
Adaptive adjustments to light in foliage and whole-plant characteristics depend on relative age in...
14
© New Phytologist (2004) 162: 683 – 696 www.newphytologist.org 683 Research Blackwell Publishing, Ltd. Adaptive adjustments to light in foliage and whole-plant characteristics depend on relative age in the perennial herb Leontodon hispidus Ülo Niinemets Department of Plant Physiology, University of Tartu, Riia 23, Tartu EE 51010, Estonia and Centro di Ecologia Alpina, Viote del Monte Bondone, 38040 (TN), Italy Summary • Growth form gradually changes during ontogeny through juvenile, reproductive, and senescent phases in perennial herbs, but resulting effects on plant structure, functioning and role in the community are poorly characterized. Here, biomass distribution, leaf structure and nutrients were studied in relation to irradiance and age state in the temperate meadow perennial Leontodon hispidus in the field. • Plant ontogeny modified plastic responses to irradiance with most plastic changes in leaf dry mass per area in reproductive and most plastic LAR alterations in young individuals. Due to age-dependent changes, light interception efficiency was maximised in young, and photosynthetic potential in reproductive specimens, while senescent plants had inferior light interception and use, but were probably more competitive for below-ground resources. • Species age spectrum shifted to senescent phases, and the fractional biomass to below-ground in sites with higher species richness. • These results collectively demonstrate a strong effect of ontogeny on dry matter partitioning, leaf architecture and functioning, and plastic adjustments in these traits and suggest that species competitive potentials change through the ontogeny. Key words: biomass partitioning, growth form, leaf architecture, nitrogen, ontogeny, species richness, Leontodon hispidus . © New Phytologist (2004) 162 : 683–696 Author for correspondence: Ülo Niinemets Tel: 372 3 366050 Fax: 372 7 366021 Email: [email protected] Received: 5 January 2004 Accepted: 17 February 2004 doi: 10.1111/j.1469-8137.2004.01071.x Introduction There are significant modifications in woody plant structure and physiological performance with their age (Day et al., 1997; Grubb, 1998; Bond, 2000; Niinemets, 2002). Specifically, young individuals have thinner leaves, and larger biomass investment in leaves, allowing maximisation of whole plant light interception efficiency, while older individuals have thicker leaves and allocate greater fractions of biomass to stem and roots (Niinemets, 2002). Biomass partitioning in herbaceous perennial species also varies largely with ageing (Rice & Bazzaz, 1989b; Coleman et al ., 1994). Given that species competitive potential depends strongly on leaf structure and biomass partitioning (Carlen et al., 1999; Ryser & Eek, 2000; Schippers & Kropff, 2001), such age- dependent changes may significantly alter species interactions in grassland vegetation (Grubb, 1977, 1998). However, structural rearrangements in herbaceous species due to ontogeny have received little attention, and species are considered as discrete entities with time-independent responses to the environment in current theoretical vegetation models (Huston & Deangelis, 1994; Tilman et al., 1997; Loreau, 1998). Most studies of age-dependent changes in grassland com- munities use absolute plant age as a variable driving temporal changes in community structure and performance (Berendse, 1993; Wedin & Tilman, 1993; Coleman et al., 1994; van der Maarel, 1996). However, the absolute age itself may be a poor predictor of competitive success in vegetation communities, because changes in physiological performance and competitive potential are a function of age-dependent morpho-physiological changes through prereproductive, reproductive and post- reproductive stages that may proceed with a differing rate depending on species and environmental conditions (Gatsuk et al., 1980; Rabotnov, 1985; Falinska, 1995). Thus, classifica- tion of the ontogeny between distinct morpho-physiological
Transcript of Adaptive adjustments to light in foliage and whole-plant characteristics depend on relative age in...

©
New Phytologist
(2004)
162
: 683–696
www.newphytologist.org
683
Research
Blackwell Publishing, Ltd.
Adaptive adjustments to light in foliage and whole-plant characteristics depend on relative age in the perennial herb
Leontodon hispidus
Ülo Niinemets
Department of Plant Physiology, University of Tartu, Riia 23, Tartu EE 51010, Estonia and Centro di Ecologia Alpina, Viote del Monte Bondone, 38040 (TN), Italy
Summary
• Growth form gradually changes during ontogeny through juvenile, reproductive,and senescent phases in perennial herbs, but resulting effects on plant structure,functioning and role in the community are poorly characterized. Here, biomassdistribution, leaf structure and nutrients were studied in relation to irradiance andage state in the temperate meadow perennial
Leontodon hispidus
in the field.• Plant ontogeny modified plastic responses to irradiance with most plastic changesin leaf dry mass per area in reproductive and most plastic LAR alterations inyoung individuals. Due to age-dependent changes, light interception efficiencywas maximised in young, and photosynthetic potential in reproductive specimens,while senescent plants had inferior light interception and use, but were probablymore competitive for below-ground resources.• Species age spectrum shifted to senescent phases, and the fractional biomass tobelow-ground in sites with higher species richness.• These results collectively demonstrate a strong effect of ontogeny on dry matterpartitioning, leaf architecture and functioning, and plastic adjustments in these traitsand suggest that species competitive potentials change through the ontogeny.
Key words:
biomass partitioning, growth form, leaf architecture, nitrogen, ontogeny,species richness,
Leontodon hispidus
.
©
New Phytologist
(2004)
162
: 683–696
Author for correspondence:
Ülo NiinemetsTel: 372 3 366050Fax: 372 7 366021Email: [email protected]
Received:
5 January 2004
Accepted:
17 February 2004
doi: 10.1111/j.1469-8137.2004.01071.x
Introduction
There are significant modifications in woody plant structureand physiological performance with their age (Day
et al
., 1997;Grubb, 1998; Bond, 2000; Niinemets, 2002). Specifically, youngindividuals have thinner leaves, and larger biomass investmentin leaves, allowing maximisation of whole plant light interceptionefficiency, while older individuals have thicker leaves and allocategreater fractions of biomass to stem and roots (Niinemets, 2002).Biomass partitioning in herbaceous perennial species also varieslargely with ageing (Rice & Bazzaz, 1989b; Coleman
et al
., 1994).Given that species competitive potential depends strongly onleaf structure and biomass partitioning (Carlen
et al
., 1999;Ryser & Eek, 2000; Schippers & Kropff, 2001), such age-dependent changes may significantly alter species interactionsin grassland vegetation (Grubb, 1977, 1998). However, structuralrearrangements in herbaceous species due to ontogeny have
received little attention, and species are considered as discreteentities with time-independent responses to the environmentin current theoretical vegetation models (Huston & Deangelis,1994; Tilman
et al
., 1997; Loreau, 1998).Most studies of age-dependent changes in grassland com-
munities use absolute plant age as a variable driving temporalchanges in community structure and performance (Berendse,1993; Wedin & Tilman, 1993; Coleman
et al
., 1994; van derMaarel, 1996). However, the absolute age itself may be a poorpredictor of competitive success in vegetation communities,because changes in physiological performance and competitivepotential are a function of age-dependent morpho-physiologicalchanges through prereproductive, reproductive and post-reproductive stages that may proceed with a differing ratedepending on species and environmental conditions (Gatsuk
et al
., 1980; Rabotnov, 1985; Falinska, 1995). Thus, classifica-tion of the ontogeny between distinct morpho-physiological

www.newphytologist.org
©
New Phytologist
(2004)
162
: 683–696
Research684
age states resulting in a relative age scale may be more inform-ative in demographic and succession studies than using theabsolute age (Orive, 1995). Rabotnov (for review Gatsuk
et al
.,1980; Rabotnov, 1985) defined a series of qualitative plantontogenetic phases that are characterized by a unique com-bination of qualitative and quantitative traits. Since then,this and similar qualitative rankings have been extensivelyused in phytosociology (Uranov, 1977; Gatsuk
et al
., 1980;Serebryakova
et al
., 1983; Zhukova & Ermakova, 1985;Zaugolnova
et al
., 1991), but the morpho-physiologicalmodifications in plants during ontogeny have not beeninvestigated quantitatively.
The primary task of this study was to characterize leafstructure and chemistry, and biomass partitioning in rela-tion to ontogenetic age state in the perennial herb
Leontodonhispidus
L. in the field. Specifically, the working hypothesiswas that juvenile establishing plants form a larger leaf areaper total biomass, while older individuals have larger biomassinvestments in below-ground parts, leading to competitivestrategy changes as outlined by Grubb (1998).
In addition to ontogeny, whole-plant biomass distributionand foliar architecture also depend on environmentalcharacteristics, in particular on incident light conditions (e.g.Meziane & Shipley, 1999; Poorter & Nagel, 2000). Laborat-ory growth experiments have provided a solid theoreticalframework of the role of plastic adjustment to light in youngherbaceous species (Shipley, 2000, 2002), but surprisingly fewfield experiments have been conducted along environmentalgradients. Given the important modifications in structure andallocation due to ontogeny, and that plasticity may vary withplant size and age (Rice & Bazzaz, 1989a,b; Schmid & Bazzaz,1990), I also hypothesised that the plasticity for leaf structureand biomass allocation changes is larger in juvenile than inreproductive and senescent plants. To test this hypothesis, plantswere sampled along a natural light gradient in a wooded meadow.The results demonstrate significant variation in plastic modi-fications in leaf morphology and whole plant leaf area to irra-diance changes as well as gradual shifts in biomass partitioningbetween above- and below-ground parts with relative age.
Because the organs and tissues with varying nutrient contentsdecompose with a different rate constant, shifts in biomasspartitioning patterns are important for understanding com-munity carbon and nutrient turnover rates. Such changes innutrient mineralization time constants significantly alter com-munity species composition and community sustainability(Wedin, 1995; Tilman
et al
., 1996; Wedin & Tilman, 1996;Hooper & Vitousek, 1998), but the possible linkages betweenchanges in species relative age spectrum and local species rich-ness have not been investigated. Given that the availability ofregeneration niches decreases with increasing species diversity(Grubb, 1977; Tilman, 1997), species-rich communities arelikely to accumulate ontogenetically older individuals withpotentially differing functional responses to environmentalvariation. As a further task of this study, I examined
L. hispidus
relative age spectrum along a natural gradient of species diversity,hypothesising that in more diverse sites, where the availabilityof regeneration niches is lower, the age spectrum is shiftedtowards ontogenetically older individuals.
Materials and Methods
The species and the age states
L. hispidus
is a dominant perennial species in calcareous temperatemeadow communities that invests large fractions of totalbiomass below-ground in rhizomes and roots. Both vegetativeand generative reproduction are important means of dispersaland population maintenance in its natural sites. In case of veget-ative reproduction, the ramets generally become separated in theyear they form or in the following year.
Methodology for age state (relative age) separation using aseries of qualitative and quantitative characteristics is providedin detail in Gatsuk
et al
. (1980), and age state descriptions fora large number of species are reported by Serebryakova
et al
.(1983) and Zhukova (1987). Plants are first divided intovegetative and reproductive individuals, and prereproductive( juvenile, immature, and virginile) and postreproductive(subsenile and senile) individuals according to the presenceand size of the rhizome (Fig. 1). Plants belonging to thejuvenile to virginile age states are always of seed origin, whilereproductive to senile age states may be of seed or rhizomeorigin (Zhukova & Ermakova, 1985). Juveniles of
L. hispidus
have nontoothed leaves, and lack a visible rhizome. Immaturesalso possess nontoothed leaves, but have a weakly developedrhizome. Virginiles already have adult foliage and a rhizome,but lack generative organs. Young reproductive plants (G1)consist of one ramet with a nonbranched generative shoot,while mature reproductive individuals (G2) generally consistof several connected ramets, each of which bears branched ornonbranched generative shoots. The rhizomes of old repro-ductive plants (G3) bear the remnants of died reproductiveramets, and have a single generative nonbranched shoot.Senile individuals are distinguished from subseniles by havingnontoothed juvenile foliage. Starting from the age state G3,further ramets are rarely formed.
Although it may be difficult to relate the relative age toabsolute age because of site-to-site differences in ontogeneticdevelopment (Zhukova, 1987), long-term field studiesdemonstrate that the whole ontogenetic cycle takes on average5–15 yr in temperate perennial species including
Leontodonhispidus
(Zhukova, 1987). According to the observationsmade at the site, seed-originated individuals of
L. hispidus
mayundergo a change from juvenile-to-immature phase during thefirst growing season, but reaching the first reproductive phase(G1) requires at least three growing seasons and postrepro-ductive age states at least five growing seasons. Due to the longduration of the ontogeny, the overall effect of seasonality onage state spectrum is minor.

©
New Phytologist
(2004)
162
: 683–696
www.newphytologist.org
Research 685
Study site
The study was conducted in the Laelatu wooded meadow,Estonia (58
°
36
′
-N, 23
°
35
′
-E; elevation 1–4 m above sealevel; Kull & Zobel, 1991). Estonian coastal meadows arehistorically young, being formed as the result of a gradualrise of land (
c
. 1 cm yr
−
1
) and concomitant recession of theseashore during the last 200–2000 yr. These communitiesare open woodlands, in which woody species colonization iscontrolled by regular mowing (Hæggström, 1983; Kukk &Kull, 1997), while the potential natural vegetation at thesecoastal areas would be broad-leaf deciduous forest. Due toabandonment of the traditional management practices in the1950s and 1960s, many of the wooded meadows turned toearly successional forests. In the specific part of the study areawhere the current experiment was conducted, the woodedmeadow community had been gradually restored 15–20 yrbefore the experiment.
Because both meadow and forest understory species growintermixed in these communities, the species richness is high,with more than 60 vascular plant species per one square meter(Kull & Zobel, 1991; Kukk & Kull, 1997). Apart from
L.hispidus
, other dominants of the herb layer are
Briza media
L.,
Carex flacca
Schreb.,
Carex tomentosa
L.,
Festuca rubra
L.,
Filipendula hexapetala
Gil.,
Scorzonera humilis
L. and
Sesleriacoerulea
(L.) Ard. (Kukk & Kull, 1997). The sparse shrub layermainly consists of 2–4-m-tall
Cornus sanguinea
L. and
Corylusavellana
L., while solitary or grouped individuals of
Betulapendula
Roth.,
Fraxinus excelsior
L. and
Quercus robur
L.constitute the tree layer (height 6–12 m). In the wooded
meadow, the overall coverage of shrubs and trees generallyvaries between 20 and 40%. However, the wooded meadowgradually changes to a broad-leaf deciduous forest, providinghabitats of
L. hispidus
with a high level of canopy shading.The soil is a rendzic leptosol that consists of two hori-
zons. A thin (17–22 cm) humus horizon (A) is lying dir-ectly on top of calcareous marine sediments and glacialmoraines (C horizon). The bedrock, a Silurian limestone, isdeeper than 1 m. Soil pH is neutral to alkaline (average
±
SE
=
6.92
±
0.02, range 6.7–7.5), and has high organic matter(humus concentration
=
9.06
±
0.29%) and total nitrogenconcentrations (0.59
±
0.01%). According to fertilisationexperiments, the herbaceous biomass is positively affected byboth N and P addition, with a strong interactive effect ofsimultaneous addition of N and P (Ü. Niinemets & K. Kull,unpublished data).
Plant sampling, and determination of incident irradiance
Plants were sampled along a gradient of incident irradiancefrom an essentially open grassland to deeply shaded forestenvironments. The size of the sample plots was 0.25 m
2
(0.5
×
0.5 m). All individuals of
L. hispidus
were carefullyexcavated from each plot, divided between the age states andtransported to the laboratory for morphological measure-ments. All reproductive-stage plants of
L. hispidus
floweredduring a 2-wk period in June, and there was no furtherformation of generative organs later in the season. Thesampling was conducted during peak flowering in mid-June,
Fig. 1 Illustration of representative plants of different age states of the perennial herb Leontodon hispidus. The age states are defined according to Gatsuk et al. (1980). According to this classification, the ontogeny is divided between juvenile (Juv), immature (Im), virginile (Vir), reproductive (young – G1, mature – G2 and old – G3), subsenile (Subs) and senile (Sen) ontogenetic phases. Each phase is characterized by a specific combination of qualitative and quantitative traits (presence of flowers, size, shape and number of leaves, balance between living and dead structures). Age states were numbered consecutively (1 = Juv, 8 = Sen) resulting in a relative age scale.

www.newphytologist.org
©
New Phytologist
(2004)
162
: 683–696
Research686
and after flowering in mid-July. At both sampling dates,
L. hispidus
from 24 plots was analysed. Sample plots wereselected randomly in every light environment, and differentplots were sampled at both sampling dates, resulting in a totalof 48 sampling plots.
Hemispherical photographs were taken above the herblayer of each sample plot to characterize incident irradiance.The fraction of penetrating diffuse irradiance (
I
) was calcu-lated from the hemispherical photographs for uniformly over-cast sky conditions as in Anderson (1964). This relative indexis used as an estimate of overall light availability. Because thetree and shrub foliage is formed as a single flush in early May,and the tree leaves are fully expanded by the end of May withno further changes in total leaf area and canopy extension,light conditions remain stable for the rest of the season. By thetime the tree foliage started to develop,
L. hispidus
individualshad one or two leaves, while the major development of theherb foliage proceeded together with or after the overstorydevelopment. Given the life-span of
L. hispidus
leaves of 3–4 wk, the leaves formed before the full development ofoverstory were senescent by the time of sampling.
Although sampling in natural gradients may always lead toconfounding the effects of plasticity and evolutionary adapta-tions (Valladares
et al
., 2002), the gap-understory gradientinvestigated in the current study was the result of manage-ment history during last 15–20 yr only, and the plots wereseparated by relatively short distances (at most 200 m) alongthe environmental gradient. Given the overall young age ofthese coastal communities, it is unlikely that the plantssampled along this gradient had developed any evolutionaryadaptations to their local light environment.
Species diversity and Ellenberg’s moisture and nitrogen numbers
Species richness was defined as the number of species in each0.5
×
0.5 m sample plot. Altogether, 111 vascular plant speciesgrew in various plots sampled for
L. hispidus
. The coverage ofevery species on the plot was also estimated, and these data wereused to calculate the coverage-weighted values of Ellenberg’smoisture and nitrogen numbers (Ellenberg, 1991) for everyplot as in ter Braak & Gremmen (1987) and Reif
et al
. (1985).Overall, these indices provide reliable measures of soil long-term water and nutrient status (Ertsen
et al
., 1998), and wereused in the current study to check for the possible soilgradients interacting with light availability.
Morphological measurements, and chemical analyses
All plants were divided into roots, rhizomes, and leaves. Ingenerative (G1–G3, Fig. 1) plants, flower stalks and flowerswere also separately analysed. Leaf area was measuredwith a computer digitizer (QD-1212, QTronix, Taiwan) andself-developed computer software. Dry mass of all biomass
fractions was determined after drying to a constant mass at70
°
C. The contribution of each age state to total
L. hispidus
leafarea index (LAI
L
) was calculated as the leaf area of all
L. hispidus
individuals from a given age state divided by the plot area.Nitrogen content was determined by standard Kjeldahl
method for all age states. Soluble carbohydrate and starchcontents were further analysed in virginiles to characterize thestorage and cumulative photosynthesis in plants growingin different light microenvironments using the anthrone methodas in Niinemets (1995). Chemical analyses were carriedout separately for leaves, and pooled fractions of roots andrhizomes as well as flower stalks and flowers. Total plant nitro-gen content was calculated using the dry mass of differentbiomass fractions and their nitrogen concentrations, allowingestimation of the fractional distribution of total nitrogenbetween various organs.
Simulation of intercepted light
The potential amount of intercepted light was calculatedaccording to the standard Lambert-Beer approach (Campbell& Norman, 1989), assuming that all leaves have a fixed anglewith respect to horizontal,
φ
M
(i.e. that the leaf angular distri-bution is conical) as is typical in rosette plants, and assumingthat leaf area dispersion is random. According to measurements,
φ
M
is 65
±
10
°
, and varies to a minor degree among the agestates. Thus, it was held constant in these simulations.Because the daily amount of light is the integrated sum overall zenith angles, plant potential light interception capacity isrelatively insensitive to the exact values of leaf inclinationangle (Campbell & Norman, 1989). Plant leaf area index(LAI) was determined from
φ
M
and the average number andsize of the leaves for each age state. The computations wereconducted for uniformly overcast sky conditions, and fulldetails are provided in Campbell & Norman (1989). Totalpotential daily intercepted light per plant dry mass (mol g
−
1
d
−
1
) is given as:
Eqn 1
where
Q
A
is the amount of light intercepted per unit groundarea, and LAR is the leaf area ratio (total leaf area per unit totalplant dry mass).
Plot age index
Using the number of plants belonging to each age state, Icalculated for each plot an age index (
∆
, Zhukova & Ermakova,1985) that increases with the overall age state of thepopulation:
Eqn 2
MA
LAILAR =
∆ ==
=
=
=
∑ ∑k m ki ii
i n
ii
i n
1 1

©
New Phytologist
(2004)
162
: 683–696
www.newphytologist.org
Research 687
where
ki is the number of individuals with an age state i, andmi is the weighting factor for each age state. The weightingfactor mi assumes that population ageing can be described bya logistic function (Partridge & Mangel, 1999) and is calculatedas (Uranov, 1977):
Eqn 3
where the index λ characterizes relative age ( Juv = −3.5,Im = −2.5 … Sen = 3.5; Fig. 1 for definition of age states).Although the weighting coefficients in Eqn 3 may bearbitrary, the index ∆ is a useful measure of the weighted ageof the population, provided the mortality rate increases withincreasing age of the individuals (Partridge & Mangel, 1999).With this set of the relative age coefficients, the weighted ageof the population at the limit (all plants are senile) is equalto 0.97.
Statistical analyses
Linear and second-order polynomial regressions were employedto test for the statistical significance of the effect of irradianceand age state on measured foliar and whole plant variables.Linear regressions between the variables and irradiance fordifferent age states were compared by covariation analyses(). A separate slope model was used first to test fordifferences in slope. Whenever the interaction term wasnot significant, the analysis was continued according to acommon slope model to test for intercept differences. Signific-ant differences between the intercepts were determined bythe Bonferroni test. The same approach was used to comparethe biomass partitioning with age state relations for Juneand July.
Although the age state is a ranked variable, it was con-sidered continuous in the statistical analyses to highlightthe trends in plant functioning with ontogeny. Given thatalternative analyses using Spearman rank correlation, andKruskal–Wallis nonparametric resulted in identicalconclusions with respect to the statistical significance of theresults, the conclusions are not biased by analysis simpli-fications. All relationships were considered significant atP < 0.05.
Results
Characterisation of the incident light gradient
Across the plots sampled, there was a weak negative cor-relation (r2 = 0.19, P < 0.05) between the Ellenberg’s averagemoisture number and relative incident irradiance (I ), but nocorrelation (r 2 = 0.04, P > 0.3) between the nitrogen numberand I. This suggests that variation in light was the primaryenvironmental gradient in our study.
Although the possible differences in shading within theherbaceous layer were not characterized, plant coverage, heightand productivity were only weakly correlated with incidentirradiance according to previous studies (Eek & Zobel,2001). This weak relationship between productivity and incid-ent irradiance was due to strong control of productivity byoverall nutrient shortage in this site, as has been generallyobserved in other natural grasslands (Semmartin & Oester-held, 2001). Furthermore, total plant coverage was independ-ent of incident irradiance in the current study as well (r2 =0.02, P > 0.4), indicating that shading within the herb layerdid not vary along the gradient of relative irradiance. Thus, Iargue that the index I provides a robust measure of plot-to-plot differences in light availability.
Distribution of L. hispidus along microsites of differing light availability
The fraction of total cover due to L. hispidus varied from 0.06to 0.38 (average ± SD = 0.18 ± 0.09), and this fraction wasindependent of incident irradiance (I, r2 = 0.04, P > 0.3). Theplot leaf area index due to L. hispidus (LAIL) ranged from 0.14to 0.65 m2 m−2 (average ± SD = 0.41 ± 0.15), and was notassociated with irradiance (r2 = 0.01, P > 0.7) suggesting thatthe competitiveness of L. hispidus did not vary along thelight gradient. Irradiance also did not significantly affectthe age index (Eqn. 2; r2 = 0.01, P > 0.6), and contributionsof young ( juveniles, immatures, virginiles; Fig. 1 for defini-tion of age states), reproductive (G1, G2, G3) and old(subseniles, seniles) individuals to LAIL (r2 = 0.00, P > 0.9for all correlations). These data collectively indicate thatthe relative age spectrum of L. hispidus did not depend onirradiance.
Dependence of leaf morphology and chemistry on irradiance and age state
Leaf dry mass per unit leaf area (MA) increased with increasingI in L. hispidus individuals of all relative ages (Fig. 2). Theintercept of MA vs I relations consistently increased withincreasing relative age, while the slope of MA vs I peaked inreproductive plants and decreased with further increases inrelative age (Fig. 3).
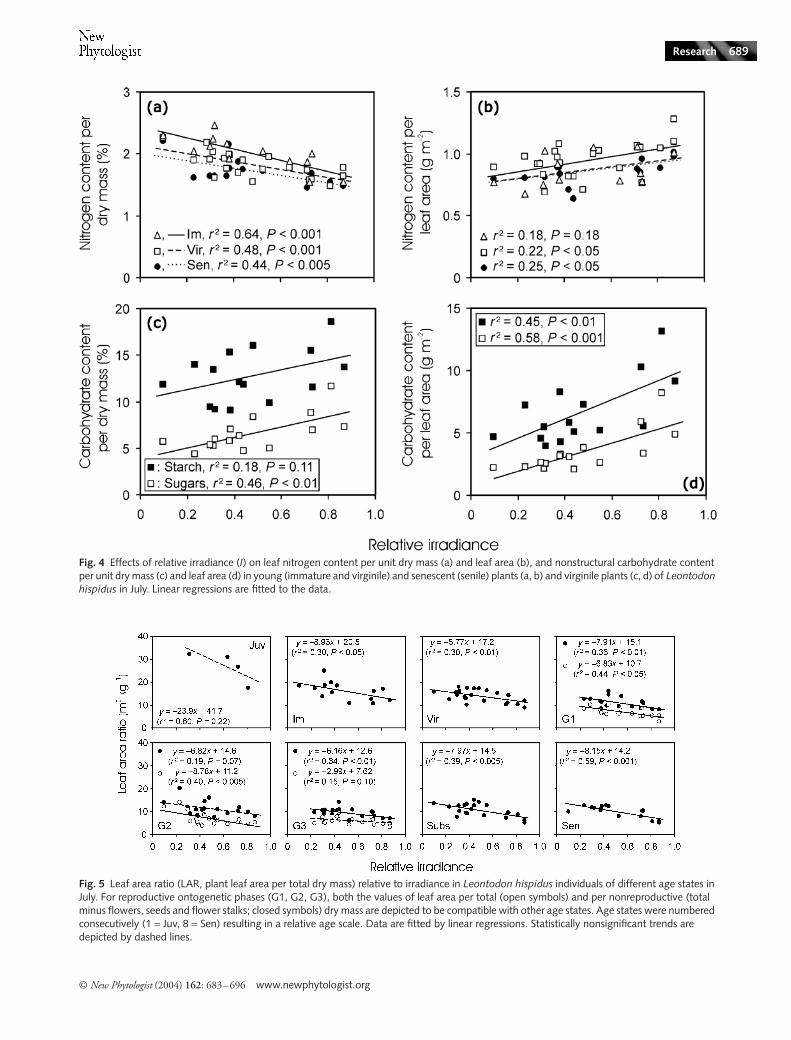
Leaf nitrogen contents of reproductive individuals wereintermediate to those of the two extremes, immatures andseniles. Thus, in the following, we demonstrate the trendswith, and differences between, the extremes. Leaf nitrogencontent per unit dry mass (NM) was negatively correlated withirradiance (Fig. 4a). However, nitrogen content per unit leafarea (NA) increased with I, except for immatures (Fig. 4b),suggesting that foliage photosynthetic potentials per areascaled positively with I.
According to separate slope analyses, NM and NAversus irradiance (Fig. 4a,b) slopes did not differ between the
mei
=
+ −1
1 λ

www.newphytologist.org © New Phytologist (2004) 162: 683–696
Research688
age states (P > 0.4). Common slope further demon-strated that immatures had a larger NM than either senileor virginile plants (P < 0.001), and virginiles had a larger NAthan seniles (P < 0.01). Thus, younger plants tended to befunctionally more active than older individuals.
Carbohydrate content was analysed in virginiles. Solublecarbohydrate contents per unit dry mass (Fig. 4c) and area(Fig. 4d) were strongly correlated with I, demonstrating thatcumulative daily photosynthesis increased with irradiance.Increases in starch content with irradiance (Fig. 4c,d) indicatean overall larger storage of photosynthates at higher light.
Leaf area construction, average and total leaf area and light interception versus irradiance and age
Leaf area ratio (LAR, total plant leaf area per total drymass) was negatively correlated with irradiance for all agestates (Fig. 5). Leaf area per plant (AT) versus I relationswere negative as well (r2 = 0.13–0.61), but not statisticallysignificant for the G3 age state (r2 = 0.13, P = 0.15).
The intercept of LAR versus I relationship was larger inyounger individuals (Fig. 6a), demonstrating that youngerplants supported a larger foliar area for a common totalbiomass. Furthermore, the slope of LAR versus I was steeper inyounger plants (Fig. 6a), suggesting that these plants exhibitmore plasticity in their biomass investments in leaf areaconstruction with changes in light regime.
Average leaf area was small in young plants, peaking invirginile and reproductive plants and decreasing in senescentstages (Fig. 6b). This pattern was essentially the same for leafnumber per plant (r2 = 0.88, P < 0.001). As the result of thesetrends, the intercept of AT versus I relation peaked in repro-ductive individuals (Fig. 6c). The slope of AT versus I relationwas also steepest, that is most negative, for the reproductiveage states (Fig. 6c). Accordingly, light-dependent changes intotal foliar area were greatest in largest plants on a wholeplant basis, despite the younger individuals producing a moreextensive foliar area for a given total plant mass.
Plant light interception capacity was largest in reproductiveindividuals due to an overall larger plant leaf area. However,the light interception capacity standardized per unit dry mass(Eqn 1, QM) to account for the size effects was the largest inyoung age states (Fig. 6d). Somewhat lower QM in reproductive
Fig. 2 The relationship between leaf dry mass per unit leaf area and relative irradiance (I) in Leontodon hispidus of different age states in July I is an index that characterizes the amount of incident irradiance relative to a completely open location, and varies due to the degree of shading by woody vegetation. Age states were numbered consecutively (1 = Juv, 8 = Sen) resulting in a relative age scale (Fig. 1). Data are fitted by linear regressions. A statistically nonsignificant trend for juvenile plants is depicted by a dashed line.
Fig. 3 Age-dependent changes in the intercepts and slopes of leaf dry mass per unit area (MA) versus relative irradiance relations in Leontodon hispidus in July (Fig. 2). Data were fitted by second-order polynomial regressions. Age states were numbered consecutively (1 = Juv, 8 = Sen) resulting in a relative age scale.
© New Phytologist (2004) 162: 683–696 www.newphytologist.org
Research 689
Fig. 4 Effects of relative irradiance (I) on leaf nitrogen content per unit dry mass (a) and leaf area (b), and nonstructural carbohydrate content per unit dry mass (c) and leaf area (d) in young (immature and virginile) and senescent (senile) plants (a, b) and virginile plants (c, d) of Leontodon hispidus in July. Linear regressions are fitted to the data.
Fig. 5 Leaf area ratio (LAR, plant leaf area per total dry mass) relative to irradiance in Leontodon hispidus individuals of different age states in July. For reproductive ontogenetic phases (G1, G2, G3), both the values of leaf area per total (open symbols) and per nonreproductive (total minus flowers, seeds and flower stalks; closed symbols) dry mass are depicted to be compatible with other age states. Age states were numbered consecutively (1 = Juv, 8 = Sen) resulting in a relative age scale. Data are fitted by linear regressions. Statistically nonsignificant trends are depicted by dashed lines.

www.newphytologist.org © New Phytologist (2004) 162: 683–696
Research690
than in senescent stages results from large plant biomassinvestment in flowers and flower stalks in reproductive agestates.
Dry matter partitioning in relation to irradiance and ontogeny
Plant dry matter partitioning studied at two different pheno-logical stages, during flowering in June and after flowering inJuly, was weakly dependent on incident irradiance. Leaf massratio (ML : MT, dry mass of leaves per total dry mass) wasnegatively associated with I for G1 (r 2 = 0.39, P < 0.01) andfor G3 (r 2 = 0.50, P < 0.05) plants in June, but there wereno significant relationships for any age states in July. Below-ground biomass (MRh+R, rhizome and roots) to total (MT)plant dry mass ratio decreased with increasing I in G2 plantsin July (r 2 = 0.26, P < 0.05), and was independent of I in allother cases. Components of MRh+R : MT ratio, rhizome tototal (MRh : MT) and root to total (MR : MT) ratios were also
generally not related to I. The only exception was a positivecorrelation between MR : MT and I in G1 plants in July(r 2 = 0.34, P < 0.01). The nitrogen content of below-grounddry mass was always negatively associated with I (Fig. 7),further suggesting that plant photosynthesis and growth wasmore nutrient-limited at higher irradiance (cf. Fig. 4a).
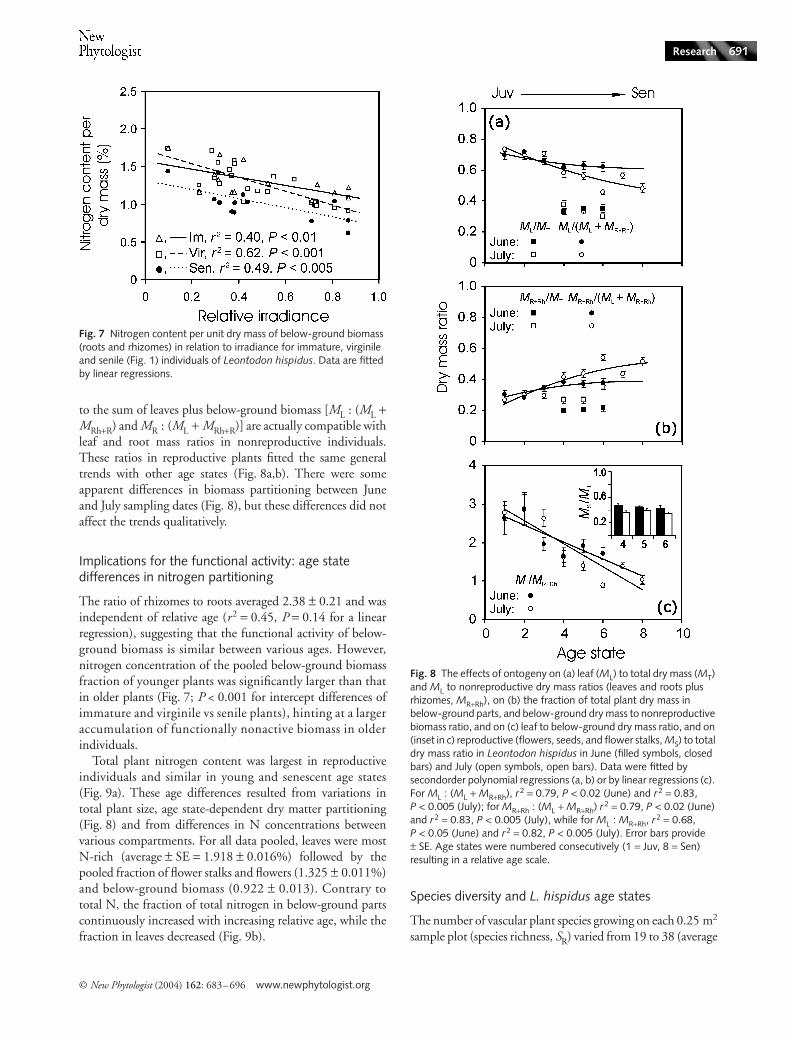
Young plants had a larger fraction of total dry mass inleaves than senescent plants (Fig. 8a), while older plants hada greater fraction of biomass in below-ground parts (Fig. 8b).As the results of these trends, the ratio of leaves to rootsmonotonically decreased with increasing relative age (Fig. 8c).This relationship did not just reflect the overall greater per-ennial part in senescent plants, but was also due to lower leafbiomass (leaf number per plant, leaf area, and mass, Fig. 6) insenescent individuals.
Because of a large biomass investment in flower stalk andflowers (inset in Fig. 8c), reproductive plants had the lowestfractions of dry mass in leaves and roots (Fig. 8a,b). However,in reproductive individuals, the ratios of leaves and roots
Fig. 6 Age-dependent modifications in (a) intercepts and slopes of leaf area ratio (LAR) versus relative irradiance (I) relations (Fig. 5), (b) average leaf area and (c) intercepts and slopes of total plant leaf area versus I relations, and (d) total intercepted light per unit whole plant dry mass in Leontodon hispidus in July. For reproductive age phases, LAR is calculated as total area per nonreproductive dry mass (Fig. 5). Potential amount of intercepted light (Eqn. 1) was calculated assuming a conical leaf angle distribution, and that plant light harvesting is not affected by other species and using a value of 36 mol m−2 d−1 for long-term seasonal incident irradiance. Data were fitted by second-order polynomial regressions. Error bars give ± SE. Age states were numbered consecutively (1 = Juv, 8 = Sen) resulting in a relative age scale.
© New Phytologist (2004) 162: 683–696 www.newphytologist.org
Research 691
to the sum of leaves plus below-ground biomass [ML : (ML +MRh+R) and MR : (ML + MRh+R)] are actually compatible withleaf and root mass ratios in nonreproductive individuals.These ratios in reproductive plants fitted the same generaltrends with other age states (Fig. 8a,b). There were someapparent differences in biomass partitioning between Juneand July sampling dates (Fig. 8), but these differences did notaffect the trends qualitatively.
Implications for the functional activity: age state differences in nitrogen partitioning
The ratio of rhizomes to roots averaged 2.38 ± 0.21 and wasindependent of relative age (r2 = 0.45, P = 0.14 for a linearregression), suggesting that the functional activity of below-ground biomass is similar between various ages. However,nitrogen concentration of the pooled below-ground biomassfraction of younger plants was significantly larger than thatin older plants (Fig. 7; P < 0.001 for intercept differences ofimmature and virginile vs senile plants), hinting at a largeraccumulation of functionally nonactive biomass in olderindividuals.
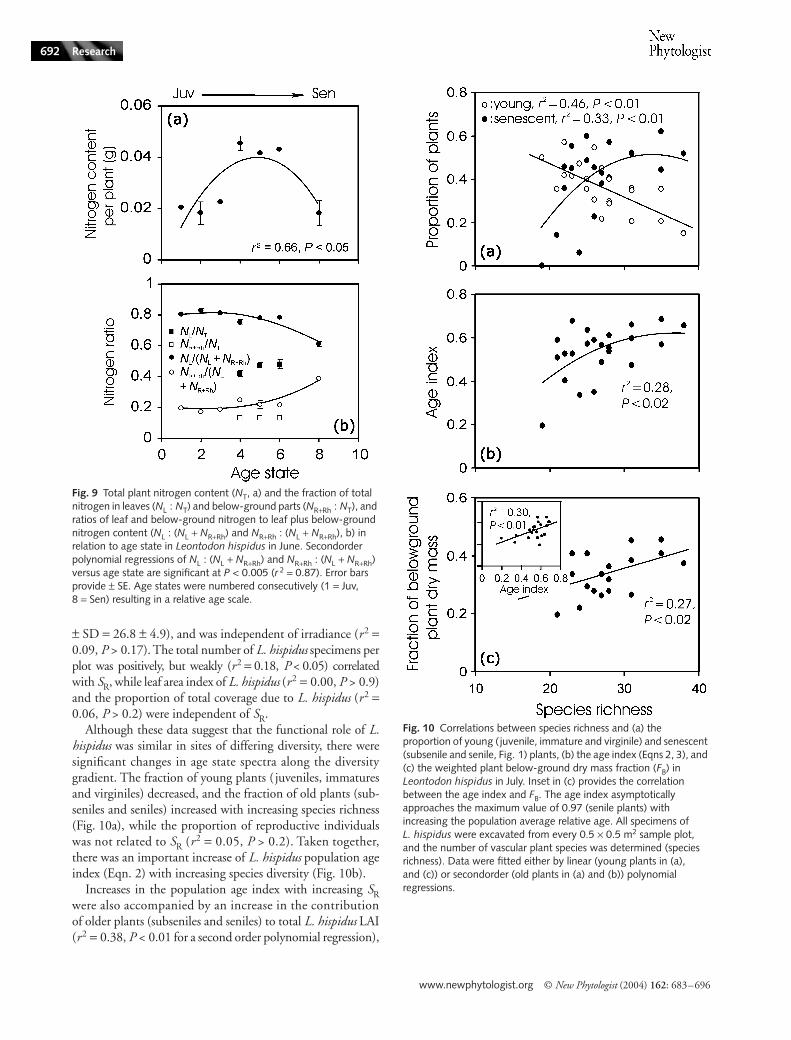
Total plant nitrogen content was largest in reproductiveindividuals and similar in young and senescent age states(Fig. 9a). These age differences resulted from variations intotal plant size, age state-dependent dry matter partitioning(Fig. 8) and from differences in N concentrations betweenvarious compartments. For all data pooled, leaves were mostN-rich (average ± SE = 1.918 ± 0.016%) followed by thepooled fraction of flower stalks and flowers (1.325 ± 0.011%)and below-ground biomass (0.922 ± 0.013). Contrary tototal N, the fraction of total nitrogen in below-ground partscontinuously increased with increasing relative age, while thefraction in leaves decreased (Fig. 9b).
Species diversity and L. hispidus age states
The number of vascular plant species growing on each 0.25 m2
sample plot (species richness, SR) varied from 19 to 38 (average
Fig. 7 Nitrogen content per unit dry mass of below-ground biomass (roots and rhizomes) in relation to irradiance for immature, virginile and senile (Fig. 1) individuals of Leontodon hispidus. Data are fitted by linear regressions.
Fig. 8 The effects of ontogeny on (a) leaf (ML) to total dry mass (MT) and ML to nonreproductive dry mass ratios (leaves and roots plus rhizomes, MR+Rh), on (b) the fraction of total plant dry mass in below-ground parts, and below-ground dry mass to nonreproductive biomass ratio, and on (c) leaf to below-ground dry mass ratio, and on (inset in c) reproductive (flowers, seeds, and flower stalks, MS) to total dry mass ratio in Leontodon hispidus in June (filled symbols, closed bars) and July (open symbols, open bars). Data were fitted by secondorder polynomial regressions (a, b) or by linear regressions (c). For ML : (ML + MR+Rh), r
2 = 0.79, P < 0.02 (June) and r2 = 0.83, P < 0.005 (July); for MR+Rh : (ML + MR+Rh) r
2 = 0.79, P < 0.02 (June) and r2 = 0.83, P < 0.005 (July), while for ML : MR+Rh, r
2 = 0.68, P < 0.05 (June) and r2 = 0.82, P < 0.005 (July). Error bars provide ± SE. Age states were numbered consecutively (1 = Juv, 8 = Sen) resulting in a relative age scale.
www.newphytologist.org © New Phytologist (2004) 162: 683–696
Research692
± SD = 26.8 ± 4.9), and was independent of irradiance (r2 =0.09, P > 0.17). The total number of L. hispidus specimens perplot was positively, but weakly (r2 = 0.18, P < 0.05) correlatedwith SR, while leaf area index of L. hispidus (r2 = 0.00, P > 0.9)and the proportion of total coverage due to L. hispidus (r2 =0.06, P > 0.2) were independent of SR.
Although these data suggest that the functional role of L.hispidus was similar in sites of differing diversity, there weresignificant changes in age state spectra along the diversitygradient. The fraction of young plants ( juveniles, immaturesand virginiles) decreased, and the fraction of old plants (sub-seniles and seniles) increased with increasing species richness(Fig. 10a), while the proportion of reproductive individualswas not related to SR (r2 = 0.05, P > 0.2). Taken together,there was an important increase of L. hispidus population ageindex (Eqn. 2) with increasing species diversity (Fig. 10b).
Increases in the population age index with increasing SRwere also accompanied by an increase in the contributionof older plants (subseniles and seniles) to total L. hispidus LAI(r2 = 0.38, P < 0.01 for a second order polynomial regression),
Fig. 9 Total plant nitrogen content (NT, a) and the fraction of total nitrogen in leaves (NL : NT) and below-ground parts (NR+Rh : NT), and ratios of leaf and below-ground nitrogen to leaf plus below-ground nitrogen content (NL : (NL + NR+Rh) and NR+Rh : (NL + NR+Rh), b) in relation to age state in Leontodon hispidus in June. Secondorder polynomial regressions of NL : (NL + NR+Rh) and NR+Rh : (NL + NR+Rh) versus age state are significant at P < 0.005 (r 2 = 0.87). Error bars provide ± SE. Age states were numbered consecutively (1 = Juv, 8 = Sen) resulting in a relative age scale.
Fig. 10 Correlations between species richness and (a) the proportion of young ( juvenile, immature and virginile) and senescent (subsenile and senile, Fig. 1) plants, (b) the age index (Eqns 2, 3), and (c) the weighted plant below-ground dry mass fraction (FB) in Leontodon hispidus in July. Inset in (c) provides the correlation between the age index and FB. The age index asymptotically approaches the maximum value of 0.97 (senile plants) with increasing the population average relative age. All specimens of L. hispidus were excavated from every 0.5 × 0.5 m2 sample plot, and the number of vascular plant species was determined (species richness). Data were fitted either by linear (young plants in (a), and (c)) or secondorder (old plants in (a) and (b)) polynomial regressions.

© New Phytologist (2004) 162: 683–696 www.newphytologist.org
Research 693
and by a greater fraction of total plant dry mass in below-ground parts (Fig. 10c). The age index and biomass allocationto below-ground parts were also strongly correlated (Fig. 10cinset).
Discussion
Irradiance effects on foliar structure, chemistry and total foliage area
Increases in leaf dry mass per unit area (MA) with irradiance(I ) are commonly observed in plant species independent ofgrowth form (Fig. 2; Meziane & Shipley, 1999; Niinemets,2001; Niinemets et al., 2003). Such a change in MA is acentral acclimation response that concentrats photo-synthetic biomass in high light environments where thephotosynthetic returns are the largest, while low MA in lowlight maximises plant leaf area and light interception(Gutschick & Wiegel, 1988). Although nitrogen content perunit dry mass (NM) decreased with increasing irradiance(Fig. 4a), the increase of MA with I (Fig. 2) generally exceededthe decrease in NM such that foliar nitrogen contents per areascaled positively with irradiance (NA = MANM; Fig. 4(b)),suggesting that foliar photosynthetic potentials did parallelthe changes in MA.
Positive correlations between carbohydrate contents and I(Fig. 4c,d) further suggested that cumulative daily photo-synthesis was maximised at high light. However, starch contentswere large, up to 20% of leaf dry mass, and increased withirradiance (Fig. 4c,d), possibly indicating limited sink activitiesfor usage of photosynthates at high irradiance. Overall,productivity responses to irradiance (Eek & Zobel, 2001)and the nutrient contents (Figs 4a and 7) indicate a greaterrequirement for nutrients at higher irradiance in the studiedgrassland, as in many natural soils where increases in light areaccompanied by decreases in foliage nutrient contents (Bazzaz& Wayne, 1994). Similar decline in plant nutrient content athigher light but at common nutrient supply has also beenobserved in many growth chamber experiments (e.g. McDonaldet al., 1992).
Leaf area per total plant biomass (leaf area ratio; LAR) isa chief determinant of species light harvesting capacity andits growth rate, especially in low irradiance (Poorter & vander Werf, 1998; Meziane & Shipley, 1999; Poorter & Nagel,2000; Ryser & Wahl, 2001). In the current study, LAR signi-ficantly increased with decreasing irradiance (Fig. 5), indicat-ing a greater light harvesting efficiency in low light as has beenconsistently observed in other studies (Meziane & Shipley,1999; Poorter, 1999; Poorter & Nagel, 2000).
Modification of plant plasticity by relative age
Changes in leaf dry mass per unit area versus irradiancerelations (Fig. 2) with the ontogeny (Figs 2 and 3)
demonstrate important changes in foliar functioning withthe age. In particular, the gradual change in the slope ofMA versus I relationships (Fig. 3) suggests that the plasticityfor structural leaf modifications changes throughout theontogeny. While young plants consistently form leaves withlower MA to capture more light, reproductive individualswith larger below-ground reserves possess greater structuralplasticity, allowing them to tune plant photosynthetic potentialto incident light conditions. By contrast, senescent plantspossessed an overall large MA (large MA versus I intercept)and a limited potential for MA adjustment to light alterations(the slope in Fig. 3). Such low plasticity is in agreementwith a limited role of nonreproducing senescent plants inpopulation maintenance.
As the result of lower MA (Figs 2 and 3) and larger frac-tional investment (ML/MT) of biomass in leaves [Fig. 8(a);LAR = (ML/MT)/MA], younger individuals had significantlyhigher LAR than older plants (Fig. 6a). In addition, youngerplants possessed greater plasticity for LAR adjustment toirradiance (Fig. 6a), collectively demonstrating that establish-ing young plants put a premium on efficient light harvesting.
Although the youngest individuals have the highest effi-ciency and plasticity for a given investment of resources, LARis a relative measure, and may not entirely reflect the resourcecapture of discrete individuals due to asymmetric competition(Anten & Hirose, 1998; Freckleton & Watkinson, 2001). Inthe current study, reproductive plants had the largest size andlargest total foliar area (Fig. 6a,b), suggesting that these plantsmay capture disproportionately more resources than youngerindividuals. However, L. hispidus is a rosette plant with allleaves inclined at a constant angle, and in the same positionin herbaceous layer irrespective of age state. This signifies thatthe effects of differences in total area itself are less than forplants with leafed stems. As the analysis of light interceptioncapacity demonstrates (Fig. 6d), potential light harvestingefficiency per total plant mass was significantly higher inyoung relative to the reproductive and senescent ages.
Perennial plant ontogeny: adaptation or inevitable ontogenetic constraints?
Overall accumulation of perennial structures in olderindividuals suggests that the trends in plasticity may reflectinevitable ontogenetic constraints. However, the life-historypatterns apparently also change throughout the ontogeny.While the premium is on survival in young plants, it shiftsto reproduction in reproductive stages. Larger MA and totalplant area and biomass in reproductive individuals can beconsidered indicative of a more aggressive tactic for resourcecapture. By contrast, young plants with the lowest plasticity forMA adjustment but largest for LAR changes exhibit a toleranttactic. Thus, the observed changes in biomass allocation andplasticity may be a part of an overall adaptive switching strategypreviously defined for woody species (Grubb, 1998).

www.newphytologist.org © New Phytologist (2004) 162: 683–696
Research694
Species is a species. Is it?
The age state significantly affected the relative fractions oftotal biomass in leaves (Fig. 8a) and roots (Fig. 8b), implyinga major modification in above-to-below-ground functionalactivity distribution. Although total N content may be similarbetween young and senescent plants (Fig. 9a), populationswith predominantly young individuals partition most Nabove-ground, while senescent populations accumulateN below-ground (Fig. 9b). These differences have majorimplications for interpreting the role of species in communityfunctioning and dynamics. Major effort has been expended inshort-term species screening for relevant ecological traits tounderstand species performance in the community (Grimeet al., 1997; Craine et al., 2002), and mechanistic ecosystem-level models of species coexistence (Tilman et al., 1997; Loreau,1998) consider every species as a discrete entity with a specificfunctional response in all situations. For instance, mechanisticexplanations of the productivity versus species richnesshypothesis state that multispecies communities are moreproductive because each of the composite species has uniqueresource requirement such that all species collectively useheterogeneously distributed environmental resources moreefficiently than a single species (Tilman et al., 1997; Hooper& Vitousek, 1998; Loreau, 1998). However, a spectrum ofresponses with respect to usage of environmental resources fora single species due to ontogeny clearly blurs the distinctdifferences between species.
Furthermore, it is important that ontogenetic changes leadto combinations of traits that may apparently seem incom-patible with specific ecosystem functioning (Lavorel &Garnier, 2002) unless one considers that such trait combina-tions may reflect adaptations to survival (young plants) orinevitable ontogenetic constraints (senescent plants). Currentmodels of species coexistence may need modification toinclude additional complexities like continuous strategychange from tolerant, high-light harvesting potential injuvenile plants to competitive, high photosynthetic potentialin reproductive plants. Such confounding aspects have beenaddressed in the past (Grubb, 1977), but are largely ignoredin current vegetation analyses.
Species diversity and population age
The importance of age modifications in species functioningfor vegetation composition and dynamics is furtherunderscored by the shift of the age state spectrum towardsontogenetically older, established individuals with increasingspecies diversity (Fig. 10a,b). This observation agrees withexperimental evidence that more diverse plant communitieshave fewer regeneration niches than less diverse communities(Tilman, 1993, 1997), and also with the data demonstratinglarger fractions of ontogenetically older individuals in morestressful environments (Zaugolnova et al., 1991). Greater
below-ground reserves in more diverse communities (Fig. 10c)may also partly explain the greater community resistance andresilience to environmental changes in more species-richhabitats (Tilman & Downing, 1994; Tilman, 1996). Further-more, larger fractions of biomass in below-ground partsdue to population ageing (Fig. 10c) imply a greater carbonstorage in more species-rich sites, as has been demonstrated inlong-term experiments (Wedin & Tilman, 1996). However,large fractional nitrogen immobilization in storage com-partments with low turnover rates further indicates that thestability of species-rich communities may partly rely on theirslow turnover of limiting resources such as nitrogen (Wedin& Tilman, 1996; Hooper & Vitousek, 1998).
Because the young plants harvest light more efficientlythan the old plants (Figs 3 and 6), decreases in the fractionaldistribution of LAIL of young individuals relative to oldplants with increasing species diversity suggest that in thecurrent study, the overall light harvesting efficiency decreasedwith increasing diversity due to age constraints. This resultcontrasts with previous observations in experimental grass-land communities (Spehn et al., 2000). However, in thelatter study, species diversity affected light harvestingonly in treatments where nitrogen availability was high dueto the abundance of legumes (Spehn et al., 2000). In oursites, total productivity was primarily constrained bynitrogen (Eek & Zobel, 2001) and the overall effect ofdiversity on productivity was minor.
Conclusions
This study demonstrates significant modifications in speciesplasticity and functioning with relative age state. Even for asingle species, there is a spectrum of morphological responsesto irradiance, implying that a species’ reaction to environ-mental variation is strongly affected by the distribution ofindividuals between various age states. Counterintuitively, themodification of biomass allocation patterns with the age is notjust due to accumulation of perennial structures with age, butalso reflects an overall change in allocation pattern of newlysynthesised biomass, compatible with changes in life historycharcteristics during ontogeny.
Although I studied a single species only, the hypothesis ofa strong linkage of species relative age spectrum, species func-tional activity and species diversity has major significancefor interpreting species coexistence patterns and calls forfurther long-term field experiments. Changes in communityperformance due to alterations in species diversity havebeen explained by large species differences in usage of environ-mental resources that are experimentally observed when speciesof common age are compared (Tilman et al., 1997; Craineet al., 2002). However, a mixture of plants with differing agesalways exists in a community, diminishing the interspecificdifferences in resource usage. Furthermore, changes in speciesage spectrum significantly modify plant nitrogen and carbon

© New Phytologist (2004) 162: 683–696 www.newphytologist.org
Research 695
distributions between pools of differing turnover rate, andthus partly determine whether specific grassland ecosystemsbehave as nutrient and carbon sinks or sources (Wedin, 1995).
Acknowledgements
I thank Kalevi Kull for stimulating discussions, MichaelBahn, Eric Garnier and the anonymous referees for insightfulcomments, and Toomas Kukk and Mari Tobias for skilledassistance with the Fig. 1. Financial support was provided bythe European Commission (CARBOMONT, EVK2-2002–0549), the Estonian Science Foundation (Grant 5702) andthe Estonian Ministry of Education and Science (grant0182468As03).
ReferencesAnderson MC. 1964. Studies of the woodland light climate. I. The
photographic computation of light conditions. Journal of Ecology 52: 27–41.
Anten NPR, Hirose T. 1998. Biomass allocation and light partitioning among dominant and subordinate individuals in Xanthium canadense stands. Annals of Botany 82: 665–673.
Bazzaz FA, Wayne PM. 1994. Coping with environmental heterogeneity: the physiological ecology of tree seedling regeneration across the gap – understory continuum. In: Caldwell MM, Pearcy RW, eds. Exploitation of environmental heterogeneity by plants. Ecophysiological processes above- and belowground. San Diego, CA, USA: Academic Press, 349–390.
Berendse F. 1993. Ecosystem stability, competition, and nutrient cycling. In: Schulze E-D, Mooney HA, eds. Biodiversity and ecosystem function. Ecological studies 99. Berlin, Germany: Springer Verlag, 409–431.
Bond BJ. 2000. Age-related changes in photosynthesis of woody plants. Trends in Plant Science 5: 349–353.
ter Braak CJF, Gremmen NJM. 1987. Ecological amplitudes of plant species and the internal consistency of Ellenberg’s indicator values for moisture. Vegetatio 69: 79–87.
Campbell GS, Norman JM. 1989. The description and measurement of plant canopy structure. In: Russell G, Marshall B, Jarvis PG, eds. Plant canopies: their growth, form and function. Society for Experimental Biology Seminar Series 31. Cambridge, UK: Cambridge University Press, 1–19.
Carlen C, Kolliker R, Nösberger J. 1999. Dry matter allocation and nitrogen productivity explain growth responses to photoperiod and temperature in forage grasses. Oecologia 121: 441–446.
Coleman JS, McConnaughay KDM, Ackerly DD. 1994. Interpreting phenotypic variation in plants. Trends in Ecology and Evolution 9: 187–191.
Craine JM, Tilman D, Wedin D, Reich P, Tjoelker M, Knops J. 2002. Functional traits, productivity and effects on nitrogen cycling of 33 grassland species. Functional Ecology 16: 563–574.
Day JS, Gould KS, Jameson PE. 1997. Vegetative architecture of Elaeocarpus hookerianus. Transition from juvenile to adult. Annals of Botany 79: 617–624.
Eek L, Zobel K. 2001. Structure and diversity of a species-rich grassland community, treated with additional illumination, fertilization and mowing. Ecography 24: 157–164.
Ellenberg H. 1991. Zeigerwerte der Gefäβpflanzen (ohne Rubus). In: Ellenberg H, Düll R, Wirth V, Werner W, Paulißen D, eds. Zeigerwerte von Pflanzen in Mitteleuropa. Scripta Geobotanica 18, Göttingen: Erich Goltze KG, 9–166.
Ertsen ACD, Alkemade JRM, Wassen MJ. 1998. Calibrating Ellenberg indicator values for moisture, acidity, nutrient availability and salinity in the Netherlands. Plant Ecology 135: 113–124.
Falinska K. 1995. Genet disintegration in Filipendula ulmaria: consequences for population dynamics and vegetation succession. Journal of Ecology 83: 9–21.
Freckleton RP, Watkinson AR. 2001. Asymmetric competition between plant species. Functional Ecology 15: 615–623.
Gatsuk LE, Smirnova OV, Vorontzova LI, Zaugolnova LB, Zhukova LA. 1980. Age states of plants of various growth forms: a review. Journal of Ecology 68: 675–696.
Grime JP, Thompson K, Hunt R, Hodgson JG, Cornelissen JHC, Rorison IH, Hendry GAF, Ashenden TW, Askew AP, Band SR, Booth RE, Bossard CC, Campbell BD, Cooper JEL, Davison AW, Gupta PL, Hall W, Hand DW, Hannah MA, Hillier SH, Hodkinson DJ, Jalili A, Liu Z, Mackey JML, Matthews N, Mowforth MA, Neal AM, Reader RJ, Reiling K, Ross-Fraser W, Spencer RE, Sutton F, Tasker DE, Thorpe PC, Whitehouse J. 1997. Integrated screening validates primary axes of specialisation in plants. Oikos 79: 259–281.
Grubb PJ. 1977. The maintenance of species-richness in plant communities: the importance of the regeneration niche. Biological Reviews of the Cambridge Philosophical Society 52: 107–145.
Grubb PJ. 1998. A reassessment of the strategies of plants which cope with shortages of resources. Perspectives in Plant Ecology, Evolution and Systematics 1: 3–31.
Gutschick VP, Wiegel FW. 1988. Optimizing the canopy photosynthetic rate by patterns of investment in specific leaf mass. American Naturalist 132: 67–86.
Hæggström C-A. 1983. Vegetation and soil of the wooded meadows in Nåtö, Åland. Acta Botanica Fennica 120: 1–66.
Hooper DU, Vitousek PM. 1998. Effects of plant composition and diversity on nutrient cycling. Ecological Monographs 68: 121–149.
Huston MA, DeAngelis DL. 1994. Competition and coexistence: the effects of resource transport and supply rates. American Naturalist 144: 954–977.
Kukk T, Kull K. 1997. Puisniidud. (Wooded Meadows), Estonia Maritima. Publications of the West-Estonian Archipelago Biosphere Reserve 2. Haapsalu, Estonia: West-Estonian Archipelago Biosphere Reserve. (In Estonian).
Kull K, Zobel M. 1991. High species richness in an Estonian wooded meadow. Journal of Vegetation Science 2: 711–714.
Lavorel S, Garnier E. 2002. Predicting changes in community composition and ecosystem functioning from plant traits: revisiting the Holy Grail. Functional Ecology 16: 545–556.
Loreau M. 1998. Biodiversity and ecosystem functioning: a mechanistic model. Proceedings of the National Academy of Sciences, USA 95: 5632–5636.
van der Maarel E. 1996. Vegetation dynamics and dynamic vegetation science. Acta Botanica Neerlandica 45: 421–442.
McDonald AJS, Lohammar T, Ingestad T. 1992. Net assimilation rate and shoot area development in birch (Betula pendula Roth.) at different steady-state values of nutrition and photon flux density. Trees: Structure and Function 6: 1–6.
Meziane D, Shipley B. 1999. Interacting determinants of specific leaf area in 22 herbaceous species: effects of irradiance and nutrient availability. Plant, Cell & Environment 22: 447–459.
Niinemets Ü. 1995. Distribution of foliar carbon and nitrogen across the canopy of Fagus sylvatica: adaptation to a vertical light gradient. Acta Oecologica 16: 525–541.
Niinemets Ü. 2001. Climatic controls of leaf dry mass per area, density, and thickness in trees and shrubs at the global scale. Ecology 82: 453–469.
Niinemets Ü. 2002. Stomatal conductance alone does not explain the decline in foliar photosynthetic rates with increasing tree age and size in Picea abies and Pinus sylvestris. Tree Physiology 22: 515–535.
Niinemets Ü, Valladares F, Ceulemans R. 2003. Leaf-level phenotypic variability and plasticity of invasive Rhododendron ponticum and non-invasive Ilex aquifolium co-occurring at two contrasting European sites. Plant, Cell & Environment 26: 941–956.

www.newphytologist.org © New Phytologist (2004) 162: 683–696
Research696
Orive ME. 1995. Senescence in organisms with clonal reproduction and complex life histories. American Naturalist 145: 90–108.
Partridge L, Mangel M. 1999. Messages from mortality: the evolution of death rates in the old. Trends in Ecology and Evolution 14: 438–442.
Poorter L. 1999. Growth responses of 15 rain-forest tree species to a light gradient: the relative importance of morphological and physiological traits. Functional Ecology 13: 396–410.
Poorter H, Nagel O. 2000. The role of biomass allocation in the growth response of plants to different levels of light, CO2, nutrients and water: a quantitative review. Australian Journal of Plant Physiology 27: 595–607.
Poorter H, van der Werf A. 1998. Is inherent variation in RGR determined by LAR at low irradiance and by NAR at high irradiance? A review of herbaceous species. In: Lambers H, Poorter H, van Vuuren MMI, eds. Inherent variation in plant growth. Physiological mechanisms and ecological consequences. Leiden, The Netherlands: Backhuys Publishers, 309–336.
Rabotnov TA. 1985. Dynamics of plant coenotic populations. In: White J, ed. The population structure of vegetation. Handbook of Vegetation Science III. Dordrecht, The Netherlands: Dr W. Junk Publishers, 121–142.
Reif A, Teckelmann M, Schulze E-D. 1985. Die Standortamplitude der Groβen Brennessel (Urtica dioica L.) – eine Auswertung vegetationskundlicher Aufnahmen auf der Grundlage der Ellenbergschen Zeigerwerte. Flora 176: 365–382.
Rice SA, Bazzaz FA. 1989a. Growth consequences of plasticity of plant traits in response to light conditions. Oecologia 78: 508–512.
Rice SA, Bazzaz FA. 1989b. Quantification of plasticity of plant traits in response to light intensity: comparing phenotypes at a common weight. Oecologia 78: 502–507.
Ryser P, Eek L. 2000. Consequences of phenotypic plasticity vs. interspecific differences in leaf and root traits for acquisition of aboveground and belowground resources. American Journal of Botany 87: 402–411.
Ryser P, Wahl S. 2001. Interspecific variation in RGR and the underlying traits among 24 grass species grown in full daylight. Plant Biology 3: 426–436.
Schippers P, Kropff MJ. 2001. Competition for light and nitrogen among grassland species: a simulation analysis. Functional Ecology 15: 155–164.
Schmid B, Bazzaz FA. 1990. Plasticity in plant size and architecture in rhizome-derived vs. seed-derived Solidago and Aster. Ecology 71: 523–535.
Semmartin M, Oesterheld M. 2001. Effects of grazing pattern and nitrogen availability on primary productivity. Oecologia 126: 225–230.
Serebryakova TI, Gatsuk LE, Zhukova LA, Kurchenko EI. 1983. Diagnoza i klyuchi vozrastnyh sostoyanii lugovyh rastenii (Diagnosis and keys of the age states of meadow plants). Metodicheskiye razrabotki dlya studentov biologicheskih spetsialnostei. Volume III. Moskva. Moskovskii Gosudarstvennyi Pedagogicheskii Institut Imeni V. I. Lenina. (In Russian).
Shipley B. 2000. Plasticity in relative growth rate and its components following a change in irradiance. Plant, Cell & Environment 23: 1207–1216.
Shipley B. 2002. Trade-offs between net assimilation rate and specific leaf area in determining relative growth rate: relationship with daily irradiance. Functional Ecology 16: 682–689.
Spehn EM, Joshi J, Schmid B, Diemer M, Körner C. 2000. Above-ground resource use increases with plant species richness in experimental grassland ecosystems. Functional Ecology 14: 326–337.
Tilman D. 1993. Species richness of experimental productivity gradients: how important is colonization limitation? Ecology 74: 2179–2191.
Tilman D. 1996. Biodiversity: population versus ecosystem stability. Ecology 77: 350–363.
Tilman D. 1997. Community invasibility, recruitment limitation, and grassland biodiversity. Ecology 78: 81–92.
Tilman D, Downing JA. 1994. Biodiversity and stability in grasslands. Nature 367: 363–365.
Tilman D, Lehman CL, Thomson KT. 1997. Plant diversity and ecosystem productivity: theoretical considerations. Proceedings of the National Academy of Sciences, USA 94: 1857–1861.
Tilman D, Wedin D, Knops J. 1996. Productivity and sustainability influenced by biodiversity in grassland ecosystems. Nature 379: 718–720.
Uranov AA. 1977. Osnovnyye cherty prostranstvenno-vremennoi organizatsii tsenopopulyatsii. (Main features of spatio-temporal organization of coenopopulations). In: Serebryakova TI, ed. Tsenopopulyatsii Rastenii. Razvitiye i Vzaimootnosheniye. Moskva: Nauka. 7–20, (In Russian).
Valladares F, Balaguer L, Martínez-Ferri E, Perez-Corona E, Manrique E. 2002. Plasticity, instability and canalization: is the phenotypic variation in seedlings of sclerophyll oaks consistent with the environmental unpredictability of Mediterranean ecosystems? New Phytologist 156: 457–467.
Wedin DA. 1995. Species, nitrogen, and grassland dynamics: the constraints of stuff. In: Jones CG, Lawton JH, eds. Linking species and ecosystems. New York, USA: Chapman & Hall, 253–262.
Wedin D, Tilman D. 1993. Competition among grasses along a nitrogen gradient: initial conditions and mechanisms of competition. Ecological Monographs 63: 199–229.
Wedin DA, Tilman D. 1996. Influence of nitrogen loading and species composition on the carbon balance of grasslands. Science 274: 1720–1723.
Zaugolnova LB, Sugorkina NS, Sherbakova YG. 1991. Zhiznennyye formy i populyatsionnoye povedeniye mnogoletnih travyanistyh rastenii. (Life forms and population behaviour of herbaceous perennials). In: Zhilov IA, ed. Ekologiya Populyatsii. Sbornik Nauchnyh Statei. Moskva: Nauka, 5–22. (In Russian).
Zhukova LA. 1987. Dinamika tsenopopulyatsii lugovyh rastenii. (Coenopopulation dynamics of meadow plants). Dr. Biol. Dissertation, Moskovskii Gosudarstvennyi pedagogicheskii Institut Imeni V.I. Lenina.
Zhukova LA, Ermakova IM. 1985. Structure and dynamics of coenopopulations of some temperate grasses. In: White J, ed. The population structure of vegetation. Handbook of Vegetation Science III. Dordrecht, The Netherlands: Dr W. Junk Publishers. 179–205.




















