
Alterations of ubiquitin ligases in human cancer and their association with the natural history of...
10
ORIGINAL ARTICLE Alterations of ubiquitin ligases in human cancer and their association with the natural history of the tumor S Confalonieri 1 , M Quarto 1 , G Goisis 2 , P Nuciforo 1 , M Donzelli 2 , G Jodice 1 , G Pelosi 2,3 , G Viale 2,3 , S Pece 1,2,3 and PP Di Fiore 1,2,3 1 IFOM, Fondazione Istituto FIRC di Oncologia Molecolare, Milan, Italy; 2 Istituto Europeo di Oncologia, Milan, Italy and 3 Dipartimento di Medicina, Chirurgia ed Odontoiatria, Universita’ degli Studi di Milano, Milan, Italy Protein ubiquitination is critical for many cellular processes, through its ability to regulate protein degradation and various signaling mechanisms. In the ubiquitin (Ub) system, substrate specificity is achieved through the E3 family of Ub ligases. Because alterations of the ubiquitination machinery have been reported in human cancers, the selective inter- ference with Ub ligases might represent a powerful therapeutic tool. Here, we report the first wide survey of misregulation of Ub ligases in cancer. We analysed 82 Ub ligases in nine types of cancer by in situ hybridization on tissue microarrays. We found 27 instances in which an Ub ligase was altered in a given type of tumor, when compared with normal tissues: 21 cases of overexpression and 6 cases of underexpression. We further analysed selected Ub ligases in large cohorts of breast and non-small-cell lung carcino- mas. In five, of six, of these extended analyses (HUWE1, CCNB1IP1, SIAH1 and SIAH2 in breast cancer and CCNB1IP1 in lung cancer), we found that the levels of Ub ligases correlated significantly with relevant prognostic factors, and with clinical outcome. Our findings show that the alteration of Ub ligases is a frequent event in cancer and identify candidate targets for molecular therapies. Oncogene (2009) 28, 2959–2968; doi:10.1038/onc.2009.156; published online 22 June 2009 Keywords: ubiquitin; ubiquitin E3 ligase; cancer; in situ hybridization; tissue microarrays Introduction Protein ubiquitination is a versatile post-translational modification that regulates numerous aspects of cell physiology. The best-characterized function of ubiquitin (Ub) is to mediate the targeting of proteins to proteasomal degradation. This is achieved through the addition of poly-Ub chains (branching from the K48 of Ub) to target proteins (Hershko and Ciechanover, 1998). In recent years, however, it has become clear that other forms of ubiquitination, mono-ubiquitination or poly-ubiquitination through non-K48 chains, are involved in many signaling processes, thus defining a vast series of non-degradative functions of Ub (Sigismund et al., 2004; Mukhopadhyay and Riezman, 2007). Not surprisingly, therefore, defects in Ub-depen- dent proteolysis and Ub-dependent signaling have been implicated in several human diseases, including cancer, neurodegenerative disease and metabolic disorders (Ciechanover and Schwartz, 2002; Ciechanover, 2003). In cancer, the subversion of the Ub system might, in principle, cause misregulation of signaling pathways, or altered degradation of critical cell-cycle regulators. For instance, the malfunction of the Ub–proteasome path- way could either enhance the effect of oncoproteins or reduce the amount of tumor suppressor proteins. Indeed, the Ub system targets several positive growth regulators, for example, N-Myc, c-Myc, c-Fos, c-Jun and Src-like proteins, to proteasomal degradation (Kim et al., 2004; Nakayama and Nakayama, 2006; Weiss et al., 2007; Xia et al., 2007; Zhao et al., 2008). Similarly, enhanced Ub–proteasome-dependent degradation of tumor suppressor proteins, such as TP53, p27 or for mutated NF2, has also been implicated in the pathogen- esis of malignancies (Gautreau et al., 2002; Sun, 2006). Thus, the ubiquitination machinery is considered a promising therapeutic target for cancer, particularly because its components are enzymes that should, in principle, be druggable. A proof of principle for this approach is provided by the efficacy of proteasome inhibitors in some types of cancer. For instance, Velcade (Millennium Pharmaceuticals, Inc., Cambridge, MA, USA) has recently been approved for the treatment of multiple myeloma and mantle cell lymphoma (Orlowski and Kuhn, 2008). However, proteasome inhibition is limited to targeting only one function of the Ub system, namely protein degradation, thus failing to target the signaling functions of the system, which might be as relevant to tumorigenesis as its degradative functions. Moreover, given the wide homeostatic significance of Ub-based degradation, its global inhibition is bound to be plagued with undesired toxicity. Thus, strategies to identify specific targets within the Ub system are critical. The complex molecular nature of the process of ubiquitination might offer keys to enter into the issue of Received 15 October 2008; revised 13 April 2009; accepted 11 May 2009; published online 22 June 2009 Correspondence: Dr S Confalonieri or Professor PP Di Fiore, IFOM Fondazione Istituto FIRC di Oncologia Molecolare, Via Adamello 16, Milan 20139, Italy. E-mails: [email protected] or pierpaolo.difi[email protected] Oncogene (2009) 28, 2959–2968 & 2009 Macmillan Publishers Limited All rights reserved 0950-9232/09 $32.00 www.nature.com/onc
-
Upload
ifom-ieo-campus -
Category
Documents
-
view
0 -
download
0
Transcript of Alterations of ubiquitin ligases in human cancer and their association with the natural history of...

ORIGINAL ARTICLE
Alterations of ubiquitin ligases in human cancer and their association
with the natural history of the tumor
S Confalonieri1, M Quarto1, G Goisis2, P Nuciforo1, M Donzelli2, G Jodice1, G Pelosi2,3,G Viale2,3, S Pece1,2,3 and PP Di Fiore1,2,3
1IFOM, Fondazione Istituto FIRC di Oncologia Molecolare, Milan, Italy; 2Istituto Europeo di Oncologia, Milan, Italy and3Dipartimento di Medicina, Chirurgia ed Odontoiatria, Universita’ degli Studi di Milano, Milan, Italy
Protein ubiquitination is critical for many cellular processes,through its ability to regulate protein degradation andvarious signaling mechanisms. In the ubiquitin (Ub) system,substrate specificity is achieved through the E3 family of Ubligases. Because alterations of the ubiquitination machineryhave been reported in human cancers, the selective inter-ference with Ub ligases might represent a powerfultherapeutic tool. Here, we report the first wide survey ofmisregulation of Ub ligases in cancer. We analysed 82 Ubligases in nine types of cancer by in situ hybridization ontissue microarrays. We found 27 instances in which an Ubligase was altered in a given type of tumor, when comparedwith normal tissues: 21 cases of overexpression and 6 casesof underexpression. We further analysed selected Ub ligasesin large cohorts of breast and non-small-cell lung carcino-mas. In five, of six, of these extended analyses (HUWE1,CCNB1IP1, SIAH1 and SIAH2 in breast cancer andCCNB1IP1 in lung cancer), we found that the levels of Ubligases correlated significantly with relevant prognosticfactors, and with clinical outcome. Our findings show thatthe alteration of Ub ligases is a frequent event in cancer andidentify candidate targets for molecular therapies.Oncogene (2009) 28, 2959–2968; doi:10.1038/onc.2009.156;published online 22 June 2009
Keywords: ubiquitin; ubiquitin E3 ligase; cancer; in situhybridization; tissue microarrays
Introduction
Protein ubiquitination is a versatile post-translationalmodification that regulates numerous aspects of cellphysiology. The best-characterized function of ubiquitin(Ub) is to mediate the targeting of proteins toproteasomal degradation. This is achieved through theaddition of poly-Ub chains (branching from the K48 ofUb) to target proteins (Hershko and Ciechanover,
1998). In recent years, however, it has become clearthat other forms of ubiquitination, mono-ubiquitinationor poly-ubiquitination through non-K48 chains, areinvolved in many signaling processes, thus defininga vast series of non-degradative functions of Ub(Sigismund et al., 2004; Mukhopadhyay and Riezman,2007). Not surprisingly, therefore, defects in Ub-depen-dent proteolysis and Ub-dependent signaling have beenimplicated in several human diseases, including cancer,neurodegenerative disease and metabolic disorders(Ciechanover and Schwartz, 2002; Ciechanover, 2003).
In cancer, the subversion of the Ub system might, inprinciple, cause misregulation of signaling pathways, oraltered degradation of critical cell-cycle regulators. Forinstance, the malfunction of the Ub–proteasome path-way could either enhance the effect of oncoproteins orreduce the amount of tumor suppressor proteins.Indeed, the Ub system targets several positive growthregulators, for example, N-Myc, c-Myc, c-Fos, c-Junand Src-like proteins, to proteasomal degradation (Kimet al., 2004; Nakayama and Nakayama, 2006; Weisset al., 2007; Xia et al., 2007; Zhao et al., 2008). Similarly,enhanced Ub–proteasome-dependent degradation oftumor suppressor proteins, such as TP53, p27 or formutated NF2, has also been implicated in the pathogen-esis of malignancies (Gautreau et al., 2002; Sun, 2006).Thus, the ubiquitination machinery is considered apromising therapeutic target for cancer, particularlybecause its components are enzymes that should, inprinciple, be druggable. A proof of principle for thisapproach is provided by the efficacy of proteasomeinhibitors in some types of cancer. For instance, Velcade(Millennium Pharmaceuticals, Inc., Cambridge, MA,USA) has recently been approved for the treatment ofmultiple myeloma and mantle cell lymphoma (Orlowskiand Kuhn, 2008). However, proteasome inhibition islimited to targeting only one function of the Ub system,namely protein degradation, thus failing to target thesignaling functions of the system, which might be asrelevant to tumorigenesis as its degradative functions.Moreover, given the wide homeostatic significance ofUb-based degradation, its global inhibition is bound tobe plagued with undesired toxicity. Thus, strategies toidentify specific targets within the Ub system are critical.
The complex molecular nature of the process ofubiquitination might offer keys to enter into the issue of
Received 15 October 2008; revised 13 April 2009; accepted 11 May 2009;published online 22 June 2009
Correspondence: Dr S Confalonieri or Professor PP Di Fiore, IFOMFondazione Istituto FIRC di Oncologia Molecolare, Via Adamello 16,Milan 20139, Italy.E-mails: [email protected] [email protected]
Oncogene (2009) 28, 2959–2968& 2009 Macmillan Publishers Limited All rights reserved 0950-9232/09 $32.00
www.nature.com/onc

specificity. The ubiquitination machinery is structured ina hierarchical cascade, in which three types of enzymes(E1, E2 and E3) act sequentially to promote the transferof Ub onto substrates (Hershko and Ciechanover,1998). In some cases, a multi-Ub chain assembly factor(E4) is also involved (Hoppe, 2005). Substrate specificityis achieved through the E3 Ub ligase family of proteins.Two major types of molecules execute this function:HECT-type E3 ligase possesses intrinsic Ub ligaseactivity and directly catalyses the transfer of Ub ontosubstrates. RING-type E3 ligase functions as specificadaptors, bridging together E2 (Ub-conjugating) en-zymes and their substrates, thereby allowing E2 tocatalyse the Ub transfer. There are more than 100known E3s. On the one hand, this projects thepossibility of specific therapeutic intervention, throughinhibition of individual E3s. On the other, with referenceto cancer, it poses the problem of compiling a preciseatlas describing which E3 is altered in which cancer, anissue that has been probed so far by one-molecule-at-a-time approaches, that is by analysing individual E3ligases in single types of cancer (see detailed referencelist in Supplementary information). In this study, wedescribe the first wide survey of misregulation of Ubligases in cancer.
Results
Study designA schematic of the study design is depicted in Figure 1a.We analysed a total of 82 Ub ligases (28 HECTs and48 RINGs, and 6 E4 enzymes, selected as described inMaterials and methods, listed and referenced inSupplementary Table 1). Since the beginning of ourscreening, other RING-E3s have been identified (Kalli-jarvi et al., 2005; Meroni and Diez-Roux, 2005;Buchwald et al., 2006; Schauvliege et al., 2006). Thus,our collection of Ub ligases is not exhaustive, althoughit contains the majority of enzymes known so far. Weused in situ hybridization (ISH) on tissue microarray(TMA) as the screening platform. This choice was basedon a number of reasons. First, a large-scale expressionstudy must necessarily be based on RNA detection,because the availability of suitable antibodies (especiallyfor in situ detection) constitutes a major limiting factorfor protein detection-based approaches. Second, amongRNA-based technologies, ISH/TMA couples the ad-vantages of a medium/high-throughput methodology(hundreds of genes can be screened on hundreds oftumors) with those of a high-resolution technology(each core can be analysed by visual inspection, therebyallowing the identification of the cellular origin of the
signal in a heterogeneous tissue). Third, ISH/TMA canbe carried out on archival paraffin-embedded samples,which is extremely advantageous for validation studieson large case collections for which patient follow-up isavailable. Fourth, we have already used ISH/TMA forlarge screening projects and showed that its resultscorrelate well with those obtainable with more quanti-tative methods, such as quantitative PCR (Nicassioet al., 2005; Capra et al., 2006; Vecchi et al., 2008).
The initial ISH/TMA screening led to the identifica-tion of 13 ligases that were misregulated in humancancers, for a total of 27 misregulation occurrences(Figure 1a). Selected ligases from this group weresubjected to further validation by analysing their abilityto predict disease outcome in two cohorts of breastcancer patients and in one cohort of non-small-cell lungcarcinomas (NSCLCs) (Figure 1a).
Alterations in the expression of Ub ligases in humancancersOur TMAs contained samples from nine types of cancer,including breast, colon-rectum, lung, prostate, larynx,stomach, uterus, kidney carcinomas and non-Hodgkinlymphomas. Together, these types of cancer account for65–70% of all cancer incidence, and for 60–65% of allcancer deaths in the United States (source NationalCancer Institute, http://seer.cancer.gov/statfacts/). Wescreened, on average, B200 tumors per gene, and aroughly equal number of normal tissues (frequently, andwhenever possible, from the same patient; details aregiven in Supplementary Table 2).
Of 82 analysed ligases, 13 (B16%) could be detected(ISH score X1, see Materials and methods) in at leastone of the analysed tissue types (Figure 1b andSupplementary Table 3; high-power views of represen-tative ISH are shown in Supplementary Figures 1–3).The remaining 69 (B84%) genes showed completelynegative or barely detectable signals in all the analysedtissues, likely due to low mRNA abundance. Theseresults are in accordance with previous reports showinglow mRNA expression of HECT-type E3s in humancells lines (Scarafia et al., 2000). It can be noted that inall cases in which antisense probes yielded positivesignals (13 genes), the corresponding sense probe, usedas a negative control, did not yield any appreciablesignal (data not show). In addition, we selectedthree representative ligases (HUWE1, SIAH2 andCCNB1IP1, for which we also carried out an extendedanalysis in breast and lung cancer, see below) andperformed RNA interference in cell lines. The interferedcells were then included in paraffin and subjected toISH, as described for the normal and tumor tissues. In
Figure 1 Misregulation of Ub ligases in human cancers. (a) A scheme of the study design is shown. Left (boxed in black), strategy andresults of the ISH/TMA screening; right (boxed in red), strategy of the various validations and of the prognostic analyses.(b) Misregulation of ligases in human cancers. The mean levels of expression in various human tumors (T) and matched normal tissues(N) are shown by a semiquantitative color code, reflecting the mean ISH scores. Actual scores (and P-values) are given inSupplementary Table 4. Asterisks mark statistically significant (Pp0.05) differences (black asterisk, upregulation; red asterisk,downregulation). (c) Examples of data summarized in panel b are shown for normal and tumor tissues. In each pair, the upper panel isa bright field (for morphological evaluation) and the lower panel is a dark field (transcripts appear as bright dots).
Ubiquitin E3 ligases deregulation in human cancerS Confalonieri et al
2960
Oncogene

HUWE1
LUNG
SYVN1
NHLBREAST
HUWE1CCNB1IP1 SIAH2 PRP19HUWE1
COLON
HUWE1 CBLC
PROSTATE
NO
RM
AL
TU
MO
R
Name T N T N T N T N T N T N T N T N T NAMFR *SYVN1 *RNF5 *HUWE1 * * * * * * *HERC1 * *MDM2 * * *SIAH1 *CCNB1IP1 *BRCA1 *CBLC *SIAH2 * * *RNF20 * *PRP19 * *
Breast Colon Lung Prostate Larynx NHL Stomach Uterus Kidney
0-0.09 0.10-0.50 0.51-1.00 1.01-1.50 1.51-2.00 > 2.00
Selected LigasesN = 82
ISH/TMA
13 misregulated ligases27 total misregulations
9 Types of tumors~ 400 specimen
Val
idat
ion
ofse
lect
ed li
gase
s
*
Validation - Breast Cancerconsecutive cohort
(N = 477)
Validation - Breast Cancercase-control cohort
(N = 444)
Correlation withdisease outcome
ISH scores
Validation - Lung Cancerconsecutive cohort
(N = 420)
Ubiquitin E3 ligases deregulation in human cancerS Confalonieri et al
2961
Oncogene

all three cases, the functional ablation of the E3 ligasecaused a marked decrease in the ISH signals vs themock-interfered cells, thus further confirming thespecificity of detection of the corresponding mRNAs(Supplementary Figure 4).
Alterations in the levels of expression of the 13detectable Ub ligases in human cancers were evaluatedthrough comparison of the average expression levelsbetween tumor cells and cells of putative origin of agiven tumor, when known, in the normal tissue counter-part (see Supplementary Methods, Figure 1b andSupplementary Table 4 for detailed information). All13 ligases were found to be significantly misregulated inat least one type of tumor. We detected 27 instancesin which one of the Ub ligases was misregulated in agiven type of tumor. Of these events, 21 representedoverexpression and 6 represented underexpression(Figure 1b, Supplementary Table 4 and SupplementaryFigures 1–3).
In conclusion, our expression survey showed mis-regulation of Ub ligases at a relatively high frequency inhuman cancers. In all cases (13) in which Ub ligasescould reliably be detected, they showed alterations in atleast one type of cancer. It can be noted that in all cases,the ISH signals of the overexpressed or underexpressedgenes were associated with the tumor cell componentand not with the adjacent or infiltrating stroma (seeFigure 1c for representative examples and Supple-mentary Table 4 for details).
Misregulation of HUWE1, CCNB1IP1, SIAH2 andSIAH1 predicts clinical outcome in breast cancerIf alterations in the expression of E3s have a role in thedevelopment or in the natural history of certain cancers,significant correlations with clinically relevant para-meters should be expected. As a proof-of-principlevalidation of this concept, we carried out an in-depthcharacterization of selected E3s in breast and lungcarcinomas, which together accounts for more thanone-third of all cancer deaths in the United States(source National Cancer Institute, http://seer.cancer.gov/statfacts/).
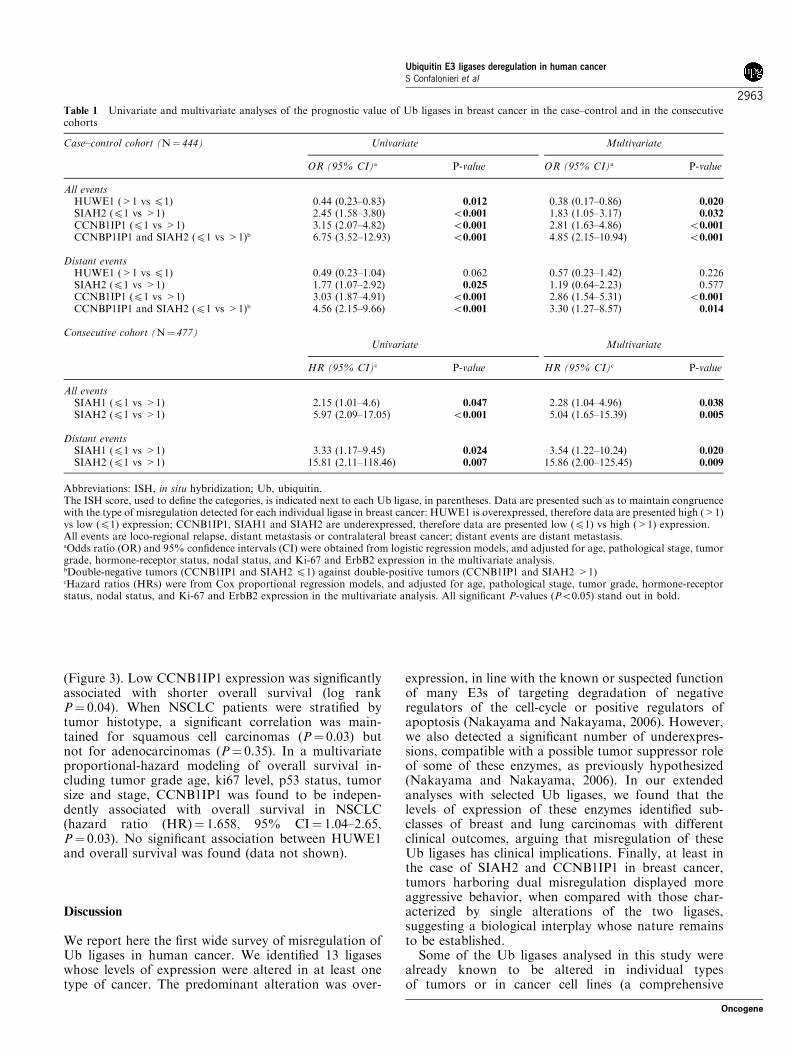
For breast studies, we initially selected three ligases:HUWE1 (overexpressed in breast cancer), and SIAH2and CCNB1IP1 (both underexpressed in breast cancer).Their expression was determined in a ‘case–control’cohort composed of 444 breast tumor samples (clinicaland pathological information is given in SupplementaryTable 5). In univariate analysis, low levels of expressionof each of the three ligases significantly correlated withhigher risks of disease recurrence (Table 1, see alsoSupplementary Table 6 for correlations between thelevels of expression of the three ligases and clinical andpathological parameters in the case–control cohort).In a multivariate analysis, in which all events wereevaluated in relation to relevant prognostic factors (age,pathological stage, tumor grade, hormone-receptorstatus, nodal status, Ki-67 and ErbB2 expression),SIAH2, HUWEI and CCNB1IP1 provided a significantindependent predictive power of a reduced disease
recurrence (Table 1). Because SIAH2 and CCNB1IP1were the most predictive genes, we investigated the effectof their coincident misregulation in the natural historyof breast cancer. As shown in Table 1, patients whosetumors displayed low expression of both genes had amuch higher risk of disease recurrence.
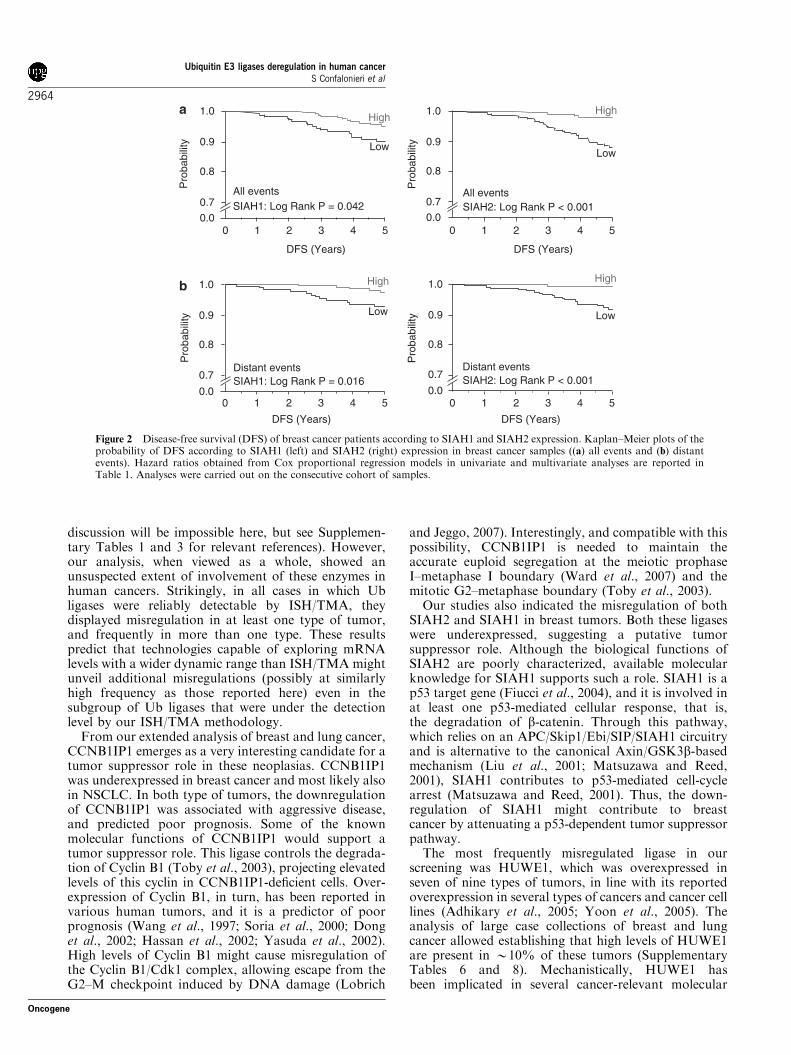
To further refine our analysis, and in order to have aprecise assessment of the probability of disease-freesurvival in time as a function of Ub ligase expression(something that cannot be assessed in a case–controlstudy), we tested the expression of SIAH2 on a secondcohort of patients (‘consecutive’ cohort), comprising 477consecutive pT1-2 patients (clinical and pathologicalinformation is given in Supplementary Table 5). We alsoscreened SIAH1 on this cohort of patients, becausethis Ub ligase—which belongs to the same familyof SIAH2—was underexpressed in breast cancer(Figure 1b). Low levels of expression of both SIAH1and SIAH2 correlated significantly with decreasedprobability of disease-free survival, both by the Kaplan–Meyer analysis (Figures 2a and b) and by univariate andmultivariate analysis (Table 1, see also SupplementaryTable 6 for correlations between the levels of expressionof the two ligases and clinical and pathologicalparameters in the consecutive cohort).
E3 ligases in lung cancerIn our initial screening, two E3s were found misregu-lated in lung cancers, HERC1 and HUWE1 (bothoverexpressed). The levels of expression of HERC1were, however, rather low, and thus this E3 wasexcluded from further analysis. In addition to HUWE1,we added CCNB1IP1 to our extended analysis, despitethe fact that this ligase was not apparently misregulatedin lung cancers in our initial survey (Figure 1b). We didso because, by closely inspecting the results of the initialscreening, we noticed that, in the normal lung,CCNB1IP1 was expressed in bronchial cells (mainly inthe basal layer) but not in alveolar cells. Conversely, inlung tumors, many tumors displayed no signal at all inany cytotype (data not shown). This finding, althoughhighlighting one of the advantages of using an in situtechnique for this kind of studies, prompted us tohypothesize that perhaps CCNB1IP1, similarly to whathappens in breast cancer, is underexpressed even in lungtumors and that tissue heterogeneity biased our initialassessment. Thus, we decided to further characterize thisligase.
A large cohort of consecutive NSCLCs (clinical andpathological features are given in SupplementaryTable 7) arrayed on TMAs was therefore characterizedfor the expression of HUWE1 and CCNB1IP1 (see alsoSupplementary Table 8 for correlations between thelevels of expression of the two ligases and clinical andpathological parameters in the NCSLC cohort). Five-year survival analysis was conducted on 357 patients forwhom follow-up was available (see SupplementaryMethods and Supplementary Table 7). We carried outKaplan–Meier estimates of overall survival as a functionof the levels of expression of CCNB1IP1 and HUWE1
Ubiquitin E3 ligases deregulation in human cancerS Confalonieri et al
2962
Oncogene
(Figure 3). Low CCNB1IP1 expression was significantlyassociated with shorter overall survival (log rankP¼ 0.04). When NSCLC patients were stratified bytumor histotype, a significant correlation was main-tained for squamous cell carcinomas (P¼ 0.03) butnot for adenocarcinomas (P¼ 0.35). In a multivariateproportional-hazard modeling of overall survival in-cluding tumor grade age, ki67 level, p53 status, tumorsize and stage, CCNB1IP1 was found to be indepen-dently associated with overall survival in NSCLC(hazard ratio (HR)¼ 1.658, 95% CI¼ 1.04–2.65,P¼ 0.03). No significant association between HUWE1and overall survival was found (data not shown).
Discussion
We report here the first wide survey of misregulation ofUb ligases in human cancer. We identified 13 ligaseswhose levels of expression were altered in at least onetype of cancer. The predominant alteration was over-
expression, in line with the known or suspected functionof many E3s of targeting degradation of negativeregulators of the cell-cycle or positive regulators ofapoptosis (Nakayama and Nakayama, 2006). However,we also detected a significant number of underexpres-sions, compatible with a possible tumor suppressor roleof some of these enzymes, as previously hypothesized(Nakayama and Nakayama, 2006). In our extendedanalyses with selected Ub ligases, we found that thelevels of expression of these enzymes identified sub-classes of breast and lung carcinomas with differentclinical outcomes, arguing that misregulation of theseUb ligases has clinical implications. Finally, at least inthe case of SIAH2 and CCNB1IP1 in breast cancer,tumors harboring dual misregulation displayed moreaggressive behavior, when compared with those char-acterized by single alterations of the two ligases,suggesting a biological interplay whose nature remainsto be established.
Some of the Ub ligases analysed in this study werealready known to be altered in individual typesof tumors or in cancer cell lines (a comprehensive
Table 1 Univariate and multivariate analyses of the prognostic value of Ub ligases in breast cancer in the case–control and in the consecutivecohorts
Case–control cohort (N¼ 444) Univariate Multivariate
OR (95% CI)a P-value OR (95% CI)a P-value
All eventsHUWE1 (>1 vs p1) 0.44 (0.23–0.83) 0.012 0.38 (0.17–0.86) 0.020
SIAH2 (p1 vs >1) 2.45 (1.58–3.80) o0.001 1.83 (1.05–3.17) 0.032
CCNB1IP1 (p1 vs >1) 3.15 (2.07–4.82) o0.001 2.81 (1.63–4.86) o0.001
CCNBP1IP1 and SIAH2 (p1 vs >1)b 6.75 (3.52–12.93) o0.001 4.85 (2.15–10.94) o0.001
Distant eventsHUWE1 (>1 vs p1) 0.49 (0.23–1.04) 0.062 0.57 (0.23–1.42) 0.226SIAH2 (p1 vs >1) 1.77 (1.07–2.92) 0.025 1.19 (0.64–2.23) 0.577CCNB1IP1 (p1 vs >1) 3.03 (1.87–4.91) o0.001 2.86 (1.54–5.31) o0.001
CCNBP1IP1 and SIAH2 (p1 vs >1)b 4.56 (2.15–9.66) o0.001 3.30 (1.27–8.57) 0.014
Consecutive cohort (N¼ 477)Univariate Multivariate
HR (95% CI)c P-value HR (95% CI)c P-value
All eventsSIAH1 (p1 vs >1) 2.15 (1.01–4.6) 0.047 2.28 (1.04–4.96) 0.038
SIAH2 (p1 vs >1) 5.97 (2.09–17.05) o0.001 5.04 (1.65–15.39) 0.005
Distant eventsSIAH1 (p1 vs >1) 3.33 (1.17–9.45) 0.024 3.54 (1.22–10.24) 0.020
SIAH2 (p1 vs >1) 15.81 (2.11–118.46) 0.007 15.86 (2.00–125.45) 0.009
Abbreviations: ISH, in situ hybridization; Ub, ubiquitin.The ISH score, used to define the categories, is indicated next to each Ub ligase, in parentheses. Data are presented such as to maintain congruencewith the type of misregulation detected for each individual ligase in breast cancer: HUWE1 is overexpressed, therefore data are presented high (>1)vs low (p1) expression; CCNB1IP1, SIAH1 and SIAH2 are underexpressed, therefore data are presented low (p1) vs high (>1) expression.All events are loco-regional relapse, distant metastasis or contralateral breast cancer; distant events are distant metastasis.aOdds ratio (OR) and 95% confidence intervals (CI) were obtained from logistic regression models, and adjusted for age, pathological stage, tumorgrade, hormone-receptor status, nodal status, and Ki-67 and ErbB2 expression in the multivariate analysis.bDouble-negative tumors (CCNB1IP1 and SIAH2 p1) against double-positive tumors (CCNB1IP1 and SIAH2 >1)cHazard ratios (HRs) were from Cox proportional regression models, and adjusted for age, pathological stage, tumor grade, hormone-receptorstatus, nodal status, and Ki-67 and ErbB2 expression in the multivariate analysis. All significant P-values (Po0.05) stand out in bold.
Ubiquitin E3 ligases deregulation in human cancerS Confalonieri et al
2963
Oncogene
discussion will be impossible here, but see Supplemen-tary Tables 1 and 3 for relevant references). However,our analysis, when viewed as a whole, showed anunsuspected extent of involvement of these enzymes inhuman cancers. Strikingly, in all cases in which Ubligases were reliably detectable by ISH/TMA, theydisplayed misregulation in at least one type of tumor,and frequently in more than one type. These resultspredict that technologies capable of exploring mRNAlevels with a wider dynamic range than ISH/TMA mightunveil additional misregulations (possibly at similarlyhigh frequency as those reported here) even in thesubgroup of Ub ligases that were under the detectionlevel by our ISH/TMA methodology.
From our extended analysis of breast and lung cancer,CCNB1IP1 emerges as a very interesting candidate for atumor suppressor role in these neoplasias. CCNB1IP1was underexpressed in breast cancer and most likely alsoin NSCLC. In both type of tumors, the downregulationof CCNB1IP1 was associated with aggressive disease,and predicted poor prognosis. Some of the knownmolecular functions of CCNB1IP1 would support atumor suppressor role. This ligase controls the degrada-tion of Cyclin B1 (Toby et al., 2003), projecting elevatedlevels of this cyclin in CCNB1IP1-deficient cells. Over-expression of Cyclin B1, in turn, has been reported invarious human tumors, and it is a predictor of poorprognosis (Wang et al., 1997; Soria et al., 2000; Donget al., 2002; Hassan et al., 2002; Yasuda et al., 2002).High levels of Cyclin B1 might cause misregulation ofthe Cyclin B1/Cdk1 complex, allowing escape from theG2–M checkpoint induced by DNA damage (Lobrich
and Jeggo, 2007). Interestingly, and compatible with thispossibility, CCNB1IP1 is needed to maintain theaccurate euploid segregation at the meiotic prophaseI–metaphase I boundary (Ward et al., 2007) and themitotic G2–metaphase boundary (Toby et al., 2003).
Our studies also indicated the misregulation of bothSIAH2 and SIAH1 in breast tumors. Both these ligaseswere underexpressed, suggesting a putative tumorsuppressor role. Although the biological functions ofSIAH2 are poorly characterized, available molecularknowledge for SIAH1 supports such a role. SIAH1 is ap53 target gene (Fiucci et al., 2004), and it is involved inat least one p53-mediated cellular response, that is,the degradation of b-catenin. Through this pathway,which relies on an APC/Skip1/Ebi/SIP/SIAH1 circuitryand is alternative to the canonical Axin/GSK3b-basedmechanism (Liu et al., 2001; Matsuzawa and Reed,2001), SIAH1 contributes to p53-mediated cell-cyclearrest (Matsuzawa and Reed, 2001). Thus, the down-regulation of SIAH1 might contribute to breastcancer by attenuating a p53-dependent tumor suppressorpathway.
The most frequently misregulated ligase in ourscreening was HUWE1, which was overexpressed inseven of nine types of tumors, in line with its reportedoverexpression in several types of cancers and cancer celllines (Adhikary et al., 2005; Yoon et al., 2005). Theanalysis of large case collections of breast and lungcancer allowed establishing that high levels of HUWE1are present in B10% of these tumors (SupplementaryTables 6 and 8). Mechanistically, HUWE1 hasbeen implicated in several cancer-relevant molecular
0.0
0.7
0.8
0.9
Pro
babi
lity
Pro
babi
lity
Pro
babi
lity
Pro
babi
lity
1.0
0 1 2 3 4 5
Low
High
DFS (Years)
SIAH1: Log Rank P = 0.042All events
Low
High
DFS (Years)
1 2 3 4 500.0
0.7
0.8
0.9
1.0
SIAH2: Log Rank P < 0.001
SIAH1: Log Rank P = 0.0160.0
0.7
0.8
0.9
1.0
0 1 2 3 4 5
DFS (Years)
Low
High
Distant eventsSIAH2: Log Rank P < 0.001
DFS (Years)
1 2 3 4 500.0
0.7
0.8
0.9
1.0
Low
High
All events
Distant events
Figure 2 Disease-free survival (DFS) of breast cancer patients according to SIAH1 and SIAH2 expression. Kaplan–Meier plots of theprobability of DFS according to SIAH1 (left) and SIAH2 (right) expression in breast cancer samples ((a) all events and (b) distantevents). Hazard ratios obtained from Cox proportional regression models in univariate and multivariate analyses are reported inTable 1. Analyses were carried out on the consecutive cohort of samples.
Ubiquitin E3 ligases deregulation in human cancerS Confalonieri et al
2964
Oncogene

circuitries that regulate: (i) p53 activity (Chen et al.,2005), (ii) p53-independent anti-proliferative pathways(Chen et al., 2005) and (iii) myc transcriptional activity(Adhikary et al., 2005). In principle, several of theputative functions of HUWE1 would be compatiblewith an oncogenic role and with its actual overexpres-
sion in tumors (Adhikary et al., 2005; Yoon et al., 2005;and this paper). It must be noted, however, that theprecise molecular functions of HUWE1 and theirbiological consequences are presently the matter ofdebate (Bernassola et al., 2008). For instance, both anti-apoptotic and pro-apoptotic effects of the functionalablation of HUWE1 have been reported (Chen et al.,2005; Zhong et al., 2005; reviewed in Bernassola et al.,2008). Thus, alternative scenarios are also worthconsidering.
One possibility, which would also be compatible withthe fact that HUWE1 is overexpressed in the greatestmajority of types of tumors, is that the misregulation ofHUWE1 is a consequence of transformation. HUWE1is involved in the degradation of CDC6 after DNAdamage, and it has been proposed to represent theprimary effector of CDC6 degradation in p53-null cells,where the canonical p53-mediated/APC-dependent me-chanism of CDC6 degradation is attenuated (Hall et al.,2007). CDC6 is required for licensing of replicationorigins before the onset of DNA replication ineukaryotic cells (Pelizon, 2003), and overexpression ofCDC6 has been observed in several tumor types(Borlado and Mendez, 2008; and reference therein),possibly contributing to genomic instability (Oehlmannet al., 2004; Lau et al., 2006). Thus, rather than being atumor-inducing event, HUWE1 overexpression mightbe a marker of p53 inactivation and CDC6 over-production in cancer cell, and its protective functionwould be to target the degradation of excess CDC6 inthe absence of a functional APC. Compatibly with thispossibility, in breast cancers, low levels of HUWE1correlate with a higher risk of recurrence and that thereis significant correlation between high levels of HUWE1and detectable p53 (a sign of mutated p53) (seeSupplementary Table 9).
Although the molecular scenarios depicted above willneed further experimental corroboration, our findingsprovide a broad atlas of the alteration of Ub ligases inhuman cancers, and point to several possible druggabletargets, thereby supporting the idea that specificinhibition of Ub ligases might prove effective in asizable fraction of human cancers. Moreover, we bringto light the prognostic value of some of these genes, thusopening the way for further studies focused on theirvalidation as cancer biomarkers.
Materials and methods
Identification and selection of Ub ligasesHECT-type E3 ligases were identified by profile-based searchesof protein databases and from literature. Initial domaininformation was collected from the SMART (http://smart.embl-heidelberg.de/) and the Pfam (http://pfam.janelia.org/)databases. The profile for the HECT domain was built usingthe HMM software starting from multiple alignment ofprotein sequences. This profile was used to search all thepublic protein databases using the HMMER software (http://hmmer.janelia.org/). After elimination of redundancies, weidentified 28 unique human proteins bearing an HECT domain(Supplementary Table 1).
0.0
0.1
0.2
0.3
0.4
0.5
0.6
0.7
0.8
0.9
1.0
10 20 30 40 50 60
Pro
babi
lity
Overall survival (Months)
Squamous cell carcinoma (SCC)
LogRank P=0.03
CCNB1IP1 Low
CCNB1IP1 High
0.0
0.1
0.2
0.3
0.4
0.5
0.6
0.7
0.8
0.9
1.0
10 20 30 40 50 60
Pro
babi
lity
Overall survival (Months)
Adenocarcinoma (ACC)LogRank P=0.35
CCNB1IP1 Low
CCNB1IP1 High
0.0
0.1
0.2
0.3
0.4
0.5
0.6
0.7
0.8
0.9
1.0
10 20 30 40 50 60
Pro
babi
lity
Overall survival (Months)
NSCLC ALL LogRank P=0.04
CCNB1IP1 Low
CCNB1IP1 High
Figure 3 Overall survival of lung cancer patients according toCCNB1IP1 expression. Kaplan–Meier plots of the 5-year overallsurvival (probability) are shown according to CCNB1IP1 expres-sion. Results are reported for 357 of 420 patients, because patientswho died after surgery within 5 months or with follow-up o5months were excluded from the analysis. Top panel: non-small-celllung carcinomas (NSCLC); middle panel: adenocarcinomas(ACC); bottom panel: squamous cell carcinomas (SSC).
Ubiquitin E3 ligases deregulation in human cancerS Confalonieri et al
2965
Oncogene

For RING-type E3s, several searches were carried out withdomain profile obtained by the SMART database. This,however, led to the identification of many ‘dubious positives’:essentially known transcription factors, displaying a RING-finger domain, for which no immediate Ub ligase functioncould be evinced from the literature. We also used severalpattern searches, using the Prosite database (http://www.expasy.ch/prosite/); in this case, patterns were derived fromliterature or directly from the Prosite database. Again, nosingle pattern could collect all known RING-type E3 ligase,and still several ‘dubious positive’ hits were selected. Thus, weturned to the literature and built a list of bona fide RING-typeligases, for which Ub ligase activity was experimentallyproved. The same procedure was used to select E4 enzymes.The complete list of selected proteins (and the relevantreferences) is reported in Supplementary Table 1.In total, we selected 82 Ub ligases (28 HECTs, 48 RINGs
and 6 E4s). We excluded some well-characterized E3s, such asthe multi-subunit SCF (Skip1/Cull/F-BOX protein) and APC/C (anaphase-promoting complex/cyclosome) E3s, becausetheir roles in cell-cycle control and human cancers have beenalready extensively characterized (Nakayama and Nakayama,2006).Representative cDNAs for each gene were retrieved from
our in-house Unigene EST clone collection (Unigene build no.106), or obtained from Invitrogen (http://clones.invitrogen.com/) or RZPD (http://www.rzpd.de/ now imaGenes http://www.imagenes-bio.de/). Multiple clones for each Ub ligasewere selected, sequence verified and used as templates to designhybridization probes for the ISH/TMA analysis. BLASTsearches were performed to identify the most specific 300 bpregion of each clone, and riboprobes were synthesized asdescribed previously (Capra et al., 2006).
Tissue samples and TMA design for the initial large-scalescreeningFor the large-scale ISH/TMA study, formalin-fixed andparaffin-embedded tumor and normal samples were providedby the Pathology Departments of Ospedale Maggiore(Novara), Presidio Ospedaliero (Vimercate), Ospedale SanPaolo (Milano) and Ospedale Sacco (Milano). Samples werearrayed in different TMAs (Supplementary Table 2), preparedessentially as previously described (Kononen et al., 1998).Briefly, two representative normal (when available) and tumorareas (diameter 0.6mm) from each sample, previouslyidentified on hematoxylin–eosin-stained sections, were re-moved from the donor blocks and deposited on the recipientblock using a custom-built precision instrument (TissueArrayer-Beecher Instruments, Sun Prairie, WI, USA). Sectionsof 2 mm thickness of the resulting recipient block were cut,mounted on glass slides and processed for ISH.
ISH/TMA analysis on large cohorts of breast and lung cancerpatientsFor the in-depth analyses of representative E3 ligases, we usedbreast and lung cancer samples from non-treated patients.Validation studies were performed on three different cohortsof samples.
Breast cancer case–control cohort. This cohort consisted ofpatients (median age 51, range 24–84 years) operated at theEuropean Institute of Oncology (Milan, Italy) for primarybreast cancer (enrollment period, 1994–1997). A total of 444women with breast cancer were selected according to thepathological characteristics of the primary tumor (nodalstatus) and upon follow-up information, in order to have a
representative number of event and event-free cases.Of these, 211 women who developed an event (loco-regionalrelapse, distant metastasis or contralateral breast cancer)within 7 years from the first breast cancer surgery constitutedthe cases, whereas 233 women who were alive and free of anyevent after 7 years of follow-up constituted the controls.Clinical and pathological characteristics are given in Supple-mentary Table 5.
Breast cancer consecutive cohort. This cohort was from asurgical trial conducted at the European Institute of Oncology(Milan, Italy) between March 1998 and December 1999(Veronesi et al., 2003) involving 447 consecutive patients(median age 55.8, range 37–75 years) with small-size primarybreast cancers (pT1 and pT2o3.0 cm in diameter) and a meanfollow-up period of 65 months. Clinical and pathologicalcharacteristics are given in Supplementary Table 5.
Lung cancer cohort. This cohort consisted of 420 NSCLCs,surgically treated at the European Institute of Oncology(Milan, Italy) between 1999 and 2002. Clinical and patholo-gical characteristics of this cohort of patients are given inSupplementary Table 7. Written informed consent for researchuse of biological samples was obtained from all patients, andthe research project was approved by the Institutional EthicalCommittee.
ISHIn situ hybridization was carried out as previously described(Capra et al., 2006). All TMAs were first analysed forthe expression of the housekeeping gene b-actin to check forthe mRNA quality of the samples. Cases showing absent orlow b-actin signal were excluded from the analysis (datanot shown). Gene expression levels were evaluated by countingthe number of grains per cell and were expressed in asemiquantitative scale (ISH score) as follows: 0 (no staining),1 (1–25 grains; weak staining), 2 (26–50 grains, moderatestaining) and 3 (>50 grains, strong staining) (Capra et al.,2006). Misregulation (upregulation or downregulation)was attributed when differences in mean expression levelsbetween the tumor and the normal groups were statisticallysignificant. To determine the clinico-pathological andsurvival correlations, samples were grouped into low-expres-sing (TMA score p1) or high-expressing (TMA score >1)tumors, based on the mean of the expression levelsof each gene between the two representative cores of the samesample.It should be noted that the number of scored cases, in some
experiments, is lower than the total number of arrayed cases.This is due to a number of reasons: (i) all cores that showedlow b-actin signal in the control hybridization were excludedfrom further consideration; (ii) in some cases, individual coresdetached from the slide during the manipulations. Forinstance, in Figure 2, SIAH1 and SIAH2 expression wasscored in 431 and 455 out of 477 samples, respectively; inFigure 3, CCNB1IP1 expression was scored in 307 out of 357samples.
Statistical analysisOverexpression or underexpression (Figure 1b and Supple-mentary Table 4) was determined using Student’s t-test. Theassociation between the clinico-pathological features of thetumors and gene expression was evaluated by Fisher’s exacttest. In the breast cancer case–control cohort, logisticregression was used to assess the association between geneexpression and the presence or absence of events after 7 years
Ubiquitin E3 ligases deregulation in human cancerS Confalonieri et al
2966
Oncogene

of follow-up. Multivariate analysis was used to assess theindependent prognostic value of gene expressions afteradjustment for recognized prognostic factors, as indicated. Inthe breast cancer consecutive cohort, disease-free interval wascalculated from the date of surgery to the date of the firstrecurrence or the date of last follow-up. Follow-up wascensored at 5 years. In the lung cancer cohort, overall survivalanalysis was carried out using 5 years censored follow-up.From the original cohort composed of 420 samples, patientswho died after surgery within 5 months and without or withfollow-upo5 months were excluded from the survival analysis(N¼ 63). The association between the clinico-pathologicalfeatures of the tumors and gene expression was evaluated byFisher’s exact test (Supplementary Table 8), consideringthe entire cohort of patients (N¼ 420). Survivalcurves according to gene expression were drawn using theKaplan–Meier method and compared by the log-rank test.Multivariate analysis was carried out by means of Cox’sproportional hazards model. Analyses were carried out usingSAS statistical software (SAS Institute Inc., Cary, NC, USA).All P-values were two-sided. A P-value p0.05 was consideredsignificant.
Abbreviations
Ub, ubiquitin; ISH, in situ hybridization; TMA, tissue micro-array.
Conflict of interest
The authors declare no conflict of interest.
Acknowledgements
We thank Maria Capra and Maria Tuttoilmondo for TMAdatabase development and TMA analysis, and Sara Voloriofor excellent technical assistance. This work was supported bygrants from AIRC (Associazione Italiana per la Ricerca sulCancro) to SC, and from AIRC and MIUR to SP and PPDF;from the European Community (VI Framework), the FerrariFoundation, the Monzino Foundation and the CARIPLOFoundation to PPDF; and from the G Vollaro Foundation toSP. PN was supported by an AIRC fellowship.
References
Adhikary S, Marinoni F, Hock A, Hulleman E, Popov N, Beier Ret al. (2005). The ubiquitin ligase HectH9 regulates transcriptionalactivation by Myc and is essential for tumor cell proliferation. Cell
123: 409–421.Bernassola F, Karin M, Ciechanover A, Melino G. (2008). The HECT
family of E3 ubiquitin ligases: multiple players in cancer develop-ment. Cancer Cell 14: 10–21.
Borlado LR, Mendez J. (2008). CDC6: from DNA replicationto cell cycle checkpoints and oncogenesis. Carcinogenesis 29:237–243.
Buchwald G, van der Stoop P, Weichenrieder O, Perrakis A, vanLohuizen M, Sixma TK. (2006). Structure and E3-ligase activity ofthe Ring–Ring complex of polycomb proteins Bmi1 and Ring1b.EMBO J 25: 2465–2474.
Capra M, Nuciforo PG, Confalonieri S, Quarto M, Bianchi M,Nebuloni M et al. (2006). Frequent alterations in the expressionof serine/threonine kinases in human cancers. Cancer Res 66:8147–8154.
Chen D, Kon N, Li M, Zhang W, Qin J, Gu W. (2005). ARF-BP1/Mule is a critical mediator of the ARF tumor suppressor. Cell 121:1071–1083.
Ciechanover A. (2003). The ubiquitin proteolytic system andpathogenesis of human diseases: a novel platform for mechanism-based drug targeting. Biochem Soc Trans 31: 474–481.
Ciechanover A, Schwartz AL. (2002). Ubiquitin-mediated degradationof cellular proteins in health and disease. Hepatology 35: 3–6.
Dong Y, Sui L, Watanabe Y, Sugimoto K, Tokuda M. (2002). Clinicalrelevance of cyclin B1 overexpression in laryngeal squamous cellcarcinoma. Cancer Lett 177: 13–19.
Fiucci G, Beaucourt S, Duflaut D, Lespagnol A, Stumptner-Cuvelette P, Geant A et al. (2004). Siah-1b is a direct transcriptionaltarget of p53: identification of the functional p53 responsiveelement in the siah-1b promoter. Proc Natl Acad Sci USA 101:3510–3515.
Gautreau A, Manent J, Fievet B, Louvard D, Giovannini M, Arpin M.(2002). Mutant products of the NF2 tumor suppressor gene aredegraded by the ubiquitin–proteasome pathway. J Biol Chem 277:31279–31282.
Hall JR, Kow E, Nevis KR, Lu CK, Luce KS, Zhong Q et al. (2007).Cdc6 stability is regulated by the Huwe1 ubiquitin ligase after DNAdamage. Mol Biol Cell 18: 3340–3350.
Hassan KA, Ang KK, El-Naggar AK, Story MD, Lee JI, Liu D et al.(2002). Cyclin B1 overexpression and resistance to radiotherapy inhead and neck squamous cell carcinoma. Cancer Res 62: 6414–6417.
Hershko A, Ciechanover A. (1998). The ubiquitin system. Annu Rev
Biochem 67: 425–479.Hoppe T. (2005). Multiubiquitylation by E4 enzymes: ‘one size’
doesn’t fit all. Trends Biochem Sci 30: 183–187.Kallijarvi J, Lahtinen U, Hamalainen R, Lipsanen-Nyman M,
Palvimo JJ, Lehesjoki AE. (2005). TRIM37 defective in mulibreynanism is a novel RING finger ubiquitin E3 ligase. Exp Cell Res
308: 146–155.Kim M, Tezuka T, Tanaka K, Yamamoto T. (2004). Cbl-c suppresses
v-Src-induced transformation through ubiquitin-dependent proteindegradation. Oncogene 23: 1645–1655.
Kononen J, Bubendorf L, Kallioniemi A, Barlund M, Schraml P,Leighton S et al. (1998). Tissue microarrays for high-throughputmolecular profiling of tumor specimens. Nat Med 4: 844–847.
Lau E, Zhu C, Abraham RT, Jiang W. (2006). The functional role ofCdc6 in S–G2/M in mammalian cells. EMBO R 7: 425–430.
Liu J, Stevens J, Rote CA, Yost HJ, Hu Y, Neufeld KL et al. (2001).Siah-1 mediates a novel beta-catenin degradation pathway linkingp53 to the adenomatous polyposis coli protein. Mol Cell 7: 927–936.
Lobrich M, Jeggo PA. (2007). The impact of a negligent G2/Mcheckpoint on genomic instability and cancer induction. Nat Rev
Cancer 7: 861–869.Matsuzawa SI, Reed JC. (2001). Siah-1, SIP, and Ebi collaborate in a
novel pathway for beta-catenin degradation linked to p53 responses.Mol Cell 7: 915–926.
Meroni G, Diez-Roux G. (2005). TRIM/RBCC, a novel class of ‘singleprotein RING finger’ E3 ubiquitin ligases. Bioessays 27: 1147–1157.
Mukhopadhyay D, Riezman H. (2007). Proteasome-independentfunctions of ubiquitin in endocytosis and signaling. Science 315:201–205.
Nakayama KI, Nakayama K. (2006). Ubiquitin ligases: cell-cyclecontrol and cancer. Nat Rev Cancer 6: 369–381.
Nicassio F, Bianchi F, Capra M, Vecchi M, Confalonieri S, Bianchi Met al. (2005). A cancer-specific transcriptional signature in humanneoplasia. J Clin Invest 115: 3015–3025.
Oehlmann M, Score AJ, Blow JJ. (2004). The role of Cdc6 in ensuringcomplete genome licensing and S phase checkpoint activation. J Cell
Biol 165: 181–190.
Ubiquitin E3 ligases deregulation in human cancerS Confalonieri et al
2967
Oncogene

Orlowski RZ, Kuhn DJ. (2008). Proteasome inhibitors incancer therapy: lessons from the first decade. Clin Cancer Res 14:1649–1657.
Pelizon C. (2003). Down to the origin: Cdc6 protein and thecompetence to replicate. Trends Cell Biol 13: 110–113.
Scarafia LE, Winter A, Swinney DC. (2000). Quantitative expressionanalysis of the cellular specificity of HECT-domain ubiquitin E3ligases. Physiol Genomics 4: 147–153.
Schauvliege R, Janssens S, Beyaert R. (2006). Pellino proteins are morethan scaffold proteins in TLR/IL-1R signalling: a role as novelRING E3-ubiquitin-ligases. FEBS Lett 580: 4697–4702.
Sigismund S, Polo S, Di Fiore PP. (2004). Signaling throughmonoubiquitination. Curr Top Microbiol Immunol 286: 149–185.
Soria JC, Jang SJ, Khuri FR, Hassan K, Liu D, Hong WK et al.(2000). Overexpression of cyclin B1 in early-stage non-small celllung cancer and its clinical implication. Cancer Res 60: 4000–4004.
Sun Y. (2006). E3 Ubiquitin ligases as cancer targets and biomarkers.Neoplasia 8: 645–654.
Toby GG, Gherraby W, Coleman TR, Golemis EA. (2003). A novelRING finger protein, human enhancer of invasion 10, alters mitoticprogression through regulation of cyclin B levels. Mol Cell Biol 23:2109–2122.
Vecchi M, Confalonieri S, Nuciforo P, Vigano MA, Capra M, BianchiM et al. (2008). Breast cancer metastases are molecularly distinctfrom their primary tumors. Oncogene 27: 2148–2158.
Veronesi U, Paganelli G, Viale G, Luini A, Zurrida S, Galimberti Vet al. (2003). A randomized comparison of sentinel-node biopsywith routine axillary dissection in breast cancer. N Engl J Med 349:546–553.
Wang A, Yoshimi N, Ino N, Tanaka T, Mori H. (1997). Over-expression of cyclin B1 in human colorectal cancers. J Cancer Res
Clin Oncol 123: 124–127.Ward JO, Reinholdt LG, Motley WW, Niswander LM, Deacon DC,
Griffin LB et al. (2007). Mutation in mouse hei10, an e3 ubiquitinligase, disrupts meiotic crossing over. PLoS Genet 3: e139.
Weiss A, Baumgartner M, Radziwill G, Dennler J, Moelling K.(2007). c-Src is a PDZ interaction partner and substrate of theE3 ubiquitin ligase Ligand-of-Numb protein X1. FEBS Lett 581:5131–5136.
Xia Y, Wang J, Xu S, Johnson GL, Hunter T, Lu Z. (2007). MEKK1mediates the ubiquitination and degradation of c-Jun in response toosmotic stress. Mol Cell Biol 27: 510–517.
Yasuda M, Takesue F, Inutsuka S, Honda M, Nozoe T, Korenaga D.(2002). Overexpression of cyclin B1 in gastric cancer and itsclinicopathological significance: an immunohistological study.J Cancer Res Clin Oncol 128: 412–416.
Yoon SY, Lee Y, Kim JH, Chung AS, Joo JH, Kim CN et al.(2005). Over-expression of human UREB1 in colorectalcancer: HECT domain of human UREB1 inhibits the activity oftumor suppressor p53 protein. Biochem Biophys Res Commun 326:7–17.
Zhao X, Heng JI, Guardavaccaro D, Jiang R, Pagano M, Guillemot Fet al. (2008). The HECT-domain ubiquitin ligase Huwe1 controlsneural differentiation and proliferation by destabilizing the N-Myconcoprotein. Nat Cell Biol 10: 643–653.
Zhong Q, Gao W, Du F, Wang X. (2005). Mule/ARF-BP1, a BH3-only E3 ubiquitin ligase, catalyzes the polyubiquitination of Mcl-1and regulates apoptosis. Cell 121: 1085–1095.
Supplementary Information accompanies the paper on the Oncogene website (http://www.nature.com/onc)
Ubiquitin E3 ligases deregulation in human cancerS Confalonieri et al
2968
Oncogene




















