
An emilin family extracellular matrix protein identified in the cochlear basilar membrane
13
An emilin family extracellular matrix protein identified in the cochlear basilar membrane Lori L. Amma, a Richard Goodyear, b Jonathan S. Faris, a Iwan Jones, a Lily Ng, a Guy Richardson, b and Douglas Forrest a, * a Department of Human Genetics, Box 1498, Mount Sinai School of Medicine, 1425 Madison Avenue, New York, NY 10029, USA b School of Biological Sciences, University of Sussex, Sussex, UK Received 7 October 2002; revised 5 February 2003; accepted 1 March 2003 Abstract The precise movement of the cochlear basilar membrane (BM) stimulates the sensory hair cells during auditory transduction. However, the molecular composition of the BM that confers its specialized properties of support and elasticity is poorly understood. A differential screen of cochlear RNA from deaf mice lacking thyroid hormone receptor was used to identify a sequence encoding a secreted protein, which is abundant in the BM and is expressed at low levels in the heart, lung, and brain. The protein possesses several domains for protein interactions and is related to emilin (elastin microfibril interface-located protein) previously isolated from aorta. This cochlear emilin-2 mRNA is expressed in the tympanic border cells underlying the BM and an antibody detected protein in the extracellular matrix surrounding the collagenous fibers in the BM. These results identify emilin-2 as a major BM component and suggest that it contributes to the developmental assembly or function of the BM. © 2003 Elsevier Science (USA). All rights reserved. Introduction The transduction of sound into neural signals by the cochlear hair cells requires the precise movement of the basilar membrane (BM), 1 a dynamic structure, on which the hair cells reside. The vibration of the BM stimulates the mechanosensitive hair cells by deflecting their apical ste- reociliary bundles against the tectorial membrane (Dallos et al., 1996; Gillespie and Walker, 2001). The BM also re- sponds to electromotile changes in the outer hair cells that amplify BM motion (Holley, 1996). Although the support and elasticity provided by the BM are critical for hearing, relatively little is known of the molecular basis for these properties. The BM consists of a specialized layer of extracellular matrix (ECM) between the overlying sensory epithelium, containing the hair cells and supporting cells, and an un- derlying layer of mesothelial cells, the tympanic border cells, adjacent to the chamber of the scala tympani (Slepecky, 1996). A basement membrane borders the BM next to the supporting cells of the epithelium (Tsuprun and Santi, 1999; Rodgers et al., 2001). Analysis by electron microscopy has shown the mammalian BM to be composed structurally of major filaments, 10 –12 nm in diameter, em- bedded in a matrix of ground substance (Iurato, 1967; Ca- bezudo, 1978; Katori et al., 1993). Critical constituent molecules of several cochlear struc- tures, including the tectorial membrane and hair cells, have been identified using approaches such as biochemical puri- fication (Legan et al., 1997), cDNA screening (Cohen- Salmon et al., 1997; Heller et al., 1998), and the study of human or murine deafness-associated genes (Resendes et al., 2001; Steel and Kros, 2001; Griffith and Friedman, 2002). However, relatively few molecular components of the BM have so far been identified. Biochemical assays and immunostaining against known ECM proteins of other tis- sues have indicated that the major BM filaments contain collagen type II (Thalmann, 1993; Tsuprun and Santi, 1999; * Corresponding author. Fax: 1-212-849-2508. E-mail address: [email protected] (D. Forrest). 1 Abbreviations used: BM, basilar membrane; ECM, extracellular ma- trix; TR, thyroid hormone receptor. Molecular and Cellular Neuroscience 23 (2003) 460 – 472 www.elsevier.com/locate/ymcne 1044-7431/03/$ – see front matter © 2003 Elsevier Science (USA). All rights reserved. doi:10.1016/S1044-7431(03)00075-7
-
Upload
independent -
Category
Documents
-
view
1 -
download
0
Transcript of An emilin family extracellular matrix protein identified in the cochlear basilar membrane

An emilin family extracellular matrix protein identified in the cochlearbasilar membrane
Lori L. Amma,a Richard Goodyear,b Jonathan S. Faris,a Iwan Jones,a Lily Ng,aGuy Richardson,b and Douglas Forresta,*
a Department of Human Genetics, Box 1498, Mount Sinai School of Medicine, 1425 Madison Avenue, New York, NY 10029, USAb School of Biological Sciences, University of Sussex, Sussex, UK
Received 7 October 2002; revised 5 February 2003; accepted 1 March 2003
Abstract
The precise movement of the cochlear basilar membrane (BM) stimulates the sensory hair cells during auditory transduction. However,the molecular composition of the BM that confers its specialized properties of support and elasticity is poorly understood. A differentialscreen of cochlear RNA from deaf mice lacking thyroid hormone receptor ! was used to identify a sequence encoding a secreted protein,which is abundant in the BM and is expressed at low levels in the heart, lung, and brain. The protein possesses several domains for proteininteractions and is related to emilin (elastin microfibril interface-located protein) previously isolated from aorta. This cochlear emilin-2mRNA is expressed in the tympanic border cells underlying the BM and an antibody detected protein in the extracellular matrix surroundingthe collagenous fibers in the BM. These results identify emilin-2 as a major BM component and suggest that it contributes to thedevelopmental assembly or function of the BM.© 2003 Elsevier Science (USA). All rights reserved.
Introduction
The transduction of sound into neural signals by thecochlear hair cells requires the precise movement of thebasilar membrane (BM),1 a dynamic structure, on which thehair cells reside. The vibration of the BM stimulates themechanosensitive hair cells by deflecting their apical ste-reociliary bundles against the tectorial membrane (Dallos etal., 1996; Gillespie and Walker, 2001). The BM also re-sponds to electromotile changes in the outer hair cells thatamplify BM motion (Holley, 1996). Although the supportand elasticity provided by the BM are critical for hearing,relatively little is known of the molecular basis for theseproperties.The BM consists of a specialized layer of extracellular
matrix (ECM) between the overlying sensory epithelium,containing the hair cells and supporting cells, and an un-
derlying layer of mesothelial cells, the tympanic bordercells, adjacent to the chamber of the scala tympani(Slepecky, 1996). A basement membrane borders the BMnext to the supporting cells of the epithelium (Tsuprun andSanti, 1999; Rodgers et al., 2001). Analysis by electronmicroscopy has shown the mammalian BM to be composedstructurally of major filaments, 10–12 nm in diameter, em-bedded in a matrix of ground substance (Iurato, 1967; Ca-bezudo, 1978; Katori et al., 1993).Critical constituent molecules of several cochlear struc-
tures, including the tectorial membrane and hair cells, havebeen identified using approaches such as biochemical puri-fication (Legan et al., 1997), cDNA screening (Cohen-Salmon et al., 1997; Heller et al., 1998), and the study ofhuman or murine deafness-associated genes (Resendes etal., 2001; Steel and Kros, 2001; Griffith and Friedman,2002). However, relatively few molecular components ofthe BM have so far been identified. Biochemical assays andimmunostaining against known ECM proteins of other tis-sues have indicated that the major BM filaments containcollagen type II (Thalmann, 1993; Tsuprun and Santi, 1999;
* Corresponding author. Fax: !1-212-849-2508.E-mail address: [email protected] (D. Forrest).1 Abbreviations used: BM, basilar membrane; ECM, extracellular ma-
trix; TR, thyroid hormone receptor.
Molecular and Cellular Neuroscience 23 (2003) 460–472 www.elsevier.com/locate/ymcne
1044-7431/03/$ – see front matter © 2003 Elsevier Science (USA). All rights reserved.doi:10.1016/S1044-7431(03)00075-7

Dreiling et al., 2002) and that these filaments are associatedwith keratan- and chondroitin-sulfate proteoglycans (Mu-nyer and Schulte, 1994). The glycoproteins fibronectin andtenascin, which are expressed in the ECM in many tissues,have been detected in the BM ground substance (Santi et al.,1989; Woolf et al., 1992; Swartz and Santi, 1999).Another approach to identifying genes for cochlear func-
tion is to exploit deaf mouse models for screening foraberrantly expressed sequences. Thrbtm1/tm1 mice lack thy-roid hormone receptor ! (TR!), a ligand-regulated tran-scription factor, and display defective auditory function(Forrest et al., 1996). TR! is expressed in the cochlearsensory epithelium (Bradley et al., 1994) and Thrbtm1/tm1mice have defects in postnatal cochlear development withimpaired hair cell function and retarded differentiation ofthe sensory epithelium (Rusch et al., 2001). Hearing losssometimes also arises in human hypothyroidism and inresistance to thyroid hormone (Brucker-Davis et al., 1996).To isolate sequences involved in cochlear maturation, weused a differential screen of cochlear RNA from Thrbtm1/tm1mice. This study identified a major component of the BM,emilin-2, that belongs to the multimerin/emilin family ofglycoproteins previously found in heart elastic fibers andother tissues (Hayward et al., 1995; Doliana et al., 1999).
Results
A cochlear cDNA encoding an emilin protein
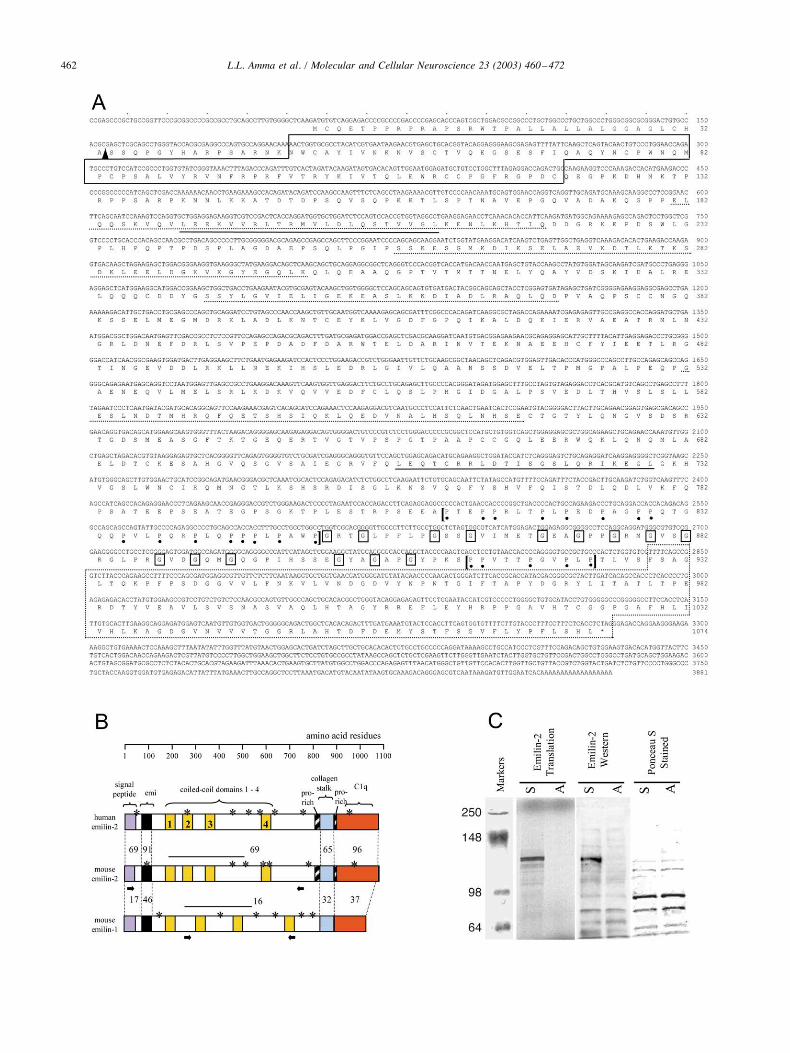
A cDNA encoding an ECM-like protein was isolatedfrom a mouse cochlear library, using a probe obtained froma differential screen of cochlear RNA of Thrbtm1/tm1 andwild-type (wt) mice at postnatal day 8 (P8). A 3.9-kb cDNAclone contained a complete 3222-bp open reading frame, apolyadenylation signal, and a poly(A) tail (Fig. 1A). Theencoded product was related to a family of proteins char-acterized by an N-terminal, cysteine-rich “Emi” domain.Emi domains have been found in emilin-1, an ECM com-ponent of elastic fibers in aorta (Doliana et al., 1999),multimerin, produced by platelets and endothelial cells(Hayward et al., 1995), EndoGlyx-1 found in endothelialtissues (Christian et al., 2001), and Emu proteins expressedin kidney and other tissues (Leimeister et al., 2002). Theemilins form a subgroup of proteins that are characterizedby several domains for protein interactions, including col-lagen and complement C1q domains (see below). The co-chlear gene product was most closely related to a humanprotein, emilin-2, that was recently isolated from kidney andaorta cDNA sources (Doliana et al., 2001). Alignments ofpredicted amino acid sequences indicated that, overall, themouse cochlear protein was 73% related to human emilin-2whereas it was only 21% related to mouse emilin-1 (Fig.1B). At the nucleotide level, the mouse cochlear cDNA was76% related to human emilin-2 but only 46% related tomouse emilin-1. Human emilin-1 and emilin-2 are "23%
related at the amino acid level (Doliana et al., 2001), acomparable degree of similarity as observed between themouse proteins. These comparisons indicate that the co-chlear cDNA encodes the mouse orthologue of human emi-lin-2.In vitro transcription and translation of the 3.9-kb cDNA
clone in reticulocyte lysates yielded a protein band of ap-proximately 120 kDa (Fig. 1C), consistent with the deducedemilin-2 sequence of 1074 amino acids with a theoreticalmolecular weight of 117 kDa. The first translation startcodon identified in the mouse emilin-2 cDNA was colinearwith that in human emilin-2 (Doliana et al., 2001) and wasconsistent with a consensus initiation site (Kozak, 1987).The 5# upstream sequences extending to the putative pro-moter region of the gene contained neither in-frame stopcodons nor other candidate start codons (see Fig. 2C), thussupporting the start codon assignment.Mouse emilin-2 contained a number of protein oligomer-
ization and interaction domains (Fig. 1B) including thecysteine-rich Emi domain common to emilin-1 and multi-merin ($43% identity across all three proteins) (Hayward etal., 1995; Doliana et al., 1999), which has been suggested tomediate protein oligomerization. Like human emilin-2, themouse sequence included an N-terminal signal peptide ofhydrophobic residues, suggesting that the protein is ex-ported. The central domain of emilin-2 contained four pre-dicted "-helical coiled-coils and the C-terminus contained acomplement C1q globular domain (residues 929–1074).C1q domains found in complement and other proteins suchas type VIII and X collagens are thought to mediate proteininteractions. The C1q domain is preceded by 15 repeats ofthe Gxy triplet typical of collagen (G % glycine, x and y %any two residues). Unlike in collagen where the triplets runcontinuously, the spacing between the triplets was irregularin emilin-2. This irregularity of the Gxy repeats also occursin human emilin-2 whereas these repeats are regular inemilin-1 (Doliana et al., 2001), suggesting that this is acharacteristic feature of emilin-2. Emilin-2 also possessedshort proline-rich segments on each side of the collagendomain. Several leucine zippers that may stabilize proteininteractions and several potential N-linked glycosylationsites were also present. The regions of least similarity be-tween mouse and human emilin-2 occurred in the interven-ing regions between the individual coiled-coil domains.
Mouse emilin-2 gene structure
The mouse emilin-2 gene contains 8 exons spanning 58kb (Fig. 2A) and is located at mouse chromosome 17E1.1(base position 57.7 million, Celera database). The humanemilin-2 gene has a related size and exon structure (Dolianaet al., 2001) and is on chromosome 18p11.3 at base position4.6 million (between markers D18S481 and D18S476). In-tron/exon boundaries were localized by comparison of themouse cDNA and genomic sequences and were in accordwith consensus splice sites (Mount, 1982) (Fig. 2B). The
461L.L. Amma et al. / Molecular and Cellular Neuroscience 23 (2003) 460–472
462 L.L. Amma et al. / Molecular and Cellular Neuroscience 23 (2003) 460–472

sizes of the introns (except the largest introns II, III, and IV)were confirmed by PCR analysis using exon-specific prim-ers on a BAC genomic clone containing the mouse emilin-2gene.
A promoter with a predicted transcription start site 72 bpupstream of exon 1 was predicted (score 1.0 of a maximumof 1.0) using the eukaryotic neural network promoter pre-diction program (Fig. 2C). This predicted RNA start sitewould add 72 bases to the available cDNA for a total lengthof 3954 bp consistent with the "4.0-kb emilin-2 mRNAsize identified by Northern blot analysis (see Fig. 3, below).Additional 5# sequences from cochlear and heart cDNAlibraries were isolated by RACE-PCR but no clone ex-tended beyond the 5# end of the original cDNA. Theseresults together with the proximity of the predicted pro-moter and the colinearity of the N-termini of mouse andhuman emilin-2 (Doliana et al., 2001) suggested that theentire reading frame was present in the original cDNAclone.
Tissue distribution and cochlear expression of emilin-2mRNA
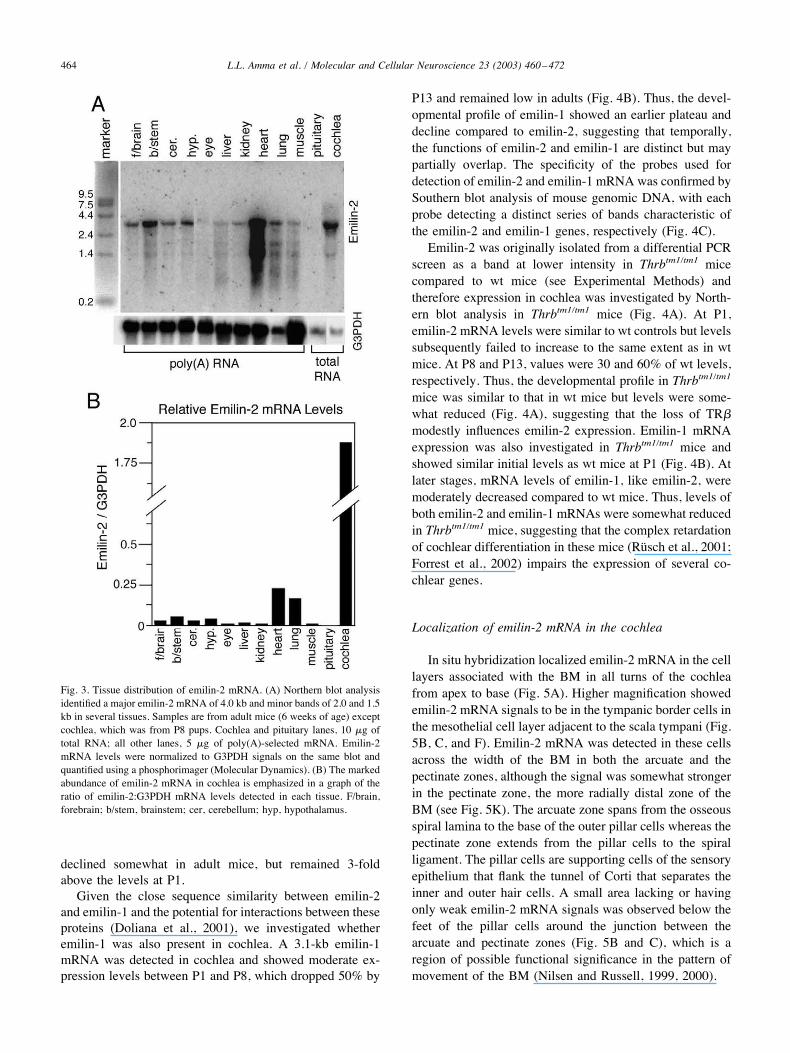
The distribution of emilin-2 mRNA was determined inwt mouse tissues by Northern blot analysis (Fig. 3A). Thehighest levels of the major 4.0-kb mRNA were detected inthe cochlea of P8 pups, followed by modest (10-fold lower)levels in adult heart and lung and much lower levels (at least47-fold lower) in forebrain, brainstem, cerebellum, and hy-pothalamus. Very low levels were detected in muscle, liver,kidney, and eye. Minor bands of 2.0 and 1.5 kb were alsopresent at low levels in most tissues. Emilin-2 was readilydetected in total RNA preparations from cochlea whereas itwas detected in other tissues using poly-A selected RNA,thus emphasizing the relative abundance of emilin-2 incochlea (Fig. 3B). Possibly consistent with these findings,human emilin-2 RNA has been reported to be detected infetal heart, lung, and placenta and at lower levels in brainand other tissues by dot blot hybridization (Doliana et al.,2001).In mice, the cochlea matures during the first 2 postnatal
weeks before the onset of auditory function at around P13(Pujol et al., 1997) and therefore the developmental expres-sion of emilin-2 was determined by Northern blot analysis.Low levels of emilin-2 mRNA were detected in cochlea inneonatal pups at P1 (Fig. 4A , wt panel). Levels increased2-fold by P5 and rose further to 16-fold at P13. Expression
Fig. 1. Mouse cochlear cDNA encoding emilin-2. (A) The nucleotide and predicted amino acid sequence of mouse emilin-2, indicating the Emi domain (solidbox), coiled-coils (dotted underline), putative leucine zippers (solid underline), C1q domain (dotted box), glycine repeats of the collagenous stalk (boxed Gs),and the flanking proline-rich regions (dots under Ps, in regions enclosed by brackets). A putative cleavage site for the signal peptide region is shown as anarrowhead. (B) Schematic comparison showing that mouse emilin-2 is more closely related to human emilin-2 than to mouse emilin-1. The percentage aminoacid similarity between domains is indicated. Putative N-linked glycosylation sites are marked (asterisks). The mouse emilin-2 domain used to raise anantiserum is indicated as a bar above the coiled-coil region and was chosen to have minimal homology with emilin-1 (&16% in this region). An emilin-1domain used for expression in vitro is also shown as a bar. Short arrows below mouse emilin-2 and emilin-1 define cDNA regions used as probes for Northernand Southern blot analyses (see Fig. 4C). (C) In vitro translation of mouse emilin-2. (Left) cDNA transcribed in sense (S) or antisense (A) directions wastranslated in reticulocyte lysates in the presence of 35S-methionine. Sense transcription yielded a protein band of the expected size of full-length emilin-2;markers in kilodaltons. (Middle), The antiserum raised against the emilin-2 antigen (see B) was used for Western blot analysis of reticulocyte lysatesprogrammed with sense (S) or antisense (A) emilin-2 transcripts (in the absence of 35S-methionine). The antiserum detected a band of the expected size ofemilin-2 in the sense strand-programmed lysates. (Right) Western blot filter stained with ponceau S, showing equivalent loading of lysates in both sense andantisense-programmed lanes.
Fig. 2. Mouse emilin-2 gene structure. (A) The 57.9-kb mouse emilin-2gene contains eight exons (solid boxes, 33–1914 bp in size) and sevenintrons (Roman numerals I–VII) (mapped from cDNA sequence in Fig.1A). (B) Intron/exon boundaries. Consensus splice sites of vertebrate genes(where n # 11 and N % any nucleotide) are depicted in bold type.Translated codons in capitals with single-letter amino acid translationabove; intron sequences in lower case. (C) 5# UTR showing predictedpromoter region (underlined), TATA box (boxed), and transcription startsite (G, enlarged, at position '72) (eukaryotic neural network promoterprediction program). This predicted RNA start adds 72 bases of 5# UTR tothe longest cDNA clone obtained, indicated by the dashed box; ATG startcodon is in bold (position !57).
463L.L. Amma et al. / Molecular and Cellular Neuroscience 23 (2003) 460–472
declined somewhat in adult mice, but remained 3-foldabove the levels at P1.Given the close sequence similarity between emilin-2
and emilin-1 and the potential for interactions between theseproteins (Doliana et al., 2001), we investigated whetheremilin-1 was also present in cochlea. A 3.1-kb emilin-1mRNA was detected in cochlea and showed moderate ex-pression levels between P1 and P8, which dropped 50% by
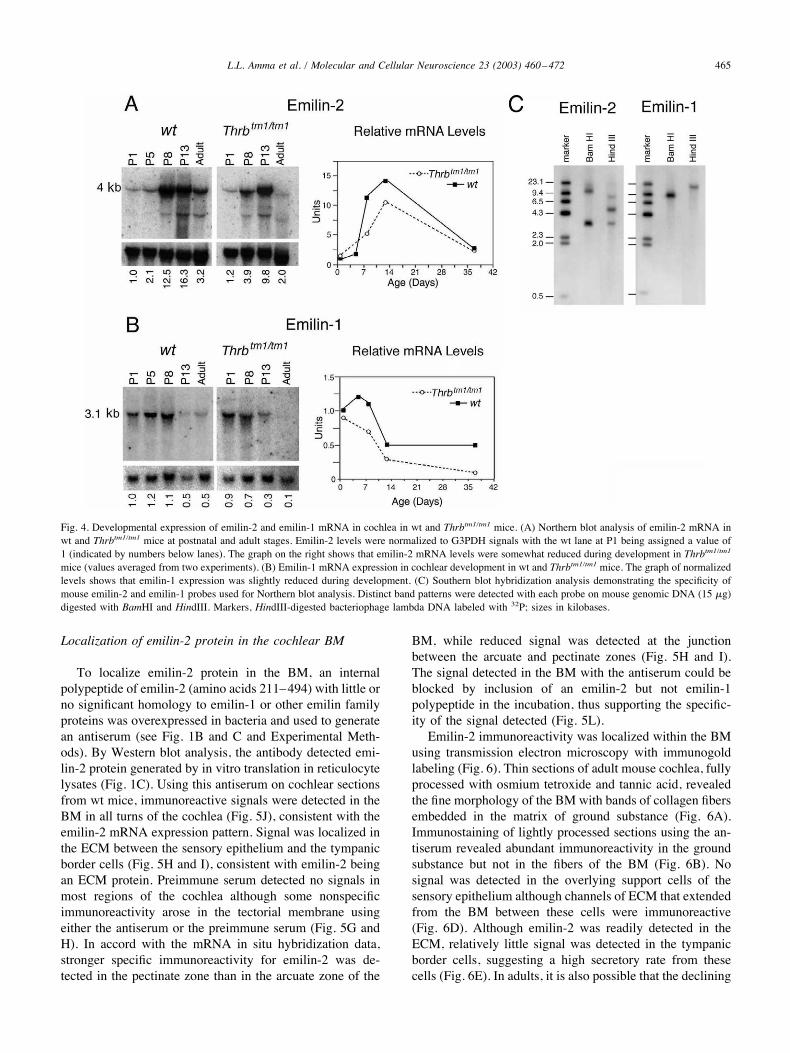
P13 and remained low in adults (Fig. 4B). Thus, the devel-opmental profile of emilin-1 showed an earlier plateau anddecline compared to emilin-2, suggesting that temporally,the functions of emilin-2 and emilin-1 are distinct but maypartially overlap. The specificity of the probes used fordetection of emilin-2 and emilin-1 mRNA was confirmed bySouthern blot analysis of mouse genomic DNA, with eachprobe detecting a distinct series of bands characteristic ofthe emilin-2 and emilin-1 genes, respectively (Fig. 4C).Emilin-2 was originally isolated from a differential PCR
screen as a band at lower intensity in Thrbtm1/tm1 micecompared to wt mice (see Experimental Methods) andtherefore expression in cochlea was investigated by North-ern blot analysis in Thrbtm1/tm1 mice (Fig. 4A). At P1,emilin-2 mRNA levels were similar to wt controls but levelssubsequently failed to increase to the same extent as in wtmice. At P8 and P13, values were 30 and 60% of wt levels,respectively. Thus, the developmental profile in Thrbtm1/tm1mice was similar to that in wt mice but levels were some-what reduced (Fig. 4A), suggesting that the loss of TR!modestly influences emilin-2 expression. Emilin-1 mRNAexpression was also investigated in Thrbtm1/tm1 mice andshowed similar initial levels as wt mice at P1 (Fig. 4B). Atlater stages, mRNA levels of emilin-1, like emilin-2, weremoderately decreased compared to wt mice. Thus, levels ofboth emilin-2 and emilin-1 mRNAs were somewhat reducedin Thrbtm1/tm1 mice, suggesting that the complex retardationof cochlear differentiation in these mice (Rusch et al., 2001;Forrest et al., 2002) impairs the expression of several co-chlear genes.
Localization of emilin-2 mRNA in the cochlea
In situ hybridization localized emilin-2 mRNA in the celllayers associated with the BM in all turns of the cochleafrom apex to base (Fig. 5A). Higher magnification showedemilin-2 mRNA signals to be in the tympanic border cells inthe mesothelial cell layer adjacent to the scala tympani (Fig.5B, C, and F). Emilin-2 mRNA was detected in these cellsacross the width of the BM in both the arcuate and thepectinate zones, although the signal was somewhat strongerin the pectinate zone, the more radially distal zone of theBM (see Fig. 5K). The arcuate zone spans from the osseousspiral lamina to the base of the outer pillar cells whereas thepectinate zone extends from the pillar cells to the spiralligament. The pillar cells are supporting cells of the sensoryepithelium that flank the tunnel of Corti that separates theinner and outer hair cells. A small area lacking or havingonly weak emilin-2 mRNA signals was observed below thefeet of the pillar cells around the junction between thearcuate and pectinate zones (Fig. 5B and C), which is aregion of possible functional significance in the pattern ofmovement of the BM (Nilsen and Russell, 1999, 2000).
Fig. 3. Tissue distribution of emilin-2 mRNA. (A) Northern blot analysisidentified a major emilin-2 mRNA of 4.0 kb and minor bands of 2.0 and 1.5kb in several tissues. Samples are from adult mice (6 weeks of age) exceptcochlea, which was from P8 pups. Cochlea and pituitary lanes, 10 $g oftotal RNA; all other lanes, 5 $g of poly(A)-selected mRNA. Emilin-2mRNA levels were normalized to G3PDH signals on the same blot andquantified using a phosphorimager (Molecular Dynamics). (B) The markedabundance of emilin-2 mRNA in cochlea is emphasized in a graph of theratio of emilin-2:G3PDH mRNA levels detected in each tissue. F/brain,forebrain; b/stem, brainstem; cer, cerebellum; hyp, hypothalamus.
464 L.L. Amma et al. / Molecular and Cellular Neuroscience 23 (2003) 460–472
Localization of emilin-2 protein in the cochlear BM
To localize emilin-2 protein in the BM, an internalpolypeptide of emilin-2 (amino acids 211–494) with little orno significant homology to emilin-1 or other emilin familyproteins was overexpressed in bacteria and used to generatean antiserum (see Fig. 1B and C and Experimental Meth-ods). By Western blot analysis, the antibody detected emi-lin-2 protein generated by in vitro translation in reticulocytelysates (Fig. 1C). Using this antiserum on cochlear sectionsfrom wt mice, immunoreactive signals were detected in theBM in all turns of the cochlea (Fig. 5J), consistent with theemilin-2 mRNA expression pattern. Signal was localized inthe ECM between the sensory epithelium and the tympanicborder cells (Fig. 5H and I), consistent with emilin-2 beingan ECM protein. Preimmune serum detected no signals inmost regions of the cochlea although some nonspecificimmunoreactivity arose in the tectorial membrane usingeither the antiserum or the preimmune serum (Fig. 5G andH). In accord with the mRNA in situ hybridization data,stronger specific immunoreactivity for emilin-2 was de-tected in the pectinate zone than in the arcuate zone of the
BM, while reduced signal was detected at the junctionbetween the arcuate and pectinate zones (Fig. 5H and I).The signal detected in the BM with the antiserum could beblocked by inclusion of an emilin-2 but not emilin-1polypeptide in the incubation, thus supporting the specific-ity of the signal detected (Fig. 5L).Emilin-2 immunoreactivity was localized within the BM
using transmission electron microscopy with immunogoldlabeling (Fig. 6). Thin sections of adult mouse cochlea, fullyprocessed with osmium tetroxide and tannic acid, revealedthe fine morphology of the BM with bands of collagen fibersembedded in the matrix of ground substance (Fig. 6A).Immunostaining of lightly processed sections using the an-tiserum revealed abundant immunoreactivity in the groundsubstance but not in the fibers of the BM (Fig. 6B). Nosignal was detected in the overlying support cells of thesensory epithelium although channels of ECM that extendedfrom the BM between these cells were immunoreactive(Fig. 6D). Although emilin-2 was readily detected in theECM, relatively little signal was detected in the tympanicborder cells, suggesting a high secretory rate from thesecells (Fig. 6E). In adults, it is also possible that the declining
Fig. 4. Developmental expression of emilin-2 and emilin-1 mRNA in cochlea in wt and Thrbtm1/tm1 mice. (A) Northern blot analysis of emilin-2 mRNA inwt and Thrbtm1/tm1 mice at postnatal and adult stages. Emilin-2 levels were normalized to G3PDH signals with the wt lane at P1 being assigned a value of1 (indicated by numbers below lanes). The graph on the right shows that emilin-2 mRNA levels were somewhat reduced during development in Thrbtm1/tm1mice (values averaged from two experiments). (B) Emilin-1 mRNA expression in cochlear development in wt and Thrbtm1/tm1 mice. The graph of normalizedlevels shows that emilin-1 expression was slightly reduced during development. (C) Southern blot hybridization analysis demonstrating the specificity ofmouse emilin-2 and emilin-1 probes used for Northern blot analysis. Distinct band patterns were detected with each probe on mouse genomic DNA (15 $g)digested with BamHI and HindIII. Markers, HindIII-digested bacteriophage lambda DNA labeled with 32P; sizes in kilobases.
465L.L. Amma et al. / Molecular and Cellular Neuroscience 23 (2003) 460–472
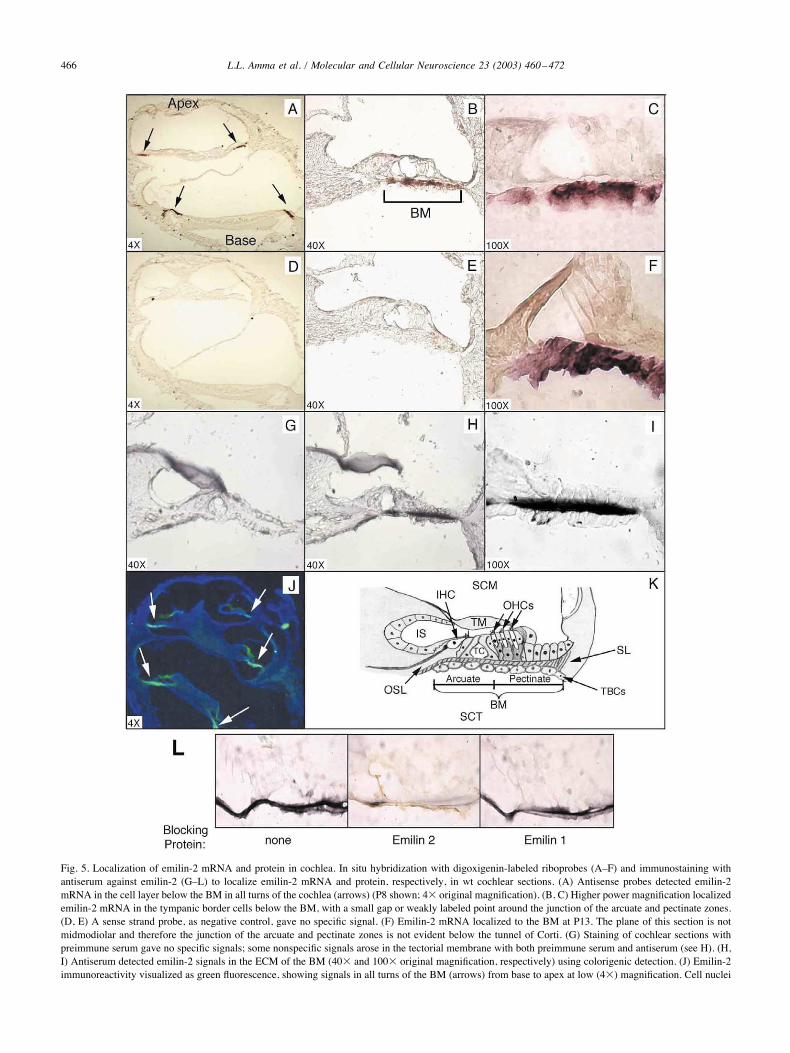
Fig. 5. Localization of emilin-2 mRNA and protein in cochlea. In situ hybridization with digoxigenin-labeled riboprobes (A–F) and immunostaining withantiserum against emilin-2 (G–L) to localize emilin-2 mRNA and protein, respectively, in wt cochlear sections. (A) Antisense probes detected emilin-2mRNA in the cell layer below the BM in all turns of the cochlea (arrows) (P8 shown; 4( original magnification). (B, C) Higher power magnification localizedemilin-2 mRNA in the tympanic border cells below the BM, with a small gap or weakly labeled point around the junction of the arcuate and pectinate zones.(D, E) A sense strand probe, as negative control, gave no specific signal. (F) Emilin-2 mRNA localized to the BM at P13. The plane of this section is notmidmodiolar and therefore the junction of the arcuate and pectinate zones is not evident below the tunnel of Corti. (G) Staining of cochlear sections withpreimmune serum gave no specific signals; some nonspecific signals arose in the tectorial membrane with both preimmune serum and antiserum (see H). (H,I) Antiserum detected emilin-2 signals in the ECM of the BM (40( and 100( original magnification, respectively) using colorigenic detection. (J) Emilin-2immunoreactivity visualized as green fluorescence, showing signals in all turns of the BM (arrows) from base to apex at low (4() magnification. Cell nuclei
466 L.L. Amma et al. / Molecular and Cellular Neuroscience 23 (2003) 460–472
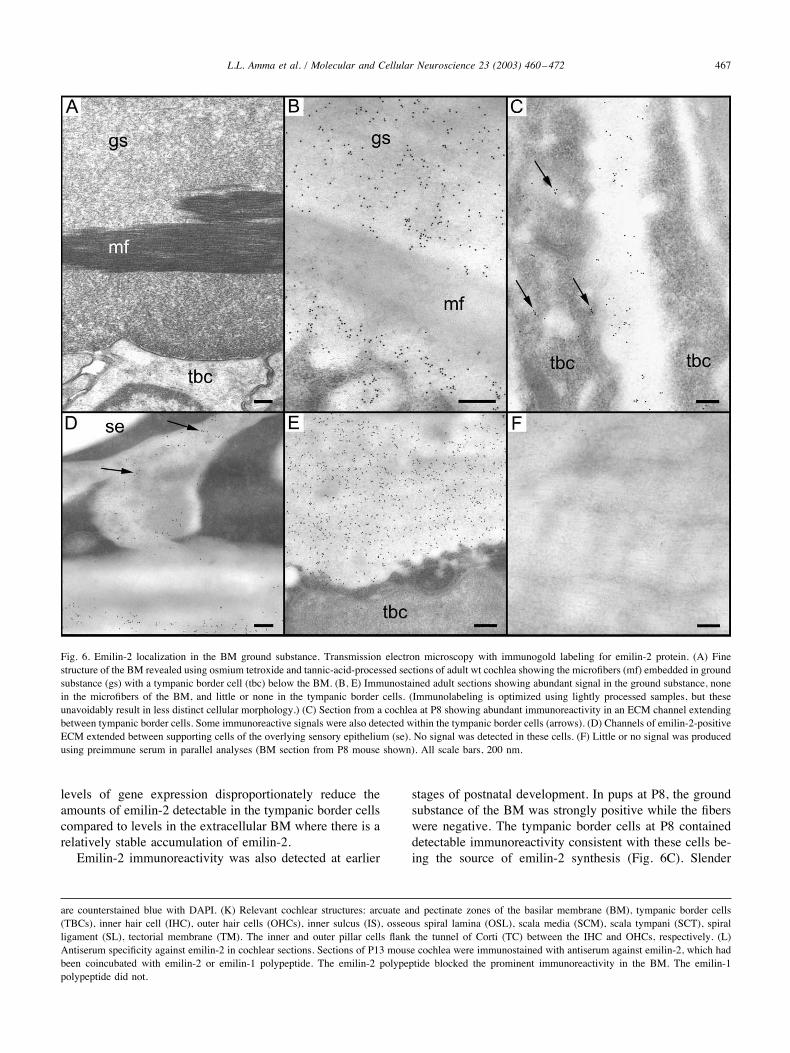
levels of gene expression disproportionately reduce theamounts of emilin-2 detectable in the tympanic border cellscompared to levels in the extracellular BM where there is arelatively stable accumulation of emilin-2.Emilin-2 immunoreactivity was also detected at earlier
stages of postnatal development. In pups at P8, the groundsubstance of the BM was strongly positive while the fiberswere negative. The tympanic border cells at P8 containeddetectable immunoreactivity consistent with these cells be-ing the source of emilin-2 synthesis (Fig. 6C). Slender
are counterstained blue with DAPI. (K) Relevant cochlear structures: arcuate and pectinate zones of the basilar membrane (BM), tympanic border cells(TBCs), inner hair cell (IHC), outer hair cells (OHCs), inner sulcus (IS), osseous spiral lamina (OSL), scala media (SCM), scala tympani (SCT), spiralligament (SL), tectorial membrane (TM). The inner and outer pillar cells flank the tunnel of Corti (TC) between the IHC and OHCs, respectively. (L)Antiserum specificity against emilin-2 in cochlear sections. Sections of P13 mouse cochlea were immunostained with antiserum against emilin-2, which hadbeen coincubated with emilin-2 or emilin-1 polypeptide. The emilin-2 polypeptide blocked the prominent immunoreactivity in the BM. The emilin-1polypeptide did not.
Fig. 6. Emilin-2 localization in the BM ground substance. Transmission electron microscopy with immunogold labeling for emilin-2 protein. (A) Finestructure of the BM revealed using osmium tetroxide and tannic-acid-processed sections of adult wt cochlea showing the microfibers (mf) embedded in groundsubstance (gs) with a tympanic border cell (tbc) below the BM. (B, E) Immunostained adult sections showing abundant signal in the ground substance, nonein the microfibers of the BM, and little or none in the tympanic border cells. (Immunolabeling is optimized using lightly processed samples, but theseunavoidably result in less distinct cellular morphology.) (C) Section from a cochlea at P8 showing abundant immunoreactivity in an ECM channel extendingbetween tympanic border cells. Some immunoreactive signals were also detected within the tympanic border cells (arrows). (D) Channels of emilin-2-positiveECM extended between supporting cells of the overlying sensory epithelium (se). No signal was detected in these cells. (F) Little or no signal was producedusing preimmune serum in parallel analyses (BM section from P8 mouse shown). All scale bars, 200 nm.
467L.L. Amma et al. / Molecular and Cellular Neuroscience 23 (2003) 460–472

channels of ECM extending from the BM between thetympanic border cells were also immunoreactive. Preim-mune serum produced little or no signal in any region of theBM in these analyses (Fig. 6F). The results indicate thatemilin-2 is present in the ground substance of the BMduring postnatal development and in the adult.
Discussion
This study identifies emilin-2 as a major component ofthe cochlear BM. Emilin-2 belongs to a class of glycopro-teins that includes other members found in aorta and endo-thelial tissues, including emilin-1 (Doliana et al., 1999,2001), multimerin (Hayward et al., 1995), and EndoGlyx-1(Christian et al., 2001). These proteins have a modularstructure indicative of self-assembly or interactive functionsthrough domains that include a cysteine-rich N-terminus, acentral region of "-helical coiled-coils, and a C-terminalC1q domain. Recently, a human emilin-2 cDNA was inde-pendently isolated from a human kidney cDNA source (Do-liana et al., 2001). Human emilin-2 is secreted from leiomy-osarcoma cells in culture but little is known of itsphysiological role. The abundance of emilin-2 in the co-chlea at considerably higher levels than in kidney or othertissues (see Fig. 3) suggests a specialized role in the devel-opment or biomechanical function of the cochlear BM.In cochlear development, emilin-2 expression rises
sharply in the postnatal period. In the mouse, the cochlearstructures are largely formed by birth and maturation occursduring the next 2 postnatal weeks before the onset of audi-tory function at around P13 (Pujol et al., 1997). The accu-mulation of emilin-2 in the BM ground substance duringthis period may provide a matrix for the assembly or cohe-sion of other components of the membrane. Emilin-2 is alsodetected in channels of ECM between the tympanic bordercells below the BM and between the cells of the epitheliumabove the BM where it could mediate cellular interactions.Such interactions could provide structural support or assistin cellular differentiation. Of interest, emilin-1 has beensuggested to modify the differentiation of chick embryoaorta cells in culture, since anti-emilin antibodies impedethe deposition of elastin fibers by these cells (Bressan et al.,1993). In the cochlea, the earlier peak and decline of emi-lin-1 expression compared to the more prolonged accumu-lation of emilin-2 may suggest a developmental role foremilin-1 and a more enduring structural role for emilin-2 inthe BM.A possible role for emilin-2 in BM elasticity is suggested
by its close similarity to emilin-1, which is found at theinterface between the elastic core and the microfibrils ofelastic fibers in aorta (Doliana et al., 1999; Colombatti et al.,2000). Tissue elasticity is often conferred by elastin-basedfibers or by microfibrils containing fibrillins or other pro-teins (Ramirez, 1996; Kielty et al., 2002). The identificationof additional constituent proteins of the BM will facilitate
further studies of how emilin-2 associates with other com-ponents involved in BM elasticity. Oligomerization pro-moted by the C1q or collagen domains (Mongiat et al.,2000) or the cysteine-rich Emi domain (Doliana et al., 2000)may be important because both emilin-1 and multimerinform trimeric complexes which can assemble as larger ag-gregates stabilized by disulfide bonds (Colombatti et al.,2000).The zonal variations in emilin-2 expression observed
across the BM may relate to the differing physiologicalfunctions of the inner and outer hair cells in auditory trans-duction (Dallos et al., 1996; Hudspeth, 1997). Emilin-2 wasexpressed across the width of the BM but its expression wasdiminished around the junction between the arcuate andpectinate zones beneath the pillar cells, which separate theinner and outer hair cells. Inner hair cells are thought toserve as the primary sensory cells and transmit auditorysignals to the brain through their abundant afferent inner-vation. In contrast, outer hair cells have less afferent inner-vation but possess an electromotility that amplifies BMmotion and modifies the signals that are relayed through theinner hair cells (Holley, 1996). The major fibers in thepectinate zone below the outer hair cells form a radial arraywhereas in the arcuate zone below the inner hair cells thefibers form a meshwork. It has been proposed that this zonaldivergence allows for a greater stretch of the BM below theouter hair cells than below the inner hair cells followingacoustic stimulation (Mikuni et al., 1995). Also, laser inter-ferometer measurements of BM movements indicate thatwhile both the arcuate and the pectinate zones vibrate, thereis an immobile flexion point beneath the pillar cells (Nilsenand Russell, 1999, 2000). If emilin-2 confers elasticity onthe BM, then less emilin-2 at this site would increase therigidity of this part of the BM. Such zonal differences in BMstructure and composition may be critical to BM functionduring auditory transduction.The function of the tympanic border cells that produce
emilin-2 is not fully defined (Slepecky, 1996). The thick-ness of this cell layer varies along the apical to basal axis ofthe cochlea, which alters the mass associated with the BMand may contribute to the frequency range of the cochlea.Another role for these cells evidently is in the biosynthesisof emilin-2 and other BM components since they have alsobeen reported to express fibronectin and tenascin which arefound in the BM (Santi et al., 1989; Woolf et al., 1992;Swartz and Santi, 1999). In older mice, emilin-2 remaineddetectable in the ECM despite a decline in mRNA levels,suggesting that emilin-2 is relatively stable and requiresonly low maintenance levels of expression in adults. Thissituation resembles that of the secreted tectorin componentsof the tectorial membrane (Rau et al., 1999).Given their defects at relatively late stages of cochlear
development, Thrbtm1/tm1 mice provide a useful model foridentifying sequences that are involved in cochlear matura-tion. Such sequences may reflect direct TR targets or, per-haps more likely, indirectly altered genes, given the range of
468 L.L. Amma et al. / Molecular and Cellular Neuroscience 23 (2003) 460–472

cochlear functions that are regulated by thyroid hormone(Deol, 1973; Uziel, 1986; Forrest et al., 2002). If emilin-2 isdirectly regulated by TR! the modest decrease in emilin-2levels in Thrbtm1/tm1 mice indicates that this regulation isminor and that other factors primarily induce the gene.Combinatorial regulation is common in vivo and TRs oftenmediate only partial or transient control over gene expres-sion (Knipper et al., 2001). However, the largely noncoin-cident expression patterns of TR!, located primarily in thesensory epithelium and other cells above the BM (Bradleyet al., 1994), and emilin-2 in cells below the BM, supportinstead an indirect influence by TR!. Also, computer anal-yses of emilin-2 genomic sequences for putative TR bindingsites failed to identify strong candidates in the inverted ordirect repeat configurations of optimal TR activation sites(Brent et al., 1991), although scattered consensus half-sitemotifs were found. Thus, in cochlear differentiation, TR!may mediate both direct actions (Rusch et al., 1998, 2001)and an indirect chain of events, perhaps involving signalingfactors or cell–cell interactions. Although the cochlear emi-lin-2 sequence was identified from a screen of TR!-defi-cient mice, we cannot exclude entirely a minor role forTR"1, since TR"1 is expressed more widely throughoutcochlear tissues (Bradley et al., 1994).To date, no known deafness-related gene has been
mapped to the murine or human emilin-2 locus. Humanautosomal recessive deafness DFNB19 maps near 18p11but in a more proximal location than the EMILIN-2 gene(R.J. Smith, personal communication; Hereditary HearingLoss Homepage, http://dnalab-www.uia.ac.be/dnalab/hhh/loci_recessive.html). Thus, mutagenesis in a model systemmay be necessary to reveal the role of emilin-2 in thecochlea.
Experimental methods
RNA and Northern blot analysis
Total RNA was prepared from pools of mouse cochlea (n% 30–80 pairs per age) and pituitary using TRI-reagent(Molecular Research Center, Inc.). Poly(A)-selected mRNAwas made from pools of lung, liver, heart, kidney, eye, andbrain regions (n # 12). Northern blots were hybridized athigh stringency (42°C) in buffer containing 50% formamideand were probed with either a 2.3-kb EcoRI fragment ofmouse emilin-2 cDNA (nucleotides 1–2327), or a 1.2-kbmouse emilin-1 cDNA (nucleotides 734–2013, AccessionNo. AK029337) (Doliana et al., 1999). Filters were washedin 0.1( SSC/ 0.2% SDS at 60°C. The mouse emilin-1 probewas isolated by RT-PCR from heart cDNA using the fol-lowing primers 5#-AGCTGCAGCTCCTGGACAACCG-3#and 5#-AGGAGCAACGTTCCACACCATC-3#, located inexon 4 of the gene. Blots were rehybridized with a probe forglyceraldehyde-3P-dehydrogenase (G3PDH) to control forRNA integrity and loading. Probes were labeled with
["-32P]dCTP using a multiprime kit (Amersham/Pharma-cia). The specificity of the mouse emilin-2 and emilin-1cDNA probes was confirmed by Southern blot hybridizationof mouse genomic DNA, digested with BamHI and HindIII.
Mouse strains
The targeted Thrb mutation, deleting all known TR!products, was maintained on a mixed background of 129/Sv( C57BL/6J strains. Thrbtm1/tm1 mice were genotyped byPCR as described (Forrest et al., 1996) and were housedusing 12-h light/12-h dark cycles. All experiments followedapproved institutional protocols.
Differential screen and isolation of cDNA and genomicclones
A differential display PCR was used to screen for se-quences that are altered in expression in Thrbtm1/tm1 versuswt pups at postnatal day 8, since these mice show retardedcochlear development (Rusch et al., 2001). Cochlear totalRNA was treated with DNase I and used to make cDNA asdescribed (Ausubel et al., 1997) with a mixture of randomhexamers or a T-anchored oligonucleotide: TA (5#-CG-GAATTCGGTTTTTTTTTTTTA-3#), TC (5#-CGGAAT-TCGGTTTTTTTTTTTTC-3#), TG (5#-CGGAATTCG-GTTTTTTTTTTTTG-3#). Each cDNA was used at twodifferent concentration in differential display-PCR (DD-PCR) (Liang et al., 1993) using 27 different arbitrary oli-gonucelotides for a total of 432 reactions, in a 20-$l volumeincluding 150 nCi ["-32P]dCTP (3000 Ci/mmol), 1 or 2 $lcDNA, 1 $M arbitrary oligonucleotide, and Tsg DNA poly-merase (Lambda Biotech). PCR conditions were as follows:3 cycles at 94°C for 1 min, 40°C for 4 min and 72°C for 1min; 25 cycles at 94°C for 45 s, 60°C for 2 min and 72°Cfor 1 min; 1 cycle at 72°C for 10 min. Products were heatedat 80°C for 4 min and analyzed on 6% acrylamide/7 M ureagels. Following autoradiography, bands appearing at differ-ent intensities in wt and Thrbtm1/tm1 lanes were excised fromthe gel and eluted in water. Approximately 50 first-roundcandidate bands were detected of which 50% were in-creased and 50% were decreased in Thrbtm1/tm1 versus wtsamples. PCR was repeated on the eluted DNA using 200$M dNTPs and 4 $M of arbitrary oligonucleotide as used inthe original reaction. PCR conditions were as follows: de-naturation at 94°C for 8 min; 35 cycles at 94°C for 30 s,61°C for 30 s and 72°C for 45 s; 72°C for 10 min. SelectedPCR products were ligated into pGEM vectors (Promega)and candidates were tested as probes in Northern blot anal-ysis of cochlear RNA from Thrbtm1/tm1 versus wt pups at P8.Clones containing ribosomal sequences were discarded.Based on Northern blot data, a candidate with reducedexpression in Thrbtm1/tm1 pups was used to screen a P6cochlear cDNA library in %ZAP (Campos-Barros et al.,2000). Bluescript plasmid subclones were isolated from%ZAP clones and sequenced in entirety. 5# RACE was
469L.L. Amma et al. / Molecular and Cellular Neuroscience 23 (2003) 460–472

performed on the P6 cochlear library and on mouse heartmarathon-ready cDNA using Advantage 2 Polymerase(Clontech). The longest 5# ends of the 8 RACE clonessequenced lay within 132 bp of the 5# end of the mouseemilin-2 cDNA (Fig. 1B).The mouse emilin-2 gene was isolated on a BAC
genomic clone using a PCR screen with primer pairs span-ning exons 1 and 2 (Incyte Genomics). The BAC clone wasshown to contain all exons by PCR analyses; the detailedstructure of the gene was confirmed using sequence data-bases (Celera and public). The mouse emilin-2 cDNA se-quence (initially named “basilin”) has GenBank AccessionNo. AF468645.
Protein translation
In vitro transcription/translation was conducted in reticu-locyte lysates (Promega, TNT). A Bluescript plasmid clonecontaining the cochlear cDNA with a complete emilin-2open reading frame was transcribed in sense and antisensedirections using T3 and T7 RNA polymerases, respectively,and translated in the presence of 35S-methionine. Productswere analyzed on 6% polyacrylamide–SDS gels, and visu-alized by autoradiography. Parallel reactions were per-formed in the absence of 35S-methionine for analysis byWestern blot using the antiserum raised against mouse emi-lin-2 (see below).
Sequence analysis
Related proteins detected by Blast searches (Zhang andMadden, 1997) were aligned using a Clustal W algorithm(version 1.8). The Expasy site was used to predict theN-terminal cleavage site (Signal P program) and coiled-coildomains (Paircoil program, Berger’s method; probability31–100%) (http://www.expasy.ch). Leucine zippers and N-glycosylation sites were identified using the GeorgetownUniversity Protein Information Resource program (http://www-nbrf.georgetown.edu). The 0.8 kb of genomic se-quence preceding exon 1 was analyzed by the eukaryoticneural network promoter prediction program (Baylor Col-lege of Medicine) (Reese and Eekman, 1995) with a lowercutoff set at 0.85 of a maximum possible score of 1.0. The15.0 kb of DNA up- and downstream of exon 1 of theemilin-2 gene was analyzed for putative TR binding sitesrelated to the consensus halfsites 5#-AGGTCA-3#.
In situ hybridization analysis
Cochleae were dissected and fixed for 2–4 h in 4%paraformaldehyde (PFA) and decalcified in 50 mM EDTAfor 2 weeks. Samples were placed in 30% sucrose over-night, embedded in OCT medium (Tissue-Tek), for cryo-sectioning at 10-$m thickness. Sections were postfixed for5 min in 4% PFA, rinsed once in 3( PBS and twice in 1(PBS, and dehydrated using graded ethanol concentrations
(30–100%). Slides were stored at '70°C prior to use.Slides were rinsed for 5 min in 1( PBS, treated withproteinase K (1.0 $g/ml) at room temperature for 5 min,fixed for 5 min in 4% PFA, rinsed in 1( PBS, 3 times for5 min, acetylated (0.1 M triethanol-amine/0.25% acetic an-hydride, pH 8) for 10 min, and treated with 1% TritonX-100 for 30 min. Slides were washed three times with 1(PBS prior to a 2-h prehybridization in 40% formamide, 10%dextran sulfate, 1( Denhardt’s solution, 4( SSC, 10 mMDTT, 1 mg/ml yeast tRNA, and 100 $g/ml of boiled salmonsperm DNA. After probe addition, slides were coverslippedand incubated in a humid chamber overnight at 61 or 67°C(both temperatures gave comparable results). Slides werewashed twice at 65°C in 50% formamide/2( SSC for 45min, twice in 25% formamide/1( SSC/0.5( PBS for 30min, and then treated with 20 mg/ml Rnase A for 30 min at37°C. After being blocked with 10% sheep serum and 1%blocking reagent in maleic acid buffer for 1 h, samples wereincubated with a 1:1000 dilution of DIG-specific antibody(Roche), rinsed 3 ( 15 min in maleic acid buffer, anddeveloped using the DIG Nucleic Acid Detection kit(Roche). A 277-bp PstI fragment (nucleotides 959–1236) ofemilin-2 cDNA, subcloned into PGem3Zf(!) (Promega),was linearized for generation of sense and antisense ribo-probes by runoff transcription using the DIG RNA Labelingkit (Roche). Probes were heated for 5 min at 85°C anddiluted with hybridization buffer to a concentration of 200ng/ml.
Antibody production and immunostaining
An 849-bp StuI–HindIII fragment of mouse emilin-2cDNA (codons 211–494) was cloned into PQE30 (Qiagen)carrying a (His)6 tag at the N-terminus. After transformationinto MC-15 Escherichia coli, the clone expressed apolypeptide of expected size (34.3 kDa, including vectortag) when induced with 2 mM IPTG. After 5 h of growth,bacteria were lysed in 6 M guanidine hydrochloride, 0.1 MNaH2PO4, 0.01 M Tris, pH 8.0. Ni-NTA resin (Qiagen) wasadded to the lysate supernatant and allowed to bind over-night. The resin was packed in a 10-ml column (Bio-Rad)and protein was eluted with 8 M urea, 0.1 M NaH2PO4, 0.01M Tris at sequential pH values of 8.0, 6.3, 5.8, 5.5, 5.0, and4.5. Most protein eluted between pH 5.8 and 5.0 and thesefractions were dialyzed (molecular weight cutoff 3500 kDa,Spectra/Por) against 15 mM Tris, pH 8.0, 10% glycerol, 50mM NaCl, 5 mM MgCl2, 5 mM DTT, with decreasing ureaconcentrations (4.0, 1.0, 0.4, 0.1, and 0.0 M) for 6 h eachchange. Dialyzed protein was used to generate antiserum ina New Zealand white rabbit (Covance Research Products,Denver, PA). After preimmune serum was obtained, therabbit was injected with 250 $g of antigen conjugated withFreund’s complete adjuvant. Boosts of 125 $g of antigenconjugated with Freund’s incomplete adjuvant were givenon days 21, 42, and 63 followed by bleeds 10 days after
470 L.L. Amma et al. / Molecular and Cellular Neuroscience 23 (2003) 460–472

each boost. ELISA was performed to determine antiserumtiters.For immunostaining, cochleae were fixed and sectioned
as described for in situ hybridization. Sections were blockedat room temperature for 30 min with 0.01% BSA, 0.4%Triton X-100, and 1.5% normal goat serum in PBS andincubated overnight with a 1:10,000 dilution of either pre-immune serum or antiserum. The following day sectionswere stained using the Vectastain ABC kit and DAB per-oxidase substrate (Vector Laboratories) for colorigenic de-tection. Some sections were stained with FITC-conjugatedanti-rabbit antibody for immunofluorescence. For antibodyblocking studies, a 944-bp PstI fragment of mouse emilin-1cDNA (codons 247–554) was cloned into PQE-31 with a(His)6 tag at the N-terminus. After transformation intoMC-15 E. coli, the clone expressed a protein of expectedsize (36.2 kDa, including vector tag), which was purified asdescribed above for emilin-2. Antiserum against emilin-2was preincubated with equivalent amounts of emilin-2 oremilin-1 polypeptides for 1 h. The mix was then overlaid oncochlear cryosections using the antiserum at 1:10,000 dilu-tion and processed as described above, using the VectastainABC Elite kit and peroxidase substrate. Emilin-1 and emi-lin-2 polypeptides were derived from similar midregions ofthe two proteins (see Fig. 1B). In a linear alignment, theseregions overlapped for 224 amino acids but possessed onlya scattered amino acid identity (36/224 residues), which wasfurther interrupted by several deletions relative to eachother.
Electron microscopy and immunogold labeling
Cochleae (from mice at P1, P8, and P21) were dissectedin PBS, fixed for 1 h in 3.7% formaldehyde buffered with0.1 M sodium phosphate, pH 7.4, washed three times withPBS, and decalcified in 0.5 M EDTA, pH 8.0, for 3 days at4°C. Following dehydration with ethanol, samples wereimbedded in Unicryl resin (British BioCell International,Cardiff, UK) as described (Thorpe et al., 1999). Ultrathinsections (90 nm) were mounted on Formvar-coated coppergrids, preblocked for 1 h with PBS! (150 mM NaCl, 10mM sodium phosphate, pH 8.2, containing 1% BSA, 0.05%Tween 20, 10 mM NaEDTA, and 0.02% sodium azide), andstained overnight with anti-basilin serum (1:500 dilution inPBS!). Following washing, bound antibodies were de-tected with 10 nm gold conjugated goat anti-rabbit antibod-ies (British BioCell International) at a dilution of 1:10 inPBS! for 30 min. Sections were counterstained with uranylacetate and lead citrate and viewed in a Hitachi 7100 trans-mission electron microscope operating at 75 kV. Full detailsof the postimbedding staining procedure are as described(Thorpe, 1999).To compare BM morphology in the lightly fixed, non-
osmicated tissues used for postimbedding immunogold la-beling with that in conventionally fixed samples, cochleaewere also fixed in 2.5% glutaraldehyde containing 1% tan-
nic acid, postfixed in 1% osmium tetroxide, and processedfor transmission electron microscopy using standard proce-dures.
Acknowledgments
We thank Drs. I. Russell and F. Ramirez for commentson the manuscript. This work was supported in part by NIH(DC-03441), the Hirschl Trust and March of Dimes BirthDefects Foundation (D.F.), and the Wellcome Trust(057410/Z/99/Z; R.G. and G.R.).
References
Ausubel, F., Brent, R., Kingston, R., More, D., Seidman, J., Smith, J.,Struhl, K., 1997. Current Protocols in Molecular Biology. Wiley, NewYork.
Bradley, D.J., Towle, H.C., Young III, W.S., 1994. " and ! thyroidhormone receptor (TR) gene expression during auditory neurogenesis:evidence for TR isoform-specific transcriptional regulation in vivo.Proc. Natl. Acad. Sci. USA 91, 439–443.
Brent, G., Moore, D., Larsen, P., 1991. Thyroid hormone regulation ofgene expression. Ann. Rev. Physiol. 53, 17–35.
Bressan, G.M., Daga-Gordini, D., Colombatti, A., Castellani, I., Marigo,V., Volpin, D., 1993. Emilin, a component of elastic fibers preferen-tially located at the elastin–microfibrils interface. J. Cell Biol. 121,201–212.
Brucker-Davis, F., Skarulis, M.C., Pikus, A., Ishizawar, D., Mastroianni,M.-A., Koby, M., Weintraub, B.D., 1996. Prevalence and mechanismsof hearing loss in patients with resistance to thyroid hormone (RTH).J. Clin. Endocrinol. Metab. 81, 2768–2772.
Cabezudo, L.M., 1978. The ultrastructure of the basilar membrane in thecat. Acta Otolaryngol. 86, 160–175.
Campos-Barros, A., Amma, L.L., Faris, J.S., Shailam, R., Kelley, M.W.,Forrest, D., 2000. Type 2 iodothyronine deiodinase expression in thecochlea before the onset of hearing. Proc. Natl. Acad. Sci. USA 97,1287–1292.
Christian, S., Ahorn, H., Novatchkova, M., Garin-Chesa, P., Park, J.E.,Weber, G., Eisenhaber, F., Rettig, W.J., Lenter, M.C., 2001. En-doGlyx-1: molecular cloning and characterization of an EMILIN-likemultisubunit glycoprotein of vascular endothelium. J. Biol. Chem. 14,14.
Cohen-Salmon, M., El-Amraoui, A., Leibovici, M., Petit, C., 1997. Oto-gelin: a glycoprotein specific to the acellular membranes of the innerear. Proc. Natl. Acad. Sci. USA 94, 14450–14455.
Colombatti, A., Doliana, R., Bot, S., Canton, A., Mongiat, M., Mun-giguerra, G., Paron-Cilli, S., Spessotto, P., 2000. The EMILIN proteinfamily. Matrix Biol. 19, 289–301.
Dallos, P., Popper, A., Fay, R., 1996. The Cochlea. Springer-Verlag, NewYork.
Deol, M.S., 1973. An experimental approach to the understanding andtreatment of hereditary syndromes with congenital deafness and hypo-thyroidism. J. Med. Genet. 10, 235–242.
Doliana, R., Bot, S., Bonaldo, P., Colombatti, A., 2000. EMI, a novelcysteine-rich domain of EMILINs and other extracellular proteins,interacts with the gC1q domains and participates in multimerization.FEBS Lett. 484, 164–168.
Doliana, R., Bot, S., Mungiguerra, G., Canton, A., Cilli, S.P., Colombatti,A., 2001. Isolation and characterization of EMILIN-2, a new compo-nent of the growing EMILINs family and a member of the EMIdomain-containing superfamily. J. Biol. Chem. 276, 12003–12011.
471L.L. Amma et al. / Molecular and Cellular Neuroscience 23 (2003) 460–472

Doliana, R., Mongiat, M., Bucciotti, F., Giacomello, E., Deutzmann, R.,Volpin, D., Bressan, G.M., Colombatti, A., 1999. EMILIN, a compo-nent of the elastic fiber and a new member of the C1q/tumor necrosisfactor superfamily of proteins. J. Biol. Chem. 274, 16773–16781.
Dreiling, F.J., Henson, M.M., Henson, O.W., 2002. The presence andarrangement of type II collagen in the basilar membrane. Hear. Res.166, 166–180.
Forrest, D., Erway, L.C., Ng, L., Altschuler, R., Curran, T., 1996. Thyroidhormone receptor ! is essential for development of auditory function.Nat. Genet. 13, 354–357.
Forrest, D., Reh, T., Rusch, A., 2002. Neurodevelopmental control bythyroid hormone receptors. Curr. Opin. Neurobiol. 12, 49–56.
Gillespie, P.G., Walker, R.G., 2001. Molecular basis of mechanosensorytransduction. Nature 413, 194–202.
Griffith, A., Friedman, T., 2002. Autosomal and X-linked auditory disor-ders, in: Keats, B., Popper, A., Fay, R. (Eds.), Genetics and AuditoryDisorders, Springer-Verlag, New York, pp. 121–227.
Hayward, C.P., Hassell, J.A., Denomme, G.A., Rachubinski, R.A., Brown,C., Kelton, J.G., 1995. The cDNA sequence of human endothelial cellmultimerin. A unique protein with RGDS, coiled-coil, and epidermalgrowth factor-like domains and a carboxyl terminus similar to theglobular domain of complement C1q and collagens type VIII and X.J. Biol. Chem. 270, 18246–18251.
Heller, S., Sheane, C.A., Javed, Z., Hudspeth, A.J., 1998. Molecular mark-ers for cell types of the inner ear and candidate genes for hearingdisorders. Proc. Natl. Acad. Sci. USA 95, 11400–11405.
Holley, M., 1996. Outer hair cell motility, in: Dallos, P., Popper, A., Fay,R. (Eds.), The Cochlea, Springer-Verlag, New York, pp. 386–434.
Hudspeth, A., 1997. How hearing happens. Neuron 19, 947–950.Iurato, S., 1967. Submicroscopic Structure of the Inner Ear. Pergamon,
Oxford.Katori, Y., Hozawa, K., Kikuchi, T., Tonosaki, A., Takasaka, T., 1993.
Fine structure of the lamina basilaris of guinea pig cochlea. ActaOtolaryngol. 113, 715–719.
Kielty, C.M., Sherratt, M.J., Shuttleworth, C.A., 2002. Elastic fibres. J. CellSci. 115, 2817–2828.
Knipper, M., Richardson, G., Mack, A., Muller, M., Goodyear, R., Lim-berger, A., Rohbock, K., Kopschall, I., Zenner, H.P., Zimmermann, U.,2001. Thyroid hormone-deficient period prior to the onset of hearing isassociated with reduced levels of beta-tectorin protein in the tectorialmembrane: implication for hearing loss. J. Biol. Chem. 276, 39046–39052.
Kozak, M., 1987. An analysis of 5# non-coding sequences from 699vertebrate messenger RNAs. Nucleic Acids Res. 15, 8125–8148.
Legan, P.K., Rau, A., Keen, J.N., Richardson, G.P., 1997. The mousetectorins. Modular matrix proteins of the inner ear homologous tocomponents of the sperm–egg adhesion system. J. Biol. Chem. 272,8791–8801.
Leimeister, C., Steidl, C., Schumacher, N., Erhard, S., Gessler, M., 2002.Developmental expression and biochemical characterization of Emufamily members. Dev. Biol. 249, 204–218.
Liang, P., Averboukh, L., Pardee, A.B., 1993. Distribution and cloning ofeukaryotic mRNAs by means of differential display: refinements andoptimization. Nucleic Acids Res. 21, 3269–3275.
Mikuni, H., Fukuda, S., Kucuk, B., Inuyama, Y., Ushiki, T., Abe, K., 1995.The three-dimensional fibrillar arrangement of the basilar membrane inthe mouse cochlea. Eur. Arch. Otorhinolaryngol. 252, 495–498.
Mongiat, M., Mungiguerra, G., Bot, S., Mucignat, M.T., Giacomello, E.,Doliana, R., Colombatti, A., 2000. Self-assembly and supramolecularorganization of EMILIN. J. Biol. Chem. 275, 25471–25480.
Mount, S.M., 1982. A catalogue of splice junction sequences. NucleicAcids Res. 10, 459–472.
Munyer, P.D., Schulte, B.A., 1994. Immunohistochemical localization ofkeratan sulfate and chondroitin 4- and 6-sulfate proteoglycans in sub-regions of the tectorial and basilar membranes. Hear. Res. 79, 83–93.
Nilsen, K.E., Russell, I.J., 1999. Timing of cochlear feedback: spatial andtemporal representation of a tone across the basilar membrane. Nat.Neurosci. 2, 642–648.
Nilsen, K.E., Russell, I.J., 2000. The spatial and temporal representation ofa tone on the guinea pig basilar membrane. Proc. Natl. Acad. Sci. USA.97, 11751–11758.
Pujol, R., Lavigne-Rebillard, M., Lenoir, M., 1997. Development of sen-sory and neural structures in the mammalian cochlea, in: Rubel, E.,Popper, A., Fay, R. (Eds.), Development of the Auditory System,Springer-Verlag, New York, pp. 146–192.
Ramirez, F., 1996. Fibrillin mutations in Marfan syndrome and relatedphenotypes. Curr. Opin. Genet. Dev. 6, 309–315.
Rau, A., Legan, P.K., Richardson, G.P., 1999. Tectorin mRNA expressionis spatially and temporally restricted during mouse inner ear develop-ment. J. Comp. Neurol. 405, 271–280.
Reese, M., Eekman, F., 1995. Novel neural network algorithms for im-proved eukaryotic promoter site recognition. The seventh internationalgenome sequencing and analysis conference, Hilton Head Island, SC.
Resendes, B.L., Williamson, R.E., Morton, C.C., 2001. At the speed ofsound: gene discovery in the auditory system. Am. J. Hum. Genet. 69,923–935.
Rodgers, K.D., Barritt, L., Miner, J.H., Cosgrove, D., 2001. The lamininsin the murine inner ear: developmental transitions and expression incochlear basement membranes. Hear. Res. 158, 39–50.
Rusch, A., Erway, L., Oliver, D., Vennstrom, B., Forrest, D., 1998. Thy-roid hormone receptor !-dependent expression of a potassium conduc-tance in inner hair cells at the onset of hearing. Proc. Natl. Acad. Sci.USA 95, 15758–15762.
Rusch, A., Ng, L., Goodyear, R., Oliver, D., Lisoukov, I., Vennstrom, B.,Richardson, G., Kelley, M.W., Forrest, D., 2001. Retardation of co-chlear maturation and impaired hair cell function caused by deletion ofall known thyroid hormone receptors. J. Neurosci. 21, 9792–9800.
Santi, P.A., Larson, J.T., Furcht, L.T., Economou, T.S., 1989. Immunohis-tochemical localization of fibronectin in the chinchilla cochlea. Hear.Res. 39, 91–101.
Slepecky, N., 1996. Structure of the mammalian cochlea, in: Dallos, P.,Popper, A., Fay, R. (Eds.), The Cochlea, Springer-Verlag, New York,pp. 44–129.
Steel, K.P., Kros, C.J., 2001. A genetic approach to understanding auditoryfunction. Nat. Genet. 27, 143–149.
Swartz, D.J., Santi, P.A., 1999. Immunolocalization of tenascin in thechinchilla inner ear. Hear. Res. 130, 108–114.
Thalmann, I., 1993. Collagen of accessory structures of organ of Corti.Connect. Tissue Res. 29, 191–201.
Thorpe, J., 1999. The application of LR Gold resin for immunogoldlabeling, in: Haijbagheri, M. (Ed.), Electron Microscopy Methods andProtocols, Humana Press, Totowa, NJ, pp. 99–110.
Thorpe, J.R., Rulten, S.L., Kay, J.E., 1999. Binding of a putative and aknown chaperone protein revealed by immunogold labeling transmis-sion electron microscopy: a suggested use of chaperones as probes forthe distribution of their target proteins. J. Histochem. Cytochem. 47,1633–1640.
Tsuprun, V., Santi, P., 1999. Ultrastructure and immunohistochemicalidentification of the extracellular matrix of the chinchilla cochlea. Hear.Res. 129, 35–49.
Uziel, A., 1986. Periods of sensitivity to thyroid hormone during thedevelopment of the organ of Corti. Acta Otolaryngol. (Stockh.) 429,23–27.
Woolf, N.K., Koehrn, F.J., Ryan, A.F., 1992. Immunohistochemical local-ization of fibronectin-like protein in the inner ear of the developinggerbil and rat. Brain Res. Dev. Brain Res. 65, 21–33.
Zhang, J., Madden, T.L., 1997. PowerBLAST: a new network BLASTapplication for interactive or automated sequence analysis and annota-tion. Genome Res. 7, 649–656.
472 L.L. Amma et al. / Molecular and Cellular Neuroscience 23 (2003) 460–472




















