaking surfaces (246, 248, 249).
Wanning of neutralising antibodies to the concentration that is no longer capable to
neutralise infection is also shown to increase severity of dengue (221, 250). The most
compelling evidence for ADE has come from studies with infants, who have passively
acquired antibodies to DENV from their mothers (221). Studies in Thailand reported
DHF/DSS peaked in populations of first-time infected infants born to dengue-immune
mothers and children who had experienced a mild or asymptomatic dengue infection
and become secondarily infected by a different dengue serotype. This study showed
DHF/DSS is 15-80 times more frequent in secondary infections than in primary
infection, and that up to 99% of DHF cases had heterotypic antibodies to the DENV
serotype causing the DHF (249). Similarly, epidemiological studies in South-East Asia
clearly link DHF/DSS to individuals who have had a previous dengue infection or who
have acquired maternal dengue antibody (251). In Taiwanese patients dengue RNA
levels even after defervescence, correlated with disease severity. Cameron et al. studied

73
Vietnamese infants and showed the majority experienced DHF when maternal anti-
DENV neutralising antibodies had declined to <1:20 (252). In vitro mechanism of ADE
with virus is studied in Fc-receptor bearing cells including U937, K562 and THP-1 cell
lines in the presence of varying antibody concentration. In the absence of antibody these
cell lines are selectively permeable to flaviviruses like DENV and ZIKV while in the
presence of non-neutralising antibody increased infection of these cell lines has been
reported (253-255).
With the recent outbreak of ZIKV the high degree of homology between DENV and
ZIKV has increased concern about enhancement of infection. Monoclonal antibody
studies show ZIKV cross-reacting and enhancing anti-DENV antibodies (256-261).
Possible outcomes of dengue and Zika immune primary and secondary infection is
shown in Figure 2.14 adapted from Culshaw et al. This figure shows that
immunopathogenesis in secondary infection is determined by interaction between
antibodies or T cells triggered during primary infection, and heterogenicity of secondary
infection virus.

74
Figure 2.13:- Antibody threshold determining immune response in Flavivirus infection
Binding of Antibody with the virus requires a threshold in number of Ab and epitopes. Multiple binding of antibody to maximum accessible
epitopes results in neutralisation of viruses while the non-neutralising concentration of Ab triggers Fc- receptor mediated entry into
macrophages resulting in enhancement of virus production(4, 5).

75
Figure 2. 14:- Adaptive immune responses in Flavivirus infection.
During primary immune response, virus-specific antibodies and T cells are activated. In secondary infection immune memory developed during primary
response can be protective or pathogenic depending on virus type involved in secondary infection. Cross-recognition within the serotypes of DENV and
flavivirus cross-reactivity (DENV-ZIKV) can be seen for both T-cells and antibodies however this elicits protective or pathogenic effects .

76
2.7. Laboratory diagnosis of Flavivirus infection
Accurate diagnosis is essential for clinical management, disease surveillance, studies of
pathogenesis, vaccine development and development of treatment strategies. Laboratory
diagnosis of flavivirus infection depends on availability of diagnostic markers. These
markers include virus and viral particles and immunological components produced in
response to infection and depend on timing of sample collection relative to time of
infection. In acute illness and during the viremic stage direct virus/viral particle
detection can be used while serological markers IgM, IgG are present at the end of the
acute phase and later in the post-infection period. Neutralising antibody has been
reported to persist for more than 60 years post infection in DENV-1 infection (224) and
70 years post DENV-1/2 infection (225).
Diagnostic markers and tests E-gene used at different times in dengue infection are
summarized in Figure 2.15.

77
Diagnosis of dengue follows two methods:- direct detection of virus and virus particles
in the course of fever and viraemia and indirect detection of infection via measurement
pf anti-DENV antibodies in the post-febrile period (7). Figure 2.16 summarises
available direct and indirect methods for dengue diagnosis. Direct detection is highly
accurate but has limitations in terms of timing of sample collection and availability of
resources while indirect diagnosis is widely used due to long term persistence of
diagnostic markers and requiring less resources.
Figure 2.15:- Course of dengue infection and timing of diagnosis tests (7)

78
2.7.1. Virus isolation
DENV was first isolated in 1943 by Ren Kimura and Susumu Hotta during a dengue
epidemic in Nagasaki, Japan(262) and in 1944 by Albert B. Sabin and Walter
Schlesinger in Hawaii, USA (263). ZIKV was isolated in April 1947 from serum of a
febrile rhesus monkey in the Ziika Forest, Uganda (127). Early virus isolation was done
by inoculation of mice and cell lines derived from mosquito and mammalian tissues.
Inoculation in newborn mice was widely applied (60) but this approach has limited
value as field strains DENV are not usually pathogenic for newborn mice and do not
consistently produce cytopathic effects (264). In 1964 recovery of DENV in a monkey
kidney cell line of grivet monkey Cercopithecus aethiope (Vero cells) was described in
attempts to use mammalian cell lines to isolate DENV from natural human infection
(265). Rosen and Gubler reported parenteral inoculation of Ae. albopictus mosquitoes in
1974 as a much more sensitive method to detect DENV present in serum from natural
infection or that had been adapted to cell lines or newborn mice (266).
Other cell lines used for DENV recovery include mosquito cell lines AP-61 (A.
pseudoscutellaris); Tra284 (Toxorynchites amboinensis;) C6/36 (Ae. albopictus); AP-64
Figure 2.16 :-Diagnosis of dengue.
Modified from Peeling, R. W. et al. Evaluation of diagnostic tests: dengue (1)

79
(clone of an A. pseudoscutellaris cell line); CLA-1 (clone of an A. pseudoscutellaris);
and mammalian cell lines LLCMK2 (monkey kidney); Vero (monkey kidney); BHK21
(Baby hamster kidney) (267). ZIKV has also been isolated in these cell lines. However,
ZIKV is neurotropic and infects a wide range of cells including neural and placental
cells. Chan et al reported productive ZIKV replication in placental (JEG-3); neuronal
(SF268); muscle (RD); retinal (ARPE19); pulmonary (Hep-2 and HFL); colonic (Caco-
2); hepatic (Huh-7) cell lines; as well as nonhuman primate (Vero and LLC-MK2); pig
(PK-15); rabbit (RK-13); hamster (BHK21); and chicken (DF-1) cell lines (268).
Isolation of ZIKV has been successful in brains of suckling mice (269).
Isolated virus may be identified by immunofluoresecences assays (IFA) using virus
specific monoclonal antibodies or polyclonal antisera, and by molecular techniques
including RT-PCR. Virus isolation requires a viremic sample and is not possible in the
post-viremic phase of infection.
2.7.2. Genome detection
Molecular diagnosis of DENV and ZIKV is based on detection of viral genomic
material. These diagnostic tests result in rapid and precise identification of infecting
virus. The most widely used techniques are reverse transcriptase PCR (RT-PCR) and
real time RT-PCR. Viral RNA is extracted from serum, body fluid, tissues including
paraffin-imbedded tissues, or culture supernatant, purified, and reverse transcribed to
complementary DNA (cDNA), which serves as template for PCR. (18, 270). Detection
and typing of DENV was first shown by Lanciotti et al using a two-step nested PCR
(271), this approach was subsequently simplified to a single tube reaction with
equivalent sensitivity (272). Recent advances in DENV RT-PCR include enhanced
RNA extraction and reverse transcription and DNA amplification methods and primer
design. Primers specific for DENV serotypes, flavivirus consensus primers located in

80
different genes including E, NS1, NS3, NS5, are widely used (267, 273-275). Real-time
RT-PCR is a more sensitive method compared to conventional PCR (276) and is
quantitative. Virus can be detected directly in blood by RT-PCR and real time RT-PCR
during the viremic phase of infection. Prolonged ZIKV detection up to 80 days in
vaginal secretions and whole blood has been reported in a case study of previously
healthy, non-pregnant travellers to United states from Honduras (277). ZIKV detection
in Urine samples has been reported for notably longer period of than for serum samples
(278). Study on travel-associated ZIKV infection imported to Italy, showed ZIKV RNA
detection in whole blood was comparable to urine with 100% detection rate till 10 days
while only 33.3% plasma samples were ZIKV RNA detection (279).
Nucleic acid slot-blot nucleic acid hybridization was also used to quantitatively detect
DENV-2 using radiolabelled cDNA probe, pVV17 (280). Reverse-transcription-loop
mediated isothermal amplification (RT-LAMP) assays target the 3’ noncoding region
for rapid detection of DENV (281). RT-LAMP has also been described for detection of
ZIKV (282). Mass spectrometry, microarray techniques, and luminescence based
techniques (Luminex) are increasingly used for detection of DENV genomes (283-285).
2.7.3. Antigen detection
The most frequently detected DENV antigen in the viremic phase is NS1 protein.
Serological detection of NS1 via EIA-based approaches is accessible in resource-poor
settings and is used worldwide for DENV surveillance whenever acute phase blood
samples are available. DENV NS1 has been detected in serum of infected person as
early as day one post infection and up to 18 days post infection. Several commercial
rapid diagnostic tests (RDTs) for NS1 antigen detection, and ELISA-based NS1 kits are
available for DENV identification. Pal et al showed that ELISA-based kits showed
higher sensitivity than antigen RDT kits (286). NS1 is a highly conserved glycoprotein

81
used to detect DENV but does not differentiate among serotypes. Recent development
of anti-NS1 monoclonal antibodies has been successful in developing assays specific to
one or all serotypes. Anti-NS1 monoclonal antibodies that differentiate DENV and
ZIKV have been developed and can be used in an immunochromatography format to
identify DENV serotypes and ZIKV without cross-reaction (287). A retrospective study
in French Guiana using the Platelia Dengue NS1 Ag kit showed no false positive
dengue NS1 test results in acute-phase ZIKV infection (288). These data indicate a role
for NS1 antigen tests in differential diagnosis of DENV and ZIKV infection. Dot blot
assays directed to the envelope/membrane (E/M) antigen are also of value in detection
of viral antigen(289). Multiplex microsphere immunoassay (MIA) for the specific
diagnosis of differential virus non-structural proteins NS1 and NS5 has been reported in
ZIKV diagnosis (290).
2.7.4. Serology
Anti-DENV antibodies persist for many years after infection. Although serology is the
most accessible technique for viral diagnosis it is most challenging. The major difficulty
for serological diagnosis is the high degree of cross-reactivity among the DENV
serotypes and other flaviviruses (286). DENV-specific antibodies can be identified by
hemagglutination inhibition (HI) (291); MAC-ELISA / IgG ELISA assays (289, 292);
immunoblotting techniques (292, 293); and serum neutralisation test (294).
HI measure total anti-viral antibody titres based on the ability of virus binding antibody
to inhibit agglutination of red blood cells (RBC) by virus. For DENV diagnosis goose
RBC are generally used. HI is traditionally used to differentiate primary and secondary
infection due to its simplicity and sensitivity (289, 291). The WHO criteria for primary
infection includes HI titre <2560; in secondary infection HI titre > 2560. This test shows
cross-reactivity between DENV and related flaviviruses (295).

82
IgM antibody-capture enzyme-linked immunosorbent assay (MAC-ELISA) is used for
detection of IgM if the sample is taken within appropriate time frame. This test aims to
identify recent infection but false positive results occur in patients with malaria or
previous dengue infections (296). IgG-ELISA is likely the most widely used sreening
assay for serodiagnosis of DENV infection in non-endemic areas. In primary infection
anti-DENV IgG appears shorty after IgM antibodies 7-10 days following onset of
symptoms; in secondary infection high levels of IgG are detectable during the acute
phase. The ratio of IgM/IgG hence can be used for differentiation of primary and
secondary DENV infection (297, 298).
Immunoblotting techniques detect anti-DENV IgM or IgG dependent on sampling time.
Commercial immunoblot kits for IgM show results comparable with IgM ELISA (299).
Comparative analysis of six DENV specific IgM and IgG kit compared sensitivity,
specificity and Kappa statistics of IgM and IgG and showed that immunoassays (EIA)
from MRL Laboratories and PanBio, a dot blot assay from Genelabs, and a dipstick EIA
from Integrated Diagnostics (INDX) are useful and reliable assays for dengue
immunoblotting (16).
Serum neutralisation tests determine anti-DENV neutralising antibody titre, which is
regarded as correlation of protection. It is also used as marker of vaccine induced
immunity (300). The gold standard test for determining neutralising antibody is the
serum dilution plaque reduction neutralisation test (PRNT). However, all field isolates
of DENV are not plaque-inducing and the traditional PRNT format has been modified
with different end-point determination techniques which includes focus reduction
neutralisation test (FRNT) (301, 302); microneutralisation test (MNT) (303, 304); and
flow cytometry (305, 306) These tests are based on measuring the capacity of anti-
DENV antibody present in patient serum or plasma to inhibit infection of a cell
monolayer. Briefly, consecutive serially diluted antiserum is incubated with a

83
standardised amount of virus and this antibody-virus complex is exposed to the cell
monolayer; the highest dilution of anti-DENV that inhibits infection by 50% (or other
endpoint) is determined as the neutralisation titre. Commonly used cell lines include
Vero, C6/36, and U937-DCSIGN (6, 307, 308). All tests listed above vary in
determination of end point. In the classical PRNT this is determined by staining the
infected monolayer with dyes including methylene blue, crystal violet and neutral red,
which stain non-infected cells leaving plaques as clear areas in the cell monolayer
which are quantified. FRNT is useful for DENV strains which are not plaque-inducing
but do infect cells. FRNT measures infectious foci on the monolayer which are detected
by an immunostaining procedure using anti-DENV monoclonal antibodies or polyclonal
antisera. Liu et al reported high correlation between PRNT and FRNT in a 96 well
format (309). MNT is the cost-effective high-throughput version of neutralisation based
the technique of ELISA. Flow cytometric determination of neutralisation uses
fluorescein-conjugated anti-DENV monoclonal antibody to determine infection of cells,
to analyse 50% virus neutralisation. Kraus et al suggested flow cytometry-based
neutralisation assays offer significant advantages over PRNT, however flow cytometry
can only be performed in selected laboratories (308). Comparative evaluation of PRNT,
MNT and flow cytometry-based tests for measurement of anti-DENV neutralising
antibodies demonstrate high reproducibility and sensitivity (310).

84
2.8. Flavivirus infection in travellers
The risk of travel-associated flavivirus infection depends on availability of the vector,
population density, immune status of the traveller. Highly viremic travellers pose a
greater risk for autochthonous transmission when the vector is present and can
contribute to the global spread of infectious disease and emergence of pathogens (311).
2.8.1. Dengue
Historically DENV has been reported to spread from Africa during travel for trade, and
troop movements in World War II. Spread of disease was due resilience of vectors
which could survive in water reservoir of sailing ships. In recent times vectors are
reported to effectively move within and between countries in automobile and truck tires.
Since the advent of frequent and cheap international airline travel travellers have
become the major cause of dengue notifications in non-epidemic areas where the
vectors may or may not be present. According to the US Geosentinel Surveillance
System, in 1997-2011 11% of travel-associated disease was dengue (311).
Travel related DENV infection has increased from 2% to 16% in 15 years from 1900s to
2000s. Within 60 years the number of countries reporting dengue increased from 9 to >
more than 100 (118). During 2003-2005, 219 imported dengue cases were reported in
Germany (312). Similarly, in 2004-2009, 16-46 travel-associated dengue cases were
diagnosed per year in Denmark (313). An incidence rate of 14.6 per 1000 person-
months was reported from the Netherlands in 2006-2007 (314). The majority of travel-
associated dengue is acquired at popular tourist destinations in the tropics and sub-
tropics.
Imported dengue is mostly acquired in Asia, particularly Southeast Asia, and the
Americas. Geosentinal Survey data from 2000-2010 reports 50% of cases were
imported from Southeast Asia (315). During 2010-2013 87-112 infective person-days

85
per 100,000 travellers from Thailand was reported each year (316). Similarly, 70% of
imported dengue cases in Sweden in 2010 were reported to be acquired in Southeast
Asia (317).
In Australia, most dengue cases are travellers from endemic/epidemic areas. Local
autochthonous DENV transmission is only reported in northern Queensland where Ae.
aegypti is present. All other dengue cases in Australia are imported cases (318), with the
exception of an apparent locally-acquired DENV infection in northern Western
Australia in 2015 (73). Ae. aegypti has not been present in WA since the 1940s
however dengue represents 40% of all Indonesian-acquired infections notified to the
West Australian Department of Health. The number dengue notifications began to
increase when cheap flights to Southeast Asia were introduced in the first decade of the
21st century. Major outbreaks of dengue occurred in Southeast Asia including Bali,
Indonesia, a popular tourist destination for WA residents, in 2012, 2015 and 2016
(319).
Travellers are also sentinels for DENV endemic areas. Study of dengue in travellers
gives geospatial distribution of strains of DENV at a given time. This also explains the
pattern of spread of disease and emergence of strains of virus (312). Emergence of new
lineages has been reported in travellers. New lineage of DENV 3 and 4 was reported
from study in Germany in 2006-2015. New lineage of DENV-2 was reported in
travellers from WA was reported in 2014 (11).
2.8.2. Zika
ZIKV infection in travellers is one of the major factor for global spread of ZIKV. The
first large outbreak in the 21st century was in Yap State, Federated States of Micronesia,
in 2007 where more than 70% of the population was infected. The virus spread
throughout the Pacific and caused an outbreak in French Polynesia; the epidemic virus

86
was of the Asian lineage and was closely related to Cambodian strain (320). A
subsequent outbreak in New Caledonia was reported in 2014; the first cases were
imported from French Polynesia (321).
The first travel-associated cases in the US were reported in two scientists returning from
working in Senegal. Possible sexual transmission was also reported in the same case
(147). In 2015, ZIKV infection in a traveller returning from Maldives in Finland was
reported (147). Cases of imported ZIKV have been reported in Europe since 2013. The
first case of ZIKV infection was reported in Germany in 2013, followed by Norway and
Italy in consecutive years. After the 2015 outbreak in Brazil the number of countries
with imported Zika has increased(320). In Australia the first travel-associated ZIKV
infection was reported in 2013 in a individual returning from Jakarta, Indonesia(322).
Increase in human mobilisation has increased spread of viruses and other pathogens.
Travellers are a medium for rapid global spread of flavivirus infection.

87
Chapter 3:- Methods
3.1. Vero cells
The Vero cell line (Vero (ATCC® CCL81™)) was used throughout. This cell line is
derived from African green monkey (Cercopithecus aethiops) kidney fibroblasts. Cells
were grown in Dulbecco's minimum essential medium (DMEM) (Invitrogen) with 1%
Penicillin-Streptomycin (GIBCO, Invitrogen, 5,000 U/mL) and 1% L-Glutamine (200
GIBCO, Invitrogen mM), supplemented with 5% fetal bovine serum. All cell cultures
were maintained in 5% CO2 at 37°C in a humidified incubator.
3.2. Virus amplification
Viruses are detailed in Chapter 4. Vero cell monolayers at 80-90% confluency was
prepared. Half of the spent media was removed and 100ul of virus suspension was
inoculated onto the monolayer. The culture was incubated overnight spent media was
removed completely, followed by addition of maintenance media (DMEM
supplemented with 2% fetal bovine serum). Appearance of CPE was noted every day;
virus was harvested when ~100% CPE was obtained or before media colour changed
acidic whichever occurred first. Growth of virus was confirmed via hemagglutination
and NS1 antigen tests (Appendix 3.1). Harvested viruses were stored as 300ul aliquots
in -800C to preserve until further use.

88
3.3. Virus Titration
3.3.1. Determination of Tissue culture infectious dose (TCID50)
Virus titration was performed using ELISA to determine 50% endpoint titre using Reed
and Muench method. This titration was done to ensure optimal growth of virus to
desired concentration. Virus with titres greater than 10-4
was used in assays. The
protocol for TCID50 determination is given in Appendix 3.2.
3.3.2. Determination of Focus forming units (FFU)
a. Preparation of monolayer
The first optimisation for establishment of the focus forming assay (FFA) was to
determine the number of cells per ml required for a 24-well plate format that would
allow the cell monolayer to remain adherent for at least 5 days to support optimal
growth of all DENV strains. A range of cell concentrations was tested; optimal input for
DENV FFA was found to be 6x104 cells/ ml.
b. Virus dilution
Virus stocks were diluted from 10-1
to 10-5
in maintenance media.
c. Test establishment
500ul of 6X104 cells/ml of Vero cell in 5% DMEM were seeded into 24 well plates to
obtain confluent monolayers in 16-24 hours. 200ul of 10-folddilutions of prepared virus
stocks was inoculated onto monolayers. After one hour a methylcellulose overlay was
added to stop virus progeny from spreading around the monolayer in a secondary
infection and allowing input virus to form infectious units at a single focus, reflecting
the number of infectious units at the time of inoculation. A plate template was prepared
to run duplicate dilutions of each virus and included no-virus cell-only controls well in
every plate. Plates were carefully monitored for formation of FFU every day under a

89
phase-contrast microscope and immunostained when infectious units were apparent.
Each strain has different time-point of maturation and size of FFU. Average time for
optimal infectious unit formation for all viruses was 3-7 days. Allowing infection to
proceed past this optimal time increased the size but not the number of infectious units.
d. Detection of FFU
i. Optimisation of Fixative
To obtain optimal immunostaining without disturbing Vero monolayers three different
fixative agents were compared: 80% Methanol; 1X PBS/ACETONE; and 4%
Formaldehyde PBS. 80% methanol interfered in spot development as spots were faint
(compared to spots with other fixatives); during the drying step cells became
excessively dry and shred from the plate surface to be removed during washing.
PBS/Acetone drying also caused cells to become detached from the plates surface. Cells
were fixed properly using 4% formaldehyde and there was no cell shredding or faint
focus unit. Therefore 4% formaldehyde was used throughout to fix cells. All chemical
waste was collected in dedicated containers and disposed as per UWA chemical safety
guidelines.
ii. Optimisation of 4G2 monoclonal antibody concentration (primary antibody):
The monoclonal antibody 4G2 was used as primary antibody for detection of DENV
infectious units. Briefly, 4G2 was produced in large volumes from a hybridoma cell line
in 10% RPMI. The optimal 4G2 titre was determined by checkerboard ELISA
(Appendix 3.3). Supernatant from the hybridoma cell culture was then filtered with a
sterile vacuum filter and stored in aliquots at -80OC. The same batch of 4G2 was used
throughout.

90
iii. Optimising secondary antibody concentration
Goat Anti-Mouse IgG (H+L)-HRP Conjugate (GAM) from Biorad (#1721011) was
used as secondary antibody for ELISA and FFA. Optimal GAM concentration was
determined along with 4G2 by checkerboard assay.
iv. Immunostaining
Visualisation of infectious units was done as previously described for mumps virus
(301). The overlay was removed, monolayers rinsed with phosphate buffered saline
(PBS, pH 7.4) and then fixed with 200 μL/well of 4% formaldehyde in PBS for 30-
45 min. 2% Casein with PBS/Tween-20 was used for blocking and permeabilization.
200ul of buffer was added and plates were gently placed on a plate rocker at the lowest
speed to ensure coverage by buffer for 30 minutes at room temperature (RT). Each well
was then incubated with 200ul of 4G2 in BB buffer (1/40 as determined by
checkerboard assay) for 1 hr at RT. Cells were then washed 2 times with PBS and
incubated with GAM-HRP secondary antibody (1/1500, determined by checkerboard
assay) for 1 hr at RT. Plates were again washed 2-times with 1X PBS followed by
incubation with 150ul of TrueBlue peroxidase substrate. Substrate was incubated for
10-20 mins maximum in the dark and the reaction was stopped with water. Blue-
coloured infectious units were observed.
FFU were counted and virus titre was determined from the mean number of infectious
foci obtained from duplicate runs. Virus dilutions giving 30–60 foci per well in a 24-
well plate were selected for ongoing experiments.

91
3.3.3. Determination of Plaque forming units (PFU)
Selected viruses, including ZIKV, which form visible plaques were also used to cross-
validate FFA neutralisation data. Establishment of plaque assay was as described for
FFA. Plaques were detected by staining monolayers with 1% Methylene Blue in 10%
formaldehyde. Stained plates were stored in the dark overnight and washed with
running water.

92
3.4. Methods for Chapters 5 and 6
3.4.1. Neutralising Antibody (NAb) titre determination
3.4.1.1. Preface
The neutralisation test is based on the principle that reaction between antibody and virus
renders the virus non-infectious and thus prevents entry of virus into host cells,
preventing or reducing infection. Known concentrations of virus are incubated with
varying dilutions of antibody as serum or plasma and added to susceptible target cells.
Virus that is not neutralised by antibody will induce infection. In the PRNT plaques
formed by infecting virus are visualised as clear areas on the cell monolayer; staining
with methylene blue dye stains only live Vero cells and enhance the appearance of
plaques. In the FFA, infection is detected by staining with virus specific monoclonal or
polyclonal antibody attached to an enzyme label; addition of the respective substrate
will visualise foci of infection.
Principals of neutralisation and methods for detection are shown in Figure 1.

93
Figure 3.1: - Neutralisation
Neutralising antibody blocks entry of virus into cells hence preventing infection. In the absence of
antibody infection can be detected by formation of infectious units or distinct plaques

94
3.4.1.2. Establishment of neutralisation test
a. Preparation of monolayer
500ul of Vero cells at a concentration of 6X104 cells/ml in 5%DMEM ml were seeded
in 24-well plates to obtain confluent monolayers in 16-24 hours.
b. Preparation of plasma sample
Plasma samples used in the study are detailed in Chapter 4. 10-fold dilution of plasma
samples were prepared and heat-inactivated for 30min in a 56oC water bath. Serial 2-
fold dilutions starting from 20, to 320 were prepared for all DENV and control samples;
the dilution scheme for ZIKV samples varied based on available sample volume.
c. Focus reduction neutralisation test (FRNT)
i. Preparation of virus dilution
Virus dilutions (as determined in 3.3.2.) were prepared to obtain dilutions that produced
30-60 FFU; this was dependent on foci size in 24 well plates.
ii. Test establishment
Titrated virus with known FFU was mixed with respective plasma dilution at equal
volume and incubated for 1 hour at 37oC in 5%CO2. 200ul of plasma / virus mixture
was then added on to the Vero monolayer and incubated for 1 hour at 37oC with 5%
CO2. In each plate, two virus control wells (virus+media only; no antisera) and two cell
control wells (no virus; media only) were included. An overlay of 1% methylcellulose
in 5% DMEM with Amphotericin –B was added. Plates were incubated for 4-5 days at
37oC with 5% CO2. All test sets were performed in duplicate in two different
experimental runs.

95
iii. Immunostaining and visualisation of FFU
Immunostaining was done as optimised in section 3.3.2 (d). FFU in virus control well
and serum/plasma dilution was noted and percentage reduction at each dilution was
calculated to determine corresponding neutralising antibody titre that gave 50,75, and
90 percent reduction.
iv. Plaque reduction neutralisation test (PRNT)
Establishment of PRNT was same as for FRNT. Virus input was determined by PFU
titration in section 3.3.3. NAb titre was determined by comparison to virus control
wells.
d. Neutralisation titre (NAb) determination
For calculation of percent reduction an Excel spread sheet was created in which the
number of FFU or PFU in test wells was subtracted from the number of FFU or PFU in
the serum/plasma dilution, divided by FFU or PFU in virus control wells; the resulting
value was converted to a percentage. Neutralisation titre is expressed as the reciprocal
of serum/plasma dilution. Percent reduction value for corresponding reciprocal of
serum/plasma dilution was entered into Graphpad Prism 7.0. Nonlinear regression curve
fit was used to interpolate NAb titre at 50%, 75% and 90% reduction. A regression
analysis approach was used because of the censored nature of data (i.e. minimum of 20,
max of 320), tobit regression with random effects (the subjects) was used.
Figure 3.2 and 3.3 shows a flow chart of neutralisation establishment and detection by
FRNT. Sample FRNT and PRNT test plates are shown in Figure 3. 4
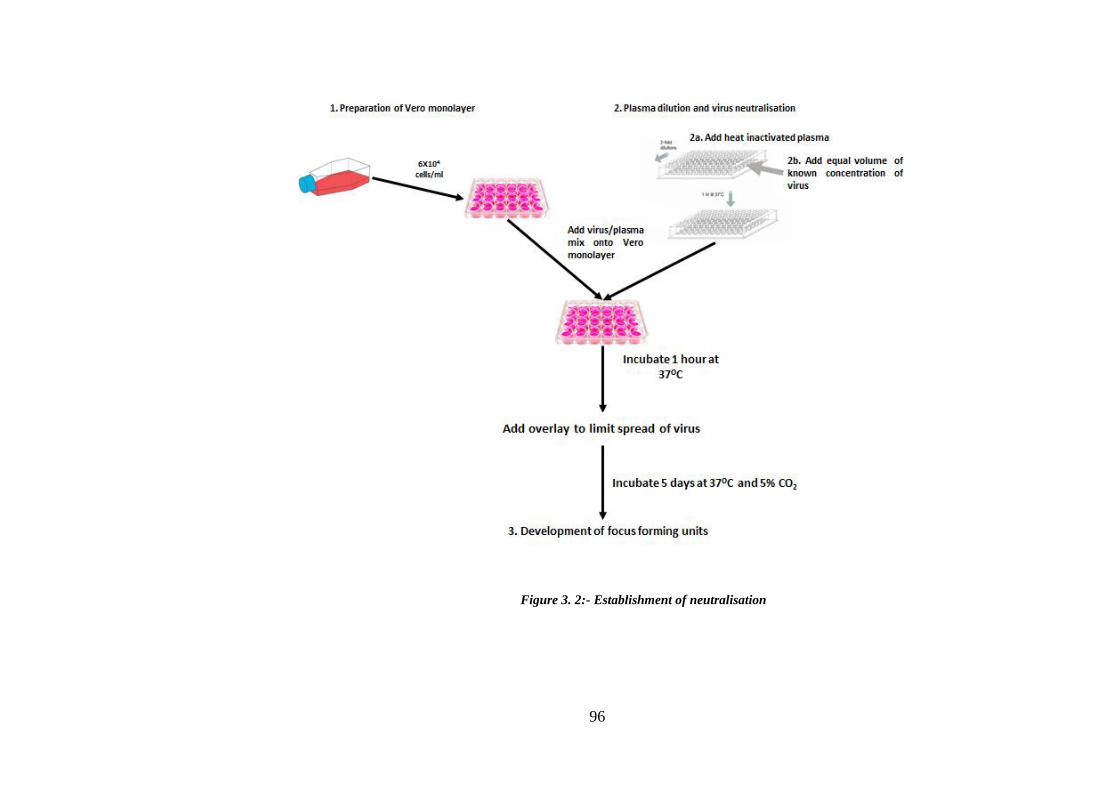
96
Flow chart for Neutralising antibody titre determination
Figure 3. 2:- Establishment of neutralisation

97
Figure 3. 3:- Immunostaining and visualisation of FFU

98
1/20
1/40
1/80
1/160
Plasma
dilution
1/320
Figure 3. 4:- FRNT and PRNT
FRNT and PRNT plates used for neutralisation assay in 24 well plate format. Virus control well
consists of virus loaded in absence of antibody representing total virus input. Each row represents
serial dilutions of plasma from 20-320-fold. Equal volume of virus was treated in each dilution. For
the virus control blank media was used as mock serum. Absence of infectious units at any dilution
represents neutralisation of virus. FRNT was used for all strains of DENV. PRNT was used in some
ZIKV cases to confirm PRNT results. Tests gave comparable results.
Virus
Control
Cell Control Virus
Control Cell Control

99
3.4.2. Hemagglutination inhibition (HI) antibody titre determination
3.4.2.1 Principle
This test is based on the principle that in the presence of antibody, binding of virus to
goose red blood cells in (GRBC) is inhibited. Virus particles attach to GRBC surface
agglutinates and form a lattice thus preventing cells falling to the bottom of V- well
plates. This phenomenon is known as hemagglutination (Figure 3.5). In the presence of
virus-specific antibody, an Ab-virus complex is formed hence no virus is available to
agglutinate GRBC, this is called hemagglutination inhibition (Figure 3.6). The
maximum dilution of Ab that inhibits hemagglutination is the antibody titre.
Figure 3. 5:- Hemagglutination of GRBC by flavivirus
Figure 3. 6:- Hemagglutination inhibition by anti-flavivirus antibody

100
3.4.2.2. Methodology to establish HI test
a. Preparation of viral antigen
Viruses for use in HI tests were amplified by cell culture. Inactivation of virus was done
with Binary Ethylenimine (BEI) as previously described (24). Virus stocks were first
centrifuged to clear dead Vero cells and treated with 0.1M Binary Ethylenimine (BEI) at
0.175N sodium hydroxide for 6-8 hours at 370C. Reaction was stopped by addition of
1M sodium thiosulphate and held overnight at 40C. After overnight incubation
inactivated virus was aliquoted and stored at -800C until further use. Preparation of all
reagents is given in Appendix 3.4.
b. Titration of inactivated virus
BEI-inactivated virus was titrated by hemagglutination assay (HA). HA was performed
in V-bottom 96 well plates. Virus was diluted in bovine albumin borate saline (BABS)
prepared from 4% bovine albumin (in saline) and borate saline at pH in 1:5 ratio. Each
virus was tested at pH 6.0, 6.2 and 6.4. Briefly, 50ul of virus dilution in BABS was
prepared in doubling dilution and 50ul of 10% Goose Red Blood cells (GRBC) was
added, followed by 1-hour incubation at room temperature. Positive reaction was
detected by a matrix of RBCs or the absence of pellet; negative reaction was indicated
by presence of a pellet.
c. Preparation of plasma sample
Non-specific inhibitors in patient serum or plasma were extracted using 20% Kaolin
suspension and 50% Goose Red Blood cells (GRBC). Briefly, 500ul of 20% Kaolin
was added to 100ul plasma samples in labelled micronic tubes. The mixture was
incubated at RT for 60 minutes with thorough mixing every 10 minutes to avoid false
positive results due to incomplete mixing. After 60 minutes tubes were centrifuged at

101
3000rpm in 40c for 10 minutes. 50ul of 50% GRBC was added to the clear supernatant
after Kaolin purification and incubated at 40C for 60 minutes mixing by inversion every
10 minutes. Mixing at this step was done carefully to avoid lysis of GRBC that can
interfere with following testing. After 60 minutes tubes were centrifuged at
3000rpm/40c for 5 minutes. Clear plasma supernatant was then aliquoted in fresh
micronic tubes and stored at 40C and used within week of preparation.
d. Establishment of test
Plate plans were prepared beforehand for 96 well v-bottom plates. On test day plates
were labelled as according to plate plan. Plasma sample prepared after kaolin treatment
was further diluted two-fold in 25ul BABS from 1:10-1:640 convalescent plasma
samples (more than two months post infection) and 1:10-1:10240 for acute phase
samples (within two months post infection). Subsequently, 25ul of viral antigen at 4HA
unit, 50ul was added to each well containing 25ul of plasma dilution. Virus-plasma
mixture was then incubated overnight at 40C. Back-titration of virus dilution was
prepared as antigen control. HI inhibition was determined by adding 10% GRBC at
appropriate pH for that particular virus as determined in part (b). Positive HI test was
detected by presence of pellet at the bottom of plate. Figure 3. 7 shows systemic
diagram for HI test performance.
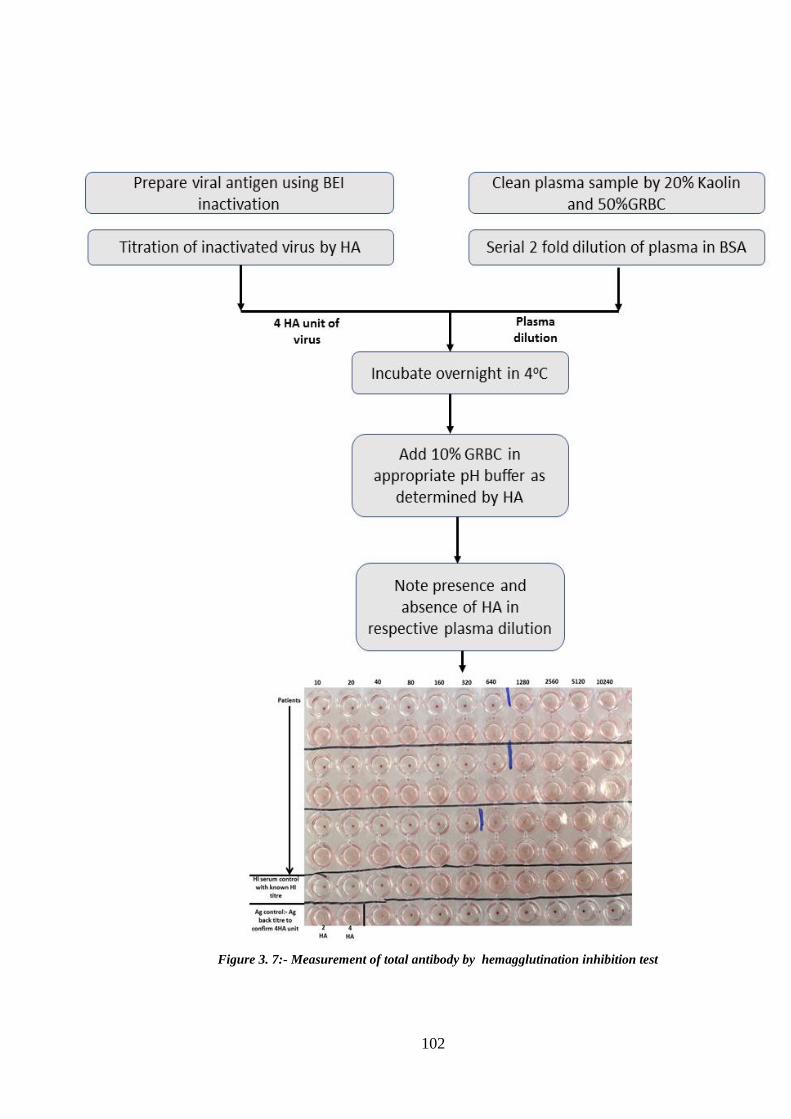
102
Figure 3. 7:- Measurement of total antibody by hemagglutination inhibition test

103
3.4.3. Mapping antigenic diversity using cartography
3.4.3.1. Preface
Neutralising antibody titres were used to create an antigenic map. Antigenic maps show
the variation in antibody responses across a range of viruses. Antigen maps were
initially used to derive antigenic distance between genetically related influenza strains
(323). In this project we expect to visualise systematic changes in neutralisation and
recognition patterns by anti-DENV antibody at different time points after infection and
to antigenically characterise DENV strains based these antibody responses.
3.4.3.2. Cartography technique
Antigenic maps were created with web based software https://acmacs-web.antigenic-
cartography.org/ designed by the Antigenic Cartography Group at the University of
Cambridge as previously described by Smith et al (323). Viruses and sera are given N-
dimensional locations on a grid in which each square side corresponds to two-fold
antiserum dilution or one antigenic unit (AU). A distance of one antigenic unit
translates to a two-fold drop in antibody titre; two square sides are four fold drop in
serum dilution, and so on (323). Briefly, the target distance from a serum to each virus
is derived by calculating the difference between the logarithm (log2) reciprocal NAb/HI
titre for that particular virus and the log2 reciprocal maximum titre achieved by that
serum (against any virus). The distance between viruses and sera in the map are
inversely proportional (323-325).

104
Figure 3. 8:- Antigenic mapping strategy

105
3.4.4. Western blot
3.4.4.1. Preface
Western blot analysis was used to investigate viral protein targets of anti-DENV and
anti-ZIKV. Vero cell lysates infected with DENV and ZIKV were gel electrophorized
through SDS-PAGE. Separated viral proteins were blotted onto nitrocellulose
membranes. Plasma from DENV- and ZIKV-infected individuals was used as primary
antibody. Viruses used in western blot analysis are ZIKV MR766 /ZIKV PRVABC59,
WHO reference strains of all four serotype of DENV (DENV-1 HW2001G4, D2-
NGCAII, D3-H87G5 and D4-H241GI) and YF17D vaccine isolate.
3.4.4.2. Methods
a. Optimisation of whole virus protein preparation
Viruses were inoculated onto Vero cell monolayers and cells were collected with a cell
scraper at 60-70% CPE. Two different techniques were used to prepare virus lysate:
radioimmunoprecipitation assay (RIPA) buffer lysis method and glutaraldehyde fixation
method, followed by sonication.
RIPA lysis buffer is widely used for preparation of protein lysates from cell culture and
to extract protein from cells and tissues for immunological assay. RIPA buffer was used
to prepare DENV lysate as previously reported (326). Scraped infected Vero cells were
lysed in RIPA buffer (ThermoFisher scientific) in the presence of 1% protease inhibitor
cocktail followed by sonication.
The glutaraldehyde fixation method was developed based on previously described
methods for DENV antigen preparation, followed by sonication to lyse fixed cells
(204). Infected cells were fixed for 20 minutes with glutaraldehyde (0.025%) at 4oC
with constant agitation in between. Cells were then sonicated at 60% pulse for 3 X 30
sec cycles or till cell suspension was clear. Comparative Coomassie stained protein

106
bands prepared by RIPA buffer lysis and glutaraldehyde fixation are shown in Figure
3.9. There are various factors that affect the stability of lysed protein. To maintain
proteins in the state at time of lysis it is critical to keep cells on ice and to use
appropriate the concentration of protease inhibitors. Based on the results obtained,
including and excluding RIPA buffer on the same batch of protein lysate,
glutaraldehyde fixation was selected for preparation of virus antigen.
Figure 3. 9:- Comparison of protein lysate preparation methods
For both methods protein samples were prepared from the same Vero cell lysates

107
b. Quantification of Protein
Protein was quantified using the bicinchoninic acid (BCA) colorimetric detection
method. Thermo Scientific™ Pierce™ BCA Protein kit was used for quantification. The
manufacturer’s protocol was followed to prepare samples. Serial dilution a of known
concentration of provided bovine serum albumin (BSA) was used as standard.
Absorbance was read using Polarstar Omega micro plate reader. MARS data analysis
software was used to construct standard curve and to quantify unknown samples.
c. Optimization of primary antibody concentration
Plasma samples from selected ZIKV and monotypic DENV subjects were used as
primary antibody. Dot blot assay was performed to optimize primary antibody
concentration for all plasma samples used. Three different concentrations of primary
antibody (1/50, 1/100 and 1/200) were tested against standardized secondary antibody
concentration (1/2000). The concentration of primary antibody that gave optimal
resolution was used in further western blot analysis. 2ul of sonicated cell lysate was
added onto nitrocellulose or PVDF membranes, allowed to dry, and the dot blot was
developed as described in the following section for immunoblot membranes.
d. SDS PAGE and Western blot
Cell lysate, prepared as described, was treated with SDS lysis buffer containing beta
mercaptoethanol (5X Laemmli sample buffer) at 95oC for 5mins. In-house prepared
12% SDS gel was used for initial optimisation of protocols. Once protein samples and
test protocol was established, a 4-15% gradient gel (Biorad TGX) was used to resolve
protein. Denatured proteins were electrophoresed along with the colour marker.
Electrophoresed protein was then blotted onto nitrocellulose membrane using trans-blot
turbo (Biorad). Semi-dry technique of transfer using homemade transfer packs was used

108
during optimisation. High background was seen for in-house transfer packs and
commercial transfer packs were used subsequently in attempts to reduce background, so
far without success. Optimisation is ongoing.
In-house transfer packs were prepared using semi-dry consumables as described in the
trans-blot turbo blotting system instruction manual. Transfer using commercial packs
was done as described by manufacturer. Transfer conditions were 30/10 mins at 25V
and 1.0A. Transfer completion was determined by complete transfer of marker.
e. Detection of viral proteins
Following transfer, membranes were dried for a few minutes to allow further protein
binding. Confirmation of transfer was done using Ponceau stain in some cases.
Membrane was then blocked for non-specific binding with 5% non-fat milk with 0.01%
Tween PBS as blocking buffer (BB) for one hour at RT on rocking platform.
Membranes were incubated with primary antibody prepared in BB (at the concentration
as determined by dot blot) overnight at 4oC on a rocking platform. The next day
membranes were washed 4X (10/5/5/5 minutes) with 0.01% tween PBS followed by
incubation with 1:2000 dilution of secondary antibody (Goat-Antihuman-HRP labelled)
for one hour at room temperature. After incubation with secondary antibody,
membranes were further washed 5X with 0.01% Tween-PBS. ECL clarity substrate
(BioRad) and Fujifilm LAS-3000 Imager was used for visualisation. The imager was set
at high precision and exposed for 30sec-1 min. Screen shots before and after exposure
was taken to determine positioning of protein bands.

109
Figure 3. 10: Flow chart of Western blot analysis

110
3.5. Method for chapter 7: - Antibody dependent enhancement
a. Cell lines
The Fc-γ receptor-bearing U937 (ATCC® CRL-1593.2™) cell line was used for
enhancement assay. The cell line was grown in RPMI (Invitrogen) supplemented with
10% fetal bovine serum, 1% Penicillin-Streptomycin (GIBCO, Invitrogen, 5,000 U/mL)
and 1% L-Glutamine (200 GIBCO, Invitrogen mM), designated as R-10. All cell
cultures were maintained in 5% CO2 at 37°C. Vero cells were used for virus titration in
plaque assays to determine enhancement of virus replication and output.
b. Virus stock and plasma samples
ADE of ZIKV PRVABC59 by anti-DENV antibody was determined by quantifying
enhanced infectious virus output from U937 cells due to the presence of antisera
obtained following DENV monotypic infection. Samples were selected to (1) assess
ADE by anti-DENV antibody at different stages of DENV infection and (2) to assess
sequential DENV and ZIKV circulation as occurred in French Polynesia in 2013.
ADE at longitudinal time points after infection was determined using a pool of
monotypic anti-DENV plasma collected within two months post infection and a pool of
plasma collected 1-6 years post-infection. Similarly, DENV-1 and DENV-3 antisera
samples collected 6 months and 1 year post-infection were selected to assess the French
Polynesia sequential DENV-1, DENV-3 and ZIKV outbreak model (327). Negative
control plasma was prepared individuals confirmed as seronegative by neutralisation
test, and the flavivirus-specific monoclonal antibody 4G2 was used as positive control.

111
c. Establishment of test
Plasma samples were serially diluted from 101 to 10
6. An equal volume of diluted
plasma mixed with ZIKV at MOI 1 was incubated for 1 hr at 37oC. The virus/plasma
mixture was then added to 2X104 U937 cells/100ul in 10% FBS RPMI in U bottom 96
well plates. Cells were infected for 18-24 hrs at 37oC. After infection cells were washed
5X with 1XPBS by centrifugation at 125g for 5 mins. Infected cells were further
incubated in the presence of lowest antibody dilution for 3 days in 10% FBS RPMI.
d. Detection of infectious units and determination of enhancement of infection.
Virus output was quantified by plaque assay. After 3 days supernatant from infected
U937 cells was harvested and inoculated onto a Vero monolayer in 24-well plate at 2-
fold and 10-1
to 10-3
dilution. Plates were incubated for 5 days until plaques were visible
in the positive control. After 5 days plates were stained overnight with 1% methylene
blue in 10% formaldehyde. PFU were counted at each sample dilution. Enhancement by
each plasma sample was tested at least twice and the average PFU/ml yield was
calculated for each dilution. A spline interpolating curve was constructed using
Graphpad prism 7.0 to graphically illustrate viral output at different plasma dilutions
and non-parametric Mann-Whitney t-test was used to determine statistical significance
of viral output in comparison with positive and negative control. Figure 3.11. shows the
flowchart to determine enhancement of ZIKV by anti-DENV antibody and Figure 3.12
shows example of PFU in the presence and absence of anti-DENV at different plasma
dilution.
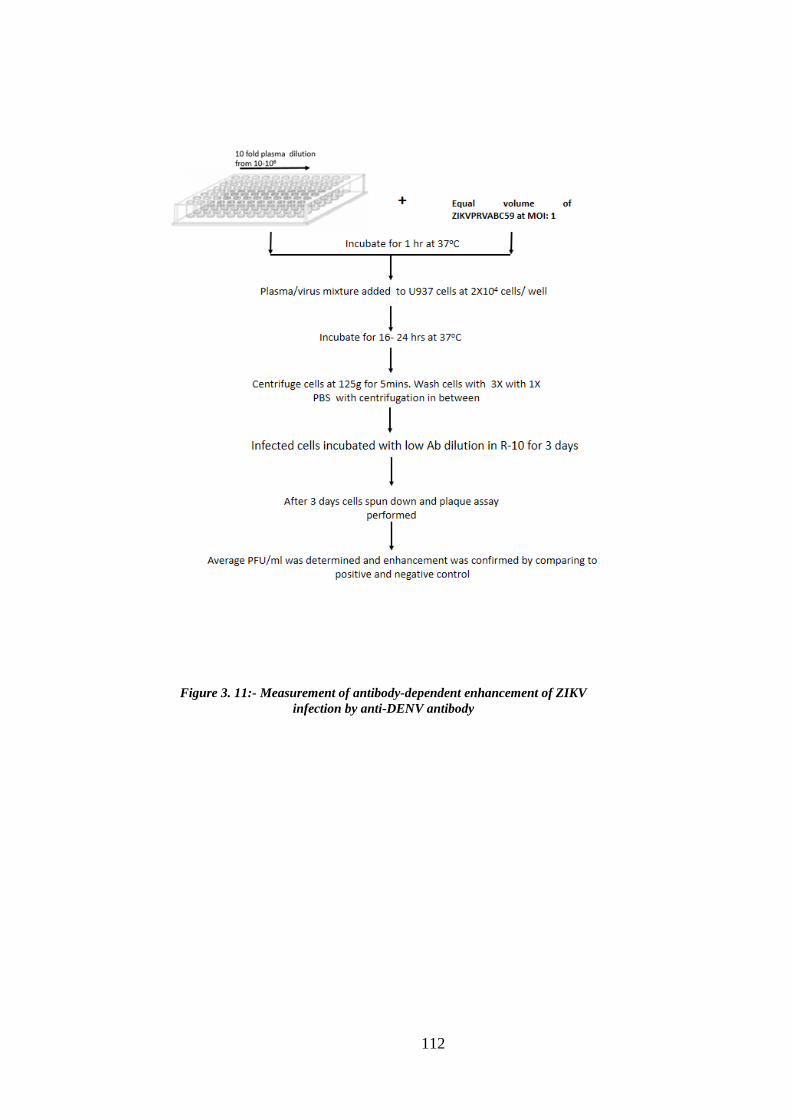
112
Figure 3. 11:- Measurement of antibody-dependent enhancement of ZIKV
infection by anti-DENV antibody

113
Figure 3. 12:- Viral load in the presence and absence of anti-DENV antibody
An example of quantification of infectious ZIKV PRVABC59 output from U937 cells in the
presence or absence of anti-DENV antibody. Supernatants from infected U937 cell lines, in the
presence of diluted antisera was inoculated on Vero monolayers to quantify virus output. Column A
is a negative control which shows virus output in the presence of varying dilutions of flavivirus-
negative plasma. Similarly, column B and C illustrate virus output in the presence of anti-DENV
antisera from monotypic travellers. Column D shows virus output in the presence of positive control
anti-flavivirus monoclonal antibody 4G2.

114
Chapter 4: Study population and viruses
4.1. Preamble
This chapter describes the study population and viruses used in this project.
Demographic and clinical information is provided. West Australian residents who travel
overseas and infected with mosquito-borne viruses are enrolled according to a study
protocol approved by the Human Research Ethics Committee, UWA. DENV field
viruses used in the study were isolated from the travellers DENV and ZIKV reference
strains and YF and JEV vaccine viruses were included in the analysis.
4.2. The cohort
Study participants were travellers returning with DENV or ZIKV infection to WA from
dengue and Zika endemic/ epidemic countries. DENV infections were monotypic.
Samples were collected from DENV subjects up to 6 years post infection and allowed
analysis of longitudinal neutralising and total antibody responses. Presentation of febrile
illness was taken as starting time of infection and time post infection was calculated
henceforth. This population of well-defined DENV monotypic infections in a non-
endemic population allows careful characterisation of DENV-specific antibody
responses over time. ZIKV-specific antibody responses were assessed in returning
travellers with monotypic infection or travellers who had previously lived in DENV-
endemic areas and who were returning to their residence in Western Australia having
acquired ZIKV overseas. Neutralising and total antibody responses were studied among
35 monotypic dengue infections, 16 ZIKV infections to generate data on specificity and
cross-reactivity against homologous heterologous and autologous viruses and
persistence and variation of these responses over time.
Traveller study volunteers donated 150ml of blood at each visit. Serum and plasma
were separated, aliquoted and stored at -80OC until use. Peripheral blood mononuclear

115
cells were isolated and cryopreserved (Figure 4.1). Information sheet and Medical/travel
history forms used during enrolment of volunteers is included in appendix 4.1- 4.3.
Figure 4. 1:- Traveller cohort establishment and virus isolation.

116
At the time of writing 90 travellers with flavivirus infection returning to WA from
different parts of the world, mostly (73%) from Bali, Indonesia were enrolled, 38 of
whom were tested in this study.
Three sera panels were assessed: A) DENV monotypic sera panel B) ZIKV sera panel
4.2.1. DENV monotypic sera panel
DENV monotypic cohort is comprised of travellers returning to WA who were infected
with DENV overseas. All participants are PCR-confirmed dengue cases. This sera panel
has 35 individuals at different time points post-infection, ranging from five days to six
years. Sera were selected randomly based on availability.
Each study subject was assigned a code: FLV and a serial number; the number of
samples is denoted by number of visits. For Eg: FLV001 V1 denotes sample from
patient with serial number 001 at first visit.
a. DENV-1 sera panel
Total and neutralising antibody from nine monotypic DENV-1 infections was studied.
Samples represent infection acquired 2010-2016 and range from nine days to 72 months
post-infection. Six of nine DENV-1 plasma samples were used to determine neutralising
and total antibody responses and three were tested for total antibody only. Eight of nine
infections were acquired in Bali and one traveller became ill after visiting Thailand.
Longitudinal samples from five individuals were included. Two individuals had history
of yellow fever vaccination. Demographic and laboratory findings are listed in Table 4.
1. Diagnostics test results were generated at PathWest Laboratory Medicine WA.

117
b. DENV-2 sera panel
Ten plasma samples were randomly selected for this sera panel from cohort of DENV-2
participants, ranging from five days to 64 months post-infection. Six of ten samples
were tested for both neutralising and total antibody response, 4 were tested for total
antibody only. Infections were acquired in Bali (7), Thailand (2) and the Philippines (1).
Two had previous history of yellow fever vaccination. Infections were acquired from
2011-2016. Summary of this panel is listed in Table 4. 2
c. DENV-3 sera panel
Ten plasma samples were selected for this sera panel, ranging from 6 to 23 months post-
infection. Six of ten samples were tested for neutralising and total antibody and four
plasma samples were tested for total antibody only. Infections were acquired after
visiting Bali (9) and Mexico, Cuba, Bahamas and USA (Florida) (1) in 2015-2016. No
individual had received yellow fever vaccine. Demographic and laboratory findings are
summarised in Table 4. 3
d. DENV-4 sera panel
Six DENV-4 sera samples were available, ranging from 6 to 72 months post-infection.
All six samples were used to measure neutralising and total antibody. Samples were
obtained in 2010, 2015 and 2016. Infections were acquired after visiting Bali (5) and
Sri Lanka followed by Philippines (1). None individual had received yellow fever
vaccine. Two of the individuals in this sera panel were tested for multiple visits at
different time points after infection. Demographic and laboratory finding of DENV-4
cohort is given in Table 4. 4

118
e. ZIKV sera panel
ZIKV panel included sera from 16 individuals. This panel is divided into confirmed and
probable cases. Eight individuals enrolled in the Traveller study and assigned a code
(FLV-) and other eight individuals tested to diagnose infection collaboration with
PathWest Laboratory Medicine WA assigned study code (ZIKV-)
Five of 16 individuals are PCR-confirmed cases and 11 are probable ZIKV cases based
on serology and clinical symptoms. The three ZIKV confirmed individuals were
enrolled in study. These cases were acquired Bali (2014), El Salvador (2015) and
Mexico (2016). The traveller returning from Bali experienced a severe febrile illness on
their return to WA; despite extensive investigation the cause of their illness was not
identified. More than one year later, in October 2015, their samples were retrieved from
storage at -20OC and tested for ZIKV by PCR (urine) and IgM IFA (serum); positive
results indicated ZIKV was circulating in Bali in December 2014. The traveller to El
Salvador was in the country for more than three months. Onset of symptoms of acute
febrile illness occurred on the day they began their journey back to WA. Traveller to
Mexico felt illness in Canada and was diagnosed there with ZIKV. Probable ZIKV
cases of enrolled in study acquired their infection in Colombia, El Salvador, Tonga and
Mexico. The travellers to El Salvador and Tonga travel there frequently to visit family;
they lived in those countries for many years prior to 2015. Three of eight enrolled
individuals have received yellow fever vaccination.
8 individuals who developed acute febrile illness after returning from India, Southeast
Asia, the Pacific, South America and West Africa, whose sera were collected for
diagnostic testing. To differentiate among flaviviruses the sera tested for ZIKV, DENV,
YF17D and IMOJEV neutralising antibody. ZIKV infection was confirmed by PCR in 2
individuals. Table 4. 5 list the demographics and laboratory finding of ZIKV panel.

119
Sample ID Gender Origin of infection Flavivirus
Vaccination
Diagnostic tests
Months post onset of illness Location Year of
infection
Duration
of Stay
(Days) Flavivirus
HI NS1
Dengue
IgM
DENV
PCR^
FLV-001 M Kuta/Bali 2010 20 No <10 + - + 24 60 72
FLV-011 F N/A 2013 N/A No <10 + - + 0.3 (9days) 34
FLV-017 F Bali 2014 10 No <10 + - + 2 20
FLV-018 F Bali 2014 10 YF, 1978 <10 + - + 2 20 31
FLV-022 F Gili Island/Bali 2015 25 No <10 + - + 14 25
FLV-026 F Bali 2015 15 No <10 + - + 14
FLV-042* M Bali 2015 3 No <10 + - + 16
FLV-076* F Legian;Canghu/Bali 2016 7 YF, 1985 <10 + - + 6
FLV-085* M Ko Samui;
Krabi/Thailand
2015 21 No <10 + - + 18
“*”: - Samples included in HI test
N/A: - information not available
“^”: Serotyping PCR specific for DENV-1
“+”: - Positive
“- “: - Negative
Table 4. 1:- Demographic and laboratory findings of individuals with DENV-1 infection

120
Sample ID Gender Origin of infection Flavi
Vaccination
Diagnostic tests Months post onset of
illness Location Year of visit Duration
of Stay
(Days) Flavi HI NS1 result Dengue
IgM
DENV
PCR^
FLV-004 M Bali 2011/2012 14 No <10 + - + 16 51 64
FLV-005 F Bali 2011 10 No <10 + + + 16 51
FLV-009 F Bali 2012 10 No 40 + + + 17 36
FLV-010 M Thailand 2012 10 No <10 + - + 19 36 48
FLV-012 F Java, Bali 2013 N/A No <10 + - + 0.1 33
FLV-014 F Bali 2013 N/A YF 2006 + - + 7 36
FLV-070* M Bali 2016 5 YF <10 + + + 7
FLV-072* M Bali 2015 10 No <10 + - + 15
FLV-073* M Thailand 2015 10 No <10 + - + 15
FLV-074* M Philippines 2015 10 No <10 + + + 13
“*”:- Samples included only in HI test
N/A: - information not available
“^”: Serotyping PCR specific for DENV-2
Table 4. 2:- Demographic and laboratory findings of individuals with DENV -2 infection
“+”: - Positive
“- “: - Negative

121
Sample ID Gender Origin of infection Flavi
Vaccination
Diagnostic tests Months post onset
of illness Location Year of
visit
Duration of Stay
(Days)
Flavi
HI
NS1
result
Dengue
IgM
DFENV
PCR^
FLV-029 M Indonesia 2015 21 No <10 + + + 12
FLV-030 F Bali 2015 8 No <10 + + + 12
FLV-033 F Bali 2015 14 No <10 + + + 14
FLV-035 F Bali 2015 >14 No <10 + + + 8
FLV-037 F Semiyak 2015 6 No <10 + - + 14
FLV-038 M Semiyak 2015 10 No <10 + - + 14 23
FLV-044* F Mexico; Cuba; Bahamas;
USA (Florida)
2015 28 No 160 + + + 8
FLV-045* M Gili Islands/ Bali 2016 11 No <10 + + + 8
FLV-046* M Manado 2015 28 No <10 + - + 18
FLV-047* F Bali 2015 14 No <10 + + + 12
Table 4. 3:- Demographic and laboratory findings of individuals with DENV -3 infection
“*”:- Samples included only in HI test
N/A: - information not available
“^”: Serotyping PCR specific for DENV-3
“+”: - Positive
“- “: - Negative
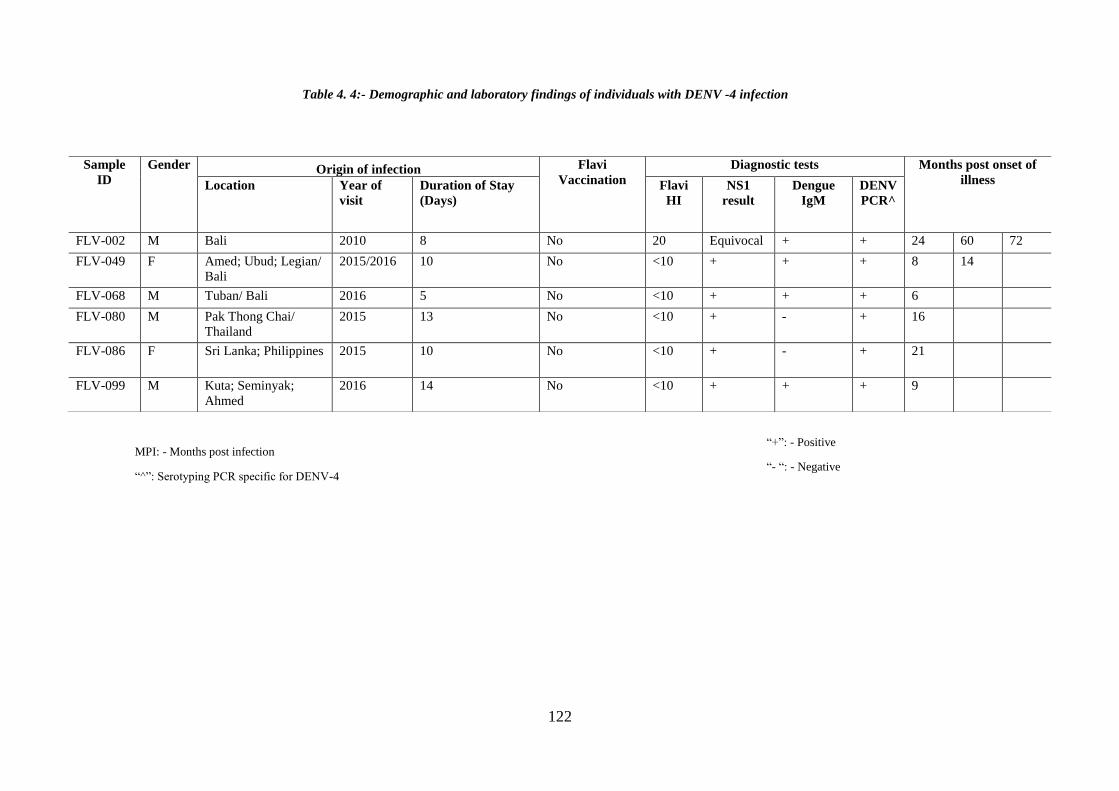
122
Sample
ID
Gender Origin of infection Flavi
Vaccination
Diagnostic tests Months post onset of
illness Location Year of
visit
Duration of Stay
(Days)
Flavi
HI
NS1
result
Dengue
IgM
DENV
PCR^
FLV-002 M Bali 2010 8 No 20 Equivocal + + 24 60 72
FLV-049 F Amed; Ubud; Legian/
Bali
2015/2016 10 No <10 + + + 8 14
FLV-068 M Tuban/ Bali 2016 5 No <10 + + + 6
FLV-080 M Pak Thong Chai/
Thailand
2015 13 No <10 + - + 16
FLV-086 F Sri Lanka; Philippines 2015 10 No <10 + - + 21
FLV-099 M Kuta; Seminyak;
Ahmed
2016 14 No <10 + + + 9
Table 4. 4:- Demographic and laboratory findings of individuals with DENV -4 infection
“+”: - Positive
“- “: - Negative
MPI: - Months post infection
“^”: Serotyping PCR specific for DENV-4

123
ZIKV case
classification
Sample
ID Gender
Origin of infection
Flavivirus
Vaccination
Diagnostic tests Samples available at
Location Year of
visit
Duration
of Stay
(Days)
Flavi HI ZIKV
IgM
ZIKV
PCR
Presentation
of illness
Days post onset
of illness
Confirmed
FLV-064 M Indonesia 2014 10 No <10 + + no 605
FLV-087 F El Salvador 2015 105 No 640 -/weakly
+ + yes 12/39/425
FLV-093 M Mexico 2016 60 YF-2015 N/A N/A + no 330
ZIKV 01 M South Pacific 2015 N/A Yes (date N/A) 20-640 + + yes 210
ZIKV 02 F Fiji 2016 N/A N/A 20 + + yes 11
Probable
FLV-032 F Colombia 2015 N/A YF-2015 160 + N/A yes 7/37/90/240/390
FLV-088 M El Salvador 2015 60 Not sure >640-320 - N/A yes 23/425
FLV-089 F El Salvador 2015 60 Not sure 320 equivocal ND yes 23/425
FLV-090 F Tonga End
2015 14 No
<10
+ ND
yes 425
FLV-091 M Mexico 2016 30 YF-2015 160 + ND yes 3/240
ZIKV 03 F Colombia 2016 N/A N/A 1280 - N/A yes 38
ZIKV 04 M Philippines 2016 N/A N/A <10 + N/A yes N/A
ZIKV 06 F Tonga End
2015 N/A N/A
<10
+ N/A
yes N/A
ZIKV 07 F Tonga End
2015 N/A N/A
<10
+ N/A
yes N/A
ZIKV 09 F India 2016 N/A N/A 10 equivocal N/A yes N/A
ZIKV011 M Thailand 2017 N/A N/A <10 + N/A yes N/A
Table 4. 5:- Demographic and Laboratory findings of individuals with ZIKV infection
MPI: - Months post infection
N/A: - information not available
ND: Not done
Flavi HI : MVE 151 antigen
ZIKV PCR: Urine
“+”: - Positive
“- “: - Negative

124
4.2. Viruses used in this study
A collection of 32 DENV isolates, two ZIKV strains, and two vaccine viruses YF17D
and IMOJEV was used to determine cross-neutralisation and cross-recognition among
flaviviruses. DENV were contemporaneous circulating viruses isolated from travellers,
and reference viruses. ZIKV strains are prototype African and Asian lineages; vaccine
viruses were included to allow assessment cross-reactivity among flaviviruses. Virus
origin and year of isolation is listed in Table 4. 6.
4.2.1 DENV Panel
DENV isolates were selected to represent genetic diversity (serotypes and genotypes),
geographic diversity (lineages) and temporal diversity (year of infection). Different
genotypes of DENV included in study are shown in Figure 4.2.
Comparison of full length E gene deduced amino acid sequences showed 98%
similarity among homologous serotypes and 60% diversity between heterologous
serotypes. Residues 1-394 in E include determinants of host range tropism, and sites
that affect binding of neutralising monoclonal antibodies (33).

125
2289 DENV-1 including 28
from WA travellers
2263 DENV-2 including 46
from WA travellers
1256 DENV-3 including 3 from
WA travellers 640 DENV-4 including 9 from
WA travellers
Figure 4. 2:- Phylogenetic tree of DENV E-sequences including isolates from travellers (11).
Virus sampled from travellers from WA is coloured in red and red boxed area represents the genotypes used in this study

126
Flavivirus
type Types
Virus definition
(Year/Location/genotype/lineage)
Code used in
study
Used for tests Genbank
accession
number FRNT HI
DENV
DENV1
2001/Hawaii/ Genotype-4 D1-HW2001G4 ▄ ● DQ672563
2010/Bali/Genotype-1/Lineage 14 D1-10BG1L14 ▄ ●
2015/Laos/Genotype-1 D1-15LG1 ▄ ●
2010/Bali/Genotype-2/Lineage 4 D1-10BG2L4 ▄ ●
2010/India/Genotype-3/Lineage 4 D1-10INDG3L4 ▄ ●
2014/Philippines/Genotype-4 D1-14PHLG4 ▄ ●
**2010/Bali/Genotype-2/Lineage 2(FLV001) D1-10BG2L2 ▄ ●
** 2014/Bali/Genotype-1/Lineage 14
(FLV018)
D1-14BG1L14 ▄ ●
**2015/Bali/ Genotype-1/Lineage H
(FLV022)
D1-15BG1LH ▄ ●
DENV2
1944/New Guinea C/ Asian II D2-NGCAII ▄ ● M29095
2010/Laos/Asian1/Lineage 9 D2-10LGA1L9 ▄ ● KM216697
2011/Bali/Cosmopolitan/Lineage 4 D2-11BGCL4 ▄ ●
*2011/Bali/Cosmopolitan/Lineage 4 D2-11BGCL4 ● KM216709
2013/Malaysia/Cosmopolitan/Lineage N D2-13MGCLN ▄ ●
2015/Bali/Cosmopolitan/Lineage 4 D2-15BGCL4 ▄ ●
2015/Bali/Cosmopolitan/Lineage N D2-15BGCLN ▄ ●
*2015/Timor
Leste/Indonesia/Cosmopolitan/Lineage G
D2-
15TLS/IDNGCLG ●
1994/ Puerto Rico/ (non-virulent strain) D2-PR1940-NV ▄ ● GQ398308
1994 /Puerto Rico/ (virulent strain) D2-PR 6913-V ▄ ● GQ398279
**2012/ Bali/Cosmopolitan/Lineage 4
(FLV004) D2-12BGCL4 ▄
**2013/ Bali/Cosmopolitan/Lineage 4
(FLV014) D2-13BGCL4 ▄
DENV3
1956/Philippines/Genotype 5 D3-H87G5 ▄ ● M93130
**2015/Bali/Genotype1 (FLV037) D3-15BG1 ▄ ●
**2015/Bali/Genotype1 (FLV038) D3-15BG1 ●
1983/Sri Lanka/(Virulent) D3-UNC3002-V ▄ ● AF547226
1989/Sri Lanka (non-virulent) D3-UNC3008-NV ▄ ● AF547232
DENV4
1956/Philippines/Genotype1 D4-H241GI ▄ ● AY947539
2010/Bali/Genotype:3/Lineage:1 D4-10BG3L1 ▄
2010/Bali/Genotype:3/Lineage:2 D4-10BG3L2 ▄ ●
*2014/Bali/Genotype:3/Lineage:2 D4-14BG3L2 ●
ZIKV ZIKV
1947/African/ Prototype ZIKV MR766 ▄ ● LC002520
2015/Puerto Rico/ Asian strain ZIKV
PRVABC59 ▄ ● KU501215
Flavi ref
strains
Yellow fever Yellow fever 17D YF-Vax YF 17D ▄ X03700
Japanese
encephalitis IMOJEV IMOJEV ▄
Kunjin Virus HI reference KUNV ●
Table 4. 6:- Flaviviruses used in study
“*”:- Tested only for total anti-DENV
antibody content
“**”:- Autologous viruses

127
a. DENV-1
Nine DENV-1 strains were used in this study. These strains represent four different
genotypes. DENV-1HW2001 used as a reference strain while all other isolates were
derived from febrile WA travellers in 2010, 2014 and 2015. Two strains represent
DENV-1 (Genotype 1) isolated in 2010 and 2015 from patients infected in Bali and
Laos respectively. Both viruses induced CPE in Vero cells although not at the same
rate: DENV-1-2015-Laos induced CPE more rapidly than DENV-1-2010-Bali. DENV-
1 (Genotype 2) acquired in Bali, 2010, was isolated from one individual. Two strains
representing DENV-1 (Genotypes 3) and DENV-1 (Genotype 4) originated in India and
Philippines in 2010 and 2014 respectively. Both viruses showed similar pattern in
inducing CPE. Three autologous viruses were isolated from individuals enrolled in the
Traveller study at the time of their acute illness; antisera were obtained from these
individuals (FLV-001; FLV-018, FLV-022) at 2 or more visits and allowed detailed
analysis of long-term immunity. Genetic diversity of DENV-1 genotypes included in
this study is shown in Figure 4.2. Comparative alignment of E-gene neutralising domain
(1-394) is shown in Figure 4.3. E gene diversity is summarised in Table 4.7.

128
Figure 4. 3:- DENV-1 E- gene (1-394) alignment
Alignment of E-gene functional domain (1-394) for DENV-1 including reference strains and patient isolates. Each virus name represents serotype year of isolation, origin,
genotype and lineage. ** :- are autologous viruses. GenBank accession: DQ672563 is strain D1-HW2001G4.

129
Amino acid position
D1-HW2001G4
D1-10BG1L14
D1-15LG1
D1-10BG2L2
D1-10INDG3L4
D1-14PHLG4
**D1-14BG1L14
**D1-15BG1LH
**D1-10BG2L4
8 N S S . . . S S .
37 D . . . N . . . .
88 T A A . A . A A .
96 F . . . . L . . .
114 I . . . L . . . .
139 V . . . . I . . .
145 N T T T T T T T T
155 T S S . . . S S .
161 I T T T . T T T .
171 S T T . . . T T .
227 S . . . . . . T .
297 I L M V V V M M T
305 S . . . . . . . P
311 E . . . D . . . .
324 V I I . . . I I .
337 F . . . I . . . .
339 S T T . T . T T .
345 V . . . . . . L .
352 V I I I I I I I .
359 T . . . . I . . .
362 E . . . . . . G .
369 A . . . T . . . .
378 I . . L . L . . .
379 V . . . . . M . .
380 V I I . I . I I .
388 K R . . . . . . .
436 V . . I . I . . .
439 I . . . V . . . .
461 I V V . V . V A .
480 I V . . . . . L .
484 M L L . . . L L .
**:- Autologous virus
Table 4. 7:- DENV-1 amino acid variation

130
b. DENV-2
This panel consists of ten strains representing two genotypes of DENV-2: Asian
1 and Cosmopolitan. DENV-2-New Guinea C was included as reference strain.
WA traveller isolates from 2010, 2011, 2013 and 2015 were used to test
antibody mediated immune response against these strains. DENV-2 (Asian 1
genotype) 2010 was acquired in Laos. DENV-2 (Cosmopolitan genotype)
strains were acquired in Bali and Malaysia 2011 and 2015. Lineage 4,
Cosmopolitan genotype is a newly identified strain that emerged in Bali,
Indonesia during a DENV-2 outbreak and was identified in WA travellers (11)
and has continued to be imported. DENV-2 isolated in Puerto Rico, PR1940 and
PR6913, represent virulent and non-virulent epidemic activity, respectively.
Non-virulent strain PR1940 induced CPE more slowly than all other DENV-2 in
this analysis. Traveller DENV-2 induced CPE at similar rates. Genetic diversity
of DENV-2 genotypes included in this study is shown in Figure 4.2. Comparison
of full length E gene sequences of viruses used this analysis is shown in Figure
4. 8. Comparative alignment of E-gene neutralising domain (1-394) is shown in
Figure 4.4. E gene diversity is summarised in Table 4.8.

131
Figure 4. 4:- DENV-2 E-gene (1-394) alignment
Functional E-domain (1-394) in all the DENV-2 isolates and reference strains. Each virus name represents serotype year of isolation, origin, genotype and
lineage. ** :- represents autologous viruses. GenBank accession: M29095.1 was used as reference sequence (D2-NGCAII)

132
Amin
o acid
Positi
on
DENV-2 sequences used in study
D2-
NGCGAII
D2-
10GAIL9
D2-
11BGCL4
D2-
11BGCL4-2
D2-
13MGCLN
**D2-
12BGCL4
**D2-
13BGCL4
D2-
15BGCL4
D2-
15BGCLN
15-
TLS/IND/GC
PR1940-
NV
PR6913
-V
47 E K K K K K K K K K K K
52 Q . H H H H H H H H . .
71 D E A A A A A A A A E E
83 N K . . . . . . . S . .
91 V . . . . . . . . . . I
126 K E E E E E E E E E E E
129 V I I I I I I I I . . .
131 Q . . . . . . . . . L L
141 I V . . . . . . . . . .
149 H . N N N N N N N N . .
164 I V V V V V V V V V . .
203 N . . . . . . . . D D D
226 T K . . . . . . . . . .
228 G E . . . . . . . . . .
308 V . . . . . . . . . I I
346 H Y . . . . . . . . . .
359 T . . . . . . . . . A .
379 I . . . . . . . . V . .
390 N . S S S S S S S S . .
402 I F F F F F F F F F F F
454 I T T T T T T T T T T T
462 I . V V V V V V V V . .
484 V I . . . . . . . . . .
491 V . . . . . . . . . A .
Table 4. 8:- DENV-2 amino acid variation
**:- Autologous virus

133
c. DENV-3
DENV-3 virus panel includes three traveller isolates obtained 2010-2015, and 3
reference strains. All DENV-3 isolated from WA travellers to date belong to Genotype-
1. The reference strain DENV3 H87 and virulent and non-virulent strains (UNC-3002
and UNC-3008) which belong Genotype 3 were also included in the analysis. Details of
virus and GenBank accession number wherever available is given in Table 4. 6.
Genetic position of DENV-3 genotype included in this study is shown in Figure 4.2.
Phylogenetic position of DENV-3 isolates shown in Figure 4. 8. Comparative alignment
of E-gene neutralising domain (1-394) is shown in Figure 4.5. E gene diversity is
summarised in Table 4.9.
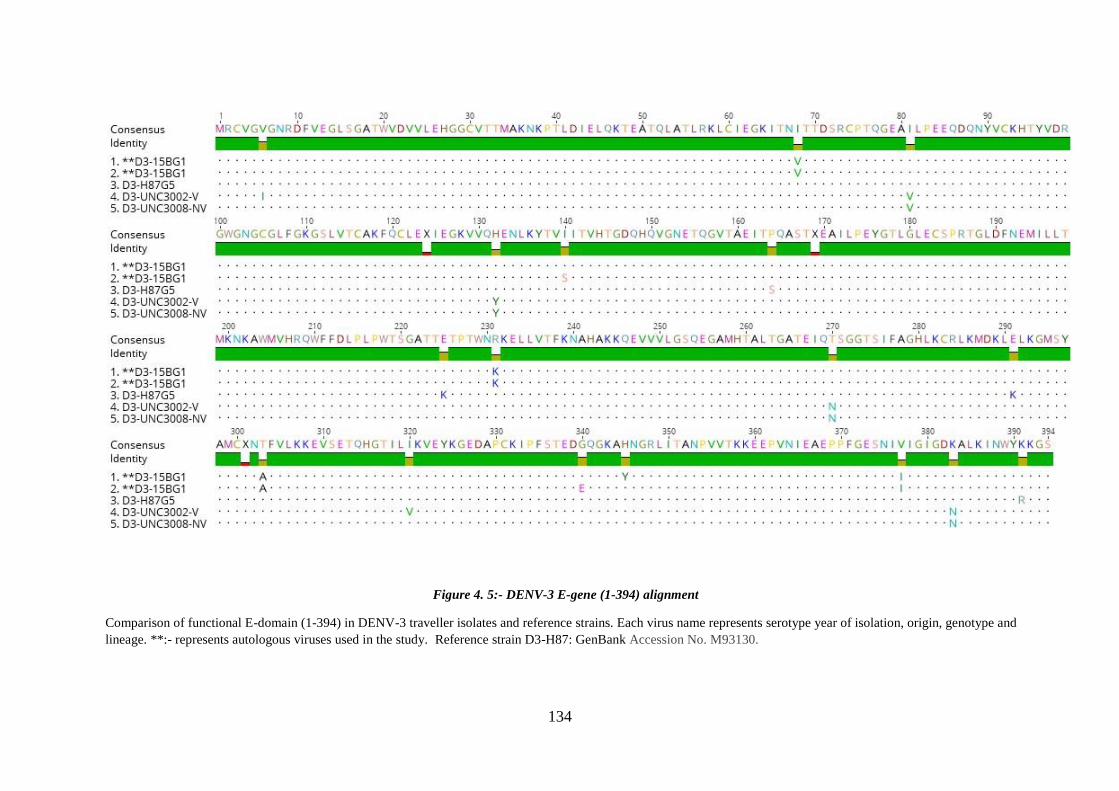
134
Figure 4. 5:- DENV-3 E-gene (1-394) alignment
Comparison of functional E-domain (1-394) in DENV-3 traveller isolates and reference strains. Each virus name represents serotype year of isolation, origin, genotype and
lineage. **:- represents autologous viruses used in the study. Reference strain D3-H87: GenBank Accession No. M93130.
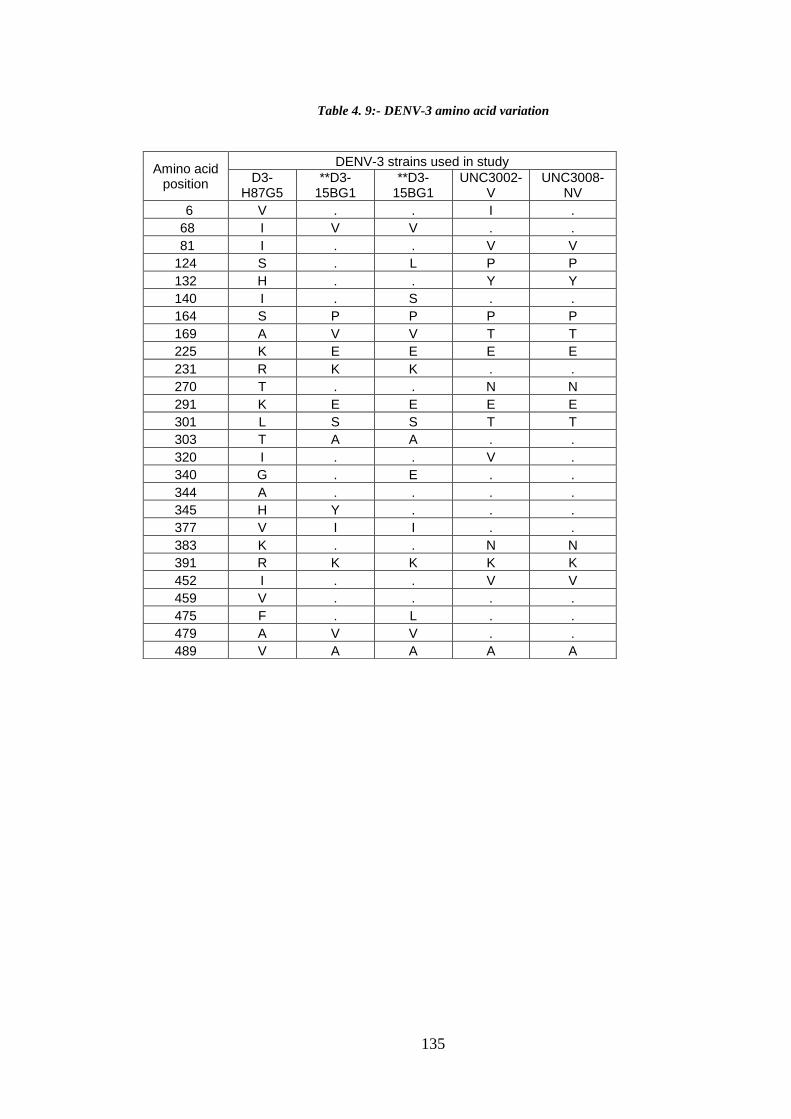
135
Table 4. 9:- DENV-3 amino acid variation
Amino acid position
DENV-3 strains used in study
D3-H87G5
**D3-15BG1
**D3-15BG1
UNC3002-V
UNC3008-NV
6 V . . I .
68 I V V . .
81 I . . V V
124 S . L P P
132 H . . Y Y
140 I . S . .
164 S P P P P
169 A V V T T
225 K E E E E
231 R K K . .
270 T . . N N
291 K E E E E
301 L S S T T
303 T A A . .
320 I . . V .
340 G . E . .
344 A . . . .
345 H Y . . .
377 V I I . .
383 K . . N N
391 R K K K K
452 I . . V V
459 V . . . .
475 F . L . .
479 A V V . .
489 V A A A A

136
d. DENV-4
Three isolates derived from travellers infected in 2010 or 2014, and one reference strain
were included. Traveller viruses belonged to Genotype 3, with 2 distinct lineages (L1
and L2). Reference strain H241 belongs to Genotype 1. Details of virus and GenBank
accession number wherever available is given in Table 4. 6. Genetic relations of DENV-
4 strains included in this study are shown in Figure 4.2. Phylogenetic distribution is
shown in Figure 4. 8. Comparative alignment of E-gene neutralising domain (1-394) is
shown in Figure 4.6. E gene diversity is summarised in Table 4.10.

137
Figure 4. 6:- DENV-4 E-gene (1-394) alignment
Alignment of E-gene functional domain (1-394) DENV-4 patient isolates and reference strains. Each virus name represents serotype year of isolation,
origin, genotype and lineage. An incomplete E-gene sequence for DENV-4 isolate (genotype 2 lineage 2) was not included in this alignment.
Reference sequence H241 GenBank accession: AY947539.1

138
Table 4. 10:- DENV-4 amino acid variation
Amino acid position
DENV-4 strains used in study
D4-H241G1
D4-10BG3L1
D4-10BG3L2
D4-14BG3L2
46 I T T T
120 S L L L
132 I . . V
155 I T T T
156 P S S S
157 N . . S
221 A T T T
233 Y N . .
265 T A A A
360 Y N N N
363 S G . .
385 S G . .
402 L F F F
429 L F F F
455 V I I I
461 F . . L
463 V . A .
494 H Q Q Q

139
e. Autologous DENV:
DENV was isolated from 7 individuals who were enrolled in the Traveller study at the
time of their acute illness. Serum samples collected at subsequent visits were assessed
for cross-reactivity and antigenicity against the same panel of homologous and
heterologous DENV serotypes, ZIKV, vaccine strains YF17D and IMOJEV, as well as
autologous DENV. Three DENV-1, two DENV-2 and two DENV-3 autologous isolates
from 2010,2012,2013,2014 and 2015 were available for study. Details of autologous
viruses are summarised in Table 4. 6.
4.2.2. ZIKV Panel
Two strains of ZIKV were included in the analysis: prototype African strain ZIKV
MR766 isolated from sentinel RHESUS monkey April 20, 1947 from Zika forest,
Entebbe, Uganda; and Asian strain ZIKV PRVABC59 isolated from a human in Puerto
Rico, 2015. NS5 sequencing and GenBank BLAST analysis confirmed that ZIKV
MR766 showed 100% homology with Uganda LC002520 and ZIKV PRVABC59 was
100% homologous with Puerto Rico KU501215. E gene diversity is summarised in
Table 4. 11. Comparative alignment of E-gene neutralising domain (1-394) is shown in
Figure 4.7.
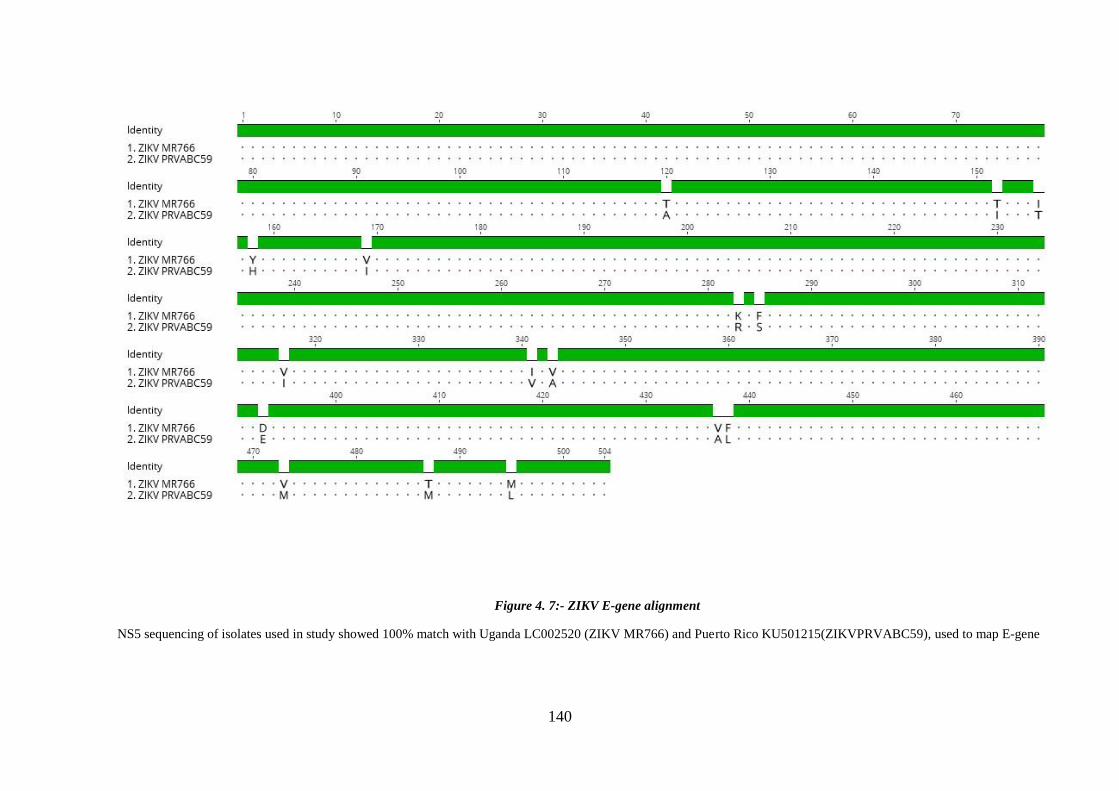
140
Figure 4. 7:- ZIKV E-gene alignment
NS5 sequencing of isolates used in study showed 100% match with Uganda LC002520 (ZIKV MR766) and Puerto Rico KU501215(ZIKVPRVABC59), used to map E-gene
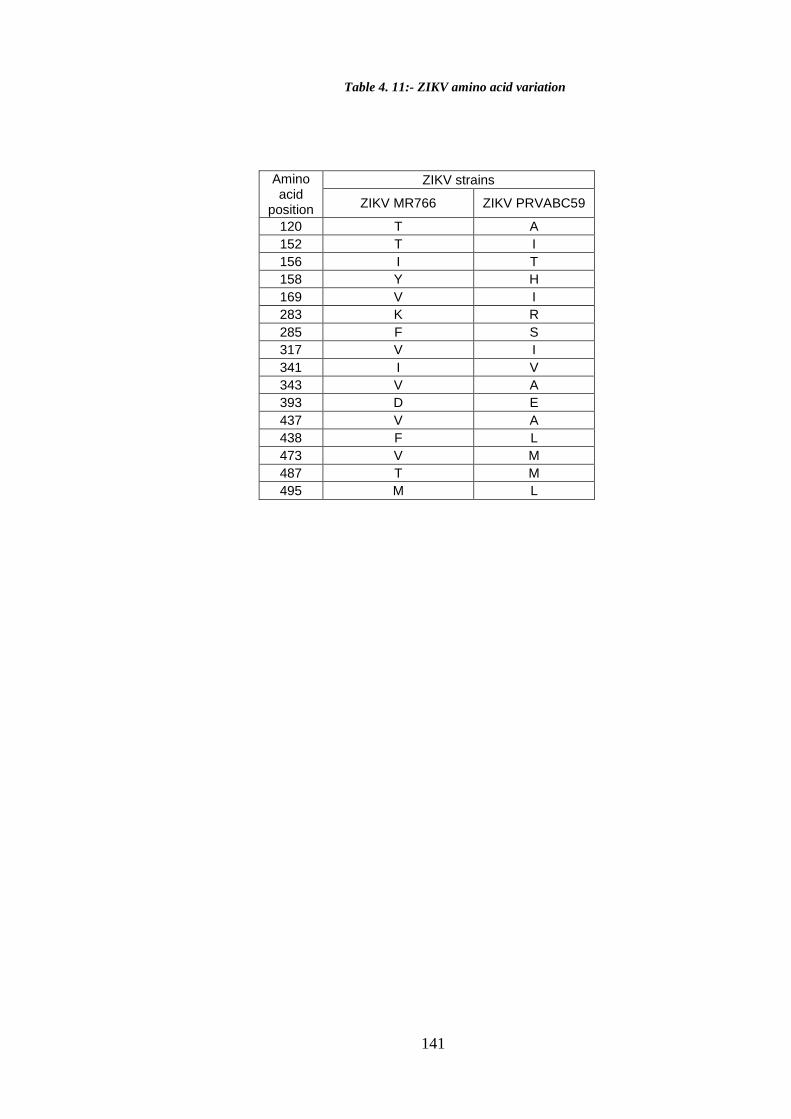
141
Amino acid
position
ZIKV strains
ZIKV MR766 ZIKV PRVABC59
120 T A
152 T I
156 I T
158 Y H
169 V I
283 K R
285 F S
317 V I
341 I V
343 V A
393 D E
437 V A
438 F L
473 V M
487 T M
495 M L
Table 4. 11:- ZIKV amino acid variation

142
4.2.3. Flavivirus vaccine strains
a. Japanese encephalitis vaccine
JEV vaccine strain was isolated from live attenuated JE vaccine “IMOJEV” (Sanofi-
Aventis Australia Pty Ltd) by inoculating Vero cells with vaccine preparation, available
lyophilised. IMOJEV is a monovalent chimeric that includes JEV prM and E coding
sequence of SA14-14-2 in a backbone of YF17D-204. JEV isolation was confirmed by
real time RT-PCR. Passage 2 of the isolate was used to determine neutralising antibody
responses to DENV- and ZIKV-infected travellers and vaccine recipients.
b. Yellow fever vaccine strain
YFV17D vaccine virus was isolated from YF-VAX, originally prepared from strain
17D-204 in living avian leucosis virus free (ALV-free) chicken embryos. The vaccine
was obtained lyophilised in sorbitol and gelatin stabilizers and was reconstituted and
inoculated onto Vero monolayer cell culture. CPE was noted every day and virus was
harvested at 5 days for first passage. In the second passage virus growth was rapid and
100 % CPE was obtained within 3 days of inoculation. YF17D isolation was confirmed
with Real time PCR. Passage 2 and Passage 3 were used for further assays.

143
Figure 4. 8:- Phylogenetic tree showing the genetic relationship among the flaviviruses strains used in this
study.
Tree was constructed using neighbour joining method with 100 bootstrap replications Different colour
represents different virus panels used in the study. Reference and vaccine strain sequences were downloaded
from NCBI GenBank. GenBank Accession numbers are listed in Table 4.1.

144
Chapter 5:- DENV-specific antibody responses in monotypic infection
5.1. Preamble
Longitudinal data on anti-DENV-specific antibody responses from the time of acute
infection to 72 months post-infection is described. Viruses and antisera were obtained
from febrile travellers returning to Western Australia and who were diagnosed with
well-characterized monotypic dengue infection upon their return. Phylogenetic analysis
of traveller DENV, originating in the Asia Pacific region (most often in Bali, Indonesia)
allowed identification of serotype, genotype and lineage; viruses represent genetic
diversity (genotypes), geographic diversity (lineages) and temporal diversity (year of
infection) within serotype. Autologous, homologous and heterologous antisera were
analysed for neutralizing and total antibody responses over time. Cross-neutralisation of
ZIKV, YF17D and IMOJEV was also assessed. Antigenic relationships among
monotypic antisera and contemporaneous circulating DENV were visualised and
quantified using antigenic cartography.
5.2. Introduction
Dengue is an acute viral infection and immunity against the infecting serotype is
thought to be lifelong (328-330). DENV-specific antibody-mediated responses post
infection include neutralising antibodies, assumed to be protective, (331) and enhancing
antibodies, assumed to be pathogenic (38, 332, 333). The high degree of genetic
homology among the four DENV serotypes means that understanding the nature of
antibody-mediated immunity is not straightforward. Although good animal models that
mirror human anti-DENV responses are lacking, IFN-knockout mice are currently one
of the best models available. Analysis in humans is complicated by the high degree of
cross-recognition among serotypes. This is particularly true for study populations in

145
dengue endemic areas, where multitypic anti-DENV responses are common once
children reach adolescence.
DENV-specific neutralising antibody (NAb) is a marker of infection and vaccine-
mediated protection. NAb epitopes are situated on the dengue envelope protein (E) on
the virion surface. Anti-E NAb block the interaction between host and virus by binding
to epitopes that facilitate virus entry into target cells (231). Flavivirus envelope proteins
consist of three non-overlapping antigenic domains DI, DII and DIII, each domain has
multiple epitopes. DIII is believed to be the specific NAb -binding epitope while cross-
reactive antibodies are also reported to bind to DII (236, 334). Conserved residues and E
protein hydrophilicity facilitates shared E epitopes and cross-neutralisation of closely
related flaviviruses. Anti-DENV neutralisation capacity in vitro is correlated with
development of E-specific humoral immune responses in vivo (335). Variation in viral
antigenic properties is due to amino acid variation that alters antibody binding site, thus
affecting binding of antibody to viral epitopes (325). Minor changes in AA orientation
can cause noticeable immunological variation, however major AA changes may not
result in immunological change (203). Seven AAs differences in E domain I and II was
proposed to result in displacement of the dominant circulating DENV-2 lineage in
Brazil, in 2007/2008 (336). Single AA residue change in DENV-1 E-gene at virus
breathing site was reported to alter neutralisation susceptibility (337). In secondary
dengue infection antibody responses directed against infecting serotype may be
predominantly derived from repertoires induced during previous flavivirus infection.
This phenomenon is termed original antigenic sin (244) and is a consequence of the
high degree of homology among flaviviruses (246).
The Study. This study, in a cohort of travellers with well-defined monotypic DENV
infection, aimed to characterize persistence of autologous, homologous and

146
heterologous DENV-specific neutralizing and total antibody responses. Antigenic
relationships among 35 monotypic antisera and 32 contemporaneous circulating DENV
were visualised and quantified using antigenic cartography.
Antigenic cartography was utilized to define antibody and antigenic diversity over time.
Antigenic cartography is derived from geographic cartography to determine relative
distance between places and derives the distance between antigenically variable
pathogens and host response based on antibody binding assay. This was first
implemented to describe emergence trends in Influenza (323). Antigens and anti-sera
are projected on a grid where corresponding distances are derived from antibody titre
obtained by total antibody (hemagglutination inhibition) or neutralising antibody. This
antigenic mapping creates a unified framework to analyse results obtained from
antibody-dependent tests, despite random error due to assay conditions and variability
among patients’ immune history (338, 339). This allows quantitative interpretation and
easy visualisation of antibody response data (340).
5.3. Aims
To characterize persistence of autologous, homologous and heterologous
DENV-specific neutralizing and total antibody responses
To determine antigenic relationships among monotypic antisera and
contemporaneous circulating DENV and to visualise and quantify antigenicity
using antigenic cartography
5.4. Results
5.4.1. Neutralising antibody
Serum neutralisation data are reported here in sections according to the infecting virus,
hereafter referred as sera panel. Each sera panel details neutralisation against (i)
infecting DENV serotype i.e. homologous serotype, (ii) other DENV serotype i.e.

147
heterologous serotype and (iii) DENV autologous virus i.e. virus isolated from patient’s
own acute sera (iv) other flavivirus serogroups i.e. ZIKV, YF17D and IMOJEV.
Data for each sera panel is presented in order of increasing months past infection.
Columns are colour coded for the same patient at different time points. Mean
neutralisation reduction for all individuals is shown in Appendix 5.1. Increase in
stringency increases serotype-specificity; however this also leads to missing results
from individual with low magnitude immune responses. Only 50% reduction did not
exclude homologous serotype NAb responses, across all serotypes (DENV-1 – DENV-
4, as shown in Tables 5.1-5.4; 5.5-5.8; 5.9-5.12; 5.13-5.16) and thus 50% reduction was
used throughout.
Neutralisation data was statistically analysed considering each FRNT 50 measurement
as a function of the person, the panel, the virus, and time since infection. Because the
measurements are restricted at both the lower and upper level, tobit regression (341)was
chosen for initial analyses. This method was derived for use with these kinds of data
where measurements cannot be taken below or above certain values. The method
assumes that there is some latent (or unobservable) variable that is being measured
imperfectly, such that below one level or above another, only the values of the levels are
known, but that between those levels the observed value equals the value of the latent
variable. The regression method then estimates the effect of various predictors (such as
time, strain similarity and their interactions) on this latent variable. In all analyses the
response variable in the regression was the natural log of the FRNT 50, with the
censoring cut-offs set to 2.99 and 5.77 as data points were between 20-320. An
alternative fairly robust method that can be (and is often) used with data restricted in
this way, is to simply replace those below the lower limit with half that value and those
above the upper limit with double that value (342).This method was also used and the

148
results compared with those obtained using tobit regression (343).Estimate of variance
on the same person at different times after infection was done with robust variance
estimation of confidence intervals for the regression coefficients. The regression
coefficients together with their robust standard errors then provide measures of the
differences in log(FRNT50) (and their statistical significance) according to: panels,
whether serotypes are homologous or heterologous, time since infection (both linear and
non-linear), whether a reference strain or not, and the individual viruses.
a. Specificity and cross-reactivity post DENV-1 infection
DENV-1 antisera showed differential neutralisation among 6 homologous and 15
heterologous strains over time. Neutralisation against homologous serotype was ~85 %
greater than against heterologous serotype. Figure 5. 1 shows an example of acute and
convalescent anti-DENV-1 NAb in a DENV-1-infected individual. Patient FLV-011
acute phase antisera neutralized all homologous DENV-1 and heterologous DENV
strains at high magnitude, whereas convalescent sera, 2 years after infection,
differentially neutralized homologous and heterologous DENV. Of note, one
homologous virus, strain D1-10INDGL4, was not neutralized by convalescent anti-
DENV-1 antiserum, whereas two heterologous DENV strains, both serotype 3, were
neutralized.
The DENV-1 sera panel consists of 6 individuals with monotypic DENV-1 infection,
sampled less than 1 week to 6 years post infection. Sera from 3 individuals were
collected within 2 months of infection and after 1 year of infection; from 2 individuals
after 1 and 2 years of infection; and from 1 individual 2, 5, and 6 years after infection
(Table 4.1 also shown below). Neutralisation titres at 50%, 75% and 90% reduction
were compared (Table 5. 1- Table 5. 3). Example of mean reduction of neutralisation is
shown in Figure 5.1.

149
Statistical verification for each FRNT-50 measurement was done using tobit regression
analysis and because there were repeated measures on the same person at different times
after infection, robust variance estimation was used to estimate confidence intervals for
regression coefficients. The regression coefficient together with robust standard errors
measures the difference in log (FRNT50) and statistical significant according to
serotypes, time post infection and individual viruses.
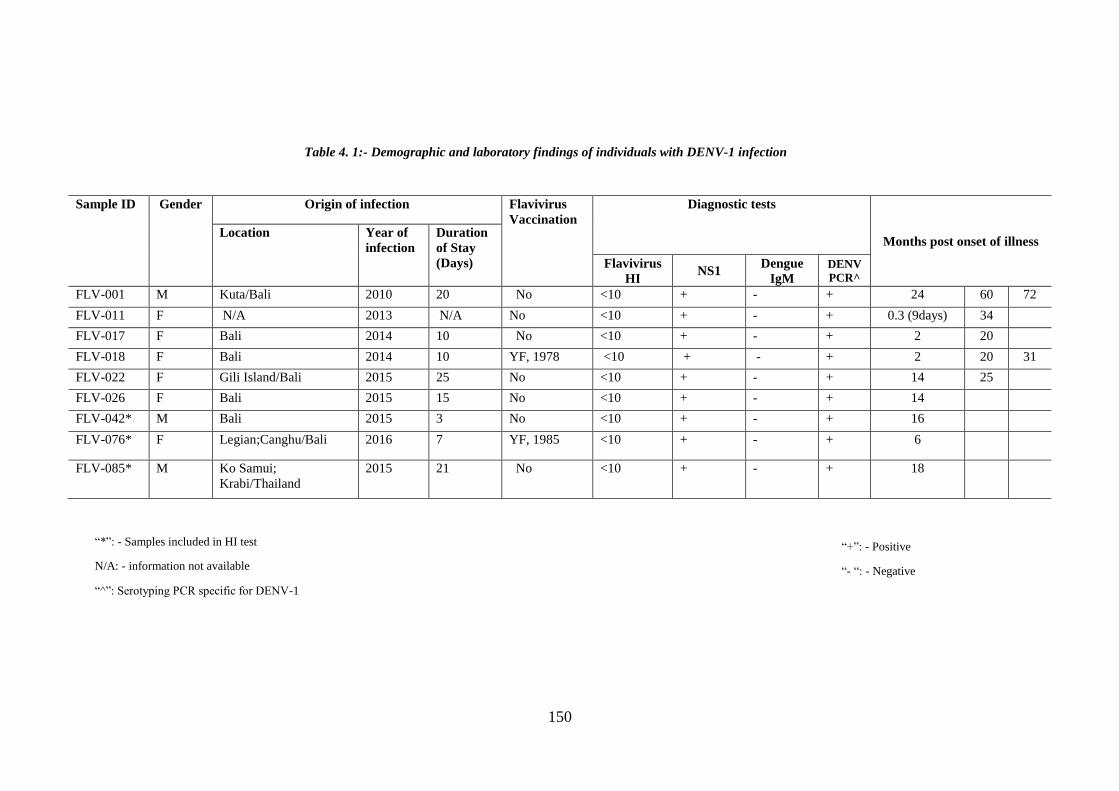
150
Sample ID Gender Origin of infection Flavivirus
Vaccination
Diagnostic tests
Months post onset of illness Location Year of
infection
Duration
of Stay
(Days) Flavivirus
HI NS1
Dengue
IgM
DENV
PCR^
FLV-001 M Kuta/Bali 2010 20 No <10 + - + 24 60 72
FLV-011 F N/A 2013 N/A No <10 + - + 0.3 (9days) 34
FLV-017 F Bali 2014 10 No <10 + - + 2 20
FLV-018 F Bali 2014 10 YF, 1978 <10 + - + 2 20 31
FLV-022 F Gili Island/Bali 2015 25 No <10 + - + 14 25
FLV-026 F Bali 2015 15 No <10 + - + 14
FLV-042* M Bali 2015 3 No <10 + - + 16
FLV-076* F Legian;Canghu/Bali 2016 7 YF, 1985 <10 + - + 6
FLV-085* M Ko Samui;
Krabi/Thailand
2015 21 No <10 + - + 18
“*”: - Samples included in HI test
N/A: - information not available
“^”: Serotyping PCR specific for DENV-1
“+”: - Positive
“- “: - Negative
Table 4. 1:- Demographic and laboratory findings of individuals with DENV-1 infection

151
Figure 5. 1:- Neutralising antibody response post DENV-1 infection, patient FLV-011
Specificity and cross-reactivity of antibody response post DENV-1 infection.

152
Note:
Value 1=<20
Value 320=>320
Table 5. 1:- Anti-DENV-1 neutralising antibody titre at 50% reduction
Note:
Value 1=<20
Value 320=>320
*MPI: Months Past Infection
*Acute: <=2 months past infection
*Convalescent: > 2 months past infection
FLV011/ 9
days
FLV018/
2
FLV017/
2
FLV022/
14
FLV026/
14
FLV018 /
20
FLV017/
20
FLV001 /
24
FLV022/
25
FLV018/
31
FLV011/
34
FLV001/
60
FLV001 /
72
D1-HW2001G4 320 320 242 320 204 263 54 320 320 278 81 320 248 227
D1-10BG1L14 320 320 320 320 320 320 193 320 320 185 320 320 320 295
D1-15LG1 320 320 65 320 223 246 52 320 108 314 256 320 320 212
D1-10BG2L4 320 135 129 320 154 122 244 320 320 89 320 320 320 218
D1-10INDG3L4 320 320 189 320 35 136 145 320 320 82 67 320 242 178
D1-14PHLG4 320 320 320 320 198 320 98 320 320 276 247 320 320 273
GMT-D1 320 277 184 320 158 219 111 320 267 179 181 320 293 230
D2-NGCGAII 320 320 240 61 1 94 39 79 161 23 1 252 70 53
D2-10LGA1L9 320 122 79 189 41 77 1 56 87 81 34 229 294 73
D2-11BGCL4 108 1 1 22 22 27 1 1 46 1 24 22 45 9
D2-13MGCLN 187 1 23 41 1 46 1 1 51 46 20 24 40 13
D2-15BGCL4 320 209 193 20 1 1 39 1 1 1 1 27 28 10
D2-15BGCLN 186 1 1 50 1 37 1 1 51 51 1 70 44 9
D2-PR6913-V 171 44 44 22 1 1 21 1 40 1 1 106 60 11
D2-PR1940-NV 320 320 122 50 1 22 21 106 139 49 47 130 89 59
GMT-D2 226 24 29 43 2 17 5 5 42 11 5 73 63 20
D3-H87G5 320 37 73 37 1 26 21 47 114 1 1 52 26 21
D3-15BG1 320 42 77 320 1 42 60 116 130 55 106 320 320 82
D3-UNC3002-V 123 21 1 41 23 35 1 70 42 1 1 40 35 13
D3-UNC3008-NV 304 66 184 282 57 134 1 40 75 30 135 54 36 62
GMT-D3 249 38 32 108 6 48 6 63 83 6 11 78 57 34
D4-H241G1 320 245 246 137 57 82 21 320 93 47 46 137 111 107
D4-10BG3L1 320 58 1 80 25 45 1 113 1 1 1 77 37 14
D4-10BG3L2 320 85 82 93 22 33 36 102 67 1 31 201 114 53
GMT-D4 320 107 27 100 31 49 9 155 18 4 11 128 78 43
ZIKV MR766 320 320 88 43 28 199 1 71 21 152 1 58 58 43
ZIKV PRVABC59 1 1 1 1 1 1 1 1 1 1 1 1 1 1
YF 17D 64 320 28 1 1 269 1 1 1 217 1 1 1 6
IMOJEV 223 1 1 1 1 1 1 1 1 1 1 1 1 2
GMT 46 18 7 3 2 15 1 3 2 13 1 3 3 5
Other
Flaviviruses
GMT-flavi
acutes: 18
(<20-320+)
GMT-flavi
convalescent: 3
(<20-320+)
GMT-flavi:5 (<20-320+)
Heterologous
serotype
GMT-
heterologous
serotype= 31
(<20-320+)
GMT-D2 strains
convalescent: 15
(<20-294)
GMT-D2 all
strains:28
(<20-320+)
GMT-D3
strains
acutes: 67
(<20-320+)
GMT-D3 strains
convalescent: 28
(<20-320+)
GMT-D4
strains
acutes: 97
(<20-320+)
GMT-D4 strains
convalescent: 33
(<20-320+)
GMT-D3 all
strains:34
(<20-320+)
GMT-D4 all
strains:43
(<20-320+)
GMT-D2
strains
acutes: 54
(<20-320+)
Study No/MPI
Virus<1 year
Homologous
serotype
GMT-D1
strains
acutes: 254
(65-320+)
1- 2 year 2- 6 yearsGMT
GMT-D1 strains
convalescent:224
(35-320+)
GMT-homologous
serotype=230 (35-320+)
Geometric Mean Titer (GMT)

153
Table 5. 2:- Anti-DENV-1 neutralising antibody titre at 75% reduction
*MPI: Months Past Infection
*Acute: <=2 months past infection
*Convalescent: > 2 months past infection
Note:
Value 1=<20
Value 320=>320
FLV011/
9 days
FLV018
/ 2
FLV017/
2
FLV022/
14
FLV026/
14
FLV018 /
20
FLV017
/ 20
FLV001/
24
FLV022/
25
FLV018/3
1
FLV011/
34
FLV001/
60
FLV001 /
72
D1-HW2001G4 306 312 121 320 75 112 28 320 320 74 38 240 98 137
D1-10BG1L14 320 320 320 320 320 320 76 198 288 98 232 150 320 230
D1-15LG1 214 103 35 320 87 130 26 245 46 161 128 320 320 125
D1-10BG2L4 320 44 43 206 42 54 82 145 211 37 112 320 320 109
D1-10INDG3L4 320 320 37 320 25 77 55 320 320 158 39 320 120 130
D1-14PHLG4 320 218 86 320 116 320 45 320 187 64 91 320 320 172
GMT-D1 297 178 76 297 79 135 47 248 194 87 88 269 223 146
D2-NGCGAII 320 274 124 28 1 1 1 42 28 1 1 143 37 15
D2-10LGA1L9 320 57 34 74 1 28 1 29 74 29 1 54 82 22
D2-11BGCL4 22 1 1 1 1 1 1 1 1 1 1 1 1 1
D2-13MGCLN 68 1 1 1 1 1 1 1 1 1 1 1 1 1
D2-15BGCL4 320 53 57 1 1 1 1 1 1 1 1 1 1 3
D2-15BGCLN 63 1 1 24 1 1 1 1 24 1 1 1 24 3
D2-PR6913-V 56 1 1 1 1 1 1 1 1 1 1 30 28 2
D2-PR1940-NV 320 72 45 23 1 1 1 47 23 1 1 46 37 11
GMT-D2 124 9 8 6 1 2 1 4 6 2 1 8 10 4
D3-H87G5 139 1 32 1 1 1 1 24 47 1 1 22 1 4
D3-15BG1 320 24 29 54 1 25 31 56 50 1 54 320 320 35
D3-UNC3002-V 37 1 1 1 1 1 1 1 1 1 1 1 1 1
D3-UNC3008-NV 115 1 1 66 1 25 1 1 37 1 1 25 21 5
GMT-D3 117 2 6 8 1 5 2 6 17 1 3 20 9 6
D4-H241G1 320 119 114 58 1 34 1 137 44 24 13 62 47 33
D4-10BG3L1 161 1 1 1 1 23 1 1 1 1 1 1 1 2
D4-10BG3L2 320 26 21 35 1 1 1 34 32 1 1 33 1 7
GMT-D4 255 15 13 13 1 9 1 17 11 3 2 13 4 8
ZIKV MR766 288 157 1 1 1 85 1 32 1 47 1 45 29 10
ZIKV PRVABC59 1 1 1 1 1 1 1 1 1 1 1 1 1 1
YF 17D 21 285 1 1 1 76 1 1 1 80 1 1 1 4
IMOJEV 35 1 1 1 1 1 1 1 1 1 1 1 1 1
GMT 22 15 1 1 1 9 1 2 1 8 1 3 2 3
Study No/MPI
Heterologous
serotype
Homologous
serotype
GMT-D4 all
strains:8
(<20-320+)
Other
Flaviviruses
<1 year 1- 2 year
GMT-flavi
acutes: 7 (<20-
288)
GMT-flavi
convalescent: 2
(<20-45)
GMT-flavi:2 (<20-288)
Geometric Mean Titer (GMT)
GMT-D1
strains
acutes:159
(37-320+)
GMT-D1 strains
convalescent:
142(25-320+)
GMT-homologous
serotype=146 (25-320+)
2-6 yearsVirus
GMT
GMT-D2
strains
acutes: 21
(<20-320+)
GMT-D2 strains
convalescent: 3
(<20-143)
GMT-D2 all
strains:5
(<20-320+)
GMT-
heterologous
serotype= 6
(<20-320+)GMT-D3
strains
acutes: 11
(<20-320+)
GMT-D3 strains
convalescent: 5
(<20-320+)
GMT-D3 all
strains:6
(<20-320+)
GMT-D4
strains
acutes: 37
(<20-320+)
GMT-D4 strains
convalescent: 5
(<20-137)
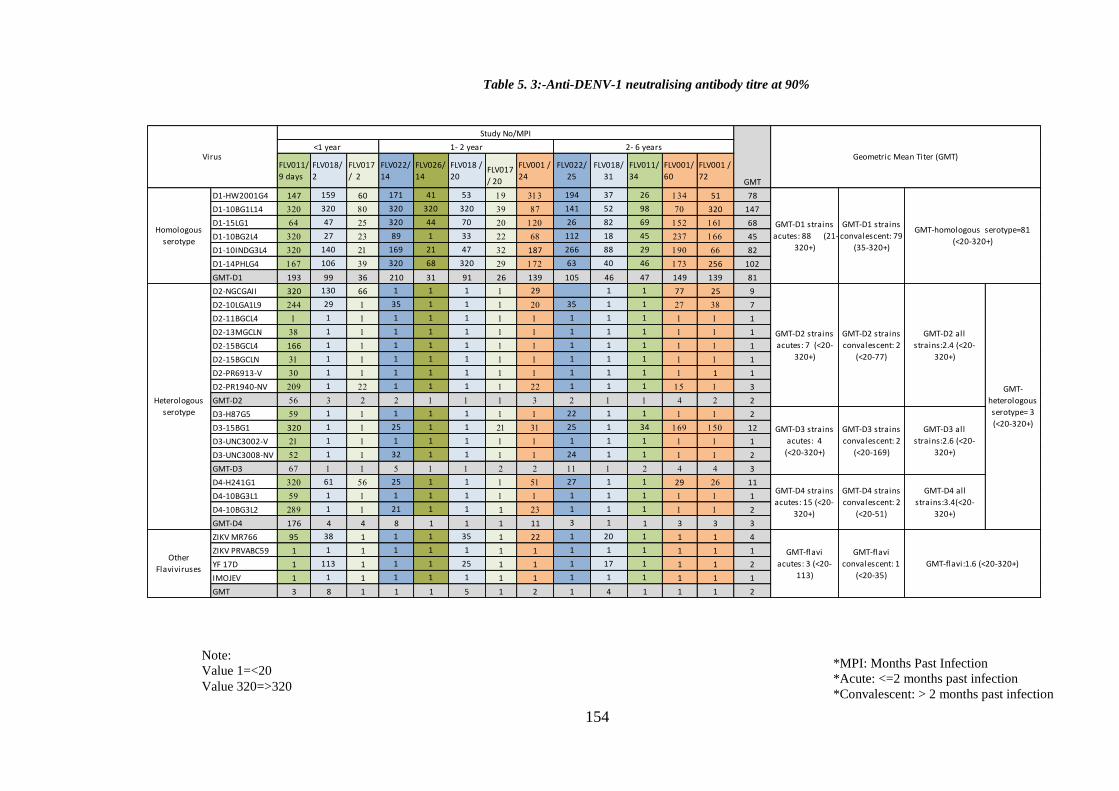
154
*MPI: Months Past Infection
*Acute: <=2 months past infection
*Convalescent: > 2 months past infection
Note:
Value 1=<20
Value 320=>320
Table 5. 3:-Anti-DENV-1 neutralising antibody titre at 90%
FLV011/
9 days
FLV018/
2
FLV017
/ 2
FLV022/
14
FLV026/
14
FLV018 /
20FLV017
/ 20
FLV001 /
24
FLV022/
25
FLV018/
31
FLV011/
34
FLV001/
60
FLV001 /
72
D1-HW2001G4 147 159 60 171 41 53 19 313 194 37 26 134 51 78
D1-10BG1L14 320 320 80 320 320 320 39 87 141 52 98 70 320 147
D1-15LG1 64 47 25 320 44 70 20 120 26 82 69 152 161 68
D1-10BG2L4 320 27 23 89 1 33 22 68 112 18 45 237 166 45
D1-10INDG3L4 320 140 21 169 21 47 32 187 266 88 29 190 66 82
D1-14PHLG4 167 106 39 320 68 320 29 172 63 40 46 173 256 102
GMT-D1 193 99 36 210 31 91 26 139 105 46 47 149 139 81
D2-NGCGAII 320 130 66 1 1 1 1 29 1 1 77 25 9
D2-10LGA1L9 244 29 1 35 1 1 1 20 35 1 1 27 38 7
D2-11BGCL4 1 1 1 1 1 1 1 1 1 1 1 1 1 1
D2-13MGCLN 38 1 1 1 1 1 1 1 1 1 1 1 1 1
D2-15BGCL4 166 1 1 1 1 1 1 1 1 1 1 1 1 1
D2-15BGCLN 31 1 1 1 1 1 1 1 1 1 1 1 1 1
D2-PR6913-V 30 1 1 1 1 1 1 1 1 1 1 1 1 1
D2-PR1940-NV 209 1 22 1 1 1 1 22 1 1 1 15 1 3
GMT-D2 56 3 2 2 1 1 1 3 2 1 1 4 2 2
D3-H87G5 59 1 1 1 1 1 1 1 22 1 1 1 1 2
D3-15BG1 320 1 1 25 1 1 21 31 25 1 34 169 150 12
D3-UNC3002-V 21 1 1 1 1 1 1 1 1 1 1 1 1 1
D3-UNC3008-NV 52 1 1 32 1 1 1 1 24 1 1 1 1 2
GMT-D3 67 1 1 5 1 1 2 2 11 1 2 4 4 3
D4-H241G1 320 61 56 25 1 1 1 51 27 1 1 29 26 11
D4-10BG3L1 59 1 1 1 1 1 1 1 1 1 1 1 1 1
D4-10BG3L2 289 1 1 21 1 1 1 23 1 1 1 1 1 2
GMT-D4 176 4 4 8 1 1 1 11 3 1 1 3 3 3
ZIKV MR766 95 38 1 1 1 35 1 22 1 20 1 1 1 4
ZIKV PRVABC59 1 1 1 1 1 1 1 1 1 1 1 1 1 1
YF 17D 1 113 1 1 1 25 1 1 1 17 1 1 1 2
IMOJEV 1 1 1 1 1 1 1 1 1 1 1 1 1 1
GMT 3 8 1 1 1 5 1 2 1 4 1 1 1 2
Other
Flaviviruses
GMT-homologous serotype=81
(<20-320+)
GMT-D2 strains
acutes: 7 (<20-
320+)
GMT-D2 strains
convalescent: 2
(<20-77)
GMT-D2 all
strains:2.4 (<20-
320+)
GMT-
heterologous
serotype= 3
(<20-320+)GMT-D3 strains
acutes: 4
(<20-320+)
GMT-D3 strains
convalescent: 2
(<20-169)
GMT-D3 all
strains:2.6 (<20-
320+)
GMT-D4 strains
acutes: 15 (<20-
320+)
GMT-D4 strains
convalescent: 2
(<20-51)
GMT-D4 all
strains:3.4(<20-
320+)
GMT-flavi
acutes: 3 (<20-
113)
GMT-flavi
convalescent: 1
(<20-35)
GMT-flavi:1.6 (<20-320+)
Heterologous
serotype
Geometric Mean Titer (GMT)
GMT-D1 strains
acutes: 88 (21-
320+)
GMT-D1 strains
convalescent: 79
(35-320+)
Virus<1 year 1- 2 year
Study No/MPI
2- 6 years
Homologous
serotype
GMT

155
Table 5. 4:- DENV-1-Sera panel statistical verification
Variable Effect (difference in
log(FRNT50)) Std. Err. Tobit p-value
(95%
Conf.interval)
Change per month -0.006 0.035 -0.18 0.868 -0.095 0.083
Homologous serotype 2.786 1.063 2.62 0.047 0.053 5.519
D1-HW2001G4 0.000
D1-10BG1L14 1.888 0.638 2.96 0.032 0.248 3.529
D1-15LG1 -0.027 0.750 -0.04 0.973 -1.954 1.901
D1-10BG2L4 0.063 1.185 0.05 0.960 -2.984 3.109
D1-10INDG3L4 -0.603 0.459 -1.31 0.246 -1.783 0.576
D1-14PHLG4 1.203 0.350 3.44 0.018 0.303 2.102
D2-NGCGAII 0.000
D2-10LGA1L9 0.045 0.682 0.07 0.949 -1.707 1.797
D2-11BGCL4 -3.270 0.600 -5.45 0.003 -4.813 -1.726
D2-13MGCLN -2.718 0.466 -5.83 0.002 -3.916 -1.519
D2-15BGCL4 -2.292 0.789 -2.91 0.034 -4.319 -0.265
D2-15BGCLN -2.615 0.520 -5.03 0.004 -3.952 -1.279
D2-PR6913-V -2.716 0.516 -5.26 0.003 -4.043 -1.388
D2-PR1940-NV -0.268 0.424 -0.63 0.556 -1.359 0.823
D3-H87G5 0.000
D3-15BG1 2.951 0.979 3.01 0.030 0.435 5.467
D3-UNC3002-V -0.831 0.583 -1.42 0.213 -2.329 0.668
D3-UNC3008-NV 1.362 0.436 3.12 0.026 0.242 2.482
D4-H241G1 0.000
D4-10BG3L1 -2.532 0.281 -9.02 <0.001 -3.253 -1.810
D4-10BG3L2 -1.339 0.427 -3.13 0.026 -2.438 -0.241
Note: Virus values are average differences from reference strain in each individual panel
Variable: Time post infection and strains of DENV tested
Effect:- Difference in log FRNT50 with censoring cut-offs set to 2.99-5.77
p-value:- Significance of effect of neutralisation compared with reference strains in each serotype
95%CI:- robust variance estimation of CI for regression coefficients.

156
i. Anti-DENV-1 antisera differentially neutralize DENV-1
Variation in neutralisation of homologous DENV-1 was observed. DENV-1 strains
were isolated from travellers infected in 2010-2015. Each strain is coded with serotype,
year of isolation, location of isolation genotype and lineage, as described in (11) as
shown in (Table 4.6). The reference virus D1-HW2001G4 isolated in 2001 DENV-1
Hawaii outbreak was included. Neutralisation against all the homologous strains was
significantly greater (p=0.047) than against heterologous viruses.
When geometric mean titres (GMT) were compared, NAb response against strain D1-
10BG1L14 was 32% greater than NAb against reference strain D1-HW2001G4, as well
as 41% greater than another isolate D1-15LG1 belonging to the same genotype and
circulating in 2015. Genotype 1 strains were more strongly neutralized than Genotype 2
or Genotype 3. Using tobit regression analysis, the difference in neutralization
(FRNT50) for 2010 Genotype 1 was statistically significant compared to reference
strain if we take p<0.05 as evidence to reject null hypothesis (there is no difference)
while difference in neutralisation for D1-15LG1, a virus belonging to the same
genotype and originating in Laos in 2015 was not statistically significant (p>0.05).
Statistical values for each dataset are given in Table 5.4. Thus, within DENV-1
serogroup the order of NAb responses against different strains of DENV-1, from highest
to lowest was D1-10BG1L14>D1-14PHLG4>D1-10BG2L4>D1-2001HWG4>D1-
15LG1>D1-10INDG3L4. Figure 5. 2 show comparative NAb response against
homologous strains.
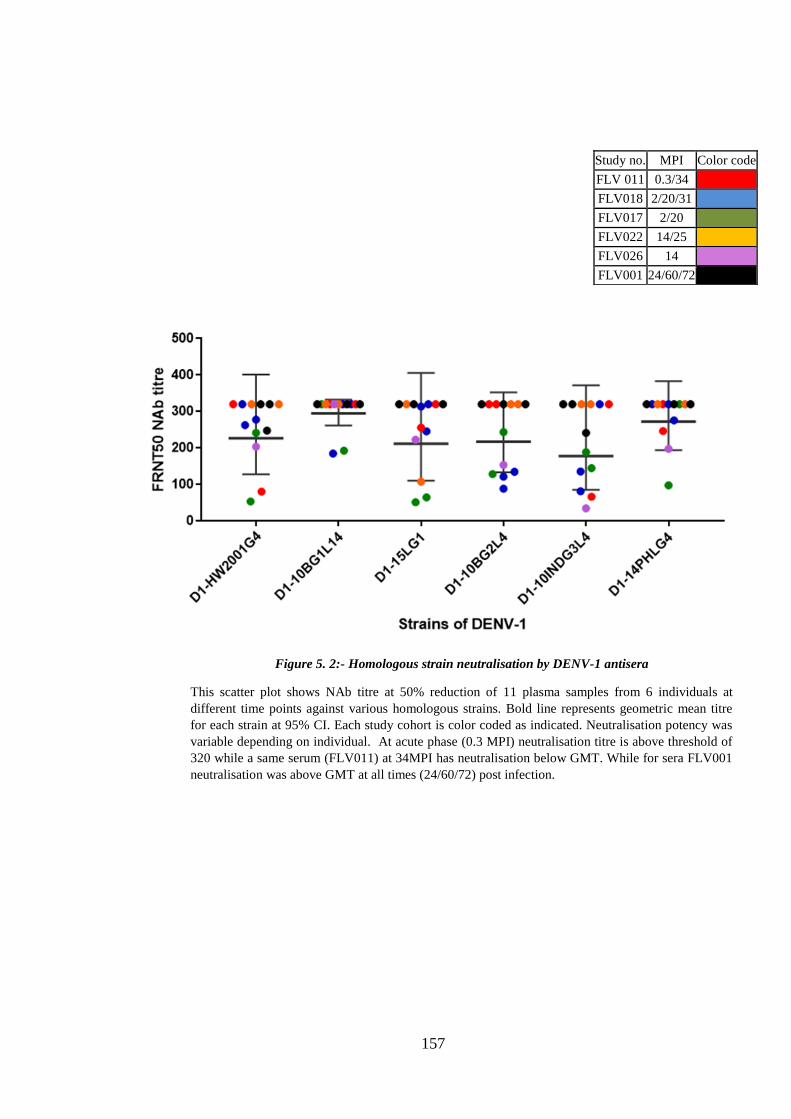
157
Study no. MPI Color code
FLV 011 0.3/34
FLV018 2/20/31
FLV017 2/20
FLV022 14/25
FLV026 14
FLV001 24/60/72
Figure 5. 2:- Homologous strain neutralisation by DENV-1 antisera
This scatter plot shows NAb titre at 50% reduction of 11 plasma samples from 6 individuals at
different time points against various homologous strains. Bold line represents geometric mean titre
for each strain at 95% CI. Each study cohort is color coded as indicated. Neutralisation potency was
variable depending on individual. At acute phase (0.3 MPI) neutralisation titre is above threshold of
320 while a same serum (FLV011) at 34MPI has neutralisation below GMT. While for sera FLV001
neutralisation was above GMT at all times (24/60/72) post infection.

158
ii. Neutralization among homologous DENV-1 decreases over time
Anti-DENV-1 acute phase sera neutralized all DENV-1 strains. Each patient for whom
there were follow-up samples demonstrated a decline in magnitude of responses,
measured as GMT, with time after infection, for example FLV011/9 days: 320,
FLV011/34 months: 181; FLV017/2 months:184, FLV/017/20 months:111. Variation
among antisera was apparent – for example patient FLV001 antisera 24 months post
infection neutralized all DENV-1 at high magnitude (>320 for all strains) whereas
patient FLV017 antisera 20 months GMT was significantly lower (111) (Table 5.1).
This pattern of differential homologous NAb responses was patient-specific: FLV017
also showed lower magnitude responses at 2 months post infection with GMT 184
whereas patient FLV018 antisera, also sampled 2 months after infection, demonstrated
GMT 277.
DENV-1 strain-specific neutralization was observed. FRNT50 titres were greatest for
strain DI-10BG1L14, with titres of 320 for 11 of 13 antisera and highest GMT; lowest
magnitude responses were seen for strain D1-10INDG3L4 (Figures 5.2 and 5.4).
Homologous neutralization persisted for all patents’ antisera however decline in
neutralization capacity that was strain-specific was apparent: Figure 5.3 shows relative
decline in neutralization against 2 strains, compared to the other 4. Despite this pattern
of patient (antisera) and strain- specific differential neutralization, overall variation
within homologous DENV-1 was not significant (p>0.05) (Table 5.4).

159
9 days after infection
2 years 10 months after infection
Figure 5. 3:- Differential neutralisation among DENV-1 homologous strains,
patient FLV-011
Acute phase sera neutralize all DENV-1 strains at high magnitude. At 2 years 10
months after infection strain-specific decline in neutralization capacity was
apparent.
1 .0 1 .5 2 .0 2 .5 3 .0
0
1 0 0
1 5 0
L o g 1 0 P la s m a D ilu tio n
Re
du
cti
on
%
50
75
90
1 .0 1 .5 2 .0 2 .5 3 .0
0
1 0 0
1 5 0
L o g 1 0 P la s m a D ilu tio n
Re
du
cti
on
%
50
75
90
D 1 -H W 2 0 0 1 G 4
D 1 -1 0 B G 1 L 1 4
D 1 -1 5 L G 1
D 1 -1 0 B G 2 L 4
D 1 -1 0 IN D G 3 L 4
D 1 -1 4 P H L G 4
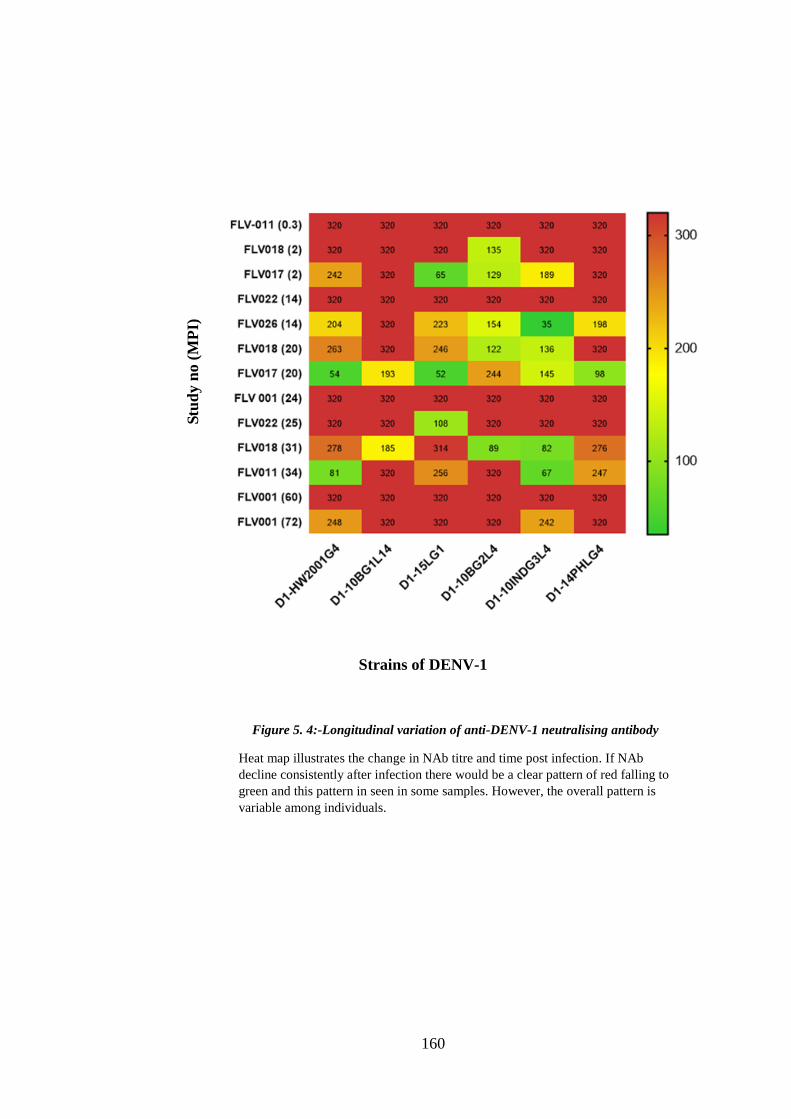
160
Figure 5. 4:-Longitudinal variation of anti-DENV-1 neutralising antibody
Heat map illustrates the change in NAb titre and time post infection. If NAb
decline consistently after infection there would be a clear pattern of red falling to
green and this pattern in seen in some samples. However, the overall pattern is
variable among individuals.
Strains of DENV-1
Stu
dy n
o (
MP
I)

161
iii. Cross-neutralisation against heterologous DENV
Cross-neutralisation against all heterologous DENV strains was seen within 2 months of
infection. Convalescent antisera more than one year after infection showed significantly
less cross-neutralization, with some exceptions that were patient- and virus-specific.
Neutralisation capacity of anti-DENV-1 antisera against 15 heterologous DENV: 8
DENV-2, 4 DENV-3 and 3 DENV-4 strains were tested (Table 5.1). Each strain
representing patient isolates are coded with serotype-date of isolation-genotype-lineage,
reference strains include DENV-2 NGC (D2-NGCAII), DENV-3 (D3-H87G5) and
DENV-4 (D4-H241G1) was used. In addition, viruses with distinct epidemiological
traits – ‘virulent’ and ‘non-virulent’ DENV-2 (PR-1940-NV and PR-6913-V) and
DENV-3 (UNC-3002-V and UNC-3008-NV) were included as heterologous strains.
DENV-4 was the most highly neutralised serotype (GMT 97) compared to DENV-2
(54) and DENV-3 (67). Differential strain-specific neutralisation was observed. Among
DENV-2 strains, strain D2-10LGA1L6, originating in Laos 2010 and belonging to the
Asian 1 Genotype, was most strongly neutralised compared to others. Similarly, within
DENV-2 Cosmopolitan genotype, strain D2-15BGCL4 originating in Bali Indonesia in
2015 is neutralized at higher magnitude than strains originating in Malaysia in 2013 and
Bali in 2015 (Figure 5.5). Non-virulent strain PR1940 was highly neutralised compared
to virulent strain PR6913, suggesting that strains with higher epidemic potential are less
susceptible to neutralisation with heterologous DENV antisera. A similar pattern was
observed for DENV-3 virulent and non-virulent strains, non-virulent UNC3002 highly
neutralised compared to virulent UNC3008. The DENV-3 strain D3-15BG1, originating
in Bali in 2015 and a representative of all DENV-3 isolated from travellers, was
strongly cross-neutralized by anti-DENV-1 antisera. DENV-4 reference strain D4-

162
H241G1 was neutralized at greatest magnitude highest neutralisation, titre compare to
DENV-4 patient isolates. Figure 5. 5 shows differential strain -specific neutralisation
by anti-DENV-1 antibody.
Variable neutralisation among heterologous strains was compared using regression
analysis. Each strain was compared with the respective reference strain.
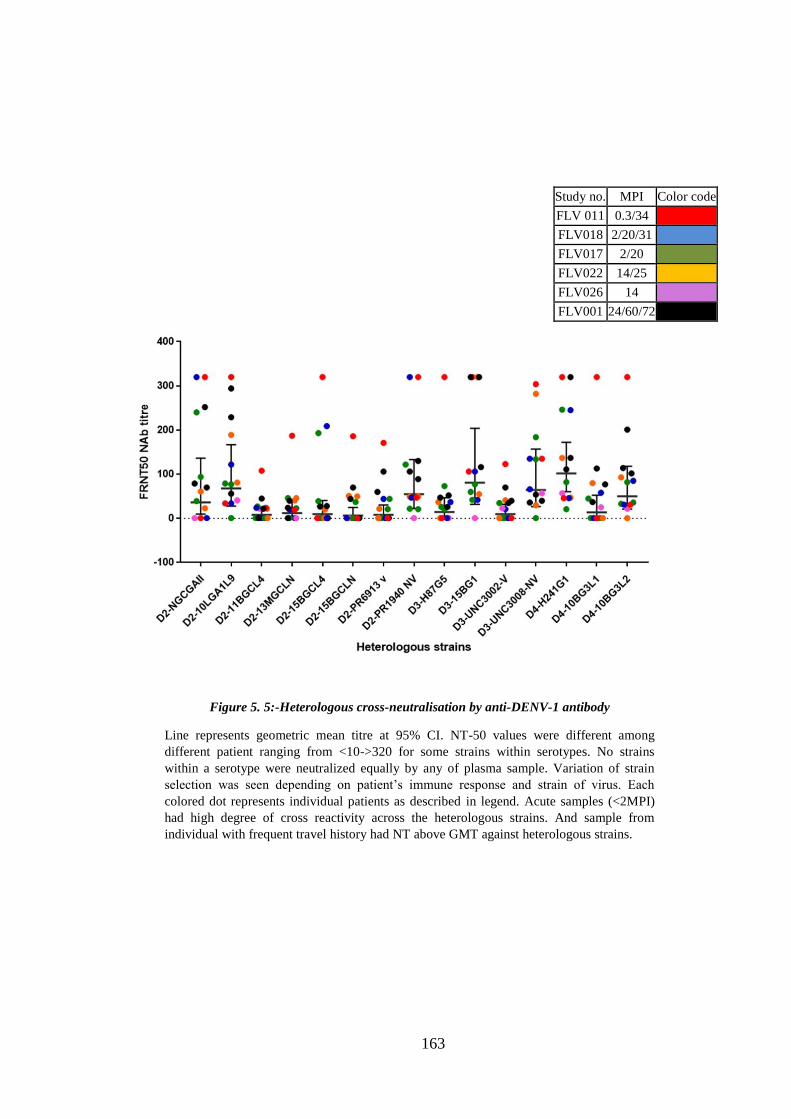
163
Study no. MPI Color code
FLV 011 0.3/34
FLV018 2/20/31
FLV017 2/20
FLV022 14/25
FLV026 14
FLV001 24/60/72
Figure 5. 5:-Heterologous cross-neutralisation by anti-DENV-1 antibody
Line represents geometric mean titre at 95% CI. NT-50 values were different among
different patient ranging from <10->320 for some strains within serotypes. No strains
within a serotype were neutralized equally by any of plasma sample. Variation of strain
selection was seen depending on patient’s immune response and strain of virus. Each
colored dot represents individual patients as described in legend. Acute samples (<2MPI)
had high degree of cross reactivity across the heterologous strains. And sample from
individual with frequent travel history had NT above GMT against heterologous strains.

164
iv. Longitudinal decrease in heterologous DENV cross-neutralization by anti-
DENV-1 antisera
An acute phase sample collected 9 days after infection (patient FLV011) cross-
neutralized all heterologous DENV at high magnitude. Convalescent antisera from the
same patient 34 and 72 months after neutralized homologous DENV at higher
magnitude than heterologous viruses (Table 5.1). A similar pattern was observed for
patients FLV018 and FLV017, for acute phase sera (2 months after infection) and
convalescent sera (31 and 20 months, respectively). However, patient FLV001 showed
an increase in heterologous cross-neutralization with development of anti-DENV-2,
DENV-3 and DENV-4 cross-neutralization at 60 months that was not present at 20
months. This individual travelled in between his 20 and 60-month visits, but not to any
countries where dengue is endemic (although it is possible that he was exposed to
DENV in Northern Territory, which neighbours dengue endemic countries and where
DENV is frequently introduced by travellers).
Serotype cross-neutralization was strain-dependent. Cross-neutralization of strain D3-
15BG1 was maintained at significant levels for 5 of 6 patients, whereas responses
against D4-10BG3L1 had declined to <20 in 3 of 4 patients with antisera available 2-6
years after infection.
Change in cross-neutralisation with the progression of time post infection is illustrated
in heat map (Figure 5. 6).

165
Figure 5. 6:- Longitudinal heterologous cross-neutralisation by anti-DENV-1 antibody
Decrease in cross-neutralization with time was observed across heterologous DENV. With some exceptions
cross-neutralization largely declines 14 months after infection.
Stu
dy
no
(M
PI)

166
v. Persistent neutralisation against autologous virus
Autologous virus isolated from acute phase sera for 3 patients from whom serum
samples were available up to 60 months (5 years) post infection were neutralised at high
magnitude. Homologous virus was also neutralized at high magnitude however in 5 of 8
cases the magnitude of the response was less than for autologous strains, within the
limits of the neutralization test endpoint dilution. Heterologous serotype cross-
neutralization was always lower than for autologous and homologous virus. In 2 of 3
patients a decline in homologous neutralization, compared to autologous neutralization,
was apparent at 25 months post infection; in the third patient this differential decline
was not apparent until 72 months. Data are shown in Figure 5.7.
All 3 autologous isolates originated in Bali, Indonesia, in 2010, 2014 and 2015 (Table
4.6).
Mo
nth
s
pos
t
inf
ecti
on
Heterologous DENV
strains

167
Figure 5. 7:- Autologous, homologous and heterologous neutralisation
Preferential neutralisation against autologous virus was observed.
FLV001 antisera were collected 24, 60 and 72 months post infection;
FLV081 2, 20 and 31 MPI, and FLV022 14 and 25 MPI.

168
vi. Cross-neutralization of ZIKV, YF17D and IMOJEV by DENV-1 antisera
No patients were known or suspected to be previously infected with ZIKV. However,
the ZIKV prototype strain MR766 was neutralized by 10 of 11 anti-DENV-1 antisera.
One of 6 patients (FLV017) showed a decline in anti-MR766 neutralization by 20
months, to undetectable levels. Despite this strong response, no antisera cross-
neutralized the 2015 epidemic strain ZIKV PRVABC59.
Cross-neutralization of the vaccine viruses YF17D and IMOJEV was mostly observed
for acute phase sera. Anti-YF17D responses were maintained in one patient, FLV018, at
20 and 31 months after infection; this patient had been vaccinated in 1978.
Change in NAb responses with time post infection is summarized in Table 5.1

169
Figure 5. 8:- Cross-neutralisation across flavivirus serogroup
Cross neutralizing responses against ZIKV MR766 but not ZIKV
PRVABC59 were observed. Cross neutralization of YF17D was a feature of
acute phase antisera.
Stu
dy
no
(M
PI)

170
b. Specificity and cross-reactivity of anti-DENV-2 antibody
DENV-2 antisera showed differential neutralisation among 8 homologous and 14
heterologous strains over time. Responses were greatest for homologous DENV-2
strains and were consistently high for acute phase antisera collected 5 days after
infection. Heterologous cross-neutralization declined with time, however persisting
responses were observed up to 64 months that were patient- and virus-specific. An
example of a DENV-2-infected 7 months and 3 years after infection is shown in Figure
5. 9 . One of 6 heterologous DENV-1 strains; 1 of 4 DENV-3 and 2 of 3 DENV-4 were
neutralized 7 months after infection. By 3 years only 1 DENV-4 strain was neutralized
at low magnitude, however 1 DENV-2 strain, D2-PR6193V, was also no longer
neutralized. D2-PR6193V is a reference strain that represents epidemic virulence, and it
is noteworthy that this virus was not neutralized whereas the non-virulent reference
strain D2-PR1940NV was neutralized. List of samples included in this sera-panel is
given in Table 4.2 (also shown below).
Non-linear regression curve was used to determine neutralisation potency at different
reduction cut-off. Neutralisation against homologous and heterologous strain (as shown
in Figure 5. 9) for all anti-DENV-2 plasma samples and non-linear regression curve at
different time points post infection are given in appendix 5.2. Antibody titre at different
cut-off is given in Table 5. 5- Table 5. 7. Statistical variables derived using tobit
regression using FRNT-50 as functional measurement and robust variance estimation is
listed in Table 5.8 and described in each section below.

171
Sample ID Gender Origin of infection Flavi
Vaccination
Diagnostic tests
Months post onset of
illness Location Year of visit Duration
of Stay
(Days) Flavi HI NS1 result Dengue
IgM
DENV
PCR^
FLV-004 M Bali 2011/2012 14 No <10 + - + 16 51 64
FLV-005 F Bali 2011 10 No <10 + + + 16 51
FLV-009 F Bali 2012 10 No 40 + + + 17 36
FLV-010 M Thailand 2012 10 No <10 + - + 19 36 48
FLV-012 F Java, Bali 2013 N/A No <10 + - + 0.1 33
FLV-014 F Bali 2013 N/A YF 2006 + - + 7 36
FLV-070* M Bali 2016 5 YF <10 + + + 7
FLV-072* M Bali 2015 10 No <10 + - + 15
FLV-073* M Thailand 2015 10 No <10 + - + 15
FLV-074* M Philippines 2015 10 No <10 + + + 13
Table 4. 2:- Demographic and laboratory findings of individuals with DENV -2 infection
“*”:- Samples included only in HI test
N/A: - information not available
“^”: Serotyping PCR specific for DENV-2
“+”: - Positive
“- “: - Negative

172
Figure 5. 9:- Neutralising antibody response by anti-DENV-2 antisera, patient FLV -014
DENV-2 antisera acute and convalescent antisera differentially neutralize homologous and
heterologous DENV.

173
*MPI: Months Past Infection
*Acute: <=2 months past infection
*Convalescent: > 2 months past infection
Note:
Value 1=<20
Value 320=>320
Table 5. 5:- Anti-DENV-2 neutralising antibody titre at 50% reduction
>5 years
FLV012
/ 5 days
FLV014
/ 7
FLV004
/ 16
FLV005
/ 16
FLV009
/ 17
FLV010
/ 19
FLV012
/ 33
FLV009
/36
FLV010
/36
FLV014
/36
FLV010
/48
FLV004
/51
FLV005
/51
FLV004/
64
D2-NGCGAII 320 232 320 320 320 320 221 40 46 138 61 176 289 51 159
D2-10LGA1L9 320 192 320 218 128 129 148 139 250 225 225 272 201 155 199
D2-11BGCL4 320 132 217 113 230 64 123 154 66 320 68 186 70 251 142
D2-13MGCLN 320 320 320 243 320 320 261 159 149 320 96 170 84 128 208
D2-15BGCL4 320 320 320 320 320 235 320 73 34 247 31 81 48 83 144
D2-15BGCLN 320 302 320 88 68 47 265 71 48 320 46 111 97 98 120
D2-PR6913 V 121 320 131 241 50 64 241 29 29 53 29 49 59 64 76
D2-PR1940 NV 320 320 224 155 163 320 320 121 74 96 88 75 96 79 149
GMT-D2 283 256 261 194 164 145 226 84 67 183 65 122 99 101 144
D1-HW2001G4 258 1 20 1 65 50 1 1 1 1 1 1 1 1 3
D1-10BG1L14 232 1 127 233 52 1 103 48 1 1 1 51 63 1 13
D1-15LG1 129 1 32 1 1 1 1 1 1 1 1 32 1 23 3
D1-10BG2L4 320 102 320 262 157 45 66 78 53 24 38 73 93 1 66
D1-10INDG3L4 320 320 66 118 43 35 1 1 1 23 1 1 1 1 9
D1-14PHLG4 210 1 1 1 1 1 1 1 1 1 1 1 1 1 1
GMT-D1 234 6 35 14 17 7 4 4 2 3 2 7 4 2 7
D3-H87G5 320 320 58 21 119 1 1 1 1 1 1 44 1 27 9
D3-15BG1 320 34 73 27 30 1 42 30 1 1 23 1 1 1 9
D3-UNC3002-V 123 34 129 76 45 1 1 1 1 1 1 1 1 24 6
D3-UNC3008-NV 320 69 320 50 46 43 30 1 1 1 30 162 29 175 30
GMT-D3 252 71 115 38 52 3 6 2 1 1 5 9 2 18 11
D4-H241G1 320 168 60 74 170 184 31 104 1 79 1 56 29 55 46
D4-10BG3L1 149 39 30 51 66 33 58 1 1 41 1 1 21 1 12
D4-10BG3L2 320 100 159 74 87 69 47 31 23 27 1 36 38 21 42
GMT-D4 248 87 66 66 99 75 44 15 3 45 1 13 29 10 28
ZIKV MR766 320 111 59 29 30 34 1 1 1 1 1 1 1 1 6
ZIKV PRVABC59 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1
YF 17D 320 245 1 1 320 1 1 100 1 191 1 1 1 1 7
IMOJEV 47 1 1 1 1 1 1 1 1 1 1 1 1 1 1
GMT 47 13 3 2 10 2 1 3 1 4 1 1 1 1 3
GMT-D4
s tra ins for
convalescent
sera : 24 (<20-
184)
GMT-D1
s tra ins for
convalescent
sera : 5 (<20-
320)
GMT-D1
stra ins for
acute sera :
234(129-320+)
GMT-D3
stra ins for
acute sera :
252(123-320+)
GMT-D3
stra ins for
convalescent
sera :8 (<20-
320)
GMT-D4
stra ins for
acute sera :
248(149-320+)
GMT-flavi
convalescent:
2 (<20-191)
Virus Geometric Mean Titre (GMT)
GMT- total sera homologous
serotype=144 (29-320+)
GMT-D1
strains
total : 7
(<20-320+)
Homologous
serotype
GMT-D2
stra ins for
acute sera:
283(121-320+)
GMT-D2
convalescent
sera: 136 (29-
320+)
Heterologous
serotype GMT-D3
strains
total : 11
(<20-320+)
GMT-D4
strains
total :28
(<20-320+)
GMT- total sera
heterologous
serotype= 10 (<20-
320+)
Other
Flaviviruses
GMT-flavi
acutes: 47
(<20-320+)
GMT-flavi:3 (<20-320+)
< 1 year 1-2 year 2-3 years
GMT
Study No./ MPI
4-5 years

174
Note:
Value 1=<20
Value 320=>320
*MPI: Months Past Infection
*Acute: <=2 months past infection
*Convalescent: > 2 months past infection
Table 5. 6:- Anti-DENV-2 neutralising antibody titre at 75% reduction
>5 years
FLV012 /
5 days
FLV014 /
7
FLV004 /
16
FLV005 /
16
FLV009 /
17
FLV010 /
19
FLV012 /
33
FLV009
/36
FLV010
/36
FLV014
/36
FLV010
/48
FLV004
/51
FLV005
/51
FLV004/6
4
D2-NGCGAII 320 139 87 320 193 320 112 27 27 72 34 75 121 32 94
D2-10LGA1L9 320 66 97 119 48 59 75 51 105 100 125 91 96 52 88
D2-11BGCL4 320 44 94 54 126 24 40 82 34 207 37 94 39 96 70
D2-13MGCLN 320 320 193 122 238 320 57 72 50 320 46 90 47 26 114
D2-15BGCL4 320 320 265 140 140 90 81 39 23 128 1 44 30 40 65
D2-15BGCLN 79 155 126 41 36 28 85 40 29 174 28 59 51 52 58
D2-PR6913 V 47 132 43 79 27 32 57 20 1 32 1 28 29 29 23
D2-PR1940 NV 320 124 90 62 78 150 33 34 1 46 38 35 49 32 47
GMT-D2 211 134 109 95 86 80 63 41 15 106 17 59 51 41 64
D1-HW2001G4 100 1 1 1 1 24 1 1 1 1 1 1 1 1 2
D1-10BG1L14 79 1 33 90 26 1 1 1 1 1 1 1 1 1 3
D1-15LG1 29 1 1 1 1 1 1 1 1 1 1 1 1 1 1
D1-10BG2L4 320 1 60 69 68 25 1 31 23 1 1 21 42 1 12
D1-10INDG3L4 320 174 36 46 27 23 1 1 1 1 1 1 1 1 6
D1-14PHLG4 50 1 1 1 1 1 1 1 1 1 1 1 1 1 1
GMT-D1 103 2 6 8 6 5 1 2 2 1 1 2 2 1 3
D3-H87G5 82 79 1 1 1 1 1 1 1 1 1 1 1 1 2
D3-15BG1 100 1 29 1 1 1 1 1 1 1 1 1 1 1 2
D3-UNC3002-V 1 1 37 21 1 1 1 1 1 1 1 1 1 1 2
D3-UNC3008-NV 65 1 94 22 23 1 20 1 1 1 1 1 1 27 5
GMT-D3 27 3 18 5 2 1 2 1 1 1 1 1 1 2 2
D4-H241G1 320 57 24 32 62 50 21 39 1 39 1 32 20 26 24
D4-10BG3L1 23 1 1 1 32 1 1 1 1 1 1 1 1 1 2
D4-10BG3L2 320 32 47 1 37 22 28 21 1 1 1 22 1 1 8
GMT-D4 132 12 11 3 42 10 8 9 1 3 1 9 3 3 7
ZIKV MR766 288 1 1 1 1 1 1 1 1 1 1 1 1 1 1
ZIKV PRVABC59 1 1 1 1 1 1 1 1 1 1 1 1 1 1
YF 17D 174 117 28 21 187 1 39 49 1 87 1 1 1 1 11
IMOJEV 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1
GMT 15 3 2 2 6 1 2 3 1 3 1 1 1 1 2
Other
Flaviviruses
< 1 year 1-2 year 2-3 years
Homologous
serotype
Geometric Mean Titre (GMT)
GMT-D2
strains for
acute sera:
211(47-320+)
GMT-D2
convalescent
sera: 58 (<20-
320+)
GMT- total sera homologous
serotype=64 (<20-320+)
Heterologous
serotype
GMT
GMT-flavi
acutes: 15
(<20-288)
GMT-flavi
convalescent: 2
(<20-117)
GMT-flavi:2 (<20-288)
Virus
GMT-D1
strains for
acute sera :
103(29-320+)
GMT-D1
strains for
convalescent
sera : 2 (<20-
174)
GMT-D1
strains
total : 3
(<20-320+)
GMT- total sera
heterologous
serotype= 3(<20-
320+)
GMT-D3
strains for
acute sera :
27(<20-100)
GMT-D3 strains
for
convalescent
sera :2 (<20-
94)
GMT-D3
strains
total : 2
(<20-100)
GMT-D4
strains for
acute sera :
132(23-320+)
GMT-D4
strains for
convalescent
sera : 5 (<20-
62)
GMT-D4
strains
total
:7(<20-
320+)
4-5 years
Study No./ MPI

175
*MPI: Months Past Infection
*Acute: <=2 months past infection
*Convalescent: > 2 months past infection
Note:
Value 1=<20
Value 320=>320
Table 5. 7:- Anti-DENV-2 neutralising antibody titre at 90% reduction
>5 years
FLV012 / 5
days FLV014 / 7
FLV004 /
16
FLV005 /
16
FLV009 /
17
FLV010 /
19
FLV012 /
33
FLV009
/36
FLV010
/36
FLV014
/36
FLV010
/48
FLV004
/51
FLV005
/51
FLV004/
64
D2-NGCGAII 268 83 38 124 85 223 49 22 1 42 24 39 59 24 44
D2-10LGA1L9 320 34 47 62 27 35 39 31 48 46 64 47 52 28 48
D2-11BGCL4 320 1 44 29 64 1 23 48 23 93 25 51 27 43 27
D2-13MGCLN 147 320 109 53 62 320 29 42 21 144 28 51 32 1 51
D2-15BGCL4 115 320 120 64 62 44 27 27 1 67 1 29 23 26 31
D2-15BGCLN 33 79 42 26 27 21 37 29 22 72 22 37 33 31 33
D2-PR6913 V 21 52 27 41 1 1 1 1 1 24 1 21 21 16 6
D2-PR1940 NV 242 48 47 37 37 51 21 1 1 30 22 24 31 18 22
GMT-D2 129 52 52 48 29 24 20 13 5 56 12 35 33 17 28
D1-HW2001G4 46 1 1 1 1 15 1 1 1 1 1 1 1 1 2
D1-10BG1L14 42 1 1 43 1 1 1 1 1 1 1 1 1 1 2
D1-15LG1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1
D1-10BG2L4 84 1 33 31 21 1 1 20 1 1 1 1 25 1 4
D1-10INDG3L4 320 84 25 23 21 1 1 1 1 1 1 1 1 1 4
D1-14PHLG4 25 1 1 1 1 1 1 1 1 1 1 1 1 1 1
GMT-D1 33 2 3 6 3 2 1 2 1 1 1 1 2 1 2
D3-H87G5 23 36 1 1 1 1 1 1 1 1 1 1 1 1 2
D3-15BG1 29 1 1 1 1 1 1 1 1 1 1 1 1 1 1
D3-UNC3002-V 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1
D3-UNC3008-NV 27 1 1 1 1 1 1 1 1 1 1 1 1 1
GMT-D3 12 2 1 1 1 1 1 1 1 1 1 1 1 1 1
D4-H241G1 293 30 1 1 32 27 1 20 1 25 1 23 1 1 6
D4-10BG3L1 320 1 1 1 18 1 1 1 1 1 1 1 1 2
D4-10BG3L2 56 20 1 1 21 1 20 1 1 1 1 1 1 3
GMT-D4 174 9 1 1 23 3 3 20 1 3 1 3 1 1 4
ZIKV MR766 95 1 1 1 1 1 1 1 1 1 1 1 1 1 1
ZIKV PRVABC59 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1
YF 17D 79 58 1 1 82 1 20 28 1 38 1 1 1 1 5
IMOJEV 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1
GMT 9 3 1 1 3 1 2 2 1 2 1 1 1 1 2
GMT-flavi
acutes: 9 (<20-
95)
GMT-flavi
convalescent: 1
(<20-82)
GMT-flavi:2 (<20-95)
Study No./ MPI
Geometric Mean Titre (GMT/NT range)
GMT-D2
strains for
acute sera:
129 (21-
320+)
GMT-D2
convalescent
sera: 25 (<20-
320+)
GMT- total sera
homologous serotype=28
(<20-320+)
GMT-D1
strains for
acute sera :
33(<20-320+)
GMT-D1
strains for
convalescent
sera : 2 (<20-
320)
GMT-D1
strains
total : 2
(<20-320+)
GMT- total
sera
heterologous
serotype= 2
(<20-320+)
GMT-D3
strains for
acute sera :
12(<20-29)
GMT-D3 strains
for
convalescent
sera :1 (<20-
36)
GMT-D3
strains
total : 1
(<20-36)
GMT-D4
strains for
acute sera :
174(56-320+)
GMT-D4
strains for
convalescent
sera : 3 (<20-
30)
GMT-D4
strains
total :4
(<20-293)
Heterologous
serotype
Other
Flaviviruses
1-2 year 2-3 years
GMT
< 1 yearVirus
4-5 years
Homologous
serotype
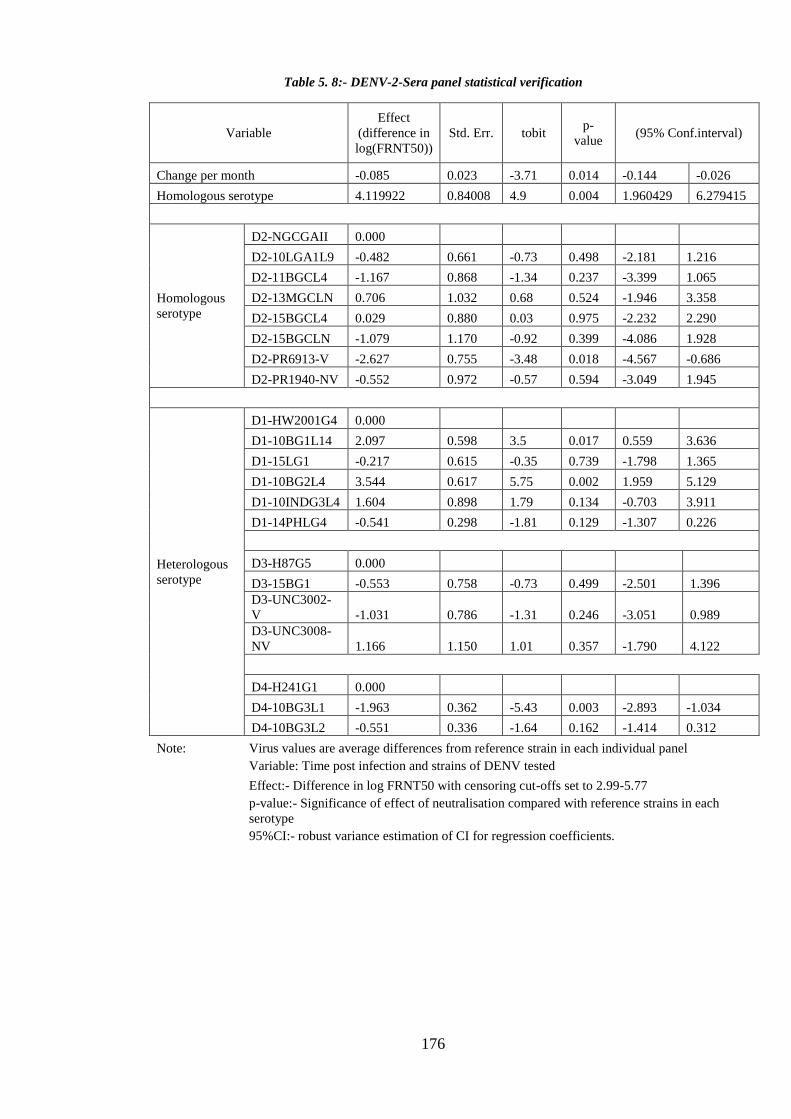
176
Table 5. 8:- DENV-2-Sera panel statistical verification
Variable
Effect
(difference in
log(FRNT50))
Std. Err. tobit p-
value (95% Conf.interval)
Change per month -0.085 0.023 -3.71 0.014 -0.144 -0.026
Homologous serotype 4.119922 0.84008 4.9 0.004 1.960429 6.279415
Homologous
serotype
D2-NGCGAII 0.000
D2-10LGA1L9 -0.482 0.661 -0.73 0.498 -2.181 1.216
D2-11BGCL4 -1.167 0.868 -1.34 0.237 -3.399 1.065
D2-13MGCLN 0.706 1.032 0.68 0.524 -1.946 3.358
D2-15BGCL4 0.029 0.880 0.03 0.975 -2.232 2.290
D2-15BGCLN -1.079 1.170 -0.92 0.399 -4.086 1.928
D2-PR6913-V -2.627 0.755 -3.48 0.018 -4.567 -0.686
D2-PR1940-NV -0.552 0.972 -0.57 0.594 -3.049 1.945
Heterologous
serotype
D1-HW2001G4 0.000
D1-10BG1L14 2.097 0.598 3.5 0.017 0.559 3.636
D1-15LG1 -0.217 0.615 -0.35 0.739 -1.798 1.365
D1-10BG2L4 3.544 0.617 5.75 0.002 1.959 5.129
D1-10INDG3L4 1.604 0.898 1.79 0.134 -0.703 3.911
D1-14PHLG4 -0.541 0.298 -1.81 0.129 -1.307 0.226
D3-H87G5 0.000
D3-15BG1 -0.553 0.758 -0.73 0.499 -2.501 1.396
D3-UNC3002-
V -1.031 0.786 -1.31 0.246 -3.051 0.989
D3-UNC3008-
NV 1.166 1.150 1.01 0.357 -1.790 4.122
D4-H241G1 0.000
D4-10BG3L1 -1.963 0.362 -5.43 0.003 -2.893 -1.034
D4-10BG3L2 -0.551 0.336 -1.64 0.162 -1.414 0.312
Note: Virus values are average differences from reference strain in each individual panel
Variable: Time post infection and strains of DENV tested
Effect:- Difference in log FRNT50 with censoring cut-offs set to 2.99-5.77
p-value:- Significance of effect of neutralisation compared with reference strains in each
serotype
95%CI:- robust variance estimation of CI for regression coefficients.

177
i. Anti-DENV-2 neutralisation specificity against homologous serotype
Anti-DENV-2 antisera neutralized all DENV-2 strains. Acute phase antisera collected 5
days after infection neutralized all homologous DENV at high magnitude as did antisera
collected 7 months after infection. Convalescent antisera (16 – 64 months post
infection) neutralized homologous DENV-2 at lower magnitude GMT 136) than acute
phase sera (GMT 283).
Most travellers returning to Western Australia with DENV-2 infection have been
infected with the same lineage (DENV-2 Cosmopolitan Genotype, Lineage 4). This
novel lineage emerged in 2010-2012 as the dominant epidemic virus in Bali, Indonesia
and has continued to be imported up to 2017. One other virus was imported, belonging
to the Asian 1 Lineage and originating in Laos (D2-10LGA1L9). Neutralization was
assessed against traveller isolates, mostly Lineage 4 or the closely related Nepal lineage
(designated N: D2-13MGCLN; D2-15BGCLN); D2-10LGA1L9; the epidemic strains
D2-PR6193V and D2-PR6193V, and the reference strain D2-NGCAII.
Field isolates imported by travellers therefore represent 2 DENV-2 genotypes, Asian I
and Cosmopolitan. Each field isolate is coded with serotype, year of isolation, genotype
and lineage.
Neutralisation against homologous DENV-1 was approximately 10-fold greater than
that against heterologous serotypes. Difference in homologous vs heterologous
neutralisation was statistically significant (p=0.004).
Within-serotype neutralization varied depending on the virus strain. Asian 1 Genotype
strain D2-10LGA1L9 was neutralized at high magnitude, as was strain D2-13MGCLN;
interestingly the other 3 closely related Lineage 4/Nepal viruses were neutralized to a

178
lesser degree. (Figure 5. 10). Among the patient isolates GMT for Asian 1 D2-
10LGA1L9 was 25% higher than GMT of Cosmopolitan isolates, as a group. Within the
4 Cosmopolitan viruses there was ~2-fold difference in GMT. D2-13MGCLN
originating in Malaysia in 2013 was neutralized with highest GMT compared to other
DENV-2. Neutralisation against the epidemic virulent PR6913-V strain was
significantly lower (p=0.018) compared to other DENV-2 strains. Reference strain
DENV-2NGC was neutralised efficiently by all the test samples.
Thus, the same pattern of patient- and virus-specific neutralization was seen for DENV-
2 as was noted for DENV-1.

179
Study no. MPI Color code FLV 012 0.1/33
FLV014 7/36
FLV004 16/51/64
FLV005 16/51
FLV009 17/36
FLV010 19/36/48
Figure 5. 10:-Homologous strain neutralisation by anti-DENV-2 antibody
Scatter plot includes result from 15 different plasma samples from 6 individuals collected at
different time points. Bar represents geometric mean at 95%CI. GMT mean titre varies with
in serotype. Each study individuals are color coded as represented in legend. Neutralisation
titre varied in each individual at different time points post infection. Antisera at acute phase
(0.1 MPI) had neutralisation above threshold, same antisera at 33 (MPI) had neutralisation
below GMT for some of the homologous strains. Neutralisation titre for FLV010 was
below GMT for 3/8 homologous strains at all time post infection. Hence the pattern of
neutralisation was variable in each individual.

180
ii. DENV-2 homologous NT decreased with time post infection
Anti-DENV-2 antisera neutralized all homologous DENV. Responses were of greatest
magnitude within the first year of infection and decreased with time. Acute phase sera
showed highest NT (283); convalescent NT were of lesser magnitude; lowest NT (65)
was 48 months post infection in one patient (Figure 5. 12) Consistently high-magnitude
NT were observed for Cosmopolitan Genotype viruses, although D2-15BGCL4 NT
were of lower magnitude, > 1 year post infection, compared to other Cosmopolitan
isolates. D2-10LGA1L9 (Asian 1 Genotype) NT were of greater magnitude than
Cosmopolitan isolates. Lowest NT were directed against the virulent epidemic reference
virus D2-PR6913V. Thus, homologous NT over time post infection is patient- and
virus-specific.

181
Figure 5. 11:- DENV-2-specific neutralisation over time, patient FLV 014
DENV-2 field isolates, epidemic strains and reference strains were
neutralized by anti-DENV-2 antisera. Magnitude of responses declined
between 7 months and 3 years in a strain-specific manner. The greatest
decline was against the virulent epidemic reference strain D2-PR6193V.
Responses against the autologous virus were maintained at the highest
magnitude over time.
3 years after infection
7 months after infection

182
iii. Cross-neutralisation against heterologous DENV
Anti-DENV-2 antisera differentially neutralized heterologous DENV. Cross-
neutralisation was assessed against 13 heterologous DENV strains. Neutralisation
against DENV-4 was approximately 3-fold higher than DENV-1 and DENV-3.
Acute phase sera cross-neutralised all heterologous strains. GMT for acute phase
sera was 320 (DENV-2); 234 (DENV-1); 252 (DENV-3); 248 (DENV-4). The
magnitude of responses declined with time, convalescent sera GMT were lower:
136 (DENV-2); 5 (DENV-1); 8 (DENV-3); 28 (DENV-4). DENV-1 D1-10BGL4
genotype-2-lineage-4 strain was neutralized to the highest magnitude among
DENV-1; non-virulent DENV-3 strain (UNC-3008) was neutralised to the highest
magnitude among DENV-3; and DENV-4 reference strain (D4-H241G1) and patient
isolate DENV-4-genotype 3-lineage 2 were neutralised to the highest magnitude
among DENV-4.

183
Figure 5. 12:- Variation of homologous strain neutralisation by anti-DENV-2.
Change in colour from red to green indicates decrease in neutralisation. The number of strains with
>200 NT decreases with time and the overall response becomes more strain-specific. Responses are
patient- and strain-dependent.
Stu
dy
no
(M
PI)

184
Study no. MPI Color code FLV 012 0.1/33
FLV014 7/36
FLV004 16/51/64
FLV005 16/51
FLV009 17/36
FLV010 19/36/48
Figure 5. 13:- Heterologous cross-neutralisation by DENV-2 antisera
Scatter plot includes result from 15 different plasma samples from 6 individuals
collected at different time points. Black bar represents geometric mean at 95%CI.
Different position of GMT bar represents variation of neutralisation across heterologous
strains of DENV. Each coloured dot represents study individuals. Heterologous NT was
seen up to 16 months post infection.

185
iv. Decreased cross-neutralisation over time
Heterologous cross-neutralization declined over time post infection. Acute phase serum
from patient FLV012 collected 5 days after infection was highly cross-reactive against
all DENV; this cross-neutralization had declined significantly by 33 months post
infection. However, even at 33 months 2 DENV-1, 1 DENV-3 and 3 DENV-4 were still
neutralized by anti-DENV-2 antisera. Persistence of cross-serotype neutralization was
seen in all 6 patients, at low magnitude. Responses were virus-specific: D1-10BG2L4
was neutralized by all DENV-2 antisera with the exception of one serum sample
collected 64 months after infection (patient FLV004); an earlier sample from this patient
collected at 51 months neutralized this strain FRNT50 titre 73 (Table 5.5) Cross
neutralisation against other heterologous strains declined completely: anti-
D1HW2001G4 was completely absent after 19 months.
Heterologous cross-neutralization was strain- and patient-specific. Responses may be
maintained up to 51 months post infection. Longitudinal variation in cross-serotype
neutralization by anti-DENV-2 antisera is shown in Figure 5.14

186
Figure 5. 14:- Decrease in heterologous neutralisation time post infection
Neutralizing capacity of anti-DENV-2 antisera against heterologous DENV strains.
Change in colour of heat map from red to green with increasing time indicates
decrease in cross reactivity. Cross-neutralization of heterologous DENV-1 isolate
D1-10BG2L4 is maintained up to 51 months post infection, whereas neutralization
of D114PHLG4 is lost by 7 months. Differential neutralization of D1-10BG1L14 is
patient antiserum dependent.
Stu
dy
no
(M
PI)

187
v. Neutralisation against autologous DENV-2
Autologous virus and antisera were available for 2 patients, FLV004 and FLV014.
Antisera were collected 14, 51 and 64 months post infection for FLV004, and 7 and 36
months for FLV014. Autologous virus was neutralized by autologous antisera at high
magnitude and this was consistent for both patients at all timepoints. Neutralization by
homologous antisera was of lesser magnitude than for autologous antisera and declined
with time; FLV004 homologous responses had declined significantly at 51 and 64
months compared to autologous responses, and also declined for FLV014. Heterologous
responses declined across all visits for both patients (86% and 94%).
Both autologous viruses belonged to the DENV-2 Cosmopolitan Genotype, Lineage 4,
as did the homologous DENV-2 strains, indicating that variation in neutralization
capacity was patient-dependent.

188
Figure 5.15:- Neutralisation against DENV-2 autologous strains
Neutralisation against autologous virus was compared with neutralisation against heterologous and
homologous virus. Neutralisation of autologous strain was maintained to the highest titre at 14,51 and 64
months post infection for patient FLV004, whereas there was a considerable decrease in homologous and
heterologous over time. Similarly, high magnitude neutralization against FLV014 autologous virus was
maintained at 7 and 36 months post infection, while heterologous and homologous neutralisation
declined.

189
vi. Neutralisation against ZIKV, YF17D and IMOJEV
Fifteen antisera from six individuals were tested against the Flavivirus serogroup.
DENV-2 antisera collected between 0.1 – 19 months post infection neutralized
prototype ZIKV MR 766 only; none neutralised the 2015 epidemic ZIKV PRVABC59,
even among the earliest acute-phase sera. The anti-MR 766 was no longer present at 33
months.
Neutralisation against YF17D vaccine virus was measured only in vaccinated
individuals, in addition to acute phase antisera sampled 1-week post infection. Two
individuals showed neutralisation against YF17D at all time points. One of them was
vaccinated 7 years before, the other individual did not recall being vaccinated. IMOJEV
was neutralised by a single, acute phase antiserum. Change in NAb with time post
infection is shown in Figure 5.16.
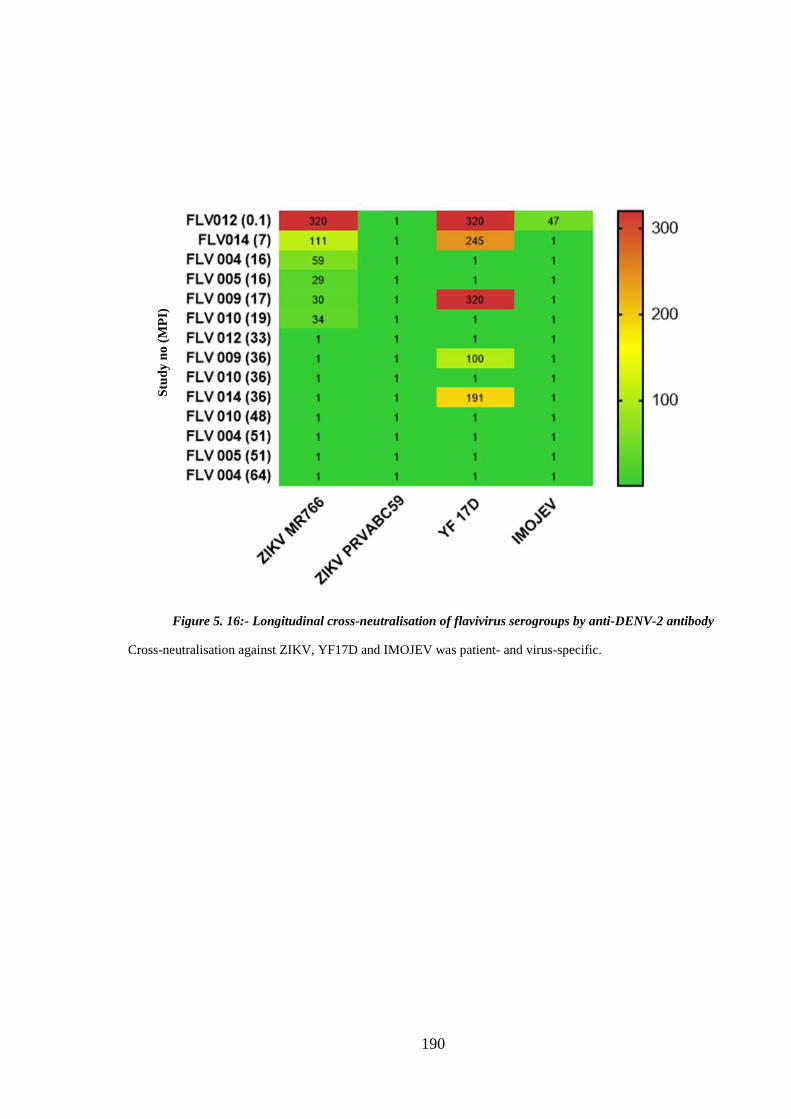
190
Figure 5. 16:- Longitudinal cross-neutralisation of flavivirus serogroups by anti-DENV-2 antibody
Cross-neutralisation against ZIKV, YF17D and IMOJEV was patient- and virus-specific.
Stu
dy
no
(M
PI)

191
c. Neutralising responses post DENV-3 infection
Anti-DENV-3 antisera collected 8-23 months post-infection neutralized all homologous
DENV-3. Heterologous cross-neutralization was observed across most heterologous
viruses within the first year of infection and diminished significantly after 12 months.
Antisera from two of 3 patients sampled in the first year after infection neutralised
DENV-1, DENV-2, or DENV-3 strains at low magnitude; no strains were neutralised by
both antisera. No antisera collected more than 1 year after infection neutralised
heterologous DENV. Compared to DENV-1 and DENV-2, DENV-3 neutralization was
more restricted to homologous viruses. The field strain D3-15GB1, representing
Genotype 1 imported by all DENV-3-infected travellers, and the epidemic non-virulent
reference virus D3-UNC3008-NV were neutralised at greatest magnitude.
Demographic details are summarised in Table 4. 3 (also shown below).
Homologous strain neutralisation by anti-DENV-3 antisera was 27-fold higher than
neutralisation against heterologous serotypes. Tobit regression and robust variance
estimation shows effect of neutralisation (FRNT-50) against homologous serotype was
significantly greater than against heterologous serotype (Table 5.9) The difference
between mean neutralisation between homologous and heterologous strain was ~96%.
No cross-neutralization of ZIKV, YF17D or IMOJEV was detected.
Neutralisation data are listed in Table 5.9-5.12.

192
Sample ID Gender Origin of infection Flavi
Vaccination
Diagnostic tests Months post onset
of illness Location Year of
visit
Duration of Stay
(Days)
Flavi
HI
NS1
result
Dengue
IgM
DFENV
PCR^
FLV-029 M Indonesia 2015 21 No <10 + + + 12
FLV-030 F Bali 2015 8 No <10 + + + 12
FLV-033 F Bali 2015 14 No <10 + + + 14
FLV-035 F Bali 2015 >14 No <10 + + + 8
FLV-037 F Semiyak 2015 6 No <10 + - + 14
FLV-038 M Semiyak 2015 10 No <10 + - + 14 23
FLV-044* F Mexico; Cuba; Bahamas;
USA (Florida)
2015 28 No 160 + + + 8
FLV-045* M Gili Islands/ Bali 2016 11 No <10 + + + 8
FLV-046* M Manado 2015 28 No <10 + - + 18
FLV-047* F Bali 2015 14 No <10 + + + 12
Table 4. 3:- Demographic and laboratory findings in individuals with DENV -3 infection
“*”:- Samples included only in HI test
N/A: - information not available
“^”: Serotyping PCR specific for DENV-3
“+”: - Positive
“- “: - Negative

193
*MPI: Months Past Infection
*Acute: <=2 months past infection
*Convalescent: > 2 months past infection
Table 5. 9:- Anti-DENV-3 neutralising antibody titre at 50% reduction
FLV035
/ 8
FLV029
/12
FLV030
/ 12
FLV033
/14
FLV037
/14
FLV038
/14
FLV038
/ 23
D3-H87G5 320 216 164 51 320 117 108 157
D3-15BG1 320 320 320 209 320 320 113 259
D3-UNC3002-V 320 167 33 48 320 78 55 102
D3-UNC3008-NV 320 227 320 107 320 159 134 208
GMT-D3 320 226 154 86 320 147 97 172
D1-HW2001G4 1 1 1 1 1 1 1 1
D1-10BG1L14 320 31 1 1 51 70 1 12
D1-15LG1 38 40 1 1 27 1 1 5
D1-10BG2L4 27 178 20 1 27 1 1 8
D1-10INDG3L4 22 48 21 1 25 1 1 7
D1-14PHLG4 39 100 1 1 1 1 1 3
GMT-D1 26 32 3 1 10 2 1 5
D2-NGCGAII 1 40 1 1 1 1 1 2
D2-10LGA1L9 184 79 25 1 36 1 1 10
D2-11BGCL4 1 1 1 1 1 1 1 1
D2-13MGCLN 29 27 1 1 83 51 1 9
D2-15BGCL4 1 29 1 1 1 1 1 2
D2-15BGCLN 52 27 35 1 1 1 1 5
D2-PR6913-V 25 1 1 1 1 1 1 2
D2-PR1940-NV 21 1 1 1 1 23 22 4
GMT-D2 11 9 2 1 3 2 1 3
D4-H241G1 36 189 32 1 31 1 1 9
D4-10BG3L1 35 44 1 1 70 60 1 9
D4-10BG3L2 77 151 23 1 84 27 1 18
GMT-D4 46 108 9 1 57 12 1 12
ZIKV MR766 1 1 1 1 1 1 1 1
ZIKV PRVABC59 1 1 1 1 1 1 1 1
YF 17D 1 1 1 1 1 1 1 1
IMOJEV 1 1 1 1 1 1 1 1
GMT 1 1 1 1 1 1 1 1
GMT-flavi:NAOther
Flaviviruses
GMT-D2 strains:3
(<20-184)
GMT-D4
strains:12
(<20-275)
Virus
Heterologous
serotype
GMT-heterologous
serotype= 5.5 (<20-
184)
Geometric Mean Titre
Study No./ MPI
Homologous
serotype
GMT- total sera homologous
serotype=172 (33-320+)
GMT-D1 strains :
5 (<20-320)
<1 year 1-2 year
GMT
Note:
Value 1=<20
Value 320=>320

194
Note:
Value 1=<20
Value 320=>320
Table 5. 10:-Anti-DENV-3 neutralising antibody titre at 75% reduction
*MPI: Months Past Infection
*Acute: <=2 months past infection
*Convalescent: > 2 months past infection
FLV035
/ 8
FLV029
/12
FLV030 /
12
FLV033
/14
FLV037
/14
FLV038
/14
FLV038
/23
D3-H87G5 320 110 85 1 306 42 43 55
D3-15BG1 320 320 320 68 320 320 58 201
D3-UNC3002-V 320 72 22 23 187 39 30 59
D3-UNC3008-NV 320 70 212 40 320 61 63 113
GMT-D3 320 115 106 16 277 75 46 93
D1-HW2001G4 1 1 1 1 1 1 1 1
D1-10BG1L14 60 1 1 1 1 1 1 2
D1-15LG1 1 1 1 1 1 1 1 1
D1-10BG2L4 1 65 1 1 1 1 1 2
D1-10INDG3L4 1 25 1 1 1 1 1 2
D1-14PHLG4 20 40 1 1 1 1 1 3
GMT-D1 3 6 1 1 1 1 1 2
D2-NGCGAII 1 1 1 1 1 1 1 1
D2-10LGA1L9 46 39 1 1 23 1 1 5
D2-11BGCL4 1 1 1 1 1 1 1 1
D2-13MGCLN 1 1 1 1 1 1 1 1
D2-15BGCL4 1 1 1 1 1 1 1 1
D2-15BGCLN 1 1 1 1 1 1 1 1
D2-PR6913-V 1 1 1 1 1 1 1 1
D2-PR1940-NV 1 1 1 1 1 1 1 1
GMT-D2 2 2 1 1 1 1 1 1
D4-H241G1 1 69 1 1 1 1 1 2
D4-10BG3L1 1 1 1 1 1 1 1 1
D4-10BG3L2 32 62 1 1 50 1 1 5
GMT-D4 3 16 1 1 4 1 1 2
ZIKV MR766 1 1 1 1 1 1 1 1
ZIKV PRVABC59 1 1 1 1 1 1 1 1
YF 17D 1 1 1 1 1 1 1 1
IMOJEV 1 1 1 1 1 1 1 1
GMT 1 1 1 1 1 1 1 1
GMT-flavi:<20
Virus
Homologous
serotype
Heterologous
serotype
Other
Flaviviruses
<1 year
Study No./ MPI
Geometric Mean Titre
GMT- total sera homologous
serotype=93 (<20-320+)
GMT-D1
strains : 2
(<20-65)
GMT-heterologous
serotype= 1(<20-69)GMT-D2
strains:1
(<20-46)
GMT-D4
strains:2
(<20-69)
1-2 year
GMT

195
Note:
Value 1=<20
Value 320=>320
*MPI: Months Past Infection
*Acute: <=2 months past infection
*Convalescent: > 2 months past infection
Table 5. 11:- Anti-DENV-3 neutralising antibody titre at 90% reduction
FLV035 /
8
FLV029
/12
FLV030 /
12
FLV033
/14
FLV037
/14
FLV038
/14
FLV038/
23
D3-H87G5 181 46 37 1 148 1 18 19
D3-15BG1 320 320 85 32 320 96 33 116
D3-UNC3002-V 170 40 1 14 96 25 21 24
D3-UNC3008-NV 320 37 93 13 320 34 38 67
GMT-D3 237 68 23 9 195 17 26 43
D1-HW2001G4 1 1 1 1 1 1 1 1
D1-10BG1L14 27 1 1 1 1 1 1 2
D1-15LG1 1 1 1 1 1 1 1 1
D1-10BG2L4 1 1 1 1 1 1 1 1
D1-10INDG3L4 1 1 1 1 1 1 1 1
D1-14PHLG4 1 23 1 1 1 1 1 2
GMT-D1 2 2 1 1 1 1 1 1
D2-NGCGAII 1 1 1 1 1 1 1 1
D2-10LGA1L9 1 23 1 1 1 1 1 2
D2-11BGCL4 1 1 1 1 1 1 1 1
D2-13MGCLN 1 1 1 1 1 1 1 1
D2-15BGCL4 1 1 1 1 1 1 1 1
D2-15BGCLN 1 1 1 1 1 1 1 1
D2-PR6913-V 34 1 1 1 1 1 1 2
D2-PR1940-NV 1 1 1 1 1 1 1 1
GMT-D2 2 1 1 1 1 1 1 1
D4-H241G1 1 30 1 1 1 1 1 2
D4-10BG3L1 1 1 1 1 1 1 1 1
D4-10BG3L2 1 1 1 1 1 1 1 1
GMT-D4 1 3 1 1 1 1 1 1
ZIKV MR766 1 1 1 1 1 1 1 1
ZIKV PRVABC59 1 1 1 1 1 1 1 1
YF 17D 1 1 1 1 1 1 1 1
IMOJEV 1 1 1 1 1 1 1 1
GMT 1 1 1 1 1 1 1 1
Other
Flaviviruses
Virus Geometric Mean Titre
GMT- total sera homologous
serotype=43(<20-320+)
GMT-D1
strains : 1
(<20-23)
GMT-heterologous
serotype= 1 (<20-34)GMT-D2
strains:1
(<20-34)
GMT-D4
strains:1
(<20-30)
GMT-flavi: (<20)
<1 year
Homologous
serotype
Heterologous
serotype
1-2 year
Study No./ MPI
GMT

196
Note: Virus values are average differences from reference strain in each individual panel
Variable: Time post infection and strains of DENV tested
Effect:- Difference in log FRNT50 with censoring cut-offs set to 2.99-5.77
p-value:- Significance of effect of neutralisation compared with reference strains in each
serotype
95%CI:- robust variance estimation of CI for regression coefficients.
Variable
Effect
(difference in
log(FRNT50))
Std. Err. tobit p-value (95%
Conf.interval)
Change per month -0.192 0.047 -4.07 0.010 -0.313 -0.071
Homologous serotype overall 3.723 1.554 2.4 0.062 -0.271 7.716
Homologous
serotype
D3-H87G5 0.000
D3-15BG1 2.687 0.866 3.1 0.027 0.462 4.912
D3-UNC3002-
V -0.781 0.420 -1.86 0.122 -1.861 0.298
D3-UNC3008-
NV 1.108 0.814 1.36 0.232 -0.984 3.199
Heterologous
serotype
D1-
HW2001G4 0.000
D1-10BG1L14 2.804 1.520 1.85 0.124 -1.103 6.711
D1-15LG1 1.005 0.587 1.71 0.148 -0.505 2.514
D1-10BG2L4 1.311 0.880 1.49 0.196 -0.951 3.573
D1-
10INDG3L4 0.867 0.505 1.71 0.147 -0.433 2.166
D1-14PHLG4 0.996 0.769 1.29 0.252 -0.982 2.973
D2-NGCGAII 0.000
D2-
10LGA1L9 1.520 0.902 1.69 0.153 -0.798 3.839
D2-11BGCL4 -0.388 0.435 -0.89 0.414 -1.506 0.730
D2-
13MGCLN 1.162 0.692 1.68 0.154 -0.617 2.942
D2-15BGCL4 -0.107 0.120 -0.89 0.414 -0.414 0.201
D2-15BGCLN 0.665 0.653 1.02 0.355 -1.013 2.343
D2-PR6913-V -0.167 0.524 -0.32 0.763 -1.515 1.181
D2-PR1940-
NV -0.022 0.572 -0.04 0.971 -1.493 1.449
D4-H241G1 0.000
D4-10BG3L1 0.003 0.867 0 0.997 -2.225 2.232
D4-10BG3L2 0.912 0.797 1.14 0.304 -1.137 2.961
Table 5. 12:- DENV-3-Sera panel statistical verification

197
i. Neutralisation among homologous DENV-3
Anti-DENV-3 antisera neutralised all homologous DENV-3, with varying magnitude.
Neutralization of four virus strains of DENV-3 serotype was assessed: WHO reference
DENV-3 strain (D3-H87G5); virulent and nonvirulent epidemic strains collected in Sri
Lanka in 1983 and 1989, respectively (UNC3002 and UNC3008); and Genotype 1
patient field isolate from 2015. A single patient isolate was tested because all DENV-3
imported by travellers to 2016 belong to Genotype 1.
Differential, strain-specific neutralisation within DENV-3 was observed (Figure 5.17).
Neutralisation of the field isolate D3-15BG1 was of significantly greater magnitude
than neutralization of the reference strain and epidemic virulent and non-virulent
strains. Comparison of confidence intervals for regression coefficients showed non-
virulent strain UNC3008 neutralisation magnitude was greater than virulent strain
UNC3002 (Table 5.9).

198
Study
no. MPI Color
code FLV 035 8 FLV029 12 FLV030 12 FLV033 14 FLV037 14 FLV038 14/23
Figure 5.17:- Homologous strain neutralisation by anti-DENV-3 antisera.
DENV-3 antisera differentially neutralized homologous DENV-3. Black lines are GMT at 95%CI. GMT is
greatest for field isolate D3-15BG1. The epidemic virulent reference strain D3-UNC3002-V was neutralized at
lowest magnitude. Among the study population, one individual (FLV033) at 14MPI has NT below GMT
against all the homologous strains. FLV038 at had NT at threshold (NT>3020) against field isolate D3-15BG1
at 14 MPI while NT dropped below GMT at 23 MPI.

199
ii. DENV-3 neutralisation over time
NT against homologous DENV-3 declined over time in the small number of antisera
tested. Patient FLV035 sampled 8 months post infection had NT for >320, two other
antisera samples at 12 months also showed NT of 320. Patient FLV038 was sampled 14
and 23 months after infection; in the second sample NT had declined from 320 (the
endpoint of the assay) to 113 (Table 5.9, Figure 5.18). This patient was infected with
D3-15BG1, the strain imported by all DENV-3 travellers and representing the dominant
strain circulating in the region at that time. Two patients sampled at 14 months
differentially neutralised DENV-3: one patient FLV033 showed low NT of 48 (against
epidemic virulent strain D3-UNC3002-V) while NT against the dominant virus
circulating in the region D3-15BG1 was maintained at the highest dilution (320); the
other patient FLV037 showed high magnitude NT at 320 against all DENV-3. As for
DENV-1 and DENV-2, DENV-3 homologous responses are patient- and virus-specific.
Figure 17 shows neutralisation of homologous strains over time. Overall comparison of
NT over time of DENV-3 antisera against DENV-3 is shown in heat map (Figure 5.19).
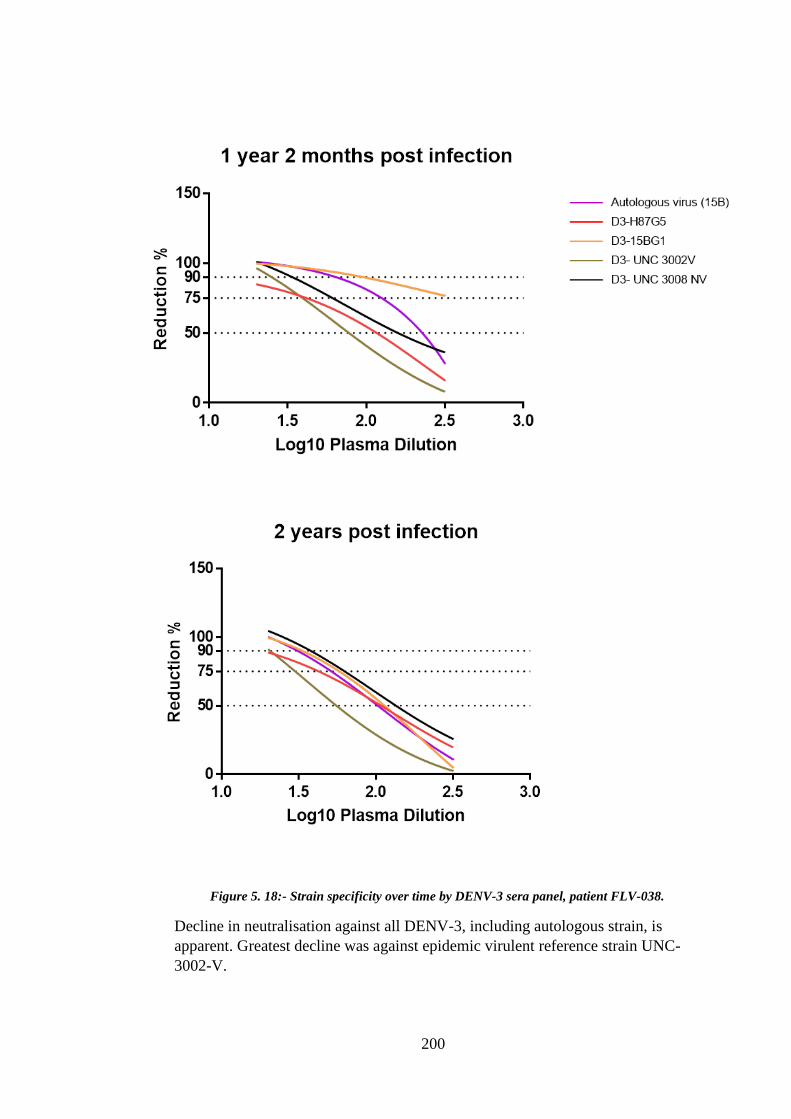
200
Figure 5. 18:- Strain specificity over time by DENV-3 sera panel, patient FLV-038.
Decline in neutralisation against all DENV-3, including autologous strain, is
apparent. Greatest decline was against epidemic virulent reference strain UNC-
3002-V.
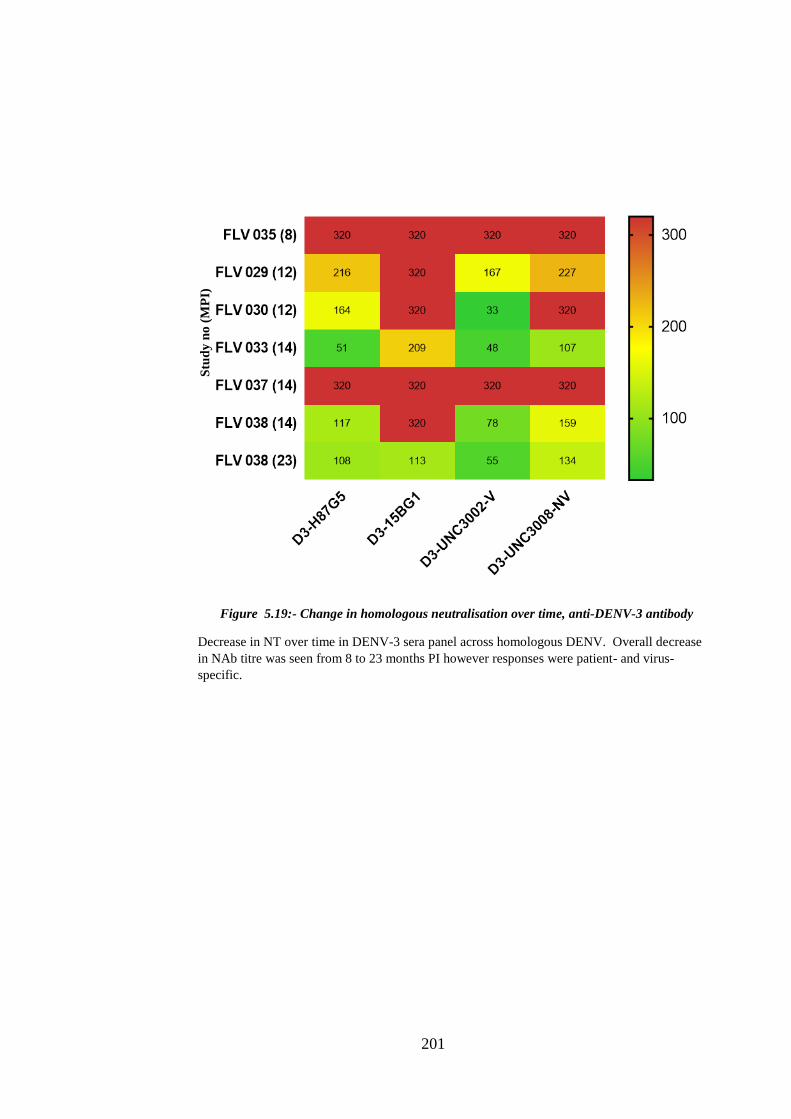
201
Figure 5.19:- Change in homologous neutralisation over time, anti-DENV-3 antibody
Decrease in NT over time in DENV-3 sera panel across homologous DENV. Overall decrease
in NAb titre was seen from 8 to 23 months PI however responses were patient- and virus-
specific.
Stu
dy
no
(M
PI)

202
iii. DENV-3 cross-neutralisation against heterologous serotypes
FLV035 antiserum collected 8 months after infection neutralised 13 of 17 heterologous
DENV. There was an overall decrease in NT (<100x) against heterologous serotypes at
12 months post infection however in 1 of 3 patients (FLV037) cross-neutralisation was
maintained at 12 months against 13 of 17 viruses. Neutralisation against DENV-4 was
2-fold higher than against DENV-1 and 4-fold higher than against DENV-2. Strain-
specific variation in cross-NT was apparent (Figure 5.20). Among strains of DENV-1,
patient isolate D1-10BG1LG1 (Genotype 1 lineage 14, originating in Bali 2010) has
high (~3-fold) neutralisation (GMT) than D1-15LG1LH (Genotype 1, lineage H,
originating Bali 2015), however reference strain D1HW2001 (Genotype 4) was not
neutralised by any anti-DENV-3 antisera. Similarly, among DENV-2 strains
neutralisation against D2-10LGA1L9 (Asian I Genotype originating in Laos 2010) and
D2-13MGCLN (Cosmpolitan Genotype originating in Malaysia 2013) was ~2-fold
higher than other strains. Three of 6 anti-DENV-3 antisera had detectable neutralisation
(NAb>20) against the DENV-2 non-virulent epidemic reference strain PR1940 while
only 1 of 6 neutralised the DENV-2 epidemic virulent reference strain PR6913. Within
DENV-2 Cosmopolitan Genotype strains there was variation in NT: D2-11BGCL4
(originating Bali 2011) was <20, ranging to 83 (D2-13MGCLN, originating Malaysia
2013). Differential neutralisation was also observed against DENV-4, with high NT
against D4-10BG3L2 (originating Bali 2010) compared to D4-10BG3L1 (originating
Bali 2010), viruses which belong to different lineages of the same genotype.
As for DENV-1 and DENV-2, cross-neutralisation of heterologous DENV was patient-
and virus-specific. Variation in DENV-3 heterologous cross-neutralisation over time is
shown in Table 5.7 and in the heat map (Figure 5.21).

203
Study
no. MPI Color
code FLV 035 8 FLV029 12 FLV030 12 FLV033 14 FLV037 14 FLV038 14/23
Figure 5. 20:- Heterologous cross-neutralisation by DENV-3 antisera
DENV-3 antisera against heterologous strains showed GMT for each group was <100 while NT for certain
strains were >320. Differential NT was also patient-specific and >100 for some individuals. DENV-3
showed less cross-NT than DENV-1 and DENV-2.
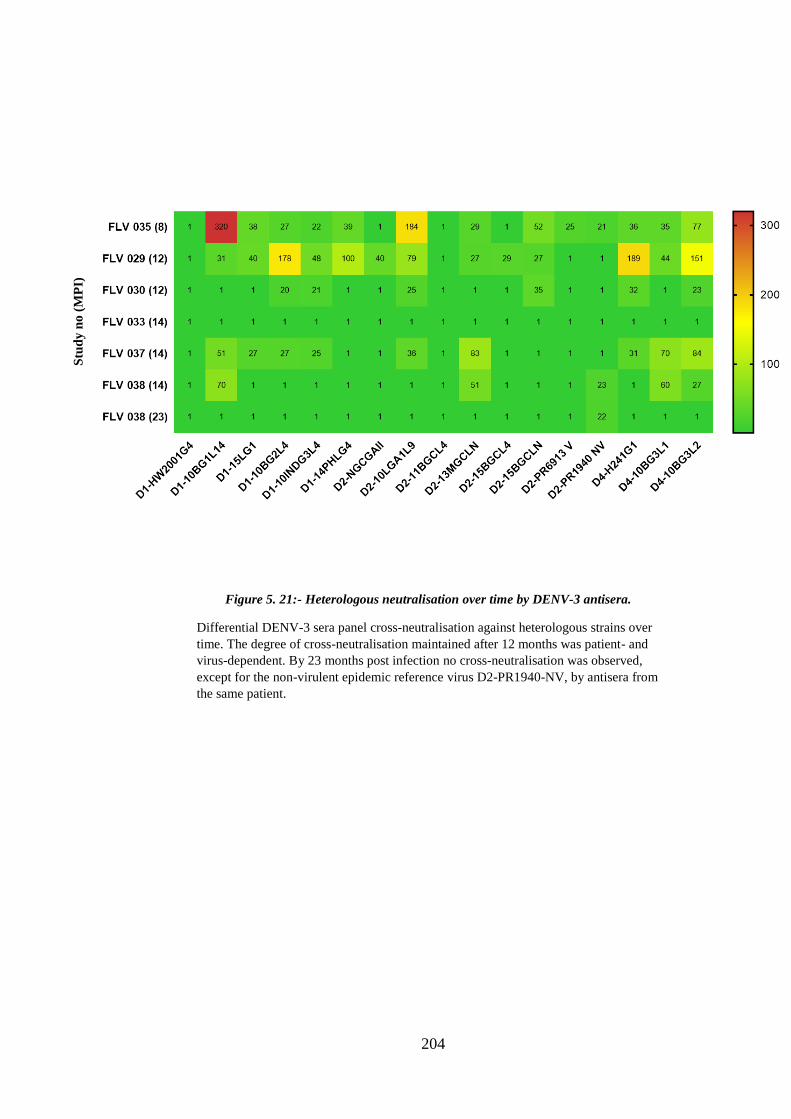
204
Figure 5. 21:- Heterologous neutralisation over time by DENV-3 antisera.
Differential DENV-3 sera panel cross-neutralisation against heterologous strains over
time. The degree of cross-neutralisation maintained after 12 months was patient- and
virus-dependent. By 23 months post infection no cross-neutralisation was observed,
except for the non-virulent epidemic reference virus D2-PR1940-NV, by antisera from
the same patient.
Stu
dy
no
(M
PI)

205
iii. Neutralisation against autologous serotype
Decrease in neutralisation against autologous and homologous strains was seen 23
months post infection in one patient.
DENV was isolated from two patients, FLV037 and FLV038, who had travelled to Bali
Indonesia in 2015. Both isolates belong to Genotype 1 (D3-15BG1). Blood samples
were obtained 14 months after infection (FLV037) and 14 and 23 months after infection
(FLV038). Autologous and homologous viruses were equally neutralised by FLV037
antisera while FLV038 showed a decline in NT at 23 months, from >320 at 14
months to 113 at 23 months.(Figure 5.22).
iv. Neutralisation against related serogroups
Cross-reactivity against ZIKV, YF17D or IMOJEV was not detected for any DENV-3
antisera.

206
Figure 5.22:- Autologous strain neutralisation by DENV-3 antisera
Autologous virus was available for 2 patients from whom antisera were obtained 14
months; and 14 and 23 months, respectively. Responses in the two patients at 14 months
were distinct: FLV017 NT were maintained at high magnitude whereas FLV038 were of
lower magnitude and declined further at 23 months. NT against heterologous DENV were
always just above or below limits of detection.

207
d. Specificity of neutralising antibody response post DENV-4 infection
DENV-4 antisera neutralised homologous viruses at high magnitude (GMT 168)
compared to heterologous DENV (GMT 2). DENV-4 sera panel showed less cross-
neutralisation than DENV-1, DENV-2, and DENV-3. No cross reactivity against any
flavivirus serogroup was observed.
Six individuals were included (Table 4.4 also shown below). Longitudinal samples
were obtained from two individuals (FLV049 and FLV002) at different times post
infection, ranging from 8 to 72 months (six years). NT at three cut-offs (50%; 75%;
90%) are listed in Table 5. 12- Table 5. 14. At 75% and 90 % reduction NT against all
heterologous strains is <30.
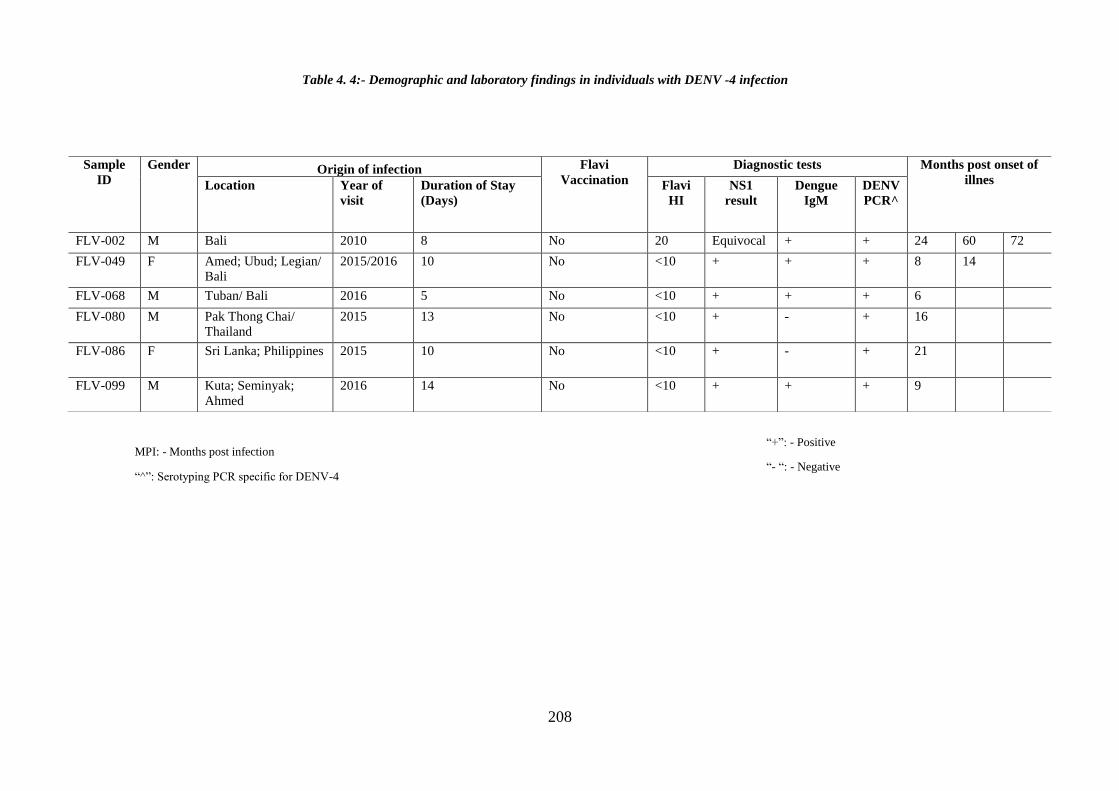
208
Sample
ID
Gender Origin of infection Flavi
Vaccination
Diagnostic tests Months post onset of
illnes Location Year of
visit
Duration of Stay
(Days)
Flavi
HI
NS1
result
Dengue
IgM
DENV
PCR^
FLV-002 M Bali 2010 8 No 20 Equivocal + + 24 60 72
FLV-049 F Amed; Ubud; Legian/
Bali
2015/2016 10 No <10 + + + 8 14
FLV-068 M Tuban/ Bali 2016 5 No <10 + + + 6
FLV-080 M Pak Thong Chai/
Thailand
2015 13 No <10 + - + 16
FLV-086 F Sri Lanka; Philippines 2015 10 No <10 + - + 21
FLV-099 M Kuta; Seminyak;
Ahmed
2016 14 No <10 + + + 9
Table 4. 4:- Demographic and laboratory findings in individuals with DENV -4 infection
“+”: - Positive
“- “: - Negative
MPI: - Months post infection
“^”: Serotyping PCR specific for DENV-4

209
Figure 5.23:- Homologous and heterologous DENV neutralisation by anti-DENV-4
antibody, Patient FLV 049
Antisera from this patient showed high NT against homologous DENV within the first year of
infection; one heterologous virus was neutralized. Fourteen months after infection (6 months
after the first sample) no serotype cross-neutralisation was observed.
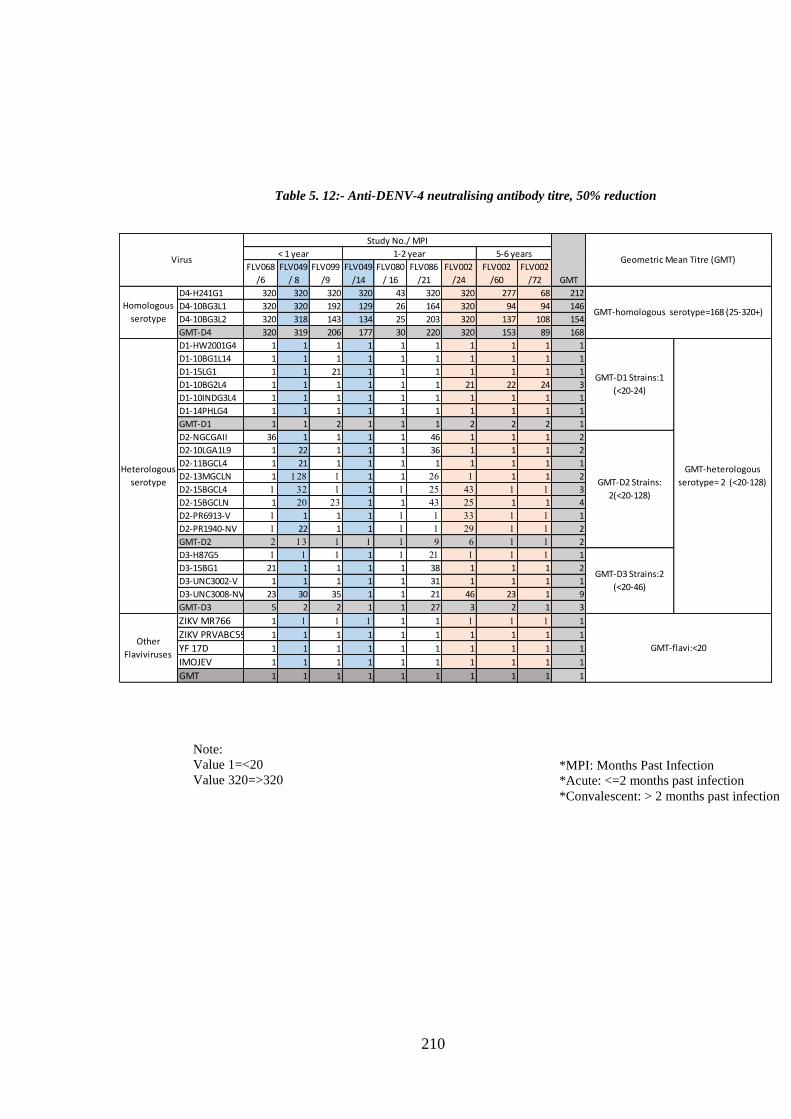
210
*MPI: Months Past Infection
*Acute: <=2 months past infection
*Convalescent: > 2 months past infection
Note:
Value 1=<20
Value 320=>320
Table 5. 12:- Anti-DENV-4 neutralising antibody titre, 50% reduction
FLV068
/6
FLV049
/ 8
FLV099
/9
FLV049
/14
FLV080
/ 16
FLV086
/21
FLV002
/24
FLV002
/60
FLV002
/72
D4-H241G1 320 320 320 320 43 320 320 277 68 212
D4-10BG3L1 320 320 192 129 26 164 320 94 94 146
D4-10BG3L2 320 318 143 134 25 203 320 137 108 154
GMT-D4 320 319 206 177 30 220 320 153 89 168
D1-HW2001G4 1 1 1 1 1 1 1 1 1 1
D1-10BG1L14 1 1 1 1 1 1 1 1 1 1
D1-15LG1 1 1 21 1 1 1 1 1 1 1
D1-10BG2L4 1 1 1 1 1 1 21 22 24 3
D1-10INDG3L4 1 1 1 1 1 1 1 1 1 1
D1-14PHLG4 1 1 1 1 1 1 1 1 1 1
GMT-D1 1 1 2 1 1 1 2 2 2 1
D2-NGCGAII 36 1 1 1 1 46 1 1 1 2
D2-10LGA1L9 1 22 1 1 1 36 1 1 1 2
D2-11BGCL4 1 21 1 1 1 1 1 1 1 1
D2-13MGCLN 1 128 1 1 1 26 1 1 1 2
D2-15BGCL4 1 32 1 1 1 25 43 1 1 3
D2-15BGCLN 1 20 23 1 1 43 25 1 1 4
D2-PR6913-V 1 1 1 1 1 1 33 1 1 1
D2-PR1940-NV 1 22 1 1 1 1 29 1 1 2
GMT-D2 2 13 1 1 1 9 6 1 1 2
D3-H87G5 1 1 1 1 1 21 1 1 1 1
D3-15BG1 21 1 1 1 1 38 1 1 1 2
D3-UNC3002-V 1 1 1 1 1 31 1 1 1 1
D3-UNC3008-NV 23 30 35 1 1 21 46 23 1 9
GMT-D3 5 2 2 1 1 27 3 2 1 3
ZIKV MR766 1 1 1 1 1 1 1 1 1 1
ZIKV PRVABC59 1 1 1 1 1 1 1 1 1 1
YF 17D 1 1 1 1 1 1 1 1 1 1
IMOJEV 1 1 1 1 1 1 1 1 1 1
GMT 1 1 1 1 1 1 1 1 1 1
Study No./ MPI
Heterologous
serotype
GMT-heterologous
serotype= 2 (<20-128)
Homologous
serotypeGMT-homologous serotype=168 (25-320+)
GMT-D1 Strains:1
(<20-24)
GMT-D2 Strains:
2(<20-128)
GMT-D3 Strains:2
(<20-46)
Virus5-6 years1-2 year
GMT
< 1 year
Other
Flaviviruses
Geometric Mean Titre (GMT)
GMT-flavi:<20

211
*MPI: Months Past Infection
*Acute: <=2 months past infection
*Convalescent: > 2 months past infection
Note:
Value 1=<20
Value 320=>320
Table 5. 13:- Anti-DENV-4 neutralising antibody titre, 75% reduction
FLV068
/36
FLV049
/ 8
FLV 099/
9
FLV049/
14
FLV080/
16
FLV086/
21
FLV002
/24
FLV002
/60
FLV002
/72
D4-H241G1 231 191 320 237 28 320 320 135 37 154
D4-10BG3L1 320 247 71 38 1 82 181 43 43 57
D4-10BG3L2 190 157 108 44 1 94 273 56 49 60
GMT-D4 241 195 135 73 3 135 251 69 43 80
D1-HW2001G4 1 1 1 1 1 1 1 1 1 1
D1-10BG1L14 1 1 1 1 1 1 1 1 1 1
D1-15LG1 1 1 1 1 1 1 1 1 1 1
D1-10BG2L4 1 1 1 1 1 1 1 1 1 1
D1-10INDG3L4 1 1 1 1 1 1 1 1 1 1
D1-14PHLG4 1 1 1 1 1 1 1 1 1 1
GMT-D1 1 1 1 1 1 1 1 1 1 1
D2-NGCGAII 1 1 1 1 1 1 1 1 1 1
D2-10LGA1L9 29 1 1 1 1 1 1 1 1 1
D2-11BGCL4 1 1 1 1 1 1 1 1 1 1
D2-13MGCLN 1 1 1 1 1 1 1 1 1 1
D2-15BGCL4 1 1 1 1 1 1 1 1 1 1
D2-15BGCLN 1 1 1 1 1 1 1 1 1 1
D2-PR6913-V 1 1 1 1 1 1 1 1 1 1
D2-PR1940-NV 1 1 1 1 1 1 1 1 1 1
GMT-D2 2 1 1 1 1 1 1 1 1 1
D3-H87G5 1 1 1 1 1 1 1 1 1 1
D3-15BG1 1 1 1 1 1 1 1 1 1 1
D3-UNC3002-V 1 1 1 1 1 1 1 1 1 1
D3-UNC3008-NV 1 1 1 1 1 1 1 1 1 1
GMT-D3 1 1 1 1 1 1 1 1 1 1
ZIKV MR766 1 1 1 1 1 1 1 1 1 1
ZIKV PRVABC59 1 1 1 1 1 1 1 1 1 1
YF 17D 1 1 1 1 1 1 1 1 1 1
IMOJEV 1 1 1 1 1 1 1 1 1 1
GMT 1 1 1 1 1 1 1 1 1 1
Geometric Mean Titre
(GMT)
Study No./ MPI
Homologous
serotype
Heterologous
serotype
Other
Flaviviruses
Virus< 1 year 1-2 year 5-6 years
GMT-flavi:NA
GMT-homologous
serotype=80(<20-320+)
GMT-D1
Strains:<20
GMT-
heterologous
serotype= 1
(<20-29)
GMT-D2
Strains: 1
(<20-29)
GMT-D3
Strains:
<20
GMT

212
Note:
Value 1=<20
Value 320=>320
*MPI: Months Past Infection
*Acute: <=2 months past infection
*Convalescent: > 2 months past infection
Table 5. 14:- Anti-DENV-4 neutralising antibody titre, 90% reduction
FLV068
/6
FLV049 /
8
FLV 099/
9
FLV049
/14
FLV080
/ 16
FLV08
6/21
FLV002
/24
FLV002
/60
FLV002
/72
D4-H241G1 107 81 215 111 22 179 238 61 25 88
D4-10BG3L1 192 113 42 25 1 46 71 29 29 34
D4-10BG3L2 72 77 57 27 1 51 118 28 30 33
GMT-D4 114 89 80 42 3 75 126 37 28 46
D1-HW2001G4 1 1 1 1 1 1 1 1 1 1
D1-10BG1L14 1 1 1 1 1 1 1 1 1 1
D1-15LG1 1 1 1 1 1 1 1 1 1 1
D1-10BG2L4 1 1 1 1 1 1 1 1 1 1
D1-10INDG3L4 1 1 1 1 1 1 1 1 1 1
D1-14PHLG4 1 1 1 1 1 1 1 1 1 1
GMT-D1 1 1 1 1 1 1 1 1 1 1
D2-NGCGAII 1 1 1 1 1 1 1 1 1 1
D2-10LGA1L9 1 1 1 1 1 1 1 1 1 1
D2-11BGCL4 1 1 1 1 1 26 1 1 1 1
D2-13MGCLN 1 1 1 1 1 25 1 1 1 1
D2-15BGCL4 1 1 1 1 1 1 1 1 1 1
D2-15BGCLN 1 1 1 1 1 1 1 1 1 1
D2-PR6913-V 1 1 1 1 1 1 1 1 1 1
D2-PR1940-NV 1 1 1 1 1 1 1 1 1 1
GMT-D2 1 1 1 1 1 2 1 1 1 1
D3-H87G5 1 1 1 1 1 1 1 1 1 1
D3-15BG1 1 1 1 1 1 1 1 1 1 1
D3-UNC3002-V 1 1 1 1 1 1 1 1 1 1
D3-UNC3008-NV 1 1 1 1 1 1 1 1 1 1
GMT-D3 1 1 1 1 1 1 1 1 1 1
ZIKV MR766 1 1 1 1 1 1 1 1 1 1
ZIKV PRVABC59 1 1 1 1 1 1 1 1 1 1
YF 17D 1 1 1 1 1 1 1 1 1 1
IMOJEV 1 1 1 1 1 1 1 1 1 1
GMT 1 1 1 1 1 1 1 1 1 1
GMT
Other
Flaviviruses
Study No./ MPI
Geometric Mean Titre (GMT)
GMT-homologous serotype=46
(<20-238+)
GMT-D1 Strains:
<20
GMT-
heterologous
serotype= 1
(<20-26)
GMT-D2 Strains:
1(<20-26)
GMT-D3
Strains:<20
GMT-flavi: (<20)
1-2 year 5-6 yearsVirus
Homologous
serotype
Heterologous
serotype
< 1 year

213
Table 5. 16:-DENV-4-Sera panel statistical verification
Variable
Effect
(difference in
log(FRNT50))
Std. Err. tobit p-value (95% Conf.interval)
Change per month -0.012 0.004 -3.27 0.022 -0.021 -0.003
Homologous serotype 3.585 1.127 3.18 0.024 0.689 6.481
Homologous
serotype
D4-H241G1 0.000
D4-10BG3L1 -2.209 0.858 -2.57 0.050 -4.415 -0.002
D4-10BG3L2 -2.214 0.972 -2.28 0.072 -4.712 0.283
Heterologous
serotype
D1-HW2001G4 0.000
D1-10BG1L14 0.000 0.000 0 1.000 0.000 0.000
D1-15LG1 0.057 0.066 0.87 0.426 -0.112 0.227
D1-10BG2L4 0.303 0.260 1.17 0.296 -0.364 0.970
D1-10INDG3L4 0.000 0.000 0 1.000 0.000 0.000
D1-14PHLG4 0.000 0.000 0 1.000 0.000 0.000
D2-NGCGAII 0.000
D2-10LGA1L9 -0.229 0.359 -0.64 0.551 -1.152 0.693
D2-11BGCL4 -0.576 0.506 -1.14 0.307 -1.878 0.725
D2-13MGCLN 0.096 0.703 0.14 0.896 -1.712 1.905
D2-15BGCL4 0.127 0.578 0.22 0.834 -1.358 1.612
D2-15BGCLN -0.010 0.402 -0.02 0.982 -1.042 1.022
D2-PR6913-V -0.351 0.631 -0.56 0.602 -1.974 1.271
D2-PR1940-NV -0.295 0.616 -0.48 0.652 -1.877 1.288
D3-H87G5 0.000
D3-15BG1 0.285 0.277 1.03 0.349 -0.426 0.997
D3-UNC3002-V 0.183 0.211 0.87 0.426 -0.360 0.725
D3-UNC3008-
NV 1.081 0.287 3.77 0.013 0.343 1.819
Note: Virus values are average differences from reference strain in each individual panel
Variable: Time post infection and strains of DENV tested
Effect: - Difference in log FRNT50 with censoring cut-offs set to 2.99-5.77
p-value: - Significance of effect of neutralization compared with reference strains in each serotype
95%CI: - robust variance estimation of CI for regression coefficients.

214
i. Longitudinal DENV-4 antisera against homologous virus
DENV-4 sera panel showed significantly greater homologous serotype neutralisation
compared with heterologous DENV. One reference strain (D4-H241G1) and two field
isolates D4-10BG3L1 and D4-10BG3L2, both originating in Bali Indonesia in 2010 and
representing lineage 1 and lineage 2 of Genotype 3 were tested. NT varied across the 3
viruses and were highest for the reference strain, which belonged to Genotype 1 (Figure
5.24); p-value for difference in log FRNT-50 was significantly lower (Table 5.16).
NTs against homologous DENV were maintained at high magnitude for antisera from 4
of 5 patients, up to 2 years after infection (Table 5.12). Antisera from the 5th
patient
(FLV080) sampled 16 months after infection neutralised all four DENV-4 at low
magnitude. This patient was infected in the Thailand in 2015, E gene sequence could
not be obtained.
Increase in specificity with increased time post infection was seen in the 2 patients for
whom consecutive samples were available and neutralisation potency per month among
the individual showed significant variation (p<0.022). Homologous neutralisation
declined overall, in a patient- and virus-specific manner (Figure 5.25andFigure 5.26)

215
Study no. MPI Color
code FLV 068 6
FLV049 8/14 FLV099 9 FLV086 21 FLV080 16 FLV002 24/60/72
Figure 5. 24:- Homologous virus neutralisation by DENV-4 antisera
DENV-4 sera panel against homologous strains shows differential neutralisation of reference vs
field strains. NT against reference virus D4-H241G1, which belongs to a different genotype
than the traveller field isolates, is of greater magnitude. Scatter plot is from 9 plasma sample
represented by dots from 6 individuals. Each dot is color coded as represented in legend. NAb
against DENV-4 reference strain was mostly >320 compare to patient isolates. Individual at 16
MPI (FLV080) showed constant low neutralisation against all homologous strains of DENV-4.

216
Figure 5. 25:- Homologous strain neutralisation by DENV-4 (FLV049)
Differential neutralisation of DENV-4 reference and field viruses by
FLV049 anti-DENV-4 antisera 8 and 14 months post infection.
8 months post infection.
14 months post infection.
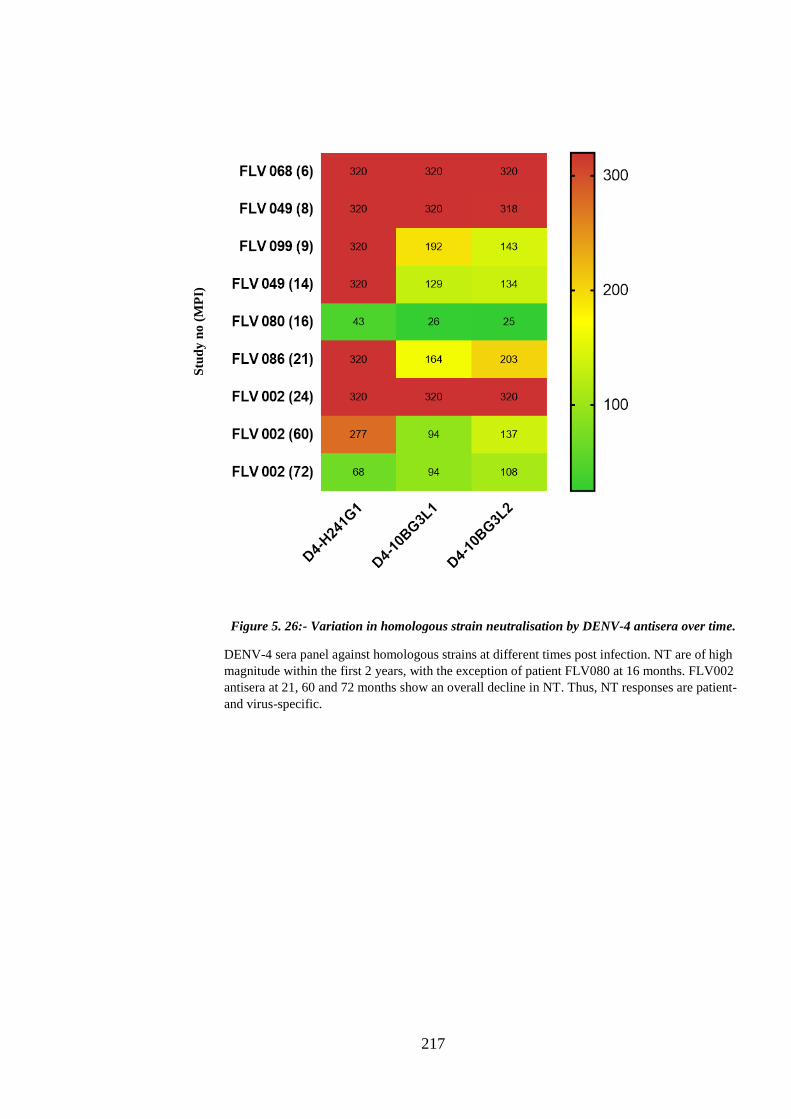
217
Figure 5. 26:- Variation in homologous strain neutralisation by DENV-4 antisera over time.
DENV-4 sera panel against homologous strains at different times post infection. NT are of high
magnitude within the first 2 years, with the exception of patient FLV080 at 16 months. FLV002
antisera at 21, 60 and 72 months show an overall decline in NT. Thus, NT responses are patient-
and virus-specific.
Stu
dy
no
(M
PI)

218
ii. Heterologous cross-neutralisation by anti-DENV-4 antisera
DENV-4 antisera sampled within the first year of infection (FLV068, 6; FLV049, 8; and
FLV099, 9 months) cross-neutralised a number of heterologous DENV in a patient-
dependent manner. FLV068 and FLV 099 differentially neutralized DENV-1, DENV-2
and DENV-3 at low magnitude (NT 21 – 36), only 1 of 1 of 5 viruses was neutralized
by all three patient antisera (Table 5.13). Patient FLV049 antisera neutralised 7 of 18
heterologous viruses; NT against DENV-2 strains were of greatest magnitude (GMT 13,
range 1-128). Between 1-2 years after infection most heterologous cross-neutralisation
had declined to undetectable levels, with the exception of FLV086 which neutralised 5
of 8 DENV-2 and 4 of 4 DENV-3. Five to 6 years after infection responses had declined
further – 3 antisera sampled from the same patient (FLV002) 24, 60, and 72 months
after infection neutralised 6, 2, and 1 virus(es) respectively, mostly against DENV-2 (4
viruses) at the earliest timepoint.
Thus, as for DENV-1, DENV-2 and DENV-3, anti-DENV-4 antisera neutralise
heterologous DENV in a patient- and virus-specific manner.
iii. Neutralisation against autologous serotype
No autologous virus was isolated from this serogroup.
iv. Neutralisation against related serogroups
The related flaviviruses ZIKV, YF17D and IMOJEV were not neutralised by any anti-
DENV-4 antisera; all NT were <20.

219
Study no. MPI Color
code FLV 068 6 FLV049 8/14 FLV099 9 FLV086 21 FLV080 16 FLV002 24/60/72
Figure 5. 27: -Heterologous strains neutralisation by anti-DENV-4 antisera
Mean neutralisation titre across DENV-4 sera panel against heterologous strains was <30 against all
strains. Interestingly, individual (FLV080) with low NT across homologous strains had higher GMT
against one of the heterologous strain of DENV-2. This individual was infected in Thailand and has
highest GMT against heterologous strain from Laos.

220
Figure 5. 28: - Heterologous neutralisation over time by anti-DENV-4 antisera.
NAb titre was <30 for all time point. No cross reactivity was observed even at 6 months post
infection. However, one of the patients at 16 MPI had NAb >100 against DENV-2 Laos
strain
Stu
dy
no
(M
PI)

221
5.4.2. Magnitude of neutralising antibody over time
Analysis of neutralising antibody responses of DENV-specific antisera against
homologous and heterologous DENV showed increasing specificity against homologous
serotype viruses over time. This pattern was seen in all the patients irrespective of
infecting serotype.
Comparison of neutralisation potency of antisera from individuals infected with any
DENV serotype was visualised using principle coordinate analysis (PCoA). The
antigenic distance between antisera is derived from differential neutralisation capacity
against all DENV strains. Each PCoA plot consists of distribution of sera over span of
six years post infection. Numbers in plot represent patient study code (eg: 11 is FLV
011). Serotype infecting individuals is represented as different coloured circle around
the patient number.
Analysis of individuals’ immune responses within a year of infection showed variation
among serotypes (Figure 5.29). DENV-1 and DENV-2 antisera clustered together;
individuals 11 & 12 were sampled within 1-week post onset of acute illness (OAI) and
showed the greatest degree of heterologous cross-neutralization. DENV-3 and DENV-4
anti-sera, collected 6 months - 1-year post-OAI, clustered within the homologous
serotype and were distant from heterologous DENV.
Twelve months after infection increased clustering of DENV-specific neutralising
responses was apparent (Figure 5.30). DENV-2 antisera consistently neutralized
homologous and heterologous DENV at high magnitude; whereas DENV-1 antisera
differentially neutralized homologous and heterologous DENV. DENV-3 antisera
neutralize homologous and heterologous DENV at a similar magnitude as DENV-4
antisera.

222
Three to 6 years after infection DENV antisera clustered by type (Figure 5.31). Antisera
from 2 individuals 5 and 6 years post-infection with DENV-1 or DENV-2 showed
similar positioning in the plot, showing that neutralization capacity remained constant.
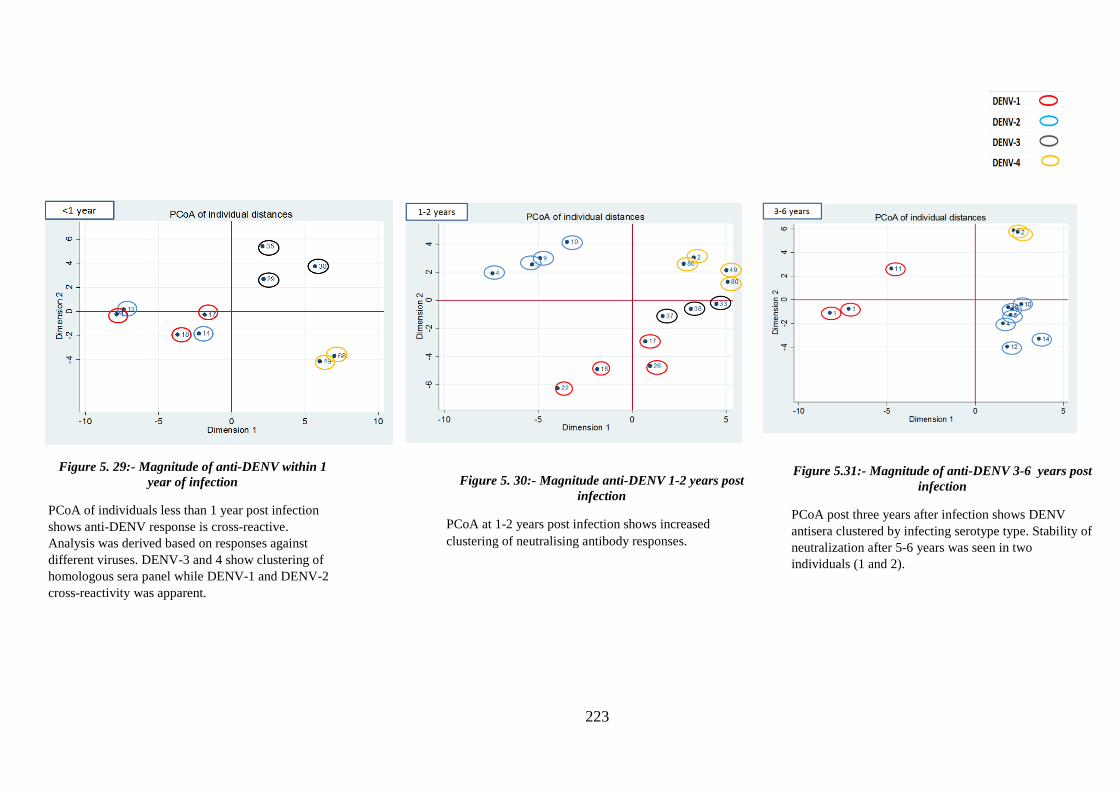
223
Figure 5.31:- Magnitude of anti-DENV 3-6 years post
infection
PCoA post three years after infection shows DENV
antisera clustered by infecting serotype type. Stability of
neutralization after 5-6 years was seen in two
individuals (1 and 2).
Figure 5. 30:- Magnitude anti-DENV 1-2 years post
infection
PCoA at 1-2 years post infection shows increased
clustering of neutralising antibody responses.
Figure 5. 29:- Magnitude of anti-DENV within 1
year of infection
PCoA of individuals less than 1 year post infection
shows anti-DENV response is cross-reactive.
Analysis was derived based on responses against
different viruses. DENV-3 and 4 show clustering of
homologous sera panel while DENV-1 and DENV-2
cross-reactivity was apparent.

224
5.4.3. Summary of neutralisation in DENV monotypic infection
Figure 5.32:- Summary of homologous and heterologous NAb responses

225
5.4.4. Total anti-DENV antibody:
Total anti-DENV antibody (TAb) responses in monotypic DENV-1-DENV-4 infection
were assessed to determine specificity and to serve as reference for parallel analysis of
neutralising antibody. Antibodies induced by DENV infection that are not neutralising
may bind to homologous and heterologous DENV.
Cross-recognition among DENV serotypes was assessed by haemagglutination
inhibition (HI) test. This test is based on the principle that sialic acid molecules on the
surface of red blood cells (RBC) bind to E glycoprotein on the DENV virion surface;
the agglutinated complex will form a lattice that appears as a smooth mat in a microtitre
plate well. Anti-DENV antibody that binds to the virion surface will inhibit
agglutination and RBC will drop to the bottom of the well, forming a red button. To
assess anti-DENV antibody titres, serum is serially diluted, and each dilution is added to
a constant amount of virus and RBC. The HI titre is the highest dilution which causes
total inhibition of agglutination.
Antisera from 34 individuals with monotypic DENV infection were tested against 25
strains of DENV. Not all the sample points of antisera tested in neutralisation were
tested for total antibody response. The closely related flavivirus Kunjin virus (KUNV)
was used as reference antigen to assess specificity.

226
a. Total antibody responses, DENV-1 infection
Twelve sera from 9 DENV-1-infected individuals sampled 9 days – 72 months post-
infection are included in the DENV-1 sera panel. Sera were tested against 6 homologous
DENV-1 strains and 18 heterologous DENV strains; data are expressed as GMT shown
in Table 5.17. Among homologous DENV-1, strain D1-HW2001G4 was recognised at
high magnitude compared to other DENV-1 strains isolated from travellers (Figure
5.33). Heat map analysis of HI titres showed that antisera from 2 individuals did not
recognise strains D1-14PHLG4 (belonging to the same genotype as D1-HW2001G4) or
D1-15LG1 (belonging to a different genotype, Genotype 1). An order of magnitude in
total antibody responses against homologous DENV-1 was apparent: D1-HW2001G4
>D1-10BG1L14>D1-10INDG3L4>D1-15LG1>D1-10BG2L4>D1-14PHLG4 that was
patient/antisera-specific.
Anti-DENV-1 antisera differentially recognised the majority of heterologous DENV
(Figure 5.34). Antisera from 4 individuals had <10 HI titre against heterologous strains.
Highest magnitude responses were directed against the non-virulent DENV-3 epidemic
reference virus D3-UNC3008NV and lowest responses were directed against D4-
14BG3L2. Five and 6 years after infection, TAb against selected strains – D2-
10LGA1L9, and the dominant Cosmopolitan strains D2-15BGCLN and D2-11BGCL4 –
were maintained at high magnitude (GMT:170) whereas TAb against related lineages -
D2-11BGCL isolated from a different traveller – were of lower magnitude (GMT:71).
Responses against the array of DENV-2, DENV-3 and DENV-4 strains varied
dependent on the virus, and the antisera, with mean GMT of 109, 155, 31 respectively.

227
Table 5. 15:- Total antibody response in DENV-1 sera panel
*= additional sample in HI test
Colour coding represents same patient at different time points
Values: 1 is <10 and 10240 is >10240
Antisera at all time points were not tested for TAb
FLV011/
9 daysFLV018/ 2 FLV017/2
*FLV076/
6
FLV022/
14
FLV026/
14
*FLV042
/16
*FLV085/
18
FLV018 /
20
FLV017/
20
FLV011/
34
FLV001/
60
FLV001 /
72
Flavi ref *KUNV 1280 320 160 160 40 40 40 40 80 40 40 80 80 89
D1-HW2001G4 10240 640 640 640 640 320 80 320 640 320 320 640 640 545
D1-10BG1L14 2560 320 160 160 320 80 40 80 120 640 80 160 160 184
D1-15LG1 640 80 40 80 80 40 240 40 40 1 20 80 80 53
D1-10BG2L4 10240 160 80 160 80 20 1 20 40 40 10 80 160 60
D1-10INDG3L4 640 160 40 120 160 20 20 40 80 80 40 160 160 83
D1-14PHLG4 640 160 80 40 80 20 1 20 20 10 20 40 40 34
GMT-D1 2032 202 101 136 160 45 16 50 76 43 40 127 143 98
D2-NGCGAII 10240 480 480 160 160 40 1 40 80 240 20 80 120 110
D2-10LGA1L9 5120 320 320 160 160 80 40 80 160 80 160 160 320 188
D2-11BGCL4 2560 320 160 80 160 40 1 20 40 80 40 80 80 71
*D2-11BGCL4 2560 640 320 160 160 80 20 80 80 160 80 240 240 170
D2-13MGCLN 1280 320 160 80 80 40 1 80 40 80 40 80 80 71
D2-15BGCL4 2560 160 160 40 160 160 20 20 40 160 40 160 160 104
D2-15BGCLN 640 640 160 40 80 40 1 20 80 80 80 160 320 79
*D2-15TLS/IDNGCLG 2560 640 320 160 160 80 80 80 80 160 80 320 20 160
D2-PR6913-V 1280 160 80 20 40 20 20 40 80 80 40 80 160 68
D2-PR1940-NV 1280 320 320 80 320 80 320 20 80 160 40 160 160 152
GMT-D2 2229 357 220 80 130 57 10 40 70 118 53 135 132 109
D3-H87G5 10240 320 320 80 320 160 640 160 40 320 160 160 160 259
D3-15BG1 2560 320 160 80 160 40 20 40 160 80 40 80 160 110
*D3-15BG1 320 240 160 80 160 40 20 40 160 80 30 80 160 90
D3-UNC3002-V 2560 320 320 80 80 40 40 40 320 80 40 80 160 123
D3-UNC3008-NV 2560 640 640 160 80 80 640 40 640 640 80 320 320 288
GMT-D3 2229 347 279 92 139 61 92 53 184 160 57 121 184 155
D4-H241G1 1280 160 320 80 160 160 1 40 20 160 40 80 80 79
D4-10BG3L2 1280 40 40 20 1 80 20 20 20 1 20 40 40 24
*D4-14BG3L2 960 40 20 20 1 40 1 1 20 20 20 20 40 16
GMT-D4 1163 63 63 32 5 80 3 9 20 15 25 40 50 31
GMT
Homologous
serotype
Heterologous
serotype
Virus
Study No/MPI
<1 year 1- 2 year 3- 6 yearsGeometric Mean Titer (GMT)
GMT-D3 strains
acutes: 599
(160-10240)
GMT-D3 strains
convalescent:
104 (20-640)
GMT-D3 all
strains:155
(20-10240)
GMT-
heterologous
serotype= 80
(<10-10240)
GMT-D2 strains
acutes: 559
(80-10240)
GMT-D2 strains
convalescent: 67
(<10-320)
GMT-D4 strains
acutes: 167
(40-1280)
GMT-D4 strains
convalescent: 17
(<10-160)
GMT-D4 all
strains:31
(10-1280)
GMT-D1 strains
acutes: 346
(40-10240)
GMT-D1 strains
convalescent:67
(<10-640)
GMT-homologous serotype=
98(<10-10240)
GMT-D2 all
strains:
109 (<10-
10240)
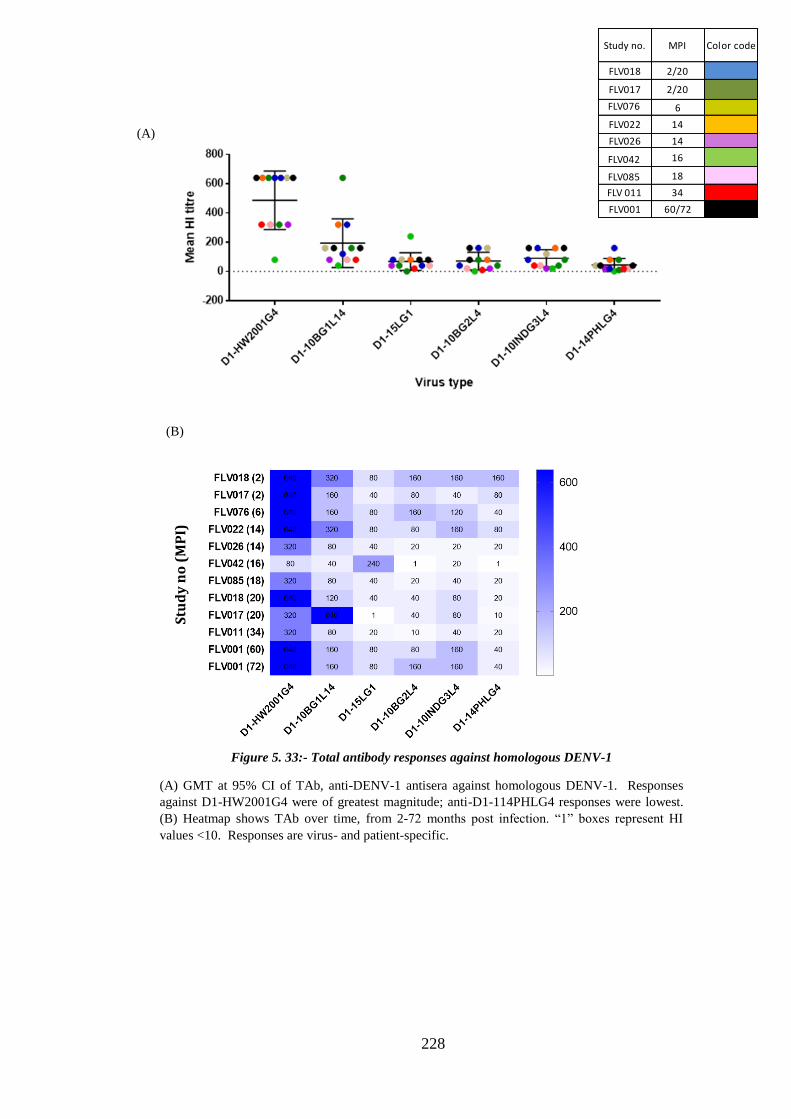
228
Figure 5. 33:- Total antibody responses against homologous DENV-1
(A) GMT at 95% CI of TAb, anti-DENV-1 antisera against homologous DENV-1. Responses
against D1-HW2001G4 were of greatest magnitude; anti-D1-114PHLG4 responses were lowest.
(B) Heatmap shows TAb over time, from 2-72 months post infection. “1” boxes represent HI
values <10. Responses are virus- and patient-specific.
(A)
(B)
Stu
dy
no
(M
PI)
Study no. MPI Color code
FLV018 2/20
FLV017 2/20
FLV076 6
FLV022 14
FLV026 14
FLV042 16
FLV085 18
FLV 011 34
FLV001 60/72

229
(B)
Figure 5. 34: - Total antibody responses, anti-DENV-1 antisera against heterologous DENV
(A) GMT at 95% CI of TAb, anti-DENV-1 antisera against heterologous DENV. Highest magnitude
responses were directed against the non-virulent DENV-3 epidemic reference virus D3-UNC3008NV
and lowest responses were directed against D4-14B. (B) Heatmap shows TAb over time, 2 – 72 months
post infection. Empty boxes represent values <10. Differential cross-recognition of heterologous DENV
is apparent 5 and 6 years after infection, with TAb titres ranging from 20 – 320. Responses are virus-
and patient-specific.
(A)
Mo
nths
post
infe
ctio
n
(A)
Study no. MPI Color code
FLV018 2/20
FLV017 2/20
FLV076 6
FLV022 14
FLV026 14
FLV042 16
FLV085 18
FLV 011 34
FLV001 60/72
Stu
dy
no
(M
PI)

230
b. Total antibody responses, DENV-2 infection
Antisera from 10 individuals infected with DENV-2 sampled five days to 51 months
post infection were assessed for total anti-DENV against 10 homologous DENV-2 and
14 heterologous DENV-1, DENV-3 and DENV-4 strains . Differential responses against
homologous DENV-2 were measured (Figure 5.35). Highest magnitude responses were
directed against the epidemic non-virulent strain D2-PR1940NV, contrasting with low
magnitude TAb against the epidemic virulent strain D2-PR6913V. Responses against
traveller field isolates varied; most (5 of 6) belonged to the Cosmopolitan Genotype, of
which 2 lineages were represented, and interestingly differential TAb magnitude against
these viruses was not clearly associated with virus lineage but was strain-dependent:
D2-11BGCL4 induced high magnitude (80-640, 204) responses whereas D2-10LGAIL9
induced low magnitude (20-160, 142) across all antisera.
All homologous DENV-2 were recognised by all antisera, albeit at levels (20) close to
the assay cut-off for D2-15BGCLN 51 months after infection. Mean TAb titres against
all homologous DENV are shown in Figure 35 (A). A heatmap analysis showing TAb
titres over time is shown in Figure 35 (B). Differential responses, that are virus- and
patient- specific, occur over time, and at the latest timepoint (51 months) a clear
distinction among virus-specific responses is apparent with high (80-160) and low (20)
magnitude homologous TAb against 4 of 10 and 3 of 10 viruses, respectively. TAb
titres for DENV-2 sera panel are shown in Table 5.18.
Responses amongst 9 of 10 individuals (excluding individual at 5 days PI) against
heterologous DENV-1, DENV-3 and DENV-4 viruses were of low magnitude with
overall GMT-17; GMT against individual viruses ranged from <10-320. Heatmap
analysis shows that most antisera produced low or negative TAb titres against

231
heterologous DENV, particularly DENV-1 (Figure 36 (B). Responses were clearly
virus-and patient-specific, but nonetheless had declined to low magnitude TAb titres
(<10 for 12 of 30 viruses; GMT 12 and 5) at 51 months post infection.

232
Table 5. 18:- Total antibody response DENV-2 sera panel
*= additional sample in HI test
Colour coding represents same patient at different time points
Values: 1 is <10 and 10240 is >10240
Not all sample time points tested for total antibody
FLV012 /
5 days
FLV014 /
7
*FLV070/
7
*FLV074/
13
*FLV072/
15
*FLV073/
15
FLV004 /
16
FLV005 /
16
FLV009 /
17
FLV010 /
19
FLV012 /
33
FLV009
/36
FLV010
/36
FLV014
/36
FLV010
/48
FLV004
/51
FLV005
/51
Flavi ref *KUNV 1280 80 30 160 30 160 20 20 40 20 40 40 20 80 20 20 20 46
D2-NGCGAII 10240 160 320 40 80 40 160 160 160 80 320 160 80 80 80 80 40 136
D2-10LGA1L9 5120 320 160 80 160 160 80 80 160 80 160 80 80 160 80 80 80 142
D2-11BGCL4 2560 160 40 40 80 40 40 40 40 20 80 40 20 80 20 40 20 53
*D2-11BGCL4 2560 640 160 160 320 80 160 160 320 80 320 160 80 640 80 160 80 204
D2-13MGCLN 1280 640 80 80 80 40 160 80 160 80 320 80 40 640 80 160 40 130
D2-15BGCL4 1280 160 80 40 10 10 40 40 80 20 160 80 20 160 20 80 40 55
D2-15BGCLN 1280 160 40 20 40 40 40 40 40 20 80 40 20 80 20 40 20 47
*D2-15TLS/IDNGLG 2560 320 160 160 480 160 160 80 160 40 320 40 40 320 80 160 160 164
D2-PR6913-V 1280 80 20 80 20 20 20 40 40 20 40 40 20 40 20 40 20 38
D2-PR1940-NV 2560 320 320 80 320 320 80 160 160 80 80 160 80 320 160 160 80 181
GMT-D2 2389 243 98 65 89 57 75 75 106 43 149 75 40 171 49 86 46 98
D1-HW2001G4 10240 160 160 40 40 40 40 40 80 40 80 80 40 80 40 80 40 80
D1-10BG1L14 10240 80 80 30 80 1 20 20 40 20 40 20 1 40 1 20 40 26
D1-15LG1 320 40 1 1 1 1 1 1 40 1 20 20 1 20 1 1 1 4
D1-10BG2L4 2560 40 120 1 1 40 1 1 20 1 20 20 1 15 1 1 1 6
D1-10INDG3L4 1280 20 120 1 1 1 1 1 20 1 1 1 1 1 1 1 1 3
D1-14PHLG4 320 40 1 1 1 1 1 1 20 1 20 20 1 20 1 1 1 4
GMT-D1 1810 50 24 3 4 3 3 3 32 3 17 15 2 16 2 3 3 9
D3-H87G5 10240 160 80 320 80 320 80 80 160 40 160 160 20 40 40 80 1 93
D3-15BG1 2560 80 20 20 20 80 20 20 40 20 40 40 1 40 1 20 20 26
*D3-15BG1 320 60 20 20 20 80 20 20 40 20 40 40 1 40 1 20 20 23
D3-UNC3002-V 5120 320 20 40 40 20 20 20 80 20 40 20 20 40 1 40 20 36
D3-UNC3008-NV 10240 160 20 40 80 20 20 20 40 1 80 20 1 80 1 40 20 28
GMT-D3 3378 132 26 46 40 61 26 26 61 13 61 40 3 46 2 35 11 35
D4-H241G1 640 40 40 80 40 40 40 80 80 30 40 40 1 40 1 80 30 35
D4-10BG3L2 640 20 40 20 1 1 20 40 40 20 20 40 1 20 1 40 1 12
*D4-14BG3L2 320 20 20 1 1 1 1 20 1 1 20 1 1 1 1 1 1 3
GMT-D4 508 25 32 12 3 3 9 40 15 8 25 12 1 9 1 15 3 11
Heterologous
serotype
Homologous
serotype
Virus
Study No./ MPI
< 1 year 1-2 year 2-3 years 4-5 years
Geometric Mean Titer (GMT)GMT
GMT-D4
strains for
acute sera :
508 (320-
640)
GMT-D4 strains
for convalescent
sera : 8 (<10-80)
GMT-D4
strains
total: 11
(<10-640)
GMT- total sera
heterologous
serotype= 15 (<10-
10240)
GMT-D3
strains for
acute sera :
3378 (320-
10240)
GMT-D3 strains
for convalescent
sera :27 (<10-320)
GMT-D3
strains
total : 35
(<10-
10240)
GMT-D1
strains for
acute sera :
1810 (320-
10240)
GMT-D1 strains
for convalescent
sera : 6 (<10-160)
GMT-D1
strains
total : 9
(<10-
10240)
GMT-D2
strains for
acute sera:
2389 (1280-
10240)
GMT-D2
convalescent
sera: 88 (10-640)
GMT- total sera homologous
serotype= 106 (10-10240)

233
Figure 5. 35:- Total antibody responses, anti-DENV-2 antisera against homologous DENV-2
(A) GMT at 95% CI of TAb against homologous DENV-2. TAb titres against 3 of 6 Cosmopolitan
genotype viruses were of greater magnitude compared to other closely-related viruses that belonged to the
same lineage. Highest magnitude responses were directed against the epidemic non-virulent reference
virus D2-PR1940NV, contrasting with low magnitude TAb against the epidemic virulent D2-PR6913V.
(B) Heatmap shows TAb from 7 – 51 months post infection. Differential responses, that are virus- and
patient- specific, occur over time, and at 51 months TAb titres against 6 0f 10 viruses have declined to
low levels.
(A)
(B)
Stu
dy
no
(M
PI)
Study no. MPI Color code
FLV014 7/36
FLV070 7
FLV074 18
FLV072 15
FLV073 15
FLV004 16/51
FLV005 16/51
FLV009 17/36
FLV010 19/36/48
FLV 012 33

234
A)
Figure 5. 36:- Total antibody responses, anti-DENV-2 antisera against heterologous DENV.
(A) GMT at 95% CI of TAb against heterologous DENV-1, DENV-3 and DENV-4. Overall, TAb titres
against DENV-3 were greater than for DENV-1 and the small number of DENV-4 assessed here. (B)
Heatmap shows TAb titres over time, from 7 - 51 months post infection. Responses are clearly patient-
and virus-specific, and anti-DENV-2 cross-recognition of heterologous viruses is below limits of detection
of the HI assay in 30% of tests. Cross-recognition of the closely related flavivirus KUNV occurs for all
antisera in a patient-specific manner, with TAb titres ranging from 20 – 160.
B)
(B)
(A)
Stu
dy
no
(M
PI)
Study no. MPI Color code
FLV014 7/36
FLV070 7
FLV074 18
FLV072 15
FLV073 15
FLV004 16/51
FLV005 16/51
FLV009 17/36
FLV010 19/36/48
FLV 012 33

235
c. Total antibody response post DENV-3 infection
Antisera from 10 individuals sampled 8 - 19 months post infection were tested against 5
homologous DENV-3 and 19 heterologous DENV-1, DENV-2 and DENV-4 viruses.
Homologous DENV-3 viruses included D3-10BG1 and D3-15BG1, which represented
the same lineage within Genotype 1, the virus imported by all DENV-3-infected
travellers during the study period 2010-2015. Two viruses, isolated from travellers
infected in 2015 and originating in Bali, Indonesia were selected. Anti-DENV-3 antisera
recognised these traveller strains with similar mean TAb titres (39, 40). TAb titres
against the epidemic virulent and non-virulent viruses D3-UNC3002V and D3-
UNC3008NV were similar. Highest magnitude responses were directed against the
prototype reference virus D3-H87G5 (Figure 5.37 (A)). Heatmap analysis of
longitudinal homologous responses showed patient-specific variation in recognition of
traveller viruses which all belonged to the same lineage (Figure 5.38 (B)). TAb titres are
shown in Table 5.19.

236
*= additional sample in HI test
Colour coding represents same patient at different time points
Values: 1 is <10 and 10240 is >10240
Not all sample time points tested for total antibody
FLV035
/ 8
*FLV044/
8
*FLV045/
8
FLV029
/12
FLV030 /
12
*FLV047/
12
FLV033
/14
FLV037
/14
FLV038
/14
*FLV046/
18
Flavi ref *KUNV 40 160 40 40 10 60 10 20 20 40 32
D3-H87G5 640 320 640 640 160 80 40 640 160 640 279
D3-15BG1 40 160 40 160 40 40 20 40 10 20 40
*D3-15BG1 40 120 40 160 40 40 20 40 10 20 39
D3-UNC3002-V 160 160 80 160 20 160 20 80 40 160 80
D3-UNC3008-NV 160 160 40 160 20 160 20 80 40 160 75
GMT-D3 121 174 80 211 40 80 23 92 30 92 76
D1-HW2001G4 160 20 10 80 80 20 320 160 20 80 57
D1-10BG1L14 160 15 1 40 40 15 240 80 20 120 35
D1-15LG1 80 1 1 20 10 40 1 80 1 40 9
D1-10BG2L4 40 20 20 20 20 10 40 20 20 40 23
D1-10INDG3L4 40 10 1 20 40 10 40 20 1 20 12
D1-14PHLG4 10 1 1 20 20 40 80 40 1 40 11
GMT-D1 57 6 2 28 28 19 46 50 4 48 19
D2-NGCGAII 160 20 10 40 40 10 320 40 20 40 37
D2-10LGA1L9 160 40 20 160 160 40 160 80 160 160 92
D2-11BGCL4 160 20 20 80 40 10 320 40 20 40 43
*D2-11BGCL4 160 20 20 80 80 20 320 40 40 80 57
D2-13MGCLN 80 10 1 40 40 20 80 20 20 40 22
D2-15BGCL4 160 20 20 40 40 20 160 40 80 160 53
D2-15BGCLN 80 20 40 20 20 1 80 20 20 40 22
*D2-15TLS/IDNGCLG 320 40 40 160 160 80 160 80 80 160 106
D2-PR6913-V 80 40 20 20 40 10 80 20 20 20 28
D2-PR1940-NV 160 40 40 80 80 80 80 160 40 40 70
GMT-D2 139 25 17 57 57 17 149 43 37 61 46
D4-H241G1 160 40 1 1 160 40 80 80 80 80 33
D4-10BG3L2 20 20 20 1 1 1 20 1 1 80 5
*D4-14BG3L2 20 20 1 1 1 1 1 20 1 20 3
GMT-D4 40 25 3 1 5 3 12 12 4 50 8
GMT Geometric Mean Titre
GMT-D1 strains
:19 (<10-320)
GMT-D2 strains:
46 (<10-320)
GMT-D4 strains: 8
(<10-160)
GMT- total sera homologous
serotype=76 (10-640)
GMT-heterologous
serotype= 19
Heterologous
serotype
VirusStudy No./ MPI
<1 year 1-2 year
Homologous
serotype
Table 5. 19:- Total antibody response of DENV-3 sera panel

237
(A)
(B)
Figure 5. 37: - Total DENV-3 antibody response against homologous DENV
(A) GMT at 95% CI of TAb anti-DENV-3 antisera against homologous virus. TAb against
traveller viruses, which both belong to Genotype 1 and to the same lineage, were of
equivalent magnitude and were lower than against the reference virus D3-H87, which
belongs to Genotype 5. TAb against the epidemic virulent and non-virulent viruses D3-
UNC3002V and D3-UNC3008NV, were similar and were of higher magnitude than against the
traveller viruses. (B) Heatmap shows TAb over time, 8 - 91 months post infection. Responses
were virus- and patient-specific, and at 19 months post infection ranged from 20 – 640.
Stu
dy
no
(M
PI)
Study no. MPIColor
code
FLV 035 8
FLV044 8
FLV045 8
FLV029 12
FLV030 12
FLV047 12
FLV033 14
FLV037 14
FLV038 14
FLV046 18

238
(B)
(A)
Figure 5. 38:- Total antibody responses, anti-DENV-3 antisera against heterologous DENV
(A) GMT at 95% CI of TAb against heterologous DENV. Responses were virus- and patient-specific,
even among closely-related viruses. DENV-1 viruses belonged to 4 different genotypes; GMT-TAb
against the two Genotype 1 viruses were distinct (35 and 9) as were GMT-TAb against the 2 Genotype 4
viruses (57 and 11). Similarly, among DENV-2, 7 of 8 belonged to Cosmopolitan genotype yet TAb
responses varied widely. (B) Heatmap analysis shows change in TAb over time, 8 - 18 months post
infection. Anti-DENV-3 antisera recognised all heterologous DENV at 18 months post infection, with
TAb ranging from 20 – 160.
Study no. MPIColor
code
FLV 035 8
FLV044 8
FLV045 8
FLV029 12
FLV030 12
FLV047 12
FLV033 14
FLV037 14
FLV038 14
FLV046 18
Stu
dy
no
(M
PI)

239
d. Total antibody responses post DENV-4 infection
Anti-DENV-4 antisera from 6 individuals sampled 6 – 72 months post infection were
tested against 3 homologous DENV-4 and 21 heterologous DENV-1, DENV-2, and
DENV-3. TAb titres against all DENV are listed in Table 5.20.
Two traveller DENV-4 viruses which belong to Genotype 3, sampled in 2010 and 2014
and originating in Bali, Indonesia, and the prototype reference virus DENV-4 H241
were assessed. In the period of study DENV-4 was rarely identified in travellers
returning form the Asia Pacific region and thus is not as well represented in this cohort
as the other 3 serotypes. TAb against traveller viruses were of lower magnitude than
TAb against the reference virus, which belonged to Genotype 1. Mean TAb is shown in
Figure 5.39 (A). Heatmap analysis shows TAb over time, 6 -60 months post infection.
Responses were patient- and virus-specific: FLV-002 TAb titres at 60 months were of
greater magnitude than antisera from the other 5 individuals, sampled within the first 2
years of infection (Figure 5.39 (B) )
Cross-recognition of heterologous DENV was of low magnitude (Figure 5.40) and was
virus-and patient-specific. Responses were of greater magnitude, overall, against
DENV-2 (5) and had declined to undetectable levels in most viruses 60 months after
infection although 4 of 21 viruses, representing 3 heterologous serotypes, were still
recognised (TAb range <10-160).
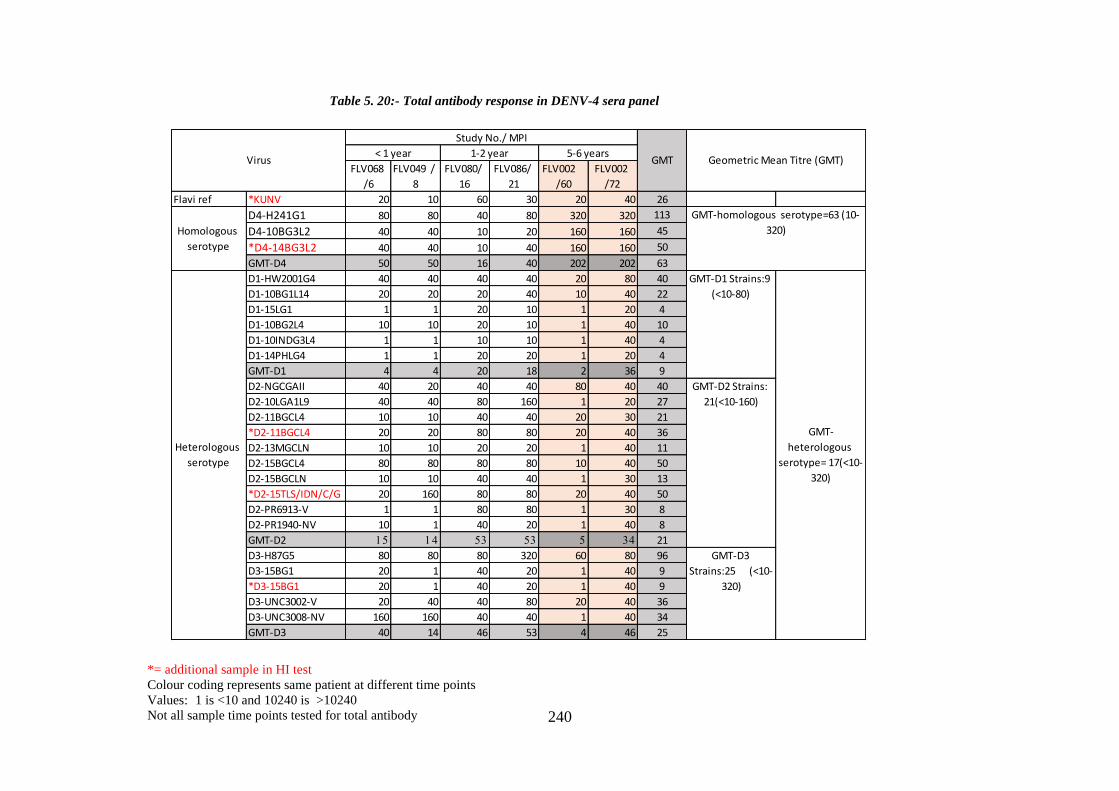
240
FLV068
/6
FLV049 /
8
FLV080/
16
FLV086/
21
FLV002
/60
FLV002
/72
Flavi ref *KUNV 20 10 60 30 20 40 26
D4-H241G1 80 80 40 80 320 320 113
D4-10BG3L2 40 40 10 20 160 160 45
*D4-14BG3L2 40 40 10 40 160 160 50
GMT-D4 50 50 16 40 202 202 63
D1-HW2001G4 40 40 40 40 20 80 40
D1-10BG1L14 20 20 20 40 10 40 22
D1-15LG1 1 1 20 10 1 20 4
D1-10BG2L4 10 10 20 10 1 40 10
D1-10INDG3L4 1 1 10 10 1 40 4
D1-14PHLG4 1 1 20 20 1 20 4
GMT-D1 4 4 20 18 2 36 9
D2-NGCGAII 40 20 40 40 80 40 40
D2-10LGA1L9 40 40 80 160 1 20 27
D2-11BGCL4 10 10 40 40 20 30 21
*D2-11BGCL4 20 20 80 80 20 40 36
D2-13MGCLN 10 10 20 20 1 40 11
D2-15BGCL4 80 80 80 80 10 40 50
D2-15BGCLN 10 10 40 40 1 30 13
*D2-15TLS/IDN/C/G 20 160 80 80 20 40 50
D2-PR6913-V 1 1 80 80 1 30 8
D2-PR1940-NV 10 1 40 20 1 40 8
GMT-D2 15 14 53 53 5 34 21
D3-H87G5 80 80 80 320 60 80 96
D3-15BG1 20 1 40 20 1 40 9
*D3-15BG1 20 1 40 20 1 40 9
D3-UNC3002-V 20 40 40 80 20 40 36
D3-UNC3008-NV 160 160 40 40 1 40 34
GMT-D3 40 14 46 53 4 46 25
GMT
GMT-
heterologous
serotype= 17(<10-
320)
GMT-D1 Strains:9
(<10-80)
GMT-D2 Strains:
21(<10-160)
GMT-D3
Strains:25 (<10-
320)
GMT-homologous serotype=63 (10-
320)
Geometric Mean Titre (GMT)
Heterologous
serotype
Homologous
serotype
Virus
Study No./ MPI
< 1 year 1-2 year 5-6 years
Table 5. 20:- Total antibody response in DENV-4 sera panel
*= additional sample in HI test
Colour coding represents same patient at different time points
Values: 1 is <10 and 10240 is >10240
Not all sample time points tested for total antibody
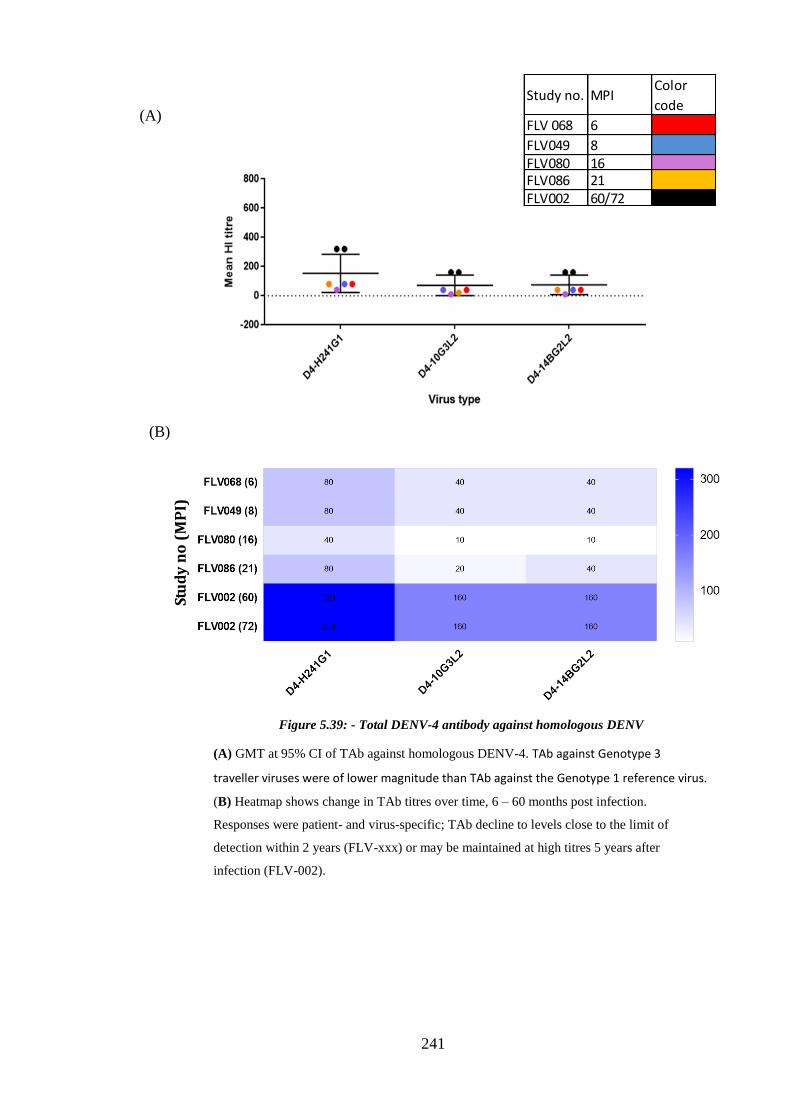
241
(A)
(B)
Figure 5.39: - Total DENV-4 antibody against homologous DENV
(A) GMT at 95% CI of TAb against homologous DENV-4. TAb against Genotype 3
traveller viruses were of lower magnitude than TAb against the Genotype 1 reference virus.
(B) Heatmap shows change in TAb titres over time, 6 – 60 months post infection.
Responses were patient- and virus-specific; TAb decline to levels close to the limit of
detection within 2 years (FLV-xxx) or may be maintained at high titres 5 years after
infection (FLV-002).
Stu
dy
no
(M
PI)
Study no. MPIColor
code
FLV 068 6
FLV049 8FLV080 16FLV086 21FLV002 60/72

242
Figure 5.40: - Total antibody responses, anti-DENV-4 antisera against heterologous DENV
(A) GMT at 95% CI of TAb against heterologous DENV-1, DENV-2, and DENV-3. Anti-DENV-4
antisera recognized heterologous DENV at low magnitude. (B) Heatmap analysis shows TAb over
time, 6 -60 months post infection. Responses were patient- and virus-specific: FLV-002 TAb titres at
60 months were of greater magnitude than antisera from the other 5 individuals, sampled within
the first 2 years of infection. TAb declined to undetectable levels in most viruses 60 months after
infection.
(B)
(A)
Stu
dy
no
(M
PI)
Study no. MPIColor
code
FLV 068 6
FLV049 8FLV080 16FLV086 21FLV002 60/72

243
5.4.5. Summary of total antibody in DENV monotypic infection
Figure 5. 41: - Summary of homologous and heterologous anti-DENV TAb

244
5.4.6. Longitudinal variation of neutralising and recognising anti-DENV antibody
Specificity and cross-reactivity of neutralising and total anti-DENV-1-4 antisera varied
over time. Acute phase (<2 months post infection) sera were highly cross-reactive
irrespective of infecting serotype. Increase in neutralisation specificity towards infecting
serotype was seen for antisera collected 1-2 years after infection although cross-
reactivity against heterologous DENV was still prevalent. By 2 years post infection
neutralisation was highly specific towards infecting serotype. Broad cross-recognition
of all four serotypes is still apparent after 2 years and after 4 years responses were
largely but not exclusively restricted to homologous serotype.
These longitudinal data was used to map antigen and antibody specificity in an
antigenic map as previously described for other viruses (323). DENV sera panels were
differentiated into five groups based on the time post infection. These maps represent
corresponding positioning of virus and antisera based on neutralisation and cross
recognition. Briefly, a target distance from a serum to each virus is derived by
calculating the difference between the logarithm (log2) reciprocal antibody titre for that
particular virus and log2 reciprocal maximum titre achieved by that serum (against any
virus). A 2-fold change in titre will equate to a single grid in a graph which is denoted
as Antigenic Unit (AU); antigens and the antisera that bind or neutralise them cluster
together, at a distance that is proportional to the magnitude of the response.
Neutralising and total antibody titre changed for each isolate at different time points of
infection. These viruses were derived from the same batch of titrated stock and used
throughout. Thus, variation in antibody responses can be attributed to differential
epitope recognition. Antigenic maps below illustrate change in specificity of
neutralising and cross-recognising antibody responses with the progression of time post
infection. These maps also show the variation of antigenicity among the DENV isolates.

245
These data show that homologous and heterologous isolates are not equally recognised
or neutralised by anti-DENV antisera.
a. Cross-reactive anti-DENV responses within 2 months post infection
Antibody repertoire activated against DENV within two months of infection does not
recognise antigenic diversity among DENV isolates. Response were highly cross-
reactive irrespective of infecting serotype. Both neutralising and total antibody showed
high antibody titres against all tested isolates.
Four plasma samples collected within 2 months of infection were assessed. The
antigenic map shows a high degree of cross-reactivity (Figure 5.42). Neutralisation
against all DENV strains was at the maximum detection limit of the FRNT (>320) and
antigenic distance could not be differentiated; this is represented as a straight line in the
map. A wider range of titres was seen for HI tests, ranging from 320-10240, and
antigenic distances were more dispersed. However, in both cases antigenic diversity
among homologous and heterologous strains could not be identified; no clustering was
observed.

246
A
Figure 5. 42:- Anti-DENV response and antigenic diversity at less than two months post infection
Maps were constructed based on the neutralising and total antibody titre of 4 travellers with in week to 2 months post infection. All the filled shapes represent virus and
clear square boxes are the plasma sample
Figure A) represents neutralisation and figure B) represents cross recognition. Both antibody responses are highly cross-reactive at this time point as shown by close
positioning of the serum in map and no clear distinction among serotypes was observed.
Total antibody
a
Neutralisation B
A

247
b. Anti-DENV antibody responses 2-12 months post infection
Antisera collected between 2 and 12 months after infection displayed greater specific
neutralisation for homologous serotype, compared to earlier acute phase antisera.
However, cross-neutralisation of heterologous serotypes still occurred.
Four plasma samples collected between 2 and 12 months after infection were tested for
NAb and 12 samples for total antibody. In the antigenic map based on the NAb titre
(Figure 43(A)) increased homologous virus neutralisation compared to acute phase
responses was seen and clusters could be distinguished. However, a number of DENV-
1, DENV-2, and DENV-3 isolates and antisera continued to cluster with heterologous
DENV. DENV-4 isolates form a distinct cluster, unlike the other 3 serotypes.
Total anti-DENV antibody responses continued to be highly cross-reactive across all
four DENV serotypes. No antigenic clustering was apparent.

248
A
A
B
A
Figure 5. 43:- Anti-DENV antibody response with a year of infection
Serotype specific cross-neutralisation increased with separation of four different serotypes as shown in neutralisation map. However, DENV-1 and DENV-2
isolates and antisera still showed close proximity to heterologous DENV. Total antibody cross-recognition was still apparent, and no clustering was apparent.
Neutralisation Total antibody
a

249
c. Anti-DENV antibody responses 1-2 years post infection
Neutralising antibody responses 1 year after infection were highly specific towards
infecting serotype. Neutralisation mapping showed clustering of homologous antisera
with infecting serotype indicating the antibody repertoire has increased specificity
compared to earlier time points. Total antibody responses also increased in specificity.
Antigenic maps were derived from 18 antisera. Four distinct clusters were observed,
however there were outliers for each serotype. D1-10BG2L14 was antigenically closer
to D2-10LGA1L9 and distant to the homologous cluster. Furthermore, DENV-2
Cosmopolitan genotype isolates do not form a distinct cluster but instead form 2 groups,
some of which are closer to DENV-1 than to the homologous DENV-2. In addition, 2
isolates of DENV-1 which both belong to Genotype 1 and isolated in different years
(2010 and 2015) and distant from each other; the 2015 virus is antigenically closer to
Genotype 4. In summary, although neutralising antibody responses become more
specific 1-2 years after infection, heterologous cross-neutralisation still occurs.
Total antibody titres derived from 24 antisera were used to construct an antigenic map
(Figure 44(B). Less antigenic overlapping among heterologous isolates was observed
compared to previous time point but there was no clustering was observed in
neutralisation maps. Total antibody mapping showed that DENV-1 and DENV-2
formed clusters with their respective antisera, unlike DENV-3 and DENV-4.

250
Figure 5. 44:- Specificity of anti-DENV antibody 1-2-year post infection
Anti-sera collected 1 year after infection showed increased neutralisation specificity towards the infecting serotype. Four distinct clusters were observed,
however there were outliers for each serotype. Total antibody was highly cross-reactive.
A
A
B
A
Neutralisation Total antibody
a

251
d. Anti-DENV antibody responses 2-4 years post infection
Neutralising and total antibody responses two year after infection were highly specific
towards infecting serotype. All antisera clustered with homologous isolates.
Five antisera (4 DENV-2 and 1 DENV-1) were available for testing at this time point.
Distinct clustering was observed for both neutralisation and total antibody maps. The
random positioning of DENV-3 and DENV-4 is due to lack of anti-DENV-3 antisera.
(Figure 5. 45 A and B).
e. Anti-DENV antibody responses 4 years after infection
Six antisera (2 each of DENV-1, DENV-2 and DENV-4) were available (Figure 5.46).
DENV-1, DENV-2 and DENV-4 formed distinct clusters in both maps. In both maps,
DENV-1 and DENV-2 clusters are closer to each other than they are to DENV-4, which
is the most distant. The random positioning of DENV-3 is due to lack of respective
antisera.
A
A

252
Total antibody B
A
Figure 5. 45:- Anti-DENV-4 antibody at 2-4 years post infection
After 2 years anti-sera are specific to infecting serotype hence in antigenic map clustered towards infecting serotypes shown by both cross-
neutralisation and cross-recognition antibody tests. At this time point, there was no DENV-3 anti-sera available. All the positioning of DENV-3
strains are based on antibody titre derived from DENV-1/2/4 antisera thus DENV-3 strains are at random positioning along with DENV-1 and 2.
A
A
Total antibody
A
A
Neutralisation
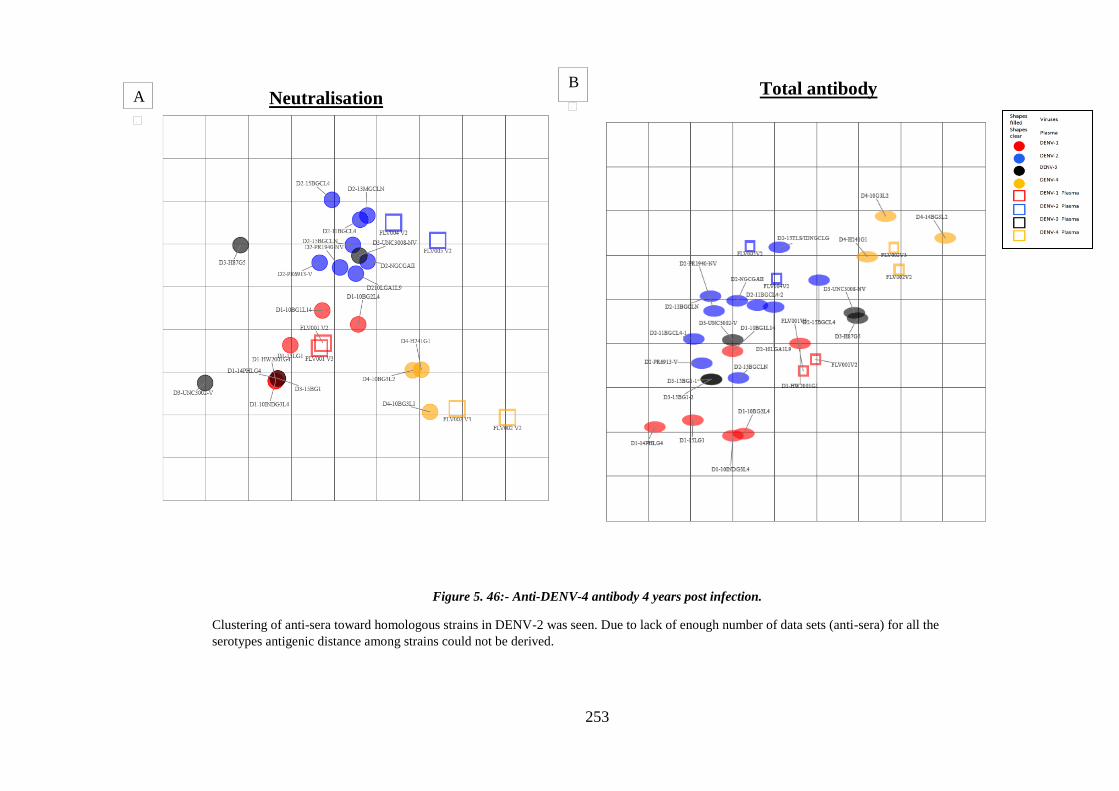
253
B
A
Neutralisation
Total antibody
Figure 5. 46:- Anti-DENV-4 antibody 4 years post infection.
Clustering of anti-sera toward homologous strains in DENV-2 was seen. Due to lack of enough number of data sets (anti-sera) for all the
serotypes antigenic distance among strains could not be derived.
A
A
B
A
A
A
Neutralisation Total antibody

254
5.5. Summary discussion
5.5.1. Neutralisation endpoint stringency and increased specificity
In this traveller cohort with well-defined monotypic DENV infection, neutralisation
titres obtained at 50% reduction of infection do not exclude heterologous serotype
cross-neutralisation. Increasing stringency to 75% and 90% reduction increased
specificity to homologous virus however there was a corresponding decrease in
sensitivity and false-negative tests were noted. Heterologous cross-reactivity decreased
by 85% for anti-DENV-1 antisera and by 70% for anti-DENV-2 antisera at 75%
reduction. Applying 90% reduction decreased cross-neutralisation across DENV-1 and
DENV-2 by 92% and 80% respectively (Table 5.1-5.6). DENV-3 and DENV-4
heterologous cross-reactivity was of low magnitude compared to DENV-1 and DENV-
2, even when 50% reduction was applied. For these reasons the 50% reduction endpoint
was applied in analysis of anti-DENV responses.
Increasing neutralisation stringency to 90% reduction differentiates the DENV and
ZIKV sero complexes in monotypic DENV infection. However, this distinction is only
relevant to ZIKV MR766; no traveller antisera neutralised ZIKV PRVABC59. Cross-
neutralisation of ZIKVMR766 at 50% reduction was serotype-specific: the majority of
anti-DENV-1 and anti-DENV-2 antisera, from acute phase up to 72 months neutralized
this ZIKV strain; no anti-DENV-3 or anti-DENV-4 antisera (up to 72 months)
neutralised this virus. The reasons for the high magnitude neutralising antibody
responses directed against the archival ZIKV MR766, but not the contemporaneous
epidemic virus ZIKV PRVABC59, are unclear but may be associated with evolution of
a distinct viral genotype and phenotype with multiple passage through a range of
animals and cell lines.

255
5.5.2. Anti-DENV antibody responses are virus- and time-dependent
Anti-DENV neutralising and total antibody responses in 35 travellers with monotypic
DENV infection varied among individuals infected with same serotype of DENV.
Antisera from each individual, at similar times post infection, differentially neutralised
homologous and heterologous viruses (Table 5.1-5.9).
Homologous anti-DENV1 antisera from 5 of 6 individuals neutralised D1-15LG1
(originating in Laos in 2015) at high magnitude while acute and convalescent antisera
from one individual consistently neutralised this virus at low magnitude (table 5.1). This
last individual was infected in Bali, Indonesia as were the other 5 people. This pattern of
differential neutralisation was also apparent among heterologous viruses. Similarly, 4
DENV-2 (Cosmopolitan genotype, lineage 4) viruses were differentially neutralised by
homologous and heterologous antisera. These data strongly suggest that the DENV-
specific antibody repertoire varies significantly between individuals infected with
highly similar strains. In addition, the non-virulent epidemic DENV-2 and DENV-3
reference strains were neutralised at greater magnitude by homologous antisera
compared to their corresponding virulent epidemic viruses, which differ by 2 and 3
amino acids, respectively, in the E proteins.
Anti-DENV-1 and anti-DENV-2 antisera cross-neutralise heterologous DENV at high
magnitude whereas anti-DENV-3 and anti-DENV-4 antisera did not. These responses
were highly strain-specific, and this study was limited by the smaller number of DENV-
3 and DENV-4 and antisera available. DENV infections imported into WA by infected
travellers are dependent on endemic and epidemic DENV activity. In recent years the
predominant viruses circulating in neighbouring Indonesia, where most travellers were
infected, were DENV-1 and DENV-2. DENV-3 and DENV-4 circulate and are
imported less frequently. Unique confirmation of DENV-4 has been reported (344) and

256
variation in virus envelope neutralising domain may contribute to the DENV-3 and
DENV-4-specific immune repertoire. However, this pattern was not observed for total
antibody responses derived by hemagglutination inhibition test, which were highly
cross-reactive across all DENV.
Acute phase monotypic anti-DENV antisera are highly cross-reactive and neutralise all
homologous and heterologous DENV. Responses restrict with time and cross-
neutralisation has largely declined by one year. However, heterologous cross-
neutralisation was maintained up to 6 years post infection; antisera differentially
neutralised and recognised distinct virus strains.
Limitation of this study is the small sample size and the convenience sampling of
antisera dictated by this traveller cohort. Enrolment into the study is ongoing and more
recently travellers have returned with more diverse DENV infections, including DENV-
3 and DENV-4 which have not circulated as widely in the past. Future research
directions will focus on characterisation of the anti-DENV B cell repertoire; data from
this present study will form the basis of this work.
5.6. Research outcomes
Anti-DENV antisera differentially neutralise homologous virus strains, including
strains that belong to the same lineage within a genotype.
Anti-DENV antisera differentially neutralise heterologous viruses including
isolates that belong to the same serotype.
Anti-DENV antisera neutralise autologous virus at greater magnitude than
homologous serotype; homologous and heterologous neutralisation declines with
time whereas autologous neutralisation is maintained in most.
Acute phase monotypic anti-DENV antisera are highly cross-reactive and
neutralise all homologous and heterologous DENV.

257
Anti-DENV responses restrict with time and cross-neutralisation largely decline
by one year. However, heterologous cross-neutralisation may be maintained up
to 6 years post infection; antisera differentially neutralise and recognise distinct
virus strains.

258
Chapter 6: Antibody immune response post ZIKV infection
6.1 Introduction
ZIKV belongs to Spondweni serogroup of genus Flavivirus. This serogroup includes
ZIKV and Spondweni virus (SPONV), both virus show high serological cross-
reactivity and similar clinical presentation (125). Discovery of ZIKV dates back 70
years, when it was first identified in the Ziika forest better known as “Zika forest” in
Uganda. First isolation was in 1947 from a sentinel rhesus monkey, followed by
isolation from Aedes africanus in 1948 from the same jungle. SPONV was first
isolated in Nigeria in 1952; first isolated strain was Chuku strain (125). This strain
was originally misidentified as ZIKV. Both viruses are transmitted by mosquito of
Aedes genus, while SPONV has been isolated from multiple other mosquito genera.
ZIKV has a wide geographic distribution; SPONV has been reported from sub-
Saharan Africa. The open reading frame (ORF) of the SPONV Chuku strain
exhibited a 68.3% -69.0% and 74.6% -75.0% nucleotide and amino acid identity
respectively, when compared to geographically and genetically distinct strains of
ZIKV (345). Genetic, geographic and vector variation between SPONV and ZIKV
support that these viruses are separate species although they exhibit misleading
serological and clinical similarities.
ZIKV is reported to exist as single serotype that has evolved into 2 major lineages
differentiated based on its geographic origin: African (MR766 prototype cluster), and
Asian genotype (140, 346). Phylogenetic study of ZIKV was done after 2007 Yap
outbreak by Lanciotti et al. African lineage diverged to East African (MR766
prototype cluster) and West African (Nigerian cluster). Asian lineage diverges to
Southeast Asia and the Pacific subclades (126, 131). Phylogenetic study based on E-
and NS5 gene sequence of 43 ZIKV strains isolated from 1947 to 2007 in Africa,
Asia and the Pacific supports the divergence of African ancestor strains to two

259
clusters, suggesting ZIKV emerged from East Africa (Uganda) and moved to West
Africa (126, 347). The same study shows the ZIKV 2007 Micronesian and Malaysian
strains belong to Asian clade. This finding was also confirmed by E-and NS3
sequence analysis. ZIKV strains within the same lineage are less divergent whereas
there is greater divergence between the two main lineages. There is less than 12%
divergence at the nucleotide level among all ZIKV strains (348). ZIKV strains
reported in the French Polynesia (FP) and Brazil epidemics were closely related and
belonged to the Asian lineage. ZIKV isolates from FP, Brazil, Columbia, and Puerto
Rico showed 99% nucleotide identity within the Asian lineage and ~89% identity to
East and West African lineages (13).
Zika was a silent infection for years, with only a small number of human cases
reported (349, 350). It has been postulated that the virus originated in East Africa and
then spread into both West Africa and Asia approximately 50–100 years ago (351).
Since the outbreak in Yap 2007, a series of outbreaks in different years have been
reported (349, 350). WHO reports till 18 Aug 2016, 70 countries and territories have
evidence of ZIKV transmission since 2007 and 53 countries reported outbreaks from
2015 onwards. In February 2016, 11 countries have reported person-to-person
transmission, 4 with possible endemic transmission or evidence of local infection
(352). Clinical symptoms of Zika include mild fever, arthralgia and conjunctivitis,
with recent reports of association with Guillain Barr Syndrome and Microcephaly
(353).
Epidemiological data suggests the epidemic Asian strain that circulated in FP has
greater virulence than the original Yap strain that emerged in 2007. This FP ZIKV
was associated with neurological complications and was also reported in epidemics
in Brazil followed to other American countries, where it is also associated with
neurological complications. Adaptive genetic changes including glycosylation

260
patterns has associated with evolution of virulent ZIKV strains (140, 351). Lack of
whole genome sequence data limits analysis of virus evolution and divergence
(348).
The spread of ZIKV to dengue-endemic areas has the potential for increased impact
in human health, owing to the high degree of homology between these two
flaviviruses and the potential for immune enhancement. ZIKV is closely related to
DENV with approximately 43% amino acid divergence across the viral polyprotein
as well as the ectodomain of the envelope (E) protein (354). ZIKV E-structure has
been shown to resemble other flaviviruses but contains a unique positive charge
adjacent to the fusion loop believed to influence host attachment (355). The
flavivirus E glycoprotein is the principal antigen in induction of neutralising
antibodies (NAb) that may contribute to protection and which are a marker for
vaccine-induced immunity. The cross-reactive but not necessarily the cross-
protective nature of E-protein-initiated antibody-mediated immune responses has
complicated assessment of ZIKV-induced immunopathology, especially in
populations where DENV is endemic.
Epitopes that elicit potent cross-neutralising antibodies against both ZIKV and
DENV have been described (356). The functional significance of such cross-reactive
antibodies has been investigated and a role for cross-neutralisation as well as
enhancement has been proposed (258, 357).
This study aimed aim to assess the capacity of anti-ZIKV antisera to cross-neutralise
and/or cross-recognise ZIKV, DENV, YFV and JEV. Antisera were obtained from
individuals with monotypic ZIKV infection; recent ZIKV infection in a background
of previous DENV infection; and ZIKV infection in individuals who received YFV
vaccine, from soon after presentation with febrile illness, to 605 days post

261
presentation. Samples were obtained from West Australian travellers who visited
countries with known ZIKV transmission or with a history of ZIKV transmission in
the past. Zika and dengue are not endemic in Western Australia. Samples were tested
against sentinel ZIKV MR766, human Puerto Rico 2015 isolate ZIKV PRVABC59);
DENV (all four serotypes:- DENV-1-HW2001, DENV-2-NGCAII, DENV-H87GV
and DENV-4-H241GI); YF 17D and IMOJEV.
6.2. Aims of study
To assess specificity of anti-ZIKV neutralising antibody responses in
monotypic ZIKV infection and in ZIKV-infected individuals with previous
DENV infection or flavivirus vaccination
To assess duration of ZIKV-specific antibody responses
To identify an anti-ZIKV protein response that may differentiate ZIKV and
DENV infection

262
6.3. Results
6.3.1. Neutralising antibody response
Antisera from 16 individuals sampled within the first month of infection and up to
605 days after presentation to their health care professional were assessed. Eight
individuals were enrolled into the WA Traveller Study (samples were coded “FLV-
[study number]”) and were available for recall and recollection, and 8 presented for
diagnosis of febrile illness (samples coded as “ZIKV-[study-no])” (Table 4.5 also
shown below).
ZIKV infection was classified as ‘confirmed’ or ‘probable’ based on the WHO case
(352). Confirmed cases were those in which ZIKV RNA was detected by PCR (in
urine); probable cases were those in which anti-ZIKV IgM and/or increased HI titres
(more than 4-fold) were detected. All individuals had a history of recent travel to
countries where ZIKV was known to be circulating (South Pacific nations;
Americas) or where sporadic Zika cases been reported (Indonesia; Philippines, India,
Thailand) and presented with clinical symptoms similar to Zika and not explained by
another etiology. ZIKV is not endemic in Western Australia.
Thirty-four serum or plasma samples from 16 individuals were tested. Nine
individuals were sampled on 2 or more occasions from acute phase (< 30 days) to
605 days post presentation (range 2-6 samples). Four individuals received yellow
fever vaccination. Serum or plasma assessed for anti-ZIKV NAb was diluted starting
at 1:10 or 1:40 depending on available sample volume and the maximum dilution
was 1:640. Unless sample volume was limited, all sera were tested against 2 strains
of ZIKV (MR766 and PRVABC59); DENV (all four serotypes when sample volume
permitted; or DENV-4 only when sample volume was restricted); YF17D; and

263
IMOJEV. Neutralising antibody titres are listed in table 6.1. Neutralisation curves for
each individual are shown in Appendix 6.1 and an example is shown in Figure 6.1.
Comparison of 50%, 75% and 90% reduction in interpretation of neutralisation titre
showed that applying 90% reduction produced false negative results in individuals
with confirmed ZIKV infection and thus FRNT-50 was routinely applied here.
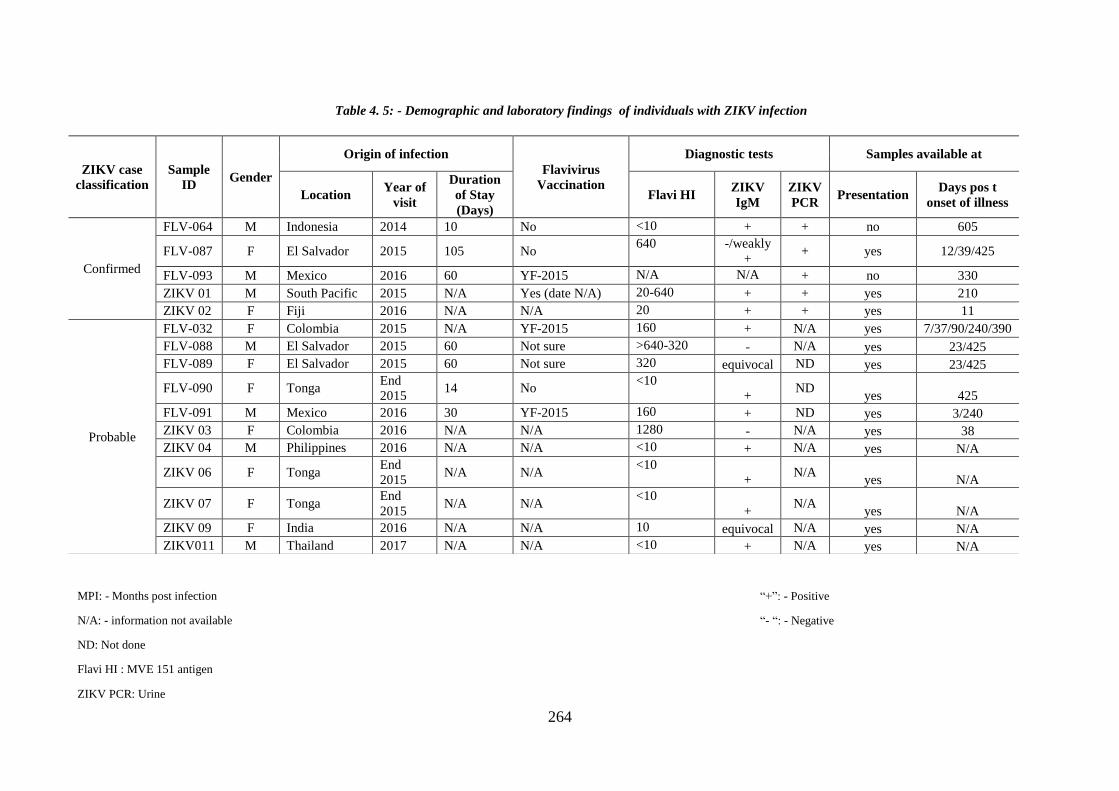
264
ZIKV case
classification
Sample
ID Gender
Origin of infection
Flavivirus
Vaccination
Diagnostic tests Samples available at
Location Year of
visit
Duration
of Stay
(Days)
Flavi HI ZIKV
IgM
ZIKV
PCR Presentation
Days pos t
onset of illness
Confirmed
FLV-064 M Indonesia 2014 10 No <10 + + no 605
FLV-087 F El Salvador 2015 105 No 640 -/weakly
+ + yes 12/39/425
FLV-093 M Mexico 2016 60 YF-2015 N/A N/A + no 330
ZIKV 01 M South Pacific 2015 N/A Yes (date N/A) 20-640 + + yes 210
ZIKV 02 F Fiji 2016 N/A N/A 20 + + yes 11
Probable
FLV-032 F Colombia 2015 N/A YF-2015 160 + N/A yes 7/37/90/240/390
FLV-088 M El Salvador 2015 60 Not sure >640-320 - N/A yes 23/425
FLV-089 F El Salvador 2015 60 Not sure 320 equivocal ND yes 23/425
FLV-090 F Tonga End
2015 14 No
<10
+ ND
yes 425
FLV-091 M Mexico 2016 30 YF-2015 160 + ND yes 3/240
ZIKV 03 F Colombia 2016 N/A N/A 1280 - N/A yes 38
ZIKV 04 M Philippines 2016 N/A N/A <10 + N/A yes N/A
ZIKV 06 F Tonga End
2015 N/A N/A
<10
+ N/A
yes N/A
ZIKV 07 F Tonga End
2015 N/A N/A
<10
+ N/A
yes N/A
ZIKV 09 F India 2016 N/A N/A 10 equivocal N/A yes N/A
ZIKV011 M Thailand 2017 N/A N/A <10 + N/A yes N/A
Table 4. 5: - Demographic and laboratory findings of individuals with ZIKV infection
MPI: - Months post infection
N/A: - information not available
ND: Not done
Flavi HI : MVE 151 antigen
ZIKV PCR: Urine
“+”: - Positive
“- “: - Negative

265
*:- Confirmed ZIKV cases
NA: “Not available” due to lack of sample these samples could not be tested against ZIKV PRVABC59
Detectable neutralising antibody response
None of anti-ZIKV antisera showed cross reactivity against IMOJEV
Table 6. 1:- Neutralising antibody response in ZIKV infection
FRNT50 FRNT90 FRNT50 FRNT90 FRNT50 FRNT90 FRNT50 FRNT 90 FRNT50 FRNT90 FRNT50 FRNT90 FRNT50 FRNT90
FLV-064* Indonesia 2014 No 605 65 22 23 11 <10 <10 <10 <10 <10 <10 <10 <10 <40 <40
<30 ≥640 89 NA NA NA NA NA NA NA NA 76 <40 <40 <40
12 ≥640 ≥640 572 <40 NA NA NA NA NA NA ≥640 ≥640 <40 <40
39 ≥640 185 224 <40 170 46 ≥640 160 ≥640 134 ≥640 160 <40 <40
425 457 109 115 33 46 11 639 153 252 46 279 49 <40 <40
FLV-093* Mexico 2016 Yes (2014) 330 322 92 48 19 <10 <10 <10 <10 <10 <10 111 <10 236 16
<30 ≥640 324 NA NA <40 <40 <40 <40 <40 <40 95 <40 ≥640 73
210 ≥640 ≥640 223 62 <40 <40 <40 <40 <40 <40 <40 <40 322 43
<30 ≥640 125 ≥640 148 NA NA NA NA NA NA 94 <40 <40 <40
11 ≥640 216 ≥640 235 NA NA NA NA NA NA 198 <40 <40 <40
<30 176 48 NA NA NA NA NA NA NA NA 55 <30 195 <40
7 619 30 NA NA NA NA NA NA NA NA <40 <40 <40 <40
37 ≥640 78 85 <40 <40 <40 <40 <40 <40 <40 <40 <40 <40 <40
90 523 116 274 46 <40 <40 <40 <40 <40 <40 <40 <40 <40 <40
240 379 94 116 37 <40 <40 <40 <40 <40 <40 <40 <40 <40 <40
390 381 92 129 38 <40 <40 <40 <40 <40 <40 <40 <40 <40 <40
<30 ≥640 ≥640 43 <40 ≥640 ≥640 ≥640 ≥640 ≥640 <40 ≥640 ≥640 <40 <40
23 ≥640 ≥640 106 <40 ≥640 ≥640 ≥640 ≥640 ≥640 98 ≥640 ≥640 <40 <40
425 431 48 57 17 ≥640 108 ≥640 123 208 18 ≥640 116 <40 <40
<30 ≥640 ≥640 194 20 ≥640 159 ≥640 271 331 58 ≥640 ≥640 <40 <40
23 ≥640 ≥640 48 <40 605 124 ≥640 102 519 40 ≥640 ≥640 <40 <40
425 ≥640 110 47 11 292 58 563 59 152 15 ≥640 198 <40 <40
<30 ≥640 ≥640 148 <40 NA NA NA NA NA NA <40 <40 <40 <40
425 82 29 40 15 <10 <10 <10 <10 <10 <10 <10 <10 <40 <40
<30 ≥640 ≥640 349 38 <40 <40 ≥640 133 63 <40 306 60 ≥640 109
3 ≥640 215 275 <40 <40 <40 348 82 40 <40 ≥640 107 618 92
240 235 85 105 30 <20 <20 <20 <20 <20 <20 193 14 ≥320 30
<30 ≥640 ≥640 252 48 ≥640 518 ≥640 626 ≥640 323 ≥640 ≥640 353 <40
38 ≥640 ≥640 107 42 ≥640 182 ≥640 470 ≥640 55 ≥640 363 <40 <40
ZIKV 04 Philippines 2016 N/A <30 ≥640 222 ≥640 <40 NA NA NA NA NA NA <40 <40 <40 <40
ZIKV 06 Tonga End 2015 N/A <30 136 36 NA NA NA NA NA NA NA NA <40 <40 <40 <40
ZIKV 07 Tonga End 2015 N/A <30 ≥640 11 NA NA NA NA NA NA NA NA <40 <40 <40 <40
ZIKV 09 India 2016 N/A <30 573 57 NA NA NA NA NA NA NA NA <40 <40 <40 <40
ZIKV011 Thailand 2017 N/A <30 ≥320 157 ≥160 23 <40 <40 <40 <40 <40 <40 <40 <40 <40 <40
ZIKV 03 Colombia 2016 N/A
FLV-090 Tonga 2015 No
FLV-091 Mexico 2016 Yes(2015)
FLV-088El
Salvador2015 No
FLV-089El
Salvador2015 No
ZIKV 02* Fiji 2016 N/A
FLV-032 Colombia 2015 Yes-2015
FLV-087*El
Salvador2015 No
ZIKV 01*South
Pacific2015
Yes(date
N/A)
Anti DENV FRNTAnti YF-17D
ZIKV MR766 ZIKV PRVABC59 Anti DENV-1-FRNT Anti DENV-2-FRNT Anti DENV-3-FRNT Anti DENV-4-FRNTSample ID Location
Date of
travel
Vaccinatio
n History
Time post
presentati
on of
illness
(days)
Anti-ZIKV FRNT
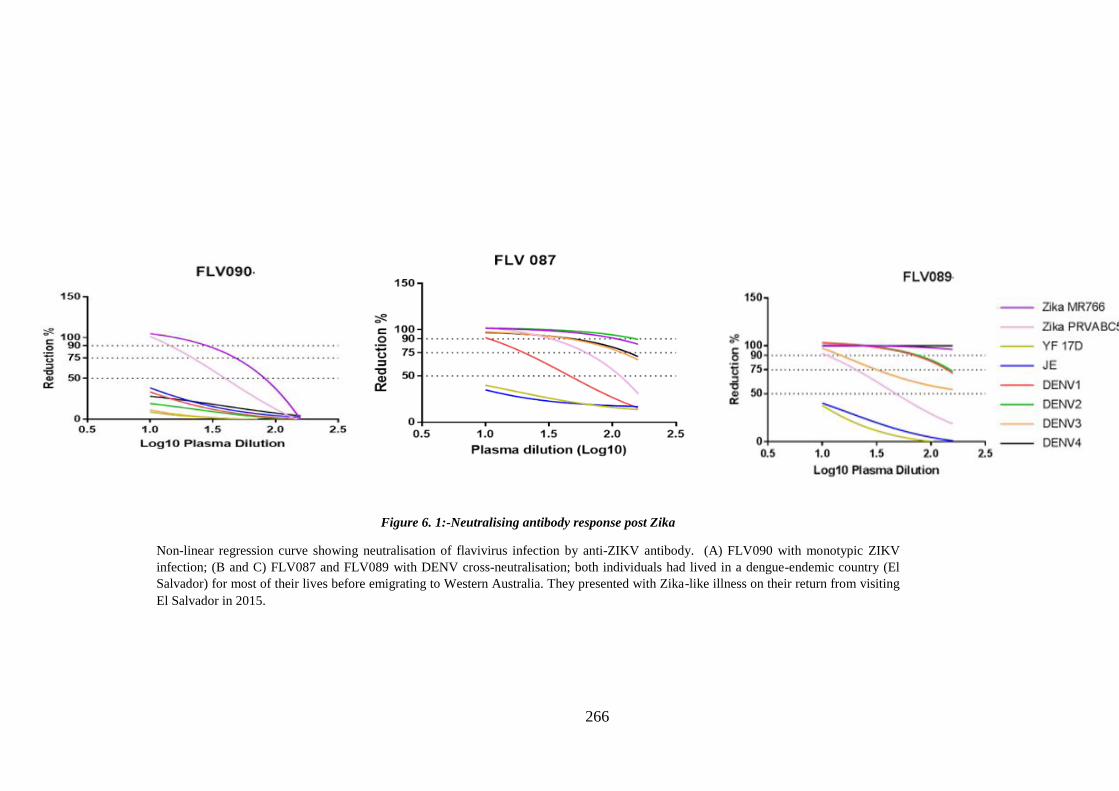
266
Figure 6. 1:-Neutralising antibody response post Zika
Non-linear regression curve showing neutralisation of flavivirus infection by anti-ZIKV antibody. (A) FLV090 with monotypic ZIKV
infection; (B and C) FLV087 and FLV089 with DENV cross-neutralisation; both individuals had lived in a dengue-endemic country (El
Salvador) for most of their lives before emigrating to Western Australia. They presented with Zika-like illness on their return from visiting
El Salvador in 2015.

267
Anti-ZIKV, anti-DENV, anti-YFV and anti-JEV antibody responses were assessed
(a) within 1 month; (b) 1-12 months; and (c) more than 12 months following
presentation with febrile illness.
a. Within 1 month
Samples were available from 11 of 16 individuals within 1 month of presentation.
High magnitude (>640) anti-ZIKV-MR766 FRNT-50 NAb titres >640 were detected
for 10 of 11 individuals. Samples from 7 individuals were available in sufficient
volume to allow assessment of anti-ZIKV-PRABC59; a minority (2 of 7) neutralised
this strain at high magnitude. Anti-ZIKV-MR766 GMT was greater than anti-ZIKV-
PRABC59 GMT (Figure 6.2). Two antisera produced low anti-ZIKV-MR766 NAb
titres; both were probable (IgM positive) cases. One of these cases submitted a
second sample 7 days later at which time NAb titre increased to 619.
ZIKV antisera from 9 of 11 individuals cross-neutralised DENV. When serum
volume permitted, all four DENV serotypes were tested; DENV-4 alone was tested
when sample volume was limited. Sera were available from 3 ZIKV confirmed cases
(FLV087; ZIKV01; ZIKV02); all 3 antisera neutralised DENV (Table 6.1). FLV087
reported a previous confirmed DENV infection while traveling in El Salvador 5
years earlier and her anti-DENV NAb respones were maintained in samples collected
following her enrolment into the Traveller Study (Table 6.1). Anti-DENV NAb titres
for ZIKV01, with no history of previous DENV infection, declined when tested at
210 days while titres were maintained at day 11 for ZIKV02, the latest sample that
was available. For both ZIKV01 and ZIKV02, FRNT90 titres were <40 and thus,
below the limits of detection of this test.

268
Anti-ZIKV antisera from 5 of 16 individuals cross-neutralised YFV17D. Of the 5
individuals, 4 were known to have previously received yellow fever vaccination.
Follow-up samples were available for 4 individuals, 3 of whom were previously
vaccinated, for between 38 – 390 days. Anti-YFV FRNT50 titres were maintained
for 210 and 240 days for 2 vaccinated individuals and declined to undetectable levels
for the other 2, one of whom was vaccinated, 7 and 38 days after the sample
collected at presentation. In both cases FRNT90 at the first, positive, sample was <40
and thus below levels of detection. Comparative FRNT50 and FRNT90 for two
strains of ZIKV is shown in figure 6.2.
b. 1 – 12 months post presentation
Sera from 6 individuals collected 30 – 365 days after presentation were assessed.
Anti-ZIKV MR766 GMT declined from 532 to 473 (12% decline), and anti-ZIKV
PRVABC59 GMT also declined, from 242 to 128 (47% decline). (Figure 6.3).
Anti-ZIKV antisera cross-neutralisation of DENV was maintained in 5 of 7 antisera
from 6 individuals with known previous DENV infection or who had lived in
dengue-endemic countries into adulthood before migrating to Western Australia.
Then 2 individuals in whom anti-DENV titres were not maintained (FLV032,
ZIKV01) were travellers who returned to WA from countries where ZIKV was
known to be circulating and who presented soon after with febrile illness, ZIKV
RNA was detected in one of their acute phase samples, and anti-ZIKV IgM was
detected in the other. Neither had a history of previous DENV infection. Samples
from all individuals neutralised DENV in the first moth after presentation.
NAb responses against YFV17D were maintained in vaccine recipients, with the
exception of FLV-032 whose anti-YFV titres had declined to undetectable levels by

269
day 37. She received her YF vaccine in Australia in October 2015, soon before
traveling to Columbia.
c. More than 12 months post presentation
Sera from six individuals were available more than one year after presentation (390 –
365 days). Five of the 6 were sampled at earlier time points.
Anti-ZIKV MR 766 FRNT50 GMT decreased from 473 (1-12 months post
presentation) to 248 and anti -ZIKV PRVABC59 GMT declined from 128 to 58
(Figure 6.4).
Serum from 3 of the 6 individuals (FLV087; FLV088; FLV089) cross-neutralised
DENV-4 with FRNT50 ranging from maximum >640 to lowest279; similarly,
FRNT90 ranged from >640-49. FLV087 was previously infected with DENV,
FLV088 and FLV089 had both lived in El Salvador for most of their lives before
migrating to WA as adults and although they do not recall being diagnosed with
dengue while living there, previous DENV infection in the endemic country is not
unlikely.
Antisera from the remaining 3 individuals (FLV064; FLV032; FLV090) did not
cross-neutralise DENV, and all 3 were never previously diagnosed with dengue and
had lived in WA all their lives. FLV064 was only sampled 605 days after presenting
with a febrile illness after returning from Indonesia, and ZIKV RNA was detected in
his urine.
Persistence of anti-ZIKV neutralising antibody in history of flavivirus exposure is
illustrated by heat map Figure 6.5 (A, B, and C).

270
Study ID
Color code
FLV-087
ZIKV 01
ZIKV 02
FLV-032
FLV-088
FLV-089
FLV-090
FLV-091
ZIKV 03
ZIKV 04
ZIKV 06
ZIKV 07
ZIKV 09
ZIKV011
Figure 6. 2:-ZIKV neutralisation within 30 days post presentation of illness.

271
Study ID Color code
FLV-087
FLV-093
ZIKV 01
FLV-032
FLV-091
ZIKV 03
Figure 6. 3:- ZIKV neutralisation 1-12 months post presentation of illness.

272
Study ID Color code
FLV-064
FLV-087
FLV-032
FLV-088
FLV-089
FLV-090
Figure 6. 4: ZIKV neutralisation more than 12 months post presentation of illness.

273
Figure 6. 5:-Variation in ZIKV neutralising antibody concentration over time based on the flavivirus background
Values on heat map represent antibody titre at respective days post infection. After one year post infection neutralisation of ZIKV in the
absence of previous flavivirus background was significantly lower than in the presence of anti-flavivirus background.
(A) Anti-ZIKV NAb from patients with ZIKV only infection 4 patients at different time points were tested (B) Anti-ZIKV NAb from
patients with DENV cross reactivity, one of the patient has confirmed DENV previous infection, while rest has DENV NAb when tested
at the given time point but no record of past DENV infection (C) NAb from patients with known history of YF 17D vaccination. NAb titre
was maintained high in individual with flavivirus exposure in compare to flavivirus naïve population.
(B) (C) ZIKV with DENV ZIKV with YF vaccination
(A) ZIKV only
Stu
dy n
o. (D
ays
post
in
fect
ion
)
(A) ZIKV only
6 4 0
6 4 0
6 4 0
6 4 0
1 7 6
6 1 9
6 4 0
6 4 0
2 3 5
3 7 9
3 2 2
3 8 1
6 4 0
2 5 2
3 4 9
2 7 5
3 8
3 0
8 5
2 2 3
1 0 5
1 1 6
4 8
1 2 9
ZIK
V M
R766
ZIK
V P
RV
AB
C59
ZIK V 0 1 (< 3 0 )
ZIK V 0 3 (< 3 0 )
F L V 0 9 1 (< 3 0 )
F L V 0 9 1 (3 )
F L V 0 3 2 (< 3 0 )
F L V 0 3 2 (7 )
F L V 0 3 2 (3 7 )
ZIK V 0 1 (2 1 0 )
F L V 0 9 1 (2 4 0 )
F L V 0 3 2 (2 4 0 )
F L V 0 9 3 (3 3 0 )
F L V 0 3 2 (3 9 0 )
2 0 0
4 0 0
6 0 0

274
6.3.2. Total anti-ZIKV antibody
Total anti-ZIKV antibody (TAb) was assessed by Hemagglutination inhibition (HI) test
to assess magnitude of responses against ZIKV MR 766 and ZIKV PRVABC59.
Sera from 14 of 16 individuals were assessed at least once post presentation. Among 4
Zika confirmed cases, 2 demonstrated a >4-fold difference in HI titres between the
strains. In Zika probable cases, 3 of 9 demonstrated a differential HI pattern. All four
showed HI titre against MR766 >4-fold greater compared to PRVABC59. Decline in HI
titres against both viruses with progression of time post infection was observed in
patients in whom consecutive samples were available.
Furthermore, high HI titre (>100) against both ZIKV MR 766 and ZIKV PRVABC59
was observed in individuals with previous DENV infection whereas HI titre was <100,
against ZIKV PRVABC59, in individuals with no history of previous DENV infection,
at the same timepoint after presentation (Table 6.2). ZIKV PRVABC59 is therefore a
more specific target for anti-ZIKV antisera induced by contemporaneous ZIKV
infection.
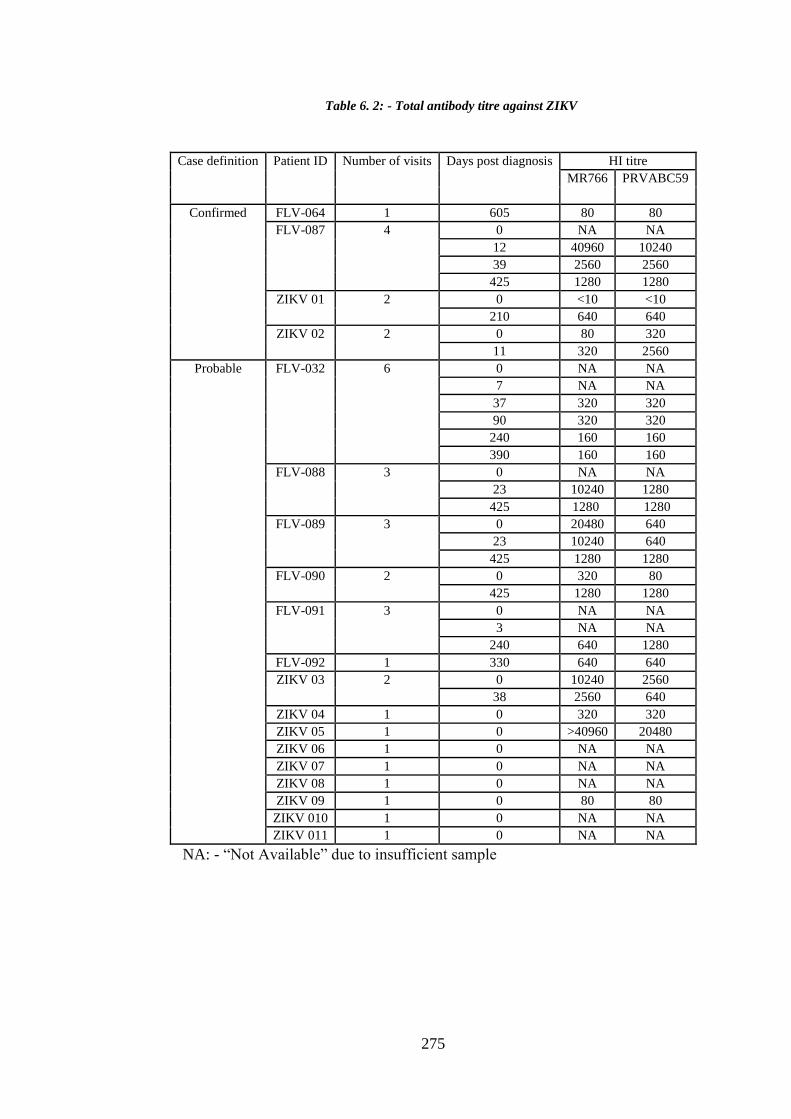
275
Case definition Patient ID Number of visits Days post diagnosis HI titre
MR766 PRVABC59
Confirmed FLV-064 1 605 80 80
FLV-087 4 0 NA NA
12 40960 10240
39 2560 2560
425 1280 1280
ZIKV 01 2 0 <10 <10
210 640 640
ZIKV 02 2 0 80 320
11 320 2560
Probable FLV-032 6 0 NA NA
7 NA NA
37 320 320
90 320 320
240 160 160
390 160 160
FLV-088 3 0 NA NA
23 10240 1280
425 1280 1280
FLV-089 3 0 20480 640
23 10240 640
425 1280 1280
FLV-090 2 0 320 80
425 1280 1280
FLV-091 3 0 NA NA
3 NA NA
240 640 1280
FLV-092 1 330 640 640
ZIKV 03 2 0 10240 2560
38 2560 640
ZIKV 04 1 0 320 320
ZIKV 05 1 0 >40960 20480
ZIKV 06 1 0 NA NA
ZIKV 07 1 0 NA NA
ZIKV 08 1 0 NA NA
ZIKV 09 1 0 80 80
ZIKV 010 1 0 NA NA
ZIKV 011 1 0 NA NA
NA: - “Not Available” due to insufficient sample
Table 6. 2: - Total antibody titre against ZIKV

276
6.3.3. Immunoblot analysis, ZIKV and DENV
The high degree of amino acid similarity among flaviviruses, including is preliminary
study to possible differential serological diagnosis post DENV and ZIKV infection. In
this study antisera from WA travellers with monotypic DENV or ZIKV infection was
applied as probe to identify viral protein targets in Western immunoblot analysis and to
assess whether a specific marker for ZIKV infection could be identified. Two ZIKV,
ZIKV MR766 and ZIKV PRVABC59; four DENV serotype reference viruses (DENV-1
HW2001G4, DENV-2-NGCAII, DENV-3-H87G5 and DENV-4-H241GI) and YF17D
were used to identify possible virus-specific binding sites.
Viral protein preparation and determination of binding sites is described in detail in
Chapter 3. Briefly, virus whole cell lysates were prepared from infected Vero cells.
Infected monolayers were scraped with cell scrappers and fixed with 0.025%
glutaraldehyde. Fixed infected cells were sonicated to prepare whole virus lysate. Cell
lysate for mock infections was prepared in a similar way as virus-infected cells. Lysates
were quantified for protein content using bicinchoninic acid assay (BCA assay). Equal
quantities (20ug/ul) of crude protein cell lysate were denatured with loading buffer (5X
laemmili sample buffer) at 950C and then subjected to 12% SDS-PAGE, followed by
transfer to nitrocellulose membrane. After transfer membrane was blocked with 5%
non-fat milk in PBST (PBS with 0.05% Tween 20, pH 7.4). Membranes were then
probed overnight with antisera as primary antibody, diluted in blocking buffer. Titration
of the correct serum dilution was determined by dot blot assay. Anti-human antibody
labelled with horseradish peroxidase was used as secondary antibody, incubated with
ECL substrate and visualised using FujiFilm Imager 3000.

277
Protein band position was visualised with Coomassie stained gels. Band size was
determined with reference to 15-250 protein ladder. Based on published data (358-361)
a band between 80-46KDa was identified as E-protein fraction and between 22-11KDa
as capsid/prM protein. No purification of protein lysate or primary antibody was done
and therefore background was high.
Two antisera from individuals (FLV064; FLV090) with confirmed ZIKV infection and
with no other detectable anti-flavivirus antibody (no vaccination and no
DENV/YF17D/IMOJEV neutralisation); one sample from an individual with previous
ZIKV and DENV infection (FLV087); one sample from an individual with ZIKV
infection, YF17D vaccination, no DENV cross-neutralisation (FLV032) and 2 samples
with monotypic DENV infection and no ZIKV cross-neutralisation (FLV030 and
FLV002) were assessed for their capacity to bind to flavivirus proteins. The flavivirus-
specific monoclonal antibody 4G2 was included as Positive control that recognises E
protein; pooled flavivirus seronegative sera served as Negative control. Presence of
protein bands was confirmed by Coomassie stain (Figure 6.6).
6.3.3.1. Positive and negative control
Purified 4G2 bound to ZIKV and DENV; a clear band at 80-56KDa was seen and there
was no binding with mock lysate (Figure 6.7 A and B).
Negative Control produced faint bands at 80KDa and numerous faint bands were seen
with the mock lysate sample.
6.3.3.2. Anti-ZIKV antisera
A band at capsid/prM protein at 22-11KDa was specific to ZIKV only and was present
for both ZIKV MR766 and ZIKV PRVABC59. In addition to this band the 80-56KDa
E-protein was observed for all the flavivirus lysates (Figure 6.8 A and B). E-protein

278
binding was seen in the mock lysate and this might be due to addition of FBS during
lysate preparation (this has been widely reported in Immunoblotting discussion sites and
in Immunoblot kit manufacturer FAQ sites); no binding to mock lysate was seen in dot
blots.
6.3.3.3. Anti-ZIKV/DENV antisera
The 22-11 KDa band specific to ZIKV capsid/prM protein was present in ZIKV lysates
alone, along with high intensity 80-56KDa E protein for all viruses (Figure 6.9).
6.3.3.4. Anti- ZIKV/YF17D
No WB bands for any viral proteins could be identified in multiple trials. The same
batch and amount of protein was used, as for other assays, but no specific bands were
seen. Binding did occur in dot blot analysis where spots were obtained at 1:50 plasma
dilution however when the same dilution was used to develop the post-transfer
membrane no bands appeared (Figure 6.10). This might be due to lack of primary
antibody binding to separated protein fragments.
The individual from whom this antiserum was obtained (FLV032) clearly developed
anti-ZIKV neutralising antibody on her return from travel to Columbia and was
vaccinated against yellow fever before she left Australia. Serum collected at her first
visit neutralised YF17D, but subsequent samples collected 7, 37, 90, 240, and 390 days
after her first visit did not. It is possible that WB is not sufficiently sensitive in this
individual’s case.
6.3.3.5. Anti-DENV
Anti-DENV-3 and DENV-4 antisera from 2 travellers with monotypic DENV-3 or
DENV-4, respectively, did not cross-neutralise ZIKV MR766 or ZIKV PRVABC59, or
heterologous DENV.

279
Anti-DENV-4 specifically bound to DENV-4 specific capsid/prM only. Faint bands
were seen across E-protein. Anti-DENV-3 sample showed a faint band at 22-11KDa
specific to DENV-3 despite the high degree of background non-specific binding in this
immunoblot (Figure 6.11).

280
Figure 6. 6:-Coomassie stain.
Coomassie stain (CS) SDS-PAGE shows lysate position after
electrophoresis

281
Figure 6. 7:-Positive and Negative control
(A) Immunoblot of purified 4G2 monoclonal antibody against DENV/ZIKV and mock lysate.
Bands produced after Coomassie stain are shown. A specific E protein band at 80-56KDa was
seen, with no non-specific binding to mock vero cell lysate. (B) Immunoblot of Negative Control
(pool of flavivirus-seronegative sera). A band at 80-46KDa was visible for mock lysate and a few
other faint bands are present.
B
A

282
Figure 6. 8:-Monotypic anti-ZIKV antisera (FLV-064/FLV090)
ZIKV-specific prM/Capsid protein band at 22-11KDa in monotypic anti-ZIKV antisera. E
protein band at 80-46KDa seen across all virus lysates. Variation in band intensity is apparent.
Dot blot at varying plasma dilution (1:50/1:100/1:200) against protein lysate ZKM (ZIKV
MR766), ZKP (ZIKV PRVABC59), DENV 1-4 (D1-D4), YF17D(YF) and Vero mock lysate
(V).
Dot blot for Plasma
titration
A

283
Figure 6. 9:- Anti-ZIKV/DENV antisera (FLV-087)
ZIKV-specific prM/Capsid protein band was visible. E protein band was
more intense than for anti-ZIKV or anti-DENV alone. Dot blot at varying
plasma dilution (1:50/1:100/1:200) against protein lysate ZKM (ZIKV
MR766), ZKP (ZIKV PRVABC59), DENV 1-4 (D1-D4), YF17D(YF) and
Vero mock lysate (V).
Dot blot plasma titration

284
Dot blot for 1o Ab titration
Figure 6.10:-Anti-ZIKV/YFV antisera (FLV-032)
A) No bands were visible in immunoblot of FLV0321 antisera, although binding was seen in dot
blot analysis using the same lysate.
B) (B) Dot blot at varying plasma dilution (1:50/1:100/1:200) against protein lysate ZKM (ZIKV
MR766), ZKP (ZIKV PRVABC59), DENV 1-4 (D1-D4), YF17D(YF) and Vero mock lysate
(V). Distinct binding at 1:50 dilution for all lysates, except mock, was obtained. It is possible
that WB is not sufficiently sensitive in this individual’s case.
Dot blot plasma titration
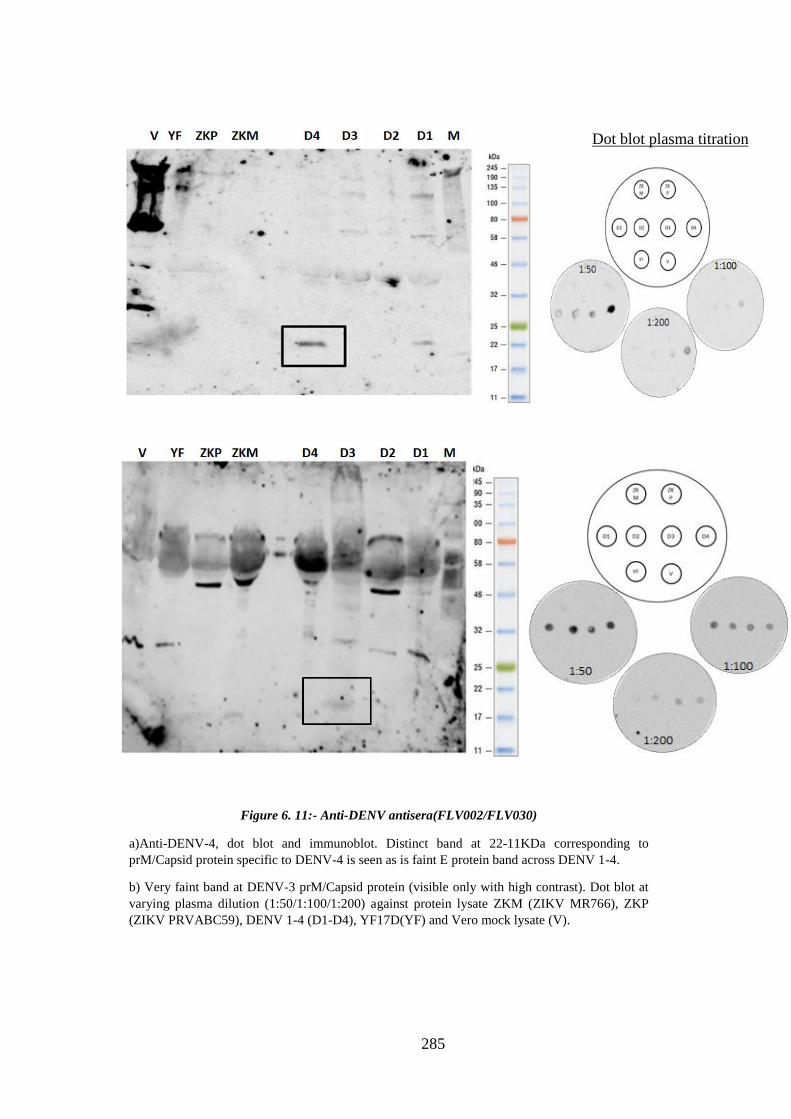
285
Figure 6. 11:- Anti-DENV antisera(FLV002/FLV030)
a)Anti-DENV-4, dot blot and immunoblot. Distinct band at 22-11KDa corresponding to
prM/Capsid protein specific to DENV-4 is seen as is faint E protein band across DENV 1-4.
b) Very faint band at DENV-3 prM/Capsid protein (visible only with high contrast). Dot blot at
varying plasma dilution (1:50/1:100/1:200) against protein lysate ZKM (ZIKV MR766), ZKP
(ZIKV PRVABC59), DENV 1-4 (D1-D4), YF17D(YF) and Vero mock lysate (V).
Dot blot plasma titration

286
6.4. Summary discussion
In this Chapter, acute and convalescent anti-ZIKV total and neutralising antibody
responses following presentation for Zika-like illness were assessed. Cross-
neutralisation was serogroup-specific. In the absence of previous DENV infection
convalescent anti-ZIKV antisera do not necessarily cross-neutralize or cross-recognize
DENV, and DENV was not cross-neutralised by anti-ZIKV antisera from individuals
who had received YFV vaccination. The magnitude of anti-ZIKV NAb responses
differentially declined with time: responses in individual with previous DENV
infection declined slowly while responses in individuals with no previous infection
declined to very low magnitude after 1 year, to titres considered negative according to
WHO criteria for ZIKV laboratory diagnosis. ZIKV MR766 and ZIKVPRVABC59
were differentially neutralised and this finding highlighted the importance of strain
selection in ZIKV diagnostic and research approaches.
6.4.1. Anti-ZIKV specificity and cross-reactivity
Sero diagnosis of ZIKV infection in dengue endemic areas is confounded by cross-
recognition of DENV antigens by anti-ZIKV antibodies. ZIKV-infected travellers who
are not from dengue-endemic areas allow investigation of sensitivity and specificity of
humoral immune responses in ZIKV infection. Cases of travel associated ZIKV
infection has been studied before. The first Zika case in Australia was reported was in a
previously healthy 52-year old Australian woman who became ill after returning from
Jarkarta (322). She was provisionally diagnosed with dengue but following a negative
dengue-specific RT-PCR test, sequencing of flavivirus-specific PCR product identified
the infecting virus as ZIKV. Prior to this, in 2008 ZIKV was first reported in USA in a
scientist returning from Senegal (361). This was also the first report of possible sexual
transmission of ZIKV. Leung et al proposed that an animal bite was a plausible route of

287
transmission in the case of Zika in a traveller returning to Australia from Indonesia,
where he was bitten by a monkey (362). However, the authors did not consider the
possibility that ZIKV was transmitted to this individual via an infected mosquito and
that ZIKV was present in Indonesia but not yet identified in humans. Other cases of
ZIKV imported to countries without known ZIKV circulation as listed in WHO reports
include Germany (2013, 2014), Canada (2014), Japan (2014), and Finland (2015) (363).
Although many traveller-associated cases have been reported there are no analyses
available from these presumably monotypic ZIKV infections regarding serum anti-
ZIKV antibody responses against DENV in vitro. Analysis of human anti-ZIKV
monoclonal antibodies (mAbs) has been reported; Wang et al (2017) described 460
mAbs directed to E-protein and compared neutralisation mechanisms and potential viral
epitopes to obtain insights for possible therapeutic approaches(364). Steller et al
isolated a panel of 119 mAbs from epidemic plasma samples, assessed their specificity
for ZIKV, and showed that the most potent neutralising antibodies were targeted against
ZIKV EDIII and NS1 protein. ZIKV EDIII monoclonal specificity has also been
reported in other studies (365).
In this study, serum samples collected from travellers with monotypic ZIKV infection
or ZIKV infection with known or likely previous DENV infection and/or YFV
vaccination were assessed for cross-neutralization and cross-recognition of DENV,
YFV and JEV, and ZIKV strain-specific responses were also examined. Using
polyclonal human sera allowed analysis of the natural history of ZIKV-specific humoral
immune responses. A major observation was that specificity of ZIKV-specific
neutralisation and binding is time- and flavivirus background-dependent. Anti-ZIKV
cross-neutralised DENV, YF17D, and JEV in early acute phase. Responses restricted
with time to become more ZIKV-specific, in the absence previous flavivirus exposure.
This pattern was patient-specific; certain individuals who had been immunised against

288
YFV maintained ZIKV-specific NAb even as their YFV-specific responses declined. A
second major observation was the dramatic decline in anti-ZIKV NAb, to titres
considered negative for ZIKV infection, more than one year after monotypic ZIKV
infection.
6.4.2. Anti-ZIKV antisera cross-neutralisation of DENV is time- and flavivirus
background-dependent
The capacity of anti-DENV antisera to cross-neutralise and cross-recognise ZIKV has
been assessed, but the capacity of anti-ZIKV antisera to cross-neutralise and cross-
recognise DENV and other flaviviruses, particularly the vaccine viruses, is not
understood. A major finding in the present study is that anti-ZIKV antisera
neutralisation and recognition of DENV is highly patient- and time-specific. Among
confirmed Zika cases cross-neutralisation against DENV was seen only in individuals
with known or likely DENV infection and similarly, cross-reactivity against YF17D
was seen only in the context of known YF vaccination history. Epitope-specific
neutralisation potency of anti-ZIKV antibody is a possible explanation for the varying
neutralisation pattern observed among our cohort. Anti-ZIKV antibody directed towards
DENV E-protein DI/DII was shown to be poorly neutralising while antibody towards
EDIII and NS1 was highly specific (365). Among 400 donors from dengue epidemic
areas of Mexico and Brazil, ZIKV and DENV neutralisation potencies varied over 2
logs. B cell clones expressed E-protein specific to ZIKV and DENV-1 neutralising
antibody in 4 out of 6 individuals (258). Analysis of the ZIKV-specific B cell repertoire
is the target of research in the WA Traveller cohort and will be based on data generated
in the current study.
In summary the present study in WA Travellers showed that convalescent anti-ZIKV
antibody response after monotypic ZIKV infection is specific to ZIKV. DENV cross-

289
neutralisation occurred in the context of previous DENV infection and was limited to
the DENV serocomplex; no cross reactivity against YFV or JEV was measured.
6.4.3. Persistence of anti-ZIKV NAb in individuals with previous flavivirus infection
High magnitude (FRNT50 >640) anti-ZIKV NAb responses were present in all
individuals in acute infection. In convalescence, ~13-14 months post presentation, high
magnitude responses against ZIKV MR766 and ZIKV PRVABC59 were measured for
antisera from individuals with YFV vaccination or previous/likely DENV infection
whereas in monotypic ZIKV infection FRNT50 titres were low (82 and ~40 respectively
against ZIKV MR766 and ZIKV PRVABC59) at 425 days for patient FLV090 and
similarly low titres for patient FLV064 (65 and 23 against ZIKV MR766 and ZIKV
PRVABC59, respectively) at 605 days. Anti-DENV-1 antibody has been proposed to
prime subsequent ZIKV infection resulting higher neutralisation, compared to a naïve
ZIKV population (258) in a mouse model.
In summary this study demonstrates that previous flavivirus exposure prolongs the anti-
ZIKV antibody response, whereas in monotypic ZIKV infection anti-ZIKV NAb titres
decline to levels considered to be negative.
6.4.4. Differential ZIKV MR766 and anti-ZIKV PRVABC59 neutralisation
Differential neutralisation against ZIKV MR766 and anti-ZIKV PRVABC59 by anti-
ZIKV antisera was observed. The majority of antisera (89%) neutralised ZIKV MR766
at high magnitude (FRNT50>640) while a minority (< 20%) showed FRNT50>640
against ZIKV PRVABC59. No individuals had undetectable neutralisation against
ZIKV MR766 (by the stringent WHO criteria for ZIKV serodiagnosis FRNT90<40)
while most did not neutralise ZIKV PRVABC59 (FRNT90<40) even during the acute
phase. To assess potential assay bias traditional PRNT was also performed for these
samples and confirmed the pattern of strain-specific neutralisation.

290
Total anti-ZIKV antibody assessed by HI test showed that a subset of antisera
differentially recognised the 2 ZIKV strains, at least 4-fold, indicating the presence of
cross-recognising but not necessarily cross-neutralising antibody elicited after ZIKV
infection. A study comparing four strains of ZIKV reported ~6-fold higher
neutralisation against ZIKV MR766 compared to ZIKV PRVABC59(366), similar to
the results of the present study. Differential neutralisation by DENV immune sera
against ZIKV HD78788 and PF13 has been reported (260). It has also been shown that
not all mAbs equally neutralise all ZIKV strains. On evaluation of 6 mAbs, 4 out of 6
showed higher FRNT 50 for African strain compared to Asian strain; 3of 6 had more
than 4-fold difference in FRNT titre (367). Phenotypic difference between Asian (CPC-
0740, SV0127-14) and African (ArD 41525) lineages of ZIKV were compared for vivo
and vitro phenotypic difference and it was reported that African isolates at low passage
had higher infection and dissemination rates compared with Asian isolates (368). In
contrast, a study on neutralisation of ZIKV MR766 and ZIKV (H/PF/2013) French
Polynesian strain by sera or plasma samples from eight ZIKV-infected individuals
showed that both ZIKV strains showed similar (< 3-fold) sensitivity to neutralisation
(369).
Identification of conserved amino acid residues that differentiate the Asian strain,
including American strain, from the ancient African strains has been reported (370).
Glycosylation in the envelope (E) protein has been suggested to contribute to
dominance of the Asian lineage in the recent Oceania epidemics (131). The recent Asian
epidemic virus contains an N-Linked glycosylation signal, whereas the majority of other
strains do not, suggesting a potential role for glycosylation in ZIKV disease severity
(371). Addition of 12 nucleotides in E, in the Asiatic lineage from the Yap ZIKV 2007
outbreak is also believed to be noteworthy as 4 E amino acids corresponding to an E
145 glycosylation motif described in flaviviruses has been associated with virulence

291
(131). Another candidate for neutralisation resistance among flaviviruses is substitution
in E Ser279 (372).
Differential cross neutralisation among the two ZIKV strains MR766 and PRVABC59
measured in this study may be a consequence of genetic variation among ZIKV strains
or differing patterns of NAb recognition of ZIKV replicating in Vero cells; maturation
of virus may also affect neutralisation. The membrane protein (prM) content of virions
may contribute to cell-type-dependent patterns of antibody neutralization (369, 373-
375).
Further epitope analysis and antibody repertoire analysis, beyond the scope of the
present study, will be undertaken as a future research to investigate differential ZIKV-
strain-specific antibody responses.
6.4.5. Virus protein binding analysis by immunoblot
The most common flavivirus antigenic target, the E protein, has more than 50% amino
acid homology between DENV and ZIKV. The E protein plays a central role in virus
entry and is major target of neutralising antibodies and therefore of vaccine
development. Other potential differential binding sites include prM and NS1 protein
(376-378).
This study aimed to assess if it was possible to differentiate DENV ZIKV binding sites
recognised by anti-ZIKV and anti-DENV polyclonal antisera. Dot blot followed by
Western immunoblot was undertaken to identify antisera binding sites against flavivirus
lysates. The 2 methods did not always align - non-specific binding was not seen against
mock cell lysate for any of the test sample in dot blot analysis while bands were visible
in mock cell lysate when electrophoresed through SDS-PAGE and blotted. Furthermore,
two anti-DENV antisera did not show any binding to ZIKV protein lysate in dot blot but
bands were seen WB after SDS-PAGE.

292
Antisera from two confirmed monotypic ZIKV infections (FLV-064/FLV090) bound to
ZIKV capsid/prM protein in both strains. E protein bands were seen for all flaviviruses.
Intensity of ZIKV-specific band at 22-11KDa varied for different antisera although the
amount of protein lysate was consistent throughout. Similarly, ZIKV capsid/prM protein
binding alone was seen in antisera from an individual with ZIKV infection and with
history of DENV infection (FLV-087); the E-protein band was of high intensity. This
individual’s DENV infection was thought to occur in 1990, 25 years before her ZIKV
infection. Anti-DENV NAb titres ranged from 46 – 640 across the four serotypes; lack
of anti-DENV antibody binding to DENV lysates may be related to assay sensitivity.
In summary this study demonstrated that antibody binding to ZIKV capsid/prM protein
can differentiate recent ZIKV infection, in individuals with previous DENV infection.
There are no reports of ZIKV capsid/prM protein-specific binding in assessment of Zika
serodiagnosis.
Differential binding to other proteins has been demonstrated. NS1 was shown to
distinguish between serogroups (DENV and WNV) (359); anti-prM antibody cross-
reacted among viruses of the same serogroup (WNV and SLEV). In the current
immunoblot system, NS1 was not visualised. Cardosa and others have shown that prM
recognition may serologically differentiate individuals infected with JEV and DENV
(376).
In the present study all antisera bound to all viral E proteins, with varying intensity.
Highly cross-reactive antibodies induced by flavivirus infection recognise heterologous
E proteins; cross-reactive E-protein was shown in the analysis of WNV, SLEV and
DENV (359) and thus recognition of E is not useful for differentiation of infecting
flavivirus.

293
Validation of the preliminary finding that ZIKV capsid/prMprotein binding can
discriminate recent ZIKV infection in the setting of previous DENV infection is
ongoing. Technical enhancements include protein (virus lysate; primary antibody)
purification. This study provides a baseline from which to explore ZIKV and DENV
protein-specific serodiagnostic approaches.
6.5. Research outcomes
Differential neutralisation between the prototype and recent strains of ZIKV was
seen.
Anti-ZIKV cross-reactivity against DENV was seen only in acute, past DENV
exposure and individuals with history of living or frequent travel to DENV
endemic areas.
Not all post ZIKV immune response is cross reactive to DENV or other
flavivirus vaccine strain.
Persistence anti- ZIKV NAb was observed to be associated with the history of
past heterologous flavivirus-specific immunity.
Possible capsid/prM protein based serological differentiation between DENV
and ZIKV.

294
Chapter 7: - Enhancement of ZIKV infection by monotypic anti-DENV antibody
7.1. Preamble
This chapter assessed the capacity for anti-DENV antibodies to enhance ZIKV
infection. Anti-DENV antisera were obtained from WA travellers with well-defined
monotypic DENV infection with all 4 serotypes. Acute and convalescent antisera were
analysed. Pools of monotypic sera representing each DENV serotype were included to
emulate ZIKV infection in populations with hyperendemic DENV transmission. The
potential risk of developing severe dengue, for WA travellers with monotypic DENV
infection and who may be reinfected during subsequent travel is not understood.
Previous studies of ZIKV enhancement have been based in dengue endemic countries
with high seroprevalence and where the sequence of infections is not known, whereas
our study population allowed pre-existing DENV immunity to be defined.
7.2 Introduction
The humoral immune response to flavivirus infection is influenced by the presence of
pre-existing antibody to related flaviviruses. Suboptimal levels of neutralizing antibody
lead to poor viral clearance and these non-neutralizing antibodies can enhance the viral
replication thereby increasing disease severity (379). However, if the cross-neutralizing
antibodies are able to block infection this results in cross-protection. Thus, humoral
immune responses in consecutive flavivirus infection have two possible outcomes: -
protection or increased disease severity. The best studied mechanism is antibody
dependent enhancement (ADE).
ADE is an immunological complication reported in several viral infections including
DENV, HIV, Influenza A, Coxsackie virus B, RSV, Ebola and also in also in protozoan
parasites like Leshmania (380). ADE also has been shown to be associated with

295
demographic and virulence factors such as age, sex, origin of population, past exposure
to infection and infecting strains (381).
7.2.1. Mechanism of ADE
ADE is based on the theory that the presence of non-neutralising antibody levels results
in opsonisation of immune complexes via Fc receptor (252). Ligation of monocyte or
macrophage with FcR receptor leads to biased immune responses enhancing
replication of infectious particles (247). Waning of primary anti-DENV Abs to the level
no longer sufficient to neutralise infection is thought to induce FcR-mediated
opsonisation. Molecular studies demonstrate FcR-mediated endocytosis triggers
cytoplasmic immuno-tyrosine activation motifs resulting in biased T-helper-1 (TH1)
response to TH-2 cells response resulting in increased infection. In THP-1 cells, after
ligation of FcRIa and FcRIIa, entry of infectious immune complexes activates
expression of the negative regulators dihydroxyaetone kinase and autophagy proteins
(cascade of down-regulatory pathways) that results in decreased production of type I
interferon-activated antiviral molecules supressing capacity of macrophage antiviral
mechanisms hence enhancing viral replication (247). Halstead et al have described two
principles of ADE: extrinsic ADE which results in FcR mediated entry of virus, and
intrinsic ADE that modifies innate and adaptive intracellular antiviral mechanisms and
enhances replication (247). Several studies have shown cross-linking of immune
complexes with FcR receptors increases cellular infectivity and results in disease
severity in dengue.
7.2.2. ADE in DENV and ZIKV infection
Anti-DENV antibodies induced in primary infection do not protect against secondary
infection with heterologous DENV (382, 383). Indeed, epidemiological observations

296
indicate that secondary infection is associated with more severe disease. ADE has been
proposed as a mechanism for disease severity in secondary infection with heterologous
DENV serotypes (384, 385). ADE has been demonstrated in vitro in FcR-receptor
bearing cell lines including U937, K562 and THP-1 (253, 254) and in vivo in non-
human primates (386).
During a 1981 DENV-2 epidemic in Cuba with 300,000 cases, 10,000 severe cases and
98% of fatalities occurred in individuals with secondary infection (249, 387). DHF in
infants during the first year of life is also associated with ADE (388). A strong
correlation between passively transferred maternal neutralizing antibody and DHF in
infants has been reported (389). Maternal antibodies are protective during the first few
months, however waning antibody titres to non-neutralizing concentrations is proposed
to induce enhancement of DENV infection in the infant (389). In a study of 20 infants
infected with DENV-2 below age of 1, 8 showed DHF/DSS and the predominance
antibody found in mothers was against DENV-1 (390). Another study done in 168
children with acute dengue aged 18 months to 14 years reported that patients with
secondary antibody responses to multiple DENV serotypes were twice likely to have
severe dengue compared to those with single infection (391). A well-characterized
paediatric cohort (2-14 years) study in Nicaragua showed that risk of severe dengue
disease is within a narrow range of pre-existing DENV antibody titres. This study shows
the risk of severe dengue in DENV-naïve children and children with high anti-DENV
antibody titres (>1:1280) was similar, while presence of 1:21 to 1:80 pre-existing
DENV-Ab increased hazard risk by 7.64-fold. Furthermore, in this paediatric cohort the
DENV-Ab half-life was estimated as 4 years, and by 3 years the decrease in Ab titre to
levels that were suggested to have exacerbated disease among 22% of children (381).
Due to these immunological complications, production of a vaccine that provides
equivalent protective immunity against all four DENV serotypes and thus does not

297
increase the risk of severe disease is a major challenge. The first dengue licensed
vaccine, Dengvaxia® (CYD-TDV) developed by Sanofi Pasteur has been licensed for
use in individuals 9-45 years living in endemic areas. Phase-III trials across Asia and
Latin America demonstrated effective immune responses for the first 25 months post
dose I while long-term follow-up raised concerns about risk of hospitalization. Among
30,000 participants aged 2-16 years vaccine efficacy against primary series was 59.2%
with 79.1% protection against severe dengue. Efficacy was questioned after
hospitalization and severe dengue in vaccine recipients, with elevated risk of
hospitalized dengue in 2-5-year olds. Vaccine efficacy in DENV seropositive
individuals at baseline was 78.2% and in individuals seronegative at baseline was
38.1% (392). Concerns have been raised that individuals of any age with no prior
dengue exposure could be at risk of hospitalization when infected with dengue post
vaccine. Even though Dengvaxia has the potential to reduce the burden of disease in
endemic areas use of the vaccine should be limited to individuals who have at least one
dengue virus infection and a simple test such as DENV-IgG was suggested to identify
seropositive individuals to be vaccinated (393, 394).
Recent studies have shown a high degree of similarity in DENV and ZIKV E-protein
structure. Dai et al (2016) reported structure of ZIKV-E at 2.0 Å resembles all the
known flaviviruses E-structure and has a unique positively charged patch adjacent to the
fusion loop region (355). Similarly, another study at 3.8 Å resolutions supports the
similarity between ZIKV and other Flaviviruses E-protein and reports ~10 amino acid
difference in E-glycoprotein that forms the icosahedral shell (395).
ZIKV shares 55.6% amino acid sequence identity with DENV, 46% with yellow fever
virus, 56.1% with Japanese encephalitis virus and 57% with West Nile virus (139).
54% E-protein sequence identity has been reported between DENV-2 and ZIKV (357).
Amino acid identity in DI, DII and DIII of E-protein of 35%, 51% and 29% between

298
ZIKV and DENV has been shown (365). This similarity is sufficient to allow cross-
reactive immune responses between DENV and ZIKV. Anti-E antibodies in DENV are
neutralising antibodies which also proposed to enhance infection in heterotypic
secondary infection. Possible cross-reactivity between anti-DENV and ZIKV has posed
significant hurdles in understanding pathogenic outcomes in populations with DENV
immune backgrounds. With the several reports of possible ZIKV enhancement in the
presence of pre-existing anti-DENV antibody, this study assessed possible potential
complications for WA travellers with history of DENV infection who may be
subsequently infected with ZIKV.

299
7.3. Results
U937 cell line (ATCC® CRL-1593.2™) was used as FcR receptor bearing cells to
detect enhancement of infection. This cell line is non-permissive to ZIKV. However, in
the presence anti-flavivirus antibody ZIKV has been reported to enter and replicate.
In this study, ZIKV PRVABC59 (Asian strain) enhancement by DENV antisera
obtained at different time points after presentation with febrile illness was tested.
Enhancement of ZIKV replication was determined by virus output detected by plaque
assay on Vero cell line. Virus production (as pfu/ml) for test antisera was compared to
virus production in the presence of the flavivirus-specific mAb 4G2 (Positive control) or
flavivirus-seronegative pooled sera (Negative control).
Mann-Whitney non-parametric t-test was used to define statistical significance of the
virus yield in terms of p-value. Difference in virus yield at different dilution for each
serum sample was compared with virus yield with positive and negative controls. The p-
values thus derived give the significance of difference (p<0.01) in enhancement. The
plasma sample with statistically significant difference from negative and insignificant
difference from positive (showing same enhancement as positive) is considered as
enhancing. Samples statistically closer to negative (insignificant p-value) are considered
non-enhancing. This statistical analysis was done to confirm enhancement as viral
output was not high compared with positive control (396-399).
Fit spline/LOWESS was used to generate curves to represent change in pfu/ml at
different plasma dilutions. Statistical analysis was done using GraphPad Prism 7.0.
Virus output from U937 cells after exposure of ZIKV in the presence of anti-DENV
antisera is listed in Table 7.2.
Six different combinations of monotypic anti-DENV antisera, and positive and negative
controls are listed in table 7.1.

300
Table 7. 1:- Description of plasma samples used in ADE
Sample group Description Infecting serotype/
Study number (MPI)
Sample ID
used in ADE
Control 4G2 monoclonal antibody from
hybridoma cell line as positive
control
4G2
Pool of three flavivirus negative
plasma samples as negative control
Negative
DENV acute plasma i) pool of plasma samples less than
two months post infection
DENV-1:-FLV-011
0.3), FLV-017(2), FLV-
018(2)
DENV-2 -FLV-012(0.1)
DENV <2
MPI
ii) Plasma sample of one of the acute
patient
FLV-018 (2) DENV-1-018
DENV convalescent
plasma pool
pool of plasma sample at 1-6 years
post infection
DENV-1 FLV 018 (31)
/FLV 001 (60)
DENV-2- FLV 014
(36)/ FLV004 (51)
DENV-3-FLV 029 (12),
FLV 030 (12)
DENV-4 -FLV 049 (14)
FLV 002 (60)
DENV >12
MPI
DENV-1 plasma DENV-1 plasma post 12 months
post infection
FLV-022 (14)
FLV-026-(14)
DENV-1-022
DENV-1-026
DENV-1 plasma sample at 6 months
post infection
FLV-076(6)
FLV- 060(6)
DENV-1-076
DENV-1- 060
DENV-3 plasma DENV-3 plasma post 12 months
post infection
FLV-038(14)
FLV-037(14)
DENV-3-038
DENV-3-037
DENV-3 plasma at 6 months post
infection
FLV-050(6)
FLV-056(6)
DENV-3-050
DENV-3-056
Pooled sera from
DENV-1 and 3
patients
Pool of DENV-1 and DENV-3 six
months post infection
FLV-076/FLV- 060+
FLV-050, FLV-056
DENV 1&3 (6
MPI)
Pooled sera from
DENV-1 and 3
patients
Pool of DENV-1 and DENV-3 one-
year post infection
FLV-022/FLV-
026+FLV-038, FLV-
037
DENV 1&3
(12 MPI)

301
Table 7. 2:- Virus output from U937 post exposure to respective anti-DENV - ZIKV immune complex
Sample group Sample ID* Virus yield at plasma dilution (pfu/ml) Average
pfu/ml
p-value
compared
with
positive
control
p-value
compared
with
negative
control 10^1 10^2 10^3 10^4 10^5 10^6
Controls 4G2 monoclonal (+ve) 7.50 x10^6 2.20 x10^6 9.00 x10^5 4.00 x10^5 1.00 x10^5 5.00 x10^3 1.85 x10^6
Negative 0 0 0 0 0 0 0
DENV acute
plasma
DENV <2 MPI 1.10 x10^4 3.30 x10^4 1.70 x10^4 1.40 x10^4 1.10 x10^4 1.30 x10^3 1.46 x10^4 0.036 0.0022
DENV-1-018 3.80 x10^5 5.10 x10^5 4.20 x10^5 4.90 x10^5 2.20 x10^5 2.00 x10^4 3.40 x10^5 0.2944 0.0022
DENV
convalescent
plasma pool
DENV >12 MPI 1.60 x10^4 1.50 x10^4 1.20 x10^4 5.00 x10^2 0 0 7.25 x10^3 0.0152 0.0152
DENV-1 plasma
DENV-1-022 3.90 x10^4 2.70 x10^4 2.80 x10^3 4.50 x10^3 0 0 1.22 x10^4 0.0087 0.0606
DENV-1-026 5.10 x10^3 2.70 x10^3 5.00 x10^2 0 0 0 1.38 x10^3 0.0043 0.1818
DENV-1-076 7.60 x10^2 0 0 0 5.20 x10^2 0 2.13 x10^2 0.0022 0.4545
DENV-1- 060 0 0 0 0 0 0 0
DENV-3 plasma
DENV-3-50 5.00 x10^2 6.00 x10^1 0 0 0 0 9.33 x10^1 0.0022 0.4545
DENV-3-056 2.00 x10^2 0 0 0 0 0 3.33 x10^1 0.0022 0.4545
DENV-3-038 1.50 x10^4 1.70 x10^4 3.70 x10^4 1.90 x10^3 5.00 x10^2 0 1.19 x10^4 0.0043 0.4545
DENV-3-037 4.40 x10^3 4.60 x10^2 0 0 0 0 8.10 x10^2 0.0152 0.0152
Pooled sera from
DENV-1 and 3
patients
DENV 1&3 (6 MPI) 1.30 x10^2 0 0 0 0 0 2.17 x10^1 0.0022 0.4545
DENV 1&3 (12 MPI) 3.10 x10^3 2.50 x10^4 0 0 0 0 4.68 x10^3 0.0043 0.4545
Note: - p<0.01 shows statistically significant distance compared with positive and negative control
Significant difference in virus output between positive control and sample: - no-enhancement
Significant difference in virus output between negative control and sample:- Enhancement
Antisera that significantly enhanced ZIKV infection are coloured in grey; non-conclusive results with significant difference from both positive and negative control are highlighted in light grey

302
7.3.1. Acute phase plasma: DENV plasma sample within two months post infection
Acute phase anti-DENV antisera collected less than 2 months after infection enhanced
ZIKV infection. Differential enhancement was seen for individual monotypic antisera
and for the pooled antisera with variable virus yield, while enhancement by both
samples was statistically significant (p=0.2944 and p=0.036). Virus output was greater
than for negative control in all cases. Acute phase plasma from a patient with
neutralising antibody against ZIKV MR766 showed greater enhancement of viral
output.
This group of anti-DENV antisera includes a pool of four plasma samples obtained <2
months post infection: 3 DENV-1 (FLV-011, FLV-017, FLV-018) and 1 DENV-2
(FLV-012) antisera. None of the antisera neutralised ZIKV PRVABC59, however anti-
ZIKV MR766 FRNT50 was > 50. No patients were infected with ZIKV and there was
no ZIKV PRVABC59-specific neutralisation with antisera collected < 2 months or at
any subsequent time.
The 2 acute phase test samples enhanced ZIKV production while no virus was produced
with the negative control pooled serum. Virus production was enhanced at 1:100
dilution of the acute pooled antiserum and the individual antiserum (FLV018). Virus
production by FLV018 antiserum was 5-fold lower than for positive control while yield
with the pooled acute antisera was more than 100-fold less but was still statistically
significant. Similarly, virus output following exposure to FLV018 antiserum was 15-
fold higher than for pooled acute antisera.

303
7.3.2. ZIKV enhancement by convalescent anti-DENV antisera collected 1-6 years post
infection
ZIKV production by pooled convalescent antisera was significantly lower than positive
control. This pool consisted of 8 anti-DENV antisera collected 1 – 6 years post
infection. Peak virus replication was obtained at the lowest antiserum dilution (1:10). At
higher dilutions of plasma (1x10^5 and 1x10^6) no viral replication was detected.
Average titre (pfu/ml) across dilutions with virus output was higher than for negative
control and was > 200-fold lower than for positive control, but the difference was not
statistically significant for either (p=0.0152).
Enhancement by pooled convalescent antisera is 2-fold lower than for acute phase
antisera. Viral yield at different plasma dilutions is illustrated in Figure 7. 1-A
Enhancement by pooled acute and pooled convalescent antisera, and positive and
negative controls, is shown in Figure 7. 1-B.
7. 3.3. DENV-1 plasma samples
No anti-DENV-1 antisera induced enhancement of replication of ZIKV PRVABC59.
Four DENV-1 plasma samples were tested representing six months and 12 months post
infection (MPI). Of two anti-DENV-1 antisera at 6 months, one induced virus yields at
10 and 10^5 dilution while no viral production was detected for the other antisera. Virus
output was obtained in both samples at 12MPI. Average pfu/ml for DENV-1-022
antisera was 9-fold higher than for anti-DENV-1-026 antisera.
Overall, anti-DENV-1 at 12 MPI had higher virus yield compared to anti-DENV at six
months post infection. Viral output in all cases was significantly lower (p=0.0022-
0.0087) than for 4G2 with more than 200-fold less virus production. Enhancement of

304
ZIKV infection was statistically comparable to negative control (p=0.0606-0.04545).
Viral output at different plasma dilution is shown as spline curve in Figure 7. 2(A) and
Enhancement by Anti-DENV-1 antisera, and controls, is shown in Figure 7. 2(B).
7.3.4. DENV-3 plasma samples
Similar to anti-DENV-1, no anti-DENV-3 antisera induced statistically significant
enhancement of ZIKV PRVABC59. This sample set consists of 4 anti-DENV-3 antisera
at six and 12 MPI. Compared to positive control, viral output is more than 200-fold less
for anti-DENV-3 antisera collected 12MPI, and more than 1000-fold less for anti-
DENV-3 antisera collected six MPI. Although there was virus output, it was not
statistically significant compared to negative control and thus, enhancement was not
induced. Viral output at different plasma dilution is shown as spline curve in Figure7.
3(A) and Figure7. 3 (B) Enhancement by Anti-DENV-3 antisera, and controls, is shown
in Figure 7. 3(B).
7.3.5. Pooled sera from DENV-1 and DENV-3 patients
a) At six months post infection
A pool of anti-DENV-1 and anti-DENV-3 antisera collected 6MPI did not induce
enhancement of ZIKV replication. Average pfu/ml was 4-fold lower compared to virus
output with individual antisera tested. Maximum yield with this pool was 130 pfu/ml
which was not statistically significant compared to 4G2.
b) At 12 months post infection
No statistically significant enhancement of infection by pooled DENV-1 and DENV-3
antisera obtained at 12 MPI. Maximum virus yield was obtained at 1:100 dilution. At
this dilution, viral output was higher than enhancement induced by 3 of 4 individual

305
antisera. Virus output with this pool was more than 400-fold less compared to 4G2.
Viral output was equivalent to negative control (p=0.4545).
Virus output at different dilution of this pool at 6 and 12 MPI is shown in Figure 7. 4
(A) and compared with positive and negative control in Figure 7. 4 (B).

306
Figure 7. 1:-ZIKV enhancement by monotypic anti-DENV pool at different stage post infection.
A) Spline curve of ZIKV output in the presence of acute phase anti-DENV antisera
(DENV<2MPI and DENV-1-018), convalescent phase anti-DENV antisera
(DENV>12MPI), 4G2 and negative pool. Highest viral output was induced by 4G2 and
lowest output was induced by convalescent antisera.
B) Viral output in the presence of anti-DENV antibody. Difference in average pfu/ml
between acute and convalescent anti-DENV antisera was not statistically significant by
Mann-Whitney non-parametric t-test. Virus production was significant for acute phase
antisera, and for convalescent antisera enhancement was inconclusive as all dilutions did not
induce virus replication.
A
B

307
A
B
Figure 7. 2:-ZIKV enhancement by anti-DENV-1 antisera
A) Spline curve of viral yield at different plasma dilutions of DENV-1 vs positive and
negative control. One of the DENV-1 at 6MPI antisera did not induce any infection
and a second anti-DENV-1 antisera showed higher virus replication at lower dilution.
Both DENV-1 at 12MPI showed yield of ZIKV.
B) No significant difference in enhancement by anti-DENV-1 antisera collected at 6
and 12 MPI. Viral output is higher than for negative control not statistically significant
as not all dilutions induced enhancement.

308
B
A
B
Figure7. 3:-ZIKV enhancement by anti-DENV-3 antisera
A) Spline curve of viral output at different plasma dilution of anti-DENV-3 antisera at 6
months and 12 months post infection. No antisera enhanced ZIKV infection.
B) No anti-DENV-3 antisera collected 6 and 12 MPI enhanced ZIKV infection.

309
B
Figure 7. 4 :-ZIKV enhancement based on French Polynesian co-circulation model.
A) Pooled anti-DENV-1 and anti-DENV-3 antisera collected 6 and 12 MPI did not enhance
ZIKV infection.
B) ZIKV production in the presence of pooled anti-DENV-1 and anti-DENV-3 antisera, 4G2
and negative control. Virus output is greater for 12 MPI antisera, compared to 6 MPI antisera,
but these values were not statistically significant compared to positive control.
A

310
7.4. Summary discussion
Re-emergence of ZIKV and spread into DENV-endemic areas has raised concerns about
possible immunopathological outcomes due to pre-existing anti-DENV antibodies.
Severe dengue is postulated to be a consequence of anti-DENV antibody-mediated
immune enhancement. Enhancement of ZIKV infection in the setting of pre-existing
anti-DENV immunity is postulated as a mechanism for the severe Zika disease that
emerged in the recent epidemics beginning in 2007. This study assessed the capacity of
well-defined anti-DENV antisera to enhance ZIKV replication in vitro. In addition, the
sequence of ZIKV circulation in French Polynesia in 2013, in the background of prior
DENV-1 and DENV-3 epidemics, was modelled to determine if anti-DENV-1 and anti-
DENV-3 antisera enhanced ZIKV infection.
7.4.1. DENV and ZIKV in French Polynesia
Transmission of mosquito-borne viruses is well documented in the Pacific. A pattern
of single DENV serotype dominance has been reported in many South Pacific island
nations. The displacement of the circulating serotype by newly introduced DENV has
been regularly observed over time (118). Epidemiological studies report periodic cycles
of 12 years of DENV and 20 years of the other 3 serotypes (400). The Pacific had
predominant circulation of DENV-1 with co-circulation of DENV-4 in 2007 along with
DENV-2 sporadic outbreaks followed by DENV-3 in 2013 (327). DENV-1 was the
documented serotype in the first recorded dengue outbreak in French Polynesia (FP) in
the 1940s. After 20 years dengue reappeared with DENV-3 as dominant serotype which
was followed by the first cases of severe dengue in the 1970s. In 2009 after decades of
circulation of DENV-1, DENV-4 reappeared in FP after 20 years (401). In March 2013
after 18 years of absence, DENV-3 re-emergence was reported in FP (327). ZIKV was
introduced into FP while DENV-3 was circulating in 2013. Following re-emergence of

311
ZIKV in Yap state in the Western Pacific in 2007 the largest ZIKV outbreak in the
Pacific occurred in FP (132). Later ZIKV infection was confirmed with sequencing
which showed ZIKV strain Cambodia as the closet strain. By 10 weeks of first case
reported 5,895 patients with suspected ZIKV infections were recorded and 294 patients
were ZIKV RT-PCR confirmed cases(132). At the same time the evidences of
concomitant circulation of ZIKV, DENV-1, and DENV-3 and increase in GBS also
raised question of impact of sequential infections and disease outcome (132).
7.4.2. ZIKV enhancement by monotypic anti-DENV from travellers
Given circulation of both ZIKV and DENV in Indonesia, presence of DENV immune
response background in WA travellers and high chances re-visit to endemic areas
influenced us to analyse possibility of DENV-1 and DENV-3 primed enhancement of
ZIKV. Furthermore, the fact that DENV-3 isolated in traveller’s population in our
cohort belongs to genotype-I encouraged us to start preliminary research on possibility
of ADE in case of non-endemic population with monotypic DENV infection based on
Pacific Island outbreak pattern. We based our study on this model to test whether either
of these serotype plasma samples from travellers may have potential to enhance ZIKV
replication in macrophages.
In this study, role of anti-DENV sera from monotypic infection to enhance ZIKV Asian
strain in vitro tested. Intrinsic ADE was tested to presume possibility of infection and
replication of virus in the presence of serum sample. Analysing total viral load output
from U937 FcR bearing cell line in the presence and absence of anti-DENV antisera
showed not all post DENV sera sample enhances viral replication. Effects of time post
infection were also studied. Viral output with acute samples pool (DENV1 and 2) was
higher than convalescent sample pool (1-6 years DENV1-4). Viral output was higher
compared to the negative tested in same experiment however was >100 fold less than

312
positive control and this difference was statistically significant. Interestingly, DENV-1
individual (FLV018) who had cross reactivity with prototype ZIKV at acute and
convalescent phase showed significant enhancement of ZIKVPRVABV59. This
individual has no cross-reactivity against ZIKVPRVABC59 when tested with
neutralisation. In vitro study showed not all DENV 1 and 3 could enhance production of
viral load, in fact some of the sera did not support viral production post exposure. No
statistical significant difference (p>0.05) was obtained between individual samples and
pooled sera sample. Although there was yield of virus in the tested samples, but the
output was >1000 fold less than positive control hence statistically closer to negative
pool. Thus, based on this pilot study on viral output post exposure with DENV-1 and 3
does not support increase in viral load in the presence of anti-DENV antibody. While
there are also reports not all anti-DENV antibody supports the enhancement of ZIKV.
However, it cannot be denied that this study includes only limited size. Differential
pattern of ADE among the samples can be explained by variable epitope sensitivity of
anti-DENV in different individuals. Epitope sensitive binding has been previously
studied in mAbs derived studies. Monoclonal antibodies derived post DENV infection
shown to selectively bind to epitope footprint that spans certain E-protein dimers (402)
Dejnirattisai et al showed DENV derived mAbs that bind to linear epitopes were able to
promote ADE (260).
In our study, significant enhancement of ZIKV viral output from U937 cell line when
exposed to virus in the presence of 4G2 monoclonal was found. A related paper showed
similar enhancement of ZIKV prototype strain by 4G2 on THP-1 cell line (403). In
contrast to our finding in non-endemic population, enhanced infection of ZIKV
PRVABC59 in FcR receptor bearing cell was reported from cases in Thailand(357). In
the study form patients in Thailand both acute/convalescent sera and plasmablast-

313
derived mAbs were reported to enhance infection. Similarly, enhancement of ZIKV by
DENV derived mAbs and plasma samples have reported in several studies (155, 257).
This is the initial phase study to generate background for possible DENV-primed ZIKV
infection in travellers hence providing important information about possible
consequences in non-endemic population. However, this study needs more data to
predict the possible outcomes of secondary ZIKV in infected traveller population.
Further this report is based on the yield of infectious virus from Fc-R bearing cells in
the presence and absence of anti-DENV antibody, still the data about enhancement of
infection needs to be analysed with molecular PCR or flow-based techniques to predict
possible consequences of pre-DENV exposure in secondary ZIKV infection in
population from non-endemic regions.
7.5. Research outcomes
Acute phase anti-DENV enhances ZIKV infection-monotypic anti-DENV-1 or
pooled acute DENV-1+DENV-2.
No convalescent (1-6 years) anti-DENV (all four) serotypes enhanced ZIKV
infection.
Anti-DENV-1 and 3 combinations did not prime ZIKV infection to U937 cells.
ADE was greatly dependent on time post infection and individual’s immune
response.

314
Chapter 8:- Summary and conclusion
Flavivirus-specific antibody responses were assessed in West Australian travellers
returning from dengue and Zika endemic or epidemic countries. The majority of study
volunteers had never lived in these countries and had well-defined monotypic infection
with DENV or ZIKV; some of them had received YFV vaccination. Dengue and Zika
do not occur in WA as the mosquito vectors are not present. Magnitude of neutralising
antibody, measured by focus reduction neutralisation test (FRNT); total antibody,
measured by haemagglutination inhibition (HI) test; as well as antibody mediated
enhancement (ADE) were assessed for antisera collected from study volunteers who
were sampled during acute phase (< 2months) up to 6 years after presentation for febrile
illness. The WA Traveller Study is not a true prospective cohort and includes
convenience samples obtained from volunteers at variable time intervals. Antisera were
tested against a collection of DENV isolates derived from travellers presenting with
febrile illness and who were diagnosed with DENV infection; the virus collection
includes representatives of all four serotypes collected between 2010 – 2015. This
collection is dependent on DENV circulating in the Asia Pacific region at the time and
thus is biased towards contemporaneous endemic or epidemic isolates, predominantly
DENV-1 and DENV-2. Phylogenetic analysis identified genotypes and lineages, and
representative isolates were used as targets. Other travellers returned from Zika
epidemic countries with confirmed or probable ZIKV infection; some had lived in
dengue endemic countries previously.
Anti-DENV responses were highly cross-reactive during acute phase and neutralized
heterologous DENV at high magnitude. Over time and up to six years after infection
responses became increasingly serotype-specific –this could be visualised on antigenic
maps - however heterologous cross-neutralisation was maintained in all individuals to

315
some degree, even at 6 years, and was virus- and patient-specific. Virus-specific
neutralisation was not genotype or lineage dependent; individual isolates were
differentially neutralised by antisera from individuals collected at the same time point
after infection. Antigenic mapping showed serotype clustering could be distinguished
after 1 year; however individual viruses were as close to heterologous serotype clusters
as they were to homologous clusters and this pattern persisted at 6 years. A second
major observation was that the magnitude of responses to homologous DENV in these
monotypic travellers also declined over time, including responses to autologous virus,
and this was true for all 4 serotypes.
The magnitude of ZIKV-specific neutralising antibody responses declined over time in
the small number of study volunteers with monotypic ZIKV infection. By 13 and 20
months after infection FRNT90 titres were below limits of detection (<40 as per WHO
criteria for sero diagnosis of ZIKV infection). However, ZIKV-infected individuals with
previous flavivirus infection showed prolonged anti-ZIKV antibody responses, up to 13
months after infection. This study demonstrated that antibody binding to ZIKV
capsid/prM protein can differentiate recent ZIKV infection, in individuals with previous
DENV infection.
Two ZIKV strains were assessed and showed a high degree of differential cross-
neutralisation, with high magnitude responses directed against the prototype ZIKV
MR766 isolate but not the contemporaneous ZIKV PRVABC59 in acute phase
monotypic DENV infection, highlighting the importance of strain selection in ZIKV
diagnostic and research approaches.
In the small number of antisera tested in the present study, ZIKV replication was
enhanced by acute phase (< 2months) antisera only. This finding contrasts with previous
reports of ZIKV enhancement by convalescent antisera, however those studies used

316
immune sera collected within 100 days of infection; convalescent antisera in the present
study were collected between 6 months and 6 years post presentation.
A major limitation of this study is the small sample size and the convenience sampling
of antisera dictated by this traveller cohort. Enrolment into the study is ongoing and
more recently travellers have returned with more diverse DENV infections, including
DENV-3 and DENV-4 which have not circulated as widely in the past. In addition,
ongoing molecular epidemiological analysis of traveller viruses has begun to delineate
genotypes and lineages and this information will be very informative for ongoing
phylogeographic antigenicity studies.
Future research directions will focus on characterisation of the anti-flavivirus B cell
repertoire; data from this present study will form the basis of this work.

317
Chapter 9:- References
1. Peeling RW, Artsob H, Pelegrino JL, Buchy P, Cardosa MJ, Devi S, et al. Evaluation of diagnostic tests: dengue. Nature reviews Microbiology. 2010;8(12 Suppl):S30-8. 2. Kuhn RJ, Zhang W, Rossmann MG, Pletnev SV, Corver J, Lenches E, et al. Structure of dengue virus: implications for flavivirus organization, maturation, and fusion. Cell. 2002;108(5):717-25. 3. Lok S-M. The Interplay of Dengue Virus Morphological Diversity and Human Antibodies. Trends in Microbiology. 2016;24(4):284-93. 4. Pierson TC, Xu Q, Nelson S, Oliphant T, Nybakken GE, Fremont DH, et al. The stoichiometry of antibody-mediated neutralization and enhancement of West Nile virus infection. Cell host & microbe. 2007;1(2):135-45. 5. Cedillo-Barrón L, García-Cordero J, Bustos-Arriaga J, León-Juárez M, Gutiérrez-Castañeda B. Antibody response to dengue virus. Microbes and Infection. 2014;16(9):711-20. 6. Laboratory Guidance and Diagnostic Testing [Internet]. Available from: https://www.cdc.gov/dengue/clinicallab/laboratory.html. 7. Halstead SB. Dengue. The Lancet. 2007;370(9599):1644-52. 8. Kuno G, Chang G-JJ, Tsuchiya KR, Karabatsos N, Cropp CB. Phylogeny of the Genus Flavivirus. Journal of virology. 1998;72(1):73-83. 9. Chen R, Vasilakis N. Dengue — Quo tu et quo vadis? Viruses. 2011;3(9):1562-608. 10. Perera R, Kuhn RJ. Structural Proteomics of Dengue Virus. Current opinion in microbiology. 2008;11(4):369-77. 11. Ernst T, McCarthy S, Chidlow G, Luang-Suarkia D, Holmes EC, Smith DW, et al. Emergence of a New Lineage of Dengue Virus Type 2 Identified in Travelers Entering Western Australia from Indonesia, 2010-2012. PLOS Neglected Tropical Diseases. 2015;9(1):e0003442. 12. Weaver SC, Vasilakis N. Molecular evolution of dengue viruses: Contributions of phylogenetics to understanding the history and epidemiology of the preeminent arboviral disease. Infection, Genetics and Evolution. 2009;9(4):523-40. 13. Lanciotti RS, Lambert AJ, Holodniy M, Saavedra S, Signor Ldel C. Phylogeny of Zika Virus in Western Hemisphere, 2015. Emerg Infect Dis. 2016;22(5):933-5. 14. Mukhopadhyay S, Kuhn RJ, Rossmann MG. A structural perspective of the flavivirus life cycle. Nat Rev Micro. 2005;3(1):13-22. 15. Kuno G, Chang GJ, Tsuchiya KR, Karabatsos N, Cropp CB. Phylogeny of the genus Flavivirus. Journal of virology. 1998;72(1):73-83. 16. Groen J, Koraka P, Velzing J, Copra C, Osterhaus AD. Evaluation of six immunoassays for detection of dengue virus-specific immunoglobulin M and G antibodies. Clinical and diagnostic laboratory immunology. 2000;7(6):867-71. 17. Calisher CH, Karabatsos N, Dalrymple JM, Shope RE, Porterfield JS, Westaway EG, et al. Antigenic relationships between flaviviruses as determined by cross-neutralization tests with polyclonal antisera. J Gen Virol. 1989;70 ( Pt 1):37-43. 18. Duane J. Gubler EEO, Subhas Vasudevan, Jeremy Farrar. Dengue and Dengue Hemorrhagic fever. 19. Scaturro P, Cortese M, Chatel-Chaix L, Fischl W, Bartenschlager R. Dengue Virus Non-structural Protein 1 Modulates Infectious Particle Production via Interaction with the Structural Proteins. PLOS Pathogens. 2015;11(11):e1005277. 20. Miller S, Kastner S, Krijnse-Locker J, Bühler S, Bartenschlager R. The Non-structural Protein 4A of Dengue Virus Is an Integral Membrane Protein Inducing Membrane Alterations in a 2K-regulated Manner. Journal of Biological Chemistry. 2007;282(12):8873-82. 21. El Sahili A, Lescar J. Dengue Virus Non-Structural Protein 5. Viruses. 2017;9(4). 22. Luo D, Vasudevan SG, Lescar J. The flavivirus NS2B-NS3 protease-helicase as a target for antiviral drug development. Antiviral Res. 2015;118:148-58. 23. Halsed SB. Dengue. Hoffman GPaSL, editor: Imperial college Press; 2008.

318
24. A AN, Berlioz-Arthaud A, Chow V, Endy T, Lowry K, Mai le Q, et al. Sustained transmission of dengue virus type 1 in the Pacific due to repeated introductions of different Asian strains. Virology. 2004;329(2):505-12. 25. Katoh H, Okamoto T, Fukuhara T, Kambara H, Morita E, Mori Y, et al. Japanese Encephalitis Virus Core Protein Inhibits Stress Granule Formation through an Interaction with Caprin-1 and Facilitates Viral Propagation. Journal of virology. 2013;87(1):489-502. 26. Rey FA, Heinz FX, Mandl C, Kunz C, Harrison SC. The envelope glycoprotein from tick-borne encephalitis virus at 2 A resolution. Nature. 1995;375(6529):291-8. 27. de Wispelaere M, Yang PL. Mutagenesis of the DI/DIII Linker in Dengue Virus Envelope Protein Impairs Viral Particle Assembly. Journal of virology. 2012;86(13):7072-83. 28. Shrestha B, Brien JD, Sukupolvi-Petty S, Austin SK, Edeling MA, Kim T, et al. The Development of Therapeutic Antibodies That Neutralize Homologous and Heterologous Genotypes of Dengue Virus Type 1. PLOS Pathogens. 2010;6(4):e1000823. 29. Crill WD, Roehrig JT. Monoclonal Antibodies That Bind to Domain III of Dengue Virus E Glycoprotein Are the Most Efficient Blockers of Virus Adsorption to Vero Cells. Journal of virology. 2001;75(16):7769-73. 30. WHO. Dengue guildelines for diagnosis, treatment, prevention and control. 9 ed2009. 31. Chu JJH, Rajamanonmani R, Li J, Bhuvanakantham R, Lescar J, Ng M-L. Inhibition of West Nile virus entry by using a recombinant domain III from the envelope glycoprotein. Journal of General Virology. 2005;86(2):405-12. 32. Bressanelli S, Stiasny K, Allison SL, Stura EA, Duquerroy S, Lescar J, et al. Structure of a flavivirus envelope glycoprotein in its low-pH-induced membrane fusion conformation. The EMBO Journal. 2004;23(4):728-38. 33. Modis Y, Ogata S, Clements D, Harrison SC. A ligand-binding pocket in the dengue virus envelope glycoprotein. Proceedings of the National Academy of Sciences. 2003;100(12):6986-91. 34. Stiasny K, Bressanelli S, Lepault J, Rey FA, Heinz FX. Characterization of a Membrane-Associated Trimeric Low-pH-Induced Form of the Class II Viral Fusion Protein E from Tick-Borne Encephalitis Virus and Its Crystallization. Journal of virology. 2004;78(6):3178-83. 35. Dejnirattisai W, Wongwiwat W, Supasa S, Zhang X, Dai X, Rouvinski A, et al. A new class of highly potent, broadly neutralizing antibodies isolated from viremic patients infected with dengue virus. Nature immunology. 2015;16(2):170-7. 36. Fernandez-Garcia M-D, Mazzon M, Jacobs M, Amara A. Pathogenesis of Flavivirus Infections: Using and Abusing the Host Cell. Cell Host & Microbe. 2009;5(4):318-28. 37. Fibriansah G, Ng T-S, Kostyuchenko VA, Lee J, Lee S, Wang J, et al. Structural Changes in Dengue Virus When Exposed to a Temperature of 37°C. Journal of virology. 2013;87(13):7585-92. 38. Cockburn Joseph JB, Navarro Sanchez ME, Fretes N, Urvoas A, Staropoli I, Kikuti Carlos M, et al. Mechanism of Dengue Virus Broad Cross-Neutralization by a Monoclonal Antibody. Structure. 2012;20(2):303-14. 39. Stiasny K, Heinz FX. Flavivirus membrane fusion. Journal of General Virology. 2006;87(10):2755-66. 40. Dowd KA, Pierson TC. Antibody-mediated neutralization of flaviviruses: A reductionist view. Virology. 2011;411(2):306-15. 41. Elshuber S, Allison SL, Heinz FX, Mandl CW. Cleavage of protein prM is necessary for infection of BHK-21 cells by tick-borne encephalitis virus. J Gen Virol. 2003;84(Pt 1):183-91. 42. Limon-Flores AY, Perez-Tapia M, Estrada-Garcia I, Vaughan G, Escobar-Gutierrez A, Calderon-Amador J, et al. Dengue virus inoculation to human skin explants: an effective approach to assess in situ the early infection and the effects on cutaneous dendritic cells. International Journal of Experimental Pathology. 2005;86(5):323-34.

319
43. Diamond MS. Evasion of innate and adaptive immunity by flaviviruses. Immunol Cell Biol. 2003;81(3):196-206. 44. Pierson TC, Fremont DH, Kuhn RJ, Diamond MS. Structural Insights into the Mechanisms of Antibody-Mediated Neutralization of Flavivirus Infection: Implications for Vaccine Development. Cell Host & Microbe. 2008;4(3):229-38. 45. Throsby M, Geuijen C, Goudsmit J, Bakker AQ, Korimbocus J, Kramer RA, et al. Isolation and Characterization of Human Monoclonal Antibodies from Individuals Infected with West Nile Virus. Journal of virology. 2006;80(14):6982-92. 46. Dengue and dengue hemorrhagic fever. Gubler DJ, Ooi, E. E., Vasudevan, S., Farrar, J., editor2014. 47. Dowd KA, Jost CA, Durbin AP, Whitehead SS, Pierson TC. A Dynamic Landscape for Antibody Binding Modulates Antibody-Mediated Neutralization of West Nile Virus. PLOS Pathogens. 2011;7(6):e1002111. 48. Parren PW, Burton DR. The antiviral activity of antibodies in vitro and in vivo. Advances in immunology. 2001;77:195-262. 49. Nybakken GE, Oliphant T, Johnson S, Burke S, Diamond MS, Fremont DH. Structural basis of West Nile virus neutralization by a therapeutic antibody. Nature. 2005;437(7059):764-9. 50. Ayala-Nunez NV, Hoornweg TE, van de Pol DP, Sjollema KA, Flipse J, van der Schaar HM, et al. How antibodies alter the cell entry pathway of dengue virus particles in macrophages. Sci Rep. 2016;6:28768. 51. Mackenzie J. Wrapping things up about virus RNA replication. Traffic (Copenhagen, Denmark). 2005;6(11):967-77. 52. Gillespie LK, Hoenen A, Morgan G, Mackenzie JM. The Endoplasmic Reticulum Provides the Membrane Platform for Biogenesis of the Flavivirus Replication Complex. Journal of virology. 2010;84(20):10438-47. 53. Reid CR, Airo AM, Hobman TC. The Virus-Host Interplay: Biogenesis of +RNA Replication Complexes. Viruses. 2015;7(8):4385-413. 54. Zybert IA, van der Ende-Metselaar H, Wilschut J, Smit JM. Functional importance of dengue virus maturation: infectious properties of immature virions. J Gen Virol. 2008;89(Pt 12):3047-51. 55. Smith CE. The history of dengue in tropical Asia and its probable relationship to the mosquito Aedes aegypti. The Journal of tropical medicine and hygiene. 1956;59(10):243-51. 56. Vasilakis N, Weaver SC. The history and evolution of human dengue emergence. Advances in virus research. 2008;72:1-76. 57. Rico-Hesse R. Molecular evolution and distribution of dengue viruses type 1 and 2 in nature. Virology. 1990;174(2):479-93. 58. Rudnick A. Ecology of Dengue Virus. Asian Journal of Infectious Diseases. 1978:156-60. 59. Messina JP, Brady OJ, Scott TW, Zou C, Pigott DM, Duda KA, et al. Global spread of dengue virus types: mapping the 70 year history. Trends in Microbiology. 2014;22(3):138-46. 60. Sabin AB, Schlesinger RW. PRODUCTION OF IMMUNITY TO DENGUE WITH VIRUS MODIFIED BY PROPAGATION IN MICE. Science. 1945;101(2634):640-2. 61. Gubler DJ, Clark GG. Dengue/dengue hemorrhagic fever: the emergence of a global health problem. Emerging Infectious Diseases. 1995;1(2):55-7. 62. FE H. The 1897 epidemic of dengue in North Queensland Australasian Medical Gazette. 1898:98-107. 63. Dengue fever [Internet]. Available from: http://www.cdc.gov.tw/english. 64. Rosen L. Dengue in Greece in 1927 and 1928 and the pathogenesis of dengue hemorrhagic fever: new data and a different conclusion. The American journal of tropical medicine and hygiene. 1986;35(3):642-53. 65. CDC. Dengue epidemiology. Available from: http://www.cdc.gov/Dengue/epidemiology/index.html.

320
66. Viennet E, Ritchie SA, Williams CR, Faddy HM, Harley D. Public Health Responses to and Challenges for the Control of Dengue Transmission in High-Income Countries: Four Case Studies. PLoS Neglected Tropical Diseases. 2016;10(9):e0004943. 67. Kuno G. Emergence of the Severe Syndrome and Mortality Associated with Dengue and Dengue-Like Illness: Historical Records (1890 to 1950) and Their Compatibility with Current Hypotheses on the Shift of Disease Manifestation. Clinical Microbiology Reviews. 2009;22(2):186-201. 68. Mackenzie JS, la Brooy JT, Hueston L, Cunningham AL. Dengue in Australia. Journal of medical microbiology. 1996;45(3):159-61. 69. NNDSS. Australia’s notifiable disease status, 2014:Annual report of the National Notifiable Diseases Surveillance System. Annual report. 2014.
70. George F Lumley FHT. Dengue: Department of health, Commonwealth of Australia; 1943. 71. Katrina Knope CG. Increasing notifications of dengue in Australia related to overseas travel, 1991 to 2012. Communicable Diseases Intelligence. 2013; 37 E55-E9. 72. Australia. DoHW. Disease Watch 2012;6:5. Bali travel behind big spike in dengue fever. (Accessed February 2018). 73. Lindsay MDA, Jardine A, Giele C, Armstrong P, McCarthy S, Whittle A, et al. Investigation of the First Case of Dengue Virus Infection Acquired in Western Australia in Seven Decades: Evidence of Importation of Infected Mosquitoes? PLOS Neglected Tropical Diseases. 2015;9(9):e0004114. 74. Kurucz WPaN. A series of exotic mosquito detections at Darwin International Airport, Northern Territory between February 2015 and January 2016. The Northern Territory Disease Control Bulletin. December 2016;23:15-9. 75. 2017 AGDoHA. Response guide for exotic mosquito detections at Australian first ports of entry. (Accessed February 2018). 76. Australia DoHW. Notifiable Infectious Diseases Reports: Dengue fever notifications. (Accessed February 2018). 77. Bhatt S, Gething PW, Brady OJ, Messina JP, Farlow AW, Moyes CL, et al. The global distribution and burden of dengue. Nature. 2013;496(7446):504-7. 78. Dengue and severe dengue [Internet]. 2017. Available from: http://www.who.int/mediacentre/factsheets/fs117/en/. 79. Duane JG, Gary GC. Dengue/Dengue Hemorrhagic Fever: The Emergence of a Global Health Problem. Emerging Infectious Disease journal. 1995;1(2):55. 80. Moncayo AC, Fernandez Z, Ortiz D, Diallo M, Sall A, Hartman S, et al. Dengue Emergence and Adaptation to Peridomestic Mosquitoes. Emerging Infectious Diseases. 2004;10(10):1790-6. 81. Murray NEA, Quam MB, Wilder-Smith A. Epidemiology of dengue: past, present and future prospects. Clinical Epidemiology. 2013;5:299-309. 82. Gubler DJ. Dengue/dengue haemorrhagic fever: history and current status. Novartis Foundation symposium. 2006;277:3-16; discussion -22, 71-3, 251-3. 83. Steinhauer DA, Domingo E, Holland JJ. Lack of evidence for proofreading mechanisms associated with an RNA virus polymerase. Gene. 1992;122(2):281-8. 84. Li D, Lott WB, Lowry K, Jones A, Thu HM, Aaskov J. Defective Interfering Viral Particles in Acute Dengue Infections. PLoS ONE. 2011;6(4):e19447. 85. Chen R, Vasilakis N. Dengue--quo tu et quo vadis? Viruses. 2011;3(9):1562-608. 86. Stoddard ST, Morrison AC, Vazquez-Prokopec GM, Paz Soldan V, Kochel TJ, Kitron U, et al. The Role of Human Movement in the Transmission of Vector-Borne Pathogens. PLoS Neglected Tropical Diseases. 2009;3(7):e481. 87. Soto SM. Human migration and infectious diseases. Clinical Microbiology and Infection. 2009;15:26-8. 88. Bäck AT, Lundkvist Å. Dengue viruses – an overview. Infection Ecology & Epidemiology. 2013;3:10.3402/iee.v3i0.19839. 89. Rothman AL. Dengue: defining protective versus pathologic immunity. Journal of Clinical Investigation. 2004;113(7):946-51.

321
90. Vaughn DW, Green S, Kalayanarooj S, Innis BL, Nimmannitya S, Suntayakorn S. Dengue viremia titer, antibody response pattern, and virus serotype correlate with disease severity. J Infect Dis. 2000;181. 91. Goncalvez AP, Escalante AA, Pujol FH, Ludert JE, Tovar D, Salas RA, et al. Diversity and evolution of the envelope gene of dengue virus type 1. Virology. 2002;303(1):110-9. 92. Villabona-Arenas CJ, Zanotto PMdA. Worldwide Spread of Dengue Virus Type 1. PLOS ONE. 2013;8(5):e62649. 93. Aviles G, Meissner J, Mantovani R, St Jeor S. Complete coding sequences of dengue-1 viruses from Paraguay and Argentina. Virus Res. 2003;98(1):75-82. 94. Vasilakis N, Tesh RB, Weaver SC. Sylvatic Dengue Virus Type 2 Activity in Humans, Nigeria, 1966. Emerging Infectious Diseases. 2008;14(3):502-4. 95. Kumar SR, Patil JA, Cecilia D, Cherian SS, Barde PV, Walimbe AM, et al. Evolution, dispersal and replacement of American genotype dengue type 2 viruses in India (1956-2005): selection pressure and molecular clock analyses. J Gen Virol. 2010;91(Pt 3):707-20. 96. El-Kafrawy SA, Sohrab SS, Ela SA, Abd-Alla AM, Alhabbab R, Farraj SA, et al. Multiple Introductions of Dengue 2 Virus Strains into Saudi Arabia from 1992 to 2014. Vector borne and zoonotic diseases (Larchmont, NY). 2016;16(6):391-9. 97. Lanciotti RS, Lewis JG, Gubler DJ, Trent DW. Molecular evolution and epidemiology of dengue-3 viruses. J Gen Virol. 1994;75 ( Pt 1):65-75. 98. King C-C, Chao D-Y, Chien L-J, Chang G-JJ, Lin T-H, Wu Y-C, et al. Comparative analysis of full genomic sequences among different genotypes of dengue virus type 3. Virology Journal. 2008;5:63-. 99. Klungthong C, Zhang C, Mammen MP, Jr., Ubol S, Holmes EC. The molecular epidemiology of dengue virus serotype 4 in Bangkok, Thailand. Virology. 2004;329(1):168-79. 100. Durbin AP, Mayer SV, Rossi SL, Amaya-Larios IY, Ramos-Castaneda J, Eong Ooi E, et al. Emergence potential of sylvatic dengue virus type 4 in the urban transmission cycle is restrained by vaccination and homotypic immunity. Virology. 2013;439(1):34-41. 101. Cleland JB, Bradley B, McDonald W. On the transmission of Australian dengue by the mosquito Stegomyia fasciata2018. 179-84 p. 102. CDC. Dengue and the Aedes aegypti mosquito: National Center for Emerging and Zoonotic Infectious Diseases. Available from: http://www.cdc.gov/dengue/entomologyecology/. 103. Diallo M, Ba Y, Sall AA, Diop OM, Ndione JA, Mondo M, et al. Amplification of the Sylvatic Cycle of Dengue Virus Type 2, Senegal, 1999–2000: Entomologic Findings and Epidemiologic Considerations. Emerging Infectious Diseases. 2003;9(3):362-7. 104. Gubler DJ. Dengue and Dengue Hemorrhagic Fever. Clinical Microbiology Reviews. 1998;11(3):480-96. 105. Gratz NG. Critical review of the vector status of Aedes albopictus. Medical and veterinary entomology. 2004;18(3):215-27. 106. Lambrechts L, Scott TW, Gubler DJ. Consequences of the Expanding Global Distribution of Aedes albopictus for Dengue Virus Transmission. PLOS Neglected Tropical Diseases. 2010;4(5):e646. 107. Mitchell CJ, Miller BR. Vertical transmission of dengue viruses by strains of Aedes albopictus recently introduced into Brazil. Journal of the American Mosquito Control Association. 1990;6(2):251-3. 108. Espinosa M, Giamperetti S, Abril M, Seijo A. VERTICAL TRANSMISSION OF DENGUE VIRUS IN Aedes aegypti COLLECTED IN PUERTO IGUAZÚ, MISIONES, ARGENTINA. Revista do Instituto de Medicina Tropical de São Paulo. 2014;56(2):165-7. 109. Grunnill M, Boots M. How Important is Vertical Transmission of Dengue Viruses by Mosquitoes (Diptera: Culicidae)? Journal of medical entomology. 2016;53(1):1-19. 110. Hirsch JF, Deschamps C, Lhuillier M. [Metropolitan transmission of dengue by accidental inoculation at a hospital]. Annales de medecine interne. 1990;141(7):629. 111. de Wazières B, Gil H, Vuitton DA, Dupond J-L. Nosocomial transmission of dengue from a needlestick injury. The Lancet. 1998;351(9101):498.

322
112. Wagner D, de With K, Huzly D, Hufert F, Weidmann M, Breisinger S, et al. Nosocomial Acquisition of Dengue. Emerging Infectious Diseases. 2004;10(10):1872-3. 113. Thaithumyanon P, Thisyakorn U, Deerojnawong J, Innis BL. Dengue infection complicated by severe hemorrhage and vertical transmission in a parturient woman. Clinical infectious diseases : an official publication of the Infectious Diseases Society of America. 1994;18(2):248-9. 114. Chye JK, Lim CT, Ng KB, Lim JM, George R, Lam SK. Vertical transmission of dengue. Clinical infectious diseases : an official publication of the Infectious Diseases Society of America. 1997;25(6):1374-7. 115. Kerdpanich A, Watanaveeradej V, Samakoses R, Chumnanvanakij S, Chulyamitporn T, Sumeksri P, et al. Perinatal dengue infection. The Southeast Asian journal of tropical medicine and public health. 2001;32(3):488-93. 116. Boussemart T, Babe P, Sibille G, Neyret C, Berchel C. Prenatal Transmission of Dengue: Two New Cases. Journal Of Perinatology. 2001;21:255. 117. Rigau-Perez JG, Vorndam AV, Clark GG. The dengue and dengue hemorrhagic fever epidemic in Puerto Rico, 1994-1995. The American journal of tropical medicine and hygiene. 2001;64(1-2):67-74. 118. Dengue and severe dengue [Internet]. WHO. 2017. Available from: http://www.who.int/mediacentre/factsheets/fs117/en/. 119. Tai AY, McGuinness SL, Robosa R, Turner D, Huang GK, Leder K, et al. Management of dengue in Australian travellers: a retrospective multicentre analysis. The Medical journal of Australia. 2017;206(7):295-300. 120. Kalayanarooj S. Standardized Clinical Management: Evidence of Reduction of Dengue Haemorrhagic FeverCase-Fatality Rate in ThailandDec-1999. Available from: http://www.who.int/iris/handle/10665/148661. 121. Deen JL, Harris E, Wills B, Balmaseda A, Hammond SN, Rocha C, et al. The WHO dengue classification and case definitions: time for a reassessment. The Lancet. 2006;368(9530):170-3. 122. WHO. Dengue hemorrhagic fever: diagnosis, treatment, prevention and control. 1997. 123. Balmaseda A, Hammond SN, Perez MA, Cuadra R, Solano S, Rocha J, et al. Short report: assessment of the World Health Organization scheme for classification of dengue severity in Nicaragua. The American journal of tropical medicine and hygiene. 2005;73(6):1059-62. 124. Phuong CX, Nhan NT, Kneen R, Thuy PT, van Thien C, Nga NT, et al. Clinical diagnosis and assessment of severity of confirmed dengue infections in Vietnamese children: is the world health organization classification system helpful? The American journal of tropical medicine and hygiene. 2004;70(2):172-9. 125. Distinguishing between Zika and Spondweni viruses [press release]. Bulletin of the World Health Organization2016. 126. Musso D, Gubler DJ. Zika Virus. Clinical Microbiology Reviews. 2016;29(3):487-524. 127. Dick GWA, Kitchen SF, Haddow AJ. Zika Virus (I). Isolations and serological specificity. Transactions of the Royal Society of Tropical Medicine and Hygiene. 1952;46(5):509-20. 128. MacNamara FN. Zika virus : A report on three cases of human infection during an epidemic of jaundice in Nigeria. Transactions of the Royal Society of Tropical Medicine and Hygiene. 1954;48(2):139-45. 129. Marchette NJ, Garcia R, Rudnick A. Isolation of Zika virus from Aedes aegypti mosquitoes in Malaysia. The American journal of tropical medicine and hygiene. 1969;18(3):411-5. 130. Olson JG, Ksiazek TG, Suhandiman, Triwibowo. Zika virus, a cause of fever in Central Java, Indonesia. Trans R Soc Trop Med Hyg. 1981;75(3):389-93.

323
131. Lanciotti RS, Kosoy OL, Laven JJ, Velez JO, Lambert AJ, Johnson AJ, et al. Genetic and Serologic Properties of Zika Virus Associated with an Epidemic, Yap State, Micronesia, 2007. Emerging Infectious Diseases. 2008;14(8):1232-9. 132. Cao-Lormeau V-M, Roche C, Teissier A, Robin E, Berry A-L, Mallet H-P, et al. Zika Virus, French Polynesia, South Pacific, 2013. Emerging Infectious Diseases. 2014;20(6):1085-6. 133. Cristiane WC, Igor ADP, Mariana K, Moreno SR, Monaise MOS, Gubio SC, et al. Outbreak of Exanthematous Illness Associated with Zika, Chikungunya, and Dengue Viruses, Salvador, Brazil. Emerging Infectious Disease journal. 2015;21(12):2274. 134. Heukelbach J, Alencar CH, Kelvin AA, de Oliveira WK, Pamplona de Goes Cavalcanti L. Zika virus outbreak in Brazil. Journal of infection in developing countries. 2016;10(2):116-20. 135. Erwin C, Margaret P-G, Pedro JB, Jorge EO, Matthew TA. Detection of Autochthonous Zika Virus Transmission in Sincelejo, Colombia. Emerging Infectious Disease journal. 2016;22(5):927. 136. Fauci AS, Morens DM. Zika Virus in the Americas — Yet Another Arbovirus Threat. New England Journal of Medicine. 2016;374(7):601-4. 137. http://apps.who.int/iris/bitstream/10665/258596/1/zika-classification-20Jul17-eng.pdf?ua=1. 138. Cohen J. Where has all the Zika gone? Science. 2017;357(6352):631. 139. Chang H-H, Huber RG, Bond PJ, Grad YH, Camerini D, Maurer-Stroh S, et al. Systematic analysis of protein identity between Zika virus and other arthropod-borne viruses. Bulletin of the World Health Organization. 2017;95(7):517-25I. 140. Haddow AD, Schuh AJ, Yasuda CY, Kasper MR, Heang V, Huy R, et al. Genetic Characterization of Zika Virus Strains: Geographic Expansion of the Asian Lineage. PLoS Neglected Tropical Diseases. 2012;6(2):e1477. 141. Musso D, Nilles EJ, Cao-Lormeau VM. Rapid spread of emerging Zika virus in the Pacific area. Clinical Microbiology and Infection. 2014;20(10):O595-O6. 142. Ledermann JP, Guillaumot L, Yug L, Saweyog SC, Tided M, Machieng P, et al. Aedes hensilli as a Potential Vector of Chikungunya and Zika Viruses. PLOS Neglected Tropical Diseases. 2014;8(10):e3188. 143. Grard G, Caron M, Mombo IM, Nkoghe D, Mboui Ondo S, Jiolle D, et al. Zika Virus in Gabon (Central Africa) – 2007: A New Threat from Aedes albopictus? PLOS Neglected Tropical Diseases. 2014;8(2):e2681. 144. Diallo D, Sall AA, Diagne CT, Faye O, Faye O, Ba Y, et al. Zika Virus Emergence in Mosquitoes in Southeastern Senegal, 2011. PLOS ONE. 2014;9(10):e109442. 145. Boorman JP, Porterfield JS. A simple technique for infection of mosquitoes with viruses; transmission of Zika virus. Trans R Soc Trop Med Hyg. 1956;50(3):238-42. 146. Panchaud A, Stojanov M, Ammerdorffer A, Vouga M, Baud D. Emerging Role of Zika Virus in Adverse Fetal and Neonatal Outcomes. Clinical Microbiology Reviews. 2016;29(3):659-94. 147. Foy BD, Kobylinski KC, Foy JLC, Blitvich BJ, Travassos da Rosa A, Haddow AD, et al. Probable Non–Vector-borne Transmission of Zika Virus, Colorado, USA. Emerging Infectious Diseases. 2011;17(5):880-2. 148. Musso D, Nhan T, Robin E, Roche C, Bierlaire D, Zisou K, et al. Potential for Zika virus transmission through blood transfusion demonstrated during an outbreak in French Polynesia, November 2013 to February 2014. Euro Surveill. 2014;19(14). 149. de Noronha L, Zanluca C, Azevedo MLV, Luz KG, dos Santos CND. Zika virus damages the human placental barrier and presents marked fetal neurotropism. Memórias do Instituto Oswaldo Cruz. 2016;111(5):287-93. 150. Calvet G, Aguiar RS, Melo ASO, Sampaio SA, de Filippis I, Fabri A, et al. Detection and sequencing of Zika virus from amniotic fluid of fetuses with microcephaly in Brazil: a case study. The Lancet Infectious Diseases.16(6):653-60. 151. Musso D, Roche C, Robin E, Nhan T, Teissier A, Cao-Lormeau VM. Potential sexual transmission of Zika virus. Emerg Infect Dis. 2015;21(2):359-61.

324
152. D’Ortenzio E, Matheron S, de Lamballerie X, Hubert B, Piorkowski G, Maquart M, et al. Evidence of Sexual Transmission of Zika Virus. New England Journal of Medicine. 2016;374(22):2195-8. 153. Filipe AR, Martins CMV, Rocha H. Laboratory infection with Zika virus after vaccination against yellow fever. Archiv für die gesamte Virusforschung. 1973;43(4):315-9. 154. Cao-Lormeau VM, Blake A, Mons S, Lastere S, Roche C, Vanhomwegen J, et al. Guillain-Barre Syndrome outbreak associated with Zika virus infection in French Polynesia: a case-control study. Lancet (London, England). 2016;387(10027):1531-9. 155. Parra B, Lizarazo J, Jiménez-Arango JA, Zea-Vera AF, González-Manrique G, Vargas J, et al. Guillain–Barré Syndrome Associated with Zika Virus Infection in Colombia. New England Journal of Medicine. 2016;375(16):1513-23. 156. dos Santos T, Rodriguez A, Almiron M, Sanhueza A, Ramon P, de Oliveira WK, et al. Zika Virus and the Guillain–Barré Syndrome — Case Series from Seven Countries. New England Journal of Medicine. 2016;375(16):1598-601. 157. Leis AA, Stokic DS. Zika Virus and Guillain–Barre Syndrome: Is There Sufficient Evidence for Causality? Frontiers in Neurology. 2016;7:170. 158. Increase in Reported Prevalence of Microcephaly in Infants Born to Women Living in Areas with Confirmed Zika Virus Transmission During the First Trimester of Pregnancy — Brazil, 2015 [press release]. MMWR Morb Mortal Wkly Rep2016;. 159. Kumar M, Ching L, Astern J, Lim E, Stokes AJ, Melish M, et al. Prevalence of Antibodies to Zika Virus in Mothers from Hawaii Who Delivered Babies with and without Microcephaly between 2009-2012. PLoS Neglected Tropical Diseases. 2016;10(12):e0005262. 160. Cuevas EL TV, Rozo N, et al. Preliminary Report of Microcephaly Potentially Associated with Zika Virus Infection During Pregnancy — Colombia. 2016. 161. Frierson JG. The Yellow Fever Vaccine: A History. The Yale Journal of Biology and Medicine. 2010;83(2):77-85. 162. Monath TP, Seligman SJ, Robertson JS, Guy B, Hayes EB, Condit RC, et al. Live virus vaccines based on a yellow fever vaccine backbone: standardized template with key considerations for a risk/benefit assessment. Vaccine. 2015;33(1):62-72. 163. Monath TP, Vasconcelos PFC. Yellow fever. Journal of Clinical Virology.64:160-73. 164. WHO. Yellow fever – Brazil. WHO-Emergencies preparedness, response. 2018. 165. Monath TP, Vasconcelos PF. Yellow fever. Journal of clinical virology : the official publication of the Pan American Society for Clinical Virology. 2015;64:160-73. 166. Front-matter A2 - Burrell, Christopher J. In: Howard CR, Murphy FA, editors. Fenner and White's Medical Virology (Fifth Edition). London: Academic Press; 2017. p. i-iii. 167. Norrby E. Yellow fever and Max Theiler: the only Nobel Prize for a virus vaccine. The Journal of Experimental Medicine. 2007;204(12):2779-84. 168. Beasley DW, McAuley AJ, Bente DA. Yellow fever virus: genetic and phenotypic diversity and implications for detection, prevention and therapy. Antiviral Res. 2015;115:48-70. 169. Converse JL, Kovatch RM, Pulliam JD, Nagle SC, Snyder EM. Virulence and Pathogenesis of Yellow Fever Virus Serially Passaged in Cell Culture. Applied Microbiology. 1971;21(6):1053-7. 170. Daffis S, Kontermann RE, Korimbocus J, Zeller H, Klenk HD, Ter Meulen J. Antibody responses against wild-type yellow fever virus and the 17D vaccine strain: characterization with human monoclonal antibody fragments and neutralization escape variants. Virology. 2005;337(2):262-72. 171. Schuh AJ, Li L, Tesh RB, Innis BL, Barrett ADT. Genetic characterization of early isolates of Japanese encephalitis virus: genotype II has been circulating since at least 1951. The Journal of General Virology. 2010;91(Pt 1):95-102. 172. Halstead SB, Jacobson J. Japanese encephalitis. Advances in virus research. 2003;61:103-38.

325
173. Khan SU, Salje H, Hannan A, Islam MA, Bhuyan AAM, Islam MA, et al. Dynamics of Japanese Encephalitis Virus Transmission among Pigs in Northwest Bangladesh and the Potential Impact of Pig Vaccination. PLOS Neglected Tropical Diseases. 2014;8(9):e3166. 174. Rosen L. The Natural History of Japanese Encephalitis Virus. Annual Review of Microbiology. 1986;40(1):395-414. 175. Solomon T, Ni H, Beasley DWC, Ekkelenkamp M, Cardosa MJ, Barrett ADT. Origin and Evolution of Japanese Encephalitis Virus in Southeast Asia. Journal of virology. 2003;77(5):3091-8. 176. Mackenzie JS, Broom AK, Hall RA, Johansen CA, Lindsay MD, Phillips DA, et al. Arboviruses in the Australian region, 1990 to 1998. Communicable diseases intelligence. 1998;22:93-9. 177. Thompson MR, Kaminski JJ, Kurt-Jones EA, Fitzgerald KA. Pattern Recognition Receptors and the Innate Immune Response to Viral Infection. Viruses. 2011;3(6):920-40. 178. Nasirudeen AM, Wong HH, Thien P, Xu S, Lam KP, Liu DX. RIG-I, MDA5 and TLR3 synergistically play an important role in restriction of dengue virus infection. PLoS Negl Trop Dis. 2011;5(1):e926. 179. Castillo Ramirez JA, Urcuqui-Inchima S. Dengue Virus Control of Type I IFN Responses: A History of Manipulation and Control. Journal of Interferon & Cytokine Research. 2015;35(6):421-30. 180. Costa VV, Fagundes CT, Souza DG, Teixeira MM. Inflammatory and Innate Immune Responses in Dengue Infection: Protection versus Disease Induction. The American Journal of Pathology. 2013;182(6):1950-61. 181. Kawai T, Akira S. Antiviral signaling through pattern recognition receptors. Journal of biochemistry. 2007;141(2):137-45. 182. Schoggins JW. Interferon-stimulated genes: roles in viral pathogenesis. Current opinion in virology. 2014;0:40-6. 183. Lin RJ, Yu HP, Chang BL, Tang WC, Liao CL, Lin YL. Distinct antiviral roles for human 2',5'-oligoadenylate synthetase family members against dengue virus infection. Journal of immunology (Baltimore, Md : 1950). 2009;183(12):8035-43. 184. Chan YK, Huang IC, Farzan M. IFITM Proteins Restrict Antibody-Dependent Enhancement of Dengue Virus Infection. PLOS ONE. 2012;7(3):e34508. 185. Morrison J, Aguirre S, Fernandez-Sesma A. Innate Immunity Evasion by Dengue Virus. Viruses. 2012;4(3):397-413. 186. Aguirre S, Maestre AM, Pagni S, Patel JR, Savage T, Gutman D, et al. DENV inhibits type I IFN production in infected cells by cleaving human STING. PLoS Pathog. 2012;8(10):e1002934. 187. Green AM, Beatty PR, Hadjilaou A, Harris E. Innate immunity to dengue virus infection and subversion of antiviral responses. Journal of molecular biology. 2014;426(6):1148-60. 188. Grant D, Tan GK, Qing M, Ng JKW, Yip A, Zou G, et al. A Single Amino Acid in Nonstructural Protein NS4B Confers Virulence to Dengue Virus in AG129 Mice through Enhancement of Viral RNA Synthesis. Journal of virology. 2011;85(15):7775-87. 189. Mazzon M, Jones M, Davidson A, Chain B, Jacobs M. Dengue virus NS5 inhibits interferon-alpha signaling by blocking signal transducer and activator of transcription 2 phosphorylation. J Infect Dis. 2009;200(8):1261-70. 190. Laurent-Rolle M, Boer EF, Lubick KJ, Wolfinbarger JB, Carmody AB, Rockx B, et al. The NS5 Protein of the Virulent West Nile Virus NY99 Strain Is a Potent Antagonist of Type I Interferon-Mediated JAK-STAT Signaling. Journal of virology. 2010;84(7):3503-15. 191. Ye J, Chen Z, Li Y, Zhao Z, He W, Zohaib A, et al. Japanese Encephalitis Virus NS5 Inhibits Type I Interferon (IFN) Production by Blocking the Nuclear Translocation of IFN Regulatory Factor 3 and NF-κB. Journal of virology. 2017;91(8):e00039-17. 192. Ashour J, Laurent-Rolle M, Shi P-Y, García-Sastre A. NS5 of Dengue Virus Mediates STAT2 Binding and Degradation. Journal of virology. 2009;83(11):5408-18.

326
193. Nesargikar PN, Spiller B, Chavez R. The complement system: history, pathways, cascade and inhibitors. European Journal of Microbiology & Immunology. 2012;2(2):103-11. 194. Stoermer KA, Morrison TE. Complement and Viral Pathogenesis. Virology. 2011;411(2):362-73. 195. Mehlhop E, Diamond MS. Protective immune responses against West Nile virus are primed by distinct complement activation pathways. The Journal of Experimental Medicine. 2006;203(5):1371. 196. Avirutnan P, Hauhart RE, Marovich MA, Garred P, Atkinson JP, Diamond MS. Complement-mediated neutralization of dengue virus requires mannose-binding lectin. mBio. 2011;2(6). 197. Mehlhop E, Ansarah-Sobrinho C, Johnson S, Engle M, Fremont DH, Pierson Theodore C, et al. Complement Protein C1q Inhibits Antibody-Dependent Enhancement of Flavivirus Infection in an IgG Subclass-Specific Manner. Cell Host & Microbe. 2007;2(6):417-26. 198. Malasit P. Complement and dengue haemorrhagic fever/shock syndrome. The Southeast Asian journal of tropical medicine and public health. 1987;18(3):316-20. 199. Avirutnan P, Punyadee N, Noisakran S, Komoltri C, Thiemmeca S, Auethavornanan K, et al. Vascular leakage in severe dengue virus infections: a potential role for the nonstructural viral protein NS1 and complement. J Infect Dis. 2006;193(8):1078-88. 200. Chung KM, Liszewski MK, Nybakken G, Davis AE, Townsend RR, Fremont DH, et al. West Nile virus nonstructural protein NS1 inhibits complement activation by binding the regulatory protein factor H. Proc Natl Acad Sci U S A. 2006;103(50):19111-6. 201. Avirutnan P, Hauhart RE, Somnuke P, Blom AM, Diamond MS, Atkinson JP. Binding of flavivirus nonstructural protein NS1 to C4b binding protein modulates complement activation. Journal of immunology (Baltimore, Md : 1950). 2011;187(1):424-33. 202. Thiemmeca S, Tamdet C, Punyadee N, Prommool T, Songjaeng A, Noisakran S, et al. Secreted NS1 Protects Dengue Virus from Mannose-Binding Lectin-Mediated Neutralization. Journal of immunology (Baltimore, Md : 1950). 2016;197(10):4053-65. 203. Williams M, Mayer SV, Johnson WL, Chen R, Volkova E, Vilcarromero S, et al. Lineage II of Southeast Asian/American DENV-2 Is Associated with a Severe Dengue Outbreak in the Peruvian Amazon. The American journal of tropical medicine and hygiene. 2014;91(3):611-20. 204. Kurane I, Innis BL, Nisalak A, Hoke C, Nimmannitya S, Meager A, et al. Human T cell responses to dengue virus antigens. Proliferative responses and interferon gamma production. The Journal of clinical investigation. 1989;83(2):506-13. 205. Turtle L, Bali T, Buxton G, Chib S, Chan S, Soni M, et al. Human T cell responses to Japanese encephalitis virus in health and disease. The Journal of Experimental Medicine. 2016;213(7):1331. 206. James EA, LaFond RE, Gates TJ, Mai DT, Malhotra U, Kwok WW. Yellow Fever Vaccination Elicits Broad Functional CD4(+) T Cell Responses That Recognize Structural and Nonstructural Proteins. Journal of virology. 2013;87(23):12794-804. 207. Co MD, Kilpatrick ED, Rothman AL. Dynamics of the CD8 T-cell response following yellow fever virus 17D immunization. Immunology. 2009;128(1 Suppl):e718-27. 208. Liang H, Lee M, Jin X. Guiding dengue vaccine development using knowledge gained from the success of the yellow fever vaccine. Cellular and Molecular Immunology. 2016;13(1):36-46. 209. Weiskopf D, Sette A. T-Cell Immunity to Infection with Dengue Virus in Humans. Frontiers in Immunology. 2014;5:93. 210. Mathew A, Kurane I, Rothman AL, Zeng LL, Brinton MA, Ennis FA. Dominant recognition by human CD8+ cytotoxic T lymphocytes of dengue virus nonstructural proteins NS3 and NS1.2a. Journal of Clinical Investigation. 1996;98(7):1684-91. 211. Spaulding AC, Kurane I, Ennis FA, Rothman AL. Analysis of Murine CD8+ T-Cell Clones Specific for the Dengue Virus NS3 Protein: Flavivirus Cross-Reactivity and Influence of Infecting Serotype. Journal of virology. 1999;73(1):398-403.

327
212. Muyanja E, Ssemaganda A, Ngauv P, Cubas R, Perrin H, Srinivasan D, et al. Immune activation alters cellular and humoral responses to yellow fever 17D vaccine. The Journal of clinical investigation. 2014;124(7):3147-58. 213. Pulendran B. Learning immunology from the yellow fever vaccine: innate immunity to systems vaccinology. Nature Reviews Immunology. 2009;9:741. 214. Khan AM, Miotto O, Nascimento EJM, Srinivasan KN, Heiny AT, Zhang GL, et al. Conservation and Variability of Dengue Virus Proteins: Implications for Vaccine Design. PLOS Neglected Tropical Diseases. 2008;2(8):e272. 215. Mathew A, Rothman AL. Understanding the contribution of cellular immunity to dengue disease pathogenesis. Immunological Reviews. 2008;225(1):300-13. 216. Mongkolsapaya J, Dejnirattisai W, Xu X-n, Vasanawathana S, Tangthawornchaikul N, Chairunsri A, et al. Original antigenic sin and apoptosis in the pathogenesis of dengue hemorrhagic fever. Nature medicine. 2003;9:921. 217. Dejnirattisai W, Duangchinda T, Lin CL, Vasanawathana S, Jones M, Jacobs M, et al. A complex interplay among virus, dendritic cells, T cells, and cytokines in dengue virus infections. Journal of immunology (Baltimore, Md : 1950). 2008;181(9):5865-74. 218. Rothman AL, Ennis FA. Immunopathogenesis of Dengue Hemorrhagic Fever. Virology. 1999;257(1):1-6. 219. Loke H, Bethell DB, Phuong CXT, Dung M, Schneider J, White NJ, et al. Strong HLA Class I–Restricted T Cell Responses in Dengue Hemorrhagic Fever: A Double-Edged Sword? The Journal of Infectious Diseases. 2001;184(11):1369-73. 220. Mongkolsapaya J, Duangchinda T, Dejnirattisai W, Vasanawathana S, Avirutnan P, Jairungsri A, et al. T Cell Responses in Dengue Hemorrhagic Fever: Are Cross-Reactive T Cells Suboptimal? The Journal of Immunology. 2006;176(6):3821. 221. Wahala WMPB, de Silva AM. The Human Antibody Response to Dengue Virus Infection. Viruses. 2011;3(12):2374-95. 222. Sa-Ngasang A, Anantapreecha S, A-Nuegoonpipat A, Chanama S, Wibulwattanakij S, Pattanakul K, et al. Specific IgM and IgG responses in primary and secondary dengue virus infections determined by enzyme-linked immunosorbent assay. Epidemiology and Infection. 2006;134(4):820-5. 223. J Groen JV, CCopra, E Balentien , V Deubel , V Vorndam.,, Osterhaus aAD. Diagnostic value of dengue virus-specific IgA and
IgM serum antibody detection. Microbes Infect. 1999;1(13):1085-90. 224. Imrie A, Meeks J, Gurary A, Suhkbaatar M, Truong TT, Cropp CB, et al. Antibody to Dengue 1 Detected More Than 60 Years after Infection. Viral immunology. 2007;20(4):672-5. 225. Ngwe Tun MM, Muta Y, Inoue S, Morita K. Persistence of Neutralizing Antibody Against Dengue Virus 2 After 70 Years from Infection in Nagasaki. BioResearch Open Access. 2016;5(1):188-91. 226. Diamond MS, Shrestha B, Marri A, Mahan D, Engle M. B cells and antibody play critical roles in the immediate defense of disseminated infection by West Nile encephalitis virus. Journal of virology. 2003;77(4):2578-86. 227. Roehrig JT, Staudinger LA, Hunt AR, Mathews JH, Blair CD. Antibody prophylaxis and therapy for flavivirus encephalitis infections. Annals of the New York Academy of Sciences. 2001;951:286-97. 228. Chambers TJ, Droll DA, Walton AH, Schwartz J, Wold WSM, Nickells J. West Nile 25A virus infection of B-cell-deficient (μMT) mice: characterization of neuroinvasiveness and pseudoreversion of the viral envelope protein. Journal of General Virology. 2008;89(3):627-35. 229. Dulbecco R, Vogt M, Strickland AGR. A study of the basic aspects of neutralization of two animal viruses, Western equine encephalitis virus and poliomyelitis virus. Virology. 1956;2(2):162-205. 230. Della-Porta AJ, Westaway EG. A Multi-Hit Model for the Neutralization of Animal Viruses. Journal of General Virology. 1978;38(1):1-19.

328
231. Westaway EG. The neutralization of arboviruses: II. Neutralization in heterologous virus-serum mixtures with four group B arboviruses. Virology. 1965;26(4):528-37. 232. Rothman AL. Immunity to dengue virus: a tale of original antigenic sin and tropical cytokine storms. Nat Rev Immunol. 2011;11(8):532-43. 233. Roehrig JT. Antigenic Structure of Flavivirus Proteins. Advances in virus research. 2003;59:141-75. 234. Roehrig JT, Bolin RA, Kelly RG. Monoclonal antibody mapping of the envelope glycoprotein of the dengue 2 virus, Jamaica. Virology. 1998;246(2):317-28. 235. Pierson TC, Diamond MS. Molecular mechanisms of antibody-mediated neutralisation of flavivirus infection. Expert reviews in molecular medicine. 2008;10:e12-e. 236. Gromowski GD, Barrett AD. Characterization of an antigenic site that contains a dominant, type-specific neutralization determinant on the envelope protein domain III (ED3) of dengue 2 virus. Virology. 2007;366(2):349-60. 237. Crill WD, Roehrig JT. Monoclonal antibodies that bind to domain III of dengue virus E glycoprotein are the most efficient blockers of virus adsorption to Vero cells. Journal of virology. 2001;75(16):7769-73. 238. Falconar AK. Identification of an epitope on the dengue virus membrane (M) protein defined by cross-protective monoclonal antibodies: design of an improved epitope sequence based on common determinants present in both envelope (E and M) proteins. Archives of virology. 1999;144(12):2313-30. 239. Wahala WMPB, Kraus AA, Haymore LB, Accavitti-Loper MA, de Silva AM. Dengue virus neutralization by human immune sera: Role of envelope protein domain III-reactive antibody. Virology. 2009;392(1):103-13. 240. da Silva ANMR, Nascimento EJM, Cordeiro MT, Gil LHVG, Abath FGC, Montenegro SML, et al. Identification of Continuous Human B-Cell Epitopes in the Envelope Glycoprotein of Dengue Virus Type 3 (DENV-3). PLOS ONE. 2009;4(10):e7425. 241. Wahala WPB, Kraus AA, Haymore LB, Accavitti-Loper MA, de Silva AM. Dengue virus neutralization by human immune sera: role of envelope protein domain III - reactive antibody. Virology. 2009;392(1):103-13. 242. Luo Y, Guo X, Yan H, Fang D, Zeng G, Zhou J, et al. Comprehensive mapping infection-enhancing epitopes of dengue pr protein using polyclonal antibody against prM. Applied Microbiology and Biotechnology. 2015;99(14):5917-27. 243. Dejnirattisai W, Jumnainsong A, Onsirisakul N, Fitton P, Vasanawathana S, Limpitikul W, et al. Cross-reacting antibodies enhance dengue virus infection in humans. Science. 2010;328(5979):745-8. 244. Halstead SB, Rojanasuphot S, Sangkawibha N. Original antigenic sin in dengue. The American journal of tropical medicine and hygiene. 1983;32(1):154-6. 245. Martina BEE, Koraka P, Osterhaus ADME. Dengue Virus Pathogenesis: an Integrated View. Clinical Microbiology Reviews. 2009;22(4):564-81. 246. Halstead SB. In Vivo Enhancement of Dengue Virus Infection in Rhesus Monkeys by Passively Transferred Antibody. The Journal of Infectious Diseases. 1979;140(4):527-33. 247. Halstead SB, Mahalingam PS, Marovich MA, Ubol S, Mosser PDM. Intrinsic antibody-dependent enhancement of microbial infection in macrophages: disease regulation by immune complexes. The Lancet infectious diseases. 2010;10(10):712-22. 248. Sellahewa KH. Pathogenesis of Dengue Haemorrhagic Fever and Its Impact on Case Management. ISRN Infectious Diseases. 2013;2013:6. 249. Guzman MG, Vazquez S. The Complexity of Antibody-Dependent Enhancement of Dengue Virus Infection. Viruses. 2010;2(12):2649-62. 250. Mathew A, West K, Kalayanarooj S, Gibbons RV, Srikiatkhachorn A, Green S, et al. B-Cell Responses During Primary and Secondary Dengue Virus Infections in Humans. The Journal of Infectious Diseases. 2011;204(10):1514-22. 251. Halstead SB, Nimmannitya S, Cohen SN. Observations related to pathogenesis of dengue hemorrhagic fever. IV. Relation of disease severity to antibody response and virus recovered. The Yale Journal of Biology and Medicine. 1970;42(5):311-28.

329
252. Simmons CP, Chau TNB, Thuy TT, Tuan NM, Hoang DM, Thien NT, et al. Maternal Antibody and Viral Factors in the Pathogenesis of Dengue Virus in Infants. The Journal of infectious diseases. 2007;196(3):416-24. 253. Littaua R, Kurane I, Ennis FA. Human IgG Fc receptor II mediates antibody-dependent enhancement of dengue virus infection. The Journal of Immunology. 1990;144(8):3183. 254. Diamond MS, Edgil D, Roberts TG, Lu B, Harris E. Infection of Human Cells by Dengue Virus Is Modulated by Different Cell Types and Viral Strains. Journal of virology. 2000;74(17):7814-23. 255. Londono-Renteria B, Troupin A, Cardenas JC, Hall A, Perez OG, Cardenas L, et al. A relevant in vitro human model for the study of Zika virus antibody-dependent enhancement. J Gen Virol. 2017;98(7):1702-12. 256. Pantoja P, Perez-Guzman EX, Rodriguez IV, White LJ, Gonzalez O, Serrano C, et al. Zika virus pathogenesis in rhesus macaques is unaffected by pre-existing immunity to dengue virus. Nature communications. 2017;8:15674. 257. Bardina SV, Bunduc P, Tripathi S, Duehr J, Frere JJ, Brown JA, et al. Enhancement of Zika virus pathogenesis by preexisting antiflavivirus immunity. Science (New York, NY). 2017;356(6334):175-80. 258. Robbiani DF, Bozzacco L, Keeffe JR, Khouri R, Olsen PC, Gazumyan A, et al. Recurrent Potent Human Neutralizing Antibodies to Zika Virus in Brazil and Mexico. Cell. 2017;169(4):597-609.e11. 259. Kam Y-W, Lee CY-P, Teo T-H, Howland SW, Amrun SN, Lum F-M, et al. Cross-reactive dengue human monoclonal antibody prevents severe pathologies and death from Zika virus infections. JCI Insight. 2017;2(8). 260. Dejnirattisai W, Supasa P, Wongwiwat W, Rouvinski A, Barba-Spaeth G, Duangchinda T, et al. Dengue virus sero-cross-reactivity drives antibody-dependent enhancement of infection with zika virus. Nature immunology. 2016;17(9):1102-8. 261. Paul LM, Carlin ER, Jenkins MM, Tan AL, Barcellona CM, Nicholson CO, et al. Dengue virus antibodies enhance Zika virus infection. Clin Trans Immunol. 2016;5:e117. 262. Hotta S. Experimental studies on dengue. I. Isolation, identification and modification of the virus. J Infect Dis. 1952;90(1):1-9. 263. Wu J-Y, Lun Z-R, James AA, Chen X-G. Dengue Fever in Mainland China. The American journal of tropical medicine and hygiene. 2010;83(3):664-71. 264. Tesh RB. A Method for the Isolation and Identification of Dengue Viruses, using Mosquito Cell Cultures*. The American journal of tropical medicine and hygiene. 1979;28(6):1053-9. 265. Halstead SB, Sukhavachana P, Nisalak A. In vitro Recovery of Dengue Viruses from Naturally Infected Human Beings and Arthropods. Nature. 1964;202:931. 266. Rosen L, Gubler D. The use of mosquitoes to detect and propagate dengue viruses. The American journal of tropical medicine and hygiene. 1974;23(6):1153-60. 267. Guzman MG, Kouri G. Advances in dengue diagnosis. Clinical and diagnostic laboratory immunology. 1996;3(6):621-7. 268. Chan JF-W, Yip CC-Y, Tsang JO-L, Tee K-M, Cai J-P, Chik KK-H, et al. Differential cell line susceptibility to the emerging Zika virus: implications for disease pathogenesis, non-vector-borne human transmission and animal reservoirs. Emerging Microbes & Infections. 2016;5(8):e93. 269. Pyke AT, Moore PR, Hall-Mendelin S, McMahon JL, Harrower BJ, Constantino TR, et al. Isolation of Zika Virus Imported from Tonga into Australia. PLoS Currents. 2016;8:ecurrents.outbreaks.849adc0ad16beec4536695281707f785. 270. Bhatnagar J, Blau DM, Shieh W-J, Paddock CD, Drew C, Liu L, et al. Molecular Detection and Typing of Dengue Viruses from Archived Tissues of Fatal Cases by RT-PCR and Sequencing: Diagnostic and Epidemiologic Implications. The American journal of tropical medicine and hygiene. 2012;86(2):335-40.

330
271. Lanciotti RS, Calisher CH, Gubler DJ, Chang GJ, Vorndam AV. Rapid detection and typing of dengue viruses from clinical samples by using reverse transcriptase-polymerase chain reaction. Journal of Clinical Microbiology. 1992;30(3):545-51. 272. Harris E, Roberts TG, Smith L, Selle J, Kramer LD, Valle S, et al. Typing of dengue viruses in clinical specimens and mosquitoes by single-tube multiplex reverse transcriptase PCR. J Clin Microbiol. 1998;36(9):2634-9. 273. Kuno G. Universal diagnostic RT-PCR protocol for arboviruses. Journal of Virological Methods. 1998;72(1):27-41. 274. Ayers M, Adachi D, Johnson G, Andonova M, Drebot M, Tellier R. A single tube RT-PCR assay for the detection of mosquito-borne flaviviruses. J Virol Methods. 2006;135(2):235-9. 275. Dash PK, Parida M, Santhosh SR, Saxena P, Srivastava A, Neeraja M, et al. Development and evaluation of a 1-step duplex reverse transcription polymerase chain reaction for differential diagnosis of chikungunya and dengue infection. Diagnostic Microbiology and Infectious Disease. 2008;62(1):52-7. 276. Chien L-J, Liao T-L, Shu P-Y, Huang J-H, Gubler DJ, Chang G-JJ. Development of Real-Time Reverse Transcriptase PCR Assays To Detect and Serotype Dengue Viruses. Journal of Clinical Microbiology. 2006;44(4):1295-304. 277. Kristy OM, Rodion G, Anna RC, Rebecca B, Lilin L, Muktha N, et al. Prolonged Detection of Zika Virus in Vaginal Secretions and Whole Blood. Emerging Infectious Disease journal. 2017;23(1):99. 278. Gourinat AC, O'Connor O, Calvez E, Goarant C, Dupont-Rouzeyrol M. Detection of Zika virus in urine. Emerging infectious diseases. 2015;21(1):84-6. 279. Rossini G, Gaibani P, Vocale C, Cagarelli R, Landini MP. Comparison of Zika virus (ZIKV) RNA detection in plasma, whole blood and urine - Case series of travel-associated ZIKV infection imported to Italy, 2016. The Journal of infection. 2017;75(3):242-5. 280. Henchal EA, Narupiti S, Feighny R, Padmanabhan R, Vakharia V. Detection of dengue virus RNA using nucleic acid hybridization. J Virol Methods. 1987;15(3):187-200. 281. Parida M, Horioke K, Ishida H, Dash PK, Saxena P, Jana AM, et al. Rapid detection and differentiation of dengue virus serotypes by a real-time reverse transcription-loop-mediated isothermal amplification assay. J Clin Microbiol. 2005;43(6):2895-903. 282. Lee D, Shin Y, Chung S, Hwang KS, Yoon DS, Lee JH. Simple and Highly Sensitive Molecular Diagnosis of Zika Virus by Lateral Flow Assays. Analytical Chemistry. 2016;88(24):12272-8. 283. Fink J, Gu F, Ling L, Tolfvenstam T, Olfat F, Chin KC, et al. Host Gene Expression Profiling of Dengue Virus Infection in Cell Lines and Patients. PLoS Neglected Tropical Diseases. 2007;1(2):e86. 284. Khan MJ, Trabuco AC, Alfonso HL, Figueiredo ML, Batista WC, Badra SJ, et al. DNA Microarray Platform for Detection and Surveillance of Viruses Transmitted by Small Mammals and Arthropods. PLoS Neglected Tropical Diseases. 2016;10(9):e0005017. 285. Glushakova LG, Bradley A, Bradley KM, Alto BW, Hoshika S, Hutter D, et al. High-throughput multiplexed xMAP Luminex array panel for detection of twenty two medically important mosquito-borne arboviruses based on innovations in synthetic biology. J Virol Methods. 2015;214:60-74. 286. Pal S, Dauner AL, Mitra I, Forshey BM, Garcia P, Morrison AC, et al. Evaluation of Dengue NS1 Antigen Rapid Tests and ELISA Kits Using Clinical Samples. PLoS ONE. 2014;9(11):e113411. 287. Bosch I, de Puig H, Hiley M, Carré-Camps M, Perdomo-Celis F, Narváez CF, et al. Rapid antigen tests for dengue virus serotypes and Zika virus in patient serum. Science Translational Medicine. 2017;9(409). 288. Séverine M, Rachida B, Bhety L, Valérie E, Laetitia B, Mirdad K, et al. Specificity of Dengue NS1 Antigen in Differential Diagnosis of Dengue and Zika Virus Infection. Emerging Infectious Disease journal. 2016;22(9):1691. 289. Organization WH. Dengue guidelines for diagnosis, treatment, prevention and control : new edition: Geneva : World Health Organization; 2009.

331
290. Wong SJ, Furuya A, Zou J, Xie X, Dupuis AP, 2nd, Kramer LD, et al. A Multiplex Microsphere Immunoassay for Zika Virus Diagnosis. EBioMedicine. 2017;16:136-40. 291. Clarke DH, Casals J. Techniques for hemagglutination and hemagglutination-inhibition with arthropod-borne viruses. The American journal of tropical medicine and hygiene. 1958;7(5):561-73. 292. Guzman MaG, Kourı G. Dengue diagnosis, advances and challenges. International Journal of Infectious Diseases. 2004;8(2):69-80. 293. Cardosa MJ, Hooi TP, Shaari NS. Development of a dot enzyme immunoassay for dengue 3: a sensitive method for the detection of antidengue antibodies. J Virol Methods. 1988;22(1):81-8. 294. Russell PK, Nisalak A, Sukhavachana P, Vivona S. A Plaque Reduction Test for Dengue Virus Neutralizing Antibodies. The Journal of Immunology. 1967;99(2):285. 295. Lukman N, Salim G, Kosasih H, Susanto NH, Parwati I, Fitri S, et al. Comparison of the Hemagglutination Inhibition Test and IgG ELISA in Categorizing Primary and Secondary Dengue Infections Based on the Plaque Reduction Neutralization Test. BioMed Research International. 2016;2016:5253842. 296. Hunsperger EA, Yoksan S, Buchy P, Nguyen VC, Sekaran SD, Enria DA, et al. Evaluation of commercially available anti-dengue virus immunoglobulin M tests. Emerg Infect Dis. 2009;15(3):436-40. 297. de Souza VA, Tateno AF, Oliveira RR, Domingues RB, Araujo ES, Kuster GW, et al. Sensitivity and specificity of three ELISA-based assays for discriminating primary from secondary acute dengue virus infection. Journal of clinical virology : the official publication of the Pan American Society for Clinical Virology. 2007;39(3):230-3. 298. Vaughn DW, Nisalak A, Solomon T, Kalayanarooj S, Nguyen MD, Kneen R, et al. Rapid serologic diagnosis of dengue virus infection using a commercial capture ELISA that distinguishes primary and secondary infections. The American journal of tropical medicine and hygiene. 1999;60(4):693-8. 299. Kuno G, Cropp CB, Wong-Lee J, Gubler DJ. Evaluation of an IgM immunoblot kit for dengue diagnosis. The American journal of tropical medicine and hygiene. 1998;59(5):757-62. 300. Timiryasova TM, Bonaparte MI, Luo P, Zedar R, Hu BT, Hildreth SW. Optimization and validation of a plaque reduction neutralization test for the detection of neutralizing antibodies to four serotypes of dengue virus used in support of dengue vaccine development. The American journal of tropical medicine and hygiene. 2013;88(5):962-70. 301. Vaidya SR, Brown DW, Jin L, Samuel D, Andrews N, Brown KE. Development of a focus reduction neutralization test (FRNT) for detection of mumps virus neutralizing antibodies. J Virol Methods. 2010;163(1):153-6. 302. Tanishita O, Takahashi Y, Okuno Y, Yamanishi K, Takahashi M. Evaluation of focus reduction neutralization test with peroxidase-antiperoxidase staining technique for hemorrhagic fever with renal syndrome virus. Journal of Clinical Microbiology. 1984;20(6):1213-5. 303. Kim SH, Jang SH, Kim J, Lee KY, Jang YS. A microneutralization assay for dengue viruses using mosquito C6/36 cells. Acta virologica. 2013;57(3):379-81. 304. Vorndam V, Beltran M. Enzyme-linked immunosorbent assay-format microneutralization test for dengue viruses. The American journal of tropical medicine and hygiene. 2002;66(2):208-12. 305. de Alwis R, de Silva AM. Measuring antibody neutralization of dengue virus (DENV) using a flow cytometry-based technique. Methods in molecular biology (Clifton, NJ). 2014;1138:27-39. 306. Lambeth CR, White LJ, Johnston RE, de Silva AM. Flow cytometry-based assay for titrating dengue virus. J Clin Microbiol. 2005;43(7):3267-72. 307. Thomas SJ, Nisalak A, Anderson KB, Libraty DH, Kalayanarooj S, Vaughn DW, et al. Dengue Plaque Reduction Neutralization Test (PRNT) in Primary and Secondary Dengue Virus Infections: How Alterations in Assay Conditions Impact Performance. The American journal of tropical medicine and hygiene. 2009;81(5):825-33.

332
308. Kraus AA, Messer W, Haymore LB, de Silva AM. Comparison of Plaque- and Flow Cytometry-Based Methods for Measuring Dengue Virus Neutralization. Journal of Clinical Microbiology. 2007;45(11):3777-80. 309. Liu L, Wen K, Li J, Hu D, Huang Y, Qiu L, et al. Comparison of Plaque- and Enzyme-Linked Immunospot-Based Assays To Measure the Neutralizing Activities of Monoclonal Antibodies Specific to Domain III of Dengue Virus Envelope Protein. Clinical and Vaccine Immunology : CVI. 2012;19(1):73-8. 310. Putnak JR, de la Barrera R, Burgess T, Pardo J, Dessy F, Gheysen D, et al. Comparative evaluation of three assays for measurement of dengue virus neutralizing antibodies. The American journal of tropical medicine and hygiene. 2008;79(1):115-22. 311. Surveillance for Travel-Related Disease — GeoSentinel Surveillance System, United States, 1997–2011 [press release]. Morbidity and Mortality Weekly Report, 19 July 2013 2013. 312. Wichmann O, Gascon J, Schunk M, Puente S, Siikamaki H, Gjø I, et al. Severe Dengue Virus Infection in Travelers: Risk Factors and Laboratory Indicators. The Journal of Infectious Diseases. 2007;195(8):1089-96. 313. Vinner L, Domingo C, Ostby AC, Rosenberg K, Fomsgaard A. Cases of travel-acquired dengue fever in Denmark 2001-2009. Clinical microbiology and infection : the official publication of the European Society of Clinical Microbiology and Infectious Diseases. 2012;18(2):171-6. 314. Baaten GG, Sonder GJ, Zaaijer HL, van Gool T, Kint JA, van den Hoek A. Travel-related dengue virus infection, The Netherlands, 2006-2007. Emerg Infect Dis. 2011;17(5):821-8. 315. Karin L, Joseph T, John SB, Mary EW, Jay SK, Elizabeth B, et al. Travel-associated Illness Trends and Clusters, 2000–2010. Emerging Infectious Disease journal. 2013;19(7):1049. 316. Polwiang S. The estimation of imported dengue virus from Thailand. Journal of travel medicine. 2015;22(3):194-9. 317. Tuiskunen A, Hjertqvist M, Vene S, Lundkvist Å. Dengue fever in returned Swedish travelers from Thailand. Infection Ecology & Epidemiology. 2011;1:10.3402/iee.v1i0.7240. 318. Huang X, Yakob L, Devine G, Frentiu FD, Fu S-Y, Hu W. Dynamic spatiotemporal trends of imported dengue fever in Australia. 2016;6:30360. 319. Megawati D, Masyeni S, Yohan B, Lestarini A, Hayati RF, Meutiawati F, et al. Dengue in Bali: Clinical characteristics and genetic diversity of circulating dengue viruses. PLOS Neglected Tropical Diseases. 2017;11(5):e0005483. 320. Saiz JC, Vazquez-Calvo A, Blazquez AB, Merino-Ramos T, Escribano-Romero E, Martin-Acebes MA. Zika Virus: the Latest Newcomer. Frontiers in microbiology. 2016;7:496. 321. Dupont-Rouzeyrol M, O'Connor O, Calvez E, Daures M, John M, Grangeon JP, et al. Co-infection with Zika and dengue viruses in 2 patients, New Caledonia, 2014. Emerg Infect Dis. 2015;21(2):381-2. 322. Kwong JC, Druce JD, Leder K. Zika virus infection acquired during brief travel to Indonesia. The American journal of tropical medicine and hygiene. 2013;89(3):516-7. 323. Smith DJ, Lapedes AS, de Jong JC, Bestebroer TM, Rimmelzwaan GF, Osterhaus ADME, et al. Mapping the Antigenic and Genetic Evolution of Influenza Virus. Science. 2004;305(5682):371. 324. Bedford T, Suchard MA, Lemey P, Dudas G, Gregory V, Hay AJ, et al. Integrating influenza antigenic dynamics with molecular evolution. eLife. 2014;3:e01914. 325. Katzelnick LC, Fonville JM, Gromowski GD, Arriaga JB, Green A, James SL, et al. Dengue viruses cluster antigenically but not as discrete serotypes. Science (New York, NY). 2015;349(6254):1338-43. 326. Friberg H, Jaiswal S, West K, O'Ketch M, Rothman AL, Mathew A. Analysis of Human Monoclonal Antibodies Generated by Dengue Virus-Specific Memory B Cells. Viral Immunology. 2012;25(5):348-59.

333
327. Cao-Lormeau V-M, Roche C, Musso D, Mallet H-P, Dalipanda T, Dofai A, et al. Dengue Virus Type 3, South Pacific Islands, 2013. Emerging Infectious Diseases. 2014;20(6):1034-6. 328. Pierson Theodore C, Kuhn Richard J. Capturing a Virus while It Catches Its Breath. Structure. 2012;20(2):200-2. 329. Moi ML, Takasaki T, Kurane I. Human antibody response to dengue virus: implications for dengue vaccine design. Tropical Medicine and Health. 2016;44(1):1. 330. Tsai W-Y, Durbin A, Tsai J-J, Hsieh S-C, Whitehead S, Wang W-K. Complexity of Neutralizing Antibodies against Multiple Dengue Virus Serotypes after Heterotypic Immunization and Secondary Infection Revealed by In-Depth Analysis of Cross-Reactive Antibodies. Journal of virology. 2015;89(14):7348-62. 331. Katzelnick LC, Montoya M, Gresh L, Balmaseda A, Harris E. Neutralizing antibody titers against dengue virus correlate with protection from symptomatic infection in a longitudinal cohort. Proceedings of the National Academy of Sciences. 2016;113(3):728-33. 332. Kliks SC, Nisalak A, Brandt WE, Wahl L, Burke DS. Antibody-dependent enhancement of dengue virus growth in human monocytes as a risk factor for dengue hemorrhagic fever. American Journal of Tropical Medicine and Hygiene. 1989;40(4):444-51. 333. Halstead SB, O’Rourke EJ. Dengue viruses and mononuclear phagocytes. I. Infection enhancement by non-neutralizing antibody. J Exp Med. 1977;146. 334. Oliphant T, Nybakken GE, Engle M, Xu Q, Nelson CA, Sukupolvi-Petty S, et al. Antibody recognition and neutralization determinants on domains I and II of West Nile Virus envelope protein. Journal of virology. 2006;80(24):12149-59. 335. Sukupolvi-Petty S, Austin SK, Purtha WE, Oliphant T, Nybakken GE, Schlesinger JJ, et al. Type- and Subcomplex-Specific Neutralizing Antibodies against Domain III of Dengue Virus Type 2 Envelope Protein Recognize Adjacent Epitopes. Journal of virology. 2007;81(23):12816-26. 336. Drumond BP, Mondini A, Schmidt DJ, Bronzoni RVdM, Bosch I, Nogueira ML. Circulation of Different Lineages of Dengue Virus 2, Genotype American/Asian in Brazil: Dynamics and Molecular and Phylogenetic Characterization. PLOS ONE. 2013;8(3):e59422. 337. Dowd KA, DeMaso CR, Pierson TC. Genotypic Differences in Dengue Virus Neutralization Are Explained by a Single Amino Acid Mutation That Modulates Virus Breathing. mBio. 2015;6(6). 338. Thomas SJ, Nisalak A, Anderson KB, Libraty DH, Kalayanarooj S, Vaughn DW, et al. Dengue plaque reduction neutralization test (PRNT) in primary and secondary dengue virus infections: How alterations in assay conditions impact performance. The American journal of tropical medicine and hygiene. 2009;81(5):825-33. 339. Hunsawong T, Wichit S, Phonpakobsin T, Poolpanichupatam Y, Klungthong C, Latthiwongsakorn N, et al. Polytopic vaccination with a live-attenuated dengue vaccine enhances B-cell and T-cell activation, but not neutralizing antibodies. Heliyon. 2017;3(3):e00271. 340. Mansfield KL, Horton DL, Johnson N, Li L, Barrett ADT, Smith DJ, et al. Flavivirus-induced antibody cross-reactivity. The Journal of General Virology. 2011;92(Pt 12):2821-9. 341. Austin PC, Escobar M, Kopec JA. The use of the Tobit model for analyzing measures of health status. Quality of life research : an international journal of quality of life aspects of treatment, care and rehabilitation. 2000;9(8):901-10. 342. Glass DC, Gray CN. Estimating mean exposures from censored data: exposure to benzene in the Australian petroleum industry. The Annals of occupational hygiene. 2001;45(4):275-82. 343. Uh HW, Hartgers FC, Yazdanbakhsh M, Houwing-Duistermaat JJ. Evaluation of regression methods when immunological measurements are constrained by detection limits. BMC immunology. 2008;9:59.

334
344. Sukupolvi-Petty S, Brien JD, Austin SK, Shrestha B, Swayne S, Kahle K, et al. Functional Analysis of Antibodies against Dengue Virus Type 4 Reveals Strain-Dependent Epitope Exposure That Impacts Neutralization and Protection. Journal of virology. 2013;87(16):8826-42. 345. Haddow AD, Nasar F, Guzman H, Ponlawat A, Jarman RG, Tesh RB, et al. Genetic Characterization of Spondweni and Zika Viruses and Susceptibility of Geographically Distinct Strains of Aedes aegypti, Aedes albopictus and Culex quinquefasciatus (Diptera: Culicidae) to Spondweni Virus. PLoS Neglected Tropical Diseases. 2016;10(10):e0005083. 346. Weaver SC, Costa F, Garcia-Blanco MA, Ko AI, Ribeiro GS, Saade G, et al. Zika virus: History, emergence, biology, and prospects for control. Antiviral Research. 2016;130:69-80. 347. Faye O, Freire CC, Iamarino A, Faye O, de Oliveira JV, Diallo M, et al. Molecular evolution of Zika virus during its emergence in the 20(th) century. PLoS Negl Trop Dis. 2014;8(1):e2636. 348. Petersen LR, Jamieson DJ, Powers AM, Honein MA. Zika Virus. New England Journal of Medicine. 2016;374(16):1552-63. 349. Duffy MR, Chen T-H, Hancock WT, Powers AM, Kool JL, Lanciotti RS, et al. Zika Virus Outbreak on Yap Island, Federated States of Micronesia. New England Journal of Medicine. 2009;360(24):2536-43. 350. Hayes EB. Zika virus outside Africa. Emerg Infect Dis. 2009;15(9):1347-50. 351. Faye O, Freire CCM, Iamarino A, Faye O, de Oliveira JVC, Diallo M, et al. Molecular Evolution of Zika Virus during Its Emergence in the 20(th) Century. PLoS Neglected Tropical Diseases. 2014;8(1):e2636. 352. WHO. Zika situation report. 2016 17-08-2016. Report No. 353. Oehler E WL, Larre P, Leparc-Goffart I, Lastère S, Valour F, Baudouin L, Mallet HP, Musso D, Ghawche F Zika virus infection complicated by Guillain-Barré syndrome – case report, French Polynesia,December 2013. Euro Surveill. 2014;19(9). 354. Lazear HM, Diamond MS. Zika Virus: New clinical syndromes and its emergence in the Western Hemisphere. Journal of virology. 2016:JVI. 00252-16. 355. Dai L, Song J, Lu X, Deng YQ, Musyoki AM, Cheng H, et al. Structures of the Zika Virus Envelope Protein and Its Complex with a Flavivirus Broadly Protective Antibody. Cell host & microbe. 2016;19(5):696-704. 356. Barba-Spaeth G, Dejnirattisai W, Rouvinski A, Vaney MC, Medits I, Sharma A, et al. Structural basis of potent Zika-dengue virus antibody cross-neutralization. Nature. 2016;536(7614):48-53. 357. Priyamvada L, Quicke KM, Hudson WH, Onlamoon N, Sewatanon J, Edupuganti S, et al. Human antibody responses after dengue virus infection are highly cross-reactive to Zika virus. Proceedings of the National Academy of Sciences of the United States of America. 2016;113(28):7852-7. 358. Se-Thoe SY, Ng MM, Ling AE. Retrospective study of Western blot profiles in immune sera of natural dengue virus infections. Journal of medical virology. 1999;57(3):322-30. 359. Oceguera LF, 3rd, Patiris PJ, Chiles RE, Busch MP, Tobler LH, Hanson CV. Flavivirus serology by Western blot analysis. The American journal of tropical medicine and hygiene. 2007;77(1):159-63. 360. Hsieh SC, Tsai WY, Nerurkar VR, Wang WK. Characterization of the ectodomain of the envelope protein of dengue virus type 4: expression, membrane association, secretion and particle formation in the absence of precursor membrane protein. PLoS One. 2014;9(6):e100641. 361. Wikan N, Suputtamongkol Y, Yoksan S, Smith DR, Auewarakul P. Immunological evidence of Zika virus transmission in Thailand. Asian Pacific journal of tropical medicine. 2016;9(2):141-4. 362. Leung GH, Baird RW, Druce J, Anstey NM. ZIKA VIRUS INFECTION IN AUSTRALIA FOLLOWING A MONKEY BITE IN INDONESIA. The Southeast Asian journal of tropical medicine and public health. 2015;46(3):460-4.

335
363. WHO. Zika infections acquired by travellers to countries without recognized virus circulation. 364. Wang Q, Yan J, Gao GF. Monoclonal antibodies against Zika virus: Therapeutics and their implications for vaccine design. Journal of virology. 2017. 365. Stettler K, Beltramello M, Espinosa DA, Graham V, Cassotta A, Bianchi S, et al. Specificity, cross-reactivity and function of antibodies elicited by Zika virus infection. Science. 2016. 366. Collins MH, McGowan E, Jadi R, Young E, Lopez CA, Baric RS, et al. Lack of Durable Cross-Neutralizing Antibodies Against Zika Virus from Dengue Virus Infection. Emerging Infectious Diseases. 2017;23(5):773-81. 367. Dai L, Song J, Lu X, Deng Y-Q, Musyoki Abednego M, Cheng H, et al. Structures of the Zika Virus Envelope Protein and Its Complex with a Flavivirus Broadly Protective Antibody. Cell Host & Microbe. 2016;19(5):696-704. 368. Smith DR, Sprague TR, Hollidge BS, Valdez SM, Padilla SL, Bellanca SA, et al. African and Asian Zika Virus Isolates Display Phenotypic Differences Both In Vitro and In Vivo. The American journal of tropical medicine and hygiene. 2017. 369. Dowd Kimberly A, DeMaso Christina R, Pelc Rebecca S, Speer Scott D, Smith Alexander RY, Goo L, et al. Broadly Neutralizing Activity of Zika Virus-Immune Sera Identifies a Single Viral Serotype. Cell Reports. 2016;16(6):1485-91. 370. Ye Q, Liu Z-Y, Han J-F, Jiang T, Li X-F, Qin C-F. Genomic characterization and phylogenetic analysis of Zika virus circulating in the Americas. Infection, Genetics and Evolution. 2016;43:43-9. 371. Zhu Z, Chan JF-W, Tee K-M, Choi GK-Y, Lau SK-P, Woo PC-Y, et al. Comparative genomic analysis of pre-epidemic and epidemic Zika virus strains for virological factors potentially associated with the rapidly expanding epidemic. Emerg Microbes Infect. 2016;5:e22. 372. Giovanetti M, Milano T, Alcantara LC, Carcangiu L, Cella E, Lai A, et al. Zika Virus spreading in South America: Evolutionary analysis of emerging neutralizing resistant Phe279Ser strains. Asian Pacific journal of tropical medicine. 2016;9(5):445-52. 373. Vogt MR, Dowd KA, Engle M, Tesh RB, Johnson S, Pierson TC, et al. Poorly neutralizing cross-reactive antibodies against the fusion loop of West Nile virus envelope protein protect in vivo via Fcgamma receptor and complement-dependent effector mechanisms. Journal of virology. 2011;85(22):11567-80. 374. Dejnirattisai W, Wongwiwat W, Supasa S, Zhang X, Dai X, Rouvinski A, et al. A new class of highly potent, broadly neutralizing antibodies isolated from viremic patients infected with dengue virus. Nature immunology. 2015;16(2):170-7. 375. Mukherjee S, Dowd KA, Manhart CJ, Ledgerwood JE, Durbin AP, Whitehead SS, et al. Mechanism and significance of cell type-dependent neutralization of flaviviruses. Journal of virology. 2014;88(13):7210-20. 376. Cardosa MJ, Wang SM, Sum MSH, Tio PH. Antibodies against prM protein distinguish between previous infection with dengue and Japanese encephalitis viruses. BMC Microbiology. 2002;2(1):9. 377. Rastogi M, Sharma N, Singh SK. Flavivirus NS1: a multifaceted enigmatic viral protein. Virology Journal. 2016;13:131. 378. Hua R-H, Liu L-K, Chen Z-S, Li Y-N, Bu Z-G. Comprehensive Mapping Antigenic Epitopes of NS1 Protein of Japanese Encephalitis Virus with Monoclonal Antibodies. PLOS ONE. 2013;8(6):e67553. 379. Tsunoda I, Omura S, Sato F, Kusunoki S, Fujita M, Park A-M, et al. Neuropathogenesis of Zika Virus Infection : Potential Roles of Antibody-Mediated Pathology. Acta medica Kinki University. 2016;41(2):37-52. 380. Flipse J, Diosa-Toro MA, Hoornweg TE, van de Pol DPI, Urcuqui-Inchima S, Smit JM. Antibody-Dependent Enhancement of Dengue Virus Infection in Primary Human Macrophages; Balancing Higher Fusion against Antiviral Responses. Scientific Reports. 2016;6:29201.

336
381. Katzelnick LC, Gresh L, Halloran ME, Mercado JC, Kuan G, Gordon A, et al. Antibody-dependent enhancement of severe dengue disease in humans. Science. 2017;358(6365):929. 382. Kurane I, Mady BJ, Ennis FA. Antibody-dependent enhancement of dengue virus infection. Reviews in Medical Virology. 1991;1(4):211-21. 383. Halstead SB, Rourke EJ. Dengue viruses and mononuclear phagocytes. I. Infection enhancement by non-neutralizing antibody. The Journal of Experimental Medicine. 1977;146(1):201. 384. Halstead SB. Dengue haemorrhagic fever — a public health problem and a field for research. Bulletin of the World Health Organization. 1980;58(1):1-21. 385. Halstead SB. Pathogenesis of dengue: challenges to molecular biology. Science. 1988;239(4839):476. 386. Halstead SB, Casals J, Shotwell H, Palumbo N. Studies on the immunization of monkeys against dengue. I. Protection derived from single and sequential virus infections. The American journal of tropical medicine and hygiene. 1973;22(3):365-74. 387. Kouri GP, Guzmán MG, Bravo JR, Triana C. Dengue haemorrhagic fever/dengue shock syndrome: lessons from the Cuban epidemic, 1981. Bulletin of the World Health Organization. 1989;67(4):375-80. 388. Chye JK, Lim CT, Ng KB, Lim JMH, George R, Lam SK. Vertical Transmission of Dengue. Clinical Infectious Diseases. 1997;25(6):1374-7. 389. Kliks SC, Nimmanitya S, Nisalak A, Burke DS. Evidence that maternal dengue antibodies are important in the development of dengue hemorrhagic fever in infants. The American journal of tropical medicine and hygiene. 1988;38(2):411-9. 390. Martinez E, Guzman MG, Valdes M, Soler M, Kouri G. [Dengue fever and hemorrhagic dengue in infants with a primary infection]. Revista cubana de medicina tropical. 1993;45(2):97-101. 391. Vaughn DW, Green S, Kalayanarooj S, Innis BL, Nimmannitya S, Suntayakorn S, et al. Dengue Viremia Titer, Antibody Response Pattern, and Virus Serotype Correlate with Disease Severity. The Journal of Infectious Diseases. 2000;181(1):2-9. 392. Updated Questions and Answers related to the dengue vaccine Dengvaxia® and its use [Internet]. 2017. Available from: http://www.who.int/immunization/diseases/dengue/q_and_a_dengue_vaccine_dengvaxia_use/en/. 393. Flasche S, Jit M, Rodríguez-Barraquer I, Coudeville L, Recker M, Koelle K, et al. The Long-Term Safety, Public Health Impact, and Cost-Effectiveness of Routine Vaccination with a Recombinant, Live-Attenuated Dengue Vaccine (Dengvaxia): A Model Comparison Study. PLoS Medicine. 2016;13(11):e1002181. 394. Aguiar M, Stollenwerk N, Halstead SB. The risks behind Dengvaxia recommendation. The Lancet Infectious Diseases. 2016;16(8):882-3. 395. Sirohi D, Chen Z, Sun L, Klose T, Pierson TC, Rossmann MG, et al. The 3.8 A resolution cryo-EM structure of Zika virus. Science. 2016;352(6284):467-70. 396. Boonnak K, Slike BM, Burgess TH, Mason RM, Wu S-J, Sun P, et al. Role of Dendritic Cells in Antibody-Dependent Enhancement of Dengue Virus Infection. Journal of virology. 2008;82(8):3939-51. 397. Wallace MJ, Smith DW, Broom AK, Mackenzie JS, Hall RA, Shellam GR, et al. Antibody-dependent enhancement of Murray Valley encephalitis virus virulence in mice. Journal of General Virology. 2003;84(7):1723-8. 398. Goncalvez AP, Engle RE, St. Claire M, Purcell RH, Lai C-J. Monoclonal antibody-mediated enhancement of dengue virus infection <em>in vitro</em> and <em>in vivo</em> and strategies for prevention. Proceedings of the National Academy of Sciences. 2007;104(22):9422. 399. Ramasamy V, Arora U, Shukla R, Poddar A, Shanmugam RK, White LJ, et al. A tetravalent virus-like particle vaccine designed to display domain III of dengue envelope proteins induces multi-serotype neutralizing antibodies in mice and macaques which

337
confer protection against antibody dependent enhancement in AG129 mice. PLOS Neglected Tropical Diseases. 2018;12(1):e0006191. 400. Aubry M, Teissier Y, Mapotoeke M, Teissier A, Giard M, Musso D, et al. High risk of dengue type 2 outbreak in French Polynesia, 2017. Eurosurveillance. 2017;22(14):30505. 401. Cao-Lormeau V-M, Roche C, Aubry M, Teissier A, Lastere S, Daudens E, et al. Recent Emergence of Dengue Virus Serotype 4 in French Polynesia Results from Multiple Introductions from Other South Pacific Islands. PLOS ONE. 2011;6(12):e29555. 402. de Alwis R, Smith SA, Olivarez NP, Messer WB, Huynh JP, Wahala WMPB, et al. Identification of human neutralizing antibodies that bind to complex epitopes on dengue virions. Proceedings of the National Academy of Sciences of the United States of America. 2012;109(19):7439-44. 403. Charles AS, Christofferson RC. Utility of a Dengue-Derived Monoclonal Antibody to Enhance Zika Infection In Vitro. PLoS Currents. 2016;8:ecurrents.outbreaks.4ab8bc87c945eb41cd8a49e127082620.

338
Appendixes
List of Appendixes
Appendix 3. 1:- Hemagglutination and NS1 antigen test
Appendix 3. 2:- TCID50 determination of virus
Appendix 3. 3:- Checker board assay for primary and secondary antibody
Appendix 3. 4:-Reagents for preparation of virus antigen
Appendix 4.1:- Information sheet
Appendix 4.2:- Travel and Medical History Questionnaire
Appendix 4.3:- Participant consent form
Appendix 5.1.:- Neutralising antibody response over time in individuals with DENV-1
infection
Appendix 5.2.:- Neutralising antibody response over time in individuals with DENV-2
infection
Appendix 5.3.:- Neutralising antibody response over time in individuals with DENV-3
infection
Appendix 5.4.:- Neutralising antibody response over time in individuals with DENV-4
infection
Appendix 6.1.:- Neutralising antibody response over time in individuals with ZIKV
infection

339
Appendixes for Chapter 3
Appendix 3.1: Hemagglutination and NS1 antigen test
Protocol followed for HA and NS1 was established in PathWest reference laboratory.
a) Hemagglutination assay (HA)
The Haemagglutination assay is a pH dependent test for the detection of viral protein.
Dilution of virus supernatant was made up in BABS buffer made from 1: 5
concentration of bovine albumin and borate saline. For the dilution 1:1 serial dilution of
virus supernatant and BABS buffer was made in 96 well v-bottom plate. Each virus
sample was tested in pH buffer solution at pH’s 6.0, 6.2 and 6.4. Goose red blood cells
at the three pH’s was made and 50 µl goose cells was added to all wells. Plates were
gently tap to shake and sealed and read after an hour for presence or absence of cell
pellet. The presence of virus is indicated by the absence of a cell pellet and no virus by
the presence of a cell pellet at the bottom of the well.
b) NS1 antigen test
This test is a very sensitive and specific test for the detection of DENV only. This was
performed to confirm virus produced is dengue virus and detection of virus in HA
negative samples. NS1 antigen was carried out at PathWest using a commercial dengue
NS1 antigen-capture ELISA kit, PLATELIA DENGUE NS1 AG (Bio-Rad). This test is
based on a one-step sandwich microplate format. The test used murine monoclonal
antibody (MAb) for capture and revelation. If NS1 antigen was present in the sample an
immune complex MAb-NS1-MAb/peroxidase would be formed. The test was
performed and calculations were made according to manufacturer’s protocol.

340
Appendix 3.2 : Tissue Culture Infective Dose 50 Assay (TCID50)
TCID50 was done to quantity of virus that will produce a cytopathic effect (CPE) in
50% of the cultures inoculated to make sure virus cultured would give optimal CPE in
further tests. End-point determination was done by ELISA using 4G2 monoclonal
antibody and goat anti-mouse antibody labelled with horse radish peroxidase. Briefly,
serial dilution of virus supernatant was grown in 96 well round bottom plates. After 7
days cells were fixed with 20% PBS/Acetone for 90 minutes in 4oC. After fixing cells
were blocked with 2% casein solution for an hour at room temperature. Cells were then
labelled with primary antibody 4G2 monoclonal antibody followed by GAM labelled
secondary antibody. Plates were washed in between primary and secondary antibody for
4X with ELISA buffer. ABTS substrate with 30% H2O2 was used as substrate and
optical densities of plate were read. 4G2 and GAM concentration was used as
previously stated in lab.
Appendix 3.3: Checker board assay for determination of 4G2 and GAM concentration
for Focus assay.
Different dilution of GAM and 4G2 was titrated to get optimal concentration that would
give visible spots in focus assay. For assay, known concentration of virus (optimal
concentration as determined by TCID50 that will give 50% CPE) was grown on 24 well
plates. Each well was inoculated with same amount of virus and 2 wells were left as cell
control (no virus). After 5-7 days plates were fixed, blocked with blocking buffer and
stained with different concentration pair of primary and secondary antibody. TrueBlue
peroxidase was used as substrate. Optimal concentration of that gives clear distinct
spots was used in further assays.

341
Appendix 3.4: Reagents for preparation of virus antigen
All the reagents were prepared following standardised protocol from PathWest.
Preparation of 0.175N sodium hydroxide
0.70g NaOH was dissolved in 100ml ultrapure water. Solution was filter sterilised with
022µ filter and stored at room temperature for <= 3 months.
Preparation of 1M sodium thiosulphate
28.82gm of Na2O2S25.H20 was dissolved in 100ml ultrapure water and sterilised with
0.22µ filter. Solution was stored at 4oC for <3 months.
Preparation of 0.1M Binary Ethylenimine (BEI)
0.20489gm of BEI was dissolved in 10 ml 0.175N NaOH. Solution was incubated at
37oC water bath for 60 mins. BEI was stored in 4
oC and used within 24hrs.

342
Participant Information Sheet
CHIEF INVESTIGATOR
Dr Allison Imrie, University of Western Australia, School of Biomedical Sciences; Telephone: (618) 6457 1377; Fax: (618) 6457 2912; email: [email protected]
PROJECT TITLE
Immune Responses in Mosquito-Borne Virus Infection
PURPOSE
You are being asked to take part in this research study because you were previously diagnosed with a mosquito-borne viral infection. Around the time of your infection, your immune system responded by producing antibodies and forming memory T and B cells (white blood cells or lymphocytes) and these cells can be found in your blood for many years after your infection. The purpose of this study is to learn more about your immune responses to the mosquito-borne viruses, and how these responses may contribute to the diseases these viruses cause. We will test antibodies and lymphocytes taken from your blood to see how they recognize mosquito-borne viruses in vitro, that is, in the laboratory.
In order to decide whether or not you wish to be in this study, you will need to know about any good or bad things that could happen to you if you decide to join. This process is called ‘informed consent’. Once you understand the study, and if you agree to take part,
you will be asked to sign a consent form. You will be given a copy to keep. EXPECTED LENGTH OF STUDY
The estimated length of this study is 5 years, however you can stop participating at any time within this 5-year period. We may ask you to give blood each year so that we can look at how your immune responses change over time.
PROCEDURES
To enter into this study, you must be at least 18 years old and be willing to sign a consent form.
First Study Visit At this visit you will:
Talk about the study with the Investigator
Read and sign the consent form after all your questions are answered
Complete a brief questionnaire on your travel history Study staff will take 150 ml of blood (about 8 tablespoons; about 1/3 the volume of a standard blood bank donation), which will be used to look at how your immune cells
School of Biomedical Sciences
The Faculty of Health and Medical Sciences
The University of Western Australia
M502, 35 Stirling Highway
Crawley WA 6009 Australia
E: [email protected] CRICOS Provider Code: 00126G
Appendix 4.1:- Information sheet
Appendix

343
respond to mosquito-borne viruses, in the laboratory. Blood will be collected from your arm vein using the same method as for routine medical blood tests. This may be performed by an experienced blood collector or a trainee under the supervision of an experienced blood collector. If the person collecting your blood is a trainee, you will be informed and have the option of electing to have a more experienced person do that task. This visit will take an estimated 45 minutes. Follow-up Visits The blood sample you donate at your first visit will be tested in the laboratory, and depending on the results we may ask you to return for a follow-up visit at which:
We may ask you to donate a large (150 ml) or small (30 ml) volume of blood. The amount you donate will depend on the kind of immune response we have measured in your blood, which we need to analyze in greater detail.
You will complete a brief questionnaire on your travel history This visit will take an estimated 30 minutes.
Depending on the results of the laboratory tests, we may ask you to return each year, for up to 5 years.
Blood collected from you will only be used for the purposes described here. Analysis of your blood sample(s) for immune responses against mosquito-borne viruses can take several weeks to complete. The results from this study will be published in medical and scientific journals and presented at national and international scientific meetings. Study participants’ names are never used in any form of report or discussion associated with this project.
RISKS AND DISCOMFORTS
Taking blood may cause some soreness, bleeding and bruising, and (very rarely) infection where the needle enters the body. A person may (very rarely) feel faint or faint when blood is taken. Although 150 ml of blood may seem like quite a large amount of blood to give, this is about one-third of the amount collected from a blood donor at the Blood Bank.
Risk of infection is slight since only sterile one-time equipment is used.
BENEFITS
You may receive no benefit from this study. However, knowledge gained from this study may, in future, benefit others who are infected with mosquito-borne viruses, such as dengue, Zika, chikungunya, Ross River, and Murray Valley. It may also help in the development of a vaccine to prevent infection in future generations.
CONFIDENTIALITY
Your research records will be confidential to the extent permitted by law. You will not be personally identified in any publication about this study. A code, which will be known only to study personnel and you, will be used instead of your name on laboratory records in this study. The code will be stored in a safe place in the Chief Investigator’s office. Personal information about your test results will not be given to anyone without your written permission. Individual results may be provided upon request and where available, though any data generated from this study are for research purposes only and not for diagnostic purposes.
COMPENSATION FOR INJURY

344
Your participation in this study does not prejudice any right to compensation, which you may have under statute or common law.
VOLUNTARY CONSENT AND CERTIFICATION
You take part in this study of your own free will and you can stop at any time for any reason. Your consent does not take away any of your legal rights in case of negligence or carelessness on the part of anyone working on this project.
ETHICS
Approval to conduct this research has been provided by the University of Western Australia, in accordance with its ethics review and approval procedures. Any person considering participation in this research project, or agreeing to participate, may raise any questions or issues with the researchers at any time. In addition, any person not satisfied with the response of researchers may raise ethics issues or concerns, and may make any complaints about this research project by contacting the Human Ethics Office at the University of Western Australia on (08) 6488 3703 or by emailing to [email protected]. All research participants are entitled to retain a copy of any Participant Information Form and/or Participant Consent Form relating to this research project.
CONTACTS To contact the research team, please call the Mosquito-Borne Virus Study Phone: (08)
6457 1129.

345
Appendix 4.2:- Travel and Medical History Questionnaire

346
Participant Consent Form
Immune Responses in Mosquito-Borne Virus Infection
Chief Investigator: Associate Professor Allison Imrie
I (the participant) have read the information provided and any questions I have asked
have been answered to my satisfaction. I agree to participate in this activity, realizing
that I may withdraw at any time without reason and without prejudice.
I understand that all attributable information that I provide is treated as strictly
confidential and will not be released by the investigator in any form that may identify
me. The only exception to this principle of confidentiality is if documents are required
by law.
I have been advised as to what data is being collected, the purpose for collecting the
data, and what will be done with the data upon completion of the research.
I have been informed that individual results may be provided upon request and where
available, though any data generated from this study are for research purposes only and
not for diagnostic purposes.
I agree that research data gathered for the study may be published provided my name or
other identifying information is not used.
________________________________________________________________________
_
Participant Name Participant Signature Date
Approval to conduct this research has been provided by the University of Western
Australia, in accordance with its ethics review and approval procedures. Any person
considering participation in this research project, or agreeing to participate, may raise any
questions or issues with the researchers at any time.
In addition, any person not satisfied with the response of researchers may raise ethics
issues or concerns, and may make any complaints about this research project by
contacting the Human Ethics Office at the University of Western Australia on (08) 6488
3703 or by emailing to [email protected].
All research participants are entitled to retain a copy of any Participant Information Form
and/or Participant Consent Form relating to this research project
School of Biomedical Sciences
The Faculty of Health and Medical Sciences
The University of Western Australia
M502, 35 Stirling Highway
Crawley WA 6009 Australia
CRICOS Provider Code: 00126G
Appendix 4. 3.:- Participant consent
form

347
Appendix for Chapter 5 : Antibody mediated immune response in monotypic
dengue
This appendix consists of nonlinear regression curve and comparative mean reduction
analysis for each DENV individuals at different time post infection. Each graph title
represents the study number followed by their infecting serotype and time of sample
collection post infection. Each sera panel is separated by a page gap between the graphs.
Graphs are arranged according to their study number. Non-linear regression curve for
each sample is illustrated as its antibody response against each serotype of DENV. In
mean reduction curve x-axis represents strains of DENV tested and y-axis represents
mean reduction percentage for each strain of virus strains.

348
Appendix 5.1.:- Neutralising antibody response over time in individuals with
DENV-1 infection
DENV-1 SERA PANEL
Number of individuals: 6
Number of individuals with multiple
sampling points:-5

349

350
D1-H
W2001G
4
D1-1
0B
G1L
14
D1-1
5L
G1
D1-1
0B
G2L
4
D1-1
0IN
DG
3L
4
D1-1
4P
HL
G4
D2-N
GC
GA
II
D2-1
0L
GA
1L
9
D2-1
1B
GC
L4
D2-1
3M
CL
N
D2-1
5B
GC
L4
D2-1
5B
CL
N
D2-
PR
6193V
D2 -
PR
1940N
V
D3-H
87G
1
D3-1
5B
G1
D3-
UN
C 3
002V
D3-
UN
C 3
008 N
V
D4-H
24G
5
D4-1
0B
G3L
1
D4-1
0B
G3L
2
ZIK
V M
R 7
66
ZIK
V P
RV
AB
C59
YF
17D
JE
IM
OJE
V
Au
tolo
go
us v
iru
s
0
1 0 0
1 5 0
F L V 0 0 1 V 2 (D 1 :~ 5 y e a rs )
V iru s ty p e
Me
an
Re
du
cti
on
%
50
75
90
D1-H
W2001G
4
D1-1
0B
G1L
14
D1-1
5L
G1
D1-1
0B
G2L
4
D1-1
0IN
DG
3L
4
D1-1
4P
HL
G4
D2-N
GC
GA
II
D2-1
0L
GA
1L
9
D2-1
1B
GC
L4
D2-1
3M
CL
N
D2-1
5B
GC
L4
D2-1
5B
CL
N
D2-
PR
6193V
D2 -
PR
1940N
V
D3-H
87G
5
D3-1
5B
G1
D3-
UN
C 3
002V
D3-
UN
C 3
008 N
V
D4-H
24G
1
D4-1
0B
G3L
1
D4-1
0B
G3L
2
ZIK
V M
R 7
66
ZIK
V P
RV
AB
C59
YF
17D
JE
IM
OJE
V
Au
tolo
go
us v
iru
s
0
1 0 0
1 5 0
F L V 0 0 1 V 3 (D 1 :~ 6 y e a rs )
V iru s ty p e
Me
an
Re
du
cti
on
%
50
75
90
D1-H
W2001G
4
D1-1
0B
G1L
14
D1-1
5L
G1
D1-1
0B
G2L
4
D1-1
0IN
DG
3L
4
D1-1
4P
HL
G4
D2-N
GC
GA
II
D2-1
0L
GA
1L
9
D2-1
1B
GC
L4
D2-1
3M
CL
N
D2-1
5B
GC
L4
D2-1
5B
CL
N
D2-P
R 6
913V
D2 -
PR
1940N
V
D3-H
87G
5
D3-1
5B
G1
D3-
UN
C 3
002V
D3-
UN
C 3
008 N
V
D4-H
24G
1
D4-1
0B
G3L
1
D4-1
0B
G3L
2
ZIK
V M
R 7
66
YF
17D
JE
IM
OJE
V
ZIK
V P
RV
AB
C59
Au
tolo
go
us v
iru
s
0
1 0 0
1 5 0
F L V 0 0 1 V 1 (D 1 :- 2 y e a rs )
Me
an
Re
du
cti
on
%
50
75
90

351

352
D1-H
W2001G
4
D1-1
0B
G1L
14
D1-1
5L
G1
D1-1
0B
G2L
4
D1-1
0IN
DG
3L
4
D1-1
4P
HL
G4
D2-N
GC
GA
II
D2-1
0L
GA
1L
9
D2-1
1B
GC
L4
D2-1
3M
CL
N
D2-1
5B
GC
L4
D2-1
5B
CL
N
D2-
PR
6193V
D2 -
PR
1940N
V
D3-H
87G
5
D3-1
5B
G1
D3-
UN
C 3
002V
D3-
UN
C 3
008 N
V
D4-H
24G
1
D4-1
0B
G3L
1
D4-1
0B
G3L
2
ZIK
V M
R 7
66
ZIK
V P
RV
AB
C59
YF
17D
JE
IM
OJE
V
0
1 0 0
1 5 0
F L V 0 1 1 V 2 (D 1 :2 y e a rs -1 0 m o n th s )
V iru s ty p e
Me
an
Re
du
cti
on
%
50
75
90
D1-H
W2001G
4
D1-1
0B
G1L
14
D1-1
5L
G1
D1-1
0B
G2L
4
D1-1
0IN
DG
3L
4
D1-1
4P
HL
G4
D2-N
GC
GA
II
D2-1
0L
GA
1L
9
D2-1
1B
GC
L4
D2-1
3M
CL
N
D2-1
5B
GC
L4
D2-1
5B
CL
N
D2-
PR
6193V
D2 -
PR
1940N
V
D3-H
87G
5
D3-1
5B
G1
D3-
UN
C 3
002V
D3-
UN
C 3
008 N
V
D4-H
24G
1
D4-1
0B
G3L
1
D4-1
0B
G3L
2
ZIK
V M
R 7
66
ZIK
V P
RV
AB
C59
YF
17D
JE
IM
OJE
V
0
1 0 0
1 5 0
F L V 0 1 1 V 1 (D 1 :9 d a y s )
V iru s ty p e
Me
an
Re
du
cti
on
%
50
75
90
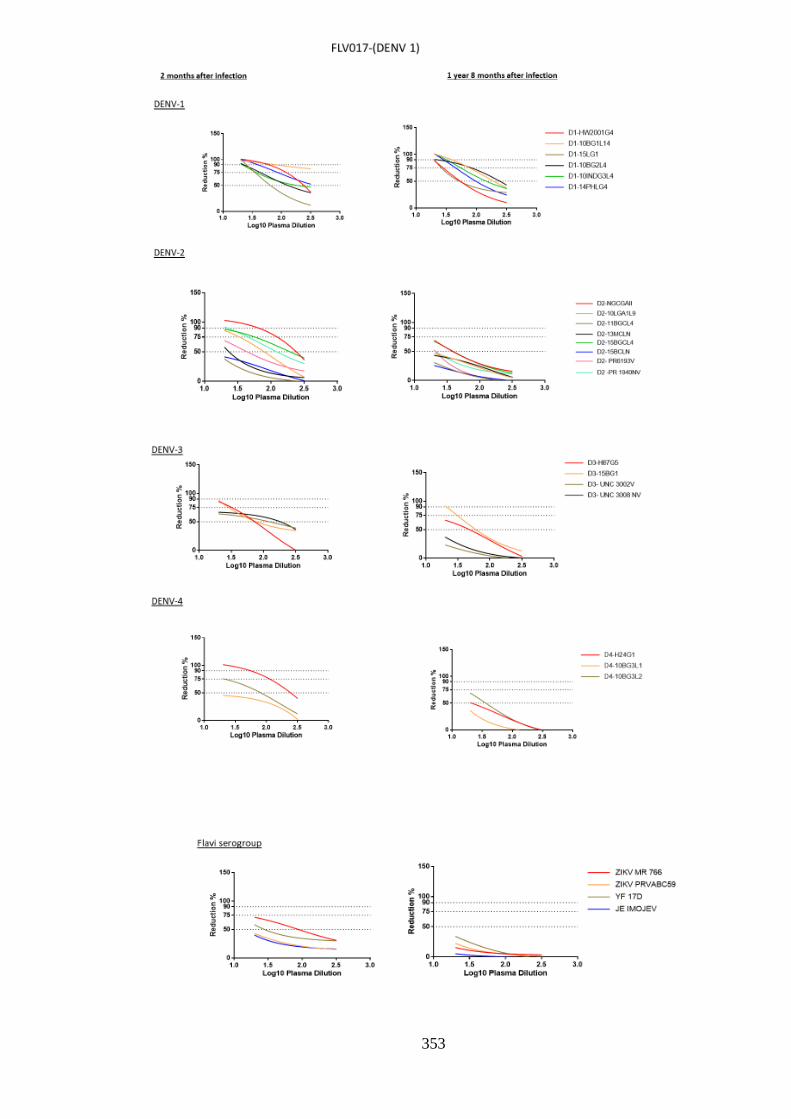
353

354
D1-H
W2001G
4
D1-1
0B
G1L
14
D1-1
5L
G1
D1-1
0B
G2L
4
D1-1
0IN
DG
3L
4
D1-1
4P
HL
G4
D2-N
GC
GA
II
D2-1
0L
GA
1L
9
D2-1
1B
GC
L4
D2-1
3M
CL
N
D2-1
5B
GC
L4
D2-1
5B
CL
N
D2-
PR
6193V
D2 -
PR
1940N
V
D3-H
87G
5
D3-1
5B
G1
D3-
UN
C 3
002V
D3-
UN
C 3
008 N
V
D4-H
24G
1
D4-1
0B
G3L
1
D4-1
0B
G3L
2
ZIK
V M
R 7
66
ZIK
V P
RV
AB
C59
YF
17D
JE
IM
OJE
V
0
1 0 0
1 5 0
F L V 0 1 7 V 2 (D 1 : 1 y e a r 8 m o n th s )
V iru s ty p e
Me
an
Re
du
cti
on
%
50
75
90
D1-H
W2001G
4
D1-1
0B
G1L
14
D1-1
5L
G1
D1-1
0B
G2L
4
D1-1
0IN
DG
3L
4
D1-1
4P
HL
G4
D2-N
GC
GA
II
D2-1
0L
GA
1L
9
D2-1
1B
GC
L4
D2-1
3M
CL
N
D2-1
5B
GC
L4
D2-1
5B
CL
N
D2-
PR
6193V
D2 -
PR
1940N
V
D3-H
87G
5
D3-1
5B
G1
D3-
UN
C 3
002V
D3-
UN
C 3
008 N
V
D4-H
24G
1
D4-1
0B
G3L
1
D4-1
0B
G3L
2
ZIK
V M
R 7
66
ZIK
V P
RV
AB
C59
YF
17D
JE
IM
OJE
V
0
1 0 0
1 5 0
F L V 0 1 7 V 1 (D 1 :2 m o n th s )
V iru s ty p e
Me
an
Re
du
cti
on
%
50
75
90

355

356
D1-H
W2001G
4
D1-1
0B
G1L
14
D1-1
5L
G1
D1-1
0B
G2L
4
D1-1
0IN
DG
3L
4
D1-1
4P
HL
G4
D2-N
GC
GA
II
D2-1
0L
GA
1L
9
D2-1
1B
GC
L4
D2-1
3M
CL
N
D2-1
5B
GC
L4
D2-1
5B
CL
N
D2-
PR
6193V
D2 -
PR
1940N
V
D3-H
87G
5
D3-1
5B
G1
D3-
UN
C 3
002V
D3-
UN
C 3
008 N
V
D4-H
24G
1
D4-1
0B
G3L
1
D4-1
0B
G3L
2
ZIK
V M
R 7
66
ZIK
V P
RV
AB
C59
YF
17D
JE
IM
OJE
V
Au
tolo
go
us V
iru
s
0
1 0 0
1 5 0
F L V 0 1 8 V 2 (D 1 - 1 y e a r 8 m o n th s )
Re
du
cti
on
%
50
75
90
D1-H
W2001G
4
D1-1
0B
G1L
14
D1-1
5L
G1
D1-1
0B
G2L
4
D1-1
0IN
DG
3L
4
D1-1
4P
HL
G4
D2-N
GC
GA
II
D2-1
0L
GA
1L
9
D2-1
1B
GC
L4
D2-1
3M
CL
N
D2-1
5B
GC
L4
D2-1
5B
CL
N
D2-
PR
6193V
D2 -
PR
1940N
V
D3-H
87G
5
D3-1
5B
G1
D3-
UN
C 3
002V
D3-
UN
C 3
008 N
V
D4-H
24G
1
D4-1
0B
G3L
1
D4-1
0B
G3L
2
ZIK
V M
R 7
66
ZIK
V P
RV
AB
C59
YF
17D
JE
IM
OJE
V
Au
tolo
go
us V
iru
s
0
1 0 0
1 5 0
F L V 0 1 8 V 3 (D 1 -2 y e a rs -7 m o n th s )
Re
du
cti
on
%
50
75
90
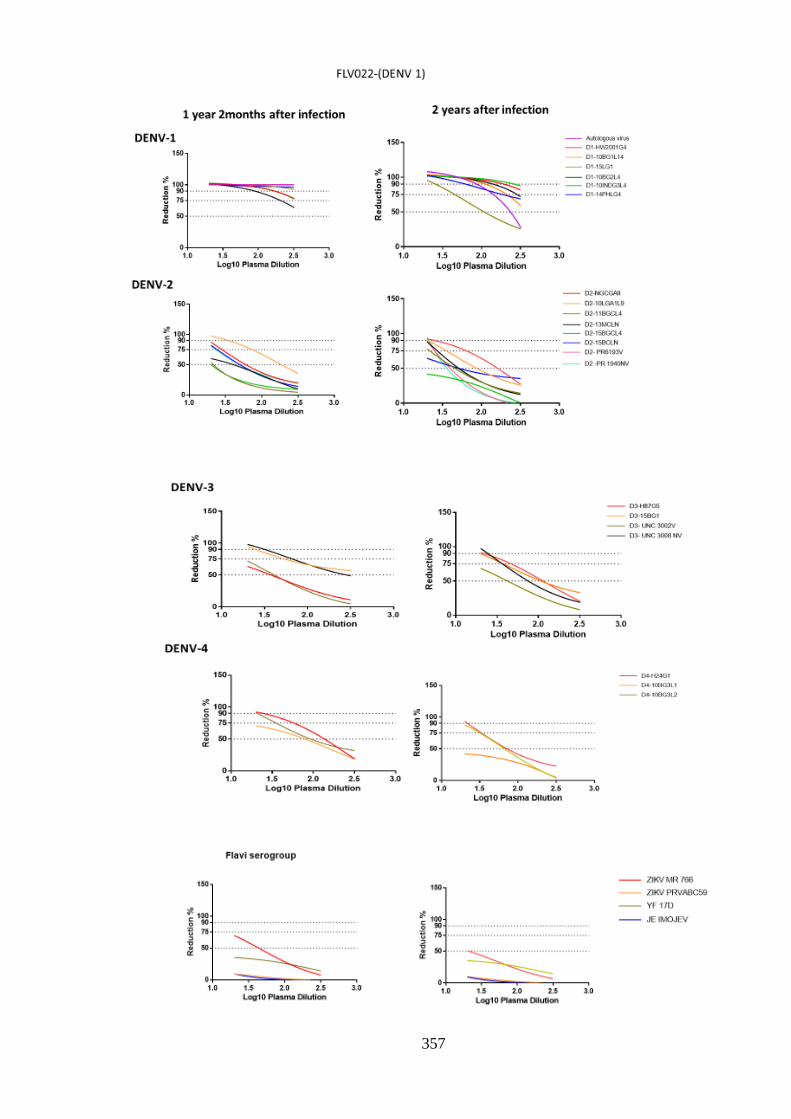
357

358
D1-H
W2001G
4
D1-1
0B
G1L
14
D1-1
5L
G1
D1-1
0B
G2L
4
D1-1
0IN
DG
3L
4
D1-1
4P
HL
G4
D2-N
GC
GA
II
D2-1
0L
GA
1L
9
D2-1
1B
GC
L4
D2-1
3M
CL
N
D2-1
5B
GC
L4
D2-1
5B
CL
N
D2-
PR
6193V
D2 -
PR
1940N
V
D3-H
87G
5
D3-1
5B
G1
D3-
UN
C 3
002V
D3-
UN
C 3
008 N
V
D4-H
24G
1
D4-1
0B
G3L
1
D4-1
0B
G3L
2
ZIK
V M
R 7
66
ZIK
V P
RV
AB
C59
YF
17D
JE
IM
OJE
V
Au
tolo
go
us v
iru
s
0
1 0 0
1 5 0
F L V 0 2 2 V 2 (D 1 -2 y e a rs p o s t in fe c t io n )
Me
an
Re
du
cti
on
%
50
75
90
D1-H
W2001G
4
D1-1
0B
G1L
14
D1-1
5L
G1
D1-1
0B
G2L
4
D1-1
0IN
DG
3L
4
D1-1
4P
HL
G4
D2-N
GC
GA
II
D2-1
0L
GA
1L
9
D2-1
1B
GC
L4
D2-1
3M
CL
N
D2-1
5B
GC
L4
D2-1
5B
CL
N
D2-
PR
6193V
D2 -
PR
1940N
V
D3-H
87G
5
D3-1
5B
G1
D3-
UN
C 3
002V
D3-
UN
C 3
008 N
V
D4-H
24G
1
D4-1
0B
G3L
1
D4-1
0B
G3L
2
ZIK
V M
R 7
66
ZIK
V P
RV
AB
C59
YF
17D
JE
IM
OJE
V
Au
tolo
go
us v
iru
s
0
1 0 0
1 5 0
F L V 0 2 2 V 1 (D 1 :1 y e a r 2 m o n th s )
V iru s ty p e
Me
an
Re
du
cti
on
%
50
75
90

359
D1-H
W2001G
4
D1-1
0B
G1L
14
D1-1
5L
G1
D1-1
0B
G2L
4
D1-1
0IN
DG
3L
4
D1-1
4P
HL
G4
D2-N
GC
GA
II
D2-1
0L
GA
1L
9
D2-1
1B
GC
L4
D2-1
3M
CL
N
D2-1
5B
GC
L4
D2-1
5B
CL
N
D2-
PR
6193V
D2 -
PR
1940N
V
D3-H
87G
5
D3-1
5B
G1
D3-
UN
C 3
002V
D3-
UN
C 3
008 N
V
D4-H
24G
1
D4-1
0B
G3L
1
D4-1
0B
G3L
2
ZIK
V M
R 7
66
ZIK
V P
RV
AB
C59
YF
17D
JE
IM
OJE
V
0
1 0 0
1 5 0
F L V 0 2 6 V 1 (D 1 :~ 1 y e a r -2 m o n th s )
V iru s ty p e
Me
an
Re
du
cti
on
%
50
75
90

360
Appendix 5.2.:- Neutralising antibody response over time in individuals with
DENV-2 infection
DENV-2 SERA PANEL
Number of individuals: 6
Number of individuals with multiple
sampling points:-6

361

362
D1-H
W2001G
4
D1-1
0B
G1L
14
D1-1
5L
G1
D1-1
0B
G2L
4
D1-1
0IN
DG
3L
4
D1-1
4P
HL
G4
D2-N
GC
GA
II
D2-1
0L
GA
1L
9
D2-1
1B
GC
L4
D2-1
3M
CL
N
D2-1
5B
GC
L4
D2-1
5B
CL
N
D2-
PR
6193V
D2 -
PR
1940N
V
D3-H
87G
5
D3-1
5B
G1
D3-
UN
C 3
002V
D3-
UN
C 3
008 N
V
D4-H
24G
1
D4-1
0B
G3L
1
D4-1
0B
G3L
2
ZIK
V M
R 7
66
ZIK
V P
RV
AB
C59
YF
17D
JE
IM
OJE
V
Au
tolo
go
us v
iru
s
0
1 0 0
1 5 0
F L V 0 0 4 V 1 (D 2 : 1 y e a r -4 m o n th s )
V iru s ty p e
Me
an
Re
du
cti
on
%
50
75
90
D1-H
W2001G
4
D1-1
0B
G1L
14
D1-1
5L
G1
D1-1
0B
G2L
4
D1-1
0IN
DG
3L
4
D1-1
4P
HL
G4
D2-N
GC
GA
II
D2-1
0L
GA
1L
9
D2-1
1B
GC
L4
D2-1
3M
CL
N
D2-1
5B
GC
L4
D2-1
5B
CL
N
D2-
PR
6193V
D2 -
PR
1940N
V
D3-H
87G
5
D3-1
5B
G1
D3-
UN
C 3
002V
D3-
UN
C 3
008 N
V
D4-H
24G
1
D4-1
0B
G3L
1
D4-1
0B
G3L
2
ZIK
V M
R 7
66
ZIK
V P
RV
AB
C59
YF
17D
JE
IM
OJE
V
Au
tolo
go
us v
iru
s
0
1 0 0
1 5 0
F L V 0 0 4 V 2 (D 2 :4 y e a r s -3 m o n th s )
V iru s ty p e
Me
an
Re
du
cti
on
%
50
75
90

363

364
D1-H
W2001G
4
D1-1
0B
G1L
14
D1-1
5L
G1
D1-1
0B
G2L
4
D1-1
0IN
DG
3L
4
D1-1
4P
HL
G4
D2-N
GC
GA
II
D2-1
0L
GA
1L
9
D2-1
1B
GC
L4
D2-1
3M
CL
N
D2-1
5B
GC
L4
D2-1
5B
CL
N
D2-
PR
6193V
D2 -
PR
1940N
V
D3-H
87G
5
D3-1
5B
G1
D3-
UN
C 3
002V
D3-
UN
C 3
008 N
V
D4-H
24G
1
D4-1
0B
G3L
1
D4-1
0B
G3L
2
ZIK
V M
R 7
66
ZIK
V P
RV
AB
C59
YF
17D
JE
IM
OJE
V
0
1 0 0
1 5 0
F L V 0 0 5 V 1 (D 2 : 1 y e a r -4 m o n th s )
V iru s ty p e
Me
an
Re
du
cti
on
%
50
75
90
D1-H
W2001G
4
D1-1
0B
G1L
14
D1-1
5L
G1
D1-1
0B
G2L
4
D1-1
0IN
DG
3L
4
D1-1
4P
HL
G4
D2-N
GC
GA
II
D2-1
0L
GA
1L
9
D2-1
1B
GC
L4
D2-1
3M
CL
N
D2-1
5B
GC
L4
D2-1
5B
CL
N
D2-
PR
6193V
D2 -
PR
1940N
V
D3-H
87G
5
D3-1
5B
G1
D3-
UN
C 3
002V
D3-
UN
C 3
008 N
V
D4-H
24G
1
D4-1
0B
G3L
1
D4-1
0B
G3L
2
ZIK
V M
R 7
66
ZIK
V P
RV
AB
C59
YF
17D
JE
IM
OJE
V
0
1 0 0
1 5 0
F L V 0 0 5 V 2 ( D 2 :4 y e a rs -3 m o n th s )
V iru s ty p e
Me
an
Re
du
cti
on
%
50
75
90

365

366
D1-H
W2001G
4
D1-1
0B
G1L
14
D1-1
5L
G1
D1-1
0B
G2L
4
D1-1
0IN
DG
3L
4
D1-1
4P
HL
G4
D2-N
GC
GA
II
D2-1
0L
GA
1L
9
D2-1
1B
GC
L4
D2-1
3M
CL
N
D2-1
5B
GC
L4
D2-1
5B
CL
N
D2-
PR
6193V
D2 -
PR
1940N
V
D3-H
87G
5
D3-1
5B
G1
D3-
UN
C 3
002V
D3-
UN
C 3
008 N
V
D4-H
24G
1
D4-1
0B
G3L
1
D4-1
0B
G3L
2
ZIK
V M
R 7
66
ZIK
V P
RV
AB
C59
YF
17D
JE
IM
OJE
V
0
1 0 0
1 5 0
F L V 0 0 9 V 1 (D 2 :~ 1 y e a r -5 m o n th s )
V iru s ty p e
Me
an
Re
du
cti
on
%
50
75
90
D1-H
W2001G
4
D1-1
0B
G1L
14
D1-1
5L
G1
D1-1
0B
G2L
4
D1-1
0IN
DG
3L
4
D1-1
4P
HL
G4
D2-N
GC
GA
II
D2-1
0L
GA
1L
9
D2-1
1B
GC
L4
D2-1
3M
CL
N
D2-1
5B
GC
L4
D2-1
5B
CL
N
D2-
PR
6193V
D2 -
PR
1940N
V
D3-H
87G
5
D3-1
5B
G1
D3-
UN
C 3
002V
D3-
UN
C 3
008 N
V
D4-H
24G
1
D4-1
0B
G3L
1
D4-1
0B
G3L
2
ZIK
V M
R 7
66
ZIK
V P
RV
AB
C59
YF
17D
JE
IM
OJE
V
0
1 0 0
1 5 0
F L V 0 0 9 V 2 (D 2 :~ 3 y e a rs )
V iru s ty p e
Me
an
Re
du
cti
on
%
50
75
90
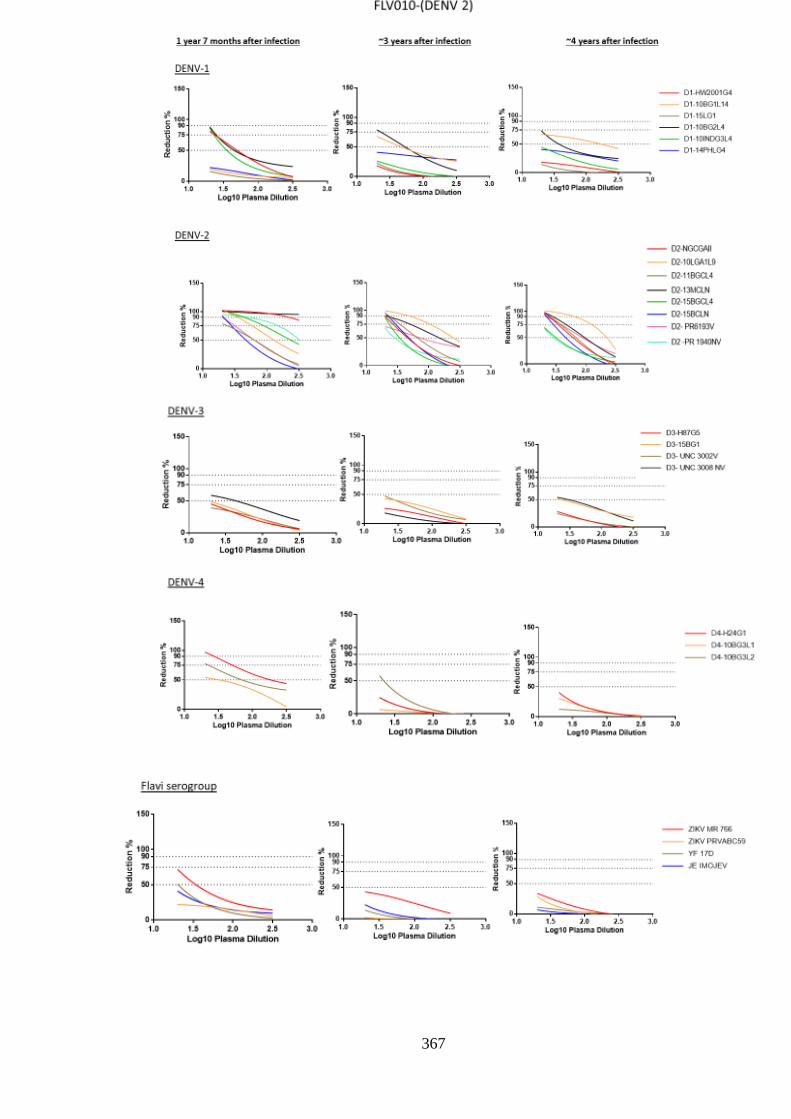
367

368
D1-H
W2001G
4
D1-1
0B
G1L
14
D1-1
5L
G1
D1-1
0B
G2L
4
D1-1
0IN
DG
3L
4
D1-1
4P
HL
G4
D2-N
GC
GA
II
D2-1
0L
GA
1L
9
D2-1
1B
GC
L4
D2-1
3M
CL
N
D2-1
5B
GC
L4
D2-1
5B
CL
N
D2-
PR
6193V
D2 -
PR
1940N
V
D3-H
87G
5
D3-1
5B
G1
D3-
UN
C 3
002V
D3-
UN
C 3
008 N
V
D4-H
24G
1
D4-1
0B
G3L
1
D4-1
0B
G3L
2
ZIK
V M
R 7
66
ZIK
V P
RV
AB
C59
YF
17D
JE
IM
OJE
V
0
1 0 0
1 5 0
F L V 0 1 0 V 1 (D 2 :1 y e a r -7 m o n th s )
V iru s ty p e
Me
an
Re
du
cti
on
%50
75
90
D1-H
W2001G
4
D1-1
0B
G1L
14
D1-1
5L
G1
D1-1
0B
G2L
4
D1-1
0IN
DG
3L
4
D1-1
4P
HL
G4
D2-N
GC
GA
II
D2-1
0L
GA
1L
9
D2-1
1B
GC
L4
D2-1
3M
CL
N
D2-1
5B
GC
L4
D2-1
5B
CL
N
D2-
PR
6193V
D2 -
PR
1940N
V
D3-H
87G
5
D3-1
5B
G1
D3-
UN
C 3
002V
D3-
UN
C 3
008 N
V
D4-H
24G
1
D4-1
0B
G3L
1
D4-1
0B
G3L
2
ZIK
V M
R 7
66
ZIK
V P
RV
AB
C59
YF
17D
JE
IM
OJE
V
0
1 0 0
1 5 0
F L V 0 1 0 V 2 (D 2 :~ 3 y e a rs )
V iru s ty p e
Me
an
Re
du
cti
on
%
50
75
90
D1-H
W2001G
4
D1-1
0B
G1L
14
D1-1
5L
G1
D1-1
0B
G2L
4
D1-1
0IN
DG
3L
4
D1-1
4P
HL
G4
D2-N
GC
GA
II
D2-1
0L
GA
1L
9
D2-1
1B
GC
L4
D2-1
3M
CL
N
D2-1
5B
GC
L4
D2-1
5B
CL
N
D2-
PR
6193V
D2 -
PR
1940N
V
D3-H
87G
5
D3-1
5B
G1
D3-
UN
C 3
002V
D3-
UN
C 3
008 N
V
D4-H
24G
1
D4-1
0B
G3L
1
D4-1
0B
G3L
2
ZIK
V M
R 7
66
ZIK
V P
RV
AB
C59
YF
17D
JE
IM
OJE
V
0
1 0 0
1 5 0
F L V 0 1 0 V 3 (D 2 :~ 4 y e a rs )
V iru s ty p e
Me
an
Re
du
cti
on
%
50
75
90

369

370
D1-H
W2001G
4
D1-1
0B
G1L
14
D1-1
5L
G1
D1-1
0B
G2L
4
D1-1
0IN
DG
3L
4
D1-1
4P
HL
G4
D2-N
GC
GA
II
D2-1
0L
GA
1L
9
D2-1
1B
GC
L4
D2-1
3M
CL
N
D2-1
5B
GC
L4
D2-1
5B
CL
N
D2-
PR
6193V
D2 -
PR
1940N
V
D3-H
87G
5
D3-1
5B
G1
D3-
UN
C 3
002V
D3-
UN
C 3
008 N
V
D4-H
24G
1
D4-1
0B
G3L
1
D4-1
0B
G3L
2
ZIK
V M
R 7
66
ZIK
V P
RV
AB
C59
YF
17D
JE
IM
OJE
V
0
1 0 0
1 5 0
F L V 0 1 2 V 1 (D 2 :5 d a y s )
V iru s ty p e
Me
an
Re
du
ctio
n %
50
75
90
D1-H
W2001G
4
D1-1
0B
G1L
14
D1-1
5L
G1
D1-1
0B
G2L
4
D1-1
0IN
DG
3L
4
D1-1
4P
HL
G4
D2-N
GC
GA
II
D2-1
0L
GA
1L
9
D2-1
1B
GC
L4
D2-1
3M
CL
N
D2-1
5B
GC
L4
D2-1
5B
CL
N
D2-
PR
6193V
D2 -
PR
1940N
V
D3-H
87G
5
D3-1
5B
G1
D3-
UN
C 3
002V
D3-
UN
C 3
008 N
V
D4-H
24G
1
D4-1
0B
G3L
1
D4-1
0B
G3L
2
ZIK
V M
R 7
66
ZIK
V P
RV
AB
C59
YF
17D
JE
IM
OJE
V
0
1 0 0
1 5 0
F L V 0 0 1 2 V 2 (D 2 :~ 2 y e a r s 9 m o n th s )
V iru s ty p e
Me
an
Re
du
cti
on
%
50
75
90

371

372
D1-H
W2001G
4
D1-1
0B
G1L
14
D1-1
5L
G1
D1-1
0B
G2L
4
D1-1
0IN
DG
3L
4
D1-1
4P
HL
G4
D2-N
GC
GA
II
D2-1
0L
GA
1L
9
D2-1
1B
GC
L4
D2-1
3M
CL
N
D2-1
5B
GC
L4
D2-1
5B
CL
N
D2-
PR
6193V
D2 -
PR
1940N
V
D3-H
87G
5
D3-1
5B
G1
D3-
UN
C 3
002V
D3-
UN
C 3
008 N
V
D4-H
24G
1
D4-1
0B
G3L
1
D4-1
0B
G3L
2
ZIK
V M
R 7
66
ZIK
V P
RV
AB
C59
YF
17D
JE
IM
OJE
V
Au
tolo
go
us v
iru
s
0
1 0 0
1 5 0
F L V 0 1 4 V 1 (D 2 :7 m o n th s )
V iru s ty p e
Me
an
Re
du
cti
on
%
50
75
90
D1-H
W2001G
4
D1-1
0B
G1L
14
D1-1
5L
G1
D1-1
0B
G2L
4
D1-1
0IN
DG
3L
4
D1-1
4P
HL
G4
D2-N
GC
GA
II
D2-1
0L
GA
1L
9
D2-1
1B
GC
L4
D2-1
3M
CL
N
D2-1
5B
GC
L4
D2-1
5B
CL
N
D2-
PR
6193V
D2 -
PR
1940N
V
D3-H
87G
5
D3-1
5B
G1
D3-
UN
C 3
002V
D3-
UN
C 3
008 N
V
D4-H
24G
1
D4-1
0B
G3L
1
D4-1
0B
G3L
2
ZIK
V M
R 7
66
ZIK
V P
RV
AB
C59
YF
17D
JE
IM
OJE
V
Au
tolo
go
us v
iru
s
0
1 0 0
1 5 0
F L V 0 1 4 V 2 (D 2 :~ 3 y e a rs )
V iru s ty p e
Me
an
Re
du
cti
on
%
50
75
90

373
Appendix 5.3.:- Neutralising antibody response over time in individuals with
DENV-3 infection
DENV-3 SERA PANEL
Number of individuals: 6
Number of individuals with multiple
sampling points:-1

374
D1-H
W2001G
4
D1-1
0B
G1L
14
D1-1
5L
G1
D1-1
0B
G2L
4
D1-1
0IN
DG
3L
4
D1-1
4P
HL
G4
D2-N
GC
GA
II
D2-1
0L
GA
1L
9
D2-1
1B
GC
L4
D2-1
3M
CL
N
D2-1
5B
GC
L4
D2-1
5B
CL
N
D2-
PR
6193V
D2 -
PR
1940N
V
D3-H
87G
5
D3-1
5B
G1
D3-
UN
C 3
002V
D3-
UN
C 3
008 N
V
D4-H
24G
1
D4-1
0B
G3L
1
D4-1
0B
G3L
2
ZIK
V M
R 7
66
ZIK
V P
RV
AB
C59
YF
17D
JE
IM
OJE
V
0
1 0 0
1 5 0
F L V 0 2 9 V 1 (D 3 :1 y e a r )
V iru s ty p e
Me
an
Re
du
cti
on
%
50
75
90

375
D1-H
W2001G
4
D1-1
0B
G1L
14
D1-1
5L
G1
D1-1
0B
G2L
4
D1-1
0IN
DG
3L
4
D1-1
4P
HL
G4
D2-N
GC
GA
II
D2-1
0L
GA
1L
9
D2-1
1B
GC
L4
D2-1
3M
CL
N
D2-1
5B
GC
L4
D2-1
5B
CL
N
D2-
PR
6193V
D2 -
PR
1940N
V
D3-H
87G
5
D3-1
5B
G1
D3-
UN
C 3
002V
D3-
UN
C 3
008 N
V
D4-H
24G
1
D4-1
0B
G3L
1
D4-1
0B
G3L
2
ZIK
V M
R 7
66
ZIK
V P
RV
AB
C59
YF
17D
JE
IM
OJE
V
0
1 0 0
1 5 0
F L V 0 3 0 V 1 (D 3 :~ 1 y e a r )
V iru s ty p e
Me
an
Re
du
cti
on
%
50
75
90

376
D1-H
W2001G
4
D1-1
0B
G1L
14
D1-1
5L
G1
D1-1
0B
G2L
4
D1-1
0IN
DG
3L
4
D1-1
4P
HL
G4
D2-N
GC
GA
II
D2-1
0L
GA
1L
9
D2-1
1B
GC
L4
D2-1
3M
CL
N
D2-1
5B
GC
L4
D2-1
5B
CL
N
D2-
PR
6193V
D2 -
PR
1940N
V
D3-H
87G
5
D3-1
5B
G1
D3-
UN
C 3
002V
D3-
UN
C 3
008 N
V
D4-H
24G
1
D4-1
0B
G3L
1
D4-1
0B
G3L
2
ZIK
V M
R 7
66
ZIK
V P
RV
AB
C59
YF
17D
JE
IM
OJE
V
0
1 0 0
1 5 0
F L V 0 3 3 V 1 (D 3 :~ 1 y e a r -2 m o n th s )
V iru s ty p e
Me
an
Re
du
cti
on
%
50
75
90
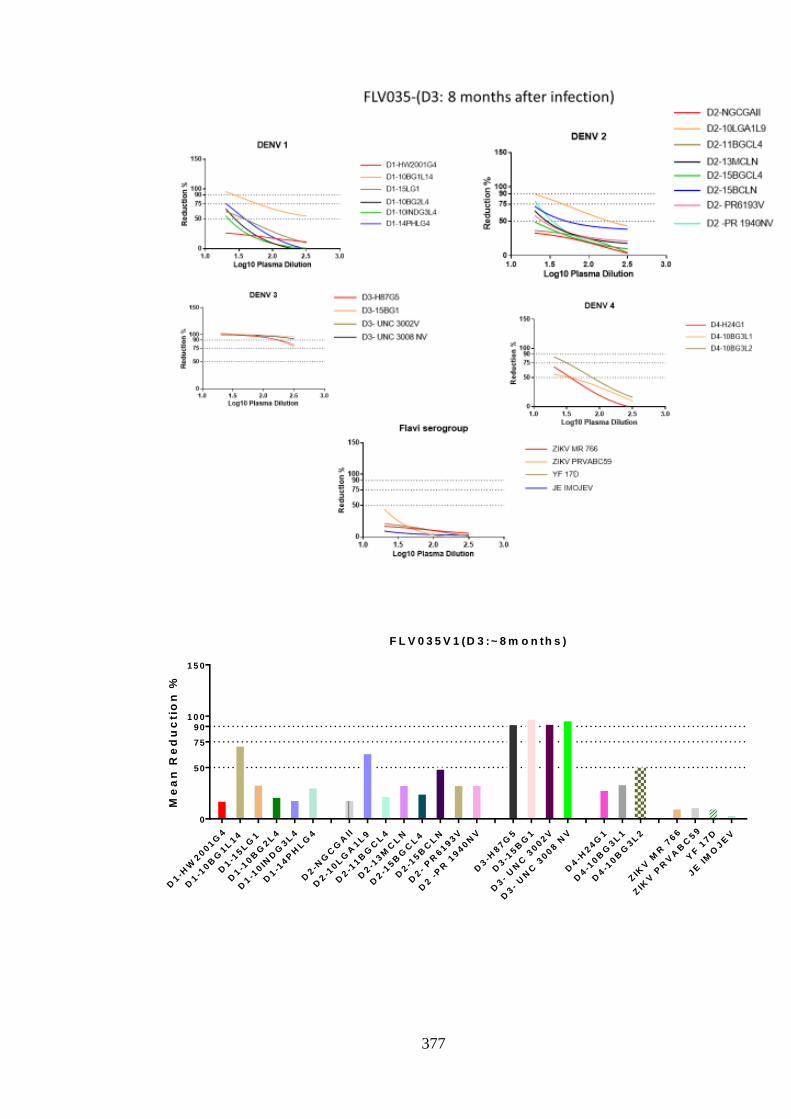
377
D1-H
W2001G
4
D1-1
0B
G1L
14
D1-1
5L
G1
D1-1
0B
G2L
4
D1-1
0IN
DG
3L
4
D1-1
4P
HL
G4
D2-N
GC
GA
II
D2-1
0L
GA
1L
9
D2-1
1B
GC
L4
D2-1
3M
CL
N
D2-1
5B
GC
L4
D2-1
5B
CL
N
D2-
PR
6193V
D2 -
PR
1940N
V
D3-H
87G
5
D3-1
5B
G1
D3-
UN
C 3
002V
D3-
UN
C 3
008 N
V
D4-H
24G
1
D4-1
0B
G3L
1
D4-1
0B
G3L
2
ZIK
V M
R 7
66
ZIK
V P
RV
AB
C59
YF
17D
JE
IM
OJE
V
0
1 0 0
1 5 0
F L V 0 3 5 V 1 (D 3 :~ 8 m o n th s )
V iru s ty p e
Me
an
Re
du
cti
on
%
50
75
90

378
D1-H
W2001G
4
D1-1
0B
G1L
14
D1-1
5L
G1
D1-1
0B
G2L
4
D1-1
0IN
DG
3L
4
D1-1
4P
HL
G4
D2-N
GC
GA
II
D2-1
0L
GA
1L
9
D2-1
1B
GC
L4
D2-1
3M
CL
N
D2-1
5B
GC
L4
D2-1
5B
CL
N
D2-
PR
6193V
D2 -
PR
1940N
V
D3-H
87G
5
D3-1
5B
G1
D3-
UN
C 3
002V
D3-
UN
C 3
008 N
V
D4-H
24G
1
D4-1
0B
G3L
1
D4-1
0B
G3L
2
ZIK
V M
R 7
66
ZIK
V P
RV
AB
C59
YF
17D
JE
IM
OJE
V
Au
tolo
go
us v
iru
s
0
1 0 0
1 5 0
F L V 0 3 7 V 1 (D 3 :~ 1 y e a r -2 m o n th s )
V iru s ty p e
Me
an
Re
du
cti
on
%
50
75
90
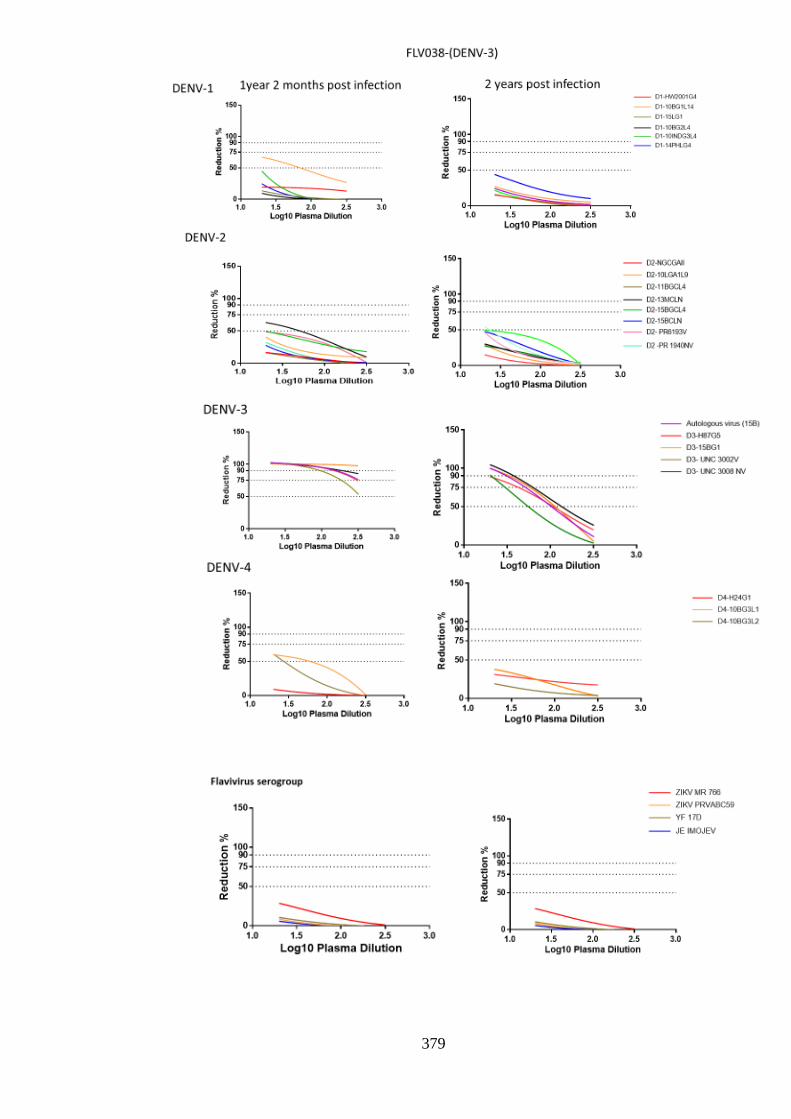
379

380
D1-H
W2001G
4
D1-1
0B
G1L
14
D1-1
5L
G1
D1-1
0B
G2L
4
D1-1
0IN
DG
3L
4
D1-1
4P
HL
G4
D2-N
GC
GA
II
D2-1
0L
GA
1L
9
D2-1
1B
GC
L4
D2-1
3M
CL
N
D2-1
5B
GC
L4
D2-1
5B
CL
N
D2-
PR
6193V
D2 -
PR
1940N
V
D3-H
87G
5
D3-1
5B
G1
D3-
UN
C 3
002V
D3-
UN
C 3
008 N
V
D4-H
24G
1
D4-1
0B
G3L
1
D4-1
0B
G3L
2
ZIK
V M
R 7
66
ZIK
V P
RV
AB
C59
YF
17D
JE
IM
OJE
V
Au
tolo
go
us v
iru
s
0
1 0 0
1 5 0
F L V 0 3 8 V 1 (D 3 :~ 1 y e a r -2 m o n th s )
V iru s ty p e
Me
an
Re
du
cti
on
%
50
75
90
D1-H
W2001G
4
D1-1
0B
G1L
14
D1-1
5L
G1
D1-1
0B
G2L
4
D1-1
0IN
DG
3L
4
D1-1
4P
HL
G4
D2-N
GC
GA
II
D2-1
0L
GA
1L
9
D2-1
1B
GC
L4
D2-1
3M
CL
N
D2-1
5B
GC
L4
D2-1
5B
CL
N
D2-
PR
6193V
D2 -
PR
1940N
V
D3-H
87G
5
D3-1
5B
G1
D3-
UN
C 3
002V
D3-
UN
C 3
008 N
V
D4-H
24G
1
D4-1
0B
G3L
1
D4-1
0B
G3L
2
Au
tolo
go
us v
iru
s (
15B
)
0
1 0 0
1 5 0
F L V 0 3 8 V 2 (D 3 :-2 y e a rs p o s t in fe c t io n )
Me
an
Re
du
cti
on
%
50
75
90

381
Appendix 5.4.:- Neutralising antibody response over time in individuals with
DENV-4 infection
DENV-4 SERA PANEL
Number of individuals: 6
Number of individuals with multiple
sampling points:-2
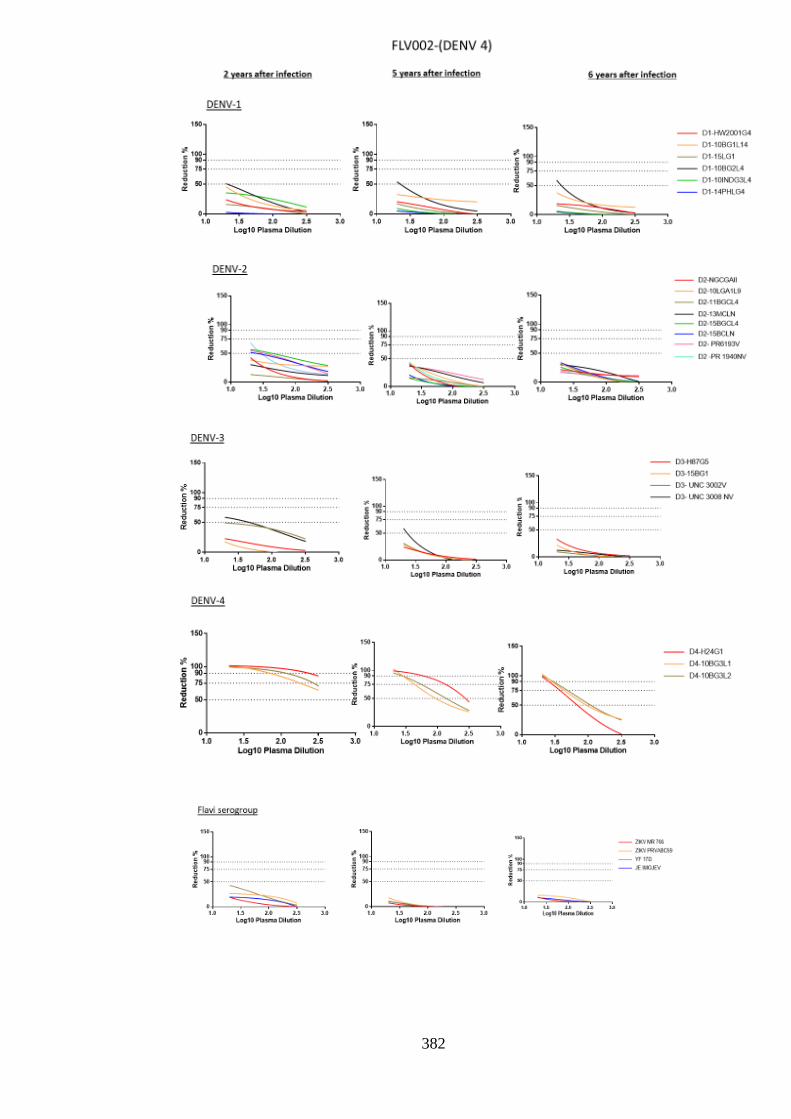
382

383
D1-H
W2001G
4
D1-1
0B
G1L
14
D1-1
5L
G1
D1-1
0B
G2L
4
D1-1
0IN
DG
3L
4
D1-1
4P
HL
G4
D2-N
GC
GA
II
D2-1
0L
GA
1L
9
D2-1
1B
GC
L4
D2-1
3M
CL
N
D2-1
5B
GC
L4
D2-1
5B
CL
N
D2-
PR
6193V
D2 -
PR
1940N
V
D3-H
87G
1
D3-H
87G
5
D3-1
5B
G1
D3-
UN
C 3
002V
D3-
UN
C 3
008 N
V
D4-H
24G
1
D4-1
0B
G3L
1
D4-1
0B
G3L
2
ZIK
V M
R 7
66
ZIK
V P
RV
AB
C59
YF
17D
JE
IM
OJE
V
0
1 0 0
1 5 0
F L V 0 0 2 V 1 (D 4 :2 y e a rs )
V iru s ty p e
Me
an
Re
du
cti
on
%
50
75
90
D1-H
W2001G
4
D1-1
0B
G1L
14
D1-1
5L
G1
D1-1
0B
G2L
4
D1-1
0IN
DG
3L
4
D1-1
4P
HL
G4
D2-N
GC
GA
II
D2-1
0L
GA
1L
9
D2-1
1B
GC
L4
D2-1
3M
CL
N
D2-1
5B
GC
L4
D2-1
5B
CL
N
D2-
PR
6193V
D2 -
PR
1940N
V
D3-H
87G
5
D3-1
5B
G1
D3-
UN
C 3
002V
D3-
UN
C 3
008 N
V
D4-H
24G
1
D4-1
0B
G3L
1
D4-1
0B
G3L
2
ZIK
V M
R 7
66
ZIK
V P
RV
AB
C59
YF
17D
JE
IM
OJE
V
0
1 0 0
1 5 0
F L V 0 0 2 V 2 (D 4 :~ 5 y e a rs )
V iru s ty p e
Me
an
Re
du
cti
on
%
50
75
90
D1-H
W2001G
4
D1-1
0B
G1L
14
D1-1
5L
G1
D1-1
0B
G2L
4
D1-1
0IN
DG
3L
4
D1-1
4P
HL
G4
D2-N
GC
GA
II
D2-1
0L
GA
1L
9
D2-1
1B
GC
L4
D2-1
3M
CL
N
D2-1
5B
GC
L4
D2-1
5B
CL
N
D2-
PR
6193V
D2 -
PR
1940N
V
D3-H
87G
5
D3-1
5B
G1
D3-
UN
C 3
002V
D3-
UN
C 3
008 N
V
D4-H
24G
1
D4-1
0B
G3L
1
D4-1
0B
G3L
2
ZIK
V M
R 7
66
ZIK
V P
RV
AB
C59
YF
17D
JE
IM
OJE
V
0
1 0 0
1 5 0
F L V 0 0 2 V 3 (D 4 :~ 6 y e a rs )
V iru s ty p e
Me
an
Re
du
cti
on
%
50
75
90

384

385
D1-H
W2001G
4
D1-1
0B
G1L
14
D1-1
5L
G1
D1-1
0B
G2L
4
D1-1
0IN
DG
3L
4
D1-1
4P
HL
G4
D2-N
GC
GA
II
D2-1
0L
GA
1L
9
D2-1
1B
GC
L4
D2-1
3M
CL
N
D2-1
5B
GC
L4
D2-1
5B
CL
N
D2-
PR
6193V
D2 -
PR
1940N
V
D3-H
87G
5
D3-1
5B
G1
D3-
UN
C 3
002V
D3-
UN
C 3
008 N
V
D4-H
24G
1
D4-1
0B
G3L
1
D4-1
0B
G3L
2
ZIK
V M
R 7
66
ZIK
V P
RV
AB
C59
YF
17D
JE
IM
OJE
V
0
1 0 0
1 5 0
F L V 0 4 9 V 1 (D 4 :~ 8 m o n th s p o s t in fe c t io n )M
ea
n R
ed
uc
tio
n %
50
75
90

386
D1-H
W2001G
4
D1-1
0B
G1L
14
D1-1
5L
G1
D1-1
0B
G2L
4
D1-1
0IN
DG
3L
4
D1-1
4P
HL
G4
D2-N
GC
GA
II
D2-1
0L
GA
1L
9
D2-1
1B
GC
L4
D2-1
3M
CL
N
D2-1
5B
GC
L4
D2-1
5B
CL
N
D2-
PR
6193V
D2 -
PR
1940N
V
D3-H
87G
5
D3-1
5B
G1
D3-
UN
C 3
002V
D3-
UN
C 3
008 N
V
D4-H
24G
1
D4-1
0B
G3L
1
D4-1
0B
G3L
2
ZIK
V M
R 7
66
ZIK
V P
RV
AB
C59
YF
17D
JE
IM
OJE
V
0
1 0 0
1 5 0
F L V 0 6 8 V 1 (D 4 :~ 8 m o n th s )
V iru s ty p e
Me
an
Re
du
cti
on
%
50
75
90

387
D1-H
W2001G
4
D1-1
0B
G1L
14
D1-1
5L
G1
D1-1
0B
G2L
4
D1-1
0IN
DG
3L
4
D1-1
4P
HL
G4
D2-N
GC
GA
II
D2-1
0L
GA
1L
9
D2-1
1B
GC
L4
D2-1
3M
CL
N
D2-1
5B
GC
L4
D2-1
5B
CL
N
D2-
PR
6193V
D2 -
PR
1940N
V
D3-H
87G
5
D3-1
5B
G1
D3-
UN
C 3
002V
D3-
UN
C 3
008 N
V
D4-H
24G
1
D4-1
0B
G3L
1
D4-1
0B
G3L
2
ZIK
V M
R 7
66
ZIK
V P
RV
AB
C59
YF
17D
JE
IM
OJE
V
0
1 0 0
1 5 0
F L V 0 8 0 V 1 (D 4 :~ 1 y e a r -4 m o n th s )
V iru s ty p e
Me
an
Re
du
cti
on
%
50
75
90
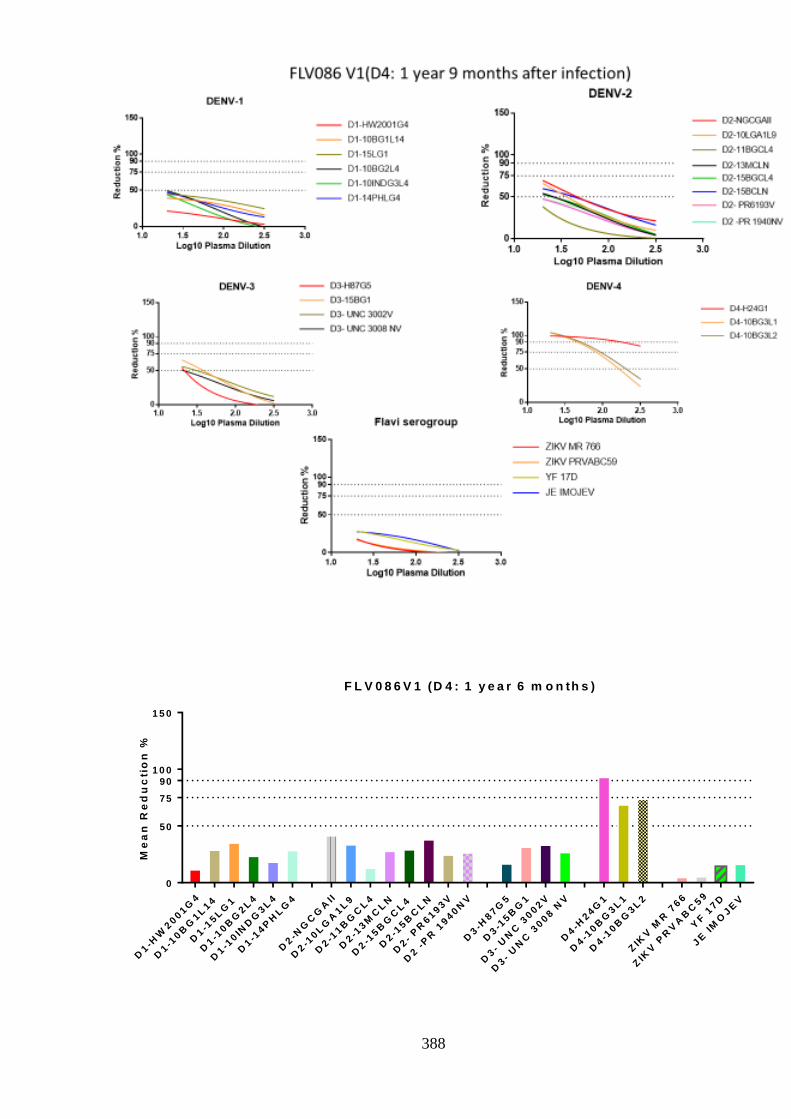
388
D1-H
W2001G
4
D1-1
0B
G1L
14
D1-1
5L
G1
D1-1
0B
G2L
4
D1-1
0IN
DG
3L
4
D1-1
4P
HL
G4
D2-N
GC
GA
II
D2-1
0L
GA
1L
9
D2-1
1B
GC
L4
D2-1
3M
CL
N
D2-1
5B
GC
L4
D2-1
5B
CL
N
D2-
PR
6193V
D2 -
PR
1940N
V
D3-H
87G
5
D3-1
5B
G1
D3-
UN
C 3
002V
D3-
UN
C 3
008 N
V
D4-H
24G
1
D4-1
0B
G3L
1
D4-1
0B
G3L
2
ZIK
V M
R 7
66
ZIK
V P
RV
AB
C59
YF
17D
JE
IM
OJE
V
0
1 0 0
1 5 0
F L V 0 8 6 V 1 (D 4 : 1 y e a r 6 m o n th s )
Me
an
Re
du
cti
on
%
50
75
90
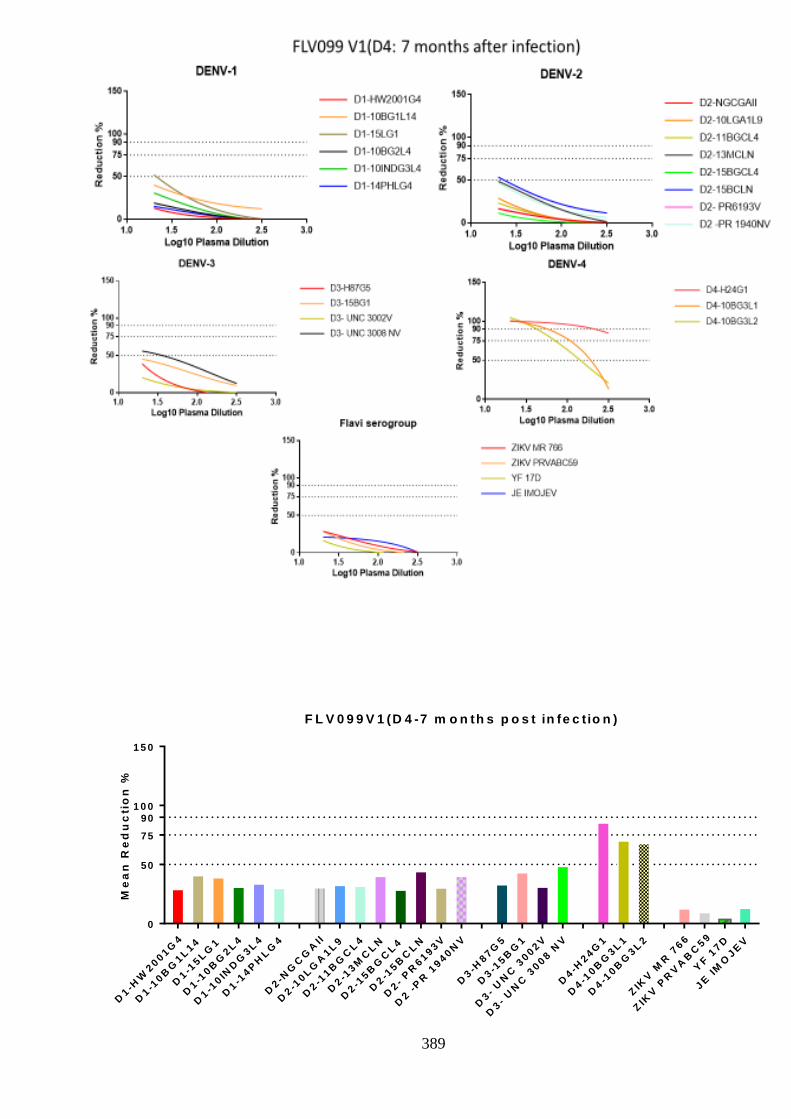
389
D1-H
W2001G
4
D1-1
0B
G1L
14
D1-1
5L
G1
D1-1
0B
G2L
4
D1-1
0IN
DG
3L
4
D1-1
4P
HL
G4
D2-N
GC
GA
II
D2-1
0L
GA
1L
9
D2-1
1B
GC
L4
D2-1
3M
CL
N
D2-1
5B
GC
L4
D2-1
5B
CL
N
D2-
PR
6193V
D2 -
PR
1940N
V
D3-H
87G
5
D3-1
5B
G1
D3-
UN
C 3
002V
D3-
UN
C 3
008 N
V
D4-H
24G
1
D4-1
0B
G3L
1
D4-1
0B
G3L
2
ZIK
V M
R 7
66
ZIK
V P
RV
AB
C59
YF
17D
JE
IM
OJE
V
0
1 0 0
1 5 0
F L V 0 9 9 V 1 (D 4 -7 m o n th s p o s t in fe c tio n )
Me
an
Re
du
cti
on
%
50
75
90

390
Appendix 6.1.:- Neutralising antibody response over time in individuals with ZIKV
infection
Each graph is the analysis of non-linear regression of neutralisation at log 10 plasma
dilution. Number of virus tested and starting point of dilution varies according to
availability of sample. Each graph title represents patient code and time point of sample
collection. Graph title with no time point represents sample taken at point of diagnosis.
50,75,90% reduction is shown in each graph. Past flavivirus history of individual is
stated for each graph. Graphs are listed in no particular order
0 .5 1 .0 1 .5 2 .0 2 .5
0
1 0 0
1 5 0
F L V 0 6 4 (6 0 5 d a y s p o s t in fe c t io n )
P la s m a d ilu tio n (L o g 1 0 )
Re
du
cti
on
%
50
75
90
Z IK V M R 7 6 6
Z IK V P R V A B C 5 9
Y F 1 7 D
J E IM O J E V
D E N V -1 (H W 2 0 0 1 )
D E N V -2 (N G C )
D E N V -3 (H 8 7 )
D E N V -4 (H 2 4 1 )
1 .0 1 .5 2 .0 2 .5
0
1 0 0
1 5 0
F L V -0 9 3 - (3 3 0 d a y s p o s t in fe c t io n )
L o g 1 0 P la s m a D ilu tio n
Re
du
cti
on
%
50
75
90
Z IK V M R 7 6 6
Z IK V P R V A B C 5 9
Y F 1 7 D
J E IM O J E V
D E N V -1 (H W 2 0 0 1 )
D E N V -2 (N G C )
D E N V -3 (H 8 7 )
D E N V -4 (H 2 4 1 )
FLV-064:- Confirmed ZIKV, no previous flavivirus exposure
FLV-093: - Probable ZIKV, with known YF17D vaccination but no
known previous DENV exposure. Frequent traveller to endemic areas.

391
1 .5 2 .0 2 .5 3 .0 3 .5 4 .0 4 .5
0
1 0 0
1 5 0
F L V 0 8 7 (3 9 d a y s p o s t in fe c tio n )
L o g 1 0 P la s m a D ilu tio n
Re
du
cti
on
%
50
75
90
Z IK V M R 7 6 6
Z IK V P R V A B C 5 9
Y F 1 7 D
J E IM O J E V
D E N V -1 (H W 2 0 0 1 )
D E N V -2 (N G C )
D E N V -3 (H 8 7 )
D E N V -4 (H 2 4 1 )
1 .5 2 .0 2 .5 3 .0 3 .5 4 .0 4 .5
0
1 0 0
1 5 0
F L V 0 8 7
L o g 1 0 P la s m a D ilu tio n
Re
du
cti
on
%
50
75
90
Z IK V M R 7 6 6
Y F 1 7 D
J E IM O J E V
D E N V -4 (H 2 4 1 )
1 .5 2 .0 2 .5 3 .0 3 .5 4 .0 4 .5
0
1 0 0
1 5 0
F L V 0 8 7 (1 2 d a y s p o s t in fe c tio n )
L o g 1 0 P la s m a D ilu tio n
Re
du
cti
on
%
50
75
90
Z IK V M R 7 6 6
Z IK V P R V A B C 5 9
Y F 1 7 D
J E IM O J E V
D E N V -4 (H 2 4 1 )
0 .5 1 .0 1 .5 2 .0 2 .5
0
1 0 0
1 5 0
F L V 0 8 7 (4 2 5 d a y s p o s t in fe c tio n )
P la s m a d ilu tio n (L o g 1 0 )
Re
du
cti
on
%
50
75
90
Z IK V M R 7 6 6
Z IK V P R V A B C 5 9
Y F 1 7 D
J E IM O J E V
D E N V -1 (H W 2 0 0 1 )
D E N V -2 (N G C )
D E N V -3 (H 8 7 )
D E N V -4 (H 2 4 1 )
FLV-087: - Confirmed ZIKV, with known DENV exposure.
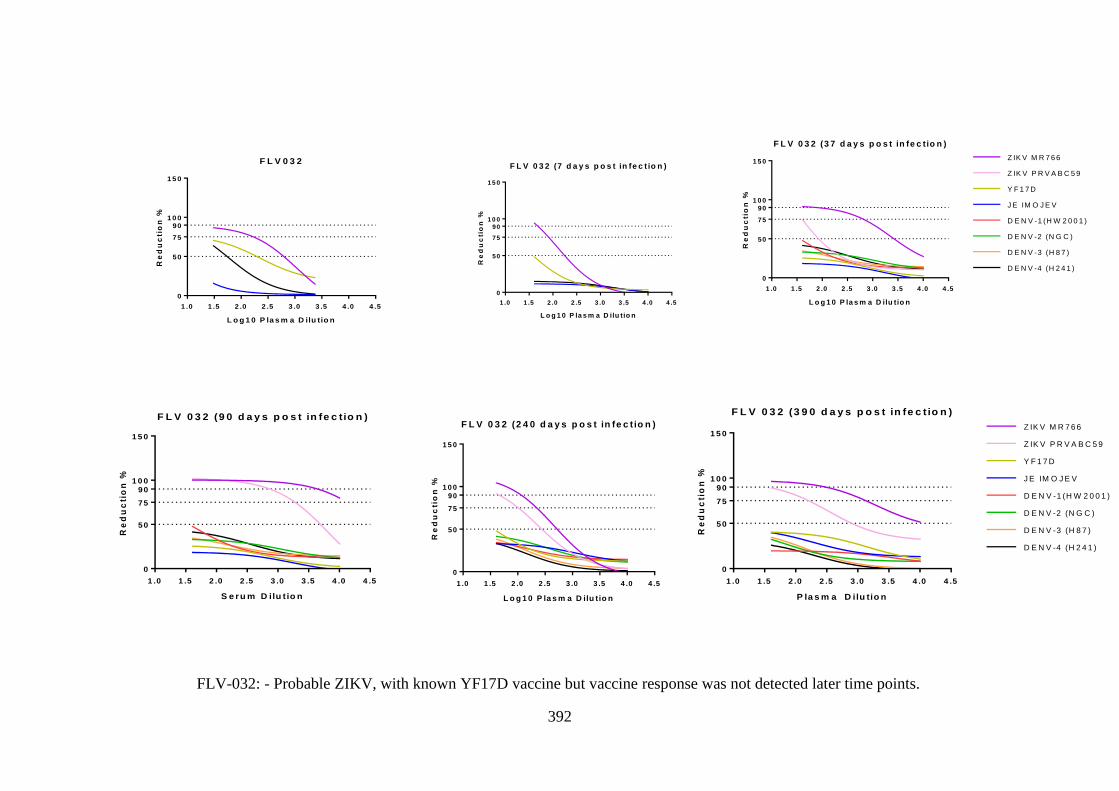
392
1 .0 1 .5 2 .0 2 .5 3 .0 3 .5 4 .0 4 .5
0
1 0 0
1 5 0
F L V 0 3 2 (7 d a y s p o s t in fe c t io n )
L o g 1 0 P la s m a D ilu tio n
Re
du
cti
on
%
50
75
90
Z IK V M R 7 6 6
Y F 1 7 D
D E N V -4 (H 2 4 1 )
J E IM O J E V
1 .0 1 .5 2 .0 2 .5 3 .0 3 .5 4 .0 4 .5
0
1 0 0
1 5 0
F L V 0 3 2
L o g 1 0 P la s m a D ilu tio n
Re
du
cti
on
%
50
75
90
Z IK V M R 7 6 6
Y F 1 7 D
D E N V -4 (H 2 4 1 )
J E IM O J E V
1 .0 1 .5 2 .0 2 .5 3 .0 3 .5 4 .0 4 .5
0
1 0 0
1 5 0
F L V 0 3 2 (3 9 0 d a y s p o s t in fe c t io n )
P la s m a D ilu tio n
Re
du
cti
on
%
50
75
90
Z IK V M R 7 6 6
Z IK V P R V A B C 5 9
Y F 1 7 D
J E IM O J E V
D E N V -1 (H W 2 0 0 1 )
D E N V -2 (N G C )
D E N V -3 (H 8 7 )
D E N V -4 (H 2 4 1 )
1 .0 1 .5 2 .0 2 .5 3 .0 3 .5 4 .0 4 .5
0
1 0 0
1 5 0
F L V 0 3 2 (3 7 d a y s p o s t in fe c tio n )
L o g 1 0 P la s m a D ilu tio n
Re
du
cti
on
%
50
75
90
Z IK V M R 7 6 6
Z IK V P R V A B C 5 9
Y F 1 7 D
J E IM O J E V
D E N V -1 (H W 2 0 0 1 )
D E N V -2 (N G C )
D E N V -3 (H 8 7 )
D E N V -4 (H 2 4 1 )
1 .0 1 .5 2 .0 2 .5 3 .0 3 .5 4 .0 4 .5
0
1 0 0
1 5 0
F L V 0 3 2 (9 0 d a y s p o s t in fe c tio n )
S e ru m D ilu t io n
Re
du
cti
on
%
50
75
90
Z IK V M R 7 6 6
Z IK V P R V A B C 5 9
Y F 1 7 D
J E IM O J E V
D E N V -1 (H W 2 0 0 1 )
D E N V -2 (N G C )
D E N V -3 (H 8 7 )
D E N V -4 (H 2 4 1 ) 1 .0 1 .5 2 .0 2 .5 3 .0 3 .5 4 .0 4 .5
0
1 0 0
1 5 0
F L V 0 3 2 (2 4 0 d a y s p o s t in fe c t io n )
L o g 1 0 P la s m a D ilu tio n
Re
du
cti
on
%
50
75
90
Z IK V M R 7 6 6
Z IK V P R V A B C 5 9
Y F 1 7 D
J E IM O J E V
D E N V -1 (H W 2 0 0 1 )
D E N V -2 (N G C )
D E N V -3 (H 8 7 )
D E N V -4 (H 2 4 1 )
FLV-032: - Probable ZIKV, with known YF17D vaccine but vaccine response was not detected later time points. FLV-032: - Probable ZIKV, with known YF17D vaccine but vaccine response was not detected later time points.
1 .0 1 .5 2 .0 2 .5 3 .0 3 .5 4 .0 4 .5
0
1 0 0
1 5 0
F L V 0 3 2 (2 4 0 d a y s p o s t in fe c t io n )
L o g 1 0 P la s m a D ilu tio n
Re
du
cti
on
%
50
75
90
Z IK V M R 7 6 6
Z IK V P R V A B C 5 9
Y F 1 7 D
J E IM O J E V
D E N V -1 (H W 2 0 0 1 )
D E N V -2 (N G C )
D E N V -3 (H 8 7 )
D E N V -4 (H 2 4 1 )

393
1 .0 1 .5 2 .0 2 .5 3 .0 3 .5 4 .0 4 .5
0
1 0 0
1 5 0
F L V 0 8 8 (2 3 d a y s p o s t in fe c tio n )
L o g 1 0 P la s m a D ilu tio n
Re
du
cti
on
%
Z IK V P R V A B C 5 9
D E N V -1 (H W 2 0 0 1 )
D E N V -2 (N G C )
D E N V -3 (H 8 7 )
D E N V -4 (H 2 4 1 )
J E IM O J E V
Y F 1 7 D
Z IK V M R 7 6 6
50
75
90
1 .0 1 .5 2 .0 2 .5 3 .0 3 .5 4 .0 4 .5
0
1 0 0
1 5 0
F L V 0 8 8
L o g 1 0 P la s m a D ilu tio n
Re
du
cti
on
%
50
75
90
Z IK V M R 7 6 6
Z ik a P R V A B C 5 9
J E IM O J E V
Y F 1 7 D
D E N V -1 (H W 2 0 0 1 )
D E N V -2 (N G C )
D E N V -3 (H 8 7 )
D E N V -4 (H 2 4 1 )
0 .5 1 .0 1 .5 2 .0 2 .5 3 .0 3 .5 4 .0
0
1 0 0
1 5 0
F L V 0 8 8 -(4 2 5 d a y s p o s t in fe c tio n )
L o g 1 0 P la s m a D ilu tio n
Re
du
cti
on
%
Z IK V M R 7 6 6
Z IK V P R V A B C 5 9
Y F 1 7 D
J E IM O J E V
D E N V -1 (H W 2 0 0 1 )
D E N V -2 (N G C )
D E N V -3 (H 8 7 )
D E N V -4 (H 2 4 1 )
50
75
90
1 .0 1 .5 2 .0 2 .5 3 .0 3 .5 4 .0 4 .5
0
1 0 0
1 5 0
F L V 0 8 9
L o g 1 0 P la s m a D ilu tio n
Re
du
cti
on
%
50
75
90
Z IK V M R 7 6 6
Z IK V P R V A B C 5 9
Y F 1 7 D
J E IM O J E V
D E N V -1 (H W 2 0 0 1 )
D E N V -2 (N G C )
D E N V -3 (H 8 7 )
D E N V -4 (H 2 4 1 )0 .5 1 .0 1 .5 2 .0 2 .5
0
1 0 0
1 5 0
F L V 0 8 9 - (4 2 5 d a y s p o s t in fe c t io n )
L o g 1 0 P la s m a D ilu tio n
Re
du
ctio
n %
50
75
90
Z IK V M R 7 6 6
Z IK V P R V A B C 5 9
Y F 1 7 D
J E IM O J E V
D E N V -1 (H W 2 0 0 1 )
D E N V -2 (N G C )
D E N V -3 (H 8 7 )
D E N V -4 (H 2 4 1 )
1 .0 1 .5 2 .0 2 .5 3 .0 3 .5 4 .0 4 .5
0
1 0 0
1 5 0
F L V 0 8 9 (2 3 d a y s p o s t in fe c tio n )
L o g 1 0 P la s m a D ilu tio n
Re
du
cti
on
%
D E N V -1 (H W 2 0 0 1 )
D E N V -2 (N G C )
D E N V -3 (H 8 7 )
Z IK V P R V A B C 5 9
50
75
90
Z IK V M R 7 6 6
Y F 1 7 D
J E IM O J E V
D E N V -4 (H 2 4 1 )
FLV-088: - Probable ZIKV, with unknown DENV frequent traveller to El Salvador.
FLV-089: - Probable ZIKV, with unknown DENV, frequent traveller to El Salvador.

394
1 .0 1 .5 2 .0 2 .5 3 .0 3 .5 4 .0 4 .5
0
1 0 0
1 5 0
F L V 0 9 1 (3 d a y s p o s t in fe c t io n )
L o g 1 0 P la s m a D ilu tio n
Re
du
cti
on
%
50
75
90
Z IK V M R 7 6 6
Z IK V P R V A B C 5 9
Y F 1 7 D
J E IM O J E V
D E N V -1 (H W 2 0 0 1 )
D E N V -2 (N G C )
D E N V -3 (H 8 7 )
D E N V -4 (H 2 4 1 )
1 .0 1 .5 2 .0 2 .5
0
1 0 0
1 5 0
F L V -0 9 1 (2 4 0 d a y s )
L o g 1 0 P la s m a D ilu tio n
Re
du
cti
on
%
50
75
90
Z IK V M R 7 6 6
Z IK V P R V A B C 5 9
Y F 1 7 D
J E IM O J E V
D E N V -1 (H W 2 0 0 1 )
D E N V -2 (N G C )
D E N V -3 (H 8 7 )
D E N V -4 (H 2 4 1 )
1 .0 1 .5 2 .0 2 .5 3 .0 3 .5 4 .0 4 .5
0
1 0 0
1 5 0
F L V 0 9 1
L o g 1 0 P la s m a D ilu tio n
Re
du
cti
on
%
50
75
90
Z IK V M R 7 6 6
Z IK V P R V A B C 5 9
Y F 1 7 D
J E IM O J E V
D E N V -1 (H W 2 0 0 1 )
D E N V -2 (N G C )
D E N V -3 (H 8 7 )
D E N V -4 (H 2 4 1 )
0 .5 1 .0 1 .5 2 .0 2 .5
0
1 0 0
1 5 0
F L V 0 9 0 - (4 2 5 d a y s p o s t in fe c t io n )
L o g 1 0 P la s m a D ilu tio n
Re
du
cti
on
%
50
75
90
Z IK V M R 7 6 6
Z IK V P R V A B C 5 9
Y F 1 7 D
J E IM O J E V
D E N V -1 (H W 2 0 0 1 )
D E N V -2 (N G C )
D E N V -3 (H 8 7 )
D E N V -4 (H 2 4 1 )
FLV-090: - Probable ZIKV only infection
FLV-091: - Probable ZIKV infection with known YF17D vaccination but no known DENV exposure. Frequent traveller to
endemic areas.
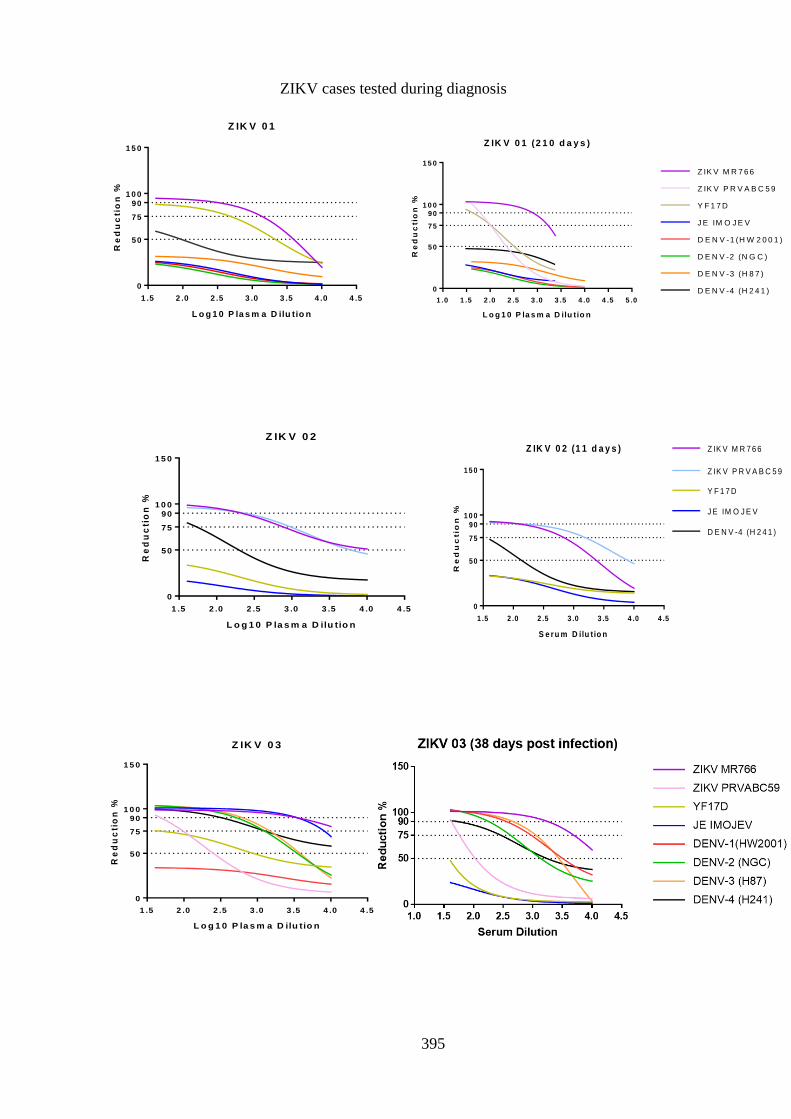
395
1 .5 2 .0 2 .5 3 .0 3 .5 4 .0 4 .5
0
1 0 0
1 5 0
Z IK V 0 3
L o g 1 0 P la s m a D ilu tio n
Re
du
cti
on
%
50
75
90
Z IK V M R 7 6 6
Z IK V P R V A B C 5 9
Y F 1 7 D
J E IM O J E V
D E N V -1 (H W 2 0 0 1 )
D E N V -2 (N G C )
D E N V -3 (H 8 7 )
D E N V -4 (H 2 4 1 )
1 .0 1 .5 2 .0 2 .5 3 .0 3 .5 4 .0 4 .5 5 .0
0
1 0 0
1 5 0
Z IK V 0 1 (2 1 0 d a y s )
L o g 1 0 P la s m a D ilu tio n
Re
du
cti
on
%
Z IK V P R V A B C 5 9
D E N V -1 (H W 2 0 0 1 )
D E N V -2 (N G C )
D E N V -3 (H 8 7 )
J E IM O J E V
D E N V -4 (H 2 4 1 )
Y F 1 7 D
Z IK V M R 7 6 6
50
75
90
1 .5 2 .0 2 .5 3 .0 3 .5 4 .0 4 .5
0
1 0 0
1 5 0
Z IK V 0 1
L o g 1 0 P la s m a D ilu tio n
Re
du
cti
on
%
50
75
90
Z IK V M R 7 6 6
Y F 1 7 D
J E IM O J E V
D E N V -1 (H W 2 0 0 1 )
D E N V -2 (N G C )
D E N V -3 (H 8 7 )
D E N V -4 (H 2 4 1 )
1 .5 2 .0 2 .5 3 .0 3 .5 4 .0 4 .5
0
1 0 0
1 5 0
Z IK V 0 2
L o g 1 0 P la s m a D ilu tio n
Re
du
cti
on
%
50
75
90
Z IK V M R 7 6 6
Z IK V P R V A B C 5 9
Y F 1 7 D
J E IM O J E V
D E N V -4 (H 2 4 1 )
1 .5 2 .0 2 .5 3 .0 3 .5 4 .0 4 .5
0
1 0 0
1 5 0
Z IK V 0 2 (1 1 d a y s )
S e ru m D ilu t io n
Re
du
ctio
n %
50
75
90
Z IK V M R 7 6 6
Z IK V P R V A B C 5 9
Y F 1 7 D
J E IM O J E V
D E N V -4 (H 2 4 1 )
ZIKV cases tested during diagnosis

396
1 .5 2 .0 2 .5 3 .0 3 .5 4 .0 4 .5
0
1 0 0
1 5 0
Z IK V 0 4
S e ru m D ilu t io n
Re
du
cti
on
%
50
75
90
Z IK V M R 7 6 6
Z IK V P R V A B C 5 9
Y F 1 7 D
J E IM O J E V
D E N V -1 (H W 2 0 0 1 )
D E N V -2 (N G C )
D E N V -3 (H 8 7 )
D E N V -4 (H 2 4 1 )
1 .0 1 .5 2 .0 2 .5 3 .0 3 .5 4 .0 4 .5
0
1 0 0
1 5 0
Z IK V 0 6
L o g 1 0 P la s m a D ilu tio n
Re
du
cti
on
%
50
75
90
Z IK V M R 7 6 6
Y F 1 7 D
J E IM O J E V
D E N V -4 (H 2 4 1 )
1 .0 1 .5 2 .0 2 .5 3 .0 3 .5 4 .0 4 .5
0
1 0 0
1 5 0
Z IK V 0 7
L o g 1 0 P la s m a D ilu tio n
Re
du
cti
on
%
50
75
90
Z IK V M R 7 6 6
Y F 1 7 D
J E IM O J E V
D E N V -4 (H 2 4 1 )
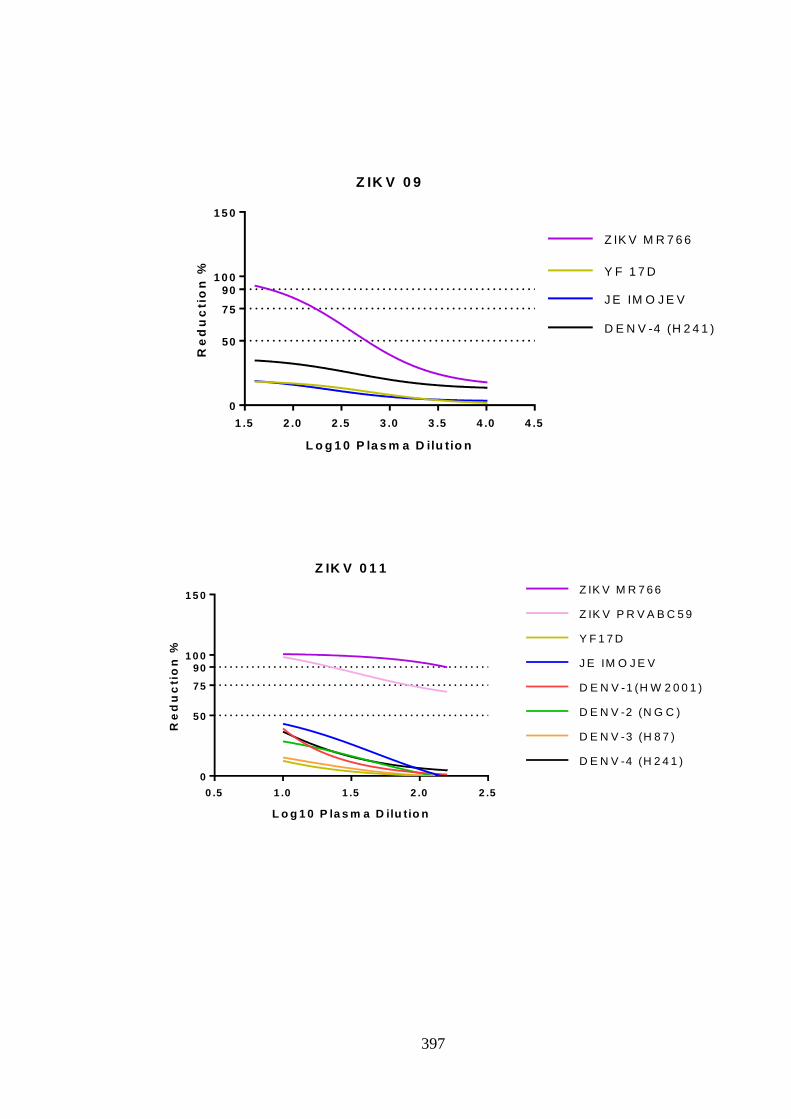
397
0 .5 1 .0 1 .5 2 .0 2 .5
0
1 0 0
1 5 0
Z IK V 0 1 1
L o g 1 0 P la s m a D ilu tio n
Re
du
cti
on
%
50
75
90
Z IK V M R 7 6 6
Z IK V P R V A B C 5 9
Y F 1 7 D
J E IM O J E V
D E N V -1 (H W 2 0 0 1 )
D E N V -2 (N G C )
D E N V -3 (H 8 7 )
D E N V -4 (H 2 4 1 )
1 .5 2 .0 2 .5 3 .0 3 .5 4 .0 4 .5
0
1 0 0
1 5 0
Z IK V 0 9
L o g 1 0 P la s m a D ilu tio n
Re
du
cti
on
%
50
75
90
Z IK V M R 7 6 6
Y F 1 7 D
J E IM O J E V
D E N V -4 (H 2 4 1 )
Copyright © 2022 FDOKUMEN