
Sequential Depletion and Acquisition of Proteins during Golgi Stack Disassembly and Reformation
Upload
ussacademyCategory
view
1download
0
Molecular Brain Research, 15 (1992) 195-206 195 © t992 Elsevier Science Publishers B.V. All rights reserved 0t69-328x/92/$05,00
BRESM ~ 4 ~
/3-Amyloid precursor protein localization in the Golgi apparatus in neurons and oligodendrocytes. An immunocytochemical structural
and ultrastructural study in normal and axotomized neurons
G. Palacios a, J.M. Palacios b, G. Mengod b and P. Frey c
a Department of Cellular Biology and Physiology, Subunit of Histology, Faculty of Medicine, Universidad Autdnoma de Barcelona, Barcelona (Spain), b Department of Neurochemistry, CID-CSIC, Barcelona (Spain) and c Sandoz Forschungsinstitut Bern, Bern (Switzerland)
(Accepted 21 April 1992)
Key words: /3-Amyloid precursor protein; Rat brain; Immunocytochemistry; Axotomy; Facial nucleus; Alzheimer's disease
We have used a polyclonal antibody raised against a synthetic peptide from the carboxyl terminal of the/3-amyloid precursor protein (APP) to examine the cellular and subcellular localization of this protein in the rat brain. Light and electron microscopic immunocytochemical techniques were used. Immunoreactivity was found throughout the brain in all the neurons examined as well as in oligodendrocytes. At the light microscopic level, a perinuclear filamentous distribution was seen in neurons, suggesting a concentration of the protein to the Golgi apparatus. Axotomy of motor neurons of the facial nucleus produced a decrease in choline acetyltransferase (CHAT) activity and an increase in the perineuronal microglial nueleoside diphosphatase (NDPase)-positive cells in addition to a hypertrophy of the GFAP immunoreactive astrocytes. On the other hand, increased APP-like immunoreactivity all over the neuronal cell bodies accompanied by a dispersion ('rete dispersion') of the Golgi apparatus labeling was demonstrated. In contrast, reactive mieroglia and hypertrophic astrocytes in axotomized facial nucleus were not immunolabeled. Oligodendrocytes showed a punctate APP immunoreactivity corresponding to the Golgi apparatus in both operated and control facial nucleus. This was further demonstrated by electron microscopic immunolabeling. These results show that the main localization of the C-terminal containing forms of the APP in the rat brain is the Golgi apparatus in both neurons and oligodendroglia and further supporting the secretory nature of these proteins. The increased synthesis of this protein after axotomy is suggestive of a role of the APPs in growth and/or regeneration.
INTRODUCTION
A charac te r i s t i c c o m p o n e n t of the seni le p l aques in
the b ra in of the pa t i en t s with A l z h e i m e r - t y p e d e m e n t i a
( A D ) is the amylo id B-p ro t e in or A 4 pro te in . This
f l -amylo id p e p t i d e is pa r t o f four p r ecu r so r po lypep-
t ides, all e n c o d e d by the same single gene loca l ized in c h r o m o s o m e 2111'17'35'45. T h e B-amylo id p r ecu r so r pro-
te in ( A P P ) gene has 18 exons, which, by a l t e rna t ive
splicing, p r o d u c e s d i f fe ren t forms of m R N A encod ing
for four p r e c u r s o r p ro t e ins o f 695, 714, 750, and 770 a m i n o acids long 10'18'20'34'46. T h e two l a rge r forms con-
ta in a d o m a i n shar ing close s imi lar i ty wi th the Kuni tz
family o f se r ine p r o t e a s e inh ib i tors (KPI domain) . Bo th
B A P P 714 and BAPP770, share an add i t i ona l sequence
o f 19 a m i n o acids wi th s imi lar i t ies to M R C OX-2
an t igen 6. The A P P has been p o s t u l a t e d to have a s ingle
m e m b r a n e - s p a n n i n g d o m a i n with a long ex t race l lu la r
N- t e rminus and a shor t in t race l lu la r C- te rminus . The
B-amylo id p e p t i d e sequence begins on the ex t race l lu la r
s ide close to the p l a s m a m e m b r a n e and ends inside o f
the pu ta t ive t r a n s m e m b r a n e reg ion 17. The A P P gene
and the c o r r e spond ing a l te rna t ive spl icing forms have also b e e n c loned in ra t and mouse 18'27'37'52'53.
Dif fe ren t funct ions have been sugges ted for the
d i f fe ren t APPs . They may m e d i a t e c e l l - c e l l or c e l l -
mat r ix in te rac t ions , act as cell sur face recep tors , p ro -
mo te neu r i t e g rowth o r have p r o t e a s e inh ib i tor activ- ity 38.
A b n o r m a l pro teolys is of A P P has b e e n p r o p o s e d as
a f u n d a m e n t a l s tep in the pa thoge ne s i s of f l -amylo id
depos i t i on into seni le p l aques 39. The p r e s e n c e of B-
Correspondence: G. Palacios, Subunidad de Histoiogia, Facultad de Medicina, Universidad Aut6noma de Barcelona, Bellaterra, Barcelona 08193, Spain.

196
amylo id co res in t he classic p l a q u e s in A D in associa-
t ion wi th n e u r o n and n e u r i t e d e g e n e r a t i o n and acti-
v a t e d gl ial cel ls has led to the p r o p o s a l o f m u l t i p l e s
sou rces for t he /3-amyloid depos i t s such as a l t e r e d
n e u r o n s 2~, a s t rocy tes 41, m i c r o g l i a 49'5°, and e n d o t h e l i a l
cel ls 3~. H o w e v e r , the o r ig in o f p l a q u e amylo id is still
u n k n o w n since in si tu h y b r i d i z a t i o n and i m m u n o c y t o -
c h e m i c a l s tud ies have d e m o n s t r a t e d tha t v i r tua l ly eve ry
cel l in t he cen t r a l and p e r i p h e r a l n e r v o u s systems,
i nc lud ing n e u r o n s , as t rocytes , mic rog l i a , o l i g o d e n -
d rog l ia and e n d o t h e l i a l cel ls as wel l as s o m e non
n e u r a l cel ls and t i ssues express and c o n t a i n t he A P P m R N A s and the i r products 1"5'15'19"27"3°'4°'41"45.
In o r d e r to u n d e r s t a n d the p a t h o g e n e s i s o f A D it is
necessa ry to o b t a i n i n f o r m a t i o n on the type o f cel ls
involved in t he m e t a b o l i s m of the A P P and in the
d e p o s i t i o n of its de r iva t ives such as the /3-amyloid
p ro te in , at bo th ce l lu l a r and subce l l u l a r levels
In the p r e s e n t s tudy we have u s e d a po lyc lona l
an t i body aga ins t a c a r b o x y - t e r m i n a l f r a g m e n t o f t he
A P P in o r d e r to e x a m i n e at the s t ruc tu ra l and u l t ra -
s t ruc tu ra l level t he l oca l i za t ion and d i s t r i bu t ion o f t he
i m m u n o r e a c t i v i t y t h r o u g h o u t t he rat bra in . W e also
have e v a l u a t e d the c h a n g e s in A P P i m m u n o r e a c t i v i t y
fo l lowing un i l a t e r a l axo tomy of t he facia l ne rve .
MATERIALS AND METHODS
Antisera The antibody against APP was obtained by immunizing New
Zealand female rabbits with a synthetic peptide fragment of the C-terminal end of APP (APP 676-695E7). The peptide was synthe- sized with an ABI synthesizer via the FMOC method. The sequence was verified by amino acid analysis and FAB mass spectrometry. The peptide was coupled to keyhole limpet hemocyanine with glutaralde- hyde. The antigen thus obtained was emulsified with Freund's Com- plete Adjuvant and injected partly intramuscularly, partly intrader- really on multiple sites on the back of the rabbit. After two weeks, the immunization was repeated with Freund's Incomplete Adjuvant. Thereafter, the rabbits were immunized in 6 weeks intervals and blood was taken 7-10 days after each injection. The antiserum thus obtained was tested in immunoblots with rat brain extracts, which had been subjected to separation on 10% acrylamide Tris/tricinc SDS gels beforehand 36. Proteins were transferred onto a nitrocellu- lose membrane by electrophoresis. After blocking non-specific bind- ing with gelatin, the filter was incubated overnight with the APP-an- tiserum, followed by incubation with a goat anti-rabbit antiserum (Sigma), to which alkaline phosphatase had been coupled. Glial fibrillary acidic protein (GFAP) and choline acetyl transferase (CHAT) antibodies were obtained commercially from Dako (Den- mark) and Boehringer Manheim (FRG), respectively.
Animals, surgery and tissue pro~ essing Thirty adult, male and female Sprague-Dawley rats were used.
Ten rats were anaesthetized with sodium pentobarbital (60 r ag / kg,i.p.) and the right facial nerve was cut near its exit from stylomas- toid foramen. At 3, 5, 7, 14, 21 days following nerve section, these animals and the other twenty control animals were perfused trans cardially under ether anesthesia with 100 ml of physiological saline and then with 4% paraformaldehyde, 0.1% glutaraldehyde and 15% saturated picric acid in 0.1M phosphate buffer pH 7.4 (PB). The brains were removed and further fixed for 4 h at 4°C with 4c~
paraformaldehyde in ILl M PB, and then were left overnight in 5% sucrose in 0.1 M PB at 4°C. Coronal sections (40 p~m) were cut with a vibratome (Lancer) throughout the brain and brainstem at the nu- cleus facialis level. The sections were then washed for 12-20 h in 0.1 M PB at 4°C.
Immunocytochemical and histochemical procedures Both the avidin-biotin-peroxidase E~' (PAP) and the peroxidase-
antiperoxidase (PAP) technique 44 were used for light and electron microscope localization of APP. The steps used with the sections processed according to the ABC method were as follows: APP antibody (diluted 1 : 1000 in PBS containing 1% normal goat serum) for 48 h at 4°C, biotinylated goat anti-rabbit IgG (Sigma Co, 1 : 100 dilution, 2 h at 21°C), ABC complex (Sigma Co, 1:400 dilution, 1 h at 21°C). The sections processed according to the PAP technique were incubated in the same way with the primary antibody followed by goat anti-rabbit IgG (Sternberger Monoclonals Inc. Baltimore, MD, USA, 1:40 dilution, 2 h at 21°C) and rabbit PAP complex (Sternberger Monoclonals Inc., Baltimore, MD USA, 1 : 100 dilution, 2 h at 21°C).
The same ABC procedure was used in some sections at the facial nuclei level to localize the GFAP, a well-defined astrocytic marker 12. The primary antibody was rabbit anti-GFAP (1:2000 dilution, 24 h at 4°C). Following sequential incubations with biotinylated goat anti- rabbit IgG (Amershan, 1:200 dilution, 1 h at 21°C) and ABC complex (Sigma Co, 1 : 100 dilution, 1 h at 21°C).
Peroxidase activity was finally visualized by incubating the sec- tions with 0.05% 3,3'diaminobenzidine (DAB) and 0.01% H20 2 in PB for 5-10 min. Some sections were processed with 1-naphthol solution for 5 rain followed by Cresyl violet basic-dye solution (I- NBD) for 2 rain 29,
Controls were obtained using sections treated in the same way except that the primary antibody was omitted. Preabsorption of APP antiserum with purified peptide, completely abolished the immuno- cytochemical staining.
Cholinergic neurons and microglial cells were visualized simulta- neously on the same vibratome section by a double immunocyto- chemical and histochemical technique as described previously 33. Other sections were mounted on slides with Glycergel (Dako) for light microscopic observation and photography of immunostained regions. For electron microscope observation, blocks obtained with punches from selected immunostained brain areas from both axo- tomized facial nucleus and control, were postfixed in 1% OsO 4 for 1 h, block-stained in 1% uranyl acetate veronal buffer, dehydrated and flat-embedded in Durcupan (Fluka). Semithin sections (1 /zm) were cut on a LKB Ultratome III and stained with 1% Toluidine blue for light-microscope photography. Ultrathin sections were also obtained and stained with alcoholic uranyl acetate for l0 rain. They were later examined and photographed with a Hitachi H-7000 electron micro- scope.
R E S U L T S
Characterization of the APP antiserum T h e speci f ic i ty o f t h e A P P - a n t i s e r u m was t e s t ed by
i n c u b a t i n g b lo ts of ra t b ra in ex t rac t s s e p a r a t e d on a
10% a c r y l a m i d e gel wi th t he a n t i s e r u m . T w o m a j o r
bands in t he e x p e c t e d m o l e c u l a r w e i g h t r a n g e o f A P P
(tool. wt. 105-115 k D a ) and o f a poss ib le f r a g m e n t o f
A P P (mol , wt. 12 k D a ) w e r e o b t a i n e d (Fig. 1, l ane A).
T h e s e bands d i s a p p e a r e d w h e n the A P P a n t i s e r u m was
p r e a b s o r b e d wi th t he syn the t ic p e p t i d e A P P 6 7 6 - 6 9 5
( excep t for a m i n o r h igh m o l e c u l a r w e i g h t c o n t a m i n a n t
at = 125 k D a ) (F ig . l , l ane B). P r e i m m u n e s e r u m was
n e g a t i v e for s t a in ing in i m m u n o b l o t (Fig. 1, l ane C).

A B C i i i i l ! !
100--- 68---
43
30 20 - - 11--
! ,
Fig. 1. Immunoblot of a crude rat brain homogenate separated on a 10% acrylamide gel with APP antiserum directed to the carboxy terminus of APP. Lane A: reacted with APP antiserum; lane B: reaction with the APP-antiserum preabsorbed with synthetic peptide APP 676-695; lane C: preimmune serum. Numbers on the left indicate the position of the molecular weight markers phosphorylase b, BSA, ovalbumin, carbonic anhydrase, soybean trypsin inhibitor
and cytochrome c.
Immunocytochemical APP localization in the central nervous system in control rats
Immunoreactivity was observed in virtually all neu- rons examined throughout the central nervous system. Similar patterns were seen with the two immunodetec- tion procedures used. Results shown are those ob- tained from sections treated with the ABC method.
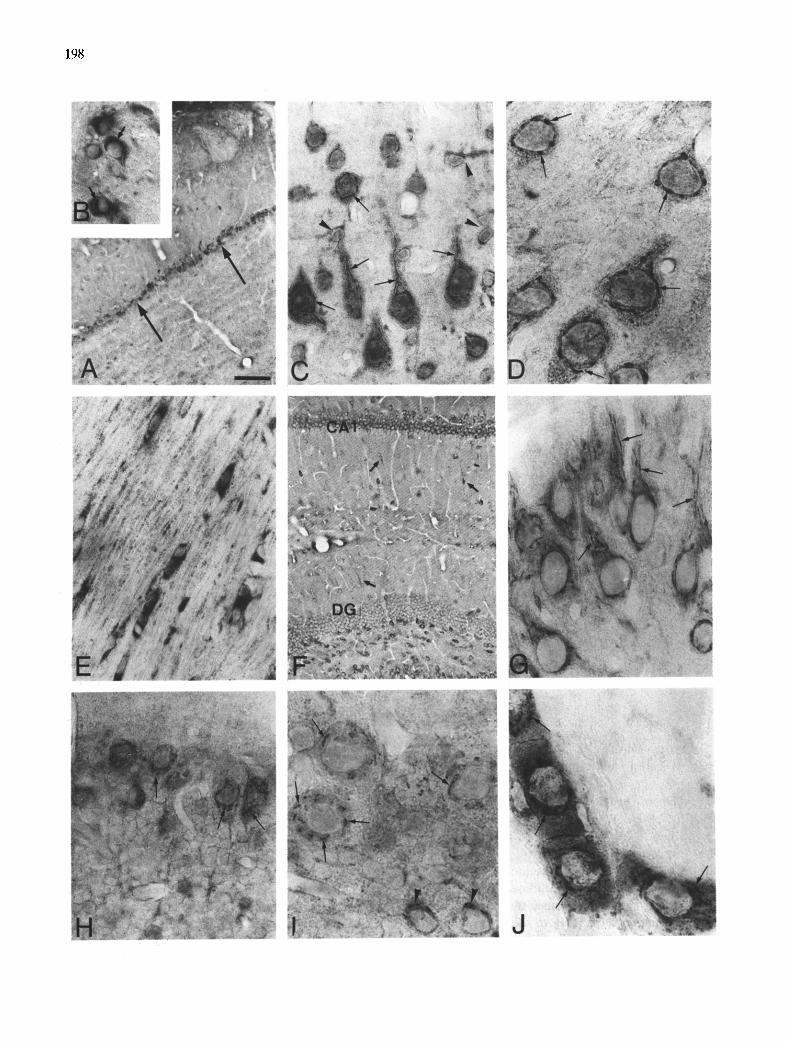
In the olfactory bulb, the most intense immunoreac- tivity was found in the mitral cell layer. These neurons presented a strong immunostained filamentous perinu- clear reticulum corresponding to the Golgi apparatus protruding over a weak and diffuse cytoplasmic im- munoreactivity. Other cells in the granular layer and periglomerular region showed a restricted punctate
197
immunoreactive structure in their cytoplasm (Fig. 2A,B).
In the neocortex the same pattern of immunoreac- tivity could be seen in the Golgi apparatus of the pyramidal neurons. These immunostained neurons were localized in all layers of the cerebral cortex (Fig. 2C,D). Fibrous astrocytes were also identified in the cortex (Fig. 2C) and specially in subpial regions. Inter- fascicular immunoreactive positive oligodendrocytes were observed forming rows between the immuno- stained fascicles of myelinated fibers in the corpus callosum. These oligodendrocytes showed intense and homogeneous cytoplasmic immunoreactivity (Fig. 2E).
Immunoreactive pyramidal neurons were also found in all regions of the hippocampus with immunostained perinuclear Golgi apparatus (Fig. 2F,G). Granule cells of the dentate gyrus exhibited a weak immunoreactivity (Fig. 2F). Also, weak immunoreactive astrocytes were scattered throughout the stratum oriens and radiatum of the hippocampus and the stratum molecular of the dentate gyrus (Fig. 2F).
In the cerebellum a dense immunoreactive Golgi apparatus in perinuclear disposition was observed in the Purkinje cells or in cells scattered in the molecular or granule layers such as basket and Golgi cells. The granule cells exhibited limited immunoreactivity (Fig. 2H,I).
In the nucleus basalis region, the neurons dis- tributed among the fiber bundles presented intense immunoreactivity with a prominent immunostained Golgi apparatus (Fig. 2J).
Cytochemical and immunocytochemical study after uni- lateral facial transectomy
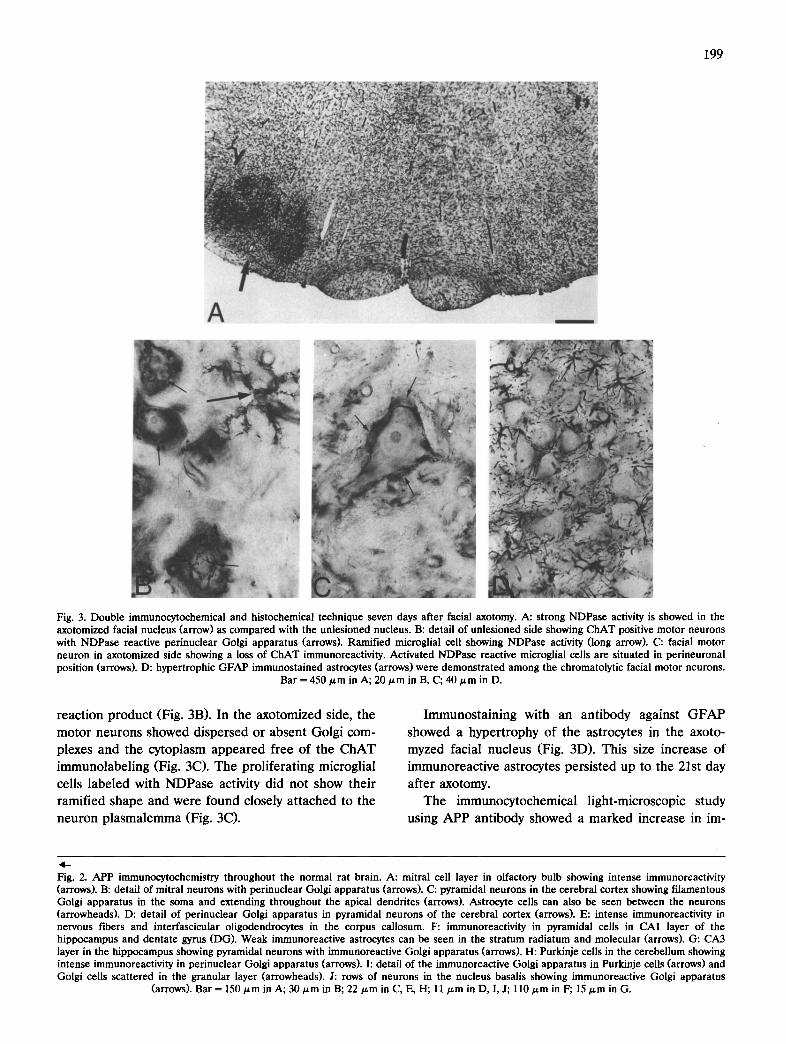
The application of a double immunocytochemical and histochemical technique using monoclonal anti- bodies raised against choline acetyltransferase (CHAT) and the nucleoside diphosphatase (NDPase) histo- chemical method, revealed significant changes follow- ing axotomy. An increase in NDPase activity and a decrease in the ChAT immunoreactivity in the axo- tomized facial nucleus were seen compared to the control side (Fig. 3A). These modifications started 3 days following axotomy and remained unchanged up to the 2P t day, the latest time-point studied. In the unle- sioned control side, motor neurons showed perinuclear complexes of the Golgi apparatus that were stained by the enzymatic reaction (Fig. 3B). These brown com- plexes were protruding over the homogeneous blue ChAT-immunoreactive cytoplasm (Fig. 3B). Microglial cells marked with NDPase activity and showing a typi- cal ramified morphology were found in the neuropil. The capillary wall was also labeled with the enzyme
198
199
Fig. 3. Double immunocytochemical and histochemical technique seven days after facial axotomy. A: strong NDPase activity is showed in the axotomized facial nucleus (arrow) as compared with the unlesioned nucleus. B: detail of unlesioned side showing ChAT positive motor neurons with NDPase reactive perinuclear Golgi apparatus (arrows). Ramified microglial cell showing NDPase activity (long arrow). C: facial motor neuron in axotomized side showing a loss of ChAT immunoreactivity. Activated NDPase reactive microglial cells are situated in perineuronai position (arrows). D: hypertrophic GFAP immunostained astrocytes (arrows) were demonstrated among the chromatolytic facial motor neurons.
Bar = 450/~m in A; 20/~m in B, C; 40/~m in D.
reaction product (Fig. 3B). In the axotomized side, the motor neurons showed dispersed or absent Golgi com- plexes and the cytoplasm appeared free of the ChAT immunolabeling (Fig. 3C). The proliferating microglial cells labeled with NDPase activity did not show their ramified shape and were found closely attached to the neuron plasmalemma (Fig. 3C).
Immunostaining with an antibody against GFAP showed a hypertrophy of the astrocytes in the axoto- myzed facial nucleus (Fig. 3D). This size increase of immunoreactive astrocytes persisted up to the 21st day after axotomy.
The immunocytochemical light-microscopic study using APP antibody showed a marked increase in im-
, D
Fig. 2. APP immunocytochemistry throughout the normal rat brain. A: mitral cell layer in olfactory bulb showing intense immunoreactivity (arrows). B: detail of mitral neurons with perinuclear Golgi apparatus (arrows). C: pyramidal neurons in the cerebral cortex showing filamentous Golgi apparatus in the soma and extending throughout the apical dendrites (arrows). Astrocyte cells can also be seen between the neurons (arrowheads). D: detail of perinuclear Golgi apparatus in pyramidal neurons of the cerebral cortex (arrows). E: intense immunoreactivity in nervous fibers and interfascicular oligodendrocytes in the corpus callosum. F: immunoreactivity in pyramidal cells in CA1 layer of the hippocampus and dentate gyrus (DG). Weak immunoreactive astrocytes can be seen in the stratum radiatum and molecular (arrows). G: CA3 layer in the hippocampus showing pyramidal neurons with immunoreactive Golgi apparatus (arrows). H: Purkinje cells in the cerebellum showing intense immunoreactivity in perinuclear Golgi apparatus (arrows). I: detail of the immunoreactive Golgi apparatus in Purkinje cells (arrows) and Golgi cells scattered in the granular layer (arrowheads). J: rows of neurons in the nucleus basalis showing immunoreactive Golgi apparatus
(arrows). Bar = 150/zm in A; 30/.~m in B; 22/~m in C, E, H; 11 p m in D, I, J; 110/~m in F; 15/.~m in G.
200
O

201
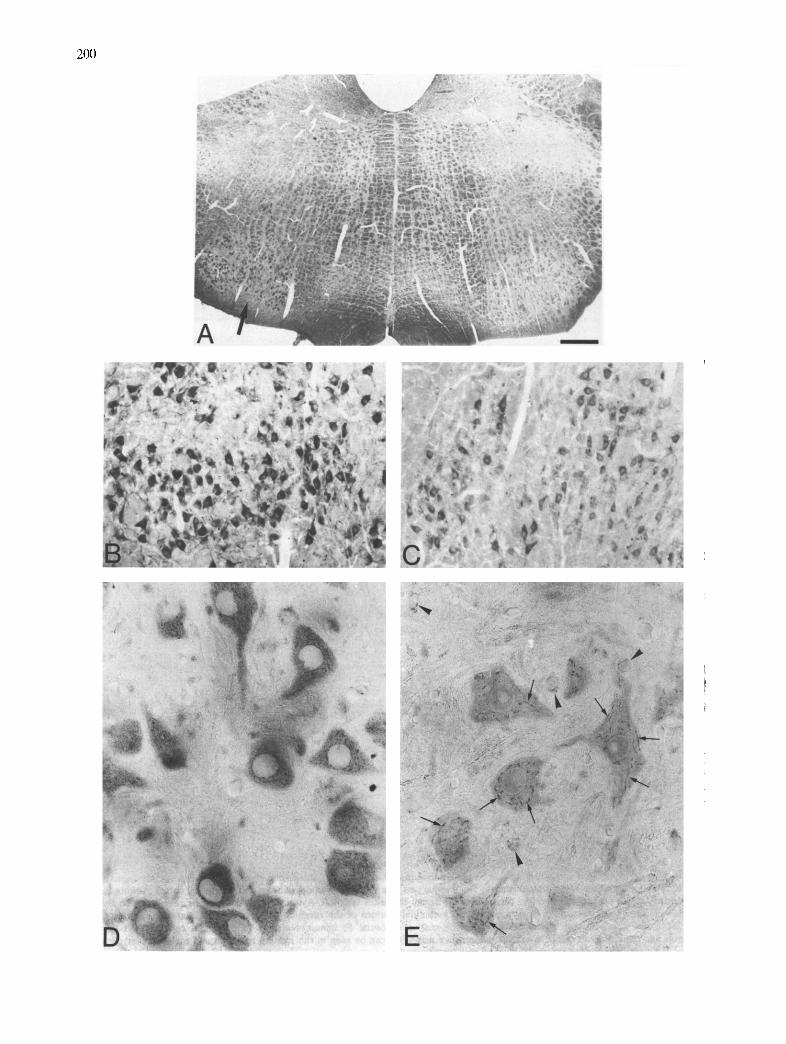
munostaining of the transectomized facial nucleus neu- rons (Fig. 4A-C). This increase was maintained throughout all experimental stages. A higher magnifi- cation of the facial immunostaining motor neurons in the unlesioned control side showed a filamentous per- inuclear Golgi apparatus with a strong APP immunore- activity (Fig, 4E). The morphological organization of the Golgi apparatus was similar to that exhibited by motor neurons incubated in the NDPase medium (Fig. 3B). The neuropil contained some round glial cells with an APP-immunoreactive punctate appearance in their cytoplasm (Fig. 4E). In the axotomyzed facial nucleus, the motor neurons showed an intense APP immunoreactivity and a 'rete dispersion' of the Golgi apparatus. Occasionally some immunostained round glial cells could be seen in the neuropil (Fig. 4D).
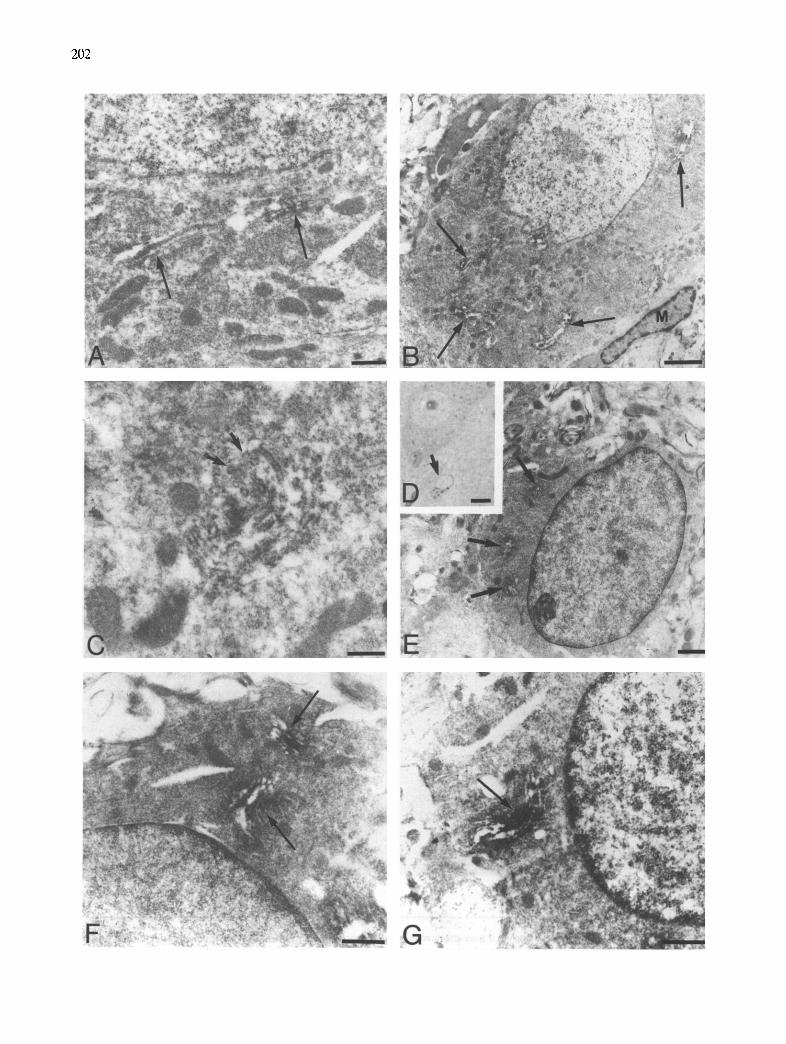
The electron microscopic study was mainly focused in the transectomized facial nucleus animals. In the control side, the motor neurons exhibited a developed Golgi apparatus arranged in the form of perinuclear complexes. APP immunoreactivity was usually located between the cisternae of all stacks of the complex and surrounding clear coated vesicles in the trans side of the Golgi complex (Fig. 5A). In the neuropil some glial ceils were identified with ultrastructural characteristics similar to oligodendrocytes. In semithin sections these cells showed a cytoplasmic punctate structure (Fig. 5D). The ultrastructural analysis demonstrated that this punctate structure corresponded to small Golgi complex (Fig. 5E). Each complex showed intense APP immunoreactivity localized between the short and wide cisternae throughout the stacks (Fig. 5E,F). No signifi- cant labeling of cellular organelles such as endoplasmic reticulum, lysosomes, mitochondria, etc., was observed. Plasma membranes were not labeled with the APP antibody.
In the axotomized side the chromatolytic motor neurons showed perineuronal microglial cells displac- ing axonal terminals from the surface of the plas- malemma (Fig. 5B). The Golgi apparatus of these neurons appeared as a dispersed complex in the cyto- plasm. These complexes showed intense labeling with the APP antibody between the cisternae and surround- ing vesicles located in the trans side (Fig. 5B,C). The oligodendroglial cells exhibited the same APP im- munoreactive Golgi apparatus as seen in the oligoden-
drocytes in the control side (Fig. 5G). No increase in the number of these cells was observed. In any case the antibody used revealed the presence of astrocytes or microglial cells in the facial nucleus.
D I S C U S S I O N
The major finding of the present immunocytochemi- cal study is that by using an antibody raised against the carboxy-terminal residues of the APP, the immunore- activity presented a wide distribution in neurons and oligodendroglial cells throughout the CNS. More re- markable is the fact that in both cell types APP im- munoreactivity was specially concentrated inside of the Golgi apparatus. Furthermore, this immunoreactivity was increased in facial axotomized motor neurons and persisted elevated for at least 3 weeks after axotomy.
Cellular and subcellular localization of APP immunore- activity
The regional and cellular distribution of APP C- terminal immunoreactivity observed in the present work is in good agreement with previous reports where other polyclonal or monoclonal antibodies were used, and also with in situ hybridization histochemistry studies. The localization of APP-immunoreactive neurons in rat olfactory bulb, cerebral cortex, hippocampus and cerebe l lum is cons is ten t with earl ier re- ports 1'5'15'19'27'30'40'41'45. Different patterns of immunola-
beling of glial cells have been reported with APP antibodies. The antibody used in the present study only labeled very weakly astrocytes in the hippocampus and cortex but not in other brain areas and nuclei. How- ever, in a light and electron microscopic study in rat by Card ans coworkers 5 using two antibodies against dif- ferent amino acid sequences located near the C- terminus of APP, they demonstrated that the im- munoreactivity was localized in the cytoplasm of the cortical neurons and astrocytes processes surrounding neurons or developing perivascular feet. The same antibodies used in the lesioned hippocampus after uni- lateral intracerebroventricular infusion of kainate, pro- duced an intense immunostaining in reactive astrocytes in ipsilateral hippocampus whereas in the contralateral side the APP immunoreactivity was exclusively local- ized within neuronal perikarya 41. Immunoelectron mi-
Fig. 4. APP immunocytochemistry in facial nucleus 7 days after axotomy. A: increase of immunostaining in the lesioned facial nucleus (arrow) as compared with the unlesioned side. B: strong immunoreaction in motor neurons in the axotomized side. C: normal immunostaining in unlesioned motor neurons. D: intense immunostaining in chromatolytic lesioned neurons. These neurons show a 'rete dispersion' of the Golgi apparatus. E: normal neurons in the control side showing a filamentous immunostained Golgi apparatus (arrows). Some punctate immunostained glial cells can
be seen scattered among the neurons (arrowheads). Bar = 450 g m in A; 110/zm in B, C; 22/~m in D, E.
202

203
croscopy applied in the former study confirmed that APP-immunostained cells are reactive astrocytes. Im- munocytochemical distribution of APP in both central and peripheral nervous tissue of mouse has been stud- ied using two different antisera against C- and N-re- sidues of A P P 19. Both antisera showed APP im- munoreactivity in neurons with a patchy appearance distributed in the cytoplasm and proximal dendrites or homogeneously distributed in the cytoplasm and cell processes of several types of glial cells specially astro- cytes, interfascicular oligodendrocytes and ependymal cells. In human central nervous system from normal and AD patients, immunocytochemical studies using polyclonal or monoclonal antibodies against parts of the APP showed the higher concentration of im- munoreactivity in neurons and glial cells of the neocor- tex, hippocampus, cerebellum and spinal cord 2'43. In- terestingly, in our study, even in the case of a reactive gliosis (axotomized facial nucleus, see bellow) we did not detect an increased astrocytic labeling.
An important result from the present study is the demonstration of an intense APP immunoreactivity inside of the Golgi complex in both neurons and oligo- dendrocytes. It has been hypothesized that APP is a membrane protein as deduced from the sequence of the APP gene 17. Previous studies have assigned APP to other subcellular compartments based on immunohis- tochemical or biochemical data. Such localizations in- cluded lysosomes and plasma membranes 2'4°. In a re- cent paper, Schubert and coworkers 37 have presented evidence that was interpreted as suggestive of presence of APP immunoreactivity in Golgi and endoplasmic reticulum structures. However, to our knowledge, our results provide the first direct evidence for the pres- ence of APP in the Golgi apparatus. This localization is not surprising since there is biochemical evidence indicating that APP is tyrosine-sulfated, and O- and N-glycosylated 3'48, postranscriptional steps which are known to occur in the Golgi apparatus 8.
Axotomized animals The morphological changes occurring after the sec-
tioning of the facial nerve have been well established previously 23 and were in agreement with the findings
of the present study. The nerve axotomy does not induce direct damage to motor neurons in the facial nucleus and a regenerating process starts after the lesion 12. The retrograde or chromatolytic changes ob- served in rat facial motor neurons corresponded to a swelling of the cell bodies and nuclear modifications. Significant changes may also involve the endomem- brane system, such as disorganization of Nissl bodies and dispersion of granular endoplasmic reticulum cis- ternae toward the periphery of the cell. In addition, a 'rete dispersion' of the Golgi apparatus has been fre- quently found in chromatolytic neurons 26. The retro- grade changes showed in motor neurons have been functionally related with the metabolic changes occur- ring in the regeneration process in these ceUs 23.
The double immunocytochemical and histochemical method used here allowed us to observe simultane- ously the activated microglial cells, the Golgi apparatus and ChAT modifications in motor neurons 33. NDPase activity has been proposed as a selective histochemical marker of the resting microglial plasma membrane 32'47. The disappearance of staining for CHAT, as seen in the present study, and for acetylcholinesterase (ACHE) have been reported following unilateral axotomy of the vagal and hypoglossal nerves ~. In the rat facial nucleus additional studies have demonstrated an AChE activity reduction one week after axotomy 7'22'23. On the other hand, a recovery of ChAT activity has been reported to occur 5 weeks after hypoglossal axotomy St. Concomi- tantly with the chromatolysis of motor neurons, mi- croglial and astrocytic changes were also seen in the present study and are in good agreement with other reports. Activated microglia involved in the process of 'synaptic stripping' and hypertrophic but not hyperpla- sic astrocytes with a strong increase in GFAP im- munoreactivity have been reported by other authors in the same experimental conditions 4'12-14,23.
The effects of axotomy in the immunocytochemical expression of APP in facial nucleus were also evalu- ated. Our results demonstrate a marked increase in immunoreactivity in the cytoplasm of the motor neu- rons in the operated side and a concomitant 'rete dispersion' of the immunostained Golgi apparatus in these neurons. These findings differ from in situ hy-
Fig. 5. APP immunoelectron micrographs in facial nucleus. A: control facial motor neuron showing an immunostained Golgi complex (arrows). B: chromatolytic motor neuron surrounded by a microglial cell (M) and showing dispersed immunostained Golgi complex (arrows). C: Golgi complex in an axotomized neuron showing APP immunoreactivity between the cisternae and surrounding vesicles localized in the trans side (arrows). D: light micrograph of neuron and oligodendrocyte in a semithin section of the control side. The oligodendrocyte shows an immunostained cytoplasmic punctate structure (arrow). E: punctate structure of the oligodendrocytes corresponding to immunostained small Golgi complex in an electron micrograph (arrows). F: detail of the oligodendrocyte Golgi complex showing APP immunoreactivity between the cisternae (arrows). (3: oligodendrocyte in the axotomized side showing the same immunostained pattern in a Golgi complex (arrow). Bar = 0.5
/,tm in A, C; 2/,~m in B; 7.5/~m in D; l ~ m in E; 0.6/~m in F, (3.

204
bridization studies reporting an increase on APP69 s m R N A levels in dorsal root ganglia neurons after crushing or transection of sciatic nerve in rats, while
APP695 m R N A levels decreased in spinal cord neu- rons 38. However, in situ hybridization studies carried
out in our laboratory (Solfi et al., in preparation)
demonstrate an increase in APP gene expression in transectomized facial nucleus. Thus, the increase in
APP immunoreactivity in these neurons can be ex-
plained by an increase in APP synthesis, although an accumulation due to a decreased release cannot be excluded. These results may also have a bearing on the possible functional role(s) of the APP proteins. The response to axotomy was a rapid and sustained stand- ing increase in the expression of the APP, in good
agreement with other experiments from our laboratory where parallel increases in APP mRNAs levels were observed, in the same model. The effect of axotomy in
different classes of proteins is well documented. While the synthesis of functional proteins such as neurotrans- mitter synthesizing enzymes (CHAT) or receptors is rapidly down regulated, structural proteins and others involved in neuronal regeneration rapidly increase as a consequence of axotomy 42. Thus, the behavior of APP
is similar to that of structural or growth-related pro-
teins. APP immunoreactivity in facial nucleus was also
localized in oligodendrocytes. In the facial nucleus, the APP immunopositive oligodendrocytes were weakly im- munostained when compared with the strongly labeled
interfascicular oligodendrocytes seen in the corpus cal- losum. The immunoreactivity was distributed in punc- rate cytoplasm structures corresponding to Golgi appa- ratus as recognized by electron microscopy. These oligodendrocytes were present in both the operated and control facial nucleus and did not show any mor- phological changes. Axotomy did not induce increase in either the number or the APP immunostaining of these oligodendrocytes. Interestingly, reactive mi-
croglia and hypertrophic astrocytes showed in axo- tomized facial nucleus were not labeled with the APP antibody. This finding is of considerable interest since it is inconsistent with the previous studies following neuronal damage and reactive gliosis 41. These authors, using a similar antiserum against the APP carboxy- terminal residues, reported APP immunoreactivity in astrocytes following experimental kainate-induced damage in the rat hippocampus. However, kainate-in- duced lesions produce neuronal degenerative changes and reactive gliosis which are different from the changes observed following facial axotomy. In kainate- induced lesions the glial response included the appear- ance of two types of phagocytic cells, microglia-like
reactive cells and brain macrophages, and hypertrophic and hyperplasic astrocytes 3~. In contrast, following axo- tomy the regenerating motor neurons were enclosed by
non-phagocytic activated microglia and by hypertrophic but not hyperplasic astrocytes 23. In both studies oligo-
dendrocytes did not reveal morphological changes. This
indicates that different types of lesions produce distinct kinds of responses of the same glial cell including expression of APP immunoreactivity.
In conclusion, by using an antiserum against the carboxy-terminal residues of APP, we have found that
immunoreactive neurons and oligodendroglial cells are widely distributed throughout the rat central nervous system. In these cells the highest level of immunoreac- tivity was restricted to the Golgi apparatus. Axotomy
resulted in an increase in the diffuse cytoplasmic APP immunoreactivity. These structural data further sup- port biochemical data 4s indicating that APP undergoes postranslational modifications which are known to oc-
cur in the Golgi apparatus. On the other hand, the constant presence of C-terminal fragments in the Golgi apparatus of all neurons and oligodendroglial cells suggested that this organelle is involved in the biosyn- thetic processing of APP and specially in the constitu- tive proteolytic cleavage that occurs during the assem- bly of these glycoproteins. Recently, mutations in the APP gene which could lead to faulty processing of proteins have been described 9'2s. Taken into account
the important role of the Golgi apparatus in the pro- cessing of these proteins it will be interesting to exam- ine APP processing in the Golgi apparatus in early phases of Alzheimer's disease.
REFERENCES
1 Bahmanyar, S., Higgins, G.A., Goldgaber, D., Lewis, D.A., Mor- rison, J.H., Wilson, M.C., Shankar, S.K. and Gaidusek, D.C., Localization of amyloid /3-protein messenger RNA in brains from patients with Alzheimer's disease, Science, 237 (1987) 77-80.
2 Benowitz, L., Rodriguez, W., Paskevich, P., Mufson, E., Schenk, D. and Neve, R.L., The amyloid precursor protein is concen- trated in neuronal lysosomes in normal and Alzheimer disease subjects, Exp. Neurol., 106 (1989) 237-250.
3 Beyreuther, K. and Masters, C.L., Amyloid genes and neuronal dysfunction, Brain Res. Re~'., 16 (1991) 86-88.
4 Blinzinger, K. and Kreutzberg, G.W., Displacement of synaptic terminals from regenerating motoneurons by microglial cells, Z. Zellforsch., 85 (1968) 145-157.
5 Card, J.P., Meade, R.P. and Davis, L.G., Immunocytochemical localization of the precursor protein for /3-amyloid in the rat central nervous system, Neuron, 1 (1988) 835-846.
6 De Sauvage, F. and Octave, J.N., A novel mRNA of the A4 amyloid precursor gene coding for a possibly secreted protein, Science, 245 (1989)651-653.
7 Engel, A.K. and Kreutzberg, G.W., Changes of acetyl- cholinesterase molecular forms in regenerating motor neurons, Neuroscience, 18 (1986) 467-473.
8 Farquhar, M.G. and Palade, G.E., The Golgi apparatus (com- plex) - - (1954-1981) - - from artifact to center stage, J. Cell BioL, 91 (1981) 77s-103s.
9 Goate, A., Chartier-Harlin, M-C. Mullan, M., Brown, J., Craw-

ford, F., Fidani, L., Giuffra, L., Haynes, A., Irving, N., James, L., Mant, R., Newton, P., Rooke, K., Roques, P., Talbot, C., Pericak-Vance, M., Roses, A., WiUiamson, R., Rossor, M., Owen, M. and Hardy, J., Segregation of missense mutation in the amyloid precursor protein gene with familial Alzheimer's disease, Nature, 349 ~991) 704-706.
10 Golde, T.E., Bstus, S., Usiak, M., Younkin, L.H. and Younkin, S.G., Expression of/3-myloid protein precursor mRNAs: recogni- tion of a novel alternatively spliced form and quantitation in Alzheimer's disease using PCR, Neuron, 4 (1990) 253-267.
11 Goldgaber, D., Lerman, M.I., Mc Bride, O.W., Saffiotti, V. and Gajdusek, D.C., Characterization and chromosomal localization of a cDNA encoding brain amyloid of Alzheimer's disease, Sci- ence, 235 (1987) 877-880.
12 Graeber, M.B. and Kreutzberg, G.W., Astrocytes increase in glial fibrillary acidic protein during retrograde changes of facial motor neurons, J. Neurocytol., 15 (1986) 363-373.
13 Graeber, M.B., Kreutzberg, G.W., Delayed astrocyte reaction following facial nerve axotomy, J. Neurocytol., 17 (1988) 209-220.
14 Graeber, M.B., Tetzlaff, W., Streit. W.J. and Kreutzberg, G.W., Microglial cells but not astrocytes undergo mitosis following rat facial nerve axotomy, Neurosci. Lett., 85 (1988) 317-321.
15 Higgins, G.A., Lewis, D.A., Bahmanyar, S., Goldgaber, D. C., Gajdusek, C., Young, W.G., Morrison, J.H. and Wilson, M.D., Differential regulation of amyloid-/3-protein mRNA expression within hippocampal neuronal subpopulations in Alzheimer's dis- ease, Proc. Natl. Acad. Sci. USA, 85 (1988) 1297-1301.
16 Hsu, S.M., Raine, L. and Fanger, H., The use of avidin-biotin- peroxidase complex (ABC) in immunoperoxidase techniques: a comparison between ABC and unlabeled antibody (peroxidase) procedures, J. Histochem. Cytochem., 29 (1981) 577-590.
17 Kang, J., Lemaire, H-G., Unterbeck, A., Salbaum, J.M., Masters C.L., Grzeschik, K-H., Multhaup, G., Beyreuther, K. and Muller- Hill, B., The precursor of Alzheimer's disease amyloid A4 protein resembles a cell-surface receptor, Nature, 325 (1987) 733-736.
18 Kang, J. and Miiller-Hill, B., Differential splicing of Alzheimer's disease amyloid A4 precursor RNA in rat tissues: pre A4695 mRNA is predominantly produced in rat and human brain, Biochem. Biophys. Res. Commun., 166 (1990) 1192-1200.
19 Kawarabayashi, T., Shoji, M., Harigaya, Y., Yamaguchi, H. and Hirai, S., Amyloid f l /A4 protein precursor is widely distributed in both the central and peripheral nervous systems of the mouse, Brain Res., 552 (1991) 1-7.
20 Kitaguchi, N., Takahashi, Y., Tokushima, Y., Shiojiri, S. and Ito, H., Novel precursor of Alzheimer's disease amyloid protein shows protease inhibitor activity, Nature, 331 (1988) 530-532.
21 Kreutzbcrg, G.W. and Barron, K.D., 5'-Nucleotidase of mi- croglial cells in the facial nucleus during axonal reaction, J. Neurocytol., 7 (1978) 601-610.
22 Kreutzberg, G.W., Tetzlaff, W. and Toth, L., Cytochemical changes of cholinesterases in motor neurons during regeneration. In M. Brzin, E.A. Barnard and D. Sket (Eds.), Cholinesterases Fundamental and Applied Aspects, de Gruyter, Berlin, 1984, pp. 273-288.
23 Kreutzberg, G.W., Graeber, M.B. and Streit, W.J., Neuron-glial relationship during regeneration of motor neurons, Metabol. Brain Dis., 4 (1989) 81-85.
24 Lams, B.E., Isacson, O. and Sofroniew, M.V., Loss of transmit- ter-associated enzyme staining following axotomy does not indi- cate death of brainstem cholinergic neurons, Brain Res., 475 (1988) 401-406.
25 Levy, E., Carman, M.D. Fernandez-Madrid, I.J., Power, M.D.,Lieberburg, I., Van Duinen, S.G., Bots, G.T.A.M., Luyendijk, W. and Frangione, B., Mutation of the Alzheimer's disease amyloid gene in hereditary cerebral hemorrhage, Dutch type, Science, 249 (1990) 1124-1126.
26 Lieberman, A.R., The axon reaction, Int. Rev. Neurobiol., 14 (1971) 49-124.
27 Manning, R.W., Reid, C.M., Lampe, R.A. and Davis, L.G., Identification in rodents and other species of an mRNA homolo- gous to the human fl-amyloid precursor, Mol. Brain Res., 3 (1988) 293-297.
205
28 Masters, C.L., Multhaup, G., Simms, G., Pottigiesser, J., Martins, R.N. and Beyreuther, K., Neuronal origin of a cerebral amyloid: neurofibrillary tangles of Alzheimers's disease contain the same protein as the amyloid of plaque cores and blood vessels, EMBO J., 4 (1985) 2757-2763.
29 Mauro, A., Germano, I., Giaccone, G., Giordana, M.T. and Schiffer, D., 1-Naphtol basic dye (1-NBD). An alternative to diaminobenzidine (DAB) in immunoperoxidase techniques, His- tochemistry, 83 (1985) 97-102.
30 Mita, S., Schon, E.A. and Herbert, J., Widespread expression of amyloid beta-protein precursor gene in rat brain, Am. J. Pathol., 134 (1989) 1253-1261.
31 Murabe, Y., Ibata, I. and Sano, Y., Morphological studies on neuroglia II Response of glial cells to kainic acid-induced lesions, Cell Tissue Res., 216 (1981) 569-580.
32 Murabe, Y. and Sano, Y., Morphological studies on neuroglia, V., Microglial cells in the cerebral cortex of the rat, with special reference to their possible involvement in synaptic function, Cell Tissue Res., 223 (1982) 493-506.
33 Palacios, G., A double immunocytochemical and histochemical technique for demonstration of cholinergic neurons and mi- croglial cells in basal forebrain and neostriatum of the rat, Neurosci. Lett., 115 (1990) 13-18.
34 Ponte, P., Gonzalez-De Whitt, P., Schilling, J., Miller, J.,Hsu, D., Greenberg, B., Davis, K., Wallace, W., Lieberburg, I., Fuller, F. and Cordell, B., A new A4 amyloid mRNA contains a domain homologous to serine proteinase inhibitors, Nature, 331 (1988) 525-527.
35 Robakis, N.K., Ramakrishna, N., Wolfe, G., and Wisniewski, H.M. Molecular cloning and characterization of a cDNA encod- ing the cerebrovascular and the neuritic plaque amyloid peptides, Proc. NatL Acad. Sci. USA, 84 (1987) 4190-4194.
36 SchS.gger, H. and von Jagow, G., Tricine-sodium dodecyl sulfate- polyacrylamide gel electrophoresis for the separation of proteins in the range of 1-100 kDa, Anal Biochem., 166 (1987) 368-379.
37 Schubert, W., Prior, R., Weideman, A., Dircksen, H., Multhap, G., Masters, C. and Beyreuther, K., Localization of Alzheimer /3A4 amyloid precursor protein at central and peripheral synaptic sites, Brain Res., 563 (1991) 184-194.
38 Scott, J.N., Parhad, I.M. and Clark, A.W., /3-Amyloid precursor protein gene is differentially expressed in axotomized sensory and motor systems, Mol. Brain Res., 10 (1991) 315-325.
39 Selkoe, D.J., The molecular pathology of Alzheimer's disease, Neuron, 6 (1991) 487-498.
40 Shivers, B.D., Hilbich, C., Multhaup, G., Salbaum, M., Beyreuther, K. and Seeburg, P.H., Alzheimer's disease amyloido- genic glycoprotein: expression pattern in rat brain suggests a role in cell contact, EMBO J., 7 (1988) 1365-1370.
41 Siman, R., Card, J.P., Nelson, R.B. and Davis, L.G., Expression of /3-amyloid precursor protein in reactive astrocytes following neuronal damage, Neuron, 3 (1989) 275-285.
42 Smith, C.B., Crane, A.M., Kadekaro, M., Agranoff, B.W., and Sokoloff, L. Stimulation of protein synthesis and glucose utiliza- tion in the hypoglossal nucleus induced by axotomy, J. Neurosci., 4 (1984) 2489-2496.
43 Stern, R.A., Otvos, L., Trojanowski, J.Q. and Lee, V.M.-Y., Monoclonal antibodies to a synthetic peptide homologous with the first 28 aminoacids of Alzheimer's disease /3-protein recog- nize amyloid and diverse glial and neuronal cell types in the central nervous system, Am. Z Pathol., 134 (1989) 973-978.
44 Sternberger, L.A., Hardy, P.H., Curculis, J.J. and Meyer, H.G., The unlabeled antibody enzyme method of immunocytochem- istry. Preparation and properties of soluble antigen-antibody complex (horseradish pcroxidase) and its use in identification of spirochetes, J. Histochem. Cytochem., 18 (1970) 315-333.
45 Tanzi, R.E., Gusella, J.F., Watkins, P.C., Bruns, G.A., St. George-Hyslop, P., Van Keuren, M.L., Patterson, D., Pagan, S., Kurmt, D.M. and Neve, R.L., Amyloid beta protein gene: cDNA, mRNA distribution, and genetic linkage near the Alzheimer locus, Science, 235 (1987) 880-884.
46 Tanzi, R.E., McClatchey, A.I., Lamperti, E.D., Villa-Romaroff, L., Gusella, J.F. and Neve, R.L., Protease inhibitor domain

206
encoded by an amyloid protein precursor mRNA associated with Alzheimer's disease, Nature, 331 (1988) 528-530.
47 Vorbrodt, A.W. and Wisniewski, H.M., Plasmalemma bound nu- cleoside diphosphatase as a cytochemical marker of central ner- vous system (CNS) mesodermal cells, J. Histochem. Cytochem., 30 (1982) 418-424.
48 Weidemann, A., K6ning, G., Bunke, D., Fischer, P., Salbaum,J.M., Masters, C.L. and Beyreuther, K., Identification, biogenesis and localization of precursors of Alzheimer's disease A4 amyloid protein, Cell, 57 (1989) 115-126.
49 Wisniewski, H.M., Weigel, J., Wang, K.C., Kujawa, M. and Lach, B., Ultrastructural studies of the cells forming amyloid fibers in classical plaques, Can. J. Neurol. Sci., 16 (1989) 535-542.
50 Wisniewski, H.M., Vorbrodt, A.W., Wegiel, J., Morys, J. and Lossinsky, A.S., Ultrastructure of the cells forming amyloid fibers
in Alzheimer disease and scrapie, Am. J. Med. Genet., Suppl., 7 (1990) 287-297.
51 Wooten, G.F., Park, D.H., Joh, T.H. and Reis, D.J., Immuno- chemical demonstration of reversible reduction in choline acetyl- transferase concentration in rat hypoglossal nucleus after hy- poglossal nerve transection, Nature, 275 (1978) 324-325.
52 Yamada, T., Sasaki, H., Furuya, H., Muyata, T., Goto, M.I. and Sakaki, Y., Complementary DNA for the mouse homolog of the human amyloid beta protein precursor, Biochem. Biophys. Res. Commun., 149 (1987) 665-671.
53 Yamada, T., Sasaki, H., Dohura, K., Goto, M.I. and Sakaki, Y., Structure and expression of the alternatively-sppliced forms of mRNA for the mouse homolog of Alzheimer's disease amyloid beta protein precursor, Biochem. Biophys. Res. Commun., 158 (1989) 906-912.
Copyright © 2022 FDOKUMEN




















