
Acid washing effect on elemental and isotopic composition of whole beach arthropods: Implications...
8
Original article Acid washing effect on elemental and isotopic composition of whole beach arthropods: Implications for food web studies using stable isotopes Oscar Serrano a , Laura Serrano a , Miguel Angel Mateo a, *, Isabella Colombini b , Lorenzo Chelazzi b , Elena Gagnarli b , Mario Fallaci b a Centro de Estudios Avanzados de Blanes, Consejo Superior de Investigaciones Cientı´ficas. Acceso a la Cala St. Francesc, 14. 17300 Blanes, Girona, Spain b Istituto per lo Studio degli Ecosistemi del CNR Sezione di Firenze, Via Madonna del Piano, Sesto Fiorentino, Florence, Italy article info Article history: Received 1 February 2008 Accepted 17 April 2008 Published online 3 June 2008 Keywords: Acidification Carbon Nitrogen Carbonates Isotopic ratios Macroinvertebrates abstract Inorganic carbon removal through acidification is a common practice prior to isotopic anal- ysis of macroinvertebrate samples. We have experimentally tested the effect of acidifica- tion on the elemental and isotopic composition of a range of beach arthropod species. Acidification resulted in a significant depletion of 7.7% and 1.2% on average for carbon and nitrogen, respectively, suggesting that acid washing affects body carbon compounds other than carbonates. With a few exceptions, d 13 C and d 15 N showed no changes following 1 N HCl attack. Based on those exceptions, our results show that only those samples with a high CaCO 3 content result in impoverished 13 C as a consequence of acidification. Those suspected to be carbonate-free are not significantly affected. Concerning d 15 N values, only high carbonate species were affected when treated with HCl. As a standard protocol, it is recommended to acidify only carbonate-rich samples prior to d 13 C analyses. When possible, muscle tissue samples should be used instead of the entire organism. ª 2008 Elsevier Masson SAS. All rights reserved. 1. Introduction Carbon and nitrogen isotopic ratios are very useful tools for the study of food web structures in aquatic ecosystems (e.g., Jennings et al., 1997; Pinnegar and Polunin, 2000; Vizzini and Mazzola, 2003; Fry, 2006). This approach helps to elucidate the origin of the ingested organic matter (Fry and Sherr, 1984; Owens, 1987; Preston, 1992) and to characterize flows of mass and energy through ecosystems (Fry, 1988; Owens, 1988; Hobson and Welch, 1992; Rau et al., 1992; Hesslein et al., 1993). Most arthropod species contain carbonates which contrib- ute to the global animal isotopic signature. It is common that samples are acidified before analysis in order to avoid the alteration of the d 13 C values by the high 13 C content of this non-dietary carbon fraction (e.g., Nieuwenhuize et al., 1994; Yokohama et al., 2005). The chemical reaction that follows the acidification treatment is: CaCO 3 þ 2HCl/CO [ 2 þ CaCl 2 þ H 2 O (1) However, the need for and the convenience of using HCl as part of the sample preparation for duel isotopic analysis is still a matter of controversy that has only recently begun to be directly addressed (e.g., Soreide et al., 2006; Mateo et al. (in press)). A few observations have reported that acidification * Corresponding author. Fax: þ34 972 337806. E-mail address: [email protected] (M.A. Mateo). available at www.sciencedirect.com journal homepage: www.elsevier.com/locate/actoec 1146-609X/$ – see front matter ª 2008 Elsevier Masson SAS. All rights reserved. doi:10.1016/j.actao.2008.04.002 acta oecologica 34 (2008) 89–96
-
Upload
independent -
Category
Documents
-
view
3 -
download
0
Transcript of Acid washing effect on elemental and isotopic composition of whole beach arthropods: Implications...

Original article
Acid washing effect on elemental and isotopic compositionof whole beach arthropods: Implications for food webstudies using stable isotopes
Oscar Serranoa, Laura Serranoa, Miguel Angel Mateoa,*, Isabella Colombinib,Lorenzo Chelazzib, Elena Gagnarlib, Mario Fallacib
aCentro de Estudios Avanzados de Blanes, Consejo Superior de Investigaciones Cientıficas. Acceso a la Cala St. Francesc,
14. 17300 Blanes, Girona, SpainbIstituto per lo Studio degli Ecosistemi del CNR Sezione di Firenze, Via Madonna del Piano, Sesto Fiorentino, Florence, Italy
a r t i c l e i n f o
Article history:
Received 1 February 2008
Accepted 17 April 2008
Published online 3 June 2008
Keywords:
Acidification
Carbon
Nitrogen
Carbonates
Isotopic ratios
Macroinvertebrates
a b s t r a c t
Inorganic carbon removal through acidification is a common practice prior to isotopic anal-
ysis of macroinvertebrate samples. We have experimentally tested the effect of acidifica-
tion on the elemental and isotopic composition of a range of beach arthropod species.
Acidification resulted in a significant depletion of 7.7% and 1.2% on average for carbon
and nitrogen, respectively, suggesting that acid washing affects body carbon compounds
other than carbonates. With a few exceptions, d13C and d15N showed no changes following
1 N HCl attack. Based on those exceptions, our results show that only those samples with
a high CaCO3 content result in impoverished 13C as a consequence of acidification. Those
suspected to be carbonate-free are not significantly affected. Concerning d15N values,
only high carbonate species were affected when treated with HCl. As a standard protocol,
it is recommended to acidify only carbonate-rich samples prior to d13C analyses. When
possible, muscle tissue samples should be used instead of the entire organism.
ª 2008 Elsevier Masson SAS. All rights reserved.
1. Introduction
Carbon and nitrogen isotopic ratios are very useful tools for the
study of food web structures in aquatic ecosystems (e.g.,
Jennings et al., 1997; Pinnegar and Polunin, 2000; Vizzini and
Mazzola, 2003; Fry, 2006). This approach helps to elucidate
the origin of the ingested organic matter (Fry and Sherr, 1984;
Owens, 1987; Preston, 1992) and to characterize flows of mass
and energy through ecosystems (Fry, 1988; Owens, 1988;
Hobson and Welch, 1992; Rau et al., 1992; Hesslein et al., 1993).
Most arthropod species contain carbonates which contrib-
ute to the global animal isotopic signature. It is common that
samples are acidified before analysis in order to avoid the
alteration of the d13C values by the high 13C content of this
non-dietary carbon fraction (e.g., Nieuwenhuize et al., 1994;
Yokohama et al., 2005). The chemical reaction that follows
the acidification treatment is:
CaCO3 þ 2HCl/CO[2 þ CaCl2 þH2O (1)
However, the need for and the convenience of using HCl as
part of the sample preparation for duel isotopic analysis is still
a matter of controversy that has only recently begun to be
directly addressed (e.g., Soreide et al., 2006; Mateo et al. (in
press)). A few observations have reported that acidification
* Corresponding author. Fax: þ34 972 337806.E-mail address: [email protected] (M.A. Mateo).
ava i lab le at www.sc ienced i rec t . com
journa l homepage : www. e lsev ier . com/ loca te /ac toec
1146-609X/$ – see front matter ª 2008 Elsevier Masson SAS. All rights reserved.doi:10.1016/j.actao.2008.04.002
a c t a o e c o l o g i c a 3 4 ( 2 0 0 8 ) 8 9 – 9 6

a c t a o e c o l o g i c a 3 4 ( 2 0 0 8 ) 8 9 – 9 690
may also alter d15N values (Pinnegar and Polunin, 1999),
supposedly as a consequence of partial loss of compounds
containing nitrogen (Bunn et al., 1995; Jacob et al., 2005) such
as chitin, proteins, and glycoprotein (Goering et al., 1990;
Shafer et al., 1994). Based on this, some authors prefer to di-
vide the sample into two aliquots, one acidified for the d13C
analysis and another one not acidified for the d15N analysis
(Polunin et al., 2001; Bouillon et al., 2002; Nyssen et al., 2002).
Other studies have reported no significant effect by HCl attack
on the isotopic signatures of marine animals, concluding that
acidification of the samples was unnecessary (e.g., Chanton
and Lewis, 1999; Grey et al., 2001; Nyssen et al., 2005). Some
authors have tried to limit the extent of the eventual effect
of acidification by acidifying samples before drying and grind-
ing (Bunn et al., 1995) or using more gentle carbonate removal
methods such as wetting samples in weak acid (0.1 N HCl; e.g.,
Hobson et al., 2002). However, only a few studies have
presented data on the elemental composition (C and N) and
discussed the effects of acidification on the elemental compo-
sition (Bunn et al., 1995). To contribute to the assessment of
these methodological effects in beach macroinvertebrates,
we evaluated the effects of acidification on C and N elemental
composition and on the d13C and d15N values of some repre-
sentative beach arthropods. The effect of acid washing on
the carbon and nitrogen composition of these species has
never been reported. To this end, the isotopic and elemental
compositions of acidified and non-acidified aliquots have
been compared.
Recently, Jacob et al. (2005) reported a positive linear rela-
tion between the effect of acidification on invertebrate d13C
values and an estimate of its carbonate content. Since then,
however, the sample’s CaCO3 content related to acid washing
effect on isotopic values has received little attention. An ex-
ception is a study by Ng et al. (2007), where they calculated
a carbonate proxy (as in Jacob et al., 2005) and recommended
acid rinsing only for carbonate-rich algae. We hypothesize
that acidification of invertebrate samples prior to analytical
procedures involves changes both in the elemental and in
the isotopic composition that can seriously confound food
web analysis based on isotopic trophic scenarios. More specif-
ically, we hypothesize that the most common acidification
procedure (i.e., 1 N HCl acid washing until bubbling cessation)
results in the removal of dietary (i.e., not from carbonates) car-
bon and nitrogen. We finally provide strong evidence support-
ing that the carbonate content of the invertebrate sample can
be one of the most useful criteria for deciding on the need for
acidifying the samples prior to isotopic analysis. To assess this
problem, we have selected various beach and semi-terrestrial
macroinvertebrate species and compared the elemental and
isotopic compositions of acid-washed to raw aliquots and
their relation with a carbonate proxy. In the light of the re-
sults, simple criteria for sample pre-treatment are proposed
for standardization.
2. Materials and methods
Sample collection was carried out in May 2005 on the beach
of Burano (N 42�2305100; E 11�2204000, Grosseto, Italy) using
pitfall cross traps that intercepted macroinvertebrates. Six
pitfall cross traps were deployed along the beach at 3-m in-
tervals from the shoreline to the base of the dune. The traps
were kept active for 24 h. The individuals captured were
stored in thermally-sealed plastic bags and frozen at the
site. Several individuals of the macroinvertebrate species
Arctosa cinerea (Araneae, Lycosidae), Geophilus sp. (Geophilo-
morpha, Geophilidae), Parallelomorphus laevigatus (Coleoptera,
Carabidae), Phaleria bimaculata (Coleoptera, Tenebrionidae),
Phaleria provincialis (Coleoptera, Tenebrionidae), Pimelia
bipunctata (Coleoptera, Tenebrionidae), Scarites buparius (Cole-
optera, Carabidae), Talitrus saltator (Amphipoda, Talitridae)
and Tylos europaeus (Isopoda, Tylidae) were selected. All
individuals were taxonomically determined, age classified,
cleaned with distilled water and oven-dried at 60 �C for
48 h. Finally, they were kept in a dry, dark place until pro-
cessing. Four replicates of each species were used for ele-
mental and isotopic analysis. Replicates consisted of 5–8
whole individuals of the same species. All replicates were
re-dried (placed in the oven overnight at 60 �C) and milled
to a fine homogeneous powder using an agate mortar and
pestle. The powder was stored in a dry environment (relative
humidity under 20%). About half of each of the replicates was
acidified by adding 1 N HCl drop-by-drop until cessation of
bubbling (Nieuwenhuize et al., 1994). Samples were then
left in excess of acid for 3 h. This procedure was selected be-
cause it is considered to be the most widely used standard.
The other half was kept untreated. The acidified sub-samples
were re-dried at 60 �C for 24 h, and milled again to a fine
homogenous powder. Samples were not rinsed with distilled
water after acidification to avoid the alteration of the isotope
values by the lixiviation of some organic carbon and nitrogen
compounds (e.g., Cloern et al., 2002).
Finally, an aliquot of 0.7 mg dry weight (�0.05 mg) of each
sample was weighed and placed in a tin capsule for solid
samples. The encapsulated samples were kept under constant
laboratory temperature (20 �C) and humidity conditions until
analysis. Elemental and isotopic composition was determined
for the gasses evolved from a single combustion using
a Finnigan Delta S isotope ratio mass spectrometer (Conflo II
interface) at the Scientific-Technical Services of the University
of Barcelona. Isotopic values are reported in the dVPDB notation
relative to the standards Vienna Pee Dee Belemnite and
atmospheric nitrogen for carbon and nitrogen, respectively
(dsample ¼ 1000 [(Rsample/Rstandard) � 1], R ¼ 13C/12C, or
R ¼ 15N/14N). Analytical precision based on the standard
deviation of internal standards (atropine, IAEA CH3, CH6,
CH7, and USGS40 – analytical grade L-glutamic acid, for car-
bon, and atropine, IAEA N1, NO3, N2, and USGS40, for nitro-
gen) ranged from 0.11 to 0.06& (mean ¼ 0.09&) for carbon,
and from 0.06 to 0.28& (mean ¼ 0.16&) for nitrogen.
All data were checked for normality (Kolmogorov–Smirnov
test) and for variance homogeneity (Levene’s test). One-way
ANOVAs were used to test the null hypothesis of no overall
difference in the elemental composition and isotopic values
between acidified and untreated samples. Post-hoc compari-
sons were then used to test specific differences between pairs
(single species, raw vs. acidified samples). A proxy was used to
estimate the content of CaCO3 and to assess its role on the ef-
fect of acid washing on the stable isotope ratios (Jacob et al.,
2005):
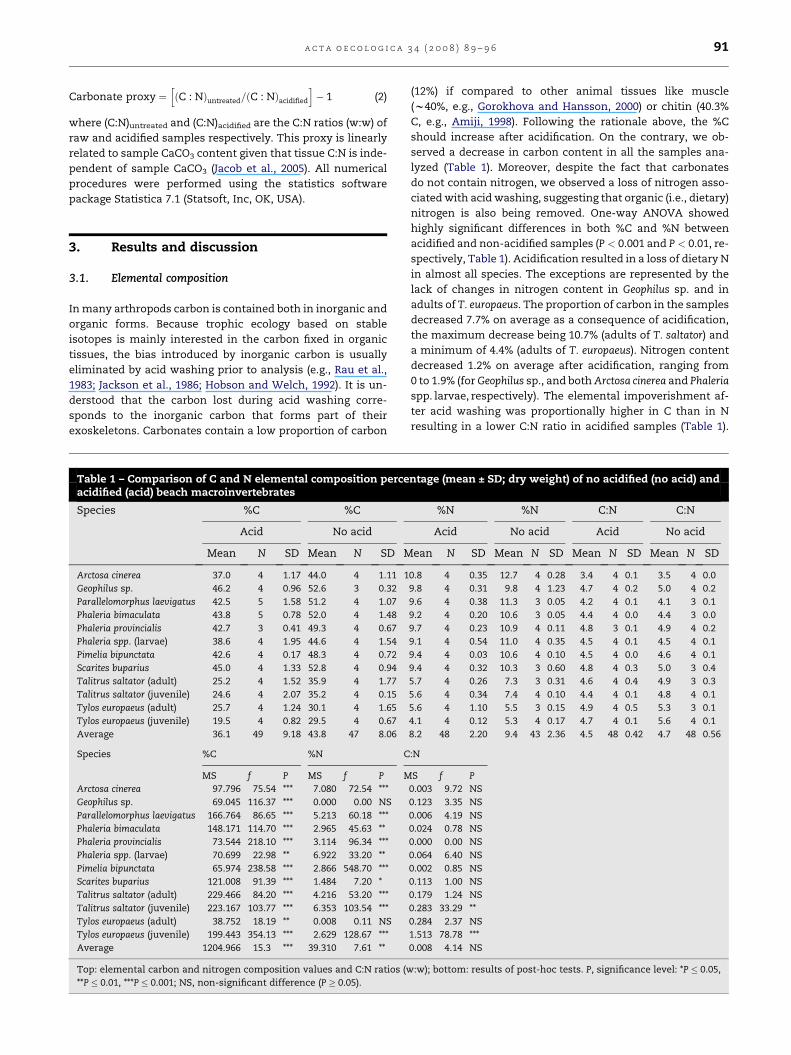
a c t a o e c o l o g i c a 3 4 ( 2 0 0 8 ) 8 9 – 9 6 91
Carbonate proxy ¼hðC : NÞuntreated=ðC : NÞacidified
i� 1 (2)
where (C:N)untreated and (C:N)acidified are the C:N ratios (w:w) of
raw and acidified samples respectively. This proxy is linearly
related to sample CaCO3 content given that tissue C:N is inde-
pendent of sample CaCO3 (Jacob et al., 2005). All numerical
procedures were performed using the statistics software
package Statistica 7.1 (Statsoft, Inc, OK, USA).
3. Results and discussion
3.1. Elemental composition
In many arthropods carbon is contained both in inorganic and
organic forms. Because trophic ecology based on stable
isotopes is mainly interested in the carbon fixed in organic
tissues, the bias introduced by inorganic carbon is usually
eliminated by acid washing prior to analysis (e.g., Rau et al.,
1983; Jackson et al., 1986; Hobson and Welch, 1992). It is un-
derstood that the carbon lost during acid washing corre-
sponds to the inorganic carbon that forms part of their
exoskeletons. Carbonates contain a low proportion of carbon
(12%) if compared to other animal tissues like muscle
(w40%, e.g., Gorokhova and Hansson, 2000) or chitin (40.3%
C, e.g., Amiji, 1998). Following the rationale above, the %C
should increase after acidification. On the contrary, we ob-
served a decrease in carbon content in all the samples ana-
lyzed (Table 1). Moreover, despite the fact that carbonates
do not contain nitrogen, we observed a loss of nitrogen asso-
ciated with acid washing, suggesting that organic (i.e., dietary)
nitrogen is also being removed. One-way ANOVA showed
highly significant differences in both %C and %N between
acidified and non-acidified samples (P < 0.001 and P < 0.01, re-
spectively, Table 1). Acidification resulted in a loss of dietary N
in almost all species. The exceptions are represented by the
lack of changes in nitrogen content in Geophilus sp. and in
adults of T. europaeus. The proportion of carbon in the samples
decreased 7.7% on average as a consequence of acidification,
the maximum decrease being 10.7% (adults of T. saltator) and
a minimum of 4.4% (adults of T. europaeus). Nitrogen content
decreased 1.2% on average after acidification, ranging from
0 to 1.9% (for Geophilus sp., and both Arctosa cinerea and Phaleria
spp. larvae, respectively). The elemental impoverishment af-
ter acid washing was proportionally higher in C than in N
resulting in a lower C:N ratio in acidified samples (Table 1).
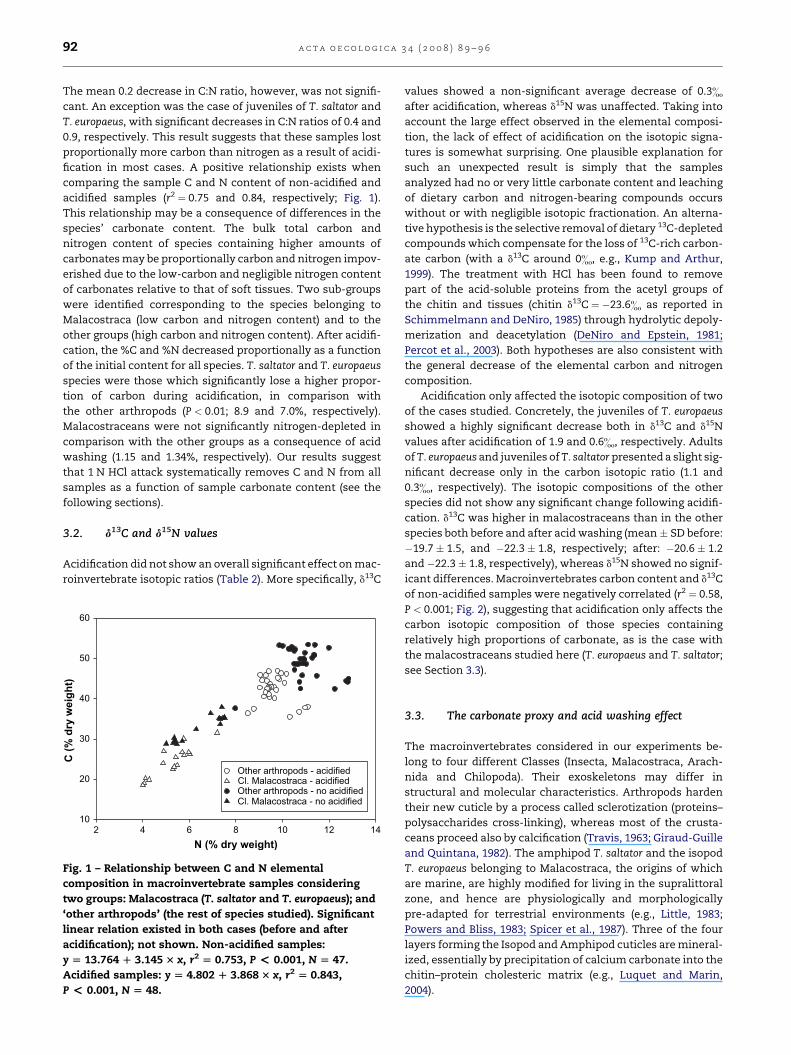
Table 1 – Comparison of C and N elemental composition percentage (mean ± SD; dry weight) of no acidified (no acid) andacidified (acid) beach macroinvertebrates
Species %C %C %N %N C:N C:N
Acid No acid Acid No acid Acid No acid
Mean N SD Mean N SD Mean N SD Mean N SD Mean N SD Mean N SD
Arctosa cinerea 37.0 4 1.17 44.0 4 1.11 10.8 4 0.35 12.7 4 0.28 3.4 4 0.1 3.5 4 0.0
Geophilus sp. 46.2 4 0.96 52.6 3 0.32 9.8 4 0.31 9.8 4 1.23 4.7 4 0.2 5.0 4 0.2
Parallelomorphus laevigatus 42.5 5 1.58 51.2 4 1.07 9.6 4 0.38 11.3 3 0.05 4.2 4 0.1 4.1 3 0.1
Phaleria bimaculata 43.8 5 0.78 52.0 4 1.48 9.2 4 0.20 10.6 3 0.05 4.4 4 0.0 4.4 3 0.0
Phaleria provincialis 42.7 3 0.41 49.3 4 0.67 9.7 4 0.23 10.9 4 0.11 4.8 3 0.1 4.9 4 0.2
Phaleria spp. (larvae) 38.6 4 1.95 44.6 4 1.54 9.1 4 0.54 11.0 4 0.35 4.5 4 0.1 4.5 4 0.1
Pimelia bipunctata 42.6 4 0.17 48.3 4 0.72 9.4 4 0.03 10.6 4 0.10 4.5 4 0.0 4.6 4 0.1
Scarites buparius 45.0 4 1.33 52.8 4 0.94 9.4 4 0.32 10.3 3 0.60 4.8 4 0.3 5.0 3 0.4
Talitrus saltator (adult) 25.2 4 1.52 35.9 4 1.77 5.7 4 0.26 7.3 3 0.31 4.6 4 0.4 4.9 3 0.3
Talitrus saltator (juvenile) 24.6 4 2.07 35.2 4 0.15 5.6 4 0.34 7.4 4 0.10 4.4 4 0.1 4.8 4 0.1
Tylos europaeus (adult) 25.7 4 1.24 30.1 4 1.65 5.6 4 1.10 5.5 3 0.15 4.9 4 0.5 5.3 3 0.1
Tylos europaeus (juvenile) 19.5 4 0.82 29.5 4 0.67 4.1 4 0.12 5.3 4 0.17 4.7 4 0.1 5.6 4 0.1
Average 36.1 49 9.18 43.8 47 8.06 8.2 48 2.20 9.4 43 2.36 4.5 48 0.42 4.7 48 0.56
Species %C %N C:N
MS f P MS f P MS f P
Arctosa cinerea 97.796 75.54 *** 7.080 72.54 *** 0.003 9.72 NS
Geophilus sp. 69.045 116.37 *** 0.000 0.00 NS 0.123 3.35 NS
Parallelomorphus laevigatus 166.764 86.65 *** 5.213 60.18 *** 0.006 4.19 NS
Phaleria bimaculata 148.171 114.70 *** 2.965 45.63 ** 0.024 0.78 NS
Phaleria provincialis 73.544 218.10 *** 3.114 96.34 *** 0.000 0.00 NS
Phaleria spp. (larvae) 70.699 22.98 ** 6.922 33.20 ** 0.064 6.40 NS
Pimelia bipunctata 65.974 238.58 *** 2.866 548.70 *** 0.002 0.85 NS
Scarites buparius 121.008 91.39 *** 1.484 7.20 * 0.113 1.00 NS
Talitrus saltator (adult) 229.466 84.20 *** 4.216 53.20 *** 0.179 1.24 NS
Talitrus saltator (juvenile) 223.167 103.77 *** 6.353 103.54 *** 0.283 33.29 **
Tylos europaeus (adult) 38.752 18.19 ** 0.008 0.11 NS 0.284 2.37 NS
Tylos europaeus (juvenile) 199.443 354.13 *** 2.629 128.67 *** 1.513 78.78 ***
Average 1204.966 15.3 *** 39.310 7.61 ** 0.008 4.14 NS
Top: elemental carbon and nitrogen composition values and C:N ratios (w:w); bottom: results of post-hoc tests. P, significance level: *P � 0.05,
**P � 0.01, ***P � 0.001; NS, non-significant difference (P � 0.05).
a c t a o e c o l o g i c a 3 4 ( 2 0 0 8 ) 8 9 – 9 692
The mean 0.2 decrease in C:N ratio, however, was not signifi-
cant. An exception was the case of juveniles of T. saltator and
T. europaeus, with significant decreases in C:N ratios of 0.4 and
0.9, respectively. This result suggests that these samples lost
proportionally more carbon than nitrogen as a result of acidi-
fication in most cases. A positive relationship exists when
comparing the sample C and N content of non-acidified and
acidified samples (r2 ¼ 0.75 and 0.84, respectively; Fig. 1).
This relationship may be a consequence of differences in the
species’ carbonate content. The bulk total carbon and
nitrogen content of species containing higher amounts of
carbonates may be proportionally carbon and nitrogen impov-
erished due to the low-carbon and negligible nitrogen content
of carbonates relative to that of soft tissues. Two sub-groups
were identified corresponding to the species belonging to
Malacostraca (low carbon and nitrogen content) and to the
other groups (high carbon and nitrogen content). After acidifi-
cation, the %C and %N decreased proportionally as a function
of the initial content for all species. T. saltator and T. europaeus
species were those which significantly lose a higher propor-
tion of carbon during acidification, in comparison with
the other arthropods (P < 0.01; 8.9 and 7.0%, respectively).
Malacostraceans were not significantly nitrogen-depleted in
comparison with the other groups as a consequence of acid
washing (1.15 and 1.34%, respectively). Our results suggest
that 1 N HCl attack systematically removes C and N from all
samples as a function of sample carbonate content (see the
following sections).
3.2. d13C and d15N values
Acidification did not show an overall significant effect on mac-
roinvertebrate isotopic ratios (Table 2). More specifically, d13C
N (% dry weight)
2 4 6 8 10 12 14
C (%
d
ry w
eig
ht)
10
20
30
40
50
60
Other arthropods - acidified Cl. Malacostraca - acidified Other arthropods - no acidified Cl. Malacostraca - no acidified
Fig. 1 – Relationship between C and N elemental
composition in macroinvertebrate samples considering
two groups: Malacostraca (T. saltator and T. europaeus); and
‘other arthropods’ (the rest of species studied). Significant
linear relation existed in both cases (before and after
acidification); not shown. Non-acidified samples:
y [ 13.764 D 3.145 3 x, r2 [ 0.753, P < 0.001, N [ 47.
Acidified samples: y [ 4.802 D 3.868 3 x, r2 [ 0.843,
P < 0.001, N [ 48.
values showed a non-significant average decrease of 0.3&
after acidification, whereas d15N was unaffected. Taking into
account the large effect observed in the elemental composi-
tion, the lack of effect of acidification on the isotopic signa-
tures is somewhat surprising. One plausible explanation for
such an unexpected result is simply that the samples
analyzed had no or very little carbonate content and leaching
of dietary carbon and nitrogen-bearing compounds occurs
without or with negligible isotopic fractionation. An alterna-
tive hypothesis is the selective removal of dietary 13C-depleted
compounds which compensate for the loss of 13C-rich carbon-
ate carbon (with a d13C around 0&, e.g., Kump and Arthur,
1999). The treatment with HCl has been found to remove
part of the acid-soluble proteins from the acetyl groups of
the chitin and tissues (chitin d13C ¼ �23.6& as reported in
Schimmelmann and DeNiro, 1985) through hydrolytic depoly-
merization and deacetylation (DeNiro and Epstein, 1981;
Percot et al., 2003). Both hypotheses are also consistent with
the general decrease of the elemental carbon and nitrogen
composition.
Acidification only affected the isotopic composition of two
of the cases studied. Concretely, the juveniles of T. europaeus
showed a highly significant decrease both in d13C and d15N
values after acidification of 1.9 and 0.6&, respectively. Adults
of T. europaeus and juveniles of T. saltator presented a slight sig-
nificant decrease only in the carbon isotopic ratio (1.1 and
0.3&, respectively). The isotopic compositions of the other
species did not show any significant change following acidifi-
cation. d13C was higher in malacostraceans than in the other
species both before and after acid washing (mean � SD before:
�19.7 � 1.5, and �22.3 � 1.8, respectively; after: �20.6 � 1.2
and �22.3 � 1.8, respectively), whereas d15N showed no signif-
icant differences. Macroinvertebrates carbon content and d13C
of non-acidified samples were negatively correlated (r2 ¼ 0.58,
P < 0.001; Fig. 2), suggesting that acidification only affects the
carbon isotopic composition of those species containing
relatively high proportions of carbonate, as is the case with
the malacostraceans studied here (T. europaeus and T. saltator;
see Section 3.3).
3.3. The carbonate proxy and acid washing effect
The macroinvertebrates considered in our experiments be-
long to four different Classes (Insecta, Malacostraca, Arach-
nida and Chilopoda). Their exoskeletons may differ in
structural and molecular characteristics. Arthropods harden
their new cuticle by a process called sclerotization (proteins–
polysaccharides cross-linking), whereas most of the crusta-
ceans proceed also by calcification (Travis, 1963; Giraud-Guille
and Quintana, 1982). The amphipod T. saltator and the isopod
T. europaeus belonging to Malacostraca, the origins of which
are marine, are highly modified for living in the supralittoral
zone, and hence are physiologically and morphologically
pre-adapted for terrestrial environments (e.g., Little, 1983;
Powers and Bliss, 1983; Spicer et al., 1987). Three of the four
layers forming the Isopod and Amphipod cuticles are mineral-
ized, essentially by precipitation of calcium carbonate into the
chitin–protein cholesteric matrix (e.g., Luquet and Marin,
2004).
a c t a o e c o l o g i c a 3 4 ( 2 0 0 8 ) 8 9 – 9 6 93
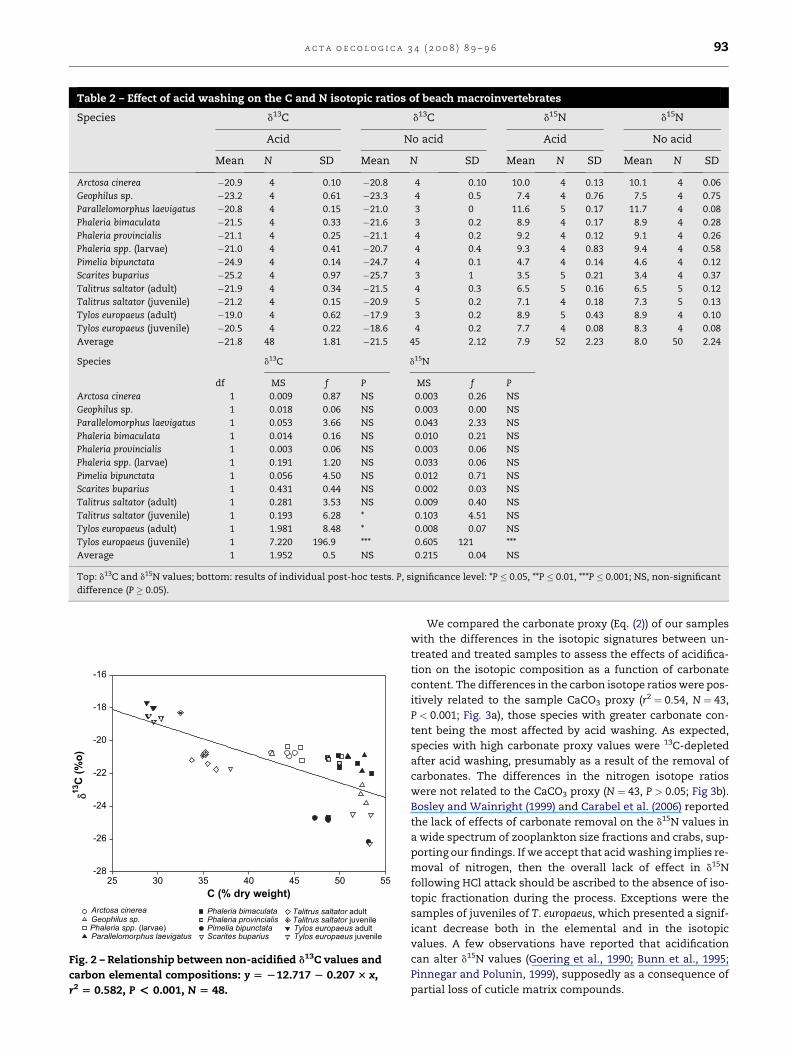
Table 2 – Effect of acid washing on the C and N isotopic ratios of beach macroinvertebrates
Species d13C d13C d15N d15N
Acid No acid Acid No acid
Mean N SD Mean N SD Mean N SD Mean N SD
Arctosa cinerea �20.9 4 0.10 �20.8 4 0.10 10.0 4 0.13 10.1 4 0.06
Geophilus sp. �23.2 4 0.61 �23.3 4 0.5 7.4 4 0.76 7.5 4 0.75
Parallelomorphus laevigatus �20.8 4 0.15 �21.0 3 0 11.6 5 0.17 11.7 4 0.08
Phaleria bimaculata �21.5 4 0.33 �21.6 3 0.2 8.9 4 0.17 8.9 4 0.28
Phaleria provincialis �21.1 4 0.25 �21.1 4 0.2 9.2 4 0.12 9.1 4 0.26
Phaleria spp. (larvae) �21.0 4 0.41 �20.7 4 0.4 9.3 4 0.83 9.4 4 0.58
Pimelia bipunctata �24.9 4 0.14 �24.7 4 0.1 4.7 4 0.14 4.6 4 0.12
Scarites buparius �25.2 4 0.97 �25.7 3 1 3.5 5 0.21 3.4 4 0.37
Talitrus saltator (adult) �21.9 4 0.34 �21.5 4 0.3 6.5 5 0.16 6.5 5 0.12
Talitrus saltator (juvenile) �21.2 4 0.15 �20.9 5 0.2 7.1 4 0.18 7.3 5 0.13
Tylos europaeus (adult) �19.0 4 0.62 �17.9 3 0.2 8.9 5 0.43 8.9 4 0.10
Tylos europaeus (juvenile) �20.5 4 0.22 �18.6 4 0.2 7.7 4 0.08 8.3 4 0.08
Average �21.8 48 1.81 �21.5 45 2.12 7.9 52 2.23 8.0 50 2.24
Species d13C d15N
df MS f P MS f P
Arctosa cinerea 1 0.009 0.87 NS 0.003 0.26 NS
Geophilus sp. 1 0.018 0.06 NS 0.003 0.00 NS
Parallelomorphus laevigatus 1 0.053 3.66 NS 0.043 2.33 NS
Phaleria bimaculata 1 0.014 0.16 NS 0.010 0.21 NS
Phaleria provincialis 1 0.003 0.06 NS 0.003 0.06 NS
Phaleria spp. (larvae) 1 0.191 1.20 NS 0.033 0.06 NS
Pimelia bipunctata 1 0.056 4.50 NS 0.012 0.71 NS
Scarites buparius 1 0.431 0.44 NS 0.002 0.03 NS
Talitrus saltator (adult) 1 0.281 3.53 NS 0.009 0.40 NS
Talitrus saltator (juvenile) 1 0.193 6.28 * 0.103 4.51 NS
Tylos europaeus (adult) 1 1.981 8.48 * 0.008 0.07 NS
Tylos europaeus (juvenile) 1 7.220 196.9 *** 0.605 121 ***
Average 1 1.952 0.5 NS 0.215 0.04 NS
Top: d13C and d15N values; bottom: results of individual post-hoc tests. P, significance level: *P � 0.05, **P � 0.01, ***P � 0.001; NS, non-significant
difference (P � 0.05).
C (% dry weight)
25 30 35 40 45 50 55
13C
(%
o)
-28
-26
-24
-22
-20
-18
-16
Arctosa cinerea
Geophilus sp.Phaleria spp. (larvae)Parallelomorphus laevigatus
Phaleria bimaculata
Phaleria provincialis
Pimelia bipunctata
Scarites buparius
Talitrus saltator adultTalitrus saltator juvenileTylos europaeus adultTylos europaeus juvenile
Fig. 2 – Relationship between non-acidified d13C values and
carbon elemental compositions: y [ L12.717 L 0.207 3 x,
r2 [ 0.582, P < 0.001, N [ 48.
We compared the carbonate proxy (Eq. (2)) of our samples
with the differences in the isotopic signatures between un-
treated and treated samples to assess the effects of acidifica-
tion on the isotopic composition as a function of carbonate
content. The differences in the carbon isotope ratios were pos-
itively related to the sample CaCO3 proxy (r2 ¼ 0.54, N ¼ 43,
P < 0.001; Fig. 3a), those species with greater carbonate con-
tent being the most affected by acid washing. As expected,
species with high carbonate proxy values were 13C-depleted
after acid washing, presumably as a result of the removal of
carbonates. The differences in the nitrogen isotope ratios
were not related to the CaCO3 proxy (N ¼ 43, P > 0.05; Fig 3b).
Bosley and Wainright (1999) and Carabel et al. (2006) reported
the lack of effects of carbonate removal on the d15N values in
a wide spectrum of zooplankton size fractions and crabs, sup-
porting our findings. If we accept that acid washing implies re-
moval of nitrogen, then the overall lack of effect in d15N
following HCl attack should be ascribed to the absence of iso-
topic fractionation during the process. Exceptions were the
samples of juveniles of T. europaeus, which presented a signif-
icant decrease both in the elemental and in the isotopic
values. A few observations have reported that acidification
can alter d15N values (Goering et al., 1990; Bunn et al., 1995;
Pinnegar and Polunin, 1999), supposedly as a consequence of
partial loss of cuticle matrix compounds.

a c t a o e c o l o g i c a 3 4 ( 2 0 0 8 ) 8 9 – 9 694
Adults and juveniles of T. europaeus are the species for
which estimated carbonate proxy was highest (mean �SD ¼ 0.13 � 0.06 and 0.21 � 0.02, respectively). Also, the car-
bonate proxy of T. saltator adults and juveniles was found to
be high (0.10 � 0.04 and 0.09 � 0.04, respectively) in relation
to the other beach macrofauna studied (average, 0.01 � 0.03,
ranging from �0.04 to 0.04). The analysis of presumably
CaCO3-free species, such as Phaleria spp. larvae, showed
negative carbonate proxy values (�0.04 � 0.03).
Carbonates contain a low proportion of carbon and do not
contain nitrogen. Malacostraceans (which yielded the highest
carbonate proxy values) also present the lowest carbon and
nitrogen relative content (mean � SD ¼ 32.7% � 3.2 and
6.4 � 1.0%, respectively). Consistently, species with low car-
bonate proxy values show higher carbon and nitrogen content
Carbonate proxy
-0,1 0,0 0,1 0,2-1,5
-1,0
-0,5
0,0
0,5
1,0
1,5
2,0
2,5
13C
cru
de -
13C
acid
-0,10 -0,05 0,00 0,05 0,10 0,15 0,20 0,25-0,4
-0,2
0,0
0,2
0,4
0,6
0,8
1,0
15N
cru
de -
15N
acid
Carbonate proxy
a
b
Arctosa cinerea
Geophilus sp.Phaleria spp. (larvae)Parallelomorphus laevigatus
Phaleria bimaculata
Phaleria provincialis
Pimelia bipunctata
Scarites buparius
Talitrus saltator adultTalitrus saltator juvenileTylos europaeus adultTylos europaeus juvenile
Fig. 3 – (a) Relationship between the carbonate proxy and
the difference in d13C between acid washed and untreated
samples ( y [ 0.025 D 0.085 3 x, r2 [ 0.54, P < 0.001,
N [ 43). (b) Relationship between the carbonate proxy and
the difference in d15N between acid washed and untreated
samples.
(48.9 � 3.9% and 11.0 � 0.9%, respectively). Due to the limited
number of species studied in our experiments, other causes
for the C/N ratio differences between groups should not be
discarded.
The relationship between carbonate content and the acid
washing effect on the carbon isotope signatures suggests
that only the carbon isotope ratios of those beach arthropods
with a carbonate proxy equal to or higher than 0.09, are
significantly altered after acidification. Jacob et al. (2005) ob-
served the same trends studying a wide range of species;
those samples with high carbonate proxies were 13C-depleted
after being acidified. Our results suggest that acid attacks
carbonates when present, but also alters chitin structure,
proteins and other tissues. Other studies report that non-
acidified whole macroinvertebrates with high proportions of
hard parts can be enriched in 13C by 2.5& compared to acid-
ified duplicates (Carabel et al., 2006). Bunn et al. (1995) also
reported that when the proportion of exoskeletons to body
mass is small, the effect of acid washing on the d13C values
may not be detectable. Soreide et al. (2006) studied the con-
troversy around the effect of the different methodologies
used prior to isotopic analysis and published C and N ele-
mental composition data of the marine species Gammarus
wilkitzkii and Thysanoessa inermis. We calculated an average
carbonate proxy including all suitable data from Soreide
et al. (2006) (mean � SD ¼ 0.77 � 0.3, N ¼ 12). The carbonate
proxy calculated for that data showed much higher CaCO3
content in comparison with our data. Aquatic crustaceans
are expected to be CaCO3-enriched because the lower impact
of gravity allows them to carry stronger and heavier protec-
tive structures (Little, 1983). Unfortunately, data on the ele-
mental composition of macroinvertebrates is very scarce in
the literature, making detailed carbonate proxy modeling
impracticable.
The carbonate proxy proposed by Jacob et al. (2005) presup-
poses that there is no effect of acidification on dietary carbon
and nitrogen. If there are no carbonates in the sample, the
quotient between the C/N ratios of raw and acidified aliquots
is 1, Eq. (2) becoming 0. It has been shown that the elemental
composition of either carbonate-free or carbonate-containing
samples is also apparently altered by acid washing. This
means that even under the assumption of a carbonate-free
sample, Eq. (2) would provide a value. A clear example of
this is represented by the carbonate proxy negative value of
Phaleria larvae in Fig. 3a,b. These samples are supposed to be
carbonate-free. A negative value strongly suggests a loss of di-
etary nitrogen due to acidification. In consequence, C:Nacidified
becomes higher than C:Nuntreated and hence Eq. (2) yields
a negative value. So in the light of our results, the carbonate
proxy becomes, in our opinion, less meaningful. In fact, it
should be considered a ‘‘carbonate þ acid-soluble com-
pounds’’ proxy, rather than a carbonate proxy alone. The
outcome of the application of the carbonate proxy to our
results (linearity with the suspected carbonate content of
the samples) indicates that, as already discussed, the isotopic
shifts are larger when the carbonate content of the sample is
high. The use of soft acidification procedures (see Section 4)
that limit the removal of dietary carbon and nitrogen would
result in a much more meaningful applicability of the carbon-
ate proxy.

a c t a o e c o l o g i c a 3 4 ( 2 0 0 8 ) 8 9 – 9 6 95
4. Conclusions
One initial conclusion is that, in the light of the results pre-
sented, the processes taking place as a consequence of acidifi-
cation are more complex than initially thought. This is
a consequence of the high chemical inter- and intra-specific
heterogeneity of the different macroinvertebrates (Welinder,
1974). In the case of semi-terrestrial crustaceans, the changes
observed in the d13C are large enough to critically confound
food web analysis. As shown, the problem associated with
acid washing (1 N HCl) is that some dietary carbon- and nitro-
gen-bearing compounds are being removed together with non-
dietary ones resulting in a substantial change in the C and N
content of the samples. The removal of carbonates from
semi-terrestrial crustaceans would require a technique that
ensured no effect on non-carbonate carbon fractions. Percot
et al. (2003) proposed and demonstrated that a concentration
of HCl of 0.25 M for 15 min was enough for complete deminer-
alization minimizing the hydrolysis of chitin polymers of the
exoskeletons, this being the most appropriate protocol for
the acid washing treatment. In order to avoid isotopic variabil-
ity due to acid washing samples we recommend using non-
acidified soft tissues whenever possible for isotopic analyses.
The results presented strongly support that acidification of
beach macroinvertebrates could be omitted in all cases except
when the study is interested in the d13C values of inverte-
brates with a carbonate proxy >0.09 (as for the case of semi-
terrestrial crustaceans in this study). In this last case, a soft
decalcification should be applied and untreated aliquots
should be analyzed separately for d15N.
Other important methodological procedures (lipid extrac-
tion, distilled water washing, gut content, etc.) critically af-
fecting isotopic signatures of marine macroinvertebrates are
reviewed in Mateo et al. (in press).
Acknowledgements
The authors are grateful to the two reviewers of this paper for
their helpful contributions. This study has been supported
partially by funds of a bilateral programme between the CNR
(Italy) and the CSIC (Spain), and partially by the EU-INCO
project WADI (CT2005-15226).
r e f e r e n c e s
Amiji, M.M., 1998. Platelet adhesion and activation on anamphoteric chitosan derivative bearing sulfonate groups.Colloids Surf B Biointerfaces 10, 263–271.
Bouillon, S., Raman, A.V., Dauby, P., Dehairs, F., 2002. Carbon andnitrogen stable isotope ratios of subtidal benthic invertebratesin an estuarine mangrove ecosystem (Andhra Pradesh, India).Estuar. Coast. Shelf Sci. 54, 901–913.
Bosley, K.L., Wainright, S.C., 1999. Effects of preservatives andacidification on the stable isotope ratios (15N:14N, 13C:12C) oftwo species of marine animals. Can. J. Fish. Aquat. Sci. 56,2181–2185.
Bunn, S.E., Loneragan, N.R., Kempster, M.A., 1995. Effects of acidwashing on stable isotope ratios of C and N in penaeid shrimp
and seagrass: implications for food-web studies usingmultiple stable isotopes. Limnol. Oceanogr. 40, 622–625.
Carabel, S., Godınez-Domınguez, E., Verısimo, P., Fernandez, L.,Freire, J., 2006. An assessment of sample processing methodsfor stable isotope analyses of marine food webs. J. Exp. Mar.Biol. Ecol. 336, 254–261.
Chanton, J.P., Lewis, F.G., 1999. Plankton and dissolved inorganiccarbon isotopic composition in a river-dominated estuary:Apalachicola Bay, Florida. Estuaries 22, 575–583.
Cloern, J.E., Canuel, E.A., Harris, D., 2002. Stable carbon andnitrogen isotope composition of aquatic and terrestrial plantsof the Sant Francisco Bay estuarine system. Limnol. Oceanogr.47 (3), 713–729.
DeNiro, M.J., Epstein, S., 1981. Isotopic composition of cellulosefrom aquatic organisms. Geochim. Cosmochim. Acta 45 (10),1885–1894.
Fry, B., Sherr, E.B., 1984. d13C measurements as indicators ofcarbon flow in marine and freshwater ecosystems. Contrib.Mar. Sci. 27, 13–47.
Fry, B., 1988. Food web structure on Georges Bank from stable C,N, and S isotopic compositions. Limnol. Oceanogr. 33 (5), 1182–1190.
Fry, B., 2006. Stable Isotope Ecology. Springer-Verlag, New York.Giraud-Guille, M.M., Quintana, C., 1982. Secondary ion
microanalysis of the crab calcified cuticle: distribution ofmineral elements and interaction with the cholesteric organicmatrix. Biol. Cell 44, 57–68.
Goering, J., Alexander, V., Haubenstock, N., 1990. Seasonalvariability of stable carbon and nitrogen isotope ratios oforganisms in a North Pacific Bay. Estuar. Coast Shelf Sci. 30,239–260.
Gorokhova, E., Hansson, S., 2000. Elemental composition of Mysismixta (Crustacea, Mysidacea) and energy costs of reproductionand embryogenesis under laboratory conditions. J. Exp. Mar.Biol. Ecol. 246, 103–123.
Grey, J., Jones, R.I., Sleep, D., 2001. Seasonal changes in theimportance of the source of organic matter to the diet ofzooplankton in Loch Ness, as indicated by stable isotopeanalysis. Limnol. Oceanogr. 46 (3), 505–513.
Hesslein, R.H., Hallard, K.A., Ramlal, P., 1993. Replacement ofsulfur, carbon, and nitrogen in tissue of growing broadwhitefish (Coregonus nasus) in response to a change in diettraced by delta d34S, delta d13C, and delta d15N. Can. J. Fish.Aquat. Sci. 50 (10), 2071–2076.
Hobson, K.A., Welch, H.E., 1992. Determination of trophicrelationships within a High Arctic marine food web using deltad13C and delta d15N analysis. Mar. Ecol. Prog. Ser. 84 (1), 9–18.
Hobson, K.A., Fisk, A., Karnovsky, N., Holst, M., Gagnon, J.,Fortier, M., 2002. A stable isotope (d13C, d15N) model for theNorth Water food web: implications for evaluatingtrophodynamics and the flow of energy and contaminants.Deep Sea Res. Part II Top. Stud. Oceanogr 49, 5131–5150.
Jackson, D., Harknes, D.D., Mason, C.F., Long, S.P., 1986. Spartinaanglica as a carbon source for salt-marsh invertebrates:a study using d13C. Oikos 46, 163–170.
Jacob, U., Mintenbeck, K., Brey, T., Knust, R., Beyer, K., 2005. Stableisotope food web studies: a case for standardized sampletreatment. Mar. Ecol. Prog. Ser. 287, 251–253.
Jennings, S., Renones, O., Morales-Nin, B., Polunin, N.C.V.,Moranta, J., Coll, J., 1997. Spatial variation in the 15N and 13Cstable isotope composition of plants, invertebrates and fisheson Mediterranean reefs: implications for the study of trophicpathways. Mar. Ecol. Prog. Ser. 146 (1-3), 109–116.
Kump, L.R., Arthur, M.A., 1999. Interpreting carbon-isotopeexcursions: carbonates and organic matter. Chem. Geol. 161,181–198.
Little, C., 1983. The Colonisation of Land: Origins and Adaptationsof Terrestrial Animals. University Press, Cambridge.

a c t a o e c o l o g i c a 3 4 ( 2 0 0 8 ) 8 9 – 9 696
Luquet, G., Marin, F., 2004. Biomineralisations in crustaceans:storage strategies. C.R. Pale vol. 3, 515–534.
Mateo, M.A., Serrano, O., Serrano, L., Michener, R.,(in press).Effects of sample preparation on stable isotope ratios of C andN in marine invertebrates: implications for food web studiesusing stable isotopes. Oecologia (in press).
Ng, J.S.S., Wai, T.-C., Williams, G.A., 2007. The effects ofacidification on the stable isotope signatures of marine algaeand molluscs. Mar. Chem. 103, 97–102.
Nieuwenhuize, J., Maas, Y.E.M., Middelburg, J.J., 1994. Rapidanalysis of organic carbon and nitrogen in particulatematerials. Mar. Chem. 45, 217–224.
Nyssen, F., Brey, T., Lepoint, G., Bouquegneau, J.-M., De Broyer, C.,Dauby, P., 2002. A stable isotope approach to the easternWeddell Sea trophic web: focus on benthic amphipods. PolarBiol. 25, 280–287.
Nyssen, F., Brey, T., Dauby, P., Graeve, M., 2005. Trophic position ofAntarctic amphipods – enhanced analysis by a 2-dimensionalbiomarker assay. Mar. Ecol. Prog. Ser. 300, 135–145.
Owens, N.J.P., 1987. Natural variations in 15N in the marineenvironment. Adv. Mar. Biol. 24, 389–451.
Owens, N.J.P., 1988. Rapid and total automation of shipboard d15Nanalysis: Examples from the North Sea. J. Exp. Mar. Biol. Ecol122 (2), 163–171.
Percot, A., Viton, C., Domard, A., 2003. Optimization of chitinextraction from shrimp shells. Biomacromolecules 4, 12–18.
Pinnegar, J.K., Polunin, N.V.C., 1999. Differential fractionation ofdelta d13C and delta d15N among fish tissues: implications forthe study of trophic interactions. Funct. Ecol 13 (2), 225–231.
Pinnegar, J.K., Polunin, N.V.C., 2000. Contributions of stable-isotope data to elucidating food webs of Mediterranean rockylittoral fishes. Oecologia 122 (3), 399–409.
Polunin, N.V.C., Morales-Nin, B., Pawsey, W.E., Cartes, J.E.,Pinnegar, J.K., Moranta, J., 2001. Feeding relationships inMediterranean bathyal assemblages elucidated by stablenitrogen and carbon isotope data. Mar. Ecol. Prog. Ser. 220, 13–23.
Powers, L.W., Bliss, D.E., 1983. Terrestrial adaptations. In:Vernberg, F.J., Vernberg, W.B. (Eds.), The Biology of Crustacea,vol. 8. Academic Press, New York, pp. 271–331.
Preston, T., 1992. The measurement of stable isotope naturalabundance variations. Plant Cell Environ 15, 1091–1097.
Rau, G.H., Mearns, A.J., Young, D.R., Olson, R.J., Schafer, H.A.,Kaplan, I.R., 1983. Animal 13C/12C correlates with trophic levelin pelagic food webs. Ecology 64, 1314–1318.
Rau, G.H., Ainley, D.G., Benqtson, J.L., Torres, J.J., Hopkins, T.L.,1992. 15N/14N and 13C/12C in Weddell Sea birds, seals, and fish:Implications for diet and trophic structure. Mar. Ecol. Prog.Ser. 84 (1), 1–8.
Shafer, T.H., Roer, R.D., Miller, C.G., Dillaman, R.M., 1994. Post-ecdysial changes in the protein and glycoprotein compositionof the cuticle of the blue crab Callinectes sapidus. J. Crust. Biol.14, 210–219.
Schimmelmann, A., DeNiro, M.J., 1985. Stable isotopic studies onchitin. Measurements on chitin/chitosan isolates and D-glucosamine hydrochloride from chitin. In: Muzzarelli, R.A.A.,Jeuniaux, C., Gooday, G.W. (Eds.), Chitin in Nature andTechnology. Plenum Press, New York.
Soreide, J.E., Tamelander, T., Hop, H., Hobson, K.A., Johansen, I.,2006. Sample preparation effects on stable C and N isotopevalues: a comparison of methods in Arctic marine food webstudies. Mar. Ecol. Prog. Ser. 328, 17–28.
Spicer, J.I., Moore, P.G., Taylor, A.C., 1987. The physiologicalecology of land invasion by the Talitridae (Crustacea:Amphipoda). Proc. R. Soc. Lond. B Biol. Sci. 232, 95–124.
Travis, D.F., 1963. Structural features of mineralization fromtissue to macromolecular levels of organization in thedecapod Crustacea. Ann. N.Y. Acad. Sci. 109, 177–245.
Vizzini, S., Mazzola, A., 2003. Seasonal variations in the stablecarbon and nitrogen isotope ratios (13C/12C and 15N/14N) ofprimary producers and consumers in a westernMediterranean coastal lagoon. Mar. Biol. 142, 1009–1018.
Yokohama, H., Tamaki, A., Harada, K., Shimoda, K., Koyama, K.,Ishihi, Y., 2005. Variability of diet-tissue isotopicfractionation in estuarine macrobenthos. Mar. Ecol. Prog. Ser.296, 115–128.
Welinder, B.S., 1974. The crustacean cuticle – I. Studies on thecomposition of the cuticle. Comp. Biochem. Physiol. 47A,779–787.




















