
Analysis of the cytotoxicity of synthetic antimicrobial peptides on mouse leucocytes: implications...
10
Journal of Antimicrobial Chemotherapy (2002) 50, 339–348 DOI: 10.1093/jac/dkf141 339 ................................................................................................................................................................................................................................................................... © 2002 The British Society for Antimicrobial Chemotherapy Analysis of the cytotoxicity of synthetic antimicrobial peptides on mouse leucocytes: implications for systemic use Sabrina Pacor 1 *, Anna Giangaspero 2 , Marina Bacac 1 , Gianni Sava 1 and Alessandro Tossi 2 Departments of 1 Biomedical Sciences and 2 Biochemistry, Biophysics and Macromolecular Chemistry, University of Trieste, Trieste I-34127, Italy Received 12 February 2002; returned 5 May 2002; revised 28 May 2002; accepted 10 June 2002 We have analysed the toxicity of highly cationic, artificial α-helical antimicrobial peptides on blood cells to assess their suitability for systemic application. Flow cytometric methods, based on the uptake of propidium iodide, were used to obtain a rapid and quantitative estimate of membrane damage to resting and concanavalin A-activated mouse lymphocytes, which was further confirmed by morphological changes as observed by scanning electron microscopy. Membrane permeabilization appeared to correlate with structural characteristics, so that the peptide L-19(9/B), which contains helix-stabilizing aminoisobutyric acid (Aib) residues and is a potent antimicrobial, was also the most lytic towards both mouse lymphocytes and human erythro- cytes. Reducing the propensity for helix formation in P19(8) resulted in a marked reduction in in vitro cytotoxicity. Changing the helical sense in D-P19(9/B) also resulted in a significant decrease in cytolytic activity towards both erythrocytes and leucocytes. A limited assessment in BALB/c mice confirmed a lower in vivo toxicity of P19(8) than L-P19(9/B). In a study of the systemic antimycotic activity of P19(8) in a mouse protection model, a modest prolongation in survival of Candida albicans-infected animals after intravenous administration was observed at 5 mg/kg peptide but not at higher doses. The implications of these observations for the systemic use of this type of peptide are discussed. Introduction Gene-encoded, ribosomally synthesized antimicrobial pep- tides (AMPs) are an important component of the innate immune response of eukaryotic organisms ranging from plants to invertebrate and vertebrate animals. 1–5 Many AMPs act directly by permeabilizing the membrane of microbial targets, with a mode of action that does not appear to depend on specific receptors and which is thus not affected by the resistance mechanisms shown by a growing number of patho- gens towards conventional antibiotics. 3,6,7 Some endogenous AMPs have also been found to stimulate cell-mediated immu- nity by recruiting leucocytes. 8–10 A considerable effort has been made to develop some classes of AMPs as pharma- ceutical agents, mostly for the treatment of external local infections such as oral mucositis associated with chemo- therapy, diabetic ulcers, chronic lung infections associated with cystic fibrosis, and ocular and bucco-dental infections, but none have as yet reached clinical use. 11–14 Although many AMPs have potent and broad-spectrum antimicrobial activity in vitro, their usefulness depends on various other factors. As they are generally cationic and amphipathic molecules, they can bind to host components such as extracellular proteins, lipoproteins, anionic constituents of cellular surfaces and of the extracellular matrix, and host cellular membranes them- selves, which reduces bioavailability. 1,15 Moreover, some of these peptides show harmful side effects, such as lysis of red blood cells at active antimicrobial concentrations. 16–18 Thus, although there are some indications that AMPs could also be used to treat systemic infections, a more detailed understand- ing of how they could affect host cells is called for. One potentially useful systemic application for AMPs concerns fungal infections, due to the increased incidence of mycoses particularly in immuno-depressed individuals. 19 As .................................................................................................................................................................................................................................................................. *Corresponding author. Tel: +39-040-5583529; Fax: +39-40-577435; E-mail: [email protected] by guest on January 7, 2015 http://jac.oxfordjournals.org/ Downloaded from
Transcript of Analysis of the cytotoxicity of synthetic antimicrobial peptides on mouse leucocytes: implications...

Journal of Antimicrobial Chemotherapy (2002) 50, 339–348DOI: 10.1093/jac/dkf141
339. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . .
© 2002 The British Society for Antimicrobial Chemotherapy
Analysis of the cytotoxicity of synthetic antimicrobial peptides on mouse leucocytes: implications for systemic use
Sabrina Pacor1*, Anna Giangaspero2, Marina Bacac1, Gianni Sava1 and Alessandro Tossi2
Departments of 1Biomedical Sciences and 2Biochemistry, Biophysics and Macromolecular Chemistry, University of Trieste, Trieste I-34127, Italy
Received 12 February 2002; returned 5 May 2002; revised 28 May 2002; accepted 10 June 2002
We have analysed the toxicity of highly cationic, artificial α-helical antimicrobial peptides onblood cells to assess their suitability for systemic application. Flow cytometric methods, based onthe uptake of propidium iodide, were used to obtain a rapid and quantitative estimate of membranedamage to resting and concanavalin A-activated mouse lymphocytes, which was furtherconfirmed by morphological changes as observed by scanning electron microscopy. Membranepermeabilization appeared to correlate with structural characteristics, so that the peptideL-19(9/B), which contains helix-stabilizing aminoisobutyric acid (Aib) residues and is a potentantimicrobial, was also the most lytic towards both mouse lymphocytes and human erythro-cytes. Reducing the propensity for helix formation in P19(8) resulted in a marked reduction in invitro cytotoxicity. Changing the helical sense in D-P19(9/B) also resulted in a significantdecrease in cytolytic activity towards both erythrocytes and leucocytes. A limited assessment inBALB/c mice confirmed a lower in vivo toxicity of P19(8) than L-P19(9/B). In a study of thesystemic antimycotic activity of P19(8) in a mouse protection model, a modest prolongation insurvival of Candida albicans-infected animals after intravenous administration was observed at5 mg/kg peptide but not at higher doses. The implications of these observations for the systemicuse of this type of peptide are discussed.
Introduction
Gene-encoded, ribosomally synthesized antimicrobial pep-tides (AMPs) are an important component of the innateimmune response of eukaryotic organisms ranging fromplants to invertebrate and vertebrate animals.1–5 Many AMPsact directly by permeabilizing the membrane of microbialtargets, with a mode of action that does not appear to dependon specific receptors and which is thus not affected by theresistance mechanisms shown by a growing number of patho-gens towards conventional antibiotics.3,6,7 Some endogenousAMPs have also been found to stimulate cell-mediated immu-nity by recruiting leucocytes.8–10 A considerable effort hasbeen made to develop some classes of AMPs as pharma-ceutical agents, mostly for the treatment of external localinfections such as oral mucositis associated with chemo-therapy, diabetic ulcers, chronic lung infections associated
with cystic fibrosis, and ocular and bucco-dental infections,but none have as yet reached clinical use.11–14 Although manyAMPs have potent and broad-spectrum antimicrobial activityin vitro, their usefulness depends on various other factors. Asthey are generally cationic and amphipathic molecules, theycan bind to host components such as extracellular proteins,lipoproteins, anionic constituents of cellular surfaces and ofthe extracellular matrix, and host cellular membranes them-selves, which reduces bioavailability.1,15 Moreover, some ofthese peptides show harmful side effects, such as lysis of redblood cells at active antimicrobial concentrations.16–18 Thus,although there are some indications that AMPs could also beused to treat systemic infections, a more detailed understand-ing of how they could affect host cells is called for.
One potentially useful systemic application for AMPsconcerns fungal infections, due to the increased incidence ofmycoses particularly in immuno-depressed individuals.19 As
. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . .
*Corresponding author. Tel: +39-040-5583529; Fax: +39-40-577435; E-mail: [email protected]
by guest on January 7, 2015http://jac.oxfordjournals.org/
Dow
nloaded from

S. Pacor et al.
340
ethical reasons restrict animal employment in the develop-ment of in vivo models of efficacy or toxicity, the preliminaryuse of in vitro methods based on the host cells likely to beencountered by the drug candidate, such as, for example,circulating leucocytes in the case of intravenous (iv) use, isdesirable. We have studied the cytotoxic activity of threeartificial α-helical AMPs on mouse primary cultures of rest-ing or activated lymphocytes. These compounds weredesigned as part of a detailed structure–activity relationshipstudy of the effect of physicochemical characteristics on thein vitro antimicrobial activity, specificity and selectivity ofthis type of small linear AMP.3,20 Peptide- and concentration-dependent membrane damage was quantitatively determinedusing flow cytometric methods and confirmed by electronmicroscopy. These data indicated the most appropriate pep-tide and dose likely to provide active protection from an infect-ing pathogen while minimizing the toxic effect on circulatinghost cells. The effectiveness of this peptide in vivo wasassessed in a limited mouse candidiasis protection study.
Materials and methods
Peptide design, synthesis and characterization
The peptides P19(8), D-P19(9/B) and L-P19(9/B) weredesigned using a ‘sequence template’ obtained by analysingnumerous naturally occurring α-helical peptides, as describedpreviously.20 Solid-phase syntheses were carried out on anautomated synthesizer (Pioneer PerSeptive Biosystems) usingPEG-PS resins (0.17–0.22 mEq/g) and a six-fold excess of1:1 Fmoc-amino acid/TBTU [where TBTU corresponds toO-(benzotriazol-1-yl)-1,1,3,3-tetramethyluronium tetrafluoro-borate] with di-isopropylethyl amine as base, as described.Peptides were purified by preparative reversed-phase (RP)HPLC [Waters (Milford, MA, USA) RadialPak, 5 µm, 300 Å,10 × 150–300 mm], and their correct structure and purity weredetermined by mass determination with an API I ion-sprayinstrument (Applied Biosystems SCIEX). Peptide concentra-tions were quantified using tyrosine absorption (ε280 = 1290M–1·cm–1). Circular dichroism (CD) spectra were obtained ona Jasco J-700 dichrometer using 2 mm path-length cuvettesand peptide concentrations of 4 × 10–5 M, in 5 mM phosphatebuffer pH 7, in the presence of 50% trifluoroethanol or 10 mMSDS, and the percentage helicity was determined as ([θ]meas –[θ]rc)/([θ]α – [θ]rc), where [θ]meas is the measured ellipticity at222 nm, [θ]rc is the ellipticity for unstructured peptide in theabsence of additives (generally close to zero) and [θ]α is theellipticity of a fully structured helix of length n, calculatedusing the relation [θ]α = 39 000 (1–4/n).21 The presence ofaminoisobutyric acid (Aib), which is achiral, was taken intoaccount in these calculations.22 Stock solutions of peptidewere obtained by resuspending lyophilized peptide in phos-phate-buffered saline (PBS) and diluting as appropriate inPBS for all experiments.
Haemolytic activity
Lysis of erythrocyte membranes was determined by monitor-ing the release of haemoglobin at 415 nm from 0.5% humanerythrocyte suspensions (0.2 mL) in PBS, in relation tocomplete (100%) haemolysis as determined by addition of0.2% Triton X-100.
Preparation of lymphocyte single-cell suspensions
Lymphocytes were obtained from spleens of healthy mice.The organ was placed on a sterile Petri dish with 5 mL ofRPMI–5% fetal calf serum (FCS) and pressed with the barrelof a plastic syringe until complete tissue disaggregation wasachieved. The cell suspension was filtered through a sterilegauze to remove debris, and centrifuged at 400g for 5 minat 4°C. Red blood cells were removed by hypotonic shock,and following a further centrifugation, lymphocytes werere-suspended with PBS, counted by the Trypan Blue exclu-sion test and diluted to the appropriate concentration withPBS or complete medium [CM: RPMI 1640 (Sigma ChemicalCo.), 10% fetal bovine serum (FBS; Euroclone), 1% penicillin/streptomycin 100× (Euroclone)] for the in vitro tests.
T-cell lymphoblasts were obtained from spleen cells,prepared as described above and dispersed at 1 × 106/mL in10 mL of CM in upright 50 mL culture flasks. After additionof 2.5 µg/mL concanavalin A (ConA), flasks were incubatedfor 72 h, followed by two washes with PBS, and activatedcells were recovered and counted by the Trypan Blue exclu-sion test.23 Lymphoblasts were identified by cell cycle,phenotypic analysis and mitochondrial respiration (JC probe,5,5′,6,6′-tetrachloro-1,1′,3,3′-tetraethylbenzimidazolyl-car-bocyanine iodide; Molecular Probes, Eugene, OR, USA).
Flow cytometric analysis
Suspensions of 106 lymphocytes in 1 mL of PBS wereexposed to the desired peptide (4–160 µM) for 30 or 60 min at37°C. Bulk phase peptide was then removed by centrifugationat 400g for 5 min at 4°C followed by double washing withPBS. Samples were prepared at least in triplicate.
Propidium iodide uptake
At the end of the incubation time washed cells were dilutedwith PBS and added to 10 µL of propidium iodide (PI) solu-tion (0.5 mg/mL in PBS) immediately prior to flow cytometricanalysis. Fluorescence data from damaged cells showingpermeability to PI were acquired in a monoparametric histo-gram (emission wavelength = 612 nm).
Dual staining FITC/PI
Cells (5 × 105) were fixed in 70% EtOH for at least 4 h, thenwashed twice and allowed to balance in PBS for 2 h. Pellets
by guest on January 7, 2015http://jac.oxfordjournals.org/
Dow
nloaded from

Cytoxicity of peptides on leucocytes
341
were resuspended in a 1 mL PBS solution containing 10 µg ofPI, 0.05 µg of fluorescein isothiocyanate (FITC) and 4 µg/mLRNase (all from Sigma Chemical Co.). Cells were stainedovernight before flow cytometric analysis.
Monoclonal antibody staining
Samples of lymphocyte suspensions obtained from 24 h invitro cultures were washed, resuspended with PBS–NaN3–bovine serum albumin (BSA) and stained with fluorescentmonoclonal antibodies (MoAb): CD3-FITC (1 µg/106 cells)and CD19-PE (phycoerythrine) (0.5 µg/106 cells) (SouthernBiotecnology Associates Inc., Birmingham, AL, USA) for30 min, and unbound antibodies removed by two washes.Samples treated with an irrelevant isotype matched and FITC/PE MoAb, run in parallel, were used to set the gates in themonoparametric histogram to include <2% aspecific fluores-cence events. All flow cytometric analyses were performedwith an XL instrument (Coulter Inst., Miami, FL, USA). Atleast 10000 events were acquired for each sample. Histo-grams were analysed with the WinMDI software (Dr J. Trot-ter, Scripps Research Institute, La Jolla, CA, USA).
Scanning electron micrographs
Lymphocyte suspensions, after challenge with the test pep-tides, as described above, were re-suspended in PBS anddiluted to 20 × 106 cells/mL. A drop of each cell suspensionwas layered on to slides coated previously with poly-L-lysinesolution (0.1 mg/mL, Sigma), and allowed to adhere for 2 h.Cells were then fixed with 2.5% glutaraldehyde in 0.1 Mcacodylate buffer overnight. All procedures were performedat 4°C. Samples were dehydrated in graded ethanol, vacuumdried and mounted on to aluminium scanning electron micro-graph (SEM) mounts. After sputter coating with gold, theywere submitted for analysis with a Leica Stereoscan 430i in-strument.
Animal studies
Animal studies were carried out according to the DHHSGuide for the care and use of laboratory animals.24 Evaluationof the in vivo toxicity was performed by a single iv injection ofthe test peptides at doses of 12.5, 25 and 50 mg/kg. Systemiccandidiasis was induced in healthy BALB/c female mice(Animal House, University of Trieste) by injection into a tailvein of 5 × 105 Candida albicans (H12 strain, ATCC56879)per mouse. This procedure leads to a lethal infection within 10days in the control group.25 Groups of 8–10 animals weretreated with the test peptide at each dose, or with PBS alone(control). For the lower dose (5 mg/kg) and control theexperiment was repeated twice. Peptide was administered as asingle dose, and was injected intravenously 6 h after the
induction of systemic candidiasis. Animal survival wasrecorded on a daily basis.
Statistical analysis
Cytometric data were submitted to computer-assistedanalysis (Instat II; Graphpad Software, version 2.05) usingthe Student/Newmann–Keuls test for analysis of variance(ANOVA), whereas animal survival data were subjected tothe non-parametric Kaplan–Meier test.
Results
In Table 1 we summarize the physicochemical characteristicsand haemolytic activities of three artificial amphipathic,α-helical peptides. L-P19(9/B) and D-P19(9/B) have the samesequence, but the first is composed of L-amino acids while thesecond is composed of D-amino acids. They were designed tohave a high charge (+9), a reasonable amphipathicity (relativehydrophobic moment of 0.6) and include three Aib residues tofavour helix formation. Although they respectively form aright-handed and left-handed helix in the presence of trifluoro-ethanol (TFE) or SDS micelles (Figure 1), they show the samedegree of structuring (∼60% estimated helix content). P19(8)also has a high net charge (8+) but a lower tendency to struc-ture (the helical content is ∼40% in 50% TFE or SDS micelles;see Figure 1 and Table 1), due to the absence of Aib residuesand the presence of glycine residues at positions 7 and 14,respectively, replacing an Aib and a norleucine (Nle) residue.None the less, it is slightly more hydrophobic than P19(9/B),as the other Aib residues are replaced by Nle.
To evaluate the effect of these AMPs on circulating bloodcells, we determined the haemolytic activity at 10 and 100 µM(Table 1) by quantifying haemoglobin release and damage tomouse leucocytes from a spleen primary cell culture, and inparticular B- and T-lymphocytes, after exposure to peptides atconcentrations ranging from 4 to 160 µM, using flow cyto-metric methods (Tables 2–4). In an initial study, membranepermeabilization was shown by PI uptake in resting cells(Figure 2 and Table 2). Both P19(8) and D-P19(9/B) showed amembrane permeabilizing effect, with the appearance of apopulation of cells characterized by stronger fluorescence(see inset to Figure 2c with respect to that in a) and by asmaller size than that of untreated cells (lower forwardscattering population in Figure 2b and c), although exposureto P19(8) resulted in a more limited effect than D-P19(9/B)under the same conditions (Figure 2 and Table 2).
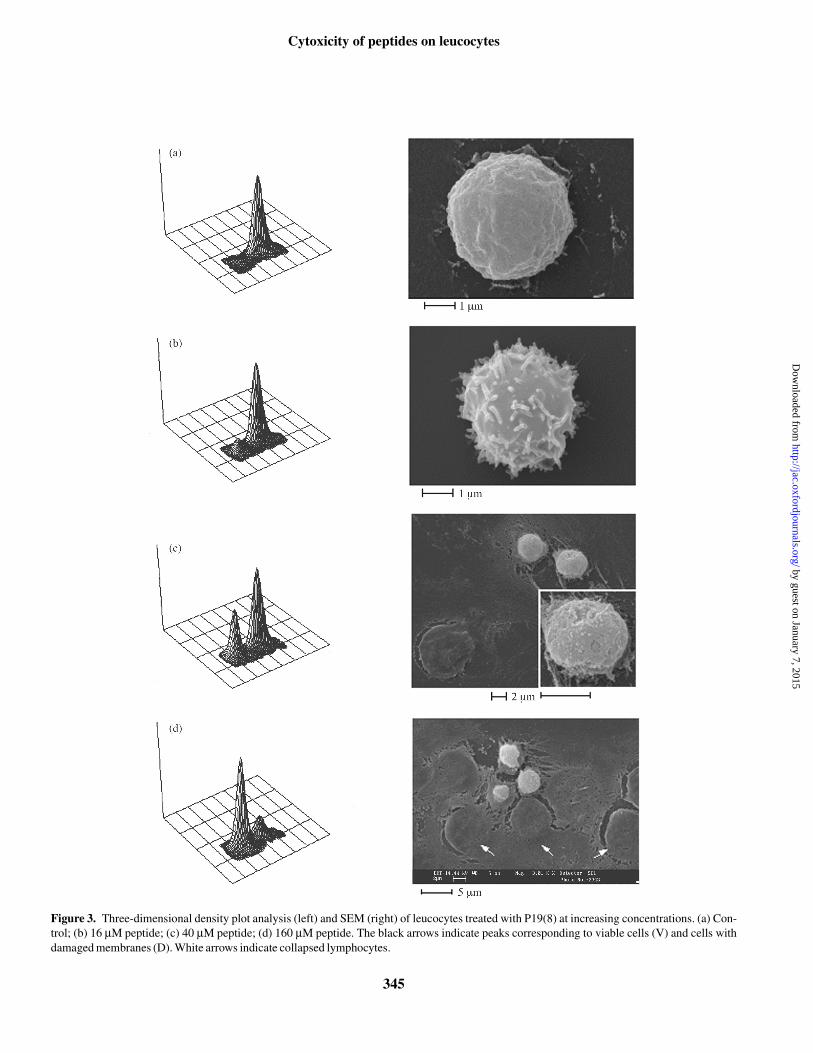
The morphological changes observed by flow cytometrycorrelate with those observed by SEM, in cells treated withP19(8) under the same conditions (Figure 3). Untreatedleucocytes have a homogeneous morphology and undamagedmembranes, as indicated by a single peak in the 3D densityplot (Figure 3a, left panel), a relatively smooth surface in
by guest on January 7, 2015http://jac.oxfordjournals.org/
Dow
nloaded from

S. Pacor et al.
342
SEM images (Figure 3a, right panel) and a low PI uptake (insetto Figure 2a). Exposure to higher concentrations of the pep-tide resulted in a second peak appearing in the 3D densityplots (Figure 3b–d, left panels), corresponding to a cell popu-lation with a smaller size (decreased forward scattering, FS)and increased fluorescence, indicative of membrane damage.After exposure to 16 µM peptide for 30 min, the majority ofcells still belong to the ‘viable’ cell population, with unalteredsize and low fluorescence, although SEM images show that asignificant alteration of the cell surface occurs, with extensiveblebbing. The smaller sized population was instead signifi-cant after exposure to 40 µM peptide and dominant at 160 µMpeptide, the highest dose used (Figure 3c and d, left panels).SEM of analogously treated cells indicated the appearanceof some completely collapsed cells at 40 µM, alongsideapparently integral cells (Figure 3c, right panel). At 160 µMpeptide most of the cells observed by SEM were collapsed,having lost any three-dimensional structure (Figure 3d, rightpanel), although a small proportion of cells with a less affectedmorphology were also present. The proportion of collapsed tointegral cells at this concentration estimated by SEM is inqualitative agreement with the number of ‘viable’ cellsobtained by flow cytometric estimation of PI exclusion(∼20%, Table 2).
A quantitative analysis of the effect of exposure to P19(8)and D-P19(9/B) on the percentage of viable cells, based on theFS signal and PI exclusion capabilities, is reported in Table 2.Damage to lymphocytes does not appear to be statisticallysignificant up to 16 µM peptide for P19(8). A moderate cyto-toxicity was instead observed at 40 µM, while most cells weredamaged at 160 µM. These effects were evident after 30 minchallenge and did not vary significantly after longer exposure.D-P19(9/B) was more cytotoxic at equivalent concentrations,and this effect was time dependent, as a substantial increase indamaged cells was observed after 1 h, even at the lowest con-
Tab
le 1
.S
eque
nces
and
pro
pert
ies
of a
rtif
icia
l pep
tide
s
a Est
imat
ed ta
king
into
acc
ount
the
pres
ence
of
thre
e ac
hira
l Aib
(B
) re
sidu
es.22
b G, G
ly; Y
, Tyr
; Z, N
le; Q
, Gln
; E, G
lu; O
, orn
ithin
e; B
, Aib
, D-a
min
o ac
ids
are
indi
cate
d in
low
er c
ase,
B r
esid
ues
are
achi
ral.
c Mea
n pe
r re
sidu
e hy
drop
hobi
city
and
rel
ativ
e am
phip
athi
citi
es.20
% α
-Hel
ixa
Hae
mol
ysis
(% ru
ptur
ed c
ells
)M
IC fo
r C
. alb
ican
s H12
Pept
ide
Seq
uenc
ebC
harg
e H
c[ µ
H]r
elc
50%
TFE
10 µ
M S
DS
10 µ
M p
eptid
e10
0 µM
pep
tide
µ
M p
eptid
e
P19(
8)GZZQOZGOOZOOZGOOZGY
8+–0
.90.
6842
37<
513
8–16
L-P
19(9
/B)
GZZOOZBOOBOOBZOOZGY
9+–1
.10.
5860
5830
702–
4D
-P19
(9/B
)GzzoozBooBooBzoozGy
9+–1
.10.
5860
5915
404
Table 2. Percentage of PI-excluding cells following in vitro treatment with peptide for 30 or 60 min
aMean ± S.E.M. for two experiments carried out in triplicate (n = 6), expressedas % viable cells with respect to untreated controls (%T/C).bStatistically different from control, P < 0.05 (ANOVA, Student/Newman–Keuls test).cStatistically different from control, P < 0.01.dStatistically different from control, P < 0.001.nd, not determined.
4 µM 16 µM 40 µM 160 µM
30 minP19(8) 100.5 ± 3.3a 92.6 ± 2.3 75.1 ± 5.3b 23.9 ± 2.4c
D-P19(9/B) 90.0 ± 1.0 81.3 ± 1.2b 61.4 ± 2.1b nd60 min
P19(8) 92.5 ± 3.0 89.8 ± 1.8 75.4 ± 4.1b 15.9 ± 1.0d
D-P19(9/B) 76.2 ± 5.9b 58.4 ± 0.5b 34.7 ± 1.4b nd
by guest on January 7, 2015http://jac.oxfordjournals.org/
Dow
nloaded from

Cytoxicity of peptides on leucocytes
343
centration used (4 µM). These results parallel the haemolyticactivity of the peptides (Table 1), indicating that D-P19(9/B)is generally more cytotoxic than P19(8).
In order to distinguish the cytotoxic effects on B- andT-lymphocytes (which we estimate to represent jointly ∼80%of the splenocyte suspension23), fluorescently labelled anti-CD19 and anti-CD3 monoclonal antibodies were used todetermine the respective percentages in recovered viablecells, 24 h after treatment with P19(8) for 30–60 min (Table
3). These cells thus resisted permeabilization in the first hourof exposure to peptide. At concentrations of up to 40 µM, thispeptide did not affect the relative proportion of undamaged B-and T-lymphocytes (roughly 1:1), as compared with anuntreated control, whereas at 160 µM, the proportion ofB-lymphocytes was significantly reduced (∼30% of the lym-phocyte population). Undamaged cells did not appear to showa different distribution among cell cycle phases from theuntreated control (based on the fluorescence characteristic of
Figure 1. CD spectra of L-P19(9/B), D-P19(9/B) and P19(8) in the presence of 10 mM SDS. The inset shows the degree of helicity (% helix content)for L-P19(9/B) and P19(8) at increasing proportions of TFE.
Figure 2. Contour plots of lymphocytes treated with antimicrobial peptides. (a) Control; (b) P19(8) at 16 µM; (c) D-P19(9/B) at 16 µM. The insetsshow the fluorescence monoparametric histograms for regions highlighted in grey, which correspond, respectively, to viable cells in (a), anddamaged, PI-permeable cells in (c). FS, forward scattering; SS, side scattering.
by guest on January 7, 2015http://jac.oxfordjournals.org/
Dow
nloaded from

S. Pacor et al.
344
cells, as assessed by PI DNA staining after ethanol per-meabilization, Table 3), independently of the challenge doseor time of exposure. Furthermore, 24 h after in vitro treatmentwith P19(8), undamaged cells had a similar proportion ofviable cells to sub-G1 cells (see the % G0/G1 cells, Table 3),and a similar protein content in viable cells to that in thecontrol (as assessed by FITC fluorescence, Table 3). Thatexposure to the peptide did not induce apoptosis was also sug-gested by the fact that the respiratory activity of mitochondriain surviving lymphocytes treated with 16 µM P19(8) for 60min was not significantly different from untreated cells asdetermined using the JC-1 probe (data not shown).
During systemic use of peptides, they are likely to encoun-ter immune cells activated by the presence of an infectiveagent. As damage to the cell membrane appears to be animportant aspect of their cytotoxic activity, this could beaffected by changes in immune cell membrane properties thatoccur after stimulation.26–28 In vitro activation of splenocyteswas effected by treatment with ConA, and the resultingT-lymphoblasts were characterized by their S-phase activa-tion, their phenotypes (CD3, CD4 and CD44) and their mito-chondrial respiration as assessed using the JC1 probe. Table 4reports the effect of exposure to P19(8), L-P19(9/B) andD-P19(9/B), for 30 or 60 min. Again, P19(8) causes the leastdamage, significantly reducing the percentage of viable cellsonly at 40 µM, in a time-independent manner. Results aresimilar to those obtained with resting cells (Table 2), suggest-ing that its cytotoxicity does not depend on the state of cellactivation. D-P19(9/B) confirmed a greater and time-depend-ent toxicity, which was more marked for activated than forresting cells. L-P19(9/B) was the most cytotoxic for ConA-activated cells; the effect at 4 µM being comparable to that ofD-P19(9/B) at 16 µM and to that of P19(8) at 40 µM. Theseresults, combined with the haemolytic activity, confirm thatthe cytotoxicity increases in the order P19(8) < D-P19(9/B)< L-P19(9/B).
In order to relate the in vitro cytotoxicity data with in vivoeffects, a limited number of toxicity experiments were per-formed on a BALB/c mouse model, administering the leastand most cytotoxic peptides, P19(8) and L-P19(9/B), at dosesvarying from 12.5 to 50 mg/kg, corresponding to estimatedblood concentrations of 80–300 µM immediately after injec-tion (respectively about half and twice the maximal dose usedin flow cytometry experiments). These concentrations arepurely indicative, as the peptide blood level is likely to berapidly reduced by binding to cellular and plasma com-ponents. Only the higher dose was lethal in both cases, while
Table 3. Evaluation of the effect of P19(8) on the cell cycle and on total protein content by dual FITC/PI staininga on surviving cells of primary lymphocyte cultures 24 h after exposure to peptide for 30 or 60 min
aMean values ± S.E.M. from triplicate samples of CD3- or CD19-positive cells, assessed in two experiments.bPercentage viable cells in the G0/G1 phase determined by PI staining of ethanol permeabilized cells, 24 h after in vitro contact with P19(8). Most of the remainingcells (>40%) were in the sub-G1 peak.cProtein content was estimated from the mean channel of fluorescence (FITC staining) of the same cells.*Statistically different from untreated control cells (P < 0.05 , ANOVA, Student/Newman–Keuls test).
30 min 60 min
Peptide concentration (µM) CD3 (%) CD19 (%)
G0/G1 cells (%)b
protein contentc CD3 (%) CD19 (%)
G0/G1 cells (%)
protein content
0 38.0 ± 1.2 30.8 ± 1.5 47.8 ± 2.5 233.4 ± 2.016 36.1 ± 0.5 31.4 ± 0.7 48.6 ± 1.8 241.7 ± 2.1 37.2 ± 0.5 35.5 ± 0.3 42.0 ± 1.8 244.7 ± 9.7
40 40.7 ± 1.7 26.1 ± 2.7 44.5 ± 1.8 248.9 ± 1.9 40.4 ± 0.3 32.5 ± 1.5 43.2 ± 0.4 238.9 ± 2.8160 57.1 ± 1.1* 10.8 ± 0.6* 42.9 ± 1.8 234.3 ± 9.5 52.0 ± 1.2* 18.9 ± 0.7* 42.0 ± 1.3 217.6 ± 2.6
Table 4. Fraction of PI-excluding T-lymphoblasts following in vitro treatment with AMPs
aMean ± S.E.M. for two experiments carried out in triplicate of % viable cellswith respect to untreated controls (%T/C).bStatistically different from control (P < 0.01, ANOVA, Student/Newman–Keuls test).
% PI-excluding cellsa
Peptide Concentration (µM) 30 min 60 min
P19(8) 4 99.1 ± 5.9 94.2 ± 1.5P19(8) 16 95.9 ± 3.8 92.0 ± 5.5P19(8) 40 77.4 ± 5.0b 78.7 ± 3.2b
D-P19(9/B) 4 83.6 ± 1.4b 57.4 ± 12.6b
D-P19(9/B) 16 59.3 ± 2.5b 45.9 ± 1.9b
D-P19(9/B) 40 30.5 ± 2.3b 19.2 ± 1.8b
L-P19(9/B) 4 68.6 ± 4.7b 50.8 ± 5.8b
L-P19(9/B) 16 24.0 ± 1.7b 20.8 ± 0.5b
L-P19(9/B) 40 12.3 ± 0.5b 11.9 ± 1.1b
by guest on January 7, 2015http://jac.oxfordjournals.org/
Dow
nloaded from
Cytoxicity of peptides on leucocytes
345
Figure 3. Three-dimensional density plot analysis (left) and SEM (right) of leucocytes treated with P19(8) at increasing concentrations. (a) Con-trol; (b) 16 µM peptide; (c) 40 µM peptide; (d) 160 µM peptide. The black arrows indicate peaks corresponding to viable cells (V) and cells withdamaged membranes (D). White arrows indicate collapsed lymphocytes.
by guest on January 7, 2015http://jac.oxfordjournals.org/
Dow
nloaded from

S. Pacor et al.
346
all animals survived at 25 mg/kg. At the lower doses both pep-tides induced visible signs of toxicity (difficulty in breathingand movement, ruffled fur), but the time of onset of sufferingwas considerably longer and that of recovery shorter forP19(8) than for L-P19(9/B), indicating a lower toxicity forP19(8) also in vivo.
On this basis, and in consideration of the in vitro effects,P19(8) was considered to be the most suitable candidate foruse in a mouse systemic candidiasis protection model. Thesurvival analysis of mice infected with C. albicans wascarried out at a dose of 5 mg/kg peptide administered iv 6 hafter the induction of infection. This dose is 10 times lowerthan the lethal dose, and in fact no visible signs of toxicitywere observed. It corresponds to an estimated blood concen-tration of ∼30 µM immediately after administration, which istwo to four times the in vitro MIC, while corresponding to arelatively contained cytotoxic activity on circulating cells asassessed by the in vitro studies. Again, this concentration ispurely indicative for the reasons discussed above. The sur-vival analysis is shown in Figure 4, and indicates a modest butstatistically significant shift of the curve towards a longersurvival. Interestingly, higher doses of the peptide (10 and20 mg/kg) did not improve survival, but conversely decreasedit with respect to untreated mice at the higher dose.
Discussion
The systematic variation of several physicochemical para-meters in α-helical AMPs, designed using a rationally derivedsequence template,3,20,29,30 has led to the development of pep-tides such as L-P19(9/B), which has potent and broad-spectrum antimicrobial activity. This results from a combina-
tion of optimized characteristics, including high charge andamphipathicity, and a stabilized α-helical structure, butunfortunately is also accompanied by a relatively highhaemolytic activity (Table 1). Modulation of helical structur-ing in P19(8) resulted in a strong reduction of haemolysis,albeit accompanied by a moderate reduction in antimicrobialpotency. A more drastic inhibition of helix formation,obtained by insertion of proline or D-amino acids, severelylimits antimicrobial activity as well.20 Another type of struc-tural modification, formation of a left-handed helix in an all-D
analogue of P19(9/B), also somewhat reduced haemolysis(Table 1), but in this case without significantly affecting anti-microbial activity.20
We have determined how these structural variations affectthe cytotoxicity of the peptides on other types of blood cells,in a murine model, with a view to evaluating their potential forsystemic applications, such as, for example, in the treatmentof systemic candidiasis. Flow cytometry is a rapid and power-ful method for the in vitro evaluation of cytotoxicity, as analy-sis of PI fluorescence allows for an accurate quantification ofdamage to circulating cells by cytolytic AMPs, and variousmolecular tools are available to distinguish this effect in dif-ferent cell types. PI uptake shows that permeabilization of thecytoplasmic membrane occurs with all three peptides tested(Tables 2 and 4), and the variation of morphological para-meters (reduction of both forward and side scattering; Figure2) would seem to indicate that this damage results in necrosis.Membrane damage is also supported by the visible morpho-logical alterations to the cellular surface observed by SEM,such as bleb formation and eventually cellular collapse (Fig-ure 3). There is, however, both a quantitative and a qualitativedifference in damage caused by the three peptides under con-sideration. (i) The proportion of damaged cells is significantlylower for P19(8) than the other two peptides; (ii) damage toresting and activated lymphocytes is comparable after expo-sure to P19(8) but greater for activated cells after exposure tothe other two peptides (Tables 2 and 4), at equivalent concen-trations; (iii) essentially no variation for the cytotoxic effectwas observed after 30 or 60 min exposure to P19(8), in con-trast to the two P19(9/B) enantiomers, where damage was sig-nificantly greater at the longer time (Table 4); (iv) the D and Lenantiomers of P19(9/B) appear to have an appreciably differ-ent effect on host cells (both erythrocytes and lymphocytes).This suggests that the mechanism of eukaryotic membranepermeabilization by the three α-helical AMPs may differsomewhat, in a manner that depends principally on the pro-pensity for helical structuring, which is lower in P19(8) thanin the P19(9/B) enantiomers, while charge, hydrophobicityand amphipathicity are substantially similar. Furthermore, italso appears to depend on the stereochemistry of the peptide.This feature of α-helical AMPs has not to our knowledge beenreported previously, and could be useful, as unlike modula-
Figure 4. Survival plot according to the Kaplan–Meier test, for micetreated iv with P19(8) 6 h after inoculation with 5 × 105 C. albicans cells.Results are based on 18 mice for the treated group and 15 mice for thecontrol group. P = 0.044 according to the Mantel–Heaenzel log-ranktest.
by guest on January 7, 2015http://jac.oxfordjournals.org/
Dow
nloaded from

Cytoxicity of peptides on leucocytes
347
tion of helix-forming propensity, changing the helical sensehas no effect on antimicrobial potency.3
The fact that P19(8) was less cytotoxic in all in vitro experi-ments, and did not result in increased damage to cells at thelonger exposure time, even at the highest concentration used,suggested that it might be the best candidate for in vivostudies, despite its somewhat lower antimicrobial potency.Notably, even at the highest concentration used (160 µM), asignificant proportion (up to 20%) of exposed cells survivedchallenge by this peptide, as observed both by flow cyto-metric analysis and SEM. This could mean that a proportionof the leucocyte population may have membranes lesssusceptible to damage by the peptide, related either to thecell type or the phase of the cell cycle. In this respect, T-lymphocytes appear to be more resistant to exposure to highconcentrations of P19(8) than B-lymphocytes, whereas thereappears to be little difference in its effect on resting or ConA-activated T-lymphocytes.
Conversely, ConA-activated lymphocytes were moresusceptible than resting cells to the more cytotoxic peptideD-19(9/B), which is reminiscent of the different cytotoxicitydisplayed by the endogenously produced human α-helicalpeptide LL-37 on resting blood cells and the proliferatingMOLT T-lymphocyte cell line.17 Another example of dif-ferential toxicity has been reported for the endogenouslyproduced α-helical bovine myeloid antimicrobial peptides(BMAP) with respect to T-lymphocytes from healthy indivi-duals and those from patients with different types of leukaemia,which were considerably more susceptible.31 Furthermore, thisstudy indicated that at low concentrations (6 µM) the BMAPpeptides induce apoptosis in activated lymphocytes. In ourstudy, after exposure to P19(8) no apparent effects were ob-served on surviving washed cells relative to unexposed cells,after 24 h (as judged by DNA and protein content), furtherindicating that under our conditions inactivation is probablyby an early necrotic effect rather than a subsequent apoptoticinduction.
On the basis of the in vitro data, a limited in vivo study wascarried out to test the toxicity and antimycotic efficacy ofP19(8) in a mouse model. The peptide appeared to be rela-tively well tolerated iv up to a single dose of 12.5 mg/kg. Wejudged that for a 5 mg/kg peptide administration, the maximalserum concentration immediately after injection (∼30 µM)might be sufficiently high for measurable antimycotic activ-ity, without resulting in marked depletion of host immunecells, even taking into account the likely rapid reduction inbioavailability of the peptide due to binding to cells andplasma components. Treatment 6 h after an infection with5 × 105 C. albicans cells did result in a modest but statisticallysignificant increase in survival, indicating that the peptidedid in fact have a protective effect, reducing the infectingcharge of the yeast cells but not sufficiently to save treated
animals. On the other hand, at higher non-lethal doses (10 and20 mg/kg), survival was, respectively, unaffected and actu-ally decreased with respect to untreated animals. This couldbe an indication that damage to leucocytes by the peptide atthese higher doses may counteract a greater antimicrobialeffect by decreasing the animals’ defence capacities.
An in vivo study on a mouse aspergillosis model has beenreported for the potent tryptophan-rich antimicrobial peptideindolicidin, which appears to be considerably more toxic thanP19(8). It was ineffective at prolonging the survival time at adose equivalent to that at which P19(8) shows some effect.16
Liposomal entrapment of this peptide, however, considerablydecreased its cytotoxicity and allowed administration ofdoses up to 40 mg/kg with a marked increase in efficacy thatresulted in 30% long-term survival. This may also be a usefulway of mitigating the cytotoxicity of α-helical AMPs.
From our study, we conclude that the in vitro cytotoxicityof the linear α-helical AMPs tested towards blood cellsdepends principally on structural characteristics. Modulatingthe structure to reduce cytotoxicity has allowed for a marginalwindow of success with respect to the in vivo use of the lesstoxic peptide in the treatment of systemic candidiasis in amouse model. While this window might be extended byfurther rationally modulating structural characteristics of thepeptides so as to boost the antimycotic activity withoutincreasing cytotoxicity, any serious consideration of theirsystemic use would also require improvement of pharmaco-logical properties by other means, such as, for example,appropriate formulation as described for indolicidin above. Ifthis is achieved, a potentially useful application of AMPs atlow concentrations could be in conjunction with clinicallyused antibiotics, especially against pathogens that havebecome resistant to them. Some studies have in fact shownthat synergy of action can exist between clinically used anti-biotics/antimycotics and AMPs.31
Acknowledgements
We are grateful to Dr D. Romeo for stimulating this work andcritically reading the manuscript. We thank Mr F. Micali foradvice with SEM sample preparation. The free availability ofthe flow cytometry facility of the Fondazione CallerioONLUS is also gratefully acknowledged. This research wasin part supported by Grants from the Italian Ministry of theUniversities and Scientific Research (PRIN MM05265243)and the EU PANAD project QLRT-2000-00411. M. Bacac issupported by a grant from the Fondazione C&D CallerioONLUS, Trieste, Italy.
References
1. Andreu, A. & Rivas, L. (1998). Animal antimicrobial peptides, anoverview. Biopolymers 47, 415–33.
by guest on January 7, 2015http://jac.oxfordjournals.org/
Dow
nloaded from

S. Pacor et al.
348
2. Hancock, R. E. & Scott, M. G. (2000). The role of antimicrobialpeptides in animal defenses. Proceedings of the National Academyof Sciences, USA 97, 8856–61.
3. Tossi, A., Sandri, L. & Giangaspero, A. (2000). Amphipathic,alpha-helical antimicrobial peptides. Biopolymers 55, 4–30.
4. Tossi, A. & Sandri, L. (2002). Molecular diversity in gene-encoded, cationic antimicrobial polypeptides. Current Pharma-ceutical Design 8, 743–61.
5. Zasloff, M. (2002). Antimicrobial peptides of multicellular organ-isms. Nature 415, 389–95.
6. Matsuzaki, K. (1999). Why and how are peptide–lipid interactionsutilized for self-defense? Magainins and tachyplesins as arche-types. Biochimica et Biophysica Acta 1462, 1–10.
7. Shai, Y. (1999). Mechanism of the binding, insertion anddestabilization of phospholipid bilayer membranes by alpha-helicalantimicrobial and cell non-selective membrane-lytic peptides. Bio-chimica et Biophysica Acta 1462, 55–70.
8. De, Y., Chen, Q., Schmidt, A. P., Anderson, G. M., Wang, J. M.,Wooters, J. et al. (2000). LL-37, the neutrophil granule- and epi-thelial cell-derived cathelicidin, utilizes formyl peptide receptor-like 1(FPRL1) as a receptor to chemoattract human peripheral bloodneutrophils, monocytes, and T cells. Journal of Experimental Medi-cine 192, 1069–74.
9. Yang, D., Chertov, O. & Oppenheim, J. J. (2001). The role ofmammalian antimicrobial peptides and proteins in awakening ofinnate host defenses and adaptive immunity. Cellular and MolecularLife Sciences 58, 978–89.
10. Nizet, V., Ohtake, T., Lauth, X., Trowbridge, J., Rudisill, J.,Dorschner, R. A. et al. (2001). Innate antimicrobial peptide protectsthe skin from invasive bacterial infection. Nature 414, 454–7.
11. Chen, J., Falla, T. J., Liu, H., Hurst, M. A., Fujii, C. A., Mosca,D. A. et al. (2000). Development of protegrins for the treatment andprevention of oral mucositis, stucture-activity relationships of syn-thetic protegrin analogues. Biopolymers 55, 88–98.
12. Hancock, R. E. W. (2000). Cationic antimicrobial peptides,towards clinical applications. Expert Opinion on Investigational Drugs9, 1723–9.
13. Lamb, H. M. & Wiseman, L. R. (1998). Pexiganan acetate.Drugs 56, 1047–52.
14. Paquette, D. W., Waters, G. S. & Stefanidou, V. L. (1997).Inhibition of experimental gingivitis in beagle dogs with topicalsalivary histatins. Journal of Clinical Periodontology 24, 216–22.
15. Sorensen, O., Bratt, T., Johnsen, A. H., Madsen, M. T. & Borre-gard, N. (1999). The human antibacterial cathelicidin, hCAP-18, isbound to lipoproteins in plasma. Journal of Biological Chemistry274, 22445–51.
16. Ahmad, I., Perkins, W. R., Lupan, D. M., Selsted, M. E. &Janoff, S. A. (1995). Liposomal entrapment of the neutrophil-
derived peptide indolicidin endows it with in vivo antifungal activity.Biochimica et Biophysica Acta 1237, 109–14.
17. Johanson, J., Gudmundsson, G. H., Rottenberg, M. E., Berndt,K. D. & Agerbeth, B. (1998). Conformation dependent antibacterialactivity of the naturally occurring human peptide LL-37. Journal ofBiological Chemistry 237, 3718–24.
18. Skerlavaj, B., Benincasa, M., Risso, A., Zanetti, M. & Gennaro,R. (1999). SMAP-29, a potent antibacterial and antifungal peptidefrom sheep leukocytes. FEBS Letters 463, 58–62.
19. Fromtling, R. A. (1998). Human mycoses and current antifungaltherapy. Drug News Perspectives 11, 185–91.
20. Giangaspero, A., Sandri, L. & Tossi, A. (2001). Amphipathicα-helical antimicrobial peptides, a systematic study of the effects ofstructural and physical properties on biological activity. EuropeanJournal of Biochemistry 268, 5589–600.
21. Chen, Y. H., Yang, J. T. & Chau, K. H. (1974). Determination ofthe helix and beta form of proteins in aqueous solution by circulardichroism. Biochemistry 13, 3350–9.
22. Gobbo, M., Bioni, L., Filira, F., Formaggio, F., Crisma, M.,Rocchi, R. et al. (1998). Helix induction potential of N-terminalα-methyl, α-amino acids. Letters in Peptide Science 5, 105–7.
23. Klaus, G. B. (1987). Lymphocytes. A Practical Approach. IRLPress, Oxford, UK.
24. Department of Health and Human Services. (1985). Guide forthe Care and Use of Laboratory Animals. pp. 23–86. DHHS Publica-tions, National Institutes of Health, Bethesda, MD, USA.
25. Polak, A. (1998). Experimental models in antifungal chemo-therapy. Mycoses 41, 1–30.
26. Lewis, R. S. & Cahalan, M. D. (1995). Potassium and calciumchannels in lymphocytes. Annual Reviews in Immunology 13, 623–53.
27. MacDonald, R. & Nabholz, M. (1986). T-cell activation. AnnualReviews in Cell Biology 2, 231–53.
28. Tossi, A., Tarantino, C. & Romeo, D. (1997). Design of syn-thetic antimicrobial peptides based on sequence analogy andamphipathicity. European Journal of Biochemistry 250, 549–58.
29. Tiozzo, E., Rocco, G., Tossi, A. & Romeo, D. (1998). Wide-spectrum antibiotic activity of synthetic amphipatic peptides. Bio-chemical and Biophysical Research Communications 249, 202–6.
30. Risso, A., Zanetti, M. & Gennaro, R. (1998). Cytotoxicity andapoptosis mediated by two peptides of innate immunity. CellularImmunology 189, 107–15.
31. van’t Hof, W., Reijnders, I. M., Helmerhorst, E. J., Walgreen-Weterings, E., Simoons-Smit, I. M., Veerman, E. C. et al. (2000).Synergistic effects of low doses of histatin 5 and its analogues onamphotericin B anti-mycotic activity. Antonie Van Leewenhoek 78,163–9.
by guest on January 7, 2015http://jac.oxfordjournals.org/
Dow
nloaded from




















