
Survey of genome organization and gene content of Corynebacterium pseudotuberculosis
Upload
filoloskibgCategory
view
0download
0
0 INSTITUT PASTEUFYELSEVIER Paris 1997
Res. Microbial. 1997, 148, 45-54
Analysis of heterogeneity of Corynebacterium diphtheriae toxin gene, tax, and its regulatory element, d&R,
by direct sequencing
H. Nakao (I), 1.K Mazurova (2), T. Glushkevich c3) and T. Popovic (l) (*)
“’ Division of Bacterial and Mycotic Diseases, National Center for Infectious Diseases, Centers for Disease Control
and Prevention, Public Health Service, U.S. Department of Health and Human Services, MS C02, C&on Rd., Atlanta, Georgia (USA),
(2) Diphtheria Reference Laboratory, Gabrichevsky Institute of Epidemiology and Microbiology, Moscow (Russia), and
f3’ Ukrainian Center of the National Sanitary and Epidemiological Surveillance,
Kiev (Ukraine)
SUMMARY
The largest diphtheria outbreak in the developed world since the 1960s is in progress in the Russian Federation. Seventy-two Corynebacferium diphtheriae strains from throughout Russia and the Ukraine, selected for temporal and geo- graphic diversity, and 6 reference and control strains were assayed by DNA direct sequencing, and DNA sequences of their diphtheria toxin gene, fox, and the regula- tory &x/7 gene, were compared to those of the Park-Williams no. 8 strain (PW8). Twenty-eight C. diphtheriae strains had entire fox sequences identical to that of the PW8 strain. Among the remaining 48 strains which were toxigenic, 4 point muta- tions were detected in the tax gene, one within the A and three within the B subunit gene. All four were silent mutations, indicating that diphtheria toxin is highly con- served at the amino acid sequence level; therefore, changes in the efficacy of the current vaccines would be unlikely to occur. Within the open reading frame of the regulatory dfxR gene, 35 point mutations were detected. Only 15 strains had entire dtxR sequences identical to that of the PW8 strain. Nine amino acid substitutions were found in the carboxyl half of &XI?: 22 and 25 strains differed from the PW8 strain in one and two amino acids, respectively. Given that naturally occurring vari- ations of dtxR might be associated with increased diphtheria toxin production, stud- ies to investigate the association of these point mutations and amino acid substitu- tions with quantified toxin production in the strains causing the current epidemic are under way.
Key-words: Diphteria, Toxin, Corynebacterbn diphtheriae; DNA sequencing, fox, &xl?, Mutations, Russia, Ukraine.
Submitted March 9, 1996, accepted June 3, 1996.
(*) Corresponding author.

46 H. NAKAO ET AL.
INTRODUCTION
Diphtheria has re-emerged in the New Inde- pendent States (NIS) of the former Soviet Union since 1990. In 1994, 47,802 cases and 1,746 deaths from diphtheria were reported, and the diphtheria incidence was still increasing in 1995 (Centers for Disease Control and Prevention, 1995). The reasons for the resurgence of the diph- theria epidemic in the NIS are not fully under- stood. Low vaccine coverage among infants and children, waning of vaccine-induced immunity in adults and increased population movement are considered the most important factors (Galazka et al., 1995), but biologic factors such as variations in the diphtheria toxin and diphtheria toxin gene, tox, must be thoroughly considered.
Diphtheria toxin is the primary virulence fac- tor of Corynebacterium diphtheriae. Diphtheria toxin is a 535-residue exotoxin, and its active form consists of two polypeptide chains linked by a disulphide bond. Its amino-terminal frag- ment (A fragment; 193 residues) contains the catalytic domain (C domain) with the ADP-ribo- syltransferase activity ; its carboxyl-terminal fragment (B fragment; 342 residues) contains the receptor-binding domain (R domain) and the transmembrane domain (T domain) (Choe et al., 1992; Silverman et al., 1994). The structural gene, tax, is carried by a family of corynebacte- riophages. The expression of tox is under the control of a chromosomal iron-dependent repressor, d&R, and a promoter/operator region. The tox promoter contains transcription and initiation sequences similar to Escherichia coli sigma 70 promoters (Greenfield, 1992). The repressor d&R gene encodes a 226-residue pro- tein with slight homology to the E. coli Fur pro- tein (25% amino acid homology) (Boyd ei ul., 1990). This protein consists of at least three domains : a transition metal-ion activation domain, a DNA-binding domain and a pro- tein/protein interaction domain (Choe et ul., 1992; Wang et al., 1994).
Recently, by using polymerase chain reac- tion/single strand conformation polymorphism (PCR-SSCP), heterogeneity of tox and dtxR was found among different strains of C. diphtheriae, isolated mostly in Russia and the Ukraine during the current diphtheria epidemic (Nakao et al., 1996). Twelve sets of primers that amplified 12 overlapping regions were constructed (8 for tox and 4 for dtxR). A single SSCP pattern was detected within 5 of 12 analysed regions, i.e., all analysed strains had an SSCP pattern identical to that of the C. diphtheriae Park-Williams no. 8 (PW8) strain that was used as a control strain. However, from 2 to 4 different SSCP patterns were detected in regions 3 (A subunit), 6 and 8 (B subunit) of the tox gene and in all four regions of the dtxR gene. In this study, we employed automated DNA sequencing of PCR-generated amplicons to determine the DNA sequences of those 7 variable regions of the ton gene and the dtxR gene to establish the extent of observed het- erogeneity and to evaluate the correlation of SSCP patterns with specific DNA sequences.
MATERIALS AND METHODS
strains
Seventy-nine C. diphtheriae strains were included in this study: 48 were from Russia, 24 were from the Ukraine and 6 were control and reference strains (table I). PW8 (ATCC 13812) was used as a control for DNA sequencing. C. diphtheriae NCTC 10648 (tax’) and NCTC 10356 (tax-) were used as PCR- positive and -negative controls, respectively. Four reference C. diphtheriae strains were provided by the Central Public Health Laboratory, London, UK: 1084 @ox-). 1085, 1086 and 1087 (fox’) as defined by the Elek assay. All strains from Russia and the Ukraine were identified and biotyped by using stan- dard microbiological procedures as described previ- ously (Efstratiou and Maple, 1994) ; 53 were of the gravis and 19 of the mitis biotype. All strains were assayed by the Elek test using the polyclonal horse serum (2,000 III/ml, Swiss Serum and Vaccine
NIS = New Independent States. ORF = open reading frame. PCR = polymerase chain reaction.
PW8 = C. diphrhriae Park-Williams no. 8. SSCP = single strand conformation polymorphism.

HETEROGENEITY OF C. DIPHTHERIAE tox AND dtxR 47
Table I. Origin and designations of 72 C. diphtheriue isolates from Russia and the Ukraine.
Ukraine 1990-1994 1985-1989
Russia 1990-1994
Strain 880, 881, 882, 883, 711,713,714, 716, 483,484, 485, 486, 489, 490, 496, designations 884,909,910, 911, 718, 719, 721, 722, 765, 855, 856, 857(*), G4168,
912, 913, 914, 915, 723(*), 724, 726, 748(*), G4169, G4170, G4172, G4174, 916(‘), 917(*), 918, 749, 752, 753(*), 756(*), G4175, G4177, G4179, G4183, 919, 920, 921, 922, G4204 G4185, G4186, G4189, G4190, 923, 924, 925, 926, G4197, G4199, G4205, G4207, 927 G4208, G4209(*), G42 11
(*) Non-toxigenic as defined by the Elek assay.
Institute, Bern, Switzerland) as described earlier (Efstratiou and Maple, 1994) and 68 of 78 strains were detected as being toxigenic. Prior to DNA sequencing, all strains were stored in 5% defibri- nated sheep blood at -70°C.
Direct DNA sequencing
Three primer pairs were used for amplification of the three variable rox regions (tax regions 3, 6 and 8), and all four dtxR regions : Dipht 3F (GTCAAAGTGACGTATC-CAGGA)-Dipht 3R (CACGGGTTTCAAAATTAATCTC) ; Dipht 6F (ATACTTCCTGGTATCGGTAGC)-Dipht 8R (TCTACCTGTGCATACTATAGC); and DtxR IF (GGGACTACAACGCAACAAGAA)-DtxR 4R (ATCTAATTTGCCGCCTTTAGT) (fig. 1). The PCR conditions used to obtain amplicons for sequencing were described previously (Nakao et al., 1996). The amplified fragments were electrophor- esed for 2 h at 100 V by using a 1.2 % agarose gel (FMC BioProducts, Maine, USA). The gels were soaked for 5 min in 20 mM Tris-HCl pH 7.0 and 1 mM EDTA, and stained by adding l-2 drops of 10 mg/ml ethidium bromide solution for an addi- tional 5 min. The amplicons were visualized under ultraviolet light. The amplicons were recovered from the gel by using “Qiaex Gel Extraction Kit” (Qiagen GmbH, Hilden, Germany), according to the manufacturer’s instructions. Each purified amplicon was dissolved in 20 ~1 of l/10 x TE, and 5 fl were then directly used in the sequence reactions with a “Prism Reaction Dye Deoxy Terminator Cycle Sequencing Kit” (The Perkin-Elmer Corp., NJ, USA) according to the manufacturer’s instructions. Nucleic acid sequencing was performed on both strands of the amplified DNA. Seven sets of oligo- nucleotide primers were used in sequencing: Dipht 3F (GTCAAAGTGACGTATCCAGGA), Dipht 3R (CACGGGTTTCAAAATTAATCTC), Dipht 6F
(ATACTTCCTGGTATCGGTAGC), Dipht 6R (CGAATCTTCAACAGTGTTCCA), Dipht 8F (GCTATAGACGGTGATGTAACT), Dipht 8R (TCTACCTGTGCATACTATAGC), DtxR 1F (GGGACTACAACGCAACAAGAA), DtxR 1R (CAACGGTTTGGCTAACTGTA), DtxR 2F (CTGAGCGTCTGGAACAATCT), DtxR 2R (TTTCACGAGCCTGCGTTCAA), DtxR 3F (TGGGAACACGTTATGAGTGA), DtxR 3R (ATCCAACACGGATGTCAGCAT), DtxR 4F (CGGATCAGTTTACACAGCTC) and DtxR 4R (ATCTAATTTGCCGCCTTTAGT) (fig. 1). The products of the sequencing reactions were separated on a 6 % polyacrylamide gel in an “ABI DNA Sequencer Model 373A” (Perkin-Elmer). The sequences were aligned by using the computer pro- gram “Wisconsin Sequence Analysis Package” (Genetics Computer Group, WI, USA) and the “Sequence Navigator” (Perkin-Elmer).
Reported d&R sequences of two wild-type C. diphtheriue strains, C7(-) and 1030(-), both non- lysogenic and non-toxigenic, were used as the base for comparison with the strains tested in this study ; they have been used as control strains in numerous studies investigating dtxR and its regulatory func- tion over the past several decades (Boyd er al., 1990, 1992). C. diphtheriue 1030(-), isolated in Austria in 1953, is of biotype belfmti, and C. diph- theriue C7(-), isolated in the US in 1950, is of bio- type gravis (ATCC 27010).
RESULTS
All strains were analysed by the Elek assay ; the PW8 strain and 68 study strains were shown to be toxigenic, while the remaining 10 were non- toxigenic. Among the 10 non-toxigenic strains,

48 H. NAKAO ET AL.
A) tox
A s&nit (645 Cp) B subunit (1026bp)
B) d&R
- abs (678 w IkRlF IMUF DtxR3F DtxR4F
IdHHH
H c-l t-u+ lkRlR lkR2R lbR3R lhR4R
Fig. 1. Location of primers for F’CR and direct DNA sequencing of 79 C. diphrheriae isolates.
2 were the control and reference strains, and 8 strains were isolated from persons identified as carriers in the Ukraine (2 strains) and Russia (6 strains).
The DNA sequence data obtained spanned a 271-base segment from positions 316 to 586 (A subunit gene), a 297-base segment Gem positions 991 to 1287 (B subunit gene), a 290-base seg- ment from positions 1462 to 175 1 (B subunit gene) of the cox gene, and an 826-base segment from positions -123 to 703 of the dtxR gene.
tar gene
Twenty-eight C. diphtheriae strains had the entire tox sequence identical to that of PW8 strain. In one strain (C. diphtheriae 718), a single point mutation within the A subunit of the tox gene was detected at position 415 (T to C). Within the B subunit of the tox gene, point muta- tions were observed in 39 strains: at positions 1083 (G to A, 4 strains), 1233 (G to A, 33 strains) and 1590 (G to A, 6 strains) (table II). Three of
these four base substitutions (1083, 1233 and 1590) occurred in wobble base, and none of the four base changes in the tox gene resulted in a change in the deduced amino acid sequences. An additional point mutation was detected within the 3’ non-coding region, position 1704 (C to T, 65 strains).
d&R gene
Within the 826-bp amplified fragment of the dtxR gene, a total of 37 point mutations were detected: 35 of them were located in the open reading frame (ORF) of the d&R gene and 2 were in the 5’ non-coding region (table III). Fifteen strains had the entire dtxR sequence identical to that of PW8. Two other strains (C. diphtheriae 718 and 719) differed from the PW8 strain by having a C to T transition at position 273. From 2 to 23 different base substitutions were detected in the remaining 61 strains. In the amino-terminal half, 14 base substitutions were detected, and none resulted in the amino acid substitutions. In the carboxyl-terminal half, 9 of 23 base substitu-
HETEROGENEITY OF C. DIPHTHERIAE tox AND dtxR 49
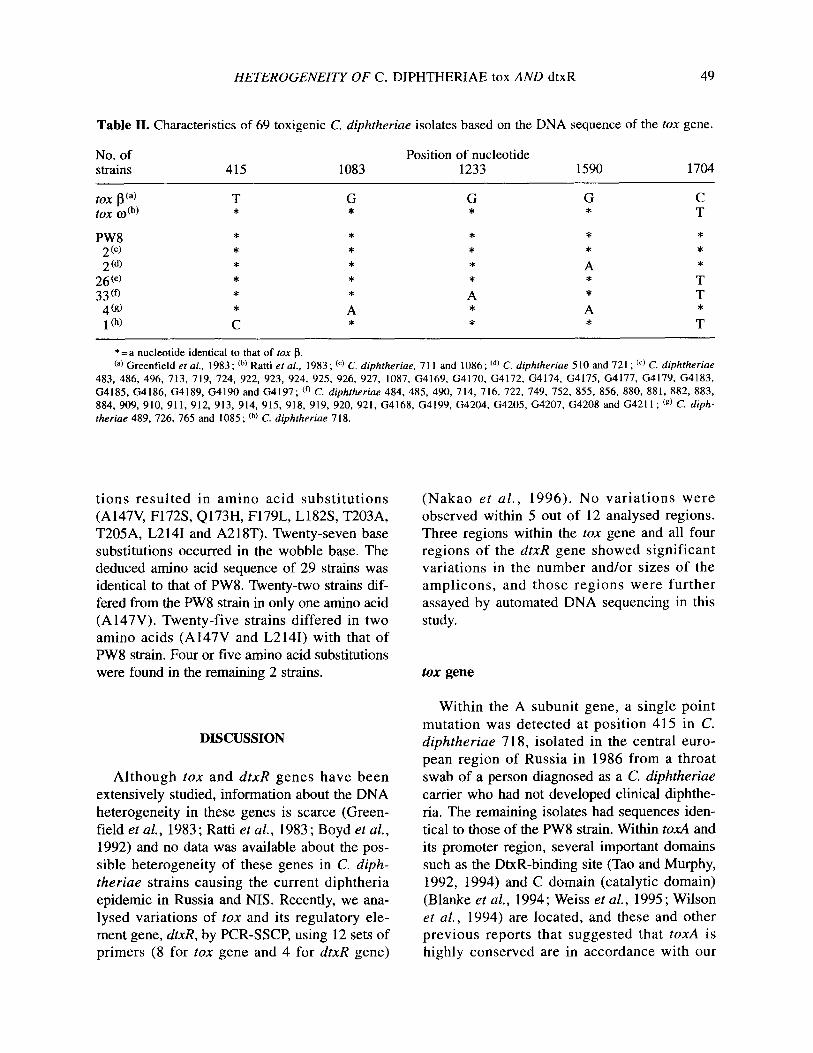
Table II. Characteristics of 69 toxigenic C. diphtheriue isolates based on the DNA sequence of the tax gene.
No. of strains 415 1083
Position of nucleotide 1233 1590 1704
tox pa) tox Kdb)
PW8 2 (4 2 Cd)
26” 33(‘,
4(9) 101)
G *
* * A * * A *
*=a nucleotide identical to that of rux p. @) Greenfield e? al., 1983 ; cb) Ratti er al., 1983 ; cc) C. diphrheriae, 7 1 1 and 1086 ; cd) C. diphfheriae 5 10 and 721 ; @) C. diphfheriae
483, 486, 496, 713, 719. 724, 922, 923, 924, 925. 926, 927, 1087, G4169. G4170, G4172, G4174, G4175, G4177, G4179, G4183, G4185, G4186, G4189, G4190 and G4197; Co C. diphfheriae 484, 485. 490, 7 14, 716, 722. 749, 752, 855, 856, 880. 881, 882, 883, 884, 909, 910. 911, 912, 913, 914, 915, 918, 919, 920. 921. G4168, G4199, G4204. G4205, G4207, G4208 and G4211; (g) C. diph- theriae 489, 726, 765 and 1085 ; ch) C. diphrherine 718.
tions resulted in amino acid substitutions (A147V, F172S, Q173H, F179L, L182S, T203A, T205A, L2 141 and A2 18T). Twenty-seven base substitutions occurred in the wobble base. The deduced amino acid sequence of 29 strains was identical to that of PW8. Twenty-two strains dif- fered from the PW8 strain in only one amino acid (A147V). Twenty-five strains differed in two amino acids (A147V and L2141) with that of PW8 strain. Four or five amino acid substitutions were found in the remaining 2 strains.
DISCUSSION
Although fox and dtxR genes have been extensively studied, information about the DNA heterogeneity in these genes is scarce (Green- field et al., 1983; Ratti et al., 1983; Boyd ef al., 1992) and no data was available about the pos- sible heterogeneity of these genes in C. diph- theriae strains causing the current diphtheria epidemic in Russia and NIS. Recently, we ana- lysed variations of tox and its regulatory ele- ment gene, dtxR, by PCR-SSCP, using 12 sets of primers (8 for fox gene and 4 for dtxR gene)
(Nakao et al., 1996). No variations were observed within 5 out of 12 analysed regions. Three regions within the fox gene and all four regions of the dtxR gene showed significant variations in the number and/or sizes of the amplicons, and those regions were further assayed by automated DNA sequencing in this study.
f&x gene
Within the A subunit gene, a single point mutation was detected at position 415 in C. diphtheriae 718, isolated in the central euro- pean region of Russia in 1986 from a throat swab of a person diagnosed as a C. diphtheriue carrier who had not developed clinical diphthe- ria. The remaining isolates had sequences iden- tical to those of the PW8 strain. Within toxA and its promoter region, several important domains such as the DtxR-binding site (Tao and Murphy, 1992, 1994) and C domain (catalytic domain) (Blanke et al., 1994 ; Weiss et al., 1995 ; Wilson et al., 1994) are located, and these and other previous reports that suggested that toxA is highly conserved are in accordance with our

50 H. NAKAO ET AL.
Table III. Characteristics of 69 toxigenic and 10 non-toxigenic C. sequence of the dtxR gene.
diphtheriae isolates, based on the DNA
No. of Position of nucleotide strains -38 -21 93 180 195 198 204 207 210 225 234 246 273 309 402 423 440 504 507
C7(-p c CTC 1030(-p * T C T
PW8 * 15(C) * 16cd) * 1 (e) *
2(f) * 21(g) *
1 (h) * 10) * 6ti) * 2 03 * 2 0)
lo(“) ; 1 (n) T
* * * * * * * * * * * * * * * * * * * * * * * * * * * * * * * * * T * * T C T
iti
* * * * * * * * * * * *
G
G A
* * * * * * * * * * * * A
c c T T
* * * * * * * * * * * * * * * * * * * * * * * * T T
T G
* * * * * * * * * * * *
G
T A T C G A
* * * * * * * * * c * * * * * * * * * * * c * *
* * ; * * * * * c * * C G A
C A
* * * * * * * * * * * * A
T C C T C G T * A T
* * * * * * * * * * * * Tt A T * * * * * * * * * * * Tt A f * * * * * A A Tt A T * * Tt A T * * Tt A T * * Tt A T * * * * * * * * * *
: C * *
C * * * * * *
C
No. of Position of nucleotide strains 515 516 519 535 544 558 564 567 579 585 600 607 613 639 640 652 654 663
C7(-)(=) T T A T C C T T C T CAACCGTT 1030(-)cb) * C * * * T * C * C G * C G T A * C
PW8 * * * * * * * * * * * * * * * * * * 15(C) * * * * * * * * * * * * * * * * * *
16cd) * c * * * * * * * * * * * * * * * * 1 (e) * * * * * * * * * * * * * * * * * * 20 * * * * * * * * * * * *
21 (g) * 1 (h)
c * * * ; 1 * ; * * * * ; Lt * ;: * * * * * * * * * * * * A * * * *
1 (1) (jti)
q c c-f c; T; * * * * * * * * * * * * * * c * * * * * * * * * * * * * * * *
204 * C***TA*T**** 2 0)
T At * C * * C***TA*T****
lo(“) T At * C *
* * * * * * * * * * * 1 (n) * * * * * T * C * C $ d; c; t it Kt * c
* = a nucleotide identical to that of C7(-) drxR; 7 = mutations that resulted in amino acid substitutions. @) Boyd et al., 1990 ; (b) Boyd et al., 1992; (‘) C. diphtheriae 483, 486, 713, 714, 716, 723, 724, 753, 756, 918, 1084, 1087, G4168.
G4170 and G4179; (d) C. diphtheriae 484, 485, 490, 749, 752, 855, G4172, G4174, G4175, G4183, G4186, G4190, G4197, G4204. G4205 and G4208 ; (‘I C. diphtkeriae 1085 ; CD C. diphtkeriae 718 and 719; (8) C. diphrheriae 880, 881, 882. 883, 884, 909, 910, 911, 912, 913, 914, 915, 919, 920, 921, 922, 923, 924. 925, 927 and G4169; ch) C. diphtheriae 721 ; (I) C. dipkrheriae 496 ; 0) C. diphrhe- riae 856, G4185, G4189, G4199. G4207 and G4211 ; ck) C. diphtkeriae 916 and 917; 0) C. diphtheriue 926 and G4177; cm) C. dip/t- rheriae 489, 510, 711. 722, 726, 748, 765, 857. 1086 and G4209 ; (“1 C. diphrkeriae 5 11.
observations. Although only 1 strain showed a single point mutation in toxA, 39 out of 68 tax+ strains showed point mutations in the ORF of toxB. From these 39 isolates, the majority, 33
isolates, had a single point mutation at position 1233 (G to A). This point mutation was more frequently detected in the gravis biotype strains (29 of 50 strains) than in the mitis biotype

HETEROGENEITY OF C. DIPHTHERIAE tox AND dtxR 51
strains (4 of 14 strains). This point mutation was also more frequently observed in the Ukrainian isolates (16 of 22 isolates) than in the Russian isolates (17 of 42 isolates). In addition, this point mutation was more frequently identified in strains isolated during the epidemic (28 of 52 strains) than in strains isolated before the epi- demic (5 of 15 strains). Another single point mutation (position 1590, from G to A) was iden- tified in two strains (C. diphtheriae 721 and 510). Two point mutations present at the same time were observed in 4 isolates (C. diphtheriae 489, 726, 765 and 1085). None of these point mutations resulted in the amino acid substitu- tion, and all of the strains, when tested by the Elek assay, were identified as toxigenic. There- fore, in spite of the observed heterogeneity at the nucleotide level, the B subunit of the tox gene that includes T domain (transmembrane domain) and R domain (receptor-binding domain) (Choe et al., 1992 ; Silverman et al., 1994) appeared to be highly conserved at the amino acid level.
Our sequencing assay also included over 70 bases downstream of the ORF of the tox gene. Two point mutations were observed at positions 1704 and 17 12. All strains tested in this study and the reported tox w sequence had C at posi- tion 1712 (Ratti et al., 1983). The DNA sequence of tox f3 has been reported only up to the nucleotide 1706 (Greenfield et al., 1983), and in this study, sequences located downstream from that position were combined with the sequence of the tox228 gene. The tox228 sequence was derived from the mutant p phage, which was isolated after i%methyl-N’-nitro-N- nitrosoguanidine mutagenesis and which encodes non-toxic cross-reaction material (CRM228) (Kaczorek et aZ., 1983). The nucleo- tide at the position 1712 is reported as G. Fifty- nine of 68 isolates had a C to T transition at posi- tion 1704, just like the PW8 strain. Finally, the sequencing data and detected point mutations in the analysed region of the tax gene clearly coin- cided with recently reported SSCP patterns of these regions (Nakao et al., 1996), suggesting that SSCP is a reliable method for detection of point mutations and could be used as a screening method prior to DNA sequencing.
d&R gene
Significantly more heterogeneity was detected within the ORF of the d&R gene. This gene is considered to consist of at least three motifs: the DNA binding motif, the metal binding motif and the protein/protein interaction motif (Wang et al., 1994). DtxR’s DNA binding ability is activated by the divalent heavy metal ions. DtxR then forms stable homodimers and inhibits transcrip- tion from the tox gene promoter and other DtxR- regulated promoters (Tao et al., 1995). A total of 35 point mutations were detected within the ORF of the dtxR gene. Eighteen strains, including PW8, differed from C. diphtheriae C7(-) at a sin- gle base, none of which resulted in the amino acid substitution. Fifteen of those 18 isolates were tox-positive when tested by the Elek assay. The remaining 3 C. diphtheriae isolates (753,756 and 1084) were non-toxigenic. Analysis of the tox gene revealed that only parts of this gene were present, resulting in the negative reaction in the Elek assay. From 2 to 12 different base substi- tutions, compared with the PW8 strain, were simultaneously detected in the remaining 60 strains; for C. diphthetiae 511, 23 different base substitutions were detected. Among these strains, base substitutions caused a subsequent amino acid substitution at 9 positions. Interestingly, all 9 amino acid substitutions were located in the car- boxyl-terminal half of the d&R gene.
Twenty-nine strains had the deduced amino acid sequence identical to that of PW8 strain. Twenty-two strains differed in only one amino acid (A147V) from the PW8 strain. Twenty-five strains differed in two amino acids (A147V, L214I) with that of PW8 strain. In spite of such base and amino acid heterogeneity, all of these strains, providing they possessed a functional tox gene, were toxigenic in the Elek assay. Studies to investigate the correlation of increased or decreased toxin production and different base or amino acid substitutions are under way.
In an earlier study, by using PCR-SSCP (Nakao et al., 1996), from 2 to 4 different SSCP patterns for four regions of dtxR were observed. A significant correlation exists between identified SSCP patterns and detected amino acid substitu-

52 H. NAkXO ET AL.
tions (table IV). In region 1, most of the strains showed the identical SSCP pattern to that of PW8 strain (SSCP pattern 1). The DNA sequence of these strains was identical to the reported d&R gene of C. diphtheriae C7(-). The strains that showed a different SSCP pattern had two C to T positions between the putative -10 region and the ribosome binding site. In region 2, 12 base substitutions that did not result in the deduced amino acid substitutions were represented by 4 different SSCP patterns. The carboxyl-terminal half had significantly more point mutations in addition to several amino acid substitutions (table IV). In~~s~gly, 21 of 24 Ukrainian iso- lates were identical and were characterized by 10 point mutations, two of which resulted in the amino acid substitutions. These data confirm the observations obtained by PCR-SSCP where a particular d&R type 5 was identified as being unique to the Ukrainian isolates.
As reported previously, C. d~~~~ue 1030(-) was found to carry 6 silent amino acid substitu-
tions in the carboxyl-terminal half of DtxR (Boyd et aZ., 1992). The amino-terminal half includes the DNA binding motif and metal binding motif (Wang et al., lQ94), but even 6 amino acid substi- tutions in the carboxyl-terminal half did not affect DtxR activity (Boyd et al., 1992). Those data are in agreement with data obtained in this study, where all amino acid substitutions had occurred in the carboxyl half. Even though all of the strains with amino acid substi~tions in regions 3 and 4 were toxigenic when analysed by the Elek assay, the effect of these substitutions on the quantity of the produced toxin remains unknown. Studies to correlate the presence of these amino acid substitutions with quantitative analysis of diphtheria toxin production are under way,
In 7 of 12 different dtxR types, a total of 9 amino acid substitutions were detected in the carboxyl half of the d&R gene. They resulted in 5 different amino acid sequences. One sequence was detected in dtxR types 5, 9 and 10 and anotherindtxRtypes2and8;dtxRtypes 1,7and
Table IV. Comparison of deduced amino acid sequences and SSCP patterns of the dtxR gene of 79 C. diphtheriae isolates.
A) d&R region 3
Position (nucleotide/amino acid) SSCP pattern @I 4401147 514-5161172 5191173 5351179 544/182
C7(-j(b) C/Ala 1030(-) Cc) C/Ala Pattern 1 C/Ala Pattern 2 T/Val Pattern 3 T/Val
m/Phe TlXYPhe ‘ITUPhe TI’Cffhe TCUSer
A/Gh A/Gin A/Gin AlGIll CMIis
Tffhe T/Phe T/Phe T/Phe CflBl
ClLeu CfLeU
c/La c/Leu T/Ser
B) dixR region 4
SSCP pattern@) ~7/203
Position (nucleotide/amino acid)
6131205 6401214 652-6541218
C7(-)@’ 1030(-)(C) Pattern 1 Pattern 2 Pattern 3 Pattern 4 G/Ala
AJSer c/Arg AlSer ABer A/Ser ClArg
CLeu T/Tyr CLeu CfLeu AAle A/Ile
GCT/Ala ACT/Thr GCT/Ala GCT/Ala GCC/Ala ACT/Thr
w Nakao et ai., 1996; cb) Boyd et al., 1990; @) Boyd et al., 1992.

HETEROGENEITY OF C. DIPHTHERIAE tox AND dtxR 53
12 had unique deduced amino acid sequences. A comparison of the predicted secondary structures of the DtxR of the wild-type strains, C. diphther- iae C7(-) and 103(-) and 5 groups of strains detected in our study, was performed by using the Peptide Structure program of the Wisconsin Sequence Analysis Package by the Genetics Computer Group, Inc. (GCG) (Jameson and Wolf, 1988). In the Chou-Fasman prediction curve, remarkable similarities were observed between C. diphtheriae C7(-) and C. diphtheriue 1030(-), with the possible exception of the region from position 168 to position 226, where some discrepancies in the form of small shifts in the extent of the helices were observed. In 3 of 5 deduced amino acid sequences (d&R type 1, dtxR types 5, 9 and 10 and dfxR types 2 and S), the predicted secondary structure of DtxR was identi- cal to that of C. diphtheriae C7(-). In d&R type 7, minimal discrepancies were detected from posi- tions 167 to 183, and in d&R type 12 fi-om posi- tion 200 to the end.
Earlier studies suggested that naturally occur- ring variants of tux and/or drxR genes might be associated with increased or decreased toxin pro- duction. Changes in the tox gene, in particular, could affect the immunogenicity of diphtheria vaccine and vaccine efficiency, and could there- fore be related to the recent re-emergence of diph- theria in Russia and the NIS. However, the DNA sequence data in this study indicate that the deduced amino acid sequences of diphtheria toxin of the analysed strains were identical to those reported to have been used for production of diphtheria toxoid. These observations suggest that decreased vaccine efficacy was not one of the primary causes of the re-emergence of diph- theria in the NIS, but that other factors such as large numbers of susceptible children and adults and low vaccination coverage may be the main reasons for this epidemic.
Another important observation from this study is that all 10 non-toxigenic strains (as determined by the Elek assay) possessed the regulatory d&R gene, which supports an earlier observation by Groman et al. (1983) that non-toxigenic strains could become toxigenic by acquiring the tox gene, assuming that their chromosomal d&R gene
is functional. Consequently, naturally occurring non-toxigenic strains with the functional dtxR gene could constitute a potential reservoir of tox- igenicity and further clarification of this issue is significant for the epidemiology of diphtheria.
Our sequence data show that the variations detected by PCR-SSCP assay correlated with the DNA sequences and that PCR-SSCP is the useful method for screening of variable regions of tax and dtxR genes. The tax gene appeared to be highly conserved, while numerous point muta- tions were detected in the dtxR gene, particularly within its carboxyl-terminal half. Studies to investigate the association of these point muta- tions and amino acid substitutions with increased or decreased toxin production are under way.
Acknowlegments
We are grateful to Dr. Robert Woblbueter, Chief, Bio- technology Core Facility Branch, CDC, for his advice and consultation regarding the predicted secondary struc- tures of DtxR.
The work is partially supported by Tbe Japan Health Science Foundation, Tokyo, Japan.
Analyse de l’hCtirogLhCiti2 du g&e de la toxine de Corynebacterium &htheriue, tax,
et de son 616ment de rhgulation, aYrR
Nous avons ttudit 72 souches de Corynebacte- rium diphtheriue isolkes en Russie et en Ukraine et 6 souches tkmoins et de rCf&ence au moyen du stquenqage ADN direct; et les sCquences de leurs g&es tax (toxine) et d&R (de rkgulation) ont CtC compar&s B celles de la souche Park-Williams no 8 (PW8). Les skquences fox de 28 C. diphtheriae sont identiques h celles de PW8. Chez 40 souches toxigknes, 4 mutations “ponctuelles” ont CtC d&e&s au niveau du g&ne fox, l’une pour la sous- unit6 A et l’autre pour la B. Ces quatre mutations montrent que la toxine diphtirique est t&s stable au niveau shuentiel des acides amin& ; c’est pourquoi des modifications de l’efficacitt vaccinale sont peu probables. Pour 1’ORF du gbne d&R, 35 points de mutation ont Ctk d&e&s. Seulement 15 souches ont des stquences d&R identiques B celles de la souche PW8. Dans la moitiC carboxyle de dtxR, 9 substitu- tions d’acides aminks ont Cte dtceltks: respective- ment 22 et 25 souches diff&nt de PW8 pour 1 ou

H. NAKAO ET AL.
2 acides amint%. &ant donnC que des variations spontankes de dtxR peuvent i%re assocites li une pro- duction accrue de toxine diphtdrique, il convient de poursuivre des etudes analysant les relations entre ces mutations ponctuelles et substitution d’acides amints et la production quantifEe de toxine par les souches responsables d’tpidtmies.
Mots-clt%: Diphtkrie, Toxine, Cotynebacterium diphtheriae ; ADN, SCquenGage ADN, tox, dtxR, Mutations, Russie, Ukraine.
References
Blanke, S.R., Huang, K., Wilson, B.A., Papini, E., Covacci, A. & Collier, R.J. (1994), Active-site muta- tions of the diphtheria toxin catalytic domain: role of histidine-21 in nicotinamide adenine dinucleotide binding and ADP-ribosylation of elongation factor 2. Biochemistry, 33,5155-5161.
Boyd, J., Oza, M.N. Br Murphy, J.R. (1990), Molecular cloning and DNA sequence analysis of a diphtheria tox iron-dependent regulatory element (d&R) from Corynebacterium diphthen’ae. Proc. Natl. Acad Sci. USA, 87, 5968-5972.
Boyd, J.M., Hall, K.C. & Murphy, J.R. (1992). DNA sequences and characterization of dtxR alleles from Corynebacterium diphtheriae PW8(-), 1030(-), and C7hm723(-). J. Bactetiol., 174, 1268-1272.
Centers for Disease Control and Prevention. (1995). Diph- theria epidemic - New Independent States of the for- mer Soviet Union, 1990-1994. Morbid Mortal. Wkly. Rep., 44, 177-181.
Choe, S., Bennett, M.J., Fujii, G., Curmi, P.M., Kantard- jieff, K.A., Collier, R.J. & Eisenberg, D. (1992), The crystal structure of diphtheria toxin. Nature (Land.), 357, 216-222.
Jameson, G.A. & Wolf, H. (1988). The antigenic index: a novel algorithm for predicting antigenic determi- nants. Computer Appl. Biosci., 4, 181-186.
Efstratiou, A. & Maple, P.A. (1994). WHO manual for the laboratory diagnosis of diphtheria. Reference no. ICP-EPI 038(C). World Health Organization, Geneva
Galazka, A.M., Robertson, S.E. & Oblapenko, G.P. (1995), Resurgence of diphtheria. Eur. J. Epidemiol., 11, 95-105.
Greenfield, L. (1992). Diphtheria toxin cloning and expression in Corynebacten’um diphtheriae. Targeted Diagn. Ther., 7, 273-305.
Greenfield, L., Bjorn, M.J., Horn, G., Fong, D., Buck, G.A., Collier, R.J. & Kaplan, D.A. (1983), Nucleo- tide sequence of the structural gene for diphtheria toxin carried by corynebacteriophage j3. Proc. Natl. Acad Sci. USA, 80, 6853-6857.
Groman, N., Cianciotto, N., Bjom, M. & Rabin, M. (1983), Detection and expression of DNA homologous to the tox gene in non-toxinogenic isolates of Corynebacte- rium diphtheriae. Infect. Imnum, 42, 48-56.
Kaczorek, M., Delpeyroux, F., Chenciner, N. & Streeck, R.E. (1983), Nucleotide sequence and expression of the diphtheria tox228 gene in Escherichia coli. Sci- ence, 221, 855-858.
Nakao, H., Pruckler, J.M., Mazurova, I.K., Narvskaia, O.V., Glushkevich, T., Marijevski, V.F., Kravetz, A.N., Fields, B.S., Wachsmuth, I.K. & Popovic, T. (19%), Heterogeneity of diphtheria toxin gene, tox, and its regulatory element, dtxR, in Corynebacterium diphtheriae strains causing epidemic diphtheria in Russia and the Ukraine. J. Clin. Microbial., 34, 171 l- 1716.
Ratti, G., Rappuoli, R. & Giannini, G. (1983), The com- plete nucleotide sequence of the gene coding for diphtheria toxin in the corynephage omega (tax’) genome. Nucleic Acids Res., 11. 6589-6595.
Silverman, J.A., Mindell, J.A., Zhan, H., Finkelstein, A. & Collier, R.J. (1994), Structure-function relationships in diphtheria toxin channels. I. Determining a mini- mal channel-forming domain. J. Membr. Biol., 137, 17-28.
Tao, X. & Murphy, J.R. (1992), Binding of the metallore- gulatory protein DtxR to the diphtheria tax operator requires a divalent heavy metal ion and protects the palindromic sequence from DNase I digestion. J. Biol. Chem., 267,21761-21764.
Tao, X. & Murphy, J.R. (1994). Determination of the min- imal essential nucleotide sequence for diphtheria tox repressor binding by in vitro affinity selection. Proc. Natl. Acad. Sci. USA, 91, 9646-9650.
Tao, X., Zeng, H.Y. & Murphy, J.R. (1995), Transition metal ion activation of DNA binding by the diphthe- ria tox repressor requires the formation of stable homodimers. Proc. Natl. Acad. Sci. USA, 92, 6803- 6807.
Wang, Z., Schmitt, M.P. & Holmes, R.K. (1994), Charac- terization of mutations that inactivate the diphtheria toxin repressor gene (&R). Infect. Immun., 62, 16CO- 1608.
Weiss, MS., Blanke, S.R., Collier, R.J. & Eisenberg, D. (1995), Structure of the isolated catalytic domain of diphtheria toxin. Biochemistry, 34, 773-78 1.
Wilson, B.A., Blanke, S.R. Reich, K.A. & Collier, R.J. (1994), Active-site mutations of diphtheria toxin - Tryptophan 50 is a major determinant of NAD aflIm- ity. J. Biol. Gem., 269, 232%-23301.
Copyright © 2022 FDOKUMEN




















