
Syngeneic Marek's Disease Virus (MDV)Specific Cell-Mediated Immune Responses against Immediate...
13
VIROLOGY 222, 87–99 (1996) ARTICLE NO. 0400 Syngeneic Marek’s Disease Virus (MDV)-Specific Cell-Mediated Immune Responses against Immediate Early, Late, and Unique MDV Proteins ABDUL R. OMAR and KAREL A. SCHAT 1 Unit of Avian Medicine, Department of Microbiology and Immunology, College of Veterinary Medicine, Cornell University, Ithaca, New York 14853 Received March 25, 1996; accepted June 12, 1996 Marek’s disease (MD) infection has been controlled effectively by vaccination using nononcogenic and/or attenuated oncogenic Marek’s disease virus (MDV) vaccines. Thus far, there is little knowledge on the role of cell-mediated immune (CMI) responses during MDV infection or vaccination. To elucidate the importance of MDV proteins in CMI responses, the pp38, Meq, ICP4, or ICP22 genes of an oncogenic strain, GA and the gB, ORF A, A41, or L1 genes of a highly oncogenic strain, RB1B were stably transfected into reticuloendotheliosis virus (REV)-transformed lymphoblastoid cells, CU-91 (MHC: B 19 B 19 ) and CU-205 (MHC: B 21 B 21 ). Cell lines positive for MDV gene transcription and/or protein expression were used in a standard 4-hr chromium release assay. Effector cells for this assay were obtained from splenocytes of chickens infected with the oncogenic strain, JM-16/13 or the nononcogenic vaccine strain, SB-1/12. Cell lines expressing MDV pp38, Meq, or gB were lysed by syngeneic but not allogeneic MDV-sensitized splenocytes obtained from chickens of B 19 B 19 and B 21 B 21 haplotypes. However, syngeneic CMI responses against ICP4 were detected only in B 21 B 21 chickens. CMI responses were not detected against B 19 B 19 and B 21 B 21 cell lines expressing A41, L1, ORF A, or ICP22. This report suggests that syngeneic CMI responses against pp38, Meq, ICP4, and gB of GA and RB1B strains, respectively, can be induced in chickens inoculated with JM16/13 or SB-1/12. The difference in CMI response to ICP4 in genetically susceptible (B 19 B 19 ) and genetically resistant (B 21 B 21 ) chickens may be an important factor in genetic resistance. q 1996 Academic Press, Inc. INTRODUCTION BamHI-H family [open reading frame (ORF) A, B, C, D, E, and F] have also been cloned and sequenced (Chen et Marek’s disease (MD), an economically important, T al., 1992; Hong and Coussens, 1994; Jones et al., 1992; cell lymphoproliferative disease in chickens is caused by Ohashi et al., 1994a, 1994b; Peng et al., 1992). Depending Marek’s disease virus (MDV), an oncogenic herpesvirus on the host major histocompatibility (MHC) genes, the (Churchill and Biggs, 1967). MDV can be divided into majority of cytotoxic T lymphocytes (CTLs) during alpha- three serotypes; MDV serotype 1 (MDV-1) consists of all herpesvirus infections in mice are directed against IE oncogenic and attenuated strains, MDV serotype 2 and/or L genes products (Banks et al., 1991; Hanke et (MDV-2) consists of naturally occurring nononcogenic al., 1991; Martin et al., 1990; Witmer et al., 1990). For strains, and MDV serotype 3 (MDV-3) consists of non- example in HSV-1 infection, ICP27 and ICP4 were target pathogenic strains from turkeys and is known as herpes- antigens for CTLs in mice of H-2 d and H-2 k haplotypes, virus of turkey (HVT) (reviewed by Schat, 1987). respectively (Banks et al., 1991; Martin et al., 1990). Con- MDV and HVT were initially classified as gammaher- versely, gB-specific CTLs were only detected in mice of pesviruses on the basis of their biological properties. H-2 b and H-2 d haplotypes (Hanke et al., 1991; Witmer et However, the overall genome structures of MDV and HVT al., 1990). are closely related to alphaherpesviruses (Buckmaster Although significant progress has been made in et al., 1988). MDV genes with homology to herpes sim- characterizing MDV genes (reviewed by Zelnik, 1995) plex virus type 1 and 2 (HSV-1 and HSV-2) genes encod- and MD infection has been controlled effectively by ing for immediate early (IE) genes, ICP4, ICP22, ICP27 vaccination of chicks at 1 day of age using one or more and late (L) genes, gB, gC, gD, gE, gH, gI, gK, and gL serotypes (Okazaki et al., 1970; Schat and Calnek, 1978; have been cloned and sequenced (Anderson et al., 1992; Witter et al., 1984), limited information is available on Binns and Ross, 1989; Brunovskis and Velicer, 1995; Ren the importance of individual MDV antigens in the induc- et al., 1994; Ross et al., 1989; Sakaguchi et al., 1992; tion of CMI responses and protective immunity. The Scott et al., 1993; Yoshida et al., 1994). Genes unique to study of cell-mediated immune (CMI) responses during MDV, pp38, pp14, Meq, A41, L1, and 1.69-kb cDNA of MD infection has been problematic due to the lack of MHC-defined target cells for use in cytotoxicity assays. Numerous investigators have used MD-tumor cell lines 1 To whom correspondence and reprint requests should be ad- dressed. Fax: (607) 253-3384. E-mail: [email protected]. as target cells demonstrating in vitro lysis of allogeneic 87 0042-6822/96 $18.00 Copyright q 1996 by Academic Press, Inc. All rights of reproduction in any form reserved.
-
Upload
independent -
Category
Documents
-
view
1 -
download
0
Transcript of Syngeneic Marek's Disease Virus (MDV)Specific Cell-Mediated Immune Responses against Immediate...

VIROLOGY 222, 87–99 (1996)ARTICLE NO. 0400
Syngeneic Marek’s Disease Virus (MDV)-Specific Cell-Mediated Immune Responsesagainst Immediate Early, Late, and Unique MDV Proteins
ABDUL R. OMAR and KAREL A. SCHAT1
Unit of Avian Medicine, Department of Microbiology and Immunology, College of Veterinary Medicine, Cornell University, Ithaca, New York 14853
Received March 25, 1996; accepted June 12, 1996
Marek’s disease (MD) infection has been controlled effectively by vaccination using nononcogenic and/or attenuatedoncogenic Marek’s disease virus (MDV) vaccines. Thus far, there is little knowledge on the role of cell-mediated immune(CMI) responses during MDV infection or vaccination. To elucidate the importance of MDV proteins in CMI responses, thepp38, Meq, ICP4, or ICP22 genes of an oncogenic strain, GA and the gB, ORF A, A41, or L1 genes of a highly oncogenicstrain, RB1B were stably transfected into reticuloendotheliosis virus (REV)-transformed lymphoblastoid cells, CU-91 (MHC:B19B19) and CU-205 (MHC: B21B21). Cell lines positive for MDV gene transcription and/or protein expression were used in astandard 4-hr chromium release assay. Effector cells for this assay were obtained from splenocytes of chickens infectedwith the oncogenic strain, JM-16/13 or the nononcogenic vaccine strain, SB-1/12. Cell lines expressing MDV pp38, Meq,or gB were lysed by syngeneic but not allogeneic MDV-sensitized splenocytes obtained from chickens of B19B19 and B21B21
haplotypes. However, syngeneic CMI responses against ICP4 were detected only in B21B21 chickens. CMI responses werenot detected against B19B19 and B21B21 cell lines expressing A41, L1, ORF A, or ICP22. This report suggests that syngeneicCMI responses against pp38, Meq, ICP4, and gB of GA and RB1B strains, respectively, can be induced in chickens inoculatedwith JM16/13 or SB-1/12. The difference in CMI response to ICP4 in genetically susceptible (B19B19) and genetically resistant(B21B21) chickens may be an important factor in genetic resistance. q 1996 Academic Press, Inc.
INTRODUCTION BamHI-H family [open reading frame (ORF) A, B, C, D, E,and F] have also been cloned and sequenced (Chen et
Marek’s disease (MD), an economically important, Tal., 1992; Hong and Coussens, 1994; Jones et al., 1992;
cell lymphoproliferative disease in chickens is caused byOhashi et al., 1994a, 1994b; Peng et al., 1992). Depending
Marek’s disease virus (MDV), an oncogenic herpesvirus on the host major histocompatibility (MHC) genes, the(Churchill and Biggs, 1967). MDV can be divided into majority of cytotoxic T lymphocytes (CTLs) during alpha-three serotypes; MDV serotype 1 (MDV-1) consists of all herpesvirus infections in mice are directed against IEoncogenic and attenuated strains, MDV serotype 2 and/or L genes products (Banks et al., 1991; Hanke et(MDV-2) consists of naturally occurring nononcogenic al., 1991; Martin et al., 1990; Witmer et al., 1990). Forstrains, and MDV serotype 3 (MDV-3) consists of non- example in HSV-1 infection, ICP27 and ICP4 were targetpathogenic strains from turkeys and is known as herpes- antigens for CTLs in mice of H-2d and H-2k haplotypes,virus of turkey (HVT) (reviewed by Schat, 1987). respectively (Banks et al., 1991; Martin et al., 1990). Con-
MDV and HVT were initially classified as gammaher- versely, gB-specific CTLs were only detected in mice ofpesviruses on the basis of their biological properties. H-2b and H-2d haplotypes (Hanke et al., 1991; Witmer etHowever, the overall genome structures of MDV and HVT al., 1990).are closely related to alphaherpesviruses (Buckmaster Although significant progress has been made inet al., 1988). MDV genes with homology to herpes sim- characterizing MDV genes (reviewed by Zelnik, 1995)plex virus type 1 and 2 (HSV-1 and HSV-2) genes encod- and MD infection has been controlled effectively bying for immediate early (IE) genes, ICP4, ICP22, ICP27 vaccination of chicks at 1 day of age using one or moreand late (L) genes, gB, gC, gD, gE, gH, gI, gK, and gL serotypes (Okazaki et al., 1970; Schat and Calnek, 1978;have been cloned and sequenced (Anderson et al., 1992; Witter et al., 1984), limited information is available onBinns and Ross, 1989; Brunovskis and Velicer, 1995; Ren the importance of individual MDV antigens in the induc-et al., 1994; Ross et al., 1989; Sakaguchi et al., 1992; tion of CMI responses and protective immunity. TheScott et al., 1993; Yoshida et al., 1994). Genes unique to study of cell-mediated immune (CMI) responses duringMDV, pp38, pp14, Meq, A41, L1, and 1.69-kb cDNA of MD infection has been problematic due to the lack of
MHC-defined target cells for use in cytotoxicity assays.Numerous investigators have used MD-tumor cell lines1 To whom correspondence and reprint requests should be ad-
dressed. Fax: (607) 253-3384. E-mail: [email protected]. as target cells demonstrating in vitro lysis of allogeneic
870042-6822/96 $18.00Copyright q 1996 by Academic Press, Inc.All rights of reproduction in any form reserved.
AID VY 8055 / 6a1c$$$$41 07-08-96 11:11:25 viras AP: Virology

88 OMAR AND SCHAT
but not syngeneic target cells by T cells obtained from expression vectors. MDV genes encoding for pp38, Meq,L1, ICP22, or gB were obtained by using DNA polymerasechickens inoculated with MDV-1 (Calnek et al., 1979;
Powell et al., 1983), MDV-2 (Schat and Calnek, 1980; chain reactions (PCRs). The templates and oligonucleo-tide primers used in amplifying these MDV genes areSchat et al., 1982b), or HVT (Sharma et al., 1977). The
use of MDV-infected chicken kidney cells (CKC) as tar- shown in Table 1. The MDV gB and ICP22 open readingframes (ORFs) were obtained from MDV-1 strains, RB1Bget cells in cytotoxicity assays is also not very success-
ful (Schat and Heller, 1985). In the avian system, synge- and GA-infected CKC DNA, respectively (Schat et al.,1982a). The L1 ORF was generated from CU-41, a RB1B-neic MHC-restricted virus-specific CD8/ CTLs have
been demonstrated in spleens of chickens infected transformed nonexpression cell line DNA (Ohashi et al.,1994a). The pp38 gene was amplified from pMD11, whichwith reticuloendotheliosis virus (REV) (Lillehoj et al.,
1988; Maccubbin and Schierman, 1986; Weinstock et contains a 5-kb fragment of BamHI-H region of MDVgenome which was kindly provided by P. J. A. Sonder-al., 1989). Recently, MHC-restricted, CD8/ virus-spe-
cific CTLs have been demonstrated during avian leuko- meijer (Intervet, Boxmeer, The Netherlands). The Meqgene was amplified from RCAS-Meq, which contains asis virus (ALV) infection (Thacker et al., 1995). Pre-
viously, we developed a system in which we were able 1395-bp ClaI fragment encoding for Meq which was agift from H-J. Kung (Case Western University, Cleveland,to transfect and express the MDV pp38 gene in REV-
transformed lymphoblastoid cell lines. These cell lines OH). As shown in Table 1, upstream and downstreamoligonucleotide primers complementary to the 5* and 3*have been used successfully to demonstrate that syn-
geneic but not allogeneic MDV-sensitized splenocytes ends of pp38 (Chen et al., 1992), Meq (Jones et al., 1992),L1 (Ohashi et al., 1994a), and ICP22 (Sakaguchi et al.,lyse these target cells (Pratt et al., 1992; Uni et al.,
1994). 1992) genes, respectively, were designed with HindIIIand XbaI sites for PCR subcloning. Upstream and down-To elucidate the importance of individual MDV proteins
in CMI responses, we cloned MDV genes pp38, Meq, stream oligonucleotide primers complementary to the 5*and 3* ends of gB gene (Ross et al., 1989) were designedgB, ICP4, ICP22, ORF A, A41, or L1 into an eukaryotic
expression vector and transfected them into REV-trans- with SpeI and NotI sites, respectively. The amplified prod-ucts of HindIII– XbaI fragments of pp38, Meq, L1, ICP22,formed lymphoblastoid cell lines, CU-91 (B19B19) and CU-
205 (B21B21). We demonstrated syngeneic but not alloge- and SpeI–NotI gB fragment were ligated into a plasmid,pBluescript (II) KS(/) [pBS/KS(/)] (Stratagene, La Jolla,neic CMI responses against pp38, Meq, and gB in chick-
ens of B19B19 and B21B21 haplotypes. Interestingly, CMI CA), termed pBS/KS-pp38, pBS/KS-Meq, pBS/KS-L1, pBS/KS-ICP22, and pBS/KS-gB, respectively, and were se-responses against ICP4 were only detected in chickens
with B21B21 haplotype, which is a MD-resistant line. MDV- quenced completely by using the dideoxy chain termina-tion method to verify their sequences (Sequenase ver-specific CMI responses were not detected against
ICP22, ORF A, A41, or L1 in chickens of both haplotypes. sion 2.0 DNA sequencing kit, United States Biochemical,Cleveland, OH).
The pBS/KS-pp38, pBS/KS-Meq, pBS/KS-L1, and pBS/MATERIALS AND METHODSKS-ICP22 plasmids were digested with HindIII and XbaI
Animals and the 938 bp of pp38, 1155 bp of Meq, 468 bp of L1,or 573 bp of ICP22 fragments were subcloned into pRc/Specific pathogen-free chickens of P2a (B19B19) andCMV to generate pRc/CMV-pp38, pRc/CMV-Meq, pRc/N2a (B21B21) lines were obtained from the departmentalCMV-L1, or pRc/CMV-ICP22, respectively. The pBS/KS-flocks at Cornell University (Weinstock and Schat, 1987).gB plasmid was digested with SpeI and NotI and the2651 bp of gB fragment was subcloned into an eukaryoticVirusesexpression vector, pRc/RSV (Invitrogen, San Diego, CA),
Nononcogenic MDV-2, SB-1 passage 12 (SB-1/12) was and termed pRc/RSV-gB. pRc/CMV-gB was obtained bypropagated in chicken embryo fibroblast (CEF) while an ligating the SpeI–NotI fragment of gB from pRc/RSV intooncogenic MDV-1, JM-16 passage 13 (JM16/13), was pRc/CMV by using a NotI linker. The plasmid pRc/CMV-propagated in chicken kidney cells (CKC) as previously A41 was generated by digesting the 641-bp A41 fragmentdescribed (Schat et al., 1982b; Uni et al., 1994). The non- from pBS/KS-A41 (Ohashi et al., 1994b) with HindIII anddefective low-virulence CS-strain of REV (cs-REV) was XbaI. The pRc/CMV-ORF A plasmid was constructed bypropagated in CKC and was used as previously de- digesting the 201-bp ORF A fragment (Peng et al., 1992)scribed (Weinstock et al., 1989). with HindIII and XbaI from pRc/RSV-ORF A, which was
kindly provided by M. Nonoyama (Tampa Bay ResearchPlasmid constructionInstitute, St. Petersburg, FL). Plasmid pRc/CMV-ICP4 wasderived by digesting the plasmid pMD145 (Anderson etAn eukaryotic expression vector, pRc/CMV (Invitrogen,
San Diego, CA) was used for the construction of MDV al., 1992), which was kindly provided by R. Morgan (Uni-
AID VY 8055 / 6a1c$$$$42 07-08-96 11:11:25 viras AP: Virology

89MDV-SPECIFIC CELL-MEDIATED IMMUNE RESPONSES
TABLE 1
Upstream and Downstream Oligonucleotide Primers Used in PCR Subcloning of Various MDV Open Reading Frames
Template Sequencea Nucleotide position MDV gene/size (bp)b
pMD11c 5*-GGGAAGCTTGTGATGGAATTCGAAGCAGAACAC*3 367 to 390 pp38/9385*GGGCCCTCTAGAAATTCGCTTAATCTCCGCCTC*3 1271 to 1292
RCAS-Meqd 5*AAAAAGCTTGAGATGTCTCAGGAGCCAGAGC*3 161 to 182 Meq/11555*GGGCCCTCTAGACTTTAACCCTTTCCTTTAT*3 1289 to 1307
CU41 DNAe 5*GCAAGCTTCCGATACACAGGCACATAG*3 702 to 720 L1/4685*CGTCTAGACGGCACTATCGGTACAACA*3 1142 to 1160
RB1B DNAf 5*GGGCCCACTAGTGCATCATTAAGATCATCTCA*3 028 to 03 gB/26515*GGCGGCCGCTCAGTTCAAATATAATAGTGCC*3 2600 to 2623
GA DNAf 5*AGGAAGCTTGGCAACATGAGTCGTGATCGA*3 2075 to 2095 ICP22/5735*CCCGGGTCTAGACTCTTCAAGTCTGGATAGTTA*3 2618 to 2638
a The oligonucleotide primers are designed based on published sequences described under Materials and Methods. The sites used in PCRsubcloning are underlined and the start codon for pp38, Meq and ICP22 are in bold type. The start codon for L1 and gB are located upstream fromthe shown sequence of the primers.
b The sizes of the amplified products are indicated in base pair (bp).c Plasmid pMD11 contains a 5-kb fragment of BamHI-H region of MDV DNA, which was kindly provided by P. J. A. Sondermeijer (Intervert,
Boxmeer, The Netherlands).d Plasmid RCAS-Meq consists of a 1395-bp ClaI fragment encoding for Meq was a gift from H-J. Kung (Case Western University, Cleveland, OH).e CU-41, a MDV-1 strain, RB1B-transformed nonexpression cell line (Ohashi et al., 1994a).f MDV-1 strains, RB1B and GA-infected chicken kidney cells DNA (Schat et al., 1982a).
versity of Delaware, Newark, DE) with BamHI and the tional in REV-transformed lymphoblastoid cell lines, areporter plasmid, pRc/CMV-lacZ was constructed by sub-4.2-kb ICP4 fragment was subcloned into the pRc/CMV
by using a NotI linker. The location and direction of tran- cloning the HindIII– XbaI fragment of the lacZ gene frompCH110 (Pharmacia, Uppsala, Sweden) into pRc/CMV.scription of MDV genes used in this study are shown in
Fig. 1. In order to determine if the CMV promoter is func- All restriction enzyme reactions and subcloning proce-dures were carried out as recommended by the manufac-turer of the restriction enzymes and as previously de-scribed (Sambrook et al., 1989).
DNA polymerase chain reaction
One hundred nanograms of template was added intoa 100-ml reaction-mixture containing 10 mM Tris–HCl(pH 9.0 at 257), 50 mM KCl, 0.1% Triton X-100, 1.5 mMMgCl2 , 0.2 mM of each deoxynucleoside triphosphate,1.0 mM of each oligonucleotide primer, and 2.0 U of TliDNA polymerase with the proof-reading mechanisms(Promega, Madison, WI). The reaction mixture was sub-jected to 35 cycles of amplification (947 for 1 min, 457 for2 min, and 727 for 1 min).
Preparation of plasmid DNA, transfection,b-galactosidase assays, and establishmentof stable cell lines
Plasmids for transfection were prepared by using QIA-GEN plasmid midi kit (Qiagen Inc., Studio City, CA).FIG. 1. Physical map of the MDV alphaherpesvirus genome structure
(Buckmaster et al., 1988). The MDV genome consists of a unique long Transfection of REV-transformed lymphoblastoid cell(UL) region, flanked by a terminal repeat long (TRL), and an internal lines was performed as previously described (Schat etrepeat long (IRL), and a unique short (US) region, flanked by an internal al., 1992). Briefly 4 1 106 REV-transformed lymphoblas-repeat short (IRS) and a terminal repeat short (TRS). Restriction sites,
toid cells, CU-91 (B19B19) an CU-205 (B21B21) were resus-B Å BamHI and E Å EcoRI. The location of the BamHI-K3, I3, A, H, I2,pended in 0.4 ml of LM base medium (Schat et al., 1992)Q2, and L fragments of MDV genome and the direction of transcription
of gB, ICP4, ICP22, pp38, ORF A, Meq, A41, and L1 are shown. with 5 mg of linearized plasmid in a 0.4-cm sterilized
AID VY 8055 / 6a1c$$$$42 07-08-96 11:11:25 viras AP: Virology

90 OMAR AND SCHAT
cuvette, transfected by electroporation using the Gene enhancer region, the polyadenylation signals, the MDVgene, or the neomycin gene of the plasmids. The di-Pulser Apparatus (Bio-Rad, Richmond, CA) set at 300 V
with capacitance extender set at 500 mF and were left gested DNA were electrophoresed on a 0.8% agaroseand transferred to Magnagraph nylon membranes (Mi-at room temperature for 10 min. Cells were resuspended
in 4 ml of LM base with 10% fetal bovine serum (FBS) cron Separations, Inc., Westborough, MA) by Southernblotting (Sambrook et al., 1989).(LM10) (GIBCO BRL, Grand Island, NY) and incubated at
417. Fourty-eight hours after transfection, 500 to 1400 mg/ A neomycin-specific probe was prepared by digestingpRc/CMV with SmaI and BstBI and was used to hybridizeml of active G418 (Life Technologies Inc., Grand Island,
NY) was added. Media were changed every 2 to 3 days. to CU-366 and CU-359 DNA. Specific probes for pp38,Meq, A41, L1, ORF A, and ICP22 were prepared by digest-Selections were carried out for 3 to 4 weeks. G418-resis-
tant cells were biologically cloned by limiting dilution ing pRc/CMV-pp38, pRc/CMV-Meq, pRc/CMV-A41, pRc/CMV-L1, pRc/CMV-ORF A, and pRc/CMV-ICP22 plas-cloning. For the transient expression of lacZ, 10 mg of
supercoiled pRc/CMV-lacZ was used and 48 hr after mids, respectively, with HindIII and XbaI and were usedto hybridize the DNA obtained from the cell linestransfection cells were pooled, fixed with acetone:metha-
nol (1:1) and stained with 1 mg/ml of 5-bromo-4-chloro-3- transfected with the corresponding MDV gene. The ICP4-specific probe was prepared by digesting the plasmidindolyl-b-D-galactopyrano-side (X-gal) (Sigma Chemical
Co., St. Louis, MO) and 5 mM K4Fe (CN)6r3H2O, and 5 pRc/CMV-ICP4 with NotI and was used to hybridize CU-373 and CU-375 DNA. CU-371 and CU-368 DNA weremM K4Fe(CN)6 . The slides were rinsed in 10 mM Tris-
buffered saline (pH 8.7) (PBS) and examined microscopi- hybridized with a gB-specific probe which was preparedby digesting pRc/CMV-gB with NotI. The digested plas-cally for blue staining cells.mids were electrophoresed on a 1.0% SeaKam Le agar-
Nomenclature of cell lines ose gel (FMC bioproducts, Rockland, ME) and the MDVfragments were gel purified by using DNA purification
Cell lines are identified following the standard nomen-QuicKit (National Scientific Supply, San Rafael, CA) and
clature for avian cell lines (Witter et al., 1979). Briefly, alabeled with [32P]dCTP by using a multiprime DNA label-
two-letter abbreviation identifies the transforming virusing kit (Amersham Corp. Amersham, UK).
(RE), and two-letter abbreviation identifies the line as achicken cell line (CC). This four-letter code is followed
Isolation of cellular RNA and reverse transcriptaseby a two-letter abbreviation for the place of origin (Cornellpolymerase chain reactionUniversity (CU)) and a sequential number (e.g., CU-91).
Stably transfected cell lines received a new number.Total cellular RNA from nontransfected and MDV-
Nontransfected and MDV-transfected REV-transformedtransfected REV-transformed lymphoblastoid cells were
lymphoblastoid cells which were used as target cells inisolated using RNAzol B method (BIOTECX Laboratories
4-hr chromium release assays (CRAs) are summarizedInc., Houston, TX). Briefly, precipitated RNA samples
in Table 2. REV-transformed lymphoblastoid cell lineswere washed with 75% ethanol and reconstituted in di-
(RECC) CU-91 and CU-205 were established from a P2aethylpyrocarbonate (DEPC)-treated water, and the RNA
(B19B19) and a N2a (B21B21) chicken, respectively (Schatconcentrations were determined by measuring the A260et al., 1992). All cell lines were maintained in LM 10 asvalue. The RNA samples were treated with 200 mg/ml of
previously described (Uni et al., 1994) and stablyribonuclease-free DNAse I (Promega, Madison, WI) at
transfected cell lines were maintained in LM 10 with 300377 for 1 hr. All RNA preparations were stored in aliquots,
mg of active G418.precipitated with an equal volume of ethanol, and storedat 0707. Approximately 2 mg of DNAse-treated RNA sam-Southern hybridizationples were used in reverse transcriptase polymerasechain reactions (RT-PCRs) using the GeneAmp RNA PCRTotal DNA was extracted from parent and MDV-
transfected cell lines (Table 2) by standard procedure kit (Perkin–Elmer–Cetus, Norwalk, CT), following theprotocol provided by the manufacturer. Samples were(Sambrook et al., 1989). Twelve micrograms of DNA from
CU-364 and CU-362 (pp38/), CU-365 and CU-376 also amplified without the RT step to demonstrate theabsence of DNA. Upstream and downstream oligonucle-(A41/), CU-371 and CU-368 (gB/), CU-374 and CU-378
(ICP22), CU-370 and CU-377 (ORF A/), and CU-366 and otide primers complementary to the internal coding re-gion of the 5* and 3* region of the MDV genes, respec-CU-359 (neo/) cell lines were digested overnight with
Bgl II. DNA from CU-363 and CU-369 (Meq/) and CU- tively, are shown in Table 3. To ensure the specificity ofthe PCR products, the amplified products were resolved372 and CU-367 (L1/) were digested overnight with XmnI
and CU-373 and CU-375 (ICP4/) DNA were digested in 2.0% SeaKam Le agarose gel (FMC bioproducts, Rock-land, ME), transferred to nylon membranes, and probedovernight with MunI. These restriction enzymes linear-
ized the transfected plasmids outside the CMV promoter/ simultaneously with the Southern blot membranes.
AID VY 8055 / 6a1c$$$$42 07-08-96 11:11:25 viras AP: Virology

91MDV-SPECIFIC CELL-MEDIATED IMMUNE RESPONSES
Antibodies and indirect immunofluorescent antibody target cells with effector/target cell (E/T) ratio of 100:1for 4 hr at 387. All cell lines were tested in two indepen-assaydent assays using splenocytes from REV-, JM16/13-, and
Indirect immunofluorescent antibody assays (IIFAs) SB-1/12-infected chickens. Following a 4-hr incubation,were used to detect the expression of MDV pp38, Meq, supernatants and pellets were separately harvested andgB, and A41 antigens. Monoclonal antibodies against counts per minute (cpm) were counted in a Beckmanpp38 (H19.47) (Yanagida et al., 1992) (1:3000) and gB Gamma 4000 counter (Beckman Instruments, Inc., Fuller-(IAN86) (Yanagida et al., 1992) (1:64) were kindly provided ton, CA) for 5 min. Each assay was performed in triplicate,by L. F. Lee (Avian Disease and Oncology Laboratory, and the spontaneous release was less than 25% of theEast Lansing, Michigan). Rabbit polyclonal antibody total release by treatment with detergent in all assays.against Meq protein (Jones et al., 1992) was kindly pro- The percentage-specific release (%SR) was calculatedvided by H-J. Kung (Case Western University, Cleveland, with the following formula:OH) and was used at 1:40 dilution. Detection of A41antigen was carried out by using a chicken polyclonal
%SRantibody (1:80) raised against glutathione S-transferasefusion proteins which contain a portion of the A41 se-quence (Ohashi et al., 1994b). Briefly, acetone:methanol Å
cpm supernatant of target cells with effectorcells from SB-1 or JM-16 infected chickens 0 X
Y 0 X(1:1)-fixed cells were incubated with monoclonal or poly-clonal antibody for 30 min at 377, washed in phosphate
1 100buffer saline (PBS), pH 7.2, for 15 min, and incubatedwith fluorescein isothiocyanate (FITC)-conjugated rabbit
in which X is the mean of the cpm in the supernatant ofanti-mouse (1:100), FITC-conjugated goat anti-rabbituninfected chickens and Y is the mean of the total re-(1:100) or FITC-conjugated rabbit anti-chicken (1:100)lease calculated by incubating an equivalent number ofsecondary antibodies (Kirkegaard & Perry Laboratories,labeled target cells and effector cells with 2% sodiumInc., Gaithersburg, MD). Cells were examined with a Leitzdodecyl sulfate. The mean percentage-specific releaseDialux fluorescence microscope equipped with epi-illu-(%SR) values were compared using Student’s t testmination.(Snedecor and Cochran, 1971).
Chromium release assayRESULTS
Nontransfected and MDV-transfected REV-transformedTransient expression of lacZ and establishment oflymphoblastoid cells were used as target cells in 4-hr chro-stably transfected REV-transformed lymphoblastoidmium release assays (CRAs). The target cells were centri-cell linesfuged 24 hr before the assay at 1500 rpm for 15 min over
a Ficoll-Hypaque gradient (Pharmacia, Uppsala, Sweden)The expression of lacZ gene could be detected be-
and seeded at 106 cells per ml. On the day of the assay, thetween 24 and 48 hr after transfection, indicating that the
target cells were centrifuged again over a Ficoll-HypaqueCMV promoter is functional in REV-transformed lymph-
gradient, washed in PBS, pH 7.2, and incubated for 45 minoblastoid cells. The level of lacZ expression was deter-
at 387 with 150–200 mCi of Na51CrO4 (sp act, 200 to 400mined by counting the number of cells stained blue over
Ci/mg, New England Nuclear, Boston, MA) per 107 cellsthe total number of cells. Transfection of CU-91 and CU-
per ml of LM20 (RPMI 1640 / 20% FBS). Afterwards cells205 with pRc/CMV-lacZ resulted in 2% and 3 to 4% of
were washed seven times in LM20 and resuspended incells expressing lacZ, respectively. For the establishment
LM20 at 0.5 1 106 cells/ml.of stable cell lines, transfected cells surviving the G418
The preparation of effector cells, conditions of CRAs,selection were examined for MDV expression by IIFAs
and the calculations of percentage-specific release haveand/or RT-PCRs. Cells that were positive for MDV tran-
previously been described (Schat et al., 1982b). Briefly,scription and/or expression were stored in liquid nitrogen
spleens were decapsulated and forced through a 60-mmand a portion of the cells were biologically cloned by
pore size mesh (Tetko, Inc., Elmsford, NY). Spleen cellslimiting dilution. Stably transfected cell lines are listed in
were washed in PBS, pH 7.2, and were centrifuged atTable 2.
1500 rpm for 20 min over a Ficoll-Hypaque gradient. Inall experiments six birds/treatment were used. Effector Detection of transfected MDV genes in MDV-cells were obtained from spleens 7 days postinfection transfected REV-transformed lymphoblastoid cellswith 104.2 tissue culture infective dose 50% of cs-REV, or1000 focus-forming units of JM16/13 or SB-1/12. Control Southern blot analyses of the transfected cell lines
confirmed the presence of the transfected plasmids ap-effector cells were obtained from uninfected hatchmates.The CRA was performed by incubating 50,000 labeled parently integrated at one or more unique sites (Fig. 2).
AID VY 8055 / 6a1c$$$$42 07-08-96 11:11:25 viras AP: Virology

92 OMAR AND SCHAT
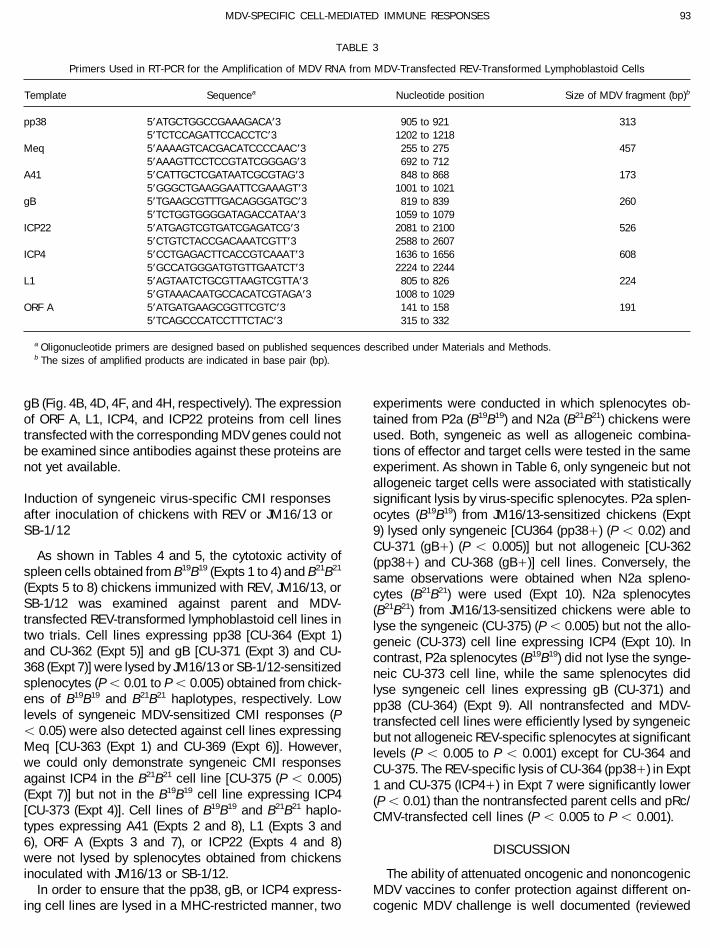
TABLE 2 3A), 457 (Fig. 3B), 173 (Fig. 3C), 260 (Fig. 3D), 526 (Fig.3E), 608 (Fig. 3F), 224 (Fig. 3G), and 191 (Fig. 3H) bpNontransfected and MDV-Transfected REV-Transformedwere detected from CU-364 and CU-362 (pp38/), CU-Lymphoblastoid Cell Lines363 and CU-369 (Meq/), CU-365 and CU-376 (A41/),
Transfected CU-371 and CU-368 (gB/), CU-374 and CU-378B19B19 cell line B21B21 cell line plasmid (ICP22/), CU-373 and CU-375 (ICP4/), CU-372 and CU-
367 (L1/), and CU-370 and CU-377 (ORF A/) RNA, re-CU-91 CU-205 nonespectively, after hybridization with the correspondingCU-366 CU-359 pRc/CMV
CU-364 CU-362 pRc/CMV-pp38 MDV genes as specific DNA probes. Bands were notCU-363 CU-369 pRc/CMV-Meq detected in PCR products from the total RNA samplesCU-365 CU-376 pRc/CMV-A41 isolated from MDV-transfected cell lines in the absenceCU-371 CU-368 pRc/CMV-gB
of reverse transcriptase or when PCR products wereCU-374 CU-378 pRc/CMV-ICP22used from the parent cell lines, CU91 and CU-205.CU-373 CU-375 pRc/CMV-ICP4
CU-372 CU-367 pRc/CMV-L1CU-370 CU-377 pRc/CMV-ORF A Expression of MDV genes in transfected REV-
transformed lymphoblastoid cell lines
Based on the IIFAs, the staining patterns of the differ-DNA extracted from nontransfected cell lines, CU-91 andCU-205, did not hybridize with probes prepared from ent MDV proteins in MDV-transfected REV-transformed
lymphoblastoid cells were the same as in MDV-infectedpp38, Meq, A41, gB, L1, ICP22, ICP4, ORF A, or neomycin[data not shown, except for pp38 (Fig. 2, lanes 1 and 2)]. CEF and MDV-infected CKC (Ohashi et al., 1994b; Yanag-
ida et al., 1992). Most cell lines expressed the MDV pro-The specificity of the Southern blots (Fig. 2) was con-firmed by the hybridization of each probe to the linearized teins in 40 to 60% of the cells. As shown in Fig. 4A, cell
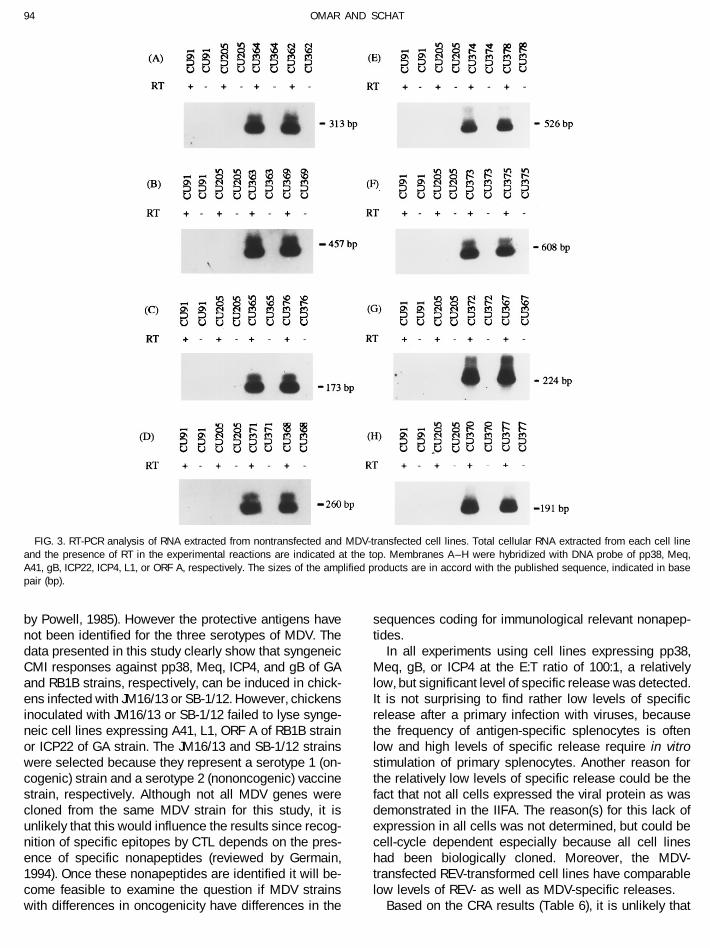
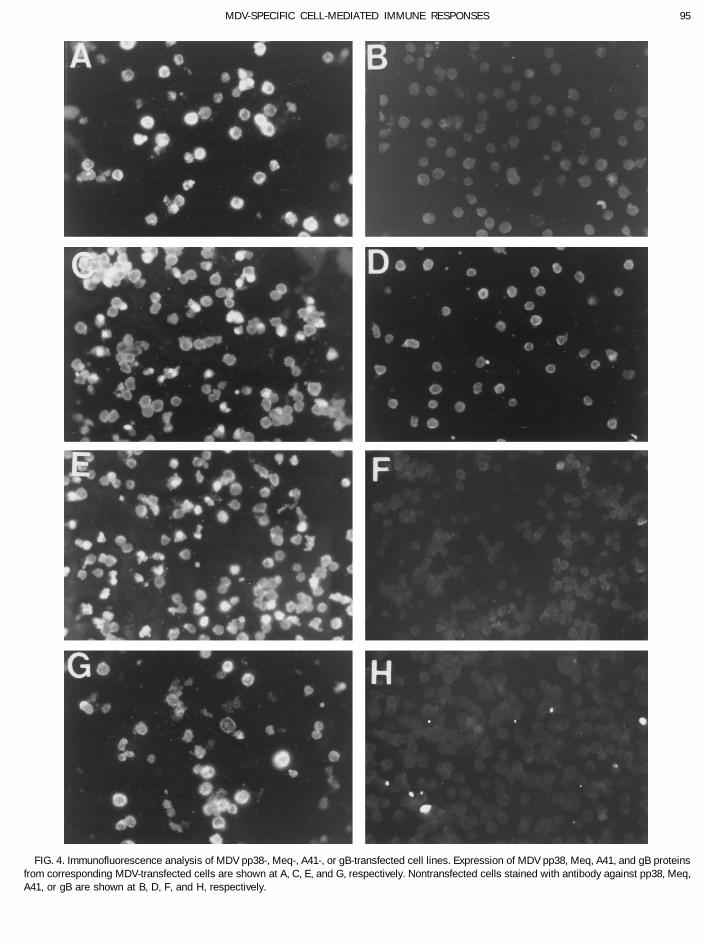
lines transfected with pp38 stained predominately in theexpression plasmid.Transcription of the individually transfected MDV cytoplasmic region while Meq protein was detected in
the nucleus (Fig. 4C). The expression of A41 protein wasgenes were confirmed by RT-PCRs using oligonucleotideprimers described in Table 3. Ten microliters of RT-PCR also localized in the cytoplasma (Fig. 4E). gB protein was
predominately expressed in the cytoplasma and had aproducts from 2 mg of total RNA isolated from cell lineswith and without reverse transcriptase were analyzed by granular appearance (Fig. 4G). Nontransfected cells did
not stain with antibodies against pp38, Meq, A41, andgel electrophoresis. As expected, fragments of 313 (Fig.
FIG. 2. Detection of integrated transfected plasmids by Southern blot analysis. Genomic DNA from each indicated cell line was digested withBgl II (lanes 1 to 5, 9 to 17, and 24 to 29), XmnI (lanes 6 to 8 and 21 to 23), or MunI (lanes 18 to 20), which cuts once within the transfectedexpression vectors. The data from nontransfected cells, CU-91 and CU-205 hybridized with Meq, A41, L1, gB, ICP22, ICP4, ORF A, or neomycinexcept pp38 were not shown. The specificity of this Southern blot was verified by the detection of fragments of expected sizes (lanes 5, 8, 11, 14,17, 20, 23, 26, and 29) from the linearized expression vectors. Molecular size markers are indicated in kilobases on the left.
AID VY 8055 / 6a1c$$$$43 07-08-96 11:11:25 viras AP: Virology
93MDV-SPECIFIC CELL-MEDIATED IMMUNE RESPONSES
TABLE 3
Primers Used in RT-PCR for the Amplification of MDV RNA from MDV-Transfected REV-Transformed Lymphoblastoid Cells
Template Sequencea Nucleotide position Size of MDV fragment (bp)b
pp38 5*ATGCTGGCCGAAAGACA*3 905 to 921 3135*TCTCCAGATTCCACCTC*3 1202 to 1218
Meq 5*AAAAGTCACGACATCCCCAAC*3 255 to 275 4575*AAAGTTCCTCCGTATCGGGAG*3 692 to 712
A41 5*CATTGCTCGATAATCGCGTAG*3 848 to 868 1735*GGGCTGAAGGAATTCGAAAGT*3 1001 to 1021
gB 5*TGAAGCGTTTGACAGGGATGC*3 819 to 839 2605*TCTGGTGGGGATAGACCATAA*3 1059 to 1079
ICP22 5*ATGAGTCGTGATCGAGATCG*3 2081 to 2100 5265*CTGTCTACCGACAAATCGTT*3 2588 to 2607
ICP4 5*CCTGAGACTTCACCGTCAAAT*3 1636 to 1656 6085*GCCATGGGATGTGTTGAATCT*3 2224 to 2244
L1 5*AGTAATCTGCGTTAAGTCGTTA*3 805 to 826 2245*GTAAACAATGCCACATCGTAGA*3 1008 to 1029
ORF A 5*ATGATGAAGCGGTTCGTC*3 141 to 158 1915*TCAGCCCATCCTTTCTAC*3 315 to 332
a Oligonucleotide primers are designed based on published sequences described under Materials and Methods.b The sizes of amplified products are indicated in base pair (bp).
gB (Fig. 4B, 4D, 4F, and 4H, respectively). The expression experiments were conducted in which splenocytes ob-tained from P2a (B19B19) and N2a (B21B21) chickens wereof ORF A, L1, ICP4, and ICP22 proteins from cell lines
transfected with the corresponding MDV genes could not used. Both, syngeneic as well as allogeneic combina-tions of effector and target cells were tested in the samebe examined since antibodies against these proteins are
not yet available. experiment. As shown in Table 6, only syngeneic but notallogeneic target cells were associated with statistically
Induction of syngeneic virus-specific CMI responses significant lysis by virus-specific splenocytes. P2a splen-after inoculation of chickens with REV or JM16/13 or ocytes (B19B19) from JM16/13-sensitized chickens (ExptSB-1/12 9) lysed only syngeneic [CU364 (pp38/) (P õ 0.02) and
CU-371 (gB/) (P õ 0.005)] but not allogeneic [CU-362As shown in Tables 4 and 5, the cytotoxic activity of (pp38/) and CU-368 (gB/)] cell lines. Conversely, the
spleen cells obtained from B19B19 (Expts 1 to 4) and B21B21
same observations were obtained when N2a spleno-(Expts 5 to 8) chickens immunized with REV, JM16/13, or cytes (B21B21) were used (Expt 10). N2a splenocytesSB-1/12 was examined against parent and MDV- (B21B21) from JM16/13-sensitized chickens were able totransfected REV-transformed lymphoblastoid cell lines in lyse the syngeneic (CU-375) (P õ 0.005) but not the allo-two trials. Cell lines expressing pp38 [CU-364 (Expt 1) geneic (CU-373) cell line expressing ICP4 (Expt 10). Inand CU-362 (Expt 5)] and gB [CU-371 (Expt 3) and CU- contrast, P2a splenocytes (B19B19) did not lyse the synge-368 (Expt 7)] were lysed by JM16/13 or SB-1/12-sensitized neic CU-373 cell line, while the same splenocytes didsplenocytes (Põ 0.01 to Põ 0.005) obtained from chick- lyse syngeneic cell lines expressing gB (CU-371) andens of B19B19 and B21B21 haplotypes, respectively. Low pp38 (CU-364) (Expt 9). All nontransfected and MDV-levels of syngeneic MDV-sensitized CMI responses (P transfected cell lines were efficiently lysed by syngeneicõ 0.05) were also detected against cell lines expressing but not allogeneic REV-specific splenocytes at significantMeq [CU-363 (Expt 1) and CU-369 (Expt 6)]. However, levels (P õ 0.005 to P õ 0.001) except for CU-364 andwe could only demonstrate syngeneic CMI responses CU-375. The REV-specific lysis of CU-364 (pp38/) in Exptagainst ICP4 in the B21B21 cell line [CU-375 (P õ 0.005) 1 and CU-375 (ICP4/) in Expt 7 were significantly lower(Expt 7)] but not in the B19B19 cell line expressing ICP4 (P õ 0.01) than the nontransfected parent cells and pRc/[CU-373 (Expt 4)]. Cell lines of B19B19 and B21B21 haplo- CMV-transfected cell lines (P õ 0.005 to P õ 0.001).types expressing A41 (Expts 2 and 8), L1 (Expts 3 and6), ORF A (Expts 3 and 7), or ICP22 (Expts 4 and 8) DISCUSSIONwere not lysed by splenocytes obtained from chickensinoculated with JM16/13 or SB-1/12. The ability of attenuated oncogenic and nononcogenic
MDV vaccines to confer protection against different on-In order to ensure that the pp38, gB, or ICP4 express-ing cell lines are lysed in a MHC-restricted manner, two cogenic MDV challenge is well documented (reviewed
AID VY 8055 / 6a1c$$$$43 07-08-96 11:11:25 viras AP: Virology
94 OMAR AND SCHAT
FIG. 3. RT-PCR analysis of RNA extracted from nontransfected and MDV-transfected cell lines. Total cellular RNA extracted from each cell lineand the presence of RT in the experimental reactions are indicated at the top. Membranes A–H were hybridized with DNA probe of pp38, Meq,A41, gB, ICP22, ICP4, L1, or ORF A, respectively. The sizes of the amplified products are in accord with the published sequence, indicated in basepair (bp).
by Powell, 1985). However the protective antigens have sequences coding for immunological relevant nonapep-tides.not been identified for the three serotypes of MDV. The
data presented in this study clearly show that syngeneic In all experiments using cell lines expressing pp38,Meq, gB, or ICP4 at the E:T ratio of 100:1, a relativelyCMI responses against pp38, Meq, ICP4, and gB of GA
and RB1B strains, respectively, can be induced in chick- low, but significant level of specific release was detected.It is not surprising to find rather low levels of specificens infected with JM16/13 or SB-1/12. However, chickens
inoculated with JM16/13 or SB-1/12 failed to lyse synge- release after a primary infection with viruses, becausethe frequency of antigen-specific splenocytes is oftenneic cell lines expressing A41, L1, ORF A of RB1B strain
or ICP22 of GA strain. The JM16/13 and SB-1/12 strains low and high levels of specific release require in vitrostimulation of primary splenocytes. Another reason forwere selected because they represent a serotype 1 (on-
cogenic) strain and a serotype 2 (nononcogenic) vaccine the relatively low levels of specific release could be thefact that not all cells expressed the viral protein as wasstrain, respectively. Although not all MDV genes were
cloned from the same MDV strain for this study, it is demonstrated in the IIFA. The reason(s) for this lack ofexpression in all cells was not determined, but could beunlikely that this would influence the results since recog-
nition of specific epitopes by CTL depends on the pres- cell-cycle dependent especially because all cell lineshad been biologically cloned. Moreover, the MDV-ence of specific nonapeptides (reviewed by Germain,
1994). Once these nonapeptides are identified it will be- transfected REV-transformed cell lines have comparablelow levels of REV- as well as MDV-specific releases.come feasible to examine the question if MDV strains
with differences in oncogenicity have differences in the Based on the CRA results (Table 6), it is unlikely that
AID VY 8055 / 6a1c$$$$43 07-08-96 11:11:25 viras AP: Virology
95MDV-SPECIFIC CELL-MEDIATED IMMUNE RESPONSES
FIG. 4. Immunofluorescence analysis of MDV pp38-, Meq-, A41-, or gB-transfected cell lines. Expression of MDV pp38, Meq, A41, and gB proteinsfrom corresponding MDV-transfected cells are shown at A, C, E, and G, respectively. Nontransfected cells stained with antibody against pp38, Meq,A41, or gB are shown at B, D, F, and H, respectively.
AID VY 8055 / 6a1c$$8055 07-08-96 11:11:25 viras AP: Virology

96 OMAR AND SCHAT
TABLE 4
Lysis of REV-Transformed Lymphoblastoid Cell Lines (B19B19 ) Transfected with a Single MDV Gene by SplenocytesObtained from P2a (B19B19) Chickens inoculated with REV or SB-1/12 or JM16/13
% specific releasea of splenocytes obtained from chickens inoculated withb
Target cells Trial 1 Trial 2
Expt Line MDV gene REV JM16 SB-1 REV JM16 SB-1
1 CU-91 None 13c 2 2 9c 1 1CU-366 pRc/CMV 14c 1 1 10c 1 NTCU-364 pp38 6d 10c 8c 6d 8c 7d
CU-363 Meq 9c 7e 5e 12c 6e 6e
2 CU-91 None 11f 2 1 13c 2 2CU-365 A41 14f 1 1 12c 1 1CU-205g None 0 1 0 1 1 0
3 CU-91 None 16c 1 2 16c 1 0CU-370 ORF A 13f 2 0 15c 3 1CU-371 gB 15f 11d 12c 16c 15c 17c
CU-372 L1 10c 3 1 11c 2 04 CU-91 None 30f 2 0 23c 2 02
CU-373 ICP4 42f 3 1 25f 3 1CU-374 ICP22 35f 3 2 20c 1 NTCU-205g None 04 2 2 1 2 0
Note. NT, not tested.a Spontaneous release from target cells did not exceed 25%.b Six chickens/group were used, E/T ratios were 100:1.c Significantly different (P õ 0.005) from control groups.d Significantly different (P õ 0.01) from control groups.e Significantly different (P õ 0.05) from control groups.f Significantly different (P õ 0.001) from control groups.g Allogeneic nontransfected target cells.
the effector cells in these assays were NK cells since Meq by inoculation with the nononcogenic SB-1 strain isof interest, since Meq has only been identified in MDV-syngeneic but not allogeneic lysis could be demon-
strated. Moreover, REV-transformed cells express high 1 (Jones et al., 1992). Probably there is a SB-1 gene withhomology for Meq which has not yet been identified.levels of MHC class I and II antigens (Hrdlickova et al.,
1994). We are currently investigating the phenotype of Even though MD has been controlled effectively byvaccination, the antigens associated with protective im-the effector cells associated with CMI responses against
cell lines expressing pp38, gB, and ICP4. In vitro deple- munity are poorly defined. It has been suggested thatmost MD vaccines protect chickens independently of thetion studies using splenocytes from SB-1/12-infected
chickens indicate that the effector cells are CD8/ and MHC gene background (Bacon and Witter, 1993). How-ever, vaccination confers superior protection in MD-re-TCRab1/ (manuscript in preparation).
The presence of CMI responses against pp38 in chick- sistant chickens compared to MD-susceptible chickens.The mechanism of MHC-related resistance to MD isens inoculated with SB-1 indicate that the gene which
encodes for pp38 is not restricted to MDV-1 as previously poorly understood. However, it is clear that the level ofsecondary cytolytic infection is markedly lower in geneti-proposed by Chen et al. (1992). Indeed, recently Ono et
al. (1994, 1995) reported that the MDV-2 strain HRPS-24 cally resistant, B21B21 chickens than the genetically sus-ceptible, B19B19 chickens (reviewed by Calnek, 1985). Itencodes for a pp38-like gene which is homologous to
pp38 of MDV-1. In addition, the MDV-3 strain FC126 en- is attractive to postulate that the induction of ICP4-spe-cific CMI responses in the B21B21 haplotype chickenscodes for a truncated form of pp38 protein which is highly
homologous to the C-terminal region of MDV-1 pp38 might play an important role in protective immunity byinhibiting the secondary cytolytic infection and the subse-(Smith et al., 1995). These findings are in good agreement
with this study and our previous studies in which specific quent development of immortalized MDV-infected T lym-phocytes.CMI responses against pp38 were demonstrated in
chickens inoculated with SB-1 or HVT (Pratt et al., 1992; The in vivo importance of CMI responses against pp38,Meq, and gB in B21B21 and B19B19 is not yet known. Nazer-Uni et al., 1994). The induction of CMI responses against
AID VY 8055 / 6a1c$$$$43 07-08-96 11:11:25 viras AP: Virology

97MDV-SPECIFIC CELL-MEDIATED IMMUNE RESPONSES
TABLE 5
Lysis of REV-Transformed Lymphoblastoid Cell Lines (B21B21) Transfected with a Single MDV Gene by SplenocytesObtained from N2a (B21B21) Chickens Inoculated with REV or SB-1/12 or JM16/13
% specific releasea of splenocytes obtained from chickens inoculated withb
Target cells Trial 1 Trial 2
Expt Line MDV gene REV JM16 SB-1 REV JM16 SB-1
5 CU-205 None 12c 3 1 13c 2 3CU-359 pRc/CMV 20d 2 2 13d 2 2CU-362 pp38 8c 9c 11c 10c 10c 9c
CU-91g None 01 1 2 0 2 26 CU-205 None 14c 2 1 15d 2 1
CU-367 L1 12c 2 0 13d 2 1CU-369 Meq 13d 7e 5 14c 6e 5e
7 CU-205 None 20d 1 2 23d 1 1CU-368 gB 25d 13c 12c 31d 12c 14c
CU-375 ICP4 8f 13c 13c 6f 14c 16c
CU-377 ORF A 22d 2 1 36d 4 0CU-91g None 04 1 2 01 02 NT
8 CU-205 None 11c 1 0 14c 0 1CU-378 ICP22 21c 3 4 14d 2 2CU-376 A41 13c 2 02 15c 1 0
Note. NT, not tested.a Spontaneous release from target cells did not exceed 25%.b Six chickens/group were used, E/T ratios were 100:1.c Significantly different (P õ 0.005) from control groups.d Significantly different (P õ 0.001) from control groups.e Significantly different (P õ 0.05) from control groups.f Significantly different (P õ 0.01) from control groups.g Allogeneic nontransfected target cells.
ian et al. (1992) reported that B2B15 chickens were pro- against gB. Recently, we have found that B19B19 chickensvaccinated with rFPV-gB also produced gB-specific CMItected against challenge after immunization with recom-
binant fowlpox virus (rFPV) vaccines expressing gB responses (Omar, Schat, and Lee, unpublished data). Itis not surprising that chickens produce both CMI re-(rFPV-gB) but not with rFPV expressing pp38. These
chickens produced virus neutralizing (VN) antibodies sponses and VN antibodies against gB since mice in-
TABLE 6
Syngeneic MHC-Restricted Lysis of Nontransfected and MDV-Transfected REV-Transformed Lymphoblastoid Cell Linesby Splenocytes Obtained from Chickens Inoculated with REV or JM16/13
% specific releasea
MHC B19B19 cell line B21B21 cell line
Effector CU-364 CU-371 CU-373 CU-362 CU-368 CU-375Expt cell Virusb CU-91 (pp38/) (gB/) (ICP4/) CU-205 (pp38/) (gB/) (ICP4/)
9 B19B19 REV 11c 10c 15c 25d 0 1 2 0JM16 1 6e 13c 2 1 1 2 3
10 B21B21 REV 01 2 0 01 15c 12c 13c 8e
JM16 02 0 0 01 2 11c 14c 18c
a Spontaneous release from target cells did not exceed 25%.b Six chickens/group were used, E/T ratios were 100:1.c Significantly different (P õ 0.005) from control groups.d Significantly different (P õ 0.001) from control groups.e Significantly different (P õ 0.02) from control groups.
AID VY 8055 / 6a1c$$$$43 07-08-96 11:11:25 viras AP: Virology

98 OMAR AND SCHAT
mice against lethal herpes simplex virus 1 infection. Proc. Natl. Acad.fected with HSV-1 developed protective gB-specific neu-Sci. USA 84, 5908–5912.tralizing antibodies as well as NK cells and CD4/ and
Chen, X., Sondermeijer, P. J. A., and Velicer, L. F. (1992). Identification ofCD8/ T cells against lethal challenge of HSV-1 (Cantin a unique Marek’s disease virus gene which encodes a 38-kilodaltonet al., 1987; Ghiasi et al., 1994). phosphoprotein and is expressed in both lytically infected cells and
latently infected lymphoblastoid tumor cells. J. Virol. 66, 85–94.The ability of JM16/13- or SB-1/12-infected chickensChurchill, A. E., and Biggs, P. M. (1967). Agent of Marek’s disease into recognize pp38, Meq, ICP4, and gB of GA and RB1B
tissue culture. Nature 215, 528–530.strains, respectively, indicate that there might be com-Germain, R. N. (1994). MHC-dependent antigen processing and peptide
mon CMI epitopes among these serotypes. Serotype- presentation: Providing ligands for T lymphocyte activation. Cell 76,cross-reactive CTL clones recognizing more than one 287–299.
Ghiasi, H., Kaiwar, R., Nesburn, A. B., Slanina, S., and Wechsler, S. L.serotype have been demonstrated in HSV (Booneau et(1994). Expression of seven herpes simplex virus type 1 glycoproteinal., 1990) and dengue virus (Zivny et al., 1995) infections.(gB, gC, gD, gE, gG, gH and gI): Comparative protection againstThe inability of JM16/13- or SB-1/12-sensitized chickenslethal challenge in mice. J. Virol. 68, 2118–2126.
to lyse syngeneic cell lines expressing A41, L1, ORF A, Hanke, T., Graham, F. L., Rosenthal, K. L., and Johnson, D. C. (1991).or ICP22 probably indicate that these genes do not play Identification of an immunodominant cytotoxic T-lymphocyte recogni-
tion site in glycoprotein B of herpes simplex virus by using recombi-a major role in inducing CMI responses.nant adenovirus vectors and synthetic peptides. J. Virol. 65, 1177–1186.
ACKNOWLEDGMENTS Hong, Y., and Coussens, P. M. (1994). Identification of an immediate-early gene in the Marek’s disease virus long internal repeat region
We thank Priscilla O’Connell and Jamie Tarigo for excellent technicalwhich encodes a unique 14-kilodalton polypeptide. J. Virol. 68, 3593–
assistance, H-J. Kung for the RCAS-meq plasmid and antibody against3603.
Meq, M. Nonoyama for the pRc/RSV-ORF A plasmid, R. Morgan for theHrdlickova, R., Nehyba, J., and Humpries, E. H. (1994). v-rel induces
pMD145 plasmid, P. J. A. Sondermeijer for the plasmid pMD11, and L.expression of three avian immunoregulatory surface receptors more
F. Lee for the monoclonal antibodies against pp38 and gB. This workefficiently than c-rel. J. Virol. 68, 308–319.
was supported in part by Grant No. 92-37204-8044 of the USDA Com-Jones, D., Lee, L. F., Liu, J. L., Kung, H-J., and Tillotson, J. K. (1992).
petitive Research Grant Program.Marek’s disease virus encodes a basic-leucine zipper gene resem-bling the fos/jun oncogenes that is highly expressed in lymphoblas-toid tumors. Proc. Natl. Acad. Sci. USA 89, 4042–4046.REFERENCES
Lillehoj, H. S., Lillehoj, E. P., Weinstock, D., and Schat, K. A. (1988).Functional and biochemical characterizations of avian T lymphocyteAnderson, A. S., Francesoni, A., and Morgan, R. W. (1992). Complete
nucleotide sequence of the Marek’s disease virus ICP4 gene. Virol- antigens identified by monoclonal antibodies. Eur. J. Immunol. 18,2059–2065.ogy 189, 657–667.
Bacon, L. D., and Witter, R. L. (1993). Influence of B-haplotype on the Maccubbin, D. B., and Schierman, L. W. (1986). MHC-restricted cyto-toxic response of chicken T cells: expression, augmentation, andrelative efficacy of Marek’s disease vaccines of different serotypes.
Avian Dis. 37, 53–59. clonal characterization. J. Immunol. 136, 12–16.Martin, S., Zhu, X., Silverstein, S. J., Courtney, R. J., Yao, F., Jenkins,Banks, T. A., Allen, E. M., Dasjupta, S., Sandri-Goldin, R., and Rouse,
B. T. (1991). Herpes simplex virus type 1-specific cytotoxic T lympho- F. J., and Rouse, B. T. (1990). Murine cytotoxic T lymphocytes specificfor herpes simplex type 1 recognize the immediate early protein ICP4cytes recognize immediate-early protein ICP27. J. Virol. 65, 3185–
3191. but not ICP0. J. Gen. Virol. 71, 2391–2399.Nazerian, K., Lee, L. F., Yanagida, N., and Ogawa, R. (1992). ProtectionBinns, M. M., and Ross, N. L. J. (1989). Nucleotide sequence of the
Marek’s disease virus (MDV) RB1B A antigen and the identification against Marek’s disease by a fowlpox virus recombinant expressingthe glycoprotein B of Marek’s disease virus. J. Virol. 66, 1409–1413.of the MDV A antigen as the herpes simplex virus-2 glycoprotein C
homologue. Virus Res. 12, 371–382. Ohashi, K., Zhou, W., O’Connell, P. H., and Schat, K. A. (1994a). Charac-terization of a Marek’s disease virus BamHI-L-specific cDNA cloneBooneau, R. H., Salvucci, L. A., Johnson, D. C., and Tevethia, S. S.
(1990). Epitope specificity of H-2Kb-restricted, HSV-1, and HSV-2 obtained from a Marek’s disease lymphoblastoid cell line. J. Virol.68, 1191–1195.cross-reactive cytotoxic T lymphocytes clones. Virology 195, 62–70.
Brunovskis, P., and Velicer, L. F. (1995). The Marek’s disease virus Ohashi, K., O’Connell, P. H., and Schat, K. A. (1994b). Characterizationof Marek’s disease virus BamHI-A-specific cDNA clones obtained(MDV) unique short region: Alphaherpesvirus-homologous, fowlpox
virus-homologous, and MDV-specific genes. Virology 206, 324–338. from a Marek’s disease lymphoblastoid cell line. Virology 199, 275–283.Buckmaster, A. E., Scott, S. D., Sanderson, M. J., Bournsnell, M. E. G.,
Ross, L. J. N., and Binns, M. M. (1988). Gene sequence and mapping Okazaki, W., Purhase, H. G., and Burmester, B. R. (1970). Protectionagainst Marek’s disease by vaccination with a herpesvirus of turkeys.data from Marek’s disease virus and herpesvirus of turkeys-implica-
tions for herpesvirus classification. J. Gen. Virol. 69, 2033–2042. Avian Dis. 14, 413–429.Omar, A. R., Schat, K. A., and Lee, L. F., unpublished data.Calnek, B. W., Carlisle, J. C., Fabricant, J., Murthy, K. K., and Schat,
K. A. (1979). Comparative pathogenesis studies with oncogenic and Ono, M., Kawaguchi, Y., Maeda, K., Kamiya, N., Tohya, Y., Kai, C., Nii-kura, M., and Mikami, T. (1994). Nucleotide sequence analysis ofnononcogenic Marek’s disease viruses and turkey herpesvirus. Am.
J. Vet. Res. 40, 541–548. Marek’s disease virus (MDV) serotype 2 homology of MDV serotype1 pp38, an antigen associated with transformed cells. Virology 201,Calnek, B. W. (1985). Genetic resistance. In ‘‘Marek’s Disease: Scientific
Basis and Methods of Control’’ (L. N. Payne, Ed.), pp. 293–327. Nij- 142–146.Ono, M., Kawaguchi, Y., Jang, H. K., Tohya, Y., Niikura, M., and Mikami,hoff, Boston, MA.
Cantin, E. M., Eberle, R., Baldick, J. L., Moss, B., Willey, D. E., Notkins, T. (1995). Expression of Marek’s disease virus (MDV) serotype 2gene which has partial homology with MDV serotype 1 pp38 gene.A. L., and Openshaw, H. (1987). Expression of herpes simplex virus
1 glycoprotein B by a recombinant vaccinia virus and protection of Virus Res. 35, 223–229.
AID VY 8055 / 6a1c$$$$43 07-08-96 11:11:25 viras AP: Virology

99MDV-SPECIFIC CELL-MEDIATED IMMUNE RESPONSES
Peng, F., Bardely, G., Tanaka, A., Lancz, G., and Nonoyama, M. (1992). Scott, S. D., Smith, G. D., Ross, N. L. J., and Binns, M. M. (1993). Identifi-cation and sequence analysis of the homologues of the herpes sim-Isolation and characterization of cDNAs from BamHI-H gene family
RNAs associated with the tumorigenicity of Marek’s disease virus. plex virus type 1 glycoprotein H in Marek’s disease virus and theherpesvirus of turkeys. J. Gen. Virol. 74, 1185–1190.J. Virol. 66, 7389–7396.
Sharma, J. M., Witter, R. L., and Coulson, B. D. (1977). Development ofPowell, P. C. (1985). Immunity. In ‘‘Marek’s Disease: Scientific Basiscell-mediated immunity to Marek’s disease tumor cells in chickensand Methods of Control’’ (L. N. Payne, Ed.), pp. 177–202. Nijhoff,inoculated with Marek’s disease vaccines. J. Natl. Cancer. Inst. 61,Boston, MA.1273–1280.Powell, P. C., Mustil, B. M., and Rennie, M. (1983). The role of histocom-
Smith, G. D., Zelnik, V., and Ross, L. J. N. (1995). Gene organization inpatibility antigens in cell-mediated cytotoxicity against Marek’s dis-herpesvirus of turkeys: Identification of a novel open reading frameease tumor-derived lymphoblastoid cell lines. Avian Pathol. 12, 461–in the long unique region and a truncated homologue of pp38 in the468.internal repeat. Virology 207, 205–216.Pratt, W. D., Morgan, R., and Schat, K. A. (1992). Cell-mediated cytolysis
Snedecor, J. L., and Cochran, W. G. (1971). ‘‘Statistical Methods,’’ 6thof lymphoblastoid cells expressing Marek’s disease virus-specificed. Iowa State Univ. Press, Ames.phosphorylated polypeptides. Vet. Microbiol. 33, 93–99.
Thacker, E. L., Fulton, J. E., and Hunt, H. D. (1995). In vitro analysis ofRen, D., Lee, L. F., and Coussens, P. M. (1994). Identification and char-a primary, major histocompatibility complex (MHC)-restricted, cyto-acterization of Marek’s disease virus genes homologous to ICP27toxic T lymphocyte response to avian leukosis virus (ALV) usingand glycoprotein K of herpes simplex virus-1. Virology 204, 242–250.target cells expressing MHC class I cDNA inserted into a recombi-Ross, L. J. N., Sanderson, M., Scott, S. D., Binns, M. M., Doel, T., andnant ALV, vector. J. Virol. 69, 6439–6444.Milne, B. (1989). Nucleotide sequence and characterization of the
Uni, Z., Pratt, W. D., Miller, M. M., O’Connell, P. H., and Schat, K. A.Marek’s disease virus homologue of glycoprotein B of herpes sim-(1994). Syngeneic lysis of reticuloendotheliosis virus-transformed cellplex virus. J. Gen. Virol. 70, 1789–1804.lines transfected with Marek’s disease virus genes by virus-specificSakaguchi, M., Urakawa, T., Hirayama, Y., Miki, N., Yamamoto, M., andcytotoxic T cells. Vet. Immunopathol. 44, 57–69.Hirai, K. (1992). Sequence determination and genetic content of an
Weinstock, D., Schat, K. A., and Calnek, B. W. (1989). Cytotoxic T lympho-8.9-kb restriction fragment in the short unique region and the internalcytes in reticuloendotheliosis virus-infected chickens. Eur. J. Immu-inverted repeat of Marek’s disease virus type 1 DNA. Virus Genesnol. 19, 267–272.6, 365–378.
Weinstock, D., and Schat, K. A. (1987). Virus specific syngeneic killingSambrook, J., Fritch, E. F., and Maniatis, T. (1989). ‘‘Molecular Cloning:
of reticuloendotheliosis virus transformed cell line target cells byA Laboratory Manual,’’ 2nd ed., Cold Spring Harbor Laboratories,
spleen cells. In ‘‘Avian Immunology’’ (W. T. Weber and D. L. Ewert,Cold Spring Harbor, NY.
Eds.), pp. 253–263. A. R. Liss, NY.Schat, K. A. (1987). Marek’s disease: A model for protection against
Witmer, L. A., Rosenthal, K. L., Graham, F. L., Friedman, H. M., Yee, A.,herpesvirus-induced tumours. Cancer Sur. 6, 1–37. and Johnson, D. C. (1990). Cytotoxic T lymphocytes specific for her-
Schat, K. A., and Calnek, B. W. (1978). Characterization of an apparently pes simplex virus (HSV) studied using adenovirus vectors expressingnononcogenic Marek’s disease virus. J. Natl. Cancer Inst. 60, 1073– HSV glycoproteins. J. Gen. Virol. 71, 387–396.1082. Witter, R. L., Kato, S., Calnek, B. W., and Powell, P. C. (1979). A proposed
Schat, K. A., and Calnek, B. W. (1980). In vitro cytotoxicity of spleen method for designating avian cell lines and transplantable tumors.lymphocytes against Marek’s disease tumor cells: induction by SB- Avian Pathol. 8, 487–498.1, an apparently nononcogenic Marek’s disease virus. In ‘‘Resistance Witter, R. L., Sharma, J. M., Lee, L. F., H. Optiz, H. M., and Henry, C. W.and Immunity to Marek’s Disease’’ (P. M. Biggs, Ed.), pp. 301–319. (1984). Field trials to test the efficacy of polyvalent Marek’s diseaseEEC Publication, Luxembourg. vaccines in broilers. Avian Dis. 28, 44–60.
Schat, K. A., Calnek, B. W., and Fabricant, J. (1982a). Characterization Yanagida, N., Ogawa, R., Lee, L. F., and Nazerian, K. (1992). Recombi-of two highly oncogenic strains of Marek’s disease virus. Avian Pa- nant fowlpox viruses expressing the glycoprotein B homology andthol. 11, 593–605. the pp38 gene of Marek’s disease virus. J. Virol. 66, 1402–1408.
Schat, K. A., and Heller, E. D. (1985). A chromium-release assay for Yoshida, S., Lee, L. F., Yanagida, N., and Nazerian, K. (1994). Identifica-the study of cell mediated immune responses to Marek’s disease tion and characterization of a Marek’s disease virus gene homolo-antigens. In ‘‘Proceedings of the International Symposium on Marek’s gous to glycoprotein L of herpes simplex virus. Virology 204, 414–Disease’’ (B. W. Calnek and J. L. Spencer, Eds.), pp. 301–316. Am. 419.Assoc. Avian Pathol., Kennett Square, PA. Zelnik, V. (1995). Marek’s disease and new approaches to its control.
Schat, K. A., Pratt, W. D., Morgan, R., Weinstock, D., and Calnek, B. W. Acta Virologica 39, 53–63.(1992). Stable transfection of reticuloendotheliosis virus-transformed Zivny, J., Kurane, I., Leporati, A. M., Ibe, M., Takiguchi, M., Zeng, L. L.,lymphoblastoid cell lines. Avian Dis. 36, 432–439. Brinton, M. A., and Ennis, F. A. (1995). A single nine-amino acid
Schat, K. A., Shek, W. R., Calnek, B. W., and Abplanalp, H. (1982b). peptide induces virus-specific, CD8/ human cytotoxic T lymphocyteSyngeneic and allogeneic cell-mediated cytotoxicity against Marek’s clones of heterogenous serotype specificities. J. Exp. Med. 182, 853–
863.disease lymphoblastoid tumor cell lines. Int. J. Cancer. 29, 187–194.
AID VY 8055 / 6a1c$$$$44 07-08-96 11:11:25 viras AP: Virology



















