
Analysis of degradation of bacterial cell division protein FtsZ by the ATP-dependent...
10
Microbiological Research 163 (2008) 21—30 Analysis of degradation of bacterial cell division protein FtsZ by the ATP-dependent zinc-metalloprotease FtsH in vitro Ramanujam Srinivasan, Haryadi Rajeswari, Parthasarathi Ajitkumar Department of Microbiology and Cell Biology, Indian Institute of Science, Bangalore-560012, India Received 30 December 2005; received in revised form 13 February 2006; accepted 6 March 2006 KEYWORDS FtsH protease; FtsZ protein; AAA family; Bacterial cell divi- sion Summary The identity of protease(s), which would degrade bacterial cell division protein FtsZ in vivo, remains unknown. However, we had earlier demonstrated that Escherichia coli metalloprotease FtsH degrades E. coli cell division protein FtsZ in an ATP- and Zn 2+ -dependent manner in vitro. In this study, we examined FtsH protease-mediated degradation of FtsZ in vitro in detail using seven different deletion mutants of FtsZ as the substrates, which lack different extents of specific regions at the N- or C- terminus. FtsH protease assay in vitro on these mutants revealed that FtsH could degrade all the seven deletion mutants irrespective of the deletions or the extent of deletions at the N- or C-terminus. These observations indicated that neither the N- terminus nor the C-terminus was required for the degradation of FtsZ, like already known in the case of the FtsH substrate s 32 protein. The recombinant clones expressing full-length FtsZ protein and FtsZ deletion mutant proteins would be useful in investigating the possibility of FtsZ as a potential in vivo substrate for FtsH in ftsH-null cells carrying ftsH suppressor function and ectopically expressed FtsH protease. & 2006 Elsevier GmbH. All rights reserved. Introduction FtsH is a membrane-bound, ATP-dependent, Zn 2+ -metalloprotease (Tomoyasu et al., 1993a,b; Akiyama et al., 1995), which is involved in the proteolytic degradation of specific integral membrane proteins (Kihara et al., 1995, 1999; Akiyama et al., 1996a, b) and cytoplasmic proteins (Herman et al., 1993, 1995; Tomoyasu et al., 1995; Herman et al., 1997). It contains a highly conserved 200 amino acid stretch, which has Walker motifs that are characteristic of the members of the ATPases Associated with diverse cellular Activities ARTICLE IN PRESS www.elsevier.de/micres 0944-5013/$ - see front matter & 2006 Elsevier GmbH. All rights reserved. doi:10.1016/j.micres.2006.03.001 Corresponding author. E-mail address: [email protected] (P. Ajitkumar).
Transcript of Analysis of degradation of bacterial cell division protein FtsZ by the ATP-dependent...

ARTICLE IN PRESS
Microbiological Research 163 (2008) 21—30
0944-5013/$ - sdoi:10.1016/j.
�CorrespondE-mail addr
www.elsevier.de/micres
Analysis of degradation of bacterial cell divisionprotein FtsZ by the ATP-dependentzinc-metalloprotease FtsH in vitro
Ramanujam Srinivasan, Haryadi Rajeswari, Parthasarathi Ajitkumar�
Department of Microbiology and Cell Biology, Indian Institute of Science, Bangalore-560012, India
Received 30 December 2005; received in revised form 13 February 2006; accepted 6 March 2006
KEYWORDSFtsH protease;FtsZ protein;AAA family;Bacterial cell divi-sion
ee front matter & 2006micres.2006.03.001
ing author.ess: [email protected]
SummaryThe identity of protease(s), which would degrade bacterial cell division protein FtsZin vivo, remains unknown. However, we had earlier demonstrated that Escherichiacoli metalloprotease FtsH degrades E. coli cell division protein FtsZ in an ATP- andZn2+-dependent manner in vitro. In this study, we examined FtsH protease-mediateddegradation of FtsZ in vitro in detail using seven different deletion mutants of FtsZas the substrates, which lack different extents of specific regions at the N- or C-terminus. FtsH protease assay in vitro on these mutants revealed that FtsH coulddegrade all the seven deletion mutants irrespective of the deletions or the extent ofdeletions at the N- or C-terminus. These observations indicated that neither the N-terminus nor the C-terminus was required for the degradation of FtsZ, like alreadyknown in the case of the FtsH substrate s32 protein. The recombinant clonesexpressing full-length FtsZ protein and FtsZ deletion mutant proteins would beuseful in investigating the possibility of FtsZ as a potential in vivo substrate for FtsHin ftsH-null cells carrying ftsH suppressor function and ectopically expressed FtsHprotease.& 2006 Elsevier GmbH. All rights reserved.
Introduction
FtsH is a membrane-bound, ATP-dependent,Zn2+-metalloprotease (Tomoyasu et al., 1993a, b;Akiyama et al., 1995), which is involved in
Elsevier GmbH. All rights rese
t.in (P. Ajitkumar).
the proteolytic degradation of specific integralmembrane proteins (Kihara et al., 1995, 1999;Akiyama et al., 1996a, b) and cytoplasmic proteins(Herman et al., 1993, 1995; Tomoyasu et al., 1995;Herman et al., 1997). It contains a highly conserved200 amino acid stretch, which has Walker motifsthat are characteristic of the members of theATPases Associated with diverse cellular Activities
rved.

ARTICLE IN PRESS
R. Srinivasan et al.22
(AAA) family of ATPases (Beyer, 1997; Ogura andWilkinson, 2001) and a sequence called SecondRegion of Homology (SRH), together which arerequired for ATP binding and hydrolysis (Karata etal., 1999, 2001; Ogura and Wilkinson, 2001). Thesecharacteristics are found conserved in the FtsHmolecules from eubacteria, archaebacteria andsome eukaryotes (Ogura et al., 1991; Tomoyasu etal., 1993a, b; Guelin et al., 1994; Nilsson et al.,1994; Ge and Taylor, 1996; Lindahl et al., 1996;Lysenko et al., 1997). The zinc-binding motif andone of the ATP-binding motifs are essential for theproteolytic activity (Akiyama et al., 1996b; Karataet al., 1999).
FtsH protease is involved in the regulation of thelevels of phage l CII protein during phage infection(Herman et al., 1993; Shotland et al., 1997) and isthe major protease involved in the degradation ofthe heat shock transcription factor s32 (Herman etal., 1995; Tomoyasu et al., 1995). FtsH is alsoinvolved in the degradation of the uncomplexedforms of an essential protein translocation subunit,SecY, in Escherichia coli (Kihara et al., 1995;Akiyama et al., 1996b). The protease has also beenfound to degrade the subunit a of F1F0 ATPase of E.coli (Akiyama et al., 1996a). FtsH has been shownto have a regulatory role not only in stress response(Deuerling et al., 1995) but also in sporulation inBacillus subtilis and protein secretion (Deuerlinget al., 1997). The FtsH of B. subtilis was found tointerfere with the expression of SpoOA protein,which is a transcriptional regulator of the initiationof sporulation (Deuerling et al., 1997). Further, a 26amino acid long peptide, SpoVM, which is essentialfor spore formation, was found to be a substrate forFtsH (Cutting et al., 1997). This protease alsoregulates sW regulon in B. subtilis (Zellmeier et al.,2003). FtsH protease has also been shown toparticipate in membrane biogenesis by regulatingthe levels of UDP-3-O-(R-3-hydroxymyristoyl)-N-acetyl glucosamine deacetylase [the lpxC (envA)gene product] involved at the committed step inthe biosynthesis of Lipid A (Ogura et al., 1999). Allthese diverse functions of FtsH demonstrate that itis a protease essential for the viability of bacterialcells.
Septation in bacteria is initiated by FtsZ, whichgets localized to the mid-cell site early in cell cycleand remains at the leading edge of the invaginatingseptum (Bi and Lutkenhaus, 1991; Ma et al., 1996).FtsZ is the prokaryotic homologue of tubulin(Erickson, 1995, 1998; Lowe and Amos, 1998),possessing GTP binding, GTPase (de Boer et al.,1992; RayChaudhuri and Park, 1992) and GTP-dependent polymerization activities (Bramhill andThompson, 1994; Mukherjee and Lutkenhaus, 1994,
1999). E. coli FtsZ protein has been reported to bestable in vivo (Garrido et al., 1993) and theconcentration of FtsZ remains constant throughoutcell cycle in E. coli and B. subtilis (Rueda et al.,2003; Weart and Levin, 2003). In Mycobacteriumtuberculosis and M. smegmatis, the levels of FtsZprotein are found reduced by 50% in late stationaryphase and upon oxygen depletion (Dziadek et al.,2002). Similarly, in Caulobacter crescentus, thelevels of FtsZ are proteolytically regulated where itis specifically degraded in the swarmer cells duringthe late stages of cell cycle (Kelly et al., 1998). InE. coli cells, FtsZ has been found to be associatedwith ClpXP (Flynn et al., 2003). Thus, unlike in thecase of other FtsH substrates such as s32 proteinand CII, whose levels are regulated by multipleproteases including FtsH protease, the identity ofthe protease(s) involved in the proteolytic regula-tion of bacterial cell division protein FtsZ remainselusive.
By using an ftsH–gfp translational fusion con-struct it was shown that FtsH protein preferen-tially accumulates at the mid-cell site of thedividing vegetative cells of B. subtilis (Wehrl etal., 2000). These authors have alluded to a possiblerole for FtsH protease in septal biogenesis.Although FtsZ is the principal protein that me-diates septal biogenesis, it is not clear as towhether FtsH has any role in the proteolyticdegradation of the cell division protein FtsZ invivo. The role of FtsH in cell division remainingunknown, we had earlier demonstrated that FtsHprotease of E. coli degrades FtsZ protein of E. coliin an ATP- and Zn2+-dependent manner in vitro(Anilkumar et al., 2001). In the present study, inorder to find out any specificity of the degradationto either of the termini of FtsZ protein, we furtherexamined degradation of FtsZ by FtsH in vitro indetail using a battery of FtsZ deletion mutantslacking different extents of N- or C-terminus. Wereport here that FtsH protease could degradeall these deletion mutants irrespective of thedeletions and the extent of deletions at the N- orC-terminus.
Materials and methods
Bacterial strains, media and plasmidconstructs
Bacterial strains and plasmid constructs usedare listed in Table 1. E. coli strains were grown inLuria Broth (Amersham Pharmacia) at 30 1C (per-missive temperature) or at 42 1C (non-permissive

ARTICLE IN PRESS
Table 1. List of bacterial strains and plasmids used
Relevant genotype Reference or source
StrainsTYE024 A derivative of MC4100, ompT::kan/F lacIq Y. Akiyama (Akiyama et al., 1995)C43 BL21 (DE3) derivative that allows the expression of
toxic proteinsJ. E. Walker (Miroux and Walker,1996)
M15 An E. coli K12 derivative, thi� lac� ara+ lon� recA+
uvr+QIAGEN
PlasmidspQE30 Cloning vector with N-terminal 6�His tag QIAGENpSTD113 EcftsH-myc-6�His under lac promoter Y. Akiyama (Akiyama et al., 1995)pQE30-ftsZ 6�His-EcftsZ under lac promoter This studypRSET-ftsZ 6�His-EcftsZ under T7 promoter This studypRSET-ftsZDC1 6�His-EcftsZDC1 under T7 promoter This studypRSET-ftsZDC1.2 6�His-EcftsZDC1.2 under T7 promoter This studypRSET-ftsZDC2 6�His-EcftsZDC2 under T7 promoter This studypRSET-ftsZDN1 6�His-EcftsZDN1 under T7 promoter This StudypRSET-ftsZDN1DC1 6�His-EcftsZDN1DC1 under T7 promoter This studypRSET-ftsZDN1DC1.2 6�His-EcftsZDN1DC1.2 under T7 promoter This studypRSET-ftsZDN2 6�His-EcftsZDN2 under T7 promoter This studypRSET-ftsZDN2DC1 6�His-EcftsZDN2DC1 under T7 promoter This study
Analysis of degradation of bacterial cell division protein FtsZ 23
temperature). E. coli strain C43 (Miroux andWalker, 1996) was grown in Luria-Bertani broth at37 1C. E. coli ftsZ gene was amplified usingpolymerase chain reaction from the genomic DNAof E. coli K12 strain and cloned in the SmaI-digestedand calf intestinal phosphatase-treated pBluescript(KS) (Stratagene) vector. The nucleotide sequenceof the ftsZ gene in the resulting plasmid pBS-ftsZwas confirmed by DNA sequencing. pBS-ftsZ wasdigested with BamHI and PstI and ligated to pQE30vector (QIAGEN) digested with BamHI and PstI toget pQE30-ftsZ. pRSET-A-ftsZ was generated byligating the BamHI-HindIII fragment from pQE30-ftsZ at the same sites in pRSET-A. EcZ-DC1 wasobtained by digesting the pRSET-A-ftsZ with EcoRVand religation. EcZ-DN2 was obtained by ligatingthe EcoRI-HindIII fragment from pQE30-ftsZ intothe same sites in pRSET-B. EcZ-DN1 was obtained byligating the KpnI-HindIII fragment from pQE30-ftsZinto the same sites in pRSET-B. EcZ-DN2DC1 wasobtained by digesting EcZ-DN2 with EcoRV andreligation. EcZ-DN1DC1 was obtained by digestingEcZ-DN2DC1 with KpnI and religation. EcZ-DC1.2was obtained by digesting EcZ-DC1 with HindIII,end-filling with T4 DNA polymerase, digesting withEcoRV, and religation. EcZ-DN1DC1.2 was obtainedby digesting EcZ-DN1DC1 with HindIII, end-fillingwith T4 DNA polymerase, digesting with EcoRV, andreligation. Ampicillin (100 mg/ml), kanamycin(25 mg/ml), tetracycline (10 mg/ml), and chloram-phenicol (10 mg/ml) were used at the concentra-tions indicated.
Purification of recombinant full-length FtsZand its deletion mutant proteins
The full-length FtsZ was purified from M15/pREP4 cells harbouring pQE30-ftsZ, while themutant proteins were purified from E. coli C43strain (Miroux and Walker, 1996) carrying appro-priate pRSET recombinant vector. Transformants ofE. coli M15/pREP4 cells carrying pQE30-ftsZ and ofC43 strain carrying deletion mutant constructs ofFtsZ in appropriate pRSET vectors were grown at37 1C in Luria–Bertani broth to an OD600 of 0.6–0.8.Expression of the recombinant proteins was in-duced with 1mM IPTG for 3 h. The recombinantproteins were purified under denaturing conditionsusing Ni2+-NTA agarose affinity chromatographyaccording to manufacturer’s instructions. Thepurified proteins were dialysed against 10mM Tris-HCl, pH 8.0, and centrifuged at 80,000 rpm in TLA120.2 Beckman tabletop ultracentrifuge rotor toremove aggregates. The purified proteins werestored in aliquots at �70 1C until further use.
Purification of recombinant FtsH protein
Recombinant FtsH protein was purified from E.coli TYE024 cells harbouring pSTD113 plasmid(Akiyama et al., 1995). Cultures were grown in LBmedium at 37 1C to an OD600 of 1.00 and expressionwas induced with 1mM IPTG. After 3 h, the cellswere harvested and resuspended in Buffer A (20mM

ARTICLE IN PRESS
R. Srinivasan et al.24
Tris-HCl, pH 8.0, 300mM KCl, 10% glycerol, 1mMPMSF, and 10mM b-mercaptoethanol). Lysozyme wasadded at a concentration of 200mg/ml, incubated onice for 30min and lysed using sonication. The celllysate was clarified by centrifugation at 12,000 rpmin SS34 (Sorvall) rotor for 15min at 4 1C. Thesupernatant thus obtained was centrifuged at40,000 rpm for 2 h in type 50Ti rotor in Beckmanultracentrifuge at 4 1C. The pellet fraction contain-ing the membrane proteins was solubilized in BufferB (0.5% NP-40, 20mM Tris-HCl, pH 8.0, 200mM KCl,10% glycerol, 1mM PMSF, 10mM b-mercaptoethanol,and 20mM imidazole) for 1 h at 4 1C. The suspensionwas centrifuged at 40,000 rpm for 30min in type50Ti rotor to remove insoluble material. The super-natant was mixed with Ni2+-NTA agarose equili-brated with Buffer B and the suspension wasincubated for 1 h at 4 1C. The bead suspension wasloaded onto a column and the beads were washedwith 20 column volumes of Buffer B. The FtsHprotein was eluted with 200mM imidazole. Thefractions containing FtsH protein were dialysedextensively against Buffer C (10mM Tris-HCl, pH8.0, 10% glycerol, 5mM MgCl2, 1mM DTT, 0.5% NP-40, and 80mM NaCl). The protein solution wasaliquoted and stored at �70 1C until further use.
Purification of r32-6�His protein
The purification protocol for s32-6�His wasessentially as described (Gamer et al., 1992), butwith slight modifications. In brief, BB2109 cellscarrying pUEH211-1 plasmid (both kind gifts from B.Bukau, University of Heidelberg, Germany) weregrown in LB medium at 30 1C to an OD600 of 0.8 andthe expression of s32-6�His protein was inducedwith 50 mM IPTG for 30min. The cells wereharvested and suspended in phosphate-bufferedsaline, pH 7.2, and lysed by sonication. The celllysate was centrifuged at 15000 rpm in SS34 rotorfor 15min at 4 1C and the supernatant was mixedwith Ni2+-NTA agarose beads. The agarose beadswere incubated for 20min at 4 1C, washed succes-sively with 50mM Tris-HCl, pH 8.0, buffer, contain-ing 5, 10, and 20mM imidazole. s32-6�His proteinwas eluted with 100mM imidazole and the fractionscontaining the purified protein were dialysedagainst 50mM Tris-HCl, pH 8.0, containing 100mMKCl and 10% glycerol.
901 light scattering assay for FtsZpolymerization activity
The 901 light scattering assay for verifyingoligomerization or polymerization of full-length
FtsZ and of FtsZ deletion mutants were carriedout essentially as described (Mukherjee and Lut-kenhaus, 1999) in the presence GTP. The proteins(5 mM) were incubated in 1�CGH buffer (1mMcitrate, 1mM glycine, 1mM HEPES, pH 6.5), 5mMMgCl2 and 50mM KCl at 30 1C. The 901 lightscattering (LS) values were initially monitored for300 s in the absence of GTP to obtain a baseline.Subsequently, 1mM GTP was added and monitoringcontinued. Reactions were carried out in theabsence of GTP also in order to confirm GTPdependence of polymerization. Unlike the conven-tional buffers, the CGH buffer was found suitablefor recording appreciable levels of polymerizationby denatured-refolded proteins using light scatter-ing assay.
FtsH protease assay
The protease assay for FtsH was carried outessentially as described (Tomoyasu et al., 1995).The reaction mixture consisted of 50mM Tris-acetate, pH 8.0, 5mM magnesium acetate,12.5 mM zinc acetate, 1.4mM b-mercaptoethanol,20mM ATP, 100 mg/ml bovine serum albumin, and142.5 mg/ml of FtsH. The reactions were carried outunder protease limiting conditions by incubation at42 1C for 3 h. Degradation of the substrates wereanalysed by SDS-PAGE followed by staining withCoomassie Blue for detecting FtsZ deletion mutantsand s32 protein or developed using anti-polyhisti-dine antibodies in the case of full-length FtsZ.
Western Blotting
Proteins were fractionated on SDS-PAGE andtransferred to PVDF membranes. The membraneswere blocked with 5% non-fat milk powder inphosphate buffered saline containing 0.2% Tween-20. Anti-FtsZ antibodies were raised in rabbits usingpurified 6�His-FtsZ and used at 1:10,000 dilutions.Anti-polyhistidine antibodies (Sigma) and anti-FtsHantisera (Tomoyasu et al., 1993a) (a kind gift fromDr. Teru Ogura) were used at dilutions of 1:6000 and1:5000, respectively. Protein A-HRP (AmershamBiosciences) was used at 1:10,000 dilution. Detec-tion was carried out using ECL detection reagentsaccording to manufacturer’s instructions.
Results
In order to investigate whether N- or C-terminussequence region of E. coli FtsZ molecule would berequired for its degradation by FtsH protease in

ARTICLE IN PRESS
Analysis of degradation of bacterial cell division protein FtsZ 25
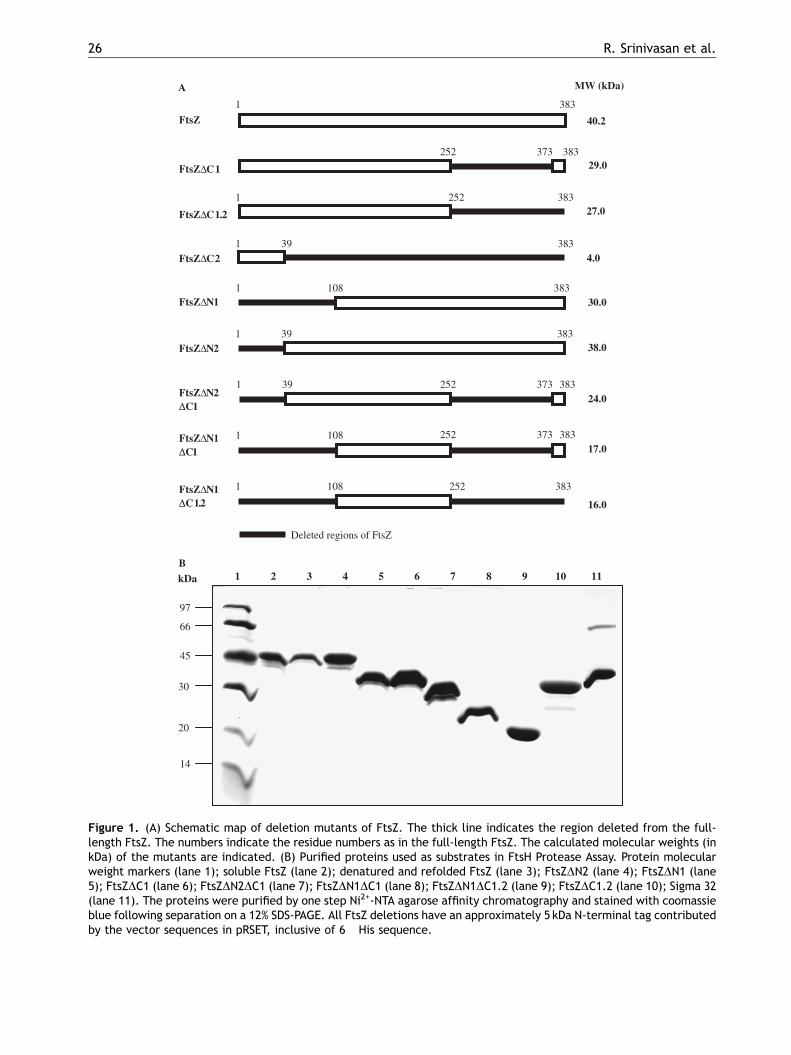
vitro, we generated eight deletion mutants of FtsZ,which lacked different extents of either N- or C-terminus or both termini, or specific segments ofthe protein (Fig. 1A). These eight FtsZ deletionmutants were used for the overexpression andpurification of the respective mutant substrateprotein for the FtsH protease assay.
Overexpression and purification of full-length FtsZ and FtsZ deletion mutants
The full-length FtsZ was overexpressed frompQE30-ftsZ construct in E. coli M15/pREP4 cells.The eight deletion mutants were independentlyoverexpressed in E. coli C43 strain (Miroux andWalker, 1996) from the respective construct.Although, several C-terminal mutants (Ma andMargolin, 1999) and few other mutants (Strickerand Erickson, 2003) of FtsZ have shown poorexpression compared to the wild-type full-lengthprotein, among the eight deletion mutants, sevenof the mutants namely, FtsZDC1, FtsZDC1.2,FtsZDN1, FtsZDN2, FtsZDN2DC1, FtsZDN1DC1, andFtsZDN1DC1.2 expressed to high yields. For reasonsnot known, the FtsZDC2 mutant did not getexpressed in spite of the correctness of the DNAclone. Therefore, this mutant could not be includedin the studies. The full-length FtsZ and the sevendeletion mutants were purified using Ni2+-NTAagarose affinity column chromatography underdenaturing conditions according to manufacturer’sinstructions and refolded by dialysis against 10mMTris-HCl buffer, pH 8.0, containing 10% glycerol(Fig. 1B).
Verification of polymerization activity of full-length FtsZ and deletion mutants
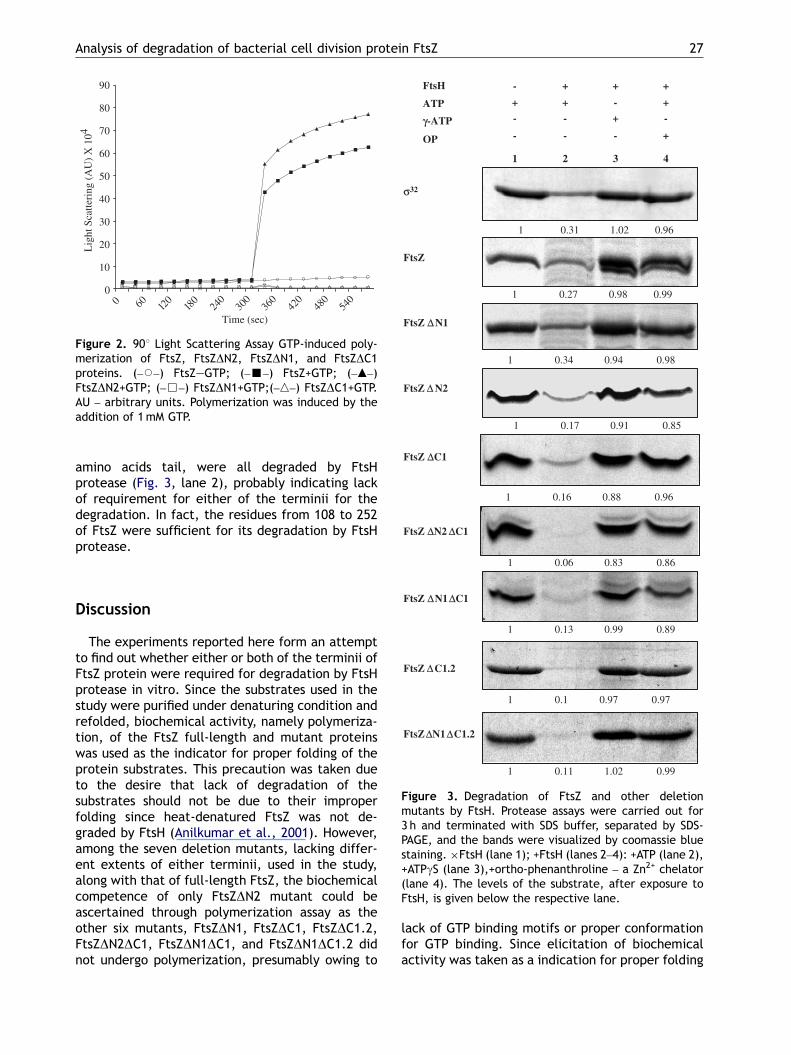
Since the full-length FtsZ protein and thedeletion mutants, which were to be subjected toprotease assay, were all purified under denaturingcondition and refolded, we wanted to ensure thatthese substrates were all properly folded andtherefore are in the native form. We took elicita-tion of biochemical activity by the proteins as theindicator for their proper folding. Therefore, inorder to ensure that the full-length FtsZ and thedeletion mutants were biochemically active, wecarried out assays for the polymerization of theproteins, which is characteristic of biochemicallyactive FtsZ molecule (Bramhill and Thompson,1994; Mukherjee and Lutkenhaus, 1994, 1999) orits mutants that can bind GTP (Gupta et al., 2004).Such a verification would not be possible for themutant proteins that do not bind GTP.
Since polymerization by FtsZ or its mutants isconditional on GTP binding, we assayed only forpolymarization and not for GTP binding per se. Inorder to monitor polymerization, we performed 901light scattering assay, which is used for FtsZpolymerization assay (Mukherjee and Lutkenhaus,1999), for full-length FtsZ and its deletion mutantsin the presence of GTP. The 901 light scatteringassays showed that full-length FtsZ and FtsZDN2mutant indeed undergo polymerization, in thepresence of GTP (Fig. 2). FtsZDN1 and FtsZDC1did not undergo polymerization (Fig. 2). Since theremaining four mutants FtsZDC1.2, FtsZDN2DC1,FtsZDN1DC1, and FtsZDN1DC1.2 were essentiallyderivatives of FtsZDC1, which did not show poly-merization, we did not assay these mutants forpolymerization. These experiments demonstratedthat the full-length FtsZ and FtsZDN2 proteins werepolymerization competent, and therefore compe-tent in GTP binding.
Degradation of FtsZ deletion mutants by FtsHin vitro
The seven deletion mutants and full-length FtsZwere tested for degradation by FtsH protease invitro. The FtsH assay system used was identical towhat has been earlier reported by us (Anilkumar etal., 2001). The conventional substrate s32 (Tomoya-su et al., 1995) was used as the positive control inthe protease assays. The full-length FtsZ and all theseven deletion mutants were efficiently degradedby FtsH protease, as in the case of the conventionalsubstrate s32 (Fig. 3, lane 2 of the respectivepanel). The degradation of full-length FtsZ, asreported earlier (Anilkumar et al., 2001), and thatof all the seven deletion mutants were ATP-dependent. The unhydrolysable analogue of ATP,ATPgS, did not support proteolysis, indicating thatATP hydrolysis was essential for the proteaseactivity (Fig. 3, lane 3 of the respective panel).Similarly, the inclusion of Zn2+ chelator, ortho-phenanthroline, inhibited proteolysis, indicatingthat the protease activity was Zn2+-dependent aswell (Fig. 3, lane 4 of the respective panel), astypical of FtsH protease activity (Tomoyasu et al.,1995). These observations demonstrate that degra-dation of full-length FtsZ and of all the sevendeletion mutants by FtsH protease proceeded withthe characteristics that are typical of classicalFtsH-mediated proteolysis in vitro (Tomoyasu etal., 1995). Thus, FtsZ deletion mutants lacking theN-terminus or the C-terminus or both the terminii(retaining only the middle portion from the residueT108 to I252), with or without the conserved 11
ARTICLE IN PRESS
MW (kDa)
40.2
Deleted regions of FtsZ
252 373
39 383
1 108
252 373
1 108 252 373
30.0
252 38327.0
17.0
1 108 252 383
16.0
4.0
FtsZ
FtsZΔC2
FtsZΔC1.2
FtsZΔN1ΔC1
FtsZΔN1 ΔC1.2
FtsZΔC1
38.0
24.0
FtsZΔN1
FtsZΔN2
383
383
383
383
383
1
391
391
1
1 383
FtsZΔN2 ΔC1
29.0
97
20
66
45
30
14
1 2 3 4 5 6 7 8 9 10 11kDa
A
B
Figure 1. (A) Schematic map of deletion mutants of FtsZ. The thick line indicates the region deleted from the full-length FtsZ. The numbers indicate the residue numbers as in the full-length FtsZ. The calculated molecular weights (inkDa) of the mutants are indicated. (B) Purified proteins used as substrates in FtsH Protease Assay. Protein molecularweight markers (lane 1); soluble FtsZ (lane 2); denatured and refolded FtsZ (lane 3); FtsZDN2 (lane 4); FtsZDN1 (lane5); FtsZDC1 (lane 6); FtsZDN2DC1 (lane 7); FtsZDN1DC1 (lane 8); FtsZDN1DC1.2 (lane 9); FtsZDC1.2 (lane 10); Sigma 32(lane 11). The proteins were purified by one step Ni2+-NTA agarose affinity chromatography and stained with coomassieblue following separation on a 12% SDS-PAGE. All FtsZ deletions have an approximately 5 kDa N-terminal tag contributedby the vector sequences in pRSET, inclusive of 6�His sequence.
R. Srinivasan et al.26
ARTICLE IN PRESS
0 60 120
180
240
300
360
420
480
540
Time (sec)
0
10
20
30
40
50
60
70
80
90L
ight
Sca
tteri
ng (
AU
) X
104
Figure 2. 901 Light Scattering Assay GTP-induced poly-merization of FtsZ, FtsZDN2, FtsZDN1, and FtsZDC1proteins. (–J–) FtsZ—GTP; (–’–) FtsZ+GTP; (–m–)FtsZDN2+GTP; (–&–) FtsZDN1+GTP;(–n–) FtsZDC1+GTP.AU – arbitrary units. Polymerization was induced by theaddition of 1mM GTP.
σ32
FtsZ
FtsZ ΔN1
FtsZ Δ N2
FtsZ ΔC1
FtsZ ΔN2 ΔC1
1 0.31 1.02 0.96
1 0.27 0.98 0.99
1 0.34 0.94 0.98
1 0.17 0.91 0.85
1 0.16 0.88 0.96
FtsH
ATP
γ-ATP
OP
3
-
-
+
+
2
-
-
+
+
4
-
+
+
+
1
-
+
-
-
Analysis of degradation of bacterial cell division protein FtsZ 27
amino acids tail, were all degraded by FtsHprotease (Fig. 3, lane 2), probably indicating lackof requirement for either of the terminii for thedegradation. In fact, the residues from 108 to 252of FtsZ were sufficient for its degradation by FtsHprotease.
FtsZ ΔN1ΔC1
FtsZ ΔC1.2
FtsZ ΔN1ΔC1.2
1 0.06 0.83 0.86
1 0.13 0.99 0.89
1 0.1 0.97 0.97
1 0.11 1.02 0.99
Figure 3. Degradation of FtsZ and other deletionmutants by FtsH. Protease assays were carried out for3 h and terminated with SDS buffer, separated by SDS-PAGE, and the bands were visualized by coomassie bluestaining. �FtsH (lane 1); +FtsH (lanes 2–4): +ATP (lane 2),+ATPgS (lane 3),+ortho-phenanthroline – a Zn2+ chelator(lane 4). The levels of the substrate, after exposure toFtsH, is given below the respective lane.
Discussion
The experiments reported here form an attemptto find out whether either or both of the terminii ofFtsZ protein were required for degradation by FtsHprotease in vitro. Since the substrates used in thestudy were purified under denaturing condition andrefolded, biochemical activity, namely polymeriza-tion, of the FtsZ full-length and mutant proteinswas used as the indicator for proper folding of theprotein substrates. This precaution was taken dueto the desire that lack of degradation of thesubstrates should not be due to their improperfolding since heat-denatured FtsZ was not de-graded by FtsH (Anilkumar et al., 2001). However,among the seven deletion mutants, lacking differ-ent extents of either terminii, used in the study,along with that of full-length FtsZ, the biochemicalcompetence of only FtsZDN2 mutant could beascertained through polymerization assay as theother six mutants, FtsZDN1, FtsZDC1, FtsZDC1.2,FtsZDN2DC1, FtsZDN1DC1, and FtsZDN1DC1.2 didnot undergo polymerization, presumably owing to
lack of GTP binding motifs or proper conformationfor GTP binding. Since elicitation of biochemicalactivity was taken as a indication for proper folding

ARTICLE IN PRESS
R. Srinivasan et al.28
of all the seven deletion mutants, which werepurified under denaturing conditions and refolding,such an assumption could not be made in the caseof six mutants, FtsZDN1, FtsZDC1, FtsZDC1.2,FtsZDN2DC1, FtsZDN1DC1, and FtsZDN1DC1.2.Therefore, in the case of such mutants, wepresumed that they were also properly folded sinceall of the mutants, which would or would notpolymerize, were all purified through denaturationand refolding under identical conditions. Thisassumption was made as there was no othermethod by which the folding of such mutants couldbe verified. Moreover, since FtsH does not degradeheat-denatured FtsZ protein (Anilkumar et al.,2001), degradation of the six mutants might beconsidered as an indication that improper foldingmight not have occurred during their purification.
The lack of requirement for either of the terminiifor degradation of FtsZ by FtsH is similar to thedegradation of the substrate s32 by FtsH. Neitherthe N-terminus nor the C-terminus was required forthe degradation of s32 by FtsH (Bertani et al., 2001;Tomoyasu et al., 2001). The deviation from suchnon-specificity with respect to the terminii of thesubstrate is found in the degradation of l phage CIIprotein and protein translocation subunit SecY.Degradation of CII protein by FtsH has suggestedan endopeptidase activity and subsequent proces-sive role for the enzyme (Shotland et al., 2000). Bycontrast, experiments to study degradation of SecYby FtsH have suggested that FtsH degrades thissubstrate in a processive manner either from the N-terminal end or the C-terminal end (Chiba et al.,2002). Although degradation of FtsZ by FtsH did notrequire either of the terminii of the protein fordegradation, we did not examine whether theproteolytic activity proceeded in an endopeptido-lytic or in a processive manner.
Finally, the physiological significance of the invitro degradation of FtsZ by FtsH remains to beinvestigated. The protease(s) that would degradeFtsZ in vivo remain hitherto unknown. Neverthe-less, the recombinant clones expressing full-lengthFtsZ protein and FtsZ deletion mutant proteinswould be useful in the investigation on thepossibility of FtsZ as a potential in vivo substratefor FtsH in ftsH-null cells carrying ftsH suppressorfunction and ectopically expressed FtsH protease.
Acknowledgements
The authors acknowledge gift of pSTD113 and E.coli TYE024 cells from Yoshinori Akiyama, Institutefor Virus Research, Kyoto University, Japan, and s32
construct pUEH211-1 from Bernd Bukau, Universityof Heidelberg, Germany. Authors thank Dr. K.Muniyappa, Department of Biochemistry, IndianInstitute of Science, Bangalore, for FluorMaxspectrofluorimeter facility, and Prabuddha Guptain our laboratory for technical help in lightscattering assays. The work was supported by aresearch grant to PA from the Department ofAtomic Energy, Government of India (2002/37/4/BRNS). The work was also supported in part by theDepartment of Biotechnology, Government of India,supported Genomics Initiative on Microbial Patho-gens – Structural Genomics Initiative in the Divisionof Biological Sciences, at Indian Institute ofScience. Authors acknowledge the infrastructureand equipment facilities supported by the FISTgrant from Department of Science and Technologyand DSA Programme from University Grants Com-mission, Government of India, and ICMR Centre forAdvanced Study in Molecular Medical Microbiology,at the Department of Microbiology and Cell Biology,Indian Institute of Science.
References
Akiyama, Y., Yoshihisa, T., Ito, K., 1995. FtsH, amembrane-bound ATPase, forms a complex in thecytoplasmic membrane of Escherichia coli. J. Biol.Chem. 270, 23485–23490.
Akiyama, Y., Kihara, A., Ito, K., 1996a. Subunit a ofproton ATPase F0 sector is a substrate of the FtsHprotease in Escherichia coli. FEBS Lett. 399, 26–28.
Akiyama, Y., Kihara, A., Tokuda, H., Ito, K., 1996b. FtsH(HflB) is an ATP-dependent protease selectively actingon SecY and some other membrane proteins. J. Biol.Chem. 271, 31196–31201.
Anilkumar, G., Srinivasan, R., Anand, S.P., Ajitkumar, P.,2001. Bacterial cell division protein FtsZ is a specificsubstrate for the AAA family protease FtsH. Micro-biology 147, 516–517.
Bertani, D., Oppenheim, A.B., Narberhaus, F., 2001. Aninternal region of the RpoH heat shock transcriptionfactor is critical for rapid degradation by the FtsHprotease. FEBS Lett. 493, 17–20.
Beyer, A., 1997. Sequence analysis of the AAA proteinfamily. Protein Sci. 6, 2043–2058.
Bi, E.F., Lutkenhaus, J., 1991. FtsZ ring structureassociated with division in Escherichia coli. Nature354, 161–164.
Bramhill, D., Thompson, C.M., 1994. GTP-dependentpolymerization of Escherichia coli FtsZ protein to formtubules. Proc. Natl. Acad. Sci. USA 91, 5813–5817.
Chiba, S., Akiyama, Y., Ito, K., 2002. Membrane proteindegradation by FtsH can be initiated from either end.J. Bacteriol. 184, 4775–4782.
Cutting, S., Anderson, M., Lysenko, E., Page, A.,Tomoyasu, T., Tatematsu, K., Tatsuta, T., Kroos, L.,

ARTICLE IN PRESS
Analysis of degradation of bacterial cell division protein FtsZ 29
Ogura, T., 1997. SpoVM, a small protein essential todevelopment in Bacillus subtilis, interacts with theATP-dependent protease FtsH. J. Bacteriol. 179,5534–5542.
de Boer, P., Crossley, R., Rothfield, L., 1992. The essentialbacterial cell-division protein FtsZ is a GTPase. Nature359, 254–256.
Deuerling, E., Paeslack, B., Schumann, W., 1995. TheftsH gene of Bacillus subtilis is transiently inducedafter osmotic and temperature upshift. J. Bacteriol.177, 4105–4112.
Deuerling, E., Mogk, A., Richter, C., Purucker, M.,Schumann, W., 1997. The ftsH gene of Bacillus subtilisis involved in major cellular processes such assporulation, stress adaptation and secretion. Mol.Microbiol. 23, 921–933.
Dziadek, J., Madiraju, M.V., Rutherford, S.A., Atkinson,M.A., Rajagopalan, M., 2002. Physiological conse-quences associated with overproduction of Mycobac-terium tuberculosis FtsZ in mycobacterial hosts.Microbiology 148, 961–971.
Erickson, H.P., 1995. FtsZ, a prokaryotic homolog oftubulin? Cell 80, 367–370.
Erickson, H.P., 1998. Atomic structures of tubulin andFtsZ. Trends Cell. Biol. 8, 133–137.
Flynn, J.M., Neher, S.B., Kim, Y.I., Sauer, R.T., Baker,T.A., 2003. Proteomic discovery of cellular substratesof the ClpXP protease reveals five classes of ClpX-recognition signals. Mol. Cell 11, 671–683.
Gamer, J., Bujard, H., Bukau, B., 1992. Physical interac-tion between heat shock proteins DnaK, DnaJ, andGrpE and the bacterial heat shock transcription factorsigma 32. Cell 69, 833–842.
Garrido, T., Sanchez, M., Palacios, P., Aldea, M., Vicente,M., 1993. Transcription of ftsZ oscillates duringthe cell cycle of Escherichia coli. EMBO J. 12,3957–3965.
Ge, Z., Taylor, D.E., 1996. Sequencing, expression, andgenetic characterization of the Helicobacter pyloriftsH gene encoding a protein homologous to membersof a novel putative ATPase family. J. Bacteriol. 178,6151–6157.
Guelin, E., Rep, M., Grivell, L.A., 1994. Sequence of theAFG3 gene encoding a new member of the FtsH/Yme1/Tma subfamily of the AAA-protein family. Yeast10, 1389–1394.
Gupta, P., Anand, S.P., Srinivasan, R., Rajeswari, H., Indi,S., Ajitkumar, P., 2004. The C-terminally truncatedMtFtsZ-DeltaC169 mutant of Mycobacterium tubercu-losis FtsZ shows GTPase and GTP-induced, GTP-specific polymerization activities in vitro. Microbiol-ogy 150, 3906–3908.
Herman, C., Ogura, T., Tomoyasu, T., Hiraga, S.,Akiyama, Y., Ito, K., Thomas, R., D’Ari, R., Bouloc,P., 1993. Cell growth and lambda phage developmentcontrolled by the same essential Escherichia coli gene,ftsH/hflB. Proc. Natl. Acad. Sci. USA 90, 10861–10865.
Herman, C., Thevenet, D., D’Ari, R., Bouloc, P., 1995.Degradation of sigma 32, the heat shock regulator in
Escherichia coli, is governed by HflB. Proc. Natl. Acad.Sci. USA 92, 3516–3520.
Herman, C., Thevenet, D., D’Ari, R., Bouloc, P., 1997.The HflB protease of Escherichia coli degrades itsinhibitor lambda cIII. J. Bacteriol. 179, 358–363.
Karata, K., Inagawa, T., Wilkinson, A.J., Tatsuta, T.,Ogura, T., 1999. Dissecting the role of a conservedmotif (the second region of homology) in the AAAfamily of ATPases. Site-directed mutagenesis of theATP-dependent protease FtsH. J. Biol. Chem. 274,26225–26232.
Karata, K., Verma, C.S., Wilkinson, A.J., Ogura, T., 2001.Probing the mechanism of ATP hydrolysis and sub-strate translocation in the AAA protease FtsH bymodeling and mutagenesis. Mol. Microbiol. 39,890–903.
Kelly, A.J., Sackett, M.J., Din, N., Quardokus, E., Brun,Y.V., 1998. Cell cycle-dependent transcriptional andproteolytic regulation of FtsZ in Caulobacter. GenesDev. 12, 880–893.
Kihara, A., Akiyama, Y., Ito, K., 1995. FtsH is required forproteolytic elimination of uncomplexed forms of SecY,an essential protein translocase subunit. Proc. Natl.Acad. Sci. USA 92, 4532–4536.
Kihara, A., Akiyama, Y., Ito, K., 1999. Dislocation ofmembrane proteins in FtsH-mediated proteolysis.EMBO J. 18, 2970–2981.
Lindahl, M., Tabak, S., Cseke, L., Pichersky, E., Anders-son, B., Adam, Z., 1996. Identification, characteriza-tion, and molecular cloning of a homologue of thebacterial FtsH protease in chloroplasts of higherplants. J. Biol. Chem. 271, 29329–29334.
Lowe, J., Amos, L.A., 1998. Crystal structure of thebacterial cell-division protein FtsZ. Nature 391,203–206.
Lysenko, E., Ogura, T., Cutting, S.M., 1997. Character-ization of the ftsH gene of Bacillus subtilis. Micro-biology 143, 971–978.
Ma, X., Ehrhardt, D.W., Margolin, W., 1996. Colocaliza-tion of cell division proteins FtsZ and FtsA tocytoskeletal structures in living Escherichia coli cellsby using green fluorescent protein. Proc. Natl. Acad.Sci. USA 93, 12998–13003.
Ma, X., Margolin, W., 1999. Genetic and functionalanalyses of the conserved C-terminal core domain ofEscherichia coli FtsZ. J. Bacteriol. 181, 7531–7544.
Miroux, D., Walker, J.E., 1996. Over-production ofproteins in Escherichia coli: mutant hosts that allowsynthesis of some membrane proteins and globularproteins at high levels. J. Mol. Biol. 260, 289–298.
Mukherjee, A., Lutkenhaus, J., 1994. Guanine nucleo-tide-dependent assembly of FtsZ into filaments.J. Bacteriol. 176, 2754–2758.
Mukherjee, A., Lutkenhaus, J., 1999. Analysis of FtsZassembly by light scattering and determination of therole of divalent metal cations. J. Bacteriol. 181,823–832.
Nilsson, D., Lauridsen, A.A., Tomoyasu, T., Ogura, T.,1994. A Lactococcus lactis gene encodes a membraneprotein with putative ATPase activity that is homo-

ARTICLE IN PRESS
R. Srinivasan et al.30
logous to the essential Escherichia coli ftsH geneproduct. Microbiology 140, 2601–2610.
Ogura, T., Tomoyasu, T., Yuki, T., Morimura, S., Begg,K.J., Donachie, W.D., Mori, H., Niki, H., Hiraga, S.,1991. Structure and function of the ftsH gene inEscherichia coli. Res. Microbiol. 142, 279–282.
Ogura, T., Inoue, K., Tatsuta, T., Suzaki, T., Karata, K.,Young, K., Su, L.H., Fierke, C.A., Jackman, J.E., Raetz,C.R., Coleman, J., Tomoyasu, T., Matsuzawa, H., 1999.Balanced biosynthesis of major membrane componentsthrough regulated degradation of the committed en-zyme of lipid A biosynthesis by the AAA protease FtsH(HflB) in Escherichia coli. Mol. Microbiol. 31, 833–844.
Ogura, T., Wilkinson, A.J., 2001. AAA+ superfamilyATPases: common stucture – diverse function. GenesCells 6, 575–597.
RayChaudhuri, D., Park, J.T., 1992. Escherichia coli cell-division gene ftsZ encodes a novel GTP-bindingprotein. Nature 359, 251–254.
Rueda, S., Vicente, M., Mingorance, J., 2003. Concentra-tion and assembly of the division ring proteins FtsZ,FtsA, and ZipA during the Escherichia coli cell cycle. J.Bacteriol. 185, 3344–3351.
Shotland, Y., Koby, S., Teff, D., Mansur, N., Oren, D.A.,Tatematsu, K., Tomoyasu, T., Kessel, M., Bukau, B.,Ogura, T., Oppenheim, A.B., 1997. Proteolysis of thephage lambda CII regulatory protein by FtsH (HflB) ofEscherichia coli. Mol. Microbiol. 24, 1303–1310.
Shotland, Y., Shifrin, A., Ziv, T., Teff, D., Koby, S., Kobiler,O., Oppenheim, A.B., 2000. Proteolysis of bacterioph-age lambda CII by Escherichia coli FtsH (HflB).J. Bacteriol. 182, 3111–3116.
Stricker, J., Erickson, H.P., 2003. In vivo characteri-zation of Escherichia coli ftsZ mutants: effects on
Z-ring structure and function. J. Bacteriol. 185,4796–4805.
Tomoyasu, T., Yamanaka, K., Murata, K., Suzaki, T.,Bouloc, P., Kato, A., Niki, H., Hiraga, S., Ogura, T.,1993a. Topology and subcellular localization of FtsHprotein in Escherichia coli. J. Bacteriol. 175,1352–1357.
Tomoyasu, T., Yuki, T., Morimura, S., Mori, H., Yamanaka,K., Niki, H., Hiraga, S., Ogura, T., 1993b. TheEscherichia coli FtsH protein is a prokaryotic memberof a protein family of putative ATPases involved inmembrane functions, cell cycle control, and geneexpression. J. Bacteriol. 175, 1344–1351.
Tomoyasu, T., Gamer, J., Bukau, B., Kanemori, M., Mori,H., Rutman, A.J., Oppenheim, A.B., Yura, T., Yama-naka, K., Niki, H., et al., 1995. Escherichia coli FtsH isa membrane-bound, ATP-dependent protease whichdegrades the heat-shock transcription factor sigma 32.EMBO J. 14, 2551–2560.
Tomoyasu, T., Arsene, F., Ogura, T., Bukau, B., 2001. TheC terminus of sigma 32 is not essential for degradationby FtsH. J. Bacteriol. 183, 5911–5917.
Weart, R.B., Levin, P.A., 2003. Growth rate-dependentregulation of medial FtsZ ring formation. J. Bacteriol.185, 2826–2834.
Wehrl, W., Niederweis, M., Schumann, W., 2000. TheFtsH protein accumulates at the septum of Bacillussubtilis during cell division and sporulation.J. Bacteriol. 182, 3870–3873.
Zellmeier, S., Zuber, U., Schumann, W., Wiegert, T.,2003. The absence of FtsH metalloprotease activitycauses overexpression of the sigmaW-controlled pbpEgene, resulting in filamentous growth of Bacillussubtilis. J. Bacteriol. 185, 973–982.




















