
Arthropods, 2013, Vol. 2, Iss. 3
59
Arthropods Vol. 2, No. 3, 1 September 2013 International Academy of Ecology and Environmental Sciences
Transcript of Arthropods, 2013, Vol. 2, Iss. 3

Arthropods
Vol. 2, No. 3, 1 September 2013
International Academy of Ecology and Environmental Sciences

Arthropods ISSN 2224-4255 Volume 2, Number 3, 1 September 2013 Editor-in-Chief WenJun Zhang Sun Yat-sen University, China International Academy of Ecology and Environmental Sciences, Hong Kong E-mail: [email protected], [email protected] Editorial Board Andre Bianconi (Sao Paulo State University (Unesp), Brazil) Anton Brancelj (National Institute of Biology, Slovenia) Hans-Uwe Dahms (Sangmyung University, Korea) A. K. Dhawan (Punjab Agricultural University, India) John A. Fornshell (Northern Virginia Community College, USA) Xin Li (Northwest A&F University, China) Oscar E. Liburd (University of Florida, USA) Ivana Karanovic (Hanyang University, Korea) Enoch A Osekre (KN University of Science and Technology, Ghana) Rajinder Peshin (Sher-e-Kashmir University of Agricultural Sciences and Technology of Jammu, India) Michael Stout (Louisiana State University Agricultural Center, USA) Eugeny S. Sugonyaev (Russian Academy of Sciences, Russia)
Editorial Office: [email protected] Publisher: International Academy of Ecology and Environmental Sciences Address: Flat C, 23/F, Lucky Plaza, 315-321 Lockhart Road, Wanchai, Hong Kong Tel: 00852-6555 7188 Fax: 00852-3177 9906 Website: http://www.iaees.org/ E-mail: [email protected]

Arthropods, 2013, 2(3): 95-104
IAEES www.iaees.org
Article
The locomotory rhythmic activity in scorpions: with a review
Michael R. Warburg
Dept. of Biology, Technion-Israel Institute of Technology, Haifa 32000, Israel
E-mail: [email protected]
Received 21 February 2013; Accepted 25 March 2013; Published online 1 September 2013
Abstract
Locomotory rhythmic behavior is entrained by the change between photophase and skotophase and to some
extent by thermal conditions. In many species studied most activity takes place during early night hours. Some
species show completely a nocturnal activity pattern, whereas a few species are entirely diurnal. There does
not appear to be a pattern related to the timing and extent of the photophase. Except perhaps for Leiurus
quinquestriatus (Hemprich and Ehrenberg, 1829) which appears to be less active at the highest temperature.
This subject was studied in 30 species of scorpions most of them buthids (53.3%), that were studied so far in
42 different studies.
Keywords Scorpiones; diel locomotory rhythm; thermal ‘Zeitgeber’.
1 Introduction
The behaviour of scorpions has received less attention than it perhaps should have (Warburg and Polis, 1990).
Most cited studies were on rhythmic activity of scorpions. Here studies were divided between those on the
optic ‘Zeitgeber’ or time giver that bring about rhythmic activity (Warburg, in preparation). However most
studies were conducted on the behaviour of locomotory rhythmic activity. The activity patterns described here
are entrained by ambient conditions largely photophase and skotophase and to a certain extent also by thermal
factors. Such entrainment factor or ‘Zeitgeber’ is the factor that brings about this rhythmic activity (Cloudsley-
Thompson, 1961, 1978).
Dube and Fleissner (1985) describe three types of movements while in the wheel-running apparatus: (1)
slow pace movements typical of a circadian rhythms of 12L/12D light regime; (2) long-lasting runs, and (3)
rapid locomotion.
It was already Wuttke (1966) who described the bimodal activity in Euscorpius carpathicus (Linnaeus,
1767). In some species no rhythmic activity was noticeable. Thus, Cloudsley-Thompson and Constantinou
(1985) studying Opisthacanthus sp which did not show any rhythm. Cloudsley-Thompson (1973) found in
Buthotus minax (Koch, 1875) that the circadian locomotory rhythm is entrained by regular transition from dark
Arthropods ISSN 22244255 URL: http://www.iaees.org/publications/journals/arthropods/onlineversion.asp RSS: http://www.iaees.org/publications/journals/arthropods/rss.xml Email: [email protected] EditorinChief: WenJun Zhang Publisher: International Academy of Ecology and Environmental Sciences

Arthropods, 2013, 2(3): 95-104
IAEES www.iaees.org
to light and by rising temperatures. A similar situation was seen in Babycurus centrurimorphus (Karsch, 1886)
by Cloudsley-Thompson (1975) who studied this buthid and found that diurnal rhythm was entrained by shifts
both from dark to light and changes in temperatures.
Many of these studies show that activity is highest during the first few hours of the night (Cloudsley-
Thompson, 1978, 1980) whereas others show activity peaking at night. Thus, Cloudsley-Thompson found a
peak of activity in Scorpio maurus, Buthus occitanus (Amoreux, 1789) and Androctonus australis (Linnaeus,
1758) during the hours of 1800-2000 PM. The latter species was largely nocturnal (Constantinou, 1980). In
(1978) Cloudsley-Thompson (Tab. 1 therein) summarized the situation in the scorpion species studied up till
then. El Bakary and Fuzeau-Braesch (1988) studied: Leiurus quinquestriatus (Hemprich and Ehrenberg, 1829)
using three methods of detecting locomotory activity. They too found bimodal onset of activity coincides with
onset of photophase and skotophases.
Benton (1992) studying Euscorpius flavicaudis (DeGeer, 1778), found them active at dusk and dawn. In
this study I shall review the subject of locomotory rhythms in scorpions. This would include only the rhythmic
activities involving locomotion and not any rhythmic physiological functions. These were discussed recently
(Warburg, 2013). This subject was studied in 30 species of scorpions that were studied so far in 42 different
studies, listed as the follows:
Buthidae (16 species): 1. Androctonus australis (Linnaeus, 1758) Constantinou (1980); Baz et al. (2009) 2. Anomalobuthus rickmersi (Kraepelin, 1900) Fet (1980) 3. Babycurus centrurimorphus Karsch, 1886 Cloudsley-Thompson (1975) 4. Buthus hottentotta Fabricius, 1787 Toye (1970) 5. Buthus occitanus (Amoreux, 1789) Constantinou (1980); Constantinou & Cloudsley-Thompson (1980) 6. Buthotus occitanus (Amoreux, 1789) Skutelsky (1996) 7. Buthotus minax (L. Koch, 1875) Cloudsley-Thompson (1963, 1973) 8. Centruroides sculpturatus Ewing, 1928 Hadley & Williams (1968); Crawford & Krehoff (1975) 9. Hottentotta judaicus (E. Simon, 1872) Warburg & Ben-Horin (1979) 10. Leiurus quinquestriatus (Hemprich & Ehrenberg, 1829) Cloudsley-Thompson (1963); Abushama (1963); El Bakary & Fuzeau-Braesch (1988); Warburg & Ben-Horin (1979) 11. Liobuthus kessleri (Birula, 1898) Fet (1980) 12. Mesobuthus gibbosus (Brullé, 1832) Kaltsas & Mylonas (2010) 13. Mesobuthus eupeus (C. Koch, 1813) Fet (1980) 14. Mesobuthus caucasicus (Nordmann, 1840) Fet (1980) 15. Orthochirus scrobilosus (Grube, 1873) Fet (1980) 16. Parabuthus villous (Peters, 1862) Harrington (1981)
Euscorpiidae (2 species):
96

Arthropods, 2013, 2(3):
IAEES www.iaees.org
17. Euscorpius carpathicus (Linnaeus, 1767) Wuttke (1966) 18. Euscorpius flavicaudis (DeGeer, 1778) Cloudsley-Thompson & Constantinou (1983); Benton (1992) Cloudsley-Thompson (1963)
Hemiscorpiidae (1 species): 19. Hadogenes bicolor (Purcell, 1899) Constantinou (1980); Constantinou & Cloudsley-Thompson (1980)
Vaejovidae (3 species): 20. Paruroctonus boreus (Girard, 1854) Tourtlotte (1974) 21. Vaejovis mesaensis probably Paruroctonus mesaensis now Smeringurus mesaensis (Stahnke, 1957) Hadley & Williams (1968); Polis (1980) 22. Vaejovis confusus Stahnke, 1940 Hadley & Williams (1968)
Scorpionidae (8 species): 23. Diplocentrus spitzeri. Stahnke, 1970 Crawford & Krehoff (1975) 24. Nebo hierichonticus (Simon, 1872) Warburg & Ben-Horin (1979) 25. Pandinus gregoryi (Pocock, 1896) Constantinou (1980); Constantinou & Cloudsley-Thompson (1980) 26. Pandinus exitialis (Pocock, 1888) Cloudsley-Thompson (1963) 27. Pandinus imperator (C.L. Koch, 1841) Toye (1970) 28. Scorpio maurus fuscus Hemprich & Ehrenberg, 1829 Warburg & Ben-Horin (1979) 29. Heterometrus swammerdami (E. Simon, 1872) Cloudsley-Thompson (1981) 30. Heterometrus fulvipes (C.L. Koch, 1838) Babu, Reddy & Kasaiah (1988)
2 Materials and Methods
Measuring activity in scorpions was carried out largely by actograph use. The actograph used here consists of a
Perspex box measuring 5 x 6 x 20cm that rotated at its mid-point on a pivot. At one side an electrical contact
closed a circuit thereby signaling a change in the position of the actograph that resulted from the scorpion
moving inside the box. These movements were recorded on a Model 712 Telrad Recorder during the 24h
experiment. This actograph apparatus was placed into a Struers Refritherm where temperature was controlled
at ±0.50C. The animals could be observed through a window in the Refritherm’s door (see Warburg and Ben-
Horin, 1979).
The scorpion species studied here were collected from the field. Four scorpion species were used here:
Scorpio maurus fuscus (Hemprich and Ehrenberg, 1829), Nebo hierichonticus (Simon, 1872) Hotenttotta
judaicus (Simon, 1872) and L. quinquestriatus.
3 Results
Diel Activity in three scorpion species is given as percentage of time spent in activity during four-six hour
watches (Fig. 1). In all three species studied here most activity (between 55.3-87.6%) was spent active during
the 1st watch (06 PM- 12). The three scorpion species differed in the amount of time spent during the
97

Arthropods, 2013, 2(3): 95-104
IAEES www.iaees.org
remainder of the day. Thus, whereas N. hierichonticus and H. judaicus had shown hardly any activity (12.3%
and 21.8% respectively), S. m. fuscus was active also during the remaining of the day (44.8%).
Fig. 1 Diel activity (%) in three scorpion species.
The number of activity runs at four temperatures (200C, 250C, 280C, 350C) was measured in four scorpion
species (Fig. 2). These scorpion species differed in their response in a temperature gradient. Thus, the peak of
activity was at the lowest temperature at 200C in L. quinquestriatus (55), and in N. hierichonticus (40.6),
whereas for S. m. fuscus and H. judaicus it was at 250C (69.1 and 41 respectively). At the highest temperature
tested here (350C) the two buthid species (L. quinquestriatus and H. judaicus) have shown the lowest activity
(5.5 and 16.3 respectively).
When the number of activity runs was compared in illuminated and dark parts of the thermo-preferendum
apparatus (Fig. 3), outstanding differences in the activity of the four scorpion species were noticeable. Thus in
N. hierichonticus and S. m. fuscus there was no marked difference between the activity in the dark and
illuminated parts of the thermo-preferendum apparatus. In the two species activity was higher in the
illuminated part. In the first species it was 29.6 at 200C, whereas in the latter species it was 29.6 at 250C. In the
two buthids activity was higher in the darkened part. Thus in H. judaicus it was highest at 250C (32.5) whereas
in L. quinquestriatus it peaked at 280C (34.4).
98

Arthropods, 2013, 2(3):
IAEES www.iaees.org
Fig. 2 Average number of activity runs at four temperatures in four scorpion species.
Lastly, the time spent in the thermo-preferendum apparatus was examined in detail in 300C thermal zones
ranging between 150C–300C (Fig. 4). This was examined in S.m. fuscus and N. hierichonticus. Both species
have shown similar results with activity rising between 210C-270C (27.8-37.3 in the first species, and 30-36.7
in the second species), dropping thereafter between 270C-300C (to 23.3 in the first species, and 8.7 in the latter).
4 Discussion
4.1 Activity during daytime
Constantinou (1980) studied four scorpion species: Pandinus gregoryi (Pocock, 1896) showed diurnal activity
most of its time (63%), Hadogenes bicolor (Purcell, 1899) with 26% activity diurnal, A. australis was largely
nocturnal, as was also B. occitanus. Constantinou and Cloudsley-Thompson (1980) studied four different
scorpion species: Scorpio maurus, B. occitanus. P. gregoryi and H. bicolor. They found the last two are
largely diurnal as was the case also with Parabuthus villosus (Peters, 1862) where Harrington (1981) found it
to be diurnal.
55
36.139.4
5.5
0
10
20
30
40
50
60
20 25 28 35
Av. n
o. ru
ns
Temp. (C)
L. quinquestriatus
20
41
24.3
16.3
05
1015202530354045
20 25 28 35
Av. n
o. ru
ns
Temp. (C)
H. judaicus
40.6
24.6
13
28.5
05
1015202530354045
20 25 28 35
Av. n
o. ru
ns
Temp. (C)
N. hierichonticus
39.8
69.1
56.9
30
01020304050607080
20 25 28 35
Av. n
o. ru
ns
Temp. (C)
S. m. fuscus
99

Arthropods, 2013, 2(3): 95-104
IAEES www.iaees.org
Fig. 3 Average number of activity runs in illuminated (grey bars) and dark arenas.
4.2 Activity during early evening
Hadley and Williams (1968) studied three species of scorpions: Vaejovis confusus (Stahnke, 1940), V.
mesaensis (Stahnke, 1957) and Centruroides sculpturatus (Ewing, 1928). All of them showed peak activity in
the evening. Similarly, Toye (1970) studying both Pandinus imperator (Koch, 1841) and Buthus hottentotta
Fabricius, used an actograph made of celluloid tubes pivoting through transverse center. He too found that the
29
9.45
1.5
26 26.7
34.4
4
05
10152025303540
20 25 28 35
Av
. n
o.
acti
vit
y ru
ns
Temp. (C)
L. quinquestriatus
4
8.5 8.66
16
32.5
15.7
10.3
0
5
10
15
20
25
30
35
20 25 28 35
Av
. n
o. o
f ac
tiv
ity
run
s
Temp. (C)
H. judaicus
29.6
13.6 1511 11
13 13.5
0
5
10
15
20
25
30
35
20 25 28 35
Av
. n
o.
of
acti
vit
y ru
ns
Temp. (C)
N. hierichonticus
22.4
29.6
4
29
17.4
11
16
26
0
5
10
15
20
25
30
35
20 25 28 35
Av
. n
o. a
ctiv
ity
run
s
Temp. (C)
S. m. fuscus
100

Arthropods, 2013, 2(3):
IAEES www.iaees.org
peak activity was between 1500-1800h in both species. Polis (1980) studying the vaejovid Paruroctonus
mesaensis (Stahnke), found that surface activity increases during early evening decrease after 1400AM.
Warburg and Ben-Horin (1979) studied effect of temperature on rhythm of three scorpion species in an
actograph. They found in S. m. fuscus high activity 1200-1800h and 1800-2400h. This activity dropped
drastically with rising temps (to 280C and 350C). N. hierichonticus showed high activity between 1800-2400hr
during 06-12 hrs, whereas H. judaicus rhythm was not affected by temperature. Babu, Reddy and Kasaiah
(1988) studied Heterometrus fulvipes (Koch, 1838) with an actograph.
Fig. 4 Percentage time spent in the thermo-preferendum apparatus.
S.m. fuscus
2023.3
47.5
2.56.7
0
10
20
30
40
50
18-21 21-24 24-27 27-30 30-34
Temp. ranges (C)
Tim
e s
pe
nt
(%)
N.hierichopnticus
51.7
43.3
4.20.8
0
10
20
30
40
50
60
18-21 21-24 24-27 27-30 30-34
Temp. ranges (C)
Tim
e s
pe
nt
(%)
L.quinquestriatus
21.7
0.8
52.5
25
0
10
20
30
40
50
60
18-21 21-24 24-27 27-30 30-34
Temp. ranges (C)
Tim
e s
pe
nt
(%)
101

Arthropods, 2013, 2(3): 95-104
IAEES www.iaees.org
4.3 Activity during night
Cloudsley-Thompson (1963) Studied three scorpion species: Pandinus exitialis (Pocock, 1888), L.
quinquestriatus and Buthotus minax (see Fig. 2 therein). All of these species were nocturnal. One of these
scorpions L. quinquestriatus, was studied in an actograph apparatus box pivoted about its median transverse
axis writing on a barograph drum that acted as a kymograph (Abushama, 1963). He too found it to be
nocturnal following an endogenous factor of the rhythm.
On the other hand, Tourtlotte (1974) who studied surface activity of Paruroctonus boreus (Girard, 1854),
found its activity peaked at 2130h. Likewise, Crawford and Krehoff (1975) studying Centruroides sculpturatus
and Diplocentrus spitzeri (Stahnke, 1970), found that surface activity in the field peaked between 2030-2230h
whereas, activity in the actograph showed nocturnal activity. Only Centruroides spitzeri showed endogenous
circadian rhythm in constant darkness. Fet (1980) studied the buthids: Orthochirus scrobilosus (Grube, 1873),
Anomalobuthus rickmersi (Kraepelin, 1900), Mesobuthus eupeus (Koch, 1813), M. caucasicus (Nordmann
1840) and Liobuthus kessleri (Birula 1898). In all these species activity peaked between 2100-0100 h.
Cloudsley-Thompson (1981) studied Heterometrus swammerdami (Simon, 1872) and found it active
especially at night. Cloudsley-Thompson and Constantinou (1983) studying E. flavicaudis described their
nocturnal habits in the field and during actograph studies.
Baz, Sallam and El-Naggar (2009) studied A. australis locomotor activity rhythm. It was synchronized
with electroretinogram rhythm starting at night. Skutelsky (1996) studying: Buthotus occitanus israelis, found
its activity dropped with full moon. Kaltsas and Mylonas (2010) studying Meobuthus gibbosus found their
activity dropped with moonlight.
There are several points that emerge from this review:
1. Most of these studies (54%) were conducted on buthids that are usually the more active species.
2. Many of the studies did not remark anything about the ecology or physiological conditions of the scorpions
studied. This is especially important since in many studies the scorpions were previously kept for sometime
before the onset of the experiment.
3 The findings of these studies are not conclusive. Under what condition does a shift in behaviour of the
scorpion's rhythmic activity take place?
Acknowledgements
The assistance of A. Ben-Horin during his M.Sc. thesis work is gratefully acknowledged. The author is
indebted to Prof. Cloudsley-Thompson for providing an outlet to early publications, and to Dr. Polis for
inviting me to write a chapter in a book he was editing.
References
Abushama EF. 1963. Bioclimate, diurnal rhythms and water-loss in the scorpion, Leiurus quinquestriatus (H
& E). Entomological Monthly Magazine, 98: 216-224
Babu K.S, Reddy TG, Kasaiah A. 1988. Daily patterns of locomotor activity in the Indian scorpion
Heterometrus fulvipes (C.L. Koch). Indian Journal of Experimental Biology, 26: 307-311
Baz E-S, Sallam A E-D, El-Naggar MHR. 2009. Development of method for monitoring the locomotor and
visual circadian rhythms in a free-moving scorpion. Egyptian Journal of Natural Toxins, 6(2): 16-30
Benton TG. 1992. The ecology of the scorpion Euscorpius flavicaudis in England. Journal of Zoology
(London), 226: 351-368
Cloudsley-Thompson JL. 1956. Studies in diurnal rhythms – VI. Bioclimatic observations in Tunisia and their
102

Arthropods, 2013, 2(3):
IAEES www.iaees.org
significance to the physiology of the fauna, especially woodlice, centipedes, scorpions and beetles. Annals
and Magazine of Natural History, 12(9): 305-329
Cloudsley-Thompson JL. 1961. Observations on the biology of the scorpion Leiurus quinquestriatus (H. & E.)
in the Sudan. Entomological Monthly Magazine, 97: 153-155
Cloudsley-Thompson JL. 1963. Some aspects of the physiology of Buthotus minax (Scorpiones: Buthidae)
with remarks on other African scorpions. Entomological Monthly Magazine, 98: 243-246
Cloudsley-Thompson JL. 1973. Entrainment of the "circadian clock" in Buthotus minax (Scorpiones:
Buthidae). Journal of Intedisciplinary Cycle Research, 4(2): 119-123
Cloudsley-Thompson JL. 1975. Entrainment of "circadian clock" in Babycurus centrurimorphus (Scorpiones:
Buthidae). Journal of Intedisciplinary Cycle Research, 6: 185-188
Cloudsley-Thompson JL. 1978. Biological clocks in Arachnida. Bulletin of the British Arachnological Society,
4(4): 184-191
Cloudsley-Thompson JL. 1981. A comparison of rhythmic locomotory activity in tropical forest Arthropoda
with that in desert species. Journal of Arid Environments, 4: 327-334
Cloudsley-Thompson JL, Constantinou C. 1983. How does the scorpion Euscorpius flavicaudis.(Deg.) manage
to survive in Britain? International Journal of Biometeorology, 27(2): 87-92
Cloudsley-Thompson JL, Constantinou C. 1985. The circadian rhythm of locomotory activity in a Neotropical
forest scorpion, Opisthacanthus sp. (Scorpionidae). International Journal of Biometeorology, 29(1): 87-89
Constantinou C. 1980. Entrainment of the circadian rhythm of activity in desert and forest inhabiting scorpions.
Journal of Arid Environments, 3: 133-139
Constantinou C, Cloudsley-Thompson JL. 1980. Circadian rhythms in scorpions. 8th Internationaler
Arachnologen Congress Wien, 53-55
Crawford CS, Krehoff RC. 1975. Diel activity in sympatric populations of the scorpions Centuroides
sculpturatus (Buthidae) and Diplocentrus spirtzeri (Diplocerntridae). Journal of Arachnology, 2: 195-204
Dube C, Fleissner G. 1985. Temporal patterns of wheel-running behaviour in the scorpion Androctonus
australis. Journal of Intedisciplinary Cycle Research, 16(4): 250-251
El- Bakary Z, Fuzeau-Braesch S. 1988. Circadian rhythms and time measurement in locomotor activity of the
scorpion Leiurus quinquestriatus (Buthidae). Chronobiology Internationl, 5(2): 167-174
Fet VY. 1980. Ecology of the scorpions (Arachnida, Scorpiones) of the southeastern Kara-Kum.
Entomologoscheskoe Obozrenie, 59: 167-170
Hadley NF, Williams SC. 1968. Surface activities of some North American scorpions in relation to feeding.
Ecology, 49(4): 726-734
Harrington A. 1981. Diurnalism in Parabuthus villosus (Peters)(Scorpiones, Buthidae). Journal of
Arachnology, 10: 85-86
Kaltsas D, Mylonas M. 2010. Locomotory activity and orientation of Mesobuthus gibbosus (Scorpiones:
Buthidae) in central Aegean Archipelago. Journal of Natural History, 44(23-24): 1445-1459
Polis GA. 1980. Seasonal patterns and age-specific variation in the surface activity of a population of desert
scorpions in relation to environmental factors. Journal of Animal Ecology, 49: 1-18
Ramakrishna T, Rao KP. 1971. State determined system of a circadian rhythm in scorpion. Proceedings of the
Indian Academy of Science Section B, 73(4): 202-207
Skutelsky O. 1996. Predation risk and state-dependent foraging in scorpions: effect of moonlight on foraging
in the scorpion Buthus occitanus. Animal Behaviour, 52: 49-57
Tourtlotte GI. 1974. Studies on the biology and ecology of the northern scorpion, Paruroctyonus boreus
(Girard). Great Basin Naturalist, 34(3): 167-179
103

Arthropods, 2013, 2(3): 95-104
IAEES www.iaees.org
Toye SA. 1970. Some aspects of the biology of two common species of Nigerian scorpions. Journal of
Zoology London, 162: 1-9
Warburg MR, Ben-Horin A. 1979. Thermal effect on the diel activity rhythm of scorpions from mesic and
xeric habitats. Journal of Arid Environments, 2: 339-346
Warburg MR, Polis GA. 1990. Behavioral responses, rhythms and activity patterns. In: The Biology of
Scorpions (Polis GA, ed). 224-246, Stanford University Press, Stanford, USA
Warburg MR. 2013. A review on the diel rhythmic activities in physiological functions of scorpions.
Entomological Science, 16(3): 278-283
Wuttke W. 1966. Untersuchungen zur Aktivitätsperiodik bei Euscorpius carpathicus. Zeitschrift der
Vergleichende Physiologie, 53: 405-448
104

Arthropods, 2013, 2(3): 105-110
IAEES www.iaees.org
Article
Two records of Macrophthalmus Desmarest, 1823 (Decapoda:
Brachyura: Thoracotremata) from the NW of the Arabian Gulf
Amaal Gh. Yasser, Ibtisam M. AbdulSahib, Murtada D. Naser, Khalid Kh. S. Al-Khafaji, Haider Sh. Darweesh Department of Marine Biology, Marine Science Centre, University of Basrah, Basrah, Iraq
E-mail: [email protected]
Received 1 March 2013; Accepted 5 April 2013; Published online 1 September 2013
Abstract
Specimens of two crabs Macrophthalmus dentipes Lucas, 1836 and Macrophthalmus laevis A. Milne-Edwards,
1867 were collected from the intertidal zone of the lower reaches of Shatt Al-Arab at Fao region, Basrah, Iraq,
2012. A note on the morphological features of these two species and a photograph is provided to confirm the
identification of the crabs.
Keywords Macrophthalmus dentipes; Macrophthalmus laevis; Brachyura; Shatt Al-Arab; Arabian Gulf.
1 Introduction
The family Macrophthalmidae Dana, 1851, is represented by two subfamilies Ilyograpsinae Števcic, 2005 and
Macrophthalminae Dana, 1851 in the Arabian Gulf. The first one subfamily Ilyograpsinae Števcic, 2005 is
represented by one species Ilyograpsus rhizophorae, While the second subfamily Macrophthalminae Dana,
1851 is represented by eight species, have been recorded from the area, all belonging to Macrophthalmus
Desmarest, 1823, represented of these: M. sinuspersici Naderloo & Türkay, 2010, Macrophthalmus graeffei
A. Milne-Edwards, 1873, M. dentipes Lucas, 1836, M. depressus Rüppell, 1830, M. grandidieri A. Milne-
Edwards, 1867, M. laevis A. Milne-Edwards, 1867, M. serenei Takeda & Komai, 1991, and M. sulcatus H.
Milne-Edwards, 1852, are important elements of the intertidal soft bottom communities (Naderloo et al., 2011).
Barnes (1970) was so far recorded M. dentipes from the Arabian Gulf at Al-Faw referred to it as M.
pectinipes Guerin-Méneville 1838. Pretzmann (1971) and Jones (1986) recorded the species from Bandar-
Abbas at the Iranian coast and from Kuwait respectively, both following Barnes (1970) using the name M.
pectinipes. Holthuis (1995) revised M. dentipes as the valid name by showing that M. pectinipes is an objective
synonym of M. dentipes, and therefore must be replaced by the latter. Apel and Türkay (1999) and Apel (2001)
listed M. dentipes from the Arabian Gulf with reference to the records by Barnes (1971) and Jones (1986).
Arthropods ISSN 22244255 URL: http://www.iaees.org/publications/journals/arthropods/onlineversion.asp RSS: http://www.iaees.org/publications/journals/arthropods/rss.xml Email: [email protected] EditorinChief: WenJun Zhang Publisher: International Academy of Ecology and Environmental Sciences

Arthropods, 2013, 2(3): 105-110
IAEES www.iaees.org
Pretzmann (1971) described M. ressli as a new species from Bandar-Abbas, on the Iranian coast near the
Straits of Hormuz. Barnes (1976) synonymised M. ressli with M. laevis and was the first to provide a detailed
description for M. laevis.
The aim of the present study is to re-describe M. dentipes from fresh specimens collected from NW of the
Arabian Gulf at Faw region and to record Macrophthalmus laevis to add to the brachyua list of Iraq.
2 Materials and Methods
Specimens of M. dentipes and M. laevis were recently collected from the intertidal muddy flats of NW of the
Arabian Gulf, Fao region (Fig. 1) on July 2012. Specimens are preserved in 70% alcohol and deposited in the
marine science centre (MSC) (collection number: 33 and 34), and M. dentipes only deposited in the Zoological
Reference Collection of the Raffles Museum of Biodiversity Research, National University of Singapore
(ZRC).
The main abiotic parameters in the study area by the time of collection were as follows: salinity 35 ppt,
water temperature 27 oC, pH 8.4.
Fig. 1 Sampling site Faw=Fao region (white dot).
3 Results and Remarks
Macrophthalmus dentipes Lucas, 1836
Systematics
Order Decapoda
Macrophthalmidae Dana, 1851
Subfamily Macrophthalminae Dana, 1851
Macrophthalmus dentipes Lucas, 1836
(Fig. 2A, B, C and D)
Macrophthalmus dentipes Lucas 1836: 551. — Holthuis 1995: 401.
Macrophthalmus pectinipes — Guerin-Méneville 1838: 1, pl. 23. — Alcock 1900: 377. — Chhapgar 1957b:
106

Arthropods, 2013, 2(3): 105-110
IAEES www.iaees.org
512. — Barnes
1970: 237, fig. 10. — Pretzmann, 1971: 31; 1974: 442. — Tirmizi 1981: 109. — Titgen 1982: 253 (in list). —
Jones 1986:
159, pl. 45. — Tirmizi & Ghani 1996: 121, fig. 46.
Macrophthalmus (Venitus) dentipes — Apel & Türkay 1999: 135. — Apel 2001: 110. — Naderloo et al.,
2011: figs. 4a–e, 5a–f, 10a–b.
Type locality
Bombay (= Mumbay), India
Material examined (msc, 33)
Carapace measurements are length × breadth respectively.
Three (38.55×60.50), (36.55×60.00), (39.00×60.50) mm collected during July 2012 from the intertidal
zones of the mudflats of lower reaches of Shatt Al-Arab at Fao.
Diagnosis
Carapace (Fig. 2 A) moderately wider than long (CB/CL = 1.6), slightly convex; large granules scattered on
entire posterior surface except in narrow median, frontal regions. Lateral margin of carapace (Fig. 2 A) with 3
distinct teeth (including exorbital tooth); first nearly subquadrate, with posterior margin smooth, curved
forward; second triangular, with smooth margin, higher than first, greatest width of carapace between second
lateral teeth; third very small, directed forwards; posterolateral margin nearly straight, slightly converging
posteriorly, with small granules, beset with long setae; posterior margin with very small granules.
Eyestalks narrow (Fig. 2A), long, but not reaching to exorbital angle.
Male abdomen (Fig. 2B) with segments 3, 4 of same length, slightly shorter than segment 5; segments 5, 6
nearly of same length, with lateral margins nearly straight; lateral margins of segment 6 with small depression
at one third distal portion; telson very slightly longer than segment 6, lateral margins strongly converging
distally, apically rounded.
Palm long (Fig. 2C), outer surface smooth without longitudinal ridge; inner surface smooth, patch of dense
setae on upper portion. Fingers remarkably curved inward distally, movable finger with upper margin smooth,
long setae densely along inner surface of upper margin, continuous on upper, outer surface, cutting edge with
subproximal differentiated tooth, small teeth distally; immovable finger narrow, with relatively large teeth on
cutting edge, long setae along inner surface.
Male G1 (Fig. 2D,E,F) moderately stout, slightly curved outward medially; distal half relatively narrowing,
with apical chitinous process remarkably long, narrow, curved outward at about 45°; distal opening large,
distinct, subdistal on dorsal portion of apical process; long feather-shaped setae densely set along lateral
margin, long setae around apical process.
Habitat
Macrophthalmus dentipes is the largest ocypodid crab, inhabiting the mid and low intertidal zones in muddy
sand/sandy mud substrates at the lower reaches of Shatt Al-Arab at Fao, it may be occur coexisting with the
grapsoid crab Metaplax indica. Macrophthalmus dentipes digs large burrows with an opening of
approximately 10 cm in diameter.
Distribution
Northern Indian Ocean: northern and eastern Arabian Gulf, Oman (Gulf of Masirah), Pakistan, west coast of
India, Iraq.
Macrophthalmus laevis A. Milne-Edwards, 1867
Systematics
107

Arthropods, 2013, 2(3): 105-110
IAEES www.iaees.org
Order Decapoda
Macrophthalmidae Dana, 1851
Subfamily Macrophthalminae Dana, 1851
Macrophthalmus laevis A. Milne-Edwards, 1867
(Fig3. A,B,C and D)
Macrophthalmus laevis A. Milne-Edwards 1867: 287. — Barnes 1976: 143, fig. 6a–c. — Titgen 1982: 150.
Macrophthalmus (Macrophthalmus) ressli Pretzmann 1971: 382, pl. 9 figs. 23.
Macrophthalmus resseli [sic!] — Pretzmann,1974: 441.
Macrophthalmus (Macrophthalmus) laevis — Barnes 1977: 277 (in key), 280 (in list); 2010: 35 (in key), 40.
— Tirmizi & Ghani 1988: 253, figs. 1–11. — Tirmizi & Ghani 1996: 109, fig. 41. — Apel & Türkay 1999:
135. — Apel 2001: 109— Naderloo et al., 2011: figs. 13a–f, 14a–e, 10e–f.
Type locality
Indian Seas.
Material examined (msc,34)
Carapace measurements are length × breadth respectively.
Two males (12.50×24.50) and (12×23.50) mm collected during July 2012 from the intertidal zones of the
mudflats of lower reaches of Shatt Al-Arab at Fao.
Carapace
Macrophthalmus laevis is a medium-size species (Fig. 3A) wider than long; posterior surface is convex, small
granules distributed on posterior surface, extensive patch of setae near posterolateral margin, long setae on
lateral margin. Regions well defined; defining gastric, epibranchial regions are remarkably deep. Lateral
margin with three teeth including exorbital angle.
Chelipeds nearly equal; merus with upper surface smooth, inner, upper margins sparsely beset with long
setae, row of long setae near inner margin.Carpus smooth with large spine- shaped tooth medially on upper
inner margin, small one behind it, two spine-shaped teeth on inner proximal margin. Movable finger long ( Fig.
3C) curved inward distally; upper margin smooth; cutting edge with differentiated subproximal tooth, large,
subquadrate, low, small denticles distal to large one along cutting edge. Immovable finger short, with median
tooth, large, extending proximally, small denticles on cutting edge, even on large tooth.
Walking legs narrow, long, anterior margin of segments bearing long setae. Merus with small subdistal
tooth on anterior margin, that of second, third legs large, last leg usually lacking this subdistal tooth ( Fig.3.A).
Male abdomen (Fig. 3B) triangular; segments 3, 4 of same length, segment 5 slightly longer; segment 6
longest with lateral margins swollen proximally, gently converging distally; telson slightly shorter than
segment 6, with margins clearly converging distally, rounded distally.
Male G1 (Fig. 3D) curved outward medially; apical chitinous process short, nearly subdistal, directed
laterally at 45°; distal opening prominent, located apically; long setae around apical part, long plumose setae
sparsely set along lateral, ventral surfaces.
Habitat
M. laevis mainly in the upper mid littoral zone on muddy silty substrata.
Distribution
North-western Indian Ocean: Persian Gulf, Gulf of Oman, Pakistan, Iraq.
108

Arthropods, 2013, 2(3): 105-110
IAEES www.iaees.org
Fig. 2 Macrophthalmus dentipes Lucas,1836, male (39.00×60.50): A, posterior view of whole crab, male; B, male ventral view. C, cheliped of male, outer surface; D- F, first gonopod, Photos taken by Murtada.D.Naser, Marine Science Centre.
Fig. 3 Macrophthalmus laevis A. Milne-Edwards 1867, male (12.5×24.5): A, posterior view of whole crab, male; B, male ventral view. C, cheliped of male, outer surface; D,first gonopod, Photos taken by Murtada.D.Naser, Marine Science Centre.
109

Arthropods, 2013, 2(3): 105-110
IAEES www.iaees.org
Acknowledgements
M.D.N. thanks Dr Peter K.L. Ng (Raffles Museum of Biodiversity Research, Department of Biological
Sciences, National University of Singapore) for confirming the identity of Macrophthalmus dentipes.
References
Alcock A. 1900. Material for a carcinological fauna of India. No. 6: The Brachyura Catometopa, or
Grapsoidea. Journal of the Asiatic Society of Bengal, 69 (2): 279-456
Apel M. 2001. Taxonomie und Zoogeographie der Brachyura, Paguridea und Porcellanidae (Crustacea:
Decapoda) des Persisch Arabischen Golfes unpublished PhD. Thesis. Johann Wolfgang Goethe-Universität,
Frankfurt am Main, Germany
Apel M, Türkay M. 1999. Taxonomic composition, distribution and zoogeographic relationships of the grapsid
and ocypodid crab fauna of intertidal soft bottoms in the Arabian Gulf. Estuarine, Coastal and Shelf
Science, 49(Suppl. A): 131-142
Al-Zaidan ASY, Kennedy H, Jones DA, et al. 2004. Role of microbial mats in Sulaibikhat Bay (Kuwait)
mudflat food webs: evidence from δ13C analysis. Marine Ecology Progress Series, 38: 27-36
Barnes RSK 1970. The species of Macrophthalmus in the collections of the British Museum (Natural History).
Bulletin of the British Museum of Natural History, 20: 203-251
Barnes RSK 1976. Contributions towards a revision of Macrophthalmus, VIII: A re-examination of the M.
telescopicus Owen complex; the status of M. laevis H. Milne-Edwards; and the affinities of M. holthuisi
Sérene. Zoologische Mededelingen, 50(10): 133-151
Barnes RSK 1977. Concluding contribution towards revision of, and a key to, the genus Macrophthalmus
(Crustacea Brachyura). Journal of Zoology London, 182: 267-280
Holthuis LB 1995. The identities of Macrophthalmus rouxii Lucas, 1836, and M. dentipes Lucas, 1836, and
the substitution of the latter name for M. pectinipes Guérin, 1838 (Decapoda, Brachyura, Ocypodidae).
Crustaceana, 68(3): 401-403
Jones DA 1986. A Field Guide to the Sea Shores of Kuwait and the Arabian Gulf. University of Kuwait,
Blandford Press, Poole, Kuwait
Lucas H. 1836. Macrophthalme, Macrophthalmus. (Crust.) In: Guérin-Meneville, F.E.: Dictionaire pittoresque
d’Histoire naturelle Vol. 4, pp. 551, pl. 315, fig. 5.
Naderloo R, Türkay M, Apel M. 2011. Brachyuran crabs of the family Macrophthalmidae Dana, 1851
(Decapoda: Brachyura: Macrophthalmidae) of the Persian Gulf. Zootaxa, 2911: 1-42
Tirmizi NM, Ghani N. 1988. The rediscovery of Macrophthalmus (Macrophthalmus) laevis A. Milne-
Edwardds, 1867, in the Arabian Sea (Decapoda Brachyura). Crustaceana, 55(3): 253-256
Tirmizi NM, Ghani N. 1996. Marine Fauna of Pakistan: 5: Crustacea: Brachyura, Brachyrhyncha, Part 1:
Xanthidae Goneplacidae, Pinnotheridae, Ocypodidae, Grapsidae. Center of Excellence, University of
Karachi, Pakistan
Titgen RH. 1982. The Systematics and Ecology of the Decapods of Dubai, and their Zoogeographic
Relationships to the Arabian Gulf and the Western Indian Ocean. Unpublished D. Phil. Thesis, Texas A &
M University, USA
110

Arthropods, 2013, 2(3): 111-125
IAEES www.iaees.org
Article
Reproductive characteristics of a brachyuran crab, Grapsus
tenuicrustatus (Herbst, 1783) (Decapoda: Grapsidae) found in Talim
Bay, Batangas, Philippines
Michael A. Clores1, Gliceria B. Ramos2 1Ateneo de Naga University, Naga City, Philippines 2Biology Department, De La Salle University, Taft Avenue, Manila, Philippines
E-mail: [email protected]
Received 7 March 2013; Accepted 10 April 2013; Published online 1 September 2013
Abstract
The study determined some reproductive characteristics of a brachyuran crab, Grapsus tenuicrustatus (Herbst,
1783), one among the most widespread and diverse groups of invertebrates. Results revealed that there were
more males (52.94%) than females (47.06%) collected at the study sites with a sex ratio of 1:1.13. Thirty
percent (30%) of the samples were ovigerous females. Ovigerous females have the largest caraface length, CL,
(31.25 ± 1.43) compared with the males (22.14 ± 0.726) and non-ovigerous females (26.63 ± 1.12). Based on
one-way ANOVA, the differences were significant. Difference between non-ovigerous and ovigerous females
was also found significant based on t-test for independent samples. There was a non-conspicuous bi-modal size
distribution for all the crabs, with non-normal distributions for all crabs and for males, but not when all
females or ovigerous females only were grouped together. The size-frequency distributions of males and
females are significantly different from each other similar with that observed between the size-frequency of
ovigerous and non-ovigerous females. There were more ovigerous crabs belonging to the first year age class
(CL = 16 - 33) (53.13 %) than those that belong to the older class (CL = 34 - 43) (46.88 %). Fecundity ranged
from 4400 (CL = 16 mm) to 26400 (CL = 43 mm) eggs. Egg volume ranged from 0.40 ml to 2.40 ml, egg
diameter from 1.1 µm to 5.0 µm with an average diameter of 3.170 µm and egg count from 4400 to 26400
with a mean of 12684 eggs. Egg number was positively correlated with female size.
Keywords Grapsus tenuicrustatus; brachyuran crab; fecundity; ovigerous; reproductive characteristics.
1 Introduction
In the Philippines, brachyurans (true crabs) are among the most widespread and diverse groups of invertebrates.
One of the interesting brachyurans are the Sally-lightfoot crab or natal sally-lightfoot or shore crab, Grapsus
tenuicrustatus, locally called “Katang”, because these are known to be harvested and processed by
Arthropods ISSN 22244255 URL: http://www.iaees.org/publications/journals/arthropods/onlineversion.asp RSS: http://www.iaees.org/publications/journals/arthropods/rss.xml Email: [email protected] EditorinChief: WenJun Zhang Publisher: International Academy of Ecology and Environmental Sciences

Arthropods, 2013, 2(3): 111-125
IAEES www.iaees.org
fermentation as a delicacy and crab roe fat.
A random search of previous studies on brachyurans in the Philippines showed a paucity of research on
their reproductive ecology and the environmental and biological constraints that influence them as a population.
Indeed, while crab population structure and reproduction of subtropical species have become major research
agenda (Spivak et al. 1991; Mouton and Felder 1995), such studies on the tropical ones are still needed (Litulo,
2005).
Understanding the breeding potential of many well-dispersed marine invertebrates, like crabs, entails
determining intraspecific variation of reproductive characteristics (Dugan, 1991). Incomplete knowledge of the
life history of the organisms that comprise communities is one of the principal problems in understanding how
marine ecosystems function. For crab populations, understanding the environment and biological constraints
that are shaping them (Oshiro, 1999; Litulo, 2005) remains one important aspect of marine ecology.
In most marine invertebrates, the newly laid eggs contain all the energy and reserves for embryonic
development (Holland, 1978; Jaeckle, 1995). In species with complex life cycles, larval survival and growth
may depend on the energy reserves that remain after hatching (Paschke, 1998; George, 1999). Consequently,
these depend on the initial egg reserves and their utilization during embryogenesis. The embryonic
development occurs in a variety of modes, e.g. free developing, encapsulated, incubated (Sastry, 1983; Levin
and Bridges, 1995), and under a particular combination of environmental factors that may affect the embryonic
energy budget and thus, larval reserves (Gimé́nez and Anger, 2001). For decapods, the marine benthos is the
typical environment. The larvae hatch from the eggs attached to the female’s abdominal appendages (pleopods)
and develop in the marine plankton for several weeks (Diesel, 1989).
The present study determined some reproductive characteristics of G. tenuicrustatus (Herbst, 1783).
Specifically, it described the following: (i) total number and sex ratios of collected crabs; (ii) the size and age
distribution of ovigerous crabs; (iii) the female size at maturity and age classes based on carapace length (CL,
mm) and their size-frequency distribution, and (iv) an estimate of population fecundity (e.g., volume, number,
and diameter of eggs; relative number of females carrying various egg stages; estimate number of age in
different size groups of ovigerous crabs, and the relationship between egg number and female size). Broadly,
results of this study could provide insights on the relative plasticity or conservation of different reproductive
characteristics and the effects of intraspecific variation in those characteristics on the reproductive potential of
a well-dispersed marine invertebrate (Dugan, 1991).
2 Materials and Methods
2.1 Study site
Samples of G. tenuicrustatus were collected in rocky areas near seagrass beds at Talim Point (130 57’ 55.43’’,
1200 36’ 20.36’’ E), a portion of Talim Bay, Barangay Ligtasin, Lian, Batangas, Philippines. Talin Bay is
located between latitude 130 58.8’ North and longitude 1200 38.0’ East of DLSU Br. Alfred Shields FSC
Marine Biology Station, approximately 200 km. south of Manila fronting South China Sea.
Laboratory activities were conducted at the Biology Laboratories of Ateneo de Naga University, Naga City,
Philippines.
2.2 Sampling method
G. tenuicrustatus were collected opportunistically with a shovel and by hand from aggregations of crabs in the
wash and surf zones of the bay following previous methods (Wenner et al., 1987; Dugan, 1990; Dugan et al.,
1991). The crabs were retained and separated from the sand by washing through mesh bags. A total number of
153 crabs were collected.
112

Arthropods, 2013, 2(3): 111-125
IAEES www.iaees.org
2.3 Female size and age
Size-frequency distributions were determined by individual measurement of carapace lengths (CL), to the
nearest millimeter, using a vernier caliper. The sex and reproductive condition of all crabs were recorded (i.e.,
ovigerous or non-ovigerous). The size-frequency distributions of female crabs were examined for modal
breaks. Since there were two non-overlapping size modes, the mode of larger ovigerous crabs were assigned to
the older overwintered age class (Years 2 and 3) and the mode of smaller ovigerous crabs were assigned to the
first year class (young of one year) following Dugan et al. (1991).
The size at maturity, and the minimum (5th percentile) and maximum (95th percentile) sizes of ovigerous
female crabs were determined from size-frequency distributions for each sample. The 5th and 95th percentile
sizes of ovigerous crabs were determined from the cumulative number of ovigerous crabs and were used as
estimates of the minimum and maximum sizes of ovigerous female crabs. For the whole sample, the
proportions of ovigerous crabs were calculated for three categories, as follows: (1) the proportion of ovigerous
crabs above the size of the smallest ovigerous crab; (2) the proportion of ovigerous crabs assigned to the older
age class; and (3) the proportion of ovigerous crabs assigned to the first year class.
The developmental stage of each clutch was determined using the method of Eickstaedt (1969), which
divides egg development into ten stages. Stages 1-4 were of most interest. The amount of cleavage and the
proportion of the egg that is free of orange yolk distinguish these stages. Stage l eggs are uncleared or in a state
of cleavage; in Stage 2 cleavage is complete; in Stage 3 up to I/4 of the egg is free of yolk, and in Stage 4 up to
1/3 of the egg is free of yolk. Stage 5 eggs have visible embryonic eye pigment and were not used in fecundity
estimates. Forty (40) eggs were randomly selected and their diameter was measured with an ocular micrometer.
Finally, descriptive statistics (e.g., mean, mode, median, standard error of the mean, minimum, maximum,
range, standard error of the sizes) of egg diameter were reported.
2.4 Population fecundity
The present study adopted Dugan’s (1991) definition of fecundity which states that fecundity refers to the
number of eggs present in a single clutch of an individual crab at the time of analysis (e.g., clutch size). The
ovigerous crabs were preserved in a mixture of ethanol, isopropyl alcohol, and acetone. Volumetric
determinations of fecundity were made on female crabs with newly extruded eggs (no eyespots evident upon
microscopic examination).
Litulo’s (2005) method of estimating fecundity was adapted in the study. Twenty (20) ovigerous females
with eggs were randomly selected for egg. Pleopods were removed from the females, placed in petri dishes
filled with water, and had their eggs detached by gradually adding a solution of sodium hypochlorite. Bare
pleopods were then discarded by being gently stirred in a beaker filled with 50 ml of seawater. With a pipette,
five sub-samples of 1 ml were taken from the water with eggs. The eggs in each sub-sample were counted
under a dissecting microscope. The average value obtained was extrapolated for the whole suspension in order
to estimate the number of eggs (Bezerraa and Matthews-Cascona, 2007).
An estimate of population fecundity was made for the whole sample. Population fecundity is defined in
this study as the number of eggs carried by a representative female crabs at or above the size of the smallest
ovigerous crab at the time of sampling. Pearson correlation was used to test the relationship between egg
volume and number of eggs with carapace length (CL, mm). To further analyze fecundity, data were analyzed
using the power function (Y = aX + b) of egg number (EN) vs. CL.
2.5 Statistical analysis
Data were analyzed using SPSS for Windows 11.0 (Copyright © SPSS Inc., 1989-2001). Descriptive statistics
(e.g., mean, mode, median, standard error of the mean, minimum, maximum, range, standard error of the sizes)
were calculated for the ovigerous, non-ovigerous and for male crabs for the purpose of comparison. T-test for
113

Arthropods, 2013, 2(3): 111-125
IAEES www.iaees.org
independent samples and one-way Analysis of Variance (ANOVA) were used to test significant differences
among groups (males vs. ovigerous females vs. non-ovigerous females; ovigerous vs. ovigerous) in terms of
mean carapace length (CL, mm). No estimates of density or abundance were made. To test the null hypothesis
that each group of sample comes from a normal distribution, one-sample Kolmogorov-Smirnov (KS)
procedure was used. This goodness-of-fit test assesses whether the observed cumulative distribution function
for a variable with a specified theoretical distribution, which in this case, normal distribution. Two-sample KS
test was also used to test the null hypothesis that male and female samples, as well as ovigerous and ovigerous
samples have the same distribution. Lastly, Pearson correlation was used to test the relationship between egg
volume and number of eggs with carapace length (CL, mm).
3 Results
3.1 Female size and age
There were more male Grapsus tenuicrustatus (52.94%, n = 81) than females (47.06%, n = 72) collected with
a sex ratio of 1:1.13. Thirty percent (30%, n = 32) of the samples were ovigerous females (Table 1).
Table 2 and Fig. 1 present the comparison of the carapace length (CL, mm) of the males and non-ovigerous
and ovigerous female G. tenuicrustatus. Ovigerous female crabs have the largest CL (mean ± SE: 31.25 ± 1.43)
compared with the males (mean ± SE: 22.14 ± 0.726) and non-ovigerous females (mean ± SE: 26.63 ± 1.12).
The differences were significant (F = 20.383, df = 2, p < 0.01). Difference between non-ovigerous and
ovigerous females was also found significant (t = 2.582, df = 70, p < 0.05). The CL (mm) of all samples
showed a mean ± SE of 25.22 ± 0.636. A sample with the smallest CL (12 mm) was found among male crabs
while the largest was from the ovigerous females (43 mm).
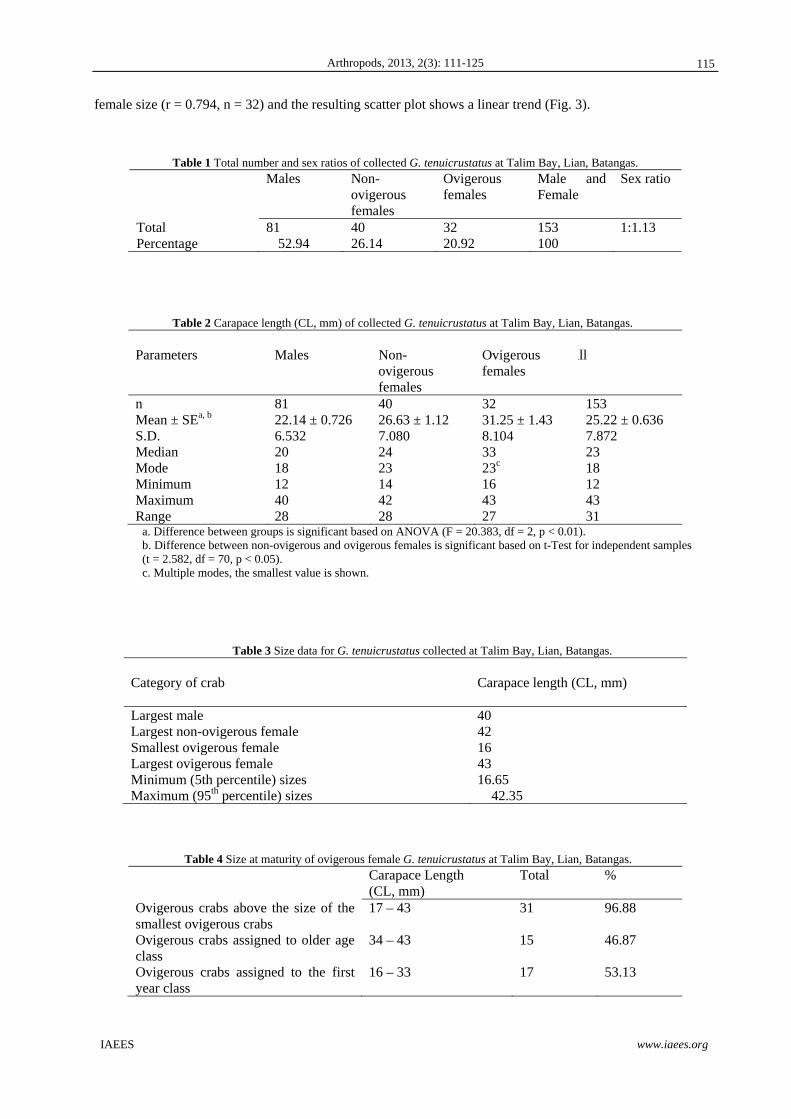
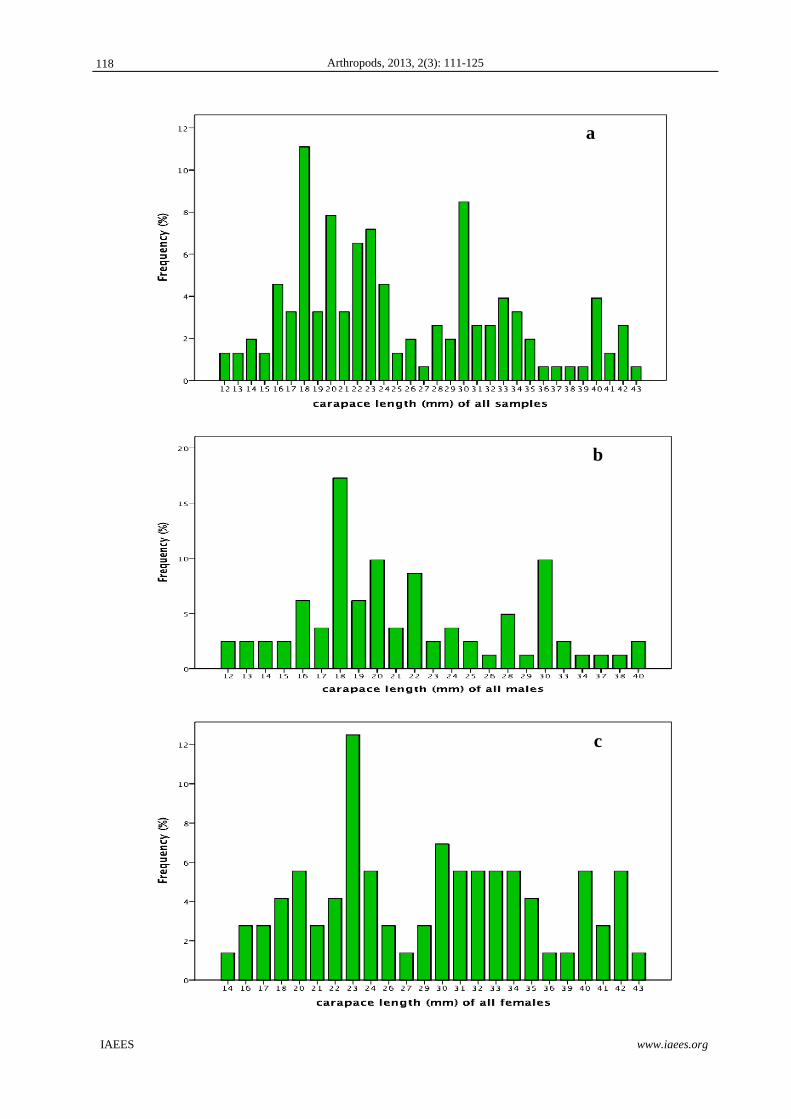
Fig. 2 (a-d) shows the size frequency distributions of all the crabs, males crabs only, female crabs only and
ovigerous females only, respectively. There was a non-conspicuous bi-modal size distribution for all the crabs,
with non-normal distributions for all crabs (KS = 1.735, p < 0.05) and for males (KS = 1.464, p < 0.05), but
not when all females or ovigerous females only were grouped together. When the size-frequency distribution
of males was compared with females, the distributions are significantly different from each other (KS = 2.582,
p < 0.0001). The same was observed between the size-frequency of ovigerous and non-ovigerous female crabs
(KS = 1502, p < 0.05).
Based on the size at maturity, and the minimum (5th percentile) and maximum (95th percentile) sizes of
ovigerous female crabs determined from size-frequency distributions for the whole sample, the largest among
the samples is an ovigerous female crab (CL = 43 mm) while the smallest is also from the same group
(CL=16). From the cumulative number of ovigerous crabs, the minimum and maximum sizes of ovigerous
female crabs are estimated as CL = 16.65 mm and CL= 42.35 mm, respectively (Table 3).
There are more ovigerous crabs belonging to the first year age class (CL = 16 - 33) (53.13 %) than those
that belong to the older class (CL = 34 – 43) (46.88 %). About 97% of the female ovigerous crabs are above
the size of the smallest crabs (CL = 17 – 43). Moreover, overlapping size modes were shown among the
ovigerous female crabs (23 mm, 34 mm, 35 mm (Table 4 and 5).
3.2 Population fecundity
The fecundity of G. tenuicrusttus ranged from 4400 (CL = 16 mm) to 26,400 (CL = 43 mm) eggs. Egg volume
ranged from 0.40 ml to 2.40 ml. The egg diameter ranged from 1.1 µm to 5.0 µm with an average diameter of
3.170 µm (Table 6; Fig. 4). There are more crabs belonging to CL class of 11-20 mm and 21-30 mm that carry
eggs at different stages (Table 7).
As shown in Table 8, egg count ranged from 4400 to 26400 with a mean of 12684 eggs. Forty-four percent
(44%) of the ovigerous crabs have eggs ranging from 4400 to 8800. Egg number was positively correlated with
114
Arthropods, 2013, 2(3): 111-125
IAEES www.iaees.org
female size (r = 0.794, n = 32) and the resulting scatter plot shows a linear trend (Fig. 3).
Table 1 Total number and sex ratios of collected G. tenuicrustatus at Talim Bay, Lian, Batangas.
Males Non-ovigerous females
Ovigerous females
Male and Female
Sex ratio
Total 81 40 32 153 1:1.13 Percentage 52.94 26.14 20.92 100
Table 2 Carapace length (CL, mm) of collected G. tenuicrustatus at Talim Bay, Lian, Batangas. Parameters
Males
Non- ovigerous females
Ovigerous females
All
n Mean ± SEa, b
S.D. Median Mode Minimum Maximum Range
81 22.14 ± 0.726 6.532 20 18 12 40 28
40 26.63 ± 1.12 7.080 24 23 14 42 28
32 31.25 ± 1.43 8.104 33 23c
16 43 27
153 25.22 ± 0.636 7.872 23 18 12 43 31
a. Difference between groups is significant based on ANOVA (F = 20.383, df = 2, p < 0.01). b. Difference between non-ovigerous and ovigerous females is significant based on t-Test for independent samples (t = 2.582, df = 70, p < 0.05). c. Multiple modes, the smallest value is shown.
Table 3 Size data for G. tenuicrustatus collected at Talim Bay, Lian, Batangas. Category of crab
Carapace length (CL, mm)
Largest male 40 Largest non-ovigerous female 42 Smallest ovigerous female 16 Largest ovigerous female 43 Minimum (5th percentile) sizes 16.65 Maximum (95th percentile) sizes 42.35
Table 4 Size at maturity of ovigerous female G. tenuicrustatus at Talim Bay, Lian, Batangas. Carapace Length
(CL, mm) Total %
Ovigerous crabs above the size of the smallest ovigerous crabs
17 – 43
31 96.88
Ovigerous crabs assigned to older age class
34 – 43
15 46.87
Ovigerous crabs assigned to the first year class
16 – 33
17 53.13
115

Arthropods, 2013, 2(3): 111-125
IAEES www.iaees.org
Table 5 Age classes of female G. tenuicrustatus collected at Talim Bay, Lian, Batangas based on modal breaks of Carapace Length (CL, mm) (n=32).
Age class Carapace Length (CL, mm)
Mode Total %
First year 16 – 33 23 17 53.13 Year 2 and 3 34 – 43 34, 35 15 46.87
Table 6 Egg volume (ml), number of eggs, and diameter of eggs sampled from collected G. tenuicrustatus at Talim Bay, Lian, Batangas.
Parameters
Carapace Length (CL, mm)
Egg volume (ml)
No. of Eggs
Egg Diameter (µm)
n Mean ± SE S.D. Median Mode Minimum Maximum Range
32 31.25±1.43a,b
8.104 33 23b
16 43 27
32 1.15±0.10a
0.592 1.05 0.60 0.40 2.40 2.00
32 12684.38±1153.04b
6522.572 11550.00 6600 4400 26400 22000
40 3.170±0.18 1.146 3.55 3.5 1.1 5.0 3.9
a, b. Positive correlation based on 2-tailed Pearson Correlation (r2 = 0.794, n =32).
Table 7 Numbers of ovigerous females of G. tenuicrustatus in different size groups (expressed as a percentage of total numbers of females collected) and relative number of females carrying various egg stages. Carapace width class (mm)
No. of females
Ovigerous females (%)
No. of ovigerous females carrying egg stage 1 2 3 4
11 - 20 5 60.0 5 4 4 2 21 - 30 7 71.4 6 5 5 0 31 - 40 8 87.5 5 0 1 1 41 - 50 3 100.0 0 1 0 0
Table 8 Estimate number of age in different size groups of ovigerous G. tenuicrustatus collected at Talim Bay, Lian, Batangas (n=32).
Estimate No. of Eggs Number of ovigerous females
%
4,400 – 8,800 14 43.75 8,801 – 13,200 6 18.75 13,201 – 17,600 4 12.5 17,601 – 22,000 4 12.5 22,001 – 26,400 4 12.5 Total 32 100
116

Arthropods, 2013, 2(3): 111-125
IAEES www.iaees.org
Fig. 1 Sizes of (a) male and female G. tenuicrustatus and (b) female ovigerous and non-ovigerous G. tenuicrustatus collected at Talim Bay, Lian, Batangas.
a
b
117
Arthropods, 2013, 2(3): 111-125
IAEES www.iaees.org
a
b
c
118

Arthropods, 2013, 2(3): 111-125
IAEES www.iaees.org
Fig. 2 Size-Frequency distribution of (a) all G. tenuicrustatus (n=153), (b) all the male G. tenuicrustatus (n=81), (c) all the female G. tenuicrustatus (n=72), and (d) all the ovigerous female G. tenuicrustatus (n=32) collected at Talim Bay, Lian, Batangas.
Fig. 3 Scatter plot for the relationship between egg number (EN) and female size (CL, mm) of G. tenuicrustatus collected at Talim Bay, Lian, Batangas.
d
119

Arthropods, 2013, 2(3): 111-125
IAEES www.iaees.org
Fig. 4 Eggs of G. tenuicrustatus collected at Talim Bay, Lian, Batangas. (a) egg still undivided, fully filled with yolk; (b to e), the free region of yolk is just visible; free area of yolk is conspicuously larger than earlier periods and ocular lobes are already visible; (f & g) overview of egg microscope.
a b
c d
g
e
f
120

Arthropods, 2013, 2(3): 111-125
IAEES www.iaees.org
4 Discussion
There are many reasons for the observed differences in total number collected and sex ratio among the G.
tenuicrustatus in the study site. First, in crustacean populations, sexual differences in distribution and mortality
may be responsible for unbalanced sex ratios (Johnson, 2003). Based on previous reports, differences between
male and female are not only exhibited by their spatial distributions and mortality rates but also on the effect of
predation on crab sex ratio (Montague, 1980; Wolf et al., 1975; Spivak et al., 1991).
Second, it was observed that the physiologic and behaviorally homeostatic crab populations living in
constant environments present a 1:1 sex ratio, or slightly male-biased. On the other hand, populations that
inhabit variable environments present deviations toward the females, in order to maximize the evolutionary
potential due to unequal selection between males and females (Geisel, 1972). Hence, it can be inferred that the
study site provided a rather male-biased environment wherein a more stable and constant conditions are
present.
In 1930, Fisher predicted that in random mating populations the evolutionary stable sex ratio would be 1:1.
Several studies supported this hypothesis. For instance, Bezerra and Matthews-Cascon (2007) found out that
the overall sex ratio of Uca thayeri population did not differ significantly from the expected 1:1 ratio and
therefore showed that this population is physiologic and behaviorally adapted to the habitat, besides also being
evolutionary stable.
Lastly, Costa (2000) observed that the low number of ovigerous females might also be due to the fact that
ovigerous females hide inside deep burrows in order to incubate their eggs. Thus, as observed in the present
study wherein the sex ratio did not differ significantly from the expected 1:1 ratio, indeed in a majority of
species is close to unity, despite some variations between populations of a species, and from year to year in the
same population (Nikolsky, 1963; Ofori-Danson, 1990).
Mantelatto and Fransozo (1996) explained that size at the onset of sexual maturity is a crucial variable to
be taken into account while investigating the reproductive ecology of a given organism. For crustaceans there
are not always outer characteristics such as color and size that clearly indicates when an individual reaches
sexual maturity In brachyuran crabs, this is easier in females due to unambiguous signals of breeding
competency, such as the presence of eggs attached to pleopods (Flores and Paula, 2002).
Among ocypodid crabs, sexual dimorphism is evidenced by males reaching larger sizes than females
(Lopez Greco et al., 2000). Females may have reduced somatic growth compared to males because they
concentrate their energy budget for gonad development. Moreover, males may reach larger sizes for successful
competition for copulation with more than one female, since larger male ocypodid crabs may have greater
chances of obtaining females for copulation and win more intra-specific fights (Christy and Salmon, 1984;
Christy, 1987). This may not be applicable in the case of G. tenuicrustatus, which was shown to have larger
females than males. Aside from reproductive pressure, other environmental and physiological factors might
explain why females are larger than males among G. tenuicrustatus.
Estimates based on the smallest egg-bearing female are dependent on the sample size and do not indicate
the average size at which females in a given population reach maturity (Lopez Greco and Rodriguez, 2004;
Ituarte et al., 2004). Hence, comparative studies using a morphological (macroscopic and histological) and
morphological analysis could be used for more precise estimates in determining the size at which males and
females reach maturity (Litulo, 2005).
The results also support the notion that size at sexual maturity and fecundity are the key parameters that
should reflect the lifetime investment in reproduction (Ramirez Llodra, 2002; Lopez Greco and Rodriguez,
2004). Certainly, fecundity and the size at the onset of sexual maturity of a species influence the periodicity
and duration of breeding season. Other important factors include temperature, salinity, food availability,
121

Arthropods, 2013, 2(3): 111-125
IAEES www.iaees.org
rainfall, photoperiod and lunar cycles. (Colpo and Negreiros-Fransozo, 2003; Costa and Negreiros-Fransozo,
2003; Litulo, 2004), but such variables were not explored in the present study and thereby merit further
investigation.
Fecundity is the number of eggs per female and a determinant of the reproductive potential of a species and
the stock size of its population (Mantelatto and Fransozo, 1997). It is an important parameter measured in
crustaceans for the estimation of the reproductive potential and future stock size of a given species or
population (Hattori and Pinheiro, 2001). Further, fecundity is directly related to life-history traits such as egg
size, age at maturity, life span and reproductive effort (Ramirez Llodra, 2002). The variation in fecundity is
very common in crab and has been reported by many workers like Erdman and Blake (1988); Melville Smith
(1987); Pauley et al. (1986); Hill et al. (1989); Gray (1969), and Ong (1966).
Hines (1982) explained that fecundity of crabs varies from species to species and also varies within the
same species due to different factors such as age, size, nourishment, ecological conditions of the water body
etc. Variation in fecundity was primarily a reflection of variation in the size of the crab at maturity.
The relationship between fecundity and size at sexual maturity depends on the life-history strategies of a
species (Ramirez Llodra, 2002). Earlier maturing species usually have a shorter generation because of the
shorter generation time needed to reach first reproduction and, the cost may be observed in a reduction in
future fecundity. In contrast, species with delayed maturity live longer, allowing them to grow larger and
therefore have a higher fecundity (Ramirez Llodra, 2002). Moreover, the longer lifespan may also give the
possibility of undergoing a higher number of lifetime fecundity and spawnings, which are characteristic in
tropical species (Emmerson, 1994).
In genus of Brachyurans, the Uca species, Thurman (1985) reported that the greatest egg amount (25012)
was registered for a female with 26.5mm of CW but concluded that the fecundity of the species in temperate
and tropical areas vary greatly, where the size and the amount of eggs are in close association with the
environmental conditions. More specifically in other studies, it was recorded that for a subtropical Uca thayeri
population that females from 23 to 26 of CW carried more than 45000 eggs, showing that in U. thayeri
fecundity is correlated with environmental conditions (Costa, 2000).
With the range of 3.9 mm in the diameter of eggs of G. tenuicrustatus as revealed by the findings of the
current study (see Table 6), the common observation that brachyuran show a great diversity of embryonic
development, especially owing to a significant variation in egg size (Hines, 1982) is supported. For blue crabs,
C. sapidus, a mean fecundity of 3.2 million eggs was revealed (Guillory et al., 1996). Shields et al. (1990)
mentioned that such variations in fecundity among brachyuran crabs may be caused by many factors including
climatic regimes, habitat and biological constraints.
Strong size-fecundity relationships are found in brachyuran families (Hines, 1982; Hartnoll, 1985). The
results of the present study are not an exception as shown by the results.
The results of the present study suggest that of an increase in number of eggs as the crabs grow larger. A
positive allometry between egg number and female size implies of an increase in fecundity of an increase of
female size. The relationship between female size and fecundity is a major characteristic of reproduction in
many crustaceans, and is related to morphological and physiological constraints in energy allocation and gonad
maturation (Ramirez Llodra, 2002). Litulo (2005) reported similar results for other brachyurans summarized
by Hines (1982) and studies done by Erdman and Blake (1988) on female golden crab, Geryon fenneri; Kumar
et al. (2000) on blue swimming crab, P. pelagicus, and Kyomo on sesarmid crab, Sesarma intermedia.
Carapace shape affects the volume reserved for gonadal development and spawn size (Hines, 1982;
Mantelatto and Fransozo, 1997; Koga, 1982). The allometric relationships between fecundity and crab size
variables is explained by the fact that egg mass is limited by the space available for the accumulation of
122

Arthropods, 2013, 2(3): 111-125
IAEES www.iaees.org
reserves as well as the gonadal development inside the cephalothorax of the crabs (Litulo, 2004). Similarly, in
the present study, it has been found that the number of eggs increased linearly with the increase of carapace
length.
5 Conclusion
The significant reproductive characteristics of G. tenuicrustatus in Talim Bay, Batangas include: (a) a slightly
male-biased ratio of 1:1.13; ovigerous female crabs having the largest CL compared with the males and non-
ovigerous females; more ovigerous crabs belonging to the first year age class than the older classes; fecundity
ranged from 4400 (CL = 16 mm) to 26,400 (CL = 43 mm) eggs; and the number of eggs increased with
increase in female size.
Acknowledgment
This work was supported by the scholarship awarded to the main author by the Advanced Science and
Technology Human Resource Development Program (ASTHRDP) of the Department of Science and
Technology (DOST) of the Philippines and Faculty and Staff Development Program of Ateneo de Naga
University, Philippines.
References
Bezerra LE, Matthews-Cascon H. 2007. Copulation and reproductive biology of the fiddler crab Uca thayeri
Rathbun, 1900 (Crustacea: Ocypodidae) in a tropical mangrove from Northeast Brazil. Acta Oecologica 31:
251-258
Christy JH, Salmon M. 1984. Ecology and evolution of mating system of fiddler crabs (genus Uca). Biological
Reviews, 59: 483-509
Christy JH. 1987. Female choice and breeding behavior of the fiddler crab Uca beebei. Journal of Crustacean
Biology, 7: 624-635
Colpo KD, Negreiros-Fransozo ML. 2003. Reproductive output of Uca vocator (Herbst, 1804) (Brachyura,
Ocypodidae) from three subtropical mangroves in Brazil. Crustaceana, 76: 1-11
Costa TM. 2000. Ecologia de caranguejos semiterrestres do geˆ nero Uca (Crustacea, Decapoda, Ocypodidae)
de uma a ́ rea de manguezal, em Ubatuba (SP). Ph.D. thesis, Universidade Estadual Paulista, Brazil
Costa TM, Negreiros-Fransozo ML. 2002. Population biology of Uca thayeri Rathbun, 1900 (Brachyura,
Ocypodidae) in a sub- tropical South America mangrove area: results from transect and catch-per-unit-
effort techniques. Crustaceana, 75: 1201-1218
Diesel R. 1989. Structure and function of the reproductive system of the symbiotic spider crab Inachus
phalangium (Decapoda: Majidae): Observations on sperm transfer, sperm storage and spawning. Journal of
Crustacean Biology, 9: 266-277
Dugan JE. 1990. Geographic and temporal variation in the lifehistory, growth and reproductive biology of the
sand crab, Emerita analoga (Stimpson). Ph.D. dissertation. University of California, Santa Barbara, USA
Dugan JE, Wenner A, Hubbard D. 1991. Geographic variation in the reproductive biology of the sand crab
Emerita analoga (Stimpson) on the California coast. Journal of Experimental Marine Biology and Ecology,
150: 63-81
Eickstaedt LL. 1969. The reproductive biology of the sand crab Emerita analoga (Stimpson). Ph.D.
Dissertation. Stanford University, Stanford, USA
Emmerson WD. 1994. Seasonal breeding cycles and sex ratios of eight species of crabs from Mgazana, a
mangrove estuary in Transkei, southern Africa. Journal of Crustacean Biology, 14: 568-578
123

Arthropods, 2013, 2(3): 111-125
IAEES www.iaees.org
Erdman RB, Blake NJ. 1988. Reproductive biology of female golden crabs Geryon fenneri Manning and
Holthuis, from souththeastern Florida. Journal of Crustacean Biology, 8: 392-400
Flores AVV, Paula J. 2002. Sexual maturity, larval release and reproductive output of two brachyuran crabs
from a rocky intertidal area in central Portugal. Invertebrate Reproduction and Development, 41: 21-34
Geisel JT. 1972. Sex ratio, rate of evolution, and environmental heterogeneity. American Naturalist, 106: 380-
387
George S. 1999. Egg quality, larval growth and phenotypic plasticity in a forcipulate seastar. Journal of
Experimental Marine Biology and Ecology, 237: 203-224
Gimé́nez L, Anger K. 2001. Relationships among salinity, egg size, embryonic development, and larval
biomass in the estuarine crab Chasmagnathus granulata Dana, 1851. Journal of Experimental Marine
Biology and Ecology, 260: 241-257
Gray GW Jr. 1969. Investigation of the Basic Life History of the Red Crab (Geryon quinquedens). Project 3-
46R Completion Report, Rhode Island Division of Conservation, USA
Guillory V, Prejean E, Bourgeois M, et al. 1996. A biological and fisheries profile of the blue crab, Callinectes
sapidus. LA. Department of Wildlife and Fisheries Management Plan Series, 8(1): 210
Hartnoll RG. 1985. Growth, sexual maturity and reproductive output. In: Factors in Adult Growth, Crustacean
Issues (Wenner AM, ed). 101-128, Balkema, Rotterdam, Netherlands
Hattori GY, Pinheiro MAA. 2001. Fecundity and embryology of Pachycheles milinifer (Dana, 1852)
(Anomura, Porcellanidae) at Praia Grande, Ubatuba, SP, Brazil. Nauplius, 9: 97-109
Hill J, Fowler DL, Van Den Avyle MJ. 1989. Species Profiles: Life Histories and Environmental
Requirements of the Coastal Fishes and Invertebrates (Mid-Atlantic)--Blue Crab. U.S. Fish and Wildlife
Service Biology Report 82 (11.100). U.S. Army Corps of Engineers, TR EL-82-4, USA
Hines AH. 1982. Allometric constraints and variables of reproductive effort in brachyuran crabs. Marine
Biology, 69: 309-320
Holland D. 1978. Lipid reserves and energy metabolism in the larvae of benthic marine invertebrates. In:
Biochemical and Biophysical Perspectives in Marine Biology (Vol. 4) (Malins D, Sargent J, eds). 85-123,
Academic Press, New York, USA
http://species-identification.org/species.php?species_group=crabs_of_japan&id=1656
Ituarte RB, Spivak ED, Luppi TA. 2004. Female reproductive cycle of the Southwestern Atlantic estuarine
crab Chasmagnathus granulatus (Brachyura: Grapsoidea: Varunidae). Scientia Marina, 68: 127-137
Jaeckle W. 1995. Variation in the size, energy content, and biochemical composition of invertebrate eggs:
correlates to the mode of larval development. In: Ecology of Marine Invertebrate Larvae (McEdward L,
ed). 49-77, CRC Press, Boca Raton, USA
Johnson PTJ. 2003. Biased sex ratios in fiddler crabs (Brachyura, Ocypodidae): a review and evaluation of the
influence of sampling method, size class and sex-specific mortality. Crustaceana, 76: 559-580
Koga T. 1998. Reproductive success and two modes of mating in the sand-bubbler crab Scopimera globosa.
Journal of Experimental Marine Biology and Ecology, 229: 197-207
Kumar M, Ferguson G, Xiao Y, et al. 2000. Studies on reproductive biology and distribution of blue swimmer
crab (Portunus pelagicus) in South Australian Waters. SARDI Research Report Series, 47
Levin L, Bridges T. 1995. Pattern and diversity in reproduction and development. In: Ecology of Marine
Invertebrate Larvae (McEdward L, ed). CRC Press, Boca Raton, USA
Litulo C. 2004. Fecundity of the Pantropical Fiddler Crab Uca annulipes (H. Milne Edwards, 1837)
(Brachyura: Ocypodidae) at Costa do Sol Mangrove, Maputo Bay, Southern Mozambique. WIOMSA, 3(1)
Litulo, C. 2005. Population structure and reproductive biology of the fiddler crab Uca inversa (Hoffman, 1874)
124

Arthropods, 2013, 2(3): 111-125
IAEES www.iaees.org
(Brachyura: Ocypodidae). Acta Oecologia, 27: 135-141
Lopez Greco LS, Hernandez JE, Bolanos J, et al. 2000. Population features of Microphrys bicornutus Latreille,
1825 (Brachyura, Majidae) from Isla Margarita,Venezuela. Hydrobiologia, 439: 151-159
Mantelatto F LM, Fransozo A. 1997. Fecundity of the crab Callinectes ornatus Ordway, 1863 (Decapoda,
Brachyura, Portunidae) from the Ubatuba region, São Paulo, Brazil. Crustaceana, 70: 214-224
Mantelatto FLM, FransozoA. 1996. Size at sexual maturity in Callinectes ornatus (Brachyura, Portunidae)
from the Ubatuba region (SP), Brazil. Nauplius, 4: 29-38
Melville-Smith R. 1987. The reproductive biology of Geryon maritae (Decapoda, Brachyura) off South West
Africa/Namibia. Crustaceana, 53: 259-275
Montague CL. 1980. A natural history of temperate western Atlantic fiddler crabs (Genus Uca) with reference
to their impact on the salt march. Contributions in Marine Science, 23: 25-55
Mouton ECJ, Felder DL. 1995. Reproduction of the fiddler crabs Uca longissignalis and Uca spinicarpa in a
Gulf of Mexico salt march. Estuaries, 18: 469-481
Nikolsky GV. 1963. The Ecology of Fishes. Academic Press, New York, USA
Ofori- Danson, P.K. 1990. Reproductive ecology of the trigger fish, Ballistes capriscus from the Ghanianain
coastal waters. Tropical Ecology, 31 (1): 1–11.
Ong KS. 1966. Observations on the post-larval life history of Scylla serrata Forskall, reared in the laboratory.
The Malaysian Agriculture Journal, 45: 429-443
Oshiro LMY. 1999. Aspectos reprodutivos do caranguejo guaia Menippe nodifrons Stimpson (Crustacea,
Decapoda, Xanthidae) da baia de Sepetiba, Rio de Janeiro, Brazil. Revista Brasileira de Zoologia, 16: 827-
834
Paschke K. 1998. Untersuchungen zum Energiestoffwechsel wa ̈hrend der Embryonalentwicklung der
Nordsee-Garnele Crangon crangon (Linnaeus 1758). Decapoda: Caridea.. PhD thesis. University of
Hamburg, Germany
Pauley GB, Amstrong DA, Heun TW. 1986. Species Profiles: Life histories and Environmental Requirements
of the Coastal Fishes and Invertebrates (Pacific-Northwest) Dungeness Crab. U.S. Fish and Wildlife
Service Biology Report 82 (11.63). U.S. Army Corps of Engineers, TR EL-82-4, USA
Ramirez Llodra E. 2002. Fecundity and life-history strategies in marine invertebrates. Advances in Marine
Biology, 43: 87-170
Sastry A. 1983. Ecological aspects of reproduction. In: The Biology of Crustacea (Vol. 8) (Bliss D, ed). 179-
269, Academic Press, New York, USA
Shields JD. 1992. The reproductive, ecology and fecundity of Cancer crabs. In: Crustacean Egg Production
(Wenner AM, Kurts AM, eds). Crustacean Issues 7. Balkema, Rotterdam, Netherlands
Spivak E, Gavio MA, Navarro CE. 1991. Life history and structure of the world’s southernmost Uca
population: Uca uruguayensis (Crustacea, Brachyura) in Mar Chiquita lagoon. Bulletin of Marine Science,
48: 679-688
Thurman, C.L., 1985. Evaporative water loss, corporal tempera- ture and the distribution of sympatric fiddler
crabs (Uca) from south Texas. Comparative. Biochemistry and Physiology A, 119: 279-286
Wenner AM, Hubbard DM, Dugan JE, et al. 1987. Egg production by sand crabs (Emerita analoga) as a
function of size and year class (Decapoda, Hippidae). Biological Bulletin, 172: 225-235
Wenner AM, Fusaro C, Oaten A. 1974. Size at onset of sexual maturity and growth rate in crustacean
populations. Canadian Journal of Zoology, 52: 1095-1106
Wolf PL, Shanholtzer SF, Reimold RJ. 1975. Population estimates for Uca pugnax (Smith, 1870) on the
Duplin estuary marsh, Georgia, U.S.A. (Decapoda, Brachyura, Ocypodidae). Crustaceana, 29: 79-91
125

Arthropods, 2013, 2(3): 126-136
IAEES www.iaees.org
Article
Biochemical properties of digestive carbohydrases from the sugar
beet weevil, Lixus incanescens (Coleoptera: Curculionidae)
Seyed Mohammad Ahsaei, Vahid Hosseininaveh, Mahdieh Bigham Department of Plant Protection, College of Agriculture, University of Tehran, Karaj 31587-77871, Iran
E-mail: [email protected],[email protected]
Received 22 April 2013; Accepted 25 May 2013; Published online 1 September 2013
Abstract
The sugar beet weevil, Lixus incanescens B., is one of the most important pests of sugar beet plant in Iran. The
petioles and leaves of sugar beet are attacked by larvae and adults of the sugar beet weevil. Chemical
application is currently used for controlling the pest. Digestion in the alimentary canal of the sugar beet weevil
is facilitated by some carbohydrases. Results of the in vitro studies indicated the presence of α-amylase, β-
glucosidase and β-galactosidase in the digestive tract of the pest. Highest activities of α-amylase, β-glucosidase
and β-galactosidase were at pH 5, pH 5 and pH 4, respectively. No significant α-glucosidase and α-
galactosidase activity was detected in the pest’s digestive system. Optimum temperatures for α-amylase, β-
glucosidase and β-galactosidase activity were determined at 45, 50 and 40 °C, respectively. α-amylase was
more stable under acidic condition (pH 4 to pH 6) than under highly acidic and alkaline condition. Na+ and K+
increased α-amylase activity, but sodium dodecyl sulfate significantly decreased amylase activity. Also, the
activity of α-amylase was inhibited by the other compounds such as MgCl2, CaCl2 and EDTA. Zymogram
analysis using native-PAGE revealed one band of α-amylase activity in Lixus incanescens. High activity of
carbohydrases in the digestive system of adults was determined and further researches are needed to be applied
to design new strategies for controlling the sugar beet weevil based on natural carbohydrase inhibitors.
Keywords α-amylases; sugar beet weevil; carbohydrases; Lixus incanescens.
1 Introduction
The sugar beet weevil, Lixus incanescens B. (Coleoptera: Curculionidae) is a key pest of sugar beet plants
causing significant damage to living plants in Iran. It has been reported from many parts of Iran and some
other countries (Davatchi et al., 1960; Aleeva, 1953). The sugar beet weevil has three generations per year.
The petioles and leaves of sugar beet plants are attacked by larvae and adults of L. incanescens. The larvae can
cause up to 75% root weight loss (Ocete et al., 1994). The most damage happens in the second and third
generations. Loss in early-planted sugar beet is less than late-planted ones (Parvizi et al., 1988). Currently,
Arthropods ISSN 22244255 URL: http://www.iaees.org/publications/journals/arthropods/onlineversion.asp RSS: http://www.iaees.org/publications/journals/arthropods/rss.xml Email: [email protected] EditorinChief: WenJun Zhang Publisher: International Academy of Ecology and Environmental Sciences

Arthropods, 2013, 2(3): 126-136
IAEES www.iaees.org
application of chemical pesticides has been a fundamental tool for pest control. However, it has had serious
consequences such as intoxication of animals and people, contamination of air, water, and soil, high
persistence in the environment, resistance in pests, and has impacted beneficial insects (Rodriguez et al., 1960;
Huignard et al., 2005). The use of such pesticides in urban areas holds special risks, as most pesticides are not
very selective, which has led to the search for safe and environmentally friendly alternatives (Breuer et al.,
2000). It seems that an integrated pest management (IPM) program, including application of selective
pesticides and some resistant cultivars bearing carbohydrase inhibitors as well as the use of field sanitation and
crop rotation would provide the best management option for control of this pest (Meiners et al., 1978).
Understanding the biochemistry of the enzymes present in the gut of target pests is the first step to design
inhibitor-transgenic crops (Oppert, 2000; Oppert et al., 2000; Wilhite et al., 2000). α-amylases (α-1,4-glucan-
4-glucanohydrolases; EC3.2.1.1) are the hydrolytic enzymes that there are in microorganisms, plants and
animals such as insects. α-amylases are hydrolytic enzymes that catalyze the hydrolysis of α-D-(1, 4) glucan
linkages in starch and related carbohydrates (Strobl et al., 1998; Ferreira et al., 1989). Α-amylases have been
detected in Coleoptera, Hymenoptera, Heteroptera, Orthoptera, Lepidoptera and Diptera (Hori, 1970;
Kanekatso, 1978; Baker, 1987; Baker, 1991; Terra et al., 1988; Schumaker et al., 1993; Ferreira et al., 1999).
Most insects depend on their amylases and glucosidases for efficient digestion of their diet. For both the
maintenance of adult longevity and optimal larval growth, carbohydrates are essential for initial hydrolyzing of
energy-producing nutrients (Dadd, 1985). α-glucosidases hydrolysis non-reducing 1,4-linked alpha-D-glucose
residues and releases α-D-glucose. α-glucosidase hydrolyzes several substrates, including maltose, sucrose ,
maltodextrin and pNP-α-Dglucopyranoside (Terra et al., 1996). β-glucosidases catalyzes hydrolysis of β-1,4
linkages between two glucoses or glucose-substituted molecules (Terra et al., 1996). α-galactosidases are exo-
acting glycoside hydrolases that cleave α-linked galactose residues from carbohydrates such as melibiose,
raffinose and gluco- or galactomannans (Meier et al., 1982). β-galactosidases are the enzyme that catalyses the
hydrolysis of β-galactosides into monosaccharides (Sezginturk et al., 2008). Our knowledge of β-
galactosidase in insects is still rudimentary.
The destructive effects of the pest L. incanescens on the production and handling of sugar beet plants have
made this insect an important target for biochemical study. There are significant variations among the
properties of insect digestive enzymes. To develop the control strategy utilizing plant-carbohydraceous
inhibitors, it is necessary to have more information on the gut enzymatic activities of insects (Wilhite et al.,
2000). In the current study, we have carried out a detailed biochemical analysis of digestive carbohydrolytic
activity in L. incanescens adults.
2 Materials and Methods
2.1 Test insect
Adults of the sugar beet weevil, Lixus incanescens, were collected from the fields of sugar beet plants at
Qazvin province, Iran and were used as the enzyme source for subsequent experiments.
2.2 Sample preparation
Whole gut of the adults were removed by dissection under a stereo-microscope in ice-cold saline buffer, and
homogenized in distilled water using a hand-held glass homogenizer. The homogenates were centrifuged at
16,000 × g for 15 min at 4°C. Resulted supernatants were removed and kept at -20 °C until needed.
2.3 α-amylase assay
α-amylase activity was assayed using dinitrosalicylic acid (DNSA) as the reagent (Strobl et al., 1998) and 1%
soluble starch (Merck) as the substrate. Ten microliters of the enzyme source was added to 5 μl substrate and
85 μl of the universal buffer (0.02 M; sodium citrate- phosphate-borate). The reaction mixture was incubated at
127

Arthropods, 2013, 2(3): 126-136
IAEES www.iaees.org
35°C for 30 min. Thereafter, 100 μl DNSA was added to the reaction mixture and heated in boiling water for
10 min. After cooling the mixture, the absorbance was measured at 540 nm. All assays were performed in
triplicate.
2.4 Glucosidase and galactosidase assay
Glucosidase and galactosidase assays was carried out by incubating 10 μl of the enzyme extract with 85 μl of
50 mM universal buffer (pH 4 to pH 11; sodium citrate-phosphate-borate) and 5 μl of each pnitrophenyl- α-D-
glucopyranoside (pNPαGlu, 5 mM) for α-glucosidase or p-nitrophenyl-β-D-glucopyranoside (pNPβGlu, 5 mM)
for β-glucosidase at 35 °C for 30 min. After the addition of NaOH (2 M) to the reaction mixture, released p-
nitrophenol was determined by measuring the absorbance at 405 nm. In the blanks, enzyme extract was added
to the reaction mixture after the addition of NaOH. The same procedure was used for detecting α- and β-
galactosidase activity but with p-nitrophenyl-α-D-galactosidase (pNPαGal, 5 mM) and p-nitrophenyl-β-D-
galactosidase (pNPβGal, 5 mM), respectively, as the substrates.
2.5 Effect of pH and temperature on enzyme activity
The effects of temperature and pH on the activity of α-amylase, β-glucosidases and β-galactosidase were
examined using enzymes extracted from the whole gut. Effect of temperature on α-amylase, β-glucosidases
and β-galactosidases activity was determined by incubating the reaction mixture at 20, 30, 35, 40, 45, 50, 60
and 70 °C for 30 min followed by measurement of the enzyme activity. The optimum pH for α-amylase, β-
glucosidases and β-galactosidase activity was determined using the universal buffer (sodium citrate-phosphate-
borate) at a pH range 3 to pH 11 with one degree interval.
2.6 pH stability of α-amylase
Stability of α-amylase was determined over a broad pH range and two incubation time periods. Enzyme extract
was mixed with buffer and incubated for 1 and 10 hr at 37°C. The substrate was then added to the buffered
enzyme extract, and α-amylase activity was determined as described above.
2.7 Effect of activators and inhibitors on α-amylase
To investigate the effect of several salts on α-amylase activity, assays were performed in the presence of
different concentrations of K+, Na+, Ca2+, and Mg2+ chloride salts as well as sodium dodecyl sulfate (SDS) and
Ethylenediaminetetraacetic acid (EDTA). The enzyme sample was pre-incubated with the compounds for 15
min.
2.8 Zymogram analysis
Activity of α-amylase present in the crude homogenates of the whole gut of the adults was visualized using
native polyacrylamide gel electrophoresis (native-PAGE). Native-PAGE was performed in a 10% (w/v)
resolving gel and a 4% stacking gel. The sample buffer contained 25% stacking buffer (0.5 M Tris–HCl; pH
6.8), 20% glycerol, 0.005% (w/v) bromophenol blue, but no mercaptoethanol and no boiling. After
electrophoresis, the gel was rinsed in distilled water and incubated for 1 h in phosphate buffer containing 1%
starch and 20 mM CaCl2 at pH 5. The gel was then rinsed with distilled water and incubated with a solution of
10 mM I2 and 14 mM KI to stop the reaction and to stain the unreacted starch background. Areas bearing α-
amylase activity appeared as light bands against a dark background.
2.9 Protein determination
Protein content was determined using the method of Lowry et al. (1951). Bovine serum albumin was used as
the standard.
3 Results
3.1 α-amylase activity
The effect of pH on amylolytic activity from the whole gut extracts is shown in Fig. 1. Relatively higher
128

Arthropods, 2013, 2(3): 126-136
IAEES www.iaees.org
activities were detected over a broad range of acidic condition (pH 4 to pH 6) with maximum activity at pH 5.
Fig. 1 The effect of pH on the activity of α-amylase extracted from the digestive system of L. incanescens adults.
Fig. 2 The effect of pH on the activity of β-glucosidases extracted from the digestive system of L. incanescens adults.
Fig. 3 The effect of pH on the activity of β-galactosidases extracted from the digestive system of L. incanescens adults.
129

Arthropods, 2013, 2(3): 126-136
IAEES www.iaees.org
3.2 Glucosidase activity
β-glucosidase activity in the gut increased steadily from pH 3 to pH 6, and then decreased with increased pH
above 6 (Fig. 2). The optimal pH for β-glucosidase activity occurred at 5.0. Activity for α–glucosidase was not
significantly detected in digestive system of L. incanescens adults.
3.3 Galactosidase activity
β-Galactosidase activity increased steadily from pH 3 to pH 4, then decreased with increased pH above 4. The
optimal pH for β-galactosidase activity occurred at pH 4.0 (Fig. 3). Activity for α–galactosidase was not
detected in digestive system of Lixus incanescens adults.
3.4 Effect of temperature on α-amylase activity
The optimum temperature for α-amylase activity was 45 °C (Fig. 4). Digestive β-glucosidase was optimally
active at 50 °C (Fig. 5). Optimum temperature for β-galactosidase activity was determined at 40 °C (Fig. 6).
Fig. 4 The effect of temperature on the activity of α-amylase extracted from the digestive system of L. incanescens adults.
Fig. 5 The effect of temperature on the activity of β-glucosidases extracted from the digestive system of L. incanescens adults.
130

Arthropods, 2013, 2(3): 126-136
IAEES www.iaees.org
Fig. 6 The effect of temperature on the activity of β-galactosidases extracted from the digestive system of L. incanescens adults.
Fig. 7 Stability of digestive α-amylase from L. incanescens at different pHs after 1 and 10 hrs incubation period.
3.5 pH stability of α-amylase
The results revealed that α-amylase was more stable in acidic to slightly acidic condition (pH 4 to pH 6) with a
short incubation period (1 hr; Fig. 7). However, α-amylase stability decreased under alkaline condition and a
longer time incubation period (10 hr; Fig. 7).
3.6 Effect of activators and inhibitors on α-amylase activity
Na+ and K+ enhanced α-amylase activity, whereas sodium dodecyl sulfate (SDS) significantly decreased α-
amylase activity. Activity of α-amylase was inhibited by other compounds such as MgCl2, CaCl2 and
ethylenediaminetetraacetic acid (EDTA) (Fig. 8).
3.7 Zymogram analysis
Further characterizations of the α-amylase activity of the gut extract from the adults of L. incanescens using
starch as the substrate under native-PAGE are shown in Fig. 9. At least one band of α-amylase activity was
revealed from the gut extract.
131

Arthropods, 2013, 2(3): 126-136
IAEES www.iaees.org
Fig. 8 Effects of different compounds on the midgut digestive amylase activity in L. incanescens adults.
Fig. 9 Zymogram of α- amylase extracted from the digestive system of L. incanescens.
132

Arthropods, 2013, 2(3): 126-136
IAEES www.iaees.org
4 Discussion
Carbohydrases play a vital role in food digestion in insects. The present study clearly showed that α-amylase,
β-glucosidase and β-galactosidase are active in the gut of L. incanescens adults. The enzymes are optimally
active in acidic condition which is congruent with the pH prevailing in the gut of Coleoptera. Stability of
amylase was also consistent with the pH value of the midgut and showed its maximum stability at acidic to
slightly acidic condition in L. incanescens. The optimal pHs for hemolymph and gut α-amylases of the red
palm weevil, Rhynchophorus ferrugineus Olivier (Col.: Curculionidae) were 5 to 6 and 4 to 5, respectively
(Saberi et al., 2012). The optimum pH for digestive α- amylase activity in the salivary glands and midgut of
Brachynema germari Kol. was determined at 6 and 5, respectively (Ramzi et al., 2010). In insects, amylases
are usually most active in neutral to slightly acidic condition (Baker, 1983; Terra et al, 1996). The optimal pH
values in larvae of various coleopterans were from 4 to 5.8 for amylases (Baker, 1983). The optimum activity
for α-amylase from the midgut of Hypera postica larvae was determined at pH 5.0 (Vatanparast et al., 2010).
Hypera postica α-amylase was more stable at pH 5 to pH 6 than highly alkaline and acidic pH in larval midgut
of the pest (Vatanparast et al., 2010). The pH in the tissues from which the enzymes are isolated usually
corresponds to the optimum pH for digestive enzyme activity. Biochemical properties of many partially
purified or purified insect gut α- glucosidases are known, and regardless of the corresponding midgut pH value,
their optimal pH range from 5 to 6.5 (Terra et al., 1994). Activity for α–glucosidase and α–galactosidase were
not detected in digestive system of Lixus incanescens adults. Insect β-glucosidases have pH optima of 4.5 to
6.5 (Terra et al, 1994; Azevedo et al, 2003). β-glucosidase activity has been reported in Diatraea saccharalis
Fabricius (Lepidoptera: Pyralidae) (Azevedo et al., 2003), Parnassius apollo ssp. frankenbergeri (Lepidoptera:
Papilionidae) (Nakonieczny et al, 2006). Also, digestive β-glucosidase activity has been reported in
Rhynchophorus palmarum L. (Coleoptera: Curculionidae) (Yapi et al., 2009). In Dysdercus peruvianus Guerin
(Hemiptera: Pyrrochoridae), galactosidases have an optimum pH of 5.0 (Silva et al., 1997). In Abracris
flavolineata De Geer (Orthoptera: Acrididae) midguts, there are two α-galactosidases and these enzyme have
maximum activity at pH 5.4 (Ferreira et al., 2001). β-galactosidase activity has been detected in Spodoptera
frugiperda (Marana et al., 2000) and Diatraea saccharalis, Tenebrio molitor (Franco et al., 2000). The highest
activities of α- and β-glucosidases of Rhynchophorus ferrugineus were at pH 5 and of α- and β-galactosidases
at pH 4 (Saberi et al., 2012). Data on the biochemical characterization of β-galactosidase in insects are not
complete.
Maximum carbohyrolytic activity was obtained at 40 to 50 °C congruent with the other insects. Higher gut
α-amylase activity of the red palm weevil was obtained at 40 to 50 °C (Saberi et al., 2012). Digestive α-
amylase from H. postica showed its maximal activity at 35 °C (Vatanparast et al., 2010). For α-amylase
activity, optimum temperature was 60 °C in Bombyx mori L. (Lepidoptera: Bombycidae) (Kanekatso, 1978).
In midgut of Brachynema germari Kol. (Hemiptera: Pentatomidae) optimum temperature for α- and β-
galactosidase was 30 °C (Ramzi et al., 2010). The sugar beet weevil β-glucosidase has optimum temperature
activity at 50 °C (Fig. 5). Also, the optimal temperature for β-galactosidase in the digestive system this pest
was 40 °C (Fig. 6). Alpha and β-glucosidases and α- and β-galactosidases from the red palm weevil had an
optimum temperature activity at 50, 50–60, 40–60, and 40 °C, respectively (Saberi et al., 2012). Activity of
digestive α- and β-glucosidases٫ α- and β-galactosidases in the digestive system of Xanthogaleruca luteola
were optimum at 60, 50, 40, 60 °C , respectively (Sharifi et al., 2011).
In the presence of Ca2+, Mg2+ and SDS, digestive α-amylase activity of Hypera postica significantly
decreased, while Na+ and K+ did not significantly affect the enzyme activity (Vatanparast et al., 2010).
Zymogram analysis showed the presence of one band of α-amylase activity in the gut of L. incanescens similar
to other coleopteran pests (Vatanparast et al., 2010). However, zymogram analysis revealed that red palm
133

Arthropods, 2013, 2(3): 126-136
IAEES www.iaees.org
weevil hemolymph and gut α-amylases had two isoforms (Saberi et al., 2012).
Producing pest-resistant transgenic plants help us to decrease the use of chemical pesticides and the
entering of these chemicals in the environment. Understand digestive enzymes of the target insect is first step
in designing a controlling program based on inhibition of digestion (Strobl et al., 1998). The biochemical
characterization of insect digestive enzymes will greatly help to design new strategies for insect control and
will facilitate the understanding of the mechanisms responsible for the inhibitory potential of the
carbohydraceous plant inhibitors.
Acknowledgements
The authors appreciate Seyed Mohammad Tabadkani and Alireza Badieneshin for their great helps with this
work. The authors have no conflict of interests to declare.
References
Aleeva MN.1953. Data on the biology of weevils (Col.: Curculionidae) injurious to sugar beet in Kazakhstan.
Ent Obozon, 33: 103-108 (In Russian)
Azevedo TM, Terra WR, Ferreira C.2003. Purification and characterization of three β-glycosidases from
midgut of the sugar cane borer, Diathraea saccharalis. Insect Biochemistry and Molecular Biology, 33:
81-92
Baker JE.1983. Properties of amylase from midgets of larvae of Sitophilus zeamais and Sitophilus granaries.
Insect Biochemistry, 13: 421-428
Baker JE.1987. Purification of isoamylases from the rice weevil, Sitophilus oryzae (L.) (Coleoptera:
Curculionidae) by high-performance liquid chromatography and their interaction with partiallypurified
amylase inhibitors from wheat. Insect Biochemistry, 17: 37-44
Baker JE.1991. Purification and partial characterization of α-amylase allozymes from the lesser grain borer,
Rhyzopertha dominica. Insect Biochemistry, 21: 303-311
Breuer M, De loof A. 2000. Laboratory trials with NeemAzal-T/S on the allergenic forest pest Thaumetopoea
processionea L. In: Practice Oriented Results on Use and Production of Neem-ingredients and Pheromones
VIII (Kleeberg H, Zebitz CPW, eds). 23-29, Hohensolms, Germany
Dadd RH. 1985. Nutrition: organisms. In: Comprehensive Insect Physiology Biochemistry and Pharmacology
8 (Kerkut GA, Gilbert LI, eds). 338-341, Pergamon Press, Oxford, UK
Davatchi A, Kheyri M. 1960. The Most Important Pest of Sugar Beet and Their Control. The Plan
Organization, Tehran, Iran
Ferreira C, Terra WR. 1989. Spatial organization of digestion, secretory mechanisms and digestive enzyme
properties in Pheropsophus aequinoctialis (Coleoptera: Carabidae). Insect Biochemistry, 19: 383-391
Ferreira C, Marana SR, Silva C, et al. 1999. Properties of digestive glycosidases and peptidases and the
permeability of the peritrophic membranes of Abracris flavolineata (Orthoptera: Acrididae). Comparative
Biochemistry and Physiology B, 123: 241-250
Ferreira AHP, Marana SR, Terra WR, et al. 2001. Purification, molecular cloning, and properties of a β-
glycosidase isolated from midgut lumen of Tenebrio molitor (Coleoptera) larvae. Insect Biochemistry and
Molecular Biology, 31: 1065-1076
Franco OL, Riggen DJ, Melo FR, et al. 2000. Activity of wheat α-amylase inhibitors towards bruchid α-
amylases and structural explanation of observed specificities. European Journal of Biochemistry, 267:
2166-2173
134

Arthropods, 2013, 2(3): 126-136
IAEES www.iaees.org
Hori K. 1970. Some properties of amylase in the salivary gland of Lygus disponsi (Hemiptera). Journal of
Insect Physiology, 16: 373-386
Huignard J, Dugravot S, Ketoh KG, et al. 2005. Use of secondary plant products to protect the seeds of a
legume, cowpea: Effects on insect pests and their parasitoids. In: Biopesticides of Plant Origin (Regnault-
Roger C, Philogène B, Vincent C eds). 123-137, Lavoisier, Paris, France
Kanekatso R. 1978. Studies on further properties for an alkaline amylase in the digestive juice of the silkworm,
Bombyx mori. Journal of Fac. Text. Science and Technology, 76: 1-21
Lowry OH, Rosembrough NJ, Farr AL, Randdall RJ.1951. Protein measurement with the Folin phenol reagent.
Journal of Biological Chemistry, 193: 267-275
Marana SR, Terra WR, Ferreira C. 2000. Purification and properties of a β-glycosidase purified from midgut
cells of Spodoptera frugiperda (Lepidoptera) larvae. Insect Biochemistry and Molecular Biology, 30:
1139-1146
Meier H, Reid JSG. 1982. Reserve polysaccharides other than starch in higher plants. In: Encyclopedia of
Plant Physiology (Loewus FA, Tanner W, eds). 418-471, Springer, New York, USA
Meiners JP, Elden TC. 1978. Resistance to insects and diseases in Phaseolus. In: Advances in Legume Science.
International Legume Conference (Summerfield RS, Bunting AH, eds). 359-364, Kew Surrey, UK
Nakonieczny M, Michalczyk K, Kedziorski A. 2006. Midgut glycosidases activities in monophagous larvae of
Apollo butterfly, Parnassius apollo ssp. frankenbergeri. C. R. Biology, 329: 765-774
Ocete R, Ocete ME, Perez-izquierdo MA , et al. 1994. Approximation to the phenology of Lixus junci B. (Col.:
Curculionidae) in La Rioja Alta: estimate of the damage it causes. Boletín de Sanidad Vegetal Plagas, 20:
611-616
Oppert B. 2000. Transgenic plants expressing enzyme inhibitors and the prospects for biopesticide
development. In: Advances in Biopesticide Research (Koul O, Dhaliwal GS, eds). 83-95, Hardwood
Academic, Amsterdam, Netherlands
Oppert B, Hartzer K, Smith CM.2000. Digestive proteinases of alfalfa weevil, Hypera postica (Gyllenhal)
(Coleoptera; Curculionidae). Transactions of Kansas Academy of Science, 103(3-4): 99-110
Parvizi R, Javanmoghaddam H. 1988. Investigation on some biological features of the sugar beet weevil (Lixus
incanescens B.). Ent Phyt Appliq., 55(1-2): 1-8 (In Persian)
Ramzi S, Hosseininaveh V. 2010. Biochemical characterization of digestive α-amylase, α-glucosidase and β-
glucosidase in pistachio green stink bug, Brachynema germari Kolenati (Hemiptera: Pentatomidae).
Journal of Asia-Pacific Entomology, 13: 215-219
Rodriguez C, Silva G, Vendramim YJ. 2003. Insecticidas de origen vegetal. In: Bases Para el Manejo Racional
de Insecticidas (Silva G, Hepp R, eds). 89-111, Universidad de Concepción, Fac. Agron. Chillán, Chile
Saberi riseh N, Ghadamyari M. 2012. Biochemical characterization of α-amylases from gut and hemolymph of
Rhynchophorus ferrugineus Olivieri (Col.: Curculionidae) and their inhibition by extracts from the
legumes Vigna radiata L. and Phaseolus vulgaris L. Research Report ISJ, 9: 72-81
Saberi riseh N, Ghadamyari M, Motamediniya B. 2012.Biochemical Characterisation of α- and β-glucosidases
and α- and β-galactosidases from Red Palm Weevil, Rhynchophorus ferrugineus (Olivier) (Col.:
Curculionidae). Plant Protection Science, 2: 85-93
Schumaker TTS, Cristofoletti PT, Terra WR. 1993. Properties and compartmentalization of digestive
carbohydrases and proteases in Scaptotrigona bipunctata (Apidae: Meliponinae) larvae. Apidologie, 24: 3-
17
Sezginturk MK, Dinckaya E. 2008. β-Galactosidase monitoring by a biosensor based on Clark electrode: Its
optimization, characterization and application. Biosensors and Bioelectronics, 23: 1799-1804
135

Arthropods, 2013, 2(3): 126-136
IAEES www.iaees.org
Sharifi M, Ghadamyari M, Mahdavi M, et al. 2011. Biochemical characterization of digestive carbohyrases
from Xanthogaleruca luteola and inhibition of its a-amylase by inhibitors extracted from the common bean.
Archives of Biological Science Belgrade, 63(3): 705-716
Silva CP, Terra WR. 1997. Alpha-galactosidase activity in ingested seeds and in the midgut of Dysdercus
peruvianus (Hemiptera: Pyrrhocoridae). Archives of Insect Biochemistry and Physiology, 34: 443-460
Strobl S, Maskos K, Wiegand G, et al. 1998. A novel strategy for inhibition of α-amylases: yellow meal worm
α-amylase in complex with Ragi bifunctional inhibitor at 2.5 Å resolution. Structure, 6: 911-921
Terra WR, Espinoza-fuentes EP, Ferreira C. 1988. Midgut amylase, lysozyme, aminopeptidase, and trehalase
from larvae and adults of Musca domestica. Archives of Insect Biochemistry and Physiology, 9: 283-297
Terra WR, Ferreira C. 1994. Insect digestive enzymes: properties, compartmentalization and function.
Comparative Biochemistry and Physiology B, 109: 1-62
Terra WR, Cristofoletti T. 1996. Midgut proteinases in three divergent species of Coleoptera. Comparative
Biochemistry and Physiology B, 113: 725-730
Terra WR, Ferreira C, Jordan BP, et al. 1996. Digestive enzymes. In: Biology of the Insect Midgut (Lehane MJ,
Billingsley PF, eds). 153-194, Chapman & Hall, London, UK
Vatanparast M, Hosseininaveh V. 2010. Digestive amylase and pectinase activity in the larvae of alfalfa weevil
Hypera postica (Coleoptera: Curculionidae). Entomological Research, 40: 328-335
Wilhite SE, Elden TC, Brzin J, et al. 2000. Inhibition of cysteine and aspartyl proteinases in the alfalfa weevil
midgut with biochemical and plant-derived proteinase inhibitors. Insect Biochemistry and Molecular
Biology, 30: 1181-1188
Yapi DYA, Gnakri D, Niamke SL, et al. 2009. Purification and biochemical characterization of a specific β-
glucosidase from the digestive fluid of larvae of the palm weevil, Rhynchophorus palmarum. Journal of
Insect Science, 9: 4
136

Arthropods, 2013, 2(3): 137-149
IAEES www.iaees.org
Article
Induced plant resistance as a pest management tactic on piercing
sucking insects of sesame crop
M. F. Mahmoud Plant Protection Department, Faculty of Agriculture, Suez Canal University, 41522 Ismailia, Egypt
E-mail: [email protected]
Received 5 April 2013; Accepted 10 May 2013; Published online 1 September 2013
Abstract
Sesame, Sesamum indicum L. is the most oil seed crop of the world and also a major oil seed crop of Egypt.
One of the major constraints in its production the damage caused by insect pests, particularly sucking insects
which suck the cell sap from leaves, flowers and capsules. Impact of three levels of potassin-F, salicylic acid
and combination between them on reduction infestation of Stink bug Nezara viridula L., Mirid bug
Creontiades sp., Green peach aphid Myzus persicae (Sulzer), Leafhopper Empoasca lybica de Berg and
Whitefly Bemisia tabaci (Gennadius) of sesame crop cultivar Shandawil 3 was carried out during 2010-2011
crop season at Experimental farm, Faculty of Agriculture, Suez Canal University, Ismailia, Egypt. Also, the
impacts of potassin-F and salicylic acid on yield production of sesame were studied. Results indicated that
percent of reduction of infestation by N. viridula, M. persicae, Creontiades sp., E. lybicae, B. tabaci and
phyllody disease were significantly higher at Level 2 (Potassin-F= 2.5 cm/l, Salicylic acid= 10−3 M and
Potassin + Salicylic= 2.5 cm/l + 10−3 M) and consequently higher seed yield per plant were obtained.
Keywords Sesamum indicum; potassin-F; salicylic acid; induced resistance; piercing sucking insects.
1 Introduction
Sesame (Sesamum indicum L.) is one of the important oil crops. It is cultivated in almost all tropical and sub
tropical Asia and African countries for its highly nutritious and edible seeds (Iwo et al., 2002). In Egypt,
sesame is considering a food crop rather than oilseed crops because most of its seeds consumed directly. It is
clear that the increase in sesame production during last ten years was mainly due to the increase in its growing
area, especially in newly reclaimed sandy soils.
Among various factors responsible for the low productivity levels of sesame, the insect pests associated
with flowering phase usually inflicts very severe damage to the crop. The physical damage may be less than
those of foliage pests, yet their impact on final yield is colossal.
Piercing sucking insects have great economic importance to sesame plants. They cause serious damage
Arthropods ISSN 22244255 URL: http://www.iaees.org/publications/journals/arthropods/onlineversion.asp RSS: http://www.iaees.org/publications/journals/arthropods/rss.xml Email: [email protected] EditorinChief: WenJun Zhang Publisher: International Academy of Ecology and Environmental Sciences

Arthropods, 2013, 2(3): 137-149
IAEES www.iaees.org
directly by sucking plant sap or indirectly by transmission of virus and mycoplasma diseases (El-Gindy, 2002).
Stink bug Nezara viridula L., Mirid bug Creontiades sp., Green peach aphid Myzus persicae (Sulzer),
Leafhopper Empoasca lybica de Berg and Whitefly Bemisia tabaci (Gennadius) are serious pests which suck
the cell sap from leaves, flowers and capsules. Due to this curling of leaf margins downward, reddening of leaf
margins, stunted growth of the plants, sickly appearance of the crop and subnormal growth of the leaf tissue
occur. The peculiar yellow spots are found on upper surface of leaves affected by whitefly. Jassid and whitefly
are also responsible to transmit phyllody and leaf curl diseases in sesame, respectively. Induced plant
resistance is very important component of integrated pest management programs, including adequate inorganic
fertilization kinds and amounts. Of all the nutrients that affect plant diseases and pests, potassium (K) is
probably the most effective. It has been considered a key component of plant nutrition that significantly
influences crop growth and some pests infestation. There are limited information about results of potassium
fertilization on reduction population of piercing sucking insects of sesame. Potassium fertilizer is negatively
associated with occurrence of Aphis glycines (Myers and Gratton, 2006) and leafhoppers and mites (Parihar
and Upadhyay, 2001). Cotton aphid population density at seedling stage was suppressed by potassium
fertilizers in proper rate (Ai et al., 2011). The high rates of potassium reduced the population density of some
piercing sucking insects on cereal, legumes and maize plants (El-Gindy, 2002, 2006).
Inducible defenses play a major role in conferring disease resistance against plant pathogens (Maleck and
Dietrich, 1999) and their effects on phytophagous insects can include increased toxicity, delay of larval
development or increased attack by insect parasitoids (Baldwin and Preston, 1999). Inducible defenses are
thought to compromise plant fitness less and may be more durable than constitutive defense mechanisms
(Agrawal, 1998). The plant nutrient status in an indicator of host plant quality plays an important role in the
population dynamics of many herbivores (Sarwar et al., 2011). Salicylic acid has been shown to have a
positive effect on some species of plants with regards to expression of dormant genes. However, no work has
yet been done to assess the level of insect attacks on the sesame plants treated with salicylic acid. Considering
the fact that some substances in association with crops under particular conditions can induce the presence of
both major and minor pests and diseases at different stages of development, the level of sucking insect
infestations on sesame in association with salicylic acid under our prevalent conditions must be properly
studied and understood. The concept of integrated pest management has taken centre stage in pest and diseases
management on a wide range of crops. This approach includes an integration of cultural, biological, chemical
and host plant resistance methods in controlling pests. Therefore, the purpose of this research was to
investigate the impact of three levels of potassin-F fertilizer and salicylic acid on reduction densities and
infestation of piercing sucking insects attacking sesame plants within two successive seasons, and to
investigate the impact of these treatments on yield production of sesame crop. Finding out the effect of
treatments on the pest load and damage can help update the control strategy of sucking insects on sesame.
2 Materials and Methods
2.1 Experimental design
This experiment was carried out at the Experimental farm, Faculty of Agriculture, Suez Canal University,
Ismailia, Egypt during the growing seasons of sesame (2010 and 2011). The soil texture of experimental site
was sandy soil (94.5% sand, 2.5% silt and 3.0% clay) with pH of 7.8.
Seeds of cultivar of sesame namely (Shandawil 3) were purchased from the Agriculture Research Center,
Giza, Egypt. These seeds were treated with Rizolex-T (3 g/kg seeds) before planting to prevent rot infection. A
randomized complete block experimental design was used and each experimental unit measuring an area of 3.5
× 4 m2. Three treatments were used at three levels and control.
138

Arthropods, 2013, 2(3): 137-149
IAEES www.iaees.org
Table 1 Potassin-F and salicylic treatment levels used in the present study.
Levels Treatments Potassin –F (P) (0 N: 8P: 30K)
Salicylic Acid (S)
*P + S
Control Level 1 Level 2 Level 3
0.0 1cm/l 2.5 cm/l 4 cm/l
0.0 10 −2 M 10 −3 M 10 −4 M
0.0 1cm/l + 10 −2 M 2.5 cm/l + 10 −3 M 4 cm/l +10 −4 M
* P= Potassin- F foliar application, S= Salicylic foliar application.
2.2 Sampling of studied insects
2.2.1 Jassid and aphid
25 sesame leaves were sampled weekly from each plot early in the morning from three different levels of the
plant. The upper and lower surfaces of the randomly chosen sesame leaves were carefully examined using lens
(5×) to count all individuals of Empoasca lybica and Myzus persicae and the data were recorded (El-Zahi et
al., 2012).
2.2.2 Whitefly
25 sesame leaves were randomly chosen from each plot and picked up weekly from the levels mentioned
above. The chosen leaves were transmitted to the laboratory in paper bags where binocular-microscope was
used to count the immature stages of B. tabaci (nymphs and pupae). The duration of sampling as mentioned
before.
2.2.3 Stink and mirid bugs
Sampling methods were made directly in the field on five randomly selected plants from each plot. All stages
were recorded. The reduction percentage of insect infestations was calculated for the seasons of 2010 and 2011.
2.3 Treatments
Sesame plants were sprayed with three levels of potassin–F and salicylic acid separately and together.
Potassin-F (P) was purchased from the Agriculture Research Center, Giza and Salicylic acid (S) was initially
dissolved in a few drops of dimethylsulfoxide and the final volume was reached using water. The
concentrations of (S) with a surfactant triton 0.1% and concentrations of (P) were sprayed twice on the whole
foliage of sesame (after 30 days from sowing and at the initiation of flowering stage). One set served as control
was sprayed with water only. The plants were sprayed with hand spray bottles10 liter capacity.
2.4 Effect of potassin-F fertilizer (P) and salicylic acid (S) on sesame production
Some parameters were measured as follows: no. of capsule/ plant, capsule weight, no. of seed/capsule, weight
of 1000 seed and seed yield/plant of sesame (Dhurve, 2008).
1- Number of capsule per plant: the number of capsule on plant was counted and expressed as capsule per
plant. Observation was made on 10 randomly selected plants per treatment.
2- Capsule weight: weight of 25 randomly selected capsules from each treatment was measured before the
harvest using an electronic balance.
3- Number of seeds per capsule: the number of seeds in a capsule was counted and expressed as seeds per
capsule. Observation was made on 10 randomly selected plants per treatment.
4- Weight of 1000 seeds: the observation was made by weighing 1000 dried seeds drawn randomly from
each treatment using an electronic balance.
5- Seed yield per plant: after maturation, the capsules from ten plants from each treatment were removed
and recorded. The seeds were separated and weight using an electronic balance and expressed in grams per
plant. Each treatment was replicated four times.
139

Arthropods, 2013, 2(3): 137-149
IAEES www.iaees.org
2.5 Statistical analysis
Data obtained in all presented experiments were subjected to analysis of variance (ANOVA) and correlation
coefficient with the honestly significant difference value calculated as Tukey’s statistic at α = 0.05 (SAS
Institute, 2002).
3 Results
Results from Fig. 1 showed five piercing sucking insects attacked sesame during growing season 2010/2011
and phyllody and healthy plant of sesame. Data presented in Fig. 2 and 3 showed the effect of three levels of
potassin-F, salicylic acid and the combination between them on population density of some piercing sucking
insects and the mean number of phyllody disease for sesame cultvar shandawil 3, during the growing season of
2010 and 2011. Data in Fig. 2 and 3 revealed that the three levels of all treatments led to reduction of
population density of jassid (Empoasca lybica), aphid (Myzus persicae), stink bug (Nezara virdula), whitefly
(Bemesia tabaci), mirid bug (Creontiades sp.) and the mean number of phyllody disease. Also, data in Fig. 2
and 3 indicated that Level 2= (Potassin-F= 2.5 cm/l, Salicylic acid= 10−3 M and Potassin + Salicylic= 2.5 cm/l
+ 10−3 M) was the more affect on reduction of population density than level 1 and level 3. Moreover, the
combination of potassin-F and salicylic acid in level 2 was the most suitable treatment to reduce population of
insects and phyllody disease.
The relationship between the reduction of infestation, % and the three levels of treatments potassin-F,
salicylic acid and the combination between them for the growing seasons of sesame, 2010 and 2011 were
computed as a coefficient of determination based on the simple linear regression Fig. 4 and 5.
Data presented in Fig. 4 indicated that level 2 of treatments (P= 2.5 cm/l, S= 10−3 M and P+S= 2.5 cm/l +
10−3 M) gave high reduction of infestation for the following insects, B. tabaci, N. viridula, and M. persicae.
The coefficient of determination (R2) was 0.993, 0.931 and 0.1, respectively. Whenever, level 1 of treatments
(P= 1 cm/l, S= 10−2 M and P+S= 1 cm/l + 10−2 M) gave high reduction of infestation of E. lybica and
Creontiades sp. R2 was 0.923 and 0.750, respectively. Finally, the level 3 (P= 4 cm/l, S= 10−4 M and P+S= 4
cm/l + 10−4 M) gave high reduction of infestation only in phyllody disease at R2 = 0.999.
Also, data presented in Fig. 5 showed that level 2 of treatments (P= 2.5 cm/l, S= 10−3 M and P+S= 2.5
cm/l + 10−3 M) gave high influence on the reduction of infestation,% for the following insects, E. lybica, N.
viridula, Creontiades sp. and M. persicae. The coefficient of determination (R2) was 0.993, 0.931, 0.1 and
0.964, respectively. While, level 3 of treatments (P= 4 cm/l, S= 10−4 M and P+S= 4 cm/l + 10−4 M) gave high
reduction of infestation of B. tabaci and phyllody disease. Results further proved that R2 for B. tabaci and
phyllody disease in the infestation of reduction was very high at values of 0.964 and 0.750, respectively.
140

Arthropods, 2013, 2(3): 137-149
IAEES www.iaees.org
Fig. 1 Some piercing sucking insects attacked sesame during growing season 2010/2011, 1- Empoasca lybica, 2- Bemesia tabaci, 3- Creontiades sp., 4- Myzus persicae, 5a- Nezara virdula eggs, 5b- N. virdula nymphs, 5c, N. viridula adults, 6- phyllody disease, 7- phyllody and healthy plant of sesame.
3
1 2
5 a 5 b 5 c
6 7
4
141
Arthropods, 2013, 2(3): 137-149
IAEES www.iaees.org
05
10152025
303540
4550
C P S P+S C P S P+S C P S P+S
Level 1 Level 2 Level 3
Treatments
Mea
n n
o. o
f ja
ssid
/25
leav
es
0
20
40
60
80
100
120
140
160
C P S P+S C P S P+S C P S P+S
Level 1 Level 2 Level 3
Treatments
Mea
n n
o. o
f ap
hid
/ 25
leav
es
0
10
20
30
40
50
60
C P S P+S C P S P+S C P S P+S
Level 1 Level 2 Level 3
Treatments
Mea
n n
o. o
f st
ink
bu
g /
5 p
lan
ts
0
10
20
30
40
50
60
70
C P S P+S C P S P+S C P S P+S
Level 1 Level 2 Level 3
Treatments
Mea
n n
o. o
f w
hit
efly
/ 25
leav
es
02468
101214161820
C P S P+S C P S P+S C P S P+S
Level 1 Level 2 Level 3
Treatments
Mea
n n
o. o
f m
irid
bu
g /
5 p
lan
ts
02468
1012141618
C P S P+S C P S P+S C P S P+S
Level 1 Level 2 Level 3
Treatments
Mea
n n
o. o
f p
hyl
lod
y / 1
00p
lan
ts
Fig. 2 Effect of three levels of potassin-F, Salicylic acid and the combination between them on the mean number of some piercing sucking insects which attack sesame and the mean number of phyllody disease during the growing season of 2010. C= Control, P= Potassin-F, S= Salicylic and P+S= Potassin-F + Salicylic; Level 1= (P= 1 cm/l, S= 10−2 M and P+S= 1 cm/l + 10−2 M); Level 2= (P= 2.5 cm/l, S= 10−3 M and P+S= 2.5 cm/l + 10−3 M); Level 3= (P= 4 cm/l, S= 10−4 M and P+S= 4 cm/l + 10−4 M).
142

Arthropods, 2013, 2(3): 137-149
IAEES www.iaees.org
0
10
20
30
40
50
C P S P+S C P S P+S C P S P+S
Level 1 Level 2 Level 3
Treatments
Mea
n n
o. o
f ja
ssid
/ 25
le
aves
0
20
40
60
80
100
120
C P S P+S C P S P+S C P S P+S
Level 1 Level 2 Level 3
Treatments
Mea
n no
. of
aphi
d / 2
5 le
aves
0
10
20
30
40
50
C P S P+S C P S P+S C P S P+S
Level 1 Level 2 Level 3
Treatments
Mea
n n
o. o
f st
ink
bu
g / 5
p
lan
ts
0
10
20
30
40
50
60
C P S P+S C P S P+S C P S P+S
Level 1 Level 2 Level 3
Treatments
Mea
n n
o. o
f w
hit
efly
/ 25
le
aves
02468
1012141618
C P S P+S C P S P+S C P S P+S
Level 1 Level 2 Level 3
Treatments
Mea
n n
o. o
f m
irid
bu
g / 5
p
lan
ts
0
5
10
15
20
25
C P S P+S C P S P+S C P S P+S
Level 1 Level 2 Level 3
Treatments
Mea
n n
o. o
f p
hyl
lod
y / 1
00
pla
nts
Fig. 3 Effect of three levels of potassin-F, Salicylic acid and the combination between them on the mean number of some piercing sucking insects which attack sesame and the mean number of phyllody disease during the growing season of 2011. C= Control, P= Potassin-F, S= Salicylic and P+S= Potassin-F + Salicylic; Level 1= (P= 1 cm/l, S= 10−2 M and P+S= 1 cm/l + 10−2 M); Level 2= (P= 2.5 cm/l, S= 10−3 M and P+S= 2.5 cm/l + 10−3 M); Level 3= (P= 4 cm/l, S= 10−4 M and P+S= 4 cm/l + 10−4 M).
143

Arthropods, 2013, 2(3): 137-149
IAEES www.iaees.org
Fig. 4 Relationship between reduction of infestation, % and different levels of potassin-F, salicylic acid and the combination between them in sesame during growing season of 2010.
Creontiades sp.
Myzus persicae
Nezara viridula Bemesia tabaci
Empoasca lybica
Phyllody
144

Arthropods, 2013, 2(3): 137-149
IAEES www.iaees.org
Fig. 5 Relationship between reduction of infestation, % and different levels of potassin-F, salicylic acid and the combination between them in sesame during growing season of 2011.
Creontiades sp.
Nezara viridula Bemisia tabaci
Empoasca lybica Myzus persicae
Phyllody
145

Arthropods, 2013, 2(3): 137-149
IAEES www.iaees.org
Results in Table 2 showed the influence of potassin-F, salicylic acid and the combination between them on
number of capsule/ plant, capsule weight, number of seed/capsule, weight of 1000 seed and seed yield (g)/
plant of sesame. Data showed significant differences in some quantitative parameters either in 2010 or 2011.
Number of capsule/ plant, capsule weight (g) and seed yield (g)/ plant in all treatments were significantly
higher compared to control in both growing seasons. In 2010 season (df= 4,35; F= 1.315; P≤ 0.0009), (df=
4,35; F= 6.381; P≤ 0.0001), (df= 4,35; F= 2.793; P≤ 0.0166), respectively. While in 2011 season (df= 4,35; F=
6.440; P≤ 0.0000), (df= 4,35; F= 4.234; P≤ 0.0013), (df= 4,35; F= 8.432; P≤ 0.0000), respectively. Number of
seed/ capsule was insignificant compared to control in both seasons. It was (df= 4,35; F= 1.315; P≤ 0.2701),
(df= 4,35; F= 1.917; P≤ 0.0878), respectively. On the other hand, weight of 1000 seed was not significant in
2010 season (df= 4,35; F= 2.271; P≤ 0.0445), and it was significant in 2011 season (df= 4,35; F= 2.708; P≤
0.0194).
The correlation coefficient (r) between seed yield (g)/ plant which produced from sesame plants treated
with different levels of potassin, salicylic and the combination between them and reduction of infestation of
different piercing sucking insects and phyllody disease was calculated of both studied seasons and presented in
Table 3. Data in this Table showed greatest direct effect on seed yield (g)/ plant produced from sesame plants
treated with potassin, with reduction of infestation of M. persicae, E. lybica, B. tabaci and N. viridula in 2011
season. It was 0.961, 0.522, 0.956 and 0.918, respectively. Also, results showed direct effect on seed yield (g)/
plant produced from sesame plants treated with salicylic, with reduction of infestation M. persicae, E. lybica
and N. viridula in 2010 season. It was 0.971, 0.642 and 0.988, respectively. And it was 0.537 with Creontiades
sp. in 2011 season. The strength of a relationship between seed yield (g)/ plant and reduction of phyllody
disease was high in both growing seasons of sesame especially with combined treatment (potassin + salicylic).
It was (0.581) in 2010 and (0.841) in 2011, respectively.
On the other hand, most of negative correlation has been recorded between seed yield (g)/ plant produced
from sesame plants treated with combined levels of potassin and salicylic and reduction infestation of M.
persicae, E. lybica, N. viridula and Creontiades sp.
Table 2 Influence of potassin-F, salicylic acid and the combination between them on no. of capsule/ plant, capsule weight, no. of seed/capsule, weight of 1000 seed and seed yield/plant of sesame.
Treatments No. of capsule/ plant Capsule weight (g) No. of seed/capsule Weight of 1000 seed
Seed yield (g)/plant
2010 2011 2010 2011 2010 2011 2010 2011 2010 2011 Control (0) 68.50 c 67.25 c 1.83 c 1.85 c 62.50 a 63.00 a 4.45 a 4.50 b 19.05 b 19.06 d Potassin- F
L1** 71.25 bc 71.75 bc 1.91 bc 2.07 abc 64.25 a 60.25 a 4.46 a 4.47 b 21.81 ab 19.26 cd
L2 77.75 abc 84.00 ab 2.00 abc 2.15 ab 68.00 a 61.00 a 4.51 a 4.52 b 23.85 ab 23.12 bcd
L3 7 7.75 abc 78.75 abc 2.08 ab 2.14 ab 66.00 a 62.50 a 4.47 a 4.57 ab 23.28 ab 22.54 bcd
Salicylic
L1 78.75 abc 75.75 abc 1.98 abc 1.99 bc 63.75 a 64.50 a 4.51 a 4.60 ab 22.53 ab 22.59 bcd
L2 81.75 a 87.75 a 2.05 ab 2.27 a 69.00 a 68.75 a 4.52 a 4.77 a 25.53 a 28.82 a
L3 81.25 a 81.50 ab 2.12 a 2.08 abc 70.00 a 68.25 a 4.58 a 4.65 ab 25.98 a 25.86 a Potassin- F + Salicylic
L1 77.25 abc 79.00 abc 2.05 ab 1.96 bc 65.00 a 62.75 a 4.57 a 4.60 ab 22.99 ab 22.83 bcd
L2 81.75 a 86.50 a 2.16 a 2.15 ab 63.25 a 64.75 a 4.50 a 4.57 ab 23.26 ab 25.59 ab
L3 79.75 ab 83.00 ab 2.10 ab 2.02 abc 64.00 a 63.50 a 4.53 a 4.57 ab 23.23 ab 24.08 abc
LSD 0.5 - 6.056 7.381 0.114 0.166 6.507 6.131 0.083 0.147 3.319 2.931
*Means followed by the same letter in a column are not statistically different by Tukey's HSD (P=0.05) ** Level 1= (P= 1 cm/l, S= 10−2 M and P+S= 1 cm/l + 10−2 M); Level 2= (P= 2.5 cm/l, S= 10−3 M and P+S= 2.5 cm/l + 10−3 M); Level 3= (P= 4 cm/l, S= 10−4 M and P+S= 4 cm/l + 10−4 M).
146

Arthropods, 2013, 2(3): 137-149
IAEES www.iaees.org
Table 3 Correlation coefficient (r) between seed yield (g)/plant produced from sesame plants treated with different levels of potassin-F, salicylic acid and the combination between them and reduction infestation of different piercing sucking insects and phyllody disease.
Seed yield (g)/plant
M. persicae E. lybica B. tabaci N. viridula
Creontiades sp.
Phyllody disease
2010 2011 2010 2011 2010 2011 2010 2011 2010 2011 2010 2011
Potassin-F 0.606 0.961 0.485 0.522 0.671 0.956 -0.949 0.918 -0.966 -0.986 -0.068 0.036
Salicylic 0.971 0.687 0.624 0.466 0.369 0.828 0.988 0.750 -0.039 0.537 0.150 -0.461
Potassin + Salicylic
-0.598 0.207 -0.547 0.281 0.561 0.281 -0.330 -0.047 -0.827 0.281 0.581 0.841
4 Discussion
In the present study, we applied potassin-F which consists of (0 Nitrogen: 8 Phosphorus: 30 Potassium). It is
well known that, potassium plays an important physiological roles including building up of resistance to insect
pests by influencing tissue of cell structures and biochemical processes. Potassium nutrition has a profound
effect on the profile and distribution of primary metabolites in plant tissues, which in turn could affect the
attractiveness of plant for insects and pathogens as well as their subsequent growth and development the plant
(Amtmann et al., 2008). A thicker cuticle in plants can be considered as a first line of defense to disease and
insect attack and increases the resistance to insect feeding, especially against sucking insects (Khattab, 2007).
Several studies conclusively indicate that phosphate is effective in controlling some important plant diseases
caused by pathogens. Action of phosphate anion is based on two mechanisms: the first is a direct toxic action
on the pathogen and the second in indirect action due to phosphite anion activates plant defence responses
(Cook et al., 2009; Avila et al., 2012).
Salicylic acid (SA), is a plant phenolic, is widely distributed throughout the plant kingdom. It is a
hormone-like substance, which plays an important role in the regulation many aspects of plant growth and
development (Raskin, 1992). However, it is especially famous for its ability to induce systemic acquired
resistance (SAR) in plants (Ryals et al., 1996) i.e. resistance in induced but also uninduced distal leaves of the
same plant.
Piercing sucking insects play an important role in reducing the production of sesame yield. The present
results revealed that sesame plants at the studied location was infested with Stink bug Nezara viridula L.,
Mirid bug Creontiades sp., Green peach aphid Myzus persicae (Sulzer), Leafhopper Empoasca lybica de Berg
and Whitefly Bemisia tabaci (Gennadius). These insects were recorded on sesame crop variety shandawil 3
during growing season of 2010 and 2011, Ismailia, Egypt (Mahmoud, 2012). Also, some of these pests were
recorded on potato plants by Parihar et al. (1996) and Kuroli (2001). On the other hand, M. percicae, A.
gossypii, A. craccivora, B. tabaci and Empoasca sp. populations were most at zero potassium fertilization in
comparison with 100, 150 and 200 kg/feddan, respectively (El-Gindy et al., 2009). Also, phyllody disease
plays a significant role in reducing yield of sesame. Leafhopper transmitted phytoplasma is the cause of
phyllody disease (Akhtar et al., 2008; Akhtar et al., 2009), which can cause up to 80% yield loss with disease
incidence of 61-80% (Kumar and Mishra, 1992).
Results in the present study confirmed significant effect of potassin-F fertilization on the insect infestations.
These findings are in agreement with those reported by (Kindler and Staples, 1970) who found that increasing
amount of potassium and phosphorus fertilizer, increased resistance against spotted alfalfa aphid in alfalfa,
green bug in sorghum (Schwessing and Wilde, 1979). Data in presented study indicated that Salicylic acid had
positive effects on plant growth, reduction infestation, % of M. persicae, B. tabaci, E. lybica, Creontiades sp.,
147

Arthropods, 2013, 2(3): 137-149
IAEES www.iaees.org
N. viridula, phyllody disease and yield components and hence could find roles to play in integrated pest
management on sesame plants.
In general, treatment of potassin-F and salicylic acid separately and together, especially at the level 2
clearly caused significant decrease in population density and percentage of infestation of N. viridula,
Creontiades sp., M. persicae, E. lybica and B. tabaci. Also, it caused reduction in the mean number of
phyllody disease during 2010 and 2011 crop seasons of sesame. Moreover, it caused significant increase in the
number of capsules on sesame stem, capsule weight (g), thousand seed wt (g) and seed yield/ plant (g).
Therefore, using potassin-F fertilization and salicylic acid is so necessary for encouraging the defense system
of plant to be more resistance or tolerant to insect injury caused by piercing sucking insects. Elden and
Kenworkthy (1995) found that the optimum or higher levels of potassium nutrition have been implicated with
decline in the incidence of disease and insect pests in several plant species.
In conclusion, potassin-F and salicylic acid provided good nutrition and resistance for the reduction of pest
load and damage and consequently, enhanced seed yield of sesame cultivar (Shandawil 3) in the reclaimed
land. Application of potassin-F (P) and salicylic acid (S) separately or combined at level 2 (P= 2.5 cm/l, S=
10−3 M and P+S= 2.5 cm/l + 10−3 M) provided the best nutrition for effective reduction of sesame pest load
and damage with increased seed production. The findings of this study also recommended that even though
application of potassin-F and salicylic acid at the level 2 (P= 2.5 cm/l, S= 10−3 M and P+S= 2.5 cm/l + 10−3 M)
reduced pest load and increased the quantity and quality of sesame, higher seed yield could be enhanced
through integrated pest management, especially if this rate is applied in conjunction with other cultural
practices such as crop rotation, inter-cropping, planting date manipulation, plant spacing and biological control.
References
Agrawal AA. 1998. Induced responses to herbivory and increased plant performance. Science, 279: 1201-1202.
Ai TC, Liu ZY, Li CR, et al. 2011. Impact of fertilization on cotton aphid population in Bt. cotton production
system. Ecological Complexity, 8: 9-14
Akhtar KP, Dickinson M, Sarwar G, et al. 2008. First report on the association of a 16SrII phytoplasma with
sesame phyllody in Pakistan. Plant Patholology, 57: 771
Akhtar KP, Sarwari G, Dickinson M, et al. 2009. Sesame phyllody disease: its symptomatology, etiology, and
transmission in Pakistan. Turkish Journal of Agriculture, 33: 477-486
Amtmann A, Troufflard S, Armengaud P. 2008. The effect of potassium nutrition on pest and disease
resistance in plants. Plant Physiology, 133: 682-691
Ávila FW, Faquin V, Lobato AKS, et al. 2012. Growth, phosphorus status, and nutritional aspect in common
bean exposed to different soil phosphate levels and foliar-applied phosphorus forms. Scientific Research
and Essays, 7(25): 2195-2204
Baldwin IT, Preston CA. 1999. The eco-physiological complexity of plant responses to insect herbivores.
Planta, 208: 137-145
Cook PJ, Landschoot PJ, Schlossberg MJ. 2009. Inhibition of Pythium spp. and suppression of Pythium Blight
of turfgrasses with phosphonate fungicides. Plant Disease, 93(8): 809-814
Dhurve SS. 2008. Impact of honey bee population on seed production of niger. MSc thesis. University of
Agriculture Sciences, Dharwad, India
Edlin TC, Kenworthy WJ. 1995. Physiological responses of an insect-resistance soubean line to light and
nutrient stress. Journal of Economic Entomology, 88(2): 430-436
El-Gindy MA, El-Refaey RM, Abou Hatab EE. 2009. Abundance of some potato homopterous pests as
148

Arthropods, 2013, 2(3): 137-149
IAEES www.iaees.org
affected by potassium fertilization level. Egyptian Academic Journal of Biological Sciences, 2(2): 179-185
El-Gindy MA. 2002. Studies on certain homopterous insect vectors of plant pathogenic diseases. PhD thesis.
Faculty of Agriculture, Zagazig University, Zagazig, Egypt
El-Gindy MA. 2006. Susceptibility of three maize cultivars to leaf hopper infestations and effect of potassium
fertilizers on leaf hoppers. Egyptian Journal of Applied Sciences, 21(10A): 302-314
El-Zahi ES, Arif SA, Jehan BA, et al. 2012. Inorganic fertilization of cotton field-plants in relation to sucking
insects and yield production components of cotton plants. Journal of American Science, 8(2): 509-517
Iwo GA, Idowu AA, Ochigbo AA. 2002. Sesame genotypes for field stability and selection in Nigeria,
Nigerian Agricultural Journal, 33: 76-82
Khattab H. 2007. The Defense Mechanism of cabbage plant against phloem-sucking aphid (Brevicoryne
brassicae L.). Australian Journal of Basic and Applied Sciences, 1(1): 56-62
Kindler SA, Staples R. 1970. Nutrients and the reaction of two alfalfa clones to the spotted alfalfa aphid.
Journal of Economic Entomology, 63: 938-940
Kumar P, Mishra US. 1992. Diseases of Sesamum indicum in Rohilkhand: intensity and yield loss. Indian
Phytopathology, 45: 121-122
Kuroli G. 2001. Change of population density of the leafhoppers Empoasca solani Curtis and E. decipiens
Paoli feeding on potatoes. Növényvédelem, 37: 225-230
Mahmoud MF. 2012. Insects associated with sesame (Sesamum indicum L.) and the impact of insect
pollinators on crop production. Pesticides and Phytomedicine, 27(2): 117-129
Maleck K, Dietrich RA. 1999. Defense on multiple fronts: how do plants cope with diverse enemies? Trends in
Plant Science, 4: 215-219
Myers SW, Gratton C. 2006. Influence of potassium fertility on soybean aphid, Aphid glycines Matsumura
(Hemiptera: Aphididae), population dynamics at a field and regional scale. Environmental Entomology, 35:
219-227
Parihar SBS, Upadhyay NC. 2001. Effect of fertilizers (NPK) on incidence of leafhoppers and mite in potato
crop. Insect Environment, 7: 10-11
Parihar SBS, Verma KD, Malik K. 1996. Evaluation of potato genotypes against Aphis gossypii damage, Insect
Environment, 2: 48-49
Raskin K. 1992. Role of salicylic acid in plants. Annual Review of Plant Physiology and Plant Molecular
Biology, 43: 439
Ryals JA, Neuenschwander UH, Willits MG, et al. 1996. Systemic acquired resistance. Plant Cell, 8: 1809-
1819
Sarwar M, Ahmad N, Tofique M. 2011. Impact of soil potassium on population buildup of aphid (Homoptera:
Aphididae) and crop yield in canola (Brassica napus L.) field. Pakistan Journal of Zoology, 43(1): 15-19
SAS Institute Inc. 2002. SAS/STAT User’s Guide (Version 9.1). SAS Institute Inc, North Carolina, USA
Schwessing FC, Wilde G. 1979. Temperature and plant nutrient effects on resistance of seedling sorghum to
the green bug. Journal of Economic Entomology, 72: 20-23
149

Arthropods
Arthropods account for more than 65% of global species and 85% of animal species. On a temperate grassland,
arthropods hold a huge biomass (1,000kg/ha), seconded to plant (20,000kg/ha) and microorganisms (7,000kg/ha)
but much higher than mammals (1.2kg/ha), birds (0.3kg/ha), and nemantodes (120kg/ha). Arthropods play the
role of both pests and beneficial organisms. Some arthropods are important crop pests but others are natural
enemies. Some arthropods are important health pests but many crustaceans are important food sources of
humankinds. Arthropods govern the structures and functions of natural ecosystems, but are always ignored by
researchers. On the global scale, the surveys of mammals, birds and vascular plants were relatively perfect
because they were economically important and easily surveyed. However, arthropods, despite their ecological
and economical importance, have not yet been fully surveyed and recorded due to their difficulties to be
sampled. The research on arthropods must be further promoted. The journal, Arthropods, is inaugurated to
provide a public and appropriate platform for the publication of studies and reports on arthropods.
ARTHROPODS (ISSN 2224-4255) is an international journal devoted to the publication of articles on various
aspects of arthropods, e.g., ecology, biogeography, systematics, biodiversity (species diversity, genetic diversity,
et al.), conservation, control, etc. The journal provides a forum for examining the importance of arthropods in
biosphere (both terrestrial and marine ecosystems) and human life in such fields as agriculture, forestry, fishery,
environmental management and human health. The scope of Arthropods is wide and embraces all
arthropods-insects, arachnids, crustaceans, centipedes, millipedes, and other arthropods. Articles/short
communications on new taxa (species, genus, families, orders, etc.) and new records of arthropods are
particularly welcome.
Authors can submit their works to the email box of this journal, [email protected]. All manuscripts
submitted to Arthropods must be previously unpublished and may not be considered for publication elsewhere
at any time during review period of this journal.
In addition to free submissions from authors around the world, special issues are also accepted. The organizer of
a special issue can collect submissions (yielded from a research project, a research group, etc.) on a specific
topic, or submissions of a conference for publication of special issue.
Editorial Office: [email protected]
Publisher: International Academy of Ecology and Environmental Sciences
Address: Flat C, 23/F, Lucky Plaza, 315-321 Lockhart Road, Wanchai, Hong Kong
Tel: 00852-6555 7188
Fax: 00852-3177 9906
E-mail: [email protected]

Arthropods ISSN 2224-4255 Volume 2, Number 3, 1 September 2013
Articles
The locomotory rhythmic activity in scorpions: with a review
Michael R. Warburg 95-104
Two records of Macrophthalmus Desmarest, 1823 (Decapoda: Brachyura: Thoracotremata)
from the NW of the Arabian Gulf
Amaal Gh. Yasser, Ibtisam M. AbdulSahib, Murtada D. Naser, et al. 105-110
Reproductive characteristics of a brachyuran crab, Grapsus tenuicrustatus (Herbst, 1783)
(Decapoda: Grapsidae) found in Talim Bay, Batangas, Philippines
Michael A. Clores, Gliceria B. Ramos 111-125
Biochemical properties of digestive carbohydrases from the sugar beet weevil, Lixus
incanescens (Coleoptera: Curculionidae)
Seyed Mohammad Ahsaei, Vahid Hosseininaveh, Mahdieh Bigham 126-136
Induced plant resistance as a pest management tactic on piercing sucking insects of sesame
crop
M. F. Mahmoud 137-149
The International Academy of Ecology and Environmental Sciences (IAEES) is a nonprofit and registered
international organization. It devotes to promote global ecology and environmental sciences and protect global
ecological environments, by publishing scientific publications, conducting research activities, launching
environmental programs, disseminating knowledge and technologies, sponsoring conferences, and providing
information and discussion spaces, etc. The responsibility for these publications rests with the International
Academy of Ecology and Environmental Sciences.
IAEES http://www.iaees.org/




















