
AFM of Ex Vivo
15
81 Pier Carlo Braga and Davide Ricci (eds.), Atomic Force Microscopy in Biomedical Research: Methods and Protocols, Methods in Molecular Biology, vol. 736, DOI 10.1007/978-1-61779-105-5_6, © Springer Science+Business Media, LLC 2011 Chapter 6 Atomic Force Microscopy of Ex Vivo Amyloid Fibrils Claudio Canale, Annalisa Relini, and Alessandra Gliozzi Abstract Here, we report a study of ex vivo amyloid fibrils formed, respectively, by the Leu174Ser Apolipoprotein A-I (ApoA-I-LS) variant and by b2-microglobulin (b2-m) (Relini et al., J. Biol. Chem. 281:16521– 16529, 2006; Relini et al., Biochim. Biophys. Acta 1690:33–41, 2004). In the work on ApoA-I-LS, the AFM has been used to characterize and compare the morphologies of amyloid fibrils isolated from two different patients, while in the study on b2-m our investigation provided important information about the factors that can promote the aggregation in vivo. Key words: AFM, Amyloid, Aggregation, Fibrillogenesis An entire class of pathologies, including Alzheimer’s disease, Parkinson’s disease, spongiform encephalopathy, Huntington Corea, and amyloidoses (1), is associated with the deposition of intracellular or extracellular fibrillar protein aggregates, character- ized by a high b-sheet content. The mechanism responsible for the conversion of soluble polypeptide chains into stable, insoluble fibrillar structures termed amyloid fibrils is not completely known. Fibrillogenesis is considered as an alternative pathway to normal protein folding, in which a destabilized polypeptide chain is sequestered by a competing path of aggregation (2). At present, at least 40 different diseases related to the deposition of amyloid or amyloid-like aggregates are known (3); representative examples are listed in Table 1. The AFM has been widely used in the last decade in the study of fibrillogenesis; the very high spatial resolution of this technique and its capability to obtain a three-dimensional image of the sample 1. Introduction
Transcript of AFM of Ex Vivo

81
Pier Carlo Braga and Davide Ricci (eds.), Atomic Force Microscopy in Biomedical Research: Methods and Protocols, Methods in Molecular Biology, vol. 736, DOI 10.1007/978-1-61779-105-5_6, © Springer Science+Business Media, LLC 2011
Chapter 6
Atomic Force Microscopy of Ex Vivo Amyloid Fibrils
Claudio Canale, Annalisa Relini, and Alessandra Gliozzi
Abstract
Here, we report a study of ex vivo amyloid fibrils formed, respectively, by the Leu174Ser Apolipoprotein A-I (ApoA-I-LS) variant and by b2-microglobulin (b2-m) (Relini et al., J. Biol. Chem. 281:16521–16529, 2006; Relini et al., Biochim. Biophys. Acta 1690:33–41, 2004). In the work on ApoA-I-LS, the AFM has been used to characterize and compare the morphologies of amyloid fibrils isolated from two different patients, while in the study on b2-m our investigation provided important information about the factors that can promote the aggregation in vivo.
Key words: AFM, Amyloid, Aggregation, Fibrillogenesis
An entire class of pathologies, including Alzheimer’s disease, Parkinson’s disease, spongiform encephalopathy, Huntington Corea, and amyloidoses (1), is associated with the deposition of intracellular or extracellular fibrillar protein aggregates, character-ized by a high b-sheet content. The mechanism responsible for the conversion of soluble polypeptide chains into stable, insoluble fibrillar structures termed amyloid fibrils is not completely known. Fibrillogenesis is considered as an alternative pathway to normal protein folding, in which a destabilized polypeptide chain is sequestered by a competing path of aggregation (2). At present, at least 40 different diseases related to the deposition of amyloid or amyloid-like aggregates are known (3); representative examples are listed in Table 1.
The AFM has been widely used in the last decade in the study of fibrillogenesis; the very high spatial resolution of this technique and its capability to obtain a three-dimensional image of the sample
1. Introduction
82 Canale, Relini, and Gliozzi
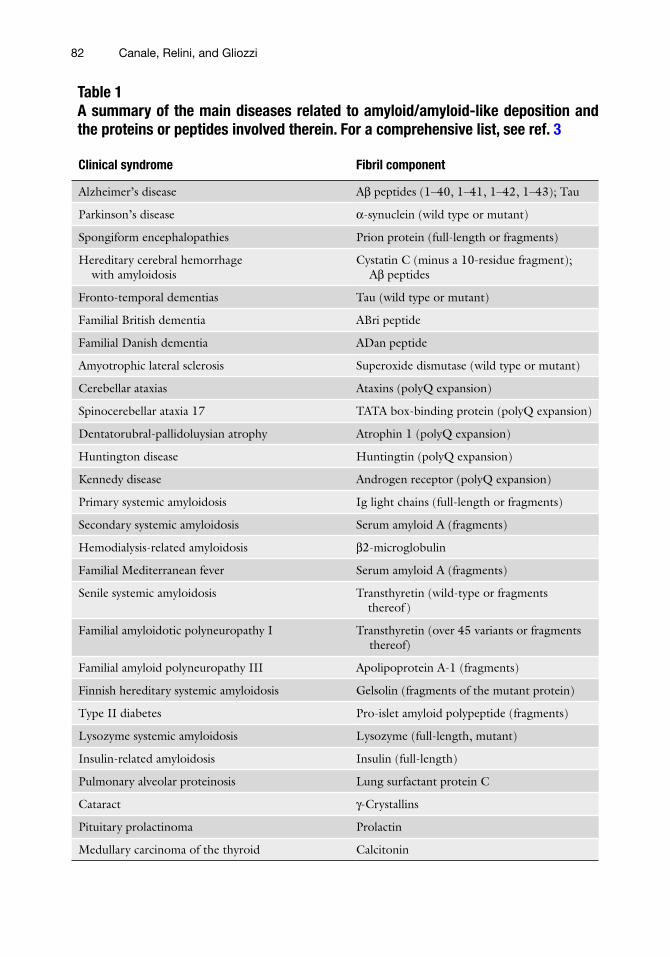
Table 1A summary of the main diseases related to amyloid/amyloid-like deposition and the proteins or peptides involved therein. For a comprehensive list, see ref. 3
Clinical syndrome Fibril component
Alzheimer’s disease Ab peptides (1–40, 1–41, 1–42, 1–43); Tau
Parkinson’s disease a-synuclein (wild type or mutant)
Spongiform encephalopathies Prion protein (full-length or fragments)
Hereditary cerebral hemorrhage with amyloidosis
Cystatin C (minus a 10-residue fragment); Ab peptides
Fronto-temporal dementias Tau (wild type or mutant)
Familial British dementia ABri peptide
Familial Danish dementia ADan peptide
Amyotrophic lateral sclerosis Superoxide dismutase (wild type or mutant)
Cerebellar ataxias Ataxins (polyQ expansion)
Spinocerebellar ataxia 17 TATA box-binding protein (polyQ expansion)
Dentatorubral-pallidoluysian atrophy Atrophin 1 (polyQ expansion)
Huntington disease Huntingtin (polyQ expansion)
Kennedy disease Androgen receptor (polyQ expansion)
Primary systemic amyloidosis Ig light chains (full-length or fragments)
Secondary systemic amyloidosis Serum amyloid A (fragments)
Hemodialysis-related amyloidosis b2-microglobulin
Familial Mediterranean fever Serum amyloid A (fragments)
Senile systemic amyloidosis Transthyretin (wild-type or fragments thereof )
Familial amyloidotic polyneuropathy I Transthyretin (over 45 variants or fragments thereof)
Familial amyloid polyneuropathy III Apolipoprotein A-1 (fragments)
Finnish hereditary systemic amyloidosis Gelsolin (fragments of the mutant protein)
Type II diabetes Pro-islet amyloid polypeptide (fragments)
Lysozyme systemic amyloidosis Lysozyme (full-length, mutant)
Insulin-related amyloidosis Insulin (full-length)
Pulmonary alveolar proteinosis Lung surfactant protein C
Cataract g-Crystallins
Pituitary prolactinoma Prolactin
Medullary carcinoma of the thyroid Calcitonin

83Atomic Force Microscopy of Ex Vivo Amyloid Fibrils
surface at the nanometric scale had a fundamental role in the morphological characterization of amyloid fibrils as well as prefi-brillar aggregates (4–8). Moreover, the ability of the AFM to work in different environments without needing any particular sample preparation procedures, such as staining, fixation, or met-allization, enables the observation of the fibrillization process in different solution conditions and/or in the presence of different cofactors (6, 7, 9).
In vitro experiments can provide important information on the mechanisms involved in the aggregation process. On the other hand, the analysis of fibrils extracted from ex vivo patho-logical deposits is fundamental for characterizing the structural and morphological features of amyloid aggregates and their pos-sible interplay with other biological factors. In spite of this, the number of studies involving in vitro fibrils is highly dominant over the analysis of ex vivo specimens, likely due to difficulties related to the availability of natural samples.
Apolipoprotein A-I (ApoA-I) is the major protein component of high-density lipoproteins (HDL). ApoA-I is responsible for cho-lesterol extraction and transport from body tissues. Variants of ApoA-I are involved in some forms of hereditary systemic amyloi-dosis. In particular, ApoA-I-LS is related to an amyloidosis form with predominant heart involvement; in this study, we examined fibrils isolated from the heart of two patients.
Amyloid fibrils of ApoA-I had not yet been characterized by AFM when we started this work, the only data available on their structure had been obtained by small angle X-ray diffraction on the ApoA-I-LS mutant (10) and by electron microscopy cross-sectional images on the Leu60Arg ApoA-I mutant (11). Characterization of the ultrastructural assembly of ApoA-I-LS in natural amyloid fibrils provided complementary information on the three-dimensional organization of this type of fibrillar deposits.
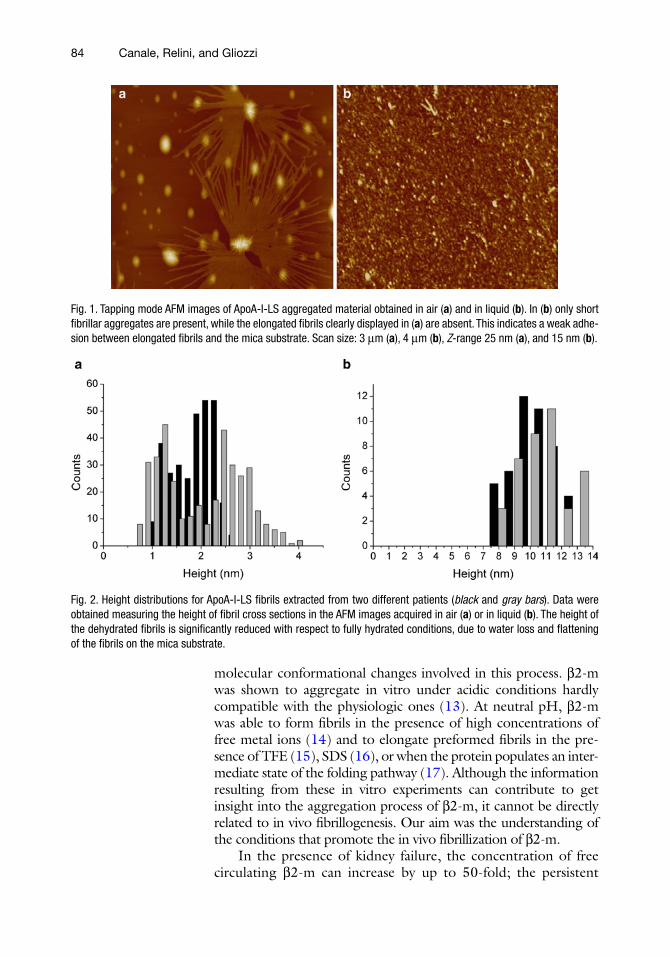
We compared the morphologies of ApoA-I-LS amyloid mate-rial obtained from the two patients and we determined the struc-tural parameters of the aggregates observed both in air and under liquid working in tapping mode. Figure 1 shows two representative areas acquired in air (Fig. 1a) or in liquid (Fig. 1b). The fibrils extracted from the two patients showed common morphologies, the same structural features and compatible sizes (Fig. 2). In addi-tion, we discussed the role of the nonfibrillar aggregates, found to coexist with the fibrils, as potential fibril precursors (12).
The deposition of insoluble fibrils of b2-m in the musculo-skeletal system is characteristic of dialysis-related amyloidosis. The aggrega-tion process of b2-m has been the object of extensive investigation since many years. Several experiments have been designed to repro-duce b2-m amyloid deposition in vitro and understand the
1.1. Morphological Characterization of Ex Vivo Amyloid Fibrils Formed by the Apolipoprotein A-I Leu174Ser Variant
1.2. The Role of Collagen in the Aggregation of b2-Microglobulin
84 Canale, Relini, and Gliozzi
molecular conformational changes involved in this process. b2-m was shown to aggregate in vitro under acidic conditions hardly compatible with the physiologic ones (13). At neutral pH, b2-m was able to form fibrils in the presence of high concentrations of free metal ions (14) and to elongate preformed fibrils in the pre-sence of TFE (15), SDS (16), or when the protein populates an inter-mediate state of the folding pathway (17). Although the information resulting from these in vitro experiments can contribute to get insight into the aggregation process of b2-m, it cannot be directly related to in vivo fibrillogenesis. Our aim was the understanding of the conditions that promote the in vivo fibrillization of b2-m.
In the presence of kidney failure, the concentration of free circulating b2-m can increase by up to 50-fold; the persistent
Fig. 1. Tapping mode AFM images of ApoA-I-LS aggregated material obtained in air (a) and in liquid (b). In (b) only short fibrillar aggregates are present, while the elongated fibrils clearly displayed in (a) are absent. This indicates a weak adhe-sion between elongated fibrils and the mica substrate. Scan size: 3 mm (a), 4 mm (b), Z-range 25 nm (a), and 15 nm (b).
Fig. 2. Height distributions for ApoA-I-LS fibrils extracted from two different patients (black and gray bars). Data were obtained measuring the height of fibril cross sections in the AFM images acquired in air (a) or in liquid (b). The height of the dehydrated fibrils is significantly reduced with respect to fully hydrated conditions, due to water loss and flattening of the fibrils on the mica substrate.

85Atomic Force Microscopy of Ex Vivo Amyloid Fibrils
increase in b2-m concentration results in amyloid deposition, preferentially localized in the musculo-skeletal system. The accu-mulation of b2-m deposits has been shown to cause arthralgias, destructive osteoarthropathies, and carpal tunnel syndrome. Although a high concentration of b2-m is a typical condition at the onset of the disease, there is not a strict correlation between the disease severity and b2-m levels (18), suggesting that other factors might be involved in b2-m amyloid deposition.
b2-m is ubiquitously present in the human body, since the peptide is released from every cell expressing the major histocom-patibility complex class I (MHCI). On the contrary, one of the most peculiar properties of DRA is its strict specificity for tissues of the skeletal system; in fact, the tissues that are primarily involved are bones and ligaments. Therefore, it might be reasonable to associate the localized protein aggregation and fibrillization to the molecular environment that is present in the skeletal system. The AFM inspection of ex vivo material has been fundamental to reveal the strict association between b2-m fibrils and collagen fibers (Fig. 3).
We performed in vitro experiments using conditions as close as possible to the conditions occurring in the periarticular tissues of patients subject to long-term hemodialysis. In the presence of a flogistic process, the local pH can decrease locally to 6.4 (18). The temperature was set to 37°C, the physiological temperature, and to 40°C, in order to mimic the extreme temperature that can be induced by an inflammatory state. We studied the aggregation
Fig. 3. Surface plots of a tapping mode AFM image (height data) of b2-m ex vivo amyloid fibrils surrounding a collagen fiber. The collagen is easily detectable from the characteristic band pattern, with a periodicity of 67 nm. The strict asso-ciation between amyloid fibrils and collagen suggested an active role of the collagen in the aggregation process of b2-m. This research was originally published by Relini et al. (6).

86 Canale, Relini, and Gliozzi
process in the absence or in the presence of type I collagen. Aggregation experiments were performed at a protein concentra-tion ranging between 0.2 and 0.6 mg/ml. Fibrillization occurred only in the presence of collagen (Fig. 4).
In this work, the analysis of ex vivo materials provided impor-tant information, not only on the morphology of amyloid aggre-gates, but also on the relation between amyloid deposition and collagen. In vitro experiments demonstrated that the latter has an active role in the fibrillogenesis of b2-m.
Fig. 4. Tapping mode AFM images of b2-m amyloid fibrils obtained in vitro at 40°C and pH 6.4 in the presence of collagen. (a) and (c) are topographical images, while (b) and (d) are amplitude images. In (a) and (b), the arrows show a collagen fibril embedded in the surrounding material partially composed of amyloid fibrils. Scan size: 880 nm (a, b), 1.5 mm (c, d), Z-range 12 nm (a), and 10 nm (c). In the absence of collagen, only a low number of globular aggregates and rare filamentous structures are found (data not shown). This research was originally published by Relini et al. (6).

87Atomic Force Microscopy of Ex Vivo Amyloid Fibrils
1. Metallic magnetic disks of 12 mm in diameter (provided by the mechanical workshop of the Physics Department at the University of Genoa).
2. Mica sheets (Agar Scientific, purchased from Assing, Monterotondo, Rome).
3. Tape and double-side adhesive tape. 4. Two-component epoxy glue.
1. Amyloid material purification buffer: 10 mM Tris, 1 mM EDTA, 140 mM NaCl, 1.5 mM phenylmethylsulfonyl fluo-ride (PhMeSO2F), and 0.1% NaN3, pH 8.0.
2. AFM imaging and sample preparation buffer: Tris 50 mM, pH 7.0.
1. A Dimension 3000 microscope (Digital Instruments-Veeco, Santa Barbara, USA), equipped with a “G” scanning head (maximum scan size 100 mm) and driven by a Nanoscope IIIa controller, was used in all the experiments.
2. V-shaped gold-coated Si3N4 cantilevers (DNP, Veeco, Santa Barbara, USA) with a nominal spring constant of 0.06 N/m and pyramidal tips having a nominal curvature radius of about 40 nm were used for imaging in liquid.
3. Single beam silicon cantilevers (type OMCL-AC160TS, Olympus, Japan) with a nominal spring constant of 40 N/m and typical tip curvature radius around 7 nm were used for imaging in air.
See Subheading 2.1.1.
1. 10 mM Tris/EDTA, containing 1.5 M phenylmethylsulfonyl fluoride.
2. Ammonium acetate 50 mM, pH 7.4.
1. AFM images were acquired using a Multimode Scanning Probe microscope (Digital Instruments-Veeco, Santa Barbara, USA) equipped with an “E” scanning head (maximum scan size 10 mm) and driven by a Nanoscope IV controller. For larger scan sizes,
2. Materials
2.1. Materials for ApoA-I Leu174Ser Variant
2.1.1. Preparation of the Substrates for the AFM Analysis
2.1.2. Chemicals
2.1.3. AFM
2.2. Materials for Aggregation of b2-Microglobulin
2.2.1. Preparation of the Substrates for the AFM Analysis
2.2.2. Chemicals
2.2.3. AFM

88 Canale, Relini, and Gliozzi
a Dimension 3000 microscope (Digital Instruments-Veeco), equipped as described in Subheading 2.1.3 was employed.
2. The same AFM cantilevers described previously in Subheading 2.1.3 were used.
The ex vivo material was purified following the methods developed by Pras et al. (19). Fibrils were extracted from the explanted hearts of two patients affected by cardiac amyloidosis. Approximately 800 mg of tissue was homogenized and centri-fuged at 60,000 × g in 2 ml of buffer containing 10 mM Tris, 1 mM EDTA, 140 mM NaCl, 1.5 mM phenylmethylsulfonyl fluoride (PhMeSO2F), and 0.1% NaN3, pH 8.0. The procedure that allowed the clearing of the non-amyloid material was stopped when the absorbance at 280 nm of the supernatant dropped under 0.1 units. The buffer was replaced by distilled water, and homogenization and centrifugation were carried out six times. Amyloid material was collected from the supernatant and the pro-tein concentration monitored by Congo red staining and thiofla-vin assay. See Obici et al. (11) for further details.
1. The Dimension 3000 sample holder is equipped with a mag-net, for this reason it is useful to use magnetic metallic disks to fix the sample.
2. Cut a piece of mica of about 1 × 1 cm2 using a pair of scissors. 3. Mica substrates must be attached to metallic disks. To per-
form imaging in air of dried samples, mica substrates can be attached before or after sample deposition and dehydration by means of double-side adhesive tape. This procedure is not suitable for liquid imaging, as usually the tape tends to detach from the metallic disk if in contact with an aqueous solution. The most convenient method to attach mica onto the metallic disks is to use a two-component epoxy glue. This is probably the best choice also for the preparation of dried samples; the only inconvenient is the long time required to dry the glue, generally more than 1 h, depending on the kind of glue. However, it is possible to prepare several substrates in advance and then to cleave mica just before sample deposition.
4. Sample deposition: the sample was diluted to a concentration of 50 mg/ml, a small aliquot of 20 ml was deposited on freshly cleaved mica. For imaging in air, the sample was dried under mild vacuum for 30 min (Note 1); for imaging in liquid, the sample was incubated on mica for 30 min and then rinsed with the same buffer to eliminate the material which had not adhered to the substrate (Note 2).
3. Methods
3.1. Methods for ApoA-I Leu174Ser Variant
3.1.1. Purification of the Ex Vivo Material
3.1.2. AFM Sample Preparation

89Atomic Force Microscopy of Ex Vivo Amyloid Fibrils
1. Mount the cantilever in the AFM holder as suggested in the user manual of the AFM.
2. Mount the holder in the AFM head and start the laser align-ment. This operation is generally described in the AFM user manuals as well. The basic concepts are the same for all AFMs, but setups produced by different companies may adopt dif-ferent procedures for laser alignment. When the laser is reflecting on the back of the cantilever and the laser spot is centered on the photodiode, the sum signal increases to a value that depends on the kind of cantilevers used. All the measurements described here were obtained using cantilevers that were metal-coated on the back side, in order to increase reflectivity; with our setup the sum was around 5–6 V.
3. When working in liquid with very soft metal-coated cantile-vers (0.03–0.4 N/m), a drift of the laser spot on the photo-detector can be clearly displayed; the vertical deflection changes with time due to cantilever bending. This pheno-menon is due to the different thermal expansion coefficients of the two materials that are in contact. Wait until the canti-lever reaches the thermal equilibrium with the environment or, alternatively, use uncoated silicon cantilevers.
4. Before engagement, the cantilever resonance frequency (fo) must be determined. Some of the AFM systems, as for exam-ple the Veeco systems, have an automatic procedure to deter-mine fo and the working amplitude of oscillation (amplitude setpoint, A). Otherwise, fo and A have to be chosen manually, following the AFM manual instructions.
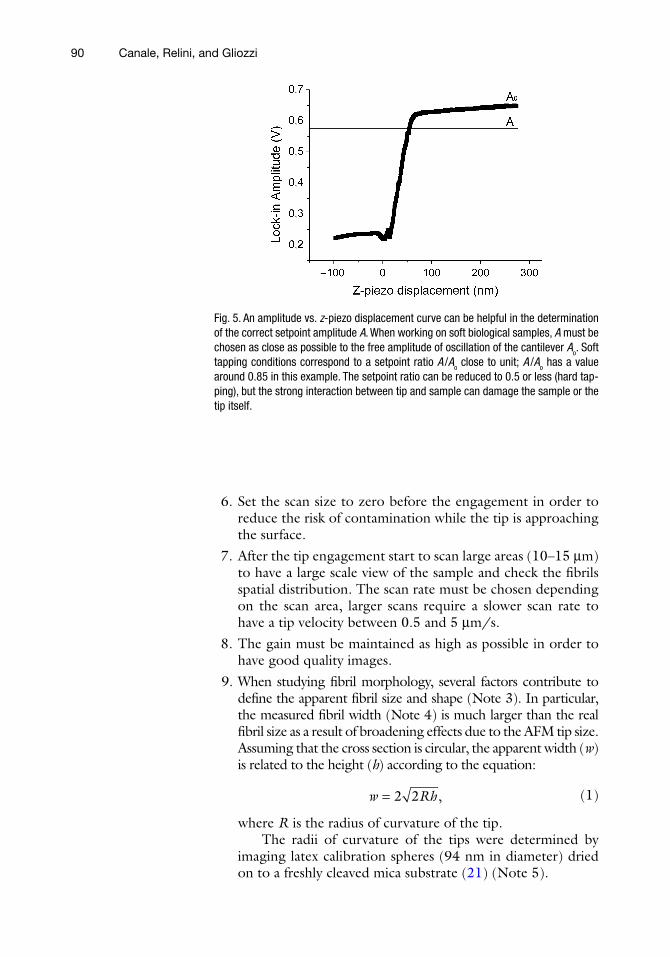
5. When working in tapping mode, a fundamental parameter is the amplitude setpoint A; lower setpoints correspond to a stronger interaction between the tip and the sample. Generally A is chosen equal to the 70–80% of the free oscillation ampli-tude Ao. A high interaction force between the AFM probe and the sample may damage the AFM tip or the sample itself, espe-cially if working on delicate biological materials. To engage softly on the sample, it is recommended to use high amplitude setpoints, more than 80% of Ao. In this case, it might happen that the tip is not really engaged to the surface. There are essentially two ways to optimize the tip engagement:
Looking at the scope mode, decrease gradually the ampli- –tude setpoint during imaging until the surface profile can be clearly seen.
In liquid, amplitude vs. – z-piezo displacement curve can help in determining the setpoint (Fig. 5); to know more about this procedure, see Lantz et al. (20). In the case of stiff silicon cantilevers used for tapping mode in air, this operation is not recommended as the strong interaction between tip and surface can easily damage the tip.
3.1.3. AFM Operation
90 Canale, Relini, and Gliozzi
6. Set the scan size to zero before the engagement in order to reduce the risk of contamination while the tip is approaching the surface.
7. After the tip engagement start to scan large areas (10–15 mm) to have a large scale view of the sample and check the fibrils spatial distribution. The scan rate must be chosen depending on the scan area, larger scans require a slower scan rate to have a tip velocity between 0.5 and 5 mm/s.
8. The gain must be maintained as high as possible in order to have good quality images.
9. When studying fibril morphology, several factors contribute to define the apparent fibril size and shape (Note 3). In particular, the measured fibril width (Note 4) is much larger than the real fibril size as a result of broadening effects due to the AFM tip size. Assuming that the cross section is circular, the apparent width (w) is related to the height (h) according to the equation:
2 2 ,w Rh= (1)
where R is the radius of curvature of the tip.The radii of curvature of the tips were determined by
imaging latex calibration spheres (94 nm in diameter) dried on to a freshly cleaved mica substrate (21) (Note 5).
Fig. 5. An amplitude vs. z-piezo displacement curve can be helpful in the determination of the correct setpoint amplitude A. When working on soft biological samples, A must be chosen as close as possible to the free amplitude of oscillation of the cantilever Ao. Soft tapping conditions correspond to a setpoint ratio A /Ao close to unit; A /Ao has a value around 0.85 in this example. The setpoint ratio can be reduced to 0.5 or less (hard tap-ping), but the strong interaction between tip and sample can damage the sample or the tip itself.

91Atomic Force Microscopy of Ex Vivo Amyloid Fibrils
1. Amyloid fibrils were isolated from the femoral head of a patient. The isolation procedure was very similar to that used for the extraction of ApoA-I-LS described in Subheading 3.1.1, details are reported in Relini et al. (6).
2. Recombinant b2-m was expressed and purified according to the procedure reported by Esposito et al. (22). The protein was dehydrated after purification.
3. When lyophilized b2-m was dissolved in a buffer at pH 6.4 at room temperature, some tangles of fibrils were observed. To avoid the presence of aggregated material, the protein was dissolved in ammonium acetate 50 mM at pH 7.4 and then the pH was adjusted to 6.4 just at the beginning of the exper-iment. The protein concentration was 2 mg/ml, and the solution was centrifuged at 16,500 × g for 1 h at the tempera-ture of 4°C in order to remove large aggregates. The super-natant was collected and filtered with 20-nm pore size filters (Anotop 10, cat. no. 6809-1002, Whatman, USA). Aggregation experiments were performed at a protein concentration of 0.2–0.6 mg/ml, after acidification to pH 6.4.
4. Type I collagen was purified from calf skin (23). SDS–PAGE was used to check the collagen purity. Fibrillar collagen was prepared by solubilizing the purified collagen in 5 mM acetic acid. The solution was incubated at 37°C for 30 min after a dilution 1:1 with 2× phosphate-buffered saline.
5. Fibrillar collagen was sonicated for 10 min in a bath sonicator (ACAD, Genoa, Italy) and washed with the ammonium ace-tate buffer. Then, it was put on a microscope slide, it was cut into several small pieces that were washed again and added to the protein solution (Note 6).
1. Subheadings 1–3 of paragraph 3.1 describe general proce-dures that have been used also in the preparation of b2-m samples. A different method has been used to deposit the sample in the presence of fibrillar collagen. In this case, a piece of collagen was collected from the solution and depos-ited onto a freshly cleaved mica substrate together with its surrounding protein solution (Note 7). The sample was dried under mild vacuum and imaged in tapping mode in air.
As the AFM settings have been already described in Part I, the present paragraph must be intended as complementary to Subheading 3.1.3.
1. Special attention must be taken when engaging on the b2-m/collagen samples. The piece of collagen dried on the mica substrate forms a relatively thick layer of very rough material. It is not possible to obtain nanometric resolution if the AFM
3.2. Methods for Aggregation of b2-Microglobulin
3.2.1. Purification of the Ex Vivo Material and In Vitro Fibrillization
3.2.2. AFM Sample Preparation
3.2.3. AFM Operation

92 Canale, Relini, and Gliozzi
probe is engaged in the center of the collagen network. To achieve a resolution allowing amyloid fibrils detection, the tip should be engaged close to the collagen edge, but in a flat area. This operation can be performed easily using the optics that is integrated in the Dimension 3000 system or using the top view optics of the Multimode system (Note 8).
The AFM technique has been successfully applied to the study of ex vivo amyloid material.
We had the rare opportunity to analyze amyloid material isolated by two patients affected by hereditary cardiac amyloido-sis. The morphological characterization of two unrelated samples of ApoA-I amyloid aggregates demonstrated that aggregate sizes and shapes are quite comparable. Our AFM inspections show that not only mature fibrils, but also the prefibrillar species display the same morphological features.
The analysis of ex vivo material extracted from the femoral head of a patient revealed the strict association between b2-m fibrils and collagen fibers. The interesting correlation between amyloid fibrils and collagen has been a starting point for a series of in vitro aggregation experiments. Following the hypothesis, suggested by the ex vivo inspection, of an active role of collagen in the aggregation process of b2-m, we have been able to obtain b2-m fibrillation in vitro under conditions which are very close to the physiopathological ones.
1. Amyloid proteins usually have a high tendency to aggregate. In this case, the drying procedure could induce an increase of the aggregation rate due to the increased protein concentra-tion as consequence of solvent evaporation. Therefore, it is important to dry the sample as quickly as possible, either using a low vacuum chamber or a dry nitrogen flow.
2. One of the most important steps for the morphological and structural characterization of a heterogeneous sample by AFM is the sample deposition procedure. During this opera-tion, part of the material composing the sample may be lost or artifacts could be generated, precluding the visualization of important components of the specimen.
When working in liquid, the materials that are adsorbed on the substrate are the only ones that can be observed by AFM, the part of the sample weakly adhering to the substrate is removed by the AFM tip during scan. In the case of ApoA-I-LS, the globular aggregates that were present in the solu-tion adsorbed onto mica and a homogeneous distribution of
3.3. Conclusions
4. Notes

93Atomic Force Microscopy of Ex Vivo Amyloid Fibrils
this material was clearly displayed (Fig. 1b), while only short fibrillar aggregates (200–250 nm in length) were observed (Fig. 1b); longer fibrils, such as those imaged in air (Fig. 1a), did not adhere to the substrate. In principle, using substrates with different characteristics (electric charge, hydrophilicity, etc.), should allow observing different species that are present in the sample, thus obtaining important information on the chemical-physical characteristics of these species.
The problem of poor sample adhesion to the substrate can be overcome working in air onto dehydrated samples, there-fore avoiding the loss of material. Moreover, when using a buf-fer the dehydration procedure gives rise to salt crystals that can completely cover the sample masking its features. In this case, it is preferable to incubate the sample on the mica substrate for some minutes and then rinse it gently with Milli-Q water.
3. Shape and size deformations induced by the dehydration pro-cess must be taken into account when measuring the size of a dried biological sample. The evaporation of water can induce a significant shrinking in a material with high water content. In addition, the drying procedure can induce a flattening of the sample on the mica surface. The decrease of fibril thickness is clearly shown in Fig. 2 for ApoA-I-LS amyloid aggregates. This result was in agreement with previous data obtained on amyloid aggregates formed by another peptide (24).
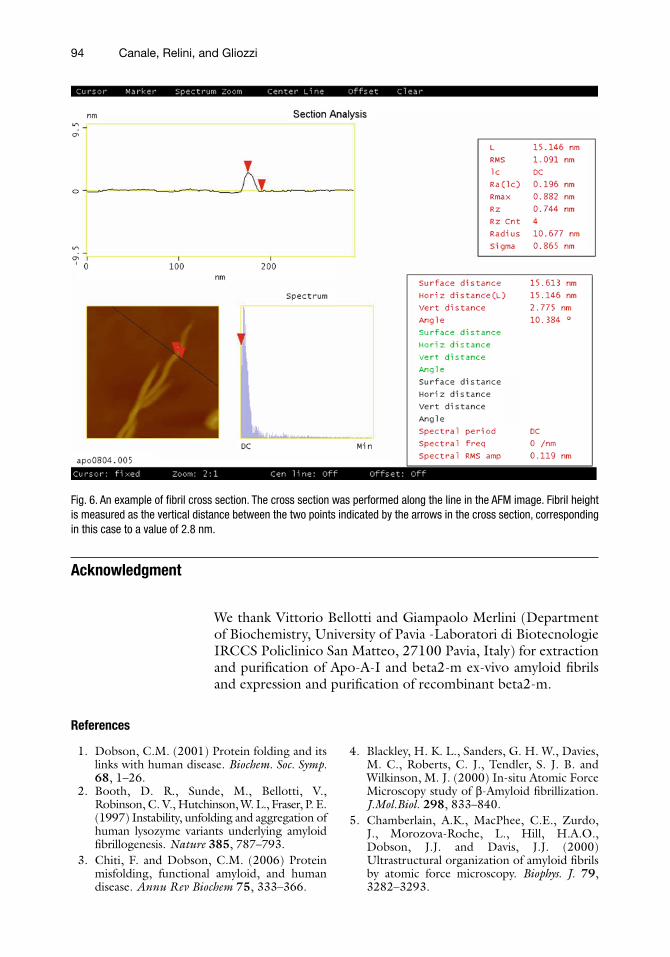
4. Fibril height and width must be measured in individual fibrils from the image sections taken perpendicularly to the fibril longitudinal axis (Fig. 6).
5. Any material with a well-known size and geometrical shape can be used to determine the AFM tip shape and size. In our work, latex spheres have been used. It is possible to use other kinds of calibration standards, for those commercially avail-able see the brief overview provided by Peter Markiewicz in 1999 (25), showing all the most common techniques that are used for AFM tip characterization.
6. Fibrillar collagen is very sticky; all the operations described at point 5 of Subheading 3.2.1 must be performed with particular care. In our case, we used Teflon tweezers to manipulate the sample. We prepared at the same time different vials containing b2-m in the conditions described in Subheading 3.2.1 and we added a single piece of collagen per vial.
7. For the AFM inspection of the sample, the collagen must be transferred from the solution to the mica surface. This opera-tion was also performed by using Teflon tweezers.
8. A great attention to sample size must be paid using a Multimode AFM head. Due to its particular head design, this instrument can be used only for the study of small samples.
94 Canale, Relini, and Gliozzi
Acknowledgment
We thank Vittorio Bellotti and Giampaolo Merlini (Department of Biochemistry, University of Pavia -Laboratori di Biotecnologie IRCCS Policlinico San Matteo, 27100 Pavia, Italy) for extraction and purification of Apo-A-I and beta2-m ex-vivo amyloid fibrils and expression and purification of recombinant beta2-m.
References
Fig. 6. An example of fibril cross section. The cross section was performed along the line in the AFM image. Fibril height is measured as the vertical distance between the two points indicated by the arrows in the cross section, corresponding in this case to a value of 2.8 nm.
1. Dobson, C.M. (2001) Protein folding and its links with human disease. Biochem. Soc. Symp. 68, 1–26.
2. Booth, D. R., Sunde, M., Bellotti, V., Robinson, C. V., Hutchinson, W. L., Fraser, P. E. (1997) Instability, unfolding and aggregation of human lysozyme variants underlying amyloid fibrillogenesis. Nature 385, 787–793.
3. Chiti, F. and Dobson, C.M. (2006) Protein misfolding, functional amyloid, and human disease. Annu Rev Biochem 75, 333–366.
4. Blackley, H. K. L., Sanders, G. H. W., Davies, M. C., Roberts, C. J., Tendler, S. J. B. and Wilkinson, M. J. (2000) In-situ Atomic Force Microscopy study of b-Amyloid fibrillization. J.Mol.Biol. 298, 833–840.
5. Chamberlain, A.K., MacPhee, C.E., Zurdo, J., Morozova-Roche, L., Hill, H.A.O., Dobson, J.J. and Davis, J.J. (2000) Ultrastructural organization of amyloid fibrils by atomic force microscopy. Biophys. J. 79, 3282–3293.

95Atomic Force Microscopy of Ex Vivo Amyloid Fibrils
6. Relini, A., Canale, C., De Stefano, S., Rolandi, R., Giorgetti, S., Stoppini, M., Rossi, A., Fogolari, F., Corazza, A., Esposito, G., Gliozzi, A. and Bellotti, V. (2006) Collagen plays an active role in the aggregation of b2-microglobulin under physiopathological con-ditions of dialysis-related amyloidosis. J. Biol. Chem. 281, 16521–16529.
7. Arimon, M., Dıez-Perez, I., Kogan, M. J., Durany, N., Giralt, E., Sanz, F. and Fernandez-Busquets, X. (2005) Fine structure study of Ab1-42 fibrillogenesis with atomic force micros-copy. FASEB J. 19, 1344–1346.
8. Natalello, A., Prokorov, V. V., Tagliavini, F., Morbin, M., Forloni, G., Beeg, M., Manzoni, C., Colombo, L., Gobbi, M., Salmona, M. and Doglia, S. M. (2008) Conformational plasticity of the Gerstmann–Sträussler–Scheinker disease peptide as indicated by its multiple aggregation pathways. J. Mol. Biol. 381, 1349–1361.
9. Ha, C., Ryu, J. and Park, C.B. (2007) Metal ions differentially influence the aggregation and deposition of Alzheimer’s b-amyloid on a solid template. Biochemistry 46, 6118–6125.
10. Mangione, P., Sunde, M., Giorgetti, S., Stoppini, M., Esposito, G., Gianelli, L., Obici, L., Asti, L., Andreola, A., Viglino, P., Merlini, G., Bellotti, V. (2001) Amyloid fibrils derived from the apolipoprotein A-I Leu174Ser vari-ant contain elements of ordered helical struc-ture. Protein Sci. 10, 187–199.
11. Obici, L., Bellotti, V., Mangione, P., Stoppini, M., Arbustini, E., Verga, L., Zorzoli, I., Anesi, E., Zanotti, G., Campana, C., Vigano, M., Merlini, G. (1999) The new apolipoprotein A-I variant Leu174Ser causes hereditary car-diac amyloidosis, and the amyloid fibrils are constituted by the 93-residue N-terminal polypeptide. Am. J. Pathol. 155, 695–702.
12. Relini, A., Rolandi, R., Bolognesi, M., Aboudan, M., Merlini, G., Bellotti, V., Gliozzi, A. (2004) Ultrastructural organiza-tion of ex vivo amyloid fibrils formed by the apolipoprotein A-I Leu174Ser variant: an atomic force microscopy study. Biochim. Biophys. Acta 1690, 33–41.
13. Kad, N.M., Myers, S.L., Smith, D.P., Smith, D.A., Radford, S.E. and Thomson, N.H. (2003) Hierarchical assembly of b2-micro-globulin amyloid in vitro revealed by atomic force microscopy. J. Mol. Biol. 330, 785–797.
14. Morgan, C. J., Gelfand, M., Atreya, C., and Miranker, A. D. (2001) Kidney dialysis-associ-ated amyloidosis: a molecular role for copper in fiber formation. J. Mol. Biol. 309, 339–345.
15. Yamamoto, S, Gejyo, F. (2005) Historical background and clinical treatment of dialysis-related amyloidosis. Biochim. Biophys. Acta 1753, 4–10.
16. Kihara, M., Chatani, E., Sakai, M., Hasegawa, K., Naiki, H., and Goto, Y. (2005) Seeding-dependent maturation of beta2-microglobulin amyloid fibrils at neutral pH. J. Biol. Chem. 280, 12012–12018.
17. Chiti, F., De Lorenzi, E., Grossi, S., Mangione, P., Giorgetti, S., Caccialanza, G., Dobson, C. M., Merlini, G., Ramponi, G., and Bellotti, V. (2001) A partially structured species of beta 2-microglobulin is significantly populated under physiological conditions and involved in fibrillogenesis, J. Biol. Chem. 276, 46714–46721.
18. Floege,J, and Ehlerding, G. (1996) Beta-2-microglobulin-associated amyloidosis. Nephron. 72, 9–26.
19. Pras, M., Schubert, D., Zucker-Franklin, D., Rimon, A., Franklin, A.C. (1968) The charac-terization of soluble amyloid prepared in water. J. Clin. Invest. 47, 924–933.
20. Lantz, M., Liu, Y. Z., Cui, X. D., Tokumoto, H., and Lindsay, S. M., (1999) Dynamic force miscroscopy in fluid. Surf. Interface Anal. 27, 354–360.
21. Ramirez-Aguilar, K.A., Rowlen, K.L. (1998) Tip characterization from AFM images of nanometric spherical particles, Langmuir 14, 2562–2566.
22. Esposito, G., Michelutti, R., Verdone, G., Viglino, P., Hernandez, H., Robinson, C.V., Amoresano, A., Dal Piaz, F., Monti, M., Pucci, P., Mangione, P., Stoppini, M., Merlini, G., Ferri, G., and Bellotti, V. (2000) Removal of the N-terminal hexapeptide from human b2-microglobulin facilitates protein aggrega-tion an fibril formation. Protein Sci 9, 831–845.
23. Giorgetti, S., Rossi, A., Mangione ,P., Raimondi, S., Marini, S., Stoppini, M., Corazza, A., Viglino, P., Esposito, G., Cetta, G., Merlini, G., Bellotti, V. (2005) b2-Micro-globulin isoforms display an heterogeneous affinity for type I collagen. Protein Sci. 14, 696–702.
24. Grégoire, C., Marco, S., Thimonier, J., Duplan, L., Laurine, E., Chauvin, J.-P., Michel, B., Peyrot, V., Verdier, J.-M. (2001) Three-dimensional structure of lithostathine protofibril, a protein involved in Alzheimer’s disease. EMBO J. 20, 3313–3321.
25. Markiewicz, P. http:// www.weizmann.ac.il/Chemical_Research_Support/surflab/peter/standard/index.htm.




















