Purification of the human blood platelet thromboxane A2/prostaglandin H2 receptor protein
Upload
khangminh22Category
view
4download
0
Indian J Physiol Phannacol1993; 37(4): 263-275
REVIEW ARTICLE
PHYSIOLOGY OF BLOOD PLATELET ACTIVATION
GUNDU H. R. RAO
Department ofLaboratory Medicine and Pathology,University ofMinnesota Medical School,UMHC Box 198420 Delaware Street S.E.,Minneapolis, Minnesota 55455 (U.S.A.)
( Received on May 10, 1993 )
Abstract: Blood platelets interact with a variety of soluble agonists such as epinephrineand adenosine diphosphale (ADP); many insoluble cell matrix components, includingcollagen and laminin, and biomateriaJs used for construction of invasive medical devices.These interactions stimulate specific receptors and glycoprotein-rich domains (integrinsand nonintegrin) on the plasma membrane and lead to the activation of intracellulareffector enzymes. The majority of regulatory events appear to require free calcium.Ionized calcium is the primary bioregulalor, and a variety of biochemical mechanismsmodulate the level and availability of free cytosolic calcium. Major enzymes that regulatethe free calcium levels via second messengers include phospholipase C, phospholipase~, and phospholipase D, together with adenylyl and guanylyl cyclases. Activation ofphospholipase C results in the hydrolysis of phosphatidyl inositol 4, 5-bisphosphateand formation of second messengers l,2-diacylglycerol and inositol l,4,5-trisphosphate(lp)). Diglyceride induces activation of protein kinase C, whereas IP) mobilizes calciumfrom internal membrance stores. Elevation of cytosolic calcium stimulates phospholipase~ and liberates arachidonic acid. Free arachidonic acid is transformed to a novelmetabolite, thromboxane A
2, by fatty acid synthetases. Thromboxane ~ is the major
metabolite of this pathway and plays a critical role in platelet recruitment, granulemobilization and secretion. Up-regulation in signalling pathways will increase the riskfor clinical complications associated with thromboembolic episodes. Down-regulationof signal transduction mechanisms may precipitate bleeding diathesis or stroke.
Key words: platelet activation signal transduction second messengers
calcium arachidonic acid phosphinositides G-proteins
INTRODUCTION
Platelets playa critical role in the recognition ofvascular injury, formation of effective hemostatic plugs,retraction of clots, and wound healing (1-14). However,when hyperactive, they can initiate events leading tomany clinical complications associated with cerebrocardiovascular problems. They lack deoxyribonucleicacid (DNA), and, therefore, do not synthesize majorbiochemical components and do not replicate (15). Theycome into the circulation well-packaged with the other
required biochemical components and have a relativelyshort half life (5-6 days).
Although completely nonsticky, in the resting slatethey can become sticky with the slightest stimulation.The degree of activation depends on the strength ofthe stimulus and the information available in theinteractive domains of the cell matrix components (16).The platelet is bestowed with multiple signal receivingmechanisms (17). It can revert back to the resting slateif the full activation is not warranted. Four well-defined
264 Rao
'*~T
Indian J Physiol Pharmacol 1993 '37(4)
Platelet Responses4) Irreversible Aggregation
tMonophasic +Biphasic
1 minute
4
..-.. /'~3) Thromboxane Synthesis and3 r ---- Seoetion of Granule Contents
j
i /. " 2 //'" 2) Development of Stickiness
~-- w1:/ I~."...........-------
Discoid Cells t" t1) Change of Shape to Spheres
"*~T - change in light transmission
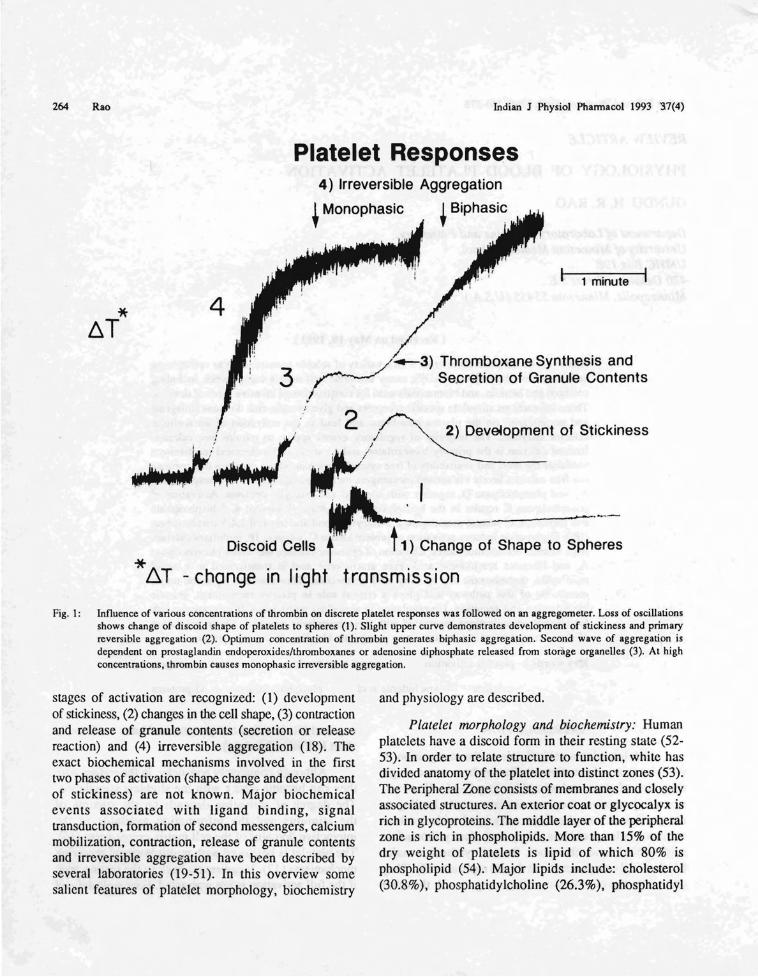
Fig. I: Influence of various concentrations of thrombin on discrete platelet responses was followed on an aggregomeler. Loss of oscillationsshows. change of di.scoid shape ?f platelets to spheres (I). Slight upper curve demonstrates development of stickiness and primaryreversible aggregallon (2). OplJ.mum concentration of thrombin generates biphasic aggregation. Second wave of aggregation isdependent.on prostagl~ndin endopcroxidcslthromboxanes or adenosine diphosphate released from storage organelles (3). At highconeentrallons, thrombm causes monophasic irreversible aggregation.
stages of activation are recognized: (1) developmentof stickiness, (2) changes in the cell shape, (3) contractionand release of granule contents (secretion or releasereaction) and (4) irreversible aggregation (18). Theexact biochemical mechanisms involved in the firsttwo phases of activation (shape change and developmentof stickiness) are not known. Major biochemicalevents associated with ligand binding, signaltransduction, formation of second messengers, calciummobilization, contraction, release of granule contentsand irreversible aggregation have been described byseveral laboratories (19-51). In this overview somesalient features of platelet morphology, biochemistry
and physiology are described.
Platelet morphology and biochemistry: Humanplatelets have a discoid form in their resting state (5253). In order to relate structure to function, white hasdivided anatomy of the platelet into distinct zones (53).The Peripheral Zone consists of membranes and closelyassociated structures. An exterior coat or glycocalyx isrich in glycoproteins. The middle layer of the peripheralzone is rich in phospholipids. More than 15% of thedry weight of platelets is lipid of which 80% isphospholipid (54). Major lipids includ~: cholesterol(30.8%), phosphatidylcholine (26.3%), phosphatidyl

Indian J Physiol Phannacol 1993; 37(4) Physiology of Blood Platelet Activation 265
ethanolamine (8.6%), sphingomyelin (11.6%),phosphatidyl serine (6.6%), and phosphatidylinositol(2.7%).
Platelet plasma membranes containtransmembrane proteins as well as glycoproteins.Glycoproteins are embedded in the lipid bilayer. Theyserve as the receptors for agonist/surface-mediatedstimuly triggering platelet activation. Platelets containintegrin as well as non-integrin domains in their plasmamembranes (55). Integrins are transmembraneglycoproteins with alpha and beta units couplednonvovalently (GPIIb-IIIa, GPla-IIa, GPIc-IIa). Theyparticipate in both cell-cell and cell-matrix interaction(55-58). In addition, there are nonintegrin glycoproteindomains on platelets capable of binding other proteinssuch as collagen and von Willebrand factor (GPIV,GPIb).
Platelets utilize energy for a variety ofbiochemical events associated with activation. Unlikemuscle. platelets do not contain creatine phosphate orcreatine phosphokinase. They contain large amountsof glycogen (30 11m/lOll cells). Platelets have aprominent glycolytic cycle and relatively fewmitochondria. Thus, the principal source of energy comesfrom the hydrolysis of ATP (59, 60).
The sol-gel zone is the matrix of the plateletcytoplasm. It contains fiber systems in various statesof polymerization. These systems support the discoidshape in resting platelets and provide a contractile systeminvolved in shape change. pseudopod extension,contraction and secretion. The contractile systemconstitutes 30% of the total platelet protein (61). Asignificant portion of this system is actin. Other proteinsof the platelet contractile system include myosin.tropomyosin, acting binding protein, a-actinin, gelsolin,profilin, vinculin and spectrin.
The organelle zone consists of granules, densebodies, peroxisomes, lysosomes, mitochondria andglycogen. This zone serves as the storage site forvarious enzymes, nonmetabolic adenine nucleotides,serotonin, a variety of proteins, calcium, andantioxidants such as ascorbic acid, glutathione andtaurine (62, 63).
The membrance system plays a major role inplatelet physiology. The dense tubular system (DTS)has been shown to be the site where calcium, animportant bioregulator, is sequestered. The DTS is alsothe site where enzymes involved in prostaglandinsynthesis are localized (64). The surface-connectedopen canalicular system (OCS) provides access to theinterior for plasma borne substances and serves as aconduit for products secreted during the release reaction(65).
Platelet responses: Platelet responses can bedivided into two categories - first, the reversibleresponses, including change of shape, development ofstickiness, adhesion, and primary reversibleaggregation; and second, contraction, release reactionand irreversible aggregation (Fig.1). When stimulatedin suspension, platelets change shape, become sticky,aggregate if brought into contact with each other bystirring, secrete the contents of their granules-densebodies, alpha granules and lysosomes. All theseresponses can be monitored on a platelet aggregometer(I 8).
Platelet shape change: Most stimuli caus~ achange in the shape of platelets. Discoidal cells, whenstimulated, extend pseudopodia or filopodia from theirperipheral zone, followed by change in their shape to aspiny sphere (dendritic) (24). The fiJ;le pseudopodia,formed upon stimulation, consist mainly of bundles ofactin filaments complexed with profilin. In addition,tropomyosin, alpha-actin, and actin binding protein alsoparticipate in the formation of pseudopodia. Inaddition to soluble physiological agonists, a variety ofcompounds such as phorbol esters and calciumionophores also cause aggregation (66, 67).Macromolecular components such as collagen fibrils,bacterial and tumor cells also cause platelet activation(67, 68). Change from a disc to an irregular sphere,constriction of microtubules and centralization of thegranules may require phosphorylation of myosin lightchain and contraction.
Platelet aggregation: Adenosine diphosphate(ADP, 311M), thrombin (0.2 U/ml), serotonin (5HT,10 11M), vasopression (100 mU/ml), epinephrine (5 11M),arachidonic acid (OA-0.6mM) and platelet activating

266 Rao Indian J Physiol Pharmacol 1993; 37(4)
factor PAP, 2 IJM) initiate aggregation within a fewseconds. By and large, all physiological agonists induceirreversible aggregation by the mediation of newlysynthesized prostaglandin endopcroxides, thromboxanesor secreted adenosine diphosphate. Two types ofresponses can be observed in vitro, a primary aggregationthat is reversible and occurs without secretion and asecondary aggregation that is accompanied by the releaseof granule contents (62, 63). Calcium or magnesiumand fibrinogen are required for agonist-mediated plateletactivation leading to aggregation (71). Several studieshave characterized a complex of two of the glyc protein(GP) on the platelet surface, GPIIb and GPIIIa as thefibrinogen receptors. Specific membrane spanningreceptors for agonists such as epinephrine, thrombin,nd thromboxane have also been characterized on the
platelet surface membrane.
Platelet adhesion: Mechanisms that underlieplatelet adhesion are n t clear (72). Platelet adhesionto glass and synthetic surfaces is probably mediatedby fibrinogen and, therefore, depends on GPIIb-IIIa.Platelets adhere to collagen, an extrac llular matrixcomponent by different specific receptors (GPIV, GPlaJIa) (73). There app ar to b a central role for vonWillebrand factor (vWF) in the adhesion of plateletsto the subendothelium (74). The vWF is rich incarbohydrates and interacts with surface membranerec ptor GPlb. Adhesion to natural and artificial surfacesis a sociated with release of granule contents and isnot inhibited by the majority of anti-platelet drugs.
Platelet release reaction: The contents of threetypes of granules are released when platelets are activatedby agonists or various natural and artificial surfaces.Weak agonists such as epinephrine, ADP and PAPdepend on formation of arachidonate metabolites,prostaglandin (PG) endoperoxides (PGG/PG~) orthromboxane for causing release of granule contents.Potent agonists such as collagen, thrombin, the calciumionophore, A23 187, and the ATPase inhibitorthapsigargin, can induce secretion of granule~independent of arachidonate metabolites (75). Thecontens of granules are released to the surface-connectedcanalicular system and extruded to the outside of thecell (62, 63).
Stimulus-response-coupling: None of thephysiological agonists can penetrate the plasma
membrane of the platelet (18, 19). They must, therefore,react at interactive domains on the platelet surfacemembrane, initiate appropriate signal, convey themessage across the plasma membrane to the interiorand activate intracellular effector enzymes (17-22).Activation of these enzymes results in the formationof second messengers that regulate the level of availablecytosolic ionized calcium (23,28,30-51).
Physiological agonists and components of theextracellular matrix such as coHagen, fibronectin andlaminin, induce platelet activation by binding at specificplasma membrane domains (receptors, integrins,nonintegrin glycoprotein-rich domains) and initiatingappropriate activating signals. The exact mechanismsinvolved in ligand binding associated signal generationat the receptor level is not clear.
Platelet receptors: A receptor for ADP has beencharacterized IIsing affinity labeled fluorosulfonylbenzyolyl adenosine (FSBA) as aggregin, a 100 Kd protein.Stimulation of this receptor induces shape change,aggregation and xposure of binding sites for fibrinogen(18, 19). Several receptors for collagen have beendescribed. In the e reports GPla and GPIV dominateas the putative receptor sites. Thromboxane ~ receptorhas been characterized using receptor antagonists andphotoaffinii.y labels. Thrombin receptor has beenidentified, although there is still considerable interestin GPV and GPIb.as possible receptor sites. Alpha
2
adrenergic receptor has been cloned and expressedand serves as interactive domains for catecholamines(43).
Signal transduction: Receptors for epinephrine,thrombin and thromboxane are members of the G-protcinreceptor family. Platelets contain phosphoinositidespecific phospholipase C(pLC), the activity of whichis modulated by G-proteins (35, 39-51). Thrombin andthromboxane also stimulate tyrosine phosphorylation,which can also initiate tyrosine phosphorylationdependent activation of PLC (34). Platelets contain highlevels of the protein tyrosine kinase (PTK) PP60C-
SRC•
Other PTKs of the SRC family, namely PP60fyn, PP55,581yn and PP61 c
•YESare also present in platelets. In addition
to these mechanisms, platelets initiate other intracellularenzymes such as phospholipase D, phospholipase A2
via G-protein mediation or independent of G-proteinsby as yet unknown mechanisms (42).

Indian J Physiol Pharmacol 1993; 37(4) Physiology of Blood Platelet Activation 267
Ionized calcium is the primary regulator of plateletactivation sequences (47,48,67, 76-80). The level offree calcium available dictates the relative state of cellactivation. In resting platelets the cytosolic-frce calciumlevels are maintained at appromimately 1 x 10-7 M byefficient calcium transport pumps and calcium-bindingproteins (80). Physiological agonists as well asantagonists exert their effect on platelets by modulatinglevels of ionized calcium through a complex chain ofsignaling mechanisms (Fig. 2). To communicate with
are then transduced across the plasma membrane toappropriate effector enzymes, such as adenylyl cyclase(AC), guanylyl cyclase (GC), phospholipase C (PLC),phospholipase ~ (PLA), and phospholipase D (PLD)via GTP-binding protein (G-protein) dependent andindependent mechanisms. Once the effector enzymesare activated, the appropriate second messengers aregenerated and these molecules further activate otherenzymes or serve as substrates for the generation ofadditional potent bioactive metabolites.
Modulation of Ionized Calciumin Human Platelets
a-AR
TxA2
ADP
Collagen
M&B22948
~-AR
PGI2
Adenosine
Forskolin
Fig. 2: Ionized calcium is the primary bioregulator. Level of cytosolic free calcium is regulated by second messengers cyclic AMP andcGMP. Agonists and antagonists act at specific plasma membrane receptors and modulate the activity of intracellular effectorenzymes, adenylyl cyclase or guanylyl cyclase and regulate the level of second messengers cAMP/cGMP.
internal effector enzymes, agonists and antagonists acton the platelet plasma membrane at discrete sites(receptors/integrins/glycoprotein-rich domains) andgcncmtc activation and inactivation signals. These signals
Platelet phospholipids: Major substrates for thephospholipases as well as for PI 3-kinase reside in themembranes (81-88). Usually threre is enough PLCactivity to hydrolyze all the inositol phospholipids. Yet,

268 Rao Indian J Physiol Phannacol 1993; 37(4)
the enzyme is unable to get access to these substrates(pI, PIP, PIP). Stimulation of cell surface by agonistsbrings about required alterations in the membrane orthe microenvironment to make these substrates availablefor the action of enymes. Furthermore, there seems tobe some specificity as to which species of the substratesis favourably predisposed for hydrolysis. Agonistmediated activation in platelets seems to favour thehydrolysis of PIP
2(84-89).
Fatty acid composition of phospholipids follow apattern. Usually stearic acid (Cia) occurs in sn-l position,arachidonic acid «20 : 4) in the sn-2 position. Inositollipids account for approximately 5-8% of the totalphospholipids, whereas phosphatidyl choline, thesubstrate for both PLD and PL~, constitute 40% ofthe platelet phospholipids (82, 82). The fatty acids aswell as the substrates for the phospholipases arepresent d in the cell to these enymes as a bilayercomprising a complex of phospholipids (pC, PE, PSand PI).
GTP-binding proteins: Activity of phospholipasesare regulated by GTP-binding proteins (40-43, 51). Bothheterotrimeric membrane associated G-proteins andmonomeric, low molecular weight (20-26 Kd), Gproteins are present in platelets. At the time of thiswriting only three G-proteins have been characterizedin terms of their function (31). They are Gs whichstimulates adenylyl cyclase, Gi which inhibits adenylylcyclase and transducin which regulates cGMPphosphodiesterase. Many other heterotrimeric G-proteinssuch as G , G , G , and G have been described. These
o p q ~
receptor coupled G-proteins share extensive homologyand contain a common a/~/y heterotrimeric structure.Alpha subunits (39-52 Kd) seem to contain interactivedomains for regulating receptors and intracellular effectorenzymes. They also have the sites for ADP ribosylationby bacterial toxins such as cholera toxin and pertussistoxin. There are large numbers of low molecular weightG-proteins in the cytosol. They are members of the rassuperfamily and belong to three major groups: ras, rho,and rab.
Current concepts on the mechanism of action ofheterotrimeric G-proteins envisages that occupation ofthe receptor by an agonist increases the rate ofdissociation of GDP from the a-subunit, facilitatingthe binding of GTP, resulting in the dissociation of G-
protein into a free a-subunit and ~/y dimer (40-43).The a-subunit with bound GTP stimulates the effectorenzyme and this stimulatory activity is terminated whenGTP is converted to GDP by the intrinsic GTPaseactivity. Alpha subunit coupled to GDP can thenreassociate with the ~/y dimer.
The G-protein that initiates hydrolysis of PI bystimulating PI-specific phospholipase C is termed G,.However, such a G-protein has not been characterizedin platelets, but conditions required to demonstrate therole of G have been described (31). According to this
p
school of thought, (a) guanine nucleotides (GN) shouldstimulate PIP
2hydrolysis in permeabilized cells, (b)
GN should mimic functional effects of agonists thatcause PI hydrolysis, (e) stable analogue of GDP (GDP~S)
should inhibit agonist-mediated hydrolysis of PI, (I)pertussis toxin should block agonist-mediated PIhydrolysis, and (g) fluoride or ALF-4 should stimulatePI hydrolysis.
G-protein-coupled receptors: Surface membrnereceptor coupled to G-protein seem to contain sevenmembrane spanning domains with three extracellularloops, equal number of intracellular loops and acytoplasmic carboxyterminal (43). There is considerablevariation in length and amino acid sequence of thethird cytoplasmic loop and the carboxyterminal. In viewof this observed variation in structure amongst knownG-protein-coupled receptors, there is considerablespeculation suggesting that these domains arc responsiblefor the selcctive interaction with the specific interactiveregions of regulatory subunits of G-proteins. In plateletessuch G-protein-coupled, membran -spanning receptorshave been characterized for agonists such as epinephrine,thrombin, and thromboxane. However, it is not clearhow other agonists such as ADP, PAF and theextracellular matrix components (collagen, fibronectin,and laminin) induce signal transduction and stimulateintracellular effector enzymes.
Phospholipid metabolism: There are four majorphospholipid species associated with platelet membranes(54). They are phosphatidyl inositol, phosphatidylcholine, phosphatidyl ethanolamine, and phosphatidylserine. Several types of phospholipases can hydrolyzephospholipids from cell membranes. Phospholipase AIspecifically removes the fatty acids from the carbon 1position, whereas phospholipase ~ exerts its effect on

Indian J Physiol Pharmacal 1993; 37(4) Physiology of Blood Platelet Activation 269
fatty acids attached to the second carbon of thephospholipids. Phospholipase C liberates phosphorylatedbases from the phospholipids, whereas phospholipaseD releases the bases such as inositol, choline,ethanolamine, and serine from the phospholipids.
In human platelets various stimuli that induceactivation stimulate phospholipase C, which is closelycoupled to specific receptors or sites of action on theplasma membrane. Platelet membranes contain largeamounts of phosphatidyl inositol (PI) and relativelysmaller amounts of phosphatidyl inositol-4 phosphate(PIP) and phosphatidyl inositol 4, 5-bisphosphate (PIP)(82, 83). All three species can serve as substrates for
the action of phospholipase C. However, physiologicalagonists seem to cause hydrolysis of PIP
2which liberates
two putative messengers, I, 2-diacyl glycerol and inositol1, 4, 5-trisphosphate (Fig. 3). If all three species arehydrolyzed, then three molecules of diglyceride areformed for every one molecule of inositol I, 4, 5tri phosphate (IPJ generated. Diglyceride produced couldbe further metabolized to form phosphatidic acid viathe action of specific kinases or acted on by a diglyceridelipase and a monoglyceride lipase release free fattyacids (82-89). Diglyceride may also playa critical rolein activation of the multifunctional enzyme protein kinaseC, whereas IP
3, which is hydrophilic, mobilizes ionized
calcium from internal membrane storage sites (17).
Phosphoinositide Metabolism and Formation of ntracelkJlar Messengers
COP - DlGlYCERIDE
PHOSPHATIDYllNOSITOl (PI)
f !
DO lIPASE
1,2 DIACYLGLYCEROL (DG)
INOSITOL 1,4.5 TRIPHOSPHATE OP3)AND CYClIC-IP3
INOSITOL 1,4 - BlSPHOSPHATE
-f !
t !
PHOSPHATIDIC ACID (PA)
PHOSPHATIDYLINOSITOL- 4.5. PHOSPHOLPASE CBISPHOSPHATE (PIP2
) .-._._._._._._._._._-_._-_._-_._.-.
PF KINASE1 1PP2 PHOSPHO-MONOESTERASE
PHOSPHATIDYLINOSITOL- 4,PHOSPHATE (PIP)
t !LYSOPHOSPHATIDIC ACID (lPA)
MONOOlYCERIDE KINASE MONOGl..YCERIJE AND_._._._._._._._._._._._._._._.-.-._. FREE FATTY ACID
Fig. 3: Agonist-mediated activation via GTP-binding proteins results in the hydrolysis of phosphatidyl inositol 4,S-bisphosphate andformation of second messengers, 1,2-diacyl glycerol (DG) and inositol 1,4,5-t.risphosphate (lP,). Diglyceride induces translocationand activation of protein kinase C, whcreas IP, mobilizes ionized calcium from internal mcmbrane storage sitcs.

270 Rao Indian J Physiol Phannacol 1993; 37(4)
Calcium metabolism: Ionized calcium is theprimary bioregulator (77-80). Resting platelets haveless than 100 nM cytosolic free calcium (80). Basedon the observation made with the use of the calciumionophore, A23187, researchers have suggested a criticalrole for calcium in platelet contraction, secretion andirreversible aggregation (67, 78). Recent studies haveprovided further support for these earlier observations(17,30,33,37). With the availability of calcium-specificfluorophores, intracellular calcium transients can bemonitored. Using such probes, several studies havedemonstrated agonist-mediated elevation in cytosoliccalcium (28, 33). Furthermore studies with agonistmediated PI hydrolysis suggest that signal transduction
from receptor to the intracellular effector enzymes iscoupled to the formation of second messengers suchas DG and IP] (37-51). Elevation of cytosolic calciumseems to promote a variety of Ca2+ -dependent eventsincluding the activation of phospholipiase Ar
Arachidonic acid metabolism: In addition tophosphatidyl inositols, phosphatidyl ethanolamine, andphosphatidyl choline may serve as a source forarachidonic acid (88). Phospholipase A
2is a membrane
associated enzyme responsible for the liberation ofarachidonic acid from these phospholipids (84, 86).Although it is capable of releasing several fatty acidsfrom the second carbon of the phospholipids, by and
Arachidonic Acid Metabolism
Membrane Phospholipids
Phospholipase A2 and C
(HETE)
6 Keto PGF;a
BLOOD VESSELS
HHT
MALONDIALDEHYDE
COOH
THROMBOXANE Az
~THROM80XANE 8.
r1'(~COOH
~~~o ''oH
IHPETE
1
ILlpoxyqenose I
PLATELETS
Ho0"
12L-Hydrolly-5,8,lO,14-eicosatetraenoic acid
Fig. 4: Elevation of cytosolic free calcium activates phospholipase A1
• Activation of this enzyme results in the liberation of arachidonicacid (AA) from membrane phospholipids. Free AA is converted by cyclooxygenases to cyclic endoperoxides, PGG1 and PGH,.These transient metabolites are further transfonned to thromboxane by thromboxane synthetase. In addition to this pathway, AA isconverted to 12-hydroperoxyeicosatetraenioc acid (12- HPETE) and 12-hydroxyeicosatetraenoic acid (12-HETE) by lipoxygenase.

Indian J Physiol Phannacol 1993; 37(4) Physiology of Blood Platelet Activation 271
large agonist-induced stimulation of platelets seems topreferentially yield arachidonic acid.The mechanismsby which phospholipase C and phospholipase A,. exerttheir specific effect on various phospholipids associatedwith the platelet membranes is not clear (85). Freearachidonic acid liberated during cell activation is themajor substrate for the synthesis of eicosanoids (88).
Arachidonic acid released during agonist-inducedactivation is rapidly converted to cyclic prostaglandinendoperoxides (pGGjPG~) by cyclooxygenases andto 12-hydroperoxy acid (HPETE) and 12-hydroxy acid12-HETE by lipoxygenase (Fig. 4). Acetylsalicylic acid(aspirin) irreversibly inhibits cyclooxygenase.Endoperoxides which are transient intermediates areas potent as thromboxane in causing platelet activation.However, these metabolites are further transformed tothromboxane A
2by thromboxane synthetase.
Thromboxane is the major metabolite of this syntheticpathway (88). In the vascular tissues prostaglandinendoperoxides (PGG/PGH2) are converted toprostacyclin (PGI) by prostacyclin synthetase.Thromboxane is a potent vasoconstrictor and plateletagonist, whereas prostacyclin is a powerful vasodilatorand platelet antagonist These two vasoactive metaboJjtesoppose each other's pharmacological actions bymodulating the activity of adenylyl cyclase and theintraeellular level of the second messenger, cAMP.This second messenger, in tum, modulates the availablelevels of the primary regulator, ionized calcium(17, 18,20,21).
Circulating adhesive proteins such as fibrinogen,components of the basement membrane, bacterialmembrane proteins, certain tumor cells, and naturaland artificial surfaces also interact with the plateletplasma membrane at discrete domains (integrin,nonintegrin). This results in activation and stimulationof various effector enzymes. Formation of secondmessengers, DG and !p], and various arachidonic acidmetabolites (PGG
2, PGH
2, TXA,.), plays a critical role
in platelet activation (development of stickiness, changeof shape, adhesion, formation of binding sites foradhesive proteins, irreversible aggregation, and secretionof granule contents). Specific mechanisms involved inthe process of centralization of granules and release oftheir contents are poorly understood.
Altered stimulus-response coupling: Severalinvestigators have attempted to correlate the in vitrofunctional response of platelets to clinical manifestationof thrombotic episodes or bleeding diathesis (7. 8, 11,12). Yet, it is not clear what functional responses ofplatelets are critical for normal hemostasis. Theavailability of functional glycoprotein Ib and lIb-lIlareceptors, ability to change shape, spread and becomesticky, irreversibly aggregate, or release granule conlcnts,are considered essential for effective hemostatic function.It has been shown that drug-induced impairment ofsignal transduction leading to altered cytosolic calciummobilization prevents agonist-mediated secretion ofgranule contents and clot retraction (33). However,unavailability or lack of significant elevation of cytosoliccalcium does not seem to prevent platelet shape change,development of stickiness, fibrinogen binding, adhesionor interaction with the vascular subendothclium.
Action of weak agonists for mediating secretionof granule contents depends upon the availability ofthromoboxane A,.. However, bovine and equine platelets,which do not aggregate in response to the action of thearachidonate metabolite, thromboxane, support normalin vivo hemostasis in these animals. Similarly, themajority of dogs have platelets that do not respond toarachidonate stimulation. On the other hand, epinephrineexposure restores the sensitivity of these refractoryplatelets so they aggregate in response to the action ofarachidonate. These observations led to the developmentof a concept explaining the role of catecholamines inmembrane and platelet function (89).
This concept was further supported by the findingthat platelets devoid of detectable cyclooxygenaseactivity supported normal hemostasis in adult individuals.Investigations from several laboratories havedemonstrated the potentiating role of adrenalin andnoradrenalin. Some reports suggest that adrenalin
, stimulation of alpha afrenoreceptors may amplify orrestore signal transduction mechanisms.
Studies by Scrutton et al and by Storomorkenand his colleagues have shown a compromised responseof platelets to adrenal in in apparently normal individuals(90,91). Weiss et al have described secretion defectsin platelets from patients with bleeding tendency (92).

272 Rao Indian J Physiol Phannacol 1993; 37(4)
White and associates have followed compromised agonistresponse of platelets of patients with diabetes, as wellas those with Hermansky-Pudlak syndrome, whoseplatelets lack calcium-rich dense bodies (93). Studiesfrom our laboratory and those of others havedemonstrated hypersensitivity of platelet of patientswith diabetes to the action of agonists. In addition, instreptozotocin-induced diabetic rats altered vascularproduction of prostacyclin and platelet synthesis ofthromboxane was demonstrated (94). However, thechanges observed in platelet and vascular metabolismof arachidonic acid metabolism was normalized by islettissue transplantation, suggesting a disease-specific effect(94). Contrary to the hyperactive state of platelet responsereported for diabetic patients, platelets of patients withHPS exhibit compromised response to the action ofagonists. This observed diminished response to agonistsis attributed to the absence of dense bodies, thatcontribute serotonin, ADP, and calcium to the releasereaction. In addition, when stimulated with weak agonistssuch as epinephrine, BPS platelets release significantlyless arachidonic acid. Studies by Hardisty et al andWare and associates have demonstrated altered signaltransduction mechanisms and suggested impairedagonist-mediated calcium mobilization as the chiefcauses of platelet dysfunction (95, 96).
A brief review of the literature related to plateletdisorders demonstrates a lack of detailed informationin the areas of signal transduction, second messengersand calcium metabolism. Investigations on both humanand animal models show that platelets can supportin vivo hemostasis effectively in spite of a lack ofsome of the essential components of signalling pathwaysobserved in human cells. Therefore, it is reasonable toconclude that platelets have multiple signalling pathwaysto regulate their discrete physiological responses.
However, loss or compromise of any signalmechanism may diminish their overall response to agiven stimulus. For instance, it has been shown in arecent stl;ldy that membrane phospholipid alterationsmay inactivate alpha
ladrenoreceptor function (97).
Modulation of this receptor plays a role in phosphatidylinositol (PI) hydrolysis. Therefore, compromisedfunction of this receptor will impair PI metabolism.Similarly, studies in our laboratory have shownthat exogenously added phospholipase A
2may
adversely affect the platelet response to epinephrine(98). Alpha
2adrenoreceptor function mediates
catecholamine responses. Alteration in this receptorfunction may compromise the mechanism of membranemodulation.
Recent studies have shown that in many malignantand inflammatory disorders plasma phospholipase A
2
activity is elevated (99). Therefore, it is possible thatvarious disease processes, oxidative stress and the normalaging process may contribute to the alterations in cellmembrane phospholipids as well as signal transductionpathways. Further detailed studies on all aspects ofsignal transduction mechanisms, second messengersystems and calcium procurement pathways in bothhuman disease models and novel animal modelsshould be explored to elucidate specific roles of theseimportant biochemical events in facilitating agonistmediated discrete functional responses in platelets(100).
ACKNOWLEDGEMENTS
The author wishes to thank Ms. Susan Schwarzefor her help in the preparation of this manuscript.
This work was supported by grants NIH HL-11880and March of Dimes FY92-0908.
REFERENCES
I. Marcus AJ, Zucker MR. The physiology of blood platelets.New York, Crune and Slrallon, 1965: 162.
2. Caen 1. Platelet aggregation. Paris, Masson and Cie, 1971:228.
3. Brinkhous KM, Shennan RW, Mostof FK. The p1atelel.Baltimore, Williams and Wilkson Co., 1971: 420.
4. Johnson SA. The circulating platelel. New York, AcademicPress, 1971: 60 I.
5. Weiss Ill. Platelets and their role in hcmostasis. Ann NewYork Acad Sci 1972; 201: 1-450.
6. Ciba Foundation. Biochcmistry and phannacology of platclets.New York, Elsevier, 1975: 352.

Indian J Physiol Pharmacol 1993; 37(4) Physiology of Blood Platelet Activation 273
7. Caen JP, Cronberg S, Kubisz P. Platelet physiology andpathology. New York, Straton Intercontinental Medical BookCo., 1977: 136.
8. deGaetano G, Garattini S. Platelets: A multidisciplinaryapproach. New York, Raven Press, 1977: 485.
9. Kwan HC, Bowie EJW. Thrombosis. Philadelphia, W.B.Saunders, 1982: 331.
10. Phillips DR, Shuman MA. Biochemistry of platelets. NewYork, Academic Press, 1986: 462.
II. Colman RW, Hirsch J, Marder VJ, Salzman EW. Hemostasisand thrombosis: basic principles and clinical practice.Philadelphia, lB. Lippincott Co., 1987: 1507.
12. Bloom AL, Thomas DP. Haemostasis and thrombosis. London,Chruchill Livingstone, 1987: 1022.
13. Marguerie G, Zwaal RFA. Biochemistry and physiopathologyof platelet membrane. L ndon, John Libbey, 1988: 189.
14. Corriveau DM, Fritsma GA. Hacmostasis and thrombosis inthe clinical laboratory. Philadelphia,l.B. Lippincott Co., 1988:443.
15. Crawford N, Scruuon Me. Biochemistry of the blood platelet.In: l-Iaemostasis and thrombosis, Bloom and Thomas, cds.,1987: 47-77.
16. Nachman RL. Thrombosis and atherogenesis: molecularconnections. Blood 1992; 79: 1897-1906.
17. Seiss W. Molecular mechanisms of platelet activation. PhysiolRev 1989; 69: 59-178.
18. Zucker MB, Nachmias VT. Platelet activation. Arteriosclerosis1985; 5: 2-18.
19. Colman RW. Platelet receptors. In: Platelets in health anddisease. Haematology Oncology Clinics ofNorth America 1990;4:27-41.
20. Haslam RJ. Signal transduction in platelet activation. In:Thrombosis and haemostasis. Verstract M, Vermylen J, LijnenHR, Arnout J, eds.). Internat Soc Thromb Haemst, Leuven,Leuven University Press, 1987: 147-173.
21. Hallam Tl, Sanchez A, Rink TJ. Stimulus-response couplingin human platelets. BiochemJ 1984; 218: 819-827.
22. Luscher EF. Mechanism of platelet aggregation. In: Antiplatelettherapy: twenty years' experience. Born GVR, Sereneri GG(eds.), Amsterdam, Excerpta Medica, 1987: 3-28.
23. Nishizuka Y. Studies and perspectives of protein kinase C.Science 1986; 233: 305-312.
24. White JG. Ultrastructural modifications in platelet membranesand cytoskeleton following activation. Blood Cells 1983; 9:237-261.
25. Hartwig JH. Mechanisms of actin rearrangements mediatingplatelet activation. J Cell Bio/ 1992; 118: 1421-1442.
26. Allen R, Zacharski L, Widirstky S, Rosenstein R. Zaitlin L.Burgess D. Transformation and motility of human platelets.J Cell Bioi 1979; 83: 126-142.
27. Nachmias VT, Yoshida K. The cytoskeleton of the bloodplatelet: a dynamic structure. Adv Cell Bioi 1988; 2: 181-211.
28. Sage S, Rink n. The kinetics of changes in intracellularcalcium concentration in Fura 2-loaded human platelets.J Bioi Chem 1987; 262: 16364-16369.
29. Stossel T. From signal to pseudopod. How cells controlcytoplasmic actin assembly. J Bioi Chem 1989; 264: 1826118264.
30. Knight DE. Calcium and diacylglycerol control of secretion.Bioscience Reports 1987; 7: 355-367.
31. Brass LF. G protein regulators of platelet activation. In: Plateletmembrane receptors. Jamieson GA (cd.), Alan R. Liss Inc.1988: 441-492.
32. Gerrard JM, McNicol A, Klassen D, Israelis Sl. Proteinphosphorylation and its relation to platelet function. In: Bloodcells and arteries in hypertension and atherosclerosis. MeyerP and Marche P (cds.), New York, Raven Press, 1989; 93114.
33. Rao GHR, Gerrard JM, Cohen I, Witkop Cl, White 10. Originand role of calcium in platelet activation-contraction-seeretioncoupling. In: Cell calcium metabolism, Gallo LL (ed.),.NewYork, Plenum Publishing, 1987: 495-505.
34. Pumigla KM, Lau L, Huang C, Burroughs S, Feinstein MB.Activation of signal transduction in platelets by tyrosinephosphate inhibitor pervanadate (vanadyl hydroperoxide).BiochemJ 1992; 286: 441-449.
35. Berridge MJ. The molecular basis of communication withinthe cell. Sci Am 1985; 253: 142-152.
36. Holmsen H. Platelet metabolism and activation. Sem Hematol1985;22:219-240.
37. Ware J, Johnson P, Smith M, Sal7.man EW. Effect of commonagonists on cytoplasmic ionized calcium concentration inplatelets. J Clin Invest 1986; 77: 878-886.
38. Lapetina EG. Inositol phospholipids and phospholipase Cin platelet receptor transduction. In: Platelet membrancereceptors. Jamieson GA (ed.), New York, Alan R, Liss, 1988:499-511.
39. Rana RS, Hokin LE. Role of phosphoinositides intransmembrane signalling. Physiol Rev ~990; 70: 115-164.
40. Kaziro Y, Itoh H, Kozasa T, Masato N, Satoh T. Structureand function of signal-transducing GTP-binding proteins. AnnRev Biochem 1991; 60: 349-400.

274 Rao Indian J Physiol Pharmacal 1993; 37(4)
41. Bimbaumer L, Abramowitz J, Brown AM. Receptor-effectorcoupling by G-proteins. Biochem Biophys Acta 1990; 1031:163-224.
42. Cockcroft S. G-protein-regulated phospholipase C, D and A2
mediated signalling in neutrophils. Biochem Biophys Acla 1992;1113: 135-160.
43. Lefkowitz RJ, Caron MG. Adrenergic receptors: models forthe study of receptors coupled to guanine nucleotide regulatoryproteins. J Bioi Chem 1988; 263: 4993-4996.
44. Berridge MI, Irvine RF. Inositol trisphosphate: a novel secondmessenger in cellular signal transduction. Nature 1984; 312:315-321.
45. Nishizuka Y. Turnover of inositol phospholipids and signaltransduction. Science 1984; 225: 1365-1370.
46. Hokin LE. Receptors and phospholinositide generated secondmessengers. Ann Rev Biochem 1985; 54: 205-235.
47. Abdel-Latiff AA. Calcium-mobilizing receptors,polyphosphoinositides, generation of second messengers andcontraction in mammalian smooth muscle: Historicalperspectives and current status. Life Sci 1989; 45: 757-786.
48. Putney JW, Takemura H, Hughes AR, Hortsman DA, ThastrupO. How does inositol phosphate regulate calcium signalling?FASEB J 1989; 3: 1899-1905.
49. Okano Y, Banno Y, Nagata K, Yamada K, Nozaya Y. Multipleforms of phosphoinositide, phospholipase C and GTP-bindingprotein in human platelets. In: The biology and medicine ofsignal transduction. Nishiguka Y (ed.), New York, RavenPress, 1990: 375-380.
50. Cau KJ, Hunyady L, Baua T. Second messengers derivedfrom inositol lipids. J Bioeng Biomed 1991; 23: 7-27.
51. Speigel AM. Heterotrimeric GTP-binding proteins: Anexpanding family of signal transducers. Med Res Rev 1992;12: 55-71.
52. Behnke O. Electron microscopic observation on the surfacecoating of human blood platelets. J Ultrastruel Res 1968; 24:51-69.
53. White JG. Platelet structural physiology: The ultrastructureof adhesion, secretion and aggregation in arterial thrombosis .Cardiovasc Clin 1987; 18: 13-23.
54. Purdon D. Phospholipid metabolism in platelets. Mod MetchPharmacol 1987; 4: 229-242.
55. Hynes RO. Integrins, a family of cell surface receptors. Cell1987;48:549-554.
56. Ruoslahti E. Integrins. J Clin Invest 1991; 87: 1-4.
57. Ruoslahti E, Pierchbacher MD. New perspectives in celladhesion. RGD and integrins. Science 1987; 238: 491-497.
58. Albelda SM, Buck CA. Integrins and other cell adhesionmolecules. FASEB J 1990; 4: 2868-2880.
59. Holmsen H, Day HJ. Adenine nucleotides and platelet function.Ser Haemalo/1971; 4: 28-58.
60. Holmsen H, Setkowsky CA, Day HJ. Effects of antimycinand 2-deoxyglucose on adenine nucleotides in hmnan platelets.Role of metabolic ATP in primary aggregation, secondaryaggregation and shape change of platelets. Biochem J 1974;144: 385-396.
61. Fox JEB. The organization of platelet contractile proteins. In:Platelet membrane glycoproteins, George IN, Nurden NJ,Phillips DR (eds.), New York, Plenum Press, 1985; 273-293.
62. Holmsen H, Day JH, Stormorken CA. 1be blood platelet releasereaction. Scand J llaenliJlol1969; 8: 1-26.
63. Day HI, Holrnsen H. Concepts of the platelet release reaction.Ser Haemalol1971; 4: 3-27.
64. Gerrard 1M, White JG, Rao GHR, Townsend D. Localizationof platelet prostaglandin production in the platelei dense tubularsystem. Am J Palho11976; 83: 283-299.
65. White JG, Oawson Cc. The surface-connected canalicularsystem of blood platelets - a fenestrated membrane system.AmJ Palho11980; 101: 353·:'64.
66. White JG, Rao GHR, Estensen RD. Investigation of the releasereaction in platelets exposed to phorbol myristate acetate. AmJ Patho11974; 75: 301-314.
67. White JG, Rao GHR, Gerrard JM. Effects of the ionophoreA23187, on blood platelets. 1. Influence on aggregation andsecretion. AmJ Palho11974; 77: 135-149.
68. Clawson CC, Rao HR, White JG. Platelet interaction withbacteria. IV. Stimulation of the release reaction. Am J Palhol1975; 81: 41l-420.
69. Zijenah LS, Barnes MJ. Platelet reactive sites in humancollagens I and m. Evidence for cell recognition sites incollagen unrelated to RGD and like sequences. ThTomb Res1990; 59: 553-566.
70. Karpatkin S, Pearlstein E. The role of platelets in tumor cellmetastasis. Annlnlern Med 1981; 95: 636-641.
71. Packham MA, Mustard J. Normal and abnormal platelet activily.In: Blood platelet function and medicinal chemistry. LassloA (ed.), New York, Elsevier Biomed, 1989,61-129.
72. Mustard IF, Packham MA. Factors influencing platelet function:Adhesion, release and aggregation. Pharmacol Rev 1970; 22:97-187.
73. Suma JJ, Nievelstein FFM, Zwaginga J, DeGroot. Adhesionof blood platelets to the extracellular matrix of cultured humanendothelial cells. In: Blood in contael with natural and artificialsurfaces. Leonard EF, Turitto vr, Vroman L (eds.), Ann NewYork Acad Sci 1987; 515: 39-52.

Indian 1 Physiol Phaffilacol 199~; 37(4) Physiology of Blood Platelet Activation 275
74. Sakariassen KS, Fressinaud E, Giffila 1, Meyer D, BaumgartnerHR. Role of platelet membrane glj'coproteins and vonwillebrand factor in adhesion of platelets to subendotheliumand collagen. In: Blood in contact with natural and artificialsurfaces. Leonard EF, Turino VT, Vroman L (eds.), Ann NewYork Acad Sci 1987; 515: 52-66.
75. Rao GHR, White IG. Platelet activating factor (PAF) causeshuman platelet aggregation through the mechanism ofmembrane modulation. Prost Leukotr Med 1982; 9: 459-472.
76. Friedman I, Detwiler TC. Stimulus-secretion coupling inplatelets. Effects of drugs on secretion of ATP. Biochem 1975;14: 1315-1320.
77. Feinman RD, Detwiler Te. Platelet secretion induced bydivalent cation ionophorcs. Nature 1974; 249: 172-173.
78. Detwiler TL, Charo IF, Feinman RD. Evidence that calciumregulates platelet function. ThrombHaemostl978; 404: 207211.
79. Feinstein Will. Release of intracellular membrane-bound calciumprecedes the onset of stimulus-induced exocytosis in platelets.Biochem Biophys Res 1980; 93: 593-600.
80. Gerrard 1M, Peterson DA, White IG. Calcium mobilization.In: Platelets in biology and pathology. Gordon 1 (cd.), Oxford,Elsevier Press, 1981: 407-436.
81. Marcus AJ, Ullman HI.. Safier LB. Lipid composition ofsubcellular particles of human blood platelets. J Lipid Res1969; 10: 108-114.
82. Menashi S, Weinstroub H, Crawford N. Characterization ofhuman platelet surface and intracellular membranes isolatedby free flow electrophoresis. J Bioi Chem 1981; 256: 40954101.
83. Lagarde M, Guichardant M, Menashi S, Crawford N. Thephospholipid and fally acid composition of human plateletsurface and intracellular membranes isolated by high voltagefree flow electrophoresis. J Bioi Chem 1982; 256: 3100-3104.
84. Bills TK, Smith JB, Silver MJ. St:!ective release of arachidonicacid from the phosphilipids of human platelets in response tothrombin. J Clin Invest 1977; 60:106.
85. Irvine RF. How is the level of free arachidonic acid controlledin mammalian cells? BiochemJ 1982; 204: 3-16.
86. Mahadevappa VG, Holub Bl. Degradation of different molecularspecies of phosphatidylinositol in thrombin-stimulatedhuman platelets. Evidence for preferemial degradation ofl-acyl-2-arachiodonyl species. J Bioi Chem 1983; 258:5337-5339.
87. Majerus PW, Wilson DB, Connolly TM, Bross TE, NeufeldEl. Phosphoinositide turnover provides a link III
stimulus-response coupling. Trends Biochem Sci 1985; 10:168-171.
88. Marcus AI. The role of lipids in platelet function: with particularreference to the arachidonic acid pathway. J Lipid Res 1978;19: 793-826.
89. Rao GHR. Epinephrine-induced membrane modulation. In:The platelet amine storage granule. Myers Ki\1', Barnes CD(eds.), Boco Raton, CRC Press, 1992: 117-141.
90. Scrullon MC, Clare KA, Bruckdorfer RR. Depressedresponsiveness to adrenalin in platelets from apparently normalhuman donor;;: a familial trait. Br J Haematol 1981; 49: 303314.
91. Stromorken H, Gogstad G, Solum NO. A new bleeding disorder:lack of platelet aggregatory response to adrenalin and lack ofsecondary aggregation to ADP and platelet activating factor(PAF). ThrombRes 1982; 29: 391-401.
92. Weiss HJ, Witte LD, Kaplan KL, Lages BA, Chernoff A,Nossel HL, Goodman DS, Baumgartner HR. Heterogeneityin storage pool deficiency: studies on granule-bound substancesin 18 patients including variants deficient in lX-granules, plateletfactor 4, ~-thrornboglobulin and platelet-derived growth factor,Blood 1979; 54: 1296-1319.
93. White JG, Rao GHR, Gerrard JM. Platelet stimulus activationcontraction-secretion coupling: a frequently fracturedchain of events. In: Hann and Taylor (eds.), The regulationof coagulation events. New York, EIsevi,er Inc., 1980:363-377.
94. Gerrard 1M, Rao GHR, Stuart MJ, White IG. Iterations inthe balance of thromboxane and prostacye1i..1 synthesis inplatelets and the vessel wall in a diabetic model. In: Theeffects of platelet active drugs on the cardiovascular system.Hirsch 1, Steele PP, Vernier RL (eds.), Boulder, Uni ersityof Colorado Press, 1981: 207 -227.
95. Hardisty RM, Machin Sl, Nokes TJC, Rink TJ, Smith SW. Anew congenital defect of platelet secretion: imp iredresponsiveness of platelets to cytoplasmic free calcium. Br JHaematol1983; 53: 543-547.
96. Ware lA, Oark BA, Smith M, Salzman EW. Abnormalitiesof cytoplasmic CA1+ .in platlets from patients with uremja.Blood 1986; 73: 172-176.
97. Shreeve SM, Valliere JE. The lX, adrenoreceptor is inactivatedby alterations in membrane phospholipids. Eur J Pharmacol1992; 226: 29-35.
98. Rao GHR, White JG. Influmce of phospholipase A1 01l humanblood platelet alpha adrenergic receptor function. Thromb Res1989; 53: 427-434.
99. Lukas R, Trif I, Cucianu M. Plasma phospholipase A, inchildren with malignant disorders. Eur J C/Ut Chem C/i"Biochem 1992; 30-127-130.
100. Rao GHR. Signal transduction second messengers and plateletfunction. J Lab Clin Med 1993; 121: 18-20.
Copyright © 2022 FDOKUMEN