
Ecophysiology of the filamentous Alphaproteobacterium Meganema perideroedes in activated sludge
12
Ecophysiology of the filamentous Alphaproteobacterium Meganema perideroedes in activated sludge Caroline Kragelund, Jeppe Lund Nielsen, Trine Rolighed Thomsen, Per Halkjær Nielsen * Section of Environmental Engineering, Department of Life Sciences, Aalborg University, Sohngaardsholmsvej 57, DK-9000 Aalborg, Denmark Received 18 June 2004; received in revised form 15 February 2005; accepted 8 March 2005 First published online 25 March 2005 Abstract A comprehensive study of the ecophysiology of the filamentous Meganema perideroedes affiliated to the Alphaproteobacteria, pos- sessing a ‘‘Nostocoida limicola Type II’’ filamentous morphology was conducted. This morphotype often causes serious bulking problems in activated sludge wastewater treatment plants, and hardly anything is known about its physiology. The study was carried out by applying a suite of in situ methods in an industrial activated sludge treatment plant with excessive growth of this species. The experiments revealed a very versatile organism able to take up a large variety of organic substrates under aerobic conditions. It had a remarkably high storage capacity forming polyhydroxyalkanoates from most substrates tested. When nitrate was present as e-accep- tor, the number of substrates to be consumed by M. perideroedes was more restricted compared to aerobic conditions. With nitrite as e-acceptor, only acetate and glucose among the substrates tested could be assimilated and used for storage and possibly growth. This indicated that M. perideroedes might be able to denitrify under certain conditions, which is unusual for filamentous bacteria in acti- vated sludge. No substrate uptake or storage was seen under anaerobic conditions. M. perideroedes was relatively hydrophobic, compared to other filamentous bacteria and microcolonies present in the sludge, indicating the presence of a hydrophobic sheath. Several excreted surface-associated exoenzymes were detected in the sludge, but M. perideroedes never showed any activity, except once after a breakdown in the production facility. This confirmed that M. perideroedes mainly grows on soluble substrates. Based on the studies of the ecophysiology of M. perideroedes, potential control strategies are suggested. Ó 2005 Federation of European Microbiological Societies. Published by Elsevier B.V. All rights reserved. Keywords: Meganema perideroedes; Microautoradiology; Ecophysiology; Activated sludge 1. Introduction Bulking and foaming due to excessive growth of fila- mentous bacteria cause problems in many activated sludge wastewater treatment plants worldwide. More than 30 different filamentous bacterial types have been described on the basis of morphology [1,2], but the true identity of many of these filamentous microorganisms is still unknown. Recently, filamentous Alphaproteobacte- ria have been detected and found in substantial quanti- ties in several activated sludge plants. Filamentous bacteria belonging to this group of Proteobacteria have never been described before. Three species named Can- didatus Alysiosphaera europaea, Candidatus Monilib- acter batavus and Candidatus Alysiomicrobium bavaricum are described, but do not yet exist in pure culture [3,4]. Candidatus Alysiosphaera europaea grows only in coexistence with yeast, while Candidatus Moni- libacter batavus and Candidatus Alysiomicrobium bavaricum could not be cultivated, and only partial 16S rDNA sequences exist [3,4]. So far, only one species has been isolated and characterized in detail [4,5], and a new genus (Meganema) and the species name of Mega- nema perideroedes were proposed. Specific oligonucleotide 0168-6496/$22.00 Ó 2005 Federation of European Microbiological Societies. Published by Elsevier B.V. All rights reserved. doi:10.1016/j.femsec.2005.03.002 * Corresponding author. Tel.: +45 96358503; fax: +45 96350558. E-mail address: [email protected] (P.H. Nielsen). www.fems-microbiology.org FEMS Microbiology Ecology 54 (2005) 111–122
Transcript of Ecophysiology of the filamentous Alphaproteobacterium Meganema perideroedes in activated sludge

www.fems-microbiology.org
FEMS Microbiology Ecology 54 (2005) 111–122
Ecophysiology of the filamentous Alphaproteobacterium Meganemaperideroedes in activated sludge
Caroline Kragelund, Jeppe Lund Nielsen, Trine Rolighed Thomsen, Per Halkjær Nielsen *
Section of Environmental Engineering, Department of Life Sciences, Aalborg University, Sohngaardsholmsvej 57, DK-9000 Aalborg, Denmark
Received 18 June 2004; received in revised form 15 February 2005; accepted 8 March 2005
First published online 25 March 2005
Abstract
A comprehensive study of the ecophysiology of the filamentousMeganema perideroedes affiliated to the Alphaproteobacteria, pos-
sessing a ‘‘Nostocoida limicola Type II’’ filamentous morphology was conducted. This morphotype often causes serious bulking
problems in activated sludge wastewater treatment plants, and hardly anything is known about its physiology. The study was carried
out by applying a suite of in situ methods in an industrial activated sludge treatment plant with excessive growth of this species. The
experiments revealed a very versatile organism able to take up a large variety of organic substrates under aerobic conditions. It had a
remarkably high storage capacity forming polyhydroxyalkanoates from most substrates tested. When nitrate was present as e-accep-
tor, the number of substrates to be consumed byM. perideroedes was more restricted compared to aerobic conditions. With nitrite as
e-acceptor, only acetate and glucose among the substrates tested could be assimilated and used for storage and possibly growth. This
indicated that M. perideroedes might be able to denitrify under certain conditions, which is unusual for filamentous bacteria in acti-
vated sludge. No substrate uptake or storage was seen under anaerobic conditions. M. perideroedes was relatively hydrophobic,
compared to other filamentous bacteria and microcolonies present in the sludge, indicating the presence of a hydrophobic sheath.
Several excreted surface-associated exoenzymes were detected in the sludge, but M. perideroedes never showed any activity, except
once after a breakdown in the production facility. This confirmed thatM. perideroedesmainly grows on soluble substrates. Based on
the studies of the ecophysiology of M. perideroedes, potential control strategies are suggested.
� 2005 Federation of European Microbiological Societies. Published by Elsevier B.V. All rights reserved.
Keywords: Meganema perideroedes; Microautoradiology; Ecophysiology; Activated sludge
1. Introduction
Bulking and foaming due to excessive growth of fila-mentous bacteria cause problems in many activated
sludge wastewater treatment plants worldwide. More
than 30 different filamentous bacterial types have been
described on the basis of morphology [1,2], but the true
identity of many of these filamentous microorganisms is
still unknown. Recently, filamentous Alphaproteobacte-
ria have been detected and found in substantial quanti-
0168-6496/$22.00 � 2005 Federation of European Microbiological Societies
doi:10.1016/j.femsec.2005.03.002
* Corresponding author. Tel.: +45 96358503; fax: +45 96350558.
E-mail address: [email protected] (P.H. Nielsen).
ties in several activated sludge plants. Filamentous
bacteria belonging to this group of Proteobacteria have
never been described before. Three species named Can-
didatus Alysiosphaera europaea, Candidatus Monilib-
acter batavus and Candidatus Alysiomicrobium
bavaricum are described, but do not yet exist in pure
culture [3,4]. Candidatus Alysiosphaera europaea grows
only in coexistence with yeast, while Candidatus Moni-
libacter batavus and Candidatus Alysiomicrobium
bavaricum could not be cultivated, and only partial
16S rDNA sequences exist [3,4]. So far, only one specieshas been isolated and characterized in detail [4,5], and a
new genus (Meganema) and the species name of Mega-
nema perideroedeswere proposed. Specific oligonucleotide
. Published by Elsevier B.V. All rights reserved.

112 C. Kragelund et al. / FEMS Microbiology Ecology 54 (2005) 111–122
probes were designed for in situ detection by fluo-
rescence in situ hybridization (FISH) [5]. Some of these
filamentous Alphaproteobacteria resemble the morpho-
type ‘‘Nostocoida limicola’’ Type II [6], and this morpho-
type is common in activated sludge wastewater
treatment plants. A recent FISH survey estimates thatup to 35% of all bulking incidences in industrial waste-
water treatment plants are caused by filamentous Alpha-
proteobacteria, and foaming problems have also been
observed [7].
In order to control filamentous bacteria in activated
sludge treatment plants not only the identity, but also
knowledge about the physiology and ecology of the bac-
teria is of importance. Little is known about the physi-ology of filamentous Alphaproteobacteria and the
morphotype ‘‘Nostocoida limicola Type II’’. A few phys-
iological characteristics of M. perideroedes are known
from the strains maintained on R2A media [5]. Growth
was not well supported on a mineral, salt and vitamin
medium supplemented with carbon sources, limiting a
detailed physiological characterization of the pure cul-
tures. The bacteria are obligate aerobic, oxidase and cat-alase-positive [5]. Most isolates stained Gram-negative,
although one strain was Gram-variable as has also been
observed in the morphotype Nostocoida limicola Type II
in activated sludge. A few investigations of the ecophys-
iology of gene probe-defined M. perideroedes have been
conducted [8]. The bacteria consumed acetate and glu-
cose under aerobic conditions and seemed to have an
unusually high substrate uptake capability. Further-more, they were found to have a high substrate affinity
for acetate (an apparent Ks of 2.4 lM). A high substrate
affinity is assumed to be a general characteristic for most
filamentous bacteria in activated sludge [1,2,9] and to-
gether with relatively low substrate uptake rates and
storage abilities, compared to floc-formers, these kinetic
properties have been used to explain the outcome of the
competition between filamentous and floc-forming bac-teria in different configurations of wastewater treatment
plants [1,2,9]. However, recent studies of the ecophysiol-
ogy of filamentous bacteria suggest that it is difficult or
impossible to make such general statements. For exam-
ple, the filamentous Microthrix parvicella is adapted to
specific substrates (lipids) and is able to take up sub-
strates under anaerobic conditions, properties which
are different from most other known bacteria, includingthe floc-formers [10]. Also Thiothrix rely on oxidation of
reduced sulfur compounds in addition to heterotrophic
growth [11]. Based on this new knowledge, our hypoth-
esis is that many of the troublesome filamentous bacte-
ria in activated sludge have a unique physiology and
ecology reflecting specific conditions in various activated
sludge systems. Thus, if such knowledge about the eco-
physiology of the most important filamentous bacteria iscombined with detailed knowledge about the process
conditions, efficient control measures may be developed.
M. perideroedes has been responsible for severe bul-
king in industrial wastewater treatment plants and is
present in many other treatment plants. In this study,
we wanted to investigate the ecophysiology of this inter-
esting filamentous Alphaproteobacterium in detail and
learn more about its physiology, eventually to find effi-cient control measures.
A number of in situ methods have been applied in
combination with FISH. Microautoradiography
(MAR) was applied to provide information regarding
substrate preference and e-acceptor conditions [8,12].
The ability of bacteria to convert carbon sources under
defined e-acceptor conditions into storage polymers, e.g.
polyhydroxyalkanoates (PHAs), is a potential advan-tage in a dynamic environment such as activated sludge.
Therefore, the PHA content in single cells was quanti-
fied using the Nile blue stain, where the fluorescence
intensity is proportional to the PHA content of the cells
[13,14]. The presence of exoenzymes can provide infor-
mation about the preferred substrates (dissolved low
molecular substrates or macromolecules which require
a hydrolytic degradation), or it can indicate unfavour-able conditions such as phosphorus limitation [15]. Exo-
enzymes located on the cell surfaces can be visualized in
the sludge by applying Enzyme-Labeled Fluorescence
substrates (ELF-97�) [10,16]. Finally, also the cell sur-
face hydrophobicity, which is of major importance in
floc formation and substrate adsorption [10], can be
determined by Microsphere Adhesion to Cells, MAC
[17].
2. Materials and methods
2.1. Activated sludge
The experiments were carried out with activated
sludge from an industrial wastewater treatment plant(WTP) in Grindsted, Denmark. The WTP is operated
at 15–20 �C with a sludge age of 10–12 days. The load
is 4200–5800 kgCOD day�1 with the following concen-
trations: COD, 5.8–8.0 g l�1; total N, 0.1–0.2 g l�1; total
P, 0.02–0.05 g l�1. Most of the COD is dissolved and
consists mainly of acetate, ethanol, propanol, glucose,
and other organic compounds, but little information is
available. Soluble sulfides are present at certain times.The salinity is high (5–7 g l�1) with a conductivity of
15–20 mS cm�1. The treatment plant mainly removes
organic matter and receives wastewater from a pharma-
ceutical plant. Nitrogen is added in the form of ammo-
nium to ensure sufficient nitrogen for growth. However,
partial nitrification/denitrification is performed during
the summertime. The plant is an aerobic, completely
mixed activated sludge system without any selector.The activated sludge was collected and stored at 4 �Cthe day before the experiments were performed. One

C. Kragelund et al. / FEMS Microbiology Ecology 54 (2005) 111–122 113
hour before the experiments were conducted, it was
acclimatized to room temperature (20–22 �C). The
sludge had a content of suspended solids (SS) of 3–
4 gSS l�1. The volatile suspended solids (VSS) were
75 ± 2% of SS. Before use, the sludge was diluted to
1 gSS l�1 with filtered nitrate and nitrite free superna-tant from the activated sludge.
2.2. Identification
The filamentous bacteria were morphologically iden-
tified using the Eikelboom classification system [6].
Furthermore, fluorescence in situ hybridization (FISH)
was applied with 16S and 23S rRNA-targeted nucleicacid probes as listed in probeBase [18], targeting the
groups of the Alpha-(ALF968), Beta-(BET42a), and
Gamma-proteobacteria (GAM42a), and Cytophaga Fla-
vobacterium group of the Bacteroidetes (CF319a). Also,
species-specific probes for M. perideroedes (Meg983
mixed with Meg1028) and Thiothrix spp. (TNI) were
used [5,19]. Development, optimization and determina-
tion of applied formamide concentration for the geneprobes Meg983 and Meg1028 was performed by
Thomsen et al. (accepted). The sequence of Meg983:
5 0-CGG GAT GTC AAA AGG TGG-3 0 and for probe
Meg1028: 5 0-CTG TCA CCG AGT CCC TTG C-3 0.
Hybridization and washing buffer were made according
to [20]. Oligonucleotides were labeled with 5(6)-car-
boxyfluorescein-N-hydroxy-succinimide ester (FLUOS)
or with the sulfoindocyanine dyes (Cy3 and Cy5)(Thermohybaid Interactive, Ulm, Germany). A Confo-
cal laser scanning microscope (LSM 510-meta, Carl
Zeiss, Oberkochen, Germany) equipped with a UV
laser (351 and 364 nm), an Ar ion laser (458 and
488 nm), and two HeNe lasers (543 and 633 nm) were
used to record fluorescent signals from the various
probes.
Combinations of MAR-FISH, PHA-FISH, ELF-FISH and MAC-FISH were used for the studies of in
situ physiology. All above-mentioned methods were
examined by confocal laser scanning microscopy
(CLSM) prior to hybridization with gene probes except
MAR-FISH. MAR-FISH hybridization with gene
probes was performed before applying the photographic
emulsion and thus before CLSM examination. Positions
of interest were recorded by an automatic stage control-ler and digital images were recorded. Subsequently, the
fresh samples used for ELF and MAC were fixed in
4% paraformaldehyde (PFA) for 3 h on the slide, before
the FISH procedure was conducted [20]. The stage con-
trol enabled relocation of the recorded microscopic field.
The PHA-FISH combination was slightly modified to
remove excess Nile blue stain prior to FISH. These
slides were pre-treated for 30 min. with 96% ethanol at4 �C, following the normal FISH procedure. The
MAC-FISH and ELF-FISH slides were washed in
dH2O prior to dehydration to remove excess micro-
spheres and precipitates from phosphate-buffered saline
medium (PBS), respectively.
2.3. Microautoradiography and MAR-FISH
The microautoradiography experiments were per-
formed using 3H-labeled and14C-labeled organic com-
pounds and 14C-labeled bicarbonate. The procedure,
which includes incubation with radiolabeled substrates
under defined e-acceptor conditions, fixation, and
hybridization with gene probes, addition of a radiosen-
sitive emulsion, exposure, processing, and microscopical
investigations, is described in detail elsewhere [11,12,21].Experiments were repeated on different occasions and
incubations were performed on two separate occasions
at least. For analysis of MAR-positive M. perideroedes,
a minimum of 30 filaments were investigated for each
incubation. In brief, two types of studies were con-
ducted. In the first study, various potential substrates
were tested for uptake under aerobic conditions to see
if M. perideroedes was a specialized or general con-sumer. For this, a selection of substrates was chosen,
representing short and long-chained fatty acids, sugars
and amino acids. In the second study, potential use of
different e-acceptors was tested by studying uptake of
the same organic substrates in the filamentous bacteria
under anaerobic conditions with or without nitrate or
nitrite present as e-acceptor. In all experiments, 2 ml di-
luted activated sludge (1 gSS l�1) were transferred toglass serum vials and incubated with a final concentra-
tion of substrate of 2 mM, and the labeled fraction
was 10 lCi per vial. Incubation time was 3 h (five for
bicarbonate). All vials for anaerobic incubations (with
or without nitrate or nitrite) were closed with thick,
gas-tight rubber stoppers and flushed with ultrapure
nitrogen gas prior to incubation. In all anaerobic exper-
iments, a pre-incubation step of 2 h was included withunlabeled organic substrate (2 mM). In this way, only
bacteria able to take up large amounts of substrate un-
der these conditions (for storage or growth) would be
MAR-positive [22]. After the pre-incubation period,
labeled and unlabeled substrate was added to a final
substrate concentration of approximately 2 mM. When
thiosulfate or nitrate was added, a final concentration
of 2 mM was used, while 0.5 mM was used for nitrite.In some experiments, a pre-incubation period of 6 h
with unlabeled acetate was applied. As a control for che-
mography, sludge was pasteurized at 70 �C for 10 min
just before the incubation under defined conditions. In
most experiments filamentous bacteria were assessed as
MAR-positive or MAR-negative by comparing the
number of silver grains on top of the filaments to the
background. In some experiments, quantitative MAR(Q-MAR) was used by enumeration of the silver grains
[8].

114 C. Kragelund et al. / FEMS Microbiology Ecology 54 (2005) 111–122
Radiochemicals [3H]acetate, [D-2-3H]glucose, [9,10(n)-3H]oleic acid, [4,5-3H]leucine, [14C]formate, [3H]glycine,
[14C]bicarbonate, [3H]mannose and [3H]galactose were
purchased from Amersham Biosciences (Amersham Bio-
science, Denmark), [1-14C]pyruvate (Na) and [1-3H]eth-
anol came from American Radiolabeled Chemical Inc.(Bio Nuclear AB, Sweden) and [1-14C]propionate (Na)
and [2,3-3H]butyric acid came from ICN Biochemicals
Inc. (Bie & Berntsen, Denmark).
2.4. Storage capabilities
Lipidic storage granules of M. perideroedes were
investigated by measuring the polyhydroxyalkanoates(PHA) content within individual filamentous bacteria
by Nile blue staining [23]. The ability of M. perideroedes
to form PHA from different carbon substrates under dif-
ferent e-acceptor conditions was examined. Samples
were incubated under similar conditions as in the
MAR experiments without addition of radioactive tra-
cer and with slightly shorter incubation length (2 h).
Prior to the experiment, all samples were aerated for1 h in order to remove easily degradable substrates
otherwise present in the sludge. After incubation, sam-
ples were spread out onto gelatine-coated glass and
stained with Nile blue. All samples were examined with
the CLSM. At least 22 filamentous M. perideroedes in
each sample were investigated, and the average fluores-
cence intensity of PHA-containing granules was ex-
pressed as intensity per filament. The intensity wasmeasured using image analysis software (ImageJ 1.33s,
Rasband W, National Institutes of Health, USA,
http://rsb.info.nih.gov/ij/). For each filament, a segment
of at least 15 lm was analysed by recording maximum
intensity in each horizontal line perpendicular to the fil-
ament image. An average of these measurements (min
100) was calculated for each analysed image of a given
incubation. At least 22 images were examined for eachsubstrate incubation and average fluorescence intensity
and standard deviations were calculated. Student�s t testwas used as statistical tool to evaluate significant PHA
formation compared to the control.
2.5. Enzyme-Labeled Fluorescence (ELF), ELF-FISH
The presence of exoenzyme activity was determinedusing Enzyme-Labeled Fluorescence (ELF-97�, Molec-
ular Probes, Eugene, OR, USA), where substrates upon
enzymatic cleavage form a fluorescent precipitate (exci-
tation 345 nm/emission 530 nm) on the surfaces of
bacteria or microcolonies within flocs [10,16]. All com-
mercially available ELF� 97 enzymes were evaluated
for activity; ELF� 97 esterase substrate (ELF� 97 ace-
tate), ELF� 97 lipase substrate (ELF� 97 palmitate),ELF� 97 b-D-galactosidase substrate (ELF� 97
b-D-galactopyranoside), ELF� 97 b-D-glucuronidase
substrate (ELF� 97 b-D-glucuronide), ELF� 97 chiti-
nase/N-acetylglucosaminidase substrate (ELF� 97
N-acetylglucosaminide; ELF� 97 NAG), and ELF� 97
Endogenous Phosphatase detection kit. Stock solutions
of enzymes were dissolved in methanol or DMSO (chiti-
nase) according to the guidelines of the manufacture.Hundred ll sludge was incubated with approx.
0.2 mM of either substrate and placed in the dark for
2–3 h. Sub-samples were spread out onto gelatine-
coated glass and examined with the CLSM prior to
FISH analysis. The enzyme samples were PFA fixed
and stored in 1 · PBS at 4 �C and could be stored with-
out significant loss in fluorescence intensity for at least 6
months.
2.6. Microsphere Adhesion to Cells, MAC-FISH
Surface properties were investigated using micro-
sphere adhesion to cells where sulfate-modified micro-
spheres rendering hydrophobic characteristics and a
diameter of 0.1 lm were applied [17,24]. Yellow-green
fluorescent polystyrene microspheres with excitationand emission properties (505 nm/515 nm) were used
(Molecular Probes, Eugene, OR, USA). The micro-
spheres were diluted in dH2O to a concentration of
0.02% (w/v) and sonicated for 100 min at 60 W. Five
ll of microspheres and 10 ll sludge were mixed with
100 ll of dH2O and vortexed for two min. Sub-samples
were spread out onto gelatine-coated cover glass and
examined immediately before drying under CLSM priorto FISH.
3. Results
3.1. Identification and description
The industrial wastewater treatment plant investi-gated has suffered from filamentous bulking for several
years with a sludge volume index (SVI) above
200 ml g�1 and often much higher. Three types of fila-
mentous bacteria have been dominating over a period
of several years as determined by FISH, and for the per-
iod of investigation, the numbers of filamentous bacteria
were: M. perideroedes constituted 60–95%, Thiothrix
spp. (5–35%), and bacteria affiliating to the Cytophaga
Flavobacterium group of the Bacteroidetes (5–10%)
(Fig. 1A and B). M. perideroedes grew as slightly coiled
filaments with a cell diameter of approx. 1.5 lm, disc-
shaped cells and a length of often up to 0.5 mm. M.
perideroedes filaments were always Gram-negative, even
though some cells within some filaments occasionally
stained Gram-positive. Some cells in the filaments con-
tained Neisser-positive granules, indicating polyphos-phate inclusions, but most cells were Neisser-negative.

Fig. 1. Identification of the predominating filaments using the Bacterial EUB-probe (green) (A) and a probe specific for the Meganema perideroedes
(red) (B). (C) Microautoradiographic image showing a positive uptake of radiolabeled acetate by the M. perideroedes in the activated sludge
investigated with oxygen as e-acceptor. (D) Shows the superimposing of (A)–(C). Image (E) and (F) show the same microscopic field with a M.
perideroedes filament as bright field and after PHA-staining by Nile blue, respectively. Images (G–I) show a MAC-FISH image of microspheres
adhesion to M. perideroedes. Hydrophobic spheres (green) (H) and FISH image of M. perideroedes (red) (G), and the overlay with a bright field
image of the same microscopic field (I). Scale bar indicates 20 lm.
C. Kragelund et al. / FEMS Microbiology Ecology 54 (2005) 111–122 115
The cells contained PHA granules as determined by Nile
blue staining. Two phenotypes of M. perideroedes were
observed in the sludge based on MAR experiments
(see later). The applied gene probes Meg983 mixed with
Meg1028 targeted both phenotypes in the sludge, so the
difference was mainly based on the differences in sub-strate uptake pattern, where one of the morphotypes
was less versatile than the other. However, it was present
in small amount and could not be distinguished system-
atically by morphology, so it was not investigated in
detail.
3.2. Ecophysiology
3.2.1. Uptake of organic substrates under aerobic
conditions
In order to determine whether M. perideroedes was a
specialized or versatile consumer of organic matter, the
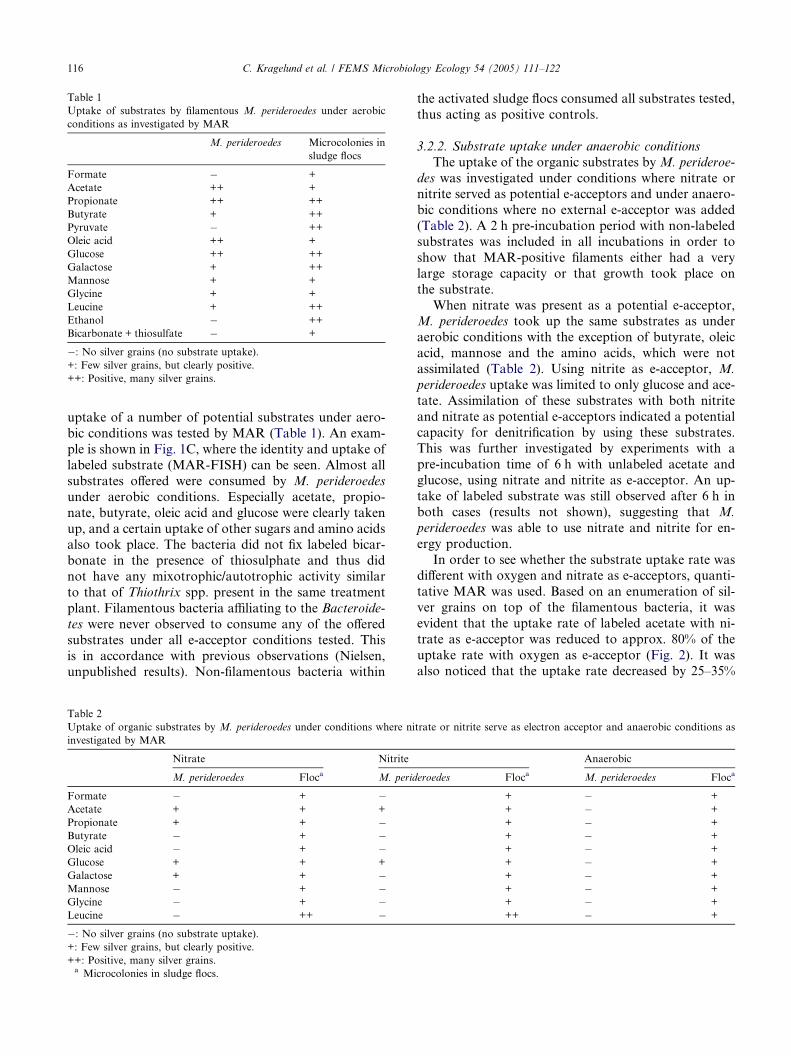
Table 1
Uptake of substrates by filamentous M. perideroedes under aerobic
conditions as investigated by MAR
M. perideroedes Microcolonies in
sludge flocs
Formate � +
Acetate ++ +
Propionate ++ ++
Butyrate + ++
Pyruvate � ++
Oleic acid ++ +
Glucose ++ ++
Galactose + ++
Mannose + +
Glycine + +
Leucine + ++
Ethanol � ++
Bicarbonate + thiosulfate � +
�: No silver grains (no substrate uptake).
+: Few silver grains, but clearly positive.
++: Positive, many silver grains.
116 C. Kragelund et al. / FEMS Microbiology Ecology 54 (2005) 111–122
uptake of a number of potential substrates under aero-
bic conditions was tested by MAR (Table 1). An exam-
ple is shown in Fig. 1C, where the identity and uptake of
labeled substrate (MAR-FISH) can be seen. Almost all
substrates offered were consumed by M. perideroedes
under aerobic conditions. Especially acetate, propio-
nate, butyrate, oleic acid and glucose were clearly taken
up, and a certain uptake of other sugars and amino acidsalso took place. The bacteria did not fix labeled bicar-
bonate in the presence of thiosulphate and thus did
not have any mixotrophic/autotrophic activity similar
to that of Thiothrix spp. present in the same treatment
plant. Filamentous bacteria affiliating to the Bacteroide-
tes were never observed to consume any of the offered
substrates under all e-acceptor conditions tested. This
is in accordance with previous observations (Nielsen,unpublished results). Non-filamentous bacteria within
Table 2
Uptake of organic substrates by M. perideroedes under conditions where ni
investigated by MAR
Nitrate Nitrite
M. perideroedes Floca M. perid
Formate � + �Acetate + + +
Propionate + + �Butyrate � + �Oleic acid � + �Glucose + + +
Galactose + + �Mannose � + �Glycine � + �Leucine � ++ ��: No silver grains (no substrate uptake).
+: Few silver grains, but clearly positive.
++: Positive, many silver grains.a Microcolonies in sludge flocs.
the activated sludge flocs consumed all substrates tested,
thus acting as positive controls.
3.2.2. Substrate uptake under anaerobic conditions
The uptake of the organic substrates by M. perideroe-
des was investigated under conditions where nitrate ornitrite served as potential e-acceptors and under anaero-
bic conditions where no external e-acceptor was added
(Table 2). A 2 h pre-incubation period with non-labeled
substrates was included in all incubations in order to
show that MAR-positive filaments either had a very
large storage capacity or that growth took place on
the substrate.
When nitrate was present as a potential e-acceptor,M. perideroedes took up the same substrates as under
aerobic conditions with the exception of butyrate, oleic
acid, mannose and the amino acids, which were not
assimilated (Table 2). Using nitrite as e-acceptor, M.
perideroedes uptake was limited to only glucose and ace-
tate. Assimilation of these substrates with both nitrite
and nitrate as potential e-acceptors indicated a potential
capacity for denitrification by using these substrates.This was further investigated by experiments with a
pre-incubation time of 6 h with unlabeled acetate and
glucose, using nitrate and nitrite as e-acceptor. An up-
take of labeled substrate was still observed after 6 h in
both cases (results not shown), suggesting that M.
perideroedes was able to use nitrate and nitrite for en-
ergy production.
In order to see whether the substrate uptake rate wasdifferent with oxygen and nitrate as e-acceptors, quanti-
tative MAR was used. Based on an enumeration of sil-
ver grains on top of the filamentous bacteria, it was
evident that the uptake rate of labeled acetate with ni-
trate as e-acceptor was reduced to approx. 80% of the
uptake rate with oxygen as e-acceptor (Fig. 2). It was
also noticed that the uptake rate decreased by 25–35%
trate or nitrite serve as electron acceptor and anaerobic conditions as
Anaerobic
eroedes Floca M. perideroedes Floca
+ � +
+ � +
+ � +
+ � +
+ � +
+ � +
+ � +
+ � +
+ � +
++ � +

Fig. 2. Quantification of labeled acetate uptake in filaments of M.
perideroedes as determined by MAR. Silver grains were enumerated
under conditions with oxygen or nitrate as e-acceptor and with or
without a 2 h pre-incubation with unlabeled acetate.
Fig. 3. Fluorescence intensity distribution of PHA in M. perideroedes
with nitrite as e-acceptor. (A) Control without addition of acetate. (B)
After 2 h incubation with acetate.
C. Kragelund et al. / FEMS Microbiology Ecology 54 (2005) 111–122 117
when a 2 h pre-incubation step with unlabeled substrate
was included (Fig. 2). This indicated that the substrateuptake rate was very high in the beginning of the incu-
bation period, probably showing the difference between
the substrate uptake rate when both storage and growth
took place, and when only growth took place. When
pre-incubation was included for nitrate, the same trend
was observed, although not statistically significant.
The uptake rate of acetate with nitrate as e-acceptor
was still approximately 80% of the rate with oxygen ase-acceptor (Fig. 2).
Anaerobic conditions without nitrate or nitrite pres-
ent showed no uptake of any of the substrates tested,
indicating thatM. perideroedes did not possess an anaer-
obic storage capability or a fermentative metabolism.
A minor subpopulation of M. perideroedes showed
less ability to take up substrates with other e-acceptors
than oxygen (results not shown). No substrates couldbe assimilated with nitrite as e-acceptor, and only propi-
onate could be consumed with nitrate as e-acceptor.
3.2.3. Storage capabilities
In fresh activated sludge samples, all M. perideroedes
filaments had small PHA-containing granules as visual-
ized by Nile blue and FISH (Fig. 1E and F). The fluores-
cence intensity among the individual cells varied,indicating some variation in the PHA content of the
individual cells within a filament. The initial intensity
was low compared to filaments after substrate uptake
(Fig. 3A and B), indicating low levels of PHA in most
cells. A number of substrates were tested for poten-
tial conversion and PHA formation under different
e-acceptor conditions by measuring the increase in fluo-
rescence intensity during 2 h incubation with the sub-
strates. For some substrates, a substantial uptake was
observed, where the individual cells in the filaments were
almost completely filled with PHA granules. There was
some variation between filaments, but with rather simi-
lar PHA content in cells along a certain filament. The
PHA content of the filaments usually showed a normal
distribution after uptake of substrate, as exemplified
by uptake of acetate with nitrite as e-acceptor (Fig. 3B).For some substrates it is likely that the standard deviation
of the PHA content was ascribed tomore than one type of
M. perideroedes, but it was not possible to systematically
assign it to a particular morphotype, since discrimination
between these was very difficult.
The distribution of the PHA content in the filaments
was recorded for all tested substrates and e-acceptor
conditions. The results are shown in Fig. 4A–C. Aerobicconversion of substrates into PHA took place for ace-
tate, propionate, oleic acid, galactose and mannose.
Glucose and leucine were not converted to PHA. With
nitrate as e-acceptor, oleic acid and galactose showed
a very big increase in the PHA content, and to a lesser
extent, but still significant, also acetate, propionate, glu-
cose, mannose and leucine formed PHA. When nitrite
was present as e-acceptor, only acetate and glucoseformed enlarged PHA granules. PHA formation under
anaerobic conditions without nitrate or nitrite was not
observed with any of the carbon sources tested. The

Fig. 4. Relative fluorescence intensity distribution of PHA in M.
perideroedes filaments for all substrates with oxygen (A), nitrate (B), or
nitrite (C) as e-acceptor.
118 C. Kragelund et al. / FEMS Microbiology Ecology 54 (2005) 111–122
difference in fluorescence intensity between control incu-bations and incubations that led to PHA formation was
at least a factor of 3. Using Student�s t test, all substratesbeing positive shown in Fig. 4 had a significantly higher
content of PHA in the cells than the control incubations.
For most substrates, except for glucose and leucine, the
formation of PHA was very similar with oxygen and ni-
trate present as e-acceptors. For some substrates, M.
perideroedes filaments formed PHA only under certaine-acceptor conditions as observed for glucose and leu-
cine with oxygen and nitrate as e-acceptor. These obser-
vations indicate that the filaments used the uptake of
substrate for either direct growth or for another type
of storage compounds not detected by PHA stain, e.g.
glycogen (Fig. 4A–C). Also, the amount of PHA that
was formed depended on e-acceptor conditions, illus-
trated with oleic acid and galactose with oxygen and ni-
trate e-acceptor (Fig. 4A–C), where much more PHA
was formed with nitrate.
When the granule sizes of PHA were measured forM.
perideroedes and Thiothrix sp., it was evident that M.
perideroedes had a much higher PHA content per cell
than Thiothrix sp. The fluorescence intensity of M.
perideroedes PHA granules (from acetate accumulation)
was approx. 3 times higher than the intensity from the
PHA granules of Thiothrix sp. (data not shown), possi-
bly due to the greater cell diameter. M. perideroedes had
2–5 granules in the individual cells, while Thiothrix sp.only had one (data not shown).
3.2.4. Exoenzyme activities
Presence and location of extracellular enzyme activ-
ity in sludge flocs and on the surface of filamentous
bacteria were investigated by enzyme-labeled fluores-
cence assays (Table 3). While microcolonies within
the flocs exhibited enzyme activity in five of the six en-zymes tested, M. perideroedes hardly ever showed any
enzyme activity during the study period. On one occa-
sion, after a shutdown of the industrial production,
approximately 15% of the M. perideroedes filaments
exhibited lipase activity. Some Thiothrix sp. filaments
tested positive in phosphatase and esterase, while fila-
mentous bacteria affiliating to the Bacteroidetes always
showed esterase activity.
3.2.5. Surface properties
Distribution of slightly or strongly hydrophobic sur-
faces in the sludge was investigated by MAC. The dom-
inating M. perideroedes was always relatively
hydrophobic in comparison with the other filaments
present in the sludge as indicated by a high degree of at-
tached hydrophobic microspheres relative to Thiothrix
sp., (Fig. 1G–I). The Bacteroidetes filaments showed
attachments of microspheres, however, in much smaller
amounts than M. perideroedes. Several parts of the
sludge flocs were shown to be relatively hydrophobic,
as many spheres attached to the floc structure, but vari-
ations among microcolonies within single flocs were
detected.
4. Discussion
This study presents the first detailed study of the eco-
physiology of a filamentous bacterium affiliating to the
Alphaproteobacteria, namely M. perideroedes, which
was recently isolated and characterized [5]. However,
very little is known about its physiology, so presentlyno specific control measures exist in full-scale treatment
plants.

Table 3
Excretion of exoenzymes by bacteria present in the sludge as investigated by Enzyme-Labeled Fluorescence, ELF
Esterase Lipase Chitinase Phosphatase Glucuronidase Galactosidase
M. perideroedes � (++) � � � �Thiothrix sp. (++) � � (++) � �Cytophaga-Flavobacterium-group of the Bacteroidetes + � � � � �Flocsa ++ � ++ ++/+++ +++ +++
�: No enzyme activity (not active).
+: Little enzyme activity, but clearly positive.
(++): Occasionally a fraction positive.
++: Clearly positive, enzyme activity.
+++: Very active, high enzyme activity.a Microcolonies in sludge flocs.
C. Kragelund et al. / FEMS Microbiology Ecology 54 (2005) 111–122 119
4.1. Substrate assimilation profile and storage capacity
The probe-defined M. perideroedes consumed almost
all organic substrates tested under aerobic conditions,
thus indicating that this bacterium was a very versatile
heterotrophic organism able to consume a range of
short and long-chain fatty acids, sugars and amino
acids. This versatility is greater than that observed forother filamentous bacteria in activated sludge systems
studied so far [10,11,21,25]. The substrate uptake pat-
tern was generally in agreement with growth patterns
observed by an isolate studied by Levantesi et al. [4].
Whether it is the same species as that investigated in this
study is unclear, as gonidia and rosette formation was
observed with their isolate, but never observed in situ
or with the isolates from Grindsted industrial WWT[5]. Using nitrate as e-acceptor, the number of substrates
utilized by M. perideroedes had diminished compared to
aerobic conditions, but the main groups of substrates
were still assimilated. When nitrite was present, only
acetate and glucose were assimilated, indicating that
storage and/or growth could take place under these con-
ditions. Under strict anaerobic conditions, no substrate
uptake took place, which is also the case for other fila-mentous bacteria in activated sludge, except M. parvi-
cella [10]. This bacterium is known to take up
substrate under anaerobic conditions as a storage com-
pound for later use with oxygen or nitrate as e-acceptor
[10,26].
M. perideroedes had a very great capacity to form a
storage compound, most probably PHA, with both oxy-
gen, nitrate or nitrite present as e-acceptors. It formed atleast 3 times more PHA within the cells than did Thio-
thrix cells in the same sludge as quantified by the Nile
blue staining. Many bacteria are able to accumulate
polymeric material which can serve as a reserve of car-
bon and/or energy during periods of unbalanced growth
due to unfavorable conditions [27]. PHA granules have
also been found in several filamentous morphotypes
from activated sludge [1]. PHA formation has, however,only in one case been reported in extensive amounts in
filamentous organisms – this was found in an unidenti-
fied ‘‘Nostocoida limicola Type II’’-like organism affiliat-
ing to the Alphaproteobacteria growing in a lab-scale
reactor [28]. These bacteria formed PHA with oxygen
and nitrate as potential e-acceptors with acetate, ethanol
and glutamic acid, while no PHA formation was ob-
served using glucose as carbon substrate. Thus, whether
an ability to form large amounts of PHA may be a com-
mon trait among filamentous Alphaproteobacteria re-mains to be investigated.
The quantification of PHA in the filaments showed
variation in the content from filament to filament, indi-
cating a range of activity levels. This is in accordance
with results with quantification of the acetate uptake
in the same gene-probe defined bacterial population
[8]. All cells within a single filament had approximately
the same activity level. This observation correspondsto our findings using PHA as activity measure with some
variation between filaments, but with rather similar
PHA content in cells along a filament.
By comparing the MAR results and the PHA quanti-
fication, it is possible to get more insight into the meta-
bolic pathways involved in the consumption and
transformation of the various substrates. With oxygen
as e-acceptor, most short chain fatty acids tested andsome sugars were converted to PHA granules. With ni-
trate as e-acceptor, the substrates taken up also accumu-
lated into PHA. Glucose was assimilated with both
oxygen, nitrate and nitrite as e-acceptors (as shown by
MAR), but PHA was only formed with nitrate or nitrite
as e-acceptors. In contrast, acetate was converted to
PHA in all cases. One explanation on this could be that
glucose was converted into glycogen because, under aer-obic conditions, this process is more energetically
favourable than conversion to PHA [29]. Unfortunately,
there is no reliable method available to study the content
of glycogen granules at single cell level, so this hypoth-
esis cannot be confirmed. Mannose formed PHA with
nitrate as e-acceptor despite no assimilation of mannose
could be detected by MAR (Table 2). This shows that
M. perideroedes only had a small storage capacity whichwas saturated during the 2 h pre-incubation period with
unlabeled mannose and that the bacterium was not able

120 C. Kragelund et al. / FEMS Microbiology Ecology 54 (2005) 111–122
to grow on this substrate with nitrate as e-acceptor. The
results emphasize, however, that although M. perideroe-
des was physiologically active with both oxygen, nitrate,
and nitrite as e-acceptor, the substrate uptake pattern
and biochemical pathways change significantly upon
change in e-acceptor conditions. Different substrates re-sulted in different quantities of formed PHA under dif-
ferent e-acceptor conditions as illustrated with glucose,
leucine, oleic acid, and galactose (with oxygen and ni-
trate). Interestingly, also the amount of PHA formed
from a specific substrate also depended on e-acceptor
condition, indicating change in metabolic pathways. In
relation to possible control strategies, it is particularly
interesting that the number of substrates became morerestricted when nitrate, and particularly nitrite, were
e-acceptors. This may explain why M. perideroedes seems
not to be common in treatment plants with denitrifica-
tion and suggests that introduction of denitrifying con-
ditions may be a way to limit its growth. A restriction
of certain substrates with nitrate as e-acceptor is also
known from isolated denitrifiers belonging to the genus
Thauera, which are present in activated sludge. T. lin-ealoolentis and T. terpenica grow on sugar compounds
under aerobic conditions, but not under denitrifying
conditions [30].
So far, no filamentous bacteria in activated sludge
have been reported to perform full denitrification to gas-
eous dinitrogen, while several filamentous bacteria are
reported to reduce nitrate to nitrite. Examples are
Microthrix parvicella [31], Thiothrix sp., and Type021N [32]. It is not known whether M. perideroedes per-
forms a full denitrification, but our results suggest that it
may be the case with acetate and glucose as substrates,
because they were able to assimilate labeled acetate after
6 h pre-incubation with unlabeled acetate with nitrate
OR nitRate as e-acceptor, indicating growth or an ex-
treme storage capacity. Furthermore, with nitrate, the
uptake rate of acetate was reduced to approximately80% of the rate with oxygen, which is typical for denitri-
fying bacteria due to the lower energy yield [33]. How-
ever, as several substrates were not consumed with
nitrite as e-acceptor, e.g. propionate and oleic acid, these
may only favour a reduction of nitrate to nitrite.
The less abundant M. perideroedes phenotype or
genotype observed in the activated sludge had a different
physiology, compared to the dominant type. It also hada versatile substrate uptake under aerobic conditions,
but was not active under any other e-acceptor condi-
tions than oxygen. This was evident in both MAR and
PHA experiments. This type could hardly be distin-
guished from the predominant type by morphology
and not by the applied gene probes. As it was present
only in small numbers, it was not studied in detail, but
shows that there are still several filamentous Alphaprote-obacteria that remain to be properly identified and
characterized.
4.2. Exoenzymatic activity and surface properties
Meganema perideroedes did not generally exhibit any
exoenzyme activity as assessed from the ELF assays.
Only in a situation after the factory shutdown, some li-
pase activity was observed, indicating that starvationand atypical conditions could induce excretion of certain
exoenzymes. Phosphate starvation has been described to
promote enhanced phosphatase expression in a biofilm
[15]. In general, we found other types of bacteria
expressing various enzymes within the flocs. It has been
hypothesized that different groups of bacteria express
different enzymes, and, for example, phosphatase in acti-
vated sludge flocs was associated mainly with bacteriaaffiliating with the Cytophaga flavobacterium group of
the Bacteroidetes [34]. Also M. parvicella, which is a spe-
cialized filamentous lipid degrader, excretes lipases on
the cell surface [10]. The lack of exoenzyme production
by M. perideroedes suggests that this bacterium mainly
consumes soluble short-chain substrates without the
need of hydrolysis before uptake. This is in agreement
with its great ability to assimilate a range of substratesand form large PHA granules. The observed potential
for excretion of lipase could be an action for utilizing
otherwise inaccessible fat (triglycerides) under starva-
tion conditions.
Cell surface hydrophobicity of bacteria in activated
sludge influences the location in the sludge floc and
the accessibility of certain substrates. If the bacteria
are very hydrophobic, they will mainly be located withinsludge flocs or they may form foam or scum. A hydro-
phobic sheath can also attract hydrophobic substrates
such as lipids [10]. M. perideroedes form very hydropho-
bic surfaces in both pure culture [5] and under in situ
conditions as assessed by MAC. It is, however, not clear
what the benefit of the hydrophobic sheath is for M.
perideroedes in the particular plant, as it apparently
grows mainly on water-soluble substrates where it isan advantage to be present in the bulk water phase
and not inside the floc. In other cases, however, we have
found this species forming foam in industrial treatment
plants, supporting a general hydrophobic nature of the
sheath.
4.3. Possible control measures
Meganema perideroedes can be characterized as a
very versatile consumer of soluble substrates with both
oxygen and nitrate as e-acceptor, it has a high substrate
affinity, and it has a very large capacity for substrate up-
take and formation of the storage polymer PHA. This is
to some extent different from the general description of
filamentous bacteria in activated sludge [1], stressing
that each species has its own specific characteristics pro-moting growth in specific treatment plants. Many indus-
trial wastewater treatment plants treat wastewater with

C. Kragelund et al. / FEMS Microbiology Ecology 54 (2005) 111–122 121
a high content of soluble organic compounds, and it is in
this type of plant that bulking problems caused by the
morphotypes represented by the Alphaproteobacteria
are reported. Usually, the most efficient control strategy
against many filamentous bacteria is the use of selectors,
where most soluble substrates are consumed by floc-forming bacteria in special selector tanks due to the
floc-formers� higher substrate uptake and growth rate
[9]. However, M. perideroedes has an extremely high
substrate uptake rate and storage capacity, which is
probably comparable to or higher than those of most
floc-forming bacteria, so a selector will probably only
work if the e-acceptor conditions are selective for the
floc-formers. Our study showed that nitrite reducedthe number of organic substrates to be consumed by
M. perideroedes significantly, compared to conditions
with oxygen or nitrate. Thus, a possible control strategy
could be to add nitrite in a selector so that a larger frac-
tion of the organic substrates will go to other denitrify-
ing bacteria, thereby outcompeting M. perideroedes.
Acknowledgements
This study was funded by the Danish Technical Re-
search Council under the research programme ‘‘Activity
and diversity in complex microbial systems’’ and by the
EU programme ‘‘Dynamics and composition of filamen-
tous micro-organism communities in industrial water
systems’’ (DYNAFILM). We thank D. Christensen,M. Kloppenborg and M. Stevenson for their technical
assistance.
References
[1] Eikelboom, D.H. (2002) Process Control of Activated Sludge
Plants by Microscopic Investigation. IWA Publishing, UK.
[2] Jenkins, D., Richard, M.G. and Daigger, G.T. (2004) Manual
on the Causes and Control of Activated Sludge Bulking,
Foaming, and Other Solids Separation Problems. IWA publish-
ing, UK.
[3] Snaidr, J., Beimfohr, C., Levantesi, C., Rossetti, S., van der
Waarde, J., Geurkink, B., Elkelboom, D., Lemaitre, M. and
Tandoi, V. (2002) Phylogenetic analysis and in situ identification
of Nostocoida limicola-like filamentous bacteria in activated
sludge from industrial wastewater treatment plants. Water Sci.
Tech. 46, 99–104.
[4] Levantesi, C., Beimfohr, C., Geurkink, B., Rossetti, S., Thelen,
K., Krooneman, J., Snaidr, J., van der Waarde, J. and Tandoi, V.
(2004) Filamentous Alphaproteobacteria associated with bulking
in industrial wastewater treatment plants. Syst. Appl. Microbiol.
27, 728–736.
[5] Thomsen, T.R., Blackall, L.L., de Muro, M.A., Nielsen, J.L. and
Nielsen, P.H. Meganema perideroedes gen. nov., sp. nov, a new
filamentous Alphaproteobacterium from activated sludge. Int. J.
Syst. Evol. Microbiol. (accepted).
[6] Eikelboom, D.H. and van Buijsen, H.J., 1983. Microscopic sludge
investigation manual. TNO Research Institute for Environmental
Hygiejne, Delft, the Netherlands. MG-TNO A 94a.
[7] van der Waarde, J., Krooneman, J., Geurkink, B., van der Werf,
A., Eikelboom, D., Beimfohr, C., Snaidr, J., Levantesi, C. and
Tandoi, V. (2002) Molecular monitoring of bulking sludge in
industrial wastewater treatment plants. Water Sci. Tech. 46, 551–
558.
[8] Nielsen, J.L., Christensen, D., Kloppenborg, M. and Nielsen,
P.H. (2003) Quantification of cell-specific substrate uptake by
probe-defined bacteria under in situ conditions by microautora-
diography and fluorescence in situ hybridization. Environ.
Microbiol. 5, 202–211.
[9] Wanner, J. (1994) Activated Sludge Bulking and Foaming
Control, 327. Technomic Publishing Company, Inc., Basel,
Switzerland.
[10] Nielsen, P.H., Roslev, P., Dueholm, T.E. and Nielsen, J.L.
(2002) Microthrix parvicella, a specialized lipid consumer in
anaerobic–aerobic activated sludge plants. Water Sci. Tech. 46,
73–80.
[11] Nielsen, P.H., de Muro, M.A. and Nielsen, J.L. (2000) Studies on
the in situ physiology of Thiothrix spp. present in activated sludge.
Environ. Microbiol. 2, 389–398.
[12] Lee, N., Nielsen, P.H., Andreasen, K.H., Juretschko, S., Nielsen,
J.L., Schleifer, K.H. and Wagner, M. (1999) Combination of
fluorescent in situ hybridization and microautoradiography – a
new tool for structure–function analyses in microbial ecology.
Appl. Environ. Microbiol. 65, 1289–1297.
[13] Page, W.J. and Tenove, C.J. (1996) Quantitation of poly-beta-
hydroxybutyrate by fluorescence of bacteria and granules stained
with Nile blue A. Biotechnol. Tech. 10, 215–220.
[14] Gorenflo, V., Steinbuchel, A., Marose, S., Rieseberg, M. and
Scheper, T. (1999) Quantification of bacterial polyhydroxyalka-
noic acids by Nile red staining. Appl. Microbiol. Biotechnol. 51,
765–772.
[15] Huang, C.T., Xu, K.D., McFeters, G.A. and Stewart, P.S. (1998)
Spatial patterns of alkaline phosphatase expression within bacte-
rial colonies and biofilms in response to phosphate starvation.
Appl. Environ. Microbiol. 64, 1526–1531.
[16] Van Ommen Kloeke, F. and Geesey, G.G. (1999) Localization
and identification of populations of phosphatase-active bacterial
cells associated with activated sludge flocs. Microb. Ecol. 38, 201–
214.
[17] Nielsen, J.L., Mikkelsen, L.H. and Nielsen, P.H. (2001) In situ
detection of cell surface hydrophobicity of probe-defined bacteria
in activated sludge. Water Sci. Tech. 43, 97–103.
[18] Loy, A., Horn, M. and Wagner, M. (2003) probeBase: an online
resource for rRNA-targeted oligonucleotide probes. Nucleic
Acids Res. 31, 514–516.
[19] Amann, R.I., Ludwig, W. and Schleifer, K.H. (1995) Phylogenetic
identification and in situ detection of individual microbial cells
without cultivation. Microbiol. Rev. 59, 143–169.
[20] Amann, R.I. (1995) In situ identification of micro-organisms by
whole cell hybridization with rRNA-targeted nucleic acid probes,
part 3.3.6 In: Molecular Microbial Ecological Manual (Akker-
mans, A.D.L., van Elsas, J.D. and de Bruijn, F.J., Eds.), pp. 1–15.
Kluwer Academic Publications, London.
[21] Andreasen, K. and Nielsen, P.H. (1997) Application of microau-
toradiography to the study of substrate uptake by filamentous
microorganisms in activated sludge. Appl. Environ. Microbiol.
63, 3662–3668.
[22] Andreasen, K. and Nielsen, P.H. (2000) Growth of Microthrix
parvicella in nutrient removal activated sludge plants: studies of in
situ physiology. Water Res. 34, 1559–1569.
[23] Ostle, A.G. and Holt, J.G. (1982) Nile blue A as a fluorescent
stain for poly-beta-hydroxybutyrate. Appl. Environ. Microbiol.
44, 238–241.
[24] Zita, A. and Hermansson, M. (1997) Effects of bacterial cell
surface structures and hydrophobicity on attachment to activated
sludge flocs. Appl. Environ. Microbiol. 63, 1168–1170.

122 C. Kragelund et al. / FEMS Microbiology Ecology 54 (2005) 111–122
[25] Thomsen, T.R., Kjellerup, B.V., Nielsen, J.L., Hugenholtz, P. and
Nielsen, P.H. (2002) In situ studies of the phylogeny and
physiology of filamentous bacteria with attached growth. Envi-
ron. Microbiol. 4, 383–391.
[26] Hesselsoe, M., Nielsen, J.L., Roslev, P. and Nielsen, P.H. (2005)
Isotope labeling and microautoradiography of active heterotro-
phic bacteria assimilation of 14CO2. Appl. Environ. Microbiol. 71,
646–655.
[27] Dawes, E.A. (1991) Storage polymers in prokaryots In: Prokary-
otic Structure and Function (Mohan, S., Dow, C. and Coles, J.A.,
Eds.), pp. 81–122. Cambridge University Press, Cambridge.
[28] Dionisi, D., Levantesi, C., Renzi, V., Tandoi, V. and Majone, M.
(2002) PHA storage from several substrates by different morpho-
logical types in an anoxic/aerobic SBR. Water Sci. Tech. 46, 337–
344.
[29] Dircks, K., Beun, J.J., van Loosdrecht, M., Heijnen, J.J. and
Henze, M. (2001) Glycogen metabolism in aerobic mixed cultures.
Biotechnol. Bioeng. 73, 85–94.
[30] Foss, S. and Harder, J. (1998) Thauera linaloolentis sp. nov. and
Thauera terpenica sp. nov., isolated on oxygen-containing mon-
oterpenes (linalool, menthol, and eucalyptol) nitrate. Syst. Appl.
Microbiol. 21, 365–373.
[31] Tandoi, V., Rossetti, S., Blackall, L.L. and Majone, M. (1998)
Some physiological properties of an Italian isolate of Microthrix
parvicella. Water Sci. Tech. 37, 1–8.
[32] Williams, T.M. and Unz, R.F. (1985) Filamentous sulfur bacteria
of activated sludge: characterization of Thiothrix, Beggiatoa, and
Eikelboom type 021N strains. Appl. Environ. Microbiol. 49, 887–
898.
[33] Nielsen, J.L. and Nielsen, P.H. (2002) Enumeration of acetate-
consuming bacteria by microautoradiography under oxygen and
nitrate respiring conditions in activated sludge. Water Res. 36,
421–428.
[34] Kloeke, F.V., Baty, A.M., Eastburn, C.C., Diwu, Z. and Geesey,
G.G. (1999) Novel method for screening bacterial colonies for
phosphatase activity. J. Microbiol. Methods 38, 25–31.




















