Additional material and information about AJP- Renal Physiology can be found at
11
doi:10.1152/ajprenal.00301.2006 292:1354-1363, 2007. First published Jan 30, 2007; Am J Physiol Renal Physiol Gamika A. Prathapasinghe, Yaw L. Siow and Karmin O You might find this additional information useful... 40 articles, 20 of which you can access free at: This article cites http://ajprenal.physiology.org/cgi/content/full/292/5/F1354#BIBL 4 other HighWire hosted articles: This article has been cited by [PDF] [Full Text] [Abstract] , August 1, 2007; 293 (2): F594-F600. Am J Physiol Renal Physiol V. Vitvitsky, A. Prudova, S. Stabler, S. Dayal, S. R. Lentz and R. Banerjee sex-dependent differences in plasma homocysteine levels Testosterone regulation of renal cystathionine beta-synthase: implications for [PDF] [Full Text] [Abstract] , January 1, 2008; 294 (1): F236-F244. Am J Physiol Renal Physiol S.-Y. Hwang, C. W. H. Woo, K. K. W. Au-Yeung, Y. L. Siow, T. Y. Zhu and K. O nuclear factor-{kappa}B activation Homocysteine stimulates monocyte chemoattractant protein-1 expression in the kidney via [PDF] [Full Text] [Abstract] , October 1, 2008; 295 (4): F912-F922. Am J Physiol Renal Physiol G. A. Prathapasinghe, Y. L. Siow, Z. Xu and K. O of pH and nitric oxide Inhibition of cystathionine-{beta}-synthase activity during renal ischemia-reperfusion: role [PDF] [Full Text] [Abstract] , July 1, 2009; 297 (1): F27-F35. Am J Physiol Renal Physiol Z. Xu, G. Prathapasinghe, N. Wu, S.-Y. Hwang, Y. L. Siow and K. O generation in the kidney Ischemia-reperfusion reduces cystathionine-{beta}-synthase-mediated hydrogen sulfide including high-resolution figures, can be found at: Updated information and services http://ajprenal.physiology.org/cgi/content/full/292/5/F1354 can be found at: AJP - Renal Physiology about Additional material and information http://www.the-aps.org/publications/ajprenal This information is current as of September 25, 2009 . http://www.the-aps.org/. American Physiological Society. ISSN: 0363-6127, ESSN: 1522-1466. Visit our website at (monthly) by the American Physiological Society, 9650 Rockville Pike, Bethesda MD 20814-3991. Copyright © 2005 by the respective cells and vasculature, as well as to the control of body fluid volume and composition. It is published 12 times a year publishes original manuscripts on a broad range of subjects relating to the kidney, urinary tract, and their AJP - Renal Physiology on September 25, 2009 ajprenal.physiology.org Downloaded from
Transcript of Additional material and information about AJP- Renal Physiology can be found at

doi:10.1152/ajprenal.00301.2006 292:1354-1363, 2007. First published Jan 30, 2007;Am J Physiol Renal Physiol
Gamika A. Prathapasinghe, Yaw L. Siow and Karmin O
You might find this additional information useful...
40 articles, 20 of which you can access free at: This article cites http://ajprenal.physiology.org/cgi/content/full/292/5/F1354#BIBL
4 other HighWire hosted articles: This article has been cited by
[PDF] [Full Text] [Abstract]
, August 1, 2007; 293 (2): F594-F600. Am J Physiol Renal PhysiolV. Vitvitsky, A. Prudova, S. Stabler, S. Dayal, S. R. Lentz and R. Banerjee
sex-dependent differences in plasma homocysteine levelsTestosterone regulation of renal cystathionine beta-synthase: implications for
[PDF] [Full Text] [Abstract], January 1, 2008; 294 (1): F236-F244. Am J Physiol Renal Physiol
S.-Y. Hwang, C. W. H. Woo, K. K. W. Au-Yeung, Y. L. Siow, T. Y. Zhu and K. O nuclear factor-{kappa}B activation
Homocysteine stimulates monocyte chemoattractant protein-1 expression in the kidney via
[PDF] [Full Text] [Abstract], October 1, 2008; 295 (4): F912-F922. Am J Physiol Renal Physiol
G. A. Prathapasinghe, Y. L. Siow, Z. Xu and K. O of pH and nitric oxide
Inhibition of cystathionine-{beta}-synthase activity during renal ischemia-reperfusion: role
[PDF] [Full Text] [Abstract], July 1, 2009; 297 (1): F27-F35. Am J Physiol Renal Physiol
Z. Xu, G. Prathapasinghe, N. Wu, S.-Y. Hwang, Y. L. Siow and K. O generation in the kidney
Ischemia-reperfusion reduces cystathionine-{beta}-synthase-mediated hydrogen sulfide
including high-resolution figures, can be found at: Updated information and services http://ajprenal.physiology.org/cgi/content/full/292/5/F1354
can be found at: AJP - Renal Physiologyabout Additional material and information http://www.the-aps.org/publications/ajprenal
This information is current as of September 25, 2009 .
http://www.the-aps.org/.American Physiological Society. ISSN: 0363-6127, ESSN: 1522-1466. Visit our website at (monthly) by the American Physiological Society, 9650 Rockville Pike, Bethesda MD 20814-3991. Copyright © 2005 by therespective cells and vasculature, as well as to the control of body fluid volume and composition. It is published 12 times a year
publishes original manuscripts on a broad range of subjects relating to the kidney, urinary tract, and theirAJP - Renal Physiology
on Septem
ber 25, 2009 ajprenal.physiology.org
Dow
nloaded from

Detrimental role of homocysteine in renal ischemia-reperfusion injury
Gamika A. Prathapasinghe,1 Yaw L. Siow,2,3 and Karmin O1,2,3
Departments of 1Animal Science and 2Physiology, University of Manitoba, 3Canadian Centre forAgri-Food Research in Health and Medicine, St. Boniface Hospital Research Centre, Winnipeg, Manitoba, Canada
Submitted 1 August 2006; accepted in final form 8 January 2007
Prathapasinghe GA, Siow YL, O K. Detrimental role of homo-cysteine in renal ischemia-reperfusion injury. Am J Physiol RenalPhysiol 292: F1354–F1363, 2007. First published January 30, 2007;doi:10.1152/ajprenal.00301.2006.—Ischemia followed by reperfusionis a major cause for renal injury in both native kidney and renalallografts. Hyperhomocysteinemia, a condition of elevated plasmahomocysteine (Hcy) level, is associated with cardiovascular diseases.Recent evidence suggests that Hcy, at higher levels, may be harmfulto other organs such as the kidney. In this study, we investigated therole of Hcy in ischemia-reperfusion-induced renal injury. The leftkidney of a Sprague-Dawley rat was subjected to either 30-min or 1-hischemia followed by 1- or 24-h reperfusion. Ischemia-reperfusioncaused a significant increase in peroxynitrite formation and lipidperoxidation in kidneys, which reflected oxidative stress. The numberof apoptotic cells in those kidneys was also markedly increased. Hcylevels were elevated 2.9- and 1.5-fold in kidneys subjected to ische-mia alone or ischemia-reperfusion, respectively. Further investigationrevealed that elevation of Hcy level in the kidney upon ischemia-reperfusion was due to reduced activity of cystathionine-�-synthase, akey enzyme in Hcy metabolism. Administration of anti-Hcy antibod-ies into the kidney not only abolished ischemia-reperfusion-inducedoxidative stress and cell death in the kidneys but also restored renalfunction after 1 h of reperfusion. However, such a protective effectwas not sustained after 24 h of reperfusion. In conclusion, ischemia-reperfusion impairs Hcy metabolism in the kidney. Hcy, at elevatedlevels, is capable of inducing oxidative stress and renal injury.Neutralization of Hcy with antibodies offers transient functionalbenefit against ischemia-reperfusion-induced oxidative stress and re-nal injury. These results suggest that Hcy may play a detrimental rolein the kidney during ischemia-reperfusion.
oxidative stress; cystathionine-�-synthase; kidney function
ISCHEMIA-REPERFUSION INJURY is one of the most common causesfor delayed function of renal allografts and is associated withpoor long-term renal function (3). Complete and prolongedinterruption of renal arterial blood flow occurs during renaltransplantation or surgical procedures such as nephro-lithotomy, parenchymal sparing surgery for renal tumors, andrenal arterial surgeries. Prolonged renal ischemia can lead toacute renal failure (2). It has been estimated that ischemicinsult, especially during renal transplantation, is responsible for20–30% primary graft dysfunction (23). Our previous studydemonstrated that ischemia-reperfusion caused oxidative stressleading to NF-�B activation and increased monocyte/macro-phage infiltration into the kidney (28). Oxidative stress repre-sents one of the important mechanisms underlying renal ische-mia-reperfusion injury (21, 28).
Hyperhomocysteinemia, a condition of elevated plasma ho-mocysteine (Hcy) level, is associated with cardiovascular and
cerebral vascular disorders (6, 10). A recent epidemiologicalstudy revealed a positive association between the plasma Hcylevel and development of chronic kidney disease in the generalpopulation (20). Hyperhomocysteinemia is a common clinicalfinding in patients with chronic kidney diseases and occursalmost uniformly in patients with end-stage renal disease (5,26). A recent study indicated that elevated plasma total Hcywas independently associated with increased kidney allograftloss in humans (36). Hcy, at elevated levels, is associated withoxidative stress in extrarenal tissue (1, 34), as well as in thekidney (7, 38). The free or reduced sulfhydryl group of Hcy ishighly reactive at the physiological pH, and in the presence ofmolecular oxygen, Hcy undergoes thiol oxidation reactions(12). Reactive oxygen species such as superoxide and hydro-gen peroxide are produced during autooxidation of Hcy (18). Ithas been suggested that autooxidation of Hcy represents one ofthe mechanisms contributing to Hcy-induced cell injury (27,33, 35). Our recent study demonstrated that levels of peroxyni-trite and oxygen-derived free radicals are significantly elevatedin kidneys isolated from hyperhomocysteinemic rats (38). Hcyis an intermediate amino acid formed during the metabolism ofmethionine. Hcy can be metabolized via two major pathways,namely, the remethylation pathway and transsulfuration path-way (8). In the remethylation pathway, Hcy is converted tomethionine by using folate or betaine as methyl group donors.In the transsulfuration pathway, Hcy is irreversibly metabo-lized to cysteine. Although the kidney contains enzymes thatare responsible for Hcy metabolism via remethylation andtranssulfuration pathways (8), it is estimated that in the kidney,the majority of Hcy is metabolized through the transsulfurationpathway (13). In mammals, cystathionine-�-synthase (CBS)catalyzes the rate-limiting step in the transsulfuration pathway(15, 29).
Although hyperhomocysteinemia is often seen in patientswith kidney disease or postrenal transplantation, it is notknown whether Hcy contributes to renal injury during ische-mia-reperfusion. The aim of the present study was to investi-gate the role of Hcy in ischemia-reperfusion-induced injury inrat kidney.
MATERIALS AND METHODS
Renal ischemia-reperfusion. Male Sprague-Dawley rats weighing250–300 g were anesthetized with an injection of pentobarbitalsodium (50 mg/kg) intraperitoneally. Rats were kept on a heat pad andthe rectal temperature was maintained at 37°C throughout the exper-imental procedure. Renal ischemia was induced by clamping the leftrenal artery for 30 min or 1 h with nontraumatic vascular clamp (FineScience Tools, Vancouver, BC) as previously described (28). After
Address for reprint requests and other correspondence: K. O, Laboratoryof Integrative Biology, CCARM, St. Boniface Hospital Research Centre,351 Tache Ave., Winnipeg, Manitoba, Canada R2H 2A6 (e-mail: [email protected]).
The costs of publication of this article were defrayed in part by the paymentof page charges. The article must therefore be hereby marked “advertisement”in accordance with 18 U.S.C. Section 1734 solely to indicate this fact.
Am J Physiol Renal Physiol 292: F1354–F1363, 2007.First published January 30, 2007; doi:10.1152/ajprenal.00301.2006.
0363-6127/07 $8.00 Copyright © 2007 the American Physiological Society http://www.ajprenal.orgF1354
on Septem
ber 25, 2009 ajprenal.physiology.org
Dow
nloaded from
the clamp was removed, reperfusion was confirmed visually andallowed for another 1 h. Both kidneys were harvested and collected inice-cold potassium phosphate buffer. In some experiments, rats weresubjected to nephrectomy of right kidney and 30-min ischemia of leftkidney followed by 1- or 24-h reperfusion. A blood sample was drawnfrom the abdominal aorta before rats were killed. Plasma was sepa-rated by centrifugation of blood at 3,000 g for 20 min at 4°C. In oneset of experiments, rats were injected with rabbit anti-Hcy polyclonalantibodies (250 �l, 1:20 dilution; Chemicon International, Temecula,CA) or rabbit IgG (250 �l, 1:20 dilution; Santa Cruz Biotechnology)through the left renal artery immediately after the induction ofischemia. Injection of anti-Hcy antibodies to the kidney via left renalartery immediately after the induction of ischemia would allowimmediate access of antibodies to the kidney to neutralize Hcygenerated inside the kidney during ischemia-reperfusion, to minimizeantibody dilution in the circulation and to minimize systemic immuneresponse since anti-Hcy antibodies were raised in rabbit. In another setof experiments, rats received L-Hcy (2.5 mg/kg) or vehicle (bufferused to prepare L-Hcy) via the left femoral vein when reperfusionstarted. L-Hcy was prepared from L-Hcy thiolactone which was
hydrolyzed with 5 M NaOH at 37°C for 5 min and neutralized with 1M KH2PO4 (31). A sham-operated group of rats was subjected to thesame surgical procedure without inducing ischemia-reperfusion andwere killed at corresponding time points. Results obtained from thisgroup were used as controls. The Hcy concentrations in the plasmaand kidney tissue homogenate were measured with the IMx Hcy assaywhich was based on fluorescence polarization immunoassay technol-ogy (Abbott Diagnostics, Abbott Park, IL) (1, 37). Plasma creatininelevel was determined by using WAKO creatinine kit (Wako ChemicalIndustries) (39). Kidneys in ice-cold potassium phosphate buffer werebisected. One-half were preserved in 10% formalin for paraffinembedding. All procedures were performed in accordance with theGuide to the Care and Use of Experimental Animals published by theCanadian Council on Animal Care and approved by the University ofManitoba Protocol Management and Review Committee.
Determination of lipid peroxidation. The degree of lipid peroxida-tion in the kidney tissue was determined by measuring malondialde-hyde (MDA) levels with thiobarbituric acid reactive substances(TBARS) assay (22, 28). Briefly, a 10% (wt/vol) kidney homogenatewas prepared in 0.1 M KCl solution containing 3 mM EDTA followedby centrifugation at 600 g for 15 min at 4°C. An aliquot of supernatantwas added to the reaction mixture containing 8% SDS, 20% aceticacid, 0.8% thiobarbituric acid and water. After incubation at 95°C for1 h, the amount of MDA formed in the reaction mixture was measuredby spectrophotometer at absorbance of 532 nm. MDA was used as thestandard, and results are expressed as a percentage of sham-operatedgroup. The amount of MDA correlates to the degree of lipid peroxi-dation produced in the tissue.
Measurement of CBS activity. The assay was based on a methoddeveloped by Mudd et al. (19) and modified by Taoka et al. (29). A10% (wt/vol) tissue homogenate was prepared in 0.05 M potassiumphosphate buffer (pH 6.9) followed by centrifugation at 18,000 g for
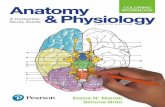
Fig. 1. Measurement of homocysteine (Hcy) concentrations and renal cysta-thionine-�-synthase (CBS) activity. A: left kidney was subjected to 1-hischemia or 1-h ischemia followed by 1-h reperfusion (I/R). Hcy level in thekidney tissue homogenate was measured. In the control group (Sham), only thelaparotomy was performed. B: plasma Hcy levels in different groups of ratswere measured. C: kidney CBS activity was determined. Results are expressedas means � SE (n � 6). *P � 0.05 when compared with the value obtainedfrom sham-operated group.
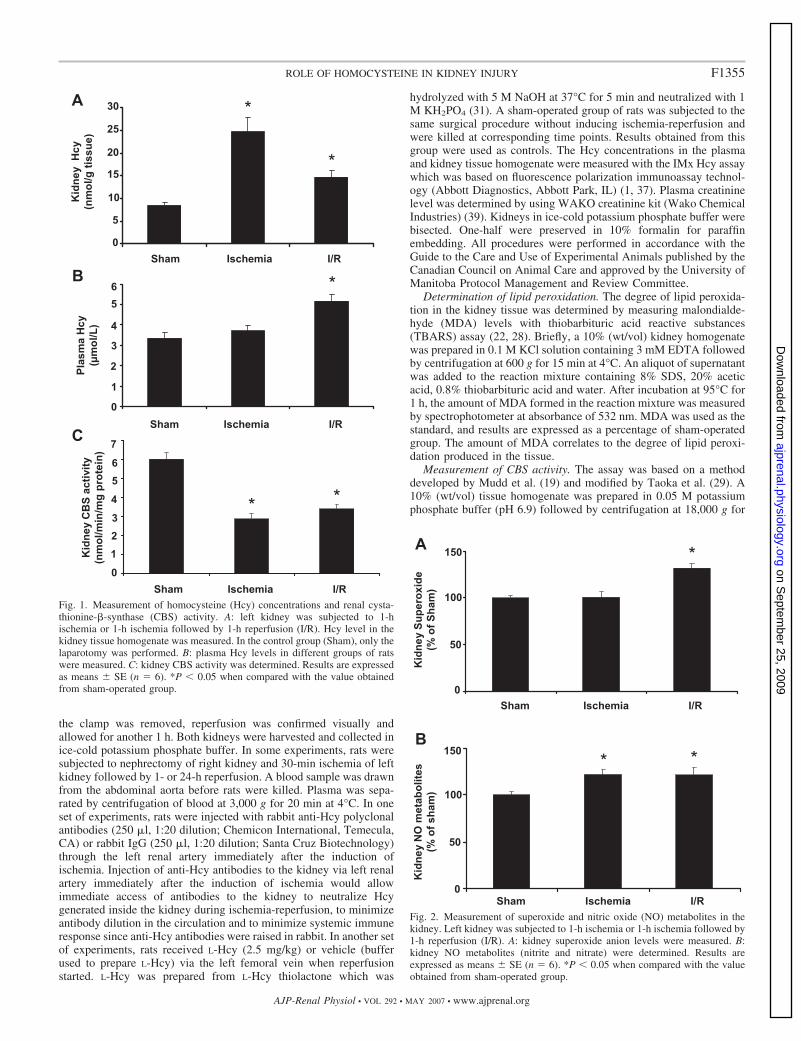
Fig. 2. Measurement of superoxide and nitric oxide (NO) metabolites in thekidney. Left kidney was subjected to 1-h ischemia or 1-h ischemia followed by1-h reperfusion (I/R). A: kidney superoxide anion levels were measured. B:kidney NO metabolites (nitrite and nitrate) were determined. Results areexpressed as means � SE (n � 6). *P � 0.05 when compared with the valueobtained from sham-operated group.
F1355ROLE OF HOMOCYSTEINE IN KIDNEY INJURY
AJP-Renal Physiol • VOL 292 • MAY 2007 • www.ajprenal.org
on Septem
ber 25, 2009 ajprenal.physiology.org
Dow
nloaded from
30 min at 4°C. The reaction was carried out in a reaction mixturecontaining 125 mM Tris �HCl (pH 8.3), 2.1 mM EDTA, 0.146 mML-cystathionine, 41.7 mM DL-Hcy, 0.316 mM S-adenosylmethionine,2.1 mM propargylglycine, and 0.42 mM pyridoxal phosphate. The
reaction was initiated by adding 30 mM [1-14C] serine to the reactionmixture. After 1-h incubation at 37°C, ice-cold 15% perchloric acidwas added to stop the reaction. An aliquot of the reaction mixture (0.5ml) was applied to a Bio-Rad AG 50W-X4 anion exchange column
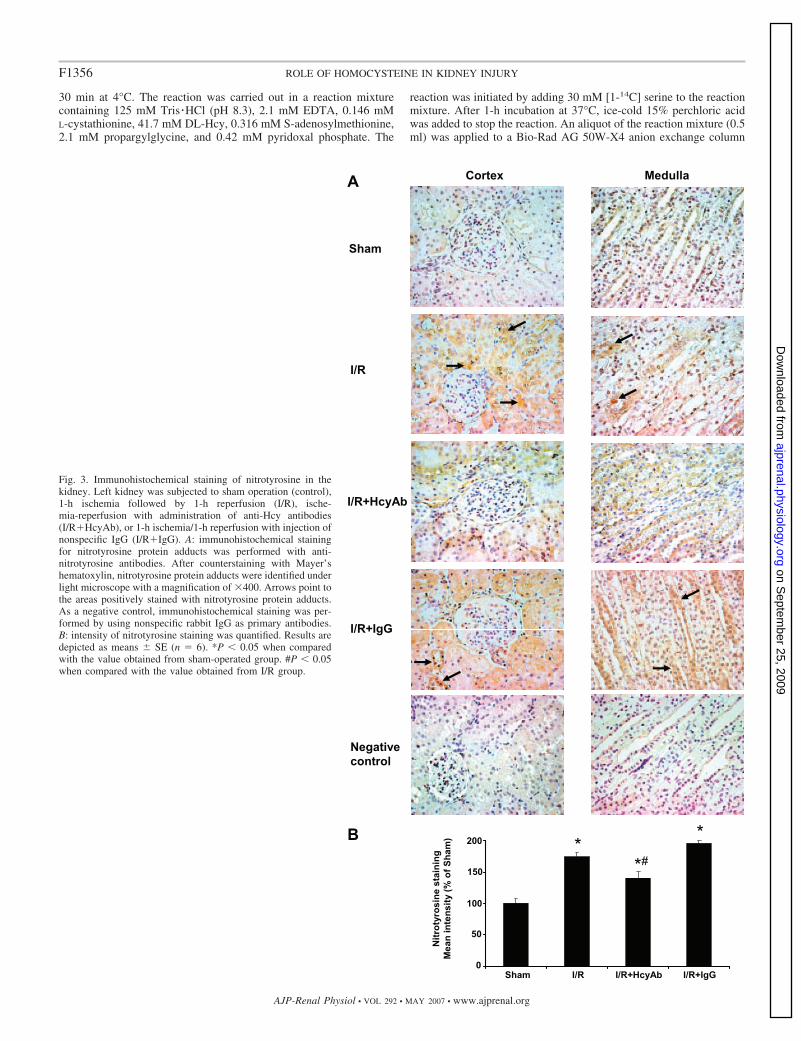
Fig. 3. Immunohistochemical staining of nitrotyrosine in thekidney. Left kidney was subjected to sham operation (control),1-h ischemia followed by 1-h reperfusion (I/R), ische-mia-reperfusion with administration of anti-Hcy antibodies(I/R�HcyAb), or 1-h ischemia/1-h reperfusion with injection ofnonspecific IgG (I/R�IgG). A: immunohistochemical stainingfor nitrotyrosine protein adducts was performed with anti-nitrotyrosine antibodies. After counterstaining with Mayer’shematoxylin, nitrotyrosine protein adducts were identified underlight microscope with a magnification of �400. Arrows point tothe areas positively stained with nitrotyrosine protein adducts.As a negative control, immunohistochemical staining was per-formed by using nonspecific rabbit IgG as primary antibodies.B: intensity of nitrotyrosine staining was quantified. Results aredepicted as means � SE (n � 6). *P � 0.05 when comparedwith the value obtained from sham-operated group. #P � 0.05when compared with the value obtained from I/R group.
F1356 ROLE OF HOMOCYSTEINE IN KIDNEY INJURY
AJP-Renal Physiol • VOL 292 • MAY 2007 • www.ajprenal.org
on Septem
ber 25, 2009 ajprenal.physiology.org
Dow
nloaded from
containing hydrogen-form resin. The column was washed with aseries of water and 1 N HCl. Finally, cystathionine was eluted with3 N NH4OH. The radioactivity associated with cystathionine wasdetermined by using a Beckman liquid scintillation counter.
Immunohistochemical staining. A portion of the kidney was im-mersion-fixed in 10% neutral-buffered formalin overnight followedby embedding in paraffin. Sequential 5-�m paraffin-embedded crosssections were prepared. Immunohistochemical staining was per-formed to detect nitrotyrosine protein adducts in the kidney (38). Inbrief, after deparaffinization, sections were incubated with primaryantibodies, mouse anti-nitrotyrosine antibodies (1:100; Zymed Labo-ratories San Francisco, CA). After overnight incubation with primaryantibodies, sections were treated with 0.3% hydrogen peroxide(H2O2) for 20 min at room temperature to inhibit the endogenousperoxidase. Sections were incubated with biotin-conjugated anti-mouse immunoglobulins (1:200, DakoCytomation, Carpinteria, CA)as secondary antibodies followed by peroxidase-conjugated strep-tavidin (Zymed). Sections were then treated with 3,3-diaminobenzi-dine (DAB)-H2O2 colorimetric substrate solution. The attached per-oxidase catalyzed the H2O2-mediated oxidation of DAB to yieldbrown color. The area displayed a brownish color, indicating thenitrotyrosine protein adducts (38). The images (5 per kidney) werecaptured using an Axioskop2 MOT microscope (Carl Zeiss Microim-aging, Thornwood, NY) with an Axiocam camera and analyzed usingPhotoshop 6.0 (Adobe, San Jose, CA). For a negative control, normalIgG was used as the primary antibody.
Determination of superoxide anion. The level of superoxide anionin the kidney was determined as previously described (40). In brief, a
portion of kidney was homogenized in 1:4 volumes of buffer contain-ing 20 mM HEPES, 1 mM EDTA, and 0.1 mM PMSF followed bycentrifugation at 3,000 g for 10 min at 4°C. The supernatant of kidneyhomogenate was incubated in a reaction mixture containing 10 �Mdihydroethidium for 30 min at 37°C. The superoxide anion in thekidney homogenate caused oxidation of dihydroethidium leading tothe formation of ethidium which was detected at an excitation of 475nm and an emission of 610 nm using a fluorometer (Spectra MaxGemini, Molecular Devices, Sunnyvale, CA). The fluorescent signalproduced by ethidium was proportional to the level of superoxideanion present in the kidney.
Determination of nitric oxide metabolites. The measurement ofnitrite and nitrate was used to assess the nitric oxide (NO) levels inkidney tissue (30). In brief, the kidney tissue was homogenized in abuffer containing 20 mM Tris (pH 7.4) and 2 mM EDTA. Thesupernatant was collected after 10 min of centrifugation at 400 g at4°C. The supernatant was deproteinized with 0.3 N NaOH and 5%ZnSO4. After deproteinization, the amount of nitrite and nitrate wasdetermined with Griess reaction method based on the azo couplingreaction (30). In brief, the supernatant was incubated with nitratereductase to reduce nitrate to nitrite. Then, 12.5 mM sulfanilamide in6 N HCl and 12.5 mM N-(1-naphthyl)ethylenediamine were added tothe nitrite solution to complete the azo coupling reaction. The diazo-amino benzene in the reaction mixture was measured by a spectro-photometer at an absorbance of 520 nm. NaNO2 served as an internalstandard.
Terminal deoxyribonucleotide transferase-mediated dUTP nick-end labeling assay. Renal ischemia-reperfusion in animal models canlead to both necrotic and apoptotic forms of cell death. Terminaldeoxyribonucleotide transferase-mediated dUTP nick-end labeling
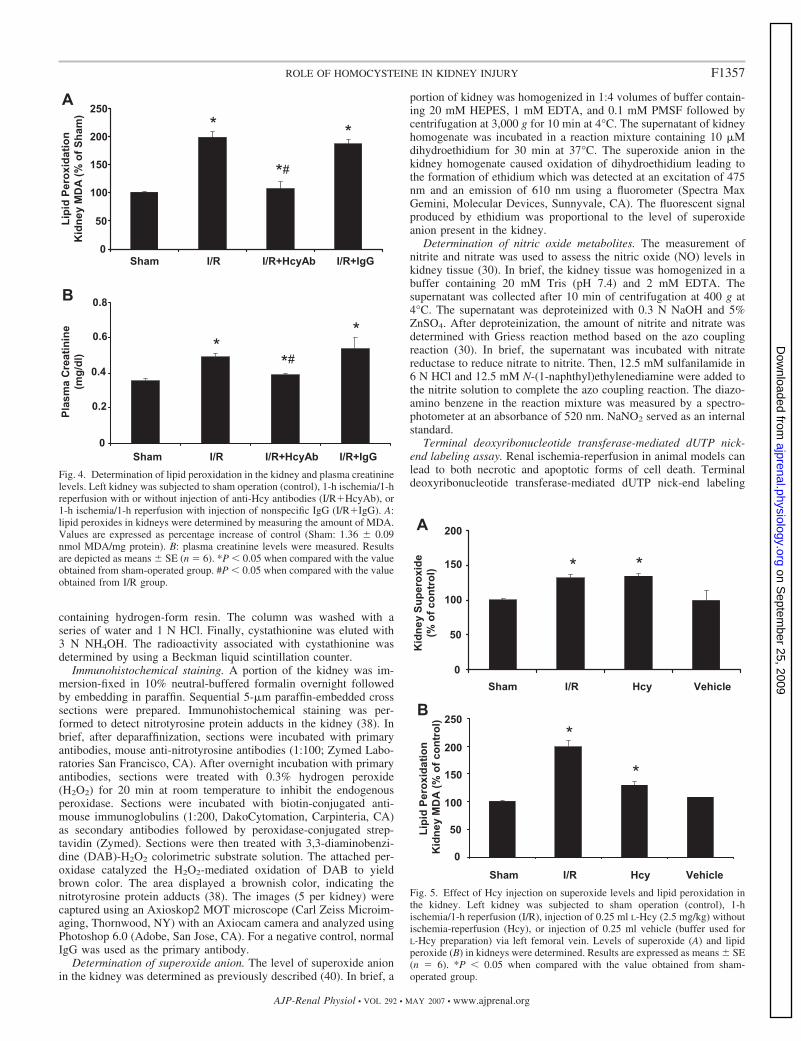
Fig. 4. Determination of lipid peroxidation in the kidney and plasma creatininelevels. Left kidney was subjected to sham operation (control), 1-h ischemia/1-hreperfusion with or without injection of anti-Hcy antibodies (I/R�HcyAb), or1-h ischemia/1-h reperfusion with injection of nonspecific IgG (I/R�IgG). A:lipid peroxides in kidneys were determined by measuring the amount of MDA.Values are expressed as percentage increase of control (Sham: 1.36 � 0.09nmol MDA/mg protein). B: plasma creatinine levels were measured. Resultsare depicted as means � SE (n � 6). *P � 0.05 when compared with the valueobtained from sham-operated group. #P � 0.05 when compared with the valueobtained from I/R group.
Fig. 5. Effect of Hcy injection on superoxide levels and lipid peroxidation inthe kidney. Left kidney was subjected to sham operation (control), 1-hischemia/1-h reperfusion (I/R), injection of 0.25 ml L-Hcy (2.5 mg/kg) withoutischemia-reperfusion (Hcy), or injection of 0.25 ml vehicle (buffer used forL-Hcy preparation) via left femoral vein. Levels of superoxide (A) and lipidperoxide (B) in kidneys were determined. Results are expressed as means � SE(n � 6). *P � 0.05 when compared with the value obtained from sham-operated group.
F1357ROLE OF HOMOCYSTEINE IN KIDNEY INJURY
AJP-Renal Physiol • VOL 292 • MAY 2007 • www.ajprenal.org
on Septem
ber 25, 2009 ajprenal.physiology.org
Dow
nloaded from
(TUNEL) staining to detect DNA fragmentation was performed inrenal sections as an index for cell death (9, 39). Briefly, a portion ofkidney was fixed in 10% neutral-buffered formaldehyde overnight andthen embedded in paraffin. DNA fragments in the sliced sections werelabeled with 3 �M biotin-conjugated dUTP (Roche Molecular Bio-chemical, Loval, Quebec) and 0.3 U/�l terminal deoxyribonucleotidetransferase (TdT; Invitrogen, Carlsbad, CA) followed by counter-stained with periodic acid-Schiff reagent. TUNEL-positive cells wererecognized by focal nuclear staining (9, 39). As a positive control, asection of sham-operated kidney was pretreated with DNase to mimicthe appearance of apoptotic cells. The number of TUNEL-positivecells in renal cortex and medulla was counted in 10 microscopic fieldsunder light microscope at a magnification of �400.
Statistical analysis. Results were analyzed using ANOVA followedby Newman-Keuls multiple comparison test. Data are presented as themeans � SE. P values �0.05 were considered significant.
RESULTS
Elevation of Hcy levels in the kidney and plasma. Ischemiaand ischemia-reperfusion were induced in the left kidney. TheHcy level in kidneys subjected to 1-h ischemia was signifi-cantly higher than that in the sham-operated group (24.68 �3.05 vs. 8.54 � 0.52 nmol/g tissue in the sham group; Fig. 1A).The Hcy level remained elevated in kidneys during the 1-hreperfusion period (12.50 � 0.69 nmol/g tissue; Fig. 1A).Ischemia alone did not elicit any significant change in theplasma Hcy level (Fig. 1B). However, the plasma Hcy levelwas significantly elevated in rats subjected to ischemia fol-lowed by reperfusion (5.18 � 0.32 vs. 3.35 � 0.29 �mol/l inthe sham group; Fig. 1B). Hcy metabolism in the kidney is
mainly through the transsulfuration pathway (13) in whichCBS catalyzes the rate-limiting reaction. To investigatewhether an increase in the Hcy level in kidneys during ische-mia-reperfusion was a result of impaired transsulfuration path-way, the CBS enzyme activity was measured. The CBS activityin kidneys subjected to ischemia alone or ischemia followed byreperfusion was significantly lower than that in the sham-operated group (Fig. 1C). Reduction of Hcy metabolism due toimpaired CBS activity during ischemia-reperfusion could leadto an elevation of Hcy level in the kidney and subsequently inthe circulation.
Generation of superoxide and NO in the kidney. Ischemia-reperfusion resulted in a significant elevation of superoxide andNO metabolite (nitrate and nitrite) levels in the kidney tissuehomogenate (Fig. 2, A and B). Superoxide can interact rapidlywith NO to form peroxynitrite, a potent oxidant. To determinewhether there was an increase in peroxynitrite formation in thekidney subjected to ischemia-reperfusion, immunohistochem-ical staining was performed to detect nitrotyrosine, a biomarkerfor peroxynitrite. A significant increase in the intensity ofnitrotyrosine protein adduct staining was found in kidneyssubjected to ischemia-reperfusion (Fig. 3, A and B), indicatingan increase in peroxynitrite formation. On the other hand, littlenitrotyrosine protein adduct was detected in sham-operatedkidneys (Fig. 3). Next, to determine whether Hcy played anyrole in ischemia-reperfusion-induced peroxynitrite formationin the kidney, anti-Hcy antibodies were injected into the leftrenal artery immediately after the induction of ischemia. Ad-ministration of anti-Hcy antibodies effectively abolished ische-mia-reperfusion-induced peroxynitrite formation in the kidney(Fig. 3). As a control, one group of rats was given nonspecificantibodies (rabbit IgG). Administration of nonspecific antibod-ies into the kidney did not attenuate ischemia-reperfusion-induced peroxynitrite formation (Fig. 3A). These results indi-cated that elevation of tissue Hcy levels might be responsiblefor increased peroxynitrite formation in the kidney upon ische-mia-reperfusion.
Increased lipid peroxidation and plasma creatinine level inthe kidney during ischemia-reperfusion. Peroxynitrite can leadto tissue damage by oxidizing lipids to form lipid peroxides.Elevation of MDA level is an indicator for lipid peroxideformation in the tissue. There was more than a twofold increasein the MDA level in kidneys subjected to ischemia-reperfusion,reflecting an increased renal lipid peroxidation (Fig. 4A). Ad-ministration of anti-Hcy antibodies into the kidney effectivelyreduced ischemia-reperfusion-induced lipid peroxidation to thebasal level found in the sham-operated group (Fig. 4A). As acontrol, one group of rats received nonspecific antibodies (IgG)and such a treatment did not reduce ischemia-reperfusion-induced lipid peroxidation in the kidney (Fig. 4A). One-hourischemia followed by 1-h reperfusion caused a significantincrease in plasma creatinine level (0.43 � 0.02 vs. 0.35 �0.01 mg/dl in the sham group). Administration of anti-Hcyantibodies reduced the plasma creatinine level to the basal levelobserved in the sham-operated group (Fig. 4B). Administrationof nonspecific antibodies (IgG) did not reduce plasma creati-nine level in rats subjected to ischemia-reperfusion (Fig. 4B).These results suggested that elevation of tissue Hcy level mightcontribute to ischemia-reperfusion-induced lipid peroxidationin the kidney and the renal dysfunction.
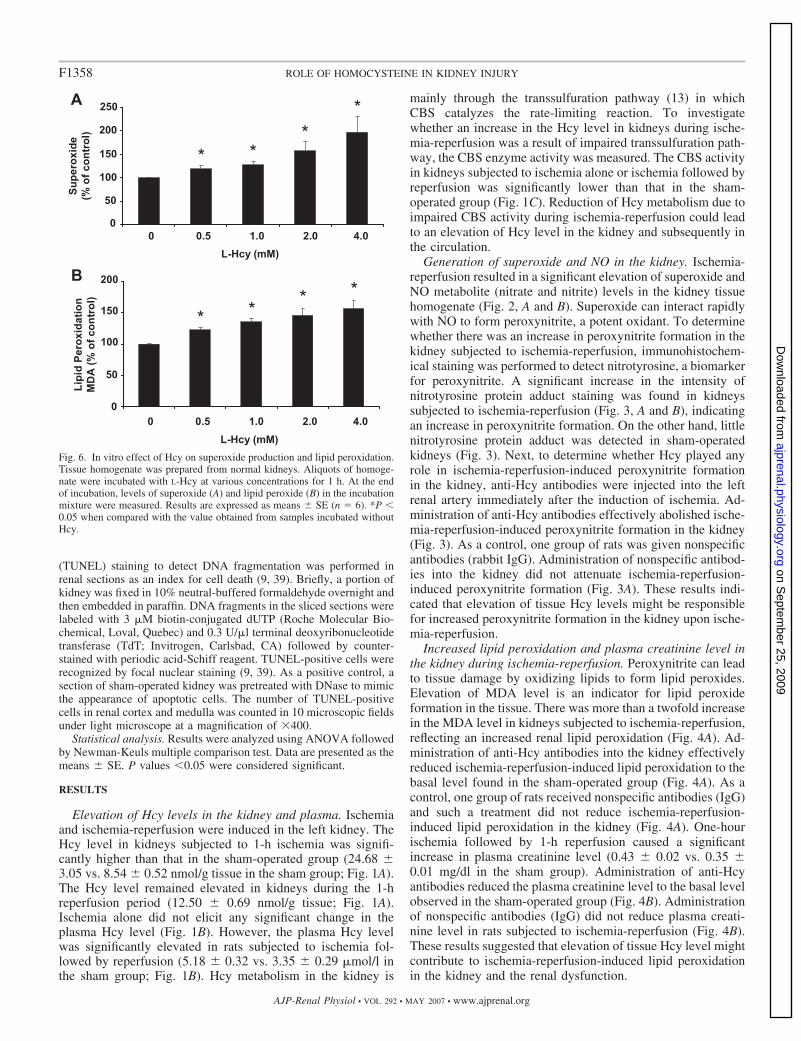
Fig. 6. In vitro effect of Hcy on superoxide production and lipid peroxidation.Tissue homogenate was prepared from normal kidneys. Aliquots of homoge-nate were incubated with L-Hcy at various concentrations for 1 h. At the endof incubation, levels of superoxide (A) and lipid peroxide (B) in the incubationmixture were measured. Results are expressed as means � SE (n � 6). *P �0.05 when compared with the value obtained from samples incubated withoutHcy.
F1358 ROLE OF HOMOCYSTEINE IN KIDNEY INJURY
AJP-Renal Physiol • VOL 292 • MAY 2007 • www.ajprenal.org
on Septem
ber 25, 2009 ajprenal.physiology.org
Dow
nloaded from
Role of Hcy in oxidative stress in the kidney. To furtherexamine the effect of Hcy on oxidative stress in the kidney,L-Hcy (2.5 mg/kg) or vehicle was injected into rats via leftfemoral vein. In the absence of ischemia-reperfusion, injectionof Hcy via intravenous route caused a significant increase inrenal superoxide (Fig. 5A) and lipid peroxide levels (Fig. 5B).Next, the in vitro effect of Hcy was examined. The addition ofL-Hcy to the tissue homogenate prepared from normal kidneyscaused a significant increase in superoxide (Fig. 6A) and lipidperoxide (Fig. 6B) levels. Such in vitro stimulatory effect byHcy was in a concentration dependent manner (Fig. 6).
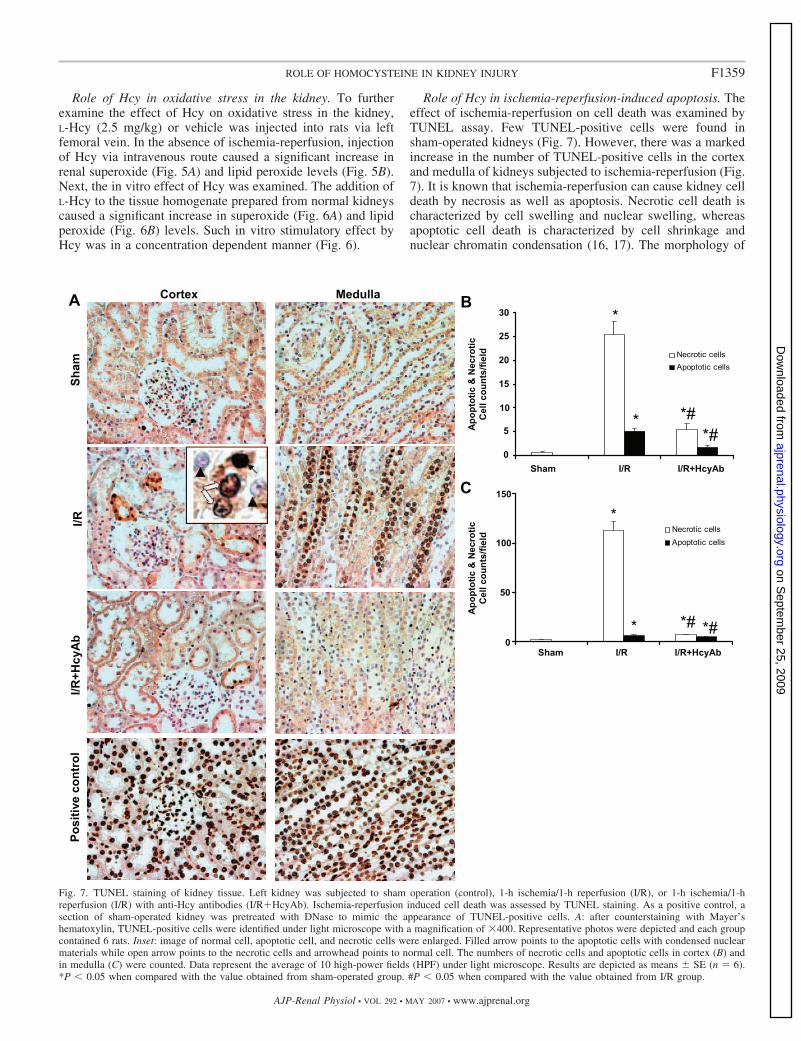
Role of Hcy in ischemia-reperfusion-induced apoptosis. Theeffect of ischemia-reperfusion on cell death was examined byTUNEL assay. Few TUNEL-positive cells were found insham-operated kidneys (Fig. 7). However, there was a markedincrease in the number of TUNEL-positive cells in the cortexand medulla of kidneys subjected to ischemia-reperfusion (Fig.7). It is known that ischemia-reperfusion can cause kidney celldeath by necrosis as well as apoptosis. Necrotic cell death ischaracterized by cell swelling and nuclear swelling, whereasapoptotic cell death is characterized by cell shrinkage andnuclear chromatin condensation (16, 17). The morphology of
Fig. 7. TUNEL staining of kidney tissue. Left kidney was subjected to sham operation (control), 1-h ischemia/1-h reperfusion (I/R), or 1-h ischemia/1-hreperfusion (I/R) with anti-Hcy antibodies (I/R�HcyAb). Ischemia-reperfusion induced cell death was assessed by TUNEL staining. As a positive control, asection of sham-operated kidney was pretreated with DNase to mimic the appearance of TUNEL-positive cells. A: after counterstaining with Mayer’shematoxylin, TUNEL-positive cells were identified under light microscope with a magnification of �400. Representative photos were depicted and each groupcontained 6 rats. Inset: image of normal cell, apoptotic cell, and necrotic cells were enlarged. Filled arrow points to the apoptotic cells with condensed nuclearmaterials while open arrow points to the necrotic cells and arrowhead points to normal cell. The numbers of necrotic cells and apoptotic cells in cortex (B) andin medulla (C) were counted. Data represent the average of 10 high-power fields (HPF) under light microscope. Results are depicted as means � SE (n � 6).*P � 0.05 when compared with the value obtained from sham-operated group. #P � 0.05 when compared with the value obtained from I/R group.
F1359ROLE OF HOMOCYSTEINE IN KIDNEY INJURY
AJP-Renal Physiol • VOL 292 • MAY 2007 • www.ajprenal.org
on Septem
ber 25, 2009 ajprenal.physiology.org
Dow
nloaded from
TUNEL-positive cells was examined (16, 17). Necrotic cellsdisplayed cell swelling with dispersed nuclear materials whileapoptotic cells were smaller in size with condensed nuclei (Fig.7A). Most of the TUNEL-positive cells found in the ischemia-reperfusion group were necrotic cells rather than apoptoticcells (Fig. 7, B and C). Taken together, these results suggestedthat Hcy, at an elevated level, contributed to ischemia-reperfusion-induced cell death both by necrosis and apoptosisin the kidney. Neutralization of tissue Hcy with anti-Hcyantibodies could attenuate 1-h ischemia/1-h reperfusion-in-duced cell death in kidneys.
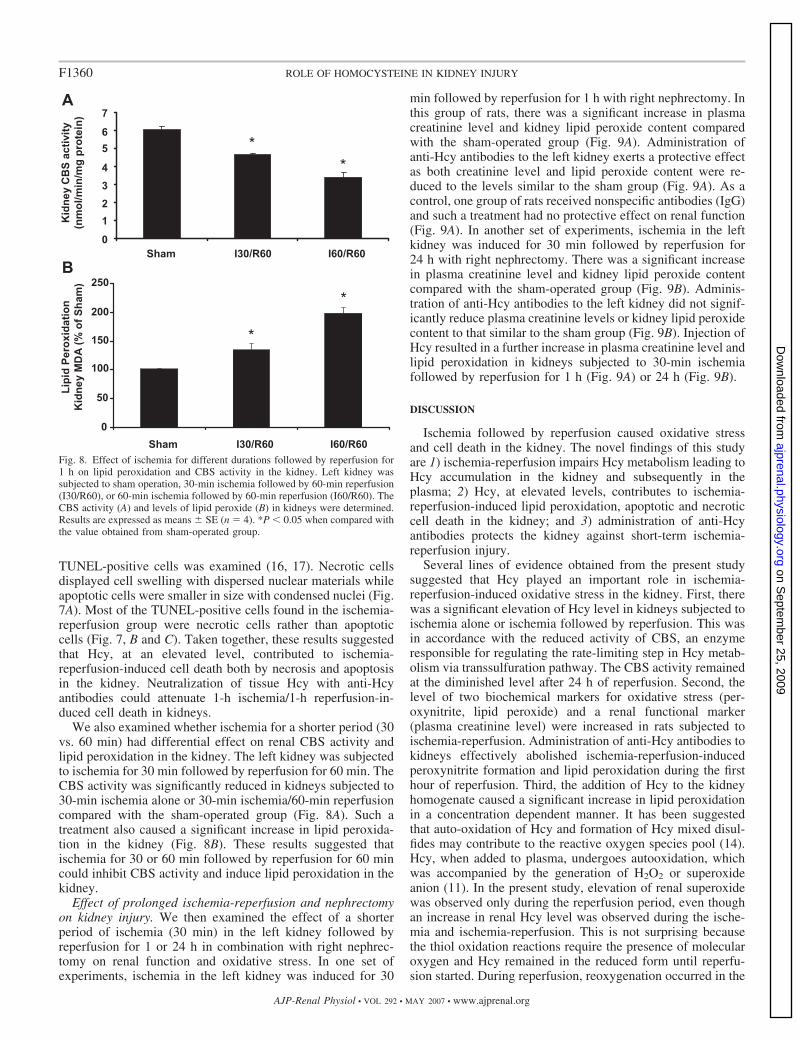
We also examined whether ischemia for a shorter period (30vs. 60 min) had differential effect on renal CBS activity andlipid peroxidation in the kidney. The left kidney was subjectedto ischemia for 30 min followed by reperfusion for 60 min. TheCBS activity was significantly reduced in kidneys subjected to30-min ischemia alone or 30-min ischemia/60-min reperfusioncompared with the sham-operated group (Fig. 8A). Such atreatment also caused a significant increase in lipid peroxida-tion in the kidney (Fig. 8B). These results suggested thatischemia for 30 or 60 min followed by reperfusion for 60 mincould inhibit CBS activity and induce lipid peroxidation in thekidney.
Effect of prolonged ischemia-reperfusion and nephrectomyon kidney injury. We then examined the effect of a shorterperiod of ischemia (30 min) in the left kidney followed byreperfusion for 1 or 24 h in combination with right nephrec-tomy on renal function and oxidative stress. In one set ofexperiments, ischemia in the left kidney was induced for 30
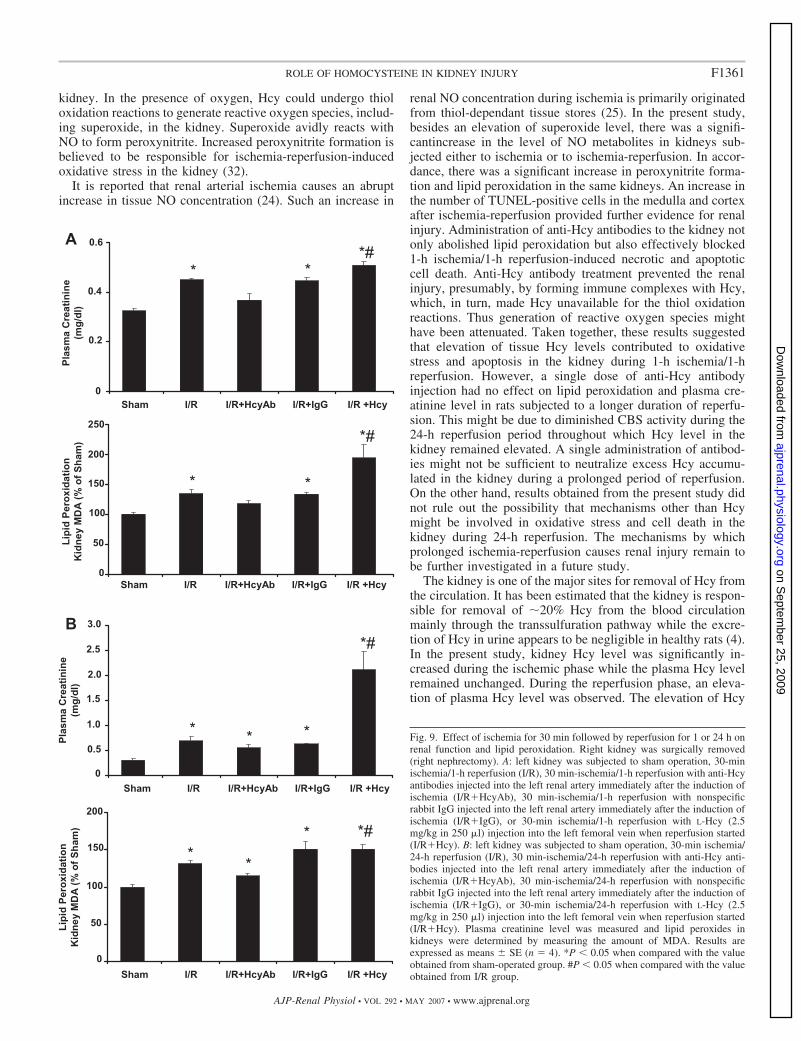
min followed by reperfusion for 1 h with right nephrectomy. Inthis group of rats, there was a significant increase in plasmacreatinine level and kidney lipid peroxide content comparedwith the sham-operated group (Fig. 9A). Administration ofanti-Hcy antibodies to the left kidney exerts a protective effectas both creatinine level and lipid peroxide content were re-duced to the levels similar to the sham group (Fig. 9A). As acontrol, one group of rats received nonspecific antibodies (IgG)and such a treatment had no protective effect on renal function(Fig. 9A). In another set of experiments, ischemia in the leftkidney was induced for 30 min followed by reperfusion for24 h with right nephrectomy. There was a significant increasein plasma creatinine level and kidney lipid peroxide contentcompared with the sham-operated group (Fig. 9B). Adminis-tration of anti-Hcy antibodies to the left kidney did not signif-icantly reduce plasma creatinine levels or kidney lipid peroxidecontent to that similar to the sham group (Fig. 9B). Injection ofHcy resulted in a further increase in plasma creatinine level andlipid peroxidation in kidneys subjected to 30-min ischemiafollowed by reperfusion for 1 h (Fig. 9A) or 24 h (Fig. 9B).
DISCUSSION
Ischemia followed by reperfusion caused oxidative stressand cell death in the kidney. The novel findings of this studyare 1) ischemia-reperfusion impairs Hcy metabolism leading toHcy accumulation in the kidney and subsequently in theplasma; 2) Hcy, at elevated levels, contributes to ischemia-reperfusion-induced lipid peroxidation, apoptotic and necroticcell death in the kidney; and 3) administration of anti-Hcyantibodies protects the kidney against short-term ischemia-reperfusion injury.
Several lines of evidence obtained from the present studysuggested that Hcy played an important role in ischemia-reperfusion-induced oxidative stress in the kidney. First, therewas a significant elevation of Hcy level in kidneys subjected toischemia alone or ischemia followed by reperfusion. This wasin accordance with the reduced activity of CBS, an enzymeresponsible for regulating the rate-limiting step in Hcy metab-olism via transsulfuration pathway. The CBS activity remainedat the diminished level after 24 h of reperfusion. Second, thelevel of two biochemical markers for oxidative stress (per-oxynitrite, lipid peroxide) and a renal functional marker(plasma creatinine level) were increased in rats subjected toischemia-reperfusion. Administration of anti-Hcy antibodies tokidneys effectively abolished ischemia-reperfusion-inducedperoxynitrite formation and lipid peroxidation during the firsthour of reperfusion. Third, the addition of Hcy to the kidneyhomogenate caused a significant increase in lipid peroxidationin a concentration dependent manner. It has been suggestedthat auto-oxidation of Hcy and formation of Hcy mixed disul-fides may contribute to the reactive oxygen species pool (14).Hcy, when added to plasma, undergoes autooxidation, whichwas accompanied by the generation of H2O2 or superoxideanion (11). In the present study, elevation of renal superoxidewas observed only during the reperfusion period, even thoughan increase in renal Hcy level was observed during the ische-mia and ischemia-reperfusion. This is not surprising becausethe thiol oxidation reactions require the presence of molecularoxygen and Hcy remained in the reduced form until reperfu-sion started. During reperfusion, reoxygenation occurred in the
Fig. 8. Effect of ischemia for different durations followed by reperfusion for1 h on lipid peroxidation and CBS activity in the kidney. Left kidney wassubjected to sham operation, 30-min ischemia followed by 60-min reperfusion(I30/R60), or 60-min ischemia followed by 60-min reperfusion (I60/R60). TheCBS activity (A) and levels of lipid peroxide (B) in kidneys were determined.Results are expressed as means � SE (n � 4). *P � 0.05 when compared withthe value obtained from sham-operated group.
F1360 ROLE OF HOMOCYSTEINE IN KIDNEY INJURY
AJP-Renal Physiol • VOL 292 • MAY 2007 • www.ajprenal.org
on Septem
ber 25, 2009 ajprenal.physiology.org
Dow
nloaded from
kidney. In the presence of oxygen, Hcy could undergo thioloxidation reactions to generate reactive oxygen species, includ-ing superoxide, in the kidney. Superoxide avidly reacts withNO to form peroxynitrite. Increased peroxynitrite formation isbelieved to be responsible for ischemia-reperfusion-inducedoxidative stress in the kidney (32).
It is reported that renal arterial ischemia causes an abruptincrease in tissue NO concentration (24). Such an increase in
renal NO concentration during ischemia is primarily originatedfrom thiol-dependant tissue stores (25). In the present study,besides an elevation of superoxide level, there was a signifi-cantincrease in the level of NO metabolites in kidneys sub-jected either to ischemia or to ischemia-reperfusion. In accor-dance, there was a significant increase in peroxynitrite forma-tion and lipid peroxidation in the same kidneys. An increase inthe number of TUNEL-positive cells in the medulla and cortexafter ischemia-reperfusion provided further evidence for renalinjury. Administration of anti-Hcy antibodies to the kidney notonly abolished lipid peroxidation but also effectively blocked1-h ischemia/1-h reperfusion-induced necrotic and apoptoticcell death. Anti-Hcy antibody treatment prevented the renalinjury, presumably, by forming immune complexes with Hcy,which, in turn, made Hcy unavailable for the thiol oxidationreactions. Thus generation of reactive oxygen species mighthave been attenuated. Taken together, these results suggestedthat elevation of tissue Hcy levels contributed to oxidativestress and apoptosis in the kidney during 1-h ischemia/1-hreperfusion. However, a single dose of anti-Hcy antibodyinjection had no effect on lipid peroxidation and plasma cre-atinine level in rats subjected to a longer duration of reperfu-sion. This might be due to diminished CBS activity during the24-h reperfusion period throughout which Hcy level in thekidney remained elevated. A single administration of antibod-ies might not be sufficient to neutralize excess Hcy accumu-lated in the kidney during a prolonged period of reperfusion.On the other hand, results obtained from the present study didnot rule out the possibility that mechanisms other than Hcymight be involved in oxidative stress and cell death in thekidney during 24-h reperfusion. The mechanisms by whichprolonged ischemia-reperfusion causes renal injury remain tobe further investigated in a future study.
The kidney is one of the major sites for removal of Hcy fromthe circulation. It has been estimated that the kidney is respon-sible for removal of 20% Hcy from the blood circulationmainly through the transsulfuration pathway while the excre-tion of Hcy in urine appears to be negligible in healthy rats (4).In the present study, kidney Hcy level was significantly in-creased during the ischemic phase while the plasma Hcy levelremained unchanged. During the reperfusion phase, an eleva-tion of plasma Hcy level was observed. The elevation of Hcy
Fig. 9. Effect of ischemia for 30 min followed by reperfusion for 1 or 24 h onrenal function and lipid peroxidation. Right kidney was surgically removed(right nephrectomy). A: left kidney was subjected to sham operation, 30-minischemia/1-h reperfusion (I/R), 30 min-ischemia/1-h reperfusion with anti-Hcyantibodies injected into the left renal artery immediately after the induction ofischemia (I/R�HcyAb), 30 min-ischemia/1-h reperfusion with nonspecificrabbit IgG injected into the left renal artery immediately after the induction ofischemia (I/R�IgG), or 30-min ischemia/1-h reperfusion with L-Hcy (2.5mg/kg in 250 �l) injection into the left femoral vein when reperfusion started(I/R�Hcy). B: left kidney was subjected to sham operation, 30-min ischemia/24-h reperfusion (I/R), 30 min-ischemia/24-h reperfusion with anti-Hcy anti-bodies injected into the left renal artery immediately after the induction ofischemia (I/R�HcyAb), 30 min-ischemia/24-h reperfusion with nonspecificrabbit IgG injected into the left renal artery immediately after the induction ofischemia (I/R�IgG), or 30-min ischemia/24-h reperfusion with L-Hcy (2.5mg/kg in 250 �l) injection into the left femoral vein when reperfusion started(I/R�Hcy). Plasma creatinine level was measured and lipid peroxides inkidneys were determined by measuring the amount of MDA. Results areexpressed as means � SE (n � 4). *P � 0.05 when compared with the valueobtained from sham-operated group. #P � 0.05 when compared with the valueobtained from I/R group.
F1361ROLE OF HOMOCYSTEINE IN KIDNEY INJURY
AJP-Renal Physiol • VOL 292 • MAY 2007 • www.ajprenal.org
on Septem
ber 25, 2009 ajprenal.physiology.org
Dow
nloaded from

level in the circulation was likely a result of excess Hcyexported from the kidney during the reperfusion phase. Theelevation of Hcy levels in the kidney upon renal ischemia-reperfusion appeared to be due to impaired Hcy metabolismthrough the transsulfuration pathway. It has been reported thatin rat kidney majority of Hcy (up to 78%) is metabolized viathe transsulfuration pathway while small amount of Hcy isremethylated to methionine (13). This finding concurred withour current observation that the activity of CBS, the enzymeresponsible for the rate-limiting reaction in the transsulfurationpathway, was markedly decreased during ischemia. CBS ac-tivity remained at the diminished level throughout the reper-fusion phase. Such a reduction in CBS activity caused Hcyaccumulation in the kidney. Taken together, these resultssuggested that accumulation of Hcy in the kidney duringischemia-reperfusion was likely caused by a reduction of CBSactivity.
To the best of our knowledge, the present study demon-strated, for the first time, that Hcy is elevated in the kidneyduring ischemia-reperfusion. In summary, our results clearlydisplay a direct link between ischemia-reperfusion-inducedelevation of tissue Hcy levels and kidney injury. Anti-Hcyantibody administration offers a transient protective effect onischemia-reperfusion-induced oxidative stress and renal dys-function. Regulation of Hcy metabolism may provide somebeneficial effect by protecting kidneys from ischemia-reperfusion-induced injury.
GRANTS
This study was supported, in part, by grants from the Heart and StrokeFoundation, Natural Sciences and Engineering Research Council of Canada,and Canadian Institutes of Health Research.
REFERENCES
1. Au-Yeung KK, Woo CW, Sung FL, Yip JC, Siow YL, O K. Hyperho-mocysteinemia activates nuclear factor-kappaB in endothelial cells viaoxidative stress. Circ Res 94: 28–36, 2004.
2. Bonventre JV. Mechanisms of ischemic acute renal failure. Kidney Int 43:1160–1178, 1993.
3. Boom H, Mallat MJ, de Fijter JW, Zwinderman AH, Paul LC.Delayed graft function influences renal function, but not survival. KidneyInt 58: 859–866, 2000.
4. Bostom A, Brosnan JT, Hall B, Nadeau MR, Selhub J. Net uptake ofplasma homocysteine by the rat kidney in vivo. Atherosclerosis 116:59–62, 1995.
5. Bostom AG, Culleton BF. Hyperhomocysteinemia in chronic renal dis-ease. J Am Soc Nephrol 10: 891–900, 1999.
6. Clarke R, Daly L, Robinson K, Naughten E, Cahalane S, Fowler B,Graham I. Hyperhomocysteinemia: an independent risk factor for vascu-lar disease. N Engl J Med 324: 1149–1155, 1991.
7. Diez N, Perez R, Hurtado V, Santidrian S. Hyperhomocysteinaemiainduced by dietary folate restriction causes kidney oxidative stress in rats.Br J Nutr 94: 204–210, 2005.
8. Finkelstein JD, Kyle W, Harris BJ. Methionine metabolism in mam-mals. Regulation of homocysteine methyltransferases in rat tissue. ArchBiochem Biophys 146: 84–92, 1971.
9. Gavrieli Y, Sherman Y, Ben-Sasson SA. Identification of programmedcell death in situ via specific labeling of nuclear DNA fragmentation.J Cell Biol 119: 493–501, 1992.
10. Hassan A, Hunt BJ, O’Sullivan M, Bell R, D’Souza R, Jeffery S,Bamford JM, Markus HS. Homocysteine is a risk factor for cerebralsmall vessel disease, acting via endothelial dysfunction. Brain 127: 212–219, 2004.
11. Heinecke JW, Rosen H, Suzuki LA, Chait A. The role of sulfur-containing amino acids in superoxide production and modification of lowdensity lipoprotein by arterial smooth muscle cells. J Biol Chem 262:10098–10103, 1987.
12. Hoffer LJ, Robitaille L, Elian KM, Bank I, Hongsprabhas P, MamerOA. Plasma reduced homocysteine concentrations are increased in end-stage renal disease. Kidney Int 59: 372–377, 2001.
13. House JD, Brosnan ME, Brosnan JT. Characterization of homocysteinemetabolism in the rat kidney. Biochem J 328: 287–292, 1997.
14. Jakubowski H, Zhang L, Bardeguez A, Aviv A. Homocysteine thiolac-tone and protein homocysteinylation in human endothelial cells: implica-tions for atherosclerosis. Circ Res 87: 45–51, 2000.
15. Kery V, Bukovska G, Kraus JP. Transsulfuration depends on heme inaddition to pyridoxal 5-phosphate. Cystathionine beta-synthase is a hemeprotein. J Biol Chem 269: 25283–25288, 1994.
16. Lieberthal W, Levine JS. Mechanisms of apoptosis and its potential rolein renal tubular epithelial cell injury. Am J Physiol Renal Fluid ElectrolytePhysiol 271: F477–F488, 1996.
17. Miller C, Kennington L, Cooney R, Kohjimoto Y, Cao LC, HoneymanT, Pullman J, Jonassen J, Scheid C. Oxalate toxicity in renal epithelialcells: characteristics of apoptosis and necrosis. Toxicol Appl Pharmacol162: 132–141, 2000.
18. Misra HP. Generation of superoxide free radical during the autoxidationof thiols. J Biol Chem 249: 2151–2155, 1974.
19. Mudd SH, Finkelstein JD, Irreverre F, Laster L. Transsulfuration inmammals. Microassays and tissue distributions of three enzymes of thepathway. J Biol Chem 240: 4382–4392, 1965.
20. Ninomiya T, Kiyohara Y, Kubo M, Tanizaki Y, Tanaka K, Okubo K,Nakamura H, Hata J, Oishi Y, Kato I, Hirakata H, Iida M. Hyperho-mocysteinemia and the development of chronic kidney disease in a generalpopulation: the Hisayama study. Am J Kidney Dis 44: 437–445, 2004.
21. Noiri E, Nakao A, Uchida K, Tsukahara H, Ohno M, Fujita T,Brodsky S, Goligorsky MS. Oxidative and nitrosative stress in acuterenal ischemia. Am J Physiol Renal Physiol 281: F948–F957, 2001.
22. Ohkawa H, Ohishi N, Yagi K. Assay for lipid peroxides in animal tissuesby thiobarbituric acid reaction. Anal Biochem 95: 351–358, 1979.
23. Ploeg RJ, van Bockel JH, Langendijk PT, Groenewegen M, van derWoude FJ, Persijn GG, Thorogood J, Hermans J. Effect of preserva-tion solution on results of cadaveric kidney transplantation. The EuropeanMulticentre Study Group. Lancet 340: 129–137, 1992.
24. Saito M, Miyagawa I. Real-time monitoring of nitric oxide in ischemia-reperfusion rat kidney. Urol Res 28: 141–146, 2000.
25. Salom MG, Arregui B, Carbonell LF, Ruiz F, Gonzalez-Mora JL,Fenoy FJ. Renal ischemia induces an increase in nitric oxide levels fromtissue stores. Am J Physiol Regul Integr Comp Physiol 289: R1459–R1466, 2005.
26. Stam F, van Guldener C, Ter Wee PM, Jakobs C, de Meer K,Stehouwer CD. Effect of folic acid on methionine and homocysteinemetabolism in end-stage renal disease. Kidney Int 67: 259–264, 2005.
27. Starkebaum G, Harlan JM. Endothelial cell injury due to copper-catalyzed hydrogen peroxide generation from homocysteine. J Clin Invest77: 1370–1376, 1986.
28. Sung FL, Zhu TY, Au-Yeung KK, Siow YL, O K. Enhanced MCP-1expression during ischemia-reperfusion injury is mediated by oxidativestress and NF-kappaB. Kidney Int 62: 1160–1170, 2002.
29. Taoka S, Ohja S, Shan X, Kruger WD, Banerjee R. Evidence forheme-mediated redox regulation of human cystathionine beta-synthaseactivity. J Biol Chem 273: 25179–25184, 1998.
30. Tarpey MM, Wink DA, Grisham MB. Methods for detection of reactivemetabolites of oxygen and nitrogen: in vitro and in vivo considerations.Am J Physiol Regul Integr Comp Physiol 286: R431–R444, 2004.
31. Uerre JA, Miller CH. Preparation of L-homocysteine from L-homocys-teine thiolactone. Anal Biochem 17: 310–315, 1966.
32. Walker LM, Walker PD, Imam SZ, Ali SF, Mayeux PR. Evidence forperoxynitrite formation in renal ischemia-reperfusion injury: studies withthe inducible nitric oxide synthase inhibitor L-N(6)-(1-Iminoethyl)lysine.J Pharmacol Exp Ther 295: 417–422, 2000.
33. Wall RT, Harlan JM, Harker LA, Striker GE. Homocysteine-inducedendothelial cell injury in vitro: a model for the study of vascular injury.Thromb Res 18: 113–121, 1980.
34. Wang G, Woo CW, Sung FL, Siow YL, O K. Increased monocyteadhesion to aortic endothelium in rats with hyperhomocysteinemia: role ofchemokine and adhesion molecules. Arterioscler Thromb Vasc Biol 22:1777–1783, 2002.
35. Welch GN, Upchurch G Jr, Loscalzo J. Hyperhomocyst(e)inemiaand atherothrombosis. Ann NY Acad Sci 811: 48 –58; discussion 58 –49, 1997.
F1362 ROLE OF HOMOCYSTEINE IN KIDNEY INJURY
AJP-Renal Physiol • VOL 292 • MAY 2007 • www.ajprenal.org
on Septem
ber 25, 2009 ajprenal.physiology.org
Dow
nloaded from

36. Winkelmayer WC, Kramar R, Curhan GC, Chandraker A, Endler G,Fodinger M, Horl WH, Sunder-Plassmann G. Fasting plasma totalhomocysteine levels and mortality and allograft loss in kidney transplantrecipients: a prospective study. J Am Soc Nephrol 16: 255–260, 2005.
37. Woo CW, Siow YL, Pierce GN, Choy PC, Minuk GY, Mymin D, O K.Hyperhomocysteinemia induces hepatic cholesterol biosynthesis and lipidaccumulation via activation of transcription factors. Am J Physiol Endo-crinol Metab 288: E1002–E1010, 2005.
38. Zhang F, Siow YL, O K. Hyperhomocysteinemia activates NF-kappaBand inducible nitric oxide synthase in the kidney. Kidney Int 65: 1327–1338, 2004.
39. Zhu T, Au-Yeung KK, Siow YL, Wang G, O K. Cyclosporine Aprotects against apoptosis in ischaemic/reperfused rat kidneys. Clin ExpPharmacol Physiol 29: 852–854, 2002.
40. Zou AP, Li N, Cowley AW Jr. Production and actions of superoxide inthe renal medulla. Hypertension 37: 547–553, 2001.
F1363ROLE OF HOMOCYSTEINE IN KIDNEY INJURY
AJP-Renal Physiol • VOL 292 • MAY 2007 • www.ajprenal.org
on Septem
ber 25, 2009 ajprenal.physiology.org
Dow
nloaded from