
Purification and characterization of an extracellular metalloprotease from Pseudomonas fluorescens
7
J. Biochenr. 121,82 88 (1997) Purification and Characterization of an Extracellular Metalloprotease from Pseudnmonds fluorescensl Hee-Jung Kimr*'t Yoshiaki Tamanoue,* Gwang-Ho Jeohn,*,r Akihiro Iwamatsuri Akira Yokota,$Yong-Tae Kim,* Takayuki Takahashi,* and Kenji Takahashi*,1''z *Deparhnent of Biophysics and. Biochemistry, Graduate Scttoot of Science, The IJniuersity of Tokyo, 7 3 l Hongo, Bunkyo-ku, Tohyo 113; lLaboratary of Molecular Biochemistry, School of Life Science, Tokyo Uniuersity of Phatmacy and Life Science, 1432 1 Horinouchi, Hachioji, Tokyo 192 03; tg"nt o, "aboratories for Key Technology, Kirin Brewery Co., Ltd., Yokohama, Kanagawa 236; and slnstitute of Mol,ecular and Cellular Biosciences, The Uniuersity of Tokyo, 1-1-1 Yayoi, Bunkyo-ku, Tohyo 113 Received for publication, August 22, 1996 An extracellular metalloprotease was purified from the culture supernatant of Pseudo- monds fluorescens strain KTl to apparent homogeneity and shown to consist of a single polypeptide chain (M 46,000-47,000). The enzyme was strongly inhibited by chelating agents such as EDTA and o-phenanthroline, and activated by certain detergents. Amongthe peptidyl 4-methylcoumaryl-7-amide (MCA) substrates examined, t-butyloxycarbonyl- Arg-Val-Arg-Arg-MCA was the best one. With this substrate, the enzyme exhibited a pH optimum of around pH 5.5 in the absence of Co2+ ions, whereas it showed two difrerent pH optima (at pHs around 5.5 and 8-9) in the presence of Co2+ ions due to remarkable activation by Co'* ions in the alkaline pH range. On the other hand, a single broad pH optimum of around 6 to 8 was obtained with some peptides in both the presence and absence of Co2+ ions, and no activation by Co'n was observed. The enzyme showed trypsin-like specificity, preferentially cleaving certain arginyl peptide bonds, and hydrolyzed the basic protein, histone, most rapidly among various proteins examined. Partial amino acid sequence analysis revealed that the enzyme is highly homologous with proteases of the serralysin family, a group of zitoLc metalloproteases. Key words: Co'* ion activation, metalloprotease, partial amino acid sequence, Pseudo- m.onas fluorescens, sequence homology. Pseudomonas sp. is ubiquitously distributed in water and soil, and frequently isolated from clinical and environmen- tal specimens. This species produces several proteases and often causes serious problems as a pathogen and/or as a source of food deterioration. Among these proteases, the alkaline metalloprotease from Pseudomonas aeruginosa has been most extensively studied (7), including determi- nation of its primary (2, 3), and tertiary (4) structures. Furthermore, the primary structures of the related metal- loproteases from Serratia rlarcescens (5) and Erwinia chrysantherni (6, 7), and the tertiary structure of the Senatia enzyrne (8) have also been reported. These 'This study was supported in part by Grants-in-Aid for Scientific Research from the Ministry of Education, Science, Sports and Culture of Japan. 'To whom correspondence should be addressed at: School of Life Science, Tokyo University of Pharmacy and Life Science, 1432-1 Horinouchi, Hachioji,, Tokyo 192 03. Tel: +87-426-76-7746, Fax: |81-426 -76-7749 Abbreviations: BAM-12P, bovine adrenal medulla dodecapeptide; Boc, f-butyloxycarbonyl; BPTI, bovine pancreatic trypsin inhibitor; DFP, diisopropylfluorophosphate; E-64, l- lrons-epoxysuccinyl-leuryi- amide- (4-guanido)butane; MCA, 4-methylcoumaryl-7-amide; MES, 2 - (morpholino) ethane sulfonate ; MOPS, 3 - (morpholino)propane sul - fonate; PAGE, polyacryIamide gei electrophoresis; PMSF, phenyl- methanesulfonyl fluoride; SBTI, soybean trypsin inhibitor; TLCK, No-tosyl-r,-lysine chioromethyi ketone; T?CK, N'-tosyl-r,-phenyl- alanine chloromethyl ketone. proteases have similar structures and thus are classified as members of the serralysin family, a family of zinc metallo- proteases (9) different from the thermolysin family. Pseudomonas fluorescens also secretes several proteases which may differ among its biotypes (10). Because this species is known as the major source of food deterioration, studies on these proteases were performed mainly on their heat stability and production control, their molecular and enzymatic charactefization having not been performed in detail (14 ll). Furthermore, no information is available about their primary structures. Therefore, it seems in- teresting to elucidate their properties in detail and to compare them with those of proteases from related species. As the first step toward this end, we isolated in this study an extracellular metalloprotease from a frild strain of P. fluorescens, and investigated its molecular and enzymatic properties, including its partial amino acid sequence. The results indicated that this protease has trypsin-like speci- ficity, preferentially cleaving certain arginyl peptide bonds, and is closely related in structure with the alkaline protease from P. d.eruginosa and related proteases ofthe serralysin family. EXPERIMENTAL PROCEDURES Materials-Peptide 4-methylcoumaryl- 7- amide (MCA) substrates, L- trans - epoxysuccinyl-leucylamide- (4-guanido) - 82 J. Biochem.
Transcript of Purification and characterization of an extracellular metalloprotease from Pseudomonas fluorescens

J. Biochenr. 121,82 88 (1997)
Purification and Characterization of an Extracellular Metalloproteasefrom Pseudnmonds fluorescensl
Hee-Jung Kimr*'t Yoshiaki Tamanoue,* Gwang-Ho Jeohn,*,r Akihiro IwamatsuriAkira Yokota,$Yong-Tae Kim,* Takayuki Takahashi,* and Kenji Takahashi*,1''z*Deparhnent of Biophysics and. Biochemistry, Graduate Scttoot of Science, The IJniuersity of Tokyo, 7 3 l Hongo,Bunkyo-ku, Tohyo 113; lLaboratary of Molecular Biochemistry, School of Life Science, Tokyo Uniuersity ofPhatmacy and Life Science, 1432 1 Horinouchi, Hachioji, Tokyo 192 03; tg"nt o,
"aboratories for Key Technology,
Kirin Brewery Co., Ltd., Yokohama, Kanagawa 236; and slnstitute of Mol,ecular and Cellular Biosciences, TheUniuersity of Tokyo, 1-1-1 Yayoi, Bunkyo-ku, Tohyo 113
Received for publication, August 22, 1996
An extracellular metalloprotease was purified from the culture supernatant of Pseudo-monds fluorescens strain KTl to apparent homogeneity and shown to consist of a singlepolypeptide chain (M 46,000-47,000). The enzyme was strongly inhibited by chelatingagents such as EDTA and o-phenanthroline, and activated by certain detergents. Amongthepeptidyl 4-methylcoumaryl-7-amide (MCA) substrates examined, t-butyloxycarbonyl-Arg-Val-Arg-Arg-MCA was the best one. With this substrate, the enzyme exhibited a pHoptimum of around pH 5.5 in the absence of Co2+ ions, whereas it showed two difrerent pHoptima (at pHs around 5.5 and 8-9) in the presence of Co2+ ions due to remarkable activationby Co'* ions in the alkaline pH range. On the other hand, a single broad pH optimum ofaround 6 to 8 was obtained with some peptides in both the presence and absence of Co2+ ions,and no activation by Co'n was observed. The enzyme showed trypsin-like specificity,preferentially cleaving certain arginyl peptide bonds, and hydrolyzed the basic protein,histone, most rapidly among various proteins examined. Partial amino acid sequenceanalysis revealed that the enzyme is highly homologous with proteases of the serralysinfamily, a group of zitoLc metalloproteases.
Key words: Co'* ion activation, metalloprotease, partial amino acid sequence, Pseudo-m.onas fluorescens, sequence homology.
Pseudomonas sp. is ubiquitously distributed in water andsoil, and frequently isolated from clinical and environmen-tal specimens. This species produces several proteases andoften causes serious problems as a pathogen and/or as asource of food deterioration. Among these proteases, thealkaline metalloprotease from Pseudomonas aeruginosahas been most extensively studied (7), including determi-nation of its primary (2, 3), and tertiary (4) structures.Furthermore, the primary structures of the related metal-loproteases from Serratia rlarcescens (5) and Erwiniachrysantherni (6, 7), and the tertiary structure of theSenatia enzyrne (8) have also been reported. These
'This study was supported in part by Grants-in-Aid for ScientificResearch from the Ministry of Education, Science, Sports and Cultureof Japan.'To whom correspondence should be addressed at: School of LifeScience, Tokyo University of Pharmacy and Life Science, 1432-1Horinouchi, Hachioji,, Tokyo 192 03. Tel: +87-426-76-7746,Fax: |81-426 -76-7749Abbreviations: BAM-12P, bovine adrenal medulla dodecapeptide;Boc, f-butyloxycarbonyl; BPTI, bovine pancreatic trypsin inhibitor;DFP, diisopropylfluorophosphate; E-64, l- lrons-epoxysuccinyl-leuryi-amide- (4-guanido)butane; MCA, 4-methylcoumaryl-7-amide; MES,2 - (morpholino) ethane sulfonate ; MOPS, 3 - (morpholino)propane sul -fonate; PAGE, polyacryIamide gei electrophoresis; PMSF, phenyl-methanesulfonyl fluoride; SBTI, soybean trypsin inhibitor; TLCK,No-tosyl-r,-lysine chioromethyi ketone; T?CK, N'-tosyl-r,-phenyl-alanine chloromethyl ketone.
proteases have similar structures and thus are classified asmembers of the serralysin family, a family of zinc metallo-proteases (9) different from the thermolysin family.Pseudomonas fluorescens also secretes several proteaseswhich may differ among its biotypes (10). Because thisspecies is known as the major source of food deterioration,studies on these proteases were performed mainly on theirheat stability and production control, their molecular andenzymatic charactefization having not been performed indetail (14 ll). Furthermore, no information is availableabout their primary structures. Therefore, it seems in-teresting to elucidate their properties in detail and tocompare them with those of proteases from related species.
As the first step toward this end, we isolated in this studyan extracellular metalloprotease from a frild strain of P.
fluorescens, and investigated its molecular and enzymaticproperties, including its partial amino acid sequence. Theresults indicated that this protease has trypsin-like speci-ficity, preferentially cleaving certain arginyl peptide bonds,and is closely related in structure with the alkaline proteasefrom P. d.eruginosa and related proteases ofthe serralysinfamily.
EXPERIMENTAL PROCEDURES
Materials-Peptide 4-methylcoumaryl- 7- amide (MCA)substrates, L- trans - epoxysuccinyl-leucylamide- (4-guanido) -
82 J. Biochem.

Metalloprotease from Pseudomonas fluorescens
butane (E-64), leupeptin, bestatin, ch),,rnostatin, bovineadrenal medulla dodecapeptide (BAM-12P), dynorphin A,neurotensin, and a-neoendorphin were purchased fromPeptide Institute. Proalbumin( 1 - 10) peptide was s1'nthe-sized in our laboratory. DEAE-cellulose (DE-52) waspurchased from Whatman, and chelating Sepharose 68,Sephacryl 5-200, and a Mono Q (HR5/5) column were fromPharmacia Biotechnology. A BCA protein assay kit wasobtained from Pierce. Fluorescamine, histone, bovineserum albumin, ovalbumin, myoglobin, 7-immunoglobu-lin, pancreatic trypsin inhibitor (Kunitz) (BPTI), soybeantrypsin inhibitor (SBTI), diisopropylfluorophosphate(DFP), N"-tosyl-r,-phenylalanine chloromethyl ketone(TLCK), .0y''-tosyl-l-Iysine chloromethyl ketone (TLCK),phenylmethanesulfonyl fluoride (PMSF), and o-phenan-throline were purchased from Sigma. Lubrol PX, sodiumdeoxycholate, histone (calf thymus), and EDTA were fromNacalai Tesque, and casein (Hammasten) from Merck. Amixture of molecular weight marker proteins for SDSpolyacrylamide gel electrophoresis (PAGE) was obtainedfrom BioRad. Other reagents used were ofthe highest gradeavailable.
Bacterial Strain-When a rat liver homogenate preparedin 50 mM triethanolamine-HCl buffer, pH 8.0, containing0.25 M sucrose was kept standing at room temperature fora few days, a bacterium was found to grow in the homo-genate and to produce an extracellular protease hydrolyzingBoc-Arg-Val-Arg-Arg-MCA. This bacterial strain, namedKT1, was isolated and identified by the conventionalmicrobiological method.
Determination of taxonomic characterisllcs: Motility wasobserved under a light microscope using cells in the loga-rithmic growth phase grown on nutrient agar. Catalaseactivity was determined by bubbling n a 33% hydrogenperoxide solution. Oxidase activity was determined by theoxidation of 1% tetramethyl-p-phenylenediamine on filterpaper. For the determination of other physiological andbiochemical characteristics, API (Appareilis et Procedes'Identification; La Balme les Grottes, Montalieu Vercieu,France) 20E was used.
Cellular fatty acid analysis: Cells harvested after cultur-ingfor 24 h in nutrient broth were freeze-dried, and then 50mg of the dried cells was suspended in 10% HCI in methanoland heated at 100"C for 3 h. Fatty acid methyl estersextracted with n-hexane were separated by TLC using asolvent system of. n-hexate/diethyl ether ( 1 : 1, v/v). Non-polar, 2-hydroxy (2-OH) and 3-hydroxy (3-OH) fatty acids,visuaiized by spraying 0.02% dichlorofluorescein in eth-anol, were extracted with diethyl ether and analyzed byGC.
DNA base composition analysis: DNA was isolated bythe method of Saito and Miura (12) . The G * C content ofDNA was determined by reversed-phase HPLC (13) afternuclease Pr and alkaline phosphatase treatment.
Analysis of isoprenoid quinones: Isoprenoid quinoneswere extracted twice with chloroform/methanol (2 :'J.,, v/v) for 4h, and purified by TLC using ru-hexane/diethylether (85 :1,5, v/v) as the solvent, and then analyzed byHPLC.
Enzyme Assay-Enzyme activity toward MCA sub-strates was determined as described by Tamanoue et al.Q$ with a slight modification. For routine assaying, f-butyloxycarbonyl (Boc) -Arg-Va1-Arg-Arg-MCA was used
Voi. 121, No. 1, 1997
83
exclusively unless otherwise specified. The assay wascarried out at 37'C for 30 min in 200 pl of a reactionmixture comprising 0.1 M Tris-HCl buffer (pH 8.0), 0.1mM MCA substrate, 1mM CoClz, and the enzyme. Whennecessary, 0.1 M 2-(morpholino)ethane sulfonate (MES)buffer (pH 5.5) was used in the absence of Co2+ ions. Thereaction was stopped by the addition of 1 ml of 0.1 Msodium acetate buffer (pH 4.3) containing 0.1 M monochlo-roacetic acid. The amount of 7-amino-4-methylcoumarinreleased was measured with a Hitachi spectrofluorometerat an excitation wavelength of 370 nm and an emissionwavelength of 460 nm. One unit of activity was defined asthe amount of enzyme releasing 1 pmol of 7-amino-4-meth-ylcoumarin per min.
Kinetic parameters (h* and K, values) for the hydroly-sis of peptide MCA substrates were determined accordingto Lineweaver-Burk (15) under the standard conditionsdescribed above except that the pH, Co'* ion concentrationand/orsubstrates were varied.
Enzyme Purification-All purification procedures \ryereperformed at {"C in the presence of 0.02% NaN.. Pseado-filonas fluorescens KT1 was grown in LB broth containing 1
mM CaCIz at 30"C for 2 days. The culture fluid was obtainedby centrifuging the whole culture (5,000 x g, 20 min). Thesupernatant was dialyzed against 10 mM l[-methyldieth-anolamine, pH 9.0 (buffer A), for 8 h and then overnightwith one change of buffer A. The dialyzed fraction wascentrifuged (5,000 x s, 30 min) and then applied to a DE-52column (3 X 29 cm) equilibrated with buffer A at a flow rateof 30 ml/h. The column was washed with 2 volumes ofbuffer A, followed by elution with a linear gradient of NaCl(0-0.25 M) in a total volume of 1,000 ml, 9.0-ml fractionsbeing collected. The active fractions (Nos. 4l-49) wercpooled and applied to a chelating Sepharose 68 column(1.8x5cm) equilibrated with 20mM Tris-HCl, pH8.0(buffer B). The column was washed with 0.5 M NaCl,followed by elution with 0.2 M glycine at a flow rate of 30ml/h, 3.0-ml fractions being collected. The active fractions(Nos. 34-37) were pooled and concentrated to 2 ml byultrafiltration. An aliquot (1 ml) of the enzyrne solution wasapplied to a Sephacryl S-200 column (1x 145 cm) equili-brated with buffer B, and elution was carried out at a flowrate of 6 ml/h, 1.5-mI fractions being collected. The activefractions (Nos. 54-57) were pooled and dialyzed against0.02 M triethanolamine-HCl buffer, pH 8.0. One-third ofthe sample was applied to a Mono Q (HR5/5) columnequilibrated with 0.02 M triethanolamine-HCl buffer, pH8.0. Elution was performed with a gradient of NaCI (0 to 0.5M) in the same buffer (15 ml) at a flow rate of 0.5 ml/min,0.3-ml fractions being collected. The active fractions werepooled.
Protein Determinatio,n-Protein was determined bymeasuring the absorbance at 280 nm of the sample solutionor by the method of Smith et al. ( 76) using the BCA reagentwith bovine albumin as a standard.
E lectrophoreszs-SDS -PA GE (12o/o acrylamide gel) wasperformed according to Laemmli (17), and. the gel wasstained with Coomassie Brilliant Blue. Electrophoresisunder non-reducing conditions was performed at pH 9.5and 4"C using 4.5% acrylamide gel containing 1 mM CoCLand 0.02 mMZnClr. The gel was sliced into 1.S-mm pieces,and each piece was immersed in 100 pl of distilled waterbefore freezing and thawing for extraction of the enzyme.
B4
The supernatant of the gel-containing solution, which hadbeen vortexed and centrifuged, was used for the determina-tion of protease activity at pH 5.5 and 8.0.
Determination of the Molecular Moss-The molecularmass was determined by SDS-PAGE using molecularweight markers (Bio-Rad). It was also determined using acalibrated column of Sephacryl 5-200.
Hydrolysis of Peptides-Eaeh peptide (1 nmol) washydrolyzed with 15 units of enzyme in 50 pl of 0.04 MTris-HCl buffer, pH 8.0 (or other buffers of different pHswhen necessary) at 37"C for 30 min. The digest was imme-diately arnlyzed.by HPLC on a TSKgeI ODS-120T column(0.46x25cm) in a Hitachi 655A-11 HPLC apparatus.Elution was performed with a gradient of acetonitrile (0 to50o/o in 60 min) in 0.1% trifluoroacetic acid at a flow rate of0.8 ml/min and monitored at 215 nm. The isolated pep-tides were hydrolyzed with 6 N HCI at 110"C for 24h inevacuated sealed tubes and then submitted to amino acidanalysis with an Applied Biosystems derivatizerf analyzer(4204/730/^-e20A).
Hydrolysis of Proteins-Each protein solution (1%) in
TABLE I. Taxonomic charaeteristics of strain KT1.
CharacteristicPseudomonas
Strain KTl aeruginosaIAM 15147
H.-J.Kim et al.
0.2 M sodium borate buffer, pH 8.0, was incubated with theenzyme aL 37"C for 30 min. The extent of hydrolysis wasanalyzed with fluorescamine as described previously (4).
Thermal Stability Measuremenf-The enzyme (24 units)was incubated at 43"C in 50 p I of 0. 1 M Tris-HCl buffer, pH8.0, or in 0.1 M MES buffer, pH 5.5, and at appropriateintervals aliquots were removed for assaying with Boc-Arg-Val-Arg-Arg-MCA as a substrate.
Determination of the NHz-Terrninal and Internal Se-quences-The NHz-terminal sequence and internal se-quences were analyzed according to Iwamatsu and Yoshida-Kubomura (18, L9). The purified enzyme was electroblot-ted onto a polyvinylidene difluoride membrane after sepa-ration by SDS-PAGE. For determination of the NHz -termi-nal sequence, the enzyme on the membrane, that had beenreduced and S-carboxymethylated, was directly submittedto amino acid sequencing with a Shimadzu RSQ- 1 gas phase
sequencer. On the other hand, for analysis of the internalsequences, the reduced and S-carbox5rmethylated enzymeon the membrane was further treated with Achromobacterprotease I (lysylendopeptidase). The released proteolyticfragments were separated by HPLC on a p-Bondasphere5 p C8-300 A column (2.2x150 cm; Waters) (data notshown) and then submitted to amino acid sequencing.
RESULTS AND DISCUSSION
Iderltificatinn of Pseudarnonas Stro.in KTl-Table Ishows the taxonomic characteristics of strain KT1. Basedon the morphological, biochemical, and chemotaxonomiccharacteristics thus determined, the isolate, KT1, was iden-tified as Pseudomonas fluorescens according to Bergey'sManual of Systematic Bacteriology (20).
Purifi.cation of a Protease from P. fluorescens The re-sults of purification of a protease from the culture super-natant of P. fluorescens strain KT1 are shown in Table II'On DE-52 column chromatography the protease activitywas eluted as a single peak at about 0.12 M NaCl. Uponsubsequent chromatography on chelating Sepharose 68with Zn2* as a ligand, the activity was eluted at 0.2 Mglycine. The enzyme was further purified by chromatog-raphy on a Sephacryl 5-200 column, followed by chro-matography on a Mono Q column. The protein and activitypeaks coincided well with each other. Through theseprocedures, the enzyme was purified about 700-fold ina60/oyield (Table II), and only a single protein band was detectedon SDS-PAGE (Fig. 1a). The molecular mass of the purifiedprotease was estimated to be approximately 47,000 Da bySDS-PAGE under both reducing and non-reducing condi-
TABLE II. Purification of the protease from the culture super'natant of Pseudomonas fl.uorescens IdTl.
Gram stainMotilityIsoprenoid quinoneCellular fatty acids":
non-poIar2-hydroxy3-hydroxy
G*C content of DNA (mol%) 60.4
NegativeMotileQ-e
16:0, 17:0Z-OH 12:03-OH 10:0
NegativeMotiIeQ-e
16:0, 16:12-OH l2:03-OH 10:067.2
Formation of:oxidasecatalaser,- arginine dihydrolaset -Iysine decarboxylaset .ornithine decarboxylasel-tryptophan deaminaseureaseindole
Hydrolysis of:o-nitrophenol
Utilization of citrateH,S formationVoges-Proskauer reactionLiquefaction of gelatinAcid formation from:
glucosemannitolinositolsorbitolL-rhamnosesaccharosemelibioseamygdalinl-arabinose
Nitrate reductionGrorth at: 16:0,
4'C4t"c
T
++
+++
+
++
+
*w
+
+
+
+
+
*w
+
+
+
Totai rotal sp9"ifi" pr.ifi.ation yieldStep protein activity
, acjivlty, i_f"ld) (V")(ms) (units) (units/mg)
Culturesupernatant
DE-52Chelating
SepharoseSephacryI
s-200Mono Q
1,440 823,500 572
24.8 569,000 22,9001.0 154,000 154,000
0.20 57,900 289,000
0.L2 47,800 398,000
1 100
40 69270 19
7.O
696 5.8
J. Biochetn.
Metalloprotease from Pseudomonas fluorescens
tions, indicating that the enz5,,rne is composed of a singlepolypeptide chain. It was also estimated by gel filtration ona column of Sephacryl 5-200 to be around 46,000 Da (datanot shown).
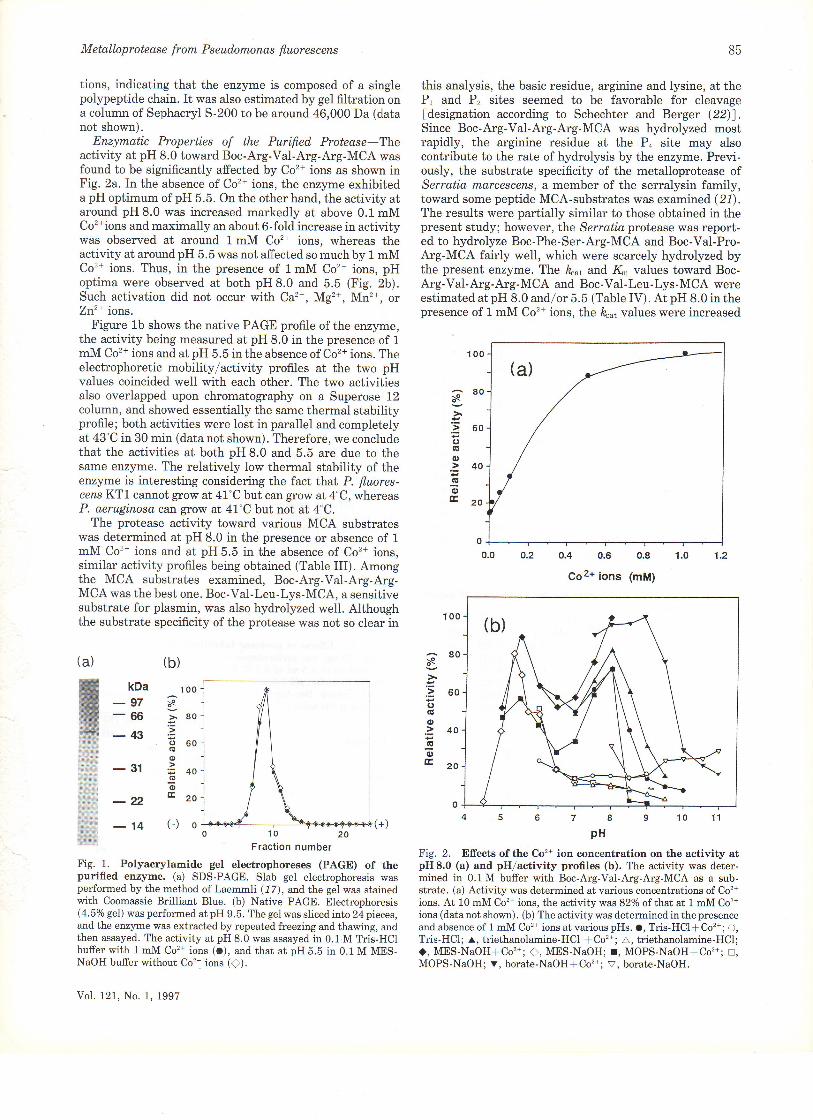
Enzymatic Properties of the Purified Protease Theactivity at pH 8.0 toward Boc-Arg-Va1-Arg-Arg-MCA wasfound to be significantly affected by Co'n ions as shown inFig. 2a. In the absence of Co2+ ions, the enzyme exhibiteda pH optimum of pH 5.5. On the other hand, the activity atarou.nd pH 8.0 was increased markedly at above 0.1 mMCo'*ions and maximally an about 6-fold increase in activitywas observed at around 1mM Co2* ions, whereas theactivity at around pH 5.5 was not affected so much by 1 mMCo2* ions. Thus, in the presence of lmM Co'* ions, pHoptima were observed at both pH 8.0 and 5.5 (Fig. 2b).Such activation did not occur with Ca2+, Mg'*, Mn2*, orZn2+ ions.
Figure 1b shows the native PAGE profile of the enzyme,the activity being measured at pH 8.0 in the presence of 1
mM Co2+ ions and at pH 5.5 in the absence of Co2+ ions. Theelectrophoretic mobility/activity profiles at the two pHvalues coincided well with each other. The two activitiesalso overlapped upon chromatography on a Superose 12column, and showed essentially the same thermal stabilityprofile; both activities were lost in parallel and completelyat 43"C in 30 min (data not shown). Therefore, we concludethat the activities at both pH 8.0 and 5.5 are due to thesame enzJrne. The relatively low thermal stability of theenzyme is interesting considering the fact Lhat P. fluores-censKTl cannot gtow at 41'C but can grow at4"C, whereasP. aeruginoso can grow at 47"C but not at 4"C.
The protease activity toward various MCA substrateswas determined at pH 8.0 in the presence or absence of 1
mM Co2+ ions and at pH 5.5 in the absence of Co'* ions,similar activity profiles being obtained (Table III). Amongthe MCA substrates examined, Boc-Arg-Va1-Arg-Arg-MCA was the best one. Boc-Val-Leu-Lys-MCA, a sensitivesubstrate for plasmin, was also hydrolyzed well. Althoughthe substrate specificity of the protease was not so clear in
kDa
-97-66-43
-31
-22-14
B5
this analysis, the basic residue, arginine and lysine, at thePr and Pz sites seemed to be favorable for cleavage[designation according to Schechter and Berger (22)].Since Boc-Arg-Val-Arg-Arg-MCA was hydrolyzed mostrapidly, the arginine residue at the P, site may alsocontribute to the rate of hydrolysis by the enzyme. Previ-ously, the substrate specificity of the metalloprotease ofSerratia rnarcescens, a member of the serralysin family,toward some peptide MCA-substrates was examined (27).The results were partially similar to those obtained in thepresent study; however, the Serratia protease was report-ed to hydrolyze Boc-Phe-Ser-Arg-MCA and Boc-Val-Pro-Arg-MCA fairly well, which were scarcely hydrolyzed bythe present enzyrne. The ft.ut and K. values toward Boc-Arg-Val-Arg-Arg- MCA and Boc- Val- Leu- Lys -MCA wereestimated at pH 8.0 and./or 5.5 (Table IV). At pH 8.0 in thepresence of 1 mM Co'* ions, the k ut values were increased
100
-80:s
:= 60(,(U
o.= 40.U
oE20
o
(a)
(b)(a)
10 20Fraction number
Fig. 1. Polyacrylamide gel electrophoreses (PAGE) of thepurified erzyme. (a) SDS-PAGE. Slab gel electrophoresis wasperformed by the method of Laemmli (17), and, the gei was stainedwith Coomassie Brilliant BIue. (b) Native PAGE. Electrophoresis(4.5o/o gel) was performed at pH 9.5. The ge1 was sliced into 24 pieces,and the enzyme was extracted by repeated freezing and thawing, andthen assayed. The activity at pH 8.0 was assayed in 0.1 M Tris-HClbuffer with 1 mM Co'* ions (O), and that at pH 5.5 in 0.1 M MES-NaOH buffer without Co'* ions (f).
Vol. 121, No. 1, 1997
o.2 0.4 0.5 0.8 1.0 1.2
Coz+ ions (mM)
pH
Fig. 2. Efrects of the Co'?+ ion concentration on the activity atpH 8.0 (a) and pHlactivity profles (b). The activity was deter-mined in 0.1M buffer with Boc-Arg.VaI-Arg.Arg-MCA as a sub-strate. (a) Activity was determined at various concentrations of Co2+
ions. At 10 mM Co'z+ ions, the activity was 82%o of that at 1 mM Co'*ions (data not shown). (b) The activity was determined in the presenceand absence of 1 mM Co2* ions at various pHs. o, Tris-HCl* Co'?+; O,Tris-HCl; r, triethanolamine-HCl * Co't ; A, triethanolamine-HCl;<), MES-NaOHlCo'*; ), MES-NaOH; r, MOPS.NaOH*Co'z+; !,MOPS-NaOH; v, borate-NaOHf Co'zt; v, borate-NaOH.
ao
:: 50(,C'o>40(!oE20
100\o
>80't,Fo60(6
o,;40(g
6Ezo0

B6
several fold as compared with those obtained in the absence
of Co2+ ions, whereas the K* values changed to a lesserextent. Therefore, the activation by Co'* ions at pH 8.0 islargely due to the increase in the k.t value. On the otherhand, at pH 5.5, these values did not change significantly inthe presence or absence of Co2+ ions. In this connection, itis interesting to note that Morihara and Tsuzuki (23)previously reported marked activation by Co'* ions at pH7.0 of P. aeruginosa. alkaline metalloprotease as to benzyl-
SubstratepH 8.0 pH 5.5
+co2* co2+ _cor*(1 mM)
H.-J.Kim et al.
100
80
60
40
20
0
100
80
60
40
20
o
Fig. 4. pH-rate profiles of the hydrolysis of peptides by theenzyme. (a) Dynorphin A, (b) BAM-12P ( ), and proalbumin (1-
10) peptide ( ). 0.04 M buffers were used: o, Tris'HCIt0.1mMCo'+ ions; o, Tris-HCI; C, MES-NaOH; x, sodium acetate. Themaximal activity was taken as 100%'
TABLE V. Efrects of protease inhibitors on the activity. The
enzyme (15 ng) was preincubated with each inhibitor at the given
concentration in 0.5 mI of 0.1 M Tris-HCI buffer, pH 8.0, or 0.1 MMES-NaOH buffer, pH 5.5, at room temperature for t h, and then theactivity toward Boc-Arg-VaI-Arg-Arg-MCA as a substrate was
measured at the same pH.
Inhibitor
Relative activity (%)
pH 5.5
- Co2+
s.;(,(g
6)
(o
oG,
Boc.Arg-Va1-Arg-ArS-MCABoc-Va1-Leu-Lys-MCABoc-GIn-Arg-Arg-MCABoc-Gly-Lys-Arg-MCABoc-Gly-Arg-Arg-MCABoc-GIn-A1a-Arg-MCABoc-Leu- Ser-Thr-Arg-MCABoc-Leu-Lys-Arg-MCABoc-Glu-Lys-Lys-MCABoc-Leu-Arg-Arg-MCABoc-Ala-GIy-Pro-Arg-MCABoc-Phe-Ser-Arg-MCABoc-Leu-Leu-Vai-Tyr-MCABoc-Gln-GIy-Arg-MCABoc-IIe-Glu-GIy-Arg-MCABoc-Leu-Thr-Arg-MCABoc-Val-Pro-Arg-MCABoc-Leu-G1y-Arg-MCASuc-Gly-Pro-MCASuc"Ala-Ala-Pro-Phe-MCA
10044JC252714
2.92.51.30.80.70.60.4o.2o.2000
1003949242tl75.72.601.3
nd0.8
ndndnd
1.0ndndndnd
100281913
7.2nd
1.12.01.27.71.30.200.30.500.10.71.00.3
4s678pH
nd, not determined; suc, succinyl.
TABLE IV. Kinetic parameters for the hydrolysis of peptideMCA substrates.
Substrate -- Co'* l?"^,Pt (1-M) (s ')
K^ h",/K^(mM) (mM'.s')
Boc-Arg-VaI-Arg-Arg-MCABoc Arg-Val-Arg-Arg-MCABoc-Arg-VaI-Arg-Arg-MCABoc-Arg-Vai-Arg-Arg-MCABoc-VaI-Leu-Lys-MCABoc-Val-Leu-Lys-MCA
0.53 0.0350.064 0.0150.39 0.0230.46 0.0230.11 0.0230.046 0.016
8.0 +8.05.5 +5.58.0 +8.0
15.14.2
16.920.14.62.9
Concen-tration(mM)
pH 8.0
* Co'* Co'*(1mM)V
I yr-Cly-Cl) -Phe-l eu-Arg Arg-lle-Arg-Pro-l ys-Leu-
Lys-Trp-Asp-Asn-Gln
YTyr-Gl1 -cly-Phe-vel-Arg- A18:\ al-Cly- Arg-Pro Llu
Y' Clu-l eu Tyr-Clu Asn I y5-Pro-Arg:ArB-Pro I yr-llc-
Leu yA rg-G ly - V a l- Phe-Arg - A rg- A sp - A I a - H i s - Ly s
Tyr- G ly-G ly- P h e- Leu -A rg- Ly s-Ty r- Pro - Ly s
(not cleaved)
Fig. 3. The sites of cleavage of some peptides by the enzyme.Each peptide was hydrolyzed with the enzyllrrc at pH 8.0 in both thepresence and absence of 1 mM Co2* ions. The digest was analyzed as
described under 'MATERIALS AND METHODS". Arrowheadsindicate the sites of cleavage. (GIu, pyT oglutamic acid residue.
DYNORPHIN
BAM- 1 2P
NEUROTENSIN
PROALBUMIN(1-r0) PEPTIDE
d -NEOENDORPHIN
NoneEDTAo-PhenanthrolinePhosphoramidoneDFPPMSFE-64N-EthyimaleimideTLCKTPCKChymostatinLeupeptinElastatinalBestatinBPTISBTI
10100.11.01.00.10.10.10.10.10.10.10.10.010.01
1000.50.8
10088837486ndndnd90ndndndnd
100*0
974
1098498nd
10182829389
707100100
1000.19.6
74918684ndndndndndndnd9591
(a)
(b)
a
\)
J. Biochem.

Metalloprotease from Pseudomonas fluorescens
NH,-terminal FAp-5 -------QASTAYS I(ASGGD_ - -GRS DAYTQVDNTLHAYARGGDELVNGH P S YTVDQAAEQ I LREQASWQKAPGDSVLTLS YS___AATTGYD HERGNG
AKAGSYTGYA HORGDG
67 90 t20F_Ap_ 5a___ _,
ir,rxpN o'rNr pwKyvs D r ys LexrsarsaooffiSffi #3;#ru, *. HFVDAGecDei
QQQAKLSLQSWAMQQAKLSLQSWA
127 150 180
l__AP_18 _
DLr EGN Es s svGGAAEAEL pDVp DALKGeiwyL r r.. r.orIliiiffi:l8iliHfuKHPATEDYGRQTTTTTTT
RNPDTDEYGRHSFTITEI
:] 210 240
FeP-eu-----KGO/EAYJASGPL
TLGLS H PGDYNAGE G DPTYADATYAEDTRAYSVMS YWEEQNTGQDFKGAYS SAPLLDDIAGDNGGHYAAAPL
GDF'KGHYSAGPL
87
Fig. 5. Comparison of the partial amino acidsequence ofthe protease with the sequences ofsome serralysin proteases. The sequences arealigned as four lines using the one letter notation,the residues in the present enzyme and theresidues identical with them being shown in boldface. First line: the partial sequences of thepresent protease; second line: the complete se-quence of Pseud,omonas aeruginosa alkaline pro-tease (J 3); third line: Setatia m,arcescensprotease (5); and fourth line: Erwinia chrysan-themi protease B (6, 7). The peptides derivedfrom the present enzyme are numbered in theorder of elution on HPLC (data not shown) exceptfor AP-18, which was obtained under differentHPLC conditions. AP stands for Achromobacterlysylendopeptidase. Small letters indicate resi-dues not fuily identified. Dashes indicate gapsintroduced to maximize the homology. Eachpeptide was partially sequenced from the NHr-terminus except for AP-8, AP-7, and AP.2b,which were sequenced completely. Each peptidewas assumed to be preceded by a iysine residuefrom the spgcificity of lysylendopeptidase. Forthe Pseudornonos a,eruginosa protease, the zincbinding motif and calcium binding sites are in-dicated by double and single underlines, respec-tively. For the Serratia ar.d Erwinia proteases,only the sequences corresponding to the partialsequences of the present enzyme are shown.
6030
24Lf-AP-3
210
FAP-2a-IFAP-9300
, KLYGAIITSTRATDTTYGENS KXEDFTSTSDKMS\,WDGGGNDtlDFscEA I QKLYCAI{LTTRT GDTVYGEITSNTERDEi SATS S S SKI,VFSVIiIDAGGIIDTLDESGFS QN
HLYGA}ITSTRTGDTVYGFIIS DF'LSTTSNSQKVITAAIIDAGGNDTFDESGYKLYGA}IMTTRTGDTVYGTNS DEYTATNSSI(ALVTSVWDAGGTDTFDFSGY
330301
KINLTAT S FSDVGGLVGNVS IAI(GYTTENAFGGSGI{DLI I GTiIqvA,.xiKGGAGI[DlIYGbxrNr,Nexar. sDvGGLKGNvs TAAGwvENAT GGscsDLL r GNDVA\TvLKGGAGND r Lyd
RTNLNEKSESDVGGLKG}I\TS IAAGW IENA,I GFR- QRLIVG}IAANLVLKGGAGTiIDVL FG
RINLNEGSESDVGGLKGNVSIAHGWTENAIGGSGND I LIGNGADN I LQGGAGDDVLYG
361AP-4
-
-- --
GGGADOTTIGG
GLGADQ1WGGAGAqT FVYGD IAES SAAAPDTLRDFVS GQDKTDLSGLDAFVNGGLVLQYV
390
450
KIDLSFENKKIDLSAFRN
hydrolyzes certain Arg-X bonds. It may binoteworthy thatthe cleavage occurred at the sites of paired arginine resi-dues. These results are largely consistent with thoseobtained with the MCA substrates.
The pH-rate profiles of the enzyme toward dynorphin A,BAM-12P and proalbumin(1-10) peptide are shown in Fig.4. Contrary to the results obtained with the peptide MCAsubstrates (Fig. 2b), the enzyme showed a broad pH/rateprofile with a single pH optimum at around pH 6 8 in boththe presence and absence of Co2+ ions. No Co'* ion activa-tion was observed and the activity was even reduced in thepresence of 1 mM Co2+ ions, as shown for dynorphin A (Fig.4a). Taken together, therefore, the apparently anomalous
FAP-2b-KIDLSGIGK
GGG}DEI.I{GG
STGADTI.TGG42L
FAP_1___--. KEGTSG--ESQrDAEAGKAGQA] LSYDAASXAGS LAI DFSGDAHADFAINL I GOATQADIW
NVTDLSVNIGGH
STTNLWLHEAGH
oxycarbonyl-Ala-Phe-Gly-Ala as a substrate; however,they found that the activation was mainly due to a decreasein the K, value rather than an increase in the hut value.
Figure 3 shows the cleavage sites of some peptides at pH8.0. The same results were obtained in both the presenceand absence of 1 mM Co2* ions. Although the peptides usedcontained several sites which are thought to be potentiallysusceptible to the enzyme, only the Arg-Ile bond in dlmor-phin, the Arg-Val bond in BAM-12P, the Arg-Arg bond inneurotensin, and the Arg-Asp bond in proalbumin(1-10)peptide were cleaved specifically, whereas no cleavage ofa-neoendorphin occurred under the conditions used. Theseresults seem to indicate that the enzyme preferentially
VoI. 121, No. 1, 1997

BB
pH-rate profile and Co'* ion activation observed with Boc-Arg-Val-Arg-Arg-MCA may be a specific phenomenon due
to the use of these small synthetic substrates.Among the various proteins examined as substrates, the
enzyrne hydrolyzed histone most rapidly at pH 8.0 in thepresence or absence of Co2* ions. The relative rates ofhydrolysis obtained under the conditions used were in thefollowing order: histone (100), myoglobin (39), bovineserum albumin (11), casein (8.9), ovalbumin (1.2), andy-immunoglobulin (0). The high activity toward histone isconsistent wth the trypsin-like specificity of the enzymefound with peptide substrates.
The inhibition profi.le of the protease with variousprotease inhibitors was examined at pH 8.0 in the presenceor absence of Co2+ ions and at pH 5.5 in the absence of Co'*ions, and the results were essentially the same, as shown inTable V. The protease was strongly inhibited by typicalmetal chelators such as EDTA and o-phenanthroline, whichindicates that it is a metalloprotease. The activity lost withthese metal chelators was restored partially (about 600/oforeach) on the addition of 1 mM Zn2+ or Co'* ions afterremoval of the chelator. Various inhibitors for serine andcysteine proteases failed to inhibit the enzyme. On theother hand, the protease was found to be markedly activat-ed by detergents such as lubrol and sodium deoxycholate inboth the presence and absence of Co'* ions; nearly three-fold activation was observed at detergent concentrations of10-3 to 1mM (data not shown). The reason for thisactivation is not certain.
NHz-Terminal and Internal Amino Acid Sequences-The NHz-terminal part of the protease and its internalsequences determined from the various fragments obtainedon lysylendopeptidase digestion are compared with thesequences of three serralysin family proteases: P. aeru-gino s a akaline protease (2, 3), S, n'LarceEcens protease ( 5),and, E. chrysanthemj protease (protease B) (6) in Fig. 5.Peptide AP-18 appears to include the first three residues ofthe zinc binding motif (His-Glu-X-X-His-X-X-GIy-X-X-His) (4 9, 24) of serralysin proteases. Three repeats of atypical calcium binding motif (Gly-Gly-X-Gly-X-Asp) werefound in the proteoly'tic fragments of the present enzyme.Overall identities of the partial sequences (total 193residues) thus determined were 65, 55, and 53% with thecorresponding sequences of the metalloproteases from P.aeruginosa, S. marcescens, and. E. crysantherni, respective-ly. Therefore, the present enzyme is suggested to belong tothis protease family, although the complete amino acidsequence and the zinc content of the present enzyme remainto be determined. Based on these data, an attempt to clonethe gene for the present enzyrne is underway.
We wish to thank Dr. Yuichi Tsuchiya for the helpful discussion on
this work.
REFERENCES
1. Maeda, H. and Morihara, K. (1995) Serralysin and relatedbacterial proteinases. Methods Enzymol. 248, 395-413
2. Okuda, K., Morihara, K., Atsumi, Y., Takeuchi, H., Kawamoto,S., Kawasaki, H., Suzuki, K., and Fukushima, J. (1990) Com-plete nucleotide sequence of the structural gene for alkalineproteinase from Pseudomonas aeruginoso IFO 3455. Infect.1mmun.58,4083 4088
H.-J.Kim et al.
3. Duong, F., Lazdunski, A., Cami, B., and Murgier, M. (1992)
Sequence of a ciuster of genes controlling s}'nthesis and secretionof alkaline protease ir Ps. aeruginoso: relationship to othersecretory pathways. Gene l2l, 47 - 54
4. Baumann, U., Wu, S., Flaherty, K. M., and McKay, B. (1993)
Three-dimensional structure of the alkaline protease of Pseudo'rhonas aeruginoso: a two-domain protein with a calcium bindingparallel beta roII motif. EMBO J. L2,3357 3364
5. Nakahama, K., Yoshimura, K., Marumoto, R., Kikuchi, M., Lee,I. S., Hase, T., and Matsubara, H. (1986) Cloning and sequencing
ol lhe Serratia protease gete. Nucleic Acids Res, 14,5843 58556. Delepelaire, P. and Wandersman, C. (1989) Protease secretion
by Eruinia chrysanthemi. Proteases B and C are synthesized andsecreted as zymogens without a signal peptide. J. Biol. Chem.
264, 9083-90897. Dahler, G.S., Bar:ras, F., andKeen, N.T. (1990) Cloningof genes
encoding extracellular metalloproteases from Erwinia chrysan'th.emiECl6. J. Bacteriol. 172, 5803 5815
8. Baumann, U. (1994) Crystal structure of the 50 kDa metallopro-tease from Serratia marcescens. J. MoL 8io1.242,244 251
9. Rawlings, N.D. and Barrett, A.J. (1995) Evolutionary famiiies ofmetallopeptidases. Methods Enzymol. 248, 183 228
10. Diermayr, P. and Klostermeyer, H. (1984) A new metallopro-teinase from Pseudornonas fluorescens biotype l. Hoppe-Seyler'sZ. Physiol. Chem..365,1345 1350
11. Diermayr, P., Kroll, S., and Klostermeyer, H. (1987) Influenceof EDTA and metal ions on a metalloproteinase from Pseudo-
monas fluorescezs biotype I. Biol. Chem. Hoppe'Seyler 368, 5761
12. Saito, H. and Miura, K. (1963) Preparation of transformingdeoxyribonucleic acid by phenol treatment. Bbchim. Biophys.Acta72,619 629
13. Mesbah, M., Premachandran, U., and Whitman, W'B. (1989)
Precise measurement of the G*C content of deoxyribonucleicacid by high-performance iiquid chromatography. Int. J. Syst.
Bacteriol. 39, 159-16714. Tamanoue, Y., Takahashi, T., and Takahashi, K' (1993) Purifica-
tion and characterization of two isoforms of serine proteinase
from the microsomal membranes of rat liver. J. Biochem. lI.3,229 235
15. Lineweaver, H. and Burk, D. (1934) The determination ofenzyme dissociation constants. J. Am. Chem. Soc' 56, 658 666
16. Smith, P.K., Krohn, R.I., Hermanson, G'T., Mallia, A.K.,Gartner, F.H., Provenzano, M.D., Fujimoto, E.K., Goeke, N.,Olson, B.J., and Klenk, D.C. (1985) Measurement of proteinusing bicinchoninic acid,. Anal. Biochem. 150, 76-85
17. Laemmli, U.K. (1970) Cleavage of structural proteins during theassembly of the head of bacteriophage "I 4. Nature 227, 680 685
18. Iwamatsu, A. (1992) S-Carboxymethylation of proteins transfer-red onto polyvinylidene difluoride membranes foilowed by ln sltzprotease digestion and amino acid sequencing. Electrophoresis 13,1,42 147
19. Iwamatsu, A. and Yoshida-Kubomura, N. (1996) Systematicpeptide fractionation of polyvinylidene difluoride (PVDF)'im-mobilized proteins prior to microsequencing. J. Biochern. 1".2O,
29"3420. Krieg, N.R. and Hott, J.G., eds. (1984) Bergey's Manual of
Sy stematic Bacteriology, Williams and Wilkins, Baltimore21. Matsumoto, K., Yamamoto, T., Kamata, R., and Maeda, H.
(1984) Pathogenesis of serratiai infection: the activation of theHageman factor-prekallikrein cascade by serratial protease' J.Biochem.96,739 749
22. Schechter, I. and Berger, A. (1967) On the size ofthe active sitein proteases. I. Papain. Biochetn. Biophys. Res. Commun. 27, 57162
23. Morihara, K. and Tsuzuki, H. (1974) Effect of cobalt ions on theenzymatic activity of Pseudomonas aeruginosa alkaline protein-ase. Agric. Biol. Chem.38,621 626
24. Hoopet, N.M. (1994) Families of zinc metailproteases' tr'XBS
Lett.354, L 6
J. Biochem.




















