
Accumulation, Activity and Localization of Cell Cycle Regulatory Proteins and the Chloroplast...
13
Accumulation, Activity and Localization of Cell Cycle Regulatory Proteins and the Chloroplast Division Protein FtsZ in the Alga Scenedesmus quadricauda under Inhibition of Nuclear DNA Replication Milada Vı´tova´ 1, 4 , Jana Hendrychova´ 1, 4 , Ma´ria C ˇ ı´ zˇ kova´ 1 , Vladislav Cepa´k 2 , James G. Umen 3 , Vile´m Zachleder 1, * and Katerˇina Bisˇova´ 1 1 Laboratory of Cell Cycles of Algae, Institute of Microbiology, ASCR, 37981 Trˇebon ˇ, Czech Republic 2 Algological Centre and Research Centre for Bioindication and Revitalization, Institute of Botany, Academy of Sciences of the Czech Republic, Trˇebon ˇ, Czech Republic 3 Plant Biology Laboratory, The Salk Institute for Biological Studies, La Jolla, CA 92037, USA Synchronized cultures of the green alga Scenedesmus quadricauda were grown in the absence (untreated cultures) or in the presence (FdUrd-treated cultures) of 5-fluorodeox- yuridine, the specific inhibitor of nuclear DNA replication. The attainment of commitment points, at which the cells become committed to nuclear DNA replication, mitosis and cellular division, and the course of committed processes themselves were determined for cell cycle characterization. FdUrd-treated cultures showed nearly unaffected growth and attainment of the commitment points, while DNA repli- cation(s), nuclear division(s) and protoplast fission(s) were blocked. Interestingly, the FdUrd-treated cells possessed a very high mitotic histone H1 kinase activity in the absence of any nuclear division(s). Compared with the untreated cultures, the kinase activity as well as mitotic cyclin B accumulation increased continuously to high values without any oscillation. Division of chloroplasts was not blocked but occurred delayed and over a longer time span than in the untreated culture. The FtsZ protein level in the FdUrd-treated culture did not exceed the level in the untreated culture, but rather, in contrast to the untreated culture, remained elevated. FtsZ structures were both localized around pyrenoids and spread inside of the chloroplast in the form of spots and mini-rings. The abun- dance and localization of the FtsZ protein were comparable in untreated and FdUrd-treated cells until the end of the untreated cell cycle. However, in the inhibitor-treated culture, the signal did not decrease and was localized in intense spots surrounding the chloroplast/cell perimeter; this was in agreement with both the elevated protein level and persisting chloroplast division. Keywords: Chloroplast fission — Cyclin-dependent kinase — Fluorodeoxyuridine — FtsZ protein — Mitotic cyclin B — Scenedesmus quadricauda. Abbreviations: BSA, bovine serum albumin; CDK, cyclin-dependent kinase; CP, commitment point; DTT, dithiothiothreitol; FdUrd, 5-fluorodeoxyuridine; FtsZ, Z-ring-forming chloroplast protein; GST, glutathione S-transferase; PBS, phosphate-buffered saline; ptDNA, chloroplast DNA. Introduction Dividing eukaryotes have to coordinate cellular and organellar divisions. While animal and fungal cells need to coordinate cell division with the division of mitochondria, plants and algae need to coordinate the divisions of the cell, mitochondria and chloroplast(s). Therefore, careful coordi- nation of cell multiplication with plastid multiplication and partition at cytokinesis is required to maintain the universal presence of plastids in the photosynthetic organisms (Heinhorst et al. 1985a, Heinhorst et al. 1985b, Coleman 1999). However, the mechanism underlining the control remains enigmatic (Miyagishima 2005). Since higher plants contain higher numbers of chlo- roplasts (reviewed in Butterfass 1995), algae containing only one chloroplast are useful models for the study of both organelle division and the coordination between the divi- sion of the cell and the chloroplast. Chloroplasts of green algae and higher plants arose from a primary endosymbiotic event between a cyanobacterium and a flagellate protist. Bacteria divide by binary fusion, directed by a Z-ring that is formed by FtsZ protein (Bi and Lutkenhaus 1991). Analogously, the chloroplasts divide by binary fission using a homolog of FtsZ protein (Lutkenhaus 1998, Margolin 2000). Homologs of bacterial FtsZ were found in many plant species (reviewed in Stokes et al. 2000) including the unicellular green algae Nannochloris bacillaris (Koide et al. 2004) and Chlamydomonas reinhardtii (Wang et al. 2003). Two types of FtsZ (FtsZ1 and FtsZ2) were iden- tified in plants (see overview in Stokes and Osteryoung 2003). Both of them are essential for chloroplast division *Corresponding author: E-mail, [email protected]; Fax, þ420-384340415. 4 These authors contributed equally to this work. Plant Cell Physiol. 49(12): 1805–1817 (2008) doi:10.1093/pcp/pcn162, available online at www.pcp.oxfordjournals.org ß The Author 2008. Published by Oxford University Press on behalf of Japanese Society of Plant Physiologists. All rights reserved. For permissions, please email: [email protected] 1805
-
Upload
independent -
Category
Documents
-
view
2 -
download
0
Transcript of Accumulation, Activity and Localization of Cell Cycle Regulatory Proteins and the Chloroplast...

Accumulation, Activity and Localization of Cell Cycle Regulatory Proteins
and the Chloroplast Division Protein FtsZ in the Alga Scenedesmus quadricaudaunder Inhibition of Nuclear DNA Replication
Milada Vıtova1, 4
, Jana Hendrychova1, 4
, Maria Cızkova1, Vladislav Cepak
2, James G. Umen
3,
Vilem Zachleder1,* and Katerina Bisova
1
1 Laboratory of Cell Cycles of Algae, Institute of Microbiology, ASCR, 37981 Trebon, Czech Republic2 Algological Centre and Research Centre for Bioindication and Revitalization, Institute of Botany, Academy of Sciencesof the Czech Republic, Trebon, Czech Republic3 Plant Biology Laboratory, The Salk Institute for Biological Studies, La Jolla, CA 92037, USA
Synchronized cultures of the green alga Scenedesmus
quadricauda were grown in the absence (untreated cultures)
or in the presence (FdUrd-treated cultures) of 5-fluorodeox-
yuridine, the specific inhibitor of nuclear DNA replication.
The attainment of commitment points, at which the cells
become committed to nuclear DNA replication, mitosis and
cellular division, and the course of committed processes
themselves were determined for cell cycle characterization.
FdUrd-treated cultures showed nearly unaffected growth
and attainment of the commitment points, while DNA repli-
cation(s), nuclear division(s) and protoplast fission(s) were
blocked. Interestingly, the FdUrd-treated cells possessed a
very high mitotic histone H1 kinase activity in the absence of
any nuclear division(s). Compared with the untreated cultures,
the kinase activity as well as mitotic cyclin B accumulation
increased continuously to high values without any oscillation.
Division of chloroplasts was not blocked but occurred delayed
and over a longer time span than in the untreated culture. The
FtsZ protein level in the FdUrd-treated culture did not exceed
the level in the untreated culture, but rather, in contrast to
the untreated culture, remained elevated. FtsZ structures
were both localized around pyrenoids and spread inside of the
chloroplast in the form of spots and mini-rings. The abun-
dance and localization of the FtsZ protein were comparable
in untreated and FdUrd-treated cells until the end of the
untreated cell cycle. However, in the inhibitor-treated culture,
the signal did not decrease and was localized in intense
spots surrounding the chloroplast/cell perimeter; this was in
agreement with both the elevated protein level and persisting
chloroplast division.
Keywords: Chloroplast fission — Cyclin-dependent kinase
— Fluorodeoxyuridine — FtsZ protein — Mitotic cyclin B
— Scenedesmus quadricauda.
Abbreviations: BSA, bovine serum albumin; CDK,
cyclin-dependent kinase; CP, commitment point; DTT,
dithiothiothreitol; FdUrd, 5-fluorodeoxyuridine; FtsZ,
Z-ring-forming chloroplast protein; GST, glutathione
S-transferase; PBS, phosphate-buffered saline; ptDNA,
chloroplast DNA.
Introduction
Dividing eukaryotes have to coordinate cellular and
organellar divisions. While animal and fungal cells need to
coordinate cell division with the division of mitochondria,
plants and algae need to coordinate the divisions of the cell,
mitochondria and chloroplast(s). Therefore, careful coordi-
nation of cell multiplication with plastid multiplication and
partition at cytokinesis is required to maintain the universal
presence of plastids in the photosynthetic organisms
(Heinhorst et al. 1985a, Heinhorst et al. 1985b, Coleman
1999). However, the mechanism underlining the control
remains enigmatic (Miyagishima 2005).
Since higher plants contain higher numbers of chlo-
roplasts (reviewed in Butterfass 1995), algae containing only
one chloroplast are useful models for the study of both
organelle division and the coordination between the divi-
sion of the cell and the chloroplast. Chloroplasts of green
algae and higher plants arose from a primary endosymbiotic
event between a cyanobacterium and a flagellate protist.
Bacteria divide by binary fusion, directed by a Z-ring that
is formed by FtsZ protein (Bi and Lutkenhaus 1991).
Analogously, the chloroplasts divide by binary fission using
a homolog of FtsZ protein (Lutkenhaus 1998, Margolin
2000). Homologs of bacterial FtsZ were found in many
plant species (reviewed in Stokes et al. 2000) including
the unicellular green algae Nannochloris bacillaris (Koide
et al. 2004) and Chlamydomonas reinhardtii (Wang et al.
2003). Two types of FtsZ (FtsZ1 and FtsZ2) were iden-
tified in plants (see overview in Stokes and Osteryoung
2003). Both of them are essential for chloroplast division
*Corresponding author: E-mail, [email protected]; Fax, þ420-384340415.
4These authors contributed equally to this work.
Plant Cell Physiol. 49(12): 1805–1817 (2008)doi:10.1093/pcp/pcn162, available online at www.pcp.oxfordjournals.org� The Author 2008. Published by Oxford University Press on behalf of Japanese Society of Plant Physiologists.All rights reserved. For permissions, please email: [email protected]
1805

(Osteryoung et al. 1998); they associate tightly to form the
Z-ring inside the inner envelope membrane, determining the
site of division (McAndrew et al. 2001, Kuroiwa et al. 2002,
Maple et al. 2005). In contrast to FtsZ2, the FtsZ1 protein is
believed not to be present in other structures (El-Kafafi
et al. 2005).
Scenedesmus quadricauda is a green monoplastidic alga
(Chlorophyceae) forming coenobia. The cells divide by
multiple fission; they grow during the G1 phase until they
reach a critical cell size for the attainment of the first
commitment point (CP). If the growth continues, the cells
can attain several additional CPs consecutively. At each
of the CPs the processes are triggered, leading, after a cer-
tain time interval, to initiation of the round (s) of DNA
replication(s) and later to nuclear division(s) (Setlık et al.
1972, Zachleder and Setlık 1990, Zachleder et al. 1997)
(Fig. 1). Therefore, the cells divide into 2n daughter cells,
where n is the number of CPs attained, and consequently
the number of doublings (Kates et al. 1968, Poyton and
Branton 1972, Setlık and Zachleder 1984).
The eukaryotic cell cycle is regulated by the activity of
cyclin-dependent kinases (CDKs) in a heterodimeric com-
plex with cyclins (for a review, see Norbury and Nurse 1992,
Nigg 1995, Pines 1996, Stern and Nurse 1996, Mironov
et al. 1999). Similarly the S. quadricauda cell cycle is regu-
lated by the activity of CDK-like kinases with histone H1
kinase activity. There are at least two different complexes
expressing CDK-like activity in S. quadricauda; one accom-
panies the attainment of CP(s), the other complex is able to
interact with suc1 protein and its activity is specific for
mitosis(es) (Bisova et al. 2000).
Scenedesmus quadricauda has been extensively studied
in an attempt to unravel the relationship between cell and
chloroplast divisions. It has been shown that chloroplast,
similarly to the nucleocytosolic compartment, undergoes
chloroplast DNA (ptDNA) replication, nucleoid division
and chloroplast fission (Zachleder and Cepak 1987b). The
processes of ptDNA replication and nucleoid division can
be uncoupled from nuclear DNA replication and division
by application of 5-fluorodeoxyuridine (FdUrd), an inhib-
itor of thymidylate synthase that in S. quadricauda specif-
ically inhibits the nuclear DNA replication and without
affecting the ptDNA replication (Zachleder 1994).
Here, we have blocked nuclear DNA replication
by FdUrd in order to uncouple nuclear and chloroplast
division cycles. We showed that chloroplast division could
be performed in the absence of nuclear and cellular divi-
sion, but it was delayed, slowed down and the number of
daughter chloroplasts often did not agree with 2n number
rule of multiple fissions (see above). Interestingly, treatment
with FdUrd caused extremely elevated steady-state levels
and activities of the cell cycle-regulating proteins mitotic
CDK-like kinase and cyclin B.
Results
Cell cycle progress
Synchronized cultures of S. quadricauda were cultivated
in either the presence or absence of FdUrd. The growth
in both variants was stopped by transfer of the cultures into
the dark after 12 h of the cell cycle. The increase in the cell
size during the light period was the same in both cultures
(Fig. 2). Also the total RNA level in the FdUrd-treated
G1
G1
G1 M G3G2 CS
C.P.
C.P.
C.P.
pS
pS S G2 G3M CG1
G2S
pS S G2 M G3 C
Classical cell cycle model
Scenedesmus binary fission
Scenedesmus multiple fission
A
B
C
M
Fig. 1 Diagrams showing different types of cell cycle phasesincluding the classical cell cycle model (A) for binary fission(Howard and Pelc 1953) and those found in the alga Scenedesmusthat divide into two (B) and four (C) daughter cells. Individual barsshow the sequence of cell cycle phases during which growth andreproductive processes take place. Whereas only one sequence ofevents leading to the duplication of cell structures occurs duringthe cell cycle of cells dividing into two daughter cells (A, B), twopartially overlapping sequences of growth and reproductive eventsoccur within a single cycle in cells dividing by multiple fission intofour daughter cells (C). Two bars (C) illustrate the simultaneouscourse of different phases from two consecutively startedsequences of growth and reproductive events. Schematic picturesof the cells indicate their size during the cell cycle, and the blackspots inside illustrate the size and number of nuclei. Large blackspots indicate the doubling of the DNA. G1: the phase duringwhich the threshold size of the cell is attained. It can be called apre-commitment period because it is terminated when thecommitment point is reached. CP: the stage in the cell cycle atwhich the cell becomes committed to triggering and terminatingthe sequence of processes leading to the duplication of reproduc-tive structures. pS: the pre-replication phase between the CP andthe beginning of DNA replication. The processes required for theinitiation of DNA replication are assumed to happen during thisphase. S: the phase during which DNA replication takes place. G2:the phase between the termination of DNA replication and the startof mitosis. Processes leading to the initiation of mitosis are assumedto take place during this phase. M: the phase during which nucleardivision occurs. G3: the phase between nuclear division and celldivision. The processes leading to cellular division are assumed totake place during this phase. C: the phase during which cellcleavage occurs. Modified after Zachleder et al. (1997).
1806 Chloroplast fission in Scenedesmus
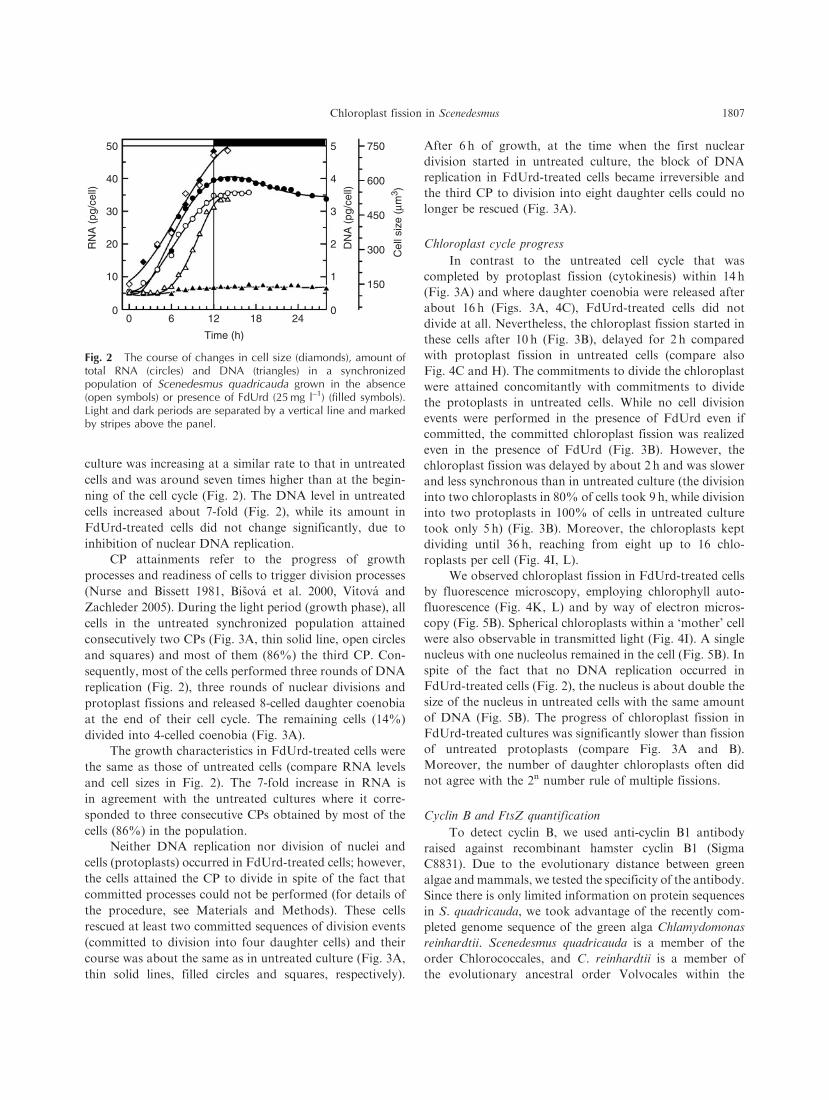
culture was increasing at a similar rate to that in untreated
cells and was around seven times higher than at the begin-
ning of the cell cycle (Fig. 2). The DNA level in untreated
cells increased about 7-fold (Fig. 2), while its amount in
FdUrd-treated cells did not change significantly, due to
inhibition of nuclear DNA replication.
CP attainments refer to the progress of growth
processes and readiness of cells to trigger division processes
(Nurse and Bissett 1981, Bisova et al. 2000, Vıtova and
Zachleder 2005). During the light period (growth phase), all
cells in the untreated synchronized population attained
consecutively two CPs (Fig. 3A, thin solid line, open circles
and squares) and most of them (86%) the third CP. Con-
sequently, most of the cells performed three rounds of DNA
replication (Fig. 2), three rounds of nuclear divisions and
protoplast fissions and released 8-celled daughter coenobia
at the end of their cell cycle. The remaining cells (14%)
divided into 4-celled coenobia (Fig. 3A).
The growth characteristics in FdUrd-treated cells were
the same as those of untreated cells (compare RNA levels
and cell sizes in Fig. 2). The 7-fold increase in RNA is
in agreement with the untreated cultures where it corre-
sponded to three consecutive CPs obtained by most of the
cells (86%) in the population.
Neither DNA replication nor division of nuclei and
cells (protoplasts) occurred in FdUrd-treated cells; however,
the cells attained the CP to divide in spite of the fact that
committed processes could not be performed (for details of
the procedure, see Materials and Methods). These cells
rescued at least two committed sequences of division events
(committed to division into four daughter cells) and their
course was about the same as in untreated culture (Fig. 3A,
thin solid lines, filled circles and squares, respectively).
After 6 h of growth, at the time when the first nuclear
division started in untreated culture, the block of DNA
replication in FdUrd-treated cells became irreversible and
the third CP to division into eight daughter cells could no
longer be rescued (Fig. 3A).
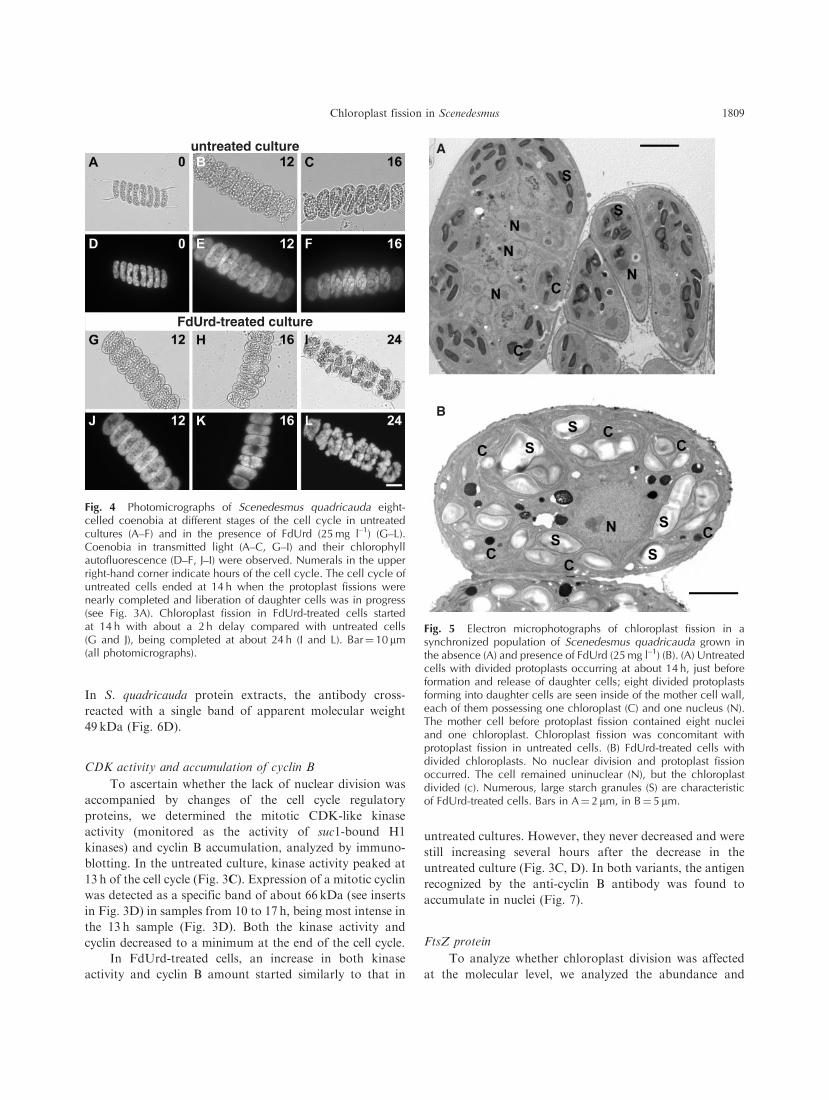
Chloroplast cycle progress
In contrast to the untreated cell cycle that was
completed by protoplast fission (cytokinesis) within 14 h
(Fig. 3A) and where daughter coenobia were released after
about 16 h (Figs. 3A, 4C), FdUrd-treated cells did not
divide at all. Nevertheless, the chloroplast fission started in
these cells after 10 h (Fig. 3B), delayed for 2 h compared
with protoplast fission in untreated cells (compare also
Fig. 4C and H). The commitments to divide the chloroplast
were attained concomitantly with commitments to divide
the protoplasts in untreated cells. While no cell division
events were performed in the presence of FdUrd even if
committed, the committed chloroplast fission was realized
even in the presence of FdUrd (Fig. 3B). However, the
chloroplast fission was delayed by about 2 h and was slower
and less synchronous than in untreated culture (the division
into two chloroplasts in 80% of cells took 9 h, while division
into two protoplasts in 100% of cells in untreated culture
took only 5 h) (Fig. 3B). Moreover, the chloroplasts kept
dividing until 36 h, reaching from eight up to 16 chlo-
roplasts per cell (Fig. 4I, L).
We observed chloroplast fission in FdUrd-treated cells
by fluorescence microscopy, employing chlorophyll auto-
fluorescence (Fig. 4K, L) and by way of electron micros-
copy (Fig. 5B). Spherical chloroplasts within a ‘mother’ cell
were also observable in transmitted light (Fig. 4I). A single
nucleus with one nucleolus remained in the cell (Fig. 5B). In
spite of the fact that no DNA replication occurred in
FdUrd-treated cells (Fig. 2), the nucleus is about double the
size of the nucleus in untreated cells with the same amount
of DNA (Fig. 5B). The progress of chloroplast fission in
FdUrd-treated cultures was significantly slower than fission
of untreated protoplasts (compare Fig. 3A and B).
Moreover, the number of daughter chloroplasts often did
not agree with the 2n number rule of multiple fissions.
Cyclin B and FtsZ quantification
To detect cyclin B, we used anti-cyclin B1 antibody
raised against recombinant hamster cyclin B1 (Sigma
C8831). Due to the evolutionary distance between green
algae andmammals, we tested the specificity of the antibody.
Since there is only limited information on protein sequences
in S. quadricauda, we took advantage of the recently com-
pleted genome sequence of the green alga Chlamydomonas
reinhardtii. Scenedesmus quadricauda is a member of the
order Chlorococcales, and C. reinhardtii is a member of
the evolutionary ancestral order Volvocales within the
50
40
30
20RN
A (
pg/c
ell)
DN
A (
pg/c
ell)
Cel
l siz
e (µ
m3 )
10
0
5 750
600
450
300
150
4
3
2
1
00 6 12
Time (h)
18 24
Fig. 2 The course of changes in cell size (diamonds), amount oftotal RNA (circles) and DNA (triangles) in a synchronizedpopulation of Scenedesmus quadricauda grown in the absence(open symbols) or presence of FdUrd (25 mg l–1) (filled symbols).Light and dark periods are separated by a vertical line and markedby stripes above the panel.
Chloroplast fission in Scenedesmus 1807

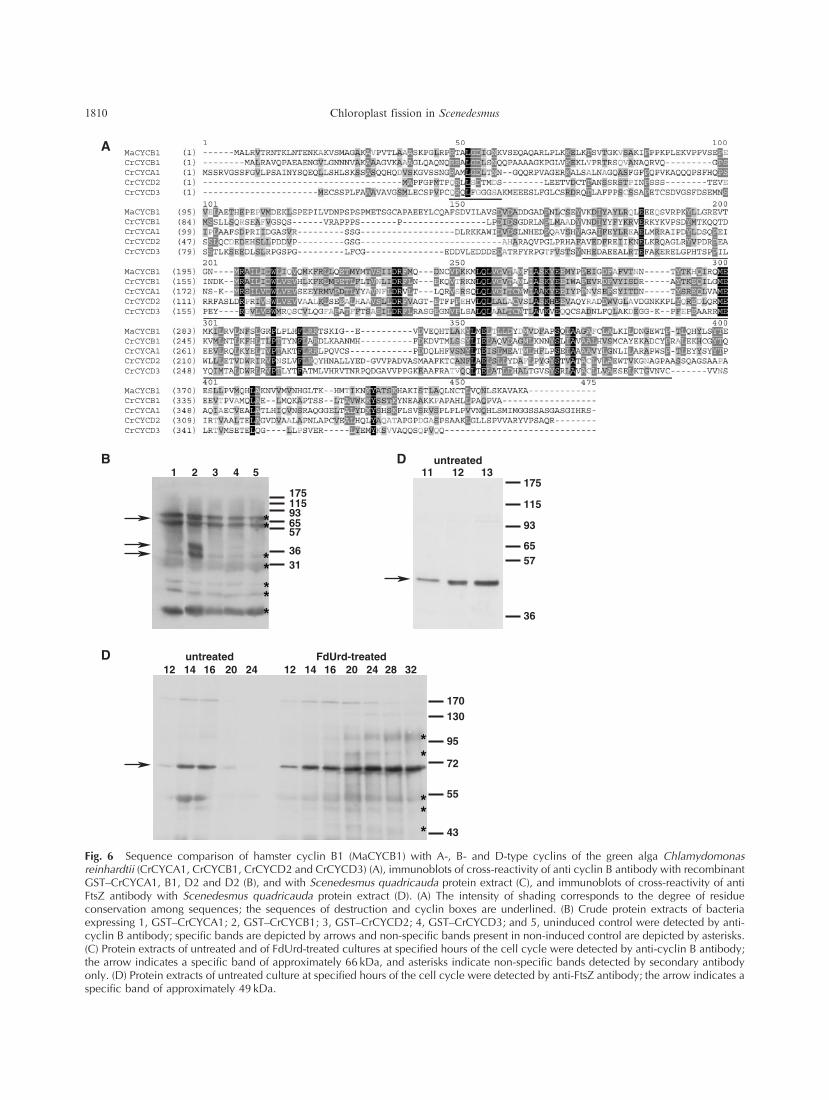
Chlorophyta division (van den Hoek et al. 1995). Sequence
comparison of hamster cyclin B1 protein sequence with
sequences of C. reinhardtii A1, B1, D2 and D3 cyclins shows
that there is high sequence similarity among all the sequences
within the cyclin box (Fig. 6A, underlined); hamster cyclin
B1 and C. reinhardtii cyclins A1 and B1 also contain
destruction boxes (Fig. 6A, underlined). There is higher
similarity between hamster cyclin B1 and C. reinhardtii
cyclins A1 and B1 as compared withC. reinhardtii cyclins D2
and D3; the highest similarity is between the two B-type
cyclins (Fig. 6A).
In order to test the antibody specificity more directly,
we produced recombinant glutathione S-transferase (GST)-
tagged versions of C. reinhardtii A1, B1, D2 and D3 cyclins.
The antibody cross-reacted only with recombinant GST–
CrCYCB1 and not with any other cyclins (Fig. 6B). It
recognized both the full-length version of GST–CrCYCB1
of predicted molecular weight 72 kDa and truncated ver-
sions of the recombinant protein of approximately 40 and
35 kDa (Fig. 6B). In S. quadricauda protein extracts, the
antibody cross-reacted with a major band of apparent
molecular weight 66 kDa (Fig. 6C).
FtsZ antibody was raised against FtsZ1 protein of
the unicellular green alga N. bacillaris (Koide et al. 2004).
100A
B
C
D
E
50
0
100
50
0
200
150
100
50
0
30
20
10
0
100
80
60
40
20
00 6 12 18
Time (h)
24 30 36
Cel
l fra
ctio
n (c
umul
ativ
e %
)K
inas
e ac
tivity
(r.u
.)C
yclin
am
ount
(r.u
.)F
tsZ
1 am
ount
(r.u
.)
Fig. 3 Time courses of the attainment of CPs to division eventsand division events themselves (mitoses, protoplast fission,daughter cell release) (A), the attainment of CPs to chloroplastfissions and chloroplast fissions themselves (B), mitotic (suc1-bound) CDK activity (C), amount of cyclin B (D), and amount ofchloroplast FtsZ protein (E) in a synchronized population ofScenedesmus quadricauda grown in the absence (open symbols)or presence of FdUrd (25 mg l–1) (filled symbols). (A) Solid thinlines: percentage of untreated (open symbols) and FdUrd-treated(filled symbols) cells that attained the commitment to divide into
two (circles), four (squares) and eight (triangles) daughter cells;solid thick lines, dotted symbols: percentage of untreated cells inwhich nuclear divisions in two (circles), four (squares) and eight(triangles) nuclei were completed; dashed lines, open symbols:percentage of untreated cells in which protoplast fissions in two(circles), four (squares) and eight (triangles) protoplasts werecompleted; dotted lines (crosses): percentage of untreated cellsthat released daughter coenobia. No nuclear division, protoplastfission and daughter cell release occurred in FdUrd-treated cells.The potential of these cells to divide (commitment point) wasdetermined in cells in which FdUrd was removed by rinsing andreplaced by an excess of thymidine prior to transfer to darkness. (B)Solid lines: percentage of FdUrd-treated cells that attained thecommitment to divide their chloroplast (in the presence of FdUrd)into two (circles), four (squares) and eight (triangles) daughterchloroplasts. Dashed lines: percentage of FdUrd-treated cellsthat contained at least two (circles), four (squares) and eight ormore (triangles) chloroplasts. (C) Mitosis-specific CDK-like kinaseactivity of kinases purified by affinity chromatography on suc1protein determined by phosphorylation of externally supplementedhistone H1 as substrate. A representative autoradiogram showingthe variation of mitotic histone H1 kinase activity for untreated(upper stripe) and FdUrd-treated (lower stripe) cultures is shownas an insert. (D) The amount of cyclin B protein detected by cyclinB-specific antibody (for specificity of the antibody see Fig. 6A–C).A representative immunoblot with the antigen (�66 kDa) variationin untreated (upper stripe) and FdUrd-treated (lower stripe) culturesis shown as an insert. (E) The amount of chloroplast FtsZ proteindetected by FtsZ-specific antibody (for specificity of the antibodysee Fig. 6D). A representative immunoblot with the antigen(�49 kDa) variation in untreated (upper stripe) and FdUrd-treated(lower stripe) cultures is shown as an insert. Light and darkperiods are separated by a vertical line and marked by stripesabove the top panel.
1808 Chloroplast fission in Scenedesmus
In S. quadricauda protein extracts, the antibody cross-
reacted with a single band of apparent molecular weight
49 kDa (Fig. 6D).
CDK activity and accumulation of cyclin B
To ascertain whether the lack of nuclear division was
accompanied by changes of the cell cycle regulatory
proteins, we determined the mitotic CDK-like kinase
activity (monitored as the activity of suc1-bound H1
kinases) and cyclin B accumulation, analyzed by immuno-
blotting. In the untreated culture, kinase activity peaked at
13 h of the cell cycle (Fig. 3C). Expression of a mitotic cyclin
was detected as a specific band of about 66 kDa (see inserts
in Fig. 3D) in samples from 10 to 17 h, being most intense in
the 13 h sample (Fig. 3D). Both the kinase activity and
cyclin decreased to a minimum at the end of the cell cycle.
In FdUrd-treated cells, an increase in both kinase
activity and cyclin B amount started similarly to that in
untreated cultures. However, they never decreased and were
still increasing several hours after the decrease in the
untreated culture (Fig. 3C, D). In both variants, the antigen
recognized by the anti-cyclin B antibody was found to
accumulate in nuclei (Fig. 7).
FtsZ protein
To analyze whether chloroplast division was affected
at the molecular level, we analyzed the abundance and
A
B
Fig. 5 Electron microphotographs of chloroplast fission in asynchronized population of Scenedesmus quadricauda grown inthe absence (A) and presence of FdUrd (25 mg l–1) (B). (A) Untreatedcells with divided protoplasts occurring at about 14 h, just beforeformation and release of daughter cells; eight divided protoplastsforming into daughter cells are seen inside of the mother cell wall,each of them possessing one chloroplast (C) and one nucleus (N).The mother cell before protoplast fission contained eight nucleiand one chloroplast. Chloroplast fission was concomitant withprotoplast fission in untreated cells. (B) FdUrd-treated cells withdivided chloroplasts. No nuclear division and protoplast fissionoccurred. The cell remained uninuclear (N), but the chloroplastdivided (c). Numerous, large starch granules (S) are characteristicof FdUrd-treated cells. Bars in A¼ 2 mm, in B¼ 5 mm.
untreated culture
FdUrd-treated culture
Fig. 4 Photomicrographs of Scenedesmus quadricauda eight-celled coenobia at different stages of the cell cycle in untreatedcultures (A–F) and in the presence of FdUrd (25 mg l–1) (G–L).Coenobia in transmitted light (A–C, G–I) and their chlorophyllautofluorescence (D–F, J–I) were observed. Numerals in the upperright-hand corner indicate hours of the cell cycle. The cell cycle ofuntreated cells ended at 14 h when the protoplast fissions werenearly completed and liberation of daughter cells was in progress(see Fig. 3A). Chloroplast fission in FdUrd-treated cells startedat 14 h with about a 2 h delay compared with untreated cells(G and J), being completed at about 24 h (I and L). Bar¼ 10 mm(all photomicrographs).
Chloroplast fission in Scenedesmus 1809
MaCYCB11 50 100
CrCYCB1CrCYCA1CrCYCD2CrCYCD3
MaCYCB1CrCYCB1CrCYCA1CrCYCD2CrCYCD3
MaCYCB1CrCYCB1CrCYCA1CrCYCD2CrCYCD3
MaCYCB1CrCYCB1CrCYCA1CrCYCD2CrCYCD3
MaCYCB1CrCYCB1CrCYCA1CrCYCD2CrCYCD3
1 2 3 4 5 11 12untreated
13175
115
93
6557
36
175115936557
3631
12 14 16untreated FdUrd-treated
20 24 12 14 16 20 24 28 32
170130
95
72
55
43
A
B
D
D
Fig. 6 Sequence comparison of hamster cyclin B1 (MaCYCB1) with A-, B- and D-type cyclins of the green alga Chlamydomonasreinhardtii (CrCYCA1, CrCYCB1, CrCYCD2 and CrCYCD3) (A), immunoblots of cross-reactivity of anti cyclin B antibody with recombinantGST–CrCYCA1, B1, D2 and D2 (B), and with Scenedesmus quadricauda protein extract (C), and immunoblots of cross-reactivity of antiFtsZ antibody with Scenedesmus quadricauda protein extract (D). (A) The intensity of shading corresponds to the degree of residueconservation among sequences; the sequences of destruction and cyclin boxes are underlined. (B) Crude protein extracts of bacteriaexpressing 1, GST–CrCYCA1; 2, GST–CrCYCB1; 3, GST–CrCYCD2; 4, GST–CrCYCD3; and 5, uninduced control were detected by anti-cyclin B antibody; specific bands are depicted by arrows and non-specific bands present in non-induced control are depicted by asterisks.(C) Protein extracts of untreated and of FdUrd-treated cultures at specified hours of the cell cycle were detected by anti-cyclin B antibody;the arrow indicates a specific band of approximately 66 kDa, and asterisks indicate non-specific bands detected by secondary antibodyonly. (D) Protein extracts of untreated culture at specified hours of the cell cycle were detected by anti-FtsZ antibody; the arrow indicates aspecific band of approximately 49 kDa.
1810 Chloroplast fission in Scenedesmus

localization of the FtsZ protein, which is known to be
involved in chloroplast division. The amount of protein
synthesized during the cell cycle was analyzed by immuno-
blotting in both the untreated and FdUrd-treated cells. The
amount of FtsZ protein increased during the growth phase.
It reached its maximum after 12 h and was decreasing
after 16 h in untreated cultures. In FdUrd-treated cultures,
FtsZ did not exceed the level in untreated cultures; it
increased more slowly and did not decrease (Fig. 3E).
The intracellular localization of FtsZ was examined
by indirect immunofluorescence staining on cross-sections.
The pattern of the fluorescence signal differed remarkably
among samples from different time periods. At the begin-
ning of the cell cycle, we observed spots and tiny rings
(Fig. 8A).
Later in the growth phase, at 7 h, at around the time
when the second nuclear division occurred in the untreated
culture, FtsZ was arranged in circles beside spots (one per
cell) in both cultures (Fig. 8B, E, arrowheads). Immunogold
labeling showed localization of the antigen at the pyrenoid
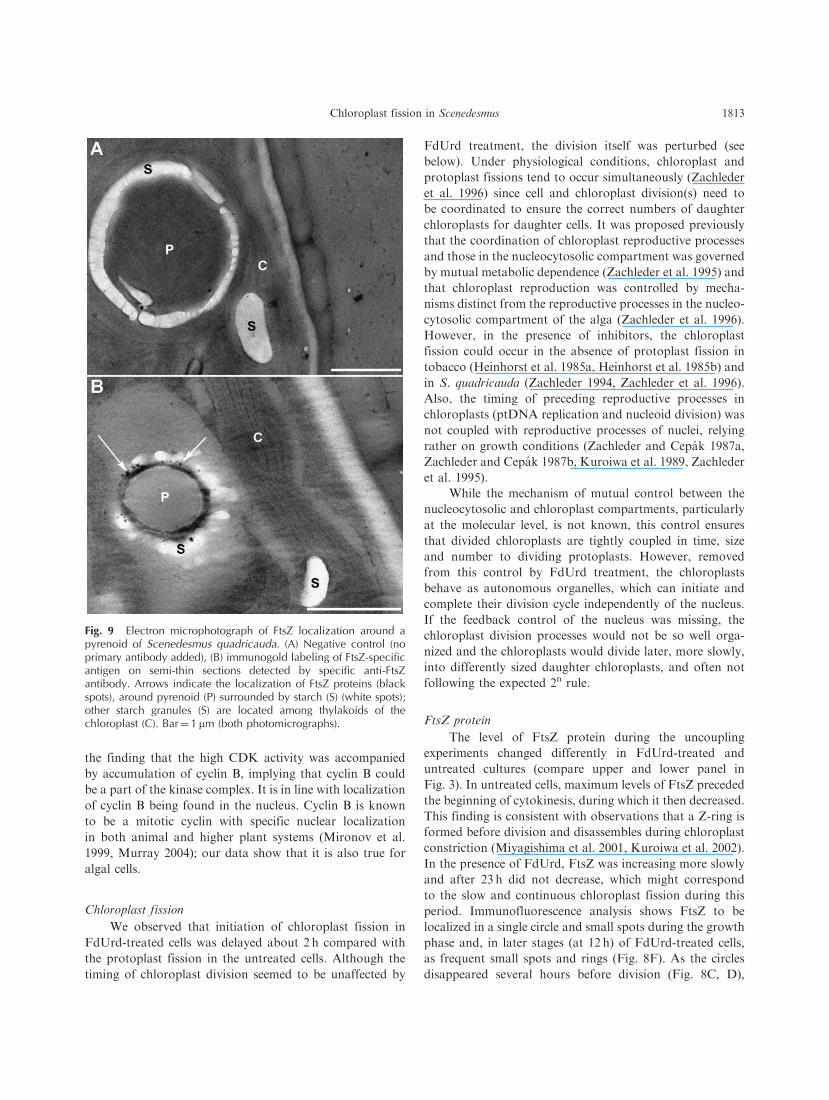
edge (Fig. 9); therefore, the circle observed in cross-sections
by immunofluorescence could represent the FtsZ structures
surrounding the pyrenoid and not real rings.
At 12 h in both the untreated and FdUrd-treated cell
cycle, the circles occurred only rarely and the small rings
became diffuse (Fig. 8C). Two hours later, the signal in
untreated cells became weak and FtsZ was localized in
dispersed fine spots (Fig. 8D), In contrast, in FdUrd-treated
cells, the signal stayed strong until 24 h, in agreement with
the continuously high protein level of FtsZ (Fig. 3E). FtsZ
was localized to intense spots surrounding the chloroplast/
cell perimeter (Fig. 8G, H).
Discussion
Processes related to the growth of the cells
The FdUrd-treated cells in our experiments grew
comparably (Fig. 3A) with untreated cells and performed
chloroplast division, whilst the DNA replication division
sequences in the nucleocytosolic compartment were not
performed (Fig. 3B). If the cells had been grown under
continuous illumination in the presence of FdUrd, they
would have attained a giant size (Zachleder et al. 1996). In
the present experiments, the growth to abnormal size was
prevented by putting the culture in darkness after 12 h of
growth in light, just at the time when liberation of daughter
cells in the untreated culture started (Fig. 3A). The size of
nuclei in FdUrd-treated cells increased during the growth
so that, at the time when untreated cell divided giving rise
to eight daughter protoplasts with eight nuclei, their size
was about double that of untreated nuclei (Fig. 5). How-
ever, the cell cycle positions of the nuclei differed. While the
untreated nuclei were at the beginning of G1 phase, the
FdUrd-treated nucleus was at the stage just before entry
into S phase which was prevented due to DNA replication
block. The nuclei in FdUrd-treated cells were in a state of
very intensive transcriptional activity since transcription
from a single genome had to ensure a level of transcription
ensured in untreated cells of similar size by 4–8 nuclei. The
size increase of nuclei during G1 phase prior to S phase
Fig. 7 Immunolocalization of cyclin B in Scenedesmus quad-ricauda in untreated (A) and FdUrd- (25 mg l–1) treated (B) culture.The position of the antigen in semi-thin sections was detected byanti-cyclin B antibody, recognizing a specific band of approxi-mately 66 kDa, and visualized by Cy3-conjugated secondaryantibody. Arrows indicate the localization of mitotic cyclin innuclei. Bar¼ 10 mm.
Chloroplast fission in Scenedesmus 1811

has also been described by other authors (for a review, see
Yen and Pardee, 1979).
CDK kinase activity and accumulation of cyclin B
Incompletely replicated or damaged DNA activates in
yeasts, mammals and higher plants a DNA damage/
replication checkpoint, which prevents mitosis by inhibition
of CDK activity (Pelayo et al. 2001, Bartek and Lukas 2007,
Calonge and O’Connell 2008). A delay in the mitotic kinase
activity after DNA damage was also reported in the uni-
cellular green alga C. reinhardtii (Slaninova et al. 2003). The
finding that the CDK activity of S. quadricauda was not
inhibited but, on the contrary, increased to a very high level
(Fig. 3C) indicated that in the presence of FdUrd the DNA
replication did not start at all and the cell therefore showed
neither incompletely replicated nor damaged DNA. The
checkpoint response to these events therefore appears to
have been intact. Indeed, it has been shown in budding
yeast that in the absence of stalled replication forks, the
completion of DNA replication prior to mitosis is not under
the surveillance of known checkpoints (Torres-Rosell et al.
2007). FdUrd blocks the synthesis of thymidine, the sub-
strate for DNA replication, but does not affect the structure
and function of DNA itself. Removing the inhibitor by
rinsing and subsequent addition of thymidine allowed
the normal progress and completion of the cell cycle of
S. quadricauda; this phenomenon was used for determina-
tion of CPs in FdUrd-treated cultures (see Materials and
Methods and Fig, 3A). Moreover, the synchronous culture,
to which FdUrd was added at the beginning of the cell cycle
together with thymidine, went through the cell cycle as
an untreated culture (unpublished results), implying that the
cell cycle progression was undisturbed without activated
checkpoints.
Mitotic CDK–cyclin complex activity in cells is inac-
tivated during mitosis due to degradation of cyclin through
ubiquitin-mediated proteolysis (Murray et al. 1989, Glotzer
et al. 1991, Murray 1995, Murray 2004). While this is a
plausible explanation for the decrease of both mitotic kinase
activity and cyclin B amount after mitosis in untreated cells,
neither a decrease of kinase activity nor a decrease of cyclin
B abundance occurred in FdUrd-treated cultures (Fig. 3D).
On the contrary, an abnormal increase in the CDK–cyclin
complex activity was observed, indicating that the mitotic
checkpoint checking degradation of cyclin after mitosis was
intact and activated (Pines 2006). This was confirmed by
untreated culture
FdUrd-treated culture
Fig. 8 Immunolocalization of FtsZ antigen in Scenedesmus quadricauda at different stages of the cell cycle in untreated cultures (A–D)and in the presence of FdUrd (25 mg l–1) (E–H). The position of antigen in semi-thin sections was detected by anti-FtsZ antibody,recognizing a specific band of approximately 49 kDa, and visualized by Cy3-conjugated secondary antibody. Numerals in the upper right-hand corner indicate the hour of the cell cycle. No substantial difference was observable within the first 12 h of the cell cycle between theuntreated and FdUrd-treated cultures. After 14 h the untreated cells divided their protoplasts including chloroplasts, and the FtsZ structureschanged into small spots or tiny rings (D). No division occurred in FdUrd-treated cells, the signal did not decrease and FtsZ was localizedto intense spots surrounding the chloroplast/cell perimeter (G and H). Arrowheads indicate minicircles surrounding pyrenoids. Bar¼ 10 mm(all photomicrographs).
1812 Chloroplast fission in Scenedesmus
the finding that the high CDK activity was accompanied
by accumulation of cyclin B, implying that cyclin B could
be a part of the kinase complex. It is in line with localization
of cyclin B being found in the nucleus. Cyclin B is known
to be a mitotic cyclin with specific nuclear localization
in both animal and higher plant systems (Mironov et al.
1999, Murray 2004); our data show that it is also true for
algal cells.
Chloroplast fission
We observed that initiation of chloroplast fission in
FdUrd-treated cells was delayed about 2 h compared with
the protoplast fission in the untreated cells. Although the
timing of chloroplast division seemed to be unaffected by
FdUrd treatment, the division itself was perturbed (see
below). Under physiological conditions, chloroplast and
protoplast fissions tend to occur simultaneously (Zachleder
et al. 1996) since cell and chloroplast division(s) need to
be coordinated to ensure the correct numbers of daughter
chloroplasts for daughter cells. It was proposed previously
that the coordination of chloroplast reproductive processes
and those in the nucleocytosolic compartment was governed
by mutual metabolic dependence (Zachleder et al. 1995) and
that chloroplast reproduction was controlled by mecha-
nisms distinct from the reproductive processes in the nucleo-
cytosolic compartment of the alga (Zachleder et al. 1996).
However, in the presence of inhibitors, the chloroplast
fission could occur in the absence of protoplast fission in
tobacco (Heinhorst et al. 1985a, Heinhorst et al. 1985b) and
in S. quadricauda (Zachleder 1994, Zachleder et al. 1996).
Also, the timing of preceding reproductive processes in
chloroplasts (ptDNA replication and nucleoid division) was
not coupled with reproductive processes of nuclei, relying
rather on growth conditions (Zachleder and Cepak 1987a,
Zachleder and Cepak 1987b, Kuroiwa et al. 1989, Zachleder
et al. 1995).
While the mechanism of mutual control between the
nucleocytosolic and chloroplast compartments, particularly
at the molecular level, is not known, this control ensures
that divided chloroplasts are tightly coupled in time, size
and number to dividing protoplasts. However, removed
from this control by FdUrd treatment, the chloroplasts
behave as autonomous organelles, which can initiate and
complete their division cycle independently of the nucleus.
If the feedback control of the nucleus was missing, the
chloroplast division processes would not be so well orga-
nized and the chloroplasts would divide later, more slowly,
into differently sized daughter chloroplasts, and often not
following the expected 2n rule.
FtsZ protein
The level of FtsZ protein during the uncoupling
experiments changed differently in FdUrd-treated and
untreated cultures (compare upper and lower panel in
Fig. 3). In untreated cells, maximum levels of FtsZ preceded
the beginning of cytokinesis, during which it then decreased.
This finding is consistent with observations that a Z-ring is
formed before division and disassembles during chloroplast
constriction (Miyagishima et al. 2001, Kuroiwa et al. 2002).
In the presence of FdUrd, FtsZ was increasing more slowly
and after 23 h did not decrease, which might correspond
to the slow and continuous chloroplast fission during this
period. Immunofluorescence analysis shows FtsZ to be
localized in a single circle and small spots during the growth
phase and, in later stages (at 12 h) of FdUrd-treated cells,
as frequent small spots and rings (Fig. 8F). As the circles
disappeared several hours before division (Fig. 8C, D),
Fig. 9 Electron microphotograph of FtsZ localization around apyrenoid of Scenedesmus quadricauda. (A) Negative control (noprimary antibody added), (B) immunogold labeling of FtsZ-specificantigen on semi-thin sections detected by specific anti-FtsZantibody. Arrows indicate the localization of FtsZ proteins (blackspots), around pyrenoid (P) surrounded by starch (S) (white spots);other starch granules (S) are located among thylakoids of thechloroplast (C). Bar¼ 1 mm (both photomicrographs).
Chloroplast fission in Scenedesmus 1813

they are probably not real Z-rings. As shown in Fig. 9, FtsZ
protein is accumulated around the pyrenoid, thus the large
rings could be a projection of an actual spherical arrange-
ment of FtsZ proteins. It is also in line with the observed
dissolution of the pyrenoids just at the time when the large
single rings disappeared. Similarly, FtsZ proteins that were
not involved in the Z-ring were previously detected in the
red alga Cyanidioschyzon merolae (Miyagishima et al. 2001).
Small spots and rings (‘mini FtsZ-rings’) were recently also
observed inside chloroplasts of Arabidopsis thaliana and
were suggested not to be associated with the chloroplast
membrane (Maple et al. 2005).
Materials and Methods
Experimental organism, culture growth conditions and cell cyclesynchronization
The chlorococcal alga S. quadricauda (Turp.) Breb., strainGreifswald/15 was obtained from the Culture Collection ofAutotrophic Microorganisms kept at the Institute of Botany,Trebon, Czech Republic. The cultures were synchronized by atleast two cycles of alternating light and dark periods (14 h : 10 h).The suspensions of synchronous cells (106 cells ml–1) were grown at308C in the inorganic nutrient medium described by Zachleder andSetlık (1982), aerated with air containing 2% (v/v) CO2 andilluminated by OSRAM L36/41fluorescent tubes; the light intensityat the surface of culture vessels was 490 mmol m–2 s–1. To inhibitDNA replication, FdUrd, the inhibitor of thymidylate synthase,was added at the beginning of the experiments after re-illuminationof the previously synchronized cultures to the final concentrationof 25mg l–1.
Determination of total DNA and RNA amount
Total nucleic acids were extracted according to Wanka(1962), as modified by Lukavsky et al. (1973). The DNA assaywas carried out as described by Decallonne and Weyns (1976), withthe modifications of Zachleder (1984); see also Zachleder (1995).
Assessment of cell cycle curves
Commitment point curves. Samples were taken from thesynchronous culture hourly and incubated under aeration at 308Cin the dark. At the end of the cell cycle, the percentages ofbinuclear daughter cells, 4- and 8-celled daughter coenobia, andundivided mother cells were assessed and plotted against the timeof the transfer in darkness. The ‘commitment curves’ illustratedthe course of attainment of individual CPs during the cell cyclein synchronous culture (John 1984, Setlık and Zachleder 1984,Zachleder et al. 1997). Because of blocked nuclear DNA repli-cation in FdUrd-treated cells, in the presence of FdUrd it was onlypossible to determine commitment to chloroplast division. Thiswas carried out as described for the commitment to nuclear andcellular division in untreated cells.
To follow the potential for cell division even in FdUrd-treatedcells, the FdUrd was washed off before transfer to darkness andexcess thymidine (250mg l–1) was added to the fresh medium tomake thymidylate synthase potentially damaged by FdUrd treat-ment dispensable.
Nuclear divisions. Nuclei were stained by SYBR Green I dye(Molecular Probes, Eugene, OR, USA) and observed througha fluorescence microscope using the technique introduced by
Vıtova et al. (2005). The percentages of mono-, bi-, tetra- andoctonuclear cells were estimated and plotted on the same graphwith the other cell cycle curves.
Cell division curves. Samples were fixed in 0.25% glutaralde-hyde. The percentage of undivided mother cells, mother cellsdivided into two, four or eight protoplasts, and daughter coenobiawere determined. Protoplast fission curves and daughter cellrelease curves were obtained by plotting the percentages as afunction of time.
Protein extraction
Samples of cell suspensions were centrifuged and the pelletswere washed with SCE buffer [100mM sodium citrate, 2.7mMEDTA-Na2, pH 7 (citric acid)]. The pellets were frozen and storedat –808C. Protein extracts were prepared as described previously(Bisova et al. 2003).
Affinity chromatography on immobilized suc1 protein
A specific binding partner of CDK, the suc1 protein (cyclin-dependent kinase subunit, CKS) (Brizuela et al. 1987) was used forisolation of the mitotic CDK complexes (Labbe et al. 1989, Bisovaet al. 2000). Affinity chromatography on the immobilized suc1protein was performed according to the method described pre-viously (Bisova et al. 2000).
Histone H1 kinase activity assay
Using histone H1 as a substrate, kinase activity bound to suc1protein was assessed employing the procedure described byMoreno et al. (1989) as modified in our laboratory (Zachlederet al. 1997, Bisova et al. 2000). Algal proteins bound to suc1–Sepharose were incubated with 10 ml of the assay buffer [20mMHEPES pH 7.5, 15mM MgCl2, 5mM EGTA, 1mM dithiothreitol(DTT), 0.1% (w/v) histone H1, 0.1mM ATP, 3.7� 106 Bq ml–1 of[�-32P]ATP (specific activity 1.5� 1014Bq mmol–1)] for 30min at308C. After separation by 12% SDS–PAGE, phosphorylatedhistone H1 bands were visualized by autoradiography. Autoradio-grams were scanned and evaluated using SigmaGel software(Jandel Scientific, San Rafael, CA, USA).
Electrophoretic separation of proteins
For the separation of proteins, SDS–PAGE was performedusing the Laemmli system (Laemmli 1970) in the Mini Protean 3Apparatus (BioRad Laboratories, Hercules, CA, USA).
Production of recombinant proteins in bacteria
cDNA coding for the C. reinhardtii A-, B- and D-type cyclins(CrCYCA1, CrCYCB1, CrCYCD2 and CrCYCD3) (Bisova et al.2005) were amplified from oligo(dT)-primed cDNA by PCR withprimers (CYCA gat for 50-CACCATGAGCTCTCGCGTCGGCTCttc-30, CYCA gat rev 50-TCACGAGCGGTGAATGCCGC-30,CYCB gat for 50-CACCATGGCTCTTCGTGCGGTTCAGCCC-30, CYCB gat rev 50-TTAAGCCACTGGCTGCGCGGGC-30,CYCD2 gat for 50-CACCATGGAGTGGCTGCTGCTGGAGACG-30, CYCD2 gat rev 50-TCACCGCTGAGCGGACGGC-30,CYCD3 gat for 50-CACCATGACGCTGGCGGTGAAGGTCG-30, CYCD3 gat rev 50-CTACTGCTGCACCGGCTGCGAC-30) and cloned into pENTR-SD/D-TOPO vector (Invitrogen,Carlsbad, CA, USA). The sequences were verified and the cyclinfragment recombined into pDEST15 (Invitrogen, Carlsbad, CA,USA), yielding pDEST15-CrCYCA1, B1, D2 or D3, respectively.The constructs were transformed into BL21-CodonPlus(DE3)-RILcells (Stratagene, La Jolla, CA, USA). Cells were grown at 378C
1814 Chloroplast fission in Scenedesmus

until an OD600 of 0.4, then transferred to 208C for 30min andinduced with 0.5mM isopropyl-b-D-thiogalactoside for 4 h. Thebacterial pellets were resuspended in 1� SDS–PAGE sample buffer(62.5mM Tris–HCl, pH 6.8, 20% glycerol, 2% SDS, 5%b-mercaptoethanol) and separated by 12% SDS–PAGE.
Sequence analyses
Sequence comparison was done using Vector NTI software(Invitrogen, Carlsbad, CA, USA). The GenBank numbers of thesequences are: MaCYCB1, P37882; CrCYCA1, XP_001693167;CrCYCB1, XP_001701288; CrCYCD2, XP_001695520; andCrCYCD3, XP_001695469.
Immunoblotting
Separated proteins were transferred by semi-dry blotting(Semi-dry blotter, Sigma-Aldrich, Prague, Czech Rep.) from thegel onto a nitrocellulose membrane (pore size 0.2mm, Protran,Schleicher and Schuell, Kent, UK) (Towbin et al. 1979) at 1mAcm–2 for 1.5 h. The dried nitrocellulose membrane was blocked ingoat serum, 5% (v/v) solution in TBS-T buffer (20mM Tris pH7.5, 0.5M NaCl, 0.05% Tween-20), overnight at 48C. Then itwas incubated with the primary antibody diluted with theserum solution, rinsed three times with TBS-T, incubated withthe secondary antibody, and washed six times for 10minwith TBS-T. Immunoreactive bands were detected by enhancedchemiluminescence (ECL; Amersham, Munich, Germany). Thefollowing primary antibodies were used: anti-FtsZ1 rabbitantiserum (diluted 1 : 1,000) raised against the FtsZ1 proteinfrom the unicellular green alga Nannochloris bacillaris (Koideet al. 2004), and the anti-cyclin IgG fraction of rabbit antiserum(diluted 1 : 1,000) raised against cyclin B1 (Sigma C8831); forantibody specificities see Results. The secondary antibodywas peroxidase-conjugated goat anti-rabbit IgG (Sigma A9169)(diluted 1 : 100,000).
Microscopy
Observations in transmitted light and fluorescence micros-copy were carried out using an Olympus BX51 microscopeequipped with a CCD camera (F-ViewII). A U-MWIBA2 filterblock (Ex/Em: 460–490/510–550 nm) was used for SYBR Green Ifluorescence and a U-MWIG2 filter block (Ex/Em: 520–550/580 nm) for Cy3 and chlorophyll fluorescence.
Immunofluorescence
Cells were fixed with 4% formaldehyde freshly preparedfrom paraformaldehyde in PEM buffer (50mM PIPES, 5mMEGTA, 1mM MgSO4�7H2O, pH 6.8), dehydrated by ethanol andembedded in LR-White resin. Semi-thin sections were attached tothe slide.
Slides were incubated for 10min in phosphate-bufferredsaline (PBS; 0.14M NaCl, 2.7mM KCl, 1.5mM KH2PO4, 6.5mMNa2HPO4�12 H2O, pH 7.4) and blocked in 1% bovine serumalbumin (BSA)/PBS in a wet chamber for 1 h at 378C. They werethen incubated with primary antibody (anti-FtsZ1 or anti-cyclin B)diluted 1 : 500 in 1% BSA/PBS for 1 h at 378C, washed three timesfor 5min with PBS, incubated with secondary antibody (Cy3-conjugated goat anti-rabbit IgG; Jackson Immunoresearch,Suffolk, UK) diluted 1 : 200 in 1% BSA/PBS for 1.5 h at 378C,and washed three times for 5min with PBS. Finally, the slides weremounted with one drop of Vectashield Hard Set MountingMedium (Vector Laboratories, Peterborough, UK) and observedusing an Olympus BX51 microscope.
Immunogold staining
Cells were fixed in 2.5% (v/v) glutaraldehyde in phosphatebuffer (160mM Na2HPO4, 40mM KH2PO4, pH 7.2), dehydratedand embedded in LRWhite’s resin. Ultrathin sections picked up onnickel grids were treated for 1.5 h with block solution [10% (v/v)goat serum, 2% (w/v) BSA, 0.2% (w/v) glycine, 0.2% (w/v) NH4Cland 0.05% (v/v) Tween-20 in PBS buffer] and then incubated for1.5 h with primary antibody against the FtsZ diluted 1 : 10 in PBS.After washing six times for 2min with PBS containing 0.05% (v/v)Tween-20, the sections were incubated for 1 h with colloidal gold10 nm particles conjugated with goat anti-rabbit antibody diluted1 : 50 in PBS and washed six times for 2min with PBS containing0.05% (v/v) Tween-20. After 10min fixation in 2.5% (v/v) glutar-aldehyde in PBS and washing three times for 2min with PBS andthree times for 2min with H2O, the sections were dried at roomtemperature overnight, stained with uranyl acetate for 30min andrinsed three times with 30% (v/v) ethanol. Dried sections werestained with lead citrate (0.03 g dissolved in 20ml of H2O and0.2ml of 10N NaOH) for 20min, rinsed with H2O and dried.Finally, the sections were stabilized by carbon coating and exam-ined using an electron microscope TEM JEOL 1010.
Funding
The Grant Agency of the Academy of Sciences of the
Czech Republic (grant Nos. B5020305, A500200614 and
A600200701); the Grant Agency of the Czech Republic
(grant No. 204/06/0102]; the Institutional Research Con-
cepts (Nos. AV0Z50200510 and AV0Z60050516) funded by
the Academy of Sciences of the Czech Republic.
Acknowledgments
We are obliged to Professor S. Kawano (Tokyo University,Kashiwa) for providing antibody raised against FtsZ proteinand to Dr. J. Nebesarova (South Bohemian University CeskeBudejovice) for her help with the electron microphotographs.
References
Bartek, J. and Lukas, J. (2007) DNA damage checkpoints: from initiationto recovery or adaptation. Curr. Opin. Cell Biol. 19: 238–245.
Bi, E. and Lutkenhaus, J. (1991) FtsZ ring structure associated with divisionin Escherichia coli. Nature 354: 161–164.
Bisova, K., Hendrychova, J., Cepak, V. and Zachleder, V. (2003) Cellgrowth and division processes are differentially sensitive to cadmium inScenedesmus quadricauda. Folia Microbiol. 48: 805–816.
Bisova, K., Krylov, D.M. and Umen, J.G. (2005) Genome-wide annotationand expression profiling of cell cycle regulatory genes in Chlamydomonasreinhardtii. Plant Physiol. 137: 1–17.
Bisova, K., Vıtova, M. and Zachleder, V. (2000) The activity of total histoneH1 kinases is related to growth and commitment points while thep13(suc1)-bound kinase activity relates to mitoses in the algaScenedesmus quadricauda. Plant Physiol. Biochem. 38: 755–764.
Brizuela, L., Draetta, G. and Beach, D. (1987) p13suc1 acts in the fissionyeast cell division cycle as a component of the p34cdc2 protein kinase.EMBO J. 6: 3507–3513.
Butterfass, T. (1995) Reproduction and continuity of chloroplasts inspermatophytes. Bot. Rev. 61: 1–27.
Calonge, T.M. and O’Connell, M.J. (2008) Turning off the G2 DNAdamage checkpoint. DNA Repair 7: 136–140.
Chloroplast fission in Scenedesmus 1815

Coleman, A.N. (1999) Temporal and spatial coordination of cells with their
plastid component. Int. Rev. Cytol. 193: 125.Decallonne, J.R. and Weyns, C.J. (1976) A shortened procedure of the
diphenylamine reaction for measurement of deoxyribonucleic acid by
using light activation. Anal. Biochem. 74: 448–456.El-Kafafi, E.S., Mukherjee, S., El-Shami, M., Putaux, J.L., Block, M.A.,
Pignot-Paintrand, I., et al. (2005) The plastid division proteins, FtsZ1 and
FtsZ2, differ in their biochemical properties and sub-plastidial localiza-
tion. Biochem. J. 387: 669–676.Glotzer, M., Murray, A.W. and Kirschner, M.W. (1991) Cyclin is degraded
by the ubiquitin pathway. Nature 349: 132–138.Heinhorst, S., Cannon, G. and Weissbach, A. (1985a) Plastid and nuclear
DNA synthesis are not coupled in suspension cells of Nicotiana tabacum.
Plant Mol. Biol. 4: 3.Heinhorst, S., Cannon, G. and Weissbach, A. (1985b) Chloroplast DNA
synthesis during the cell cycle in cultured cells of Nicotiana tabacum:
inhibition by nalidixic acid and hydroxyurea. Arch. Biochim. Biophys.
239: 475–479.Howard, A. and Pelc, S.R. (1953) Synthesis of deoxyribonucleic acid in
normal and irradiated ceils and its relation to chromosome breakage.
Heredity Suppl. 6: 261–273.John, P.C.L. (1984) Control of the cell division cycle in Chlamydomonas.
Microbiol. Sci. 1: 96–101.Kates, J.R., Chiang, K.S. and Jones, R.F. (1968) Studies on DNA
replication during synchronized vegetative growth and gametic differ-
entiation in Chlamydomonas reinhardtii. Exp. Cell Res. 49: 121–135.Koide, T., Yamazaki, T., Yamamoto, M., Fujishita, M., Nomura, H.,
Moriyama, Y., et al. (2004) Molecular divergence and characterization of
two chloroplast division genes, FTSZ1 and FTSZ2, in the unicellular
green alga Nannochloris bacillaris (Chlorophyta). J. Phycol. 40: 546–556.Kuroiwa, H., Mori, T., Takahara, M., Miyagishima, S. and Kuroiwa, T.
(2002) Chloroplast division machinery as revealed by immunofluores-
cence and electron microscopy. Planta 215: 185–190.Kuroiwa, T., Nagashima, H. and Fukuda, I. (1989) Chloroplast division
without DNA synthesis during the life cycle of the unicellular alga
Cyanidium caldarium M-8 as revealed by quantitative fluorescence
microscopy. Protoplasma 149: 120–129.Labbe, J.C., Capony, J.P., Caput, D., Cavadore, J.C., Derancourt, J.,
Kaghad, M., et al. (1989) MPF from starfish oocytes at first meiotic
metaphase is a heterodimer containing one molecule of cdc2 and one
molecule of cyclin B. EMBO J. 8: 3053–3058.Laemmli, U.K. (1970) Cleavage of structural proteins during the assembly
of the head of bacteriophage T4. Nature 227: 680–685.Lukavsky, J., Tetık, K. and Vendlova, J. (1973) Extraction of nucleic acid
from the alga Scenedesmus quadricauda. Arch. Hydrobiol. Algol. Stud. 9:
416–426.Lutkenhaus, J. (1998) Organelle division: from coli to chloroplasts. Curr.
Biol. 8: R619–R621.Maple, J., Aldridge, C. and Moller, S.G. (2005) Plastid division is mediated
by combinatorial assembly of plastid division proteins. Plant J. 43:
811–823.Margolin, W. (2000) Organelle division. Self-assembling GTPases caught in
the middle. Curr. Biol. 10: 328–330.McAndrew, R.S., Froelich, J.E., Vitha, S., Stokes, K.D. and
Osteryoung, K.W. (2001) Colocalization of plastid division proteins in
the chloroplast stromal compartment establishes a new functional
relationship between FtsZ1 and FtsZ2 in higher plants. Plant Biol. 127:
1656–1666.Mironov, V., De Veylder, L., Van Montagu, M. and Inze, D. (1999) Cyclin-
dependent kinases and cell division in plants—the nexus. Plant Cell 11:
509–529.Miyagishima, S. (2005) Origin and evolution of the chloroplast division
machinery. J. Plant Res. 118: 295–306.Miyagishima, S.Y., Takahara, M., Mori, T., Kuroiwa, H., Higashiyama, T.
and Kuroiwa, T. (2001) Plastid division is driven by a complex
mechanism that involves differential transition of the bacterial and
eukaryotic division rings. Plant Cell 13: 2257–2268.Moreno, S., Hayles, J. and Nurse, P. (1989) Regulation of p34cdc2 protein-
kinase during mitosis. Cell 58: 361–372.
Murray, A. (1995) Cyclin ubiquitination: the destructive end of mitosis. Cell
81: 149–152.Murray, A.W. (2004) Recycling the cell cycle: cyclins revisited. Cell 116:
221–234.Murray, A.W., Solomon, M.J. and Kirschner, M.W. (1989) The role of
cyclin synthesis and degradation in the control of maturation promoting
factor activity. Nature 339: 280–286.Nigg, E.A. (1995) Cyclin-dependent protein kinases: key regulators of the
eukaryotic cell cycle. BioEssays 17: 471–480.Norbury, C. and Nurse, P. (1992) Animal cell cycles and their control.
Annu. Rev. Biochem. 61: 441–470.Nurse, P. and Bissett, Y. (1981) Gene required in G1 for commitment to
cell cycle and in G2 control of mitosis in fission yeast. Nature 292:
558–560.Osteryoung, K., Stokes, K., Rutherford, S., Percival, A. and Lee, W. (1998)
Chloroplast division in higher plants requires members of two
functionally divergent gene families with homology to bacterial ftsZ.
Plant Cell 10: 1991–2004.Pelayo, H., Lastres, P. and de la Torre, C. (2001) Replication and G2
checkpoints: their response to caffeine. Planta 212: 444–453.Pines, J. (1996) Cell cycle reaching for a role for the Cks proteins. Curr. Biol.
6: 1399–1402.Poyton, R.O. and Branton, D. (1972) Control of daughter-cell number
variation in multiple fission—genetic versus environmental determinants
in Prototheca. Proc. Natl Acad. Sci. USA 69: 2346–2350.Setlık, I., Berkova, E., Doucha, J., Kubın, S., Vendlova, J. and
Zachleder, V. (1972) The coupling of synthetic and reproduction
processes in Scenedesmus quadricauda. Arch. Hydrobiol. Algolog. Stud.
7: 172–217.Setlık, I. and Zachleder, V. (1984) The multiple fission cell reproductive
patterns in algae. In The Microbial Cell Cycle. Edited by Nurse, P. and
Streiblova, E. pp. 253–279. CRC Press Inc., Boca Raton, FL.Slaninova, M., Nagyova, B., Galova, E., Hendrychova, J., Bisova, K.,
Zachleder, V., et al. (2003) The alga Chlamydomonas reinhardtii UVS11
gene is responsible for cell division delay and temporal decrease in histone
H1 kinase activity caused by UV irradiation. Mut. Res. DNA Repair 2:
737–750.Stern, B. and Nurse, P. (1996) A quantitative model for the cdc2 control of
S phase and mitosis in fission yeast. Trends Genet. 12: 345–350.Stokes, K.D., McAndrew, R.S., Figueroa, R., Vitha, S. and
Osteryoung, K.W. (2000) Chloroplast division and morphology are
differentially affected by overexpression of FtsZ1 and FtsZ2 genes in
Arabidopsis. Plant Physiol. 124: 1668–1677.Stokes, K.D. and Osteryoung, K.W. (2003) Early divergence of the FtsZ1
and FtsZ2 plastid division gene families in photosynthetic eukaryotes.
Gene 320: 97–108.Torres-Rosell, J., De Piccoli, G., Cordon-Preciado, V., Farmer, S.,
Jarmuz, A., Machin, F., et al. (2007) Anaphase onset before complete
DNA replication with intact checkpoint responses. Science 315:
1411–1415.Towbin, H., Staehelin, T. and Gordon, J. (1979) Electrophoretic
transfer of proteins from polyacrylamide gels to nitrocellulose
sheets: procedure and some applications. Proc. Natl Acad. Sci. USA
76: 4350–4354.van den Hoek, C., Mann, D.G. and Jahns, H.M. (1995) In Algae, an
Introduction to Phycology. Cambridge University Press, Cambridge.Vıtova, M., Hendrychova, J., Cepak, V. and Zachleder, V. (2005)
Visualization of DNA-containing structures in various species of
Chlorophyta, Rhodophyta and Cyanophyta using SYBR green I dye.
Folia Microbiol. 50: 333–340.Vıtova, M. and Zachleder, V. (2005) Points of commitment to reproductive
events as a tool for analysis of the cell cycle in synchronous cultures of
algae. Folia Microbiol. 50: 141–149.Wang, D., Kong, D., Wang, Y., Hu, Y., He, Y. and Sun, J. (2003) Isolation
of two plastid division ftsZ genes from Chlamydomonas reinhardtii and its
evolutionary implication for the role of FtsZ in plastid division. J. Exp.
Bot. 54: 1115–1116.Wanka, F. (1962) Die Bestimmung der Nucleinsauren in Chlorella
pyrenoidosa. Planta 58: 594–619.
1816 Chloroplast fission in Scenedesmus

Yen, A.B. and Pardee, A.B. (1979) Role of nuclear size in cell growthinitiation. Science 204: 1315–1317.
Zachleder, V. (1984) Optimization of nucleic acids assay in green and blue-green algae: extraction procedures and the light-activated reaction forDNA. Arch. Hydrobiol. Algolog. Stud. 36: 313–328.
Zachleder, V. (1994) The effect of hydroxyurea and fluorodeoxyuridine oncell cycle events in the chlorococcal alga Scenedesmus quadricauda(Chlorophyta). J. Phycol. 30: 274–279.
Zachleder, V. (1995) Regulation of growth processes during the cell cycle ofthe chlorococcal alga Scenedesmus quadricauda under a DNA replicationblock. J. Phycol. 30: 941–947.
Zachleder, V. and Cepak, V. (1987a) The effect of light on the number ofchloroplast nucleoids in daughter cells of the alga Scenedesmusquadricauda. Protoplasma 138: 37–44.
Zachleder, V. and Cepak, V. (1987b) Variations in chloroplast nucleoidnumber during the cell cycle in the alga Scenedesmus quadricauda grownunder different light conditions. Protoplasma 141: 74–82.
Zachleder, V., Kawano, S. and Kuroiwa, T. (1995) The course of chloroplastDNA replication and its relationship to other reproductive processes inthe chloroplast and nucleocytoplasmic compartment during the cell cycleof the alga Scenedesmus quadricauda. Protoplasma 188: 245–251.
Zachleder, V., Kawano, S. and Kuroiwa, T. (1996) Uncoupling ofchloroplast reproductive events from cell cycle division processes by5-fluorodeoxyuridine in the alga Scenedesmus quadricauda. Protoplasma192: 228–234.
Zachleder, V., Schlafli, O. and Boschetti, A. (1997) Growth-controlledoscillation in activity of histone H1 kinase during the cell cycle ofChlamydomonas reinhardtii (Chlorophyta). J. Phycol. 33: 673–681.
Zachleder, V. and Setlık, I. (1982) Effect of irradiance on the course ofRNA synthesis in the cell cycle of Scenedesmus quadricauda. Biol. Plant.24: 341–353.
Zachleder, V. and Setlık, I. (1990) Timing of events in overlapping cellreproductive sequences and their mutual interactions in the algaScenedesmus quadricauda. J. Cell Sci. 97: 631–638.
(Received September 5, 2008; Accepted October 26, 2008)
Chloroplast fission in Scenedesmus 1817




















