
Filamentous actin is a substrate for protealysin, a metalloprotease of invasive Serratia...
11
Filamentous actin is a substrate for protealysin, a metalloprotease of invasive Serratia proteamaculans Olga Tsaplina 1 , Tatiana Efremova 1 , Ilya Demidyuk 2 and Sofia Khaitlina 1 1 Institute of Cytology, RAS, St Petersburg, Russia 2 Institute of Molecular Genetics, RAS, Moscow, Russia Keywords aluminium fluoride; bacterial proteases; cytoskeleton; ECP32 ⁄ grimelysin; F-actin dynamics Correspondence S. Khaitlina, Institute of Cytology, Russian Academy of Sciences, Tikhoretsky av. 4, 194064 St Petersburg, Russia Fax: +7 812 297 03 41 Tel: +7 812 297 42 96 E-mail: [email protected] (Received 2 August 2011, revised 3 November 2011, accepted 4 November 2011) doi:10.1111/j.1742-4658.2011.08420.x Homologous bacterial metalloproteases ECP32 ⁄ grimelysin from Serratia grimesii and protealysin from Serratia proteamaculans are involved in the invasion of the nonpathogenic bacteria in eukaryotic cells and are sug- gested to translocate into the cytoplasm [Bozhokina ES et al. (2011) Cell Biol Int 35, 111–118]. The proteases have been characterized as actin- hydrolyzing enzymes with a narrow specificity toward intact cell proteins. However, cleavage of filamentous actin (F-actin) (i.e. the main actin species in the cell) and the properties of the cleaved F-actin have not been investi- gated previously. In the present study, we revealed the presence of proteal- ysin in the cytoplasm of 3T3-SV40 cells infected with S. proteamaculans or recombinant Escherichia coli expressing the protealysin gene. We also show for the first time that purified protealysin and the lysates of the recombi- nant E. coli producing protealysin cleave 20–40% of F-actin. Cleavage lim- ited predominantly to the bond Gly42-Val43 efficiently increases the steady-state ATPase activity (dynamics) of F-actin. AlF 4 abolishes this effect and promotes the nucleation of protealysin-cleaved Mg-globular- actin even in the absence of 0.1 M KCl, most likely as a result of the stabi- lization of lateral intermonomer contacts of actin subunits. The results obtained in the present study suggest that F-actin can be a target for protealysin upon its translocation into the host cell. Structured digital abstract l Protealysin cleaves Actin by protease assay ( View interaction) Introduction Actin is involved in the majority of cell processes, ranging from chromatin remodeling and transcription regulation to intracellular transport, cell organization and various types of cell movements and muscle con- traction. It is natural, therefore, that the actin cyto- skeleton is a target for drugs and toxins, as well as for viruses and pathogenic bacteria interacting with eukaryotic cells [1–5]. The effectors modify cytoskele- ton dynamics through the modulation of small GTPases and actin-binding proteins, or by direct interaction with actin. Many bacterial effectors promote the acti- vation of small GTPases Rho, Rac and Cdc42, follo- wed by the activation of signaling cascades resulting in actin polymerization [6–8]. Thereby bacterial effectors often mimic natural activators of small GTPases or directly stimulate the host signaling pathways by a functional mimicry of host GTPases [7,8]. Other bacterial effectors stimulate the efficient uptake of bacteria in host cells by the formation of complexes with actin-binding proteins [9], or by mediating Abbreviations F-actin, filamentous actin; FITC, fluorescein isothiocyanate; G-actin, globular actin; P i, inorganic phosphate. 264 FEBS Journal 279 (2012) 264–274 ª 2011 The Authors Journal compilation ª 2011 FEBS
-
Upload
independent -
Category
Documents
-
view
0 -
download
0
Transcript of Filamentous actin is a substrate for protealysin, a metalloprotease of invasive Serratia...

Filamentous actin is a substrate for protealysin,a metalloprotease of invasive Serratia proteamaculansOlga Tsaplina1, Tatiana Efremova1, Ilya Demidyuk2 and Sofia Khaitlina1
1 Institute of Cytology, RAS, St Petersburg, Russia
2 Institute of Molecular Genetics, RAS, Moscow, Russia
Keywords
aluminium fluoride; bacterial proteases;
cytoskeleton; ECP32 ⁄ grimelysin; F-actin
dynamics
Correspondence
S. Khaitlina, Institute of Cytology, Russian
Academy of Sciences, Tikhoretsky av. 4,
194064 St Petersburg, Russia
Fax: +7 812 297 03 41
Tel: +7 812 297 42 96
E-mail: [email protected]
(Received 2 August 2011, revised 3
November 2011, accepted 4 November
2011)
doi:10.1111/j.1742-4658.2011.08420.x
Homologous bacterial metalloproteases ECP32 ⁄ grimelysin from Serratia
grimesii and protealysin from Serratia proteamaculans are involved in the
invasion of the nonpathogenic bacteria in eukaryotic cells and are sug-
gested to translocate into the cytoplasm [Bozhokina ES et al. (2011) Cell
Biol Int 35, 111–118]. The proteases have been characterized as actin-
hydrolyzing enzymes with a narrow specificity toward intact cell proteins.
However, cleavage of filamentous actin (F-actin) (i.e. the main actin species
in the cell) and the properties of the cleaved F-actin have not been investi-
gated previously. In the present study, we revealed the presence of proteal-
ysin in the cytoplasm of 3T3-SV40 cells infected with S. proteamaculans or
recombinant Escherichia coli expressing the protealysin gene. We also show
for the first time that purified protealysin and the lysates of the recombi-
nant E. coli producing protealysin cleave 20–40% of F-actin. Cleavage lim-
ited predominantly to the bond Gly42-Val43 efficiently increases the
steady-state ATPase activity (dynamics) of F-actin. AlF�4 abolishes this
effect and promotes the nucleation of protealysin-cleaved Mg-globular-
actin even in the absence of 0.1 M KCl, most likely as a result of the stabi-
lization of lateral intermonomer contacts of actin subunits. The results
obtained in the present study suggest that F-actin can be a target for protealysin
upon its translocation into the host cell.
Structured digital abstractl Protealysin cleaves Actin by protease assay (View interaction)
Introduction
Actin is involved in the majority of cell processes,
ranging from chromatin remodeling and transcription
regulation to intracellular transport, cell organization
and various types of cell movements and muscle con-
traction. It is natural, therefore, that the actin cyto-
skeleton is a target for drugs and toxins, as well as for
viruses and pathogenic bacteria interacting with
eukaryotic cells [1–5]. The effectors modify cytoskele-
ton dynamics through the modulation of small GTPases
and actin-binding proteins, or by direct interaction
with actin. Many bacterial effectors promote the acti-
vation of small GTPases Rho, Rac and Cdc42, follo-
wed by the activation of signaling cascades resulting in
actin polymerization [6–8]. Thereby bacterial effectors
often mimic natural activators of small GTPases or
directly stimulate the host signaling pathways by a
functional mimicry of host GTPases [7,8]. Other
bacterial effectors stimulate the efficient uptake of
bacteria in host cells by the formation of complexes
with actin-binding proteins [9], or by mediating
Abbreviations
F-actin, filamentous actin; FITC, fluorescein isothiocyanate; G-actin, globular actin; Pi, inorganic phosphate.
264 FEBS Journal 279 (2012) 264–274 ª 2011 The Authors Journal compilation ª 2011 FEBS

phosphorylation ⁄dephosphorylation of an actin-bind-
ing protein [10–13].
The other way in which many toxins and bacterial
factors modify the cytoskeleton is the direct interaction
of the effectors with actin. Latrunculin from sponges
reversibly and specifically disrupts actin cytoskeleton
by altering the actin monomer interface and thus
inhibiting polymerization [3,14]. Marine macrolide tox-
ins disrupt the cytoskeleton because they target both
globular actin and actin filaments with a high affinity
and either sterically prevent polymerization or mimic
fragmentation of filamentous actin (F-actin) by sever-
ing proteins [15,16]. Clostridium botulinum C2 toxin
and ADP-ribosyltransferase SpvB from Salmonella ent-
erica cause ADP-ribosylation of actin at Arg177,
thereby inhibiting actin polymerization [17,18]. A
unique mechanism of actin filament depolymerization
is exhibited by Vibrio cholerae RTX toxin, which cova-
lently cross-links actin monomers, thus interfering with
the formation of functional filaments [19]. On the
other hand, the bacterial protein SipA from Salmo-
nella typhimurium inhibits depolymerization of actin fila-
ments mechanically stabilizing the filament by
tethering actin subunits in opposing strands [20,21].
Even these few examples demonstrate the variability of
the mechanisms developed by bacteria and other
organisms to subvert actin dynamics for facilitating
the corresponding invasion or protection processes.
Previously, we reported that spontaneously isolated
nonpathogenic bacteria Serratia grimesii and Serratia
proteamaculans synthesize thermolysin-like metallopro-
teases ECP32 ⁄ grimelysin [22,23] or protealysin [24–26],
which are characterized by a high specificity toward
actin. The appearance of the proteases correlated with
the capability of the bacteria to invade eukaryotic cells
and remodel their cytoskeleton [26,27]. Moreover, non-
invasive E. coli transformed by the grimelysin or the
protealysin gene confer the invasive phenotype [28].
We have also shown that protealysin introduced into
the culture medium does not induce the internalization
of non-invasive E. coli [28], which led us to suggest
that the enzyme translocates into the eukaryotic cell
where it modifies cytoplasmic proteins.
Proteases ECP32 ⁄grimelysin and protealysin are
actin-hydrolyzing enzymes with a narrow specificity
toward intact cell proteins [22,26]. Limited proteolysis
of globular actin (G-actin) by protease ECP32 abol-
ishes or slows down polymerization [29,30] and
enhances the dynamics of the filaments formed by
the cleaved monomers [31], thus producing changes in the
actin cytoskeleton favorable for bacteria to enter
the host cell. However, under physiological conditions,
actin cannot exist as a free monomer but forms fila-
ments (i.e. F-actin) whose accessibility to proteolysis is
very low [32,33]. The properties of cleaved F-actin
have not been investigated previously. In the present
study, we took advantage of using antibodies against
protealysin, purified recombinant protealysin and
lysates of recombinant E. coli expressing the protealy-
sin gene, aiming to elucidate the localization of prote-
alysin in the infected cell culture and the effects of
protealysin on F-actin dynamics.
The results obtained demonstrate for the first time
that the infection of eukaryotic cells with protealysin-
producing bacteria is accompanied by the translocation
of protealysin into the host cells. Furthermore, we
show that up to 40% of F-actin is digested with prote-
alysin in vitro within a time approximately correspond-
ing to that of the invasion experiments. F-actin is
cleaved by protealysin mainly at Gly42-Val43 within
the DNase I-binding loop, which strongly enhances the
filament dynamics. AlF�4 stabilizes the filaments and
promotes the nucleation step of actin polymerization,
most likely enhancing the effect of magnesium ions on
the lateral intermonomer contacts along the actin fila-
ment. These properties of protealysin-cleaved actin did
not depend upon whether purified protealysin or the
lysates of E. coli expressing the protealysin gene were
used. Therefore, we conclude that protealysin appears
to be the only component of the bacterial lysates to
affect actin dynamics. Taken together, these results
suggest that, as protealysin translocates into the host
cell, both G-actin and F-actin are targets for the
enzyme.
Results
Localization of protealysin in the infected cells
In invasion experiments, cultured cells are incubated
with the fluorescein isothiocyanate (FITC)-conjugated
bacteria, fixed with formaldehyde, stained with rhoda-
mine–phalloidin, and examined under a confocal
microscope to visualize bacteria and cytoskeleton
structures [26,28]. We used a similar protocol to reveal
the localization of protealysin in the infected cells by
staining the enzyme with anti-protealysin serum. Fig-
ure 1 represents the results of a typical experiment in
which cultured Balb 3T3 SV40 cells were incubated
either with the wild-type protealysin-producing bacte-
ria S. proteamaculans 94 (Fig. 1A) or with recombi-
nant E. coli BL21 (DE3) (pProPlnHis6) expressing the
protealysin gene (Fig. 1B). Lysates of these bacteria
demonstrated efficient and specific proteolytic activity
toward G-actin (Fig. 1D). As a control, we used
recombinant E. coli BL21 (DE3) transformed with
O. Tsaplina et al. Cleavage of F-actin with protealysin
FEBS Journal 279 (2012) 264–274 ª 2011 The Authors Journal compilation ª 2011 FEBS 265

plasmid pET23b not carrying the protealysin gene
(Fig. 1C). Lysates of E. coli BL21 (DE3) (pET23b) did
not exhibit any actin-hydrolyzing activity (Fig. 1D). In
line with our previous results [26–28], within 2 h of
incubation, the actin-containing structures within the
cells were rearranged, indicating that the stress fibres
had been destroyed (Fig. S1) and � 10% of the cells
were invaded by either the wild-type or recombinant
bacteria. Anti-protealysin serum revealed the enzyme
in both the cells infected by S. proteamaculans
(Fig. 1A) and those infected by the recombinant E. coli
synthesizing protealysin (Fig. 1A,B). In addition, colo-
calization of antibodies with bacteria was sometimes
observed. By contrast, E. coli BL21 (DE3) (pET23b)
not carrying the protealysin gene did not enter 3T3
SV40 cells and, in these samples, no protealysin could
be detected (Fig. 1C). These data imply that invasion
of the protealysin-producing bacteria is accompanied
by translocation of the enzyme into eukaryotic cells.
Limited proteolysis of F-actin
Previously, we showed that protealysin cleaves globu-
lar actin between Gly42 and Val43 within the DNase
I-binding loop, generating two fragments of 36 and
5 kDa. At a high enzyme ⁄ actin ratio, further proteoly-
sis of the 36-kDa fragment yields a 33-kDa product as
a result of cleavage of the peptide bonds Thr66-Leu67
and Gly63-Ile64 in the nucleotide cleft [26]. In the
actin polymer (F-actin), the sites Gly42-Val43 and
Gly63-Ile64 are involved in the monomer–monomer
contacts [34,35], which can protect them from
A B C
D
a b
Fig. 1. Localization of protealysin during the invasion of protealysin-producing bacteria in eukaryotic cells. S. proteamaculans 94 grown for
45 h at 30 �C with aeration (A, a), E. coli BL21 (DE3) (pProPlnHis6) carrying the protealysin gene (B, b) and E. coli BL21 (DE3) (pET23b) not
carrying the protealysin gene (C) grown for 18 h at 37 �C with aeration were stained with FITC as described in the Materials and methods.
Balb 3T3-SV40 fibroblasts were incubated with bacteria for 2 h, fixed with formaldehyde and stained with polyclonal rabbit anti-protealysin
serum (blue) and rhodamine-phalloidin (red). (a, b) Higher magnification of the regions delimited by the white frames in (A) and (B), respec-
tively. Images of the optical sections were taken over the middle of the cells. Scale bar = 15 lm. (D), Limited proteolysis of G-actin with
lysates of S. proteamaculans 94 (a), E. coli BL21 (DE3) (pProPlnHis6) (b) and E. coli BL21 (DE3) (pET23b) (c), corresponding to the invasion
experiments shown in (A, a), (B, b) and (C), respectively. G-actin (1 mgÆmL)1) was incubated with an equal volume of the lysate for 2 h at
22 �C. Ac, nondigested actin.
Cleavage of F-actin with protealysin O. Tsaplina et al.
266 FEBS Journal 279 (2012) 264–274 ª 2011 The Authors Journal compilation ª 2011 FEBS
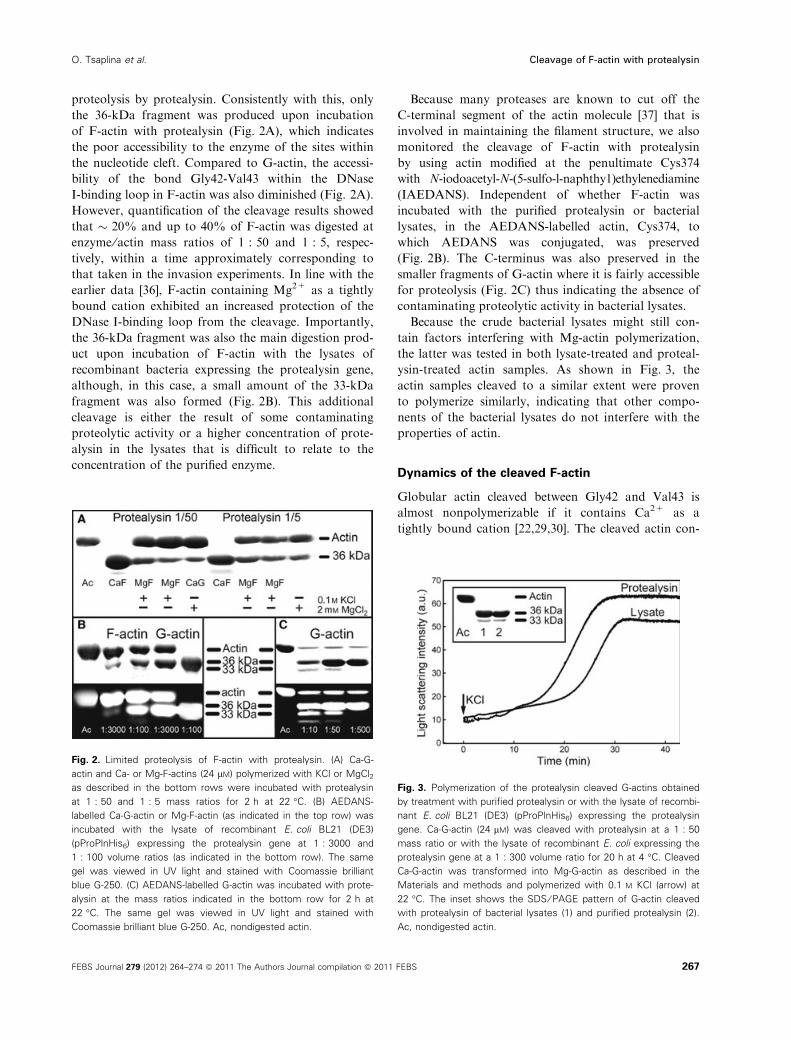
proteolysis by protealysin. Consistently with this, only
the 36-kDa fragment was produced upon incubation
of F-actin with protealysin (Fig. 2A), which indicates
the poor accessibility to the enzyme of the sites within
the nucleotide cleft. Compared to G-actin, the accessi-
bility of the bond Gly42-Val43 within the DNase
I-binding loop in F-actin was also diminished (Fig. 2A).
However, quantification of the cleavage results showed
that � 20% and up to 40% of F-actin was digested at
enzyme ⁄ actin mass ratios of 1 : 50 and 1 : 5, respec-
tively, within a time approximately corresponding to
that taken in the invasion experiments. In line with the
earlier data [36], F-actin containing Mg2+ as a tightly
bound cation exhibited an increased protection of the
DNase I-binding loop from the cleavage. Importantly,
the 36-kDa fragment was also the main digestion prod-
uct upon incubation of F-actin with the lysates of
recombinant bacteria expressing the protealysin gene,
although, in this case, a small amount of the 33-kDa
fragment was also formed (Fig. 2B). This additional
cleavage is either the result of some contaminating
proteolytic activity or a higher concentration of prote-
alysin in the lysates that is difficult to relate to the
concentration of the purified enzyme.
Because many proteases are known to cut off the
C-terminal segment of the actin molecule [37] that is
involved in maintaining the filament structure, we also
monitored the cleavage of F-actin with protealysin
by using actin modified at the penultimate Cys374
with N-iodoacetyl-N-(5-sulfo-l-naphthy1)ethylenediamine
(IAEDANS). Independent of whether F-actin was
incubated with the purified protealysin or bacterial
lysates, in the AEDANS-labelled actin, Cys374, to
which AEDANS was conjugated, was preserved
(Fig. 2B). The C-terminus was also preserved in the
smaller fragments of G-actin where it is fairly accessible
for proteolysis (Fig. 2C) thus indicating the absence of
contaminating proteolytic activity in bacterial lysates.
Because the crude bacterial lysates might still con-
tain factors interfering with Mg-actin polymerization,
the latter was tested in both lysate-treated and proteal-
ysin-treated actin samples. As shown in Fig. 3, the
actin samples cleaved to a similar extent were proven
to polymerize similarly, indicating that other compo-
nents of the bacterial lysates do not interfere with the
properties of actin.
Dynamics of the cleaved F-actin
Globular actin cleaved between Gly42 and Val43 is
almost nonpolymerizable if it contains Ca2+ as a
tightly bound cation [22,29,30]. The cleaved actin con-
Fig. 2. Limited proteolysis of F-actin with protealysin. (A) Ca-G-
actin and Ca- or Mg-F-actins (24 lM) polymerized with KCl or MgCl2
as described in the bottom rows were incubated with protealysin
at 1 : 50 and 1 : 5 mass ratios for 2 h at 22 �C. (B) AEDANS-
labelled Ca-G-actin or Mg-F-actin (as indicated in the top row) was
incubated with the lysate of recombinant E. coli BL21 (DE3)
(pProPlnHis6) expressing the protealysin gene at 1 : 3000 and
1 : 100 volume ratios (as indicated in the bottom row). The same
gel was viewed in UV light and stained with Coomassie brilliant
blue G-250. (C) AEDANS-labelled G-actin was incubated with prote-
alysin at the mass ratios indicated in the bottom row for 2 h at
22 �C. The same gel was viewed in UV light and stained with
Coomassie brilliant blue G-250. Ac, nondigested actin.
Fig. 3. Polymerization of the protealysin cleaved G-actins obtained
by treatment with purified protealysin or with the lysate of recombi-
nant E. coli BL21 (DE3) (pProPlnHis6) expressing the protealysin
gene. Ca-G-actin (24 lM) was cleaved with protealysin at a 1 : 50
mass ratio or with the lysate of recombinant E. coli expressing the
protealysin gene at a 1 : 300 volume ratio for 20 h at 4 �C. Cleaved
Ca-G-actin was transformed into Mg-G-actin as described in the
Materials and methods and polymerized with 0.1 M KCl (arrow) at
22 �C. The inset shows the SDS ⁄ PAGE pattern of G-actin cleaved
with protealysin of bacterial lysates (1) and purified protealysin (2).
Ac, nondigested actin.
O. Tsaplina et al. Cleavage of F-actin with protealysin
FEBS Journal 279 (2012) 264–274 ª 2011 The Authors Journal compilation ª 2011 FEBS 267
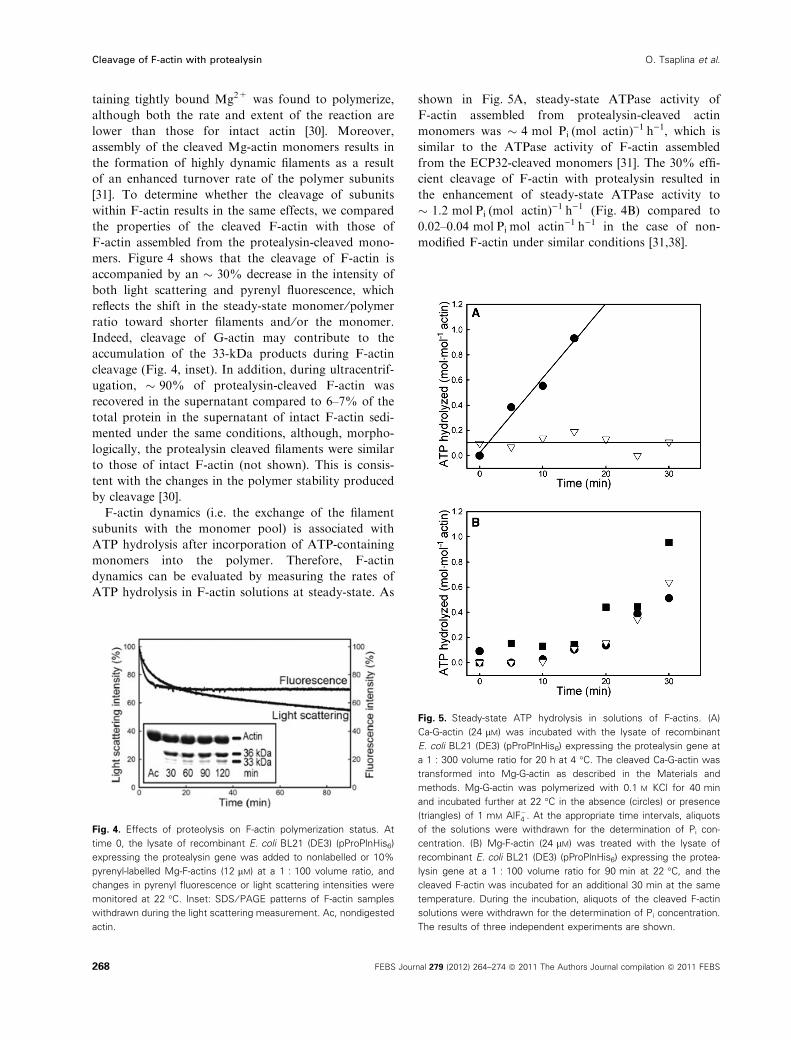
taining tightly bound Mg2+ was found to polymerize,
although both the rate and extent of the reaction are
lower than those for intact actin [30]. Moreover,
assembly of the cleaved Mg-actin monomers results in
the formation of highly dynamic filaments as a result
of an enhanced turnover rate of the polymer subunits
[31]. To determine whether the cleavage of subunits
within F-actin results in the same effects, we compared
the properties of the cleaved F-actin with those of
F-actin assembled from the protealysin-cleaved mono-
mers. Figure 4 shows that the cleavage of F-actin is
accompanied by an � 30% decrease in the intensity of
both light scattering and pyrenyl fluorescence, which
reflects the shift in the steady-state monomer ⁄polymer
ratio toward shorter filaments and ⁄or the monomer.
Indeed, cleavage of G-actin may contribute to the
accumulation of the 33-kDa products during F-actin
cleavage (Fig. 4, inset). In addition, during ultracentrif-
ugation, � 90% of protealysin-cleaved F-actin was
recovered in the supernatant compared to 6–7% of the
total protein in the supernatant of intact F-actin sedi-
mented under the same conditions, although, morpho-
logically, the protealysin cleaved filaments were similar
to those of intact F-actin (not shown). This is consis-
tent with the changes in the polymer stability produced
by cleavage [30].
F-actin dynamics (i.e. the exchange of the filament
subunits with the monomer pool) is associated with
ATP hydrolysis after incorporation of ATP-containing
monomers into the polymer. Therefore, F-actin
dynamics can be evaluated by measuring the rates of
ATP hydrolysis in F-actin solutions at steady-state. As
shown in Fig. 5A, steady-state ATPase activity of
F-actin assembled from protealysin-cleaved actin
monomers was � 4 mol PiÆ(mol actin))1Æh)1, which is
similar to the ATPase activity of F-actin assembled
from the ECP32-cleaved monomers [31]. The 30% effi-
cient cleavage of F-actin with protealysin resulted in
the enhancement of steady-state ATPase activity to
� 1.2 molÆPiÆ(mol actin))1Æh)1 (Fig. 4B) compared to
0.02–0.04 molÆPiÆmol actin)1Æh)1 in the case of non-
modified F-actin under similar conditions [31,38].
Fig. 4. Effects of proteolysis on F-actin polymerization status. At
time 0, the lysate of recombinant E. coli BL21 (DE3) (pProPlnHis6)
expressing the protealysin gene was added to nonlabelled or 10%
pyrenyl-labelled Mg-F-actins (12 lM) at a 1 : 100 volume ratio, and
changes in pyrenyl fluorescence or light scattering intensities were
monitored at 22 �C. Inset: SDS ⁄ PAGE patterns of F-actin samples
withdrawn during the light scattering measurement. Ac, nondigested
actin.
Fig. 5. Steady-state ATP hydrolysis in solutions of F-actins. (A)
Ca-G-actin (24 lM) was incubated with the lysate of recombinant
E. coli BL21 (DE3) (pProPlnHis6) expressing the protealysin gene at
a 1 : 300 volume ratio for 20 h at 4 �C. The cleaved Ca-G-actin was
transformed into Mg-G-actin as described in the Materials and
methods. Mg-G-actin was polymerized with 0.1 M KCl for 40 min
and incubated further at 22 �C in the absence (circles) or presence
(triangles) of 1 mM AlF�4 . At the appropriate time intervals, aliquots
of the solutions were withdrawn for the determination of Pi con-
centration. (B) Mg-F-actin (24 lM) was treated with the lysate of
recombinant E. coli BL21 (DE3) (pProPlnHis6) expressing the protea-
lysin gene at a 1 : 100 volume ratio for 90 min at 22 �C, and the
cleaved F-actin was incubated for an additional 30 min at the same
temperature. During the incubation, aliquots of the cleaved F-actin
solutions were withdrawn for the determination of Pi concentration.
The results of three independent experiments are shown.
Cleavage of F-actin with protealysin O. Tsaplina et al.
268 FEBS Journal 279 (2012) 264–274 ª 2011 The Authors Journal compilation ª 2011 FEBS
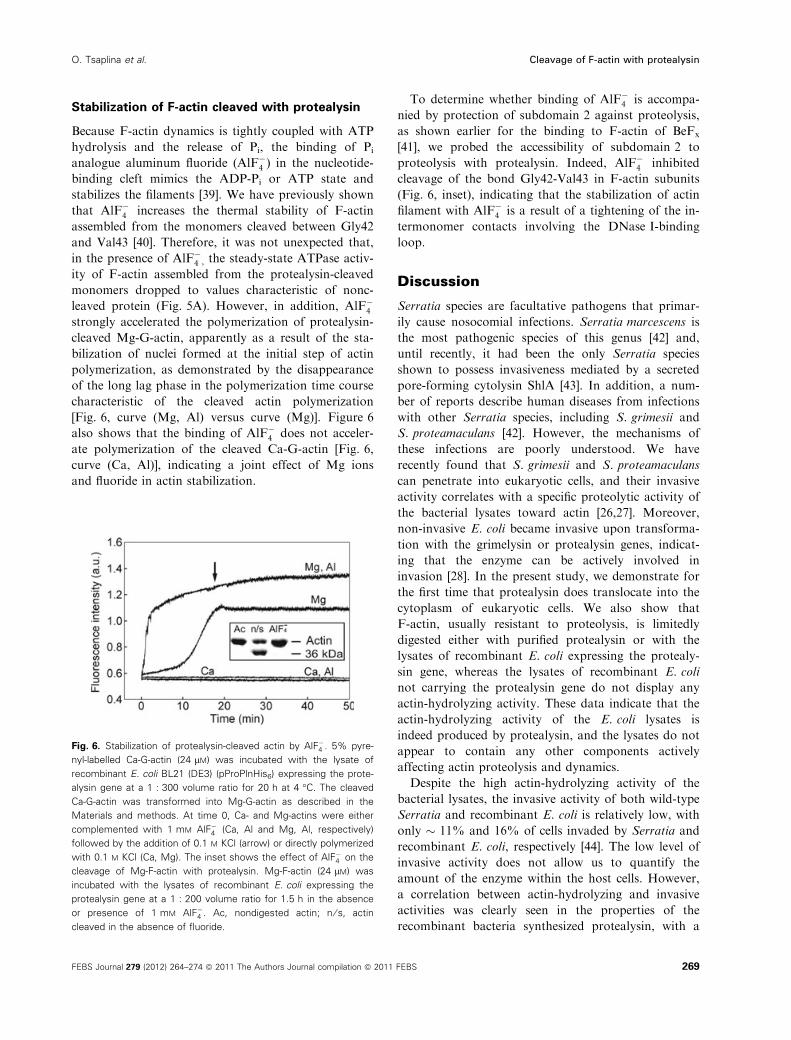
Stabilization of F-actin cleaved with protealysin
Because F-actin dynamics is tightly coupled with ATP
hydrolysis and the release of Pi, the binding of Pi
analogue aluminum fluoride (AlF�4 ) in the nucleotide-
binding cleft mimics the ADP-Pi or ATP state and
stabilizes the filaments [39]. We have previously shown
that AlF�4 increases the thermal stability of F-actin
assembled from the monomers cleaved between Gly42
and Val43 [40]. Therefore, it was not unexpected that,
in the presence of AlF�4 , the steady-state ATPase activ-
ity of F-actin assembled from the protealysin-cleaved
monomers dropped to values characteristic of nonc-
leaved protein (Fig. 5A). However, in addition, AlF�4strongly accelerated the polymerization of protealysin-
cleaved Mg-G-actin, apparently as a result of the sta-
bilization of nuclei formed at the initial step of actin
polymerization, as demonstrated by the disappearance
of the long lag phase in the polymerization time course
characteristic of the cleaved actin polymerization
[Fig. 6, curve (Mg, Al) versus curve (Mg)]. Figure 6
also shows that the binding of AlF�4 does not acceler-
ate polymerization of the cleaved Ca-G-actin [Fig. 6,
curve (Ca, Al)], indicating a joint effect of Mg ions
and fluoride in actin stabilization.
To determine whether binding of AlF�4 is accompa-
nied by protection of subdomain 2 against proteolysis,
as shown earlier for the binding to F-actin of BeFx
[41], we probed the accessibility of subdomain 2 to
proteolysis with protealysin. Indeed, AlF�4 inhibited
cleavage of the bond Gly42-Val43 in F-actin subunits
(Fig. 6, inset), indicating that the stabilization of actin
filament with AlF�4 is a result of a tightening of the in-
termonomer contacts involving the DNase I-binding
loop.
Discussion
Serratia species are facultative pathogens that primar-
ily cause nosocomial infections. Serratia marcescens is
the most pathogenic species of this genus [42] and,
until recently, it had been the only Serratia species
shown to possess invasiveness mediated by a secreted
pore-forming cytolysin ShlA [43]. In addition, a num-
ber of reports describe human diseases from infections
with other Serratia species, including S. grimesii and
S. proteamaculans [42]. However, the mechanisms of
these infections are poorly understood. We have
recently found that S. grimesii and S. proteamaculans
can penetrate into eukaryotic cells, and their invasive
activity correlates with a specific proteolytic activity of
the bacterial lysates toward actin [26,27]. Moreover,
non-invasive E. coli became invasive upon transforma-
tion with the grimelysin or protealysin genes, indicat-
ing that the enzyme can be actively involved in
invasion [28]. In the present study, we demonstrate for
the first time that protealysin does translocate into the
cytoplasm of eukaryotic cells. We also show that
F-actin, usually resistant to proteolysis, is limitedly
digested either with purified protealysin or with the
lysates of recombinant E. coli expressing the protealy-
sin gene, whereas the lysates of recombinant E. coli
not carrying the protealysin gene do not display any
actin-hydrolyzing activity. These data indicate that the
actin-hydrolyzing activity of the E. coli lysates is
indeed produced by protealysin, and the lysates do not
appear to contain any other components actively
affecting actin proteolysis and dynamics.
Despite the high actin-hydrolyzing activity of the
bacterial lysates, the invasive activity of both wild-type
Serratia and recombinant E. coli is relatively low, with
only � 11% and 16% of cells invaded by Serratia and
recombinant E. coli, respectively [44]. The low level of
invasive activity does not allow us to quantify the
amount of the enzyme within the host cells. However,
a correlation between actin-hydrolyzing and invasive
activities was clearly seen in the properties of the
recombinant bacteria synthesized protealysin, with a
Fig. 6. Stabilization of protealysin-cleaved actin by AlF�4 . 5% pyre-
nyl-labelled Ca-G-actin (24 lM) was incubated with the lysate of
recombinant E. coli BL21 (DE3) (pProPlnHis6) expressing the prote-
alysin gene at a 1 : 300 volume ratio for 20 h at 4 �C. The cleaved
Ca-G-actin was transformed into Mg-G-actin as described in the
Materials and methods. At time 0, Ca- and Mg-actins were either
complemented with 1 mM AlF�4 (Ca, Al and Mg, Al, respectively)
followed by the addition of 0.1 M KCl (arrow) or directly polymerized
with 0.1 M KCl (Ca, Mg). The inset shows the effect of AlF�4 on the
cleavage of Mg-F-actin with protealysin. Mg-F-actin (24 lM) was
incubated with the lysates of recombinant E. coli expressing the
protealysin gene at a 1 : 200 volume ratio for 1.5 h in the absence
or presence of 1 mM AlF�4 . Ac, nondigested actin; n ⁄ s, actin
cleaved in the absence of fluoride.
O. Tsaplina et al. Cleavage of F-actin with protealysin
FEBS Journal 279 (2012) 264–274 ª 2011 The Authors Journal compilation ª 2011 FEBS 269

point Glu113 ⁄Ala mutation in the active site that pre-
vented removal of the propeptide from the protealysin
precursor [45]. The recombinant bacteria synthesizing
inactivated protealysin were not found to be invasive
unless a small amount of actin-hydrolyzing activity
could be detected in their lysates at the late post-loga-
rithmic growth stage [28]. As the time of infection
increased, the efficiency of the invasion augmented
concomitantly with the enhancement of actin cleavage.
Furthermore, changes in cell morphology and cytoskel-
eton rearrangements accompanying the invasion of
E. coli synthesizing wild-type protealysin were more
pronounced than those produced by the E. coli synthe-
sizing mutant enzyme [28]. These results suggest that
cleavage-produced modifications of the properties of
actin promote bacterial internalization.
Limited proteolysis of G-actin within the DNase I-
binding loop has been shown to impair polymerization
[22,29,46] and enhance the polymer dynamics [31], as
well as decrease the affinity of gelsolin [47], myosin
subfragment 1 [48] and tropomyosin [49] to actin. It
has been suggested that the properties of F-actin trea-
ted with protease would be similar to those of F-actin
assembled from the proteolytically modified mono-
mers. The data reported in the present study provide
experimental support for this suggestion, which is
important in view of the translocation of the proteo-
lytic enzymes into the eukaryotic cell where F-actin is
a major actin species. Most importantly, cleavage of
F-actin with protealysin is accompanied by an efficient
enhancement of the filament dynamics. On the other
hand, aluminium fluoride, known to mimic a stable
ADP-Pi or ATP conformation of actin filaments
[39,41], accelerates the polymerization of protealysin-
cleaved actin monomers whose nucleation is impaired
by cleavage. Together with the increased dynamics of
the cleaved F-actin, these effects provide mechanisms
that may facilitate the invasion of eukaryotic cells by
bacteria.
Actin dynamics is determined by the polarity of the
actin filament where the protein subunits are added
predominantly to the ‘barbed end’, and dissociate
mainly from the other end, termed the ‘pointed end’.
Cleavage of actin between Gly42 and Val43 resulted in
enhancement of the turnover rate of polymer subunits
by one order of magnitude, primarily as a result of a
substantial increase in the rate constants for the disso-
ciation of actin subunits from the filament ends. The
dissociation rate constants were found to be approxi-
mately ninefold and more than fivefold higher for the
cleaved actin at the barbed and pointed ends, respec-
tively [31]. These effects appear to be determined by
the direct involvement of the cleavage site in the
intermonomer contacts of the filament and changes in
the overall conformation of subdomain 2 [31]. In addi-
tion, the recently published 3D structure of the pointed
end of the actin filament may shed light on the acceler-
ated release of the cleaved ADP-actin subunits from
the pointed end of the actin filament [50]. In this
model, the terminal subunit is tilted relative to the heli-
cal symmetry of the whole actin filament in such a
way that the hydrophobic loop of this subunit and the
DNase I-binding loop of the adjacent subunit are rear-
ranged to form a tighter contact, which may be
responsible for the low dissociation rate constants at
the pointed end of the actin filament [50]. Impairment
of the DNase I-binding loop by proteolysis may pre-
vent the tightening of the contact and result in faster
dissociation of the cleaved subunits and accelerated
filament dynamics.
Another important side of actin dynamics is the sta-
bilization of filaments by actin-binding proteins or low
molecular weight compounds. Recently, we have
shown that aluminium fluoride greatly increased the
thermal stability of F-actin assembled from the cleaved
actin monomers [40]. In the present study, we show
that this effect is associated with the inhibition by
AlF�4 of the filament dynamics and is likely a result of
tightening of the intermonomer contacts involving the
DNase I-binding loop. This is evident from the strong
protection of the protealysin cleavage site between
Gly42 and Val43 in subunits of AlF�4 -F-actin and is
consistent with the stabilization of actin subdomain 2
by BeFx, as demonstrated by the 3D reconstruction of
the filament structure [51] and biochemical approaches
[41].
The penetration of bacteria into eukaryotic cells
involves the disassembly of actin structures at the site
of bacterial–cell contact followed by actin polymeriza-
tion in cell surface protrusions, which rise around the
bacterial body and allow its engulfment in a macropin-
ocytic-like process [11,52]. The results obtained in the
present study show that F-actin can be limitedly
cleaved with protealysin within the DNase I-binding
loop, and that the cleavage strongly enhances the
dynamics of actin filaments and promotes their depoly-
merization. These effects can lead to the disassembly
of the actin cytoskeleton at the site of bacterial–cell
contacts and provide actin monomers for filament
assembly in cell surface protrusions. During this latter
step, the reversibility of the effects is of primary impor-
tance: assembly of the cleaved actin monomers into fila-
ments may be promoted by the low molecular weight
effectors mimicking Pi (present study) and actin-bind-
ing proteins [47,53] that stimulate actin polymerization
and stabilize the filaments. Thus, the limited cleavage
Cleavage of F-actin with protealysin O. Tsaplina et al.
270 FEBS Journal 279 (2012) 264–274 ª 2011 The Authors Journal compilation ª 2011 FEBS

of F-actin with bacterial enzymes allows actin to pre-
serve its functional properties and produce changes in
the actin cytoskeleton that could be used by bacteria
to enter the host cell. This specific activity of the
enzyme toward actin, taken together with its narrow
substrate specificity toward intact cell proteins [54,55],
suggests that, upon translocation of protealysin into
the host cell, actin proteolysis is involved in the inter-
nalization of bacteria.
Materials and methods
Reagents
Tris, ATP (disodium salt), sodium azide, AlCl3, NaF, Triton
X-100, reagents for electrophoresis, FITC, rhodamine–phal-
loidin, gelatin and Mounting medium were purchased from
Sigma Chemical Co. (St Louis, MO, USA). Peptone and
yeast extract were obtained from Difco (Franklin Lakes, NJ,
USA). IAEDANS and N-(1-pyrenyl)iodoacetamide were
from Molecular Probes Inc. (Carlsbad, CA, USA).
Bacterial strains, cell culture and growth
conditions
S. proteamaculans 94 [24] were grown in LB medium at
30 �C, with aeration. The recombinant E. coli BL21 (DE3)
(pProPlnHis6) expressing the protealysin gene [45] and con-
trol recombinant E. coli BL21 (DE3) (pET23b) not carrying
the protealysin gene [45] were grown in LB medium at
37 �C, with aeration times specified as appropriate.
The cell line Balb 3T3-SV40 (SV40 transformed fibro-
blasts 3T3) was obtained from the Russian Cell Culture
Collection (Institute of Cytology, St Petersburg, Russia).
Cells were grown on glass cover slips in culture dishes in
the antibiotic-free DMEM (Biolot, Moscow, Russia) con-
taining 10% fetal bovine serum (Sigma) in a incubator at
37 �C supplemented with 5% CO2 for the time required to
form a monolayer (� 48 h).
Invasion assay
Bacteria were grown as described above until the actinase
activity of their extracts could be detected [26,28]. Thirty
minutes before the experiment, FITC (1 mgÆmL)1 of the
bacterial suspension) was added to the bacterial culture to
visualize bacteria. Bacteria were pelleted at 9600 g for
10 min; the pellets were washed with DMEM and added to
the host cells in a fresh portion of DMEM. The host cells
and bacteria were co-cultivated at 37 �C in 5% CO2 for the
times indicated as appropriate.
Rabbit polyclonal antibodies against protealysin precur-
sor were produced by Biotest Systems Ltd (Moscow,
Russia) in accordance with a standard protocol. For
immunization, recombinant protealysin precursor carrying
mutation Glu113 to Ala (inhibiting the precursor matura-
tion) and C-terminal His6-tag was purified as described by
Gromova et al. [45]. To visualize protealysin, 3T3-SV40
cells infected with S. proteamaculans 94, E. coli BL21
(DE3) (pProPlnHis6) expressing the protealysin gene or
control recombinant E. coli BL21 (DE3) (pET23b) that did
not carry the protealysin gene were washed with NaCl ⁄Pi,
fixed with 3.7% formaldehyde for 10 min and incubated
with 0.1% Triton X-100 for 5 min. After washing three
times with NaCl ⁄Pi, the samples were incubated in 1%
BSA for 30 min, and stained with polyclonal rabbit anti-
protealysin antibodies dissolved in 1% BSA, for 18 h at
4 �C. The samples were then washed with 0.05% Tween in
NaCl ⁄Pi three times and incubated for 1 h at 4 �C with
Alexa647-conjugated anti-rabbit IgG secondary serum
(Jackson ImmunoResearch Inc., Bar Harbor, ME, USA).
To visualize actin cytoskeleton, the antibody-stained sam-
ples were washed with 0.05% Tween in NaCl ⁄Pi three times
and with once with NaCl ⁄Pi and incubated with rhoda-
mine–phalloidin for 15 min at 37 �C in the dark. After
washing with NaCl ⁄Pi, the samples were mounted in the
mounting medium and analyzed with a confocal laser scan-
ning microscope.
Protein preparations
Rabbit skeletal muscle actin was isolated from acetone
dried muscle powder according to a standard procedure
[56]. G-actin was stored in buffer G (0.2 mM ATP, 0.1 mM
CaCl2, 5 mM Tris–HCl, pH 7.5, 0.02% NaN3) on ice for
1 week or as 0.2-mL aliquots (0.5–1.0 mgÆmL)1) frozen at
)20 �C for a single use. ATP-Ca-G-actin was transformed
into ATP-Mg-G-actin by 3–5 min of incubation with
0.2 mM EGTA ⁄ 0.1 mM MgCl2 at room temperature. Actin
labelled with N-(1-pyrenyl)iodoacetamide at Cys374 was
prepared as described previously [57]. Pyrenyl-labelled actin
was lyophilized in the presence of 2 mM sucrose and stored
at )70 �C. Before use, the lyophilized pyrenyl-labelled actin
was dissolved in buffer G and dialyzed against the same
buffer overnight. Actin labelled with IAEDANS at Cys374
was prepared as described previously [58]. To stabilize actin
with AlF�4 , G- or F-actin (24 lM) was incubated with 1 mM
ATP and 5 mM NaF for 10 min, which followed by the
addition of 1 mM AlCl3 [40]. The concentration of G-actin
was determined spectrophotometrically using an absorption
coefficient of 0.63 mLÆmg)1Æcm)1 at 290 nm [40] or by the
microbiuret method [59].
Protealysin was purified from E. coli BL21 (DE3)
(pProPlnHis6) cell lysate by sequential metal chelate affinity
chromatography on Ni2+-NTA-agarose (Qiagen, Valencia,
CA, USA) and gel filtration on a Superdex 75 HR 10 ⁄ 30column (Amersham Biosciences, Piscataway, NJ, USA) as
described previously [45]. Protealysin was stored in 25 mM
Tris–HCl buffer (pH 8.0) at 4 �C.
O. Tsaplina et al. Cleavage of F-actin with protealysin
FEBS Journal 279 (2012) 264–274 ª 2011 The Authors Journal compilation ª 2011 FEBS 271

Limited proteolysis assay
To estimate the susceptibility of F-actin to protealysin,
actin was polymerized under different conditions known to
affect the polymer structure [36]. G-actin (0.5 mgÆmL)1) in
buffer G containing Ca2+ as a tightly-bound cation (Ca-G-
actin) was polymerized with 0.1 M KCl for 2 h at room
temperature. G-actin containing Mg2+ as a tightly bound
cation (Mg-G-actin) was polymerized with 0.1 M KCl or
2 mM MgCl2 for 2 h at room temperature. F-actin was
incubated with protealysin (0.3 mgÆmL)1 in 25 mM Tris–
HCl, pH 8.0) at various enzyme ⁄protein mass ratios at
22 �C. At different time points, the digestion was stopped
by the addition of an equal volume of the electrophoresis
sample buffer containing 4% SDS, 125 mM Tris–HCl (pH
6.8) followed by 3 min of boiling. The digestion products
were analyzed by SDS ⁄PAGE [60]. The actinase activity
was determined by the appearance of specific actin frag-
ments. The small (5 kDa) product of the cleavage was not
seen on the gels.
To determine the ability of the protealysin-containing
bacterial extracts to cleave actin, bacteria were grown as
described above (usually for 18–24 h) and harvested by cen-
trifugation at 9600 g for 10 min. The pellets were resus-
pended in buffer G, and the bacteria were lysed by five
cycles of freezing and thawing. The lysates (bacterial
extracts) were clarified by centrifugation at 9600 g for 10
min. F-actin in the corresponding buffer (0.5 mgÆmL)1) was
mixed with an equal volume of the clarified lysate and incu-
bated for 2 h at room temperature or for 18 h at 4–6 �C.The reaction was stopped by the addition of the electropho-
resis sample buffer followed by 3 min of boiling. The diges-
tion products were analyzed by SDS ⁄PAGE as described
above.
ATP hydrolysis measurements
Protealysin-cleaved F-actin or Mg-G-actin, polymerized for
40 min to steady-state, were incubated at 22 �C. In aliqu-
ots of the F-actin solutions withdrawn after various time
intervals, the ATPase reaction was quenched by the addi-
tion of an equal volume of 0.6 M ice-cold perchloric acid,
precipitated protein was removed by centrifugation, and
released Pi was determined by the Malachite green method
[61].
Fluorescence microscopy
Cell samples were examined under a confocal scanning
microscope (Leica TCS SL; Leica Microsystems, Wetzlar,
Germany) using the argon ion (488 nm; green fluorescence)
and helium ⁄ neon (532 nm; red fluorescence; 633 nm, blue
fluorescence) laser system to visualize the FITC-stained
bacteria, rhodamine–phalloidin-stained cytoskeleton and
protealysin, respectively.
Fluorescence and light-scattering measurements
All fluorescence and light-scattering measurements were
performed in a Fluorat-02-Panorama spectrofluorimeter
(Lumex, St Petersburg, Russia). The fluorescence intensity of
pyrenyl-labelled actin was monitored at 407 nm after excita-
tion at 365 nm [35]. The intensity of light scattered at 90� wasmeasured at 350 nm. To visualize AEDANS-labelled fluores-
cence peptides, gels were photographed in UV light before
staining.
Acknowledgements
We thank Dr Ilya Nevzorov for discussions and his
valuable help in the preparation of the manuscript, as
well as the anonymous reviewers for their helpful
comments. The work was supported by the Russian
Foundation for Basic Research grants 11-04-00393 and
09-04-00734 and by the Program for Molecular and
Cell Biology of the Russian Academy of Sciences.
References
1 Wada S, Matsunaga S, Saito S, Fusetani N & Watabe
S (1998) Actin-binding specificity of marine macrolide
toxins, mycalolide B and kabiramide D. J Biochem 123,
946–952.
2 Chen YH, Chen SH, Jong A, Zhou ZY, Li W, Suzuki
K & Huang SH (2002) Enhanced Escherichia coli inva-
sion of human brain microvascular endothelial cells is
associated with alternations in cytoskeleton induced by
nicotine. Cell Microbiol 4, 503–514.
3 Fenteany G & Zhu S (2003) Small-molecule inhibitors
of actin dynamics and cell motility. Curr Top Med
Chem 3, 593–616.
4 Gouin E, Welch MD & Cossart P (2005) Actin-based
motility of intracellular pathogens. Curr Opin Microbiol
8, 35–45.
5 Favoreel HW & Enquist LW& (2007) Actin and Rho
GTPases in herpesvirus biology. Trends Microbiol 15,
426–433.
6 Adam T, Giry M, Boquet P & Sansonetti P (1996)
Rho-dependent membrane folding causes Shigella entry
into epithelial cell. EMBO J 15, 3315–3321.
7 Stebbins CE & Galan JE (2001) Structural mimicry in
bacterial virulence. Nature 412, 701–705.
8 Huang Z, Sutton SE, Wallenfang AJ, Orchard RC, Wu
X, Feng Y, Chai J & Alto NM (2009) Structural
insights into host GTPase isoform selection by a family
of bacterial GEF mimics. Nat Struct Mol Biol 16,
853–860.
9 Bourdet-Sicard R, Rudiger M, Jockusch BM, Gounon
P, Sansonetti PJ & Nhieu GT (1999) Binding of the
Shigella protein IpaA to vinculin induces F-actin
depolymerization. EMBO J 18, 5853–5862.
Cleavage of F-actin with protealysin O. Tsaplina et al.
272 FEBS Journal 279 (2012) 264–274 ª 2011 The Authors Journal compilation ª 2011 FEBS

10 Bierne H, Gouin E, Roux P, Caroni P, Yin HL &
Cossart P (2001) A role for cofilin and LIM kinase in
Listeria-induced phagocytosis. J Cell Biol 155, 101–112.
11 Dai S, Sarmiere PD, Wiggan O, Bamburg JR & Zhou
D (2004) Efficient Salmonella entry requires activity
cycles of host ADF and cofilin. Cell Microbiol 6, 459–
471.
12 Athman R, Fernandez M-I, Gounon P, Sansonetti P,
Louvard D, Philpott D & Robine S (2005) Shigella
flexneri infection is dependent on villin in the mouse
intestine and in primary cultures of intestinal epithelial
cells. Cell Microbiol 7, 1109–1116.
13 Selbach M & Backert S (2005) Tyrosine-phosphorylated
bacterial effector proteins: the enemies within. Trends
Microbiol 13, 181–189.
14 Morton WM, Ayscough KR & McLaughlin PJ (2000)
Latrunculin alters the actin-monomer subunit interface
to prevent polymerization. Nat Cell Biol 2, 376–378.
15 Klenchin VA, Allingham JS, King R, Tanaka J, Marri-
ott G & Rayment I (2003) Trisoxazole macrolide toxins
mimic the binding of actin-capping proteins to actin.
Nat Struct Biol 10, 1058–1063.
16 Allingham JS, Klenchin VA & Rayment I (2006) Actin-
targeting natural products: structures, properties and
mechanisms of action. Cell Mol Life Sci 63, 2119–2134.
17 Margarit SM, Davidson W, Frego L & Stebbins CE
(2006) A steric antagonism of actin polymerization by a
salmonella virulence protein. Structure 14, 1219–1229.
18 Barth H & Aktories K (2011) New insights into the
mode of action of the actin ADP-ribosylating virulence
factors Salmonella enterica SpvB and Clostridium botu-
linum C2 toxin. Eur J Cell Biol 90, 944–950.
19 Kudryashov DS, Cordero CL, Reisler E & Satchell KJ
(2008) Characterization of the enzymatic activity of the
actin cross-linking domain from the Vibrio cholerae
MARTX Vc toxin. J Biol Chem 283, 445–452.
20 Zhou D, Mooseker MS & Galan JE (1999) Role of the
S. typhimurium actin-binding protein SipA in bacterial
internalization. Science 283, 2092–2095.
21 Lilic M, Galkin VE, Orlova A, VanLoock MS, Egel-
man EH & Stebbins CE (2003) Salmonella SipA poly-
merizes actin by stapling filaments with nonglobular
protein arms. Science 301, 1918–1921.
22 Khaitlina SY, Smirnova TD & Usmanova AM (1988)
Limited proteolysis of actin by a specific bacterial
protease. FEBS Lett 228, 172–174.
23 Bozhokina ES, Khaitlina SY & Adam T (2008)
Grimelysin, a novel metalloprotease from Serratia
grimesii, is similar to ECP32. Biochem Biophys Res
Commun 367, 888–892.
24 Demidyuk IV, Kalashnikov AE, Gromova TY, Gasa-
nov EV, Safina DR, Zabolotskaya MV, Rudenskaya
GN & Kostrov SV (2006) Cloning, sequencing, expres-
sion, and characterization of protealysin, a novel
neutral proteinase from Serratia proteamaculans
representing a new group of thermolysin like proteases
with short N-terminal region of precursor. Protein Expr
Purif 47, 551–561.
25 Demidyuk IV, Gromova TY, Polyakov KM, Melik-Ad-
amyan WR, Kuranova IP & Kostrov SV (2010) Crystal
structure of protealysin precursor: insights into propep-
tide function. J Biol Chem 285, 2003–2013.
26 Tsaplina OA, Eferemova TN, Kever LV, Komissarchik
YY, Demidyuk IV, Kostrov SV & Khaitlina SY (2009)
Probing for actinase activity of protealysin. Biochemis-
try (Mosc) 74, 648–654.
27 Efremova TN, Ender NA, Brudnaja MS, Komissarchik
YY & Khaitlina SY (2001) Specific invasion of trans-
formed cells by Escherichia coli A2 strain. Cell Biol Int
25, 557–561.
28 Bozhokina ES, Tsaplina OA, Efremova TN, Kever LV,
Demidyuk IV, Kostrov SV, Adam T, Komissarchik YY
& Khaitlina SY (2011) Bacterial invasion of eukaryotic
cells can be mediated by actin-hydrolyzing metallopro-
teases grimelysin and protealysin. Cell Biol Int 35,
111–118.
29 Khaitlina SY, Collins JH, Kusnetsova IM, Pershina
VP, Synakevich IG, Turoverov KK & Usmanova AM
(1991) Physico-chemical properties of actin cleaved with
bacterial protease from E. coli A2 strain. FEBS Lett
279, 49–51.
30 Khaitlina SY, Moraczewska J & Strzelecka-Golas-
zewska H (1993) The actin- actin interactions involving
the N-terminal portion of the DNase-I-binding loop are
crucial for stabilisation of the actin filament. Eur J
Biochem 218, 911–920.
31 Khaitlina SY & Strzelecka-Goaszewska H (2002) Role
of the DNase-I-binding loop in dynamic properties of
actin filament. Biophys J 82, 321–334.
32 Rich SA & Estes JE (1976) Detection of conformational
changes in actin by proteolytic digestion: evidence for a
new monomeric species. J Mol Biol 104, 777–792.
33 Mantulenko VB, Khaitlina SY & Shelud’ko NS (1982)
A high-molecular-weight proteolysis-resistant actin frag-
ment. Biochemistry (Mosc) 48, 61–66.
34 Holmes KC, Popp D, Gebhard W & Kabsch W (1990)
Atomic model of the actin filament. Nature 347, 44–49.
35 Oda T, Iwasa M, Aihara T, Maeda Y & Narita A
(2009) The nature of the globular- to fibrous-actin tran-
sition. Nature 457, 441–445.
36 Strzelecka-Goaszewska H, Wozniak A, Hult T & Lind-
berg U (1996) Effects of the type of divalent cation,
Ca2+ or Mg2+, bound at the high-affinity site and of
the ionic composition of the solution on the structure
of F-actin. Biochem J 316, 713–721.
37 Mornet D & Ue K (1984) Proteolysis and structure of
skeletal muscle actin. Proc Natl Acad Sci USA 81,
3680–3684.
38 Carlier MF, Pantaloni D & Korn ED (1984) Evidence
for an ATP cap at the ends of actin filaments and its
O. Tsaplina et al. Cleavage of F-actin with protealysin
FEBS Journal 279 (2012) 264–274 ª 2011 The Authors Journal compilation ª 2011 FEBS 273

regulation of the F-actin steady state. J Biol Chem 259,
9983–9986.
39 Combeau C & Carlier MF (1988) Probing the mecha-
nism of ATP hydrolysis on F-actin using vanadate and
the structural analogs of phosphate BeF3– and AlF4
–.
J Biol Chem 263, 17429–17436.
40 Pivovarova AV, Khaitlina SY & Levitsky DI (2010)
Specific cleavage of the DNase-I -binding loop dramati-
cally decreases the thermal stability of actin. FEBS J
277, 3812–3822.
41 Muhlrad A, Cheung P, Phan BC, Miller C & Reisler E
(1994) Dynamic properties of actin: structural changes
induced by beryllium fluoride. J Biol Chem 269,
11852–11858.
42 Grimont F & Grimont P (2006) The genus Serratia. In
The Prokaryotes (Dworkin M ed.), pp. 219–244.
Springer, New York.
43 Hertle R & Schwarz H (2004) Serratia marcescens inter-
nalization and replication in human bludder epithelial
cells. BMC Infect Dis 4, 16–30.
44 Bozhokina ES, Vachromova EA, Gamaley IA &
Khaitlina SY (2011) Bacterial invasion as a model to
study the effects of antioxidants N-acethylcysteine and
lipoic acid on the transformed cells. Tsitologiya 53,
927.
45 Gromova TY, Demidyuk IV, Kozlovskiy VI, Kuranova
IP & Kostrov SV (2009) Processing of protealysin
precursor. Biochimie 91, 639–645.
46 Schwyter D, Phillips M & Reisler E (1989) Subtilisin-
cleaved actin: polymerization and interaction with myo-
sin subfragment 1. Biochemistry 28, 5885–5895.
47 Khaitlina SY & Hinssen H (1997) Conformational
changes in actin induced by its interaction with gelsolin.
Biophys J 73, 929–937.
48 Wawro B, Khaitlina SY, Galinska-Rakoczy A &
Strzelecka-Golaszewska H (2005) Role of DNase-I-
binding loop in myosin subfragment 1-induced actin
polymerization. Implications to the polymerization
mechanism. Biophys J 88, 2883–2896.
49 Moraczewska J, Gruszczyncka-Bielaga J, Redowicz MJ,
Khaitlina SY & Strzelecka-Golaszewska H (2004) The
DNase-I-binding loop of actin may play a role in the
regulation of actin-myosin interaction by tropomyo-
sin ⁄ troponin. J Biol Chem 279, 31197–31204.
50 Narita A, Oda T & Maeda Y (2011) Structural basis
for the slow dynamics of the actin filament pointed end.
EMBO J 30, 1230–1237.
51 Orlova A & Egelman EH (1992) Structural basis for the
destabilization of F-actin by phosphate release follow-
ing ATP hydrolysis. J Mol Biol 227, 1043–1053.
52 Cossart P & Sansonetti PJ (2004) Bacterial invasion: the
paradigms of enteroinvasive pathogens. Science 304,
242–248.
53 Khaitlina SY & Strzelecka-Golaszewska H (2003)
Effects of tropomyosin on dynamics of actin filaments.
Biophys J 84, 480a.
54 Matveyev VV, Usmanova AM, Morozova AB & Khait-
lina SY (1996) Purification and characterization of the
proteinase ECP-32 from Escherichia coli A2 strain.
Biochim Biophys Acta 1296, 55–62.
55 Morozova AV, Khaitlina SY & Malinin AY (2011)
Heat shock protein DnaK – substrate of actin-specific
bacterial protease ECP32. Biochemistry (Mosc) 76,
455–461.
56 Spudich JA & Watt S (1971) The regulation of rabbit
skeletal muscle contraction. I. Biochemical studies of
the interaction of the tropomyosin troponin complex
with actin and the proteolytic fragments of myosin.
J Biol Chem 246, 4866–4871.
57 Kouyama T & Mihashi K (1981) Fluorimetry study of
N-(1-pyrenyl)iodoacetamide-labeled F-actin. Local
structural change of actin protomer both on polymeri-
zation and on binding of heavy meromyosin. Eur J
Biochem 114, 33–48.
58 Strzelecka-Goaszewska H, Moraczewska J, Khaitlina S
& Mossakowska M (1993) Localization of the tightly
bound divalent-cation-dependent and nucleotide-depen-
dent conformation changes in G-actin using limited
proteolytic digestion. Eur J Biochem 211, 731–742.
59 Itzhaki RF & Gill DM (1964) A Micro-biuret method
for estimating proteins. Anal Biochem 9, 401–410.
60 Laemmli UK (1970) Cleavage of structural proteins
during the assembly of the head of bacteriophage T4.
Nature 227, 680–685.
61 Kodama T, Fukui K & Kometani K (1986) The initial
phosphate burst in ATP hydrolysis by myosin and sub-
fragment-1 as studied by a modified Malachite green
method for determination of inorganic phosphate.
J Biochem 99, 1465–1472.
Supporting information
The following supplementary material is available:
Fig. S1. Cytoskeleton rearrangements accompanying
the invasion of 3T3-SV40 cells with the protealysin-
producing bacteria.
This supplementary material can be found in the
online version of this article.
Please note: As a service to our authors and readers,
this journal provides supporting information supplied
by the authors. Such materials are peer-reviewed and
may be re-organized for online delivery, but are not
copy-edited or typeset. Technical support issues arising
from supporting information (other than missing files)
should be addressed to the authors.
Cleavage of F-actin with protealysin O. Tsaplina et al.
274 FEBS Journal 279 (2012) 264–274 ª 2011 The Authors Journal compilation ª 2011 FEBS




















